◄ Carnets Geol. 21 (14) ►
![]()
Outline:
[1. Introduction]
[2. Stratigraphy]
[3. Methodology]
[4. Systematic palaeontology]
[5. Concluding remarks]
and ... [Bibliographic references]
Department of Geography and Geology, The University of the
West Indies, Mona, Kingston (Jamaica)
or
Published online in final form (pdf) on July 7, 2021
DOI 10.2110/carnets.2021.2113
![]()
[Editor:
Robert W. Scott; technical editor: Bruno R.C. Granier]
![]()
The lower Campanian (Upper Cretaceous) Clifton Limestone of Jamaica yields three species of hippuritid bivalve: Barrettia ruseae Chubb, Whitfieldiella luceae sp. nov. and Vaccinites vermunti Mac Gillavry, and the plagioptychid: Plagioptychus sp. The hippuritids are described in detail using statistics. Barrettia ruseae is demonstrated to be a more primitive species of Barrettia than B. monilifera Woodward or B. multilirata Whitfield, and the species Whitfieldiella luceae is shown to be a more primitive species of Whitfieldiella than W. gigas Chubb. The specimens of Vaccinites from the Clifton Limestone are compared with populations of Vaccinites from elsewhere in the Americas, and five species (probably representing a single evolutionary lineage) are recognized: V. alencasteri sp. nov. (?late Turonian-?Coniacian), V. martini Mac Gillavry (probably early to mid Santonian), V. macgillavryi Palmer (probably mid to late Santonian), V. vermunti Mac Gillavry (earliest Campanian), and V. temazcali sp. nov. (late early Campanian). The Vaccinites species can be distinguished using statistical techniques. The ages of the Clifton Limestone and the five Vaccinites species are reviewed. This research demonstrates the value of using hippuritids for biostratigraphy in the Upper Cretaceous of the Americas.
•
Barrettia;
• Whitfieldiella;
• Hippuritidae;
• rudist bivalve
statistics;
• Upper Cretaceous;
• Turonian-Campanian rudist
biostratigraphy;
• Lucea Inlier, Jamaica
Mitchell S.F. (2021).- Rudist bivalves (Hippuritoidea) from the Clifton Limestone (Lower Campanian) of western Jamaica and a reassessment of the genus Vaccinites in the Americas.- Carnets Geol., Madrid, vol. 21, no. 14, p. 315-341.
Bivalves rudistes (Hippuritoidea) de la Formation du Calcaire de Clifton (Campanien inférieur) de Jamaďque occidentale et réexamen du genre Vaccinites aux Amériques.- La Formation du Calcaire de Clifton en Jamaďque d'âge Campanien inférieur (Crétacé supérieur) renferme trois espčces de bivalves hippuritides : Barrettia ruseae Chubb, Whitfieldiella luceae sp. nov. and Vaccinites vermunti Mac Gillavry, et un plagioptychidé : Plagioptychus sp. Les hippuritides sont décrits en détail au moyen de statistiques. Barrettia ruseae est reconnue comme étant une espčce plus primitive de Barrettia que B. monilifera Woodward ou B. multilirata Whitfield, et l'espčce Whitfieldiella luceae apparaît comme étant une espčce plus primitive de Whitfieldiella que W. gigas Chubb. Les spécimens de Vaccinites du Calcaire de Clifton sont comparés aux populations de Vaccinites d'autres localités des Amériques, et cinq espčces (représentant probablement une męme lignée évolutive) sont reconnues : V. alencasteri sp. nov. (Turonien supérieur ? - Coniacien ?), V. martini Mac Gillavry (probablement Santonien inférieur ŕ moyen), V. macgillavryi Palmer (probablement Santonien moyen ŕ supérieur), V. vermunti Mac Gillavry (Campanien basal) et V. temazcali sp. nov. (partie supérieure du Campanien inférieur). Les espčces du genre Vaccinites peuvent ętre caractérisées au moyen de techniques statistiques. Les âges du Clifton Limestone et des cinq espčces du genre Vaccinites sont révisés. Ce travail démontre l'utilité des hippuritides en biostratigraphie pour le Crétacé supérieur des Amériques.
• Barrettia ;
• Whitfieldiella ;
•
Hippuritidae ;
• statistiques de
bivalves rudistes ;
• Crétacé supérieur ;
• biostratigraphie des
rudistes du Turonien-Campanien ;
• Fenętre de Lucea,
Jamaďque
Rudist bivalves were discovered in the limestones at Clifton in the parish of Hanover, Jamaica, by the Ruseas High School Geology Class from Lucea (Dixon & Chubb, 1960). The specimens were subsequently described by Chubb (1968, 1971), who recognised two species: Barrettia ruseae Chubb and Vaccinites eyrei Chubb. Dommelen (1971) revised the species and placed V. eyrei in synonymy with Vaccinites vermunti Mac Gillavry, 1937. During the mapping of the Lucea Inlier by the present author from 2003 to 2006, a number of rudist bivalves were collected from the lower Campanian Clifton Limestone and this enables a revision of these two species and the addition of a third new hippuritid rudist to the fauna.
The rudist bivalves of the Santonian and early Campanian of the Americas have not been studied as fully as those of the mid Campanian to Maastrichtian. It is therefore important to describe the faunas of the early Campanian of Jamaica and to place them into their appropriate phylogenetic position. The Clifton Limestone yields the only specimens so far discovered of the genus Vaccinites in Jamaica. These specimens also enable a revision of the species of Vaccinites that are found in the Americas using statistical analysis. Abbreviations used for rudist bivalves in the text and figures are explained in Table 1.
Table 1: Abbreviations used in description of species.
| AM - Anterior myophore |
| AS - Anterior socket |
| AT - Anterior tooth |
| CT - Central tooth |
| LV - Left valve |
| P0 - Ligamental infold |
| P1 - First pillar/infold |
| P2 - Second pillar/infold |
| PM - Posterior myophore |
| PS - Posterior socket |
| PT - Posterior tooth |
| RV - Right valve |
The geology of the Lucea Inlier has been described by Grippi (1980) and Schmidt (1988) each of whom proposed quite different schemes. Subsequent investigations (e.g., Donovan et al., 2006; Mitchell, 2006) have generally followed the Grippi (1980) scheme. Mitchell (2020a) reviewed the previous work and presented a revision of the geology of the inlier based on new geological mapping; his study showed that the scheme of Grippi (1980) was preferable to that of Schmidt (1988).
The
Lucea inlier can be divided into three blocks (southern, central and northern)
separated by the Maryland and Fat Hogs Quarters fault zones (Grippi,
1980). The Clifton Limestone forms a scarp in the central block (Fig. 1 ![]() ).
Biostratigraphic data was reviewed by Mitchell
(2020a), who concluded that the Clifton Limestone was lower lower Campanian.
).
Biostratigraphic data was reviewed by Mitchell
(2020a), who concluded that the Clifton Limestone was lower lower Campanian.
|
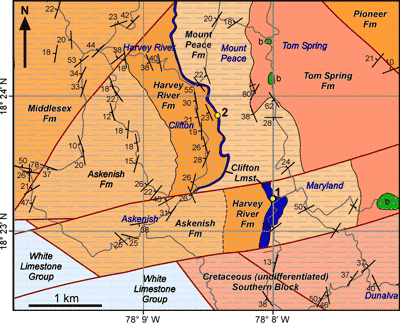
Figure 1: Geological map of part of the Central Block
(and northern part of the
Southern Block) of the Lucea Inlier, Jamaica (based on new geological mapping)
showing the outcrop pattern of the Clifton Limestone (blue band) and the rudist
collection localities (1. Maryland; 2, Clifton Cave). Faults in Red, roads in
grey. b, basalt. |
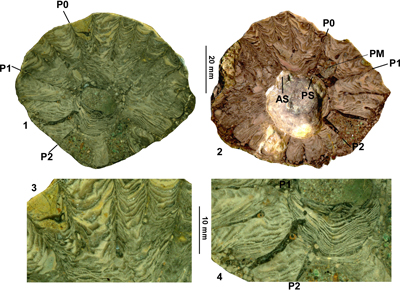
Rudist bivalve specimens from the Clifton Limestone were collected in the field and returned to the laboratory. Most specimens are encased in rock, so that external details cannot be determined. The specimens were cleaned as far as possible and sections (mostly parallel to the commissure) were cut with a rock saw. The cut surfaces were polished using successively finer grades of corundum grit to achieve a good polish. Specimens were then wetted with water and scanned at high resolution on a desk top scanner.
Measurements
were made relating to the positions of the pillars and the myocardinal
arrangement (Fig. 2 ![]() ). The measurements of populations of specimens from the
Clifton Limestone are compared with populations, and single specimens, from
other levels in the stratigraphic succession of Jamaica and elsewhere in the
American bioprovince. Material in various museum collections was used for this
study and the locations of deposited specimens is indicated in the systematic
section. Measurements are tabulated and univariate statistical characters (min,
mean, max, standard deviation) were calculated and tabulated. The various
measurements are compared by use of scatter plots to show similarities and
differences between different populations and different species. This work has
confirmed that significant differences between genera and species can be
determined by measurements and statistical treatment. A brief outline of the
various measurements is provided in the following sections.
). The measurements of populations of specimens from the
Clifton Limestone are compared with populations, and single specimens, from
other levels in the stratigraphic succession of Jamaica and elsewhere in the
American bioprovince. Material in various museum collections was used for this
study and the locations of deposited specimens is indicated in the systematic
section. Measurements are tabulated and univariate statistical characters (min,
mean, max, standard deviation) were calculated and tabulated. The various
measurements are compared by use of scatter plots to show similarities and
differences between different populations and different species. This work has
confirmed that significant differences between genera and species can be
determined by measurements and statistical treatment. A brief outline of the
various measurements is provided in the following sections.
|
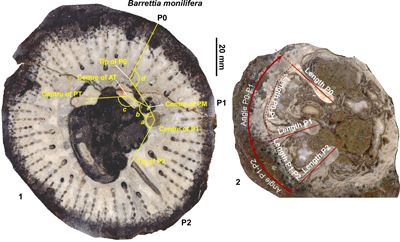
Figure 2: Diagram showing the measurements made on hippuritid rudist bivalves in this
study. 1. Barrettia monilifera Woodward
showing measurements relating to the angles of the myocardinal arrangement and P0.
2. Vaccinites vermunti Mac
Gillavry showing measurements for lengths and angles between pillars
and lengths of pillars. |
Two
different approaches have been used to investigate the relationship between the
spacing of the principal pillars (P0, P1, P2) in American Vaccinites.
One approach (e.g., Dommelen, 1971) is to measure the distance around the interior circumference of the inner
layer between the mid-points of the base of each pillar, reported here as Length
P0-P1 and Length P1-P2 (Fig. 2 ![]() ). The numbers generated are clearly dependent on
the size of the individual and will be affected by ontogeny. This approach does
not take any account of the angle that each pillar has as it extends into the
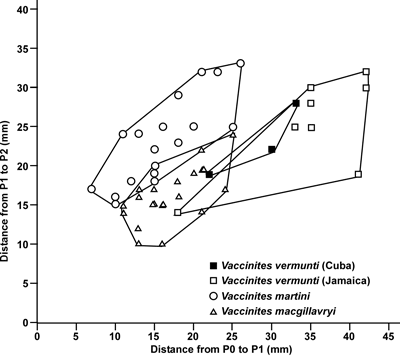
body cavity/inner shell layer of the rudist. Dommelen
(1971) used this measurement to distinguish three species of Vaccinites
(which he referred to Pseudovaccinites)
and his diagram shows that three species (V.
martini Mac Gillavry, V.
macgillavryi Palmer and V.
vermunti Mac Gillavry) plot in
different fields (Fig. 3
). The numbers generated are clearly dependent on
the size of the individual and will be affected by ontogeny. This approach does
not take any account of the angle that each pillar has as it extends into the
body cavity/inner shell layer of the rudist. Dommelen
(1971) used this measurement to distinguish three species of Vaccinites
(which he referred to Pseudovaccinites)
and his diagram shows that three species (V.
martini Mac Gillavry, V.
macgillavryi Palmer and V.
vermunti Mac Gillavry) plot in
different fields (Fig. 3 ![]() ).
).
|
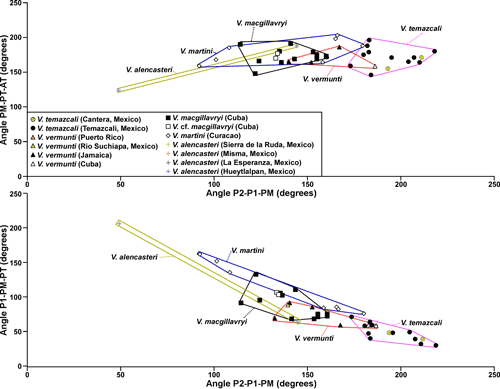
Figure 3: Scatter plot
of distances of P1 to P2 versus P0 to P1 (in mm) redrawn from Dommelen
(1971, Fig. 3) showing the relationships between Vaccinites
martini, V. macgillavryi and V. vermunti based on specimens from
Curaçao, Cuba and Jamaica. Some
of these specimens are preserved in Naturalis, The Netherlands, but others are
apparently lost. |
An
alternative approach is to use the angles between the pillars (e.g.,
Chubb, 1971; Grubić,
2004) using a line drawn between the tip and mid-point of the base of each
pillar (Fig. 2 ![]() ). These angles are reported as Angle P0-P1 and Angle P-P2 in this
work. Pillar angle is less dependent (but does change: Dommelen,
1971, Fig. 2) on growth than using length measurements around the inner shell
layer. Rather than the pillar axes themselves, some authors have taken the
angular projection from the centre of the shell to the base of the pillar when
considering angles (e.g., Dommelen, 1971; Oviedo,
2005), although this
approach is not taken here.
). These angles are reported as Angle P0-P1 and Angle P-P2 in this
work. Pillar angle is less dependent (but does change: Dommelen,
1971, Fig. 2) on growth than using length measurements around the inner shell
layer. Rather than the pillar axes themselves, some authors have taken the
angular projection from the centre of the shell to the base of the pillar when
considering angles (e.g., Dommelen, 1971; Oviedo,
2005), although this
approach is not taken here.
The
relative lengths of pillars can also be measured. The length of the pillar is
determined from its base (on a line that would encircle the inside of the body
cavity if the pillars were not there) to its tip (Fig. 2.2 ![]() ) and is recorded here
as Length P0, Length P1 and Length P2. Other workers have measured along the
mid-width (i.e., a straight, curved or
irregular line) of each pillar to get the length (e.g.,
Steuber, 1999), but this approach is not
used here.
) and is recorded here
as Length P0, Length P1 and Length P2. Other workers have measured along the
mid-width (i.e., a straight, curved or
irregular line) of each pillar to get the length (e.g.,
Steuber, 1999), but this approach is not
used here.
3.1. Angles between the principal myocardinal-pillar elements
Boissevane
& Mac Gillavry (1932) and Dommelen (1971) used the
angle formed between the lines drawn between the centres of PT and AT and the
centres of the PT and MP to ascertain the degree of development of Barrettia.
Although their measurements suggested an unreasonable lineage (B.
sparcilirata to Barrettia sp. (of Douvillé,
1926) to B. monilifera to B.
multilirata) that was not in line with the stratigraphic succession of the
species involved (some of which are now placed in different genera), the use of
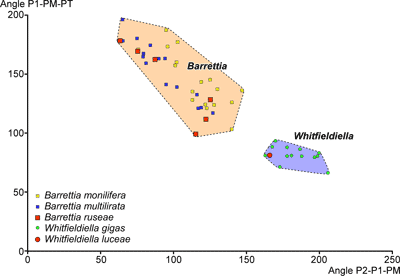
angles to understand relationships in hippuritids clearly has some merit. Mitchell
(2010) expanded on this and used three angles, defined by lines drawn between
the tip of P2, the centre of P1 (or half the width of P1 in from the tip if not
circular), the centre of the PM, the centre of the PT and the centre of the AT (Fig.
2 ![]() ). The three angles P2-P1-PM, P1-PM-PT and PM-PT-AT (the last one being
the same as used by Boissevane & Mac Gillavry,
1932, and Dommelen, 1971) were determined
for species of Barrettia monilifera Woodward,
B. multilirata Whitfield,
Whitfieldiella gigas (Chubb)
and Praebarrettia sparcilirata (Whitfield).
This showed that Barrettia, Whitfieldiella and Praebarrettia
plotted in different fields of a P1-PM-PT versus P2-P1-PM scatter plot. Further
studies (Götz & Mitchell, 2009; Mitchell,
2020b) expanded on this by adding fields for the genera Laluzia
and Caribbea.
). The three angles P2-P1-PM, P1-PM-PT and PM-PT-AT (the last one being
the same as used by Boissevane & Mac Gillavry,
1932, and Dommelen, 1971) were determined
for species of Barrettia monilifera Woodward,
B. multilirata Whitfield,
Whitfieldiella gigas (Chubb)
and Praebarrettia sparcilirata (Whitfield).
This showed that Barrettia, Whitfieldiella and Praebarrettia
plotted in different fields of a P1-PM-PT versus P2-P1-PM scatter plot. Further
studies (Götz & Mitchell, 2009; Mitchell,
2020b) expanded on this by adding fields for the genera Laluzia
and Caribbea.
Pons et al. (2019), subsequently, dismissed the use of angles and argued that the correlation between P1-PM-PT versus P2-P1-PM was negative and synonymised Whitfieldiella with Barrettia. However, because of the arrangement of the rudist myocardinal-pillar system, only a limited space in P1-PM-PT versus P2-P1-PM scatter plots is possible, and this must show a negative correlation, because the four features defining the angles must be situated close to the inner shell layer. While the area of possible P1-PM-PT versus P2-P1-PM scatter plots is completely occupied when looking at plots of Tethyan Vaccinites, Hippurites and Hippuritella (where there are numerous lineages), the New World (American) hippuritids are clearly partitioned into separate space on P1-PM-PT versus P2-P1-PM scatter plots. The fact that other characteristics (such as, the pore systems) can also be used to distinguish genera, indicates that a separation of the genera Barrettia and Whitfieldiella (which have different pore systems: Mitchell, 2010) based on P1-PM-PT versus P2-P1-PM scatter plots is useful, and therefore warranted. The fact that the early early Campanian species of Barrettia and Whitfieldiella described in this paper fall into the same fields as the late middle Campanian Barrettia and Whitfieldiella fields described by Mitchell (2010) indicates that these fields are conservative in these genera. In contrast, American species of Vaccinites plot in separate overlapping fields that probably indicate an evolutionary progression based on the limited amount of dating evidence of the species involved.
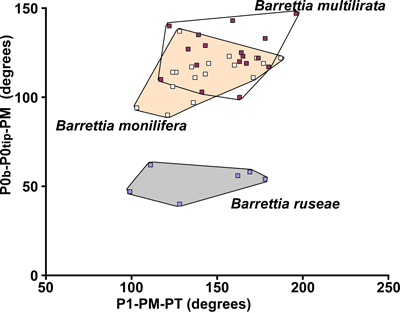
3.2. Ligament and posterior myophore relationship
The
length of the ligamental infold (P0) and the position of the posterior myophore
(PM) is also of significance in understanding the evolution of some hippuritids,
notably the multiple folded hippuritids Barrettia
and Whitfieldiella. This can be
assessed by an angle measured between a line drawn between the tip and the
mid-point of the base of P0 and the centre of the PM (Fig. 2 ![]() ). This angle is
found to be sensitive to understanding the evolution of these genera, with early
forms having long, commonly strongly curved, P0s and consequently smaller P0-PM
angles and more advanced forms having shorter POs (almost the same as the 'normal' rays) and larger P0-PM angles. Because different lineages have
different positions of the PM relative to the body chamber, this criterion can
only be used to assess the degree of 'advancement' within each lineage.
). This angle is
found to be sensitive to understanding the evolution of these genera, with early
forms having long, commonly strongly curved, P0s and consequently smaller P0-PM
angles and more advanced forms having shorter POs (almost the same as the 'normal' rays) and larger P0-PM angles. Because different lineages have
different positions of the PM relative to the body chamber, this criterion can
only be used to assess the degree of 'advancement' within each lineage.
Higher level systematics follow Skelton (2013) with further details from Skelton (2018). The classification of the Hippuritidae into families follows Mitchell (2020b). Material is stored in the following repositories:
CNHM: the Cuban Natural History Museum in Havana, Cuba,
IGM: the Geological Museum at the University of Mexico, Mexico City,
NL: Naturalis, Leiden, The Netherlands,
UPR: the Geology Museum at the University of Puerto Rico,
USNM: United States National Museum, Smithsonian, Washington D.C.,
UT: The Invertebrate Collections of the Jackson School of Geoscience at the University of Texas in Austin, USA,
UWIGM: University of the West Indies Geology Museum, Kingston, Jamaica.
Order HIPPURITIDA Newell, 1965
Suborder HIPPURITIDINA Newell, 1965
Superfamily RADIOLITOIDEA Orbigny, 1847
Family HIPPURITIDAE Gray, 1848
Subfamily BARRETTIINAE Chubb, 1971
Genus Barrettia Woodward, 1862
Type species. Barrettia monilifera Woodward, 1862, from the mid Campanian Back Rio Grande Limestone by original designation (Woodward, 1862).
Barrettia ruseae Chubb, 1968
. 1968 Barrettia rusaea n.sp.; Chubb, p. 30.
v. 1971 Barrettia rusaea Chubb; Chubb, p. 213, Pl. 55, fig. 1.
v. 1971 Barrettia ruseae Chubb; Dommelen, p. 84, Pls. 5-6.
v. 2004 Barrettia ruseae Chubb; Grubić, p. 159, Pl. 6, fig. 1.
non 2007 Barrettia monilifera Woodward; Oviedo et al., p. 310.
non 2010 Barrettia cf. ruseae Chubb; Pons et al., p. 981, Fig. 10.1-10.4.
|
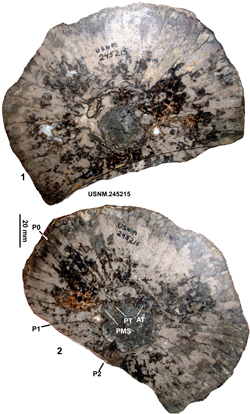
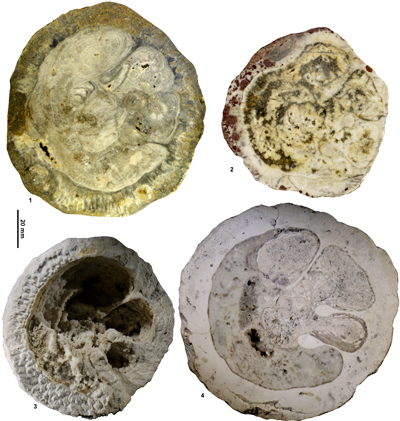
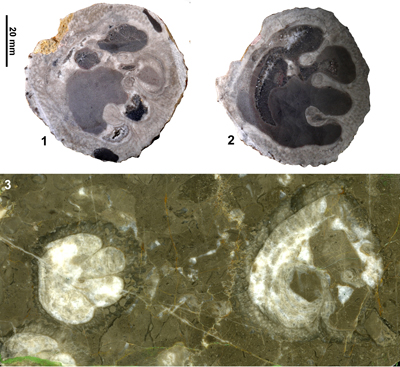
Figure 4: Views of the holotype (USNM.245215) of Barrettia
ruseae Chubb from the Clifton
Limestone at Clifton.1, RV, transverse section, adapical view. 2, RV, transverse
section, abapical view. |
|
Figure 5:
Barrettia ruseae Chubb
from the Clifton Limestone. 1-2 (UWIGM.2020.1), Maryland Locality; 1, RV,
transverse section, adapical view; 2, detail showing coarse vesicular inner
shell layer. 3 (UWIGM.RUD.2006.2), RV, transverse section, adapical view. 4
(UWIGM.RUD.2006.1), RV, transverse section, abapical view. 3-4, Clifton Cave
locality. |
|
Figure 6:
Barrettia ruseae Chubb
from the Clifton Limestone. 1 (UWIGM.RUD.2006.3), RV, transverse section,
adapical view. 2 (UWIGM.RUD.2006.4), RV, transverse section, abapical view.
Clifton Cave locality. |
Diagnosis. A moderately large species of Barrettia with a long ligamental pillar, submoniliform rays and a coarse, cellular infill in the inter-ray spaces.
Type specimen. The type specimen is preserved in the collections of the USNM under
the number USNM.245215 (formerly UWI 1190), from the Clifton Limestone at
Clifton, Jamaica. It was figured by Chubb (1971, Pl. 55, fig. 1) and a drawing
based on this photograph was presented by Grubić
(2004, Pl. 6, fig. 1). Although Chubb
(1971, p. 213-214) clearly had more than one specimen of this species, he
clearly designated UWI 1190 as the type, and as such it is the holotype. Chubb's
other material is neither in the US National Museum nor in the UWI Geology
Museum and must be considered lost. The holotype consists of a single slab and
two new photographs of it are presented in Figure 4 ![]() .
.
Material. The available material for study consists of three collections. Firstly, the holotype. Secondarily, eight specimens (J3390-1, J3390-2, J4009-1, J4009-2, J4010-2, J4010-3, J4012-1, J4012-2) that were studied by Dommelen (1971), none of which preserves the myocardinal arrangement, that are preserved in Naturalis, Leiden, The Netherlands. Thirdly, five specimens (UWIGM.RUD.2006.1 to 2006.4 from Clifton Cave, and UWIGM.RUD.2020.1 from Maryland) collected from the Clifton Limestone (all of which preserve the myocardinal features) during geological mapping of the Lucea Inlier in 2006.
Description. The largest incomplete adult right valve seen has a diameter of about 140 mm and a height of 130 mm. The right valve (e.g., J3390-2: Dommelen, 1971, Pl. V.1-2) initially increases rapidly in diameter in youth before becoming generally disc shaped and then low cylindrical. The outer shell layer of the right valve is dark grey to black and has a thickness of about 1 to 1.5 mm (according to Dommelen, 1971); in many specimens (all the new material) it has been destroyed either by clionid sponge borings (Entobia isp.) or by pressure solution. The outer layer of the right valve is folded into the inner layer as a series of rays. The number of rays is difficult to count, but the holotype has about 66 rays (with a diameter of 141 mm), UWIGM.RUD.2006.2 has about 53 rays (with a diameter of 120 mm), and UWIGM.RDU.2020.1 is estimated to have had 44 rays (with a diameter of 98 mm). P0 is represented by a very long narrow ray which is distinctly curved at its end. P1 is represented by a ray with a broadly circular knob with a diameter of about 6 mm. P2 is represented by a ray with a radially elongated, parallel-sided to lens-shaped knob with a length of about 14 to 18.5 mm and a thickness of about 2.5 to 4 mm. The rays vary from almost parallel sided to distinctly submoniliform. The more parallel-sided rays have widths of about 1 mm or a little less and show only slight changes in width comparable to the beads seen in the more submoniliform rays. For the submoniliform rays, the beads are rather irregular, radially elliptical with widths from 1 to 2 mm and lengths of about 2.5 to 3 mm; beads are spaced at distances (from centre to centre of beads) of about 4 to 5 mm along rays. The terminal knob on each ray is larger with a width of up to 3 mm. The inner shell layer is formed by anastomosing lamellae that produce a vesicular fill with the vesicles having widths of about 1.5 to 3 mm. About three vesicles are developed concentrically between rays. In some specimens the vesicles are arranged in arcs that presumably reflect the original furrow that would have been present on the apertural surface between each pair of rays (cf. Parastroma guitarti as illustrated in Mitchell, 2020b). There are no inter-ray partitions. The left valve is poorly known. Dommelen (1971, p. 85) described a specimen with part of the left valve with the inner layer showing the suggestion of pallial canals. UWIGM.RUD.2006.4 also preserves a portion of the left valve and appears to show two pallial canals in the inner layer of each buttress between adjacent rays.
Details of the myocardinal arrangement are preserved in some of the specimens. The anterior tooth is about twice the size of the posterior tooth. The teeth fit into slots formed by grooves on the walls of the shell interior. The posterior myophore has a rather irregular, radial orientated, ellipsoidal form.
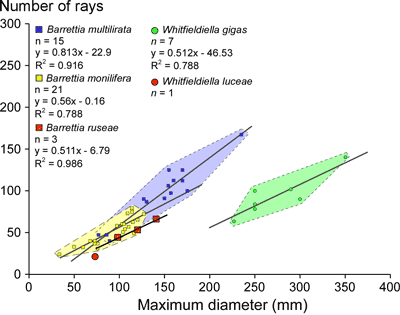
Measurements.
The
number of rays versus diameter (Fig. 7 ![]() ) shows that this species is similar to
other species of Barrettia (Mitchell, 2010),
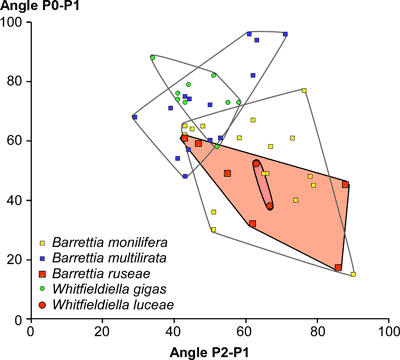
but has slightly fewer rays for a given diameter. The angle between P0 and P1
ranges from 17° to 59°, and the angle from P1 to P2 ranges from 43° to 88° (Fig.
8
) shows that this species is similar to
other species of Barrettia (Mitchell, 2010),
but has slightly fewer rays for a given diameter. The angle between P0 and P1
ranges from 17° to 59°, and the angle from P1 to P2 ranges from 43° to 88° (Fig.
8 ![]() ).
On P2-P1-PM vs. P1-PM-PT scatter plots Barrettia ruseae plots in a similar field to Barrettia monilifera
(Fig. 9
).
On P2-P1-PM vs. P1-PM-PT scatter plots Barrettia ruseae plots in a similar field to Barrettia monilifera
(Fig. 9 ![]() ). The relatively long P0 produces a
small P0t-P0tip-MP angle which contrasts with the larger angle seen in B.
monilifera and B. multilirata (Fig. 10
). The relatively long P0 produces a
small P0t-P0tip-MP angle which contrasts with the larger angle seen in B.
monilifera and B. multilirata (Fig. 10 ![]() ).
).
|
Figure 7: Graph showing number of rays versus diameter for species of Barrettia and Whitfieldiella
from Jamaica. |
|
|
|
|
|
|
Discussion. Barrettia ruseae is a distinctive early early Campanian species of Barrettia. It differs from both the late middle Campanian B. monilifera and B. multilirata by its longer P0, its lack of inter-ray partitions, and the morphology of the beads on the rays. In both B. monilifera and B. multilirata the beads have a well-defined, elliptical form with the 'string' connecting the beads being almost hairline-thin (Mitchell, 2010, Fig. 8). In contrast, the beads in B. ruseae are much more poorly defined and in some specimens is almost undefined, and the 'string' of the ray connecting the beads is thick and irregular. Additionally, the teeth of B. ruseae are of unequal size, with the posterior tooth being about half the size of the anterior tooth, whereas in B. monilifera and B. multilirata the teeth are of more-or-less equal size (Mitchell, 2010, Fig. 6). The small posterior tooth would appear to be an ancestral character, as small posterior tooth is regarded as a primitive character in rudist bivalves (e.g., Skelton & Smith, 2000; Skelton, 2018).
Barrettia coatesi (Chubb) is a similar early Campanian form to B. ruseae. The former is, however, smaller, has almost parallel-sided rays with very ill-defined beads, and the right valve inner layer shell structure appears to be coarser. Barrettia dommeleni Grubić, B. philipy Grubić and B. macgillavryi Grubić (Grubić, 2004) are too poorly known to compare with B. ruseae, and any comparison will have to await a re-description of these Puerto Rican and Cuban forms.
Barrettia cf. ruseae from the lower Campanian of Mexico was described by Pons et al. (2010) and would appear to belong to the group of B. ruseae - B. coatesi or to a primitive form of Whitfieldiella. Unfortunately, the material is not very well preserved. Judging from the figures and description, it is a large 'species' and the ligament is relatively long and thin as in the B. ruseae - B. coatesi group. Unfortunately, the myocardinal-pillar arrangement is not possible to determine. However, the inner layer shell structure seems to consist of much smaller vesicles than in B. ruseae and is closer to that of Whitfieldiella luceae n.sp. (described below), and it is doubtful if this material belongs to B. ruseae. Better material will have to be collected to understand the true affinity of this form.
Genus Whitfieldiella Mitchell, 2010
Type species. Barrettia gigas Chubb from the upper middle Campanian of Jamaica (by original designation: Mitchell, 2010).
Discussion. Mitchell (2010) established Whitfieldiella for barrettine hippuritids that had a distinct pore system consisting of robust pustule-like sieve plates that were separated from adjacent sieve plates by a wide imperforate band, with the distinctive left vale morphology not being seen in other hippuritids. The genus could also be determined by its smaller number of rays (when compared with Barrettia), which is due to the presence of the imperforate bands, by its different right valve inner shell structure and by its myocardinal-pillar arrangement. The distinctive left valve pore system is present in at least two species (both awaiting description) in Jamaica and contrasts with the pore systems of other New World hippuritids, such as, Barrettia, Parastroma, Laluzia and Praebarrettia (Mitchell, 2010, 2020b). The genus is therefore well founded and represents a parallel evolution to Barrettia as demonstrated by the new species described here.
Whitfieldiella luceae sp. nov.
(Fig. 11 ![]() )
)
|
|
Origin of name. After the Lucea Cretaceous Inlier of western Jamaica where the species is found.
Diagnosis. A small species of Whitfieldiella with parallel-sided to submoniliform rays, a long ligamental (P0) infold and lacking inter-ray partitions.
Material. One specimen (Holotype: UWIGM.RUD.2020.2) from the Clifton Limestone of the Lucea Inlier Jamaica. An additional specimen is also available from the Cotui Limestone (lower Campanian) of Puerto Rico (designated Paratype: UWIGM.RUD.2020.3) and will be described elsewhere.
Description. A small species of Whitfieldiella with a maximum diameter of 98 mm (height not determined). The outer shell layer of the right valve is poorly preserved but appears to be compact with a thickness of 0.5 mm, although in places it shows extensive clionid sponge borings (Entobia isp.). The outer shell layer of the right valve is folded into the left valve as a series of infoldings (rays), which number 22 in the holotype. The rays are arranged in about three cycles with rays of each subsequent cycle being shorter and penetrating less far into the inner shell layer. The rays range from almost parallel sided to weakly moniliform. Parallel-sided rays have thicknesses of 0.8 - 1 mm, whereas weakly submoniliform rays have beads with thicknesses of 0.9 to 1.1 mm separated by ray strings with thicknesses of 0.4 to 0.5 mm. The insipient beads are irregular and separated by distances (bead centre-to-bead centre) of 3.9 to 4.8 mm. The terminal knob on secondary rays (i.e., other than for P0, P1 and P2) has a thickness of 1.2 to 2.0 mm and is generally irregularly tear-shaped. P0 is a long ray which is curved at its tip. P1 is a ray with a large, almost circular (but weakly tear-shaped) knob which has a diameter of 4.2 mm. P2 is a ray with an elongate knob, which is tear shaped, it has a length of 9.6 mm and gradually increases in width reaching a maximum width of 1.7 mm at 2.7 mm from its tip. The inner layer of the right valve is formed by closely spaced tabulae, which do not or only rarely anastomose, in both the body chamber and in the inter-ray areas. The tabulae in the inter-ray areas are arcuate and appear to indicate that the original inter-ray area was occupied by a furrow (in which the buttresses of the left valve would fit) between the ridges formed by the rays (as in P. guitarti: Mitchell, 2020b). Details of the left valve are largely unknown other than for the posterior myophore that is preserved in its socket in the right valve. The sockets for the teeth can be determined on the polished apertural surface of the holotype, and the posterior myophore is represented by a radial elongate blade that is preserved in the posterior myophore cavity.
Measurements.
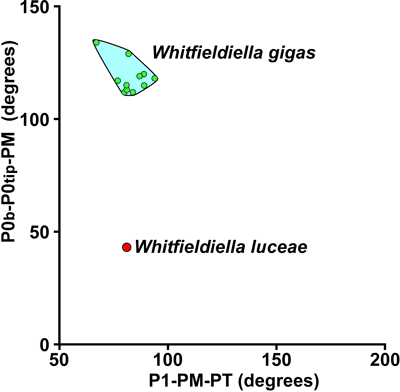
The
Holotype plots in the Whitfieldiella
field on P2-P1-PM versus P1-PM-PT scatter plots
(Fig. 9 ![]() ), has slightly fewer
rays for the same diameter as species of Barrettia
(Fig. 7
), has slightly fewer
rays for the same diameter as species of Barrettia
(Fig. 7 ![]() ), and plots in the same field as B.
ruseae and B. monilifera on plots
of angles P0-P1 versus P1-P2 (Fig.
8
), and plots in the same field as B.
ruseae and B. monilifera on plots
of angles P0-P1 versus P1-P2 (Fig.
8 ![]() ). On a scatter plot for the angles
P0b-P0tip-PM versus P1-PM-PT (Fig. 12
). On a scatter plot for the angles
P0b-P0tip-PM versus P1-PM-PT (Fig. 12 ![]() ), W.
luceae plots in a similar position relative to W. gigas as B. ruseae does
in relationship to B. monilifera (Fig. 10
), W.
luceae plots in a similar position relative to W. gigas as B. ruseae does
in relationship to B. monilifera (Fig. 10 ![]() ).
).
|
|
Discussion. Whitfieldiella luceae shares many characters with Barrettea ruseae, notably the form of the weakly moniliform rays and the lack of inter-ray partitions. The two species are distinguished on the basis of their dentition (Whitfieldiella-like versus Barrettia-like) and the structure of the inner shell layer (W. luceae has closely spaced tabulae whereas B. ruseae has a coarse vesicular structure). W. gigas (the type species and only other described species of the genus) clearly differs from W. luceae in being much larger, having fully moniliform rays on a hair-line string, a shorter ligamental (P0) ray, and in possessing inter-ray partitions.
Subfamily Hippuritinae Gray, 1848
Genus Vaccinites Fischer, 1887
Type species. Hippurites cornuvaccinum Bronn, 1831, from the Upper Cretaceous of Untersberg on the border between Germany and Austria.
Discussion. Many species names have been used for New World (American) Vaccinites including names carried over from the Old World (e.g., V. giganteus (D'Hombras-Firmas), V. grossouvrei (Douvillé), V. gosaviensis (Douvillé), V. praegiganteus Toucas: Alencáster et al., 1987) as well as names established for New World species (e.g., V. martini Mac Gillavry, V. macgillavryi Palmer, V. vermunti Mac Gillavry, V. eyrei Chubb). Only limited systematic statistical treatment of the group has used pillar angles (Mac Gillavry, 1937) or pillar distances (Dommelen, 1971), and the relationships or synonymies among species have not been sufficiently investigated. A large amount of material resides in museum collections, and much of this has been directly studied for this work. Some specimens are lost, and in such cases, original figures, or drawings, were used for this study. Additionally material is also available in publications and theses. For this study, both previously used criteria as well as the angles described in the methodology section are used to investigate the range of variation in New World Vaccinites and to put the Clifton Limestone Vaccinites into an American Province perspective.
Mac
Gillavry
(1937) used a series of angles in describing species of Vaccinites
in Cuba, including the angle between line joining PT-AT and P0, the angle
AT-PT-PM, and the lengths P0-P1 and P1-P2. Dommelen
(1971, Fig. 3, reproduced here as Fig. 3 ![]() ) demonstrated that plots of the
perimeter distance of the pillars in Vaccinites
(his Pseudovaccinites) fell in three
fields that separated populations of V.
martini, V. macgillavryi and V.
vermunti, although there was a little overlap of fields. Subsequent authors
(e.g., Oviedo, 2005; Pons et
al., 2010) have maintained V. vermunti
and V. macgillavryi as separate species.
) demonstrated that plots of the
perimeter distance of the pillars in Vaccinites
(his Pseudovaccinites) fell in three
fields that separated populations of V.
martini, V. macgillavryi and V.
vermunti, although there was a little overlap of fields. Subsequent authors
(e.g., Oviedo, 2005; Pons et
al., 2010) have maintained V. vermunti
and V. macgillavryi as separate species.
One of the largest problems in dealing with Vaccinites, is the lack of data on the myocardinal arrangement in many specimens. This is particularly true of some of the (probably) older species (?Turonian-?Coniacian) from Mexico. Despite this, locations are treated as separate entities and material is assessed depending on the location it came from. A further complication is the relatively wide plasticity of characters in American Vaccinites, which makes analysing single specimens difficult or impossible.
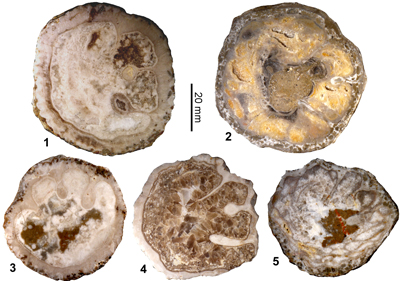
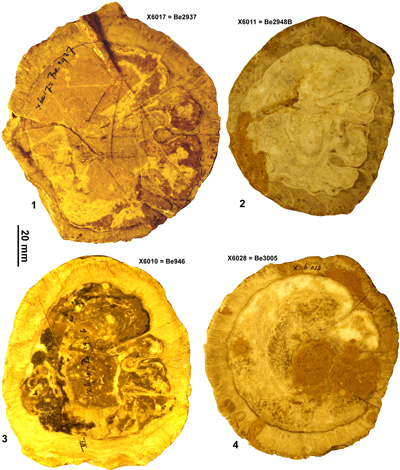
Vaccinites alencasteri sp. nov.
(Fig. 13 ![]() )
)
. 1987 Vaccinites praegiganteus Toucas; Alencáster et al., p. 30, Figs. 1(4-6)
. 1987 Vaccinites gosaviensis (Douvillé); Alencáster et al., p. 32, Figs. 1(1-3)
. 1987 Vaccinites grossouvrei (Douvillé); Alencáster et al., p. 31, Figs. 2(8-9)
. 1987 Vaccinites giganteus (D'Hombras-Firmas); Alencáster et al., p. 32, Figs. 2(4-7)
. 2005 Vaccinites macgillavryi Palmer; Oviedo, p. 72-77, Figs. 53(1-9), 54(1-14), 55(1-6).
Origin of name. After the late Dr Gloria Alencáster Ybarra who described the first specimens, and for her work on the rudist bivalves of Mexico.
|
|
Diagnosis. A small/medium species of Vaccinites with P2 distinctly shorter than P1.
Type specimens. Holotype: IGM 4311 (figured as Vaccinites grossouvrei by Alencáster et al., 1987); Paratypes: IGM 4309 (figured as Vaccinites paragiganteus by Alencáster et al., 1987) and IGM 4312 (figured as Vaccinites gossaviensis by Alencáster et al., 1987); all from Hueytlalpan, Guerrero State, Mexico.
Material. Extensive material of this species is available, but the myocradinal features are only preserved in two specimens. The material used in this study is the specimens in the IGM, Mexico, figured by Alencáster et al. (1987), illustrations of specimens from four localities (La Esperanza, Hueytlalpan, Sierra de la Ruda (Santa Isabel), and Misma Localidad, all in the state of Guerrero) have also been figured by Oviedo (2005), and these illustrations have been measured to provide biometric data for statistical analysis.
Description. A small- to medium-sized Vaccinites with a maximum diameter reaching 62 mm. The RV is conical to cylindro-conical and the exterior surface is marked with longitudinal costae separated by distances of 3.5 to 5.5 mm. The outer shell layer is up to 8.5 mm thick, and there are very weak to weak orimentary folds on its inner surface. The outer shell layer is folded into the shell as three principal pillars. P0 often has a broad triangular base, and may taper, be parallel sided, or have a slightly broader tip; the inner end is typically truncated to some degree. P1 is short, moderately to strongly pinched and has a radially elongate to subcircular knob. P2 is longer than P1, is moderately to strongly pinched, and has a radially elongate elliptical knob. The inner shell layer is very thin, and the body cavity fill appears to be cellular. The LV (IGM 4310) is low and convex, and shows typical radial furrows. The myocardinal arrangement is poorly understood and is only seen in two specimens.
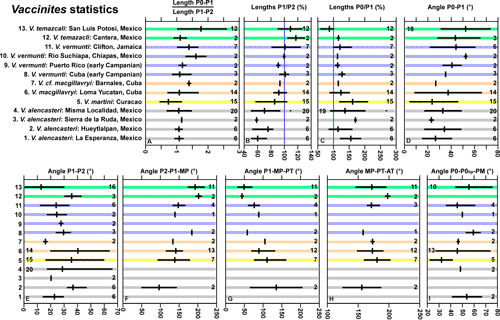
Measurements.
Because of the limited number of specimens (2) that show the myocardinal
arrangement, it is difficult to compare this species with other species.
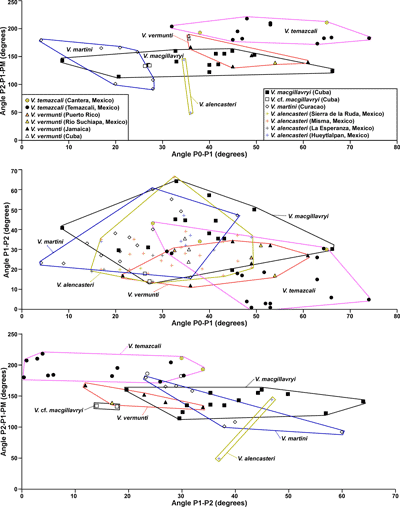
Univariate measurements are summarized in Figure 14 ![]() . On the Angle PM-PT-AT versus Angle P2-P1-PM scatter
plots (Fig. 15
. On the Angle PM-PT-AT versus Angle P2-P1-PM scatter
plots (Fig. 15 ![]() ), the two specimens plot in different places, but more specimens
are required to understand whether these are single large fields or more than
one species may be represented. In scatter plots of Angle P2-P1-PM versus Angle
P0-P1, the field of V. alencasteri (2
specimens) partially overlaps that of V.
macgillavryi (Fig. 16
), the two specimens plot in different places, but more specimens
are required to understand whether these are single large fields or more than
one species may be represented. In scatter plots of Angle P2-P1-PM versus Angle
P0-P1, the field of V. alencasteri (2
specimens) partially overlaps that of V.
macgillavryi (Fig. 16 ![]() ). On a scatter plot of angle P1-P2
versus angle P0-P1, V. alencasteri
plots in the same field as V. martini
and V. macgillavryi (Fig. 16
). On a scatter plot of angle P1-P2
versus angle P0-P1, V. alencasteri
plots in the same field as V. martini
and V. macgillavryi (Fig. 16 ![]() ). On a
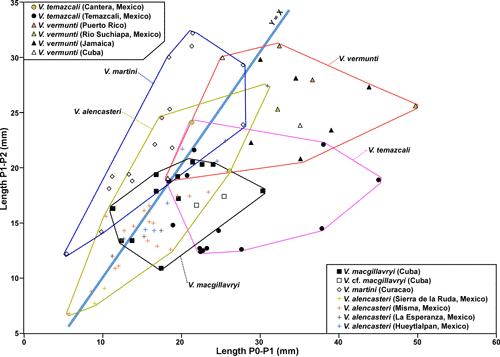
scatter plot of Length P1 versus Length P0, V.
alencasteri plots in a similar field to V.
macgillavryi, but a different field to V.
martini (Fig. 17
). On a
scatter plot of Length P1 versus Length P0, V.
alencasteri plots in a similar field to V.
macgillavryi, but a different field to V.
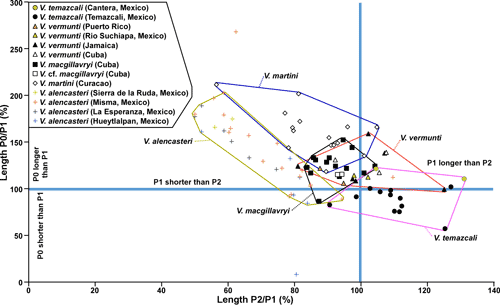
martini (Fig. 17 ![]() ). The most definitive scatter plot is Length P0/P1 versus Length P1/P2 (Fig. 18
). The most definitive scatter plot is Length P0/P1 versus Length P1/P2 (Fig. 18 ![]() ), where V.
alencasteri plots in a field with only minimal overlap with other species (although
there are two outliers), indicating that this species can be distinguished
statistically from other species of American Vaccinites.
), where V.
alencasteri plots in a field with only minimal overlap with other species (although
there are two outliers), indicating that this species can be distinguished
statistically from other species of American Vaccinites.
|
|
|
|
|
|
|
|
|
|
Remarks.
The paucity of details of the myocardinal arrangement make a clear assessment of
V. alencasteri difficult. It shares
many characters in common with V. martini
and V. macgillavryi, but can be
distinguished by the relative lengths of its pillars and specimens largely plot
in a separate field from the last two species on a scatter plot of P0/P1 versus P2/P1
(Fig. 18 ![]() ). Even on this plot two outliers may represent
deformed individuals, or maybe they represent other species. Because of the
distinctiveness of the population on this scatter
plot, it is considered a separate species.
). Even on this plot two outliers may represent
deformed individuals, or maybe they represent other species. Because of the
distinctiveness of the population on this scatter
plot, it is considered a separate species.
Age and Distribution. The age of this species is difficult to determine because it occurs on carbonate platforms with few associated biostratigraphically useful fossils. The only associated rudist is found at Hueyitlalpan, where it is associated with Durania curasavica (Oviedo, 2005), which appears to be long ranging species. In the Guerrero-Morelos basin in southern Mexico V. alencasteri (under the name V. gosaviensis - identified by Gloria Alencáster) occurs in the upper part of the Cuautla Formation, which was assigned to the Marginotruncana sigali planktic foraminiferal zone of late Turonian age (Aguilera-Franco, 2003; Aguilera-Franco & Hernández-Romano, 2004, Fig. 8). The species can therefore be considered as of probable late Turonian or early Coniacian age.
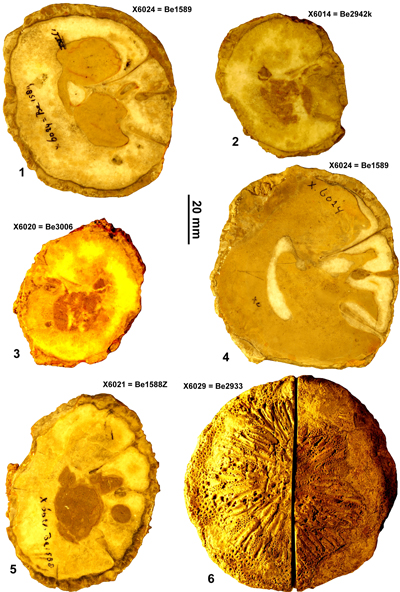
Vaccinites martini Mac Gillavry, 1932
. 1932 Vaccinites martini n.sp.; Mac Gillavry, p. 381-382, Figs. 1-2, Pl. 1, figs. 1-6.
. 1971 Pseudovaccinites martini Mac Gillavry; Dommelen, p. 22-24, Fig. 2.A-D, Pl. 1, figs. 1-2.
Diagnosis. A small/medium sized species of Vaccinites with a relatively short length P0-P1 (mm) versus length P1 to P2 (mm) and a shorter P1 compared to P2 (mm).
Type specimens. The holotype was not found in the collections of Naturalis, Leiden, The Netherlands when I searched in 2010 and must be assumed to be lost. Mac Gillavry (1932) stated that he had five specimens. None of these are in the collections at Naturalis. Dommelen (1971) provided measurements for an additional 16 specimens, of which 11 are still present in Naturalis. Ten specimens are figured in this paper for recording purposes. Specimen number X6024 (= Be1589) has the best preserved myocardinal arrangement and is designated neotype here.
|
|
|
|
Description. A small- to medium-sized species of Vaccinites with a maximum diameter of up to 123 mm and length of 100 mm. The RV is conical to cylindro-conical and the transverse section is generally slightly elliptical. The outer surface of the RV is ornamented with triangular, longitudinal ribs with a spacing between ribs of about 2 mm. The outer shell layer is relatively thick (up to 13 mm), compact, with three well-defined pillars. P0 is long, narrow, generally parallel-sided, and gently curves away from the other pillars; its termination is generally rounded and the inner end is only occasionally slightly expanded. P1 is strongly pinched and has a radially elongate to nearly circular, large knob at the end. P2 is more elongate than P1, is strongly pinched, and has a radially elongate knob, which is thicker towards its innermost end. The inner surface of the RV may be either smooth, or bear weak orimentary folds. The inner shell layer is very thin. The lower part of the body cavity is filled with inner shell layer material; in most specimens this is recrystallized, but in some specimens it shows an indistinct tabulate to coarsely vesicular structure. The fill of the body cavity forms the enclosed sockets for the teeth and the cavity for the posterior myophore. The left valve has a reticulate pore system with the pores divided by trabeculae into porules in well-preserved specimens.
Measurements.
Univariate and bivariate (Scatter) plots of characters for V.
martini are presented in Figure 14 ![]() .
In scatter plots of angle P2-P1-PM versus angle P0-P1 (Fig. 16
.
In scatter plots of angle P2-P1-PM versus angle P0-P1 (Fig. 16 ![]() ) and Angle P1-PM-PT versus Angle
P2-P1-PM (Fig. 15
) and Angle P1-PM-PT versus Angle
P2-P1-PM (Fig. 15 ![]() ), V. martini only
partially overlaps the field of V.
macgillavryi and does not come close to fields of any other species. A
scatter plot of Angle PM-PT-AT versus Angle P2-P1-PM (Fig. 15
), V. martini only
partially overlaps the field of V.
macgillavryi and does not come close to fields of any other species. A
scatter plot of Angle PM-PT-AT versus Angle P2-P1-PM (Fig. 15 ![]() ) greatly overlaps
fields of other species. However, in scatter plots of Length
P1 versus Length P0 (Fig. 17
) greatly overlaps
fields of other species. However, in scatter plots of Length
P1 versus Length P0 (Fig. 17 ![]() ) and Lengths P0/P1 versus P1/P2 (Fig. 18
) and Lengths P0/P1 versus P1/P2 (Fig. 18 ![]() )
the species is in distinct fields with limited overlap.
)
the species is in distinct fields with limited overlap.
Discussion.
V. martini is most easily
distinguished from other species based on the lengths and relative lengths of
its pillars (Figs. 17 ![]() - 18
- 18 ![]() ). In terms of its myocardinal arrangement it shares
similar features with V. macgillavryi
(Fig. 15
). In terms of its myocardinal arrangement it shares
similar features with V. macgillavryi
(Fig. 15 ![]() ).
).
Age and Distribution. Vaccinites martini has only been found in the Seru Treinchi Limestone in Curaçao. Here it is associated with Durania curasavica (Martin). The geological succession on Curaçao consists of an older Curaçao Lava Formation with plateau affinity overlain unconformably by a thick sedimentary succession belonging to the Knip Group (Beets in Matsumoto, 1966; Beets, 1972; Wright & Wyld, 2010; Loewen et al., 2013). The Curaçao Lava Formation has generally been attributed to the Caribbean Large Igneous Province (CLIP), but recent 40Ar/39Ar dates cluster around 92 Ma (early Turonian), 88-84 Ma (early Coniacian to mid Santonian), 80-75 Ma (mid Campanian) and 70-63 Ma (mid Maastrichtian to early Paleocene) (Loewen et al., 2013), but these ages conflict with ages derived from fossils from the Knip Group. The Knip Group rests unconformably on the Curaçao Lava Formation above a soil layer indicating prior subaerial exposure (Wright & Wyld, 2010). The lower part of the Knip Group contains lenses of limestone in different places, three of which have yielded fossils (Beets in Matsumoto, 1966; Beets, 1972). Beets (in Matsumoto, 1966; 1972) discussed in some detail the stratigraphy of the Knip Group of Curaçao, which yielded the rudist material. The Zevenbergen Limestone lense contains a rudist fauna including Torreites tschoppi and fragments of Radiolites sp. and Plagioptychus sp. (Beets in Matsumoto, 1966; Beets, 1972, p. 27; Dommelen, 1971). The Casabao Limestone lenses yield abundant examples of the larger benthic foraminiferan Pseudorbitoides curacaoensis (Krijnen, 1967, 1972). Both of these limestone lenses were interpreted as forming in place by Beets (1972). The Zevenbergen Limestone contains V. martini and D. curacivici (Mac Gillavry, 1932; Beets in Matsumoto, 1966; Beets, 1972; Dommelen, 1971), and Beets (1972, p. 66-67) considered these lenses as slide blocks as they were associated with slump breccias. P. curacaoensis is a primitive form of Pseudorbitoides that has a long primary spire; it is more primitive than P. chubbi (Krijnen, 1972) that occurs in the late early Campanian (zone CC19a) in Jamaica (Krijnen et al., 1993; Jiang, 1993). The presence of P. curacaoensis in the Casabao Limestone lenses therefore indicates an early to mid-early Campanian age. The lack of larger benthic foraminifers in the limestone lenses with the rudists suggests that these limestones are older. The presence of Vaccinites, by analogy with the similar European species V. inaequicostatus has been used to argue a Campanian age (Mac Gillavry, 1937; Dommelen, 1971), but V. inaequicostatus ranges from Santonian to Lower Campanian in Europe (Steuber, 1999). It would therefore seem that the rudist-bearing limestones lenses are most likely Santonian and the Pseudorbitoides limestone lens is probably of early early Campanian age. So T. tschoppi (=T. cf. sanchezi of Chubb, 1971 = T. chubbi of Grubić, 1979) is probably of Santonian age in Curaçao and Cuba and of early early Campanian age in Jamaica and Puerto Rico. In fact, the size of specimens of T. tschoppi increases progressively from Curaçao to Cuba to Jamaica/Puerto Rico, which if stratigraphically related (as seems plausible) may suggest that the Curaçao locality is the oldest, followed by the Cuban locality and with the Jamaican/Puerto Rican localities being the youngest. This is in line with changing features in the species of Vaccinites found at each locality and suggests that they could form a single evolving lineage. V. martini is therefore difficult to assign an age to, but an early to mid Santonian age seems most probable.
Vaccinites macgillavyri Palmer, 1933
. 1933 Vaccinites macgillavryi n.sp.; Palmer, p. 97-98, Pl. 4, fig. 1; Pl. 5, figs. 3-4.
. 1937 Vaccinites inaequicostatus macgillavryi (Palmer); Mac Gillavry, p. 111-118, Pl. 4, figs. 1-8; Pl. 10, fig. 2.
. 1937 Vaccinites sp.; Vermunt, p. 265-266, Fig. 2.k.
. 1937 Vaccinites cf. inaequicostatus macgillavryi (Palmer); Mac Gillavry, p. 118.
. 1971 Pseudovaccinites inaequicostatus macgillavryi (Palmer); Dommelen, p. 24, Figs. 4.B-D, .G.
|
|
|
|
Diagnosis. A small/medium sized species of Vaccinites with a relatively short medium length from P0-P1 (mm) and a relatively shorter lenth from P1-P2 (mm).
Type specimens. The type specimen (P222) was figured by Palmer (1933).
Material. Mac Gillavry (1937) had 22 specimens from the type locality, Loma Yucatán, but only nine specimens were found in the collections at Leiden, The Netherlands. The other specimens are presumably lost.
Description. A relatively small species of Vaccinites with a maximum diameter of 70 mm, with an elongate conical to cylindrical RV, and a low elevated conical LV. The surface of the RV is ornamented with strong triangular ribs spaced at 2-5 mm. Transverse sections are round, weakly elliptical or somewhat irregular. The outer shell layer of the RV is thick, apparently compact, and there are three well defined pillars. P0 is long, straight to gently curved (away from P1 and P2), and generally has a rounded tip. It may be weakly conical, parallel-sided or swell a little towards its inner end. P1 is moderately short, has a strongly rounded tip and ranges from slightly pinched to strongly pinched. P2 is generally longer than P1, but shorter than P0, has a rounded tip and is generally moderately to strongly pinched. The inner surface of the RV generally has very gentle orimentary folds. The inner shell layer is very thin. The lower part of the body cavity is filled with inner shell-layer material, but has suffered too much recrystallization to determine if it was vesicular or not. The fill of the body cavity forms the enclosed sockets for the teeth and the cavity for the posterior myophore. Complete LVs are not known, but the myocardinal features are preserved in place in most specimens. The PT is about 25% the size of the AT and about 50% the size of the MP. The teeth are broadly rounded and the PM is elliptical to triangular, with the one side of the triangle parallel to the inner wall of the body chamber; sometimes it may be a little forked as in Torreites tschoppi. From sections cut through incrusted specimens, Mac Gillavry (1937, p. 112) concluded that this species had subreticulate pores.
Measurements.
Vaccinites macgillavryi is the most
difficult species of Vaccinites to
distinguish based on statistics because its field overlaps with other species (particularly
V. martini and V.
vermunti) in the various scatter plots (Figs. 15 ![]() -
16
-
16 ![]() -
17
-
17 ![]() -
18
-
18 ![]() ). It is most easily
distinguished from these two species in Dommelen's
(1971, Fig. 3, reproduced here as Fig. 3
). It is most easily
distinguished from these two species in Dommelen's
(1971, Fig. 3, reproduced here as Fig. 3 ![]() ) plotting lengths between P0-P1 against
P1-P2, where there is relatively little overlap (although re-measurements of
specimens preserved in Naturalis are less distinct: Fig. 17
) plotting lengths between P0-P1 against
P1-P2, where there is relatively little overlap (although re-measurements of
specimens preserved in Naturalis are less distinct: Fig. 17 ![]() ).
).
Discussion.
Vaccinites cf. macgillavryi of Mac Gillavry
(1937) (=Vaccinites sp. Vermunt,
1937) falls within the same fields as V.
macgillavryi in most scattered plots (Figs. 15 ![]() -
16
-
16 ![]() -
17
-
17 ![]() -
18
-
18 ![]() ), only falling just
outside the field for V. macgillavryi in the plot of angle P2-P1-PM
versus angle P0-P1 (Fig. 16
), only falling just
outside the field for V. macgillavryi in the plot of angle P2-P1-PM
versus angle P0-P1 (Fig. 16 ![]() ). It does not fall close to the fields of any other
American species of Vaccinites when
all the plots are considered, and the tentative assignment of this species to V.
macgillavryi by Mac Gillavry (1937),
and definite assignment by Dommelen
(1971), seems to be correct.
). It does not fall close to the fields of any other
American species of Vaccinites when
all the plots are considered, and the tentative assignment of this species to V.
macgillavryi by Mac Gillavry (1937),
and definite assignment by Dommelen
(1971), seems to be correct.
Age and Distribution. Vaccinites macgillavryi is recorded from Loma Yucatán, Camaguey Province (Palmer, 1933; Mac Gillavry, 1937; Dommelen, 1971; Rojas et al., 1996), and V. cf. macgillavryi from near Bramales, Pinar del Rio Province (Vermunt, 1937; Mac Gillavry, 1937), Cuba. In Loma Yucatán it is associated with Torreites tschoppi Mac Gillavry, 'Pironaea' corrali Palmer, Durania lopeztrigoi (Palmer), D. curasavica (Martin) and plagioptychids, but there is a notable absence of orbitoidal foraminifers and Sulcoperculina (Palmer, 1933; Mac Gillavry, 1937; Dommelen, 1971). Mac Gillavry (1937, p. 118) assigned this locality to the Campanian. Iturralde Vinent & de la Torre (1990) referred the Loma Yucatán locality to the Santonian. Rojas et al. (1996, p. 280) also assigned the Loma Yucatán locality to the Santonian, based on an association of V. macgillavryi with Santonian ammonites in the Las Moscas Member of the Jarao Formation, Sancti Spiritus Province, Cuba. The occurrence of V. macgillavryi near Bramales does not help with determining the age, as these specimens were recovered from a conglomerate (probably at the base of the Maastrichtian: Mac Gillavry, 1937) and are undoubtedly reworked from older deposits. It would appear therefore that both V. martini and V. macgillavryi are of Santonian age.
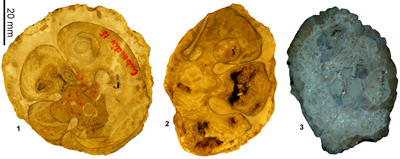
Vaccinites vermunti Mac Gillavry, 1937
v. 1937 Vaccinites macgillavryi Palmer; Vermunt, p. 266, Fig. 2.g-h.
*v. 1937 Vaccinites inaequicostatus vermunti Mac Gillavry, p. 118, Pl. 5, fig. 2.
v. 1968 Vaccinites eyrei n.sp.; Chubb, p. 29.
v. 1971 Vaccinites eyrei Chubb; Chubb, p. 206, Pl. 50, figs. 1-3.
v. 1971 Pseudovaccinites inaequicostatus vermunti (MacGillavry); Dommelen, p. 25, Fig. 4.E-F.
. 1990 Vaccinites inaequicostatus vermunti Mac Gillavry; Alencáster, p. 64, Pl. 2, fig. 6.
v. 1990 Vaccinites inaequicostatus vermunti Mac Gillavry; Alencáster & Michaud, p. 180, Pl. 1, figs. 1-3; Pl. 2, figs. 1-2.
|
|
|
|
|
|
Diagnosis. A large sized species of Vaccinites with a large length P0-P1 (mm) versus a medium length P1-P2 (mm).
Type specimens. Mac Gillavry (1937, p. 119)
designated specimen W 11 from locality H, 0.5 km S. of San Diego de los Bańos,
Pinar del Rio Province, Cuba, as the type specimen and is therefore the holotype.
This specimen was not found in the collection at Naturalis, Leiden, and is
presumably lost. As such, specimen W10 (Fig. 24.2 ![]() ) from locality M 966, 3 km WSW
of San Diego de los Bańos, Pinar del Rio Province, Cuba, is designated as
neotype, because it formed part of the type series of Mac
Gillavry (1937) and is the only other specimen of known province (the
third and only other specimen in the 'type series' was collected by Dr. Tschopp
from an unknown locality).
) from locality M 966, 3 km WSW
of San Diego de los Bańos, Pinar del Rio Province, Cuba, is designated as
neotype, because it formed part of the type series of Mac
Gillavry (1937) and is the only other specimen of known province (the
third and only other specimen in the 'type series' was collected by Dr. Tschopp
from an unknown locality).
Material. Vaccinites vermunti seems to be the most widespread species of Vaccinites in the Americas and has been collected from Cuba (material of Mac Gillavry, 1937; and material in NHMC), Puerto Rico (material from the Cotui and lower Parguera limestones preserved in UPR), Jamaica (Clifton Limestone, material in the USNM, NL and UWIGM) and in Mexico (material in IGM). This material was examined for this work.
Description. A large species of Vaccinites which can reach a diameter of 150 mm and has a conical to cylindrical RV. The outer shell layer of the RV is ornamented with moderately sharp, longitudinal, triangular ridges with a spacing (crest-to-crest) of between 3 and 6 mm. The outer shell layer is up to 13 mm thick and is infolded into the shell as three pillars. P0 is very variable ranging from thin, straight and parallel side, to parallel-sided and curved, to having a strongly swollen tip resembling the other pillars. P1 is relatively long, but not as long as P2; it is strongly pinched with a long thin section and a weakly elliptical, radially elongated knob. P2 is very similar in shape to P1, but is slightly longer. Orimentary folds on the inside of the outer shell layer range from absent to rather strongly developed. The inner shell layer is thin and the lower part of the body cavity is filled with vesicular inner shell material. The LV is gently convex and has a well-developed reticular pore system (specimens from Puerto Rico). The myocardinal arrangement is preserved in specimens from Cuba, Mexico and Jamaica. The PT is smaller than the AT and the PM is triangular to fork-shaped as in T. tschoppi.
Measurements.
Univariate and bivariate plots for measured characters of V.
vermunti are shown in Figures 14 ![]() -
15
-
15 ![]() -
16
-
16 ![]() -
17
-
17 ![]() -
18
-
18 ![]() .
On scatter plots of Angle P2-P1-PM versus Angle P0-P1 (Fig. 16
.
On scatter plots of Angle P2-P1-PM versus Angle P0-P1 (Fig. 16 ![]() ), and Length P1-P2 versus Length
P0-P1 (Fig. 16
), and Length P1-P2 versus Length
P0-P1 (Fig. 16 ![]() ), V. vermunti plots in
a field that overlaps only partially the field of V. macgillavryi. On a scatter plot of pillar length rations (P0/P1
versus P1/P2: Fig. 18
), V. vermunti plots in
a field that overlaps only partially the field of V. macgillavryi. On a scatter plot of pillar length rations (P0/P1
versus P1/P2: Fig. 18 ![]() ), V. vermunti
falls in a different field from V. temazcali.
), V. vermunti
falls in a different field from V. temazcali.
Remarks.
V. vermunti shows a close relationship
with V. macgillavryi, but the
population from the Clifton Limestone falls in a different field from the
population of V. macgillavryi from
Loma Yucatán on scatter plots of Angle P2-P1-PM
versus Angle P0-P1 (Fig. 15 ![]() ), and Length P0-P1 versus Length P1-P2
(Fig. 17
), and Length P0-P1 versus Length P1-P2
(Fig. 17 ![]() ).
The
Mexican specimens from Rio Suchiapa (Alencáster
& Michaud, 1990) plot within the same field as specimens of V.
vermunti from Jamaica in all the plots and clearly represent an equivalent
population (Figs. 14
).
The
Mexican specimens from Rio Suchiapa (Alencáster
& Michaud, 1990) plot within the same field as specimens of V.
vermunti from Jamaica in all the plots and clearly represent an equivalent
population (Figs. 14 ![]() -
15
-
15 ![]() -
16
-
16 ![]() -
17
-
17 ![]() -
18
-
18 ![]() ). The two Puerto Rican specimens lack details of the
myocardinal arrangements, but plot within the field of V.
vermunti from Jamaica (as well as the field of V.
macgillavryi from Cuba) in plots of Angle P1-P2 versus Angle P0-P1 (Fig. 16
). The two Puerto Rican specimens lack details of the
myocardinal arrangements, but plot within the field of V.
vermunti from Jamaica (as well as the field of V.
macgillavryi from Cuba) in plots of Angle P1-P2 versus Angle P0-P1 (Fig. 16 ![]() ), Length P1
versus Length P0
(Fig. 17
), Length P1
versus Length P0
(Fig. 17 ![]() ), relative lengths
of P0/P1 versus P1/P2 (Fig. 18
), relative lengths
of P0/P1 versus P1/P2 (Fig. 18 ![]() ), and are best attributed to V.
vermunti based on their size and their faunal associations.
), and are best attributed to V.
vermunti based on their size and their faunal associations.
Age and Distribution. Vaccinites vermunti from Cuba comes from location H961 (Vermunt, 1937), but is not associated with other rudists. The Jamaican forms from the Clifton Limestone are associated with submoniliform hippuritids (B. ruseae and W. luceae) and the Clifton Limestone has been attributed to the lower lower Campanian (Mitchell, 2020b). The specimens from Rio Suchiapa were also found isolated without other rudists (Alencáster & Michaud, 1990). In Puerto Rico, the material attributed to V. vermunti comes from the Cotui Limestone (Krushensky, 2001), where it is associated with Barrettia sp. (small), W. luceae, Torreites tschoppi, radiolitids and antillocaprinids. V. vermuti is also in the lower (Bahía Fosforescente) member of the Parguera Limestone (Volckmann, 1984), where it is associated with Barrettia sp. ind. and antillocaprinids. The similarity of the rudist assemblages between the Cotui Limestone and Clifton limestone (both containing W. luceae and V. vermunti) is notable, suggesting a similar age.
Vaccinites temazcali n.sp.
(Fig. 26 ![]() )
)
. 2005 Vaccinites vermunti Mac Gillavry; Oviedo, p. 77-80, Fig. 58.1-3.
. 2005 Vaccinites n.sp.; Oviedo, p. 80-82, Fig. 60.1-13.
. 2006 Vaccinites n.sp.; Oviedo et al., p. 62.
. 2007 Vaccinites n.sp.; Oviedo et al., p. 310.
. 2010 Vaccinites vermunti Mac Gillavry; Pons et al., p. 983-987, Figs. 12.1-12.8, 13.1-13.10.
|
|
Origin of name. From the locality of Temazcal in Mexico from where the species was described under the name V. vermunti by Pons et al. (2010).
Diagnosis. A medium sized species of Vaccinites with a long P1, relatively short P0 and by the large Angle P2-P1-PM.
Type specimens. Holotype: IGM 9537 (PUAB 0919; figured by Pons et al., 2010, Fig. 12.1-7); Paratypes: PUAB 81015, 'Hacienda El Durazno'; PUAB 81004, 'Arroyo Temazcal'; PUAB 1288 717, 'Hacienda El Durazno'; PUAB 81010, 'Cerro La Mesa'; PUAB 1288 715, 'Hacienda El Durazno'; PUAB 81014, 'Cerro La Mesa'; PUAB 81003, 'Arroyo Temazcal'; PUAB 0919 685, 'Hacienda El Durazno'; PUAB 1288 716, 'Hacienda El Durazno'; PUAB 0921 649, 'El Potrero.'; figured by Pons et al., 2010, Fig. 13.1-10, respectively). All specimens from the 'Temazcal' Limestone, about 100 km NNE of the city of San Luis Potosi, San Luis Potosi State, Mexico (Pons et al., 2010).
Description. A medium sized species of Vaccinites with a diameter up to 75 mm. RV conical to cylindro-conical and ornamented with longitudinal ridges with a spacing of 3.5 to 4.5 mm. The RV inner shell layer has a thickness of up to about 8 mm and is folded into the shell as three strong pillars. P0 is relatively short, shorter than P1, is moderately wide, curved away from P1 and P2, and generally has a swollen radially elongated end, which is rounded. P1 is equal to or longer than P2, strongly pinched, with an almost circular knob. P2 is similar to P1. The interior ranges from smooth to showing very weak orimentary folds. The inner shell layer is thin and the lower part of the body cavity is filled with vesicular shell structure. The sockets for the teeth are formed from the vesicular fill of the body chamber as is the cavity for the PM. The LV is a low dome and has a reticulate pore system (Pons et al., 2010). The PT is about 25% of the size of the AT and about the same size as the PM. The PM may be radially elongate to triangular shaped. The long P1 and position of PM close to the posterior shell wall means that the angle P2-P1-PM is large, and this is a distinctive feature of the species.
Measurements.
Univariate summary statistics are show in Figure 14 ![]() and bivariate statistics, in Figures 15
and bivariate statistics, in Figures 15 ![]() -
16
-
16 ![]() -
17
-
17 ![]() -
18
-
18 ![]() . The type population of V.
temazcali falls in distinct fields on plots of Lengths of P0/P1 versus P1/P2
(Fig. 18
. The type population of V.
temazcali falls in distinct fields on plots of Lengths of P0/P1 versus P1/P2
(Fig. 18 ![]() ), because of the relatively long P1 and short
P0, and Angles P2-P1-PM vs. P0-P1 (Fig. 16
), because of the relatively long P1 and short
P0, and Angles P2-P1-PM vs. P0-P1 (Fig. 16 ![]() ), Angles P1-PM-PT
versus P0-P1 (Fig. 16
), Angles P1-PM-PT
versus P0-P1 (Fig. 16 ![]() ), Angles PM-PT-AT versus P2-P1-PM
(Fig. 15
), Angles PM-PT-AT versus P2-P1-PM
(Fig. 15 ![]() ), and Angles P1-PM-PT versus P2-P1-PM
(Fig. 15
), and Angles P1-PM-PT versus P2-P1-PM
(Fig. 15 ![]() ), due to the
large Angle P2-P1-PM. The small population of specimens from Cantera (Oviedo,
2005) plots in the same field as the type population in scatter plots of Angles P2-P1-PM versus P0-P1
(Fig. 16
), due to the
large Angle P2-P1-PM. The small population of specimens from Cantera (Oviedo,
2005) plots in the same field as the type population in scatter plots of Angles P2-P1-PM versus P0-P1
(Fig. 16 ![]() ), Angles PM-PT-AT
versus P2-P1-PM (Fig. 15
), Angles PM-PT-AT
versus P2-P1-PM (Fig. 15 ![]() ), and
Angles P1-PM-PT versus P2-P1-PM (Fig. 15
), and
Angles P1-PM-PT versus P2-P1-PM (Fig. 15 ![]() ), and close to the same field in the
plot of and Angles P2-P1-PM vs. P0-P1 (Fig. 16
), and close to the same field in the
plot of and Angles P2-P1-PM vs. P0-P1 (Fig. 16 ![]() ). However, in plots of Lengths of
P0/P1 versus P1/P2 (Fig. 18
). However, in plots of Lengths of
P0/P1 versus P1/P2 (Fig. 18 ![]() ), the Cantera population plots within the
field of V. vermunti because P0 is
longer than P1. The Cantera population may therefore be transitional between V.
vermunti and V. temazcali, but is attributed to V. temazcali based on the large Angle P2-P1-PM.
), the Cantera population plots within the
field of V. vermunti because P0 is
longer than P1. The Cantera population may therefore be transitional between V.
vermunti and V. temazcali, but is attributed to V. temazcali based on the large Angle P2-P1-PM.
Remarks. Vaccinites temazcali is easily distinguished from all other American species of Vaccinites based on its long P1 and relatively short P0 and by the large Angle P2-P1-PM.
On
scatter plots of the length of P1 versus the length of P0 (Fig. 18 ![]() ), V.
temazcali n.sp. plots in a distinct field with large P2/P1 and relatively
small P0/P1. On scatter plots of Pm-PT-AT vs. P2-P1-PM and P1-PM-PT vs. P2-P1-PM
(Fig. 15
), V.
temazcali n.sp. plots in a distinct field with large P2/P1 and relatively
small P0/P1. On scatter plots of Pm-PT-AT vs. P2-P1-PM and P1-PM-PT vs. P2-P1-PM
(Fig. 15 ![]() ), V. temazcali
n.sp. plots in a field with large P2-P1-PM angles
which only minimally overlaps with V.
vermunti.
), V. temazcali
n.sp. plots in a field with large P2-P1-PM angles
which only minimally overlaps with V.
vermunti.
There
is a little overlap in characters of V.
temazcali n.sp. with specimens of V.
vermunti from Cuba. The two remaining syntypes of V. vermunti from Cuba and an additional specimen (NHMC no. 126:
Fig. 24.3 ![]() ) plot just within the field of V.
temazcali n.sp. on scatter plots of
Angle PM-PT-AT versus Angle P2-P1-PM (Fig. 15
) plot just within the field of V.
temazcali n.sp. on scatter plots of
Angle PM-PT-AT versus Angle P2-P1-PM (Fig. 15 ![]() ) and Angle P1-PM-PT versus Angle P2-P1-PM
(Fig. 15
) and Angle P1-PM-PT versus Angle P2-P1-PM
(Fig. 15 ![]() ), or between the
fields of V. vermunti from Jamaica and
V. temazcali n.sp. on scatter plots of Angle P1-PM-PT versus
Angle P0-P1 (Fig. 16
), or between the
fields of V. vermunti from Jamaica and
V. temazcali n.sp. on scatter plots of Angle P1-PM-PT versus
Angle P0-P1 (Fig. 16 ![]() ). The Cuban specimens plot clearly within the field of
Jamaican specimens in plots of Angle P1-P2 versus Angle P0-P1 (Fig. 16
). The Cuban specimens plot clearly within the field of
Jamaican specimens in plots of Angle P1-P2 versus Angle P0-P1 (Fig. 16 ![]() ), Length
P1-P2 versus Length P0-P1 (Fig. 17
), Length
P1-P2 versus Length P0-P1 (Fig. 17 ![]() ) and relative lengths of P0/P1 and
P2/P1 (Fig. 18
) and relative lengths of P0/P1 and
P2/P1 (Fig. 18 ![]() ). Thus the Cuban specimens are better associated with the
Jamaica population, and V. temazcali
n.sp. is considered a distinct species with only minimal overlap with V. vermunti.
). Thus the Cuban specimens are better associated with the
Jamaica population, and V. temazcali
n.sp. is considered a distinct species with only minimal overlap with V. vermunti.
Age and Distribution. This species has only been reported from Mexico, with populations from the 'Temazcal' Limestone in the state of St Luis Potosi (Pons et al., 2010), and Cantera in the state of Chiapas (Ovideo, 2005). The fauna from the Temazcal Limestone includes Potosites tristantorresi (Pons et al.), Radiolites acutocostata (Adkins), 'Barrettia' cf. ruseae and Torreites sanchezi (Douvillé), together with poorly preserved plagioptychids and antillocaprinids, and was attributed to the early Campanian by Pons et al. (2010). The presence of T. sanchezi, which is recorded from the mid Campanian of Cuba (Mac Gillavry, 1937; Dommelen, 1971; Rojas et al., 1996) and Jamaica (Jung, 1971) in association with the advanced V. temazcali indicates a higher stratigraphic level than the early Campanian assemblages with T. tschoppi and V. vermunti found in Puerto Rico and Jamaica. This suggests that V. temazcali is probably of late early Campanian age, whereas V. vermunti is of early early Campanian age.
Family PLAGIOPTYCHIDAE Douvillé, 1888
Genus PLAGIOPTYCHUS Matheron, 1842
Type species. Plagioptychus paradoxus Matheron, 1842 (=Caprina aguilloni Orbigny, 1840, after Douvillé, 1888), from the Santonian? of Le Beausset, Bouches-du-Rhône, France.
Plagioptychus sp. indet.
Discussion. A single fragmentary specimen of Plagioptyus was seen in limestone at Clifton; it was not collectable or in a suitable position to photograph. Dommelen (1971, p. 25) also reported the presence of Plagioptychus in the Clifton Limestone. Further discussion will have to wait until suitable material is collected.
One of the strange things about the Clifton Limestone is the abundance and relatively high diversity of hippuritids, but a complete lack of radiolitids. This is also true of the Peters Hill Limestone (Mitchell, 2003, 2013), which contains abundant examples of Barrettia coatesi (Chubb, 1971), but only very rare examples of radiolititids (Mitchell, 2009). In contrast, the Cotui Limestone has only rare examples of huppuritids, but common radiolitids and antillocaprinids. The environmental controls on the spatial distribution of hippuritids and radiolitids are not known, but clearly different carbonate platforms in the earliest Campanian were inhabited by different assemblages of rudist bivalves.
The revision of the rudists from the Clifton Limestone and a revision of the genus Vaccinites in the Americas enables a discussion of rudist assemblages from the Turonian to mid Campanian. Dating is relatively problematic for some of the assemblages with Vaccinites, and this might be sorted out by the use of strontium isotopes (e.g., Steuber et al., 2002) since the pillars of Vaccinites are constructed of compact calcite and if diagenetically screened it should be possible to produce reliable ages. Table 2 shows rudist assemblages for the Antillean islands and Mexico with the most likely age of the different assemblages. Lower case letters are used to indicate the relative uncertainty of levels since formal division of the Santonian and Campanian have not been proposed.
Table 2: Rudist assemblages (mainly hippuritids) from Mexico and the Antilles from the Turonian to the mid Campanian with confirmed and suggested levels.
| Probable level | Rudist Assemblage | Locations |
| Campanian (middle) | Moniliform Barrettia (including B. monilifera and B. multilirata, as well as undescribed forms); moniliform Whitfieldiella (W. gigas); Parastroma sanchezi and P. trechmanni; Torreites sanchezi | Jamaica (Back Rio Grande, Stapleton and Haulton Hall limestones). Puerto Rico (upper member Parguera Limestone). Cuba ('Barrettia monilifera fauna'). Mexico (Chiapas) |
| Campanian (upper lower) | Submoniform Barrettia (maybe B. dommeleni); submoniliform Whitfieldiella (maybe including B. cf. ruseae of Pons et al., 2010); Torreites sanchezi; Vaccinites temazcali, D. curasavica | Jamaica (Dry Hill Limestone). Puerto Rico (Botijas Limestone). Mexico (San Luis Potosi and Cantera). |
| Campanian (lower lower) | Submoniform Barrettia (B. coatesi, B. ruseae); submoniliform Whitfieldiella (W. luceae); Torreites tschoppi; Vaccinites vermunti | Jamaica (Clifton and Peters Hill limestones). Mexico (Rio Suchiapa). Cuba (San Diego de los Bańos). Puerto Rico (Cotui and lower Parguera limestones). |
| Santonian (mid? or upper?) | 'Pironaea' coralli; Torreites tschoppi; Vaccinites macgillavryi; Durania curasavica | Cuba (Loma Yucatán). |
| Santonian (lower? or mid?) | Vaccinites martini, D. curasavica | Curaçao (Seru Treinchi Limestone) |
| upper Turonian to lower? Coniacian | Vaccinites alencasteri; Radiolites | Mexico (Guerrero) |
| Turonian (mid? to upper?) | Hippurites mexicanus, Radiolites | Mexico (Guerrero) |
I thank Richard Coutou and Jason Fisher for help with collecting material in the field. I thank the following for access to collections in their care: Jann Thompson and Brian Huber (Smithsonian Institution); the late Ann Molineux (Non-Vertebrate Paleontology Collections Manager, Texas Natural Science Center, The University of Texas at Austin); Reinaldo Rojas-Consuegra (The National Natural History Museum, Havana, Cuba); Dra María del Carmen Perrillat and Violeta A. Romero Mayén (Museum of Paleontology, Institute of Geology, Universidad Nacional Autónoma de México, Ciudad Universitaria, México); W. J. Wildenberg (Naturalis, Leiden, The Netherlands); Hernán Santos (Geological Museum, University of Puerto Rico, Mayagüez; Ian Brown and Sherene James-Williamson (The University of the West Indies Geological Museum, Kingston, Jamaica). Peter Skelton took some of the photographs in the Smithsonian during our joint trip there in 2010, and I am indebted to him for allowing their use here, and the enjoyable discussions on rudist bivalves oven the last 20 years. Part of this work was undertaken during two sabbaticals at the University of the West Indies (August 2010 to July 2011 and August 2019 to January 2021), which is thanked for its support. Funding for the trip to the Non-Vertebrate Paleontology Collections Manager, Texas Natural Science Center, The University of Texas at Austin, in 2009 was supported by NSF Grant No. 0646468 awarded to the Texas Natural Science Center at the University of Texas. Travel to other museums was funded from travel and study grants from the University of the West Indies (Mona, Jamaica). I thank the reviewers (José Maria Pons and Robert Scott) and editor (Robert Scott) for their suggestions for improvement of the original manuscript.
Aguilera-Franco N. (2003).- Cenomanian - Coniacian zonation (foraminifers and calcareous algae) in the Guerrero - Morelos basin, southern Mexico.- Revista Mexicana de Ciencias Geológicas, Ciudad de Mexico, vol. 20, no. 3, p. 202-222.
Aguilera-Franco N. & Hernández-Romano U. (2004).- Cenomanian-Turonian facies succession in the Guerrero-Morelos Basin, Southern Mexico.- Sedimentary Geology, vol. 170, p. 135-162.
Alencáster G., Hernandez-Garcia R. & Garcia-Villegas F. (1987).- Rudistas hippuritidos (Bivalvia-Hippuritacea) del Cretácico Superior de la parte central del estado de Guerrero.- Revista de la Sociedad Mexicana de Paleontologia, Ciudad de Mexico, vol. 1, no. 1, p. 24-39 (2 Pls.).
Alencáster G. & Michaud F. (1990).- Rudista (Bivalvia-Hippuritacea) del Cretécico Superior de la región de Tuxla Gutiérrez, Chiapas (México).- Actas Facultad de Ciencias Tierra UANL, Nuevo León, vol. 4, p. 175-193 (2 Pls.).
Beets D.J. (1972).- Lithology and stratigraphy of the Cretaceous and Danian succession of Curaçao.- Natuurwetenschappelijke Studiekring vor Suriname en de Nederlandse Antillen, Utrecht, no. 70, 153 p. (6 Pls.)
Boissevane H. & Mac Gillavry H.J. (1932).- Some remarks on Barrettia sparcilirata Whitfield and Chiapasella radiolitiformis (Trechmann).- Koninklijke Akademie van Wetenschappen te Amsterdam, Proceedings of the Section of Sciences, vol. 35, p. 1303-1312.
Bronn H.G. (1831).- Hippurites. In: Ersch J.S. & Gruber J.G. (eds.), Allgemeine Encyklopaedie der Wissenschaften und Künste, Leipzig, vol. 2 (8), p. 371-376.
Chubb L.J. (1968).- New rudist species from the Cretaceous rocks of Jamaica.- Journal of the Geological Society of Jamaica, Kingston, vol. 9 (1967), p. 24-31.
Chubb L.J. (1971).- Rudists of Jamaica.- Palaeontographica Americana, Ithaca - NY, vol. 7, p. 157-257.
Dixon I. & Chubb L.J. (1960).- Research at Rusea's. Letter and reply.- Geonotes, Kingston, vol. III, p. 109-111.
Dommelen H. van (1971).- Ontogenetic, phylogenetic and taxonomic studies of the American species of Pseudovaccinites and of Torreites and the multiplefold hippuritids.- Thesis, University of Amsterdam, 125 p.
Donovan S.K., Mitchell S.F. & Lewis D. (2006).- The irregular echinoid Nucleopygus Agassiz from the Upper Cretaceous of western Jamaica-. Cretaceous Research, vol. 27, p. 577-583.
Douvillé H. (1888).- Études sur les caprines.- Bulletin de la Société géologique de France (3e Série), Paris, vol. 16, p. 699-730.
Douvillé H. (1926).- Quelques fossiles du Crétacé supérieur de Cuba.- Compte rendu sommaire des Séances et Bulletin de la Société géologique de France (4e Série), Paris, vol. 26, p. 127-138.
Fischer P. (1887).- Manuel de conchyliologie et de paléontologie conchyliologique ou histoire naturelle des mollusques vivants et fossiles.- F. Savy, Paris, 1369 p. (23 Pls.).
Götz S. & Mitchell S.F. (2009).- The Laluzia armini (gen. et spec. nov.) ecosystem: Understanding a deeper-water rudist lithosome from the Early Maastrichtian of Mexico.- Facies, Erlangen, vol. 55, p. 539-551.
Gray J.E. (1848).- On the arrangement of the Brachiopoda.- Annals and Magazine of Natural History (serie 2), London, vol. 2, p. 435-440.
Grippi J. (1980).- Geology of the Lucea Inlier, western Jamaica.- Journal of the Geological Society of Jamaica, Kingston, vol. 19, p. 1-24.
Grubić A. (1979).- Torreites milovanovici sp. nov. from Oman, T. coxi sp. nov. and T. chubbi sp. nov. from Jamaica, a new description of the genus Torreites Palmer, and a reference to the significance of its palaeogeographic distribution.- Zavod za geoloska i geofizicka Istrazivanja, Vesnik Geologija, Beograd, vol. 37, p. 81-95 (2 Pls).
Grubić A. (2004).- Revision of the rudists subfamily Barrettinae Chubb.- Bulletin de l'Académie serbe des sciences et des arts, Classe des sciences mathématiques et naturelles, Sciences naturelles, Beograd, t. CXXVIII, no. 42, p. 139-197.
Iturralde-Vinent M. & Torre A. de la (1990).- Posición estratigráfica de los rudistas de Camagüey, Cuba. In: Larue D.K. & Draper G. (eds.), Transactions of the 12th Caribbean Geological Conference, St. Croix - U.S. Virgin Islands.- Miami Geological Society, p. 59-67.
Jiang M.M. (1993).- Campanian calcareous nannofossils in the Sunderland Inlier, western Jamaica. In: Wright R.M. & Robinson E. (eds)., Biostratigraphy of Jamaica.- Geological Society of America, Boulder - CO, Memoir 182, p. 19-28.
Jung P. (1971).- Torreites sanchezi (Douvillé) from Jamaica.- Palaeontographica Americana, Ithaca - NY, vol. 7, p. 1-13.
Krijnen J.P. (1967).- Pseudorbitoid Foraminifera from Curaçao.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (B), Amsterdam, vol. 70, p. 144-164
Krijnen J.P. (1972).- Morphology and phylogeny of pseudorbitoid foraminifera from Jamaica and Curaçao, a revisional study.- Scripta Geologica, Leiden, vol. 8, p. 1-133 (27 Pls.).
Krijnen J.P., Mac Gillavry H.J. & Dommelen H. van (1993).- Review of Upper Cretaceous orbitoidal larger foraminifera from Jamaica, West Indies, and their connection with rudist assemblages. In: Wright R.M. & Robinson E. (eds.), Biostratigraphy of Jamaica.- Geological Society of America, Boulder - CO, Memoir, 182, p. 29-63.
Krushensky R.K. (2001).- Geologic map of Puerto Rico with correlation chart and map unit descriptions. In: Bawiec W.J., Marsh S.P., Learned R.E., Krushensky R.D., Schellekens J.H., Griscom A., Kitchen N.E., Cox G.E., McKelvey G.E., Paidakovich M.E., Handler A., Alonzo R., Lipin B.R., Page N.J., Meier A.L., Scanlon K.M., Trias J.L., Rodriguez R.W., Delorey C.M., Poppe L.J., Commeau J.A., Luepke G. & Weaver J.N. (authors), Geology, geochemistry, geophysics, mineral ccurrences, and mineral resource assessment for the Commonwealth of Puerto Rico.- U.S. Geological Survey Open-File Report, OF-98-38, scale 1:100,000.
Loewen M.W., Duncan R.A., Kent, A.J.R. & Krawl K. (2013).- Prolonged plume volcanism in the Caribbean Large Igneous Province: New insights from Curaçao and Haiti.- Geochemistry Geophysics Geosystems, vol. 14, no. 10, p. 4241-4259.
Mac Gillavry H.J. (1932).- The rudist fauna of Seroe Teintje limestone (northern Curacao).- Koninklijke Akademie van Wetenschappen te Amsterdam, Proceedings of the Section of Sciences, vol. 35, p. 381-392.
Mac Gillavry H.J. (1937).- Geology of the province of Camaguey, Cuba, with revisional studies in rudist paleontology.- Geographische en geologische Mededeelingen, Utrecht, vol. 14, 168 p.
Matheron P. (1842).- Catalogue méthodique et descriptif des corps organisés fossiles du département des Bouches-du-Rhône et lieux circonvoisins.- Répertoire des travaux de la Société de Statistique de Marseille, vol. 6, p. 81-341.
Matsumoto T. (1966).- A Cretaceous ammonite from the Island of Curaçao, Netherlands Antilles: With Appendix. Stratigraphic position and age of a Cretaceous ammonite from Curaçao, Netherlands Antilles by D.J. Beets.- Memoirs of the Faculty of Science, Kyūsyū University (Series D, Geology), vol. XVII, no. 3, p. 277-294.
Mitchell S.F. (2003).- Sedimentary and tectonic evolution of central Jamaica. In: Bartolini C., Buffler R.T. & Blickwede J.F. (eds.), The Circum-Gulf of Mexico and the Caribbean: Hydrocarbon habitats, basin formation, and plate tectonics.- American Association of Petroleum Geologists Memoir, Tulsa - OK, vol. 79, p. 605-623.
Mitchell S.F. (2006).- Timing and implications of Late Cretaceous tectonic and sedimentary events in Jamaica.- Geologica Acta, Barcelona, vol. 4, p. 171-178.
Mitchell S.F. (2009).- A new rudist from the Santonian of Jamaica.- Caribbean Journal of Earth Science, Kingston, vol. 40, p. 15-20.
Mitchell S.F. (2010).- Revision of three large species of Barrettia from Jamaica.- Caribbean Journal of Earth Science, Kingston, vol. 41, p. 1-16.
Mitchell S.F. (2013).- The lithostratigraphy of the Central Inlier, Jamaica.- Caribbean Journal of Earth Science, Kingston, vol. 46, p. 31-42.
Mitchell S.F. (2020a).- Cretaceous geology and tectonic assembly of Jamaica. In: Davison I., Hull J.N.F. & Pindell J. (eds.), The basins, orogens and evolution of the southern Gulf of Mexico and northern Caribbean.- Geological Society, London, Special Publications, vol. 504, p. 1-27.
Mitchell S.F. (2020b).- Exceptionally well-preserved silicified hippuritid rudist bivalves from the lower Maastrichtian of Puerto Rico.- Carnets Geol., vol. 20, no. 18, p. 333-366.
Newell N.D. (1965).- Classification of the Bivalvia.- American Museum Novitates, New York, no. 2206, 25 p.
Orbigny A. d' (1840).- Note sur le genre Caprine.- Revue zoologique par la Société Cuvierienne, Paris, 1839, p. 168-170.
Orbigny A. d' (1847).- Sur les Brachiopodes ou Palliobranches (deuxičme mémoire).- Comptes rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, vol. 25, p. 266-269.
Oviedo A. (2005).- Rudistas del Cretácico superior del centro al sureste de México (Recuento sistemático de rudistas americanos).- Ph.D. Thesis, Universitat Autňnoma de Barcelona, 123 p. URL: http://www.tdx.cat/TDX-0620106-232433
Oviedo A., Pons J.M. & Alencáster G. (2006).- Rudistas hippurítidos del centro y sureste de México. In: Memoria del X Congreso Nacional de Paleontología.- Publicaciones Especiales del Instituto de Geología, Universidad Nacional Autónoma de México - UNAM, 5, p. 62.
Oviedo A., Pons J.M., Pichardo-Barrón Y., Aguilar-Pérez J. & Alencáster G. (2007).- Stratigraphical distribution of the hippuritid rudists in the Upper Cretaceous of central Mexico.- In E. Díaz-Martínez & I. Rábano, eds., 4th European Meeting on the Palaeontology and Stratigraphy of Latin America.- Cuadernos del Museo Geominero 8. Instituto Geológico y Minero de Espańa, Madrid, p. 309-313.
Palmer R.H. (1933).- Nuevos rudistas de Cuba.- Revista de Agricultura, Comercio y Trabajo, La Habana, vol. 14(15-16), p. 95-125.
Pons J.M., Vicens E. & García-Barrera P. (2019).- Campanian and Maastrichtian hippuritid rudists (Hippuritida, Bivalvia) of the Chiapas Central Depression (southern Mexico) and implications for American multiple-fold hippuritid taxonomy.- Journal of Paleontology, Tulsa - OK, vol. 93, p. 291-311.
Pons J.M., Vicens E., Pichardo Y., Aguilar J., Oviedo A., Alencáster G. & García-Barrera P. (2010).- A new Early Campanian Rudist fauna from San Luis Potosi in Mexico and its taxonomic and stratigraphic significance.- Journal of Paleontology, Tulsa - OK, vol. 84, p. 974-995.
Rojas R., Iturralde-Vinent M. & Skelton P.W. (1996).- Stratigraphy, composition and age of Cuban rudist-bearing deposits. In: Alencáster G. & Buitrón-Sánchez B.E. (eds.), Third International Conference on Rudists.- Revista mexicana de Ciencias geológicas, Ciudad de Mexico, vol. 12 (1995), p. 272-291.
Schmidt W. (1988).- Stratigraphy and depositional environment of the Lucea Inlier, western Jamaica.- Journal of the Geological Society of Jamaica, Kingston, vol. 25, p. 15-35.
Skelton P.W. (2013).- Rudist classification for the revised Bivalvia volumes of the Treatise on Invertebrate Paleontology.- Caribbean Journal of Earth Science, Kingston, vol. 45, p. 9-33.
Skelton P.W. (2018).- Part N, Volume 1, Chapter 26A: Introduction to the Hippuritida (rudists): Shell structure, anatomy, and evolution.- Treatise Online, Lawrence - KS, vol. 104, p. 1-37.
Skelton P.W. & Smith A.B. (2000).- A preliminary phylogeny of rudist bivalves: Sifting clades from grades. In: Harper E.M., Taylor J.D. & Crame J.A. (eds.), The evolutionary biology of the Bivalvia.- Special Publications of the Geological Society, London, vol. 177, p. 97-127.
Steuber T. (1999).- Cretaceous rudists of Boeotia, central Greece.- Special Papers in Palaeontology, London, vol. 61, p. 1-229.
Steuber T., Mitchell S.F., Buhl D., Gunter G. & Kasper H.U. (2002).- Catastrophic extinction of Caribbean rudist bivalves at the Cretaceous-Tertiary boundary.- Geology, Boulder - CO, vol. 30, p. 999-1002.
Vermunt L.W.J. (1937).- Cretaceous rudists of Pinar del Rio province, Cuba.- Journal of Paleontology, Tulsa - OK, vol. 11, p. 261-275.
Volckmann R.P. (1984).- Geologic map of the Cabo Rojo and Parguera quadrangles, southwest Puerto Rico.- U.S. Geological Survey Miscellaneous Investigations Series, Map I-1557, scale 1:20,000.
Woodward S.P. (1862).- Some account of Barrettia, a new and remarkable fossil shell from the hippurite limestone of Jamaica.- The Geologist: A popular illustrated monthly Magazine of Geology, London, vol. V (October 1862), p. 372-377 (Pls. XX-XXI).
Wright J.E. & Wyld S.J. (2010).- Late Cretaceous subduction initiation on the eastern margin of the Caribbean-Colombian Oceanic Plateau: One Great Arc of the Caribbean (?).- Geosphere, Boulder - CO, vol. 7, p. 468-493.
| Nomenclatural note: Life Sciences Identifier (LSID) http://zoobank.org/References/AACF25AB-6C09-414A-BBA3-477E47CABD01 • Vaccinites alencasteri Mitchell, 2021 http://zoobank.org/NomenclaturalActs/B7C32F9B-9DF8-400C-9A04-86B9E8C2E029 • Vaccinites temazcali Mitchell, 2021 http://zoobank.org/NomenclaturalActs/0D5377E0-8D5D-4B9D-A9FB-6E940F9650A7 • Whitfieldiella luceae Mitchell, 2021 http://zoobank.org/NomenclaturalActs/DFB581B7-F0DC-4B41-BFA9-E25F80027EF9 |