◄ Carnets Geol. 21 (8) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Results and discussion]
[5. Conclusions] and ... [Bibliographic references]
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Corresponding author;
Research Center in Historical, Cultural and Natural Heritage;
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Departmento de Biología y Geología, Física y Química Inorgánica, ESCET, Universidad Rey Juan Carlos, c/Tulipán, s/n, 28933, Móstoles (Spain)
Departmento de Biología y Geología, Física y Química Inorgánica, ESCET, Universidad Rey Juan Carlos, c/Tulipán, s/n, 28933, Móstoles (Spain);
Instituto de Investigaciones Científicas y Tecnológicas (IDICTEC-UDA), Universidad de Atacama, Avenida Copayapu, 485, Copiapó (Chile)
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Research Center in Historical, Cultural and Natural Heritage;
Departmento de Ciencias de la Tierra, Universidad de Huelva, Avenida 3 de marzo, 21071 Huelva (Spain)
Published online in final form (pdf) on April 1, 2021
DOI 10.2110/carnets.2021.2108
![]()
[Editor: Bruno Granier;
topic editor: Francesco Sciuto; language editor:
Stephen Eagar]
![]()
During the Neogene, the Betic Strait was one of the gateways that connected the Atlantic Ocean and the Mediterranean Sea. In this paper, we have analyzed the ostracod faunas of samples collected from sediments crossed by a long borehole in southwestern Spain. These sediments were deposited in the Betic strait just before the Messinian Salinity Crisis. During the middle Messinian (6.8-6.0 Ma), the scarce and low diversified ostracod assemblages (Krithe, Parakrithe, Henryhowella) are typical of upper bathyal palaeoenvironments (200-400 m water depth). This period includes a short transition (6.26-6.25 Ma) to outer neritic palaeoenvironments, coinciding with a glaciation and characterized by the presence of Acanthocythereis hystrix (Reuss, 1850) and the disappearance of Krithe and Parakrithe. The most abundant species have a wide biostratigraphic distribution, most of them ranging from the Tortonian until the Holocene.
• Betic Strait;
• SW Spain;
• Messinian;
• upper bathyal-outer shelf ostracods;
• palaeoenvironmental evolution
Romero V., Ruiz F., González-RegaladoM.L., Tosquella J., Abad M., Izquierdo T., Toscano A. & Gómez P. (2021).- Messinian ostracodes from the western Betic Strait (SW Spain).- Carnets Geol., Madrid, vol. 21, no. 8, p. 181-192.
Ostracodes messiniens du Détroit Bétique occidental (Sud-Ouest de l'Espagne).- Au Néogčne, le Détroit Bétique est l'un des passages entre l'Océan Atlantique et la Mer Méditerranée. Dans cet article, nous analysons les faunes d'ostracodes provenant d'un forage réalisé dans le sud-ouest de l'Espagne et, plus précisément, situé sur le trajet du détroit. Ce forage a traversé des sédiments déposés au cours de la période immédiatement antérieure ŕ la crise de salinité messinienne. Au cours du Messinien moyen (6,8-6,0 Ma), les associations d'ostracodes, rares et peu diversifiés (Krithe, Parakrithe, Henryhowella), sont typiques de paléo-environnements bathyaux supérieurs (de 200 ŕ 400 m de profondeur). Cette période comprend une courte transition (6,26-6,25 Ma) jusqu'ŕ des paléo-environnements néritiques externes. Elle coďncide avec un épisode de glaciation et l'association est caractérisée par la présence d'Acanthocythereis hystrix (Reuss, 1850) et la disparition des genres Krithe et Parakrithe. Les espčces les plus fréquentes ont une large distribution biostratigraphique, la plupart étant présentes du Tortonien ŕ l'Holocčne.
• Détroit Bétique ;
• Sud-Ouest de l'Espagne ;
• Messinien ;
• ostracodes bathyaux-néritiques ;
• évolution du paléo-environnement
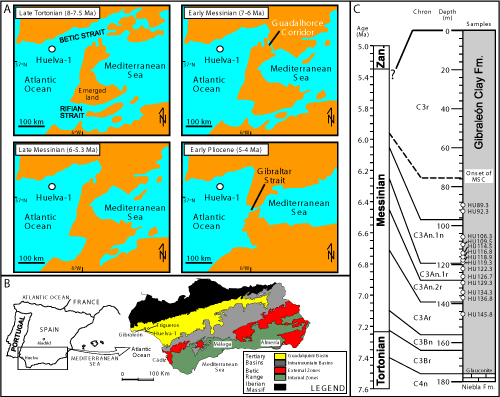
During the Tortonian and early Messinian, the Atlantic Ocean and the Mediterranean Sea were connected through the northern Betic Strait, which crossed the current Guadalquivir Basin (southern Spain), and the Rifian Corridor, located north of Morocco (Fig. 1.A ![]() ; Fleker et al., 2015). These connections were closed during the so-called Messinian Salinity Crisis (MSC; Hsü et al., 1973, 1977), with an onset dated at 5.97 Ma and a later partial isolation of the Mediterranean Sea (5.59-5.33 Ma; Krijgsman et al., 1999). These closures have been attributed to eustatic changes, climatic events or tectonic processes (Duggen et al., 2003; Leroux et al., 2018), although this discussion still persists (Vai, 2016; Sternai et al., 2017).
; Fleker et al., 2015). These connections were closed during the so-called Messinian Salinity Crisis (MSC; Hsü et al., 1973, 1977), with an onset dated at 5.97 Ma and a later partial isolation of the Mediterranean Sea (5.59-5.33 Ma; Krijgsman et al., 1999). These closures have been attributed to eustatic changes, climatic events or tectonic processes (Duggen et al., 2003; Leroux et al., 2018), although this discussion still persists (Vai, 2016; Sternai et al., 2017).
The timing of closure of the Betic Strait is still subject to debate. According to Martín et al. (2009) and Pérez-Asensio et al. (2014), the Guadalhorce Corridor was the last seaway of the Betic Strait and it was closed during the early-middle Messinian (Fig. 1.A ![]() : ~6.18 Ma). Nevertheless, new research proposes that the Betic Strait was closed during the late Tortonian (Schee et al., 2018). Other studies postulate that the Gibraltar Corridor was the sole Atlantic gateway during the Messinian (Krijgsman et al., 2018).
: ~6.18 Ma). Nevertheless, new research proposes that the Betic Strait was closed during the late Tortonian (Schee et al., 2018). Other studies postulate that the Gibraltar Corridor was the sole Atlantic gateway during the Messinian (Krijgsman et al., 2018).
During the late Tortonian and early Messinian, these palaeogeographic, palaeoclimatic and/or palaeoceanographic changes affected the benthic faunas of the western Betic Strait. In a general overview, the analysis of the benthic foraminiferal assemblages indicates a progressive decrease in depth during this interval, together with remarkable variations in oxygen contents (González-Regalado & Ruiz, 1996; González-Regalado et al., 2019).
In this paper, we study the ostracod fauna coming from samples of marine sediments crossed by the Huelva-1 borehole. These sediments constitute the Neogene infill of the Guadalquivir Basin (SW Spain). Results are compared with those obtained from the foraminiferal assemblages of the same core (González-Regalado et al., 2019) to recognize the main palaeoenvironmental changes that took place in this sector prior to the MSC.
In the southwestern sector of the Guadalquivir basin (Fig. 1.B ![]() ), three main geological formations are exposed on a Palaeozoic-Mesozoic basement. The Upper Tortonian Niebla Formation (Baceta & Pendón, 1999) is composed of fluvial conglomerates, littoral sands and shallow marine calcarenites, this latter with a remarkable palaeontological record (echinoderms, nummulitids, red algae, bryozoans). The overlying Gibraleón Clay Formation (Civis et al., 1987) consists of gray-blue marls and clays, with a condensed, silty glauconitic layer near the base. This unit presents a very rich micropalaeontological record (mainly foraminifera and calcareous nannoplankton). The planktonic foraminiferal fauna indicates a late Tortonian to Messinian age for these sediments (Sierro, 1985), with Tortonian-Messinian boundary located just above the glauconitic level.
), three main geological formations are exposed on a Palaeozoic-Mesozoic basement. The Upper Tortonian Niebla Formation (Baceta & Pendón, 1999) is composed of fluvial conglomerates, littoral sands and shallow marine calcarenites, this latter with a remarkable palaeontological record (echinoderms, nummulitids, red algae, bryozoans). The overlying Gibraleón Clay Formation (Civis et al., 1987) consists of gray-blue marls and clays, with a condensed, silty glauconitic layer near the base. This unit presents a very rich micropalaeontological record (mainly foraminifera and calcareous nannoplankton). The planktonic foraminiferal fauna indicates a late Tortonian to Messinian age for these sediments (Sierro, 1985), with Tortonian-Messinian boundary located just above the glauconitic level.
|
|
Figure 1: A. palaeogeographical evolution of the Atlantic-Mediterranean connections during the Upper Neogene (modified from Martín et al., 2009); B. Main geological domains of southern Spain and location of the Huelva-1 borehole and some Messinian sections; C. Geochronology and sampling of the Huelva-1 borehole (modified from Larrasoańa et al., 2008, 2014). MSC: Messinian Salinity Crisis. |
The Pliocene sedimentation of this area is represented by the Huelva Sand Formation (Civis et al., 1987) consisting of massive, bioturbated sandy layers alternating with lumachellic layers attributed to storm action in an open bay palaeoenvironment (González-Regalado et al., 2009). These deposits include a new glauconitic layer with a rich fauna of selachians (Ruiz et al., 1998). A broader regional analysis can be consulted in Viguier (1977).
The Huelva-1 borehole (Fig. 1.C ![]() ; UTM PB818265) was drilled by the IGME (Spanish Geological Survey) and it encompasses the upper part of the Niebla Formation (4 m) and most of the Gibraleón Clay Formation (Fig. 1.C
; UTM PB818265) was drilled by the IGME (Spanish Geological Survey) and it encompasses the upper part of the Niebla Formation (4 m) and most of the Gibraleón Clay Formation (Fig. 1.C ![]() : 172 m). The magnetostratigraphic datings indicate that this borehole spans from the latest Tortonian (C3Br.2r, ca. 7.4 Ma) to the latest Messinian (uppermost C3r, ca. 5.4 Ma) (Larrasoańa et al., 2008, 2014). Its geological record presents a cyclicity that started at 7.16 Ma, coinciding with the first sign of Mediterranean-Atlantic gateway restriction (Berg et al., 2018).
: 172 m). The magnetostratigraphic datings indicate that this borehole spans from the latest Tortonian (C3Br.2r, ca. 7.4 Ma) to the latest Messinian (uppermost C3r, ca. 5.4 Ma) (Larrasoańa et al., 2008, 2014). Its geological record presents a cyclicity that started at 7.16 Ma, coinciding with the first sign of Mediterranean-Atlantic gateway restriction (Berg et al., 2018).
Fourteen samples (25 g) for micropalaeontological analysis were selected between 145.8 m and 89.3 m (Fig. 1.C ![]() : HU145.8 to HU89.3). These samples include the four chrons previous to the MSC and chron C3r, within which this crisis developed. The approximate time interval is included between 6.83 Ma (HU145.8) and 6.01 Ma (HU89.2) (Fig. 2
: HU145.8 to HU89.3). These samples include the four chrons previous to the MSC and chron C3r, within which this crisis developed. The approximate time interval is included between 6.83 Ma (HU145.8) and 6.01 Ma (HU89.2) (Fig. 2 ![]() ). This time interval was calculated according to the magnetostratigraphy deduced by Larrasoańa et al. (2008, 2014) for this borehole. These samples were wet sieved (63 μm mesh) and dried in an oven at 70°C. At the end all ostracodes were picked in each sample.
). This time interval was calculated according to the magnetostratigraphy deduced by Larrasoańa et al. (2008, 2014) for this borehole. These samples were wet sieved (63 μm mesh) and dried in an oven at 70°C. At the end all ostracodes were picked in each sample.
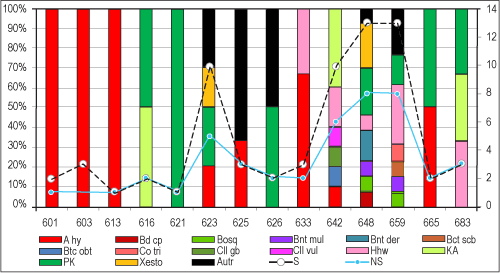
The Huelva-1 borehole presents a badly-preserved and scarce ostracod fauna (Fig. 2 ![]() ). The total number of specimens is very low (69), with a maximum of thirteen specimens per sample in two samples (HU-134.3 and HU-129.3). On the contrary, sample HU-92.3 is barren (Figs. 1
). The total number of specimens is very low (69), with a maximum of thirteen specimens per sample in two samples (HU-134.3 and HU-129.3). On the contrary, sample HU-92.3 is barren (Figs. 1 ![]() - 2
- 2 ![]() ). The ostracod fauna is poorly diversified through the entire studied section of this core. A total of 21 taxa have been identified, including nine of them in open nomenclature. In a general overview, the ostracod fauna is dominated by Acanthocythereis hystrix (Reuss, 1850), Henryhowella partenopaea Bonaduce et al., 1999, Krithe gr. K. iniqua Abate et al., 1993, and Parakrithe group P. dactylomorpha Ruggieri, 1962 (Fig. 2
). The ostracod fauna is poorly diversified through the entire studied section of this core. A total of 21 taxa have been identified, including nine of them in open nomenclature. In a general overview, the ostracod fauna is dominated by Acanthocythereis hystrix (Reuss, 1850), Henryhowella partenopaea Bonaduce et al., 1999, Krithe gr. K. iniqua Abate et al., 1993, and Parakrithe group P. dactylomorpha Ruggieri, 1962 (Fig. 2 ![]() ).
).
Bosquetina carinella (samples HU134.3 to HU129.3) and two species of Buntonia [B. dertonensis (Ruggieri, 1954) and B. multicostata Ruggieri, 1962)] have certain vertical continuity in some sections of the Huelva-1 borehole. Other species not included in Fig. 2 ![]() are Cytheropteron cf. C. sulcatum Bonaduce, Ciampo & Masoli, 1975 (sample HU129.3), Loxoconcha sp. (sample HU119.3), Uroleberis sp. (sample HU118.9), Occultocythereis cf. O. scipionis Bonaduce et al., 1992 (sample HU118.9), Propontocypris sp. (sample HU116.8) and Aglaiocypris? sp. (sample HU116.8).
are Cytheropteron cf. C. sulcatum Bonaduce, Ciampo & Masoli, 1975 (sample HU129.3), Loxoconcha sp. (sample HU119.3), Uroleberis sp. (sample HU118.9), Occultocythereis cf. O. scipionis Bonaduce et al., 1992 (sample HU118.9), Propontocypris sp. (sample HU116.8) and Aglaiocypris? sp. (sample HU116.8).
These low diversities and densities have also been verified in other Messinian sections of the Gibraleón Clay Formation, such as Gibraleón or Trigueros (Fig. 1.B ![]() ; González-Regalado & Ruiz, 1988, 1990). The ostracod assemblages of these sections are very similar to those of the Huelva-1 borehole, with Cytherella, Krithe, Parakrithe, Henryhowella and Costa as the most representative genera.
; González-Regalado & Ruiz, 1988, 1990). The ostracod assemblages of these sections are very similar to those of the Huelva-1 borehole, with Cytherella, Krithe, Parakrithe, Henryhowella and Costa as the most representative genera.
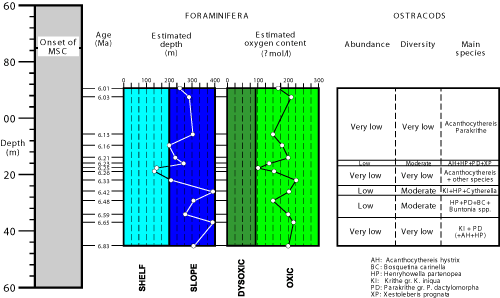
As mentioned above, the main ostracod assemblage of the Huelva-1 borehole is composed of Cytherella spp., Henryhowella partenopaea Bonaduce et al., 1999, Krithe spp. and Parakrithe spp. This assemblage characterizes Neogene to Recent upper slope environments of France, Morocco and the Mediterranean Sea (Puri et al., 1969; Peypouquet, 1979; Llano, 1981; Carbonel, 1985). This overall palaeoenvironment is confirmed by other key species with bathyal affinity, such as Retibythere (Bathybythere) scaberrima (Brady, 1887), Bythocythere obtusata or Costa tricostata (Reuss, 1850) (Sciuto, 2014, 2015). In addition, the estimated palaeodepth calculated on the basis of the recorded benthic foraminiferal assemblages agrees with an upper bathyal palaeoenvironment (Fig. 3 ![]() : 200-400 m depth) for most samples (González-Regalado et al., 2019).
: 200-400 m depth) for most samples (González-Regalado et al., 2019).
In these scenarios, there is usually a high variability in ostracod diversity (Fanget et al., 2013), although the presence of low to moderately diversified ostracod assemblages (<15 species/sample in most cases) is frequent (Benson, 1973a; Sciuto, 2014; Sciuto & Rosso, 2015). The low ostracod diversity observed through the entire studied section of Huelva-1 borehole is comparable with those reported in other upper bathyal Messinian sections of the western Betic Strait (mean: <8 species/sample; González-Regalado & Ruiz, 1990, 1991).
|
|
Figure 2: Density (black dotted line), diversity (blue line) and abundance of the main species (in %). Horizontal axis: samples. Abbreviations: A hy: Acanthocythereis hystrix; Bd cp: Bairdoppilata conformis; Bosq: Bosquetina carinella; Bnt mul: Buntonia multicostata; Bnt der: Buntonia dertonensis; Bct scb: Retibythere scaberrima; Btc obt: Bythocypris obtusata; Co tri: Costa tricostata; Cll gb: Cytherella gibba; Cll vul: Cytherella vulgata; Hhw: Henryhowella partenopea; K.A: Krithe gr. K. iniqua; Xesto: Xestoleberis prognata; Autr.: other species. |
Both density (2-3 individuals/25 g) and diversity (2-3 species/sample) decrease significantly between samples HU122.3 and HU118.9 (Figs. 1 ![]() - 2
- 2 ![]() : 6.33-6.25 Ma). Acanthocythereis hystrix (Reuss, 1850), an allochtonous outer neritic species in the remaining samples, is the most representative species of this interval, together with the presence of some individuals of the allochtonous genera Uroleberys and Loxoconcha should also be highlighted (HU119.3-HU118.9; 6.26-6.25 Ma). Species of these two genera are generally collected together in recent and past shallow marine areas (Zao & Wang, 1988; Safak et al., 2015; Eglington, 2019). These features would indicate a regression, with a transition from upper epibathyal to outer neritic palaeoenvironments, a process also detected in the foraminiferal assemblages (Fig. 3
: 6.33-6.25 Ma). Acanthocythereis hystrix (Reuss, 1850), an allochtonous outer neritic species in the remaining samples, is the most representative species of this interval, together with the presence of some individuals of the allochtonous genera Uroleberys and Loxoconcha should also be highlighted (HU119.3-HU118.9; 6.26-6.25 Ma). Species of these two genera are generally collected together in recent and past shallow marine areas (Zao & Wang, 1988; Safak et al., 2015; Eglington, 2019). These features would indicate a regression, with a transition from upper epibathyal to outer neritic palaeoenvironments, a process also detected in the foraminiferal assemblages (Fig. 3 ![]() ; 130-150 m water depth; González-Regalado et al., 2019). This period coincided with a Miocene glacial period deduced from the isotopic record of deep ODP cores in the North Atlantic (Hodell et al., 2001).
; 130-150 m water depth; González-Regalado et al., 2019). This period coincided with a Miocene glacial period deduced from the isotopic record of deep ODP cores in the North Atlantic (Hodell et al., 2001).
|
|
Figure 3: Comparison between the palaeoenvironmental conditions deduced from the benthic foraminifera and the ostracod fauna of the Huelva-1 borehole. |
In the remaining samples, the presence of these last species and others with shallower distributions [e.g., Bosquetina carinella (Reuss, 1850), Loxoconcha sp., Cytheropteron cf. C. sulcatum Bonaduce et al., 1975] can be attributed to post-mortem transport (Hastrup & Thomsen, 2005; Salihoglu et al., 2018). Neritic species are frequently found in upper bathyal palaeoenvironments because of downslope contamination by neritic sediments (e.g., Guernet & Fourcade, 1988).
Brief considerations about the most significant species, according to their palaeoenvironmental significance or biostratigraphic distribution. They are listed in alphabetical order in each section.
C.1. Autochtonous species
One specimen of Bairdoppilata conformis (Terquem, 1878) was found in sample HU129.3 (6.48 Ma). This species has a wide bathymetrical distribution in both the Mediterranean Sea (Bonaduce et al., 1983; Montenegro et al., 1998) and the Atlantic Ocean (Whatley & Coles, 1987; Yasuhara & Okahashi, 2014), ranging from circalittoral to bathyal environments. It is a long-ranging species widely distributed within the Cenozoic, but this species has also been collected in recent sediments of the Mediterranean Sea (Harten & Droste, 1988; Bossio et al., 2006; Sciuto, 2012; Sciuto & Rosso, 2015).
Two specimens of Buntonia dertonensis (Ruggieri, 1954) were extracted in sample HU129.3 (Figs. 1 ![]() - 2
- 2 ![]() : 6.48 Ma). This species has been collected (also as Buntonia sublatissima dertonensis Ruggieri, 1954) from Miocene to Recent marine sediments in Austria (Szczechura & Aiello, 2003), Malta (Barra & Bonaduce, 2001), Italy (Sciuto, 2014), and Spain (González-Delgado et al., 1982). This species lives in lower circalittoral to epibathyal environments in the Mediterranean Sea (Sciuto, 2014; see review in Stow et al., 2013).
: 6.48 Ma). This species has been collected (also as Buntonia sublatissima dertonensis Ruggieri, 1954) from Miocene to Recent marine sediments in Austria (Szczechura & Aiello, 2003), Malta (Barra & Bonaduce, 2001), Italy (Sciuto, 2014), and Spain (González-Delgado et al., 1982). This species lives in lower circalittoral to epibathyal environments in the Mediterranean Sea (Sciuto, 2014; see review in Stow et al., 2013).
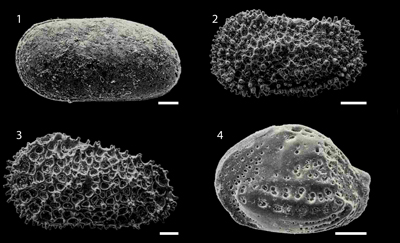
Buntonia multicostata Ruggieri, 1962 (Fig. 4.4 ![]() ), has been recovered (as Buntonia sublatissima multicostata) from Miocene to Pleistocene sediments in Spain (González-Regalado & Ruiz, 1990; Ruiz et al., 2008b), Italy (Ruggieri, 1962; Colalongo et al., 1990), and Malta (Bonaduce & Barra, 2002). It is a open-shelf/deep-water species (sensu Russo et al., 2012), cited in lower circalitoral to bathyal palaeoenvironments of Algeria (>100 m depth; Carbonnel & Courme-Rault, 1997). This species is present in two consecutive samples (Figs. 1
), has been recovered (as Buntonia sublatissima multicostata) from Miocene to Pleistocene sediments in Spain (González-Regalado & Ruiz, 1990; Ruiz et al., 2008b), Italy (Ruggieri, 1962; Colalongo et al., 1990), and Malta (Bonaduce & Barra, 2002). It is a open-shelf/deep-water species (sensu Russo et al., 2012), cited in lower circalitoral to bathyal palaeoenvironments of Algeria (>100 m depth; Carbonnel & Courme-Rault, 1997). This species is present in two consecutive samples (Figs. 1 ![]() - 2
- 2 ![]() ; HU134.3-HU129.3: 6.59-6.48 Ma).
; HU134.3-HU129.3: 6.59-6.48 Ma).
One specimen of Bythocypris obtusata (Sars, 1866) has been collected in sample HU126.7 (6.42 Ma). It is mainly found in bathyal (palaeo-)environments, both in the Mediterranean Sea (300-2905 m depth; Puri et al., 1964; Bonaduce & Pugliese, 1979; Sciuto, 2014) and the Atlantic Ocean (Benson et al., 1983), although this species has been also observed in neritic areas of the North Atlantic (Sars, 1928). Specimens of this species has been collected from Miocene to Recent sediments in Italy (Colalongo & Pasini, 1980; Sciuto & Rosso, 2015), Turkey (Ertekin & Tunoglu, 2008), and Greece (Sissingh, 1972).
|
|
Figure 4: Main species of the Huelva-1 borehole. 1: Cytherella vulgata Ruggieri, 1962 (sample HU126.7); 2: Henryhowella partenopea Bonaduce et al., 1999 (sample 145.8); 3: Acanthocythereis hystrix (Reuss, 1850) (sample 136.8); 4: Buntonia multicostata Ruggieri, 1962 (sample 134.3). Scale bar: 100 μm. |
Costa tricostata (Reuss, 1850) is a long-ranging species from palaeogene to Neogene sediments in France (Ducasse & Mondain-Monval, 1984), Austria (Zorn, 2004), Italy (Dall'Antonia, 2002; Sciuto, 2014), and Spain (Ruiz et al., 2011). This species was typical of sediments referred to upper bathyal palaeoenvironments in Italy (Ruggieri, 1992; Bossio et al., 2006; Sciuto, 2014) and Spain (Abad et al., 2005). One specimen of this species was found in sample HU134.3 (Figs. 1 ![]() - 2
- 2 ![]() : 6.59 Ma).
: 6.59 Ma).
Cytherella gibba Aiello et al., 1996, has been described from Tortonian to Pleistocene in Italy (Aiello et al., 1996; Faranda et al., 2007). This species inhabited at moderate water depths, exceeding 150 m, in upper Italian bathyal palaeoenvironments (Faranda et al., 2007; Cipollinari et al., 2009; Baldanza et al., 2013). One specimen of this species was collected in sample HU126.7 (Figs. 1 ![]() - 2
- 2 ![]() : 6.42 Ma).
: 6.42 Ma).
Cytherella vulgata Ruggieri, 1962 (Fig. 4.1 ![]() ), is frequent from Miocene to Recent in the Mediterranean Sea (Ruggieri, 1962; Sissingh, 1972; Carbonnel & Courme-Rault, 1997) and adjacent Atlantic areas (Abad et al., 2011). This species inhabits in lower circalittoral to epibathyal environments, at water depths exceeding usually 125 m in the Mediterranean Sea (Puri et al., 1969; Bonaduce et al., 1975; Aranki, 1987) and the Moroccan Atlantic shelf (Llano, 1981). This species was only collected (1 specimen) in sample HU126.7 (Figs. 1
), is frequent from Miocene to Recent in the Mediterranean Sea (Ruggieri, 1962; Sissingh, 1972; Carbonnel & Courme-Rault, 1997) and adjacent Atlantic areas (Abad et al., 2011). This species inhabits in lower circalittoral to epibathyal environments, at water depths exceeding usually 125 m in the Mediterranean Sea (Puri et al., 1969; Bonaduce et al., 1975; Aranki, 1987) and the Moroccan Atlantic shelf (Llano, 1981). This species was only collected (1 specimen) in sample HU126.7 (Figs. 1 ![]() - 2
- 2 ![]() : 6.42 Ma).
: 6.42 Ma).
Eleven specimens of Henryhowella partenopea Bonaduce et al., 1999 (Fig. 4.2 ![]() ), were collected in six basal samples of the Huelva-1 borehole (Figs. 1
), were collected in six basal samples of the Huelva-1 borehole (Figs. 1 ![]() - 2
- 2 ![]() : HU145.8 to HU118.9, except two samples; 6.83-6.25 Ma). It is a common species [as Henryhowella asperrima (Reuss, 1850) in most cases] in Tortonian to Recent marine sediments from the Mediterranean Sea (Bonaduce et al., 1999) and the adjacent Atlantic zones (Ruiz et al., 2008a). This species has been found in recent shelf sediments of the Mediterranean Sea (40-170 m depth; Bonaduce et al., 1999), although this species has been also found in Neogene upper slope palaeoenvironments of the Betic Strait (Ruiz & González-Regalado, 1996).
: HU145.8 to HU118.9, except two samples; 6.83-6.25 Ma). It is a common species [as Henryhowella asperrima (Reuss, 1850) in most cases] in Tortonian to Recent marine sediments from the Mediterranean Sea (Bonaduce et al., 1999) and the adjacent Atlantic zones (Ruiz et al., 2008a). This species has been found in recent shelf sediments of the Mediterranean Sea (40-170 m depth; Bonaduce et al., 1999), although this species has been also found in Neogene upper slope palaeoenvironments of the Betic Strait (Ruiz & González-Regalado, 1996).
Krithe gr. K. iniqua Abate et al., 1993, and Parakrithe gr. P. dactylomorpha Ruggieri, 1962, are very similar to their reference-species, but the minimum differences are probably due to taphonomic processes. Krithe iniqua Abate et al., 1993, has been extracted from Tortonian to Pleistocene sediments in Italy (Ciampo, 1980, 1986; Abate et al., 1993; Aiello & Barra, 2001) and Langhian-Serravalian limestones and clays in Malta (Bonaduce & Barra, 2002). This species and others very similar have been collected in upper bathyal palaeoenvironments of Italy (Sciuto & Rosso, 2008; Sciuto & Baldanza, 2020). Seven specimens of this species have been collected in four samples, ranging from 6.83 Ma (sample HU145.8) to 6.16 Ma (sample HU106.3).
Parakrithe dactylomorpha Ruggieri, 1962, has been found in Miocene to Pleistocene sediments from Spain (González-Regalado & Ruiz, 1990), Switzerland (Brinkmann et al., 2019), Italy (Ruggieri, 1962), Croatia (Hajek-Tadesse & Prtoljan, 2011), and Greece (Sissingh, 1972; Hastrup & Thomsen, 2005). This species inhabited in lower circalittoral to epibathyal sediments both in the Betic Strait (Benson, 1972; González-Regalado & Ruiz, 1991) and the Mediterranean Sea (Hastrup & Thomsen, 2005). It is the most abundant species of the Huelva-1 borehole, with fourteen specimens extracted in nine samples distributed throughout the interval studied.
Retibythere (Bathybythere) scaberrima (Brady, 1887) is a common species (as Bythoceratina scaberrima) in Mediterranean bathyal palaeoenvironments from Miocene to the early Pleistocene (review in Sciuto, 2015). This species has even been described in Holocene sequences of the NE Atlantic Ocean (Whatley & Ayress, 1988) and recent bathyal sediments of the Atlantic Ocean (see review in Yasuhara et al., 2014). One specimen of this species was found in sample 134.3 (Figs. 1 ![]() - 2
- 2 ![]() : 6.59 Ma)
: 6.59 Ma)
Xestoleberis prognata Bonaduce & Danielopol, 1988, has been collected in Tortonian to Pleistocene sediments from Italy (Abate et al., 1994) and Spain (Ruiz et al., 2004). In Italy, this species is frequent in bathyal palaeoenvironments (Abate et al., 1994; Violanti et al., 2009). Five specimens of this species were found in samples HU129.3 (6.48 Ma) and HU114.5 (6.23 Ma).
To sum up, the species found present a wide biostratigraphic range and indicate a Tortonian-Holocene age for the studied samples. This age is refined from the magnetostratigraphic analysis of Huelva-1 borehole. Ages of the fourteen studied samples ranges from 6.83 Ma (HU145.8) to 6.01 Ma (HU89.3) (Larrasoańa et al., 2008, 2014).
C.2. Allochtonous species
Thirteen specimens of Acanthocythereis hystrix (Reuss, 1850) (Fig. 4.3 ![]() ) were collected in eight samples (Figs. 1
) were collected in eight samples (Figs. 1 ![]() - 2
- 2 ![]() : HU136.8 to HU89.3; 6.65-6.01 Ma). This species was previously known from the Middle Miocene of Austria (Zorn, 2004), Miocene-Pleistocene of Greece (Hastrup & Thomsen, 2005; Faranda et al., 2008) and Plio-Pleistocene of Tunisia (Temani et al., 2016). This allochtonous species generally inhabits (or inhabited) shelf environments located between the external infralittoral to the inner circalittoral zones in the Eastern Mediterranean Sea (Nazik, 2001; Parlak & Nazik, 2016) and Italy (Bonaduce & Pugliese, 1979; Montenegro et al., 1998).
: HU136.8 to HU89.3; 6.65-6.01 Ma). This species was previously known from the Middle Miocene of Austria (Zorn, 2004), Miocene-Pleistocene of Greece (Hastrup & Thomsen, 2005; Faranda et al., 2008) and Plio-Pleistocene of Tunisia (Temani et al., 2016). This allochtonous species generally inhabits (or inhabited) shelf environments located between the external infralittoral to the inner circalittoral zones in the Eastern Mediterranean Sea (Nazik, 2001; Parlak & Nazik, 2016) and Italy (Bonaduce & Pugliese, 1979; Montenegro et al., 1998).
Bosquetina carinella (Reuss, 1850). This species was found from Eocene to Holocene both in the Mediterranean Sea (Parlak & Nazik, 2016; Temani et al., 2016; Salihoglu et al., 2018) and the Atlantic Ocean (Ruiz & González-Regalado, 1996). This species inhabited mostly circalittoral palaeoenvironments in Austria (Zorn, 2007) and Spain (Ruiz et al., 2008b), although it has also been cited in infralittoral areas of Portugal (Antunes et al., 1996).
The mentioned autochtonous ostracod assemblage (Retibythere, Krithe, Cytherella, Henryhowella, Bythocypris) is typical of bathyal palaeoenvironments with low to very low temperatures (Benson, 1973b; Sciuto, 2015). Species of Henryhowella and Krithe were found at temperatures below 10°C (Benson, 1973b; Nazeer et al., 2019), whereas Retibythere scaberrima lives even in polar environments (Yasuhara et al., 2014).
The foraminiferal assemblages indicate the presence of a well-oxygenated palaeoenvironment during the period studied (Messinian: 6.8-6.0 Ma; González-Regalado et al., 2019), with a small drop between 6.26-6.25 Ma. This interval coincides with a decrease in both the density and diversity of the ostracod fauna (Fig. 2 ![]() ). In addition, the frequent presence of Parakrithe is associated with high productivity (Peypouquet, 1979; Bassetti et al., 2010).
). In addition, the frequent presence of Parakrithe is associated with high productivity (Peypouquet, 1979; Bassetti et al., 2010).
During the middle Messinian (6.8-6.0 Ma), the western sector of the Betic Strait was occupied by upper bathyal palaeoenvironments (200-400 m depth), with a general decreasing depth trend during this period. These upper slope scenarios were dominated by Henryhowella partenopaea Bonaduce et al., 1999, Krithe gr. K. iniqua Abate et al., 1993, and Parakrithe gr. P. dactylomorpha Ruggieri, 1962, with minor contributions of the genera Cytherella, Buntonia and Xestoleberis. A timely transition to outer neritic palaeoenvironments is characterized by the presence of Acanthocythereis hystrix (Reuss, 1850), which is the main allochtonous species in most of samples. This general overview coincides with the results previously obtained from the benthic foraminiferal assemblages. The biostratigraphic distribution of the main species (mainly Tortonian-Holocene) agrees with the ages obtained by magnetostratigraphic studies (6.8-6.0 Ma). Small drops of the oxygen levels caused a decline in ostracod populations during a glaciation that occurred between 6.26-6.25 Ma.
Funds have come from Andalusian Government (RNM-238). It is a contribution to the Research Center in Historical, Cultural and Natural Heritage (CIPHCN) of the University of Huelva. We thank Dr Bruno Granier for his editorial review. The manuscript also benefited from the constructive reviews of Dr Pierre Carbonel and Dr Francesco Sciuto.
Abad M., Ruiz F., Pendón J.G., González-Regalado M.L. & Tosquella J. (2005).- Tortonian ostracodes of southwestern Spain.- Geobios, Villeurbanne, vol. 38, p. 563-573.
Abad M., Toscano A., García E.X.M., González-Regalado M.L., Ruiz F., Civis J., González-Delgado J.A., Jiménez E., Tosquella J., Álvarez G., Velo D. & Molin M. (2011).- The palaeontological record of a condensed section (Sands of Huelva Fm., Lower Pliocene, SW Spain).- Abstracts Joint RCMNS - RCANS Interim Colloquium, Salamanca, p. 52-53.
Abate S., Barra D., Aiello G. & Bonaduce, G. (1993).- The genus Krithe Brady, Crosskey & Robertson, 1875 (Crustacea: Ostracoda) in the Pliocene-Early Pleistocene of the M. San Nicola Section (Gela, Sicily).- Bolletino della Societŕ Paleontologica Italiana, Milano, vol. 32, p. 349-366.
Abate S., Barra D. & Bonaduce, G. (1994).- The deep-water Xestoleberidinae Sars, 1928 (Crustacea, Ostracoda) in the Pliocene-Early Pleistocene of the M. San Nicola Section (Gela, Sicily).- Revista Espańola de Micropaleontología, Madrid, vol. 26, p. 43-47.
Aiello G. & Barra D. (2001).- Pliocene ostracod assembiages at the M PI 3 - M PI 4 boundary in the Capo Rossello borehoie (Agrigento, Sicily).- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 40, p. 97-103
Aiello G., Barra D., Bonaduce G. & Russo A. (1996).- The genus Cytherella Jones, 1849 in the Italian Tortonian-Recent.- Revue de Micropaléontologie, Paris, vol. 39, p. 171-190.
Antunes M.T., Legoinha P., Nascimiento A. & Pais J. (1996).- The evolution of the Lower Tagus basin (Lisbon and Setúbal Peninsula, Portugal) from Lower to early Middle Miocene.- Géologie de la France, Orléans, vol. 4, p. 59-77.
Aranki F.J. (1987).- Marine lower Pliocene ostracodes of southern Spain with notes on the Recent fauna.- Bulletin of the Geological Institutions of the University of Upssala, vol. 13, p. 1-93.
Baceta J.I. & Pendón J.G. (1999).- Estratigrafía y arquitectura de facies de la Formación "Niebla", Neógeno Superior, sector occidental de la Cuenca del Guadalquivir.- Revista de la Sociedad Geológica de Espańa, Madrid, vol. 12, p. 419-438.
Baldanza A., Bizzarri R., Famiani F., Garassino A., Hyzny M. & Pasini G. (2013).- The bathyal decapod crustacean community from the Poggio i Sodi quarries (Siena Basin, Tuscany, Italy).- Boletín de la Sociedad Geológica Mexicana, Ciudad de México, vol. 65, p. 335-353.
Barra D. & Bonaduce G. (2001).- Some new and poorly know Middle Miocene ostracods of Malta Isle.- Bolletino della Societŕ Paleontologica Italiana, Milano, vol. 40, p. 55-74.
Bassetti M.A., Carbonel P., Sierro F.J., Pérez-Folgado M., Jouët G. &. Berné S. (2010).- Response of ostracods to abrupt climate changes in the Western Mediterranean (Gulf of Lions) during the last 30 kyr.- Marine Micropaleontology, vol. 77, p. 1-14.
Benson R.H. (1972).- Miocene deep-sea Ostracodes of the Iberian Portal and the Balearic Basin.- Marine Micropaleontology, vol. 1, p. 249-262.
Benson R.H. (1973a).- Psychrospheric and continental Ostracoda from ancient sediments in the floor of Mediterranean.- Initial Report of the Deep-Sea Drilling Project, College Station - TX, vol. 8, p. 1002-1009.
Benson R.H. (1973b).- An ostracodal view of the Messinian salinity crisis. In: Drooger C.W. (Ed.), Messinian Events in the Mediterranean.- Geodynamics Scientific Reports, Amsterdam, vol. 7, p. 235-242.
Benson R.H., Del Grosso R.M. & Steineck P.L. (1983).- Ostracode distribution and biofacies, Newfoundland continental slope and rise.- Micropaleontology, Flushing - NY, vol. 29, p. 430-453.
Berg B.C.J. van der, Sierro F.J., Hilgen F.J., Flecker R., Larrasoańa J.C., Krijgsman W., Flores J.A. & Mata M.P. (2018).- Imprints of Messinian Salinity Crisis events on the Spanish Atlantic margin.- Newsletters on Stratigraphy, vol. 51, p. 93-115.
Bonaduce G. & Barra D. (2002).- The ostracods in the palaeoenvironmental interpretation of the late Langhian - Early Serravallian section of Ras il-Pellegrin (Malta).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 108, p. 211-222.
Bonaduce G., Barra D. & Aaiello G. (1999).- The genus Henryhowella Puri, 1957 (Crustacea, Ostracoda) in the Atlantic and Mediterranean from Miocene to Recent.- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 38, p. 59-72.
Bonaduce G., Ciampo G. & Masoli M. (1975).- Distribution of Ostracoda in the Adriatic Sea.- Publicazioni della Stazione Zoologica di Napoli, vol. 40, p. 1-304.
Bonaduce G., Ciliberto B., Masoli M., Minichelli G. & Pugliese N. (1983).- The deep-water benthic ostracodes of the Mediterranean. In: Maddocks R.F. (ed.), Application of Ostracoda.- Department of Geosciences, University of Houston, p. 459-471.
Bonaduce G. & Pugliese N. (1979).- Benthic ostracods as depth indicators.- Rapports de la Commission Internationalse sur la Mer Méditerranée, Monte Carlo, vol. 25/26, p. 167-169.
Bossio A., Dall'Antonia B., Da Prato S., Foresi L.M. & Oggiano G. (2006).- Preliminary stratigraphical investigations on the Miocene successions of the Porto Torres basin (northern Sardinia, Italy).- Atti della Societŕ Toscana di Scienze Naturali, Pisa, vol. 111, p. 67-74.
Brinkmann L., Pipperr M. & Reichenbacher B. (2019).- A new well-preserved ostracod fauna from the middle Burdigalian (lower Miocene) of the North Alpine Foreland Basin.- Geobios, Villeurbanne, vol. 56, p. 65-93.
Carbonel P. (1985).- Neogene. In: Oertli H. (ed.), Atlas des Ostracodes de France.- Bulletin du Centre de Recherche et Exploitation Production elf-Aquitaine, Pau, vol. 9, p. 313-335.
Carbonnel G. & Courme-Rault M.D. (1997).- Ostracodes miocčnes d'Algérie (systématique, biostratigraphie, distribution palinspatique).- Mémoires du Muséum d'Histoire naturelle de Lyon, vol. 1, 131 p.
Ciampo G. (1980).- Ostrcodi miocenici (Tortoniano-Messiniano) della regione di Ragusa (Sicilia).- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 19, p. 5-20.
Ciampo G. (1986).- Ostracodi del limite Tortoniano/Messiniano in alcune sezioni italiane.- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 24, p. 29-110.
Cipollari P., Cosentino D., Faranda C., Gennari R., Gliozzi E. & Grossi F. (2009).- The Messinian/Zanclean boundary at Maccarone (Marche): Integrated biostratigraphy and palaeoenvironmental evolution.- Abstracts 2009, Gruppo Informale di Ricerche Micropaleontologiche e Malacologiche, Roma, p. 1-2.
Civis J., Sierro F.J., González-Delgado J.A., Flores J.A., Andrés I., Porta J. & Valle M.F. (1987).- El Neógeno marino de la Provincia de Huelva: Antecedentes y definición de sus unidades litoestratigráficas In: Civis J. (ed.), Paleontología del Neógeno de Huelva (W Cuenca del Guadalquivir).- Universidad de Salamanca, p. 5-16.
Colalongo M.L. & Pasini G. (1980).- La ostracofauna plio-pleistocenica della Sezione Vrica in Calabria (con considerazioni sul limite Neogene/Quaternario). Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 19, p. 44-126.
Colalongo M.L., Pasini G., Poluzzi A. & Sprovieri R. (1990).- Relationship between the benthic foraminifers and the ostracodes in the Pliocene-Pleistocene Tyrrhenian deep-sea record (ODP leg 107, site 654).- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 107, p. 479-493.
Dall'Antonia B. (2002).- Short palaeoecological notes on the middle Serravalian-basal Tortonian ostracods from the Tremiti Islands.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 108, p. 289-296.
Ducasse O. & Mondain-Monval J.Y. (1984).- Les genres Costa et Paleocosta (Ostracodes) du Paléogčne aquitain : Les structures, de populations, témoins des paléoenvironnements. Description de Paleocosta aquitanica nov. sp.- Geobios, Villeurbanne, vol. 17, p. 549-565.
Duggen S., Hoernle K., Van den Bogaard P., Rupke L. & Morgan J.P. (2003).- Deep roots of the Messinian salinity crisis.- Nature, vol. 422, p. 602-606.
Eglington C. (2019).- Marine ostracoda (Crustacea) from the Late Oligocene Gellibrand marl, Otway Basin, Victoria, Australia.- Proceedings of the Royal Society of Victoria, Melbourne, vol. 131, no. 2, p. 53-73.
Ertekin I.K. &. Tunoglu C. (2008).- Pleistocene-Holocene marine ostracods from Mersin offshore sediments, Turkey, Eastern Mediterranean.- Revue de Micropaléontologie, Paris, vol. 51, p. 309-326.
Fanget A.S., Bassetti A., Berné S. & Arnaud M. (2013).- Epi-bathyal ostracod assemblage in Holocene Rhone deltaic sediments (Gulf of Lions) and their palaeoecological implications.- Revue de Paléobiologie, Genčve, vol. 32, p. 589-606.
Faranda C., Cipollinari P., Cosentino D., Gliozzi E. & Pipponzi G. (2008).- Late Miocene ostracod assemblages from eastern Mediterranean coral reef complexes (central Crete, Greece).- Revue de Micropaléontologie, Paris, vol. 51, p. 287-308.
Faranda C., Gliozzi E. & Mazzini I. (2007).- Palaeoenvironmental evolution of the Plio-Pleistocene Monte Mario succession (Rome, Italy) inferred from ostracod assemblages.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 113, p. 473-485.
Flecker R., Krijgsman W., Capella W., Martins C., Mayser J.P., Marzocchi A., Modestu S., Ochoa D., Simon D., Tulbure M., Berg B.V.D., Schee M.V.D., De Lange G., Ellan R., Govers R., Guthir R., Hilgen F., Kouwenhoven T., Lofi J., Mejier P., Sierro F.J., Bachiri N., Barhoun N., Chakor A., Chacón B., Flores J. A., Gregory J., Howard J., Lunt D., Ochoa M., Pancost R., Vincent S. & Youfin M. Z. (2015).- Evolution of the Late Miocene Mediterranean-Atlantic gateways and their impact on regional and global environmental change.- Earth-Science Reviews, vol. 150, p. 365-392.
González-Delgado J.A., Civis J. & Sierro F.J. (1982).- Ostracofauna del Neógeno de Niebla (Huelva).- Stvdia Geologica Salmanticensia, vol. 17, p. 43-65.
González-Regalado M.L., Romero V., Abad M., Tosquella J., Gómez P., Clemente M.J., Toscano A., Rodríguez Vidal J., Cáceres L.M., Muńoz J.M., Prudencio M.I., Dias M.I., Marques R., García E.X.M., Monge G., Carretero M.I. & Ruiz F. (2019).- Late Tortonian-middle Messinian palaeoenvironmental changes in the western Betic Strait (SW Spain).- Ameghiniana, Buenos Aires, vol. 56, p. 336-360.
González-Regalado M.L. & Ruiz F. (1988).- Precisiones sobre la ostracofauna del Neógeno de Trigueros (Huelva).- Actas IV Jornadas de Paleontología, Salamanca, vol. 1, p. 46-47.
González-Regalado M.L. & Ruiz F. (1990).- Los ostrácodos del tramo inferior de la Formación "Arcillas de Gibraleón" (Gibraleón, provincial de Huelva, S.W. Espańa).- Revista de la Sociedad Geológica de Espańa, Madrid, vol. 3, p. 23-31.
González-Regalado M.L. & Ruiz F. (1991).- Significado paleoecológico y bioestratigráfico de los ostrácodos del Neógeno Superior de la sección de Huelva.- Revista Espańola de Paleontología, Madrid, vol. 6, p. 107-116.
González-Regalado M.L. & Ruiz F. (1996).- Les foraminiféres benthiques de la baie du sud-ouest de l'Espagne pendant le Néogčne supérieur : Le Mio-Pliocčne de Huelva.- Revue de Paléobiologie, Genčve, vol. 15, p. 109-120.
González-Regalado M.L., Ruiz F., Abad M., Civis J., González-Delgado J.A., Pendón J.G. & Toscano A. (2009).- Impact of storms on Pliocene benthic foraminiferal assemblages of southwestern Spain.- Ameghiniana, Buenos Aires, vol. 46, p. 345-360.
Guernet C. & Fourcade É. (1988).- Cenozoic ostracodes from Hole 628A, ODP LEG 101, Bahamas.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 101, p. 139-151.
Hajek-Tadesse V. & Prtoljan B. (2011).- Badenian Ostracoda from the Pokupsko area (Banovina, Croatia).- Geologica Carpathica, Bratislava, vol. 62, p. 447-461.
Harten D. van & Droste H.J. (1988).- Mediterranean deep-sea ostracods, the species poorness of the eastern basin as a legacy of an early Holocene anoxic event.- Developments in Palaeontology and Stratigraphy, vol. 11, p. 721-737.
Hastrup A. & Thomsen E. (2005).- Paleoenvironmental interpretation of the Plio-Pleistocene Kallithea Bay section, Rhodes, Greece based on ostracods.- Cushman Foundation for Foramiferal Research, Lawrence - KS, vol. 39, p. 159-191.
Hodell D.A., Curtis J.H., Sierro F.J. & Raymo M.E. (2001).- Correlation of late Miocene to early Pliocene sequences between the Mediterranean and North Atlantic.- Paleoceanography, vol. 16, p. 164-178.
Hsü K.J., Montadert L., Bernoulli D., Cita M.B., Erickson A., Garrison R.E., Kidd R.B., Meliéres F., Muller C. & Wright R. (1977).- History of the Messinian salinity crisis.- Nature, vol. 267, p. 399-403.
Hsü K.J., Ryan W. & Cita M.B. (1973).- Late Miocene dessiccation of the Mediterranean.- Nature, vol. 242, p. 240-244.
Krijgsman W., Capella W., Simon D., Hilgen F.J., Kouwenhoven T.J., Meijer P.T., Sierro F.J., Tulburé M.A., Berg B.V.D., Schee M.V.D. & Flecker R. (2018).- The Gibraltar Corridor: Watergate of the Messinian salinity crisis.- Marine Geology, vol. 403, p. 238-246.
Krijgsman W., Hilgen F.J., Raffi I., Sierro F.J., & Wilson D.S. (1999).- Chronology, causes and progression of the Messinian salinity crisis.- Nature, vol. 400, p. 652-655.
Larrasoańa J.C., González-Delgado J.A., Civis J., Sierro F.J., Alonso-Gavilán G. & Pais J. (2008).- Magnetostratigraphic dating and environmental magnetism of Late Neogene marine sediments recovered at the Huelva-1 and Montemayor-1 boreholes (lower Guadalquivir basin, Spain).- Geotemas, Madrid, vol. 10, p. 1175-1178.
Larrasoańa J.C., Liu Q., Hu P., Roberts A.P., Mata P., Civis J., Sierro F.J. & Pérez-Asensio J.N. (2014).- Paleomagnetic and paleoenvironmental implications of magnetofossil occurrences in late Miocene marine sediments from the Guadalquivir Basin, SW Spain.- Frontiers in Microbiology, Washington D.C., vol. 5, p. 71.
Leroux E., Aslanian D., Rabineau M., Pellen R. & Moulin M. (2018).- The late Messinian event: A worldwide tectonic revolution.- Terra Nova, vol. 30, p. 207-214.
Llano M. (1981).- Intéręt des ostracodes dans l'interpretation des phénomčnes hydrologiques sur les plateaux continentaux : La plate-forme atlantique marocaine.- Ph.D. Thesis, Université de Bordeaux, 256 p.
Martín J.M., Braga J.C., Aguirre J. & Puga-Bernabéu A. (2009).- History and evolution of the North-Betic Strait (Prebetic Zone, Betic Cordillera): A narrow, early Tortonian, tidal-dominated, Atlantic-Mediterranean marine passage.- Sedimentary Geology, vol. 216, p. 80-90.
Montenegro M.E., Pugliese N. & Bonaduce G. (1998).- Shelf ostracods distribution in the Italian seas. In: Crasquin-Soleau S., Braccini E. & Lethiers F. (eds.), What about Ostracoda.- Bulletin du Centre Recherche elf Exploration Production, Pau, p. 91-101.
Nazeer M.N., Radhakrishnan K., Hussain S.M., Sivapriya V. & Rajkumar A. (2019).- Genus Krithe (Ostracoda) as a proxy to decipher paleoceanography: A global review of the genus.- Oceanography & Fisheries, Irvine - CA, vol. 10, no. 3, 3 p.
Nazik A. (2001).- Ostracode faunas of bottom sediments from the continental shelf, south Marmara Sea, NW Turkey, and their comparison with other shelf environments in the Mediterranean and Aegean regions.- Geological Journal, vol. 36, p. 111-123.
Parlak D. & Nazik A. (2016).- Ostracodes of the Mediterranean (the Gulf of Antalya) and the Aegean Sea (Ayvalik and Kusadasi) and their biogeographical distributions.- Bulletin of the Mineral Research and Exploration, Ankara, vol. 152, p. 63-83.
Pérez-Asensio J.N., Aguirre J., Schmiedl G. & Civis J. (2014).- Messinian productivity changes in the northeastern Atlantic and their relationship to the closure of the Atlantic-Mediterranean gateway: Implications for Neogene palaeoclimate and palaeoceanography.- Journal of the Geological Society, London, vol. 171, p. 389-400.
Peypouquet J.P. (1979).- Ostracodes et paléoenvironnements. Méthodologie et applications aux domaines profonds du Cénozoďque.- Bulletin du Bureau de Recherches Géologiques et Miničres, Orléans, vol. 4, p. 3-79.
Puri H.S., Bonaduce G. & Malloy J. (1964).- Ecology of the Gulf of Naples.- Pubblicazioni della Stazione Zoologica di Napoli, vol. 3, p. 87-199.
Puri H.S., Bonaduce G. & Gervasio A.M. (1969).- Distribution of Ostracoda in the Mediterranean. In: Neale J.W. (ed.), The taxonomy, morphology and rcology of Recent Ostracoda.- Oliver & Boyd, Edimburgh, p. 356-412.
Ruggieri G. (1954).- Iconografia degli Ostracodi marine del Pliocene e Pleistocene italiani. Parte II.- Atti della Societŕ Italiana di Scienze Naturali, Milano, vol. 93, p. 561-575.
Ruggieri G. (1962).- Gli Ostracodi marini del Tortoniano (Miocene medio-superiore) de Enna nella Sicilia orientale.- Palaeontographia Italica, Pisa, vol. LVI (Nuova Serie, vol. XXVI) for 1961, p. 1-68 (Pls. XI-XVII [I-VII]).
Ruggieri G. (1992).- Considerazioni tassonomiche su Ostracodi neogenici e pleistocenici risultate dalla revisione di vecchi lavori dello scrivente.- Bolletino della Societŕ Paleontologica Italiana, Milano, vol. 31, p. 175-188.
Ruiz F., Abad M., Pendón J.G., González-Regalado M.L. & Tosquella J. (2004).- Ostrácodos tortonienses del sector central de la Cuenca del Guadalquivir (Sur de Espańa).- Geogaceta, vol. 35, 131-134.
Ruiz F., Abad M., Toscano A., González-Regalado M.L. & García E.X.M. (2011).- Messinian ostracodes of southwestern Spain.- Abstracts Joint RCMNS - RCANS Interim Colloquium, Salamanca, p. 212-213.
Ruiz F., Clauss F.L. & González-Regalado M.L. (1998).- Primeras consideraciones sobre los Condrichtios de la Formación "Arenas de Huelva".- Stvdia Geologica Salmanticensia, vol. 32, p. 129-139.
Ruiz F. & González-Regalado M.L. (1996).- Les ostracodes du Golfe Mio-Pliocene du sud-ouest de l'Espagne.- Revue de Micropaléontologie, Paris, vol. 39, p. 137-151.
Ruiz F., González-Regalado M.L., Abad M. & Civis J. (2008a).- Ostrácodos neógenos marinos del suroeste de Europa. In: González-Delgado J.A. & Civis J. (eds.), The Atlantic Neogene in the International Year of the Earth Planet.- Abstracts Fourth Congress RCANS, Buenos Aires, p. 51.
Ruiz F., González-Regalado M.L., Abad M., Civis J., González-Delgado J.A., García E.X.M., Prudencio M.I., Dias M.I. (2008b).- Pliocene ostracods of Southwestern Europe.- Geobios, Villeurbanne, vol. 41, p. 845-859.
Russo A., Pugliese N. & Serventi P. (2012).- Miocene ostracodes of cold seep settings from northern Apennines (Italy).- Revue de Micropaléontologie, Paris, vol. 55, p. 29-38.
Safak U., Ocakollu F. & Acikalin S. (2015).- Ostracoda assemblage and the environmental characteristics of the Eocene succession of the Central Sakarya Region.- Micropaleontology, Flushing - NY, vol. 61, p. 49-68.
Salihoglu R., Akcer-on S., Langer M.R. & Viehberg F.A. (2018).- Holocene environmental reconstruction of Lake Bafa (SW, Anatolia) by statistical analyses of ostracod and benthic foraminifer assemblages.- 20th EGU General Assembly, EGU2018, Proceedings from the conference held 4-13 April, 2018 in Vienna, p. 1108.
Sars G.O. (1928).- An account of the Crustacea of Norway vol. IX: Ostracoda.- CD book, Bergen Museum, 119 Pls.
Schee, M. van der, Berg, B.C.J. van der, Capella W., Simon D., Sierro F.J., & Krijgsman W. (2018).- New age constraints on the western Betic intramontane basins: A late Tortonian closure of the Guadalhorce Corridor?.- Terra Nova, vol. 30, p. 325-332.
Sciuto F. (2012).- Bythocythere solisdeus n. sp. and Cytheropteron eleonorae n. sp. (Crustacea, Ostracoda) from the Early Pleistocene Bathyal sediments of Cape Milazzo (NE, Sicily).- Geosciences, Basel, vol. 2, pp. 147-156.
Sciuto F. (2014).- Ostracods of the Upper Pliocene-Pleistocene Punta Mazza succession (NE Sicily) with special focus on the Family Trachyleberididae Sylvester-Bradley, 1948, and description of a new species.- Carnets Geol., Madrid, vol. 14, no. 1, p. 1-13.
Sciuto F. (2015).- Palaeoenviromental and palaeogeographical traits on deep-water ostracod assemblages from the Lower Pleistocene sediments along the Ionian side of Mount Etna (Sicily, Italy).- Carnets Geol., Madrid, vol. 15, no. 14, p. 179-197.
Sciuto F. & Baldanza A. (2020).- Full restoration of marine conditions after the late Messinian Mediterranean Lago-Mare phase in Licodia Eubea and Villafranca Tirrena areas (east Sicily).- Carnets Geol., Madrid, vol. 20, no. 6, p. 107-123.
Sciuto F. & Rosso A. (2008).- Distribution pattern of deep-water ostracod assemblages from Lower Pleistocene sediments from Furnari, Sicily.- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 47, p. 33-43.
Sciuto F. & Rosso A. (2015).- Bathyal ostracods from the Santa Maria di Leuca deep-water coral province (northern Ionian Sea).- Palaeontologia Electronica, vol. 18.2.22a, p. 1-17.
Sierro F.J. (1985).- Estudio de los foraminíferos planctónicos, bioestratigrafía y cronoestratigrafía del Mioceno-Plioceno del borde occidental de la cuenca del Guadalquivir (SO de Espańa).- Stvdia Geologica Salmanticensia, vol. 21, p. 7-85.
Sissingh W. (1972).- Late Cenozoic Ostracoda of the South Aegean Island Arc.- Utrech Micropaleontological Bulletins, vol. 6, p. 1-188.
Sternai P., Caricchi L., García-Castellanos D., Jolivet L., Sheldrake T.E. & Castelltort S. (2017).- Magmatic pulse driven by sea-level changes associated with the Messinian Salinity Crisis.- Nature Geosciences, vol. 10, p. 783.
Stow D.A.V., Hernández-Molina F.J., Álvarez-Zarekian C. & The Expedition 339 Scientists (2013).- Site U1387.- Proceedings IODP, College Station - TX, vol. 339, 133 p.
Szczechura J. & Aiello G. (2003).- The ostracod genus Nipponocythere Ishizaki, 1971 from the Middle Miocene of the Fore-Carpathian Depression, Central Paratethys; its origin and paleoenvironment.- Geologica Carpathica, Bratislava, vol. 54, p. 9-20.
Temani R., Nachite D., Sciuto F., Razgallah S., Bekkali R., Hayet K. & Gaaloul N. (2016).- Les Ostracodes plio-pléistocčnes des séries sédimentaires de la bordure orientale du Cap Bon (coupe de l'Oued Lebna, Tunisie orientale).- Carnets Geol., Madrid, vol. 16, no. 18, p. 431-447.
Vai G.B. (2016).- Over half a century of Messinian salinity crisis. Boletín Geológico y Minero, Madrid, vol. 127, p. 625-641.
Viguier C. (1977).- Les grands traits de la tectonique de Basin néogčne du Bas Guadalquivir.- Boletín Geológico y Minero, Madrid, vol. 88, p. 39-44.
Violanti D., Trenkwalder S., Lozar F. & Gallo L.M. (2009).- Micropalaeontological analyses of the Narzole core: Biostratigraphy and palaeoenvironment of the late Messinian and early Zanclean of Piedmont (Northwestern Italy).- Bolletino della Societŕ Paleontologica Italiana, Milano, vol. 48, p. 167-181.
Whatley R. & Ayress M. (1988).- Pandemic and endemic distribution pattern in Quaternary deep sea ostracods. In: Hanai T., Ikeya N. & Ishizaki K. (eds.), Evolutionary biology of Ostracoda. Its fundamentals and applications.- Developments in Paleontology and Stratigraphy, vol. 11, p. 739-755.
Whatley R. & Coles G. (1987).- The late Miocene to Quaternary Ostracoda of Leg 94, Deep Sea Drilling Project.- Revista Espańola de Micropaleontologia, Madrid, vol. 19, p. 33-97.
Yasuhara M., Stepanova A., Okahashi H., Cronin T.M. & Brouwers E.M. (2014).- Taxonomic revision of deep-sea Ostracoda from the Arctic Ocean.- Micropaleontology, Flushing - NY, vol. 60, p. 399-444.
Zao Q. & Wang P. (1988).- Distribution of Modern Ostracoda in the shelf seas off China.- Developments in Paleontology and Stratigraphy, vol. 11, p. 805-821.
Zorn I. (2004).- Ostracoda from the Lower Badenian (Middle Miocene) Grund Formation (Molasse Basin, Lower Austria).- Geologica Carpathica, Bratislava, vol. 55, p. 179-189.
Zorn I. (2007).- Ostracoda from the holostratotype of the Badenian (Middle Miocene), Central Paratethys, Austria.- Poster 6th European Ostracodologists-Meeting, Frankfurt.