◄ Carnets Geol. 21 (13) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Materials and methods]
[4. Systematic palaeontology]
[5. Taphonomy] [6. Biostratigraphic
considerations]
[7. Palaeobiogeographical implications] [8. Conclusions]
and ... [Bibliographic references]
Department of Geology and Meteorology, Institute of Geography and Earth
Sciences, Faculty of Sciences, The University of Pécs, 6 Ifjúság útja,
H-7624, Pécs (Hungary); ORCID: 0000-0001-9699-7711
MSc student, The University of Pécs, Faculty of Sciences, 6 Ifjúság útja,
H-7624, Pécs (Hungary); ORCID: 0000-0002-4337-0410
BSc student, The University of Pécs, Faculty of Sciences, 6 Ifjúság útja,
H-7624, Pécs (Hungary)
BSc student, The University of Pécs, Faculty of Sciences, 6 Ifjúság útja,
H-7624, Pécs (Hungary)
BSc student, The University of Pécs, Faculty of Sciences, 6 Ifjúság útja,
H-7624, Pécs (Hungary)
BSc student, The University of Pécs, Faculty of Sciences, 6 Ifjúság útja,
H-7624, Pécs (Hungary)
Published online in final form (pdf) on July 4, 2021
DOI 10.2110/carnets.2021.2113
![]()
[Editor: Bruno R.C. Granier; language editor: Nigel D. Chapman]
![]()
A new collection at Zengővárkony (Mecsek Mountains, Hungary) provided a rich and diverse but poorly preserved cephalopod-dominated fossil assemblage representing the Kimmeridgian and the lower Tithonian. The material came from mixed scree, soil, and amongst roots affected by weathering processes having been exposed to the elements for a long time. The nautiloid Pseudaganides strambergensis is the first record from the Mecsek Mountains. Due to the weathering, the ammonite fauna consists of mainly fragmentary and dissolved individuals that comprises 528 specimens belonging to 34 species and 30 genera out of which 20 species and 15 genera are reported for the first time from the Mecsek Mountains. The fauna includes specimens of known taxa. No new taxa are introduced. Based on the comparison with other faunas, this assemblage most closely resembles the fauna of the Venetian Alps (Italy). Additional faunal elements include aptychi (Laevaptychus latus, Lamellaptychus murocostatus), belemnites (Hibolithes semisulcatus), and an indetermined brachiopod. The first record of Spiraserpula spirolinites, an encrusting fossil polychaete preserved on the internal mould of a Taramelliceras shell fragment indicates favourable bottom conditions for the epifauna. The presence of Aspidoceras caletanum, Gravesia aff. gigas, and Pseudowaagenia inerme indicates faunal connections with the Submediterranean Province of the Tethys, which is in line with the tectonic and palaeogeographical position of the Mecsek Zone during the Late Jurassic. The ammonite assemblage represents elements of five Tethysian ammonite zones of the Kimmeridgian and Tithonian. The lower Kimmeridgian Herbichi Zone is indicated by Streblites tenuilobatus and Praesimoceras herbichi. The upper Kimmeridgian Acanthicum Zone is indicated by Aspidoceras acanthicum, and the Cavouri Zone by Mesosimoceras cavouri and Aspidoceras caletanum. The upper Kimmeridgian Beckeri Zone is suggested by Hybonoticeras pressulum and Pseudowaagenia inerme. Whereas Gravesia aff. gigas, Lithacoceras aff. siliceum, and Malagasites? denseplicatus are faunal elements characterising the Early Tithonian Hybonotum Zone. Phylloceratid and lytoceratid specimens account only for 12% of the fauna, while the majority of the specimens belong to the Oppeliidae and Ataxioceratidae (60%).
• taxonomy;
• Upper Jurassic;
• ammonites;
• Tisza Mega-unit;
• Submediterranean influence;
• serpulid encrustation
Bujtor L., Albrecht R., Farkas C., Makó M., Maróti D. & Miklósy Á. (2021).- Kimmeridgian and early Tithonian cephalopods from the Kisújbánya Limestone Formation, Zengővárkony (Mecsek Mountains, southern Hungary), their faunal composition, palaeobiogeographic affinities, and taphonomic character.- Carnets Geol., Madrid, vol. 21, no. 13, p. 265-314.
Les céphalopodes du Kimméridgien et du Tithonien inférieur de la Formation du Calcaire de Kisújbánya, Zengővárkony (Massif du Mecsek, Hongrie méridionale), leur composition faunistique, leurs affinités paléobiogéographiques et leur caractéristiques taphonomiques.- Un nouvel échantillonnage ŕ Zengővárkony (Massif du Mecsek, Hongrie) a fourni un assemblage fossile dominé par les céphalopodes, riche et diversifié mais mal préservé, représentant le Kimméridgien et le Tithonien inférieur. Le matériel provient d'un mélange d'éboulis, de sols et de racines, le tout ayant été exposés aux éléments pendant une longue période et affecté par les processus d'altération. Le nautiloďde Pseudaganides strambergensis est signalé pour la premičre fois dans le Massif du Mecsek. En raison de l'altération, l'ammonitofaune consiste principalement en des éléments fragmentés et dissous qui représentent 528 spécimens appartenant ŕ 34 espčces et 30 genres parmi lesquels 20 espčces et 15 genres sont signalés pour la premičre fois dans le Massif du Mecsek. La faune ne comporte que des spécimens de taxons déjŕ connus. Aucun nouveau taxon n'y est reconnu. En se fondant sur la comparaison avec d'autres faunes, cet assemblage ressemble trčs fortement ŕ la faune des Alpes vénitiennes (Italie). Des éléments faunistiques additionnels incluent des aptychi (Laevaptychus latus, Lamellaptychus murocostatus), des bélemnites (Hibolithes semisulcatus) et un brachiopode indéterminé. Le premier signalement de Spiraserpula spirolinites, un polychčte fossile encroűtant conservé sur le moulage interne d'un fragment de coquille de Taramelliceras, indique des conditions de fond favorables ŕ l'épifaune. La présence de Aspidoceras caletanum, Gravesia aff. gigas et de Pseudowaagenia inerme indique des connexions avec la province sub-méditerranéenne de la Téthys, qui dans le prolongement tectonique et paléogéographique de la Zone du Mecsek pendant le Jurassique supérieur. L'assemblage d'ammonites comporte des éléments de cinq ammonitozones téthysiennes du Kimméridgien et du Tithonien. La Zone ŕ Herbichi du Kimméridgien inférieur est indiquée par Streblites tenuilobatus et Praesimoceras herbichi. La Zone ŕ Acanthicum du Kimméridgien supérieur est caractérisée par Aspidoceras acanthicum, et la Zone ŕ Cavouri par Mesosimoceras cavouri et Aspidoceras caletanum. La Zone ŕ Beckeri du Kimméridgien supérieur est suggérée par Hybonoticeras pressulum et Pseudowaagenia inerme, tandis que Gravesia aff. gigas, Lithacoceras aff. siliceum et Malagasites ? denseplicatus sont des éléments faunistiques caractérisant la Zone ŕ Hybonotum du Tithonien inférieur. Les spécimens de phyllocératides et de lytocératides représentent seulement 12% de la faune, tandis que la majorité des spécimens appartient aux Oppeliidae et aux Ataxioceratidae (60%).
• taxonomie ;
• Jurassique supérieur ;
• ammonites ;
• Méga-unité de Tisza ;
• influence sub-méditerranéenne ;
• algues calcaires ;
• incrustation de serpulides
Kimmeridgian and early Tithonian ammonites are widely known and abundant in Western Tethysian settings. Kimmeridgian ammonites have been studied in detail from many different localities of the Hungarian Transdanubian Middle Range (Vigh, 1961, 1984; Főzy, 1993a; Főzy et al., 2011, 2013, among others). Kimmeridgian ammonites from the Mecsek Mountains were first trivially reported by Peters (1862, p. 281). Böckh (1880, p. 17), who carried out geological mapping in the Zengővárkony region, reported with neither description nor figuration Pygope diphya from the Tithonian limestones of the abandoned quarry of Pusztafalu (today referred to as Zengővárkony), that nevertheless suggested for the first time the presence of Upper Jurassic strata in 1874. Later Vadász (1935, p. 63) proved this Jurassic terrain by listing the following taxa from Zengővárkony, the only locality that provided Kimmeridgian ammonites from the Mecsek Mountains:
Ptychophylloceras cf. ptychoicum Quenstedt
Holcophylloceras cf. polyolcum Benecke
Sowerbiceras cf. protortisulcatum Pompeckj
Lytoceras cf. sutile Oppel
Taramelliceras nobilis Neumayr
Taramelliceras cf. compsa Oppel
Taramelliceras cf. trachynota Oppel
Taramelliceras cf. pseudoflexuosum Favre
Taramelliceras succedens Oppel
Pseudowaagenia cf. haynaldi Herbich
Pseudowaagenia cf. pressulum Neumayr
Aspidoceras cf. acanthicum Oppel
Aspidoceras cf. neuburgensis Oppel
Aspidoceras cf. cyclotum Oppel
Aspidoceras cf. binodosum Oppel
Aspidoceras cf. liparum Oppel
Acanthosphaerites longispinum Sowerby
Simoceras sp.
Virgatosphinctes sp.
Perisphinctes div. sp.
The majority of the determinations reflect the poor preservation (whorl fragments, dissolution), and scarcity of collectible ammonite specimens. Vadász (1935, p. 62) concluded that this fauna represents the Streblites tenuilobatus 'level', which partly corresponds to the Strombecki and Herbichi zones of the current Mediterranean zonation. Regarding the Upper Jurassic-Lower Cretaceous lithostratigraphy of the Mecsek Mountains, Fülöp (1967) indicated a 160 m thick, continuous succession from the top Callovian to the Berriasian. It includes a 7 m thick Tithonian-Berriasian sequence at Zengővárkony with an unconformity at the T/B boundary and distribution of rich microfauna (Globochaete, Cadosina, Tintinnida, Saccocoma, and Radiolaria) around the boundary beds. At the beginning of the 1960s the Hungarian Geological Survey started 1:10.000 mapping in the region referring to a continuous Upper Jurassic sequence at Zengővárkony (Hetényi et al., 1968) and mentioned a Hybonoticeras hybonotum (Oppel) from the Kimmeridgian at Zengővárkony. Nagy (1971, p. 324) added nominally some previously unknown ammonite taxa, with neither illustration or inventory numbers for his ammonites: Hemihaploceras nobilis [sic!] (Neumayr), Taramelliceras trachinotum (Oppel), Ataxioceras polyplocum (Reinecke), and Aulacostephanus yo (Orbigny). Wein (1974, p. 61) summarized the geological structure of the Mecsek Mountains, but only proved the faunal list of Vadász (1935). In the 1990s Főzy (1993b) conducted field work in the Mecsek Mountains and collected some Jurassic/Cretaceous ammonites concluding (Főzy, 1993b, p. 198) that Kimmeridgian ammonites are only found at Zengővárkony. Főzy (1993b, p. 198) practically replicated the faunal list of Vadász (1935, p. 63) adding two new taxa: Taramelliceras pugile (Neumayr) and ?Orthosphinctes sp. Főzy (1993b, p. 199) also referred to the occurrence of Nebrodites cavouri (Gemmellaro) absent from his faunal list, included in the 'Simoceras' sp. of Vadász (1935) but without the repository number thwarting attempted retrieval of this specimen from the old collection of MBFSz (Mining and Geological Survey of Hungary). Based on his observations, and the earlier work of Vadász (1935), Főzy (1993b, p. 203) summarized that the Kimmeridgian fauna of Zengővárkony represents the upper Kimmeridgian Beckeri and Cavouri zones of the Mediterranean zonal scheme.
Tithonian strata can be found on the surface at a handful of places in the Mecsek Mountains (Wein, 1965; Főzy, 1993b, Fig. 2) representing the lower Tithonian albeit yielding only a poor ammonite fauna. Vadász (1935, p. 63) listed the following lower Tithonian ammonites from Zengővárkony: Lytoceras cf. montanum Oppel, Aspidoceras cf. binodum Oppel, A. cf. cyclotum Oppel, and Perisphinctes cimbricus Neumayr (= Biplisphinctes cimbricus, cf. Schlögl & Zorn, 2012). Nagy (1971, p. 326) added some interesting forms: Prorasenia witteana (Oppel), Virgatosphinctes [= Paraulacosphinctes] transitorius (Oppel), Substreblites zonarius (Oppel). Nagy and Raucsik (in Főzy, 2012) mentioned Haploceras cf. elimatum and Usseliceras sp. from the Kisújbánya Limestone Fm., but lacking description, illustration and accession numbers rendering specimens unavailable for this study. Recently Bujtor et al. (2021) reported a lower Tithonian ammonite from Zengővárkony (Volanoceras volanense), which indicates the Ponti Zone. Before the present authors started their field work, our knowledge on the Kimmeridgian/early Tithonian ammonite fauna of the Mecsek Mountains was based on faunal lists that reflected a hundred-year-old knowledge. It hinted at the presence of some lower and upper Kimmeridgian ammonite zones, as well as Tithonian strata. However systematic descriptions, taxonomic evaluations and zonal subdivisions were unavailable for the sequence.
The present authors conducted field work in July 2018 and revisited the locality in May 2019 and July 2020 for further collecting. The field work provided some 500 ammonites that represent the Kimmeridgian and lower Tithonian from the investigated locality, and provided many ammonite taxa previously unknown from the Mecsek Mountains. We also studied the old collection of the Mining and Geological Survey of Hungary, referred to hereafter as MBFSz and assessed deposited and unpublished material collected during field work by Andor Semsey, János Böckh (1874-1881), and Elemér Vadász (1930-1933). The aim of the present paper is to describe and figure for the first time the Kimmeridgian and lower Tithonian ammonites of Zengővárkony (Mecsek Mountains) that have been known since 1862 yet remained unpublished so far. Notwithstanding the poor preservation of this Kimmeridgian ammonite fauna from the Zengővárkony locality in the Mecsek Mountains these important specimens were collected to provide insights into the fauna and stratigraphy of the Upper Jurassic of this biogeographically and tectonically interesting unit.
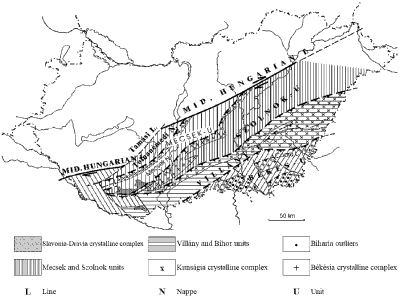
As the northernmost unit of the tectonic domain, the Mecsek Mountains (Fig. 1 ![]() ) belong to the Tisza Mega-Unit (Haas & Péró,
2004) which is considered a microplate (Vörös,
1993; Csontos & Vörös,
2004) having a nappe structure. The Late Jurassic sedimentary succession of the Mecsek Mountains (Fig. 2
) belong to the Tisza Mega-Unit (Haas & Péró,
2004) which is considered a microplate (Vörös,
1993; Csontos & Vörös,
2004) having a nappe structure. The Late Jurassic sedimentary succession of the Mecsek Mountains (Fig. 2 ![]() ) is characterized by calm, pelagic limestone sedimentation (Raucsik in Főzy,
2012) with ammonitico rosso type red, nodular, sometimes cherty and thin Oxfordian and Kimmeridgian condensed beds with regular dissolution (Nagy, 1964). These formations (e.g., Fonyászó Limestone Fm., Kisújbánya Limestone Fm.) are less than 50 metres in thickness. The youngest formation (Márévár Limestone Fm.) is characterized by a 100 m thick limestone succession with somehow elevated sea floor that has a maiolica facies character in the upper part. Around the T/B boundary, the first indication of the later pronounced volcanic activity (Mecsekjánosi Basalt Fm.) appears, which is intensified in the late Berriasian and related to a continental rifting of the tectonic zone. However, there are indications for the earlier (Oxfordian-Kimmeridgian) volcanic activities in the region (Császár & Turnšek,
1996). The Late Jurassic sedimentation in the Mecsek Mountains provided a thin, continuous, and at maximum 200 m thick limestone succession, which has a poorly preserved macrofauna dominated by ammonites accompanied by belemnites and very scarce
brachiopods; irregular echinoids also occur.
) is characterized by calm, pelagic limestone sedimentation (Raucsik in Főzy,
2012) with ammonitico rosso type red, nodular, sometimes cherty and thin Oxfordian and Kimmeridgian condensed beds with regular dissolution (Nagy, 1964). These formations (e.g., Fonyászó Limestone Fm., Kisújbánya Limestone Fm.) are less than 50 metres in thickness. The youngest formation (Márévár Limestone Fm.) is characterized by a 100 m thick limestone succession with somehow elevated sea floor that has a maiolica facies character in the upper part. Around the T/B boundary, the first indication of the later pronounced volcanic activity (Mecsekjánosi Basalt Fm.) appears, which is intensified in the late Berriasian and related to a continental rifting of the tectonic zone. However, there are indications for the earlier (Oxfordian-Kimmeridgian) volcanic activities in the region (Császár & Turnšek,
1996). The Late Jurassic sedimentation in the Mecsek Mountains provided a thin, continuous, and at maximum 200 m thick limestone succession, which has a poorly preserved macrofauna dominated by ammonites accompanied by belemnites and very scarce
brachiopods; irregular echinoids also occur.
|
Figure 1: Structural position of the Mecsek Mountains in Hungary with the indication of the major tectonic units. Map based on Haas and Péró
(2004), simplified. |
|
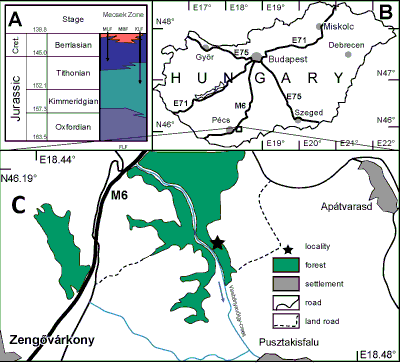
Figure 2: Locality map and the lithostratigraphic subdivision of the Upper Jurassic-Lower Cretaceous (pars) strata mentioned in the text. A. Lithostratigraphy of the Mecsek zone for the study period. Numerical ages in million years after Cohen et al.
(2013). B. Simplified map of Hungary. Black rectangle indicates the study area northeast from Pécs. C. The Zengővárkony area indicating the abandoned quarry. The star indicates the locality on the left (=Eastern) bank viewed looking south - which is downstream of the Vasbányavölgy creek, an abandoned small quarry from where the ammonites were collected. Legend: FLF: Fonyászó Limestone Fm; KLF: Kisújbánya Limestone Fm; MBF: Mecsekjánosi Basalt Fm; MLF: Márévár Limestone Fm. Arrow indicates the direction of water flow. Lithostratigraphic units after Császár
(1996) for the Cretaceous and Főzy
(2012) for the Jurassic, simplifying and unifying their diagrams. |
Studied sections: The shallow valley of the Vasbányavölgy-creek (Fig. 2.C ![]() ) cuts through the Middle Jurassic-Lower Cretaceous continuous limestone succession from the Oxfordian (Nagy,
1964) to the upper Berriasian (Grabowski et al.,
2016), however Callovian strata is also present (Molnár,
1961). South from the Zengővárkony lime-kilns, Early and Middle Jurassic strata are also exposed (Sidó,
1966). An abandoned quarry called 'Várkonyi mészkemence' (=lime-kiln of Várkony) on the right bank (west) of the Vasbányavölgy creek exposes the upper Kimmeridgian to the lower Berriasian strata of the Márévár Limestone Formation. In the old literature this was the source area for ammonites collected by Andor Semsey, János Böckh, Karl Zittel, and Elemér Vadász (Böckh,
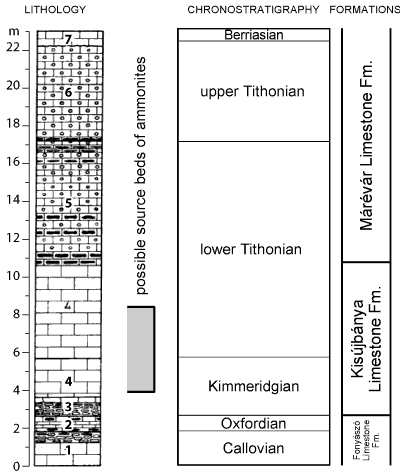
1880; Vadász, 1935). However, on the left side (east) of the creek there is a smaller, also abandoned quarry that exposes an older formation (Kisújbánya Limestone Fm.) of typical red coloured, condensed horizons representing the lower Kimmeridgian. These typical ammonitico rosso strata (bed 3, Fig. 3
) cuts through the Middle Jurassic-Lower Cretaceous continuous limestone succession from the Oxfordian (Nagy,
1964) to the upper Berriasian (Grabowski et al.,
2016), however Callovian strata is also present (Molnár,
1961). South from the Zengővárkony lime-kilns, Early and Middle Jurassic strata are also exposed (Sidó,
1966). An abandoned quarry called 'Várkonyi mészkemence' (=lime-kiln of Várkony) on the right bank (west) of the Vasbányavölgy creek exposes the upper Kimmeridgian to the lower Berriasian strata of the Márévár Limestone Formation. In the old literature this was the source area for ammonites collected by Andor Semsey, János Böckh, Karl Zittel, and Elemér Vadász (Böckh,
1880; Vadász, 1935). However, on the left side (east) of the creek there is a smaller, also abandoned quarry that exposes an older formation (Kisújbánya Limestone Fm.) of typical red coloured, condensed horizons representing the lower Kimmeridgian. These typical ammonitico rosso strata (bed 3, Fig. 3 ![]() ) also provided a useful fauna for study representing the Strombecki and Herbichi zones, in accordance with Vadász (1935). Above the red coloured, nodular limestone strata, bed 4 (on Fig. 3
) also provided a useful fauna for study representing the Strombecki and Herbichi zones, in accordance with Vadász (1935). Above the red coloured, nodular limestone strata, bed 4 (on Fig. 3 ![]() ) evidences a colour change to yellowish-white, and ammonites disappear. The succession is then covered by scree, debris and soil. After careful field work and trial-channelling, a remarkably rich albeit poorly preserved, fragmentary ammonite assemblage was found in the scree, soil and among roots. These ammonites are weathered and dissolved being accumulated for thousands of years in the soil, while the host limestone beds were eroded. The ammonites represent the Kimmeridgian and the lower Tithonian strata, with the same lithological features (coarse-grained, yellowish-white or grey, poorly stratified limestone) that is typical for the Kisújbánya Limestone Formation. In Figure 3
) evidences a colour change to yellowish-white, and ammonites disappear. The succession is then covered by scree, debris and soil. After careful field work and trial-channelling, a remarkably rich albeit poorly preserved, fragmentary ammonite assemblage was found in the scree, soil and among roots. These ammonites are weathered and dissolved being accumulated for thousands of years in the soil, while the host limestone beds were eroded. The ammonites represent the Kimmeridgian and the lower Tithonian strata, with the same lithological features (coarse-grained, yellowish-white or grey, poorly stratified limestone) that is typical for the Kisújbánya Limestone Formation. In Figure 3 ![]() , we indicated the possible source rocks of the collected fauna, that is described in this paper. Careful analysis of the accession labels of older material housed in MBFSz, let us discriminate those ammonites that were collected here and six ammonite specimens from older repositories were included in this study. The present paper therefore describes the ammonites collected from scree and from among roots and mixed debris and soil from the so-called 'small quarry beside the lime-kiln of Várkony' indicated by the star on Figure 2.C
, we indicated the possible source rocks of the collected fauna, that is described in this paper. Careful analysis of the accession labels of older material housed in MBFSz, let us discriminate those ammonites that were collected here and six ammonite specimens from older repositories were included in this study. The present paper therefore describes the ammonites collected from scree and from among roots and mixed debris and soil from the so-called 'small quarry beside the lime-kiln of Várkony' indicated by the star on Figure 2.C ![]() . The locality (Fig. 4
. The locality (Fig. 4 ![]() ) picture shows the accumulated material as cobbles and fragments during our field work that provided the remarkably rich and diverse ammonite fauna described herein. Ammonites were not sampled bed-by-bed, but were collected from the scree.
) picture shows the accumulated material as cobbles and fragments during our field work that provided the remarkably rich and diverse ammonite fauna described herein. Ammonites were not sampled bed-by-bed, but were collected from the scree.
|
Figure 3: The upper Jurassic-lower Cretaceous section at Zengővárkony, Mecsek Mountains traversing the abandoned quarries. Lithological column after Nagy (1961, 1964), simplified. Shaded stripe indicates the supposed stratigraphical position of the ammonites included in this study. Legend: 1: light coloured yellowish-grey thick bedded limestone; 2: thin bedded limestone; 3: marly-clayey, red coloured, patchy and nodular limestone; 4: light coloured yellowish-grey, thick bedded limestone; 5: light coloured yellowish-grey brecciated limestone with light grey flint nodules; 6: yellowish-white brecciated limestone; 7: brownish-yellow clayey limestone. Chronostratigraphic subdivision of the section is based on micropalaeontological investigations of Nagy (1961, 1964). |
|
Figure 4: Locality exposing the Kisújbánya Mészkő Formation. Ammonites were collected from scree, and from the mixed soil, debris and roots. One of the authors (Á.M.) points to the mixed soil and debris material from which the specimens appeared as cobbles.
GPS coordinates: 46°11'03.5"N 18°27'34.2"E. |
The cephalopods from Zengővárkony available for our study comprise 525 specimens (including belemnites and aptychi). Of these 519 were collected by the authors and the other 6 are housed in the historical collection of the MBFSz with hand-written information on the original labels of János Böckh and Andor Semsey from the 1870-1880s. All six specimens referred to on the labels as 'small quarry beside the lime-kiln of Várkony' are included in this paper.
Table 1: Higher taxa of macrofossils with numbers of specimens and percentages in the Zengővárkony fauna examined in the present paper.
| Taxa | number of specimens | percentage |
| Ammonoidea | 507 | 96.0 |
| Annelida | 1 | 0.2 |
| Aptychi | 12 | 2.3 |
| Belemnoidea | 6 | 1.1 |
| Brachiopoda | 1 | 0.2 |
| Nautiloidea | 1 | 0.2 |
| Total | 528 | 100.0 |
Macrofossils with the respective numbers of specimens and percentages are shown in Table 1 of the Zengővárkony Late Jurassic fauna examined in the present paper. The ammonite fauna is dominated by ataxioceratids, aspidoceratids and oppeliids, that represent 84% of the specimens but the less abundant phylloceratids and lytoceratids are also present. Abbreviations: FAD: First Appearance Datum; Fm: formation; D: diameter of the conch; Wb: width of the conch; Wh: height of the last whorl; U: diameter of the umbilicus or umbilical lobe - up to the context; L: lateral lobe; N/2: number of primary ribs on the last half whorl; n.d.: no data due to the fragmentary status or poor preservation of the specimen. Dimensions are given in mm. Measurements were acquired by a manual caliper. Measurements in brackets refer to estimated data due to poor preservation. Scale bars indicate 1 cm, otherwise as noted. Rib curves are not prepared due to poor preservation of the specimens. Zonal subdivisions of Kimmeridgian and Tithonian follows Zeiss (2003) with the emendations of Wierzbowski et al. (2016) and Hesselbo et al. (2020).
Abbreviations for aptychi: L: distance between the terminal point and the umbilicus projection; S: distance between the apex and the terminal point; Lat: distance between the terminal point and the point of maximum valve width (Měchová et al., 2010).
Photos were taken by L.B. with a Nikon D3500 DSLR camera under natural sunlight. Due to the Covid-19 curfew the university facilities (laboratories, microscopes etc.) were permanently inaccessible, therefore ammonium-chloride coating of the fossils and laboratory photographic conditions were not available.
Repositories and institutional abbreviations. All specimens included in this study are deposited in the MBFSz palaeontological collection (of the Mining and Geological Survey of Hungary, Budapest); figured and measured specimens with the inventory numbers starting with "J" come from the historical MBFSz collection of MBFSz with specimens donated by János Böckh and Andor Semsey. Figured and measured specimens collected by the present authors are labelled with prefix "J 2020".
Taxonomic remarks. Systematics of Phyllo- and Lytoceratina are in accordance with Arkell et al. (1957); systematics of the Ammonitina (excl. Perisphinctoidea) follows Donovan and Callomon (1980), and systematics of Perisphinctoidea are from Énay and Howarth (2019). Recently Parent et al. (2020) proposed to raise aspidoceratid ammonites to superfamily level: Their proposal is followed here. Fischer and Zeiss (1987) and later Scherzinger et al. (2006) classified the subfamily Gravesiinae as incertae sedis at the family level according to the uncertain affinities of Gravesia. Their views were rejected by Énay and Howarth (2019), and Gravesia is considered belonging to Ataxioceratinae, which is accepted here. Due to the limited number of specimens per taxa and their poor preservation subspecies names are not assigned in this study. King and Evans (2019) also suggested modified higher ranked systematics for nautiloids, which is followed here. Systematics of Serpulidae are from Ippolitov (2007).
Phylum Annelida Lamarck, 1809
Class Polychaeta Grube, 1850
Subclass Canalipalpata Rouse & Fauchald, 1997
Order Sabellida Fauchald, 1977
Family Serpulidae Rafinesque, 1815
Subfamily Serpulinae Rafinesque, 1815
Genus Spiraserpula Regenhardt, 1961
Type species: Spiraserpula spiraserpula Regenhardt, 1961
Spiraserpula spirolinites (Münster in Goldfuss, 1831)
(Fig. 5.A1-A2 ![]() )
)
1831. Serpula Spirolinites Münster, Münster in Goldfuss, p. 229, Pl. 68, fig. 5.a-c.
1996. Serpula (Dorsoserpula) spirolinites Münster in Goldfuss, Gerasimov et al., p. 50, Pl. 6, fig. 10.a-b.
Material. One fairly preserved specimen on an internal mould of a whorl fragment of a Taramelliceras (J 2020.291.1).
Dimensions. The only specimen has 6 mm maximum diameter.
Description. Small sized planispiral structure on the midflank of an ammonite internal mould. There are five, densely spaced spiral whorls that are firmly attached to each other. The spiral is three-dimensional and deepened into the mould. The width of the spiral ridge at the innermost part is 160 μm and continuously increasing till the end of the spiral where it has 500 μm width.
Remarks. The holotype consists of three, densely coiled individual spirals of 5 whorls that continued (Goldfuss, 1831, Pl. 68, fig. 5.b) or discontinued (ibidem, Pl. 68, fig. 5.a upper left) in meandering tubes. These tubes are positive calcareous fossils on a shell surface, while the present specimen is a negative imprint that originally was attached to the internal wall of the shell of the host Taramelliceras (Franz Fürsich personal comm.). The spiral structures are close to the holotype in the number of spirals, densely packed coils and diameter; however, the meandering continuation is not present but this is not necessarily preserved. After final burial the aragonite shell dissolved, but the infilling lime mud filled the conch and preserved the negative imprint of the originally internally attached serpulid tube. First record from Hungary.
Stratigraphic and geographic distribution. Callovian (Ippolitov, 2007) to Kimmeridgian-lower Tithonian of Mecsek Mountains, Hungary, Central Russia (Gerasimov et al., 1996), Bavaria, Germany (Goldfuss, 1831), Switzerland (Tribolet, 1873).
|
Figure 5: Upper Kimmeridgian-lower Tithonian annelids, nautiloids and ammonoids from Zengővárkony, Mecsek Mountains, Hungary. A. Spiraserpula spirolinites (Münster in Goldfuss,
1831) on the upper flank of a Taramelliceras sp. whorl fragment. A1. specimen J 2020.291.2; A2. close-up view of the polychaete spiral tube imprint on the internal mould. B. Pseudaganides strambergensis (Oppel,
1865), specimen J 2020.265.1. B1. lateral view; B2. apertural view. C. Calliphylloceras benacense (Catullo,
1847), specimen J 2020.327.1. D. Sowerbyceras loryi (Munier-Chalmas in Hébert,
1875), specimen J 2020.267.1. D1. lateral view; D2. cross-section; D3. ventral view. E. Lytoceras polycyclum Neumayr, 1871a; I1. specimen J
2020.289.1. Scale bars indicate 1 cm except A2, which indicates 0.5 mm. |
Phylum Mollusca Linnaeus, 1758
Class Cephalopoda Cuvier, 1797
Subclass Nautilia Wade, 1988 [nom. corr. King & Evans, 2019]
Order Nautilida Agassiz, 1847
Family Nautilidae Orbigny, 1840
Subfamily Pseudaganinae Kummel, 1956
Genus Pseudaganides Spath, 1927
Type species: Nautilus kutchensis Waagen, 1873
Pseudaganides strambergensis (Oppel, 1865)
(Fig. 5.B1-B2 ![]() )
)
1865. Nautilus Strambergensis Opp., Oppel, p. 546.
1868. Nautilus Strambergensis Opp., Zittel, p. 42, Pl. 2, figs. 8-11.
1916. Nautilus strambergensis Zittel, Jekelius, p. 265, Pl. 6, fig. 3.
1956. Pseudaganides strambergensis (Oppel), Kummel, p. 392, fig. 15.J.
non 1973. Nautilus strambergensis Zittel, Preda, Pl. 3, fig. 1.
2013. Pseudaganides strambergensis (Oppel), Főzy et al., p. 29, Fig. 6.
Material. One fairly preserved internal mould.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.265.1. | 47.6 | (20) | (28) | (3) | 0.714 | 0.588 | 0.06 |
Description. Small sized, fairly preserved phragmocone; compressed, involute with very small umbilicus; rounded umbilical shoulder, passing to convex flanks then converting into a rounded, smooth venter. Septums placed 6 mm from each other on the early, and 7 mm on later whorls. There is a shallow ventral lobe, and a high lateral lobe and a shallow saddle.
Remarks. The present specimen is most similar to the specimen of Jekelius (1916, Pl. 6, Fig. 3), the only difference is that the latter has body chamber on the last half whorl, while the present specimen is thoroughly septate. Its cross section is identical to the cross section of the specimen illustrated by Kummel (1956, p. 393, Fig. 15.J) Suture of the present specimen is similar to P. schneidi (Schairer & Barthel, 1977, Pl. 11, fig. 3.a) however in cross section it is markedly different compared to the present specimen. P. franconicus has shallower lobes as Jekelius (1916) had already noted. Preda (1973, Pl. 3, fig. 1) figured the ventral view of a very wide, depressed specimen, which is out of the range of this species. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Upper Jurassic of Central Europe (Czech Republic: Kummel, 1956; Tithonian of Gerecse Mountains, Hungary: Főzy et al., 2013; Romania: Jekelius, 1916).
Subclass Ammonoidea Zittel, 1884
Order Ammonitida Fischer, 1882
Suborder Phylloceratina Arkell, 1950
Family Phylloceratidae Zittel, 1884
Subfamily Phylloceratine Zittel, 1884
Genus Phylloceras Suess, 1865
Type species: Ammonites heterophyllus J. Sowerby, 1820
Phylloceras sp. ind.
Material. 13 fragmentary internal moulds (J 2020.174.2, 198.1, 203.1, 204.1, 214.1, 233.1, 266.1 and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.91.1. | 43.6 | 17.2 | 25.3 | 2.5 | 0.680 | 0.580 | 0.057 |
| J 2020.138.1. | 58.8 | (21) | (38) | 1.7 | 0.553 | 0.646 | 0.029 |
| J 2020.198.1. | (77) | 24.3 | (46) | (10) | 0.528 | 0.597 | 0.130 |
| J 2020.232.1. | 79.3 | 26.5 | 44.5 | 9.6 | 0.595 | 0.561 | 0.121 |
| J 2020.263.1. | 63.6 | 23.4 | 39.9 | 6.5 | 0.586 | 0.627 | 0.102 |
| J 2020.285.1. | 70.3 | (22) | (42) | (4) | 0.524 | 0.597 | 0.057 |
Subfamily Calliphylloceratinae Spath, 1927
Genus Calliphylloceras Spath, 1927
Type species: Phylloceras disputabile Zittel, 1869
Calliphylloceras benacense (Catullo, 1847)
(Fig. 5.C ![]() )
)
1847. Ammonites benacensis, Cat., Catullo, p. 9 [seconda appendice], Pl. 13, fig. 1.a-b.
1871a. Phylloceras Benacense Catullo, Neumayr, p. 336, Pl. 15, fig. 3.
? 1878. Phylloceras Békasense nov. sp., Herbich, p. 112, Pl. 3, fig. 1.
1879. Ammonites (Phylloceras) Benacensis, Catullo, Favre, p. 14, Pl. 2, fig. 1.
1959. Calliphylloceras benacense Cat., Collignon, Pl. 97, fig. 368.
1976. Calliphylloceras benacense (Catullo), Joly, p. 192, Pl. 10, fig. 1; Pl. 44, fig. 1.a-b.
1984. Calliphylloceras benacense (Catullo), Verma & Westermann, p. 28, Pl. 1, fig. 2.
1986. Calliphylloceras benacense (Catullo), Sarti, p. 485, Pl. 1, fig. 1.
2007. Calliphylloceras benacense (Catullo), Cecca & Savary, p. 514, Fig. 4.A.
2013. Calliphylloceras benacense (Catullo), Grigore, p. 85, Pl. 1, figs. 1, 3.
Material. One fairly preserved internal mould.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.327.1. | 65.8 | (21) | (38) | 4.5 | 0.553 | 0.577 | 0.068 |
Description. Fairly preserved, middle sized conch representing phragmocone and body chamber. Coiling involute, umbilicus small, shallow. Umbilical shoulder short, rounded. No ventral shoulder. Conch compressed, flanks convex, converging. Venter rounded. Fine, narrow, shallow line situated on the mid-siphonal region. There are six, shallow and wide constrictions on the conch that rise from the umbilical shoulder radially; shortly above the umbilical shoulder they become prorsiradiate and travel radially on the flanks. On the upper flank at ľ flank height the constrictions slightly bend forward. Suture, aperture not seen.
Remarks. The present specimen lacks the fine riblets depicted on the holotype, however other representatives of the species (cf. Collignon, 1959; Joly, 1976) did not show them. This is due to the poor preservation of the studied ammonite. Herbich (1878, p. 112) introduced his new species (P. bekasense) based on a single, poorly preserved specimen without differential diagnosis. His specimen is identical with Catullo's holotype; therefore, its specific independence is questionable as Grigore (2013, p. 85) already underlined. First record from Hungary.
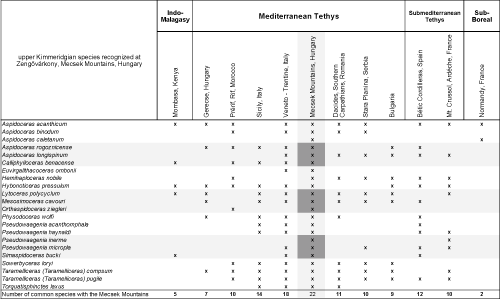
Stratigraphic and geographic distribution. Calliphylloceras benacense is reported from the upper Oxfordian to the uppermost Kimmeridgian. Geographically it has a wide geographic distribution in the Tethys (Mediterranean: Cecca & Savary, 2007; Submediterranean and Indo-Malagasy Provinces: Collignon, 1959; Verma & Westermann, 1984).
Genus Holcophylloceras Spath, 1927
Type species: Phylloceras mediterraneum Neumayr, 1871a
Holcophylloceras sp. ind.
Material. Six internal moulds (J 2020.152.1, 172.1, 207.1, 212.1, 231.1, 236.2).
Genus Sowerbyceras Parona & Bonarelli, 1895
Type species: Ammonites tortisulcatus Orbigny, 1840
Sowerbyceras loryi (Munier-Chalmas in Hébert, 1875)
(Fig. 5.D1-D3 ![]() )
)
1875. Phylloceras Loryi, Munier-Chalmas, p. 388.
1877. Ammonites (Phylloceras) Loryi Munier-Chalmas, Favre, p. 19, Pl. 1, figs. 14-15.
1907. Phylloceras (Sowerbyceras) Loryi Munier-Chalmas, Pervinquičre, p. 15, Pl. 1, figs. 1-2.
1979. Sowerbyceras loryi (Munier-Chalmas in Hébert), Sapunov, p. 36, Pl. 4, figs. 4-6.
1981. Sowerbyceras loryi (Munier-Chalmas), Parishev & Nikitin, p. 20, Pl. 6, fig. 7.
1989. Sowerbyceras loryi (Munier-Chalmas), Alkaya, p. 62, Pl. 1, fig. 1.
1993. Sowerbyceras loryi (Munier-Chalmas in Pillet & de Fromental), Sarti, p. 54, Fig. 14, Pl. 1, figs. 3-4.
2011. Sowerbyceras loryi loryi (Munier-Chalmas), Grigore, p. 197, Pl. 2, figs. 7, 9.
2011. Sowerbyceras loryi (Munier-Chalmas in Pillet & de Fromental), Reháková et al., Pl. 7, fig. 5.
2013. Sowerbyceras loryi loryi (Munier-Chalmas), Grigore, p. 89, Pl. 2, figs. 7, 9.
2017. Sowerbyceras loryi, Sarti, Pl. 2, figs. E-I; Pl. 3, figs. A-D, G.
Material. 19 fragmentary internal moulds (J 2020.106.1, 270.1, 264.1, 279.1, 288.1, 314.1, 315.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.192.1. | 64.1 | (28) | (31) | (13) | 0.903 | 0.484 | 0.203 |
| J 2020.267.1. | 48.8 | 21.7 | (21) | 8.0 | 1.033 | 0.430 | 0.164 |
| J 2020.280.1. | 70.8 | 25.8 | (34) | (14) | 0.759 | 0.480 | 0.198 |
| J 2020.281.1. | 58.6 | (15) | (22) | (13) | 0.682 | 0.375 | 0.222 |
| J 2020.282.1. | (57) | 19.2 | (27) | (11) | 0.711 | 0.474 | 0.193 |
| J 2020.299.1. | 62.4 | (23) | (34) | (14) | 0.676 | 0.545 | 0.224 |
| J 2020.316.1. | 59.6 | (21) | (29) | (11) | 0.724 | 0.486 | 0.185 |
| J 2020.347.1. | (53) | (20) | (25) | (10) | 0.800 | 0.472 | 0.189 |
| J 2020.353.1. | 56.1 | (21) | (26) | (12) | 0.807 | 0.463 | 0.214 |
| J 2020.356.1. | 57.4 | (24) | (26) | (11) | 0.923 | 0.453 | 0.192 |
| J 2020.357.1. | 64.8 | (22) | (28) | (14) | 0.786 | 0.432 | 0.216 |
| J 2020.583.1. | 75.3 | (27) | (34) | 13.9 | 0.794 | 0.451 | 0.185 |
Description. Coiling involute, shell compressed, flanks parallel of small and medium sized specimens. Umbilicus shallow, umbilical wall oblique, umbilical shoulder rounded. No sculpture preserved except for some constrictions that are shallow, and cross the venter without interruption. Constrictions prorsiradiate on the upper flank, bending gently backward on the ventral shoulder and cross the venter perpendicularly with shallow constriction. There are 3 constrictions per half whorl. Suture not seen.
Remarks. Due to the fragmentary status and poor preservation of the specimens, important features (coiling, number of constrictions per whorl) are not seen, however the ventral part is well preserved that allows a positive separation from other Sowerbyceras species. As Sarti stated (1993, p. 53) discrimination of S. loryi from S. silenum is possible based on 5 criteria out of which one is present here: In the case of S. loryi constrictions cross the venter gently to form only a shallow band, while in the case of S. silenum there is a strong, upfolding constriction crossing the venter. The ventral constriction shown by the present specimens is typical of S. loryi (Sarti, 1993, Pl. 1, fig. 4). Apart from these difficulties the present specimens most resemble the specimens of Favre (1877, Pl. 1, fig. 14.a, .c) and Sapunov (1979, Pl. 4, fig. 6.a-b). First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Known from the lower Kimmeridgian Herbichi Zone to the upper Kimmeridgian Beckeri Zone: In addition loryi is occasionally reported from the lower Tithonian Hybonotum Zone (Sapunov, 1979, p. 37). Geographically there are records from the Upper Kimmeridgian Acanthicum Zone, Eastern Carpathians: Grigore, 2013; Ukraine Pieniny Klippen Belt. Upper Kimmeridgian Sardinia, Veneto, Italy: Sarti, 1993; Sicily, Tunisia: Pervinquičre, 1907; Eastern Carpathians, Bulgaria: Sapunov, 1979; Hungary, Romania: Grigore, 2011, Turkey: Alkaya, 1989. Sowerbyceras loryi is also reported from the Submediterranean Province; Crussol, France (Favre, 1877), Fribourg (Switzerland), Andalucia (Spain).
Suborder Lytoceratina Hyatt, 1889
Family Lytoceratidae Neumayr, 1875
Subfamily Lytoceratinae Neumayr, 1875
Genus Lytoceras Suess, 1865
Type species: Ammonites fimbriatus J. Sowerby, 1817
Lytoceras sp. ind.
Material. 13 internal moulds (J 2020.127.1, 171.1, 247.1, 298.1, 301.1, 302.1, 312.1, 313.1, 328.1, 329.1, 330.1, 352.1, 358.1).
Lytoceras polycyclum Neumayr, 1871b
(Fig. 5.E ![]() )
)
1871b. Lytoceras polycyclum nov. sp., Neumayr, p. 24.
1873. Lytoceras polycyclum Neumayr, Neumayr, p. 160 [20], Pl. 31, fig. 4.
1878. Lytoceras polycyclum, Neum., Gemmellaro, p. 179, Pl. 2, fig. 5.
1907. Lytoceras polycyclum Neumayr, Pervinquičre, p. 17, Pl. 1, fig. 4.A-B.
? 1961. Lytoceras polycyclum Neumayr, Rakus, p. 146, Pl. 11, fig. 1.
1966. Lytoceras polycyclum camertinum Canavari, Anđelković, p. 22, Pl. 2, figs. 1, 6, 8.
1973. Lytoceras polycyclum Neumayr, Preda, Pl. 4, figs. 3-4.
1979. Lytoceras polycyclum Neumayr, Sapunov, p. 39, Pl. 5, figs. 3-4.
1984. Lytoceras polycyclum (Neumayr), Rossi, p. 88, Pl. 31, fig. 5.
1996. Lytoceras cf. polycyclum Neumayr, Maisch, p. 53, Fig. 1.A-B.
2005. Lytoceras polycyclum (Neumayr), Boughdiri et al., p. 309. [in lit.]
2011. Lytoceras polycyclum Neumayr, Baudouin et al., Pl. 12, fig. 6.
2011. Lytoceras polycyclum Neumayr, Reháková et al., Pl. 7, fig. 4.
2013. Lytoceras polycyclum (Neumayr), Főzy & Scherzinger, p. 171, Pl. 4, figs. 1, 4.
Material. Nine internal moulds (J 2020.112.1, 130.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.125.1. | 112.4 | (43) | 46.4 | (38) | 0.927 | 0.413 | 0.338 |
| J 2020.134.1. | 85.6 | (34) | (33) | 33.1 | 1.030 | 0.385 | 0.387 |
| J 2020.142.1. | 74.7 | 24.6 | 27.8 | 27.6 | 0.885 | 0.372 | 0.369 |
| J 2020.177.1. | 89.4 | (28) | (31) | 39.2 | 0.903 | 0.347 | 0.438 |
| J 2020.268.1. | 64.9 | (25) | (24) | 26.1 | 1.042 | 0.370 | 0.402 |
| J 2020.289.1. | (45) | (15) | (14) | 22.0 | 1.071 | 0.311 | 0.489 |
| J 2020.345.1. | 44.9 | (12) | (16) | 19.5 | 0.750 | 0.356 | 0.434 |
Description. Small to medium sized, worn, and crushed conchs of typical lytoceratid (=serpenticone) coiling with wide and shallow umbilicus representing mostly the phragmocone. Cross section circular to subcircular. If subcircular, then flanks inflated, convex. Umbilical shoulder rounded, smooth. Flanks convex, rounded. Venter smooth. All specimens worn, partly dissolved, suture weathered and reduced. If early whorls are preserved, they are crushed and flattened, but three whorls seen. No sculpture, constriction noted.
Remarks. If preservation is good, or even original shell remains preserved, radial, straight ribs thoroughly persist on the conch as Grigore (2013, p. 91) discussed. Naturally, in our case there is no trace of ribbing. The specimen of Rakus (1961, Pl. 11, fig. 2.) is evolute: Inner whorls bear wide ribs but coiling is not of lytoceratid but rather serpenticone, which questions this determination. Specimen J 2020.177.1. is most similar to specimen J464 of Sapunov (1979, Pl. 5, fig. 4) and MM i5862 of Rossi (1984, Pl. 31, fig. 5) representing the stock of low expansion rate, while J 2020.125.1. resembles cru104 of Baudouin et al. (2011, Pl. 12, fig. 6) and represents the stock of higher expansion rate of the last whorl (from Wb/Wh=1 to Wb/Wh<1). First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Generally, it is reported from the Kimmeridgian, but occasionally also known from the Tithonian. Geographically it has a typical Tethysian distribution: Italy, Sicily, Tunisia (Pervinquičre, 1907; Boughdiri et al., 2005), Hungary, Serbia: Anđelković, 1966; Bulgaria: Sapunov, 1979; Eastern Carpathians (Ukraine and Romania: Preda, 1973), however it also inhabited peri-Tethysian territories (Submediterranean Province: France (Crussol: Baudouin et al., 2011), Austria, and southern Germany: Maisch, 1996).
Suborder Ammonitina Hyatt, 1889
Superfamily Haploceratoidea Zittel, 1884
Family Oppeliidae Douvillé, 1890
Subfamily Taramelliceratinae Spath, 1928
Genus Taramelliceras Del Campana, 1904
Subgenus Taramelliceras Del Campana, 1904
Type species: Ammonites trachinotus Oppel, 1862
Taramelliceras div. sp.
Material. 98 poorly preserved, fragmentary or complete internal moulds (J 2020.103.1, 128.1, 154.1, 157.1, 163.1-166.1, 182.1, 213.1, 216.1, 220.1, 222.1, 226.1, 244.1, 277.1, 284.1, 369.1-447.1, 569.2, and below).
Remarks. These fragments definitely represent different species of Taramelliceras, most possibly T. (T.) compsum, T. (T.) pugile, T. (T.) trachinotum, and T. (T.) pseudoflexuosum, however the poor preservation hinders specific assignments.
Taramelliceras (Taramelliceras) compsum (Oppel, 1863)
(Fig. 6.A1-A3 ![]() )
)
1863. Ammonites compsus Opp., Oppel, p. 215, Pl. 57, fig. 1.a-b.
1872. Oppelia compsa, Opp., Gemmellaro, p. 144, Pl. 1, fig. 3.
1878. Oppelia compsa Oppel, Herbich, p. 150 [132], Pl. 5.
1879. Oppelia compsa Oppel, Fontannes, p. 34, Pl. 5, fig. 1.
1909. Neumayria cfr. compsa Opp., Koch, p. 267. [in lit.]
1916. Oppelia compsa Opp., Jekelius, p. 270, Fig. 12.
1935. Taramelliceras cf. compsa Opp., Vadász, p. 62. [in lit.]
1955. Taramelliceras (Taramelliceras) compsum (Oppel), Hölder, p. 110, Pl. 19, fig. 22.
1959. Taramelliceras compsum Opp. var. crassa nov. var., Collignon, Pl. 112, fig. 414.
1966. Taramelliceras (Taramelliceras) compsum (Oppel), Anđelković, p. 27, Pl. 6, figs. 1-2; Pl. 7, fig. 4; Pl. 10, figs. 3, 7; Pl. 24, fig. 1; Pl. 26, fig. 1.
1973. Oppelia (Taramelliceras) compsa Oppel, Preda, Pl. 9, fig. 5.
1977. Taramelliceras (Taramelliceras) compsum compsum (Oppel), Sapunov, Pl. 4, fig. 1.
1978. Taramelliceras (Taramelliceras) compsum (Oppel), Olóriz, p. 83, Pl. 6, fig. 3.
1979. Taramelliceras (Taramelliceras) compsum compsum (Oppel), Sapunov, p. 48, Fig. 5/1, Pl. 9, figs. 1-2.
1986. Taramelliceras (Taramelliceras) compsum (Oppel), Sarti, p. 496, Pl. 2, fig. 1.A-B.
1989. Taramelliceras (Taramelliceras) compsum holbeini Oppel, Alkaya, p. 63, Pl. 1, fig. 2.
1994. Taramelliceras (Taramelliceras) compsum (Oppel), Wierzbowski, Pl. 4, fig. 12.
1998. Taramelliceras (Taramelliceras) compsum (Oppel), Howarth, p. 37, Pl. 1, fig. 8.
1999. Taramelliceras (T.) aff. T. (T.) compsum (Oppel), Fatmi & Zeiss, p. 43, Pl. 6, fig. 2.
2005. Taramelliceras sp. gr. compsum (Oppel), Boughdiri et al., Pl. 1, fig. 2.
2011. Taramelliceras compsum (Oppel), Baudouin et al., p. 626, Pl. 1, figs. 1-4; Pl. 2, figs. 3-10; Pl. 3, figs. 1-7; Pl. 4, figs. 1-8; Pl. 5, figs. 1-6; Pl. 6, figs. 1-6; Pl. 7, figs. 1-11; Pl. 8, figs. 1-10.
2011. Taramelliceras compsum (Oppel), Főzy et al., p. 418, Fig. 3.1-2.
2013. Taramelliceras compsum (Oppel), Főzy & Scherzinger, p. 173, Pl. 2, figs. 2, 6.
Material. 33, variously, but poorly preserved internal moulds (J 2078, J 2020.102.1, 141.1, 155.1, 319.1, 325.1, 338.1, 344.1, 349.1, 350.1, 351.1, 354.1, 360.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 702 | 76.4 | (23) | (39) | (11) | 0.590 | 0.510 | 0.144 |
| J 2020.224.1. | (57) | (14) | (27) | (12) | 0.518 | 0.474 | 0.210 |
| J 2020.243.1. | (51) | (12) | (21) | (14) | 0.571 | 0.411 | 0.274 |
| J 2020.250.1. | 71.0 | (17) | (35) | (16) | 0.486 | 0.493 | 0.225 |
| J 2020.278.1. | 118.5 | (28) | (55) | (22) | 0.509 | 0.464 | 0.185 |
| J 2020.287.1. | 93.7 | 26.3 | (49) | (17) | 0.537 | 0.523 | 0.181 |
| J 2020.292.1. | (72) | (20) | (38) | (11) | 0.523 | 0.528 | 0.153 |
| J 2020.306.1. | 72.5 | 14.0 | 37.5 | (10) | 0.373 | 0.517 | 0.138 |
| J 2020.307.1. | 72.6 | (20) | (39) | (12) | 0.513 | 0.537 | 0.165 |
| J 2020.308.1. | 72.4 | (19) | (40) | (11) | 0.475 | 0.552 | 0.152 |
| J 2020.309.1. | 75.2 | (19) | (38) | (16) | 0.500 | 0.505 | 0.213 |
| J 2020.310.1. | 71.9 | (21) | (38) | (15) | 0.553 | 0.529 | 0.209 |
| J 2020.311.1. | (74) | (17) | (36) | (16) | 0.472 | 0.486 | 0.216 |
| J 2020.318.1. | 88.0 | 22.2 | (45) | (18) | 0.493 | 0.511 | 0.204 |
| J 2020.320.1. | 132.0 | (30) | (63) | (23) | 0.476 | 0.477 | 0.174 |
| J 2020.321.1. | 114.8 | 31.4 | (52) | (19) | 0.604 | 0.453 | 0.165 |
| J 2020.323.1. | 92.2 | (27) | (51) | (17) | 0.529 | 0.553 | 0.184 |
| J 2020.326.1. | 101.0 | (21) | (50) | (12) | 0.420 | 0.495 | 0.119 |
| J 2020.336.1. | 61.0 | 16.8 | (33) | (10) | 0.509 | 0.541 | 0.164 |
| J 2020.337.1. | 80.5 | (22) | (38) | (16) | 0.579 | 0.472 | 0.199 |
| J 2020.341.1. | (75) | (20) | (33) | (18) | 0.606 | 0.440 | 0.240 |
Description. Small to medium sized phragmocone conchs. Umbilicus small, shallow. Umbilical wall short, oblique. Umbilical shoulder rounded. Flanks convex, converging, high oval. Ventral shoulder rounded. Venter rounded, smooth. From the midflank equal, prorsiradiate ribs travel toward the ventral shoulder. These ribs originate from the umbilical shoulder, however due to poor preservation it is rarely seen. On the early whorls, ribs end in elongated tubercles at the ventral shoulder, while on later whorls every third to fifth one ended in tubercles; majority of the primaries end on the ventral shoulder. On early whorls there is a row of fine tubercles on the mid-siphonal line that later diminish. Above 25 mm whorl height, the mid-siphonal line is devoid of tubercles, but primaries cross the venter, joining corresponding primaries. On the last half whorl there are 10 tubercles on the ventral shoulder. Intraspecific variation is expressed in the changing number of tubercles, their size, and the number of primary ribs between them. Sutural elements present but aperture not seen.
Remarks. This highly variable species was excellently discussed by Baudouin et al. (2011). However, when the preservation is poor and/or fragmentary, there are difficulties in discriminating compsum from T. (T.) pugile. Notwithstanding, based on statistically significant material, some general guidelines could be outlined as Baudouin et al. (2011, p. 633) demonstrated. Therefore, it is suggested here that the separation of poorly preserved specimens is reliable. The present specimens exhibit high intraspecific variation.
Stratigraphic and geographic distribution. Taramelliceras compsum is reported from the lower Kimmeridgian (Herbichi Zone) and upper Kimmeridgian (Acanthicum Zone). Outside the Western Tethys it occurs in younger Kimmeridgian strata. Geographically it has a wide distribution in the Western Tethys (Italy, Sicily: Gemmellaro, 1872; Tunisia: Boughdiri et al., 2005; Hungary, Eastern Carpathians: Preda, 1973; Bulgaria: Sapunov, 1977; Turkey: Alkaya, 1989; Yemen: Howarth, 1998) and the adjacent Submediterranean Province (Betic Cordilleras Spain: Olóriz, 1978; Crussol, Ardéche France, South Germany). It is also known from the northeastern Tethysian margin (Baluchistan: Fatmi & Zeiss, 1999) and the Indo-Malagasy Province in the southern Tethys (Madagascar: Collignon, 1959).
|
Figure 6: Upper Kimmeridgian-lower Tithonian ammonoids from Zengővárkony, Mecsek Mountains, Hungary. A. Taramelliceras (Taramelliceras) compsum (Oppel, 1863), A1. specimen J 2078; A2. specimen J 2020.141.1; A3-A5: specimen J 702. A3, A5. lateral view; A4. cross-section. B. Taramelliceras (Taramelliceras) pugile (Neumayr,
1871b), specimen J 2020.133.1. B1. lateral view; B2. ventral view., specimen J 2020.225.1. C. Taramelliceras (Taramelliceras) cf. trachinotum (Oppel, 1863), specimen J 2020.190.1; oblique ventral view. D. Taramelliceras (Metahaploceras) strombecki (Oppel, 1857), specimen J 2020.223.1. D1. lateral view; D2. apertural view. E. Hemihaploceras nobile (Neumayr,
1873), specimen J 2020.577.1. E1. lateral view; E2. cross-section of specimen J 2020.582.1. F. Streblites tenuilobatus (Oppel, 1863), specimen J 2020.115.1. F1. lateral view; F2. cross-section of specimen J 2020.114.1. Scale bars indicate 1 cm. |
Taramelliceras (Taramelliceras) pugile (Neumayr, 1871b)
(Fig. 6.B1-B2 ![]() )
)
1871b. Oppelia pugilis nov. sp., Neumayr, p. 24.
1871a. Oppelia pugilis Neumayr, Neumayr, p. 167 [27], Pl. 32, figs. 1-2.
1872. Oppelia pugilis, Neum., Gemmellaro, p. 143, Pl. 1, fig. 2.
1879. Oppelia pugilis Neumayr, Fontannes, p. 45, Pl. 7, figs. 1-2.
1959. Taramelliceras pugile (Neumayr), Berckhemer & Hölder, p. 76, Figs. 46-47, Pl. 16, fig. 77; Pl. 20, fig. 100.
1966. Taramelliceras (Taramelliceras) pugile (Neumayr), Anđelković, p. 30, Fig. 23; Pl. 5, fig. 7; Pl. 24, fig. 4.
1973. Oppelia (Taramelliceras) pugilis Neumayr, Preda, Pl. 7, fig. 2.
1977. Taramelliceras (Taramelliceras) pugile (Neumayr), Sapunov, Pl. 5, fig. 3.A-B.
1978. Taramelliceras (Taramelliceras) pugile pugile (Neumayr), Olóriz, p. 94, Pl. 7, fig. 2.A-B.
1979. Taramelliceras (Taramelliceras) pugile (Neumayr), Sapunov, p. 51, Pl. 10, figs. 5-7; Pl. 11, figs. 1-2.
1986. Taramelliceras (Taramelliceras) pugile (Neumayr), Sarti, p. 494, Pl. 2, fig. 3.A-B.
1993b. Taramelliceras pugile (Neum.), Főzy, p. 198. [in lit.]
1993. Taramelliceras (Taramelliceras) pugile pugile (Neumayr), Sarti, p. 66, Pl. 4, fig. 1.
1995. Taramelliceras pugile (Neumayr), Főzy, p. 135, Pl. 21, fig. 2.
1997. Taramelliceras (Taramelliceras) pugile (Neumayr), Benzaggagh & Atrops, Pl. 3, fig. 2.
1999. Taramelliceras (Hemihaploceras) pugile (Neumayr), Fatmi & Zeiss, p. 40, Pl. 7, fig. 1.
2004. Taramelliceras pugile pugiloides (Canavari), Marino et al., p. 368, Pl. 2, fig. 5.
2011. Taramelliceras (Taramelliceras) pugile pugiloide Canavari, Reháková et al., Pl. 7, fig. 1.
Material. 11 poorly preserved internal moulds (J 2020.221.3, 295.1, 324.2, 339.1, 339.2, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.133.1. | 112.8 | (25) | (54) | 19.4 | 0.463 | 0.479 | 0.172 |
| J 2020.225.1. | 106.0 | (27) | (47) | (28) | 0.574 | 0.443 | 0.264 |
| J 2020.249.1. | 110.6 | (23) | (41) | (31) | 0.561 | 0.371 | 0.280 |
| J 2020.317.1. | 114.0 | (26) | 48.1 | (28) | 0.540 | 0.422 | 0.245 |
| J 2020.340.1. | 97.2 | (25) | 51.6 | (18) | 0.484 | 0.531 | 0.185 |
| J 2020.343.1. | 90.6 | (19) | (36) | (26) | 0.528 | 0.397 | 0.287 |
Description. Medium to big sized conchs (to D=115 mm) representing phragmocone and body chamber. Conchs crushed, dissolved, fragmented. Umbilicus wide, shallow. Umbilical wall short, rounded. Flanks parallel or slightly converging. Ventral shoulder rounded. Venter rounded, smooth, sometimes forming a gentle and short keel. At the ventral shoulders corresponding rows of tubercles rise; there are 10 tubercles on the last half whorl. On the mid-siphonal line, in between the ventral shoulder row of tubercles, another row of tubercles rises. The height and length of these tubercles are equal or smaller than the tubercles on the ventral shoulder. No other sculpture. Suture, aperture not seen.
Remarks. Regarding the separation of specimens belong to T. (T.) compsum from the present species, see Baudouin et al. (2011). Although the present specimens are poorly preserved, important features (e.g., umbilicus, external tubercles, keel) are well seen, therefore specific determination is justified. Specimen J 2020.133.1. is closest to specimen F.G7.10.1 of Olóriz (1978, Pl. 7, fig. 2.a-b); specimen J 2020.249.1 resembles specimen no. 168 of Sarti (1986, Pl. 2, fig. 3.a-b), and both represents the stock with robust mid-siphonal and ventral shoulder tubercles.
Stratigraphic and geographic distribution. Taramelliceras pugile is reported from the upper Kimmeridgian with wide geographic distribution in the western Tethys (Italy, Sicily: Gemmellaro, 1872; Hungary, Eastern Carpathians: Preda, 1973; Serbia: Anđelković, 1966; Bulgaria: Sapunov, 1977; Tunisia: Benzaggagh & Atrops, 1997), peri-Tethysian territories (Submediterranean: Betic Cordilleras Spain: Olóriz, 1978; Crussol France: Fontannes, 1879; southern Germany), and northern Tethys (Baluchistan: Fatmi & Zeiss, 1999).
Taramelliceras (Taramelliceras) cf. trachinotum (Oppel, 1863)
(Fig. 6.C ![]() )
)
1863. Ammonites trachinotus Opp., Oppel, p. 214, Pl. 56, fig. 4.a-b.
1870. Oppelia trachynota Opp., Zittel, p. 70, Pl. 5, fig. 3.
1875. Oppelia trachynota Oppel, Waagen, p. 54, Pl. 10, fig. 6.
1877. Ammonites (Oppelia) trachynotus Oppel, Favre, p. 34, Pl. 3, fig. 2.
1878. Oppelia trachynota Oppel, Herbich, p. 154 [136], Pl. 3, fig. 2.
1879. Oppelia trachynota Oppel, Fontannes, p. 36, Pl. 5, fig. 2.
1929. Oppelia trachynota Oppel, Wegele, p. 19 (113), Pl. 26, figs. 8-9.
1935. Taramelliceras cf. trachynota Opp., Vadász, p. 62. [in lit.]
1955. Taramelliceras (Taramelliceras) trachinotum (Oppel), Hölder, p. 102, Pl. 18, fig. 21.
1959. Taramelliceras trachinotum Opp., Collignon, Pl. 114, fig. 426.
1966. Taramelliceras trachinotum (Oppel), Anđelković, p. 31, Pl. 4, fig. 2; Pl. 7, fig. 1.
1973. Oppelia (Taramelliceras) trachynota Oppel, Preda, Pl. 15, fig. 3.
1976. Taramelliceras aff. trachinotum (Opp.), Fülöp, p. 190, Pl. 33, fig. 2.
1976. T. (Taramelliceras) trachynotum (Oppel), Lillo Bevia, p. 463, Pl. 2, figs. 5-6.
1977. Taramelliceras (Taramelliceras) trachinotum (Oppel), Sapunov, Pl. 3, fig. 1.
1978. Taramelliceras (Taramelliceras) trachinotum Oppel, Olóriz, p. 80, Pl. 9, figs. 2.A-B.
1979. Taramelliceras (Taramelliceras) trachinotum (Oppel), Sapunov, p. 46, Pl. 8, fig. 1.
1984. Taramelliceras (Taramelliceras) trachynotum (Oppel), Verma & Westermann, p. 35, Pl. 3, fig. 2.A-B.
1993. Taramelliceras (Taramelliceras) trachinotum (Oppel), Sarti, p. 59, Pl. 3, fig. 1.
1997. Taramelliceras (Taramelliceras) trachinotum (Oppel), Benzaggagh & Atrops, Pl. 2, figs. 3-4.
2013. Taramelliceras cf. trachinotum (Oppel), Főzy & Scherzinger, p. 172, Pl. 14, fig. 1.
Material. 11 whorl fragments (J 2020.190.1, 201.1, 215.1, 246.1, 275.1, 291.1, 322.1, 324.2, 342.1, 355.1, 361.1) of internal moulds.
Dimensions. Due to poor preservation dimensions are not given.
Description. All fragments represent the ventral region from the midflank to the venter of body chambers. Cross section trapezoidal, flanks converging. Ventral shoulder gently rounded. On the ventral shoulder massive spines rise correspondingly. In the middle of the venter, along the mid-siphonal line, another row of tubercles rises. Length, height, and shape of the ventrolateral and mid-siphonal tubercles are equal. Apart from tubercles the conch is smooth, and unsculptured. Suture not seen.
Remarks. The holotype (Oppel, 1863, Pl. 56, fig. 4.a-b) has more frequent mid-siphonal tubercles and fine ribs than the present specimens. Although there are fine ribs on the flanks of the present species, if the specimens are worn, these ribs are not preserved (cf. Anđelković, 1997, Pl. 4, fig. 2). The present specimens are comparable to the specimen KNMI-MA31 of Verma and Westermann (1984, Pl. 3, fig. 2.b), the only notable difference is that the latter has stronger tubercles. The present specimens are also close to specimen F.G14.5.16 of Olóriz (1978, Pl. 9, fig. 2.b), however the latter is more inflated and presents fine ribs on the mould. Notwithstanding, the present specimens do not show ribs, most probably due to the poor preservation.
Stratigraphic and geographic distribution. Taramelliceras trachinotum is reported from the upper Kimmeridgian. Geographically widespread from the Tethys (Italy: Sarti, 1993; Tunisia: Benzaggagh & Atrops, 1997; Hungary, Serbia: Anđelković, 1966; Bulgaria: Sapunov, 1979; Eastern Carpathians: Preda, 1973) and from peri-Tethysian territories (Submediterranean: Betic Cordilleras, Alicante Spain: Olóriz, 1978; Crussol France: Fontannes, 1879; southern Germany). It is also known from the Indo-Malagasy Province of the southern Tethys (Kenya: Verma & Westermann, 1984; and Madagascar: Collignon, 1959).
Subgenus Metahaploceras Spath, 1925
Type species: Ammonites lingulatus nudus Quenstedt, 1849
Taramelliceras (Metahaploceras) aff. strombecki (Oppel, 1857)
(Fig. 6.D1-D2 ![]() )
)
1849. Ammonites lingulatus nudus, Quenstedt, p. 130, Pl. 9, fig. 8.
1857. Ammonites Strombecki n. sp., Oppel, p. 687.
1878. Oppelia Strombecki Oppel, Herbich, p. 148 [130], Pl. 4, fig. 1.
1907. Neumayria cf. Strombecki Oppel, Pervinquičre, p. 19. [in lit.]
1955. Taramelliceras (Metahaploceras) strombecki (Oppel), Hölder, p. 135, Figs. 157-161.
1973. Oppelia (Taramelliceras) strombecki Oppel, Preda, Pl. 6, fig. 1.
1978. Taramelliceras (Metahaploceras) strombecki (Oppel), Olóriz, p. 110, Pl. 10, fig. 1.
1993. Taramelliceras (Metahaploceras) strombecki (Oppel), Sarti, p. 58, Pl. 1, fig. 6.A-B.
1994. Taramelliceras (Metahaploceras) strombecki (Opp.), Schlegelmilch, p. 40, Pl. 11, fig. 3.
2003. Taramelliceras (Metahaploceras) cf. strombecki (Oppel), Lukeneder et al., p. 226, Pl. 1, figs. 7-8.
2011. Taramelliceras strombecki (Oppel), Főzy et al., p. 417, Fig. 2.3-2.4.
2013. Metahaploceras strombecki (Oppel), Főzy & Scherzinger, p. 174, Pl. 2, fig. 7.
Material. Three poorly preserved whorl fragments and internal moulds (J 2020.156.1, 158.1, 223.1).
Dimensions. Due to fragmentary status, dimensions are not given.
Description. These whorl fragments most probably belonged to large-sized (?adult) ammonites and represent body chambers. Umbilical region not preserved. Middle and upper flank sculptured with fine, s-shaped ribs, occasionally ending in fine tubercles at the ventral shoulder. Sometimes the fine ribs cross the venter continuously forming a forwardly convex gentle bend. Specimen J 2020.223.1 most similar to Herbich's specimen (1878, Pl. 4, fig. 1) with smooth flank, fine ribs and occasional fine and short tubercles. Cross section is compressed, oval, venter rounded. Suture not seen.
Remarks. Although coiling, umbilical region and midflank area are uncertain, the smooth upper flank, fine sculpture and cross section definitely associate this species with the strombecki species. T. (M.) nodosiusculum (Fontannes, 1879) is also comparable however the latter has more frequent and less sinuous ribbing (cf. Olóriz, 1978, p. 115). The poor preservaton justifies the uncertain assignment. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Taramelliceras strombecki is reported from the Strombecki Zone, lower Kimmeridgian, however it occurs in younger strata (e.g., Herbichi Zone: Főzy et al., 2011). Geographically it is typical of the Tethysian (Mediterranean Province: Italy: Sarti, 1993; Hungary, Eastern Carpathians: Preda, 1973; Algeria: Bouchemla et al., 2020; Tunisia: Pervinquičre, 1907), and peri-Tethysian settings (Submediterranean Province: Betic Cordilleras Spain: Olóriz, 1978; southern Germany: Schlegelmilch, 1994).
Genus Hemihaploceras Spath, 1925
Type species: Oppelia nobilis Neumayr, 1873
Hemihaploceras nobile (Neumayr, 1873)
(Fig. 6.E1-E2 ![]() )
)
1873. Oppelia nobilis nov. sp., Neumayr, p. 167 [27], Pl. 32, Figs. 3-4.
1877. Ammonites (Oppelia) nobilis Neumayr, Favre, p. 36, Pl. 2, fig. 14.
1879. Oppelia nobilis Neumayr, Fontannes, p. 46, Pl. 3, fig. 4.
1935. Taramelliceras nobilis Neum., Vadász, p. 62. [in lit.]
1959. Taramelliceras (Hemihaploceras) nobile (Neumayr), Berckhemer & Hölder, p. 77, Pl. 16, fig. 74.
1966. Hemihaploceras nobilis (Neumayr), Anđelković, p. 42, Pl. 4, fig. 3; Pl. 6, fig. 6.
1973. Oppelia (Taramelliceras) nobilis Neumayr, Preda, Pl. 17, fig. 14.
1977. Hemihaploceras nobile (Neumayr), Sapunov, Pl. 6, fig. 4.
1978. Hemihaploceras nobile (Neumayr), Olóriz, p. 117, Pl. 7, fig. 5.
1979. Hemihaploceras nobile (Neumayr), Sapunov, p. 59, Pl. 13, figs. 1-4.
1989. Hemihaploceras nobile (Neumayr), Főzy, Pl. 4, fig. 1.
1995. Hemihaploceras nobile (Neumayr), Főzy, p. 136, Pl. 21, fig. 1.
1997. Taramelliceras (Hemihaploceras) nobile (Neumayr), Benzaggagh & Atrops, Pl. 3, fig. 1.
1999. Hemihaploceras sp. cf. nobile (Neumayr), Caracuel & Olóriz, Fig. 5.3.
2010. Hemihaploceras (Hemihaploceras) nobile (Neumayr), Mandl et al., p. 94.
2018. Hemihaploceras nobile (Neumayr), Metodiev, p. 98, Fig. 1.c-d.
Material. 12 poorly preserved internal moulds of whorl fragments (J 2020.147.1, 362.1, 577.1-582.1, 585.1-588.1).
Dimensions. Due to fragmentary status, dimensions are not given.
Description. Worn, dissolved fragments of internal mould. Umbilicus, inner whorls lost. Flanks convex, converging. Cross section compressed, oval, through the ventral shoulder tubercles, rectangular. Venter flat, smooth, rounded, no tubercles on the mid-siphonal ridge. On the ventral shoulder strong, wide, paired tubercles rise that correspond to each other. Below the tubercles on the upper flank, fine, wide, shallow, s-shaped double ribs rise and travel toward the lower flank. Suture, aperture not seen.
Remarks. Lack of ventral tubercles on mid-siphonal ridge, characteristic sculpture, cross section, and coiling provide an unmistakeable and easy-to-recognize species (Olóriz, 1978 p. 119). Its intraspecific variety is restricted to coiling, and strength of sculpture. Specimen J 2020.577.1 is most similar to the holotype of Neumayr (1873, Pl. 32, fig. 3.a) and represents the more evolute and strongly sculptured morphs of the species. Specimen J 2020.581.1 most similar to specimen J182 of Sapunov (1979, Pl. 13, fig. 3) and represents the faintly sculptured stock with well-developed tubercles.
Stratigraphic and geographic distribution. Hemihaploceras nobile is reported from the upper Kimmeridgian Cavouri and Beckeri zones from the northern and southern margins of the western Tethys.
Subfamily Streblitinae Spath, 1925
Genus Streblites Hyatt, 1900
Type species: Ammonites tenuilobatus Oppel, 1857
Streblites tenuilobatus (Oppel, 1857)
(Fig. 6.F1-F2 ![]() )
)
1857. Ammonites tenuilobatus, n. sp., Oppel, p. 686.
1863. Ammonites tenuilobatus Opp., Oppel, p. 160, Pl. 50, fig. 1.a-b.
1867. Ammonites tenuilobatus Oppel, Pictet, p. 235, Pl. 36, fig. 10.
1876. Ammonites tenuilobatus Oppel, Dumortier & Fontannes, p. 52, Pl. 7, figs. 1-2.
1877. Ammonites (Oppelia) tenuilobatus Oppel, Favre, p. 26, Pl. 2, fig. 5.
1878. Oppelia tenuilobata, Opp., Gemmellaro, p. 186, Pl. 2, fig. 6.
1879. Oppelia tenuilobata Oppel, Fontannes, p. 22, Pl. 3, figs. 5-6.
1907. Oppelia tenuilobata Opp., Toula, p. 22, Pl. 5, fig. 6.
1929. Streblites tenuilobatus Oppel, Wegele, p. 11 [105], Pl. 25, figs. 7-9.
1973. Oppelia (Streblites) tenuilobatus Oppel, Preda, Pl. 10, fig. 5.
1977. Streblites tenuilobatus (Oppel), Ziegler, Pl. 3, fig. 6.
1978. Streblites tenuilobatus (Oppel), Olóriz, p. 44, Pl. 4, figs. 2-3.
1979. Streblites tenuilobatus (Oppel), Sapunov, p. 62, Pl. 14, fig. 1.A-B.
1988. Streblites tenuilobatus frotho (Oppel), Abdulkasumzade, p. 105, Pl. 12, fig. 8.
2000. Streblites tenuilobatus (Oppel), Grigore, Pl. 1, fig. 4.
2000. Streblites tenuilobatus (Oppel), Gygi, p. 73, Pl. 14, fig. 4.
2011. Streblites cf. tenuilobatus (Oppel), Főzy et al., p. 418, Fig. 3.3.
2018. Streblites tenuilobatus (Oppel), Grigore, p. 16, Pl. 1, figs. 8, 11-12.
Material. Two relatively well-preserved internal moulds.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.114.1. | 86.4 | (24) | 49.7 | 8.2 | 0.483 | 0.575 | 0.095 |
| J 2020.115.1. | 83.6 | 23 | 48.2 | 7.3 | 0.477 | 0.576 | 0.087 |
Description. Highly involute, strongly compressed shell. Phragmocone cross section is oxyconic (cross section of living chamber is more rounded, with inflated flank as Olóriz (1978, p. 54) demonstrated). Umbilicus small, shallow. Umbilical wall vertical or oblique. Umbilical shoulder slightly rounded. At the umbilical shoulder, fine, radial ribs rise that become shortly prorsiradiate and disappear at the lower third of the midflank. There are 9 ribs on half whorl. Upper flank and venter dissolved; no sculpture preserved. Venter acute in cross section, and slightly rounded.
Remarks. The preservation is poor, therefore the lateral tubercles and the fine secondary ribs on the upper flank are not preserved. The present specimens are most similar to material illustrated by Dumortier and Fontannes (1876, Pl. 7, fig. 2) and Olóriz (1978, Pl. 4, fig. 3). Although the sculpture is poorly preserved, the U/D ratio is decisive for the specific designation as Olóriz (1978, p. 47) demonstrated. In the case of S. tenuilobatus s. str. it is 8-10%. Our specimens fall into this range (8.7% and 9.5%). Due to the poor preservation, any ventral fine ribbing is uncertain. The year of erecting this species by Oppel varies in literature. Many authors (e.g., Dumortier & Fontannes, 1876; Gygi, 2000; Pictet, 1867; Sapunov, 1979) refer it to 1858, others even to 1863 (e.g., Olóriz, 1978; Főzy et al., 2011). Correctly it is 1857 when Oppel published his second volume of Juraformation ('Jahreshefte' 13), pp. 439-694. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Streblites tenuilobatus is a typical faunal element of the Kimmeridgian Herbichi Zone with stray individuals from the Acanthicum Zone. Geographically widespread in the western Tethys (Mediterranean: Sicily, Hungary, Eastern Carpathians, Bulgaria; and Submediterranean: Betic Cordilleras Spain, Crussol France, southern Germany; Provinces), also along the northern margin of the Tethys (Azerbaijan).
Superfamily Perisphinctoidea Steinmann, 1890
Family Perisphinctidae Steinmann, 1890
Perisphinctidae gen. et sp. ind.
Material. 11 poorly preserved internal moulds (J 2020.174.1, 176.1, 239.1, 248.1, 557.1-562.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.237.1. | 86.4 | (24) | (23) | (50) | 1.043 | 0.266 | 0.579 |
Subfamily Passendorferiinae Meléndez, 1989
Genus Praesimoceras Sarti, 1990 [emend. Gygi, 2003, p. 91]
Type species: Ammonites nodulatus (Quenstedt, 1888)
Praesimoceras cf. herbichi (Hauer, 1866)
(Fig. 7.A1-A2 ![]() )
)
1866. Ammonites Herbichi n. sp., Hauer, p. 194 [24]
1873. Simoceras Herbichi v. Hauer, Neumayr, p. 186 [46], Pl. 40, figs. 1-2.
1877. Ammonites (Simoceras) Herbichi v. Hauer, Favre, p. 55, Pl. 6, fig. 2; Pl. 7, fig. 3.
1879. Simoceras herbichi von Hauer, Fontannes, p. 83, Pl. 11, fig. 11.
1959. Nebrodites (Mesosimoceras) herbichi (von Hauer), Ziegler, Pl. 1, fig. 21.
1966. Pseudosimoceras herbichi (v. Hauer), Anđelković, p. 96, Pl. 10, fig. 2.
1973. Pseudosimoceras herbichi (Hauer), Preda, Pl. 16, fig. 2.
1978. Nebrodites (Mesosimoceras) herbichi (v. Hauer), Olóriz, p. 182, Fig. 2.A-B, Pl. 16, fig. 1.
1999. Nebrodites (Mesosimoceras) cf. N. (M.) herbichi (von Hauer), Fatmi & Zeiss, p. 50, Pl. 37, fig. 3.
2000. Presimoceras herbichi (Hauer), Grigore, Pl. 2, fig. 1.
2003. Praesimoceras cf. herbichi (von Hauer), Gygi, p. 92, Fig. 100.
2003. Presimoceras cf. herbichi (Hauer), Rasser et al., Pl. 1, fig. 9-11.
2010. Presimoceras herbichi (von Hauer), Grigore, p. 287, Pl. 1, figs. 2, 4, 6-7.
2010. Nebrodites (Mesosimoceras) herbichi (von Hauer), Mandl et al., p. 95, Pl. 14, fig. 18.
2011. Presimoceras herbichi (Hauer), Petti et al., p. 166, Pl. 2, fig. 2.A-B.
2013. Presimoceras cf. herbichi (von Hauer), Főzy & Scherzinger, p. 177, Pl. 3, figs. 2, 5-6.
Material. Two poorly preserved, fragmented internal moulds (J 2020.109.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.117.1. | (175) | 28.8 | 42.3 | (100) | 0.681 | 0.242 | 0.571 |
Description. Whorl fragments of big sized conch representing body chamber. Strongly evolute, serpenticone coiling. Early whorls lost. Umbilical wall rounded, smooth, oblique. Flanks converging. Ventral shoulder rounded, venter and its mid-siphonal region eroded and dissolved; not well preserved. Cross section high subtrapezoidal. At the umbilical shoulder strong, widely spaced, radial or slightly prorsiradiate ribs occur. There are 22 ribs on specimen J 2020.117.1, which represents just less than a half whorl. Intercalatory ribs and constrictions may occur. Neither suture nor aperture seen.
Remarks. The ratios of conch of the present specimen fits remarkably well with the figured specimen of Neumayr (1873) (U/D= 0.57 here, 0.61 by Neumayr; Wh/D= 0.24 here, 0.22 for Neumayr, at diameter 175 mm here, and 140 mm in Neumayr (1873, p. 186 [46]). Number of ribs varies as the diameter increases: At greater diameters, the number of ribs per half whorl decreases. This is well documented by the authors who have given detailed palaeontological descriptions or well-described accounts of specimens (Neumayr, 1873; Ziegler, 1959; Preda, 1973; Olóriz, 1978; Grigore, 2010) and varies between 12-33 with gradual decrease as the diameter increases. The poor preservation (loss of internal whorls, presence of only less than a half whorl with dissolved ventral region) creates pitfalls of assigning these collected fragments into any distinct species. The lateral view of the present specimens may refer to herbichi but also P. teres and P. planulascinctum are comparable, however their ventral region cannot be compared. The present specimens are tentatively assigned to herbichi based on whorl section and ribbing. Fragments collected are considered to represent middle whorls of this perisphinctid. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Praesimoceras herbichi is reported from the upper part of the Strombecki and the Herbichi zones of the lower Kimmeridgian with a wide geographic distribution in the western Tethysian and peri-Tethysian (Submediterranean Province) territories and also from the northern Tethys (Baluchistan).
|
Figure 7: Upper Kimmeridgian-lower Tithonian ammonoids from Zengővárkony, Mecsek Mountains, Hungary. A. Praesimoceras herbichi (Hauer,
1866), specimen J 2020.117.1. A1. lateral view; A2. cross-section. B. Mesosimoceras cavouri (Gemmellaro,
1872), specimen J 2020.290.1. B1. lateral view; B2. cross-section. C. Ataxioceratidae sp., specimen J 2020.105.1. C1. lateral view; C2. apertural view; specimen J 2020.107.1. C3. lateral view; C4. cross-section., C5. specimen J 2020.572.1. D. Crussoliceras sp. ind., specimen J 2020.269.1. D1. lateral view; D2. cross-section. E. Progeronia sp. ind., specimen J 2020.368.1. E1. lateral view; E2. cross-section of specimen J 2020.571.1. |
Genus Mesosimoceras Spath, 1925
Type species. Simoceras Cavouri Gemmellaro, 1872
Mesosimoceras cavouri (Gemmellaro, 1872)
(Fig. 7.B1-B2 ![]() )
)
1872. Simoceras Cavouri Gemm., Gemmellaro, p. 151, Pl. 2, figs. 3-4.
1966. Mesosimoceras covouri (Gemmellaro), Anđelković, p. 98, Pl. 8, fig. 2; Pl. 11, fig. 7; Pl. 20, fig. 5.
1978. Nebrodites (Mesosimoceras) cavouri (Gemmellaro), Olóriz, p. 178, Pl. 15, fig. 5.
1979. Nebrodites (Mesosimoceras) cavouri (Gemmellaro), Sapunov, p. 118, Pl. 32, fig. 2.
1989. Nebrodites (Mesosimoceras) cavouri (Gemmellaro), Főzy, p. 152, Pl. 1, fig. 3.
1990. Mesosimoceras cavouri (Gemmellaro), Sarti, p. 47, Fig. 6.A.
2000. Mesosimoceras cavouri (Gemmellaro), Grigore, Pl. 3, fig. 5.
2011. Nebrodites cavouri (Gemmellaro), Főzy et al., p. 419, Fig. 4.
2013. Mesosimoceras cavouri (Gemmellaro), Főzy & Scherzinger, p. 176, Pl. 3, fig. 8.
2016. Mesosimoceras cavouri (Gemmellaro), Scherzinger et al., p. 146, Fig. 2.
Material. One incomplete, fairly preserved whorl fragment.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.290.1. | n.d. | 14.5 | 20.4 | n.d. | 0.711 | n.d. | n.d. |
Description. Fairly preserved whorl fragment. Based on the fragment coiling very much evolute (=serpenticone). Umbilicus not seen. Umbilical wall very short, rounded. Simple, strong, radial ribs rise at umbilical wall, cross the umbilical shoulder and travel radially through flanks. Venter not preserved. Cross section compressed, oval, flanks convex, converging. Suture, aperture not preserved.
Remarks. Although the collected fragment represents only a short whorl segment, the cross section, simple ribbing and coiling allows assignation to the characteristic cavouri biospecies. There are only two species assigned to this genus (Sarti, 1990): M. cavouri and M. risgoviensis. Based on the differential analysis of Sarti (1990, p. 41) the present specimen is tentatively assigned to the former on the grounds that the collected specimen shows hardly overlapping coils, no constrictions, nor tubercles on the ventrolateral shoulder. Whereas the collected specimen differs from risgoviensis by fainter sculpture and simple ribbing. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. FAD in the lower part of Kimmeridgian Cavouri Zone; it is known to the end of Beckeri Zone in the Mediterranean Tethys, with stray occurrences in the Submediterranean Province.
Family Ataxioceratidae Buckmann, 1921
Subfamily Ataxioceratinae Buckmann, 1921
Ataxioceratidae div. sp.
(Fig. 7.C1-C5 ![]() )
)
Material. 118 poorly preserved internal moulds, mainly whorl fragments (J 2020.88.1, 92.1, 94.1, 99.1, 100.1, 104.1, 107.1, 108.1, 113.1, 121.1-124.1, 126.1, 135.1, 137.1, 139.1, 140.1, 173.1, 175.1, 179.1, 180.1, 181.1, 183.1, 187.1, 188.1, 189.1, 195.1-197.1, 199.1, 200.1, 206.1, 208.1, 209.1, 210.1, 218.1, 234.1, 235.1, 238.1, 240.1, 241.1, 245.1, 303.1.-305.1, 497.1-556.1, 584.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.118.1. | 103.3 | (28) | (32) | (44) | 0.875 | 0.310 | 0.426 |
| J 2020.151.1. | (95) | (30) | (31) | (47) | 0.968 | 0.326 | 0.495 |
| J 2020.178.1. | (71) | (21) | 21.3 | 30.4 | 0.986 | 0.300 | 0.428 |
| J 2020.194.1. | (68) | 20.6 | (22) | (33) | 0.936 | 0.323 | 0.485 |
| J 2020.572.1. | (91) | (29) | (30) | (41) | 0.967 | 0.330 | 0.451 |
| J 2020.573.1. | (76) | (22) | (26) | 33.4 | 0.846 | 0.342 | 0.439 |
| J 2020.574.1. | (107) | (23) | (31) | (48) | 0.742 | 0.290 | 0.448 |
| J 2020.575.1. | (110) | (30) | (34) | 44.6 | 0.882 | 0.309 | 0.405 |
| J 2020.576.1. | 73.4 | 23.2 | (19) | 39.6 | 1.221 | 0.259 | 0.539 |
Description. Poorly preserved fragments of conchs of body chamber and phragmocone. In better preserved specimens, umbilicus wide, shallow, 2-3 whorls seen if present. Whorl section circular, subcircular to oval. Finer or stronger, radial to prorsiradiate ribs rise at umbilical shoulder being prorsiradiate on flanks and cross the venter continuously. Ribs bifurcate on the upper flanks. Single ribs, constrictions and virgatotomous ribs may be present. Suture, aperture not seen.
Remarks. The various collected whorl fragments definitely belong to many different Ataxioceratidae species and genera. Ataxioceratid ammonites are abundant and important biogeographical and biostratigraphical indicators in the Submediterranean Province of the Tethys and adjacent areas.
Genus Crussoliceras Énay, 1959
Type species. Ammonites Crusoliensis Fontannes, 1879
Crussoliceras sp.
(Fig. 7.D1-D2 ![]() )
)
Material. One poorly preserved internal mould (J 2020.269.1).
Dimensions. Due to the fragmentary status, dimensions are not given.
Description. Poorly preserved, fragmentary internal mould of a body chamber with less than 30 mm whorl height. Only 4 ribs preserved of the whorl fragment. Cross section oval, slightly compressed. Strong, rectiradiate ribs rise from the umbilical region that bend forward and radial on midflank. Ribs slightly prorsiradiate on the upper flank. Venter not preserved. Ribs highest on midflank. Ribs may bifurcate on upper flank. Suture, umbilicus, peristome not preserved.
Remarks. The strong, coarse and projected, sparsely placed, sometimes bifurcating ribbing that raised above the flanks and the oval cross section is typical for the genus. Unfortunately, the paucity, poor preservation and fragmentary status restricted further determination. The typical bifurcation of ribs on the upper flank is well visible on the third rib. The present specimen shows similarities toward the inner whorls of C. crusoliense Fontannes, C. geyeri Sapunov and C. sayni Camus & Thieuloy, however the fragmentary status hinders specific determination. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Crussoliceras is typical of the Kimmeridgian. Geographically it occurs in the Submediterranean Province (SE France, Spain, French and Swiss Jura Mountains), however it is reported from the Mediterranean Province (Morocco, Bulgaria), also by Énay and Howarth (2019).
Genus Progeronia Arkell, 1953
Type species: Perisphinctes progeron Ammon, 1875
Progeronia sp.
(Fig. 7.E1-E2 ![]() )
)
Material. Two poorly preserved and fragmentary internal moulds (J 2020.368.1, 571.1).
Dimensions. Due to fragmentary status dimensions are not given.
Description. Poorly preserved whorl fragments of phragmocone. Conch rather evolute, compressed. Umbilicus wide, profound. Three whorls seen. Umbilical wall vertical on inner, and oblique on outer whorls. Umbilical shoulder rounded. Flanks convex, converging. Venter rounded. Fine, simple, rectiradiate ribs rise on the umbilical wall, bending forward upward and become radial on the umbilical shoulder. Passing the umbilical shoulder bend forward and become prorsiradiate; ribs travel straightforward on the flank and bifurcate on the upper third. Bifurcated ribs continuously cross the venter. Number of primaries per half whorl acquired at D=20 mm: 21; D=45 mm: 27; D=100 mm: 36 [estimated]. Cross section compressed, rounded trapezoidal. Neither suture nor aperture seen.
Remarks. Progeronia is an important genus in the Kimmeridgian (Olóriz, 1978, p. 399). The poor and fragmentary collected material prevented specific determination. Rib curve not presented. Due to the poor material available, only the number of ribs is counted per half whorl at estimated diameters. Based on the very estimated rib curve of specimen J 2020.368.1, this may belong to the P. unicompta - P. simplex group (cf. Sarti, 1993, Fig. 23). First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Progeronia species are typical for the Kimmeridgian; geographically the species noted above are reported from the western Tethys and Submediterranean Province (Énay & Howarth, 2019).
Genus Gravesia Salfeld, 1913
Type species: Ammonites Gravesianus Orbigny, 1847
Gravesia aff. gigas (Zieten, 1830)
(Fig. 8.A1-A3 ![]() )
)
1830. Ammonites gigas, Zieten, p. 17, Pl. 13, fig. 1.a-b.
1959. Gravesia cf. gigas (Zieten), Berckhemer & Hölder, p. 66, Fig. 35.B, G, Pl. 15, fig. 71.
1963. Gravesia gigas (Zieten), Hahn, p. 97, Pl. 9, figs. 1-4; Pl. 10, fig. 1-2.
1966. Gravesia gigas (Zieten), Énay, p. 7, Pl. A, fig. 1.A-B.
1967. Gravesia gigas (Zieten), Cope, p. 12, Pl. 1, fig. 1.
1977. Gravesia gigas (Zieten), Ziegler, Pl. 6, fig. 5.
1989. Gravesia gigas gigas (Zieten), Hantzpergue, p. 191, Figs. 48.A-F, 49.A-B, 50.A-C, 57-58, 128.16; Pl. 17, figs. A-C.
1994. Gravesia gigas (Zieten), Fischer, p. 186, Pl. 85, fig. 2; Pl. 86, fig. 1.
2010. Gravesia cf. gigas, Gallois & Etches, Fig. 8.a-b.
2015. Gravesia gigas (Zieten), Comment et al., Pl. 5, fig. 1.
2016. Gravesia gigas intermedia Hantzpergue, Scherzinger & Schweigert, Figs. 4.A-F, 5.A-B, .D, .G-H, 6.A-E.
Material. One worn internal mould from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.101.1. | 113.9 | 44.1 | (34) | 39.1 | 1.297 | 0.298 | 0.343 |
Description. Small-sized Gravesia with rather involute, compressed conch. Umbilicus is wide and shallow. Umbilical wall vertical and high. Umbilical shoulder, flanks and venter all rounded. At the umbilical shoulder or a little higher (uncertainty due to the poor preservation) low tubercles rise. Prorsiradiate ribs emanate from the low tubercles that bifurcate below the midflank. Ribs travel toward the venter, however due to poor preservation their crossing the venter is not visible. The phragmocone is worn and dissolved; suture only partly preserved and dissolved but approximately resembling the suture of Figure 48.C in Hantzpergue (1989, p. 194).
Remarks. Reliable identification of Gravesia species is based on the analysis of coiling, cross section (cf. Gallois & Etches, 2010, Fig. 9) and sculpture. The present specimen has similar fine sculpture and dense ribbing comparable to G. irius (Orbigny, 1850), as seen on the specimen illustrated by Schweigert (1999, p. 34, Fig. 4), however the cross section of the latter one is much more depressed. Gravesia gravesiana and Gravesia gigas intermedia are more evolute than the present specimen. The collected specimen compares favourably to a specimen of Gravesia gigas illustrated by Comment et al. (2015, Pl. 1, fig. 1.a-b) based on their cross sections and ribbing. The cross section of the present specimen is perhaps closest to the specimen of Gravesia cf. gigas in Berckhemer and Hölder (1959, p. 64, Fig. 35.a). However, although there are common characters with G. gigas, and G. irius, the poor preservation prevented a precise determination. First record from Hungary.
Stratigraphic and geographic distribution. Reported from the lower Tithonian (middle Hybonotum Zone) in the Franco-German Bioma, England. Its FAD indicates the Tithonian (Comment et al., 2015) and Portlandian (Hantzpergue, 1989). Geographically Gravesia gigas is only reported from the Subboreal and the Submediterranean Provinces in western Europe therefore the Hungarian occurrence indicates the extreme southern limit of its distribution.
|
|
Subfamily Lithacoceratinae Zeiss, 1968
Genus Lithacoceras Hyatt,
1900
[= Discosphinctoides Olóriz, 1978; Silicisphinctes Schweigert & Zeiss,
1999]
Type species: Ammonites Ulmensis Oppel, 1858
? Lithacoceras sp.
(Fig. 8.B1-B2 ![]() )
)
Material. Two poorly preserved internal moulds from the quarry floor amongst debris (J 2020.97.1, 367.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D | N/2 |
| J 2020.97.1. | 151 | 32 | 45 | 72 | 0.711 | 0.298 | 0.477 | 32 |
Description. Big sized, poorly preserved conch representing the phragmocone and body chamber. Coiling serpenticone, three whorls seen. Umbilicus wide, shallow. Umbilical wall short, vertical. Umbilical shoulder rounded. Flanks parallel or slightly converge. No ventral shoulder. Venter rounded. At the umbilical shoulder densely placed, fine, prorsiradiate ribs rise. Ribs bifurcate on the upper flank, around 2/3 of the whorl height, where the new whorl touches the previous. Bifurcation regular, no single ribs observed. Ribs cross the venter radially. Whorl section compressed oval. Suture, aperture and constrictions not seen.
Remarks. The original designation of the genus by Olóriz (1978, p. 481) fits to the present specimen. Unfortunately, further designation is impossible. On one hand, there are many species included in this genus: L. ardescicus, L. geron, L. modestus, L. neohispanicum, L. praenuntians, L. rhodaniforme, L. rigida, L. roubyanus, L. stenocyclus, L. subborealis, L. vandellii, L. virgulatiformis. These species show fine distinctions and variations of their rib density and thickness, bifurcation, trifurcation, number of intercalatory ribs and whorl section. Careful analysis would require construction of rib curves for complete specimens and of good preservation but on the other hand, the poor preservation of the present specimen prevents positive assignation to any of these species however there are certain similarities with L. roubyanus, and L. praenuntians. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Reported from the upper Kimmeridgian of the Submediterranean Province (Olóriz, 1978) and Indo-Malagasy Province (Énay & Howarth, 2019).
Lithacoceras sp. juv.
(Fig. 8.C1-C2 ![]() )
)
Material. One fragmentary and poorly preserved internal mould.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D | N/2 |
| J 2020.365.1. | n.d. | 34.4 | (36) | (52) | 0.955 | n.d. | n.d. | 40 |
Description. Poorly preserved and crushed internal mould of half whorl segment with a small fragment of inner whorl attached. Coiling evolute, umbilicus wide, shallow. Four whorls partly seen. Umbilical shoulder vertical, short, smooth. Umbilical shoulder rounded. Flanks convex, converging. Venter rounded, no mid-siphonal ridge or band seen. Cross section rounded trapezoidal; conch slightly compressed. Fine, densely spaced ribs rise from the umbilical edge. Ribs rectiradiate on the umbilical wall, bending forward on the umbilical shoulder, then flex on the flank. Ribs bifurcate at 2/3 of the flank. Bifurcating ribs equally spaced and uniform, crossing the venter continuously.
Remarks. The poor preservation and fragmentary status did not allow for the precise counting of ribs per whorl or half whorl; therefore rib curve analysis was not undertaken. Whorl section of the present specimen is very close to the holotype of Lithacoceras delcampanai (Sarti, 1993, Fig. 37, no. A413V). The only difference is that the collected specimen has slightly inflated flanks, forming a transition between the holotype and the specimen no. ME479 (illustrated by Sarti, 1993) having subcircular cross section. Number of ribs is difficult to assess. Only two quarter whorls are preserved moderately enough to estimate a rib count. The specimens of Sarti (1993, Fig. 39.W-X) show 60-70 primaries at a diameter of 110 mm. Calculation for the collected specimen at a comparable diameter based on the last quarter whorl approximates to 76 (19x4), however this is an overestimation because the earlier whorls show fewer primaries (cf. Sarti, 1993, Fig. 39). The poor preservation therefore did not justify a specific assignment.
Stratigraphic and geographic distribution. As for Lithacoceras sp. above.
Lithacoceras aff. siliceum (Quenstedt, 1857)
(Fig. 8.D1-D2 ![]() )
)
1857. Amm. planulatus siliceous, Quenstedt, p. 775, Pl. 95, fig. 27.
1888. Amm. planulatus siliceous, Quenstedt, p. 1073, Pl. 125, fig. 2.
1959. Perisphinctes siliceus (Quenstedt), Berckhemer & Hölder, p. 41, Pl. 14, figs. 69-70; Pl. 16, fig. 73.
1959. Perisphinctes siliceus (Quenstedt), Hölder & Ziegler, p. 187, Pl. 17, fig. 2.
1968. Usseliceras (Subplanitoides) siliceum (F.A. Quenstedt), Zeiss, p. 64, Pl. 4, fig. 1.
1993. Subplanitoides siliceum (Quenstedt), Olóriz et al., p. 281, Pl. 2, fig. 3.
1994. Usseliceras (Subplanitoides) siliceum (Qu.), Schlegelmilch, Pl. 43, fig. 4.
2018. Lithacoceras (Silicisphinctes) sp. gr. siliceus (Quenstedt), Énay, p. 83, Pl. 7, fig. 3.
Material. One poorly preserved and fragmentary internal mould (J 2020.185.1).
Dimensions. Due to the fragmentary status, dimensions are not given.
Description. Small sized phragmocone fragment. Inner whorl segments also visible. Umbilicus wide and shallow. Smooth umbilical wall. Umbilical shoulder rounded. Flanks converging. No ventral shoulder. Venter rounded. Oval cross section. Densely spaced and fine radial or prorsiradiate ribs emanate from the umbilical shoulder. Radial ribs gently bend forward at about 1/3 of the flank and travel through the upper whorl side and cross the venter. A mid-siphonal bend or groove is unobserved. Ribs appear to bifurcate on the upper whorl and bidichotomous ribbing may occur. Secondary ribs may appear on the upper flank. Primaries and secondaries have a comparable sculpture. There are 26 ventral ribs and 14 primaries on the last quarter whorl. Neither suture nor peristome are seen.
Remarks. Although the present specimen is poorly preserved and fragmentary, the dense ribbing, bifurcation and bidichotomous ribs (also characteristic for many other genera) most probably refers the collected specimen to Lithacoceras. Almost all the figured specimens depict adult microconchs with well-developed lappets creating difficulties when analysing early growth stages. Collected whorl fragment is closest to specimen IGM-6187 illustrated by Olóriz et al. (1993, Pl. 2, fig. 3) with its rather straight and sometimes polyplocoid ribbing. Zeiss (1968, Pl. 4, fig. 1) and Schlegelmilch (1994, Pl. 43, fig. 4) refigured the holotype specimen (Quenstedt, 1857, Pl. 95, fig. 27) of an adult microconch with lappets. The poor preservation and juvenile growth stage of the present specimens prevented analysis of important characters (number of ribs, adult stage, complete whorls) and the specimen is only tentatively allied to the group siliceum as Lithacoceras aff. siliceum. First record from Hungary.
Stratigraphic and geographic distribution. Lithacoceras siliceum has been reported from the lower Tithonian (Hybonotum Zone) of Mexico, southern Germany, France and Hungary.
Genus Euvirgalithacoceras Zeiss et al., 1996 [= Subplanites Spath, 1925]
Type species: Virgatosphinctes supremus Schneid, 1915
Euvirgalithacoceras? sp.
(Fig. 8.E1-E2 ![]() )
)
Material. 2 fragmentary, poorly preserved internal moulds from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.105.1. | (90) | 27.7 | (36) | (35) | 0.769 | 0.400 | 0.389 |
| J 2020.111.1. | (103) | (25) | (36) | (42) | 0.694 | 0.350 | 0.408 |
Description. Middle sized, rather evolute conch. Umbilicus wide, shallow. Umbilical wall inclined inward, forming a folded umbilical shoulder. Cross section rounded trapezoidal with inwardly inclined umbilical shoulder. Strong, simple, and prorsiradiate ribs appear at the umbilical shoulder that branch on the upper part of the flank (roughly at three quarter of whorl height). Branching of the primaries bipartiate. Virgatotomous branching occurs, too. At the height of branching intercalating secondary ribs may appear. Venter, suture, constrictions not seen.
Remarks. The presence of virgatotomous ribs on both specimens suggests on one hand that these specimens may belong to Virgatosphinctinae and probably represent early, internal whorls of Euvirgalithacoceras or Pachysphinctes. Assigning the present specimens to Euvirgalithacoceras is supported by the fact that some of the ribs are virgatotomous. On the other hand, important features (venter, number of primaries per one or half whorl) are not preserved, therefore even generic assignment is tentative. Imlay (1981, Pl. 8, fig. 15) figured a very similar specimen from the Boreal Faunal Realm (Alaska). First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Euvirgalithacoceras is an upper Kimmeridgian-lower Tithonian taxon with ubiquitous distribution both in Boreal and Tethysian Faunal Realms.
Euvirgalithacoceras aff. ombonii (Del Campana, 1905)
(Fig. 8.F1-F2 ![]() )
)
1905. Perisphinctes ombonii, Del Campana, p. 83, Pl. 3, fig. 8.
1993. Subplanites ombonii (Del Campana), Sarti, p. 88, Figs. 33, 35.H, Pl. 11, fig. 3.
Material. One poorly preserved whorl fragment (J 2020.589.1).
Dimensions. Due to fragmentary status, dimensions are not given.
Description. The tiny whorl fragment represents partially the umbilical shoulder and flank up to the venter. Umbilicus not preserved. Strong, flexuous, distantly spaced primary ribs rise from the umbilical shoulder. Ribs bifurcate at 2/3 of the flank. Ribs polygirate, sometimes intercalatory ribs appear. Ribs bifurcate and trifurcate.
Remarks. Cross section of the present fragment is comparable to specimen A4V illustrated by Sarti (1993, Fig. 33.A). Similar species are E. moernsheimensis and E. reisi, however Sarti (1993, p. 89) made a detailed differential diagnosis that is only partially applicable to the present specimen. On one hand it strengthens its position in this species; but on the other hand, the present specimen is only a whorl fragment, hence umbilicus and coiling are uncertain, therefore doubts justify assignment to Euvirgalithacoceras aff. ombonii. First record from Hungary.
Stratigraphic and geographic distribution. Stratigraphically Euvirgalithacoceras ombonii occurs in the upper part of the Beckeri Zone of the Mediterranean Tethys.
Subfamily Torquatisphinctinae Tavera, 1985
Genus Torquatisphinctes Spath, 1924
Type species: Ammonites torquatus J. de C. Sowerby, 1840
Torquatisphinctes aff. laxus Olóriz, 1978
(Fig. 9.A ![]() )
)
1978. Torquatisphinctes laxus n. sp., Olóriz, p. 454, Pl. 40, fig. 4.
1993. Torquatisphinctes laxus Oloriz, Sarti, p. 87, Pl. 10, fig. 1.
1994. Torquatisphinctes laxus Oloriz, Schlegelmilch, p. 90, Pl. 41, fig. 3.
2004. Torquatisphinctes gr. laxus Oloriz, Marino et al., Pl. 2, fig. 4.
Material. Two poorly preserved internal moulds.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D | N/2 |
| J 2020.144.1. | 100.7 | (27) | (29) | 48.4 | 0.931 | 0.288 | 0.481 | 31 |
| J 2020.364.1. | (110) | (30) | (35) | 49.7 | 0.857 | 0.318 | 0.452 | 31 |
Description. Medium size, evolute conch fragments of the body chamber. Cross section compressed, rounded subrectangular. Umbilical wall deep, smooth, vertical. Umbilical shoulder rounded. Strong, slightly prorsiradiate primary ribs originate at the umbilical shoulder crossing the venter continuously. On the midflank or higher (usually 2/3 of the flank) ribs bifurcate and also cross the venter continuously. There are 31 primary ribs on the last half whorl. At least 2 primary ribs are unbifurcated. There may be more; however, the poor preservation restricted analysis. Neither suture nor constrictions visible.
Remarks. Published specimens do not allow thorough comparisons, however the number of primary ribs on the last whorl can elucidate the intraspecific variation of the ribbing. Due to the poor preservation of the present specimens, only the number of primary ribs of the last half whorl is calculated. These numbers can vary between 25-31 as follows: 25 in Sarti (1993, Pl. 10, fig. 1); F.G12.20.3: 27 (holotype, Olóriz, 1978, Pl. 40, fig. 4); 29 in Marino et al. (2004, Pl. 2, fig. 4); and E51: 31 (Schlegelmilch, 1994, p. 216, Pl. 41, fig. 3). Number of primary ribs per half whorl of the collected specimens falls in this range, however, all the measured parameters and the general features of our material suggests referral to this species. Related species (T. intermedius and T. regularis) either bear more single primary ribs, or the prorsiradiate primaries bend pronouncedly forward or the umbilicus is wider. Unfortunately, these features are uncertain for the Hungarian specimens, which are tentatively assigned to Torquatisphinctes laxus. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Torquatisphinctes laxus is reported from the upper Kimmeridgian (Beckeri Zone) to the lower Tithonian Hybonotum Zone of Betic Cordilleras Spain, southern Germany, Hungary, the Eastern Carpathians and Sicily.
|
|
Genus Pachysphinctes Dietrich, 1925 [= Balochistaniceras Fatmi & Zeiss, 1999]
Type species: Perisphinctes africogermanus Dietrich, 1925
Pachysphinctes sp.
(Fig. 9.B1-B4 ![]() )
)
Material. 12 poorly preserved internal moulds (J 2020.184.1, 211.1, 366.1, 563.1-568.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D | N/2 |
| J 2020.148.1. | 87.6 | (20) | (27) | 44.5 | 0.741 | 0.308 | 0.508 | 24 |
| J 2020.286.1. | (67) | (24) | (21) | (30) | 1.143 | 0.313 | 0.448 | 27 |
| J 2020.363.1. | 104.3 | (27) | 29.4 | 51.1 | 0.918 | 0.282 | 0.490 | 26 |
Description. Poorly preserved, fragmented, evolute conch. Umbilicus wide, shallow. Umbilical wall rounded, inclined, short and smooth. Umbilical shoulder rounded. Flanks convex, rounded. Venter rounded, no ventral shoulder. Distinct, dense, radial or slightly prorsiradiate ribs rise from the umbilical shoulder. Ribs regularly bifurcate just above the midflank. Ribs cross the venter without interruption. Rarely, constrictions are present. Suture and aperture not seen.
Remarks. The generic diagnosis emended by Olóriz (1978, p. 461) clearly stated that there is no trifurcation of ribs s. str. which is a generic feature for the genus. The present specimens show only bifurcation, which strengthens the designation of these specimens to Pachysphinctes. P. bathyplocus (Waagen) figured by Howarth (1998, Pl. 5, fig. 1.a), which has the same number of primary ribs per half whorl shows a U/D range of 0.45-0.55. This is the same for P. adelus (Gemmellaro) having 25 primary ribs per half whorl (Olóriz, 1978, Pl. 54, fig. 2), where the U/D range is 0.51-0.52. Having poorly preserved fragmentary and incomplete specimens to hand any assignment to species-level would be poorly justified: Pachysphinctes sp. is an adequate initial determination. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. It is reported from the upper Kimmeridgian in the Tethys (Indo-Malagasy Province: Énay & Howarth, 2019).
Subfamily Virgatosphinctinae Spath, 1923
Genus Malagasites Énay, 2009
Type species. Perisphinctes (Virgatosphinctes) Haydeni Uhlig, 1910
Malagasites? denseplicatus (Waagen, 1875)
(Fig. 9.C ![]() )
)
1875. Perisphinctes denseplicatus Waagen, n. sp., Waagen, p. 201, Pl. 46, fig. 3; Pl. 55, figs. 1-2.
1910. Perisphinctes (Virgatosphinctes) denseplicatus Waagen, Uhlig, p. 313, Pl. 53, fig. 3, Pl. 54, fig. 1; Pl. 55, figs. 1-3; Pl. 56, fig. 1.
1935. Virgatosphinctes sp., Vadász, p. 62. [in lit.]
1943. Virgatosphinctes cf. V. denseplicatus (Waagen), Imlay, p. 535, Pl. 89, figs. 1-4.
1979. Virgatosphinctes aff. denseplicatus (Waagen), Thomson, p. 16, Fig. 5, Pls. 3.j-k, 4.a.
1989. "Virgatosphinctes" sp. aff. V. denseplicatus (Waagen), Myczyński, p. 99, Pl. 8, fig. 1; Pl. 12, fig. 5.
non 1991. "Virgatosphinctes" denseplicatus (Waagen), Olóriz & Tintori, p. 471, Pl. 22, fig. 2.
non 2004. Virgatosphinctes denseplicatus (Waagen), Yin & Énay, p. 673, Fig. 4.4.
2004. Virgatosphinctes denseplicatus (Waagen), Yin & Énay, p. 673, Fig. 4.3.
2016. Virgatosphinctes denseplicatus (Waagen), Pandey et al., p. 148, Figs. 6.A-C, 7, Pl. 1, fig. 5; Pl. 2, fig. 1.
Material. Two poorly preserved internal moulds, whorl fragments from Zengővárkony.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D | N/2 |
| J 711 | 94.6 | 29.4 | (33) | 35.8 | 0.891 | 0.349 | 0.378 | 44 |
| J 2020.150.1. | 93.8 | (28) | (27) | 39.9 | 1.037 | 0.288 | 0.425 | 43 |
Description. Mid-sized conchs representing body chambers lacking the early whorls, which are crushed and/or dissolved. Umbilicus wide, shallow. Umbilical wall low. Umbilical shoulder rounded. Flanks converging, subparallel. No ventral shoulder. Venter rounded. Cross section oval to rounded trapezoidal. Sculpture consists of fine, densely placed ribs that rise on the lower part of the umbilical wall, flexing backward and cross the upper flank; on the lower flank bending forward to become radial on the midflank then start to bend backward again on the upper flank and crossing the mid-siphonal line perpendicularly. Ribs bifurcate at upper 2/3 of flank. Rarely, bidichotomous ribs usually form around constrictions. No band or groove seen on the mid-siphonal line. Constrictions are present following the shape of the primaries numbering one constriction per half whorl. Neither suture nor aperture are visible.
Remarks. The Virgatosphinctes sp. referred to by Vadász (1935, p. 62) is the same specimen under label J 711 collected by János Böckh in 1877, which is described and illustrated here. There are no ribs joining at the umbilical shoulder, and the absence of virgatotomous ribbing refers the specimen to this species as Myczyński (1989, p. 98) underlined. The present specimens most resemble the KG.712.68 specimen illustrated by Thomson (1979). Olóriz and Tintori (1991) and Yin and Énay (2004, p. 673, Fig. 4.4.) illustrated morphs with virgatotomous ribbing. Specimen J 711 compares favourably to specimen USNM 103363b of Imlay (1943, Pl. 89, fig. 4) and represents the finer ribbed and more compressed variety of the species as Imlay (1943, p. 535) noted. When Énay (2009) introduced this genus, he included forms described from the Indo-Malagasy province (Énay & Howarth, 2019). The present specimens are only tentatively placed in Malagasites.
Stratigraphic and geographic distribution. According to Bardhan et al. (2007, p. 381) Virgatosphinctes (including V. denseplicatus) is a typical Indo-Madagascan form and typically restricted to peri-Gondwanan regions. From the Western Tethys it is reported from Mexico, Cuba and now Hungary in the early Tithonian.
Superfamily Aspidoceratoidea Zittel, 1895
Family Aspidoceratidae Zittel, 1895
Subfamily Aspidoceratinae Zittel, 1895
Genus Aspidoceras Zittel, 1868
Type species: Ammonites Rogoznicensis Zejszner, 1846
Aspidoceras div. sp.
Material. 54 poorly preserved, dissolved, mostly fragmentary specimens (J 2020.129.1, 132.1, 167.1-170.1, 193.1, 202.1, 219.1, 227.1, 242.1, 276.1, 300.1, 456.1-2020.496.1, 569.1).
Dimensions. Due to poor preservation dimensions are not given.
Remarks. These fragments definitely represent a range of different Aspidoceras species; however, the poor preservation prevented specific designations. The fragments may belong to A. acanthicum, A. apenninicum, A. binodum, A. sesquinodosum, or perhaps A. uninodosum.
Aspidoceras acanthicum (Oppel, 1863)
(Fig. 9.D1-D2 ![]() )
)
1863. Ammonites acanthicus Opp., Oppel, p. 219.
1872. Aspidoceras acanthicum, Opp., Gemmellaro, p. 148, Pl. 2, figs. 8-9.
1873. Aspidoceras acanthicum Oppel, Neumayr, p. 195 [55], Pl. 41.
1876. Ammonites acanthicus Oppel, Dumortier & Fontannes, p. 125, Pl. 18, figs. 4-5.
1878. Aspidoceras acanthicum Oppel, Herbich, p. 174 [156], Pl. 16, fig. 2.
1879. Aspidoceras acanthicum Oppel, Fontannes, p. 88, Pl. 12, fig. 5.
1886. Aspidoceras acanthicum Opp., Pavlow, p. 75, Pl. 2, fig. 2.
1931. Aspidoceras aff. acanthicum (Oppel), Spath, p. 624, Pl. 123, fig. 7.
1935. Aspidoceras cf. acanthicum Opp., Vadász, p. 62. [in lit.]
1959. Aspidoceras acanthicum Opp., Collignon, Pl. 128, figs. 479-480.
1966. Aspidoceras acanthicum (Oppel), Anđelković, p. 75, Pl. 28, fig. 1.
1973. Aspidoceras acanthicum Oppel, Preda, Pl. 10, fig. 11; Pl. 18, fig. 8.
1976. Euaspidoceras sp. gr. acanthicum, Lillo Bevia, p. 26, Pl. 6, figs. 1-2.
1977. Aspidoceras acanthicum (Oppel), Ziegler, Pl. 4, fig. 1.
1978. Aspidoceras acanthicum acanthicum (Oppel), Olóriz, p. 301, Pl. 23, fig. 3.
1984. Aspidoceras (Aspidoceras) acanthicum (Oppel), Rossi, p. 107, Pl. 34, fig. 5.
1984. Aspidoceras cf. A. acanthicum (Oppel), Verma & Westermann, p. 63, Pl. 14, figs. 1-3, Pl. 15, fig. 1.
1985a. Aspidoceras acanthicum (Oppel), Checa, p. 74, Fig. II.3.5, .10, Pl. 8, fig. 1.
1986. Aspidoceras acanthicum (Oppel), Sarti, p. 505, Pl. 6, fig. 3.A-B.
1991. Aspidoceras acanthicum (Oppel), Krishna & Pathak, p. 11, Pl. 1, fig. 2.
1993. Aspidoceras acanthicum (Oppel), Sarti, p. 124, Pl. 26, fig. 2.
1997. Aspidoceras acanthicum (Opp.), Hantzpergue et al., Pl. 24, fig. 8.
1999. Aspidoceras (A.) acanthicum acanthicum (Oppel), Fatmi & Zeiss, p. 73, Pl. 34, fig. 3; Pl. 35, fig. 2.
2011. Aspidoceras acanthicum (Oppel), Baudouin et al., Pl. 11, fig. 6.
2013. Physodoceras acanthicum (Oppel), Főzy & Scherzinger, p. 185, Pl. 11, fig. 3; Pl. 12, fig. 1.
Material. Two whorl fragments of large sized specimens (J 2020.131.1, 228.1).
Dimensions. Due to fragmentary status, dimensions not given.
Description. Coiling not seen. Fragments represent phragmocones, septae not seen. Cross section is oval, convex, midflanks converging or parallel. Umbilical wall deep, smooth and vertical. At the umbilical shoulder big, well developed tubercles are erected. On the midflank, or on the upper midflank a second row of tubercles are situated above the umbilical tubercles. There are 3 tubercles per quarter whorl. Tubercles may interconnect by fine, short rib like folds. Venter smooth and rounded, no ventral shoulder. Cross section compressed, usually higher than wide. Suture not seen.
Remarks. This is a frequent species with high intraspecific variability and usually to large diameters (D>200 mm). The holotype specimen illustrated by Oppel is also a substantially sized (D=200 mm) specimen. The present whorl fragments also represent large individuals. Checa (1985a, Pl. 8, fig. 1) published an atypical specimen with one row of tubercles at the umbilical shoulder. The present specimens are closest to the specimens of Baudouin et al. (2011, Pl. 11, fig. 6), Collignon (1959, Pl. 128, fig. 479), Dumortier & Fontannes (1876, Pl. 18, fig. 4), Herbich (1878, Pl. 16, fig. 2), Rossi (1984, Pl. 34, fig. 5). Checa (1985a, Pl. 8, fig. 1), Olóriz (1978, Pl. 23, fig. 3) and Sarti (1993, Pl. 26, fig. 2), whereas Spath (1931, Pl. 123, fig. 7.a) depicted less heavily ornamented varieties of this species. Whorl section of specimen J 2020.228.1 is comparable to the cross section of the holotype refigured by Checa (1985a, Fig. II.3.5.C).
Stratigraphic and geographic distribution. Aspidoceras acanthicum is reported from the Acanthicum Zone of the upper Kimmeridgian. It has a wide geographic distribution in the Tethysian (Mediterranean, Submediterranean, and Indo-Malagasy Provinces) and the Boreal Faunal Realms (Subboreal Province).
Aspidoceras binodum (Oppel, 1863)
(Fig. 9.E1-E2 ![]() )
)
1863. Ammonites binodus Opp., Oppel, p. 217.
1907. Aspidoceras binodum Opp. sp., Toula, p. 64, Pl. 14.
1929. Physodoceras binodum Oppel, Wegele, p. 89 (183), Pl. 11, fig. 3.
1931. Aspidoceras cf. binodum (Oppel), Spath, p. 637, Pl. 119, fig. 2.a-b.
1935. Aspidoceras binodosum Opp., Vadász, p. 62. [in lit.]
1959. Aspidoceras aff. binodum Opp., Collignon, Pl. 129, fig. 482.
1966. Physodoceras binodum (Oppel), Anđelković, p. 86, Pl. 25, fig. 1.
1973. Aspidoceras binodum Oppel, Preda, Pl. 18, figs. 3-5.
1978. Aspidoceras binodum (Oppel), Olóriz, p. 289, Pl. 24, figs. 2-4.
1985a. Aspidoceras binodum (Oppel), Checa, p. 54, Fig. II.3.1, .6, Pl. 1, fig. 1; Pl. 2, figs. 2-5.
1994. Aspidoceras cf. binodum (Oppel), Wierzbowski, Pl. 4, figs. 10-11.
1999. Aspidoceras (Aspidoceras) aff. binodum (Oppel), Fatmi & Zeiss, p. 76, Pl. 26, fig. 2.
2003. Aspidoceras binodum (Oppel), Gygi, p. 141, fig. 162.
Material. Seven fairly preserved internal moulds from the MBFSz collection and from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2097 | 57.1 | 30.4 | (25) | 16.3 | 1.216 | 0.438 | 0.285 |
| J 2020.119.1. | (77) | (40) | (38) | 19.0 | 1.053 | 0.493 | 0.247 |
| J 2020.149.1. | (60) | (26) | (20) | 19.9 | 1.300 | 0.333 | 0.332 |
| J 2020.297.1. | 54.5 | (24) | (22) | (17) | 1.091 | 0.404 | 0.312 |
| J 2020.333.1. | (61) | (26) | (24) | (22) | 1.083 | 0.393 | 0.361 |
| J 2020.335.1. | 76.2 | (31) | (31) | (22) | 1.000 | 0.407 | 0.289 |
| J 2020.348.1. | (90) | (42) | (38) | 27.5 | 1.105 | 0.422 | 0.305 |
Description. Small to medium sized phragmocone conchs. Umbilicus moderate, deep, umbilical wall vertical, smooth and profound. Umbilical shoulder rounded; flanks convex. No ventral shoulder. Venter rounded, smooth. Cross section depressed, subcircular. Stronger or weaker tubercles are located at the umbilical shoulder. There are 8 peri-umbilical tubercles on the last half whorl. A second row of tubercles appear on the lower flank, that sometimes correspond to the peri-umbilical row of tubercles. They are densely placed close to each other or sometimes adherent. On the last half whorl there are fewer tubercles in the upper row than at the umbilical shoulder. Rarely, on the well-preserved septate parts, delicate, fine and shallow folds rise from some tubercles of the lower flank, that travel toward the venter, but disappear at the upper flank. Remains of suture lines are preserved; but no aperture seen.
Remarks. According to Checa (1985a, p. 56) A. binodum and A. rogoznicense show morphological overlapping characters, however their markedly different stratigraphic position helps to properly discriminate between them. Based on lithological characters the specimens are representative of these two species. Specimen J 2020.119.1. is closest to Ch.G27.1.7 of Checa (1985a, Pl. 2, fig. 5) representing the morphospecies with well-developed tubercles. Specimen J2097 is closest to No. 93 of Spath (1931, Pl. 119, fig. 2a) and represents morphs with fainter tubercles.
Stratigraphic and geographic distribution. Aspidoceras binodum is reported from the lower Kimmeridgian Bimammatum to Acanthicum zones. It has a wide geographic distribution in the western Tethys and adjacent territories in the Submediterranean Province and also from the northern Tethys (Baluchistan) and the southern Tethys (Madagascar).
Aspidoceras caletanum (Oppel, 1863)
(Fig. 9.F1-F2 ![]() )
)
1863. Ammonites Caletanus Opp., Oppel, p. 220.
1877. Ammonites (Aspidoceras) Caletanus d'Orbigny, Favre, p. 60, Pl. 7, fig. 6.
1881. Ammonites caletanus Oppel, Gosselet, Pl. 12, fig. 10.
1886. Aspidoceras Caletanum Opp., Pavlow, p. 73, Pl. 8, fig. 1.a-b.
1985a. Aspidoceras caletanum (Oppel), Checa, p. 94, Fig. II.3.14, .17, Pl. 15, fig. 2.
1989. Aspidoceras caletanum (Oppel), Hantzpergue, p. 330, Figs. 110.A-K, 111-112, 118-119; Pl. 43, figs. A-E; Pl. 44, figs. A-B.
1994. Aspidoceras caletanum (Oppel), Fischer, p. 178, Pl. 81, figs. 1-2; Pl. 82, fig. 1.
1997. Aspidoceras caletanum (Opp.), Hantzpergue et al., Pl. 23, fig. 8.
1997. Aspidoceras ex gr. caletanum (Oppel), Kutek & Zeiss, Pl. 14, fig. 1.
1999. Aspidoceras caletanum (Oppel), Schweigert, p. 30, fig. 2.
2015. Aspidoceras caletanum (Oppel), Comment et al., Pl. 4, figs. 2, 4-5; Pl. 5, figs. 3, 5.
Material. One internal mould resting on the quarry floor amongst debris.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.87.1. | 98.4 | 33.4 | (33) | 35.7 | 1.012 | 0.335 | 0.362 |
Description. Comparatively well-preserved half whorl represents part of the phragmocone is septate throughout. Coiling evolute, umbilicus wide and shallow. Umbilical wall vertical, umbilical shoulder rounded. Flanks parallel, ventral shoulder rounded; venter smooth. At the umbilical shoulder fine, radial tubercles rise, that diminish rapidly toward midflank. Shallow, fine tubercles are located at the umbilical shoulder. There are 9 ventrolateral tubercles per half whorl. Cross section oval, with parallel flanks. Suture not seen.
Remarks. According to Hantzpergue (in Fischer, 1994, p. 178) A. caletanum differs from A. longispinum with its subcircular cross section and more evolute coiling, which is typical of the present specimen, which compares favourably to the specimen illustrated by Comment et al. (2015, Pl. 5, fig. 3.a-b). It is identical in cross section to the specimen of Checa (1985a, Fig. II.3.14.B). There are the same number of tubercles (nine) on the ventral shoulders compared to specimens illustrated by Gosselet (1881, Pl. 12, fig. 10), Hantzpergue (1989, Pl. 44, fig. B) and Pavlow (1886, Pl. 8, fig. 1.a). The collected specimen differs from the others only in having ventrolateral tubercles upper, slightly below on the ventrolateral shoulder. Specimens of this biospecies usually exceed 140 mm reaching 250 mm in diameter; however, the range stretches from 64 mm (Hantzpergue, 1989) to 252 mm (Checa, 1985a). First record from Hungary.
Stratigraphic and geographic distribution. Aspidoceras caletanum is reported from the upper Kimmeridgian Eudoxus Zone (Caletanum Subzone) from the Subboreal Province with occasional occurrences within the Submediterranean Province.
Aspidoceras ex gr. longispinum (Sowerby, 1825)
(Fig. 9.G ![]() )
)
1873. Aspidoceras longispinum Sowerby, Neumayr, p. 196 [56], Pl. 42, fig. 1.
1874. Ammonites longispinus Sowerby, Loriol & Pellat, p. 276, Pl. 2, fig. 2.
1877. Ammonites (Aspidoceras) longispinus Sowerby, Favre, p. 60, Pl. 7, figs. 7-8.
1879. Aspidoceras longispinum Sowerby, Fontannes, p. 87, Pl. 12, fig. 4.
1907. Aspidoceras longispinum Sow., Toula, p. 67, Pl. 15.
1935. Acanthosphaerites longispinum Sow., Vadász, p. 62. [in lit.]
1959. Aspidoceras longispinum Sow. var. antsavolensis nov. var., Collignon, Pl. 110, figs. 404-405.
1963. Aspidoceras longispinum (Sowerby), Pachucki, p. 2, Pl. 1, fig. 1.
1971. Aspidoceras longispinum (J. de C. Sowerby), Callomon & Cope, p. 174, Pl. 12, figs. 1-3.
1973. Aspidoceras longispinum (Sowerby), Preda, Pl. 17, fig. 3.
1978. Aspidoceras sp. aff. A. longispinum (Sowerby), Olóriz, p. 293, Pl. 24, fig. 1.
1979. Aspidoceras longispinum (J. de C. Sowerby), Sapunov, p. 140, Pl. 42, fig. 2.
1979. Aspidoceras longispinum (Sowerby), Schairer & Barthel, p. 18, Figs. 3-4, Pl. 3, figs. 3-8; Pl. 4.
1985a. Aspidoceras longispinum (Sowerby), Checa, p. 76, Fig. II.3.10-11, .15, Pl. 9, figs. 1-3; Pl. 10, fig. 2; Pl. 14, fig. 2; Pl. 15, fig. 1.
1986. Aspidoceras longispinum (Sowerby), Sarti, p. 504, Pl. 5, fig. 4.
1989. Aspidoceras longispinum (Sowerby), Checa & Martin-Ramos, Fig. 1.A.
2006. Physodoceras cf. longispinum (Sowerby), Scherzinger & Mitta, p. 230, Fig. 3.3.
2009. Aspidoceras sp. aff. A. longispinum (Sowerby), Énay, p. 194, Pl. 46, fig. 1.a-b.
2011. Aspidoceras cfr. longispinum (Sowerby), Petti et al., p. 169, Pl. 1, fig. 1.A-B.
Material. One poorly preserved whorl fragment (J 2020.274.1).
Dimensions. Due to fragmentary status, dimensions are not given.
Description. The whorl fragment represents the midflank region of a large sized specimen. Three tubercles seen. The height of the tallest tubercle is 13 mm. Tubercles are paired and widely spaced. Shell is smooth otherwise. Suture, venter and umbilicus not seen.
Remarks. Callomon and Cope (1971) refigured the holotype and emphasized the important specific characters: Rather widely spaced and paired, not very prominent tubercles that are directed outwards perpendicular to the plane of the spiral. Although the orientation of the fragment is somehow dubious, based on the above description a specific determination is possible.
Stratigraphic and geographic distribution. Aspidoceras longispinum has a long stratigraphic range from the lower Kimmeridgian Herbichi Zone to the lower Tithonian Hybonotum Zone. Geographically it has a remarkably widespread distribution from the Mediterranean and Submediterranean Provinces of the Tethysian Realm (including Nepal) with stray individuals venturing into the Subboreal Province of the Boreal Realm.
Aspidoceras rogoznicense (Zejszner, 1846)
(Fig. 9.H1-H3 ![]() )
)
1846. Ammonites Rogoznicensis, Zejszner, Pl. 4, fig. 4.a-d.
1868. Ammonites (Aspidoceras) Rogoznicensis Zeuschn., Zittel, p. 116, Pl. 24, figs. 4-5.
1870. Aspidoceras Rogoznicense Zeuschn., Zittel, p. 79, Pl. 7, fig. 1.A-B.
1871a. Aspidoceras Rogoznicense Zeuschn., Gemmellaro, p. 241, Pl. 12, figs. 7-9.
1976. Aspidoceras rogoznicense (Zeuschn.), Fülöp, p. 194, Pl. 35, fig. 7.
1978. Aspidoceras rogoniscense (Zeuschner), Olóriz, p. 279, Pl. 23, fig. 4; Pl. 24, fig. 6.
1979. Aspidoceras rogoznicense (Zejszner), Sapunov, p. 139, Fig. 14/1, Pl. 41, fig. 2.
? 1981. Aspidoceras rogoznicensis (Zeuschner), Parishev & Nikitin, p. 70, Pl. 36, fig. 4.
1984. Aspidoceras rogoznicense irregulare nov. ssp., Vigh, Pl. I, fig. 4.
1985a. Aspidoceras rogoznicense (Zeuschner), Checa, p. 98, Fig. II.3.18-19, .21, Pl. 16, figs. 1-4.
1986. Aspidoceras rogoznicense (Zeuschner), Sarti, p. 505, Pl. 6, fig. 1.A-B.
? 1989. Aspidoceras rogoznicense (Zeuschner), Főzy, Pl. 4, fig. 3.
1990. Aspidoceras rogoznicense (Zeuschner), Főzy, Pl. 2, figs. 4-6.
1998. Aspidoceras rogoznicense (Zejszner), Howarth, p. 63, Pl. 10, fig. 1.
1999. Aspidoceras rogoznicense (Zeuschner), Caracuel & Olóriz, Fig. 7.3.
2005. Aspidoceras rogoznicense (Zeuschner), Énay et al., Fig. 4.1.
2015. Aspidoceras cf. rogoznicense (Zeuschner), Berger, p. 50, Fig. 27.
2018. Aspidoceras rogoznicense (Zejszner), Vašíček et al., p. 190, Fig. 3.C-D.
Material: Ten moderately preserved complete or fragmentary internal moulds (J 2020.110.1, 2020.259.1, 2020.271.1, 2020.273.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.120.1. | 73.2 | 38.2 | 30.2 | 25.2 | 1.265 | 0.412 | 0.344 |
| J 2020.143.1. | 80.2 | (36) | 32.6 | 24.1 | 1.104 | 0.406 | 0.300 |
| J 2020.255.1. | 75.8 | (40) | (27) | 23.2 | 1.481 | 0.356 | 0.306 |
| J 2020.283.1. | (101) | (45) | (44) | 27.4 | 1.023 | 0.435 | 0.271 |
| J 2020.334.1. | 87.8 | 46.9 | (36) | (27) | 1.303 | 0.410 | 0.307 |
| J 2020.346.1. | (72) | (32) | (27) | (21) | 1.185 | 0.375 | 0.292 |
Description. Coiling rather involute, whorls inflated, always depressed or strongly depressed. Specimens represent phragmocone and body chamber. Coiling cadicone, cross section strongly depressed, always strongly wider then high. Umbilical wall high, vertical, smooth. Umbilical shoulder rounded. Flanks inflated, convex, short, ventral shoulder may exist, then rounded. Venter smooth. Strong, erected tubercles rise from the umbilical shoulder. There are 7-8 tubercles per half whorl. Below the midflank another row of tubercles appears that corresponds to the tubercles situated at the umbilical shoulder. These tubercles may overgrow to form short but wide ribs. Suture poorly preserved, but generally corresponds to the specimens of Checa (1985a, Fig. II.3.21.A-D).
Remarks. Cross section of specimen J 2020.110.1. is typical of this species: Besides the sculpture and ornamentation short, depressed, cadicone whorl sections characterise this species closely resembling the cross section of the internal whorls of specimen F.Gg.10.6 illustrated by Checa (1985a, p. 102, Fig. II.3.18.A). Specimen J 2020.120.1. is almost identical with the specimen of Énay et al. (2005, Fig. 4.1) and represents the inflated, strongly ornamented stock of the species. Whereas specimen J 2020.143.1 more resembles specimen F.Gg.10.6 of Checa (1985a, Pl. 16, fig. 4) and represents the less pronounced, closely developed tuberculated stock. Főzy (1989) figured a worn specimen with poorly preserved tubercles without cross section and description that raises ambiguity about its determination. The specimen of Parishev and Nikitin (1981) has a wide umbilicus and distantly placed tubercles, which also raises questions about their determination. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Aspidoceras rogoznicense has a long stratigraphic range from the uppermost Kimmeridgian (Beckeri Zone) to the lower Berriasian (Jacobi Zone). Geographically it has a global Tethysian distribution.
Subfamily Physodoceratinae Schindewolf, 1925
Genus Orthaspidoceras Spath, 1925
Type species: Ammonites orthocera Orbigny, 1847
Orthaspidoceras cf. ziegleri Checa, 1985a
1878. Aspidoceras liparum Oppel, Herbich, p. 174, Pl. 16, fig. 1.
1985a. Orthaspidoceras ziegleri sp. nov., Checa, p. 150, Fig. II.3.37-38, .43, Pl. 1, fig. 5; Pl. 28, figs. 1-3, Pl. 29, figs. 1-2.
1987. O. ziegleri, Pavia et al., p. 71. [in lit.]
1987. Orthaspidoceras ziegleri, Checa, p. 273, Fig. 1.C
1993. Orthaspidoceras ziegleri Checa, Sarti, p. 127. [in lit.]
1997. Orthaspidoceras gr. ziegleri Checa, Hantzpergue et al., p. 96. [in lit.]
2007. Orthaspidoceras ziegleri, Cecca & Savary, p. 511. [in lit.]
Material. Three poorly preserved and worn internal moulds (J 2020.98.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.93.1. | 72.8 | (30) | (35) | 11.2 | 0.857 | 0.481 | 0.154 |
| J 2020.116.1. | 65.0 | n.d. | (28) | 12.3 | n.d. | 0.430 | 0.189 |
Description. Small and medium sized conchs representing phragmocone and part of the body chamber. Conch involute, umbilicus narrow and profound. Umbilical wall vertical, high, unsculptured. Umbilical shoulder is rounded. Flanks convex, converging, unsculptured. No ventral shoulder. Venter is rounded, smooth. Cross section compressed, usually higher than wide. At the umbilical shoulder short, wide, gentle tubercles rise. There are 9 tubercles per half whorl. Suture partly preserved. L and U deep, trifid, frilled. Cross section compressed.
Remarks. Checa (1985a, p. 151) placed A. liparum non Oppel (Herbich, 1878, Pl. 16, fig. 1) into his new species based on the more frequent periumbilical tubercles than A. liparum. His view is accepted here. This species is uncommon in the literature and the present paper is only the third to figure typical material. Number of tubercles on the last whorl usually varies between 8-11, herein we note 9. Specimen J 2020.116.1. is closest to K.M2.20.4 of Checa (1985a, Pl. 28, fig. 2) and represents the larger umbilicated variety with finer tubercles. Due to the poor preservation none of the present specimens show the whole umbilicus, which causes uncertainty regarding the umbilical region. There are similarities with other species: Physodoceras altenense is superficially similar, however it has a narrower umbilicus with less well-developed tubercles. Aspidoceras circumspinosum has similar periumbilical tubercles, however those tubercles are spiny and sparsely placed. First record from Hungary.
Stratigraphic and geographic distribution. Orthaspidoceras ziegleri has a limited stratigraphic distribution in the Kimmeridgian from the Herbichi to the basal Cavouri zones. It occurs in the Mediterranean and Submediterranean Provinces of the Tethysian Faunal Realm.
|
|
Genus Physodoceras Hyatt, 1900
Type species. Ammonites circumspinosus Quenstedt, 1849
Physodoceras wolfi (Neumayr, 1873)
1873. Aspidoceras Wolfi nov. sp., Neumayr, p. 195 [55], Pl. 38, fig. 5.
1878. Aspidoceras Wolfi Neumayr, Herbich, p. 170 [152].
1878. Aspidoceras Báthori nov. sp., Herbich, p. 170 [152], Pl. 19, fig. 4.a-b.
1878. Aspidoceras Deáki nov. sp., Herbich, p. 175 [157], Pl. 14-15, fig. 2.
1886. Aspidoceras Deaki Herb., Pavlow, p. 76, Pl. 3, figs. 2.a-d, 3.a-b, 4.a-b.
1963. Aspidoceras (Pseudowaagenia) bathori Herbich, Geyer p. 191, Pl. 17, fig. 1.
1970. Aspidoceras (Aspidoceras) aff. wolfi Neumayr, Bernoulli & Renz, p. 598, Pl. 6, fig. 1.a-b.
1985a. Physodoceras wolfi (Neumayr), Checa, p. 134, Figs. II.3.30-31, .33, Pl. 24, fig. 5; Pl. 25, figs. 1-4.
1994. Ph. wolfi (Neumayr), Schlegelmilch, p. 128, Pl. 69, fig. 4.
1999. Physodoceras wolfi, Sarti, p. 326, Fig. 11.E-H.
2007. Physodoceras cf. wolfi (Neumayr), Cecca & Savary, p. 537, Fig. 12.A.
2011. Physodoceras wolfi (Neumayr), Főzy et al., p. 417, Fig. 2.1-2.
Material. One poorly preserved and worn internal mould from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.89.1. | 72.0 | (36) | (31) | 20.5 | 1.161 | 0.431 | 0.285 |
Description. Small sized, moderately preserved, rather compressed phragmocone conch of typical aspidoceratid coiling. Two whorls are visible. Umbilicus rather wide, deep. Umbilical wall deep, smooth, vertical. Umbilical shoulder rounded. Flanks convex, converging. No ventral shoulder. Venter rounded, smooth. Cross section subcircular. From the umbilical shoulder short, gentle tubercles rise. There are 7 tubercles on the last half whorl. Except for these tubercles no other sculpture is preserved. Neither suture nor aperture seen.
Remarks. When Neumayr (1873, p. 195) established this species he clearly stated that it is a 'strongly inflated and entirely smooth form' as well seen on his original plate. Five years later, when Herbich set up his new species (A. bathori and A. deaki), he collected from the same locality as Neumayr. Herbich was clear and strictly followed the view of Neumayr regarding this species to nominate his new species based on peri-umbilical tubercles. His view was followed by Pavlow (1886). Comparable morphospecies include Schaireria neumayri Checa, 1985a, and S. fluegeli Zeiss, 1994. The holotype of the latter is a plaster cast taken from an imprint of a large sized specimen. S. fluegeli has strongly erected tubercles ending in spines that originated from the umbilical shoulder. The other species is S. neumayri Checa, 1985a. Morphologically these species are similar, however S. neumayri have smaller, and less well-developed peri-umbilical tubercles than S. fluegeli.
Presently this species is considered to subsume the other sculptured forms with a peri-umbilical row of tubercles. The content of this species (P. wolfi) is shifting from Checa (1985a) to D'Arpa and Meléndez (2002). It would be worthwhile to make a new collection at the type locality, to morphologically discriminate these species (P. bathori, P. deaki, P. wolfi) following the original view of Neumayr. Specimen Ch.G12.R (Checa, 1985a, Pl. 24, fig. 6) represents the sculpture of P. wolfi sensu Neumayr, while L.38/5 (Checa, 1985a, Pl. 25, fig. 4) is Neumayr's refigured holotype. These two specimens represent P. wolfi according to the original species concept of Neumayr. The other specimens of Checa (1985a, Pl. 24, fig. 5; Pl. 25, figs. 1-3) reveal the peri-umbilical tubercles that vary in strength, number, and placement along the umbilical shoulder. Regarding the other figured specimens, Schlegelmilch (1994) copied Neumayr's original plate, while the specimens of Cecca and Savary (2007), and Főzy et al. (2011) represent the smooth varieties, reflecting the original species concept of Neumayr. First record from the Mecsek Mountains.
Stratigraphic and geographic distribution. Although Physodoceras wolfi is a rare species, it has a long stratigraphic range and wide geographic distribution. It is reported from the Kimmeridgian Bimammatum to the Acanthicum zones in the Submediterranean, Subboreal and Mediterranean Provinces.
Genus Pseudowaagenia Spath, 1931
Type species: Aspidoceras haynaldi Herbich, 1868
Pseudowaagenia div. sp.
Material. Seven fragmentary and poorly preserved internal moulds (J 2020.448.1-454.1, and below).
Remarks. Apart of the poor preservation, these specimens may belong to some different P. species. But lacking complete whorls, a specific assignment is unwise.
Pseudowaagenia acanthomphala (Zittel, 1870)
1870. Aspidoceras acanthomphalus Zitt., Zittel, p. 79, Pl. 5, fig. 4.A-B.
1878. Aspidoceras microplum Oppel, Herbich, p. 172, Pl. 14-15, fig. 4.
1931. Pseudowaagenia carpathica, sp. nov., Spath, p. 621, Pl. 123, fig. 5.a-c.
1966. Aspidoceras (Pseudowaagenia) serbicum n. sp., Anđelković, p. 81, Pl. 21, fig. 2; Pl. 22, fig. 2; Pl. 25, fig. 5.
1978. Aspidoceras (Pseudowaagenia) acanthomphalum acanthomphalum (Zittel), Olóriz, p. 316, Pl. 26, figs. 4-5.
1985a. Pseudowaagenia acanthomphala (Zittel), Checa, p. 127, Fig. II.3.25-26, .28-29, Pl. 23, figs. 1-4.
1989. Pseudowaagenia acanthomphala (Zittel), Főzy, Pl. 3, fig. 5.
1993. Pseudowaagenia acanthomphala (Zittel), Sarti, p. 130, Pl. 27, fig. 3.
1993b. Pseudowaagenia acanthomphala (Zitt.), Főzy, p. 199. [in lit.]
1999. Pseudowaagenia acanthomphala (Zittel), Caracuel & Olóriz, Fig. 7.9.
2011. Pseudowaagenia acanthomphala (Zittel), Főzy et al., Fig. 3.4-.5.
Material: Eight internal moulds, one specimen from the quarry floor amongst debris, others from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.90.1. | 24.8 | (8) | 10.5 | (9) | 0.762 | 0.423 | 0.363 |
| J 2020.145.1. | 73.3 | (19) | 23.4 | 32.5 | 0.812 | 0.319 | 0.443 |
| J 2020.146.1. | 86.4 | 20.6 | (25) | 38.6 | 0.824 | 0.289 | 0.447 |
| J 2020.260.1. | 90.6 | 21.4 | (27) | (45) | 0.793 | 0.298 | 0.497 |
| J 2020.261.1. | 70.6 | (18) | (21) | 32.7 | 0.857 | 0.297 | 0.463 |
| J 2020.272.1. | 70.5 | (25) | (28) | (25) | 0.893 | 0.397 | 0.355 |
| J 2020.332.1. | 94.4 | (23) | (28) | (47) | 0.821 | 0.297 | 0.498 |
| J 2020.455.1. | 101.2 | (21) | (30) | 47.2 | 0.700 | 0.296 | 0.466 |
Description. Small and medium sized specimens representing septate phragmocones and body chambers. Coiling evolute, one to three whorls visible. Umbilicus wide and shallow. Umbilical wall high, smooth and vertical on the inner whorls, while shallow and oblique on the outer whorls. Umbilical shoulder not rounded in the inner and fairly rounded on the last whorl. On the umbilical shoulder of inner whorls, fine tubercles emerge that are elongated rectoradiately. There are 14-16 tubercles per half whorl on the inner whorls. Tubercles become less frequent toward the body chamber and usually disappear beyond the last septum. Otherwise, tubercles become fainter and are located farther apart. Flanks smooth, parallel or convex, unsculptured. On the last whorl, close to the aperture and beyond the last septum, fine ribs emerge from the tubercles and travel through the flanks, crossing the venter continuously as Herbich (1878, p. 140 - Hungarian version) noted. Cross section rounded rectangular, compressed. Suture not seen.
Remarks. Checa (1985a, p. 129) included Aspidoceras (Pseudowaagenia) serbicum n. sp. of Anđelković (1966, p. 81) in this species based on homologous phenotypic characters and the stratigraphic revision of its locus typicus by Sapunov and Ziegler (1976, p. 28). The Aspidoceras microplum of Herbich (1878, p. 172, Pl. 14-15, fig. 4) was also included in this species by Checa (1985a, p. 127) based on its phenotypic characters, and his opinion is accepted here. Főzy et al. (2011, Fig. 3.4-.5) refigured the specimen of Főzy (1989, Pl. 3, fig. 5) in better quality. Specimen J 2020.146.1 is closest to F.G23.11.8 of Olóriz (1978, Pl. 26, fig. 4) whereas specimen J 2020.145.1. is closer to specimen M.1631/b illustrated by Anđelković (1966, Pl. 21, fig. 2).
Microconchs with well developed lappets have been reported rarely among aspidoceratine ammonites (e.g., Scherzinger et al., 2018, Fig. 3.A, .C), therefore even in the case of numerous specimens the macro- or microconchs are difficult to recognize.
Stratigraphic and geographic distribution. Pseudowaagenia acanthomphala is reported from the upper Kimmeridgian (Cavouri to Beckeri zones) from the western Tethys (Mediterranean Province) and adjacent territory (Submediterranean Province).
Pseudowaagenia cf. haynaldi (Herbich, 1868)
1868. Ammonites Haynaldi Herb. nov. sp., Herbich, p. 33.
1873. Aspidoceras Haynaldi Herbich, Neumayr, p. 194 [54], Pl. 42, fig. 3.a-b.
1876. Ammonites Haynaldi Herbich, Dumortier & Fontannes, p. 122, Pl. 17, fig. 4, Pl. 18, figs. 2-3.
1878. Aspidoceras Hajnaldi Herbich, Herbich, p. 137, Pl. 14-15, fig. 1.A-B.
1931. Pseudowaagenia haynaldi (Herbich MS), Spath, p. 621, Pl. 91, fig. 1.a-b; Pl. 118, fig. 11.
1935. Pseudowaagenia cf. haynaldi Herb., Vadász, p. 62. [in lit.]
1936. Aspidoceras haynaldi Herbich, Besairie, p. 132, Pl. 9, fig. 2.
1978. Aspidoceras (Pseudowaagenia) haynaldi haynaldi (Herbich), Olóriz, p. 308, Pl. 27, fig. 1.
1985a. Pseudowaagenia haynaldi (Herbich), Checa, p. 126, Fig. II.3.25, .27, Pl. 21, figs. 7-10
1999. Aspidoceras (Pseudowaagenia) cf. haynaldi (Herbich), Fatmi & Zeiss, p. 87, Pl. 24, fig. 2.
2018. Aspidoceras (Pseudowaagenia) haynaldi, Énay, p. 77, Pl. 1, fig. 8.
Material. One internal mould.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.191.1. | 50.7 | 21.7 | (21) | (13) | 1.033 | 0.414 | 0.256 |
Description. Small sized conch representing the phragmocone and partial body chamber. Umbilicus wide, shallow. Umbilical wall short, vertical, unsculptured. Umbilical shoulder rounded. Flanks parallel, unsculptured. No ventral shoulder, venter rounded, smooth. Cross section high oval, with flattened flanks. Sculpture consists of fine, distantly spaced short tubercles at the umbilical shoulder. Tubercles only seen on last half whorl.
Remarks. Herbich (1868, p. 33) introduced this new species with only a short qualifying remark: 'Perarmatus family, next to A. eucyphus Opp. but definitely differs. From grey marl at Gyilkos-kő.' Neumayr (1873, Pl. 42, fig. 3.a-b) refigured the original specimen of Herbich, that reveals a wide umbilicus and a row of fine peri-umbilical tubercles, showing 10 of them on the last half whorl. According to the analysis of Checa (1985a, p. 127) this species shows characters transitional between P. micropla and P. acanthomphala, however there is strong subjectivity between different authors when describing internal whorls and juvenile stages. The specimen MNHN.F.J07898 of Besairie (1936) has 7 tubercles on the last half whorl distributing the same fine radial ribbing as the holotype figured by Neumayr (1873). These characters are only poorly preserved on the collected specimen.
Stratigraphic and geographic distribution. Pseudowaagenia haynaldi is reported from the upper Kimmeridgian (Acanthicum to Beckeri zones) mainly from the Submediterranean Province with occasional occurrences in the Mediterranean Province and from the northern Tethys, however stray individuals are also reported from the Indo-Malagasy Province.
Pseudowaagenia inerme Énay, 2018
2018. Aspidoceras (Pseudowaagenia) inerme n. sp., Énay, p. 78, Pl. 1, figs. 9-10; Pl. 2, figs. 2-3.
Material. One poorly preserved internal mould.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.570.1. | n.d. | 24.1 | (29) | (41) | 0.831 | n.d. | n.d. |
Description. Medium sized conch representing phragmocone and body chamber. Coiling evolute, two whorls seen, inner whorls lost. Umbilicus wide, shallow. Umbilical wall short, vertical, smooth. Umbilical shoulder rounded. Flanks convex, converging. No ventral shoulder. Venter rounded, smooth. Cross section compressed, high oval. Fine radial folds rise at midflank to become prorsiradiate on upper flank and cross the venter continuously. Thirteen folds are seen on the better-preserved ventral part, however there are more that are lost to the poor preservation. Folds are unequal and randomly placed. Neither suture not aperture apparent.
Remarks. Our Hungarian specimen is comparable to the holotype illustrated by Énay (2018, Pl. 1, fig. 9.a-c, FSL 341349): By comparison their cross section, fine ribbing and rate of coiling are nearly identical. But the collected specimen has poorer preservation and therefore fewer folds are discernible. First record from Hungary.
Stratigraphic and geographic distribution. Upper Kimmeridgian Beckeri Zone, Jura Mountains, France.
Pseudowaagenia micropla (Oppel, 1863)
1863. Ammonites microplus Opp., Oppel, p. 218, Pl. 58, fig. 4.a-b.
1877. Ammonites (Aspidoceras) microplus Oppel, Favre, p. 63, Pl. 7, fig. 4.
non 1878 Aspidoceras microplum Oppel, Herbich, p. 172, Pl. 14-15, fig. 4. [=P. acanthomphala]
1878. Aspidoceras microplum Opp., Gemmellaro, p. 248, Pl. 15, fig. 11.
1879. Aspidoceras microplum Oppel, Fontannes, p. 92, Pl. 12, figs. 11-13.
1931. Pseudowaagenia micropla (Oppel), Spath, p. 622, Pl. 122, fig. 3.a-c.
1966. Aspidoceras (Pseudowaagenia) microplus (Oppel), Anđelković, p. 80, Pl. 30, fig. 5.
1978. Aspidoceras (Pseudowaagenia) microplum (Oppel), Olóriz, p. 313, Pl. 27, fig. 2.
1984. Aspidoceras (Pseudowaagenia) cfr. microplum (Oppel), Rossi, p. 109, Pl. 34, fig. 2.
1985a. Pseudowaagenia micropla (Oppel), Checa, p. 116, Fig. II.3.24, .27, Pl. 21, figs. 2-6.
1986. Aspidoceras (Pseudowaagenia) microplum (Oppel), Sarti, p. 506, Pl. 6, fig. 2.
1987. Pseudowaagenia micropla (Oppel), Pavia et al., Pl. 3, fig. 3.
Material. Four internal moulds (J 2020.359.1, and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.153.1. | 50.7 | 19.6 | (21) | (13) | 0.933 | 0.414 | 0.256 |
| J 2020.205.1. | (57) | (18) | (22) | (13) | 0.818 | 0.368 | 0.228 |
| J 2020.590.1. | 53.8 | 24.3 | (23) | (17) | 1.056 | 0.427 | 0.316 |
Description. Poorly preserved small sized conchs representing the phragmocone. Umbilicus moderate, shallow. Suture, aperture not seen. Umbilical wall short, smooth and oblique. Umbilical shoulder rounded. Flanks converge, smooth. No ventral shoulder, venter smooth and rounded. Cross section is high oval. Fine tubercles rise at the umbilical shoulder. There are 11, tiny, pearl-like peri-umbilical tubercles on the last whorl. Exactly on midflank, another row of small tubercles is occasionally apparent. If present, they usually number 5 on the last whorl. Rarely, some of the tubercles are radially elongated toward the upper flank. In some cases, very fine, radial ribs rise on the upper flank from some of the midflank tubercles and travel radially toward the venter. Venter poorly preserved; sutures and aperture not apparent.
Remarks. Intraspecific variability within this species lays in the changing number and dimensions of periumbilical tubercles on the last whorl (it is 13 for Oppel, 12 for Checa [1985a, Pl. 21, fig. 3], 8 noted by Favre and Spath, 6 by Rossi) and presence/absence and numbers of midflank tubercles as thoroughly discussed and documented by Checa (1985a). Specimen J 2020.205.1. is closer to the holotype of Oppel (1863, Pl. 58, Fig. 4.a-b), the only differences are that the type specimen has more periumbilical nodes with a more slender cross section. Whorl section of our specimen is closest to specimen F.G12.6.8 of Checa (1985a, Fig. II.3.24.C) representing the attenuated variety of the species. Specimen J 2020.153.1. is closest to specimen M54 (1950 I 97) of Checa (1985a, Pl. 21, fig. 6), the only difference is that the Hungarian specimen has 5 tubercles on the midflank of the last half whorl, while it is 6 for M54; otherwise they are comparable. First record from Hungary.
Stratigraphic and geographic distribution. Pseudowaagenia micropla has a long stratigraphical range from the Bimammatum to the Acanthicum zones of the Kimmeridgian in the western Tethys (Mediterranean Province) and adjacent territories (Submediterranean Province).
Genus Simaspidoceras Spath, 1925
Type species. Aspidoceras argobbae Dacqué, 1905
Simaspidoceras bucki Checa, 1985a
1985a. Simaspidoceras bucki sp. nov., Checa, p. 175, Fig. II.3.42, .45-47, Pl. 36, figs. 1-2; Pl. 37, figs. 1-2.
1987. Simaspidoceras bucki Checa, Pavia et al., Pl. 7, fig. 1.
Material. One half whorl of an internal mould from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.186.1. | 89.6 | (40) | (42) | (21) | 0.952 | 0.469 | 0.234 |
Description. Fairly preserved, middle sized conch fragment representing both body chamber and phragmocone. Umbilicus wide, shallow. Umbilical wall oblique, smooth. Umbilical shoulder rounded. Flanks convex, converging, smooth. No ventral shoulder. Venter rounded, smooth, flat. Cross section rounded triangular. At the umbilical shoulder, occasional, not regular, gentle, short tubercles rise. Tubercles continue in short, wide, gentle ribs ending above the midflank. Four fine ribs seen on half whorl. Upper flank and venter smooth, ribs disappear. Suture, aperture not seen.
Remarks. In the evolutionary summary of the aspidoceratine Checa and Olóriz (1987, Fig. 3) described this species as 'development of wide, expanded at the lateroventral portion, ribs.' The present specimen corresponds well with this description being closest to specimen F.X.3 of Checa (1985a, Pl. 36, fig. 2) with even and finer tubercles and ribbing. The closest morphospecies is Simaspidoceras bauschi Checa, 1985b (op. cit., Pl. 1; specimen BSP 1980 XV 13) having stronger umbilical tubercles, wider ribs of significantly bigger size. Pavia et al. (1987, Pl. 7, fig. 1.a-b) published a specimen with stronger ribbing and tubercles. Cross section of the collected specimen is closest to specimen F.G2.22.11 of Checa (1985a, Fig. II.3.42.C), however the Hungarian example is more compressed, which is a decisive character for the species (Checa, 1985b, p. 24). First record from Hungary.
Stratigraphic and geographic distribution. Simaspidoceras bucki has a limited stratigraphic distribution in the Kimmeridgian (Herbichi to Acanthicum zones). Geographically it rarely occurs in the Mediterranean (Italy) and the Submediterranean Provinces (Spain, Hungary) and it is abundant in the Indo-Malagasy Province (Yemen, Somalia, Kenya, Malagasy) as noted by Énay and Howarth (2019).
Genus Hybonoticeras Breistroffer, 1947
Type species. Ammonites Hybonotum Oppel, 1863
Hybonoticeras pressulum (Neumayr, 1871b)
1871b. Asp. pressulum nov. sp., Neumayr, p. 25.
1873. Aspidoceras pressulum Neumayr, Neumayr, p. 201 [61], Pl. 37, figs. 2-3.
1878. Aspidoceras Verestoicum nov. sp., Herbich, p. 181 [163], Pl. 14-15, fig. 3.
1935. Pseudowaagenia cf. pressulum Neum., Vadász, p. 62. [in lit.]
1959. Hybonoticeras pressulum (Neumayr), Berckhemer & Hölder, p. 20, Fig. 2, Pl. 1, figs. 6, 8; Pl. 2.
? 1963. Hybonoticeras cf. pressulum (Neumayr), Pachucki, p. 3, Pl. 1, fig. 6.
1978. Hybonoticeras pressulum (Neumayr), Barthel & Schairer, p. 18, Pl. 2, figs. 4-7.
1978. Hybonoticeras (Hybonoticeras) pressulum pressulum (Neumayr), Olóriz, p. 355, Pl. 32, fig. 5.
1984. Hybonoticeras pressulum (Neumayr), Verma & Westermann, p. 75, Pl. 19, fig. 1.
1984 Hybonoticeras (Hybonoticeras) pressulum verestoicum (Herbich), Rossi, p. 118, Pl. 34, figs. 1, 4.
1989. Hybonoticeras pressulum subsp. B Berckhemer & Hölder, Malinowska, Pl. 5, fig. 7.
1991. Hybonoticeras pressulum (Neumayr), Krishna & Pathak, p. 11, Pl. 1, fig. 1; Pl. 2, fig. 4.
1993. Hybonoticeras pressulum (Neumayr), Sarti, p. 132, Pl. 28, fig. 1.A-B.
1995. Hybonoticeras pressulum (Neumayr), Főzy, p. 138, Pl. 22, figs. 6, 8.
1996. Hybonoticeras pressulum (Neumayr), Schweigert el al., p. 353, Fig. 3.a-b.
1999. Hybonoticeras sp. gr. pressulum (Neumayr), Caracuel & Olóriz, p. 586, Fig. 6.6.
2013. Hybonoticeras pressulum (Neumayr), Főzy & Scherzinger, p. 188, Pl. 10, figs. 2-3.
Material. 15 fragmentary, poorly preserved specimens of internal moulds from old collection (MBFSz) and from mixed debris and soil (J 2087, J 2020.95.1, 96.1, 229.1, 251.1-254.1, 294.1, 296.1, 331.1, 591.1 and below).
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2088 | 70.1 | 21.4 | 24.2 | 28.5 | 0.884 | 0.345 | 0.406 |
| J 2020.262.1. | 75.4 | (13) | (25) | (35) | 0.520 | 0.332 | 0.464 |
| J 2020.271.1. | 88.3 | 21.1 | (26) | (37) | 0.812 | 0.294 | 0.419 |
Description. Medium sized conch, however estimation of the whorl fragments indicates that the diameter of the entire conch was up to 20 cm representing both body chambers and phragmocones. Coiling evolute, however only one crushed specimen has one entire whorl but none of them show preserved inner whorls. Cross section rounded trapezoidal, compressed. Umbilicus wide, shallow. Umbilical wall steep, vertical, smooth. Umbilical shoulder rounded; flanks slightly converge. Ventral shoulder rounded. Venter bears a longitudinal shallow groove. At the umbilical shoulder strong, pronounced, vertically placed spines appear on early growth stages. Later the spines become slightly rectoradiate and give rise to smooth, falcoid ribs that cross the flank. Suture not seen.
Remarks. Due to the poor preservation and lack of the early growth stages of inner whorls, the juvenile bituberculate stage (sensu Schweigert et al., 1996, p. 354) is not evident for us. H. pressulum is very close to P. acanthomphala the only difference is the absence/presence of the ventral groove as remarked upon by Schweigert and Schlampp (2020, p. 30). The species palaeontological designation involving the missing holotype and erroneously designated lectotype has been excellently discussed by Schweigert et al. (1996, p. 354), as well as morphologically inseparable Hybonoticeras verestoicum (Herbich), which is included in H. pressulum by these authors. Their view is accepted and followed here, although there is some dissent from those authors who continue to recognize separate and independent species (e.g., Olóriz & Villaseńor, 1999; Boughdiri et al., 2009; Grigore, 2011). However, some etymological remarks are worthy of mention here. Neumayr (1871b) collected his specimen at Gyilkos-kő in Nagy-Hagymás Mountains, and from the same bed, Herbich (1878) collected his specimen described as Aspidoceras verestoicum. The original Hungarian phrase 'Veres-tó' means red lake in English (in Romanian Lacul Roşu) referring to the lake in neighborhood of Gyilkos-kő. The locality is referred to as Lacul Roşu in the recent Romanian literature. Specimens J 2020.95.1 and 2020.96.1 are most comparable to the specimen of Neumayr (1873, Pl. 37, fig. 3.a-b) representing the big sized, smooth stage of ontogeny with elongated umbilical tubercles and ventral groove. Pachucki (1963) reported a specimen with strong, S-shaped prorsiradiate ribs that fall out of specific variation with this species, rendering the designation as ambiguous.
Stratigraphic and geographic distribution. Hybonoticeras pressulum has a limited stratigraphic range in the upper Kimmeridgian (Beckeri Zone), however it has a wide geographical distribution throughout the entire Tethys.
|
|
Superfamily incertae sedis
Ammonite gen. et sp. ind.
Material. Two poorly preserved internal moulds (J 2020.136.1, 2020.217.1) from mixed debris and soil.
Dimensions.
| specimen | D | Wb | Wh | U | Wb/Wh | Wh/D | U/D |
| J 2020.136.1. | 85.9 | (23) | (28) | (31) | 0.821 | 0.326 | 0.361 |
Descriptions.
Specimen J 2020.136.1. (Fig. 11.B2 ![]() ). The early whorls of this specimen superficially resemble to Simoceratidae (?Volanoceras) however the outer whorls are either lost or dissolved and fractured.
). The early whorls of this specimen superficially resemble to Simoceratidae (?Volanoceras) however the outer whorls are either lost or dissolved and fractured.
Specimen J 2020.217.1. (Fig. 11.B1 ![]() ). The whorl fragment refers to an involute, coarsely ribbed specimen with no recognized affinity.
). The whorl fragment refers to an involute, coarsely ribbed specimen with no recognized affinity.
Remarks. Preservation of these specimens are so poor, that even suprageneric assignment is dubious. On the other hand, they are interesting and present such features that may refer to rare/not yet described taxa, therefore they are included in this study.
Other faunal elements
Although ammonites dominate the assemblage, other faunal elements also occur. Belemnites are present, as well as aptychi. Regarding the occurrence of aptychi it is remarkable that aptychi in the most cases are found fossilized together with Taramelliceras. Solitary aptychi are rare (Table 2). Belemnites are always worn and fragmented specimens that may refer to reworking. Other faunal elements are extremely rare.
Aptychi. Table 2 summarizes the recognized aptychi that represent two of the seven aptychi types of Parent et al.
(2014), i.e., lamellaptychi and laevaptychi. This is not surprising because Lamellaptychus are associated with haploceratoids, while Laevaptychus with aspidoceratoids. These ammonite groups dominate the Zengővárkony assemblage (61%). It is remarkable that Strigogranulaptychus was not collected, however it is associated with perisphinctoids that have a total share of 31.5% in the ammonite assemblage. On the other hand, it is noteworthy that Praestriaptychus, which is typical of perisphinctoids are mainly known from clayey facies (Mikhail
Rogov personal comm.). Vadász (1935, p. 62-62) listed a Laevaptychus latus (Parkinson, 1811), a Lamellaptychus sp. and a Punctaptychus? punctatum (Voltz, 1837). These records are partly confirmed by our observations (Fig. 11.C-D ![]() ). In the case of Lamellaptychus, a species was recognized: L. sp. aff. murocostatus Trauth, 1938 (Fig. 11.D
). In the case of Lamellaptychus, a species was recognized: L. sp. aff. murocostatus Trauth, 1938 (Fig. 11.D ![]() ) being the first record from the Mecsek Mountains. Nagy (1971) listed other species: Punctaptychus cf. punctatus (Voltz), Aptychus crassicauda Quenstedt, and Lamellaptychus beyrichi (Oppel). These data are not confirmed by our findings.
) being the first record from the Mecsek Mountains. Nagy (1971) listed other species: Punctaptychus cf. punctatus (Voltz), Aptychus crassicauda Quenstedt, and Lamellaptychus beyrichi (Oppel). These data are not confirmed by our findings.
Table 2: Aptychi from the Zengővárkony upper Jurassic locality: Dimensions, orientation of valves, and accompanying fossils.
| specimen | Taxon | orientation of valve | Dimensions | accompanying fossils |
| J 2020.92.2 | Aptychus sp. ind. | right, inside | L: - mm; S: - mm; Lat: 17 mm. | Ataxioceratidae gen. et sp. ind. |
| J 2020.161.1 | Laevaptychus latus | left, outside | L: - mm; S: 29 mm; Lat: 24.7 mm. | no |
| J 2020.221.1 | Lamellaptychus sp. | left, outside | L: 9.3 mm; S:8.7 mm; Lat: 4.2 mm. | Taramelliceras sp. |
| J 2020.221.2 | Lamellaptychus sp. | right, outside | L: 8.6 mm; S: 7.7 mm; Lat: 3.8 mm. | Taramelliceras sp. |
| J 2020.230.1 | Lamellaptychus murocostatus | left, outside | L: - mm; S: 47 mm; Lat: 26 mm. | no |
| J 2020.244.1 | Laevaptychus latus | right, inside | L: - mm; S: 47 mm; Lat: 27 mm. | no |
| J 2020.257.1 | Lamellaptychus murocostatus | left, outside | L: 53.7 mm; S: 48.4 mm; Lat: 23 mm. | no |
| J 2020.258.1 | Laevaptychus latus | left, outside | L: 59 mm; S: 48 mm; Lat: 39 mm. | no |
| J 2020.277.2 | Lamellaptychus sp. | left, outside | L: 39 mm; S: 32 mm; Lat: 17 mm. | Taramelliceras sp. |
| J 2020.293.1 | Laevaptychus latus | right, outside | L: 65 mm; S: 58.9 mm; Lat: 38.1 mm. | no |
| J 2020.303.1 | Lamellaptychus sp. | left, outside | L: 36 mm; S: 32 mm; Lat: 21 mm. | Ataxioceratidae gen. et sp. ind. |
| J 2020.339.3 | Lamellaptychus sp. | left, outside | L: 28 mm; S: 23 mm; Lat: 11 mm. | Taramelliceas pugile |
Belemnites. Poorly preserved belemnite guards are also collected (J 2020.159.1, 160.1, 162.1, 256.1, 304.2, 490.2). The figured specimen (Fig. 11.ED ![]() ) most probably a Hibolithes semisulcatus Münster.
) most probably a Hibolithes semisulcatus Münster.
Brachiopods. A Kimmeridgian brachiopod from the Mecsek Mountains (J 2020.244.3) is recorded as a weathered small sized specimen fossilized together with an aptychus and a Taramelliceras sp. (Fig. 11.F1-F2 ![]() ).
).
The studied material clearly shows the signals of prolonged surface weathering, with exposure resulting in abrasion and erosion. Therefore, analysis of the original taphonomic marks is difficult and needs a very optimistic albeit careful approach. It is also noteworthy that due to the absence of stratigraphic control all conclusions are time-averaged for the whole stratigraphic interval. Condensation or reworking is only surely recognized when the weathered cobbles are preserving more ammonites. Figure 11.G ![]() shows a nice example of certain condensation of ammonites, but specimens J 2020.324.1-2 are of two taramelliceratids. From the taphonomic point of view, the most interesting finding is the encrusting serpulid tube fossil, Spiraserpula spirolinites (Fig. 5.A
shows a nice example of certain condensation of ammonites, but specimens J 2020.324.1-2 are of two taramelliceratids. From the taphonomic point of view, the most interesting finding is the encrusting serpulid tube fossil, Spiraserpula spirolinites (Fig. 5.A ![]() ). It is noteworthy, that serpulid and sabellid tube worms often encrusted the inner wall of cephalopods, usually nautilids (Luci et al.,
2013) but ammonoids too (Aguirre-Urreta & Rawson,
2010) as in our case. Later the shell was redeposited by storm agitation or marine mass movements into the basin, as Radwańska
(2004) has demonstrated. The majority of ammonites are fragments, but this is probably due to later surficial erosion of the host rocks and not related to the primary reworking or multiple transportation.
). It is noteworthy, that serpulid and sabellid tube worms often encrusted the inner wall of cephalopods, usually nautilids (Luci et al.,
2013) but ammonoids too (Aguirre-Urreta & Rawson,
2010) as in our case. Later the shell was redeposited by storm agitation or marine mass movements into the basin, as Radwańska
(2004) has demonstrated. The majority of ammonites are fragments, but this is probably due to later surficial erosion of the host rocks and not related to the primary reworking or multiple transportation.
The reported ammonite taxa in this study with known stratigraphic distributions from other localities is summarized in Figure 12 ![]() . Based on the distributional data of the species to which we compare, this assemblage could represent six ammonite zones of the Kimmeridgian and the lower Tithonian. The standard Tethysian ammonite zones are applied here sensu Wierzbowski et al.
(2016), and Hesselbo et al.
(2020).
. Based on the distributional data of the species to which we compare, this assemblage could represent six ammonite zones of the Kimmeridgian and the lower Tithonian. The standard Tethysian ammonite zones are applied here sensu Wierzbowski et al.
(2016), and Hesselbo et al.
(2020).
|
|
Although the localities on the left bank of the Vasbányavölgy creek exposes a continuous sequence of Kimmeridgian strata, the present material did not contain elements from the Silenum Zone. Situated 25 m distant to the NE from the present locality there is an outcrop that corresponds to the wall of the abandoned small quarry and exposes the older part of the Kimmeridgian.
Lower Kimmeridgian Metahaploceras strombecki Zone
This zone was introduced by Olóriz (1978) assigning Taramelliceras (Metahaploceras) strombecki as a zonal marker. The presence of some specimens of the zonal index in the assemblage clearly indicates the presence of this zone as the oldest possible age for the assemblage. It corresponds more or less to the standard Tethysian Ataxioceras hypselocyclum Zone.
Lower Kimmeridgian Praesimoceras herbichi Zone
This zone was introduced by Sarti (1993) for the Venetian Alps, Italy. According to Sarti (1993, p. 41) this is an assemblage zone of Praesimoceras, species of which are ubiquitous and abundant. The presence of some fairly preserved Streblites tenuilobatus and Praesimoceras cf. herbichi clearly indicate this zone. It corresponds to the standard Tethysian Crussoliceras divisum Zone.
Upper Kimmeridgian Aspidoceras acanthicum (standard Tethysian) Zone
This zone was introduced by Neumayr (1873) and emended in a modern sense by Ziegler (1964). It is only represented by some specimens of A. acanthicum. According to Pavia et al. (1987) the zone is characterized by the FADs of Aspidoceras longispinum and Ceratosphinctes rachistrophus. The presence of these two referred Aspidoceras species acceptably indicates the presence of the Acanthicum Zone. All the other ammonites refer only partly to this zone.
Upper Kimmeridgian Mesosimoceras cavouri Zone
The Cavouri Zone was introduced by Olóriz (1978) as a taxon-range zone for the Submediterranean Province. Mesosimoceras cavouri FAD is at a somewhat higher level than the beginning of the zone, and appears in the Beckeri Zone. Hemihaploceras nobile also appears first in the Cavouri Zone. However, the presence of Aspidoceras caletanum supports the presence of the Cavouri Zone here. The ammonite species A. caletanum is a subzonal index species in the Submediterranean Faunal Province (Hantzpergue, 1989) and in South Germany as the middle subzone of the Eudoxus Zone. It is more or less equivalent to the standard Tethysian Aulacostephanus eudoxus Zone.
Upper Kimmeridgian Hybonoticeras beckeri (standard Tethysian) Zone
This zone was originally introduced by Neumayr (1873). Although the zonal marker ammonite was not revealed in our study, some other characteristic ammonites were collected: Only Hybonoticeras pressulum and Pseudowaagenia acanthomphala represent this zone. In addition the occurrence of the rare and newly described species, Pseudowaagenia inerme also strengthens the presence for the Beckeri Zone in the Zengővárkony area.
Lower Tithonian Hybonoticeras hybonotum (standard Tethysian) Zone
This zone was introduced by Benecke (1866) by the FAD of its zonal marker species. Although H. hybonotum was not found, the Tithonian is surely indicated by the presence of Silicisphinctes, Malagasites, and Gravesia. Although Aspidoceras rogoznicense is generally considered a Tithonian species, it appears already in the Beckeri Zone, therefore its presence in this mixed fauna is not decisive for proving the Tithonian. Among Ataxioceratidae div. sp. there are certainly other stratigraphically important taxa, however the fragmentary and poorly preserved specimens did not allow a reliable designation. Further collecting can probably equivocally solve the question of the presence of younger Early Tithonian ammonites in this assemblage. The most interesting faunal element is the Gravesia aff. gigas that indicates the basal Tithonian, however in certain places in Germany it already appears in the upper Kimmeridgian (Schweigert, 1999). In the Franco-German Biome it also appears in the uppermost Kimmeridgian (Autissiodorensis Subzone, Autissiodorensis Zone: Cope & Etches, 2020). G. gigas is therefore a very important species for long-distance correlation (Schweigert, 2005).
Table 3: Ammonite suprageneric taxa (families) and specimen numbers of the Zengővárkony fauna examined in the present paper.
| Taxa | Number of specimens | Percentage |
| Phylloceratidae | 40 | 7.9 |
| Lytoceratidae | 22 | 4.3 |
| Oppeliidae | 170 | 33.6 |
| Perisphinctidae | 22 | 4.3 |
| Ataxioceratidae | 136 | 26.8 |
| Aspidoceratidae | 115 | 22.7 |
| Superfamily incertae sedis | 2 | 0.4 |
| TOTAL | 507 | 100.0 |
Figure 13 ![]() summarizes the palaeobiogeographical affinities of the Zengővárkony Kimmeridgian ammonite fauna compared to some selected, mainly Tethysian localities. The late Kimmeridgian ammonite fauna of the Mecsek Mountains has a Mediterranean character based on qualitative comparison and being similar to the fauna of the Venetian Alps, northern Italy. It is noteworthy that the phylloceratid and lytoceratid specimens contribute only 12% to the total ammonite assemblage
(Table 3). Other Mediterranean localities (Sicily, Serbia, Bulgaria, Dacides: Romania, and the Gerecse Mountains, Hungary) are similar too, however almost the same number of common species are recognized with the Submediterranean localities in SE France, and SE Spain. The most remarkable feature of the Mecsek late Kimmeridgian ammonite fauna is the slight influence of the Submediterranean Province, which is reflected in
Figure 13
summarizes the palaeobiogeographical affinities of the Zengővárkony Kimmeridgian ammonite fauna compared to some selected, mainly Tethysian localities. The late Kimmeridgian ammonite fauna of the Mecsek Mountains has a Mediterranean character based on qualitative comparison and being similar to the fauna of the Venetian Alps, northern Italy. It is noteworthy that the phylloceratid and lytoceratid specimens contribute only 12% to the total ammonite assemblage
(Table 3). Other Mediterranean localities (Sicily, Serbia, Bulgaria, Dacides: Romania, and the Gerecse Mountains, Hungary) are similar too, however almost the same number of common species are recognized with the Submediterranean localities in SE France, and SE Spain. The most remarkable feature of the Mecsek late Kimmeridgian ammonite fauna is the slight influence of the Submediterranean Province, which is reflected in
Figure 13 ![]() by A. caletanum, however, other Subboreal/Submediterranean elements are there, too: Gravesia gigas (for the earliest Tithonian) and Pseudowaagenia inerme for the late Kimmeridgian. It reflects the connections of the Mecsek "Zone" towards its original European roots inherited from the Triassic and Early Jurassic. Further collection may elucidate this recognized biogeographical connection deeper. The weak biogeographical similarities between the Mecsek Mountains and the geographically close Gerecse Mountains of the Transdanubian Middle Ranges are not surprising and strengthens the previous palaeogeographical reconstructions for the Jurassic (Yilmaz et al.,
1996; Hinsbergen et al., 2020). At this time the separation of the Tisza microplate or megaunit (including Mecsek tectonic unit) from stable Europe was an ongoing process by the Alpine Atlantic Ocean (Gawlick & Missoni,
2019) that may have filtered or decreased the chance of invasion of deep water phyllo- and lytoceratid ammonites to occupy this tectonic unit. Due to the shallower water depth the ecological conditions were not yet ideal to these deep-water cephalopods, however deepening basin later resulted the increase of phyllo- and lytoceratid ratio during the Early Cretaceous (Bujtor,
1993). It is noteworthy that during the Late Oxfordian - Early Kimmeridgian the water depth was comparatively deep as demonstrated by Bujtor and Albrecht
(2021) on the existence of brachiopods (Nucleata and Pygope) preferred deep-water habitats.
by A. caletanum, however, other Subboreal/Submediterranean elements are there, too: Gravesia gigas (for the earliest Tithonian) and Pseudowaagenia inerme for the late Kimmeridgian. It reflects the connections of the Mecsek "Zone" towards its original European roots inherited from the Triassic and Early Jurassic. Further collection may elucidate this recognized biogeographical connection deeper. The weak biogeographical similarities between the Mecsek Mountains and the geographically close Gerecse Mountains of the Transdanubian Middle Ranges are not surprising and strengthens the previous palaeogeographical reconstructions for the Jurassic (Yilmaz et al.,
1996; Hinsbergen et al., 2020). At this time the separation of the Tisza microplate or megaunit (including Mecsek tectonic unit) from stable Europe was an ongoing process by the Alpine Atlantic Ocean (Gawlick & Missoni,
2019) that may have filtered or decreased the chance of invasion of deep water phyllo- and lytoceratid ammonites to occupy this tectonic unit. Due to the shallower water depth the ecological conditions were not yet ideal to these deep-water cephalopods, however deepening basin later resulted the increase of phyllo- and lytoceratid ratio during the Early Cretaceous (Bujtor,
1993). It is noteworthy that during the Late Oxfordian - Early Kimmeridgian the water depth was comparatively deep as demonstrated by Bujtor and Albrecht
(2021) on the existence of brachiopods (Nucleata and Pygope) preferred deep-water habitats.
|
|
Regarding the puzzling presence of ?Malagasites denseplicatus in the Mecsek Mountains as an element of the Indo-Madagascan Province, is more doubtful and perhaps unlikely. It is noteworthy that other faunal connections were present during the Kimmeridgian between these two regions as Caswell (1958) has already presented. According to Caswell (1958, p. 27): "The Kimmeridgian ammonite fauna of Kenya has more in common with that of Europe than that of India, a feature shared by the entire mollusk and brachiopod assemblage. Having only two, poorly preserved specimens, their real taxonomic relationships cannot be surely recognized".
The Kimmeridgian and early Tithonian ammonite dominated assemblage collected from scree and mixed soil and debris from the Kisújbánya Limestone Formation exposed at Zengővárkony (Mecsek Mountains, Hungary) provided 528 specimens of a poorly preserved but diverse, cephalopod dominated assemblage. It could represent the lower Kimmeridgian Metahaploceras strombecki and Praesimoceras herbichi zones, all the upper Kimmeridgian ammonite zones and the Hybonoticeras hybonotum Zone of the lower Tithonian. The fauna has a truly Mediterranean character (however phyllo- and lytoceratid specimens account for only 12% of the total collected ammonite assemblage) showing greatest affinities with the fauna of the Venetian Alps, Italy. Typical Submediterranean elements also occur either in the Kimmeridgian (Aspidoceras caletanum, Pseudowaagenia inerme) or in the lower Tithonian (Gravesia aff. gigas). These results strengthen the palaeogeographical position of the Tisza microplate/megaunit (including the Mecsek tectonic zone) along the northern margin of the Tethys close to its stable European original position at the beginning of the Jurassic. Later tectonic processes (continental rifting) have detached this microcontinent from stable Europe, however it remained accessible for both Mediterranenan and Submediterranean ammonite populations. Besides ammonites, a nautiloid species (Pseudaganides strambergensis) and a serpulid (Spiraserpula spirolinites) were also recognized as first records from the Mecsek Mountains. Our results highlight the importance of this long-time neglected ammonite fauna, that is diverse, interesting and provided many previously unknown taxa from the Mecsek Mountains let alone from the rest of Hungary. The faunal assemblage comprises 30 genera (out of which 15 are first records from the Mecsek Mountians) and 34 species (out of which 20 are first records from this region). Aptychi (Laevaptychus latus, and Lamellaptychus murocostatus) and a belemnite (Hibolithes semisulcatus) are also recognized. Although some data from old literature indicated further interesting ammonite species, only more field work can confirm them. This tectonic unit still hides interesting discoveries and will illuminate stunning palaeobiogeographical connections towards the Submediterranean region. Newly collected and very much interesting ammonite fauna of Late Oxfordian - Early Kimmeridgian is under analysis now also showing Mediterranean and Submediterranean ammonites that path the way to a quantitative analysis of the ammonite assemblages of the Mecsek Mountains through the Late Jurassic - Early Cretaceous. It seems to be plausible that the Mecsek tectonic zone could have provided a stepping stone in the distribution of ammonite faunas along the northern margin of the Tethys during the Upper Jurassic.
Study conception and design; analysis and interpretation of data; drafting of manuscript: L.B.
Acquisition of data: L.B., R.A., Cs.F., B.M., D.M., Á.M.
This research was supported by the project 99öu02 of the Österreichische-Ungarische Aktion Gesellschaft (Osztrák-Magyar Akció Alapítvány). The authors are thankful to the owner of the land, Mr. Ferenc Vogl for his kind permission to collect specimens. We are indebted to László Makádi (MBFSz) for the access to old collections and for permission to deposit our material with the MBFSz. We are also thankful to Olga Piros (MBFSz) for providing valuable original publications. Our special thanks are due to Alfred Uchman, Franz Fürsich and Mikhail Rogov who clarified the nature of the serpulid and interpreted the mould. We highly acknowledge and value the precise and careful reviews of Raymond Énay, Horacio Parent, Mikhail Rogov, and Christian Salazar. Their high-quality reviews, all supportive remarks and positive attitudes greatly encouraged us during the review process. Their contributions vastly improved the quality of this study. Last but not least we deeply acknowledge editor Nigel D. Chapman's brilliant linguistic and professional comments that further improved our work.
We dedicate this paper to the eternal memory of Sigrid Missoni (1973-2020) who lost her battle against terminal illness. We spent unforgettable times with her in the field and learned much from a seasoned and disciplined scientist. Rest in Peace.
Abdulkasumzade M.R. (1988).- Верхния юра Малого Кавказа в пределах Азербайджанской ССР (стратиграфия и аммонитовая фауна) [Upper Jurassic of Lesser Caucasus at the border of Azerbaijani SSR (stratigraphy and ammonite fauna)].- Издательство Элм, Баку, Elm Publisher, Baku, 180 p. (20 Pls.)
Agassiz L. (1847).- An introduction to the study of Natural History, in a series of lectures delivered in the hall of the College of Physicians and Surgeons.- Greeley & McElrath, New York, 58 p.
Aguirre-Urreta M.B. & Rawson P.F. (2010).- Lower Cretaceous ammonites from the Neuquén Basin, Argentina: The neocomitids of the Pseudofavrella angulatiformis Zone (upper Valanginian).- Cretaceous Research, vol. 31, p. 321-343.
Alkaya F. (1989).- Kimmeridgian-Lower Tithonian ammonite fauna and stratigraphy of the Soğukçam (Bolu) area.- Bulletin of Earth Sciences Application and Research Center of Hacettepe University, Ankara, vol. 15, p. 55-73.
Ammon L. von (1875).- Die Jura-Ablagerungen zwischen Regensburg und Passau.- Th. Ackermann, München, x + 200 p. (4 Pls.)
Anđelković M.Ž. (1966).- Die Ammoniten aus den Schichten mit Aspidoceras acanthicum des Gebirges Stara Planina in Ostserbien (Jugoslavien).- Palaeontologia Jugoslavica, Beograd, vol. 6, p. 1-135 (31 Pls.).
Arkell W.J. (1950).- A classification of the Jurassic ammonites.- Journal of Paleontology, Cambridge, vol. 24, p. 354-364.
Arkell W.J. (1953).- Seven new genera of Jurassic ammonites.- Geological Magazine, Cambridge, vol. 90, no. 1, p. 36-40.
Arkell W., Furnish W., Kummel B., Miller A., Moore R., Schindewolf O., Sylvester-Bradley P. & Wright W. (1957).- Introduction to Mesozoic Ammonoidea, part L. In: Moore R.C. (ed.). Treatise on Invertebrate Paleontology, Part L, Mollusca 4, Cephalopoda-Ammonoidea.- Geological Society of America, Boulder - CO; University of Kansas Press, Lawrence - KS, L490 p.
Bardhan S., Shome S. & Roy P. (2007).- Biogeography of Kutch ammonites during the latest Jurassic (Tithonian) and a global paleobiogeographic overview. In: Landman N.H., Davis R.A. & Mapes R.H. (eds.), Cephalopods Present and Past: New insights and fresh perspectives.- Springer, p. 375-395.
Barthel K.W. & Schairer G. (1978).- Das Alter einiger Korallenriff- und Stotzenkalke des Oberjura entlang der Donau in Bayern.- Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, München, vol. 18, p. 11-27.
Baudouin C., Boselli P. & Bert D. (2011).- The Oppeliidae of the Acanthicum Zone (Upper Kimmeridgian) from Mount Crussol (Ardčche, France): Ontogeny, variability and dimorphism of the genera Taramelliceras and Streblites (Ammonoidea).- Revue de Paléobiologie, Genčve, vol. 30, no. 2, p. 619-684 (14 Pls.).
Benecke E.W. (1866).- Über Trias und Jura in den Südalpen.- Geognostisch-Paläontologische Beiträge, München, Band I, Heft I, 204 p.
Benzaggagh M. & Atrops F. (1997).- Stratigraphie et association de faune d'ammonites des zones du Kimméridgien, Tithonien et Berriasien basal dans le Prérif interne (Rif, Maroc).- Newsletters on Stratigraphy, Stuttgart, vol. 35, no. 3, p. 127-163.
Berckhemer F. & Hölder H. (1959).- Ammoniten aus dem Oberen Weißen Jura Süddeutschlands.- Beihefte zum Geologischen Jahrbuch, Hannover, vol. 35, p. 1-135 (27 Pls.).
Berger G. (2015).- Ammoniten aus dem oberen Malm (Tithonium) von Ruhpolding (Obb.).- Natur und Mensch, Nürnberg, 2014, p. 35-60.
Bernoulli D. & Renz O. (1970).- Jurassic carbonate facies and new ammonite faunas from western Greece.- Eclogae Geologicae Helvetiae, Basel, vol. 63, no. 2, p. 573-607 (6 Pls.).
Besairie H. (1936).- Fossiles kimméridgiens. In: Recherches géologiques ŕ Madagascar. Premičre suite. La géologie du Nord-Ouest.- Mémoires de l'Académie Malgache, Tananarive, vol. 21, p. 130-133.
Böckh J. (1880).- Adatok a Mecsekhegység és dombvidéke jurakorbeli lerakodásainak ismeretéhez. I. Stratigraphiai rész.- Értekezések a Természettudományok Köréből, Budapest, vol. 10, no. 10, p. 3-50.
Bouchemla I., Bendella M., Benyoucef, M., Lagnaoui A., Ferré B., Scherzinger A. & Bel Haouz W. (2020).- The Upper Jurassic Faďdja Formation (Northwestern Algeria): Sedimentology, biostratigraphy and ichnology.- Journal of African Earth Sciences, vol. 169, article 103874, 22 p.
Boughdiri M., Olóriz F., Lopez-Marques B., Layeb, M., de Matos J. & Sallouhi H. (2005).- Upper Kimmeridgian and Tithonian ammonites from the Tunisian "Dorsale" (NE Tunisia): Updated biostratigraphy from the Jebel Oust.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 111, no. 2, p. 305-316.
Boughdiri M., Sallouhi, H., Haddad S., Cordey F. & Soussi M. (2009).- Integrated biostratigraphy and regional correlations of Upper Jurassic-lowermost Cretaceous series in northern Tunisia.- GFF, Stockholm, vol. 131, no. 1-2, p. 71-81.
Breistroffer M. (1947).- Notes de Nomenclature paléozoologique. Procčs-verbal Mensuel de la Société Scientifique du Dauphiné (26th year), vol. 195, p. 99-103.
Buckmann S.S. (1909-1930).- (Yorkshire) Type ammonites. Volumes 1-7.- William Wesley and Son, London, 790 p. (67 Pls.).
Bujtor L. (1993).- Valanginian ammonite fauna from the Kisújbánya Basin (Mecsek Mts., South Hungary) and its palaeobiogeographical significance.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 188, no. 1, p. 103-131.
Bujtor L. & Albrecht R. (2021).- Oxfordian brachiopods from the ammonitico rosso-type Fonyászó Limestone Formation at Zengővárkony, Mecsek Mountains, Hungary and their palaeoecological, palaeobiogeographical and palaeopathological significance.- Paläontologische Zeitschrift, Heidelberg, 15 p. DOI: https://doi.org/10.1007/s12542-021-00560-z
Bujtor L., Albrecht R., Maróti D. & Miklósy Á. (2021).- Lower Tithonian and lower Berriasian brachiopods from the Márévár Limestone Formation, Zengővárkony (Mecsek Mountains Hungary), and remarks on their palaeoenvironment.- Paläontologische Zeitschrift, Heidelberg, vol. 95, no. 1, p. 85-95.
Callomon J.H. & Cope J.C.W. (1971).- The stratigraphy and ammonite succession of the Oxford and Kimmeridge Clays in the Warlingham Borehole.- Bulletin of the Geological Survey of Great Britain, London, vol. 36, p. 147-176.
Caracuel J.E. & Olóriz F. (1999).- Recent data on the Kimmeridgian-Tithonian boundary in the Sierra Norte of Mallorca (Spain), with notes on the genus Hybonoticeras Breistroffer.- Géobios, Villeurbanne, vol. 32, no. 4, p. 575-591.
Caswell P.V. (1958).- Geology of the Kilifi-Mazeras area.- Ministry of Environment and Natural Resources, Mines and Geological Department, Kenya. Report, Nairobi, vol. 34, p. 1-54.
Catullo T.A. (1847).- Prodromo Geognosia Paleozoica delle Alpi Venete.- Tipografia Sicca, Padova, 158 p. (13 Pls.). + p. 1-8 [Appendice] + p. 9-16 [Seconda Appendice].
Cecca F. & Savary B. (2007).- Palaeontological study of Middle Oxfordian - Early Kimmeridgian (Late Jurassic) ammonites from the Rosso Ammonitico of Monte Inici (north-western Sicily, Italy).- Geodiversitas, Paris, vol. 29, no. 4, p. 507-548.
Checa A. (1985a).- Los Aspidoceratiformes en Europa (Ammonitina, fam. Aspidoceratidae: Subfamilias Aspidoceratinae y Physodoceratinae).- Tesis doctoral, Universidad de Granada, 413 p. (42 Pls.).
Checa A. (1985b).- Simaspidoceras bauschi sp. nov. (Ammonitina, Physodoceratinae). The evolutive meaning of Submediterranean Simaspidoceras.- Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, München, vol. 25, p. 21-26.
Checa A. (1987).- Sutural simplification in Physodoceratinae (Aspidoceratidae, Ammonitina).- Estudios geologicos, Madrid, vol. 43, p. 271-278.
Checa A. & Martin-Ramos M. (1989).- Growth and function of spines in the Jurassic ammonite Aspidoceras.- Palaeontology, London, vol. 32, no. 1, p. 645-655.
Checa A. & Olóriz F. (1987).- Oxfordian and Kimmeridgian "Aspidoceras" in the Mediterranean. A methodological approach.- Estudios geologicos, Madrid, vol. 43, p. 513-520.
Cohen K.M., Finney, S.C., Gibbard P.L. & Fan. J.-X. (2013).- The ICS International Chronostratigraphic Chart.- Episodes, Seoul, vol. 36, p. 199-204.
Collignon M. (1959).- Atlas des fossiles caractéristiques de Madagascar. Fascicule V (Kimméridgien).- Service géologique, Tananarive, Pls. 96-133.
Comment G.A., Lefort J., Koppka P. & Hantzpergue P. (2015).- Le Kimméridgien d'Ajoie (Jura, Suisse) : Lithostratigraphie et biostratigraphie de la Formation de Reuchenette.- Revue de Paléobiologie, Genčve, vol. 34, no. 2, p. 161-194 (5 Pls.).
Cope J.C.W. (1967).- The palaeontology and stratigraphy of the lower part of the upper Kimmeridge Clay of Dorset.- Bulletin of the British Museum (Natural History) Geology, London, vol. 15, no. 1, p. 1-79 (33 Pls.).
Cope J.C.W. & Etches S.M. (2020).- Subdivision of the Kimmeridgian Autissiodorensis Zone (Upper Jurassic, UK).- Proceedings of the Geologists' Association, London, vol. 131, no. 5, p. 528-534.
Császár G. (ed.) (1996).- Magyarország litosztratigráfiai alapegységei. Kréta.- Magyar Állami Földtani Intézet, Budapest, 163 p.
Császár G. & Turnšek D. (1996).- Vestiges of atoll-like formations in the Lower Cretaceous of the Mecsek Mountains, Hungary.- Cretaceous Research, vol. 17, no. 4, p. 419-442.
Csontos L. & Vörös A. (2004).- Mesozoic plate tectonic reconstruction of the Carpathian region.- Palćogeography, Palćoclimatology, Palćoecology, vol. 210, p. 1-56.
Cuvier G. (1797).- Tableau élémentaire de l'Histoire naturelle des animaux.- Imprimeur Baudouin, Paris, 302 p.
D'Arpa C. & Meléndez G. (2002).- Gregoryceras fouquei (Kilian, 1889). In: Pavia G. & Cresta S. (eds.), Revision of Jurassic ammonites of the Gemmellaro Collections.- Quaderni del Museo Geologico "G.G. Gemmellaro", Palermo, vol. 6, p. 317-319.
Dacqué E. (1905).- Beiträge zur Geologie des Somalilandes. II. Teil. Oberer Jura.- Beiträge zur Paläontologie und Geologie Oesterreich-Ungarns, Wien, vol. 17, no. 3-4, p. 119-160 (Pls. XIV-XVIII).
Del Campana D. (1904).- Faunula del Giura superiore di Collalto di Solagna (Bassano).- Bollettino della Societŕ geologica Italiana, Roma, vol. 23, no. 2, p. 239-268.
Del Campana D. (1905).- Fossili del Giura superiore dei Sette Comuni in provincia di Vicenza.- Pubblicazioni del Reale Istituto di Studi Superiori Pratici e di Perfecionamento in Firenze, sezione di Scienze Fisiche e Naturali, vol. 28, p. 3-140 (6 Pls.).
Dietrich W.O. (1925).- Über eine dem mittleren Sauriermergel am Tendaguru äquivalente, rein marine Kimmeridgebildung in Mahokondo, Deutsch-Ostafrika.- Palaeontographica (Series 2), Cassel, Supplement 7, Part 1, p. 1-23.
Donovan D.T. & Callomon J.H. (1980).- Classification of the Jurassic Ammonoidea. In: House M.R. & Senior J.R. (eds.), The Ammonoidea.- Systematics Association Special Volume 18, Academic Press London, New York, p. 101-155.
Douvillé H. (1890).- Sur la classification des Cératites de la Craie.- Bulletin de la Société Géologique de France (3e série), Paris, vol. 18, p. 275-292.
Dumortier E. & Fontannes F. (1876).- Description des ammonites de la Zone ŕ Ammonites tenuilobatus de Crussol (Ardčche) et de quelques autres fossiles jurassiques nouveaux ou peu connus.- Mémoires de l'Académie royale des Sciences, Belles-Lettres et Arts de Lyon, Section des Sciences, t. XXI, 159 p. (19 Pls.).
Énay R. (1959).- La stratigraphie du Jurassique supérieur dans la chaîne anticlinale Faucille-Grand Cręt d'Eau.- Comptes rendus de l'Académie des Sciences, Paris, vol. 248, p. 125-128.
Énay R. (1966).- Le genre Gravesia (Ammonitina, Jurassique) dans le Jura français et les chaînes subalpines.- Annales de Paléontologie, Invertébrés, Paris, vol. 52, no. 1, p. 95-105.
Énay R. (2009).- Les faunes d'ammonites de l'Oxfordien au Tithonien et la biostratigraphie des Spiti-shales (Callovien supérieur-Tithonien) de Thakkhola, Népal central.- Documents des Laboratoires de Géologie de Lyon, Villeurbanne, vol. 166, p. 1-351.
Énay R. (2018).- Lithostratigraphie, biostratigraphie et faunes d'ammonites du complexe corallien d'âge kimméridgien supérieur du Jura méridional et de l'Ile Crémieu (France).- Revue de Paléobiologie, Genčve, vol. 37, no. 1, p. 41-108.
Énay R. & Howarth M.K. (2019).- Part L, Volume 3B, Chapter 7: Systematic descriptions of the Perisphinctoidea.- Treatise Online, Lawrence - KS, vol. 120, p. 1-184.
Énay R., Gallois R. & Etches S. (2014).- Origin of the Kimmeridgian-Tithonian Boreal perisphinctid faunas: Migration and descendants of the Tethyan genera Crussoliceras and Garnierisphinctes.- Revue de Paléobiologie, Genčve, vol. 33, no. 2, p. 299-377.
Énay R., Hantzpergue P., Soussi M. & Mangold Ch. (2005).- La limite Kimméridgien-Tithonien et l'âge des formations du Jurassique supérieur de la Dorsale tunisienne, comparaisons avec l'Algérie et Sicily.- Géobios, Villeurbanne, vol. 38, p. 437-450.
Fatmi A.N. & Zeiss A. (1999).- First Upper Jurassic and Lower Cretaceous (Berriasian) ammonites from the Sembar Formation (Belemnite shales), Windar Nai, Lasbela - Balochistan, Pakistan.- Geological Survey of Pakistan, Memoir, Quetta, vol. 19, p. 1-114 (57 Pls.).
Fauchald K. (1977).- The polychaete worms, definitions and keys to the orders, families and genera.- Natural History Museum of Los Angeles County: Los Angeles, CA (USA), Science Series, vol. 28, 188 p.
Favre E. (1877).- La zone ŕ Ammonites acanthicus dans les Alpes de la Suisse et de la Savoie.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 4, p. 1-114 (9 Pls.).
Fischer P. (1880-1887).- Manuel de conchyliologie et de paléontologie conchyliologique ou histoire naturelle des Mollusques vivants et fossiles.- 11 fascicules, Paris, 1369 p. (23 Pls.).
Fischer J.-C. (ed.) (1994).- Révision critique de la paléontologie française d'Alcide d'Orbigny. Volume I. Céphalopodes jurassiques.- Muséum National d'Histoire Naturelle, Paris, 340 p.
Fischer R. & Zeiss A. (1987).- Zwei neue Gravesienfunde (Ammonoidea) aus dem Mittleren Kimmeridge von Wolfsburg, Niedersachsen (Praegravesia rolkei nov. gen., nov. sp.).- Geologica et Palaeontologica, Marburg, vol. 21, p. 227-235.
Fontannes F. (1876).- Sur les Ammonites de la zone ŕ A. tenuilobatus de Crussol-Ardčche.- Bulletin de la Société Géologique de France (3e Série), Paris, vol. 5, p. 33-39.
Fontannes F. (1879).- Description des ammonites des calcaires du Chateau de Crussol - Ardčche - (Zones ŕ Oppelia tenuilobata et Oppelia tenuilobata et Waagenia beckeri).- Librairie Georg, Lyon, xi + 122 p. (XIII Pls.). URL: https://archive.org/details/descriptiondeamm00font
Főzy I. (1989).- Felsőjura ammonitesz biosztratigráfia a Bakony hegységben.- Földtani Közlöny, Budapest, vol. 119, p. 133-156.
Főzy I. (1990).- Ammonite succession from three Upper Jurassic sections in the Bakony Mts. (Hungary). In: Pallini G., Cecca F., Cresta S. & Santantonio M. (eds.), Fossili, evolutione, ambiente.- Atti IIo Convegno Internazionale F.E.A. Pergola, 87, p. 323-339.
Főzy I. (1993a).- Upper Jurassic ammonite biostratigraphy in the Gerecse and Pilis Mts. (Transdanubian Central Range, Hungary).- Földtani Közlöny, Budapest, vol. 123, no. 4, p. 441-464.
Főzy I. (1993b).- Upper Jurassic ammonite biostratigraphy of the Mecsek Mts., Hungary.- Földtani Közlöny, Budapest, vol. 123, no. 2, p. 195-205.
Főzy I. (1995).- Upper Jurassic ammonites from Seno di Guidaloca (Western Sicily).- Hantkeniana, Budapest, vol. 1, p. 131-143.
Főzy I. (ed., 2012).- Magyarország litosztratigráfiai alapegységei. Jura.- Magyarhoni Földtani Társulat, Budapest, 235 p.
Főzy I. & Scherzinger A. (2013).- Systematic descriptions of Kimmeridgian ammonites of the Gerecse and Pilis Mountains, p. 167-206. In: Főzy I. (ed.), Late Jurassic-Early Cretaceous fauna, biostratigraphy, facies and deformation history of the carbonate formations in the Gerecse and Pilis Mountains (Transdanubian Range, Hungary).- GeoLitera Publishing House, Szeged, 422 p.
Főzy I., Janssen, N.M.M. & Price G.D. (2011).- High-resolution ammonite, belemnite and stable isotope record from the most complete Upper Jurassic section of the Bakony Mts (Transdanubian Range, Hungary).- Geologica Carpathica, Bratislava, vol. 62, no. 5, p. 413-433.
Főzy I., Meléndez G., Scherzinger A., Szinger B. & Szives O. (2013).- Upper Jurassic-lowermost Cretaceous fossil localities of the Gerecse and Pilis Mountains (rocks, fossils and stratigraphy), p. 21-94. In: Főzy I. (ed.), Late Jurassic-Early Cretaceous fauna, biostratigraphy, facies and deformation history of the carbonate formations in the Gerecse and Pilis Mountains (Transdanubian Range, Hungary).- GeoLitera Publishing House, Szeged, 422 p.
Fülöp J. (1967).- A júra-kréta határ kérdéséről.- Magyar Állami Földtani Intézet, Budapest, 28 p.
Fülöp J. (1976).- The Mesozoic basement horst blocks of Tata.- Geologica Hungarica, series Geologica, Budapest, vol. 16, p. 1-229 (52 Pls.).
Gallois R.W. & Etches S.M. (2010).- The distribution of the ammonite Gravesia (Salfeld, 1913) in the Kimmeridge Clay Formation (late Jurassic) in Britain.- Geoscience in South-West England, Exeter, vol. 12, p. 240-249.
Gawlick H.-J & Missoni S. (2019).- Middle-Late Jurassic sedimentary mélange formation related to ophiolite obduction in the Alpine-Carpathian-Dinaridic Mountain Range.- Gondwana Research, vol. 74, p. 144-172.
Gemmellaro G.G. (1871a).- Studii paleontologici sulla fauna del calcare a Terebratula Janitor del Nord di Sicilia.- Giornale di Scienze Naturali ed Economiche, Palermo, vol. VI, parte I, p. 153-186 + 237-252. URL: https://www.biodiversitylibrary.org/item/110507#page/11/mode/1up
Gemmellaro G.G. (1871b).- Studii paleontologici sulla fauna del calcare a Terebratula Janitor del Nord di Sicilia.- Giornale di Scienze Naturali ed Economiche, Palermo, vol. VII, parte I, p. 73-108 + 149-157. URL: https://www.biodiversitylibrary.org/item/110526#page/9/mode/1up
Gemmellaro G.G. (1872).- Sopra i Cefalopodi della zona con Aspidoceras acanthicum Opp. sp. di Burgilamuni presso Favara, provincia di Girgenti.- Giornale di Scienze Naturali ed Economiche, Palermo, vol. VIII, parte I, p. 137-159. URL: https://www.biodiversitylibrary.org/item/109639#page/164/mode/1up
Gemmellaro G.G. (1878).- Sopra i Cefalopodi della zona inferiore degli strati con Aspidoceras acanthicum di Sicilia.- Atti dell' Accademia Gioenia di Scienze Naturali in Catania (Serie Terza), vol. 12, p. 153-250. URL: https://archive.org/details/attidellaaccadem3121878acca/page/n7/mode/2up
Gerasimov P.A., Mitta V.V., Kochanova M.D. & Tesakova E.M. (1996).- Ископаемые келловейского яруса Центральной России [Natural resources of the Callovian Stage in Central Russia].- VNIGNI, Moscow, 127 p. (57 Pls.).
Geyer O.F. (1963).- Beiträge zur Stratigraphie und Paläontologie des Jura von Ostspanien.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 118, no. 2, p. 182-196 (18 Pls.).
Goldfuss A. (1826-1831).- Petrefacta Germanić. Erster Theil.- Arnz & Comp., Düsseldorf, 252 p. (71 Pls.).
Gosselet M.J. (1881).- Esquisse géologique du Nord de la France et des contrées voisines. 2e fascicule. Terrains Secondaires. Planches.- Imprimerie Six-Horemans, Lille, 21 Pls.
Grabowski J., Császár G., Márton, E., Pszczólkowski A. & Lodowski D. (2016).- Upper Berriasian magnetostratigraphy in the Mészkemence section, Mecsek Mts (southern Hungary).- Publications of the Institute of Geophysics, Polish Academy of Sciences, Warszawa, vol. 423 (C112), p. 45-46.
Grigore D. (2000).- Kimmeridgian and lower Tithonian sequences from East and South Carpathians - Romania.- Anuarul Institutului Geologic al Romaniei, Bucureşti, vol. 72, no. 2, p. 37-45.
Grigore D. (2010).- Idoceratinae (Presimoceras, Trenerites and Lessiniceras genera) from "Acanthicum beds" of the Hăghimas Mts. (the Eastern Carpathians - Romania).- Muzeul Olteniei Craiova. Oltenia. Studii si comunicări. Științele Naturii, vol. 26, no. 2, p. 287-295.
Grigore D. (2011).- Phylloceratids from the Upper Jurassic deposits of Hăghimas Mts. (the Eastern Carpathians - Romania).- Muzeul Olteniei Craiova. Oltenia. Studii si comunicări. Științele Naturii, vol. 27, no. 2, p. 191-202.
Grigore D. (2013).- Revised ammonites fauna (Phylloceratids, Lytoceratids and Aspidoceratids-Sutneria species) from "Acanthicum beds" of the Hăghimas Mts. (Eastern Carpathians - Romania).- Revue Roumaine de Géologie, Bucureşti, vol. 57, no. 1-2, p. 81-110.
Grigore D. (2018).- Oppelid and Haploceratid ammonites from the Upper Jurassic deposits of Haghimas Mts. (The Eastern Carpathians - Romania).- Muzeul Olteniei Craiova. Oltenia. Studii si comunicari. Științele Naturii, vol. 34, no. 2, p. 15-22.
Grube A.E. (1850).- Die Familien der Anneliden.- Archiv für Naturgeschichte, Berlin, vol. 16, no. 1, p. 249-364.
Gygi R.A. (2000).- Integrated stratigraphy of the Oxfordian and Kimmeridgian (Late Jurassic) in northern Switzerland and adjacent southern Germany.- Memoirs of the Swiss Academy of Sciences, Basel, vol. 104, p. 1-151 (15 Pls.).
Gygi R.A. (2003).- Perisphinctacean ammonites of the Late Jurassic in northern Switzerland: A versatile tool to investigate the sedimentary geology of an epicontinental sea.- Schweizerische Paläontologische Abhandlungen, Basel, vol. 123, p. 1-232.
Haas J. & Péró Cs. (2004).- Mesozoic evolution of the Tisza Mega-unit.- International Journal of Earth Sciences, vol. 93, no. 2, p. 297-313.
Hauer F. (1866).- Sitzung am 4. December (1866).- Verhandlungen der k. k. geologischen Reichsanstalt, Wien, vol. 16, no. 4, p. 171-197.
Hahn W. (1963).- Die Gattung Gravesia Salfeld (Ammonoidea) im Oberjura Mittel- und Nordwesteuropas.- Palaeontographica, Abt. A., Cassel, vol. 122, no. 1-3, p. 90-110 (13 Pls.).
Hantzpergue P. (1989).- Les ammonites kimméridgiennes du haut-fond d'Europe occidentale : Biochronologie, systématique, évolution, paléobiogéographie.- Cahiers de Paléontologie, Paris, 428 p. (45 Pls.).
Hantzpergue P., Atrops F. & Énay R. (1997).- Kimméridgien. In: Cariou E. & Hantzpergue P. (eds.), Biostratigraphie du Jurassique ouest-européen et mediterranéen : Zonations parallčles et distribution des invertébrés et microfossiles.- Bulletin du Centre de Recherche elf Exploration Production, Mémoire, Pau, vol. 17, p. 87-96 (Pls. 23-24).
Hébert E. (1875).- Observations sur le travail de M. Pillet relatif ŕ la colline de Lémenc.- Bulletin de la Société géologique de France (3e Série), Paris, vol. 3, p. 387-388.
Herbich F. (1868).- Beiträge zur Paläontologie Siebenbürgens.- Verhandlungen und Mitteilungen des siebenbürgischen Vereins für Naturwissenschaften zu Hermannstadt, Sibiu, vol. 19, no. 2, p. 24-36.
Herbich F. (1878).- Das Széklerland mit Berücksichtigung der angrenzenden Landestheile, geologisch und paläontologisch beschrieben.- Jahrbuche der königlichen ungarischen geologischen Anstalt, Budapest, vol. 5, no. 2, p. 111 [93] - 186 [168] (33 Pls.).
Hesselbo S.P., Ogg J.G. & Ruhl M. (2020).- The Jurassic Period. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale, 2020.- Elsevier, Amsterdam, p. 955-1021.
Hetényi R., Hámor G. & Nagy I. (1968).- Magyarázó a Mecsek hegység földtani térképéhez 10 000-es sorozat.- Magyar Állami Földtani Intézet, Budapest, 55 p.
Hinsbergen D.J.J. van, Torsvik T.H., Schmid S.M., L.C. Matenco L.C., Maffione M., Vissers R.L.M., Gürer D. & Spakman W. (2020).- Orogenic architecture of the Mediterranean region and kinematic reconstruction of its tectonic evolution since the Triassic.- Gondwana Research, vol. 81, p. 79-229.
Hölder H. (1955).- Die Ammoniten-Gattung Taramelliceras im Südwestdeutschen Unter- und Mittelmalm. Morphologische und Taxionomische Studien an Ammonites flexuosus Buch (Oppeliidae).- Palaeontographica Abt. A., Cassel, vol. 106, no. 3-6, p. 37-158 (19 Pls.).
Hölder H. & Ziegler A. (1959).- Stratigraphische und faunistische Beziehungen im Weißen Jura (Kimeridgien) zwischen Süddeutschland und Ardčche.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 108, no. 2, p. 150-214 (22 Pls.).
Howarth M.K. (1998).- Ammonites and nautiloids from the Jurassic and Lower Cretaceous of Wadi Hajar, southern Yemen.- Bulletin of the natural History Museum, London (Geology), vol. 54, no. 1, p. 33-107 (24 Pls.).
Hyatt A. (1889).- Genesis of the Arietidae.- Smithsonian Contributions to Knowledge, Washington D.C., no 673, xi + 238 p.
Hyatt A. (1900).- Cephalopoda. In: Zittel K.A. (ed.), Textbook of Palaeontology, 1st English ed., translated by C.R. Eastman.- Macmillan, London & New York, p. 502-592.
Imlay R.W. (1943).- Upper Jurassic ammonites from the Placer de Guadalupe District, Chihuahua, Mexico.- Journal of Paleontology, Cambridge, vol. 17, no. 5, p. 527-543.
Imlay R.W. (1981).- Late Jurassic ammonites from Alaska.- Geological Survey Professional Paper, vol. 1190, iv + 40 p. (12 Pls.)
Ippolitov A.P. (2007).- Contribution to the Revision of Some Late Callovian Serpulids (Annelida, Polychaeta) of Central Russia: Part 2.- Paleontological Journal, Moscow, vol. 41, no. 4, p. 429-436.
Jekelius E. (1916).- A brassói hegyek mezozoós faunája III. - VII. A brassói dogger és malmfauna.- A Magyar Királyi Földtani Intézet Évkönyve, Budapest, vol. 24, no. 3, p. 217-314 (6 Pls.).
Joly B. (1976).- Les Phylloceratidae malgaches au Jurassique. Généralités sur les Phylloceratidae et quelques Juraphyllitidae.- Documents des Laboratoires de Géologie de Lyon, Villeurbanne, vol. 67, 471 p. (61 Pls.). URL: https://www.persee.fr/doc/geoly_0076-1672_1976_mon_67_1
King A.H. & Evans D.H. (2019).- High-level classification of the nautiloid cephalopods: A proposal for the revision of the Treatise Part K.- Swiss Journal of Palaeontology, Zürich, vol. 138, p. 65-85.
Koch N. (1909).- A tatai Kálváriadomb földtani viszonyai.- Földtani Közlöny, Budapest, vol. 39, no. 5, p. 255-275.
Krishna J. & Pathak. D.B. (1991).- Ammonoid biochronology of the upper Jurassic Kimmeridgian stage in Kachchh, India.- Journal of the Palaeontological Society of India, Lucknow, vol. 36, p. 1-13.
Kummel B. (1956).- Post-Triassic nautiloid genera.- Bulletin of the Museum of Comparative Zoology at Harvard College, Cambridge - MA, vol. 114, no. 7, p. 321-494 (28 Pls.).
Kutek J. & Zeiss A. (1997).- The highest Kimmeridgian and lower Volgian in Central Poland; their ammonites and biostratigraphy.- Acta Geologica Polonica, Warszawa, vol. 47, no. 3-4, p. 107-198 (42 Pls.).
Lamarck J.B.A.M. de (1809).- Philosophie zoologique, ou exposition des considérations relatives ŕ l'histoire naturelle des animaux ; ŕ la diversité de leur organisation et des facultés qu'ils en obtiennent ; aux causes physiques qui maintiennent en eux la vie et donnent lieu aux mouvemens qu'ils exécutent ; enfin, ŕ celles qui produisent les unes le sentiment, et les autres l'intelligence de ceux qui en sont doués.- Bailličre, Paris, 475p.
Lillo Bevia J. (1976).- Ammonites del Oxfordense Superior y Kimmeridgense de la Sierra Crevillente (Alicante).- Boletín Geológico y Minero, Logrońo, vol. 87-V, p. 462-479 (7 Pls.).
Linnaeus C. (1758).- Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata.- Holmiae [Stockholm], Laurentii Salvii, Tomus I, p. 1-824.
Loriol P. de & Pellat A. (1874).- Monographie paléontologique et géologique des étages supérieurs de la formation jurassique des environs de Boulogne-sur-Mer.- Mémoires de la Société de Physique et d'Histoire naturelle de Genéve, vol. 23, no. 2, p. 253-426 (10 Pls.).
Luci L., Garberoglio R.M. & Lazo D.G. (2013).- Serpulids and other calcareous tube-dwelling encrusting polychaetes from the Early Cretaceous Agrio Formation (Neuquén Basin, Argentina).- Géobios, Villeurbanne, vol. 46, p. 213-224.
Lukeneder A., Krystyn L., Rasser M.W. & Marzendorfer G. (2003).- A unique ammonoid fauna from the Upper Jurassic Loser section (Northern Calcareous Alps, Salzkammergut). In: Piller W.E. (ed.), Stratigraphia Austriaca.- Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftliche Kommission, Wien, vol. 16, p. 217-229 (2 Pls.).
Maisch M.W. (1996).- Revision der Lytoceraten (Ammonoidea, Lytocerataceae) aus dem Oberjura Südwestdeutschlands. Teil 1: Erstnachweis des tethyalen Lytoceraten Lytoceras cf. polycyclum Neumayr, 1873 im Unteren Kimmeridgium.- Jahresheften der Gesellschaft für Naturkunde in Württemberg, vol. 152, p. 47-55.
Malinowska L. (1989).- Biostratigraphy and paleozoogeography of the lowermost Tithonian in the Extra-Carpathian Poland.- Bulletin of the Polish Academy of Sciences, Warsaw, vol. 37, no. 1-2, p. 1-25 (5 Pls.).
Mandl G.W., Dulai A., Schlögl J., Siblik M., Szabó J., Szente I. & Vörös A. (2010).- First results on stratigraphy and faunal content of the Jurassic between Bad Mitterndorf and Toplitzsee (Salzkammergut, Austria).- Abhandlungen der Geologischen Bundesanstalt, Wien, vol. 65, p. 77-134 (17 Pls.).
Marino M.C., Andreini G., Baldanza A., D'Arpa C., Mariotti N., Pallini G., Parisi G. & Petti F.M. (2004).- Middle Jurassic-Early Cretaceous integrated biostratigraphy (ammonites, calcareous nannofossils and calpionellids) of the Contrada Diesi section (South-Western Sicily, Italy).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 110, no. 1, p. 357-372 (3 Pls.).
Měchová L., Vašíček Z. & Houša V. (2010).- Early Cretaceous ribbed aptychi - a proposal for a new systematic classification.- Bulletin of Geosciences, Pilsen, vol. 85, no. 2, p. 219-274.
Meléndez G. (1989).- El Oxfordiense en el sector central de la Cordillera Iberica (provincias de Zaragoza y Teruel).- Institucion Fernando el Catolico, Instituto de Estudios Turolenses, Zaragoza-Teruel, 418 p. (62 Pls.).
Metodiev L. (2018).- New indication of the Hybonoticeras beckeri ammonite Zone (Kimmeridgian) of the Teteven area (Central Fore-Balkan) and its significance.- Review of the Bulgarian Geological Society, Sofia, vol. 79, no. 3, p. 97-98.
Molnár J. (1961).- A zengővárkonyi vasérckutatás.- Bányászati Lapok, Budapest, vol. 94, p. 187-194.
Myczyński R. (1989).- Ammonite biostratigraphy of the Tithonian of Western Cuba.- Annales Societatis Geologorum Poloniae, Kraków, vol. 59, p. 43-125 (17 Pls.).
Nagy I. (1961).- Examen microbiofacial du complexe du Malm en affleurement ŕ Zengővárkony (Montagne Mecsek).- A Magyar Állami Földtani Intézet Évi Jelentése az 1961. évről, Budapest, p. 97-108.
Nagy I. (1964).- A Zengővárkonynál feltárt malm rétegösszlet mikrobiofácies-vizsgálata.- Magyar Állami Földtani Intézet Évi Jelentése az 1961. évről, Budapest, p. 97-108.
Nagy I. (1971).- Расчленение верхнеюрских отложений гор Мечек по ископаемым организмам.- A Magyar Állami Földtani Intézet Évkönyve, Budapest, vol. 54, no. 2, p. 319-332.
Neumayr M. (1871a).- Jurastudien. 3. Die Phylloceraten des Dogger und Malm.- Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, Wien, vol. 21, no. 3, p. 297-354.
Neumayr M. (1871b).- Jurastudien. 4. Die Vertretung der Oxfordgruppe im östlichen Theile der mediterranen Provinz.- Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, Wien, vol. 21, no. 3, p. 355 [59] - 378 [82].
Neumayr M. (1871c).- Die Fauna der Schichten mit Aspidoceras acanthicum Oppel im Nagy-Hagymás-Gebirge in Siebenbürgen.- Verhandlungen der Geologischen Bundesanstalt, Wien, vol. 1871(2), p. 21-25.
Neumayr N. (1873).- Die Fauna der Schichten mit Acanthoceras acanthicum.- Abhandlungen der kaiserlich-königlichen geologischen Reichsanstalt, Wien, vol. 5, no. 6, p. 141-257 (43 Pls.).
Neumayr M. (1875).- Die Ammoniten der Kreide und die Systematik der Ammonitiden.- Zeitschrift der Deutschen Geologischen Gesellschaft, Berlin, vol. 27, p. 854-942.
Olóriz F. (1978).- Kimmeridgiense-Tithonico inferior en el sector central de las Cordilleras Beticas (Zona Subbetica). Paleontologia. Bioestratigrafia.- Tesis doctoral, Universidad de Granada, xvii + 730 p. (57 Pls.).
Olóriz F. & Tintori A. (1991).- Upper Jurassic (Tithonian) ammonites from the Spiti shales in western Zanskar (NW Himalayas).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 96, no. 4, p. 461-486.
Olóriz F. & Villaseńor A.B. (1999).- New microconchiate Hybonoticeras from Mexico.- Géobios, Villeurbanne, vol. 32, no. 4, p. 561-573.
Olóriz F., Lara L., Mora A. de la, Villaseńor A.B. & González-Arreola C. (1993).- The Kimmeridgian/Tithonian boundary in the "Barranquito del Alacrán" section at Cuencamé (Durango, Mexico); its biostratigraphy and ecostratigraphic interpretation.- Acta Geologica Polonica, Warszawa, vol. 43, no. 3-4, p. 273-288.
Oppel A. (1856-1858).- Die Juraformation Englands, Frankreichs und des südwestlichen Deutschlands.- Württembergische naturwissenschaftliche Jahreshefte, Stuttgart, vol. 12, p. i-iv + 1-438 [1856]; vol. 13, p. 439-694 [1857]; vol. 14, p. 695-857 [1858].
Oppel A. (1862-1863).- Ueber jurassische Cephalopoden.- Palaeontologische Mittheilungen aus dem Museum des königlichen bayerische Staates, Stuttgart, vol. 3, p. 127-162 [1862]; p. 163-266 [1863].
Oppel A. (1865).- Die tithonische Etage.- Zeitschrift der Deutschen geologischen Gesellschaft, Berlin, vol. 17, no. 3, p. 535-568.
Orbigny A. d' (1842-1851).- Paléontologie française. Terrains oolithiques ou jurassiques. I. Céphalopodes.- Masson, Paris, 642 p.
Pachucki C. (1963).- Die Ammoniten-Fauna des unteren Bononiens (Bonebeds) und des oberen Kimmeridge in Belchatow und Tuszyn.- Annales Universitatis Mariae Curiae-Sklodowska Lublin-Polonia Sectio B, Lublin, vol. 18, no. 1, p. 1-21.
Pandey D.K., Alberti M., Fürsich F.T., Bhaumik S. & Ayoub-Hannaa W. (2016).- A review of the Tithonian ammonites from the Kachchh Basin, Western India.- Journal of the Palaeontological Society of India, Lucknow, vol. 61, no. 2, p. 141-173 (11 Pls.).
Parent H., Westermann G.E.G. & Chamberlain J.A. jr (2014).- Ammonite aptychi: Functions and role in propulsion.- Géobios, Villeurbanne, vol. 47, p. 45-55.
Parent H., Schweigert G. & Scherzinger A. (2020).- A review of the classification of Jurassic aspidoceratid ammonites - the Superfamily Aspidoceratoidea.- Volumina Jurassica, Warsaw, vol. 18, p. 47-52.
Parona C.F. & Bonarelli G. (1895).- Sur la faune du Callovien inférieur (Chanasien) de Savoie.- Mémoires de l'Académie Royale des Sciences, Belles-lettres et Arts de Savoie (Série 4), Chambéry, vol. 6, p. 1-183.
Parishev A.V. & Nikitin. I.I. (1981).- Головоногие моллюски Юры Украины.- Киев Наукова Думка, 144 p. (66 Pls.).
Pavia G., Benetti A. & Minetti C. (1987).- Il Rosso Ammonitico dei Monti Lessini Veronesi (Italia NE). Faune ad Ammoniti e discontinuitŕ stratigrafiche nel Kimmeridgiano inferiore.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 26, no. 1-2, p. 63-92 (7 Pls.).
Pavlow A. (1886).- Les Ammonites de la zone ŕ Aspidoceras acanthicum de l'Est de la Russie.- Mémoires du Comité Géologique, St-Pétersbourg, vol. 2, no. 3, p. 1-91 (10 Pls.).
Pervinquičre L. (1907).- Études de paléontologie tunisienne I. Céphalopodes des terrains secondaires.- F.R. de Rudeval, Paris, 438 p. (27 Pls.).
Peters K.F. (1862).- Über den Lias von Fünfkirchen.- Sitzungsberichte der kaiserlich Akademie der Wissenschaften Wien, Mathematisch-Naturwissenschaftliche Klasse, Wien, vol. 46, p. 241-293 (1 Pl.).
Petti F.M., Sarti C., Bernardi M., Deflorian M.C., Ferretti P., Todesco R. & Avanzini M. (2011).- Le ammoniti del Giurassico Superiore di Cima Campo (Trentino - Alto Adige) nelle collezioni paleontologiche del Museo Tridentino di Scienze Naturali.- Studi Trentini di Scienze Naturali, Trento, vol. 88, p. 159-185 (6 Pls.).
Pictet J.F. (1867).- Études paléontologiques sur la faune ŕ Terebratula diphyoides de Berrias (Ardéche).- Mélanges paléontologiques, Genčve, 2e livraison, p. 99-103 (Pls. XXIII-XXIV) + 106-108 (Pls. XXV-XXVI); 3e livraison, p. 135-202 (Pls. XXIX-XXXIV).
Preda I. (1973).- Die variationfazies und die Biostratigraphie der oberen Jura aus dem Hăghimaş-Gebirge.- Le Musée des Sciences Naturelles - Piatra Neamţ Études et Recherches de Géologie-Géographie-Biologie (Série Géologie-Géographie), vol. II, p. 19-41 (8 Pls.).
Quenstedt F.A. (1849).- Petrefactenkunde Deutschlands. Der Ersten Abtheilung Erster Band. Cephalopoden.- Ludwig Friedrich Fues, Tübingen, 581 p. (36 Pls.).
Quenstedt, F.A. (1857).- Der Jura.- Tübingen, 842 p. (100 Pls.).
Quenstedt F.A. (1888).- Die Ammoniten des Schwäbischen Jura. Band III. Der Weisse Jura.- E. Schweizerbart'sche Verlagshandlung (E. Koch), Stuttgart, p. 817-1140 (54 Pls.).
Radwanska U. (2004).- Tube-dwelling polychaetes from the Upper Oxfordian of Wapienno/Bielawy, Couiavia region, north-central Poland.- Acta Geologica Polonica, Warszawa, vol. 54, no. 1, p. 35-52.
Rafinesque C.S. (1815).- Analyse de la nature ou Tableau de l'univers et des corps organisés.- Palermo, p. 1-224 (self-published).
Rakus M. (1961).- Die Ammonitenfauna aus den roten knolligen Kalken der Manín-Serie.- Geologické práce, Zprávy, Bratislava, vol. 24, p. 143-151.
Rasser M.W., Lukeneder A., Krystyn L. & Märzendorfer G. (2003).- Ein aussergewöhnlicher Ammonoideen-fund aus dem Ober-Jura des Salzkammergutes.- Der steierische Mineralog, Graz, vol. 13-18, p. 4-6.
Regenhardt H. (1961).- Serpulidae (Polychaeta Sedentaria) aus der Kreide Mitteleuropas, ihre ökologische, taxonomische und stratigraphische Bedeutung.- Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, vol. 30, p. 5-115.
Reháková D., Matyja B.A., Wierzbowski A., Schlögl J., Krobicki M. & Barski M. (2011).- Stratigraphy and microfacies of the Jurassic and lowermost Cretaceous of the Veliky Kamenets section) Pieniny Klippen Belt, Carpathians, Western Ukraine).- Volumina Jurassica, Warsaw, vol. 9, p. 61-104.
Rouse G.W. & Fauchald K. (1997).- Cladistics and polychaetes.- Zoologica Scripta, Hoboken - NJ, vol. 26, no. 2, p. 139-204.
Rossi F. (1984).- Ammoniti del Kimmeridgiano superiore-Berriasiano inferiore del Passo del Furlo (Appennino Umbro-Marchigiano).- Memoria della Societŕ Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, vol. 23, no. 3, p. 75-136 (Pls. XXX-XXXVII).
Salfeld H. (1913).- Certain upper Jurassic strata of England.- Quarterly Journal of the Geological Society of London, vol. 69 (275), p. 423-430.
Sapunov I.G. (1977).- Ammonite stratigraphy of the upper Jurassic in Bulgaria. III. Kimmeridgian: Substages, zones and subzones.- Geologica Balcanica, Sofia, vol. 7, no. 1, p. 63-80.
Sapunov I.G. (1979).- Les fossiles de Bulgarie. III. 3. Jurassique supérieur. Ammonoidea.- Académie Bulgare des Sciences, Sofia, 237 p. (59 Pls.).
Sapunov I.G. & Ziegler B. (1976).- Stratigraphische Probleme im Oberjura des westlichen Balkangebirges.- Stuttgarter Beiträge der Naturkunde Serie B (Geologie und Paläontologie), vol. 18, p. 1-47 (3 Pls.).
Sarti C. (1986).- Fauna e biostratigrafia del Rosso Ammonitico del Trentino centrale (Kimmeridgiano-Titoniano).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 23, no. 2, p. 473-514 (7 Pls.).
Sarti C. (1990).- Taxonomic revision of the Kimmeridgian (Upper Jurassic) genus Mesosimoceras (Ammonoidea) and institution of the new genus Presimoceras (Ammonitina, Idoceratinae).- Paläontologische Zeitschrift, Heidelberg, vol. 64, no. 1-2, p. 39-55.
Sarti C. (1993).- Il Kimmeridgiano delle Prealpi Veneto-Trentine: Faune e biostratigrafia.- Memorie del Museo Civico di Storia naturale di Verona (II Serie) sezione Scienze della Terra, Verona, vol. 5, p. 1-145 (28 Pls.).
Sarti C. (1999).- Whorl width in the body chamber of ammonites as a sign of dimorphism. In: Olóriz F. & Rodríguez-Tovar F.J. (eds.), Advancing research on living and fossil Cephalopods.- Springer, New York, p. 315-332.
Sarti C. (2017).- Sea-level changes in the Kimmeridgian (Late Jurassic) and their effects on the phenotype evolution and dimorphism of the ammonite genus Sowerbyceras (Phylloceratina) and other ammonoid faunas from the distal pelagic swell area of the "Trento Plateau" (Southern Alps, Northern Italy.- Annali del Museo Civico di Rovereto Sezione: Archeologia, Storia, Scienze Naturali, vol. 31, p. 297-347 (3 Pls.).
Schairer G. & Barthel K.W. (1977).- Die Cephalopoden des Korallenkalks aus dem Oberen Jura von Laisacker bei Neuburg a. d. Donau.- Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, München, vol. 17, p. 115-124.
Scherzinger A. & Mitta V.V. (2006).- New data on ammonites and stratigraphy of the Upper Kimmeridgian and Lower Volgian (Upper Jurassic) of the middle Volga Region (Russia).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 241, p. 225-251.
Scherzinger A. & Schweigert G. (2016).- The ammonite genera Gravesia Salfeld and Pseudogravesia Hantzpergue in the Tithonian of S Germany and their correlation value with Western Europe.- Proceedings of the Geologists' Association, London, vol. 127, p. 288-296.
Scherzinger A., Schweigert G. & Parent H. (2006).- New considerations on dimorphism and aptychus in Gravesia Salfeld (Ammonoidea; Late Jurassic).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 241, no. 2, p. 269-286.
Scherzinger A., Schweigert G. & Főzy I. (2016).- First record of the Mediterranean zonal index Mesosimoceras cavouri (Gemmellaro, 1872) in the Upper Jurassic (Pseudomutabilis Zone, semicostatum γ horizon) of SW Germany and its stratigraphical significance.- Volumina Jurassica, Warsaw, vol. 14, p. 145-154.
Scherzinger A., Parent H. & Schweigert G. (2018).- A new species of the ammonite genus Physodoceras Hyatt (Aspidoceratidae) from the Hybonotum Zone (Lower Tithonian) of Southern Germany, with comments on the phylogeny of the genus.- Boletín del Instituto de Fisiografía y Geología, Rosario, vol. 88, p. 11-24.
Schindewolf O.H. (1925).- Entwurf einer Systematik der Perisphincten.- Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Abteilung B, Stuttgart, Beilage-Band LV, p. 497-517 (Pl. XIX).
Schlegelmilch R. (1994).- Die Ammoniten des süddeutschen Malms.- Gustav Fischer Verlag, Stuttgart, 298 p. (73 Pls.).
Schlögl J. & Zorn I. (2012).- Revision of the Jurassic Cephalopod holotypes in the collections of the Geological Survey of Austria in Vienna.- Jahrbuch der Geologischen Bundesanstalt, Wien, vol. 152, no. 1-4, p. 159-200 (15 Pls.).
Schneid T. (1915).- Die Geologie der fränkischen Alb zwischen Eichstätt und Neuburg a. Donau. I. Stratigraphischer Teil 1.- Geognostischen Jahresheften, München, vol. 27, p. 59-172.
Schweigert G. (1999).- Neue biostratigraphischen Grundlagen zur Datierung des nordwestdeutschen höheren Malm.- Osnabrücker Naturwissenschaftliche Mitteilungen, vol. 25, p. 25-40.
Schweigert G. (2005).- Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany - First results and open questions.- Zitteliana, München, vol. B26, p. 22-23.
Schweigert G. & Schlampp V. (2020).- Hypowaagenia nov. gen., a rare genus of giant aspidoceratid ammonite from the Upper Jurassic of Southern Germany.- Volumina Jurassica, Warsaw, vol. 18, p. 23-36.
Schweigert G. & Zeiss A. (1999).- Lithacoceras ulmense (Oppel) (Ammonitina) - eine wichtige Leitart des Ober-Kimmeridgiums.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 211, no 1-2, p. 49-73.
Schweigert G., Krishna J., Pandey B. & Pathak D.B. (1996).- A new approach to the correlation of the Upper Kimmeridgian Beckeri Zone across the Tethyan Sea.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlugen, Stuttgart, vol. 202, no. 3, p. 345-373.
Sidó M. (1966).- Mikropaläontologische Untersuchungen am Lias-Dogger-profil von Zengővárkony.- A Magyar Állami Földtani Intézet Évi Jelentése az 1964. évről, Budapest, p. 31-51.
Sowerby J. & Sowerby J. de C. (1812-1846).- The Mineral Conchology of Great Britain. 113 parts in 7 volumes.- Meredith, London, 1353 p.
Spath L.F. (1923).- On Ammonites from New Zealand.- Quarterly Journal of the Geological Society of London, vol. 79, no. 3, p. 286-312.
Spath L.F. (1924).- On the Blake collection of ammonites from Kachh, India.- Memoirs of the Geological Survey of India, Palaeontologia Indica (new series), Calcutta, vol. IX, no. 1, p. 1-29.
Spath L.F. (1925).- The Collection of Fossils and Rocks from Somaliland made by Messrs. Wyllie and Smellie, VII. Ammonites and Aptychi.- Monographs of the Geological Department of the Hunterian Museum, Glasgow, vol. 1, p. 111-164.
Spath L.F. (1927).- Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch), Part I.- Memoirs of the Geological Survey of India, Palaeontologia Indica (new series), Calcutta, vol. IX, no. 2, p. 1-71.
Spath L.F. (1928).- Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch), Part II. Memoirs of the Geological Survey of India, Palaeontologia Indica (new series), Calcutta, vol. IX, no. 2, p. 73-161.
Spath L.F. (1931).- Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch). Part V.- Memoirs of the Geological Survey of India, Palaeontologia Indica (new series), Calcutta, vol. IX, no. 2, p. 551-658.
Steinmann G. (1890).- Elemente der Paläontologie.- Verlag von Wilhelm Engelmann, Leipzig, 844 p.
Suess E. (1865).- Über Ammoniten.- Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften in Wien, Mathematisch-Naturwissenschaftliche Klasse, Band 52 (Abt. 1), p. 71-89.
Tavera J.M. (1985).- Los Ammonites del Tithonico Superior-Berriasense de la zona Subbetica (Cordilleras Beticas).- Tesis doctoral, Universidad de Granada, 382 p. (49 Pls.).
Thomson M.R.A. (1979).- Upper Jurassic and Lower Cretaceous ammonite faunas of the Ablation Point area, Alexander Island.- British Antartic Survey Scientific Reports, Cambridge, vol. 97, p. 1-37 (7 Pls.).
Toula F. (1907).- Die Acanthicus-Schichten im Randgebirge der Wiener Bucht bei Giesshübl (Mödling WNW).- Abhandlungen der kaiserlich und königlichen Geologischen Reichsanstalt, Wien, vol. 16, no. 2, p. 1-120 (19 Pls.).
Tribolet G. de. (1873).- Recherches géologiques et paléontologiques dans le Jura supérieur neuchâtelois.- Zurcher et Furrer, Zurich, 71 + 21 p. (3 Pls.).
Uhlig V. (1910).- Himalayan Fossil. The Fauna of the Spiti Shales.- Palaeontologia Indica (Series 15), Calcutta, vol. 4. fasc. 3, p. 307-395 (XLIX-LXXVI Pls.).
Vadász E. (1935).- Das Mecsek-Gebirge. Geologische Beschreibung Ungarischer Landschaften I.- Königlich Ungarischen Geologischen Anstalt, Budapest, 180 + xxv p.
Vašíček Z., Skupien P. & Jagt J.W.M. (2018).- Current knowledge of ammonite assemblages from the Štramberk Limestone (Tithonian-lower Berriasian) at Kotouč Quarry, Outer Western Carpathians (Czech Republic).- Cretaceous Research, vol. 90, p. 185-203.
Verma H.M. & Westermann G.E.G. (1984).- The Ammonoid fauna of the Kimmeridgian-Tithonian boundary beds of Mombasa, Kenya.- Royal Ontario Museum Life Sciences Contributions, Toronto, vol. 135, p. 1-123 (19 Pls.).
Vigh G. (1961).- A Gerecsehegység Ny-i felének földtani vázlata.- A Magyar Állami Földtani Intézet Évkönyve, Budapest, vol. 49, no. 2, p. 185 [445] - 202 [462].
Vigh G. (1984).- Néhány bakonyi (titon) és gerecsei (titon-berriázi) lelőhely ammonites-faunájának biosztratigráfiai értékelése. II. A lókúti-domb titon ammonites-faunájának biosztratigráfiai értékelése.- A Magyar Állami Földtani Intézet Évkönyve, Budapest, vol. 67, no. 1, p. 1-210 (7 Pls.).
Vörös A. (1993).- Jurassic microplate movements and brachiopod migrations in the western part of the Tethys.- Palćogeography, Palćoclimatology, Palćoecology, vol. 100, p. 125-145.
Wade M. (1988).- Nautiloids and their descendants: Cephalopod classification in 1986.- New Mexico Bureau of Mines and Mineral Resources, Memoir, Albuquerque, vol. 44, p. 15-25.
Waagen W. (1873-1875).- Jurassic Cephalopoda of Kutch. Volume I. The Cephalopoda.- Palaeontologia Indica, Calcutta, vol. 1, no. 1-4, p. 1-247 (60 Pls.).
Wegele L. (1929).- Stratigraphische und faunistische untersuchungen im Oberoxford und Unterkimmeridge Mittelfrankens. II. Palaeontologischer Teil.- Palaeontographica, Cassel, vol. 72, p. 1 [95] - 94 [188].
Wein Gy. (1965).- A Mecsek hegység "Északi pikkely"-ének földtani felépítése.- A Magyar Állami Földtani Intézet Évi Jelentése az 1963. évről, Budapest, p. 35-52.
Wierzbowski A. (1994).- Late Middle Jurassic to earliest Cretaceous stratigraphy and microfacies of the Czorsztyn Succession in the Spisz area, Pieniny Klippen Belt, Poland.- Acta Geologica Polonica, Warszawa, vol. 44, no. 3-4, p. 223-249.
Wierzbowski A., Atrops F., Grabowski J., Hounslow M.W., Matyja B.A., Olóriz F., Page K.N., Parent H., Rogov M.A., Schweigert G., Wierzbowski H. & Wright J.K. (2016).- Towards a consistent Oxfordian-Kimmeridgian global boundary: Current state of knowledge.- Volumina Jurassica, Warsaw, vol. 14, p. 14-49.
Yilmaz P.O., Norton I.O., Leary D.A. & Chuchla R.J. (1996).- Tectonic evolution and paleogeography of Europe.- Houston Geological Society Bulletin, vol. 39, no. 2, p. 1-10.
Yin J. & Énay R. (2004).- Tithonian ammonoid biostratigraphy in eastern Himalayan Tibet.- Géobios, Villeurbanne, vol. 37, p. 667-686.
Zeiss A. (1968).- Untersuchungen zur Paläontologie der Cephalopoden des Unter-Tithon der Südlichen Frankenalb.- Verlag der Bayerischen Akademie der Wissenschaften, München, 190 p. (27 Pls.).
Zeiss A. (1994).- Neue Ammonitenfunde aus dem oberen Malm Süddeutschlands.- Abhandlungen der Geologischen Bundesanstalt in Wien, vol. 50, p. 509-528 (4 Pls.).
Zeiss A. (2003).- The Upper Jurassic of Europe: Its subdivision and correlation.- Geological Survey of Denmark and Greenland Bulletin, Copenhagen, vol. 1, p. 75-114.
Zeiss A., Schweigert G. & Scherzinger A. (1996).- Hegovisphinctes n. gen. neue Ammonitengattung aus dem Unter Tithonium des nördlichen Hegau und einige Bemerkungen zur Taxonomie des Lithacoceratinae.- Geologische Blatte NO Bayern, Erlangen, vol. 46, p. 127-144.
Zejszner L. (1846).- Nowe lub niedokładnie opisane gatunki skamieniałości tatrowych odkrił i opisał.- Strabskiego, Warszawa, 32 p. (4 Pls.).
Ziegler B. (1959).- Idoceras und verwandte Ammoniten Gattungen im Oberjura Schwabens.- Eclogae Geologicae Helvetiae, Basel, vol. 52, no. 1, p. 19-56.
Ziegler B. (1964).- Das untere Kimeridgien in Europa. Colloque du Jurassique, Luxembourg, 1962.- Comptes Rendus et Mémoires, Institut grand-ducal, section des Sciences naturelles, physiques et mathématiques, Luxembourg, p. 345-354.
Ziegler B. (1977).- The "White" Upper Jurassic in Southern Germany.- Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie), vol. 26, p. 1-79 (11 Pls.).
Zieten C.H. von. (1830).- Die Versteinerungen Württembergs. Erklaerung der Abbildungen.- Schweizerbart, Stuttgart, 97 p. (72 Pls.).
Zittel K.A. (1868).- Palaeontologische Studien über die Grenzschichten der Jura- und Kreide-Formation im Gebiete der Karpathen, Alpen und Apenninen. I. Abtheilung. Die Cephalopoden der Stramberger Schichten.- Verlag von Ebner & Seubert, Stuttgart, 139 p.
Zittel K.A. (1869).- Geologische Bobachtungen aus den Central-Apenninen. In: Benecke E.W. (ed.), Geognostisch-Paläontologische Beiträge, vol. 2, number 2.- R. Oldenbourg, München, p. 88-178 (Pls. 13-15).
Zittel K.A. (1870).- Palaeontologische Studien über die Grenzschichten der Jura- und Kreide-Formation im Gebiete der Karpathen, Alpen und Apenninen. II. Abtheilung. Die Fauna der aeltern Cephalopodenfuehrenden Tithonbildungen.- Palaeontographica, Supplement, Cassel, ix + 310 p. (15 Pls.).
Zittel K.A. (1884).- Cephalopoda. In: Zittel K.A. (ed.), Handbuch der Palaeontologie, Band 1, Abt. 2, Lief 3.- Oldenbourg, Munich & Leipzig, p. 329-522.
Zittel K.A. (1895).- Grundzüge der Palaeontologie.- Oldenbourg, Munich & Leipzig, viii + 971 p.