◄ Carnets Geol. 15 (14) ►
![]()
Contents
[1. Introduction]
[2. Materials and methods]
[3. Results]
[4. Systematics]
[5. Discussion and conclusions]
[Bibliographic references] [Plates] and ... [Table]
Palaeoecological Research Group, Dipartimento di Scienze
Biologiche, Geologiche ed Ambientali, Universitŕ di Catania, Corso Italia 55, I-95129 Catania (Italy)
Published online in final form (pdf) on November 11, 2015
[Editor: Bruno ; language editor:
Donald E. ]
![]()
The ostracod associations of the Lower Pleistocene sedimentary succession cropping out at Serra San Biagio (Catania, Sicily NE) have been investigated. The ostracod fauna from nine samples is poorly diversified but well-preserved: A total of forty taxa of ostracods have been identified, thirty-three at specific level and seven at genus level or doubtful species. Thirty species are considered as constituents of the in situ original assemblages. The associations consist almost exclusively of bathyal taxa such as Bythocypris obtusata (), Anchistrocheles antemacella , Henryhowella ex H. hirta () group, Quasibuntonia radiatopora (), Retibythere (Bathybythere) scaberrima () and Bathycythere vansraateni . Also the Krithe group is well-represented with Krithe compressa () and K. pernoides (). Almost all species, some, particularly interesting from the palaeoecological and palaeogeographical point of view, have been here described, illustrated and commented on, including a species belonging to the genus Cytherella , 1849, found in all samples of the section, which is here proposed as new.
Marine ostracods; bathyal; palaeogeography; palaeoenvironment; new species; Early Pleistocene; blue clays; Mount Etna; Sicily.
F. (2015).- Palaeoenviromental and palaeogeographical traits on deep-water ostracod assemblages from the Lower Pleistocene sediments along the Ionian side of Mount Etna (Sicily, Italy).- Carnets Geol., Madrid, vol. 15, nº 14, p. 179-197.
Caratteristiche paleoambientali e paleogeografiche dell'associazione ad ostracodi di sedimenti del Pleistocene Inferiore affioranti lungo il versante ionico del Monte Etna.- Sono state studiate le associazioni ostracodi della successione sedimentaria del Pleistocene Inferiore affiorante a Serra San Biagio (Catania, Sicilia NE). La fauna č spesso poco diversificata ma ben conservata: sono state determinati 40 taxa, 33 a livello specifico, 7 a livello generico. L'associazione consiste quasi esclusivamente di taxa batiali come Bythocypris obtusata (), Anchistrocheles antemacella , Henryhowella ex H. hirta () group, Quasibuntonia radiatopora (), Retibythere (Bathybythere) scaberrima () e Bathycythere vansraateni . Anche il gruppo dei Krithe č ben rappresentato con Krithe compressa () e K. pernoides (). Nel presente lavoro, inoltre, sono state descritte, illustrate e commentate piů in dettaglio alcune specie particolarmente significative dal punto di vista paleoecologico e paleogeografico. Tra di esse una, rinvenuta lungo tutta la sezione ed appartenente al genere Cytherella , 1849, viene proposta come nuova.
Ostracodi marini; batiale; paleogeografia; paleoambiente; specie nuove; Pleistocene Inferiore; argille azzurre; Etna; Sicilia.
Caractéristiques paléoenvironnementales et paléogéographique des associations d'ostracodes des sédiments d'âge Pléistocčne inférieur du versant ionien de l'Etna (Sicile, Italie).- Nous avons étudié les associations d'ostracodes de la série sédimentaire d'âge Pléistocčne inférieur affleurant ŕ Serra San Biagio (Catane, NE Sicile). La faune d'ostracodes de neuf échantillons est peu diversifiée mais bien préservée : 40 taxons y ont été identifiés, trente-trois au niveau de l'espčce et sept au niveau du genre ou d'espčces discutables. Trente espčces sont considérées comme appartenant aux associations primaires autochtones. Les associations se composent presque exclusivement de taxons bathyaux tels que Bythocypris obtusata (), Anchistrocheles antemacella , Henryhowella ex groupe H. hirta (), Quasibuntonia radiatopora (), Retibythere (Bathybythere) scaberrima () et Bathycythere vanstraateni . Le groupe Krithe est aussi bien représenté avec Krithe compressa () et K. pernoides (). Presque toutes les espčces, dont certaines, particuličrement intéressantes du point de vue paléoécologique et paléogéographique, ont été ici décrites, illustrées et commentées ; une de ces espčces, attribuée au genre Cytherella , 1849, et observée dans tous les échantillons de la coupe, est nouvelle et fait l'objet d'une description.
Ostracodes marins ; étage bathyal ; paléogéographie ; paléoenvironnement ; nouvelle espčce ; Pléistocčne inférieur ; argiles bleues ; Etna ; Sicile.
Bathyal Plio-Pleistocene sediments crop out widely in Sicily. Only some of their ostracod associations have been so far studied in detailed, namely those of the "Trubi" and blue clay outcrops at Mont St. Nicola ( et al., 2000), those of "Trubi" of Capo Rossello ( & , 2001) and those of Plio-Pleistocene sediments of Capo Milazzo (, 2014, inter alias), Furnari ( & , 2008) and Centuripe ( et al., 2003).
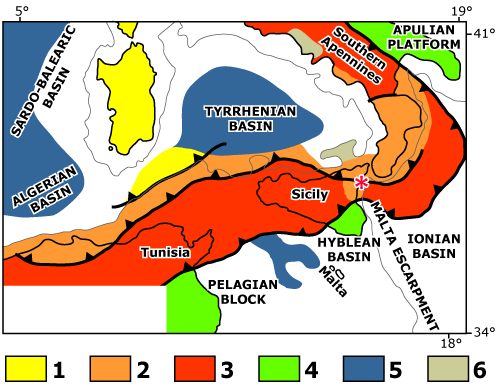
Quaternary sediments occur also on the slopes of the Mount Etna, particularly along his Ionian side, where they crop out discontinuously at different heights above the present-day sea level. This area is located in a complex geological and geodynamic context just at the contact between the volcanic products of the Mount Etna to the South and the thrust front of the Kabylo - Calabride Units to the North. Environments and palaeo-environments are strongly controlled by active tectonics (Fig. 1 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 1: Tectonic scheme of the central Mediterranean area with the site of the study area. 1 = Corsica - Sardinia; 2= Calabrian Arc, Kabylians and "Internal" Flysch sequence ophiolites; 3 = Maghrebian-Sicilian-Southern Apennine nappes and deformed foreland; 4 = foreland and mildly folded foreland (Tunisia, Hyblean plateau, Apulia); 5 = areas with superimposed extension; 6 = Plio-Quaternary volcanoes; * = studied area (after et al., 1996).
The Quaternary sediments, were deposited unconformably on the Apennine-Maghrebian units during the Plio-Pleistocene ( et al., 1999) and, in their turn, constitute the substratum for lava flows of the Etna volcano ( et al., 1979), Holocene lava debris and alluvial deposits. Quaternary marine sediments have greatly variable thickness ( et al., 1999) and generally consist of calcarenites grading upward to clays and sandy silts. The clays are known as "argille grigio-azzurre" formation and crop out discontinuously in Acitrezza, Vambolieri, Ficarazzi, Nizzeti, Cannizzaro, Vena, Santa Venera, Poggio Cibali and Catira areas, below the Mount Etna lavas.
Several authors of the nineteenth and twentieth centuries have analyzed the invertebrate faunas of the blue clays from these localities located in the hills near Catania; one of the first was , whose mollusc collection from the Nizzeti clays, was described and published by (1858); posteriorly (1900) inter alias, based on the molluscan fauna, referred these sediments to the "Post Pliocene" or Piacenziano auct. (, unpublished data). Recently, the mollusc faunas of the Monte Vamboleri have been analysed by (2000, and unpublished data) who highlighted that the numerous Early Pleistocene mollusc species identified presumably lived in bathyal environments located at depths exceeding 400 meters. Microfaunas from thee sediments cropping out at Mt. Catira were analysed by (1862).
The same Pleistocene sedimentary succession is also well exposed norhwards of the Acitrezza area, along the Ionian coast of Mount Etna, near Fiumefreddo, on Serra San Biagio hill, along the right side of the Torrente Minissale (Fig. 1 ![]() ). In this locality the Pleistocene sedimentary succession cropping out with a thickness of about 80 m includes both the calcarenites and the clays.
The age of these sediments has been referred to the Early-Middle Pleistocene by et al.
(1999), whereas &
(2002) suggest a time interval between the Sicilian and the Ionian Stage
(MNN 19e and MNN 19f zones).
). In this locality the Pleistocene sedimentary succession cropping out with a thickness of about 80 m includes both the calcarenites and the clays.
The age of these sediments has been referred to the Early-Middle Pleistocene by et al.
(1999), whereas &
(2002) suggest a time interval between the Sicilian and the Ionian Stage
(MNN 19e and MNN 19f zones).
There are no recent publications on the ostracod fauna coming from the "argille grigio azzurre" out cropping along the Jonian side of the Mout Etna, located in a complex geological context characterized by intense active tectonics ( et al., 1996). Therefore, a study of ostracod associations from the Pleistocene sediments cropping out at Serra San Biagio hill (Fiumefreddo, Catania) was undertaken, as part of a program on systematic, stratigraphic and palaeoecological study of fossil (, 2003, 2005; & , 2008, inter alias) and Recent (, 2014; et al., 2010) bathyal faunas. The results are given in the present paper.
The study of ostracod associations focuses on the sandy-silty sedimentary succession, six metres thick, cropping in a quarry on the Serra San Biagio hill, along the right side of the Torrente Minissale (Tav. Fiumefreddo, F. 262, II NO; 37°48'17.52"N 15°13'35.38"E), near the village of Fiumefreddo (CT) (Figs. 2 ![]() - 3
- 3 ![]() ).
).
Click on thumbnail to enlarge the image.
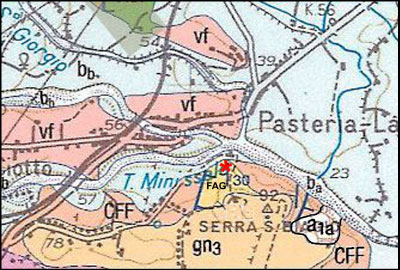
Figure 2: Geological map of the study area (from Carta geologica d'Italia 1:50.000, Foglio 613 Taormina). CFF =Pleistocene calcarenites and FAG = Pleistocene clays. vf = Volcanics. gn3 = Old alluvial deposits. Bb = Recent alluvial deposits. a1a = Landslide deposits.

Click on thumbnail to enlarge the image.
Figure 3: General view of the North side of Serra San Biagio hill and location of the quarry.
Nine samples collected along the quarry were analysed (Figs. 4 ![]() - 5
- 5 ![]() ). Ostracods were studied from 300 cm3 samples. The sediments were routinely washed and ostracod specimens picked up from the > 63 microns fraction. The specimens were examined and measured under a stereomicroscope and photographed by a LMU Tescan Vega II Scanning Electron Microscope.
). Ostracods were studied from 300 cm3 samples. The sediments were routinely washed and ostracod specimens picked up from the > 63 microns fraction. The specimens were examined and measured under a stereomicroscope and photographed by a LMU Tescan Vega II Scanning Electron Microscope.

Click on thumbnail to enlarge the image.
Figure 4: The investigated sedimentary succession exposed in the quarry of Serra San Biagio hill. Sample locations indicated in figure. Note: the bulldozer blade created the vertical grooves oblique to the stratification.

Click on thumbnail to enlarge the image.
Figure 5: The Lower Pleistocene sandy-silty sediments in the quarry of Serra San Biagio hill. Scale: the diameter of the coin is 21 mm.
The presence of carapaces and valves is indicated in Table 1. The presence of juvenile valves was also recorded. Distinction between autochthonous and allochthonous specimens was defined on the basis of the knowledge of the ecological distribution of species, ontogenetic structure, taphonomic features and palaeoecological inferences (cf. , 2003; et al., 2003).
Species have been grouped based on their distribution in the context of the benthic zones ( & , 1989; et al., 1998; & , 2002, inter alias): species restricted to the bathyal zone (bathyal group); species typically found from the deepest outer shelf (circalittoral zone) to the bathyal (shelf edge-bathyal group); species ranging from the shallow circalittoral zone to the bathyal (circalittoral-bathyal group) and widely distributed species (ubiquitous group).
The material is housed in the Paleontological Museum of Catania University. The repository number of Holotype, Paratypes and figured specimens is given in the systematic accounts.
The examined sediments are mainly terrigenous and their texture falls within silts and very fine sands. The carbonate component is very slight and consists predominantly of foraminifera tests, ostracods and rare molluscs (Fig. 5 ![]() ).
).
Click on thumbnail to enlarge the image.
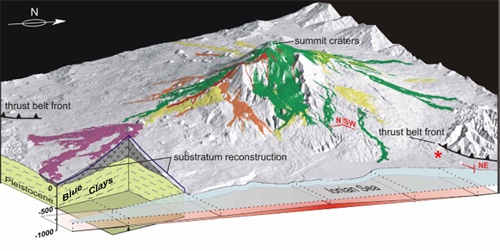
Figure 6: Schematic block diagram shows the sedimentary substrate below the lava flows of Mt. Etna (after et al., 2014). * studied area.
A total of forty ostracod taxa have been identified, thirty at specific level and ten at genericlevel or doubtful species (Table 1). Thirty species are considered as constituents of the in situ original assemblages. The remaining taxa are rare everywhere, and includ Aurila, Celtia and Urocythereis species. They were considered displaced and constitute the allochthonous component of the fauna.
The lower part of the section (samples 1-5) is characterized by the presence of most of the in situ ostracod taxa (29 taxa), that are usually abundant. The most abundant and common species are Anchistrocheles antemacella , 1969, Bairdoppilata conformis (, 1878), Bythocypris obtusata (, 1866), Cytherella carmela n. sp., less abundant but still frequent are the Krithe group species, Quasibuntonia radiatopora sculpta (, 1880), Retibythere (Bathybythere) scaberrima (, 1887), Henryhowella ex gr. H. hirta (, 1853) and Macrosarisa bensoni , 1990. Finally, the less abundant and less frequent species are Argilloecia acuminata , 1894, Sclerochilus contortus , 1866, and species belonging to Cytheropteron. The upper part of the section (samples 6-9) is characterized by less abundant and less frequent species with a total of 24 taxa.
Among all the species found along the stratigraphic section, some particularly significant from the palaeoecological point of view are described, commented and figured below. Furthermore, a species belonging to the genus Cytherella , 1849, found in all the samples, is here proposed as new.
Class Ostracoda , 1806
Subclass Podocopa , 1866
Order Platycopida , 1866
Superfamily Cytherelloidea , 1866
Family Cytherellidae , 1866
Genus Cytherella , 1849
Type species: Cythere ovata (, 1841)
Cytherella carmela n. sp.
(Pl. 1 ![]() ,
figs. A-H)
,
figs. A-H)
1978 Cytherella vulgata , 1962: , Pl. 2, fig. 3;
2009 Cytherella robusta s.l. & (1980): et al., p. 884, Pl. 1, figs. 7-12.
Derivatio nominis: from the name Carmela, my mother.
Material: eight right valves and nine left valves from samples 1 and 2.
Holotype: a complete carapace from sample 1 figured in Pl. 1 ![]() ,
fig. E (L = 890 µm; H = 480 µm; l max = 220 µm) (PMC.O13 H 06/11/2014).
,
fig. E (L = 890 µm; H = 480 µm; l max = 220 µm) (PMC.O13 H 06/11/2014).
Paratypes: the two left valves figured in Pl. 1 ![]() ,
fig. A, D, the two right valves figured in Pl. 1
,
fig. A, D, the two right valves figured in Pl. 1 ![]() ,
figs. B-C (PMC.O48-51 P. 06.11.2014) and other valves not figured.
,
figs. B-C (PMC.O48-51 P. 06.11.2014) and other valves not figured.
Type locality: the quarry along the right side of Torrente Minissale, near Fiumefreddo (Tav. Fiumefreddo, F. 262, II NO, 37°48'17.52"N 15°13'35.38"E), in the Lower Pleistocene blue sandy silts.
Stratigraphic range: Early Pleistocene, Sicilian stage.
Diagnosis: Cytherella carmela n. sp. is characterized by a large bottleneck in the median part of the carapace and by prominent and acute spines in the posterior area.
Description: Carapace medium-sized, which shows, in lateral view, the typical shape of the genus
(Pl. 1 ![]() , figs. A-E). Anterior and posterior margins regularly arched. Dorsal margin concave toward the front, ventral margin slightly convex.
, figs. A-E). Anterior and posterior margins regularly arched. Dorsal margin concave toward the front, ventral margin slightly convex.
In dorsal view the central part of the carapace is characteristically skimmed (compressed)
(Pl. 1 ![]() , fig. E), posterior area with rounded margin more inflated than the anterior; anterior margin slightly acute.
, fig. E), posterior area with rounded margin more inflated than the anterior; anterior margin slightly acute.
Outer surface ornamented by numerous tiny papillae distributed especially around the central part to form a thin festooned structure
(Pl. 1 ![]() ,
figs. A-C, G). In the anterior margin the papillae form some curved lines parallel to the outer margin
(Pl. 1
,
figs. A-C, G). In the anterior margin the papillae form some curved lines parallel to the outer margin
(Pl. 1 ![]() ,
fig. G). In the posterior area the papillae become progressively thicker towards the outer rim, been replaced by acute and ribbed spines
(Pl. 1
,
fig. G). In the posterior area the papillae become progressively thicker towards the outer rim, been replaced by acute and ribbed spines
(Pl. 1 ![]() ,
fig. F). Normal pore canals few, simple, and distributed regularly on the carapace
(Pl. 1
,
fig. F). Normal pore canals few, simple, and distributed regularly on the carapace
(Pl. 1 ![]() ,
figs. A-C). Closure of the valve (Pl. 1
,
figs. A-C). Closure of the valve (Pl. 1 ![]() ,
fig. D) and muscle scars (Pl. 1
,
fig. D) and muscle scars (Pl. 1 ![]() ,
figs. D, H) as for the genus.
,
figs. D, H) as for the genus.
Remarks: In lateral view the specimens of Serra S. Biagio (Pl. 1 ![]() ) show general morphologic features similar to C. robusta & ,
1980 [particularly to the specimen from the Pleistocene of the Vrica section, figured by &
(1980, Pl. 6, fig. 4), and the specimen figured by et al.
(1996: Pl. 2, fig. 9)]. Nevertheless, the posterior area is characterised by a different ornamentation. Indeed in C. robusta the ornamentation in the posterior area consists of simple and stumpy papillae or tubercles. Conversely in C. carmela n. sp. prominent and acute, in some cases sometimes ribbed
(Pl. 1
) show general morphologic features similar to C. robusta & ,
1980 [particularly to the specimen from the Pleistocene of the Vrica section, figured by &
(1980, Pl. 6, fig. 4), and the specimen figured by et al.
(1996: Pl. 2, fig. 9)]. Nevertheless, the posterior area is characterised by a different ornamentation. Indeed in C. robusta the ornamentation in the posterior area consists of simple and stumpy papillae or tubercles. Conversely in C. carmela n. sp. prominent and acute, in some cases sometimes ribbed
(Pl. 1 ![]() ,
fig. F), spines are present. In dorsal view C. carmela n. sp. shows a concavity in the central area of the carapace more marked than C. robusta. The specimens found at 1798 m wd in the Western North Atlantic by et al.
(2009) were referred to the Latest Pleistocene-Holocene. The specimen figured by
(1978), found in the Alghero-Provencal Basin, is referred to the Piacentian (Late Pliocene, now Early Pleistocene sensu & ,
2008).
,
fig. F), spines are present. In dorsal view C. carmela n. sp. shows a concavity in the central area of the carapace more marked than C. robusta. The specimens found at 1798 m wd in the Western North Atlantic by et al.
(2009) were referred to the Latest Pleistocene-Holocene. The specimen figured by
(1978), found in the Alghero-Provencal Basin, is referred to the Piacentian (Late Pliocene, now Early Pleistocene sensu & ,
2008).
Order Podocopida , 1866
Family Bairdiidae , 1865
Genus Bairdoppilata et al., 1935
Type species: Bairdoppilata martyni et al., 1935
Bairdoppilata subdeltoidea (, 1830) var. conformis (, 1878)
(Pl. 2 ![]() ,
fig. A)
,
fig. A)
1878 Bairdia subdeltoidea V. var. conformis , p. 93, Pl. 10, figs. 17 a-c;
1883a Bairdia subdeltoidea V. : , p. 58;
1883b Bairdia subdeltoidea V. : , p. 124, Pl. 1, fig. 8;
1975 Bairdia conformis (): et al., p. 22, Pl. 6, figs. 5-10;
1980 Bairdia conformis (): & , p. 52, Pl. 1, fig. 5;
2000 Bairdoppilata conformis (): et al., p. 85, Pl. 1, fig. 2;
2008 Bairdia conformis (): & , p. 35, Pl. 1, fig 2;
2014 Bairdoppilata conformis (, 1878): , p. 4.
The muscle scars of the figured specimen in Pl. 2 ![]() ,
fig. A, are very similar to those figured by
(1969: p. 15, fig. 3i) and fit well within the genus Bairdoppilata. In the Recent this species has a wide ecological distribution. In the Mediterranean it is reported from 50 m wd down to the Bathyal Zone ( et al.,
1983) and in the Atlantic from 1245 m wd. Bairdoppilata subdeltoidea var. conformis is also known as fossil from Plio-Pleistocene sediments deposited in bathyal environments such as those of the Monte San Nicola section ( et al.,
2000), the Vrica section ( & ,
1980) and the Capo Milazzo area (,
2014). The stratigraphic range of this species is from Lower Pleistocene to Recent.
,
fig. A, are very similar to those figured by
(1969: p. 15, fig. 3i) and fit well within the genus Bairdoppilata. In the Recent this species has a wide ecological distribution. In the Mediterranean it is reported from 50 m wd down to the Bathyal Zone ( et al.,
1983) and in the Atlantic from 1245 m wd. Bairdoppilata subdeltoidea var. conformis is also known as fossil from Plio-Pleistocene sediments deposited in bathyal environments such as those of the Monte San Nicola section ( et al.,
2000), the Vrica section ( & ,
1980) and the Capo Milazzo area (,
2014). The stratigraphic range of this species is from Lower Pleistocene to Recent.
Family Bythocyprididae , 1969
Genus Bythocypris , 1880
Type species: Bythocypris reniformis , 1880
Bythocypris obtusata (, 1866)
1866 Bairdia obtusata , p. 24;
1880 Bairdia obtusata var. producta, , p. 191;
1880 Cytheridea lucida , p. 290, Pl. 16, fig. 51;
1987 Bythocypris lucida (): , p. 48, Pl. 2, fig. 6;
1972 Bythocypris obtusata (): , p. 79, Pl. 3, figs. 10-11;
1972 Bythocypris lucida (): , p. 78, Pl. 3, figs. 8-9;
1975 Bairdia obtusata : , p. 205, Pl. 2, fig. 9;
1980 Bythocypris obtusata (): & , p. 52, Pl. 2, figs. 9-10;
1981 Bythocypris obtusata (): , p. 83, Pl. 4, figs. 1-3;
2008 Bythocypris obtusata (): & , p. 35, Pl. 1, fig. 3;
2014 Bythocypris obtusata (): , p. 4.
B. obtusata has been reported from the Recent in the Atlantic-Mediterranean Region between 600 and 2669 m wd ( et al., 1983), particularly at 300 m wd in the Gulf of Naples ( & , 1979), between 145 and 165 m wd off Norwegian and British coasts (, 1928) and between 150 and 2905 m in localities of the Mediterranean Sea ( et al., 1969).
Fossils records of this species are from sediments referable to bathyal environments including the Tyrrhenian Sea ( et al., 1988, 1990), Le Castella section (, 1965), Capo Milazzo (, 2003, 2005), Furnari ( & , 2008) and the Vrica section ( & , 1980). The stratigraphic range of B. obtusata is from the Miocene ( & , 1985; , 1972) to the Recent.
Genus Anchistrocheles & , 1889
Type species: Anchistrocheles fumata , 1890
Anchistrocheles antemacella , 1969
(Pl. 2 ![]() ,
figs. B-C)
,
figs. B-C)
1969 Anchistrocheles antemacella , p. 116, Figs. 60 i-l;
1974 Zabythocypris antemacella (, 1969): et al., p. 173;
1980 Zabythocypris antemacella (, 1969): & , p. 68, Pl. 30, figs. 9-10;
1988 Zabythocypris antemacella (, 1969): & , p. 282;
1990 Zabythocypris antemacella (, 1969): et al., p. 488.
The Serra S. Biagio specimens are very similar to those figured by & (1980) in pl. 30, figs. 9-10. In particular, the H/L ratio is roughly comparable and is diagnostic to distinguish the specimens of A. antemacella from A. interrupta, described and figured by et al. (1996), and from A. tenera (, 1975).
This species is so far unkown as living from the Mediterranean Sea. The stratigraphic range entirely covers the Pleistocene as A. antemacella has been reported from bathyal sediments of the Vrica section ( & , 1980), from the ODP LEG 107, Site 654, in the Tyrrhenian sea ( et al., 1990) and from Monasterace ( et al., 1974). This species has been also found subfossil in the Mozambico Channel in submerged sediments deeper than 1800 m wd (, 1969).
Family Macrocyprididae , 1912
Genus Macrosarisa , 1990
Type species: Macrosarisa bensoni , 1990
Macrosarisa bensoni , 1990
(Pl. 2 ![]() ,
figs. D-F)
,
figs. D-F)
1972 Macrocypris sp.1, , p. 79, Pl. 3, fig. 14;
1990 Macrosarisa bensoni , p. 88, Figs. 12 (8, 9) et alias;
2004 Macrosarisa bensoni , p. 166;
2009 Macrosarisa bensoni et al., p. 887.
Until now, M. bensoni has been not reported as living in the Mediterranean Sea, whereas it thrives in West Indies and North Brazil (, 1880) and in South Atlantic at 834-939 m wd (, 1990), and the Gulf of Mexico at 1079 m wd ( et al., 2009). This species is known as fossil from the bathyal sediments of the Calabrian Stage tage of Rodi (, 1972).
Genus Macrocyprissa , 1960
Type species: Macrocyprissa cylindracea (, 1855)
Macrocyprissa arcuata ( & , 1980)
(Pl. 2 ![]() ,
fig. G)
,
fig. G)
1965 Macrocypris sp. , p. 89, Pl. 1, fig. 7;
1980 Paramacrocypris arcuata & , p. 106, Pl. 25, figs. 1-8;
1990 Macrocyprissa arcuata ( & , 1980): , p. 85, Pl. 30, figs. 7-10;
2000 Macrocyprissa arcuata ( & , 1980): et al., p. 89.
M. arcuata is reported as living from the northeastern Atlantic Ocean at 1491-1500 m wd (, 1990). Fossils of this species are known from bathyal sediments of Le Castella (, 1965), the Vrica Section ( & , 1980) and from the Porcupine Basin at 700-750m wd ( et al., 1996).
Family Krithidae , 1958
Genus Krithe et al., 1874
Type species: Ilyobates praetexta , 1866
Krithe frutex et al., 1993
1993 Krithe frutex et al., Pl. 4, figs. 1-4, Pl. 6, fig. 15.
This species seems to be rare in the Mediterranean area and presently reported only from the Monte san Nicola section from the Lower Pleistocene ( et al., 1993).
Krithe pernoides (, 1855)
(Pl. 2 ![]() ,
fig. H)
,
fig. H)
1855 Bairdia pernoides , p. 358, Pl. 20, fig. 7;
1986 Krithe sinuosa , p. 87, Pl. 17, figs. 13-53;
1993 Krithe pernoides (): et al., p. 362, Pl. 2, figs. 6-12; Pl. 3, figs. 9-10;
2008 Krithe pernoides (): & , Pl. 1, fig 7.
The specimen here figured share its morphological features with the specimen figured by et al. (1993) in Pl. 2, fig. 7. Strong affinities are also present with K. padovani & , 1980, as figured in their Pl. 1, fig 5 ( & , 1980).
In the Mediterranean area K. pernoides () is known from the Lower Miocene to the LowerEarly Pleistocene. In the Atlantic Ocean this species is known from Miocene to Quaternary in deep-water sediments and also in the Recent with a wide depth range between 185 and 5440m ( et al., 1994).
Krithe marialuisae et al., 1993
(Pl. 2 ![]() ,
fig. I)
,
fig. I)
1993 Krithe marialuisae et al., p. 361, Pl. 2, figs. 1-6; Pl. 6, fig. 5;
1999 Krithe marialuisae et al., 1993: et al., p. 6, figs. 3 I-J & 8 G-H.
K. marialuisae is not reported as living in the Mediterranean sea. Valves of this species have been reported from the Tasman Sea and off southern Australia, at depth 1066 and 2860 m ( et al., 1999). Fossil distribution ranges from the Upper Pliocene to the Lower Pleistocene ( et al., 1993).
Krithe perpulchra et al., 1993
1993 Krithe perpulchra et al., p. 364, Pl. 3, figs. 6-9; Pl. 6, fig. 13;
1999 Krithe perpulchra et al.: et al., p. 10, Figs. 5C, 5E, 8R.
This species is known as fossil in the Mediterranean basin from the Upper Pliocene to the Lower Pleistocene ( et al., 1993). In the Australian continental slope valves of K. perpulchra were found at depths of 759, 773 and 1321 m ( et al., 1999).
Krithe undecimradiata , 1974
(Pl. 2 ![]() ,
fig. L)
,
fig. L)
1974 Krithe undecimradiata : et al., p. 175-177, Figs. 6, 3a, 3b, 4a, 4b;
1986 Krithe undecimradiata : , Pl. 18, figs. 2-3.
(1974) described K. undecimradiata from the Calabrian stage of Monasterace and considered this species as a psychrosphaeric taxon.
Krithe aequabilis , 1986
(Pl. 2 ![]() ,
figs. M-N)
,
figs. M-N)
1986 Krithe aequabilis , p. 87, Pl. 17, figs. 1-2;
1990 Krithe aequabilis , 1986: et al., p. 488, Pl. 1, figs. 5-6;
1994 Krithe aequabilis , 1986: et al., p. 78, Pl. 1, figs. 7-12;
2009 Krithe aequabilis , 1986: , p. 5.
In the Mediterranean area this species is known only as fossil. In the Holocene of North Atlantic it is known between 1200 to 4000 m wd ( et al., 1994). In subpolar North Atlantic Ocean this species has been reported from MIS 6 up to the Lower Holocene , 2009).
Krithe compressa (, 1880)
(Pl. 2 ![]() ,
fig. O)
,
fig. O)
1880 Ilyobates compressa , p. 325, Pl. 17, figs. 30, 30A;
1974 Krithe compressa (, 1880): et al., p. 173;
1988 Krithe compressa (, 1880): & , p. 285;
1993 Krithe compressa (, 1880): et al., p. 354, Pl. 3, figs. 1-5; Pl. 6, figs. 6-8;
1994 Krithe reversa (, 1958) (pars): et al., p. 77, Figs. 3 A, D;
1999 Krithe compressa (, 1880): et al., p. 6, Figs. 2A-B, 3A-D, 8A-B;
2003 Krithe compressa (, 1880): , p. 273.;
2008 Krithe compressa (, 1880): & , p. 222, Pl. 5, figs. 1-5;
2008 Krithe compressa (, 1880): & , p. 35, Pl. 1, fig. 6;
2014 Krithe compressa (, 1880): , p. 4.
This species is not known as living in the Mediterranean Sea. In the Holocene, valves of K. compressa have been reported from the Tasman Sea and from off southern Australia at water depths ranging from 759 to 3281 m ( et al., 1999). K. compressa is distributed from the Miocene to the Quaternary in North Atlantic ( et al., 1994) and from the Miocene to the Pleistocene in the Mediterranean basin ( et al., 2000).
Family Trachyleberididae -, 1948
Genus Quasibuntonia , 1958
Type species: Quasibuntonia radiatopora (, 1880)
Quasibuntonia radiatopora sculpta (, 1880)
(Pl. 2 ![]() ,
fig. P)
,
fig. P)
1880 Cythere radiatopora sculpta , p. 193;
1954 Buntonia radiatopora sculpta : , p. 562, Figs. 17-17a;
1958 Quasibuntonia radiatopora sculpta (): , Figs. 22-22a;
1965 Buntonia (Quasibuntonia) radiatopora sculpta (): , p. 100, Pl. 12, fig. 3;
1973 Quasibuntonia radiatopora (): , p. 67, Fig. 3 (4);
1980 Quasibuntonia sculpta (): & , p. 64, Pl. 27, figs. 3-5;
2008 Quasibuntonia radiatopora (): & , p. 35, tab. 1, Pl. 1, fig. 10;
2014 Quasibuntonia radiatopora sculpta (): , p. 5,
Pl. 1 ![]() , figs. C-D.
, figs. C-D.
(2005) considered this subspecies as a simple morphological variant of Q. radiatopora (), because differences between the two subspecies are minimal and, above all, because the two morphotypes are commonly found associated. Here, following (1954, 1958), the distinction between the two subspecies is maintained based on the presence or absence of the ornamentation in the anterior part of the carapace, that cannot be considered minimal. The genus Quasibuntonia () is known as a bathyal taxon (, 1958), and Q. radiatopora was reported among the psychrospheric ostracods by (1972, 1973). The species is known from the Lower Pliocene (, 1954, 1958) to the Lower Pleistocene ( & , 1988) and commonly reported from bathyal sediments of the Mediterranean Basin. At Serra San Biagio this species is common all along the section. In the Recent Q. radiatopora has benn recorded from between 600 and 2669 m wd in the Atlantic Ocean.
Genus Bathycythere , 1971
Type species: Bathycythere vanstraateni , 1971
Bathycythere vanstraateni , 1971
1971 Bathycythere vanstraateni , p. 410, Pl. 1, 2, figs. 2-4;
1971 Xandarosina n. nudum & -, p. 68, Figs. 3 a-b;
1975 Bathycythere vanstraateni , p. 213, Pl. 4, fig. 20;
1980 Bathycythere vanstraateni : & , p. 52, Pl. 4, figs. 7-8;
1983 Bathycythere vanstraateni : et al., p. 463;
1985 Bathycythere vanstraateni et al., Pl. 1, fig. 7;
1987 Bathycythere vanstraateni & , p. 81, Pl. 6, fig. 3;
1988 Bathycythere vanstraateni : & , p. 283;
1988 Bathycythere vanstraateni : & , p. 732, Pl. 1, fig. 1;
1990 Bathycythere vanstraateni : et al., p. 483;
1997 Bathycythere vanstraateni : & , p. 18;
2004 Bathycythere vanstraateni : , p. 48, Figs. 25 K-N.
B. vanstraateni is listed among the abyssal ostracods living in the Atlantic Ocean. It seems to be extinct in the Recent Mediterranean, from where it is known as fossil at shallower depths (, 1990). In the study on the Vrica section, B. vanstraateni is reported among psychrospheric ostracods by & (1997). In the Adriatic Sea (1975) found this species in sediments of the Upper Pleistocene including the Allerřd and the Younger Dryas climatic stages. In the Santa Maria di Leuca Coral Province this species is poorly represented and it seems restricted to the Framework Corals facies (FC) and to hardground fragments colonized by solitary coral growths at 775 m wd ( & , 2015). According to et al. (1990), in Hole 654 A (Tyrrhenian Sea), the last occurrence of B. vanstraateni occurs within the Glacial Pleistocene above the Pseudoemiliania lacunosa LAD, therefore the specimens of (1975) in the Adriatic sea have been reworked. According to & (1988) this species, as well as Krithe monosteracensis and Macropyxis adriatica, disappeared from the eastern Mediterranean about 9 Ka ago following an anoxic event related to sapropel deposition. According to - et al. (1985) Bathycythere vanstraateni and Macropyxis adriatica are cold-climatic markers.
Family Bythocytheridae , 1866
Genus Retibythere , 1981
Subgenus Bathybythere , 1987
Type species: Retibythere (Bathybythere) scaberrima (, 1887)
Retibythere (Bathybythere) scaberrima (, 1887)
(Pl. 2 ![]() ,
fig. R )
,
fig. R )
1972 Bythoceratina scaberrima (): , p. 68, Figs. 1, 3;
1973 Bythoceratina scaberrima (): , p. 1003, Fig. 2 B;
1974 Bythoceratina scaberrima (): et al., p. 173;
1987 Bythoceratina scaberrima (): & , p. 80.;
1990 Bythoceratina scaberrima (): & , p. 223;
2002 Bythoceratina scaberrima (): et al., p. 203;
2002 Bythoceratina scaberrima (): et al., Fig. 2 B;
2005 Bythoceratina scaberrima (): , p. 222;
2014 Retibythere (Bathybythere) scaberrima (): , p. 4;
2015 Bythoceratina scaberrima (): & , Fig. 5.C
This species is extinct in the modern Mediterranean Sea but it is considered a characteristic Mediterranean psycsthrospheric ostracod from Miocene to the Early Pleistocene (Early Calabrian, Santernian age, with the first appearance of Hyalinea balthica) (, 1972). Pleistocene findings include Le Castella section (, 1965), Vrica section ( & , 1980), Monte S. Nicola section ( et al., 2000) and Capo Milazzo (, 2005, 2014). & (1988) found this species in the Quaternary of Atlantic, Indian and Pacific Oceans and in the Holocene of the NE Atlantic Ocean.
According to et al. (2002), Retibythere scaberrima could be a relatively warm water-bathyal species characteristic of the post glacial period in Late Quaternary Arctic Ocean. The occurrence of this species would be allowed by an increase of the thermohaline circulation in the Nordic seas and an influx of warmer Atlantic deep water into the Arctic Ocean.
Palaeobiogeography
The nine samples analyzed from the Pleistocene section of the Serra S. Biagio quarry showed very similar ostracod associations. In all samples, the ostracod allochthonous component is very scarce, both in terms of abundance and species richness. The studied samples include a Pleistocene ostracod association that, in addition to a strong bathyal affinity, shows a remarkable Atlantic and/or oceanic affinity as well. Today, indeed, some of the species found have an Atlantic - Mediterranean distribution; they are Bairdoppilata conformis, Bythocypris obtusata, Argilloecia acuminata, Pseudocythere caudata, Henryhowella hirta. Other species, on the other hand, are no longer reported in the Recent of the Mediterranean Sea, and presently have either an Atlantic distribution (Cytherella carmela n. sp., Krithe aequabilis, Bathycythere vanstraateni, Quasibuntonia radiatopora, Retibythere (Bathybythere) scaberrima, Macrocyprissa arcuata) or an even wider (Atlantic-Pacific, Indo-Pacific) distribution (Krithe perpulchra, K. marialuisae, K. compressa, Macrosarisa bensoni, Anchistrocheles antemacella).
The data related to the Atlantic provenance ostracods are in agreement with the general trend of the invertebrate Mediterranean faunas during the Pliocene and the Pleistocene. According to et al. (1989), et al. (1996), and & (1998), there was a progressive establishment of invertebrate taxa of Atlantic provenance in the Mediterranean basin. This is allegedly presented as a consequence of the temperature lowering during the Pleistocene cold periods. However, how did the passage of the benthic fauna from the Atlantic to the Mediterranean happen and why have the deep-Mediterranean waters become so cold (4°C) are still a matter of controversy. The assessment of the only area where the Atlantic Ocean and the Mediterranean waters come into contact might therefore shed some new light on the issue.
According to (1972), et al. (1989), and et al. (1996), during the Pliocene, the threshold at Gibraltar was deeper than today due to the raising of the sea level (or even the absence of a real threshold at Gibraltar). This allowed the Mediterranean to be widely connected to the Atlantic and its deep-water masses to participate in the global oceanic circulation with the development of a psychrosphera with water temperature ranging between 4°C and 6°C in more than 1000 m depth. Deep and cold waters of polar origin, coming from the Atlantic (, 2005, inter alias) and entering the deep Mediterranean, allowed its colonization by some stenotermic and stenobathic taxa of Atlantic origin. This scheme explains the presence of psychrospheric taxa, such as Agrenocythere pliocenica, Quasibuntonia radiatopora, Krithe undecimradiata and Retibythere (Bathybythere) scaberrima in deep-water Pliocene sediments.
According to & (2004, 2008), during the Pleistocene the depth of the Mediterranean Sea was comparable to that of the present-day, the threshold at Gibraltar was located at about 300 m wd, and this prevented or limited the entry of the deep benthic faunas of Atlantic origin. Furthermore, during this time interval, benthic faunas were affected by Quaternary climatic fluctuations ( et al., 1989; et al., 1997). The migration of the Atlantic species in the Mediterranean basin was mainly determined by the change of the current system between the Atlantic Ocean and the Mediterranean Sea across the Strait of Gibraltar.
During glacial period, the Atlantic boreal-arctic cold stenothermic taxa both eurybathic and stenobathic (shelf taxa), such as Cytheropteron testudo and other Northern Guests colonized the Mediterranean basin ( & , 2011), while the psychrospheric, deeper-cold taxa either disappeared, as Agrenocythere pliocenica, or remained as residual faunas, as it is possibly the case for Bathycythere vanstraateni (, 2005). Presently, psychrospheric Atlantic waters cannot enter in the Mediterranean basin because the Gibraltar threshold is too shallow (, 1984). This causes the disappearance of psychrospheric and bathyal stenothermic taxa from the Mediterranean Sea.
According to 's study of the bathyal sharks of Serra san Biagio (2007), this palaeogeographic pattern can be extended to the bathyal Pleistocene vertebrate fauna.
Palaeodepth
This autochthonous association is significantly indicative of bathyal environments, although species that show a wide depth range are present. This datum is also supported by the presence of the Krithe group, one of the most common deep-sea taxa in the open ocean, and one that also occurs in abyssal assemblages (, 1978; , 2009, inter alias). According to (1984), the association structure shows features that are typical of bathyal environments, being constituted by few dominant species accompanied by many scarcely represented species. The similarities between the Serra S. Biagio ostracod fauna and the Monasterace ostracod fauna studied by et al. (1974) are particularly interesting. In fact, the species reported from both localities (Retibythere (Bathybythere) scaberrima, Quasibuntonia radiatopora, Anchistrocheles antemacella, Bathycythere vanstraateni and Krithe undecimradiata) are extremely significant from an ecological point of view and point to a paleodepth deeper than 800 m. Therefore, studied associations testify a sedimentary paleobasin with typically oceanic paleo-environmental conditions, characterized by very low temperatures, and located in the Bathyal Zone, approximately at a depth of about a thousand metres. Consequently, Cytherella carmela n.sp. can be considered as a bathyal stenothermic taxon of Atlantic affinity and it is expected that further findings will confirm this assumption.
This datum agrees with the sedimentation depth of the Pleistocene sediments cropping out in the Strait of Messina area that, while belonging to different palaeoenvironmental domains, contain a rich bathyal association of Atlantic affinity invertebrates ( et al., 1989; , 1995).
Stratigraphic remarks
The occurrence of the benthic foraminifer Hyalinea balthica in all samples suggests deposition during a Pleistocene cold period, not earlier than the Emilian Age. All the detected ostracod species have stratigraphical distributions, that although wider, include this stage. In particular, Quasibuntonia radiatopora, Krithe pernoides, K. marialuisae, K. undecimradiata, K. perpulchra and Retibythere (Bathybythere) scaberrima have been reported up to the Globigerina cariacoensis Zone ( et al., 1993), whereas Krithe compressa and Bathycythere vanstraateni are known up to the close of the Pleistocene and Krithe aequabilis until the early Holocene.
Some other species, even if considered allochthonous, may provide some useful stratigraphic information; among these, Aurila latisolea has been reported from the Pliocene to the Sicilian Stage.
The author is grateful to the reviewers and the editors for detailed suggestions and comments to the manuscript. Thanks are also due to Donald for language editing and to Mr. Alfio (University of Catania) for SEM photos assistance. Catania Palaeoecological Research Group contribution nº 401.
S., D., G. & G. (1993).- The genus Krithe , & , 1874 (Crustacea: Ostracoda) in the Pliocene-Early Pleistocene of the M. San Nicola section (Gela, Sicily).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 32, nº 3, p. 349-366.
G. & D. (2001).- Pliocene ostracod assemblages at the MPl 3-MPl 4 boundary in the Capo Rossello borehole (Agrigento, Sicily).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 40, nº 1, p. 97-103.
G., D. & G. (2000).- Systematics and biostratigraphy of the Ostracoda of the Plio-Pleistocene Monte S. Nicola section (Gela, Sicily).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 39, nº 1, p. 83-112.
G., D., G. & A. (1996).- The genus Cytherella , 1849 (Ostracoda) in the Italian Tortonian-Recent.- Revue de Micropaléontologie, Paris, vol. 39, nº 3, p. 171-190.
C.A. (2009).- Data report: late Quaternary ostracods at IODP Site U1314 (North Atlantic Ocean).- Proceedings of the Integrated Ocean Drilling Program, College Station, vol. 303-306, p. 1-22.
J.F. (1987).- Marine lower Pliocene Ostracoda of southern Spain with notes on the recent fauna.- Bulletin of the Geological institutions of the University of Uppsala, (N.S.), vol. 13, p. 1-94.
M., T., V. & R.C. (1996).- New Bythocytherid and Cytherurid ostracode species from the deep sea Australia, with enigmatic dorsal expansion.- Geobios, Villeurbanne, vol. 29, p. 73-90.
P.J. (1971).- Die Ostracoden des Kuestenbereiches von Naxos (Griechenland) und ihre Lebensbereiche.- Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, vol. 67, p. 255–326.
P., I., C., M. & H. (1989).- Présence de faunes bathyales atlantiques dans le Pliocene et le Pleistocene de Méditerranée (Détroit de Messine, Italie.- Bulletin de la Societé géologique de France, Paris, vol. 5, nº 4, p. 787-796.
R.H. (1972).- Ostracodes as indicators of threshold depth in the Mediterranean during the Pliocene. In: D.J. (ed.), The Mediterranean Sea: a natural sedimentation laboratory.- Dowden, Hutchinson & Ross, Stroudsburg, p. 63-73.
R.H. (1973).- Psychrospheric and continental Ostracoda from ancient sediments in the floor of Mediterranean.- Initial Report of the Deep-Sea Drilling Project, College Station, vol. 8, p. 1002-1009.
R.H. (1975).- The origin of the psychrosphere as recorded in changes of deep-sea ostracodes assemblages.- Lethaia, Oslo, vol. 8, p. 69-83.
R.H. (1978).- The paleoecology of the ostracods of DSDP Leg 42 A.- Initial Reports of Deep Sea Drilling Project, College Station, vol. 42, p. 777-787.
R.H. (1984).- Estimating greater paleodepths with ostracodes, especially in past thermospheric oceans.- Palæogeography, Palæoclimatology, Palæoecology, vol. 48, p. 107-141.
R.H. & P.C. (1971).- Deep-sea Ostracodes and the transformation of ocean to sea in the Tethys.- Bulletin du Centre de Recherches de Pau (SNPA), vol. 5, p. 63-92.
W.A. van den (1958).- Ostracoda from the Brasso Formation of Trinidad.- Micropaleontology, New York, vol. 4, nº 4, p. 391-418.
G., G. & M. (1975).- Distribution of Ostracoda in the Adriatic Sea.- Pubblicazioni della Stazione Zoologica di Napoli, Napoli, vol. 40 (Suppl.), 304 p.
G., B., M., G. & N. (1983).- The deep-water benthic ostracodes of the Mediterranean. In: R.F. (ed.), Applications of Ostracoda.- University of Houston, p. 459-471.
G. & N. (1979).- Benthic ostracods as depth indicators.- Rapport de la Commission International sur la Mer Méditerranée, Monaco, vol. 25/26, nº 4, p. 167-169.
G. & A. (1985).- The Miocene Ostradodes of Sardinia.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 23, nº 2, p. 421-437.
G.S. (1880).- Report on the Ostracoda dredged by H.M.S. Challenger during the years 1873-1876.- Report on the scientific results of the voyage of H.M.S. Challenger during the years 1873-1876, London, (Zoology), vol. I, part III, 184 p. (XLIV Pls.). URL: https://escholarship.org/uc/item/1287v4h5#page-196
G.S. (1887).- Notes on Entomostraca.- Report of the Fishery Board of Scotland, Glasgow, vol. 5, Appendix 11, p. 328-330.
G.S. (1890).- XIV. On Ostracoda collected by H.B. in the South Sea islands.- Transactions of the Royal Society of Edinburgh, vol. 35, p. 489-525 (Pls. I-IV).
G.S., H.W. & D. (1874).- A monograph of the post-tertiary Entomostraca of Scotland including species from England and Ireland. URL: https://doi.org/10.5962/bhl.title.84825
G.S. & A.M. (1889).- A monograph of the marine and freshwater Ostracoda of the North Atlantic and of the Northwestern Europe.- Section I. Podocopa.- The Scientific Transactions of the Royal Dublin Society, (Series II), vol. IV, p. 63-270 (Pls. VIII-XXIII). URL: http://www.biodiversitylibrary.org/part/59836#/summary
S.N. (2004).- Brazilian deep-sea Macrocyprididae , 1912 (Crustacea, Ostracoda, Macrocypridoidea).- Arquivos do Museu Nacional, Rio de Janeiro, vol. 62, nº 2, p. 151-172.
H. (1855).- Die mikroskopische Fauna des Septarienthones von Hermsdorf bei Berlin.- Zeitschrift der Deutschen Geologischen Gesellschaft, Berlin, Band 7, p. 307-371.
E. (1975).- Ostracodes in a bottom core from the deep southeastern basin of the Adriatic Sea. I, II.- Koninklijke Nederlandse Akademie van Wetenschappen, Procedings, Amsterdam, (Ser. B), vol. 78, p. 198-218.
R., P., A. & F.P. (1996).- Paleogeography and structure of the central Mediterranean: Sicily and its offshore area.- Tectonophysic, vol. 260, p. 291-323.
G. (1986).- Ostracodi del limite Tortoniano-Messiniano in alcune sezione italiane.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 24, nº 1, p. 29-110.
M.L. (1965).- Gli Ostracodi della serie di Le Castella (Calabria).- Giornale di Geologia, Bologna, vol. XXXIII, p. 83-123.
M.L. & G. (1980).- La ostracofauna plio-pleistocenica della sezione della Vrica in Calabria (con considerazioni sul limite Neogene-Quaternario).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 19, nº 1, p. 44-126.
M.L. & G. (1988).- Ostracofauna plio-pleistocenica batiale rinvenuta nel Pozzo 654A dell'ODP Leg 107 (Mar Tirreno occidentale).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 27, nº 3, p. 277-289.
M.L., G., A. & R. (1990).- Relationship between the benthic foraminifers and the ostracodes in the Pliocene-Pleistocene Tyrrhenian deep-sea record (ODP LEG 107, site 654).- Proceedings of the Ocean Drilling Program Scientific Results, College Station, vol. 107, p. 479-493.
G.P., N.R., R.C. & R.W. (1996).- Foraminifera and Ostracoda from Quaternary carbonate mounds associated with gas seepage in the Porcupine Basin, offshore Western Ireland.- Revista Espańola de Micropaleontologia, Madrid, vol. 28, nº 2, p. 113-151.
H.N., C.H. & P.H. (1935).- Bairdoppilata, a new genus of Ostracoda, with two new species.- American Museum Novitates, New-York, nº 777, p. 1-5.
O.G. (1853).- Paleontologia del regno di Napoli. Parte III.- Giorgio Franz, Monaco, 196 p.
R., F., G. & R. (1979).- Integrazione dei dati geologici, geofisici e petrografici per la stesura di un profilo crostale in corrispondenza dell'Etna.- Bollettino della Societŕ Geologica Italiana, Roma, vol. 98, p. 239-247.
T.M., I., G.S. & J. (2002).- Ostracoda and paleoceanography. In: J.A. & A.R. (eds.), The Ostracoda, application in Quaternary research.- Geophysical Monograph, Washington, vol. 131, p. 99-119.
G., S., S. & M. (2014).- New evidence for Late Quaternary deformation of the substratum of Mt. Etna volcano (Sicily, Italy): clues indicate active crustal doming.- Bulletin of Volcanology, vol. 76, nº 816, p. 1-13.
I. (1995).- Benthic assemblages of the Plio-Quaternary soft and hard substrata in the Strait s of Messina area. In: L., A. & E. (eds.), The Straits of Messina ecosystem.- Proceedings of the symposium held in Messina (4-6 April 1991), Messina, p. 105-118.
I. (2000).- Faune quaternarie di Acitrezza.- Cutgana, Catania, p. 1-16.
I., R., R., a. & R. (1997).- Cooling evidence from Pleistocene shelf assemblages in SE Sicily. In: M.B. (ed.), Climates, Past and present.- Geological Society, Special Publication, London, vol. 181, p. 113-120.
I., R. & A. (1996).- The Plio-Quaternary evolution of the deep-sea Mediterranean benthos: an outline. In: Proceedings of the Interim Colloquium "La Méditerranée : variabilité climatique, environnement et biodiversité", p. 286-291.
A. & S. (2002).- Long-term uplift rate of the Etna volcano basement (Southern Italy) based on biochronological data from Pleistocene sediments.- Terra Nova, vol. 14, p. 61-68.
C., H.A. & J.P. (2002).- Late Quaternary deep-sea ostracodes in the polar and subpolar North Atlantic: paleoecological and palaeoenvironmental implications.- Palæogeography, Palæoclimatology, Palæoecology, vol. 184, p. 195-212.
R.V. & A.R. (1990).- Benthic ostracods and deep water-masses in the Atlantic Ocean.- Palæogeography, Palæoclimatology, Palæoecology, vol. 80, p. 213-235.
C. & P. (2004).- The Mediterranean deep-sea fauna: historical evolution, bathymetric variations and geographical changes.- Carnets Geol., Madrid, vol. 4, nº A01 (G2004_A01), 10 p.
C. & P. (2008).- Les échanges de la faune profonde en mer Méditerranée. In: C. & P. (eds.), La Méditerranée autour de ses iles.- Actes des Congrčs nationaux des Sociétés historique et scientifiques, Paris, p. 31-43.
C. & E. (2011).- A revision of the "northern guest" Ostracoda (Crustacea) occurrence in the Quaternary of the Mediterranean area.- Il Quaternario, Roma, vol. 24, nº 1, p. 75-92.
P. & K.M. (2008).- Global chronostratigraphical correlation table for the last 2.7 million years.- Episodes, Beijing, vol. 31, nº 2, p. 243-247.
A., G. & R. (1974).- La sezione calabriana di Monasterace (Calabria).- Bollettino della Societŕ Geologica Italiana, Roma, vol. 93, p. 151-179.
C. (2005).- Ostracodes et stratigraphie du Néogčne et du Quaternaire méditerranéens.- Revue de micropaléontologie, Paris, vol. 48, p. 83-121.
C. & F. (1989).- Ostracodes et recherche des milieux anciens : Possibilités et limites.- Bulletin de la Société géologique de France, Paris, vol. 8, nº 5, p. 577-588.
D. van (1984).- A model of estuarine circulation in the Pliocene Mediterranean based on new ostracod evidence.- Nature, vol. 312, p. 359-361.
D. van (1986).- Use of ostracodes to recognize downslope contamination in paleobathymetry and a preliminary reappraisal of the paleodepth of Prasas Marls (Pliocene), Crete, Greece.- Geology, vol. 14, p. 856-859.
D. van (1990).- Modern abyssal ostracod faunas of the eastern Mid-Atlantic Ridge area in the North Atlantic and a comparison with the Mediterranean. In: R. & C. (eds.), Ostracoda and global events.- Chapman & Hall, London, p. 321-328.
D. van & H.J. (1988).- Mediterranean deep-sea ostracods, the species poorness of the Eastern Basin as a legacy of an Early Holocene anoxic event. In: T., N. & K. (eds.), Evolutionary biology of Ostracoda.- Proceeding of the Ninth International Symposium on Ostracoda, Shizuoka, p. 721-737.
T.R. (1849).- A monograph of the Entomostraca of the Cretaceous formation in England.- Palæontological Society, London, 39 p. (VII Pls.). URL: https://archive.org/details/monographofentom00jone
G., A. & M. (1999).- Retroricoprimento medio Pleistocenico di Argille Scagliose a Serra San Biagio (Sicilia Orientale): evidenze stratigrafiche e tettoniche.- Rendiconti dell'Accademia dei Lincei, Roma, vol. 10, p. 63-80.
C. (1858).- On the structure of lavas which consolidated on steep slopes with remarks on the mode of origin of Mount Etna and on the theory of craters of elevation.- Philosophical Transaction of the Royal Society of London, London, vol. 148, p. 703-786. URL: http://rstl.royalsocietypublishing.org/content/148/703.full.pdf+html
R.F. (1969).- Revision of Recent Bairdiidae (Ostracoda).- United States Natural Museum Bulletin, Washington, vol. 295, p. 1-126.
R. (1990).- Living and fossil Macrocyprididae (Ostracoda).- The University of Kansas Paleontological Contributions, Lawrence, vol. 2, 404 p.
R., M.L. & F.R. (2009).- Podocopan ostracoda (Crustacea) from the Gulf of Mexico. In: L. & K. (eds.), Gulf of Mexico origin, waters and biota. Vol. 1, Biodiversity.- Texas University Press, Austin, p. 877-894.
M.I. (1958).- New ostracode genera and species.- Trudy VNIGRI, New Series, 115 (Microfauna SSSR, 9), p. 232-299.
S. (2007).- A new bathyal shark fauna from the Pleistocene sediments of Fiumefreddo (Sicily, Italy).- Geodiversitas, Paris, vol. 29, nº 2, p. 229-247.
I. (2004).- Taxonomy, biogeography and ecology of Quaternary benthic Ostracoda (Crustacea) from circumpolar deep water of the Emerald Basin (Southern Ocean) and the S Tasman Rise (Tasman Sea).- Senckenbergiana Maritima, Frankfurt-am-Main, vol. 35, p. 1-119.
M., B., F., G. & P. (1985).- Paleoclimatic record from 4 cores (Gulf of Taranto, Ionian Sea). Evidence from Foraminifera and Ostracoda.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 23, nº 1, p. 21-51.
M.E., N. & G. (1998).- Shelf ostracods distribution in the Italian seas. In: - S., E. & F. (eds.), What about Ostracoda!- Bulletin des Centres de Recherche Exploration Production elf-Aquitaine, Pau, vol. 20, p. 91-101.
G.W. (1894).- Die ostracoden des Golfes von Neapel und der angrenzenden meeresabschnitte.- Verlag von R. Friedländer und Sohn, Berlin, 743 p.
G.W. (1912).- Ostracoda.- Verlag von R. Friedländer und Sohn, Berlin, 437 p.
G. von (1830).- Ueber einige fossile Arten Cypris (, .) und Cythere (, , ).- Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, Heidelberg, vol. 1, p. 60-67.
A.M. (1862).- Contributions to British Carcinology. II. On species of Ostracoda new to Great Britain.- Annals and Magazine of Natural History, London, (Third Series 3), vol. IX, p. 43-52 (Pls. II-III).
G. & M.L. (1997).-The Pliocene Pleistocene boundary stratotype at Vrica, Italy. In: J. van (ed.), The Pliocene-Pleistocene boundary and the beginning of the Quaternary.- Cambridge University Press, Cambridge, MA, p. 15-45.
H.S., G. & A.M. (1969).- Distribution of Ostracoda in the Mediterranean. In: J.W. (ed.), The taxonomy, morphology and ecology of Recent Ostracoda.- Oliver & Boyd, Edinburgh, p. 358-411.
F.A. (1840-1841).- Die Versteinerungen des nordeutschen Kreidegebietes.- Hahn'schen Hofbuchhandlung, Hannover, 145 p. (XVI Pls.). URL: https://archive.org/details/dieversteinerung00roem
A. & I. (1998).- Deep-sea Pleistocene Bryozoa of Southern Italy.- Geobios, Villeurbanne, vol. 30, nº 3, p. 303-317.
A., A., I., R., F., R., D., C., M., F. & A. (2010).- Hard- and soft-bottom thanatofacies from the Santa Maria di Leuca deep-water coral province, Mediterranean Sea.- Deep-Sea Research. Part II. Topical Studies in Oceanography, vol. 57, p. 360-379.
G. (1954).- Iconografia degli ostracodi marini del Pliocene e del Pleistocene italiani.- Atti della Societŕ Italiana di Scienze Naturali, Milano, vol. 93, nº 3-4, p. 561-575.
G. (1958).- Alcuni Ostracodi del Neogene Italiano.- Atti della Societŕ Italiana di Scienze Naturali, Milano, vol. 97, nº 2, p. 127-149.
G. (1962).- Alcuni ostracodi quaternari e recenti pertinenti al genere Costa Neviani.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 1, p. 3-9.
G. (1974).- Revisione della ostracofauna marina di Imola (Bologna).- Revista Espańola de Micropaleontologia, Madrid, vol. 6, p. 419-446.
G.O. (1866).- Oversigt af Norges marine Ostracoder.- Forhandlinger i Videnskabs - Selskabet i Christiania, vol. 7, p. 1-30.
G.O. (1922-1928).- An account of the Crustacea of Norway. Vol. IX: Ostracoda.- Bergen Museum, 277 p.
S. (1900).- Revisione della fauna post-pliocenica dell'argilla di Nizzeti presso Acicastello (Catania).- Atti dell'accademia Gioenia di Scienze Naturali in Catania, vol. 13, p. 1-26.
E.I. (1981).- Ostracods of the Family Bythocytheridae of Far Eastern seas.- Nauka, Moscow, p. 278.
F. (2003).- Dati preliminari sulla ostracofauna pliocenica di Capo Milazzo (Sicilia NE).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 42, nº 1-2, p. 179-184.
F. (2005).- Ostracodi batiali pleistocenici di Capo Milazzo (Sicilia NE) ed implicazioni paleoambientali.- Rendiconti della Societŕ Paleontologica Italiana, Modena, vol. 2, p. 219-227.
F. (2014).- Ostracods of the Upper Pliocene - Pleistocene Punta Mazza succession (NE Sicily) with special focus on the Family Trachyleberididae , 1948, and description of a new species.- Carnets Geol., Madrid, vol. 14, nº 1, p. 1-13.
F. & A. (2008).- Distribution pattern of deep-water ostrad assemblages in the Lower Pleistocene sediments from Furnari, Sicily.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 47, nº 1, p. 33-43.
F. & A. (2015).- Bathyal ostracods from the Santa Maria di Leuca deep-water coral province.- Palaeontologia Electronica, vol. 18, nº 2, 18.2.22A, 17 p.
F., A. & S. (2003).- Associazioni ad ostracodi del Pliocene di Centuripe (Sicilia): interpretazione paleoambientale.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 42, nº 3, p. 267-280.
G. (1862).- Prime ricerche intorno ai Rizopodi fossili delle argille pleistoceniche dei dintorni di Catania.- Atti dell'Accademia Gioenia di Scienze Naturali in Catania, vol. 18, 42 p.
G. (1879-1880).- Le formazioni terziarie nella provincia di Reggio (Calabria).- Regia Accademia dei Lincei, Memorie, Roma, Classe di scienza fisiche matematiche e naturali, (Serie 3a), vol. VI, 446 p. (XV Pls.). URL: https://archive.org/details/leformazioniterz00segu
G. (1883a).- Gli ostracodi dei periodi terziarii e quaternarii viventi nel mare di Messina.- Bollettino della Societŕ Geologica Italiana, Roma, vol. 2, p. 57-63.
G. (1883b).- Il Quaternario di Rizzolo. II Gli Ostracodi.- Naturalista Siciliano, Palermo, 1883, anno II vol. 11, p. 256-259.
W. (1971).-Tricostate Trachileberidinae (Ostracoda) from Neogene-Recent deposits of Europe.- Koninklijke Nederlandse Akademie van Wetenschappen, Procedings, Amsterdam, (Ser. B), vol. 74, p. 195-205.
W. (1972).- Late Cenozoic Ostracoda in the South Aegean Island Arc.- Utrecht Micropaleontological Bulletins, vol. 6, 187 p. (12 Pls.). URL: http://dspace.library.uu.nl/handle/1874/205810
A.J. & D.J. (2002).- Ecology of marine, marginal marine and nonmarine Ostracodes.- In: J.A. & A.R. (eds.), The Ostracoda: applications in Quaternary research.- Geophysical Monograph Series, Washington D.C., vol. 131, p. 37-64.
P.C. (1948).- The ostracode genus Cythereis.- Journal of Paleontology, vol. 22, p. 792-797.
O. (1878).- Les foraminifčres et les entomostracés-ostracodes du Pliocčne supérieur de l'Ile de Rhodes.- Deuxičme Section (ostracodes).- Mémoires de la Société géologique de France, Paris, vol. 3, p. 81-135.
V. (1981, unpublished).- Contribution to the study of Pleistocene of Zakinthos Island, W Greece (Ostracoda - Palaeoenvironment).- PhD thesis, University of Patras, 134 p.
R. & G. (1987).- The Late Miocene to Quaternary ostracoda of leg 94, deep sea drilling project.- Revista Espańola de Micropaleontologia, Madrid, vol. 19, nº 1, p. 33-97.
R. M. (1988).- Pandemic and endemic distribution pattern in Quaternary deep sea ostracods, In: T., N. & K. (eds.), Evolutionary biology of Ostracoda. Its fundamentals and applications.- Developments in Paleontology and Stratigraphy, vol. 11, p. 739-755.
M. & H. (2015).- Late Quaternary deep-sea ostracod taxonomy of the eastern North Atlantic Ocean.- Journal of Micropalaeontology, London, vol. 34, nº 1, p. 21-50.
M., H. & T. (2009).- Taxonomy of Quaternary deep-sea ostracods from the Western North Atlantic Ocean.- Palaeontology, London, vol. 52, nº 4, p. 879-931.
C., F., P. & M. (2011).- Distribution des ostracods dans les sédiments de subsurface de la Sebkha el-Guettiate (Skhira, Golge de Gabčs). Intéręt pour la reconstitution des paléo-environnements de l'Holocčne.- Carnets Geol., Madrid, vol. 11, nº A03 (G2011_A03), p. 63-81.
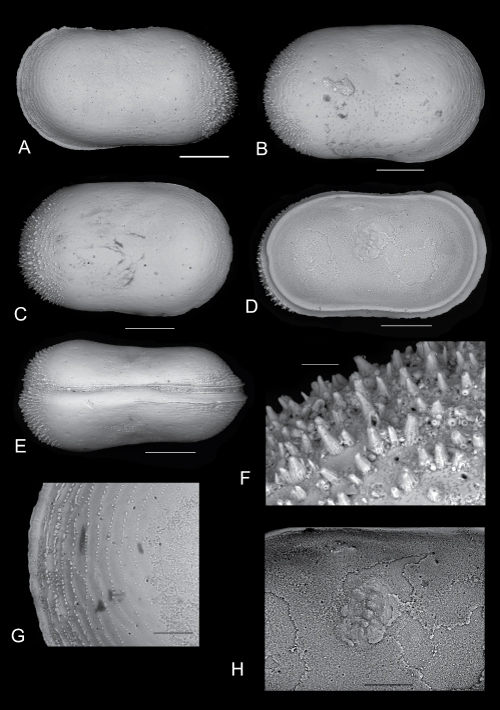
Click on thumbnail to enlarge the image.
Cytherella carmela n. sp.
A - Left valve. Paratype PMC. O48 P. 06.11.2014 external lateral view (scale bar 200µm);
B - Right valve. Paratype PMC. O49 P. 06.11.2014 external lateral view (scale bar 200µm);
C - Right valve. Paratype PMC. O50 P. 06.11.2014 external lateral view (scale bar 200µm);
D - Left valve. Paratype PMC. O51 P. 06.11.2014 internal lateral view (scale bar 200µm);
E - Complete carapace. Hoolotype PMC.O13 H 06/11/2014. Dorsal view (scale bar 200µm);
F - Detail of the posterior part of the carapace ornamented by spines. (scale bar 20µm);
G - Detail of the anterior part of the carapace ornamented by papillae festooned (scale bar 50µm);
H - Muscle scars (scale bar 100µm).
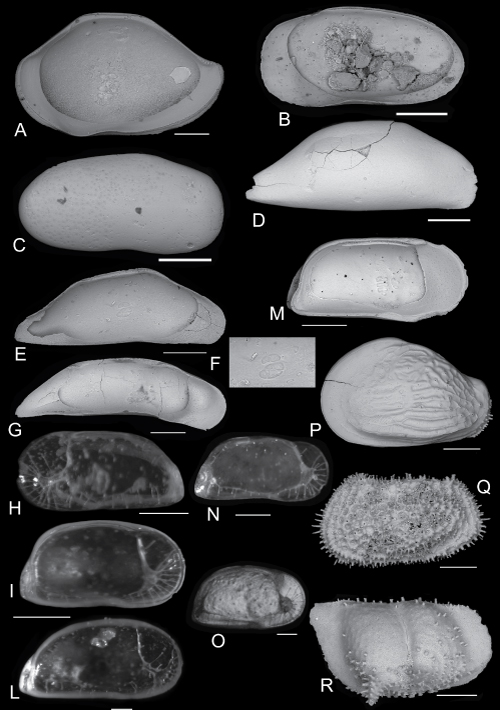
Click on thumbnail to enlarge the image.
A - Bairdoppilata subdeltoidea (, 1830) var. conformis (, 1878). Right valve internal lateral view (scale bar 200µm);
B-C - Anchistrocheles antemacella , 1969. B - right valve internal lateral view. C - right valve external lateral view (scale bar 200µm);
D-F Macrosarisa bensoni , 1990. D - Right valve external lateral view. E - left valve internal lateral view (scale bar 200µm). F - muscle scars (scale bar 100µm);
G - Macrocyprissa arcuata ( & , 1980). Left valve internal lateral view (scale bar 200µm);
H - Krithe pernoides (, 1855). Left valve external lateral view (scale bar 200 µm);
I - Krithe marialuisae et al., 1993. Left valve internal lateral view (scale bar 200 µm);
L - Krithe undecimradiata , 1974. Left valve internal lateral view (scale bar 100 µm);
M-N - Krithe aequabilis , 1986. Left valve internal lateral view. N - right valve external lateral view (scale bar 200 µm);
O - Krithe compressa (, 1880). Right valve external lateral view (scale bar 200 µm);
P - Quasibuntonia radiatopora sculpta (, 1880). Left valve external lateral view (scale bar 200 µm);
Q - Henryhowella ex H. hirta (, 1853) group. Left valve external lateral view (scale bar 200 µm);
R - Retibythere (Bathybythere) scaberrima (, 1887). Right valve external lateral view (scale bar 200 µm).
Table 1: List of the Ostracod species (in order of appearance) found in the Serra San Biagio quarry (X = 1-5 specimens; XX= 5-10 specimens; XXX= 10-20 specimens; XXXX = > 20 specimens).
| Species | Samples | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| IN SITU ASSEMBLAGES | ||||||||||
| Anchistrocheles antemacella (, 1969) | xxx | xx | xx | xx | xx | x | x | |||
| Bairdoppilata conformis , 1878 | xx | xx | xx | xx | x | x | x | x | ||
| Cytherella carmela n.sp. | xx | xx | x | xx | x | x | x | x | x | |
| Krithe compressa (, 1880) | xx | xx | x | x | x | x | x | x | ||
| Krithe marialuisae et al., 1993 | x | x | x | x | x | x | x | x | ||
| Cytheropteron bifidum & , 1980 | x | x | x | x | x | x | ||||
| Bythocypris obtusata (, 1866) | xxx | xx | xx | x | x | |||||
| Cytheropteron omega et al., 1996 | x | x | ||||||||
| Pseudocythere caudata , 1866 | x | x | ||||||||
| Argilloecia spp. | x | x | x | x | ||||||
| Retibythere (Bathybythere) scaberrima (, 1887) | x | x | x | x | x | x | x | xx | ||
| Henryhowella ex H. gr. hirta (, 1853) | x | x | x | x | x | |||||
| Quasibuntonia radiatopora sculpta , 1880 | x | x | xx | xx | xx | x | x | |||
| Macrosarisa bensoni 1990 | x | x | x | x | x | x | x | |||
| Paracytherois striata , 1894 | x | x | ||||||||
| Paradoxostoma simile , 1894 | x | |||||||||
| Krithe undecimradiata , 1974 | xxxx | x | x | x | ||||||
| Macrocyprissa arcuata ( & , 1980) | x | xx | x | x | ||||||
| Argilloecia acuminata , 1894 | x | |||||||||
| Monoceratina oblita et al., 1975 | x | |||||||||
| Krithe frutex et al., 1993 | x | |||||||||
| Krithe pernoides (, 1855) | x | |||||||||
| Pterygocythereis sp. | x | x | x | |||||||
| Bathycythere vanstraateni , 1971 | x | |||||||||
| Krithe monosteracensis , 1885 | xxx | |||||||||
| Pontocypris acuta et al., 1976 | x | |||||||||
| Rectobuntonia sp. | x | x | ||||||||
| Krithe perpulchra et al., 1993 | xxx | |||||||||
| Bythocypris sp. | x | |||||||||
| Sclerochilus contortus , 1866 | x | |||||||||
| ALLOCHTONOUS | ||||||||||
| Aurila spp. | x | x | x | x | x | x | x | x | ||
| Celtia sp. | x | x | ||||||||
| Costa punctatissima , 1961 | x | x | ||||||||
| Aurila (Cimbaurila) latisolea , 1975 | x | |||||||||
| Cytherelloidea beckmanni , 1971 | x | |||||||||
| Graptocythere hscripta (, 1849) | x | x | x | x | ||||||
| Loxoconcha minima , 1894 | x | x | x | x | x | x | x | x | ||
| Mutilus sp. | x | x | x | |||||||
| Tenedocythere prava , 1850 | x | x | x | |||||||
| Urocythereis favosa , 1838 | x | x | x | |||||||