◄ Carnets Geol. 21 (12) ►
![]()
Outline:
[1. Introduction]
[2. Methodology and conventions]
[3. Paleontological study]
[4. Conclusions] [Bibliographic references] and ...
[Plates]
16, rue Frédéric Mistral, 26200 Montélimar (France);
Centre d'Études Méditerranéennes, 04170 St-André-les-Alpes (France)
Département des Sciences de la Terre, Université de Nice-Sophia-Antipolis, Faculté des Sciences, 28 avenue Valrose, 06108 Nice Cedex 2 (France);
Centre d'Études Méditerranéennes, 04170 St-André-les-Alpes (France)
Fachbereich Geowissenschaften, Universität Bremen, Klagenfurter Strasse 4, 28359 Bremen (Germany)
Groupement d'Intérêt Paléontologique, Science et Exposition, 60 bd Georges Richard, 83000 Toulon (France)
Chemin de Dardène, La Barthelasse, 84000 Avignon (France);
Centre d'Études Méditerranéennes, 04170 St-André-les-Alpes (France)
Grand rue, 04330 Barrême (France)
Published online in final form (pdf) on June 24, 2021
DOI 10.2110/carnets.2021.2112
![]()
[Editor: Bruno R.C. Granier]
![]()
The occurrence of the nautiloid genus Anglonautilus Spath in France was limited to the report of Anglonautilus dorsoplicatus (Wiedmann) from the Albian of Escragnolles (Alpes-Maritimes) and Anglonautilus sp. from the Aptian of Les Ferres (Alpes-Maritimes). Here we document the presence of the successive species Anglonautilus praeundulatus Lehmann et al., Anglonautilus undulatus (Sowerby) and Anglonautilus dorsoplicatus (Wiedmann) from the Cretaceous of France. The stratigraphic range of Anglonautilus Spath, hitherto unknown before Aptian times is extended downward to the Hauterivian. The Hauterivian species Nautilus begudensis Kilian & Reboul, sometimes assigned to the genus Anglonautilus Spath, is revised and re-assigned to the genus Cymatoceras Hyatt.
Nautiloidea;
Nautilidae;
Anglonautilus;
Cymatoceras;
Lower Cretaceous
Baudouin C., Delanoy G., Lehmann J., Frau C., Gonnet R. & Vermeulen J. (2021).- The Cretaceous nautiloid genus Anglonautilus Spath, 1927, in France.- Carnets Geol., Madrid, vol. 21, no. 12, p. 235-263.
Les nautiloïdes crétacés du genre Anglonautilus Spath, 1927, en France.- La répartition du genre Anglonautilus Spath en France était jusqu'à présent limitée à la présence d'Anglonautilus dorsoplicatus (Wiedmann) dans l'Albien d'Escragnolles (Alpes-Maritimes) et d'Anglonautilus sp. dans l'Aptien de Les Ferres (Alpes-Maritimes). Nous rapportons ici la présence des espèces successives Anglonautilus praeundulatus Lehmann et al., Anglonautilus undulatus (Sowerby) et Anglonautilus dorsoplicatus (Wiedmann) dans le Crétacé de France. Le genre Anglonautilus Spath, connu jusqu'alors uniquement à partir de l'Aptien, est présent dès l'Hauterivien. L'espèce hauterivienne Nautilus begudensis Kilian & Reboul, parfois attribuée au genre Anglonautilus Spath, est révisée et transférée au genre Cymatoceras Hyatt.
Nautiloidea ;
Nautilidae ;
Anglonautilus ;
Cymatoceras ;
Crétacé inférieur
The extinct genus Anglonautilus Spath, 1927, remains a rather neglected nautiloid taxon from the Lower Cretaceous of both, the Tethys and the Boreal realms. Its first occurrence is probably in the upper Valanginian, and it certainly ranges through the Hauterivian into the Albian, with the species Anglonautilus praeundulatus Lehmann et al., 2017, A. karpinskyi (Karakasch, 1907), A. undulatus (Sowerby, 1813), A. farringdonensis (Sharpe, 1854), and A. dorsoplicatus (Wiedmann, 1960). Its presence in the Upper Cretaceous is based on the species A. dorsoplicatus (Wiedmann), with an occurrence in the Middle Cenomanian of France and possibly in the Lower Cenomanian of Spain, England and Iran, and ? A. catarinae Sundberg, 1984, from the Maastrichtian of Baja California (Mexico). There are also doubtful records in the basal Paleocene [A. somovitensis (Tzankov, 1941)] and the Eocene (Ţibuleac & Ajdanlijsky, 2017, p. 59) of Bulgaria, but their generic affiliation is in need of revision.
Lehmann et al. (2017) recently attempted to reconstruct the evolutionary history of the genus, and also introduced the new species Anglonautilus praeundulatus Lehmann et al. It is worth noting that the Upper Cretaceous species Anglonautilus japonicus Matsumoto & Takahashi, 1982, Anglonautilus mamiyai Matsumoto & Miyauchi, 1983, and Anglonautilus suciensis (Whiteaves, 1879) have been excluded from that genus by Lehmann et al. (2017) since they share only superficial similarities with the type species A. undulatus (Sowerby).
The collection of new specimens from the Cretaceous of France improves our understanding of the genus Anglonautilus Spath, especially regarding its earliest members.
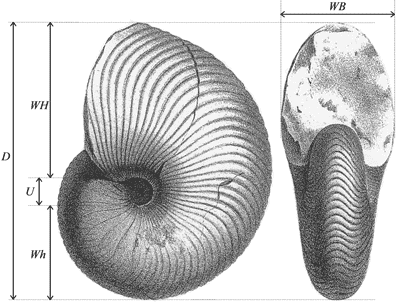
The measurements are given in millimeters;
measurements of height (WH), width of
the whorls (WB) and diameter of the
umbilicus (U) were made at the largest
diameter (D); Wh
is taken at a position diametrically opposed to that of WH (Fig. 1 ![]() ) and α is
the angle of that part of whorl where the undulations typical of Anglonautilus
Spath, 1927, are present.
) and α is
the angle of that part of whorl where the undulations typical of Anglonautilus
Spath, 1927, are present.
|
Figure 1:
Explanatory scheme for the measurements made on the studied
specimens (illustration from Orbigny, 1840-1842, modified). |
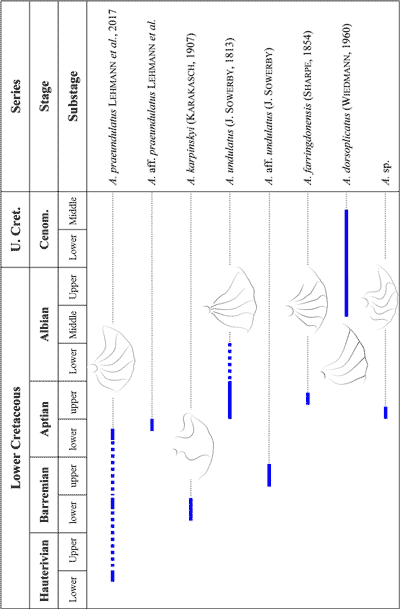
The biostratigraphic scheme of the Lower Cretaceous followed herein (Fig. 2 ![]() ) is that developed by the IUGS Lower Cretaceous Ammonite Working Group (Reboulet
et al., 2018) for the northern and western Tethys.
) is that developed by the IUGS Lower Cretaceous Ammonite Working Group (Reboulet
et al., 2018) for the northern and western Tethys.
|
Figure 2:
Biozonation of the Lower Cretaceous used in this work, adopted
from Reboulet et al. (2018). |
Table 1: List of abbreviations used in the text.
| CF | Camille Frau coll., Muséum d'Histoire Naturelle d'Aix-en-Provence (France) |
| GACO | Esparrac coll., Musée de Paléontologie de Provence, Aix-Marseille University (France) |
| GD | Gérard Delanoy coll. |
| GSC, GSM | British Geological Survey, Nottingham (UK) |
| GSUB | Geowissenschaftliche Sammlung, Bremen University (Germany) |
| PMAL | Pierre Martel coll., Réserve Naturelle Géologique du Luberon, Apt (France) |
| PU | Museo Regionale di Scienze Naturali, Turin (Italy) |
| RG | Roland Gonnet coll. |
| UJF-ID | Observatoire des Sciences de l'Univers de Grenoble, Saint-Martin d'Hères, Isère (France) |
| ULP | Cyril Baudouin coll. |
The suprageneric classification follows that of Lehmann et al. (2017), who have confirmed that the genus Anglonautilus Spath is closely related to Cymatoceras Hyatt, 1884, and is probably derived from it, which leads its classification within the Nautilidae Blainville, 1825 (Shimansky, 1975; Tintant & Kabamba, 1983; Tintant, 1993).
Order Nautilida Agassiz, 1847
Superfamily Nautiloidea Blainville, 1825
Family Nautilidae Blainville, 1825
Genus Anglonautilus Spath, 1927 (=Palelialia Shimansky, 1955)
Type species: Nautilus undulatus Sowerby, 1813, by original designation of Spath (1927, p. 21).
Diagnosis: The genus Anglonautilus Spath, 1927, is characterised by an involute coiling and a subtrapezoidal to suboval whorl section, compressed to moderately depressed. The ornamentation consists of broad undulations present at the top of the flanks and especially on the ventral region, on a part of the shell ranging from α = 60° to 200°, generally at the end of the phragmocone and/or at the beginning of the body chamber. Their strength is highly variable, ranging from discrete to very marked. These undulations are sometimes followed by cymatoceratid ribs towards the end of the body chamber.
The suture line is generally slightly sinuous, with a more or less marked lateral lobe and sometimes a weak ventral lobe. If exposed, the siphuncle is seen to be located close to the middle of the section, slightly shifted dorsally.
Geographical and
stratigraphical distribution: The genus Anglonautilus Spath,
1927, appears in the Early Hauterivian (Fig. 3 ![]() ) and probably even in the Late
Valanginian in Germany (Lehmann, unpublished data). However,
it is rare in the Hauterivian and Barremian. At the current state of knowledge,
only two specimens are known in the Hauterivian: the specimen figured by Karakasch
(1907, Pl. II, fig. 9) as Nautilus
pseudoelegans Orbigny, 1840, from the "Hauterivian sandstone of
Biasala" (Crimea), which can be attributed to the Lower Hauterivian, and a
recently discovered specimen of Anglonautilus praeundulatus Lehmann
et al., 2017, from the Lower
Hauterivian (Lyticoceras nodosoplicatum
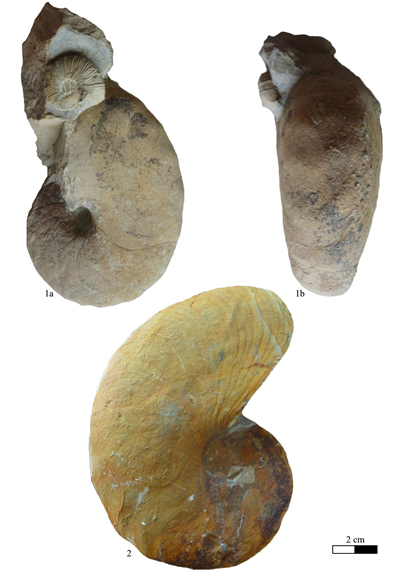
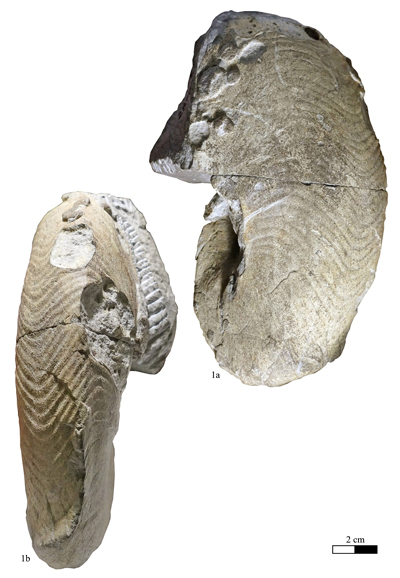
Zone) of Curnier (Drôme, France) (RG2007, figured here on Pl. I
) and probably even in the Late
Valanginian in Germany (Lehmann, unpublished data). However,
it is rare in the Hauterivian and Barremian. At the current state of knowledge,
only two specimens are known in the Hauterivian: the specimen figured by Karakasch
(1907, Pl. II, fig. 9) as Nautilus
pseudoelegans Orbigny, 1840, from the "Hauterivian sandstone of
Biasala" (Crimea), which can be attributed to the Lower Hauterivian, and a
recently discovered specimen of Anglonautilus praeundulatus Lehmann
et al., 2017, from the Lower
Hauterivian (Lyticoceras nodosoplicatum
Zone) of Curnier (Drôme, France) (RG2007, figured here on Pl. I ![]() , fig. 1.a-b;
Pl. II
, fig. 1.a-b;
Pl. II ![]() , fig.
1.a-b).
, fig.
1.a-b).
|
Figure 3:
Stratigraphic position and suture line of the different species
of the genus Anglonautilus
Spath, 1927. |
The
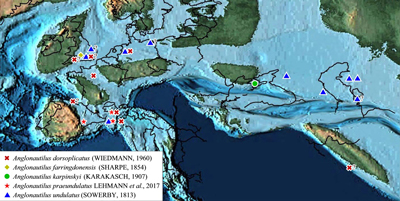
genus occurs in Europe (England, France, Germany, Spain, Bulgaria?) and Asia
(Crimea, Russia (northern Caucasus), Kazakhstan (Mangyshlak), Iran, Turkmenistan?), as well as possibly in Baja California, Mexico (Karakasch,
1907;
Tzankov, 1941; Shimansky,
1975; Kennedy et
al., 1979; Sundberg, 1984; Mutterlose,
1996; Weber, 1996; Wilmsen,
2000; Kennedy, 2002; Lehmann et al., 2017) (Fig. 4 ![]() ).
).
|
Figure 4: Paleogeographical distribution of the different species of the genus Anglonautilus Spath, 1927. Paleogeographical map of the Early Albian from Scotese, 2013, used here as an approximation of the Hauterivian-Cenomanian paleogeography. Data from Karakasch, 1907; Shimansky, 1975; Kennedy et al., 1979; Mutterlose, 1996; Weber, 1996; Wilmsen, 2000; Kennedy, 2002, and Lehmann et al., 2017. |
Discussion: As currently defined, the genus Anglonautilus Spath, 1927, includes the following species:
A. dorsoplicatus (Wiedmann, 1960)
A. farringdonensis (Sharpe, 1854)
A. karpinskyi (Karakasch, 1907)
A. praeundulatus Lehmann et al., 2017
A. subalbensis (Sinzow, 1913)
A. undulatus (Sowerby, 1813)
Doubtful are:
The lower Barremian taxon Nautilus karpinskyi Karakasch, 1907, type-species of the genus Palelialia Shimansky, 1955, has been revised by Shimansky (1975), who, in the same publication, designated specimen no. 103/107 (p. 130, figured by Karakasch, 1907, Pl. VIII, fig. 10) as well as specimen no. 103/106 as lectotype (p. 200, caption in Pl. XXIX, fig. 1; specimen figured by Karakasch, 1907, Pl. II, fig. 2, and refigured by Shimansky, 1975). Therefore, it is unclear which specimen Shimansky intended as lectotype and this designation is therefore invalid (ICZN art. 74.5). Subsequently, Tintant (1980, p. 64) designated specimen no. 103/106 as lectotype.
An
understanding of the taxon Nautilus
karpinskyi Karakasch and thus the genus Palelialia
Shimansky is therefore based on the lectotype no. 103/106 (Karakasch,
1907, Pl. II, fig. 2). This specimen develops an ornamentation composed of
distinct undulations, followed by cymatoceratid ribbing, which
is typical of the genus Anglonautilus Spath. Furthermore, the suture
line of this species, rather sinuous with a distinctive lateral lobe, has also
been observed in other representatives of the genus Anglonautilus Spath
(this work,
Pl. IV ![]() , fig. 2;
Pl. XII
, fig. 2;
Pl. XII ![]() , fig. 1.a-b) and does not in itself justify
a generic distinction. We therefore consider the genus Palelialia
Shimansky as a junior synonym of Anglonautilus Spath.
Pending re-examination, the species P.
alpanensis (Rouchadzé, 1938) and P.
imerica (Rouchadzé, 1938) are kept separate from Anglonautilus
Spath as they lack of ornamentation and their suture lines differ from
that of the genus. Their generic affinities remain to be investigated.
, fig. 1.a-b) and does not in itself justify
a generic distinction. We therefore consider the genus Palelialia
Shimansky as a junior synonym of Anglonautilus Spath.
Pending re-examination, the species P.
alpanensis (Rouchadzé, 1938) and P.
imerica (Rouchadzé, 1938) are kept separate from Anglonautilus
Spath as they lack of ornamentation and their suture lines differ from
that of the genus. Their generic affinities remain to be investigated.
Interpretation
of the species A. farringdonensis (Sharpe) was thus far based on the original hand-drawing and description of Sharpe
(1854). Three specimens assigned to this species from the Faringdon Sponge
Gravels (upper Aptian, Parahoplites
nutfieldiensis Zone) of Faringdon (Berkshire, England) are kept in the
collections of the British Geological Survey (Nottingham, England). They are
here figured on Plates XII ![]() ,
XIII
,
XIII ![]() and XIV
and XIV ![]() . Specimen no. GSC7552 (Pl. XII
. Specimen no. GSC7552 (Pl. XII ![]() ,
fig. 2.a-b; Pl. XIII
,
fig. 2.a-b; Pl. XIII ![]() , fig. 2) corresponds
to the specimen depicted by Sharpe
(British Geological Survey comm.). It is herein designated as lectotype.
The species is characterised by weak ornamentation, with only slight ventral
undulations, and by a slightly sinuous suture line.
, fig. 2) corresponds
to the specimen depicted by Sharpe
(British Geological Survey comm.). It is herein designated as lectotype.
The species is characterised by weak ornamentation, with only slight ventral
undulations, and by a slightly sinuous suture line.
The species A. catarinae Sundberg, from the Lower Maastrichtian of Baja California (Mexico), bears low undulations on the venter in later growth stages, present on only a few specimens (Sundberg, 1984, p. 46). Its suture line is slightly sinuous, with a broad and weak lateral lobe. Its indistinct undulations question its generic attribution.
The species A. somovitensis (Tzankov) is based on two specimens from the Lower Paleocene (Danian/Montian) of Somovit (Bulgaria). The poor quality illustration does not permit recognition of its main features. The species likely develops a few irregular undulations at the beginning of the body chamber. A re-examination of the material is needed to confirm its generic assignment.
The Upper Cretaceous taxa A. hewaidyi Moneer et al., 2021, A. japonicus Matsumoto & Takahashi, 1982, A. mamiyai Matsumoto & Miyauchi, 1983, and A. suciensis (Whiteaves, 1879) develop a peculiar ornamentation, composed of undulations arising from cymatoceratid ribbing (Chirat & Bucher, 2006, p. 62) and not independently as in the Lower Cretaceous forms. These taxa should be excluded from Anglonautilus Spath (Lehmann et al., 2017, p. 74).
The taxon Nautilus begudensis Kilian & Reboul, 1915, from the Lower Hauterivian of the Alpes-de-Haute-Provence (France), has been assigned to the genus Anglonautilus Spath by Kummel (1956) and Shimansky (1975); it is revised in this work and re-assigned into the genus Cymatoceras Hyatt, 1884, in agreement with Thieuloy (1972) and Lehmann et al. (2017).
When the shell is not preserved, the study of nautiloids with rib ornamentation can also be complicated by the different types of mould preservation, as noted by Malchyk et al. (2017). The thickness and structure of nautiloid shells, with different internal and external ornament, induces different types of mould preservation (composite moulds; Crampton, 2004; Wilmsen, 2016). Therefore, the preserved ornamentation might represent a mixture of external and internal shell features. However, a study of a sufficient number of representatives of the genus Anglonautilus Spath allows us to confirm that, in this genus, one never observes ornamentation of the Cymatoceras type in association with the typical undulations of Anglonautilus Spath in that stage and that the two genera are indeed distinct, although certainly phyletically closely related.
Anglonautilus praeundulatus Lehmann et al., 2017
(Pl. I ![]() , fig. 1.a-b;
Pl. II
, fig. 1.a-b;
Pl. II ![]() , fig. 1.a-b;
Pl. III
, fig. 1.a-b;
Pl. III ![]() , fig. 1.a-c;
, fig. 1.a-c;
Pl. IV ![]() ,
figs. 1.a-b, 2;
Pl. V
,
figs. 1.a-b, 2;
Pl. V ![]() ,
figs. 1, 2.a-b;
Pl. VI
,
figs. 1, 2.a-b;
Pl. VI ![]() , fig. 1)
, fig. 1)
2017. Anglonautilus praeundulatus sp. nov. - Lehmann et al., p. 69, Fig. 2.
2019. Anglonautilus sp. - Bersac & Bert, Pl. 3, figs. 7-8.
Material (n = 6): Specimen no. RG2007, Lower Hauterivian, Lyticoceras nodosoplicatum Zone, Curnier (Drôme, France); no. RG2004, lower Barremian, Kotetishvilia compressissima Zone ("Combe-Petite" Horizon), Lure Mountain (Alpes-de-Haute-Provence, France); no. RG2005, upper Barremian, Lure Mountain (Alpes-de-Haute-Provence, France); no. CF-B-EM.5, lower Aptian, lower part of the Deshayesites deshayesi Zone, bed 160, Conte Quarry, Cassis (Bouches-du-Rhône, France); specimens no. GD28800, GD28801, lower Aptian, Deshayesites deshayesi Zone, Deshayesites grandis Subzone, bed 107, Les Graoux, Les Ferres (Alpes-Maritimes, France).
Type: Holotype is specimen no. GSUB C7505, deposited at the Geowissenschaftliche Sammlung, University of Bremen, Germany. It originates from the Forcall Formation, Deshayesites deshayesi Zone, lower Aptian, Castellote (Teruel, Spain).
Geographical and stratigraphical distribution: Based on our new data, Anglonautilus praeundulatus Lehmann et al., 2017, ranges from the Lower Hauterivian (Lyticoceras nodosoplicatum Zone) to the lower Aptian (Deshayesites deshayesi Zone) and is known from eastern Spain and southeastern France.
Dimensions (in mm)
Table 2: Measurements of Anglonautilus praeundulatus Lehmann et al., 2017.
| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| RG2004 | 100.1 | 10.3 | 58.2 | 33.7 | 55.1 | 150 | 0.10 | 0.58 | 1.73 | 0.55 | 0.95 |
| RG2005 | 119.8 | 9.1 | 63.3 | 47.2 | - | 110 | 0.08 | 0.53 | 1.34 | - | - | 96.2 | 7 | 57.1 | 30.4 | 46.6 | 0.07 | 0.59 | 1.88 | 0.48 | 0.82 |
| RG2007 | 145.6 | 4.2 | 84.6 | 51.6 | 65.8 | 150 | 0.03 | 0.58 | 1.64 | 0.45 | 0.78 |
| CF-B-EM.5 | 129.4 | 17.9 | 68.4 | 41.9 | - | c120 | 0.14 | 0.53 | 1.63 | - | - |
| GSUB C7505 | 112.5 | 15 | 55 | 35 | 48 | 90 | 0.13 | 0.49 | 1.57 | 0.43 | 0.87 |
| GD28800 | 149 | 16.5 | 87 | 46.5 | - | - | 0.11 | 0.58 | 1.87 | - | - |
| GD28801 | c114 | - | - | - | - | - | - | - | - | - | - |
Description: The six specimens at our disposal show an involute coiling, with a U/D ratio between 0.03 and 0.14, the earlier specimens appearing to be more involute. Most specimens are affected by sediment compaction; the best-preserved specimens RG2004, RG2005 and RG2007 show a subtrapezoidal and moderately compressed whorl section. The flanks, which are slightly rounded in the inner whorls, become convex during growth to give the section a suboval shape beyond a diameter of about 100 mm. The venter is moderately broad and rounded. The peristome, preserved in specimens RG2004, RG2007 and CF-B-EM.5, is slightly sinuous, with a rounded indentation on the ventral region.
The suture line is slightly sinuous, with a shallow ventral lobe, slightly deeper on specimen CF-B-EM.5. The position of the siphuncle is unknown.
The ornamentation is composed of undulations on the ventral region, appearing at about D = 70-80 mm, on the last half or quarter whorl of the phragmocone. These undulations occur throughout an interval of between α = 90° and α = 150° and, on the largest specimens, disappear at about D = 90-100 mm to give way to an ornamentation of cymatoceratid ribs. As observed on specimens RG2005 and RG2007, the change from undulations to ribs is quite progressive, since undulations gradually lose their width and increase in density. The ribs are weak and mainly visible on the ventral region, but also on the flanks of specimens RG2007 and CF-B-EM.5 at the end of the body chamber. On specimen CF-B-EM.5, this change is more abrupt and the cymatoceratid ribbing appears as soon as the undulations disappear at the end of the phragmocone; they are thin on the main part of the body chamber and progressively strengthen when approaching the peristome.
Discussion and comparisons: Anglonautilus praeundulatus Lehmann et al., 2017, is the earliest species of the genus Anglonautilus Spath, 1927.
The species is closely allied to Anglonautilus undulatus (Sowerby, 1813). It can be distinguished by a much narrower ventral region and a more compressed section, with a WB/WH ratio between 0.87 and 0.95 in A. praeundulatus Lehmann et al. and about 1.23 in the neotype of A. undulatus (Sowerby). In addition, in A. praeundulatus Lehmann et al. the undulations are less pronounced and occupy a smaller portion of the shell [between α = 90° and α = 150° in A. praeundulatus Lehmann et al. versus α = 180° in A. undulatus (Sowerby)].
The earlier species, A. praeundulatus Lehmann et al. is from the lower Aptian, Deshayesites deshayesi Zone and probably ranges into the Dufrenoyia furcata Zone (Anglonautilus aff. praeundulatus Lehmann et al., infra), whereas it was succeeded by A. undulatus (Sowerby) in the upper Aptian, Epicheloniceras martini Zone.
Anglonautilus praeundulatus Lehmann et al. can be easily distinguished from the Barremian species Anglonautilus karpinskyi (Karakasch, 1907) by its compressed whorl section [WB/WH about 1.19 in the lectotype of A. karpinskyi (Karakasch)] and by its much less sinuous suture line, with a shallower lateral lobe.
Anglonautilus praeundulatus Lehmann et al. differs from Anglonautilus farringdonensis (Sharpe, 1854) by a more compressed section and a stronger ornamentation, the undulations being very attenuated in A. farringdonensis (Sharpe).
Finally, Anglonautilus praeundulatus Lehmann et al. can be distinguished from Anglonautilus dorsoplicatus (Wiedmann, 1960) by a longer stage with undulations [α = 60° to 70° in A. dorsoplicatus (Wiedmann)]. It also occupies a different stratigraphic position since A. dorsoplicatus (Wiedmann) is only known from the Middle Albian to the Middle Cenomanian.
Anglonautilus aff. praeundulatus Lehmann et al., 2017
(Pl. VII ![]() , fig. 1.a-b;
Pl. VIII
, fig. 1.a-b;
Pl. VIII ![]() , fig. 1.a-b)
, fig. 1.a-b)
2018. Anglonautilus sp. - Delanoy et al., Fig. 17.B-C.
Material (n = 1): Specimen no. GD28773, lower Aptian, Dufrenoyia furcata Zone, Dufrenoyia dufrenoyi Subzone, bed 110, Les Graoux, Les Ferres (Alpes-Maritimes, France).
Geographical and stratigraphical distribution: Anglonautilus aff. praeundulatus Lehmann et al., 2017, is only known from bed 110 of Les Graoux, Les Ferres (Alpes-Maritimes, France), assigned to the lower Aptian, Dufrenoyia furcata Zone, Dufrenoyia dufrenoyi Subzone.
Dimensions (in mm)
Table 3: Measurements of Anglonautilus aff. praeundulatus Lehmann et al., 2017.
| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| GD28773 | 170 | - | - | - | - | 110 | - | - | - | - | - |
Description: Specimen GD28773 is strongly distorted, and its whorl section is difficult to characterise; it is seemingly moderately compressed, suboval on the body chamber. The flanks are relatively flat on the phragmocone and become rounded on the body chamber. The ventral region is narrow and rounded. The suture line is rather sinuous and shows a distinctive lateral lobe. The position of the siphuncle is not known.
The ornamentation appears on the phragmocone at D = 50-60 mm. It is first composed of well-marked, distant undulations, visible only on the ventral region where they form a chevron. At the transition between the phragmocone and the body chamber, at D = 85 mm, the undulations disappear and give way to a dense and strong cymatoceratid ribbing, present on both flanks and venter of the body chamber. These ribs follow a convex path on the flanks and cross the ventral region strongly curved backwards, forming a chevron of the same shape as the undulations on the phragmocone.
Discussion and comparisons: Anglonautilus aff. praeundulatus Lehmann et al., 2017, is close to A. praeundulatus Lehmann et al.; it mainly differs by the presence of strong cymatoceratid ribs on the body chamber. This ribbing can be observed, although much less developed, on specimens CF-B-EM.5 and GD28800 of A. praeundulatus Lehmann et al. A. aff. praeundulatus Lehmann et al. can therefore be interpreted as a form showing strongly developed cymatoceratid ribbing of the type seen in A. praeundulatus Lehmann et al. However, given the uniqueness of this specimen, its rather poor preservation and its slightly later stratigraphic position, it seems preferable to leave its true affinities open to questions.
Anglonautilus undulatus (Sowerby, 1813)
(Pl. IX ![]() ,
figs. 1.a-b, 2;
Pl. X
,
figs. 1.a-b, 2;
Pl. X ![]() ,
figs. 1-2, 3.a-b;
Pl. XI
,
figs. 1-2, 3.a-b;
Pl. XI ![]() , fig. 1.a-b)
, fig. 1.a-b)
1813. Nautilus undulatus sp. nov. - Sowerby, p. 87, Pl. XL, upper figure.
non 1853. Nautilus undulatus Sowerby - Sharpe, p. 15, Pl. V, fig. 4.a-c [? = Anglonautilus dorsoplicatus (Wiedmann, 1960)].
1913. Nautilus subalbensis sp. nov. - Sinzow, p. 113, Pl. VI, fig. 7, 7a.
non 1951. Anglonautilus undulatus (Sowerby) - C.W. Wright & E.V. Wright, p. 12, Pl. V, fig. 4 [? = Anglonautilus dorsoplicatus (Wiedmann, 1960)].
1956. Anglonautilus undulatus (Sowerby) - Kummel, p. 430, Fig. 26, Pl. 20, figs. 1-2.
1960. Anglonautilus undulatus (Sowerby) - Shimansky, p. 244, Pl. VII, fig. 1.а-в.
1960. Angulithes (Pseudocenoceras) undulatus (Sowerby) - Wiedmann, p. 176.
1964. Anglonautilus undulatus (Sowerby) - Kummel, p. K453, Fig. 334, 3.a-b.
1975. Anglonautilus subalbensis (Sinzow) - Shimansky, p. 120, Pl. XXVII, fig. 2.а- б.
1975. Anglonautilus undulatus (Sowerby) - Shimansky, p. 119, Pl. XXVII, fig. 1.а-б.
non 1979. Anglonautilus undulatus (Sowerby) - Kennedy et al., p. 7, Pl. 1, fig. 1 [? = Anglonautilus dorsoplicatus (Wiedmann, 1960)].
1983. Anglonautilus undulatus (Sowerby) - Kabamba, Pl. IV, fig. 3.a-b.
? 1983. Anglonautilus ou Cymatoceras - Kabamba, p. 59, Pl. IV, fig. 4.
1996. Anglonautilus undulatus (Sowerby) - Mutterlose, Pl. 6, fig. 2.
1996. Anglonautilus undulatus (Sowerby) - Weber, Pl. 9, fig. 3.
non 2002. Anglonautilus undulatus (Sowerby) - Kennedy, p. 231, Fig. 10.1I [? = Anglonautilus dorsoplicatus (Wiedmann, 1960)].
? 2016. Anglonautilus ? sp. - Lehmann et al., Pl. 5, fig. 2; Pl. 10, fig. 3.
Material (n = 5): Specimen no. CF-B-PM.4.13, upper Aptian, lower part of the Epicheloniceras martini Zone, bed 191?, La Marcouline gate, Cassis (Bouches-du-Rhône, France); no. CF-B-LM.12, upper Aptian, upper part of the Epicheloniceras martini Zone, bed M52, La Marcouline quarry, Cassis (Bouches-du-Rhône, France); no. CF-FcFb.65, CF-FcFb.106, CF-CUG.1, upper Aptian, upper part of the Epicheloniceras martini Zone (equivalent to bed M52), Fontblanche Forest, Roquefort-la-Bédoule (Bouches-du-Rhône, France).
Type: According to Kummel (1956, p. 431), the type specimen of Sowerby is lost. The original illustration is idealised and makes it difficult to appreciate its diagnostic features. Indeed, the ventral undulations described by Sowerby are not illustrated in his drawing. Interpretation of this species was subsequently based on the topotype B.M.N.H. 66336, illustrated by Kummel (1956, Pl. 20). In order to define Sowerby's species much more comprehensively than the original illustration allows, we herein retain specimen no. B.M.N.H. 66336 as neotype of A. undulatus (Sowerby, 1813). It originates from the type locality (Nutfield, Surrey, England) and is assigned to the upper Aptian, Parahoplites nutfieldiensis Zone; it is stored in the collections of the Natural History Museum in London.
Geographical and stratigraphical distribution: The species is recorded from the upper Aptian of England, southeastern France, Germany, Russia (northern Caucasus) and Kazakhstan (Mangyshlak). Anglonautilus subalbensis (Sinzow, 1913) [= Anglonautilus undulatus (Sowerby, 1813)] is also recorded in Turkmenistan (Tuarkyr mountains) according to Shimansky (1975), but the corresponding specimen lacks illustration. This species would extend up into the Lower Albian according to Shimansky (1975); the Albian specimen from Hunstanton (England) figured by Kabamba (1983, Pl. IV, fig. 4) as "Anglonautilus or Cymatoceras" could also be a representative of Anglonautilus undulatus (Sowerby). Specimens attributed to Anglonautilus undulatus (Sowerby) by Sharpe (1853), C.W. Wright & E.V. Wright (1951) and Kennedy et al. (1979) from the Lower Cenomanian of England and Iran are here tentatively re-assigned to Anglonautilus dorsoplicatus (Wiedmann, 1960).
Dimensions (in mm)
Table 4: Measurements of Anglonautilus undulatus (Sowerby, 1813).
| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| CF-B-LM.12 | c134.7 | 15.8 | 69 | c48.2 | - | 180 | 0.12 | 0.51 | 1.43 | - | - |
| CF-B-PM.4.13 | 94.5 | - | - | - | - | 200 | - | - | - | - | - |
| CF-FcFb.65 | - | - | 46.3 | - | - | - | - | - | - | - | - |
| CF-FcFb.106 | 145.8 | 18.5 | 75.1 | 56.4 | 66.9 | c170 | 0.13 | 0.52 | 1.33 | 0.46 | 0.89 |
| CF-CUG.1 | c146.2 | c13.8 | 95 | - | - | - | 0.09 | 0.65 | - | - | - |
Description: The five specimens at our disposal are distorted by sedimentary compaction, and only specimen no. CF-FcFb.106 shows a whorl section suitable for measurement. The coiling is moderately involute, with a relatively open umbilicus (U/D ratio greater than 0.10). The section is suboval, inflated, with increasingly rounded flanks during growth and a relatively wide, rounded ventral region. The peristome, preserved on specimens no. CF-FcFb.106 and CF-CUG.1, is sinuous, with a ventral indentation, following the same shape as the ribs. The suture line, as those seen in specimen no. CF-B-LM.12, is not very sinuous, with a wide and shallow lateral lobe. The position of the siphuncle remains unknown.
The ornamentation is first composed of relatively strong undulations, widely spaced, visible over about α = 180°, present on the ventral region and the top of the flanks of the last part of the phragmocone; they are also present on the first part of the body chamber of specimen no. CF-CUG.1. Specimen no. CF-FcFB.106 develops a few thin and very attenuated riblets, overlapping the undulations. These undulations are followed, on the body chamber of specimens no. CF-FcFb.106, CF-B-LM.12 and CF-CUG.1, by an ornamentation of relatively weak cymatoceratid ribs, attenuated on the lower half of the flanks; they are more clearly marked on the umbilical wall and the upper part of the flanks, especially near the peristome. They are convex on the flanks and then curve backwards on the upper third of the flanks. In specimen CF-FcFb.106, they cross the venter where they are attenuated, forming a rounded chevron. On specimen no. CF-B-LM.12 there are also fine growth striae superimposed on the ribs and of the same pattern, visible on the upper half of the flanks of the last half of the body chamber.
Discussion and comparisons: The species Anglonautilus subalbensis (Sinzow, 1913), from the upper Aptian of Kazakhstan (Mangyshlak), is similar to Anglonautilus undulatus (Sowerby, 1813), both regarding general morphology and ornamentation. Shimansky (1975) indicates that A. subalbensis (Sinzow) has a more trapezoidal whorl section and that the undulations of this species are only present on the ventral region. However, the section, which is obviously very close to that of A. undulatus (Sowerby), is only partially visible on the original figure of A. subalbensis (Sinzow), and the undulations seem to be present at the top of the flanks of the type-specimen. These differences do not seem to justify a specific distinction and A. subalbensis (Sinzow) is thus regarded as a junior synonym of A. undulatus (Sowerby).
Anglonautilus undulatus (Sowerby) closely resembles Anglonautilus praeundulatus Lehmann et al., 2017. Differences between the two species have been discussed above.
Anglonautilus undulatus (Sowerby) differs from Anglonautilus farringdonensis (Sharpe, 1854) by its distinct undulations, present over about half a whorl, whereas these are discrete in A. farringdonensis (Sharpe).
Anglonautilus undulatus (Sowerby) is also closely related to Anglonautilus dorsoplicatus (Wiedmann, 1960). A. undulatus (Sowerby) differs from it in having a much wider section, a slightly less sinuous suture line and, above all, much more developed undulations, present for a range of about α = 180°, whereas they are present for only about α = 60°-70° in A. dorsoplicatus (Wiedmann).
Anglonautilus aff. undulatus (Sowerby, 1813)
(Pl. I ![]() , fig. 2.a-b;
Pl. II
, fig. 2.a-b;
Pl. II ![]() , fig. 2.a-b)
, fig. 2.a-b)
Material (n = 1): Specimen no. RG2006, upper Barremian, Lure Mountain (Alpes-de-Haute-Provence, France).
Geographical and stratigraphical distribution: Anglonautilus aff. undulatus (Sowerby, 1813) is only known in the upper Barremian of the Lure Mountain (Alpes-de-Haute-Provence, France).
Dimensions (in mm)
Table 5: Measurements of Anglonautilus aff. undulatus (Sowerby, 1813).
| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| RG2006 | 83.7 | 9.1 | 50.4 | 26.2 | 50.5 | 170 | 0.11 | 0.60 | 1.92 | 0.60 | 1.00 |
Description: Specimen no. RG2006 is small-sized, well preserved, complete with its body chamber and peristome. The coiling is involute and the section is subtrapezoidal, with slightly rounded flanks converging to a broadly rounded ventral region. The peristome is sinuous, with a slight ventral indentation. Neither the suture line nor the position of the siphuncle are discernable.
Ornamentation, absent on the phragmocone, appears at the beginning of the body chamber, formed by widely spaced undulations on the ventral region well-marked and of irregular strength. The undulations are slightly rursiradiate and cross the ventral region forming a weak adapical sinus. In the last half of the body chamber, feebly convex, slightly marked and irregularly shaped cymatoceratid ribs appear on the flanks, becoming somewhat more pronounced towards the peristome.
Discussion and comparisons: Specimen no. RG2006 shows great affinities to Anglonautilus undulatus (Sowerby, 1813) by its wide whorl section and distinctive undulations, present over about α = 170°. However, it differs from it by the irregularity of these undulations and their presence on the body chamber only, whereas in A. undulatus (Sowerby) they are mainly present on the last 180° of the phragmocone, sometimes at the beginning of the body chamber. It also occupies a much earlier stratigraphic position, in the upper Barremian, whereas A. undulatus (Sowerby) is only known from the upper Aptian. The peculiarities and uniqueness of specimen no. RG2006 do not allow us to integrate it into the Sowerby's species; it may either represent an atypical form because of its lower stratigraphic position, which needs to be supported by more material, or it may represent an undescribed species.
Specimen no. RG2006 shares some similarities with the lower Barremian species Anglonautilus karpinskyi (Karakasch, 1907) by the shape of its whorl section. However, the lectotype of A. karpinskyi (Karakasch) can be easily distinguished by the undulations present on its phragmocone and the early appearance of a distinctive cymatoceratid ribbing. The lack of the body chamber in the lectotype of A. karpinskyi (Karakasch) and the impossibility to observe the suture line on specimen no. RG2006 prevent further comparison.
Specimen no. RG2006 can be distinguished from Anglonautilus praeundulatus Lehmann et al., 2017, by its distinctly wider whorl section and by the well-marked undulations, present only on the body chamber, being more persistent (α = 170° instead of α = 90° to 150° in A. praeundulatus Lehmann et al.).
Anglonautilus dorsoplicatus (Wiedmann, 1960)
(Pl. VI ![]() , fig. 2.a-c;
Pl. XI
, fig. 2.a-c;
Pl. XI ![]() , fig. 2.a-c;
Pl. XIV
, fig. 2.a-c;
Pl. XIV ![]() , fig. 2.a-c)
, fig. 2.a-c)
pars 1853. Nautilus largilliertianus d'Orbigny - Sharpe, p. 16, Pl. VI, fig. 2 ?; non Pl. VI, fig. 1 [= Pseudocenoceras largilliertianum (Orbigny)].
? 1853. Nautilus undulatus Sowerby - Sharpe, p. 15, Pl. V, fig. 4.a-c.
1897. Nautilus largilliertianus ? d'Orbigny - Parona & Bonarelli, p. 76, Pl. 10, fig. 6.
? 1951. Anglonautilus undulatus (Sowerby) - C.W. Wright & E.V. Wright, p. 12, Pl. V, fig. 4.
1960. Angulithes (Pseudocenoceras) dorsoplicatus nov. sp. - Wiedmann, p. 176.
? 1979. Anglonautilus undulatus (Sowerby) - Kennedy et al., p. 7, Pl. 1, fig. 1.
pars 1999. Pseudocenoceras cf. fittoni (Sharpe) - Wittler et al., p. 25, Fig. 32.a-b, non Fig. 30a, b [= Pseudocenoceras fittoni (Sharpe)], 31.a-b (= ? Eutrephoceras sp.).
? 2000. Pseudocenoceras dorsoplicatus (Wiedmann) - Wilmsen, p. 41, Pl. 5, figs. 13, 26.
? 2002. Pseudocenoceras dorsoplicatus (Wiedmann) - Kennedy, p. 223, Fig. 10.1F.
? 2002. Anglonautilus undulatus (Sowerby) - Kennedy, p. 231, Fig. 10.1I.
2017. Anglonautilus dorsoplicatus (Wiedmann) - Lehmann et al., Fig. 3.
Material (n = 3): Cast of the holotype no. GSUB C7523, Middle Albian, Escragnolles (Alpes-Maritimes, France); specimen no. PMAL 340, Upper Albian, Saint-Etienne-les-Orgues (Alpes-de-Haute-Provence, France); no. ULP01, Middle Cenomanian, Acanthoceras rhotomagense Zone, Turrilites costatus Subzone, Saint-Ulphace (Sarthe, France).
Type:
The holotype is the
specimen no. PU20409 figured by Parona
& Bonarelli (1897, Pl. 10, fig. 6) and refigured here Pl. XI ![]() , fig.
2.a-c; it is housed at the Museo Regionale di Scienze Naturali in
Turin (Italy) and comes from the Middle Albian of Escragnolles (Alpes-Maritimes,
France); it originates from beds having a condensed fauna including ammonites
indicative of the Lyelliceras lyelli, Hoplites
spathi and Anahoplites intermedius
subzones (Lehmann et al., 2017,
p. 68). We have access to a cast, stored as no. GSUB C7523 in the
Geowissenschaftliche Sammlung of the University of Bremen (Germany) that was
already figured by Lehmann et al. (2017, Fig. 3).
, fig.
2.a-c; it is housed at the Museo Regionale di Scienze Naturali in
Turin (Italy) and comes from the Middle Albian of Escragnolles (Alpes-Maritimes,
France); it originates from beds having a condensed fauna including ammonites
indicative of the Lyelliceras lyelli, Hoplites
spathi and Anahoplites intermedius
subzones (Lehmann et al., 2017,
p. 68). We have access to a cast, stored as no. GSUB C7523 in the
Geowissenschaftliche Sammlung of the University of Bremen (Germany) that was
already figured by Lehmann et al. (2017, Fig. 3).
Geographical and stratigraphical distribution: The species ranges from the Middle Albian to the Middle Cenomanian of France and Germany and is possibly present in the Lower Cenomanian of northern Cantabria, Spain. The Lower Cenomanian specimens from England and Iran figured by Sharpe (1853, Pl. VI, fig. 2 only) as Pseudocenoceras largilliertianum (Orbigny, 1840) and by Sharpe (1853), C.W. Wright & E.V. Wright (1951) and Kennedy et al. (1979) as Anglonautilus undulatus (Sowerby, 1813) probably belong to Anglonautilus dorsoplicatus (Wiedmann, 1960).
Dimensions (in mm)
Table 6: Measurements of Anglonautilus dorsoplicatus (Wiedmann, 1960).| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| GSUB C7523 | 60.5 | 6.8 | 37.3 | 22.6 | 32.9 | 60 | 0.11 | 0.62 | 1.65 | 0.54 | 0.88 |
| PMAL 340 | 69.6 | - | 39.2 | 25.9 | - | 70 | - | 0.56 | 1.51 | - | - |
| ULP01 | 63 | 6.8 | 34.6 | 22 | 26.8 | - | 0.11 | 0.55 | 1.57 | 0.43 | 0.77 |
Description: The three specimens show a fairly involute coiling and a compressed subtrapezoidal section, with slightly rounded flanks and a relatively narrow, rounded ventral region. On the holotype and specimen no. PMAL 340, only the very beginning of the body chamber is preserved; specimen no. ULP01 is entirely septate. The suture line is weakly sinuous on the holotype and specimen no. ULP01, with a broad, shallow lateral lobe and the absence of a ventral lobe. The lateral lobe is less marked on specimen no. PMAL 340. According to Parona & Bonarelli (1897, p. 76), the siphuncle is located on the inner quarter of the septal surface in the holotype; it is not visible on cast no. GSUB C7523. However, it is also visible on the inner quarter of the septal surface of specimen no. ULP01 (Pl. XIV, fig. 2c).
Ornamentation, comprising a few undulations, is present only during a short stage (about α = 60°-70°) at the end of the phragmocone, appearing at the top of the flanks and especially visible on the ventral region. They are distinctly rursiradiate and form a more or less angular sinus in the ventral region.
Discussion and comparisons: The Lower Cenomanian specimens figured by Sharpe (1853), C.W. Wright & E.V. Wright (1951) and Kennedy et al. (1979) are doubtfully re-assigned here to Anglonautilus dorsoplicatus (Wiedmann, 1960) since they show some differences to the holotype. The position of the siphuncle in the very fragmentary specimen of Sharpe (1853, Pl. VI, Fig. 2), referred to as Nautilus largilliertianus Orbigny, 1840, shown at the base of the septal surface, differs markedly from that of the holotype, where it is situated in the lower quarter of that surface. However, the position of the siphuncle as shown in Sharpe's drawing is questioned by Parona & Bonarelli (1897, p. 76). On the specimen figured by Sharpe (1853) and C.W. Wright & E.V. Wright (1951) as Anglonautilus undulatus (Sowerby, 1813), the undulations show a similar development to those of A. dorsoplicatus (Wiedmann) but are located on the last part of the body chamber, whereas they are present on the phragmocone of the holotype of A. dorsoplicatus (Wiedmann). Finally, on the specimen figured by Kennedy et al. (1979) as A. undulatus (Sowerby), the section appears distinctly different, wider and subcircular.
Anglonautilus dorsoplicatus (Wiedmann) differs from Anglonautilus undulatus (Sowerby) by its much more compressed section and by its undulations being present for a much shorter stage, about α = 60-70° instead of α = 180°.
Anglonautilus dorsoplicatus (Wiedmann) is also close to Anglonautilus praeundulatus Lehmann et al., 2017, but is essentially different by its less developed undulations, present over about α = 60°-70° instead of α = 90°-150° in A. praeundulatus Lehmann et al. It also occupies a different stratigraphic position.
Anglonautilus dorsoplicatus (Wiedmann) differs from Anglonautilus farringdonensis (Sharpe, 1854) by a much more compressed section and by its more pronounced undulations, which are barely perceptible in A. farringdonensis (Sharpe).
Anglonautilus dorsoplicatus (Wiedmann) is also close to the Cenomanian species Pseudocenoceras largilliertianum (Orbigny), from which it differs only by the presence of weak undulations on the ventral region. Affinities between the genus Anglonautilus Spath, 1927, and P. largilliertianum (Orbigny) have already been noted by Tintant (1993, p. 365), and A. dorsoplicatus (Wiedmann) has frequently been attributed to the genus Pseudocenoceras Spath, 1927 (Wiedmann, 1960; Wilmsen, 2000; Kennedy, 2002). The phyletic relationships between A. dorsoplicatus (Wiedmann) and P. largilliertianum (Orbigny) need to be clarified.
Anglonautilus sp.
(Pl. XII ![]() , fig. 1.a-b)
, fig. 1.a-b)
Material (n = 1): Specimen no. GACO.033, upper Aptian, base of the Epicheloniceras martini Zone, La Marcouline quarry, Cassis (Bouches-du-Rhône, France).
Geographical and stratigraphical distribution: Anglonautilus sp. is based on specimen no. GACO.033, from the upper Aptian (base of the Epicheloniceras martini Zone) of Roquefort-la-Bédoule (Bouches-du-Rhône, France).
Dimensions (in mm)
Table 7: Measurements of Anglonautilus sp.
| no. | D | U | WH | Wh | WB | α | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| GACO.033 | 137.2 | 19 | 73.6 | 55.6 | 41.6 | c110 | 0.14 | 0.54 | 1.32 | 0.30 | 0.57 |
Description: Specimen no. GACO.033 is complete with its body chamber. It shows an involute coiling and a compressed section, with slightly rounded flanks and a narrow, rounded ventral region. The very sinuous suture line shows a very wide and deep lateral lobe. The position of the siphuncle is unknown.
The ornamentation consists of well-marked undulations, present over at least α = 110°, on the entire visible part of the phragmocone. The undulations appear at the top of the flanks and are mainly visible on the ventral region where they are strong, widely spaced and clearly rursiradiate, forming a rounded adapical sinus. On the body chamber, the undulations disappear and the ornamentation is composed only of faint growth striae visible on the flanks. The peristome is preserved on one flank; it is sinuous, with a lappet in the upper third of the flank and then strongly retroverse as it approaches the ventral region.
Discussion and comparisons: Specimen no. GACO.033 shows some affinities to Anglonautilus undulatus (Sowerby, 1813), notably by the strong development of its undulations, present over at least α = 110°. However, it differs from that species by its much narrower section and especially by its very sinuous suture line with a well-developed lateral lobe, very different from the suture line of most other species of the genus Anglonautilus Spath, 1927.
The suture line of specimen GACO.033 is however similar to that of the lectotype of Anglonautilus karpinskyi (Karakasch, 1907), which also shows a very wide and deep lateral lobe. However, the shell morphology of A. karpinskyi (Karakasch) differs by its much wider section, undulations present for a much shorter stage, and the appearance of strongly developed cymatoceratid ribs on the phragmocone.
The unusual features of specimen no. GACO.033 do not allow attribution to any Anglonautilus species. It could represent a new species, but this could only be confirmed by additional material.
Genus Cymatoceras Hyatt, 1884
Type species: Nautilus pseudoelegans Orbigny, 1840, by original designation of Hyatt (1884, p. 301).
Cymatoceras begudensis (Kilian & Reboul, 1915)
(Pl. XV ![]() , fig. 1.a-c;
Pl. XVI
, fig. 1.a-c;
Pl. XVI ![]() ,
figs. 1.a-c, 2.a-b)
,
figs. 1.a-c, 2.a-b)
1915. Nautilus begudensis sp. nov. - Kilian & Reboul, p. 227, Pl. X, fig. 1.
1956. Anglonautilus begudensis (Kilian & Reboul) - Kummel, p. 431.
1972. Anglonautilus ? begudensis (Kilian & Reboul) - Thieuloy, p. 22.
1975. Anglonautilus begudensis (Kilian & Reboul) - Shimansky, p. 119.
2017. "Anglonautilus" begudensis (Kilian & Reboul) - Lehmann et al., p. 72.
Material (n = 3): Specimens no. UJF-ID 434, UJF-ID 435, UJF-ID 25328, Lower Hauterivian, Lyticoceras nodosoplicatum Zone, La Bégude, La-Palud-sur-Verdon (Alpes-de-Haute-Provence, France).
Type: The lectotype, designated by Thieuloy (1972), is specimen no. UJF-ID 434 (lectotype designation before 2000, CINZ art. 74.5), figured on Pl. XVI, fig. 1.a-c.
Geographical and stratigraphical distribution: Cymatoceras begudensis (Kilian & Reboul, 1915) only occurs in the Lower Hauterivian, Lyticoceras nodosoplicatum Zone of La-Palud-sur-Verdon (Alpes-de-Haute-Provence, France). The species is also reported (without illustration) from the Lower Hauterivian, Crioceratites loryi Zone, Olcostephanus jeannoti Subzone of Rottier (Drôme) and from Rougon (Alpes-de-Haute-Provence) by Thieuloy (1972), but these unnumbered specimens have not been found in the collections of the Observatoire des Sciences de l'Univers de Grenoble.
Dimensions (in mm)
Table 8: Measurements of Cymatoceras begudensis (Kilian & Reboul, 1915).
| no. | D | U | WH | Wh | WB | U/D | WH/D | WH/Wh | WB/D | WB/WH |
| UJF-ID 434 | 95.3 | 10 | 56.7 | - | 43.8 | 0.10 | 0.59 | - | 0.46 | 0.77 |
| UJF-ID 435 | 87.8 | - | 49.2 | - | 42.9 | - | 0.56 | - | 0.49 | 0.87 |
| UJF-ID 25328 | 124.9 | 16.9 | 64.1 | - | 59 | 0.14 | 0.51 | - | 0.47 | 0.92 |
Description:
The three specimens studied here show a compressed (WB/WH
close to 0.80-090) and relatively evolute shell (U/D between 0.10 and 0.14). The section is oval, with slightly
rounded flanks. The suture line, visible on specimen no. UJF-ID 435 (Pl. XVI ![]() ,
fig. 2.a-b), is only slightly
sinuous. The position of the siphuncle is unknown.
,
fig. 2.a-b), is only slightly
sinuous. The position of the siphuncle is unknown.
The ornamentation is only visible from about D = 55 mm. It is first composed of sinuous and slightly concave ribs. They are weak on the flanks, then become stronger in the upper third and bend abruptly backwards to cross the ventral region forming a clear concave sinus.
From about D = 70-80 mm, there is a short stage where some ribs (about one in two) strengthen very strongly as they pass through the ventral region while conversely, the remaining ribs weaken.
Beyond about D = 100 mm, the ornamentation becomes regular again. The ribs are well marked on the flanks where they are straight to slightly concave, then bend backwards to cross the ventral region in equal strength.
Discussion and comparisons: The ornamentation of Cymatoceras begudensis (Kilian & Reboul, 1915), with a short stage showing strengthened ribs on the ventral region, led Kummel (1956) to assign the species to Anglonautilus Spath, 1927. However, as noted by Thieuloy (1972) and Lehmann et al. (2017), these ribs are superimposed on a typical cymatoceratid ribbing as defined by Chirat & Bucher (2006) and do not conform to the stage with independent undulations as observed in Anglonautilus Spath. These strengthened ribs are also very different from the undulations observed in typical representatives of the genus Anglonautilus Spath, with a much higher, narrow and angular shape. As such, the species of Kilian & Reboul compares better with typical Cymatoceras Hyatt, 1884.
Thieuloy (1972) suggested that Cymatoceras begudensis (Kilian & Reboul) may correspond to a peculiar morphotype of Cymatoceras bifurcatus (Ooster, 1858). This species remains little studied. Note that the type specimens of Ooster come from the Albian and the "Neocomian" of Switzerland and are probably not conspecific. As a consequence, Thieuloy's hypothesis remains unclear and a revision of the taxonomy and age of C. bifurcatus (Ooster) is needed. However, the particular ornamental characters and the restricted stratigraphic position of C. begudensis (Kilian & Reboul) suggest that this may be a distinct species.
Cymatoceras begudensis (Kilian & Reboul) is also closely allied to Cymatoceras neocomiense (Orbigny, 1840), a species present in the Hauterivian and Barremian. It is distinguished by its much more compressed section and its short stage with strengthened ribs in the ventral region, by contrast to the regular cymatoceratid ribbing of C. neocomiense (Orbigny).
The study of new material of the genus Anglonautilus Spath, 1927, from southeastern France has allowed us to identify the stratigraphic succession of the species A. praeundulatus Lehmann et al., 2017, A. undulatus (Sowerby, 1813) and A. dorsoplicatus (Wiedmann, 1960) in the Lower Cretaceous and base of the Upper Cretaceous and also to define their specific characteristics. In addition, we have been able to show that the genus Anglonautilus Spath, present in the Early Hauterivian, was most certainly derived from the genus Cymatoceras Hyatt, 1884. The illustration of the lectotype and two other topotypes of Nautilus farringdonensis Sharpe, 1854, allowed us to confirm that this species also belongs to the genus Anglonautilus Spath. However, a study of typical specimens of Nautilus begudensis Kilian & Reboul, 1915, led us to exclude this species from the genus Anglonautilus Spath and to attribute it to the genus Cymatoceras Hyatt instead.
The genus Anglonautilus Spath is still present in the Upper Cretaceous but remains poorly known, especially regarding the post-Cenomanian taxa. In particular, the phyletic links of Early Cretaceous species with the latest Cretaceous (Maastrichtian) species ? A. catarinae Sundberg, 1984, and Paleocene (Danian/Montian) ? A. somovitensis (Tzankov, 1941) remain to be clarified. The generic assignment of these two species to the genus Anglonautilus Spath still has to be confirmed.
For their help and photographs, we are indebted to Alain Bonnot (Université de Bourgogne, Dijon, Côte-d'Or, France), Annalaura Pistarino (Museo Regionale di Scienze Naturali, Torino, Italy), Frederick A. Sundberg (Museum of Northern Arizona, Flagstaff, Arizona, USA), Simon J. Harris and Paul J. Shepherd (British Geological Survey, Nottingham, England). We warmly thank Fabienne Giraud (Observatoire des Sciences de l'Univers de Grenoble, Saint-Martin d'Hères, Isère, France) and Stéphane Legal (Réserve Naturelle Géologique du Luberon, Apt, Vaucluse, France) for their welcome during our visit in the collections in their care. We also warmly thank our reviewers Markus Wilmsen (Senckenberg Naturhistorische Sammlungen, Dresden, Germany) and Oksana Malchyk (Polish Academy of Sciences, Warszawa, Poland) for their constructive remarks. We are very thankful for H. Mark Bayliss (London, England) for a linguistic correction of the manuscript.
Bersac S. & Bert D. (2019).- The lower Aptian ammonites of the Les Ferres Aptian Basin (Lower Cretaceous, Southeast of France). Part I: Introduction and biostratigraphy.- Carnets Geol., Madrid, vol. 19, no. 9, p. 149-183.
Chirat R. & Bucher H. (2006).- Shell microstructure and morphogenesis of the ornamentation in Cymatoceras Hyatt, 1883, Cretaceous Nautilida. Systematic implications.- Lethaia, Oslo, vol. 39, p. 57-64.
Crampton J. (2004).- Shell composition, cryptic costae, complex composite molds, and taphonomic chicanery in Mytiloides (Inoceramidae, Bivalvia).- Journal of Paleontology, Cambridge, vol. 78, no. 6, p. 1091-1096.
Delanoy G., Baudouin C., Pictet A., Delattre P. & Pénagé E. (2018).- Ancyloceratidae (Ammonoidea, Ancyloceratoidea) nouveaux ou peu connus de l'Aptien inférieur des Alpes-Maritimes (Sud-Est de la France).- Annales du Muséum d'Histoire Naturelle de Nice, vol. XXXII, p. 35-90.
Hyatt A. (1883-1884).- Genera of fossil cephalopods.- Proceedings of the Boston Society of Natural History, vol. 22 (1882-1883), p. 253-338.
Kabamba M. (1983).- Les Nautiles du crétacé supérieur de Madagascar : Contribution à l'étude de l'évolution des nautiloïdes du Crétacé; systématique, phylogénie, biostratigraphie, paléoécologie, évolution.- Thèse 3ème cycle, Géologie, Dijon University, 136 p.
Karakasch N.I. (1907).- Le Crétacé inférieur de la Crimée et sa faune.- Travaux de la Société impériale des Naturalistes de St.-Pétersbourg, vol. XXXII, livr. 5, 482 p.
Kennedy W.J. (2002).- Nautiloids. In: Smith A.B. & Batten D.J. (Eds.), Fossils of the Chalk. Second edition, revised and enlarged.- The Palaeontological Association, London, p. 219-231.
Kennedy W.J., Chahida M.R. & Djafarian M.A. (1979).- Cenomanian cephalopods from the Glauconitic limestone southeast of Esfahan, Iran.- Acta Palaeontologica Polonica, Warsaw, vol. 24, no. 1, p. 3-50.
Kilian W. & Reboul P. (1915).- Contribution à l'étude des faunes paléocrétacées du Sud-Est de la France. II. Sur quelques ammonites de l'Hauterivien de La Bégude (La Bégüe) [Basses-Alpes]. Matériaux pour l'étude de la faune de l'Hauterivien des environs de Moustiers-Sainte-Marie, La Palud et Châteauneuf-les-Moustiers (Basses-Alpes).- Mémoires pour servir à l'explication de la carte géologique détaillée de la France, no. 14, Paris, p. 223-288.
Kummel B. (1956).- Post-Triassic nautiloid genera.- Bulletin of the Museum of comparative Zoology, Cambridge, vol. 114, no. 7, 494 p.
Kummel B. (1964).- Nautiloidea-Nautilida. In: Moore R.C. (Ed.), Treatise on Invertebrate Paleontology, part K, Mollusca 3.- University of Kansas Press, Lawrence - KS; Geological Society of America, Boulder - CO, p. K383-466.
Lehmann J., Maisch M.W., Baudouin C. & Salfinger-Maisch A. (2017).- Origin and evolutionary history of Anglonautilus (Nautilida, Cymatoceratidae) and a new species from the lower Aptian of Spain.- Cretaceous Research, vol. 72, p. 66-80.
Lehmann J., Hoffmann R., Owen H. & Beckert W. (2016).- Cephalopoden aus unterkreidezeitlichen Geschieben der Region um Wolgast-Hohendorf, Vorpommern.- Archiv für Geschiebekunde, Hamburg/Greifswald, vol. 7, no. 7-8, p. 401-530.
Malchyk O., Machalski M., Waksmundzki B. & Duda M. (2017).- Shell ornament, systematic position and hatching size of Epicymatoceras vaelsense (Nautilida): New insights based on specimens in mould preservation from the Upper Cretaceous of Poland.- Cretaceous Research, vol. 80, p. 1-12.
Moullade M., Granier B. & Tronchetti G. (2011).- The Aptian stage: Back to fundamentals.- Episodes, Bangalore, vol. 34, no. 3, p. 148-156.
Mutterlose J. (1996).- Die Litho- und Biostratigraphie des Apt der Tongruben Schnermann am Rothenberge.- Geologie und Paläontologie in Westfalen, Münster, no. 45, p. 41-73.
Orbigny A. d' (1840-1842).- Paléontologie française, description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France. Terrains Crétacés, Vol. I. Céphalopodes.- Masson, Paris, 662 p.
Parona C.F. & Bonarelli G. (1897).- Fossili Albiani d'Escragnolles del Nizzardo e della Liguria occidentale.- Palaeontographia Italica, Memorie di paleontologia, Pisa, vol. II (1896), p. 53-112.
Reboulet S., Szives O., Aguirre-Urreta B., Barragán R., Company M., Frau C., Kakabadze M.V, Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S.N., Vaíček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Scotese C.R. (2013).- Map Folio 25, Early Cretaceous (early Albian, 110 Ma), PALEOMAP PaleoAtlas for ArcGIS, volume 2, Cretaceous paleogeographic, paleoclimatic and plate tectonic reconstructions.- PALEOMAP Project, Evanston - IL, 27 p.
Sharpe D. (1853).- Description of the fossil remains of Mollusca found in the Chalk of England, part I.- Palaeontographical Society, London, p. 1-26.
Sharpe D. (1854).- On the age of the fossiliferous sands and gravels of Farringdon and its neighbourhood.- The Quarterly Journal of the Geological Society of London, vol. 10, no. 1-2, p. 176-198.
Shimansky V.N. (1960).- Nautilids. In: Druschits V.V. & Kudryavtsev M.P. (Eds.), Atlas of the Lower Cretaceous fauna of Northern Caucasus and Crimea.- Vsesoyuznyi Nauchno-Issiedovatel'skii Institut Prirodnykh Gazov, Moscow, p. 239-248.
Shimansky V.N. (1975).- Cretaceous Nautiloids.- Academy of Sciences of the USSR, Transactions of the Palaeontological Institute, Moscow, vol. 150, 208 p.
Sinzow I. (1913).- Beiträge zur Kenntnis der unteren Kreideablagerungen des Nord-Kaukasus.- Travaux du Musée Géologique Pierre le Grand près l'Académie Impériale des Sciences de St. Petersbourg, t. VII, p. 93-117.
Sowerby J. (1812-1815).- The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth, vol. I.- London, p. 9-234.
Spath L.F. (1927).- Revision of the Jurassic cephalopod fauna of Kachh (Cutch).- Memoirs of the Geological Survey of India, Paleontologia Indica, Calcutta, vol. 9, memoir 2, 71 p.
Sundberg F.A. (1984).- Two Cretaceous nautiloids from Baja California, Mexico and southern California.- Bulletin of the Southern California Academy of Sciences, Los Angeles, vol. 83, issue 1, p. 43-52.
Thieuloy J.-P. (1972).- Biostratigraphie des lentilles à pérégrinelles (brachiopodes) de l'Hauterivien de Rottier (Drôme, France).- Géobios, Villeurbanne, no. 5, p. 5-53.
Tintant H. (1980).- Un cas de parallélisme évolutif synchrone chez les Nautiles à côtes du Jurassique.- Societa Geologica de Portugal, Boletim, Lisboa, vol. 22, p. 63-69.
Tintant H. (1993).- L'évolution itérative des Nautiles post-triasiques.- Géobios, Villeurbanne, Mémoire spécial no. 15, p. 359-372.
Tintant H. & Kabamba M. (1983).- Le Nautile, fossile vivant ou forme cryptogène ? Essai sur l'évolution et la classification des Nautilacés.- Bulletin de la Société Zoologique de France, Paris, vol. 108, p. 569-579.
Tzankov V. (1941).- Les céphalopodes fossiles du Danien en Bulgarie.- Review of the Bulgarian Geological Society, Sofia, vol. 13, no. 1, p. 71-76.
Ţibuleac P. & Ajdanlijsky G. (2017).- First report of the genus Euciphoceras (Nautiloidea, Mollusca) from the latest Eocene rocks of the North-western Thrace Basin (Bulgaria).- Analele Stiintifice ale Universitatii "Al. I. Cuza" din Iasi, Seria Geologie, vol. 63, no. 1-2, p. 51-63.
Weber M. (1996).- Die Tongrube 4 der Ziegelei Schnermann in Rothenberge: Litho- und Biostratigraphie eines Ober-Aptprofils in NW-Deutschland.- Geologie und Paläontologie in Westfalen, Münster, no. 45, p. 75-103.
Wiedmann J. (1960).- Zur Systematik jungmesozoischer Nautiliden.- Palaeontographica Abteilung A, Stuttgart, vol. A115, p. 144-206.
Wilmsen M. (2000).- Late Cretaceous nautilids from northern Cantabria, Spain.- Acta Geologica Polonica, Warsaw, vol. 50, no. 1, p. 29-43.
Wilmsen M. (2016).- Early Late Cretaceous nautilids from the Elbtal-group (Saxony, Germany). In: Niebuhr B., Wilmsen M., Kunzmann L. & Stefen C. (Eds.), 87th Annual Conference of the Paläontologische Gesellschaft e.V. Programme.- Abstracts, Field trip guides, Dresden 2016, p. 165.
Wittler F.A., Roth R. & Legant J. (1999).- Die Nautiliden der oberen Kreide (Cenoman-Campan) vom Süd- und Westrand des Münsterländer Beckens.- Arbeitskreis Paläontologie Hannover, vol. 27, no. 1, p. 1-52.
Wright C.W. & Wright E.V. (1951).- A survey of the fossil Cephalopoda at the Chalk of Great Britain: Primarily a nomenclatorial revision of Daniel Sharpe's "Description of the fossil remains of Mollusca found in the Chalk of England. Part I, Cephalopoda".- Palaeontographical Society, London, 40 p.
Plate
I: |
|
|
Plate
II: |
|
|
Plate
III: |
|
|
Plate
IV: |
|
|
Plate
V: |
|
|
Plate
VI: |
|
|
Plate
VII: |
|
|
Plate
VIII: |
|
|
Plate
IX: |
|
|
Plate
X: |
|
|
Plate
XI: |
|
|
Plate
XII: |
|
|
Plate
XIII: |
|
|
Plate
XIV: |
|
|
Plate
XV: |
|
|
Plate
XVI: |
|