◄ Carnets Geol. 21 (10) ►
![]()
Outline:
[1. Introduction]
[2. Geological and palaeontological framework]
[3. Systematics]
[4. General discussion] and ... [Bibliographic references]
Dipartimento di Scienze
della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Museo di Storia Naturale, Universitŕ di Pisa, via
Roma 79, 56011 Calci (Italy)
Dipartimento di Scienze
della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Corso di Dottorato Regionale in Scienze della Terra "Pegaso", via Santa Maria 53, 56126 Pisa
(Italy)
Gruppo Avis Mineralogia e Paleontologia
Scandicci, p.za Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (Italy)
Dipartimento di Scienze
dell'Ambiente e della Terra, Universitŕ degli Studi di Milano-Bicocca, Piazza della Scienza 4, Milano
20126 (Italy)
Gruppo Avis Mineralogia e Paleontologia
Scandicci, p.za Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (Italy);
Studio Tecnico Geologia e Paleontologia, via F.lli Rosselli 4, S. Casciano Val di Pesa,
50026 Firenze (Italy)
Published online in final form (pdf) on June 24, 2021
DOI 10.2110/carnets.2021.2110
![]()
[Editor:
Bruno R.C. Granier; language editor: Stephen Carey]
![]()
Aetobatus (Myliobatiformes: Aetobatidae) is a living genus of eagle rays that occurs in shallow-marine, tropical and subtropical environments of the Atlantic, Pacific, and Indian Oceans. Nowadays, Aetobatus does not inhabit the cool- to warm-temperate European and Mediterranean waters, though it is known from this broad region by virtue of several fossil teeth ranging chronostratigraphically from the lower Palaeogene to the upper Neogene. The present paper reports on a fossil aetobatid tooth discovered in mid-Pliocene (upper Zanclean to lower Piacenzian, 3.82-3.19 Ma) marine deposits exposed in the vicinities of Certaldo (Tuscany, Italy) and identified as belonging to †Aetobatus cf. cappettai. This specimen comprises the youngest occurrence of Aetobatus along the coasts of mainland Europe; furthermore, together with previous finds from roughly coeval deposits of Mallorca (Balearic Islands, Spain), it represents the most recent record of this genus in the whole Euro-Mediterranean region. In light of the environmental preferences of extant Aetobatus spp., our discovery suggests palaeoenvironmental conditions favourable to the persistence of tropical/subtropical taxa of "Miocene survivors" along the Pliocene coasts of Tuscany. In addition, it raises the question of whether or not the Messinian Salinity Crisis really resulted in the complete collapse of the Mediterranean marine biota and in the subsequent recolonisation of the Mediterranean Basin from the adjoining Atlantic waters and/or scattered marginal intrabasinal refugia at the beginning of the Pliocene. The possibility of Aetobatus recolonising the Mediterranean Sea through the Suez Canal in the near future is discussed.
• Aetobatidae;
• climate change;
• conservation palaeobiology;
• Myliobatiformes;
• palaeobiogeography;
• palaeoecology;
• palaeoichthyology;
• Tuscany
Collareta A., Merella M., Casati S., Coletti G. & Di Cencio A. (2021).- Another thermophilic "Miocene survivor" from the Italian Pliocene: A geologically young occurrence of the pelagic eagle ray Aetobatus in the Euro-Mediterranean region.- Carnets Geol., Madrid, vol. 21, no. 10, p. 203-214.
Un autre "survivant miocčne" thermophile du Pliocčne italien : Une occurrence précoce de l'aigle de mer pélagique Aetobatus dans la région euro-méditerranéenne.- Le genre Aetobatus (Myliobatiformes : Aetobatidae) est un genre actuel de raies aigles vivant dans les environnements tropicaux et subtropicaux marins peu profonds des océans Atlantique, Pacifique et Indien. De nos jours, Aetobatus n'habite plus les eaux tempérées de l'Europe et de la Méditerranée, bien qu'il soit connu dans cette vaste région par le recensement de plusieurs dents fossiles dont la distribution chronostratigraphique s'étale du Paléogčne inférieur au Néogčne supérieur. Le présent article décrit une dent fossile d'Aetobatidae, identifiée comme appartenant ŕ †Aetobatus cf. cappettai, découverte dans les dépôts marins du Pliocčne moyen (3,82-3,19 Ma, Zancléen supérieur - Piacenzien inférieur) affleurant dans les environs de Certaldo (Toscane, Italie). Ce spécimen représente l'occurrence la plus récente d'Aetobatus le long des côtes d'Europe continentale ; en outre, avec les découvertes antérieures de gisements plus ou moins contemporains de Majorque (Baléares, Espagne), il représente l'enregistrement fossile le plus récent de ce genre dans toute la région euro-méditerranéenne. Compte tenu des préférences environnementales des espčces actuelles d'Aetobatus, notre découverte évoque des conditions paléo-environnementales favorables ŕ la persistance de taxons "survivants miocčnes" ŕ affinités tropicales/subtropicales le long des côtes du Pliocčne de la Toscane. En outre, cela soulčve la question de savoir si la crise de salinité messinienne a abouti ou non ŕ l'effondrement complet du biote marin méditerranéen et ŕ la recolonisation ultérieure de ce bassin ŕ partir des eaux atlantiques voisines et/ou des refuges intra-bassinaux marginaux dispersés au début du Pliocčne. La possibilité de recoloniser la mer Méditerranée ŕ travers le canal de Suez dans un futur proche est enfin discutée pour ce qui concerne le genre Aetobatus.
• Aetobatidae ;
• changement climatique ;
• paléobiologie
de la conservation ;
• Myliobatiformes ;
• paléo-biogéographie ;
• paléoécologie ;
• paléo-ichtyologie ;
• Toscane
Among extant stingrays (Myliobatiformes: Myliobatoidei), those assigned to the pelagic eagle ray genus Aetobatus occur in shallow tropical and subtropical waters of the Atlantic, Pacific, and Indian Oceans (Weigmann, 2016; Last et al., 2016). Aetobatus mainly differs from the allied genera Aetomylaeus and Myliobatis in having a deeply notched internasal flap, spiracles that are placed on the dorsolateral region of the head, pectoral fins that join the head at the level of the eyes, broadly rounded free rear tips of the pectoral fins, and a single median dental file on each jaw at all growth stages (Capapé & Quignard, 1975; Compagno & Last, 1999; Cappetta, 2012; White & Moore, 2013; White, 2014; White & Naylor, 2016). Long regarded as belonging to the eagle ray family Myliobatidae, Aetobatus is now placed in its own family, the currently monotypic Aetobatidae (White & Naylor, 2016).
The Pliocene transitional and marine deposits of Tuscany (central Italy) have been the setting of remarkable discoveries of vertebrate fossils for several centuries (Collareta et al., 2020b, and references therein). Vertebrate finds from these Tuscan Pliocene successions include marine mammals (both odontocete and mysticete cetaceans as well as sea cows and rarer pinnipeds), sea turtles, bony and cartilaginous fishes, and even terrestrial forms (e.g., Dominici et al., 2018, and references therein). In spite of a long history of palaeontological research, discoveries of new taxa or somewhat "exotic" vertebrate specimens are still relatively frequent from the Tuscan Pliocene marine and paralic sediments (e.g., Spadini & Manganelli, 2015; Collareta et al., 2017a, 2017b, 2018, 2020a, 2020c, 2020d, 2021; Bianucci et al., 2019; Manganelli & Spadini, 2019).
Here, we report on the discovery of a fossil eagle ray tooth attributed to Aetobatus from mid-Pliocene marine deposits of Tuscany and briefly discuss its relevance with respect to palaeoecology, palaeobiogeography and conservation palaeobiology.
The myliobatoid fossil tooth
described herein was discovered by two of the authors (S.C. and A.D.C.) at an
outcrop in the vicinity of Certaldo village (Florence Province, Tuscany), in the
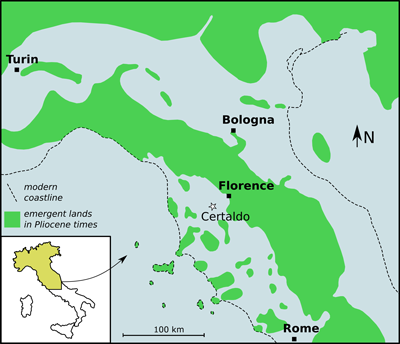
Valdelsa Basin (Fig. 1 ![]() ). The Universal Transverse Mercator (=UTM) geographic
coordinates of the finding site are: 32N 668021 4826620 (note that this outcrop
is located ca. 4 km east of that investigated by Collareta et al.,
2020d, 2021, and therein indicated as the "Certaldo
quarry").
). The Universal Transverse Mercator (=UTM) geographic
coordinates of the finding site are: 32N 668021 4826620 (note that this outcrop
is located ca. 4 km east of that investigated by Collareta et al.,
2020d, 2021, and therein indicated as the "Certaldo
quarry").
|
Figure 1:
Schematic palaeogeographic map of central and northern
Italy in Pliocene times, showing the location of the site where the aetobatid
tooth described herein was found. Palaeogeographical reconstruction redrawn and
modified after Vai (1989). |
The succession cropping out at this locality belongs to the S3 Synthem recognised in the upper Miocene to lower Pleistocene Valdelsa succession by Benvenuti et al. (2014). S3 is a large-scale depositional sequence that mostly consists of an alternation of sands and mudstones, interpreted as deltaic, overlain by shelf mudstones (Benvenuti et al., 2014). According to Benvenuti et al. (2014), the planktonic foraminiferal assemblages from the S3 mudstones belong to Mediterranean Pliocene (MPl) zone 4 of Cita (1975), whose bounding bioevents have been calibrated to 3.98 and 3.19 Ma (Violanti, 2012). Furthermore, the calcareous nannofossil assemblages from the same strata are indicative of Calcareous Nannofossil Plio-Quaternary (CNPL) zone 4 of the more recent Neogene biozonation scheme of Backman et al. (2012), whose bounding bioevents have been calibrated to 3.82 and 2.76 Ma (Benvenuti et al., 2014). In light of these considerations, the deposits cropping out at the finding site are referable to the 3.82-3.19 Ma interval, which is consistent with a late Zanclean to early Piacenzian time span.
At the study site, strata belonging
to S3 crop out along an artificial cliff. These deposits
consist of interbedded sands
and clayey sands. Vertebrate fossils are concentrated in a few decimeter-thick
horizons that feature abundant remains of bivalves (including veneroids,
ostreids and pectinids), gastropods, echinids and barnacles. Besides the
aetobatid specimen described herein, vertebrate fossils from the discovery site
include several teeth and dermal elements of bony and cartilaginous fishes,
including Carcharhinus brachyurus, Carcharhinus cf. falciformis,
Carcharias taurus, †Cosmopolitodus cf. plicatilis,
Dasyatidae indet., †Megascyliorhinus miocaenicus, Myliobatis
sp., Rhizoprionodon sp., Sparidae indet., Squatina sp. and
Trichiuridae indet. (Fig.
2 ![]() ), as well as some indeterminate otoliths.
), as well as some indeterminate otoliths.
|
Figure 2:
Selection of fossil fish remains (dental and dermal
elements) associated with the aetobatid specimen GAMPS-00967 from the Pliocene
locality of Certaldo (Tuscany, central Italy). a, Carcharias taurus,
b, †Cosmopolitodus cf. plicatilis, c, Dasyatidae
indet., d, Sparidae indet., e, Carcharhinus brachyurus, f,
Myliobatis sp., g, †Megascyliorhinus miocaenicus, h,
Carcharhinus cf. falciformis, i, Squatina sp., j,
Rhizoprionodon sp., k, Trichiuridae sp. Specimens a, b, e, g, h, i
and j are depicted in lingual view, specimens c, d, and f are depicted in apical
view, specimen k is depicted in profile view. All the figured specimens are
currently stored at the GAMPS. |
ELASMOBRANCHII Bonaparte, 1838
BATOMORPHII Cappetta, 1980
MYLIOBATIFORMES Compagno, 1973
MYLIOBATOIDEI Compagno, 1973
AETOBATIDAE Agassiz, 1858
Aetobatus Blainville, 1816
†Aetobatus cappettai Antunes & Balbino, 2006
†Aetobatus cf. cappettai
(Fig. 3 ![]() )
)
Referred material. One incomplete upper tooth, currently stored at Badia a Settimo (Scandicci, Italy), in the permanent exhibition of "Gruppo AVIS Mineralogia e Paleontologia Scandicci" (=GAMPS), under accession number GAMPS-00967.
Occurrence. Mid-Pliocene (3.82-3.19 Ma) marine deposits exposed in the vicinities of Certaldo, Tuscany, central Italy (see Section 2 above).
Remarks.
GAMPS-00967 measures 15.2 mm in maximum preserved width, 4.3 mm in maximum
preserved length, and 5.7 mm in maximum preserved height. Based on comparisons
with fossil and Recent teeth of Aetobatus, it is here identified as
comprising the leftmost third or quarter of an upper tooth, including the
well-preserved left lateral margin. The occlusal (Fig. 3.d ![]() ) and lingual
(Fig. 3.a
) and lingual
(Fig. 3.a ![]() )
faces of GAMPS-00967 are mostly pristine, whereas the basal (Fig.
3.c
)
faces of GAMPS-00967 are mostly pristine, whereas the basal (Fig.
3.c ![]() ) and labial
(Fig. 3.b
) and labial
(Fig. 3.b ![]() ) faces are locally damaged and the latter is slightly abraded. The
well-preserved lingual crown face of this specimen is vertical and gently
ornamented by means of weak costulae. These costulae are short, irregular and
subhorizontal in the vicinity of the inner ledge for tooth interlocking that
marks the crown base, whereas they appear as longer and more erect close to the
occlusal face (Fig. 3.a
) faces are locally damaged and the latter is slightly abraded. The
well-preserved lingual crown face of this specimen is vertical and gently
ornamented by means of weak costulae. These costulae are short, irregular and
subhorizontal in the vicinity of the inner ledge for tooth interlocking that
marks the crown base, whereas they appear as longer and more erect close to the
occlusal face (Fig. 3.a ![]() ). Such an ornamentation pattern strongly recalls that
observed on the labial crown face of upper teeth of extant members of Aetobatus
(Herman et al., 2000,
Pl. 26;
A.C., personal observation on uncatalogued materials kept at Universitŕ di
Pisa) while differing from the stronger ornamentation (consisting of pustules,
pits or wrinkles) that is found on the lingual crown face of most members of the
allied myliobatoid genera Myliobatis, Aetomylaeus (including
species that have previously been assigned to Pteromylaeus; White,
2014), and Rhinoptera (Herman et
al., 2000; Purdy et al.,
2001;
Cappetta, 2012; Hovestadt & Hovestadt-Euler,
2013).
). Such an ornamentation pattern strongly recalls that
observed on the labial crown face of upper teeth of extant members of Aetobatus
(Herman et al., 2000,
Pl. 26;
A.C., personal observation on uncatalogued materials kept at Universitŕ di
Pisa) while differing from the stronger ornamentation (consisting of pustules,
pits or wrinkles) that is found on the lingual crown face of most members of the
allied myliobatoid genera Myliobatis, Aetomylaeus (including
species that have previously been assigned to Pteromylaeus; White,
2014), and Rhinoptera (Herman et
al., 2000; Purdy et al.,
2001;
Cappetta, 2012; Hovestadt & Hovestadt-Euler,
2013).
Antunes & Balbino (2006, p. 44) diagnosed †Aetobatus cappettai as follows:
"Upper teeth narrow, long, slightly convex; crown low, thickened in the central part; lateral extremities prominent; root higher than crown, decreasing in height from the centre to the edges; blades on the labial face are almost smooth, lacking marked grooves; grooves on the lingual face very marked, they persist almost to the crown; basal surface of the root with narrow, deep grooves as well as blades, all of the same width; lower teeth arched and labially convex, extremely flat; basal angle ca 30°, with very oblique basal face; root high, thinning from the central part to the lateral edges; blades wide and separated by deep, wide grooves."
The same authors (op. cit., p. 44) characterised the upper teeth of †A. cappettai in these terms:
"The upper teeth are narrow, long and slightly convex. The crown is low and thicker in its central part. Lateral extremities are prominent; in occlusal view, the lateral extremities constitute a lateral border that is not affected by abrasion. The vertical, lingual face of the crown is separated from the root by a thin edge. The root is thicker than the crown; the thickness regularly decreases from the central part towards the borders (in lateral view). The basal surface presents alternating (narrow and shallow) grooves and blades. Width is the same in all blades, whose basal surface is flat. The tooth's labio-lingual section is nearly straight."
These descriptive notes are also largely
applicable to the morphology of the incomplete tooth GAMPS-00967. In particular,
the latter resembles the upper teeth of †A. cappettai in displaying: i)
a gentle posterior bending (Fig. 3.c-d ![]() ); ii) an apicobasally low crown
(Fig. 3.a-b
); ii) an apicobasally low crown
(Fig. 3.a-b ![]() ); iii) a distinct thickening of the crown close to the lateral margin of the
tooth (Fig. 3.a-b
); iii) a distinct thickening of the crown close to the lateral margin of the
tooth (Fig. 3.a-b ![]() ); iv) a lateral border of the crown that is well-defined, not
significantly abraded, nor laterally pointed (Fig. 3.c-d
); iv) a lateral border of the crown that is well-defined, not
significantly abraded, nor laterally pointed (Fig. 3.c-d ![]() ); v) a root whose
apicobasal thickness decreases progressively towards the lateral margin of the
tooth (Fig. 3.a-b
); v) a root whose
apicobasal thickness decreases progressively towards the lateral margin of the
tooth (Fig. 3.a-b ![]() ). Strong similarities are observed particularly between
GAMPS-00967 and the specimens depicted by Antunes & Balbino
(2006)
in their figures 1 and 2, the latter featuring the holotype of †A.
cappettai [we note here that the identifications of the lingual and labial
sides provided by Antunes and Balbino
(2006)
for these teeth may not be correct].
). Strong similarities are observed particularly between
GAMPS-00967 and the specimens depicted by Antunes & Balbino
(2006)
in their figures 1 and 2, the latter featuring the holotype of †A.
cappettai [we note here that the identifications of the lingual and labial
sides provided by Antunes and Balbino
(2006)
for these teeth may not be correct].
In light of the above considerations, GAMPS-00967 is here assigned to the genus Aetobatus. Owing to the incomplete nature of the specimen and the scarcity of species-diagnostic characters in the teeth of myliobatoids in general (Hovestadt & Hovestadt-Euler, 2013) and aetobatids in particular (Bor et al., 2012), it is here placed in open nomenclature as belonging to †Aetobatus cf. cappettai.
Recently, Reinecke et al. (2011), Bor et al. (2012), Cappetta (2012) and Fialho et al. (2019) have confirmed the validity of †A. cappettai. However, a deeper understanding of the alpha-diversity of living aetobatids, as well as a better knowledge of their dental morphology and variability, is nonetheless needed for reassessing the taxonomic status of several nominal species that are based on isolated fossil teeth whose morphology is closely comparable to the general dental design of extant Aetobatus, including †A. cappettai (Hovestadt & Hovestadt-Euler, 2013).
Hovestadt and Hovestadt-Euler (2013) highlighted several morphological similarities between the teeth of †A. cappettai and those of juveniles of extant A. narinari. GAMPS-00967 is alike in this regard. A juvenile status for GAMPS-00967 might be supported by its relatively small size, distinct from the larger dimensions of adult teeth of extant members of Aetobatus (A.C., personal observation on uncatalogued materials kept at Universitŕ di Pisa).
|
Figure 3:
†Aetobatus cf. cappettai (GAMPS-00967),
incomplete upper tooth from the Pliocene locality of Certaldo (Tuscany, central
Italy). a, lingual view; b, labial view; c, basal view; d, apical view. |
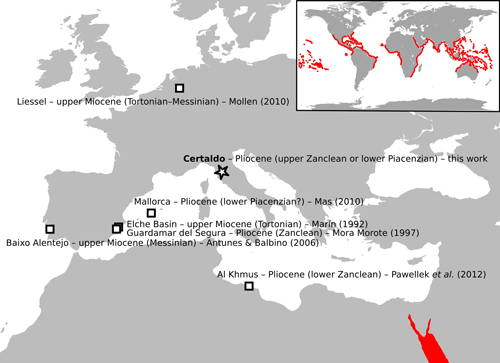
The eagle rays of the genus Aetobatus do
not currently inhabit the cool- to warm-temperate European and Mediterranean
waters (Serena, 2005) (Fig. 4 ![]() ). However, Aetobatus
has a long and relatively conspicuous palaeontological record in the
Euro-Mediterranean region, its earliest fossils including occurrences from early
and middle Eocene deposits of England, Belgium, Morocco and Egypt
(proto-Mediterranean area) (Marramŕ et al., 2018,
and references therein). During the Miocene, this genus was widespread in the
North Sea Basin (e.g., Bor et al., 2012),
western Paratethys (e.g., Höltke et al.,
2020),
central Paratethys (e.g., Villafańa et al.,
2020),
Atlantic western Europe (e.g., Jonet,
1968),
and Mediterranean Sea (e.g., Menesini, 1969),
though it declined in the European high latitudes starting from the middle
Miocene, possibly because of decreasing seawater temperatures (Reinecke et
al., 2011). As regards the upper Neogene
fossil record of Europe and the Mediterranean Basin, Aetobatus is a rare
genus, reported only from the Tortonian of continental Spain (Marín,
1992),
the Tortonian-Messinian of The Netherlands (Mollen, 2010),
the Messinian of Portugal (Antunes & Balbino,
2006),
the basal Pliocene of Libya (Pawellek et al., 2012), the Pliocene
of continental Spain (Mora Morote, 1997),
and the upper Pliocene of the Spanish island of Mallorca (Mas,
2010) (Fig.
4
). However, Aetobatus
has a long and relatively conspicuous palaeontological record in the
Euro-Mediterranean region, its earliest fossils including occurrences from early
and middle Eocene deposits of England, Belgium, Morocco and Egypt
(proto-Mediterranean area) (Marramŕ et al., 2018,
and references therein). During the Miocene, this genus was widespread in the
North Sea Basin (e.g., Bor et al., 2012),
western Paratethys (e.g., Höltke et al.,
2020),
central Paratethys (e.g., Villafańa et al.,
2020),
Atlantic western Europe (e.g., Jonet,
1968),
and Mediterranean Sea (e.g., Menesini, 1969),
though it declined in the European high latitudes starting from the middle
Miocene, possibly because of decreasing seawater temperatures (Reinecke et
al., 2011). As regards the upper Neogene
fossil record of Europe and the Mediterranean Basin, Aetobatus is a rare
genus, reported only from the Tortonian of continental Spain (Marín,
1992),
the Tortonian-Messinian of The Netherlands (Mollen, 2010),
the Messinian of Portugal (Antunes & Balbino,
2006),
the basal Pliocene of Libya (Pawellek et al., 2012), the Pliocene
of continental Spain (Mora Morote, 1997),
and the upper Pliocene of the Spanish island of Mallorca (Mas,
2010) (Fig.
4 ![]() ). The Libyan Pliocene occurrence, from near Al Khums and identified in
open nomenclature as belonging to Aetobatus sp., is found in a
transgressive lag on top of an erosional unconformity that is regarded as
representing the Messinian event (Pawellek et al.,
2012).
The shark and ray assemblage studied by Pawellek et al.
(2012)
is unique in featuring a high abundance of tropical elements (including the
lemon shark Negaprion and the snaggletooth shark Hemipristis,
neither of which is known from any other Plio-Quaternary site of the
Mediterranean Basin) as well as the coexistence of the large- to mega-toothed
mackerel sharks †Carcharocles, Carcharodon and †Cosmopolitodus
(Pawellek et al., 2012). On
the other hand, it is worth noting that the taxonomic composition of this Libyan fossil
fauna is reminiscent of the
purportedly Pliocene Angolan shark and ray assemblages studied by Antunes
(1977, 1978).
Teeth of †Aetobatus cf. arcuatus (Agassiz,
1843)
were described by Mora Morote (1997) from
the Pliocene of Guardamar del Segura, near Alicante, southern Spain. These teeth
are part of a diverse elasmobranch assemblage (see Mora Morote,
1996)
that likely originated in the ~5.3-4.19 Ma time span, corresponding
to the lower to mid-Zanclean (see Discussion in Collareta et al.,
2020d).
A third record of Aetobatus from the Mediterranean Pliocene was provided
by Mas (2010) and consists of some teeth
of Aetobatus sp. from calcarenite deposits regarded as representative of
the basal portion of the Sant Jordi Formation exposed in Mallorca, Balearic
Islands, Spain (e.g., Capó & Garcia,
2019).
A Piacenzian depositional age has been inferred for this unit on the sole basis
of the detection of Zanclean microfossils in the underlying Son Mir Formation,
whose upper strata pass gradually into the lowermost horizons of the Sant Jordi
Formation (Pomar et al., 1983).
This means that a Piacenzian age estimate is at best tentative for the
elasmobranch assemblage studied by Mas (2010).
Given also that the fossil-bearing deposits directly overlie finer sediments
assigned to the Zanclean (Mas, 2010), the
finds of Aetobatus from the Pliocene of Mallorca are possibly not younger
than early Piacenzian, and as such, indistinguishable in age from the better
chronostratigraphically constrained Italian fossil studied herein. In light of
these considerations, GAMPS-00967 comprises
the youngest occurrence of Aetobatus along the coast of mainland Europe
as well as in the central Mediterranean Basin. Furthermore, together with the
Mallorcan finds described by Mas (2010),
it represents the most recent record of this genus
in the whole Euro-Mediterranean region. Differing from previous palaeofaunistic
analyses (e.g., Marsili, 2008), our find corroborates the presence of the family
Aetobatidae in the Mediterranean Sea during the Pliocene, thus
improving our knowledge of the late Neogene Mediterranean biodiversity.
). The Libyan Pliocene occurrence, from near Al Khums and identified in
open nomenclature as belonging to Aetobatus sp., is found in a
transgressive lag on top of an erosional unconformity that is regarded as
representing the Messinian event (Pawellek et al.,
2012).
The shark and ray assemblage studied by Pawellek et al.
(2012)
is unique in featuring a high abundance of tropical elements (including the
lemon shark Negaprion and the snaggletooth shark Hemipristis,
neither of which is known from any other Plio-Quaternary site of the
Mediterranean Basin) as well as the coexistence of the large- to mega-toothed
mackerel sharks †Carcharocles, Carcharodon and †Cosmopolitodus
(Pawellek et al., 2012). On
the other hand, it is worth noting that the taxonomic composition of this Libyan fossil
fauna is reminiscent of the
purportedly Pliocene Angolan shark and ray assemblages studied by Antunes
(1977, 1978).
Teeth of †Aetobatus cf. arcuatus (Agassiz,
1843)
were described by Mora Morote (1997) from
the Pliocene of Guardamar del Segura, near Alicante, southern Spain. These teeth
are part of a diverse elasmobranch assemblage (see Mora Morote,
1996)
that likely originated in the ~5.3-4.19 Ma time span, corresponding
to the lower to mid-Zanclean (see Discussion in Collareta et al.,
2020d).
A third record of Aetobatus from the Mediterranean Pliocene was provided
by Mas (2010) and consists of some teeth
of Aetobatus sp. from calcarenite deposits regarded as representative of
the basal portion of the Sant Jordi Formation exposed in Mallorca, Balearic
Islands, Spain (e.g., Capó & Garcia,
2019).
A Piacenzian depositional age has been inferred for this unit on the sole basis
of the detection of Zanclean microfossils in the underlying Son Mir Formation,
whose upper strata pass gradually into the lowermost horizons of the Sant Jordi
Formation (Pomar et al., 1983).
This means that a Piacenzian age estimate is at best tentative for the
elasmobranch assemblage studied by Mas (2010).
Given also that the fossil-bearing deposits directly overlie finer sediments
assigned to the Zanclean (Mas, 2010), the
finds of Aetobatus from the Pliocene of Mallorca are possibly not younger
than early Piacenzian, and as such, indistinguishable in age from the better
chronostratigraphically constrained Italian fossil studied herein. In light of
these considerations, GAMPS-00967 comprises
the youngest occurrence of Aetobatus along the coast of mainland Europe
as well as in the central Mediterranean Basin. Furthermore, together with the
Mallorcan finds described by Mas (2010),
it represents the most recent record of this genus
in the whole Euro-Mediterranean region. Differing from previous palaeofaunistic
analyses (e.g., Marsili, 2008), our find corroborates the presence of the family
Aetobatidae in the Mediterranean Sea during the Pliocene, thus
improving our knowledge of the late Neogene Mediterranean biodiversity.
|
|
Figure 4: Distribution of fossil remains of Aetobatus in the upper Miocene and Pliocene of the Euro-Mediterranean region. Red shading indicates the modern range of Aetobatus in the Red Sea (main map) and worldwide (top right panel) as reported by Kyne et al. (2006). See the main text for further information. |
At present, Aetobatus accounts for five or six living species that are distributed in the warm waters of the Atlantic, Pacific and Indian Oceans (White et al., 2013; Last et al., 2016; Weigmann, 2016; Sales et al., 2019). In the Atlantic Ocean (including the Caribbean and Gulf of Mexico), Aetobatus is represented by the spotted eagle ray Aetobatus narinari and, possibly, by Aetobatus latirostris (a species whose validity is currently regarded as dubious; White & Naylor, 2016; Sales et al., 2019). Along the eastern Atlantic coast, A. narinari does not extend northwards of Cabo Verde and Senegal, ca. 19°N (Kyne et al., 2006; Sales et al., 2019). Interestingly, south of 20-22°N, the coast of western Africa is currently home to some mollusc species [e.g., Gastrana lacunosa (Chemnitz, 1782) and Tugonia anatina (Gmelin, 1791)] that inhabited the Mediterranean Sea until ca. 3 Ma, when they were extirpated from this basin following a major interval of climate cooling (Monegatti & Raffi, 2001). Considering the thermophilic nature of Aetobatus, its presence in the S3 deposits of Certaldo indicates tropical or subtropical affinities for the northwestern Mediterranean Sea during the mid-Pliocene, ca. 3.8-3.2 Ma. This is also suggested by the recent discovery of rostral spines of the knifetooth sawfish Anoxypristis from shelf mudstones, belonging to the same synthem, exposed at the nearby locality of Tegoliccio (Collareta et al., 2017a). Recent palaeothermometric estimates for the Mediterranean Sea during the Zanclean and early Piacenzian support surface seawater temperatures ca. 2-3°C higher than today (e.g., Prista et al., 2015; Ragaini et al., 2019; Coletti et al., 2021), that is, in line with the environmental preferences of the extant species A. narinari (Froese & Pauly, 2019). The eventual extirpation of Aetobatus from the Mediterranean Sea is likely to have occurred during some late Pliocene or early Pleistocene episode of climate change, as hypothesised for other thermophilic marine vertebrates (including Anoxypristis) that inhabited the Mediterranean Basin during the first part of the Pliocene and disappeared afterward (e.g., Sorbi et al., 2012; Collareta et al., 2017a).
Among elasmobranchs, and along with pristids, the aetobatids represent some of the most recently (re)discovered lineages of "Miocene survivors" in the Mediterranean Pliocene. Similar cases have also been pointed out among thermophilic invertebrates that were thought to have disappeared from the Mediterranean Sea at the end of the Miocene (e.g., the lingulid brachiopods; Di Cencio et al., 2021). This raises the question of whether or not the latest Miocene Messinian Salinity Crisis did in fact result in the collapse of the Mediterranean marine biota and in the subsequent recolonisation of the Mediterranean Basin from the adjoining Atlantic waters and/or scattered marginal intrabasinal refugia at the beginning of the Pliocene. Research focused on the bony fish record has recently questioned the reliability of such a reconstruction, revealing instead a remarkable degree of ecological homogeneity throughout the Messinian Salinity Crisis, with a nearly continuous presence of marine steno- and euryhaline taxa (Carnevale et al., 2006, 2008, 2018, 2019). Although data on the cartilaginous fishes are more fragmentary, and very few data exist regarding the Mediterranean elasmobranch fauna during the Messinian (e.g., Arambourg, 1927), major discontinuities are unlikely to emerge between the late Miocene and early to mid-Pliocene shark and ray assemblages of the Mediterranean Sea. More research and new sampling campaigns at stratigraphically constrained, elasmobranch-rich horizons from the upper Miocene and lower Pliocene of the Mediterranean region are definitely needed to shed further light on this palaeontological vexata quaestio.
As a possible consequence of current anthropogenic global warming, many thermophilic marine organisms are expanding their range towards the higher latitudes of both hemispheres (e.g., Carlton, 2000). This might ultimately lead pelagic eagle rays to re-enter the Mediterranean Basin. Indeed, the occurrence of A. narinari off remote islands such as Bermuda have led to speculation that this fish is capable of extensive journeys (Bigelow & Schroeder, 1953), and the extant whitespotted eagle rays living along the western coast of Florida are known to move over large distances along shallow-marine, nearshore migratory corridors to meet their environmental (mostly thermal) preferences (DeGroot et al., 2021). Rather than via the Gibraltar Strait, which is quite far from the northernmost border of the modern range of Aetobatus along the western coast of Africa, the pelagic eagle rays might re-enter the Mediterranean Basin from the Red Sea (which is inhabited by various aetobatid species; Kyne et al., 2006, 2016; White, 2006), through the Suez Canal, in the frame of the so-called "lessepsian migration" (e.g., Por, 1978) that also accounts for the occurrence of the dasyatid myliobatoid Himantura uarnak in the eastern Mediterranean Sea (Başusta et al., 1998). By displaying values of seawater temperature that are likely even slightly higher than those associated with the northwestern Mediterranean during the early and mid-Pliocene (Langer et al., 2012; Coletti et al., 2021), the present-day eastern Mediterranean would indeed prove climatically suitable for the dispersal of Aetobatus. However, most living elasmobranchs are increasingly imperilled due to overfishing, habitat loss, and other typologies of human-induced pressure, and Aetobatus is no exception in this respect (Sales et al., 2019), its worldwide population being currently estimated as declining (Kyne et al., 2006, 2016; White, 2006). Considering also that several elasmobranch fisheries of the Red Sea are at present unsustainable (Spaet & Berumen, 2015), a recolonisation of the Mediterranean Sea by the pelagic eagle rays is here regarded as possible but not very likely.
Alice Pieri is kindly acknowledged for her help during field activities at Certaldo. We are also sincerely indebted to Frederick H. Mollen and Jürgen Pollerspöck, whose thorough reviews greatly contributed to improve an early draft of this paper. Last but not least, Bruno Granier and Stephen Carey provided most valuable editorial support: Thank you very much!
Agassiz L. (1833-1843).- Recherches sur les poissons fossiles, vol. 3.- Petitpierre, Neuchâtel, 392 p.
Agassiz L. (1858).- [A new species of skate from the Sandwich islands].- Proceedings of the Boston Society of Natural History, vol. 6, p. 385.
Antunes M.T. (1977).- Late Neogene fish faunas from Angola, their age and significance.- Journal of the Palaeontological Society of India, Lucknow, vol. 20, p. 224-229.
Antunes M.T. (1978).- Faunes ichthyologiques du Néogčne supérieur d'Angola, leur âge, remarques sur le Pliocčne marin en Afrique australe.- Cięncias da Terra (UNL), Lisbon, vol. 4, p. 59-90.
Antunes M.T. & Balbino A.C. (2006).- Latest Miocene myliobatids (Batoidei, Selachii) from the Alvalade Basin, Portugal.- Cainozoic Research, Leiden, vol. 4, p. 41-49.
Arambourg C. (1927).- Les poissons fossiles d'Oran.- Matériaux pour la Carte Géologique de l'Algérie (1re série), Paléontologie (sér. 1), Alger, vol. 6, 298 p.
Backman J., Raffi I., Rio D., Fornaciari E. & Pälike H. (2012).- Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes.- Newsletters on Stratigraphy, Stuttgart, vol. 45, p. 221-244.
Başusta N., Erdem U. & Kumlu M. (1998).- Two new fish records for the Turkish seas: Round stingray Taeniura grabata and skate stingray Himantura uarnak (Dasyatidae).- Israel Journal of Zoology, Tel Aviv, vol. 44, p. 65-66.
Benvenuti M., Del Conte S., Scarselli N. & Dominici S. (2014).- Hinterland basin development and infilling through tectonic and eustatic processes: Latest Messinian-Gelasian Valdelsa Basin, northern Apennines, Italy.- Basin Research, vol. 26, p. 387-402.
Bianucci G., Pesci F., Collareta A. & Tinelli C. (2019).- A new Monodontidae (Cetacea, Delphinoidea) from the lower Pliocene of Italy supports a warm-water origin for narwhals and white whales.- Journal of Vertebrate Paleontology, vol. 39, article #e1645148.
Bigelow H.B. & Schroeder W.C. (1953).- Sawfishes, guitarfishes, skates and rays. Part 2. Fisheries of the Western North Atlantic.- Memoirs of the Sears Memorial Foundation for Marine Research, vol. 1, 514 p.
Blainville H.M.D. de (1816).- Prodrome d'une nouvelle distribution systématique du rčgne animal.- Bulletin de la Société Philomathique de Paris, vol. 8, p. 105-112, 121-124.
Bonaparte C.L.J.L. (1838).- Selachorum tabula analytica.- Nuovi Annali delle Scienze Naturali, Bologna, vol. 1, p. 195-214.
Bor T., Reinecke T. & Verschueren S. (2012).- Miocene Chondrichthyes from Winterswijk-Miste, The Netherlands.- Palaeontos, vol. 21, 136 p.
Capapé C. & Quignard J.P. (1975).- Contribution ŕ la systématique et ŕ la biologie de Pteromylaeus bovinus (Geoffroy Saint-Hilaire, 1817), (Pisces, Myliobatidae) des côtes tunisiennes.- Bulletin du Muséum National d'Histoire Naturelle de Paris, Zoologie (sér. 3), no. 338, p. 1329-1347.
Capó A. & Garcia C. (2019).- Basin filling evolution of the central basins of Mallorca since the Pliocene.- Basin Research, vol. 31, p. 948-966.
Cappetta H. (1980).- Les sélaciens du Crétacé supérieur du Liban. II. Batoďdes.- Palaeontographica Abteilung A, Paläozoologie-Stratigraphie, Stuttgart, vol. 168, p. 149-229.
Cappetta H. (2012).- Chondrichthyes. Mesozoic and Cenozoic Elasmobranchii. Teeth. In: Schultze H.P. (ed.), Handbook of Paleoichthyology, vol. 3E.- Dr. Pfeil Verlag, Munich, 512 p.
Carlton J.T. (2000).- Global change and biological invasions in the oceans. In: Mooney H.A. & Hobbs R.J. (eds.), Invasive Species in a Changing World.- Island Press, Covelo, p. 31-53.
Carnevale G., Caputo D. & Landini W. (2006).- Late Miocene fish otoliths from the Colombacci Formation (Northern Apennines, Italy): Implications for the Messinian 'Lago-mare' event.- Geological Journal, London, vol. 41, p. 537-555.
Carnevale G., Dela Pierre F., Natalicchio M. & Landini W. (2018).- Fossil marine fishes and the 'Lago Mare' event: Has the Mediterranean ever transformed into a brackish lake?- Newsletters on Stratigraphy, Stuttgart, vol. 51, p. 57-72.
Carnevale G., Gennari R., Lozar F., Natalicchio M., Pellegrino L. & Dela Pierre F. (2019).- Living in a deep desiccated Mediterranean Sea: An overview of the Italian fossil record of the Messinian salinity crisis.- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 58, p. 109-140.
Carnevale G., Longinelli A., Caputo D., Barbieri M. & Landini W. (2008).- Did the Mediterranean marine reflooding precede the Mio-Pliocene boundary? Paleontological and geochemical evidence from upper Messinian sequences of Tuscany, Italy.- Palæogeography, Palæoclimatology, Palæoecology, vol. 257, p. 81-105.
Chemnitz J.H. (1782).- Neues Systematischen Conchylien Cabinet, vol. 6.- Gabriel Nicolaus Raspe, Nürnberg, 375 p.
Cita M.B. (1975).- Studi sul Pliocene e gli strati di passaggio dal Miocene al Pliocene. VII. Planktonic foraminiferal biozonation of the Mediterranean Pliocene deep-sea record. A revision.- Rivista Italiana di Paleontologia e Stratigrafia, Milan, vol. 81, p. 527-544.
Coletti G., Bosio G. & Collareta A. (2021).- Lower Pliocene barnacle facies of western Liguria (NW Italy): A peek into a warm past and a glimpse of our incoming future.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), Milan, vol. 127, p. 103-131.
Collareta A., Casati S., Catanzariti R. & Di Cencio A. (2017b).- First record of the knifetooth sawfish Anoxypristis (Elasmobranchii: Rhinopristiformes) from the Pliocene of Tuscany (central Italy).- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 284, p. 289-297.
Collareta A., Casati S. & Di Cencio A. (2017a).- A pristid sawfish from the lower Pliocene of Lucciolabella (Radicofani basin, Tuscany, central Italy).- Atti della Societŕ Toscana di Scienze Naturali, Memorie (ser. A), Pisa, vol. 124, p. 49-55.
Collareta A., Casati S. & Di Cencio A. (2018).- The porbeagle shark, Lamna nasus (Elasmobranchii: Lamniformes), from the late Pliocene of the central Mediterranean Basin.- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 287, p. 307-316.
Collareta A., Casati S., Zuffi M.A.L. & Di Cencio A. (2020a).- First authentic record of the freshwater turtle Mauremys from the upper Pliocene of Italy, with a new occurrence of the rarely reported ichnotaxon Thatchtelithichnus holmani.- Carnets Geol., Madrid, vol. 20, no. 16, p. 301-313.
Collareta A., Collareta M., Berta A. & Bianucci G. (2020b).- On Leonardo and a fossil whale: A reappraisal with implications for the early history of palaeontology.- Historical Biology, London, https://doi.org/10.1080/08912963.2020.1787403
Collareta A., Merella M., Casati S. & Di Cencio A. (2020c).- Did titanic stingrays wander the Pliocene Mediterranean Sea? Some notes on a giant-sized myliobatoid stinger from the Piacenzian of Italy.- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 298, p. 155-164.
Collareta A., Merella M., Mollen F.H., Casati S. & Di Cencio A. (2020d).- The extinct catshark Pachyscyllium distans (Probst, 1879) (Elasmobranchii: Carcharhiniformes) in the Pliocene of the Mediterranean Sea.- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 295, p. 129-139.
Collareta A., Mollen F.H., Merella M., Casati S. & Di Cencio A. (2021).- Remarkable multicuspid teeth in a new elusive skate (Chondrichthyes, Rajiformes) from the Mediterranean Pliocene.- Paläontologische Zeitschrift, Stuttgart, vol. 95, p. 117-128.
Compagno L.J.V. (1973).- Interrelationships of living elasmobranchs. In: Greenwood P.H., Miles R.S. & Patterson C. (eds.), Interrelationships of fishes.- Zoological Journal of the Linnean Society, London, vol. 53, suppl. 1, p. 15-61.
Compagno L.J.V. & Last P.R. (1999).- Order Myliobatiformes. In: Carpenter K.E. & Niem V.H. (eds.), FAO species identification guide for fisheries purposes. The living marine resources of the western central Pacific. Volume 3. Batoid fishes, chimaeras and bony fishes. Part 1 (Elopidae to Linophrynidae).- FAO, Rome, p. 1467-1529.
DeGroot B.C., Bassos-Hull K., Wilkinson K.A., Lowerre-Barbieri S., Poulakis G.R. & Ajemian M.J. (2021).- Variable migration patterns of whitespotted eagle rays Aetobatus narinari along Florida's coastlines.- Marine Biology, vol. 168, p. 1-21.
Di Cencio A., Dulai A., Catanzariti R., Casati S. & Collareta A. (2021).- First record of the brachiopod Lingula? from the Pliocene of Tuscany (Italy): The youngest occurrence of lingulides in the Mediterranean Basin.- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 299, p. 237-249.
Dominici S., Danise S. & Benvenuti M. (2018).- Pliocene stratigraphic paleobiology in Tuscany and the fossil record of marine megafauna.- Earth-Science Reviews, vol. 176, p. 277-310.
Fialho P., Balbino A. & Antunes M.T. (2019).- Langhian rays (Chondrichthyes, Batomorphii) from Brielas, Lower Tagus Basin, Portugal.- Geologica Acta, Barcelona, vol. 17, p. 1-16.
Froese R. & Pauly D., eds. (2020).- FishBase. World Wide Web electronic publication (version 12/2020).- Accessed on 21 January 2021. https://www.fishbase.org
Gmelin J.F. (1791).- Vermes. In: Gmelin J.F. (ed.), Caroli a Linnaei Systema Naturae per Regna Tria Naturae, Ed. 13, tome 1(6).- G.E. Beer, Leipzig, p. 3021-3910.
Herman J., Hovestadt-Euler M., Hovestadt D.C. & Stehmann M. (2000).- Part B: Batomorphii 4c: Order Rajiformes - Suborder Myliobatoidei - Superfamily Dasyatoidea - Family Dasyatidae - Subfamily Dasyatinae - Genus: Urobatis, Subfamily Potamotrygoninae - Genus: Paratrygon, Superfamily Plesiobatoidea - Family Plesiobatidae - Genus: Plesiobatis, Superfamily Myliobatoidea - Family Myliobatidae - Subfamily Myliobatinae - Genera: Aetobatus, Aetomylaeus, Myliobatis and Pteromylaeus, Subfamily Rhinopterinae - Genus: Rhinoptera and Subfamily Mobulinae - Genera: Manta and Mobula. Addendum 1 to 4a: erratum to Genus Pteroplatytrygon. In: Stehmann M. (ed.), Contributions to the study of the comparative morphology of teeth and other relevant ichthyodorulites in living supraspecific taxa of Chondrichthyan fishes.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Biologie, Brussels, vol. 70, p. 5-67.
Höltke O., Unger E., Pollerspöck J. & Rasser M.W. (2020).- The elasmobranch fauna from the Upper Marine Molasse (lower Miocene, Burdigalian) of Ursendorf (SW-Germany).- Palaeontos, Antwerp, vol. 33, p. 3-55.
Hovestadt D.C. & Hovestadt-Euler M. (2013).- Generic assessment and reallocation of Cenozoic Myliobatinae based on new information of tooth, tooth plate and caudal spine morphology of extant taxa.- Palaeontos, Antwerp, vol. 24, p. 1-66.
Jonet S. (1968).- Notes d'ichthyologie miocčne portugaise. V. - Quelques batoďdes.- Revista da Faculdade de Cięncias da Universidade de Lisboa, Lisbon, vol. 15, p. 233-258.
Kyne P.M., Dudgeon C.L., Ishihara H., Dudley S.F.J. & White W.T. (2016).- Aetobatus ocellatus.- The IUCN Red List of Threatened Species, article #e.T42566169A42566212.- Accessed on 21 January 2021. http://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T42566169A42566212.en
Kyne P.M., Ishihara H., Dudley S.F.J. & White W.T. (2006).- Aetobatus narinari.- The IUCN Red List of Threatened Species, article #e.T39415A10231645.- Accessed on 21 January 2021. https://dx.doi.org/10.2305/IUCN.UK.2006.RLTS.T39415A10231645.en
Langer M.R., Weinmann A.E., Lötters S. & Rödder D. (2012).- "Strangers" in paradise: Modeling the biogeographic range expansion of the foraminifera Amphistegina in the Mediterranean Sea.- Journal of Foraminiferal Research, Lawrence, vol. 42, p. 234-244.
Last P.R., White W.T., Carvalho M.R. de, Séret B., Stehmann M.F.W. & Naylor G.J.P. (2016).- Rays of the World.- CSIRO Publishing, Melbourne, 800 p.
Manganelli G. & Spadini V. (2019).- Megascyliorhinus miocaenicus (Chondrichthyes, Galeomorphii) from the Zanclean (early Pliocene) of San Quirico d'Orcia, central Italy.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 58, p. 165-170.
Marín J.M. (1992).- Paleoictiología de algunos yacimientos neógenos de la provincia de Alicante (II).- Cidaris, Rivista Ilicitana de Paleontología y Mineralogía, Elche, vol. 1, p. 4-24.
Marramŕ G., Carnevale G., Engelbrecht A., Claeson K.M., Zorzin R., Fornasiero M. & Kriwet J. (2018).- A synoptic review of the Eocene (Ypresian) cartilaginous fishes (Chondrichthyes: Holocephali, Elasmobranchii) of the Bolca Konservat-Lagerstätte, Italy.- Paläontologische Zeitschrift, Stuttgart, vol. 92, p. 283-313.
Marsili S. (2008).- Systematic, paleoecologic and paleobiogeographic analysis of the Plio-Pleistocene Mediterranean elasmobranch fauna.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (ser. A), Pisa, vol. 113, p. 81-88.
Mas G. (2010).- Ictiofauna del Pliocč del barranc de sa Talaia (Mallorca, Illes Balears, Mediterrŕnia Occidental). Implicacions paleoambientals.- Bolletí de la Societat d'Histňria Natural de les Balears, Palma de Mallorca, vol. 53, p. 43-70.
Menesini E. (1969).- Ittiodontoliti miocenici di Terra d'Otranto.- Palaeontographia Italica, vol. 65, p. 1-61.
Mollen F.H. (2010).- A partial rostrum of the porbeagle shark Lamna nasus (Lamniformes, Lamnidae) from the Miocene of the North Sea Basin and the taxonomic importance of rostral morphology in extinct sharks.- Geologica Belgica, Brussels, vol. 13, p. 61-76.
Monegatti P. & Raffi S. (2001).- Taxonomic diversity and stratigraphic distribution of Mediterranean Pliocene bivalves.- Palæogeography, Palæoclimatology, Palæoecology, vol. 165, p. 171-193.
Mora Morote P. (1996).- Peces Galeomorfos y Squatinomorfos en el Plioceno de Guardamar del Segura (Alicante).- Cidaris - Rivista Ilicitana de Paleontología y Mineralogía, Elche, vol. 5, p. 98-124.
Mora Morote P. (1997).- Peces Myliobatiformes y Pristiophoriformes en el Plioceno de Guardamar del Segura (Alicante).- Cidaris - Rivista Ilicitana de Paleontología y Mineralogía, Elche, no. 11-12, p. 48-63.
Pawellek T., Adnet S., Cappetta H., Métais E., Salem M., Brunet M. & Jaeger J.-J. (2012).- Discovery of an earliest Pliocene relic tropical fish fauna in a newly detected cliff section (Sabratah Basin, NW Libya).- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 266, p. 93-114.
Pomar L., Marzo M. & Barón A. (1983).- El Terciario de Mallorca. In: Pomar L., Obrador O., Fornós J.J. & Rodríguez-Perea A. (eds.), El Terciario de las Baleares (Mallorca-Menorca). Guía de las Excursiones del X Congreso Nacional de Sedimentología.- Institut d'Estudis Baleŕrics and Universitat de Palma de Mallorca, Palma de Mallorca, p. 21-44.
Por F.D. (1978).- Lessepsian migrations. The influx of Red Sea biota into the Mediterranean by way of the Suez Canal.- Springer, Heidelberg, 228 p.
Prista G.A., Agostinho R.J. & Cachăo M.A. (2015).- Observing the past to better understand the future: A synthesis of the Neogene climate in Europe and its perspectives on present climate change.- Open Geosciences, Berlin, vol. 7, p. 65-83.
Purdy R.W., Schneider V.P., Applegate S.P., McLellan J.H., Meyer R.L. & Slaughter B.H. (2001).- The Neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. In: Ray C.E. & Bohaska D.J. (eds.), Geology and paleontology of the Lee Creek Mine, North Carolina, vol. III.- Smithsonian Contributions to Paleobiology, Washington D.C., vol. 90, p. 71-202.
Ragaini L., Ficini F., Zanchetta G., Regattieri E., Perchiazzi N. & Dallai L. (2019).- Mineralogy and oxygen isotope profile of Pelecyora gigas (Veneridae, Bivalvia) from Tuscan Pliocene.- Alpine and Mediterranean Quaternary, Rome, vol. 32, p. 5-13.
Reinecke T., Louwye S., Havekost U. & Moths H. (2011).- The elasmobranch fauna of the late Burdigalian, Miocene, at Werder-Uesen, Lower Saxony, Germany, and its relationships with early Miocene faunas in the North Atlantic, Central Paratethys and Mediterranean.- Palaeontos, Antwerp, vol. 20, 170 p.
Sales J.B.L., Oliveira C.N. de, Santos W.C.R. dos, Rotundo M.M., Ferreira Y., Ready J., Sampaio I., Oliveira C., Cruz V.P., Lara-Mendoza R.E. & Silva Rodrigues-Filho L.F. da (2019).- Phylogeography of eagle rays of the genus Aetobatus: Aetobatus narinari is restricted to the continental western Atlantic Ocean.- Hydrobiologia, The Hague, vol. 836, p. 169-183.
Serena F. (2005).- Field identification guide to the sharks and rays of the Mediterranean and Black Sea. In: FAO species identification guide for fishery purposes.- FAO, Rome, 97 p.
Sorbi S., Domning D.P., Vaiani S.C. & Bianucci G. (2012).- Metaxytherium subapenninum (Bruno, 1839) (Mammalia, Dugongidae), the latest sirenian of the Mediterranean Basin.- Journal of Vertebrate Paleontology, Lawrence, vol. 32, p. 686-707.
Spadini V. & Manganelli G. (2015).- A megachasmid shark tooth (Chondrichthyes, Lamniformes) from the Zanclean (early Pliocene) of San Quirico d'Orcia, central Italy.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 54, p. 67-70.
Spaet J.L.Y. & Berumen M.L. (2015).- Fish market surveys indicate unsustainable elasmobranch fisheries in the Saudi Arabian Red Sea.- Fisheries Research, vol. 161, p. 356-364.
Vai G.B. (1989).- A field trip guide to the Romagna Apennine geology. The Lamone Valley.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 28, p. 343-367.
Villafańa J.A., Marramŕ G., Klug S., Pollerspöck J., Balsberger M., Rivadeneira M. & Kriwet J. (2020).- Sharks, rays and skates (Chondrichthyes, Elasmobranchii) from the Upper Marine Molasse (middle Burdigalian, early Miocene) of the Simssee area (Bavaria, Germany), with comments on palaeogeographic and ecological patterns.- Paläontologische Zeitschrift, Stuttgart, vol. 94, p. 725-757.
Violanti D. (2012).- Pliocene Mediterranean foraminiferal biostratigraphy: A synthesis and application to the paleoenvironmental evolution of northwestern Italy. In: Elitok Ö. (ed.), Stratigraphic analysis of layered deposits.- Intech Open, London, p. 123-160.
Weigmann S. (2016).- Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity.- Journal of Fish Biology, Oxford, vol. 88, p. 837-1037.
White W.T. (2006).- Aetobatus flagellum.- The IUCN Red List of Threatened Species, article #e.T60119A12306888.- Accessed on 21 January 2021. https://dx.doi.org/10.2305/IUCN.UK.2006.RLTS.T60119A12306888.en
White W.T. (2014).- A revised generic arrangement for the eagle ray family Myliobatidae, with definitions for the valid genera.- Zootaxa, Auckland, vol. 3860, p. 149-166.
White W.T., Furumitsu K. & Yamaguchi A. (2013).- A new species of eagle ray Aetobatus narutobiei from the northwest Pacific: An example of the critical role taxonomy plays in fisheries and ecological sciences.- PLOS ONE, vol. 8, article #e83785.
White W.T. & Moore A.B.M. (2013).- Redescription of Aetobatus flagellum (Bloch & Schneider, 1801), an endangered eagle ray (Myliobatoidea: Myliobatidae) from the Indo-West Pacific.- Zootaxa, Auckland, vol. 3752, p. 199-213.
White W.T. & Naylor G.J.P. (2016).- Resurrection of the family Aetobatidae (Myliobatiformes) for the pelagic eagle rays, genus Aetobatus.- Zootaxa, Auckland, vol. 4139, p. 435-438.