◄ Carnets Geol. 21 (15) ►
![]()
Outline:
[1. Introduction]
[2. Geological overview]
[3. Biostratigraphy]
[4. Material and methods]
[5. Systematics]
[6. Discussion]
[7. Conclusions]
[Bibliographic references] and ...
[Plates]
This work is dedicated to the memory of Rolf Schroeder, an outstanding micropalaeontologist who recently passed away. His numerous works on the Orbitolinidae are basic literature for their taxonomy and his name will always be linked to this group of larger benthic foraminifera.
Lerchenauerstr. 167, D-80935 München (Germany)
Babeş-Bolyai University, Department of Geology and Center for Integrated Geological Studies, M. Kogălniceanu str., 1, 400084 Cluj-Napoca (Romania)
Published online in final form (pdf) on August 17, 2021
DOI 10.2110/carnets.2021.2115
![]()
[Editor:
Daniela Basso; language editor: Stephen Eagar;
technical editor: Bruno R.C. Granier]
![]()
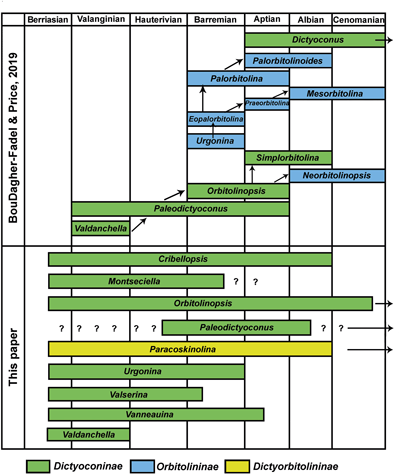
New data from the Carpatho-Balkanides of eastern Serbia evidence the more or less near-simultaneous "explosive" first appearances of several genera of the Orbitolinidae in the late Berriasian. Most of the observed taxa were previously recorded from strata not older than the Late Hauterivian (= classical Urgonian of southeastern France), evidence that these ages refer to local first appearance data. The diversified assemblage from Serbia includes representatives of the subfamilies Dictyoconinae: genera Cribellopsis Arnaud-Vanneau, Montseciella Cherchi & Schroeder, Orbitolinopsis Henson, Urgonina Foury & Moullade, Valserina Schroeder & Conrad, Vanneauina Schlagintweit, and Dictyorbitolininae: genus Paracoskinolina Moullade. Representatives of the Orbitolininae (with complex embryo) have not been observed. They appeared later in the fossil record seemingly during the Late Hauterivian-early Barremian. All together 17 taxa are reported, of which three in open nomenclature. A new species is described as Cribellopsis sudari n. sp. The majority of the observed species display medium- to high-conical tests and a rather simple exoskeleton lacking horizontal partitions (rafters). The new data contradict a phylogenetic evolution of distinct genera displaying different internal test structures one after the other in time (= ancestor-descendant relationships) as postulated by some authors. The explosive radiation ("early burst") of the Orbitolinidae in the late Berriasian is accompanied by the first appearance date of several other large benthic foraminifera including mostly agglutinating (e.g., Ammocycloloculina, Choffatella, Drevennia, Eclusia, Moulladella, Pfenderina, Pseudotextulariella) but also complex porcelaneous taxa (Pavlovcevina) providing evidence for a bioevent in this time period that exceeds the number of taxa originating in the previous (Tithonian) and the following stage (Valanginian). The early evolutionary history of the Orbitolinidae can be considered a classical example of adaptive radiation within the clade's history.
• Foraminifera;
• Orbitolinidae;
• taxonomy;
• phylogeny;
• biostratigraphy
Schlagintweit F. & Bucur I.I. (2021).- The late Berriasian early evolutionary burst of the Orbitolinidae: New insights into taxonomy, origin, diversification and phylogeny of the family based on data from eastern Serbia.- Carnets Geol., Madrid, vol. 21, no. 15, p. 343-382.
La diversification précoce des Orbitolinidae au Berriasien supérieur : Nouvelles perspectives sur la taxonomie, l'origine, la diversification et la phylogénie de la famille basée sur des données de Serbie orientale.- De nouvelles données provenant des Carpatho-Balkanides de Serbie orientale témoignent de premičres apparitions "explosives" plus ou moins simultanées de plusieurs genres d'Orbitolinidae dans le Berriasien supérieur. Précédemment la plupart des taxons observés étaient répertoriés dans des couches gučre plus anciennes que l'Hauterivien supérieur (= Urgonien classique du sud-est de la France), démontrant que ces âges de premičre apparition n'ont de signification que localement. L'assemblage diversifié de Serbie comporte des représentants des sous-familles Dictyoconinae avec les genres Cribellopsis Arnaud-Vanneau, Montseciella Cherchi & Schroeder, Orbitolinopsis Henson, Urgonina Foury & Moullade, Valserina Schroeder & Conrad et Vanneauina Schlagintweit, et Praedictyorbitolininae avec le genre Paracoskinolina Moullade. Aucun représentant des Orbitolininae (ŕ embryon complexe) n'a été observé, cette sous-famille apparaissant plus tardivement dans le registre fossile, apparemment durant l'Hauterivien supérieur-Barrémien inférieur. Au total, 17 taxons sont reconnus, parmi lesquels trois sont laissés en nomenclature ouverte. Une nouvelle espčce est décrite : Cribellopsis sudari n. sp. La majorité des espčces observées montre des tests coniques moyens ŕ hauts et un exosquelette plutôt simple dépourvu de cloisonnettes horizontales ("rafters"). Les nouvelles données contredisent une évolution phylogénétique des différents genres montrant différentes structures internes de test se succédant dans le temps (= relations ancętre-descendant) comme certains auteurs le postulent. L'explosion radiative ("diversification précoce") des Orbitolinidae au Berriasien supérieur s'accompagne de l'événement de premičre apparition (FAD) de plusieurs autres grands foraminifčres benthiques comprenant des taxons essentiellement agglutinants (e.g., Ammocycloloculina, Choffatella, Drevennia, Eclusia, Moulladella, Pfenderina et Pseudotextulariella) mais aussi porcelanés complexes (Pavlovcevina), constituant la preuve d'un bio-événement majeur ŕ cette époque qui dépasse le nombre de taxons apparaissant dans les étages précédents (Tithonien) et suivant (Valanginien). L'histoire évolutive initiale des Orbitolinidae peut ętre considérée comme un exemple classique de radiation adaptative au sein de l'histoire d'un clade.
• foraminifčres ;
• Orbitolinidae ;
• taxinomie :
• phylogénie ;
• biostratigraphie
Lower Cretaceous shallow-water
carbonates of the Carpatho-Balkanides (e.g.,
Haas et al., 2011, Fig. 1) are
outcropping in a wide area south of the Danube River extending from Southern
Romania to Eastern Serbia and to Western Bulgaria. Studies of these
Urgonian-type limestones were made by Jankičević
(1978, 1996)
who focused on the rudist assemblages. Micropalaeontological associations (calcareous
algae, benthic foraminifera) have been investigated by Radoičić
(1978), Bucur et al. (1995,
2020), Sudar et al. (2008), Polavder
and Radulović (2005), Polavder (2014) and Schlagintweit
et al. (2019a, 2019b). The stratigraphy of these shallow-water
carbonates was a matter of discussion in the literature. Part of the limestones
was considered to be Barremian-Aptian in the Geological map sheet Aleksinac (Krstić
et al., 1978, 1980). In contrast
hereto, Radoičić (1978) described calcareous algae from this
area highlighting a "Neocomian" age (see also Bucur et
al., 1995). Polavder (2014) studied the rich orbitolinid assemblages
from the Kamenica section of the Kurilovo anticline. As some of the identified
species were previously known with their "first appearance dates" from the Upper Hauterivian-lower Barremian strata of southeastern France (Clavel et
al., 2014), Polavder (2014) concluded that the upper part of the
studied carbonate sequence belongs to the Upper Hauterivian (Fig.
1 ![]() ). The most
contradictionary fact is the coexistence of typically Berriasian-Valanginian
algal and benthic foraminiferal assemblages together with these "Upper
Hauterivian-lower Barremian" orbitolinids. Starting from these contradictions,
the study of some geological sections from the western part of the
Carpatho-Balkanides (Eastern Serbia) was continued in order to clarify the age
of these deposits through detailed sampling and micropalaeontological
investigations. Bucur et al.
(2020) documented a well diversified assemblage of calcareous algae, benthic
foraminifera, associated with rare calpionellids that indicate a late
Berriasian age. In the framework of this study, rich material of orbitolinids
was obtained from the thin-sections studied but lacking systematic description.
Belonging to the oldest known records of the Orbitolinidae, they provide many
new data and insights on the taxonomy, biostratigraphy, and phylogeny of the
whole group. These results are presented herein.
). The most
contradictionary fact is the coexistence of typically Berriasian-Valanginian
algal and benthic foraminiferal assemblages together with these "Upper
Hauterivian-lower Barremian" orbitolinids. Starting from these contradictions,
the study of some geological sections from the western part of the
Carpatho-Balkanides (Eastern Serbia) was continued in order to clarify the age
of these deposits through detailed sampling and micropalaeontological
investigations. Bucur et al.
(2020) documented a well diversified assemblage of calcareous algae, benthic
foraminifera, associated with rare calpionellids that indicate a late
Berriasian age. In the framework of this study, rich material of orbitolinids
was obtained from the thin-sections studied but lacking systematic description.
Belonging to the oldest known records of the Orbitolinidae, they provide many
new data and insights on the taxonomy, biostratigraphy, and phylogeny of the
whole group. These results are presented herein.
The Lower Cretaceous deposits of the
Getic domain which outcrop in the Southern Carpathians (Romania) in the Reşiţa-Moldova
Nouă zone also continues within the Carpatho-Balkanides (Eastern Serbia)
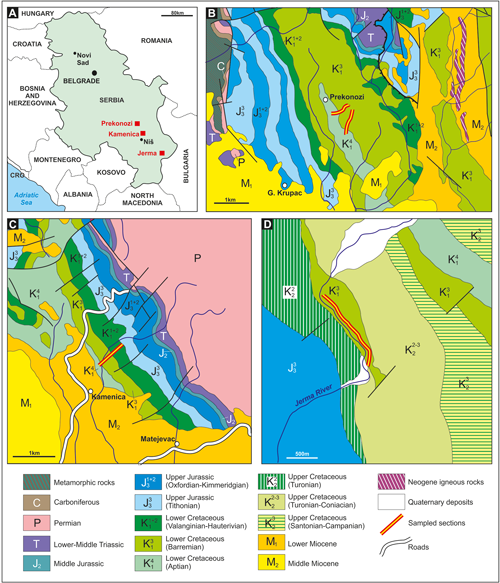
between the Danube and Western Bulgaria (Fig. 1.A ![]() ). The westernmost part of this
zone comprises the folded geological structures Kurilovo and Ozren made of
Paleozoic and Mesozoic rocks. According to the geological map 1:100.000 (sheet
Aleksinac, Krstić et al., 1978,
1980) this area belongs to the NNW-SSE oriented Gornjak-Stuva Planina Unit.
). The westernmost part of this
zone comprises the folded geological structures Kurilovo and Ozren made of
Paleozoic and Mesozoic rocks. According to the geological map 1:100.000 (sheet
Aleksinac, Krstić et al., 1978,
1980) this area belongs to the NNW-SSE oriented Gornjak-Stuva Planina Unit.
The
Kurilovo folded structure (Petković, 1935), including the studied
Kamenica section (Fig. 1.C ![]() ) consists of an asymmetrical anticline associated with
several minor folds (Krstić et
al., 1978, 1980). The core of this structure is made of Paleozoic deposits (Devonian,
Carboniferous, and Permian) being flanked by Mesozoic (Triassic, Jurassic, and
Lower Cretaceous) sequences. According to Krstić et
al. (1980), the Lower Cretaceous deposits are represented by
Valanginian-Aptian carbonates succeded by a sequence of Aptian sandy limestones.
Towards the NW, Lower Cretaceous deposits are outcropping near to Prekonozi (belonging
to the Ozren folded structure) characterized by similar lithostratigraphic
features as in Kurilovo (Fig. 1.B
) consists of an asymmetrical anticline associated with
several minor folds (Krstić et
al., 1978, 1980). The core of this structure is made of Paleozoic deposits (Devonian,
Carboniferous, and Permian) being flanked by Mesozoic (Triassic, Jurassic, and
Lower Cretaceous) sequences. According to Krstić et
al. (1980), the Lower Cretaceous deposits are represented by
Valanginian-Aptian carbonates succeded by a sequence of Aptian sandy limestones.
Towards the NW, Lower Cretaceous deposits are outcropping near to Prekonozi (belonging
to the Ozren folded structure) characterized by similar lithostratigraphic
features as in Kurilovo (Fig. 1.B ![]() ). The age of a part of the limestones from both
structures (considered as Barremian-Aptian on the 1:100,000 geological map-sheet
Aleksinac, Krstić et al., 1978,
1980) was reconsidered by Polavder & Radulović
(2005) and Polavder (2014) as Upper Hauterivian-lower Barremian based
on orbitolinid assemblages. In SE, close to the Serbian-Bulgarian border, the
Lower Cretaceous carbonates outcrop in the Jerma River Canyon. On the geological
map of this area (sheet Breznik, Andelković et
al., 1975, 1977), they have been attributed to the Barremian, forming a
narrow strip, flanked by two faults between an Upper Cretaceous sedimentary
sequence. Based on the dasycladales and benthic foraminifera, the age of the
Jerma River Canyon limestones (Fig. 1.D
). The age of a part of the limestones from both
structures (considered as Barremian-Aptian on the 1:100,000 geological map-sheet
Aleksinac, Krstić et al., 1978,
1980) was reconsidered by Polavder & Radulović
(2005) and Polavder (2014) as Upper Hauterivian-lower Barremian based
on orbitolinid assemblages. In SE, close to the Serbian-Bulgarian border, the
Lower Cretaceous carbonates outcrop in the Jerma River Canyon. On the geological
map of this area (sheet Breznik, Andelković et
al., 1975, 1977), they have been attributed to the Barremian, forming a
narrow strip, flanked by two faults between an Upper Cretaceous sedimentary
sequence. Based on the dasycladales and benthic foraminifera, the age of the
Jerma River Canyon limestones (Fig. 1.D ![]() ) was revised by Bucur et
al. (1995) and assigned to the Berriasian-Valanginian. For the lithological
evolution of the three sections, stratigraphic and micropalaeontological data
reference is made to Bucur et al.
(2020). In all three sections, the
orbitolinids occur together with rare calpionellids in the upper part of the
succession. Generally, the higher energy deposits (e.g., grain-suported fabrics) from the Prekonozi section reflect an
outer platform setting compared to the inner platform facies (e.g.,
mud-supported fabrics) of the Kamenica section. This trend is also suggested by
a less diverse assemblage of orbitolinids. Therefore, the main data included in
the systematic part and the discussion on other aspects such as phylogeny refers
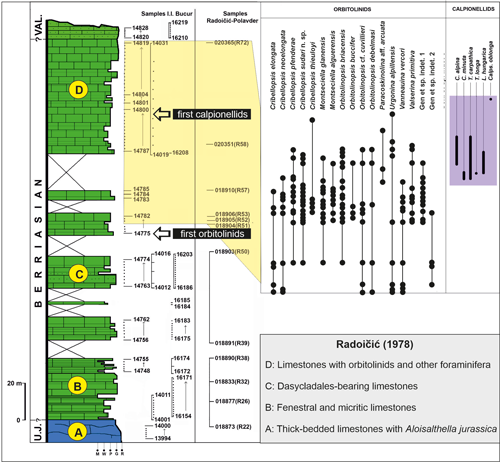
essentially to the Kamenica section (Fig. 2
) was revised by Bucur et
al. (1995) and assigned to the Berriasian-Valanginian. For the lithological
evolution of the three sections, stratigraphic and micropalaeontological data
reference is made to Bucur et al.
(2020). In all three sections, the
orbitolinids occur together with rare calpionellids in the upper part of the
succession. Generally, the higher energy deposits (e.g., grain-suported fabrics) from the Prekonozi section reflect an
outer platform setting compared to the inner platform facies (e.g.,
mud-supported fabrics) of the Kamenica section. This trend is also suggested by
a less diverse assemblage of orbitolinids. Therefore, the main data included in
the systematic part and the discussion on other aspects such as phylogeny refers
essentially to the Kamenica section (Fig. 2 ![]() ).
).
|
Figure 1:
Location of the studied material (from Bucur
et al., 2020, Fig. 1). A. General
sketch map of Serbia with location of the three studied sections. B. Geological
map of the Prekonozi area. Part of the Basic Geological Map of SFRY, 1:100,000,
Sheet Aleksinac (K 34-20) (Kristić et
al., 1978), modified. C. Geological map of the Kamenica area. Part of the
Basic Geological Map of SFRY, 1:100,000, Sheet
Aleksinac (K 34-20) (Kristić et
al., 1978), modified. D. Geological map of the Jerma River Canyon area,
redrawn and modified from Bucur et
al. (1995). |
|
Figure 2:
Kamenica section showing distribution of Orbitolinidae and
Calpionellidae (modified from Bucur et
al., 2020). Lithologic units (A to D) in accordance with Radoičic
(1978). Note: Gen. et sp. indet 3 is absent in the Kamenica section. |
For the studied Lower Cretaceous orbitolinid-bearing sections of eastern Serbia, there is no calibration with ammonite zones, unlike southeastern France (e.g., Clavel et al., 2014). There, the biostratigraphy of the occurring orbitolinids is well constrained based on ammonoids and calpionellids due to their occurrence in resediments within platform-to-basin depositional settings. In the Serbian sections, numerous benthic foraminifera and Dasycladales have been observed that are also occurring in the Lower Cretaceous of Switzerland and southeastern France (e.g., Darsac, 1983; Blanc, 1996; Granier, 2019). The biostratigraphic setting of the studied sections has been discussed in detail by Bucur et al. (2020). Therefore, only some remarks are provided herein.
Biostratigraphic important Dasycladales
are represented for example by Salpingoporella
steinhauseri Conrad et al.
or Falsolikanella campanensis
(Azéma & Jaffrezo). The latter species has recently been
defined as an upper Berriasian marker taxon by Granier (2019). Although
it has also been observed in the lower part of the Kamenica section that might
be early Berriasian in age, F. campanensis
has never been observed in Valanginian strata. Among the benthic foraminifera
the occurrence of Danubiella gracilima
Neagu, Pfenderina neocomiensis (Pfender) (Fig.
3.5-3.8 ![]() ), Pseudotextulariella courtionensis Brönnimann is worth
mentioning because the FAD of P.
neocomiensis consistently indicates the middle part of the upper Berriasian
(base alpillensis subzone) (Darsac,
1983; Boisseau, 1987; Virgone,
1997; Granier, 2019).
According to Blanc (1996, Fig. 58), the FAD's of Pfenderina neocomiensis (Pfender) and the calpionellid Lorenziella
hungarica Knauer & Nagy (= hungarica
Zone) are contemporaneously recorded from the base of the alpillensis ammonoid zone referring to the middle part of the upper
Berriasian. Both taxa are also recorded from upper Berriasian sections studied
herein. In the Kamenica section, the first but undeterminable orbitolinids are
recorded from sample 14776 (Fig. 2
), Pseudotextulariella courtionensis Brönnimann is worth
mentioning because the FAD of P.
neocomiensis consistently indicates the middle part of the upper Berriasian
(base alpillensis subzone) (Darsac,
1983; Boisseau, 1987; Virgone,
1997; Granier, 2019).
According to Blanc (1996, Fig. 58), the FAD's of Pfenderina neocomiensis (Pfender) and the calpionellid Lorenziella
hungarica Knauer & Nagy (= hungarica
Zone) are contemporaneously recorded from the base of the alpillensis ammonoid zone referring to the middle part of the upper
Berriasian. Both taxa are also recorded from upper Berriasian sections studied
herein. In the Kamenica section, the first but undeterminable orbitolinids are
recorded from sample 14776 (Fig. 2 ![]() ). Higher in the section, the Orbitolinidae
appear contemporaneously with different taxa. P. neocomiensis appears about 20 m above the first orbitolinids and
persists throughout the uppermost part of the section. Calpionellids (incl. Lorenziella hungarica) are recorded higher up in the section. The
whole calpionellid assemblage (see Bucur et al.,
2020) corresponds to the Calpionellopsis zone, and, most probably to the oblonga subzone that
characterizes the upper Berriasian (Remane et al.,
1986; Pop, 1994; Blau & Grün,
1997; Reháková & Michalík,
1997; Lakova & Petrova,
2013; Benzaggagh, 2020). The overall succession with fenestral limestones (intertidal) to subtidal
foraminiferan-algal wackestones associated with rare calionellids in the upper
part reflects a shift from internal to external platform settings. This might
explain the time discrepancy between the offset occurrences of P.
neocomiensis and L. hungarica in the Kamenica section (Fig. 2
). Higher in the section, the Orbitolinidae
appear contemporaneously with different taxa. P. neocomiensis appears about 20 m above the first orbitolinids and
persists throughout the uppermost part of the section. Calpionellids (incl. Lorenziella hungarica) are recorded higher up in the section. The
whole calpionellid assemblage (see Bucur et al.,
2020) corresponds to the Calpionellopsis zone, and, most probably to the oblonga subzone that
characterizes the upper Berriasian (Remane et al.,
1986; Pop, 1994; Blau & Grün,
1997; Reháková & Michalík,
1997; Lakova & Petrova,
2013; Benzaggagh, 2020). The overall succession with fenestral limestones (intertidal) to subtidal
foraminiferan-algal wackestones associated with rare calionellids in the upper
part reflects a shift from internal to external platform settings. This might
explain the time discrepancy between the offset occurrences of P.
neocomiensis and L. hungarica in the Kamenica section (Fig. 2 ![]() ). The thickness of the
orbitolinid-bearing strata in the Kamenica section is about 78 m, interrupted by
two passages in the field without exposed rocks (Fig. 2
). The thickness of the
orbitolinid-bearing strata in the Kamenica section is about 78 m, interrupted by
two passages in the field without exposed rocks (Fig. 2 ![]() ). The first (not further
determined) specimens occur in sample 14776 and slightly higher up, almost all
the different taxa reported herein have been observed. There might be minor
differences in the appearances of individual species in the studied section, but
these are difficult to verify. Taking into account sample biasing,
incompleteness due to exposure gaps etc., all forms are treated as more or less
contemporaneous herein, and assigned to the late Berriasian. In previous works,
the upper orbitolinid-bearing part of the Kamenica section was attributed to the
Late Hauterivian (Polavder, 2014; with kind assistance of B.
Clavel, see
acknowledgements therein). This assignment was due to the occurrence of several
Orbitolinidae that have their "first appearance datum" (FAD) in the Late Hauterivian
times (e.g., Clavel et
al., 2014). This assignment was state-of-the-art because these taxa have so
far nowhere else been reported from older strata. The published data on the
recovered orbitolinids from eastern Serbia, however, are in contradiction to our
results, i.e., a late Berriasian age. Except those described as new or in open
nomenclature herein, all other orbitolinid species from eastern Serbia are well
known from the classical Urgonian of southeastern France (Arnaud-Vanneau,
1980; Clavel et al.,
2009a, 2009b, 2010, 2014; Granier et al.,
2013a, 2013b, 2017). Based on numerous hemipelagic sections comprising ammonites
and redeposited orbitolinids, a new detailed range chart ("nouvelle
biostratigraphie des orbitolinidés") for 37 orbitolinid species [excluding
the pfenderinid Moulladella jourdanensis
(Foury & Moullade); see Bucur & Schlagintweit,
2018; Fig. 3.1-3.4
). The first (not further
determined) specimens occur in sample 14776 and slightly higher up, almost all
the different taxa reported herein have been observed. There might be minor
differences in the appearances of individual species in the studied section, but
these are difficult to verify. Taking into account sample biasing,
incompleteness due to exposure gaps etc., all forms are treated as more or less
contemporaneous herein, and assigned to the late Berriasian. In previous works,
the upper orbitolinid-bearing part of the Kamenica section was attributed to the
Late Hauterivian (Polavder, 2014; with kind assistance of B.
Clavel, see
acknowledgements therein). This assignment was due to the occurrence of several
Orbitolinidae that have their "first appearance datum" (FAD) in the Late Hauterivian
times (e.g., Clavel et
al., 2014). This assignment was state-of-the-art because these taxa have so
far nowhere else been reported from older strata. The published data on the
recovered orbitolinids from eastern Serbia, however, are in contradiction to our
results, i.e., a late Berriasian age. Except those described as new or in open
nomenclature herein, all other orbitolinid species from eastern Serbia are well
known from the classical Urgonian of southeastern France (Arnaud-Vanneau,
1980; Clavel et al.,
2009a, 2009b, 2010, 2014; Granier et al.,
2013a, 2013b, 2017). Based on numerous hemipelagic sections comprising ammonites
and redeposited orbitolinids, a new detailed range chart ("nouvelle
biostratigraphie des orbitolinidés") for 37 orbitolinid species [excluding
the pfenderinid Moulladella jourdanensis
(Foury & Moullade); see Bucur & Schlagintweit,
2018; Fig. 3.1-3.4 ![]() ] has been provided by Clavel et al. (2014) for this area. The first species are recorded from the
lowermost Hauterivian (Sayni ammonite zone), and the latest from the lower
Aptian (top Furcata zone). Among the numerous orbitolinids, just a few are
considered as "marker species", such as Valserina
primitiva for the Upper Hauterivian (Charollais et al.,
2009, Fig.
2; Clavel et al., 2014, Fig. 5). Others like Paracoskinolina arcuata have a longer range, from the
Late Hauterivian to the early Aptian (Clavel et al., 2014, Fig. 5). For the individual ranges of the orbitolinids
(and other taxa), Granier et al. (2013a, p. 140;
2017, p. 118) highlighted the
"calibration
of the First Appearance Datum - FAD - (and eventually the Last Appearance
Datum - LAD ...)". Both, the FAD and LAD define the total range (zone)
of a given taxon (e.g., Nichols, 2009), that should ideally correspond to its origination and extinction (Holland
& Patzowsky, 2002, for details; Guex et
al., 2016, p. 109: first phyletic appearance of a species).
Lithostratigraphically, the first orbitolinids of southeastern France are
recorded from the "Urgonien jaune" (e.g.,
Clavel et al., 2014, Fig. 39),
nowadays replaced by the name Vallorbe Formation (Strasser et al.,
2016). The
Lower Hauterivian is represented by hemipelagic
marls and other lithologies unsuitable for the proliferation of orbitolinids, in
particular dictyoconinids (Strasser et al., 2016, e.g., Grand
Essert Formation). In our opinion it concludes that the appearances of the
orbitolinids in the Upper Hauterivian in southeastern France are
facies-controlled, and cannot be considered as FADs per definition as the first
(oldest) appearances in the geological record. That in turn means that the "new
biostratigraphy" from southeastern France (Clavel et al., 2014) consists of regional and not supraregional individual
ranges, and not taxon range zones. The transferation of these ranges from
southeastern France to other areas (with differing lithostratigraphic evolution)
led to the dating of the orbitolinid-bearing strata of the Kamenica section as
Late Hauterivian (Polavder, 2014). This was based on the occurrence
mainly of the "marker" species Valserina
primitiva. Referring to the numerous publications of Clavel and co-workers,
Polavder (2014, p. 12) stated that the "results are in agreement with
recent integrated studies carried out in SE France and adjacent French-Suiss
Jura". Following the general subdivision of the Lower Cretaceous strata by Radoičić
(1978), the Kamenica section comprises peritidal "fenestral and micritic
limestones" (unit B), followed by "Dasycladales-bearing limestones" (unit
C), and finally "limestones with orbitolinids and other foraminifera" (unit
D). If unit B appears as not a very favourable environment for the orbitolinids,
at least the subtidal inner platform facies of unit C can be considered as
suitable facies realm. We therefore assume that the base of unit B is very close
to the real FADs of the species described herein being aware that "the first
occurrence of a species in any given local section is likely to postdate the
time of origination of the species, and the last occurrence is likely to predate
the extinction" (Holland & Patzkowsky,
2002, p. 134). In any
case, the stratigraphic ranges of several species have to be enlarged
significantly. V. primitiva now spans
about two stages (upper Berriasian to Upper Hauterivian), but still remains one
of the stratigraphically more restricted species of the studied assemblage. For V.
primitiva, the given interval most likely corresponds to the total taxon
range. Others, like Cribellopsis
neoelongata, ranging over several stages, from the upper Berriasian to the
lower Aptian. In this case, the correspondance of the youngest record to the LAD
(see Clavel et al., 2014, Fig.
5) is uncertain since the last orbitolinid occurrences in the late lower Aptian
in southeastern France coincides with environmental changes, i.e., to platform
drowning and demise of shallow water carbonates. Summarizing, the detailed orbitolinid ranges in southeastern France (Clavel et
al., 2014) are in no case doubted. But the transferation of these data by
subsequent workers to other regions should (i) keep in mind that these are local
orbitolinid ranges and (ii) the usage of the terms FAD/LAD [First/Last
Appearance Datum] or FO/LO [First/Last Occurrence] (e.g.,
Masse & Fenerci-Masse, 2013) may be misleading. As Guex
et al. (2016, p. 109) summed up: "datum lost its original meaning" and is treated
"synonymous with first
local appearance and first local disppearance of a species" by many workers.
] has been provided by Clavel et al. (2014) for this area. The first species are recorded from the
lowermost Hauterivian (Sayni ammonite zone), and the latest from the lower
Aptian (top Furcata zone). Among the numerous orbitolinids, just a few are
considered as "marker species", such as Valserina
primitiva for the Upper Hauterivian (Charollais et al.,
2009, Fig.
2; Clavel et al., 2014, Fig. 5). Others like Paracoskinolina arcuata have a longer range, from the
Late Hauterivian to the early Aptian (Clavel et al., 2014, Fig. 5). For the individual ranges of the orbitolinids
(and other taxa), Granier et al. (2013a, p. 140;
2017, p. 118) highlighted the
"calibration
of the First Appearance Datum - FAD - (and eventually the Last Appearance
Datum - LAD ...)". Both, the FAD and LAD define the total range (zone)
of a given taxon (e.g., Nichols, 2009), that should ideally correspond to its origination and extinction (Holland
& Patzowsky, 2002, for details; Guex et
al., 2016, p. 109: first phyletic appearance of a species).
Lithostratigraphically, the first orbitolinids of southeastern France are
recorded from the "Urgonien jaune" (e.g.,
Clavel et al., 2014, Fig. 39),
nowadays replaced by the name Vallorbe Formation (Strasser et al.,
2016). The
Lower Hauterivian is represented by hemipelagic
marls and other lithologies unsuitable for the proliferation of orbitolinids, in
particular dictyoconinids (Strasser et al., 2016, e.g., Grand
Essert Formation). In our opinion it concludes that the appearances of the
orbitolinids in the Upper Hauterivian in southeastern France are
facies-controlled, and cannot be considered as FADs per definition as the first
(oldest) appearances in the geological record. That in turn means that the "new
biostratigraphy" from southeastern France (Clavel et al., 2014) consists of regional and not supraregional individual
ranges, and not taxon range zones. The transferation of these ranges from
southeastern France to other areas (with differing lithostratigraphic evolution)
led to the dating of the orbitolinid-bearing strata of the Kamenica section as
Late Hauterivian (Polavder, 2014). This was based on the occurrence
mainly of the "marker" species Valserina
primitiva. Referring to the numerous publications of Clavel and co-workers,
Polavder (2014, p. 12) stated that the "results are in agreement with
recent integrated studies carried out in SE France and adjacent French-Suiss
Jura". Following the general subdivision of the Lower Cretaceous strata by Radoičić
(1978), the Kamenica section comprises peritidal "fenestral and micritic
limestones" (unit B), followed by "Dasycladales-bearing limestones" (unit
C), and finally "limestones with orbitolinids and other foraminifera" (unit
D). If unit B appears as not a very favourable environment for the orbitolinids,
at least the subtidal inner platform facies of unit C can be considered as
suitable facies realm. We therefore assume that the base of unit B is very close
to the real FADs of the species described herein being aware that "the first
occurrence of a species in any given local section is likely to postdate the
time of origination of the species, and the last occurrence is likely to predate
the extinction" (Holland & Patzkowsky,
2002, p. 134). In any
case, the stratigraphic ranges of several species have to be enlarged
significantly. V. primitiva now spans
about two stages (upper Berriasian to Upper Hauterivian), but still remains one
of the stratigraphically more restricted species of the studied assemblage. For V.
primitiva, the given interval most likely corresponds to the total taxon
range. Others, like Cribellopsis
neoelongata, ranging over several stages, from the upper Berriasian to the
lower Aptian. In this case, the correspondance of the youngest record to the LAD
(see Clavel et al., 2014, Fig.
5) is uncertain since the last orbitolinid occurrences in the late lower Aptian
in southeastern France coincides with environmental changes, i.e., to platform
drowning and demise of shallow water carbonates. Summarizing, the detailed orbitolinid ranges in southeastern France (Clavel et
al., 2014) are in no case doubted. But the transferation of these data by
subsequent workers to other regions should (i) keep in mind that these are local
orbitolinid ranges and (ii) the usage of the terms FAD/LAD [First/Last
Appearance Datum] or FO/LO [First/Last Occurrence] (e.g.,
Masse & Fenerci-Masse, 2013) may be misleading. As Guex
et al. (2016, p. 109) summed up: "datum lost its original meaning" and is treated
"synonymous with first
local appearance and first local disppearance of a species" by many workers.
The present study is based on
thin-sections coming from three localities or sections (Fig. 1 ![]() ): Kamenica and
Prekonozi (situated north of Niš city) and Jerma River Canyon (south of Pirot
town, easternmost Serbia). More than 280 samples were collected (most of them,
198, from the Kamenica section) for thin-section preparation. In the framework
of the present study, also 114 thin-sections collected from Kamenica section
belonging to Rajka Radoičić's collection were reanalysed. All of the used thin-sections are deposited in
Rajka
Radoičić's private collection numbers RR 3412 to RR 4049 and
in Ioan I. Bucur collection (14001 to 14034; 14713 to 14739; 14748 to
14838; 16149 to 16227, and 16319 to 16370) from the Department of Geology (Babeş-Bolyai
University in Cluj-Napoca).
): Kamenica and
Prekonozi (situated north of Niš city) and Jerma River Canyon (south of Pirot
town, easternmost Serbia). More than 280 samples were collected (most of them,
198, from the Kamenica section) for thin-section preparation. In the framework
of the present study, also 114 thin-sections collected from Kamenica section
belonging to Rajka Radoičić's collection were reanalysed. All of the used thin-sections are deposited in
Rajka
Radoičić's private collection numbers RR 3412 to RR 4049 and
in Ioan I. Bucur collection (14001 to 14034; 14713 to 14739; 14748 to
14838; 16149 to 16227, and 16319 to 16370) from the Department of Geology (Babeş-Bolyai
University in Cluj-Napoca).
The upper Berriasian orbitolinid fauna shows striking similarities to the one from the Urgonian type-area in southern Switzerland and southeastern France. Hence, the numerous works (including type descriptions) of (in alphabetical order) A. Arnaud-Vanneau (Arnaud-Vanneau, 1976, 1980; Arnaud-Vanneau & Thieuloy, 1972; Arnaud-Vanneau & Schroeder, 1976), B. Clavel (Charollais et al., 2009; Clavel in Granier et al., 2013a, 2013b, 2017; Clavel et al., 2014), G. Foury (Foury & Moullade, 1966; Foury, 1968), M. Moullade (Moullade, 1960, 1966), and R. Schroeder (Schroeder & Charollais, 1966; Schroeder et al., 1969; Cherchi & Schroeder, 1978, 1999) should be highlighted for determination purposes.
The orbitolinid assemblage from the upper Berriasian of eastern Serbia comprises 17 taxa belonging to seven genera. Three taxa were described in open nomenclature. The majority of the observed genera belong to the subfamily Dictyoconinae Moullade, and a single genus of the Dictyorbitolininae Schroeder: Paracoskinolina Moullade. No Orbitolininae (with complex embryo) were observed. According to present knowledge, they appear in the Late Hauterivian [Schroeder & Cherchi, 2002; Clavel et al., 2014: Eopalorbitolina pertenuis (Foury)]. For each species, the occurrences in the studied three sections of eastern Serbia (Kamenica, Jerma, Prekonozi) are indicated with semi-quantitative abundances (very rare, rare, common, abundant). These informal statements however must bear in mind that the data base (e.g., number of thin-sections per studied section) is not balanced (see Material and Methods). For the already known species, reference is made to detailed descriptions available in the literature. Some additional remarks/observations from the Serbian material are provided along with informations on the biometric data (D = test diameter, H = test height).
The
high-rank classification follows Pawlowski et al.
(2013). For the low-rank classification, see Kaminski
(2014). For glossary, report to Hottinger
(2006). Note: orientation of structures (e.g.,
pillars) with respect to chamber (floor versus ceiling or roof) in direction of
growth (see Hottinger & Drobne,
1980, p. 205). The common
agglutinated conical species Moulladella (former
Paracoskinolina) jourdanensis (Foury & Moullade) is not treated
herein due to its recent transfer to the Pfenderinidae (see Bucur & Schlagintweit,
2018) (Fig. 3.1-3.3 ![]() ).
).
|
Figure 3:
Pfenderinids Moulladella
jourdanensis (Foury & Moullade) (1-4) and Pfenderina
neocomiensis (Pfender) (5-8)
from the upper Berriasian of the Kamenica section, Eastern Serbia. 1-2:
Subaxial sections of adult and juvenile microspheric specimens. Thin-section
14729, 14032. 3: Axial section of a
megalospheric specimen. Thin-section RR 4029. 4:
Transverse section. Thin-section 14808. 5:
Tangential section. Thin-section 14830. 6:
Transverse section. Thin-section 14803. 7:
Subaxial section. Thin-section 14805. 8:
Axial section. Thin-section 14792. |
Phylum Foraminifera Orbigny, 1826
Class Globothalamea Pawlowski et al., 2013
Order Loftusiida Kaminski & Mikhalevich, 2004
Suborder Loftusiina Kaminski & Mikhalevich, 2004
Superfamily Orbitolinoidea Martin, 1890
Family Orbitolinidae Martin, 1890
Subfamily Dictyoconinae Schubert, 1912
Genus Cribellopsis Arnaud-Vanneau, 1980
Remarks: All
species of Cribellopsis show more or
less identical subdivision of the marginal zone: no horizontal partitions, and
one to three intercalary beams between the main beams (Arnaud-Vanneau,
1980; Chiocchini, 1989). The reticulate central zone may be well
developed or reduced in different species. This means in turn that species are
defined mainly by size and/or general morphology and in cases by indefinite
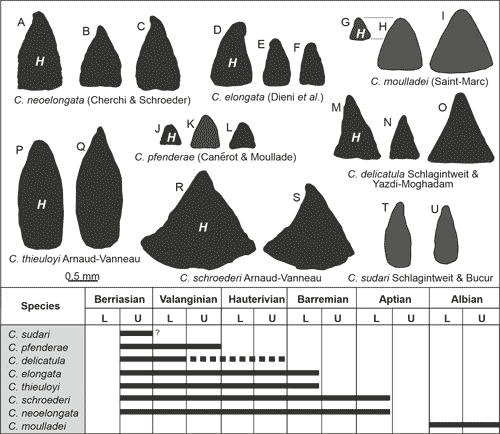
features (more ..., less ...) (Fig. 4 ![]() ).
).
|
Figure 4:
Morphological variety of different Cribellopsis
species and known stratigraphic ranges. |
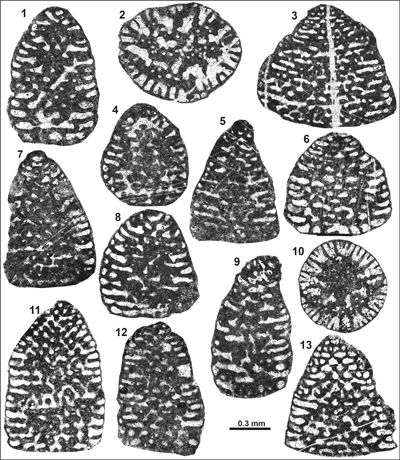
Cribellopsis elongata (Dieni et al., 1963)
(Pl. 1 ![]() , figs. 1-9)
, figs. 1-9)
1963 Orbitolinopsis elongatus n. sp. - Dieni et al., p. 4; Pl. 1, figs. 1-2, 4
1980 Cribellopsis elongata (Dieni, Massari & Moullade) nov. comb. - Arnaud-Vanneau, p. 678; Pl. 96, figs. 3-8 (see comments of Becker, 1999, below)
2009 Cribellopsis elongata - Charollais et al.; Pl. 5, fig. 21
2010 Cribellopsis elongata - Clavel et al.; Pl. 1, specimen 6892-1b; Pl. 5, specimen GC13.4a; Pl. 7, specimen 161.29-1b, 471-16a, 387.4b1, 471-1b; Pl. 9, specimens 387.9-3b, 387.9-2b, 387.14-7a; Pl. 14, specimen 407.3c3; Pl. 20, specimens Ai31-11a, Ai19-12a, Ai19-1b; Pl. 27, 218.5-5b; Pl. 29, specimen 226.22-5e
2013a Cribellopsis elongata - Clavel in Granier et al., p. 151; Pl. 5, figs. 1-5, 18, 20
2014 Cribellopsis elongata - Clavel et al.; Pl. 6, specimen MC233-6b; Pl. 8, specimen 511-12a; Pl. 10, specimen 400-7a; Pl. 12, specimen 470-5c; Pl. 14, specimen 464-6a; Pl. 16, specimen 493.3-24a; Pl. 21, specimen 6870-6a; Pl. 23, specimen 410-17a; Pl. 27, specimen LEST27-17a, Pl. 28 (topotypes!), specimens 7343-2a, 7343-1b, 7341-42a, 7341-12a; Pl. 29, specimens 7871-5b, 7871-8a; Pl. 31, specimens 6816-2a, 6816-1a, SANF1-1b, SANF1-1a, SANF10-1a
2015 Cribellopsis elongata - Bonvallet; Pl. 4.2, figs. 8-9
2017 Cribellopsis elongata - Clavel in Granier et al., p. 130; Pl. 5, figs. 1-5, 18, 20
2017 Dicytoconid foraminifera - Ungureanu et al. 2017; Fig. 7.n
non 2018 Cribellopsis elongata - Ghafor & Mohialdeen; Pl. 2, fig. c (= ?Coscinoconus sp., see Granier, 2020)
Remarks: The species has witnessed differing views on its taxonomy and validity that is shortly summarized herein. Dieni et al. (1963) described two new species as Orbitolinopsis elongatus and O. subkiliani from the Barremian of the region of Orosei, Sardinia. All specimens illustrated are axial or subaxial sections; no transverse sections have been provided. From the four specimens of O. elongatus, only the holotype is from the type-locality, while the three paratypes are from southeastern France (Drôme and La Bédoule areas). It is worth mentioning here, that from the lower Barremian of Cassis-La Bédoule, Clavel et al. (2014, Fig. 28) illustrated four topotype specimens (Pl. 28, therein, see synonymy). From the Orosei area (incl. the type-locality of O. subkiliani), Cherchi and Schroeder (1978) were studying the orbitolinid fauna, including the two forms described by Dieni et al. (1963). The authors state (p. 161) that O. elongatus and O. subkiliani should only differ from their dimensions (diameter, height, ratio d/h) and the number of chambers in the rectilinear stage. Sumarizing, Cherchi and Schroeder (1978) concluded that O. elongatus represents a synonym of O. subkiliani. Arnaud-Vanneau (1980, p. 678) transferred O. elongatus to the genus Cribellopsis including the holotype, and two of the three paratypes of Dieni et al. (1963). Arnaud-Vanneau (1980) studying the type-material of Dieni et al. (1963) concluded that the internal structure of O. elongata and O. subkiliani is "sensiblement différente" thereby rejecting the synonymization proposed by Cherchi and Schroeder (1978). Whether O. subkiliani is a valid species has not been further considered by Arnaud-Vanneau. In any case it is lacking in the comparative table of Orbitolinopsis species occurring in the Vercors and Chartreuse areas of southeastern France (Arnaud-Vanneau, 1980, Fig. 226). Becker (1999) followed the view of Cherchi & Schroeder (1978) treating O. elongatus a synonym of O. subkiliani. She also noted that the specimens illustrated by Arnaud-Vanneau (1980) as Cribellopsis elongatus are not identical to the species described by Dieni et al. (1963) because of the presence of intercalary beams. The latter are not visible in the specimens from Sardinia (Becker, 1999, p. 412), but this is, in our opinion, just due to the axial/subaxial section planes. Clavel et al. (2010, p. 6) finally observed the structural identity ("structure absolument identique") of O. subkiliani and O. kiliani. Based on observations on a large number of specimens from southeastern France, Clavel et al. (2010) rejected the synonymization of O. elongata with O. subkiliani, as proposed by Cherchi & Schroeder (1978) and Becker (1999). Instead, these show generic differences (here: Cribellopsis vs. Orbitolinopsis, see Arnaud-Vanneau, 1980), a view that is followed here.
Dimensions: D = 0.35-0.55 mm; H = 0.5-0.85 mm; D/H 0.59-0.72
Occurrences: Rare in the Kamenica and Jerma sections.
Cribellopsis neoelongata (Cherchi & Schroeder, 1978)
(Pl. 1 ![]() , figs. 10-12;
Pl. 2
, figs. 10-12;
Pl. 2 ![]() , figs. 1-12)
, figs. 1-12)
1966 Orbitolinopsis sp. (Gruppe1) - Schroeder & Charollais, p. 103; Pl. 1, figs. 1-10
1978 Orbitolinopsis (?) neoelongata n. sp. - Cherchi & Schroeder, p. 162; Pl. 2, figs. 5-10
1980 Cribellopsis neoelongata (Cherchi & Schroeder) nov. comb. - Arnaud-Vanneau, p. 669; Pl. 59, figs. 4-7; Pl. 95, figs. 7-14, Fig. 228
2000 Cribellopsis neoelongata (Cherchi & Schroeder) - Bernaus, p. 64; Pl. 5, figs. 15-18
non 2000 Cribellopsis neoelongata (Cherchi & Schroeder) - Hughes; Pl. 6, figs. 4 (= Praechrysalidina infracretacea) - 5 (= Voloshinoides murgensis)
2002 Cribellopsis neoelongata (Cherchi & Schroeder) - Bernaus et al.; Fig. 5.G
2009a Cribellopsis neoelongata - Clavel et al.; Pl. 1, specimen DL101a; Pl. 2, specimens 6922-19a, 6922-22a, 6921-8a, 6923-23a, 6922, b14, 6923-1c
2010 Cribellopsis neoelongata (Cherchi & Schroeder) - Clavel et al.; Pl. 14, specimen 407.3-1c; Pl. 20, specimen Ai31-15b; Pl. 27, specimen 218.5-2a; Pl. 29, specimen 226.22-3b
2013a Cribellopsis neoelongata (Cherchi & Schroeder) - Clavel in Granier et al.; p. 151, Pl. 5, figs. 9-14; Pl. 9, fig. 9; Pl. 10, fig. 10
2014 Cribellopsis neoelongata (Cherchi & Schroeder) - Clavel et al.; Pl. 9, specimen GF15-1c; Pl. 11, specimen 7833-9a; Pl. 13, specimen 470-8b; Pl. 14, specimen 464-44b; Pl. 17, specimen 575-1a; Pl. 22, specimen 407.3-1c; Pl. 23, specimen 409b43; Pl. 25, specimen 229.2-1b3; Pl. 26, specimen LEST54-31a; Pl. 28, specimen 7341-41a; Pl. 30, specimen 6943-10a; Pl. 31, specimen SANF50-1a2015
2014 Cribellopsis neoelongata (Cherchi & Schroeder) - Polavder; Fig. 5a-l
2015 Cribellopsis neoelongata - Bonvallet; Pl. 4.3, figs. 8-10
2017 Cribellopsis neoelongata (Cherchi & Schroeder) - Clavel in Granier et al.; p. 130, Pl. 5, figs. 9-14; Pl. 9, fig. 9; Pl. 10, fig. 10
2019 Cribellopsis neoelongata (Cherchi & Schroeder) - Bonvallet et al.; Figs. 10.27, 10.38-10.39
Remarks: A detailed description of the species has been provided by Schroeder in Schroeder & Charollais (1966, p. 103-105) in open nomenclature as Orbitolinopsis sp. (Gruppe 1). Systematically, it has been described as Orbitolinopsis (?) neoelongata by Cherchi & Schroeder (1978), later revised by Arnaud-Vanneau (1980). For instance, typical features are the often observed slight inclination of the rather acute apical part, a flat to slightly convex base, and the overall cylindro-conical morphology, resulting from the decreasing chamber diameter of the youngest chambers in adult specimens (Cherchi & Schroeder, 1978, p. 162, "prima parte ... é conica, ... stadio tardoontogenetico č cilindrico"). Adult specimens consist of 20 to 30 uniserial chambers. C. neoelongata is one of the most common taxa in the Kamenica section.
Dimensions: D = 0.70-0.95 mm; H = 0.80-1.35 mm; D/H 0.55-1.1
Occurrences: Common in the Kamenica and rare in the Jerma sections.
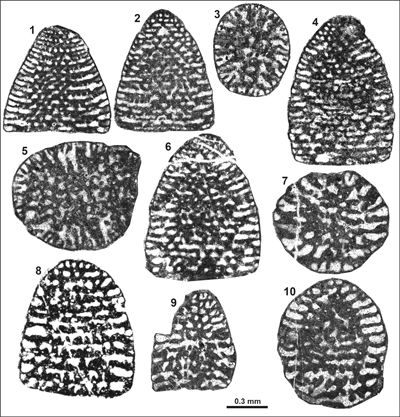
Cribellopsis pfenderae (Canérot & Moullade, 1971) nov. comb.
(Pl. 3 ![]() ,
figs. 1-11)
,
figs. 1-11)
1971 Paracoskinolina pfenderae n. sp. - Canérot & Moullade, p. 216; Pl. 2, figs. 8-20
1976 Paracoskinolina pfenderae Canérot & Moullade - Peybernčs; Pl. 14, figs. 17-20
1984 Paracoskinolina pfenderae Canérot & Moullade - Canérot; Pl. 1, figs. 14-16
1995 Paracoskinolina pfenderae Canérot & Moullade - Bucur et al.; Pl. VII, figs. 10-11
2004 Paracoskinolina pfenderae Canérot & Moullade - Ivanova & Koleva-Rekalova; Pl. 4, figs. 8-12
2018 Paracoskinolina? pfenderae Canérot & Moullade - Bucur & Schlagintweit; Figs. 7e, h
2019 Paracoskinolina pfenderae Canérot & Moullade - Granier & Clavel; Figs. 5M-O
Remarks:
The central zone of the small-sized "Paracoskinolina"
pfenderae was originally described as
possessing triangular pillars that are tapering towards the test base ("pointe
en base"). The single slightly oblique transverse section does in our opinion
not show pillars but radial main partitions reaching into the central part of
the test where they anastomose (Canérot & Moullade, 1971: Pl.
2, fig. 12). They alternate between subsequent chambers (e.g., Canérot & Moullade, 1971:
Pl. 2,
figs. 10,
13, 19). The exoskeleton lacks horizontal partitions and exhibits one to three
intercalary beams between two main partitions (Pl. 3 ![]() , figs. 4, 7). Tests are
medium-conical with up to 12 chambers in adult specimens. As revised by Arnaud-Vanneau
(1980), the genus Paracoskinolina Moullade
possesses pillars in the central zone that are aligned between subsequent
chambers. Due to the inner test structure as defined above, the species is here
transferred to the genus Cribellopsis Arnaud-Vanneau
becoming its smallest representative (see Fig. 4
, figs. 4, 7). Tests are
medium-conical with up to 12 chambers in adult specimens. As revised by Arnaud-Vanneau
(1980), the genus Paracoskinolina Moullade
possesses pillars in the central zone that are aligned between subsequent
chambers. Due to the inner test structure as defined above, the species is here
transferred to the genus Cribellopsis Arnaud-Vanneau
becoming its smallest representative (see Fig. 4 ![]() ). All reported occurrences are
from the former Northern Neotethysian margin stretching from Spain to the west
to Bulgaria-Serbia to the east (see synonymy).
). All reported occurrences are
from the former Northern Neotethysian margin stretching from Spain to the west
to Bulgaria-Serbia to the east (see synonymy).
Dimensions: D = 0.30-0.45 mm; H = 0.30-0.50 mm; D/H = 0.9-1.1.
Occurrences: Common in the Kamenica and rare in the Jerma sections.
Cribellopsis thieuloyi Arnaud-Vanneau, 1980
(Pl. 3 ![]() , figs. 12-17)
, figs. 12-17)
1980 Cribellopsi thieuloyi n. sp. - Arnaud-Vanneau, p. 675; Pl. 96, figs. 13-19
2010 Cribellopsis thieuloyi Arnaud-Vanneau - Clavel et al.; Pl. 14, specimen DL105a; Pl. 20, specimen Ai31-8a; Pl. 24, specimen 6923-3a; Pl. 30, specimen 275.4-1a
2013a Cribellopsis thieuloyi Arnaud-Vanneau - Clavel in Granier et al., p. 151; Fig. 9A, Pl. 5, figs. 6-8, 17
2014 Cribellopsis thieuloyi Arnaud-Vanneau - Clavel et al.; Pl. 8, specimen 7847-12a; Pl. 10, specimen 597-13a; Pl. 14, specimen 398-18b; Pl. 16, specimen 500.9d12; Pl. 19, specimen 263-28a; Pl. 21, specimen DL105a; Pl. 23, specimen 410-15b; Pl. 27, specimen LEST88.2-1b; Pl. 28, specimen 7341-19a, 7341-11a; Pl. 29, specimen 7339-15b
2017 Cribellopsis thieuloyi Arnaud-Vanneau - Clavel in Granier et al., p. 131; Fig. 9A, Pl. 5, figs. 6-8, 17
Remarks: C. thieuloyi represents one of the largest species of the genus known so far only from southeastern France. It is of cylindro-conical morphology with acutely tapering apex; adult specimens consist of up 40 chambers. C. thieuloyi is comparably rare in the upper Berriasian orbitolinid assemblage of eastern Serbia.
Dimensions: D = 0.7-1.0 mm; H = 1.7-2.15 mm; D/H: 0.39-0.46; number of chambers last 0.5 mm: 5-8.
Occurrences: Common in the Kamenica and rare in the Jerma sections.
Cribellopsis sudari Schlagintweit & Bucur n. sp.
(Fig. 5 ![]() ,
Pl. 4
,
Pl. 4 ![]() ,
figs. 1-13; Pl. 5
,
figs. 1-13; Pl. 5 ![]() ,
figs. 1-15)
,
figs. 1-15)
Etymology: The species is dedicated to Milan Sudar for his contribution to the study of Mesozoic deposits from Serbia.
Holotype: Specimen illustrated in
Pl. 4 ![]() , fig. 5,
thin-section 16209-2.
, fig. 5,
thin-section 16209-2.
Paratypes:
Pl. 4 ![]() ,
figs. 1 (14791-4) - 2 (14020), 10 (14796); Pl. 5
,
figs. 1 (14791-4) - 2 (14020), 10 (14796); Pl. 5 ![]() ,
figs. 1 (14792-19) - 2
(14019).
,
figs. 1 (14792-19) - 2
(14019).
Type horizon: Upper Berriasian carbonates, not yet assigned lithostratigraphically to any formation.
Type
locality: Kamenica section located north of Kamenica (Fig.
1.A, .C ![]() ). The locality can be found on the geological map of Yugoslavia 1:100.000
(sheet K34-20, Aleksinac) (Kristic et al., 1978). GPS coordinates:
43°13'48.2"N
21°33'38.5"E.
). The locality can be found on the geological map of Yugoslavia 1:100.000
(sheet K34-20, Aleksinac) (Kristic et al., 1978). GPS coordinates:
43°13'48.2"N
21°33'38.5"E.
Description: High-conical to cylindroconical test displaying an acute tapering apex where the subspherical, simple embryo (diameter 0.06-0.13 mm) is positioned. It is followed by a high trochospirally coiled part of a few whorls, while the main part consists of up to 25 uniserial, slightly convexe (in growth direction) chambers. The marginal zone is subdivided by beams and two to three intercalary beams; no rafters are present. The thickness of the intercalary beams equals the one of the marginal chamber wall. The beams are widening inwards where they form a coarse reticulate network. This central part (or zone) amounts roughly one third of the total diameter. In shallow tangential sections, a rectangular pattern, higher than wide, is present. Inwards, the partitions are widening at the base and top producing a rounded-triangular to trapezoidal pattern with foramina arranged cross-wise obliquely in diagonal lines. The wall is finely agglutinated. Test dimorphism has not been observed.
Dimensions: D = 0.39-0.51; H = 0.99-1.25 mm; D/H = 0.39-0.44; numbers of chambers last 0.5 mm: 8-12.
Comparisons: Shallow tangential sections (from
apex towards base) showing the rectangular pattern of compartments (Pl. 4 ![]() , fig.
10) can be confused with Paracoskinolina
maynci (Chevalier). In the latter species the vertical elements (beams,
pillars) are arranged in continuity; in C. sudari they are alternating (Pl. 4
, fig.
10) can be confused with Paracoskinolina
maynci (Chevalier). In the latter species the vertical elements (beams,
pillars) are arranged in continuity; in C. sudari they are alternating (Pl. 4 ![]() , fig. 10, middle part), and
only in some chambers (e.g., lower and
upper part of the same specimen) they appear as being aligned. The alternating
arrangement becomes evident when the section plane passes from the random to the
central parts in these specimens (Pl. 4
, fig. 10, middle part), and
only in some chambers (e.g., lower and
upper part of the same specimen) they appear as being aligned. The alternating
arrangement becomes evident when the section plane passes from the random to the
central parts in these specimens (Pl. 4 ![]() , fig. 1). The general test morphology
compares to some extent with C. neoelongata,
often showing a bell-shaped morphology. C.
sudari has a more slender cylindro-conical test expressed by a lower D/H
ratio and a high trochospirally coiled initial part. The upper test of C.
sudari rarely and only slightly bends. C.
thieuloyi with its bullet-like test morphology is distinctly larger (D, H),
also having chambers of greater height.
, fig. 1). The general test morphology
compares to some extent with C. neoelongata,
often showing a bell-shaped morphology. C.
sudari has a more slender cylindro-conical test expressed by a lower D/H
ratio and a high trochospirally coiled initial part. The upper test of C.
sudari rarely and only slightly bends. C.
thieuloyi with its bullet-like test morphology is distinctly larger (D, H),
also having chambers of greater height.
Occurrences: Rare in the Jerma and Prekonozi sections.
|
Figure 5:
Evolution of Cretaceous Orbitolinidae, with special reference to the
Dictyoconinae. Above: Concept of "gradual morphological changes" (redrawn after BouDagher-Fadel & Price,
2019, Fig. 4). Note: Urgonina is not a
member of the Orbitolininae but the Dictyoconinae (e.g., Loeblich & Tappan,
1987). Below
(this work): Sudden burst evolution due to adaptive radiation in the upper
Berriasian. Note that Valdanchella has
been recorded from the upper Berriasian of southern Spain (Bádenas et al.,
2004, Fig. 2) and southwestern Switzerland (Granier et
al., 2014, Fig. 2). |
Genus Montseciella Cherchi & Schroeder, 1999
Remarks: In contrast to the classification of Kaminski (2014), the genus Montseciella Cherchi & Schroeder with its simple embryo is removed from the Orbitolininae and considered a representative of the Dictyoconinae here.
Montseciella glanensis (Foury, 1968)
(Pl. 6 ![]() )
)
1968 Paleodictyoconus glanensis n. sp. - Foury, p. 146; Pl. 18, figs. 1-12; Pl. 2, fig. 19 (fide Schroeder & Cherchi, 1982)
1969 Urgonina cf. alpillensis (Foury) - Schroeder et al., p. 93; Pl. 2, figs. 1-6; Pl. 3, fig. 6
1982 "Paleodictyoconus" glanensis Foury - Schroeder et al.; Pl. 4, figs. 1, 4, 6
1999 Montseciella glanensis (Foury) n. gen., n. comb. - Cherchi & Schroeder, p. 9; Pl. 1, figs. 1-6; Pl. 2, figs. 1-7
2010 Montseciella glanensis (Foury) - Charollais et al.; Pl. 4, figs. 1-7; Pl. 2, figs. 1-5, 22; Pl. 6, figs. 1-4
2010 Montseciella glanensis (Foury) - Clavel et al.; Pl. 5, specimens PA16.17-2a and CA1-6a; Pl. 8, specimens 471-2a, 387.12-2b, 471-13a, 387.12-1a, 387.12-2a, 161.29-1e, 387.11-6a and 387.11-8a; Pl. 13, specimen PL19.0-4d; Pl. 15, specimens DL80a, DL78a, DL88a, DL84b and DL88a; Pl. 19, specimen Ai19-8b; Pl. 21, specimens Ai31-16a and Ai31-3b; Pl. 25, specimens 6922-13a, 6923-4a and 6923-13a; Pl. 27, specimen 218.5-4a; Pl. 29, specimen 226.22-4a and 226.22-11a; Pl. 30, specimen 275.4-5a, 275.3-7a, 275.3-6b and 275.5-2b
2013a Montseciella glanensis (Foury) - Clavel in Granier et al., p. 152; Pl. 6, figs. 13, 16-17, 19-21; Pl. 8, fig. 13; Pl. 9, fig. 5
2014 Montseciella glanensis (Foury) - Clavel et al., 2014; Pl. 4, specimen 8504-9b; Pl. 7, specimens RH 352-51 and MC233-5b; Pl. 8, specimen GF15-3a; Pl. 12, specimen 471-2a; Pl. 15, specimen 7864-9a; Pl. 16, specimen 500.2-2b; Pl. 19, specimen 7402-1a; Pl. 21, specimen DL60a; Pl. 23, specimen 375-4a; Pl. 27, specimen LEST84.5-4b; Pl. 28, specimen 7341-44b and 7341-c6; Pl. 29, specimen 6933-a2; Pl. 31, specimen 6812-4c, 6816a5 and SANF10-2b
2014 Montseciella glanensis (Foury) - Polavder; Fig. 9.A
2017 Montseciella glanensis (Foury) - Clavel in Granier et al., p. 132; Pl. 6, figs. 13, 16-17, 19-21; Pl. 8, fig. 13; Pl. 9, fig. 5
Remarks: It was initially described as Paleodictyoconus glanensis by Foury (1968) from the Barremian of southeastern France noting structural differences to P. cuvillieri that "might finally justify the creation of a generic or subgeneric taxonomic unit" (Foury, 1968, p. 148, translated). This taxon represents the type-species of the genus Montseciella Cherchi & Schroeder, 1999. Cruz-Abad (2018) noted structural differences of the specimens from Sardinia that were included in the description and diagnosis of Montseciella by Cherchi and Schroeder (1999). In fact, we note that none of the specimens of Paleodictyoconus glanensis as described by Foury (1968) shows a very prominent initial coil that is defined as one typical feature of the genus Montseciella Cherchi & Schroeder. "Paleodictyoconus" glanensis should possess a slightly eccentric embryo followed by only a small spire, accepted by Cruz-Abad (2018, p. 86). Cherchi and Schroeder (1999, p. 10) on the other hand interpreted that this embryo "laterally of the apical region" with a "thick semicircular sparitic wall" absolutely unusual for Orbitolinidae…" does not belong to the figured specimen"… and "could be a transversal section of an encrusting organism, perhaps an annelid tube". This view is accepted herein. Any further taxonomic discussion based solely on our material from eastern Serbia however is beyond the scope of the present paper. Contrasting M. alguerensis, M. glanensis only occasionally has "a vertical plate" (= intercalary beam) in the marginal zone (Cherchi & Schroeder, 1999). Cherchi and Schroeder (1999, p. 10) noted that "horizontal plates are lacking in our specimens", but there seems to be rare individual rafters in adult chambers in the type-material (Foury, 1968, Pl. 18, fig. 9, penultimate chamber, left side; re-illustrated herein in Pl. 6, fig. 2, arrow). In the upper Berriasian material, M. glanensis has preferentially been observed in grainstones.
Dimensions: D up to 1.2 mm, H up to 1.1 mm; due to the majority of incomplete and/or oblique sections, dimensional data are not provided.
Occurrences: Common in the Kamenica and Prekonozi sections.
Montseciella alguerensis Cherchi & Schroeder, 1999
(Pl. 7 ![]() )
)
1999 Montseciella alguerensis n. sp. - Cherchi & Schroeder, p. 10; Pl. 3, figs. 1-3; Pl. 4, figs. 1-5
1999 Paleodictyoconus? leridanus n. sp. - Becker, p. 409; Pl. 14, figs. 1-6; Pl. 15, figs. 1-2, 4
2014 Montseciella alguerensis Cherchi & Schroeder - Clavel et al.; Pl. 8, specimen OV288a; Pl. 10, specimen 524-4a; Pl. 15, specimen 7864-4b; Pl. 16, specimen 500.9m85; Pl. 19, specimen 275.5-2b; Pl. 21, DL67b
Remarks:
The species was described separately as Paleodicytoconus?
leridanus and Montseciella alguerensis in the same year (Becker,
1999; Cherchi
& Schroeder, 1999). In this case, the ICZN (1999, article 24.2.2)
regulates that the "first reviewer" stating the synonymy may fix the name in
a published work (see Cherchi & Schroeder in Schroeder et
al., 2000, p. 22). According to Cherchi & Schroeder
(1999,
p. 11), it is "a species of Montseciella characterized by the regular occurrence of horizontal plates within the
marginal zone". Medium,
rarely high-conical tests with up to 25 chambers in adult specimens (Pl.
7 ![]() , fig.
1). There is one short rafter, and one to three intercalary beams in the
marginal zone. The central zone consists of vermicular partitions that form a
labyrinthic structure (Pl. 7
, fig.
1). There is one short rafter, and one to three intercalary beams in the
marginal zone. The central zone consists of vermicular partitions that form a
labyrinthic structure (Pl. 7 ![]() , figs. 6, 8). The structure of the central zone
("structural elements ... forming an imperfect network", Becker,
1999, p. 409 ... unvollkommenes Geflecht; Foury, 1968:
"pseudoréticulaire")
was the reason for Becker (1999) to assign the specimens only with
reservation to Paleodictyoconus Moullade.
The pores in the central area show a predominantly vertical arrangement. A
chronological order (1, M. glanensis; 2, M. alguerensis) at the beginning of
the "Montseciella - Rectodictoyconus
lineage" (Cherchi & Schroeder,
1999; Schroeder et al.,
2002) cannot be verified in the upper Berriasian of eastern
Serbia. Also the belonging of Dictyoconus?
arabicus Henson, 1948, to the
genus Montseciella is highly debatable;
again this topic is beyond the scope of the present paper.
, figs. 6, 8). The structure of the central zone
("structural elements ... forming an imperfect network", Becker,
1999, p. 409 ... unvollkommenes Geflecht; Foury, 1968:
"pseudoréticulaire")
was the reason for Becker (1999) to assign the specimens only with
reservation to Paleodictyoconus Moullade.
The pores in the central area show a predominantly vertical arrangement. A
chronological order (1, M. glanensis; 2, M. alguerensis) at the beginning of
the "Montseciella - Rectodictoyconus
lineage" (Cherchi & Schroeder,
1999; Schroeder et al.,
2002) cannot be verified in the upper Berriasian of eastern
Serbia. Also the belonging of Dictyoconus?
arabicus Henson, 1948, to the
genus Montseciella is highly debatable;
again this topic is beyond the scope of the present paper.
Dimensions: D = up to 1.3 mm, H = up to 1.5 mm. Like with Orbitolinopsis buccifer, Vanneauina vercori, Montseciella glanensis, and other taxa reported herein also the dimensions of the upper Berriasian specimens are reduced in comparisons to the respective younger type-material.
Occurrences: Rare to common in the Kamenica section.
Genus Orbitolinopsis Henson, 1948
Orbitolinopsis cf. briacensis Arnaud-Vanneau, 1980
(Pl. 8 ![]() ,
figs. 1-9)
,
figs. 1-9)
1978 Orbitolinopsis gr. flandrini Dieni, Massari & Moullade - García-Hernández; Pl. 25, figs. 13-14
1980 Orbitolinopsis briacensis n. sp. - Arnaud-Vanneau, p. 644; Pl. 10, figs. 8-10; Pl. 94, figs. 1-8, Fig. 225
2013a Orbitolinopsis briacensis Arnaud-Vanneau - Clavel in Granier et al., p. 152; Pl. 2, figs. 17, 20; Pl. 9, fig. 4 (cf.); Pl. 10, figs. 1-3
2014 Orbitolinopsis briacensis Arnaud-Vanneau - Clavel et al.; Pl. 13, specimen 470-3a; Pl. 14, specimen 464-14a; Pl. 15, specimen 7864-19a; Pl. 18, specimens 500.9-19a and 500.9-44b; Pl. 20, specimen 336-2c; Pl. 24, specimen 410-14a; Pl. 26, specimen LEST84.5-a19; Pl. 30, specimens 7339-1a and 6937-2b; Pl. 32, specimens SANF48-1b and SANF46-1a
2015 Orbitolinopsis briacensis Arnaud-Vanneau - Bonvallet; Pl. 4.6, figs. 5, 6?, 8
2017 Orbitolinopsis briacensis Arnaud-Vanneau - Clavel in Granier et al., p. 132; Pl. 2, figs. 17, 20; Pl. 9, fig. 4 (cf.); Pl. 10, figs. 1-3
2019 Orbitolinopsis briacensis Arnaud-Vanneau - Bonvallet et al.; Fig. 10/57, 10/60?, 10/61
non 2018 Orbitolinopsis briacensis - Ghafor & Mohialdeen; Pl. 1a, any litho- or bioclast?
Remarks: Medium-sized species of Orbitolinopsis, superficially similar to O. kiliani with intercalary beams in the marginal zone (Arnaud-Vanneau, 1980, Fig. 226; Clavel in Granier et al., 2013a, 2017). The test structure and many parameters of the Serbian specimens agree with the type-material of southeastern France except that the chamber height is distinctly reduced and the maximum value of the test diameter (see below); there are 14-15 chambers in the last 0.5 mm of the test in the former compared to 8-9 (Arnaud-Vanneau, 1980). A separation of a new species only based on this feature would not make sense in our opinion. It might be just a phylogenetic increase in the chamber height from the late Berriasian to the Late Hauterivian-Barremian.
Dimensions: D = 0.45-0.62 mm (Arnaud-Vanneau, 1980: up to 1.0 mm); H = 0.5-0.95 mm (Arnaud-Vanneau , 1980: up to 1.0 mm); D/H: 0.65-0.9; number of chambers last 0.5 mm: 14-15.
Occurrences: Common to frequent in the Kamenica section, common in the Prekonozi section, rare in the Jerma section.
Orbitolinopsis cf. cuvillieri Moullade, 1960
(Pl. 8 ![]() , figs. 10-17)
, figs. 10-17)
1960 Orbitolinopsis cuvillieri n. sp. - Moullade, p. 190; Pl. 2, figs. 1-7
1980 Orbitolinopsis cuvillieri Moullade - Arnaud-Vanneau, p. 634; Pl. 60, figs. 1-3; Pl. 93, figs. 1-13, fig. 223
2010 Orbitolinopsis cuvillieri Moullade - Clavel et al.; Pl. 15, specimens DL 81a and DL86a; Pl. 25, specimen 6922-25a; Pl. 29, specimen 226.22-9a
2013a Orbitolinopsis cuvillieri Moullade - Clavel in Granier et al., p. 152; Pl. 2, figs. 1-5, 8-10, 12, 16, 21; Pl. 9, figs. 16-19
2014 Orbitolinopsis cuvillieri Moullade - Clavel et al.; Pl. 9, specimen MM319az; Pl. 11, specimen 400-10c; Pl. 13, specimen 470-2a; Pl. 14, specimen 398-8a; Pl. 15, specimen 462-8a; Pl. 18, specimens 500.9-13a and 500.9-22c; Pl. 20, specimens 348-2b? And 348-1a; Pl. 22, specimens DL86a and 407.2-19b; Pl. 24, specimens 409-2a, 375-36a and 410-19b; Pl. 25, specimen 229.1-2a Pl. 26, specimen LEST27-2a and LEST26.6-7a; Pl. 30, specimens 6935-12ab, 7339-4a and 7339a3; Pl. 32, specimens SANF4-1a (cf.), SANF46-1b and SANF48-1c
2017 Orbitolinopsis cuvillieri Moullade - Clavel in Granier et al., p. 132; Pl. 2, figs. 1-5, 8-10, 12, 16, 21; Pl. 9, figs. 16-19
Remarks: There are two very close forms:
the type-species Orbitolinopsis kiliani
Silvestri, 1932, and O. cuvillieri
Moullade, 1960. In the literature there are differing views on the
validity of both. Schroeder et al.
(1969, p. 97) considered O. cuvillieri
as a junior synonym of O. kiliani
since "structural differences between both species are not recognizable".
And even Moullade (1966), who was describing the former species, reported
other specimens later as Orbitolinopsis
gr. kiliani-cuvillieri. Arnaud-Vanneau
(1980, p. 636) remarked that both are "quite close" but treated them as
separate species. According to Arnaud-Vanneau (1980), O.
cuvillieri is smaller, possesses a reduced central part, and the cupules,
being more spaced-away, have a more regular shape. Arnaud-Vanneau (1980)
assumed that O. kiliani was deriving from O. cuvillieri. According to the studies of Clavel et
al. (2014, Fig. 5), both have their first local appearances in southeastern
France at the base of the Barremian. The upper Berriasian specimens from Serbia
are directly comparable to those from the Barremian of France, namely the two (subaxial)
sections of O. cuvillieri by Moullade
(1960, Pl. 2, figs. 2-3). Unlike O.
briacensis, O. cuvillieri exhibits
a less complex marginal zone lacking intercalary beams (Pl. 8 ![]() , figs.
13-15).
, figs.
13-15).
Dimensions: D = 0.6-0.8 mm, H = 0.48-0.9 mm, D/H = 0.9-1.3
Occurrences: Common in the Kamenica, and rare in the Prekonozi sections.
Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy, 1971
(Pl. 9 ![]() )
)
1970 Orbitolinopsis n. sp. - Fourcade; Pl. 34, figs. 3-4
1972 Orbitolinopsis buccifer n. sp. - Arnaud-Vanneau & Thieuloy; Pl. 1, figs. 1-9; Pl. 2, figs. 1-11
1974 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Canérot; Pl. 16, Figs. 1-2
1978 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - García-Hernández; Pl. 25, figs. 15-16
1979 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Azéma et al.; Pl. 4, fig. 7
1980 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Arnaud-Vanneau, p. 636; Pl. 60, fig. 6; Pl. 92, figs. 9-12
1984 Paleodictyoconus? n. sp. - Canérot; Pl. 1, fig. 20
1988 Orbitolinopsis buccifer - Bucur & Cociuba; Pl. 2, figs. 12-13
2001 Orbitolinopsis buccifer - Castro et al.; Fig. 5.G
2008 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Sudar et al.; Fig. 9.9-9.11
2012 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Léonide et al.; Fig. 7.C
2013a Orbitolinopsis buccifer - Clavel in Granier et al., p. 152; Pl. 2, figs. 11, 13-14, 18-19; Pl. 9, figs. 11-15; Pl. 11, fig. 7
2014 Orbitolinopsis buccifer - Clavel et al.; Pl. 9, specimen OV177b; Pl. 11, specimen 7862-1a; Pl. 13, specimen 470-9a; Pl. 14, specimen 464-31a; Pl. 18, specimen 500.9-18b; Pl. 22, specimens DL77a, 257.4-37a; Pl. 24, specimens 6874-2a, 410a18; Pl. 25, specimen 229.2-3b; Pl. 26, specimen LEST54-2a; Pl. 30, specimen 7338-d2; Pl. 32, specimens 6816-2c, SANF46-1a, SANF43-1b
2015 Orbitolinopsis buccifer - Bonvallet; Pl. 4.6, figs. 7, 11
2017 Orbitolinopsis buccifer - Clavel in Granier et al., p. 132; Pl. 2, figs. 11, 13-14, 18-19; Pl. 9, figs. 11-15; Pl. 11, fig. 7
non 2018 Orbitolinopsis buccifere - Ghafor & Mohialdeen; Pl. 1, fig. B (= any calcitic bioclast?); Pl. 2, fig. E (= superficial ferrugineous ooid, see Granier, 2020)
2019 Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy - Bonvallet et al., Figs. 10.55-56
Remarks: Among the different Orbitolinopsis species, O. buccifer is outstanding due to its morphology and size. The test is generally rather broad, low- to medium-conical. The adult chambers may be sigmoseptal to annular with margins bending upwards (= reflexed convexo-concave type of Henson, 1948: Fig. 12u-v). Like with Orbitolinopsis briacensis, the upper Berriasian specimens of O. buccifer do not attain the large sizes as the younger type-material (Upper Hauterivian-lower Aptian) from southeastern France, especially the microspheric generation.
Dimensions: D = up to 2.1 mm; H = up to 1.3 mm.
Occurrences: Rare in the Kamenica and Prekonozi sections.
Orbitolinopsis debelmasi Moullade & Thieuloy, 1965
(Pl. 10 ![]() )
)
1960 Orbitolinopsis kiliani - Moullade, p. 190; Pl. 3, Figs. 6-12
1965 Orbitolinopsis debelmasi n. sp. - Moullade & Thieuloy, p. 4178
1974 Orbitolinopsis debelmasi Moullade & Thieuloy - Canérot; Pl. 16, fig. 3
1980 Orbitolinopsis debelmasi Moullade & Thieuloy - Arnaud-Vanneau, p. 642; Pl. 60, fig. 7; Pl. 94, figs. 9-16, Fig. 225
2006 Orbitolinopsis kiliani? - Albrich et al.; Pl. 6, fig. 14
2010 Orbitolinopsis debelmasi Moullade & Thieuloy - Clavel et al.; Pl. 15, specimen DL139a; Pl. 24, specimen 6923-9a; Pl. 27, specimen 218.5-5b
2013a Orbitolinopsis debelmasi Moullade & Thieuloy - Clavel in Granier et al.; p. 152 Pl. 7, figs. 1-5
2014 Orbitolinopsis debelmasi Moullade & Thieuloy - Clavel et al.; Pl. 4, specimen 8509-1b; Pl. 6, specimen EB H22.4; Pl. 8, specimen OV275; Pl. 10, specimen 524-5a; Pl. 13, specimen 458-7a; Pl. 16, specimen 500.6-1b; Pl. 19, specimen 7401-5a; Pl. 21, specimen 258.15-1a; Pl. 23, specimen 409-4a; Pl. 27, specimen LEST54-23a; Pl. 28, specimens 7341c5, 7341-48a and 7341-52a; Pl. 29, specimens 7339-18a, 7338-13a and 7338-10b; Pl. 31, specimens 6812-4a and SAN21-1a
2014 Orbitolinopsis debelmasi Moullade & Thieuloy - Polavder; Fig. 7A-C
2015 Orbitolinopsis debelmasi - Bonvallet; Pl. 4.2, figure 10
2019 Orbitolinopsis debelmasi - Bonvallet et al.; Fig. 10.25
Remarks: Initially,
the type-specimens were illustrated as Orbitolinopsis
kiliani Prever by Moullade
(1960, Pl. 3, figs. 6-12), later
described as a new species, Orbitolinopsis
debelmasi by Moullade & Thieuloy
(1965). At the
type-locality, it occurs in high-energetic deposits (grainstones; "Facičs
microconglomératique", Moullade, 1960, p. 188). In the studied
sections of eastern Serbia, O. debelmasi
has been observed in comparable facies, grainstones with ooids, e.g., in the upper parts of the Prekonozi and Jerma sections. O.
debelmasi is characterized by its large-sized test of high-conical (apical
angles 40 to 70 degrees) morphology (often with an ultimate cylindrical part, Pl. 10 ![]() , figs. 8-9), a wide central zone with a pattern of regularly alternating
cupules, and vertical plates (intercalary beams) in the marginal zone. The
pattern of the foramina cross-wise obliquely arranged
in diagonal lines is well discernible (Pl. 10
, figs. 8-9), a wide central zone with a pattern of regularly alternating
cupules, and vertical plates (intercalary beams) in the marginal zone. The
pattern of the foramina cross-wise obliquely arranged
in diagonal lines is well discernible (Pl. 10 ![]() , figs. 4, 6, 12). Adult specimens
consist of up to 35 uniserial chambers.
, figs. 4, 6, 12). Adult specimens
consist of up to 35 uniserial chambers.
Dimensions: D = 0.7-0.95 mm, H = 1.0-1.5 mm, D/H = 0.5-0.75; number of chambers last 0.5 mm = 9-11.
Occurrences: Rare in the Kamenica section, common in the Prekonozi and Jerma sections.
Genus Urgonina Foury & Moullade, 1966
Remarks: Correctly, the genus Urgonina is classified as belonging to the Dictyoconinae (Loeblich & Tappan, 1987; Kaminski, 2014), and not to the Orbitolininae (BouDagher-Fadel & Price, 2019, p. 6, Fig. 4).
Urgonina alpillensis (Foury, 1963)
(Pl. 11 ![]() , figs. 1-9)
, figs. 1-9)
1963 Coskinolina alpillensis n. sp. - Foury; Pl. 1, fig. 7, non 8; Pl. 3, figs. 1-9
1966 Urgonina protuberans n. sp. - Foury & Moullade, p. 252; Pl. 1, figs. 7-19; Pl. 2, figs. 1-11
1969 Urgonina cf. alpillensis (Foury) - Schroeder et al., p. 93-96; non Pl. 2, figs. 1-6 [= Montseciella glanensis (Foury) fide Cherchi & Schroeder, 1999]; Pl. 3, fig. 6
1980 Urgonina alpillensis (Foury) - Arnaud-Vanneau, p. 617; Pl. 91, figs. 1-9
2007 Urgonina alpillensis (Foury) - Bucur et al.; Fig. 4/2
2006 Orbitolinopsis? sp. - Albrich et al.; Pl. 5, fig. 4
2009a Urgonina alpillensis - Clavel et al.; Pl. 2, specimen 6922-22a
2010 Urgonina alpillensis - Clavel et al.; Pl. 12, specimens PL18.3-2c, PL19.1-5a; Pl. 14, specimen 407.1-2b; Pl. 19, specimens Ai19-1a, Ai19-6d, Ai19-17d; Pl. 24, specimen 6923-1b
2013a Urgonina alpillensis (Foury) - Clavel in Granier et al., p. 153; Pl. 6, figs. 1-3; Pl. 9, fig. 6
2014 Urgonina alpillensis (Foury) - Clavel et al.; Pl. 6, specimen 372-43a; Pl. 8, specimen 7847-24a; Pl. 10, specimen 527-14a; Pl. 12, specimens 387.4-3a and 470-9b; Pl. 15, specimen 7865-11b; Pl. 16, specimen 493.3-3b; Pl. 19, specimen 263a1; Pl. 23, specimen 375-35b; Pl. 27, specimen LEST54-25c; Pl. 28, specimens 7341a17, 7341-14b, 7341-33c, 7341-38a; Pl. 29, specimens 6935-21a and 7338c 2; Pl. 31, specimens 6812-2b, SAN16-1c, SAN21-1c and SANF1-1c
Remarks: The
synonymy between Coskinolina alpillensis
Foury and Urgonina protuberans Foury
& Moullade was discussed by Schroeder et al. (1969) leading to the new combination Urgonina alpillensis (Foury)
(see also Clavel in Granier
et al., 2013a, 2017) and the monotypy
of the genus. It is worth mentioning that Arnaud-Vanneau (1980) raises the
question of two sub-species, alpillensis
protuberans and alpillensis
alpillensis with different stratigraphic abundances in the Vercors area, SE
France. U. alpillensis is
characterized by a well-developed initial spire (Pl. 11 ![]() ,
fig. 1), massive test
elements (septa, marginal chamber wall, pillar-like vertical elements) and
vertical pores. Shallow tangential sections show an undivided marginal zone (for
test reconstruction see Arnaud-Vanneau, 1980, Fig. 219). The upper
Berriasian specimens from eastern Serbia are smaller in dimensions than the
Barremian type-material (e.g., H = 1.0
to 1.875 mm according to Foury & Moullade,
1966). We consider
this variation as a simple intraspecific evolution (without any structural
differences) or example of Cope's rule (e.g., Guex,
2020).
,
fig. 1), massive test
elements (septa, marginal chamber wall, pillar-like vertical elements) and
vertical pores. Shallow tangential sections show an undivided marginal zone (for
test reconstruction see Arnaud-Vanneau, 1980, Fig. 219). The upper
Berriasian specimens from eastern Serbia are smaller in dimensions than the
Barremian type-material (e.g., H = 1.0
to 1.875 mm according to Foury & Moullade,
1966). We consider
this variation as a simple intraspecific evolution (without any structural
differences) or example of Cope's rule (e.g., Guex,
2020).
Dimensions: D = 0.6-0.9 mm, H = 0.85-1.0 mm, D/H = 0.9 (1 value)
Occurrences: Common in the Kamenica section, rare in the Jerma section (in grainstones).
Genus Valserina Schroeder & Conrad in Schroeder et al., 1967
Remarks: Although described more than half a century ago, the suprageneric status of Valserina is still pending in our opinion. In Loeblich & Tappan (1987) it is included in the subfamily Orbitolininae ("complex embryonal apparatus", p. 163 therein), whereas in the genus diagnosis (p. 168, therein), an "embryonic apparatus of simple protoconch and deuteroconch" is mentioned, that would consequently lead to its attribution to the Dictyoconinae Schubert. The current classification of Kaminski (2014) also lists Valserina among the Orbitolininae (see also Clavel in Granier et al., 2013a, 2017). This is obviously due to the fact that the type-species V. broennimanni has a simple embryo (without partitions) while Valserina turbinata (Foury, 1968) an embryo with partitions. Trivially, a genus cannot have species that belong to different subfamilies. It is noteworthy that Eygalierina turbinata Foury is treated as belonging to Palorbitolina in Schroeder [1993: Palorbitolina turbinata (Foury)] followed by Becker (1999). In the same work, Eopalorbitolina charollaisi Schroeder in Schroeder & Conrad, 1968, is newly combined as Valserina charollaisi (Schroeder) and Eopalorbitolina regarded as a junior synonym of Valserina. Later, Eopalorbitolina is again regarded as an individual genus different from Valserina and Eopalorbitolina charollaisi reinstated (Schroeder et al., 2010). A further increased taxonomic complexity arises from the reinstatement of Eygalierina turbinata Foury in Schroeder et al. (2002) and the giving up of its former transfer to Valserina. Last but not least, the validity of the three genera Valserina Schroeder & Conrad, Alpillina Foury, 1968, and Eygalierina Foury, 1968, has also been discussed by Arnaud-Vanneau (1980, p. 680) concluding that Eygalierina represents a junior synonym of Alpillina. Excluding the species turbinata (with complex embryo) from Valserina would speak for the inclusion of the latter into the Dictyoconinae that is provisionally accepted herein.
Valserina primitiva Schroeder et al., 1969
(Fig. 6 ![]() , Pl. 11
, Pl. 11 ![]() , figs. 10-18)
, figs. 10-18)
1969 Valserina brönnimanni primitiva n. ssp. - Schroeder et al., 99; Pl. 3, figs. 1-5
1999 Valserina primitiva Schroeder, Charollais & Conrad - Becker, p. 393; Pl. 5, figs. 1-4
1999 Valserina primitiva Schroeder, Charollais & Conrad - Schroeder et al.; Fig. 2
2002 Valserina primitiva Schroeder, Charollais & Conrad - Schroeder et al.; Pl. 2, fig. 9
2009a Valserina primitiva Schroeder, Charollais & Conrad - Clavel et al.; Pl. 1, specimen PL19.2-2a
2009 Valserina primitiva Schroeder, Charollais & Conrad - Charollais et al.; Pl. 4, figs. 12-16; Pl. 5, figs. 6-7; Pl. 6, figs. 8-10, 14-15
2010 Valserina primitiva Schroeder, Charollais & Conrad - Clavel et al.; Pl. 2, specimen 6894-3a; Pl. 4, specimens BC8Ga, BA19a.02-1a; Pl. 5, specimens BC23.05b, PA16.15a; Pl. 7, specimens 161.29-1b, 471-16a, 387.4b1; Pl. 12, specimen PL18.3-1c
2014 Valserina primitiva Schroeder, Charollais & Conrad - Clavel et al.; Pl. 6, specimen 7878-31a; Pl. 8, specimen GF9-2a; Pl. 10, specimen 447-21a; Pl. 12, specimen 161.29-1b; Pl. 21, specimen PL18.3-1c
2014 Valserina primitiva Schroeder, Charollais & Conrad - Polavder; Fig. 3.a-i
Remarks: The
radial zone of Valserina (incl. the
type-species V. broennimanni and V.
primitiva) is well developed with meandering septules that are thickening
inwards and foramina arranged in linear rows (Fig. 6.2 ![]() ,
Pl. 11
,
Pl. 11 ![]() ,
figs. 12-13,
15-17). The reticular zone shows meshes with polygonal to elongated-branchy
outlines. In the reticular (or central) zone, the chambers are connected by
cross-wise oblique pores arranged in diagonal line, as in the genus Orbitolinopsis.
The
subaxial section illustrated in Figure 6
,
figs. 12-13,
15-17). The reticular zone shows meshes with polygonal to elongated-branchy
outlines. In the reticular (or central) zone, the chambers are connected by
cross-wise oblique pores arranged in diagonal line, as in the genus Orbitolinopsis.
The
subaxial section illustrated in Figure 6 ![]() shows a specimen of a Chinese hat
morphology ("chapeau chinois", Arnaud-Vanneau, 1980, legend Fig. 206), that due to its low apical angle and size is interpreted as a microspheric
specimen. The same morphotype of a microspheric form has also been observed from
the type-species Valserina broennimanni
has been reported by Schroeder in Schroeder et
al. (1968, Fig. 4). The upper Berriasian specimen consists of 25 chambers;
it has a test diameter of ~1.85 mm and a height (almost complete) of ~1.2 mm.
There are 22 chambers for the last mm axial test length.
shows a specimen of a Chinese hat
morphology ("chapeau chinois", Arnaud-Vanneau, 1980, legend Fig. 206), that due to its low apical angle and size is interpreted as a microspheric
specimen. The same morphotype of a microspheric form has also been observed from
the type-species Valserina broennimanni
has been reported by Schroeder in Schroeder et
al. (1968, Fig. 4). The upper Berriasian specimen consists of 25 chambers;
it has a test diameter of ~1.85 mm and a height (almost complete) of ~1.2 mm.
There are 22 chambers for the last mm axial test length.
Dimensions: D = 0.8-1.0 mm, H = 0.95-1.6 mm (megalospheric specimens)
Occurrences: Rare to common in the Kamenica section.
|
Figure 6:
Valserina primitiva Schroeder
& Charollais from the upper Berriasian of the Kamenica section,
eastern Serbia. 1. Tangential section of a low conical specimen,
presumably microspheric; thin-section 16208,
Kamenica section. 2. Subaxial
section, specimen of Chinese hat morphology ("chapeau chinois", Arnaud-Vanneau,
1980, legend fig. 206), thin-section 14790. Abbreviations: m. z. = marginal
zone, rad. z. = radial zone, ret. z. = reticular zone. |
Genus Vanneauina Schlagintweit, 2020
Vanneauina vercorii (Arnaud-Vanneau, 1980)
(Pl. 12 ![]() )
)
1980 Dictyoconus? vercorii n. sp. - Arnaud-Vanneau, p. 653; Pl. 61, figs. 1-8; Pl. 97, figs. 1-10, Fig. 227
2011 Dictyoconus (collection Deloffre, origine inconnue) - Mathieu et al.; Photos 8-9
2013a Dictyoconus? vercorii Arnaud-Vanneau - Clavel in Granier et al., p. 151; Pl. 8, figs. 10-12; Pl. 9, fig. 7; Pl. 10, fig. 4
2014 Dictyoconus? vercorii Arnaud-Vanneau - Clavel et al.; Pl. 8, specimen 7847-11a; Pl. 10, specimen 7833-1c; Pl. 12, specimen 6898-2a; Pl. 17, specimen 575-1b; Pl. 22, specimen 407.2-16b. Pl. 25, specimen 229.1-1a1; Pl. 26, specimen LEST54-36a; Pl. 29, specimen 6935-4b; Pl. 32, specimen SANF10-2c
2020 Vanneauina vercorii (Arnaud-Vanneau) n. gen., n. comb. - Schlagintweit, p. 2; Fig. 2A-G
Remarks: This taxon is characterized by its cylindroconical test with a small initial spire, solid coalescing pillars (often largely filling the central chamber lumen and appearing as micritic masses or patches), and the lack of rafters (Arnaud-Vanneau, 1980, p. 650). Incompatible with Dictyoconus Blanckenhorn, it has been described as the new genus Vanneauina by Schlagintweit (2020). The dimensions of the upper Berriasian specimens are reduced compared to the Lower Cretaceous type-material of southeastern France (Arnaud-Vanneau, 1980). Specimens with comparable dimensions however have been illustrated by Clavel in Granier et al. (2013a, Pl. 8, figs. 10-12; 2017, Pl. 8, figs. 10-12) or Clavel et al. (2014, e.g., Pl. 8, specimen 7847-11a) from the Barremian-lower Aptian of the same area (see synonymy).
Dimensions: D = 0.65-1.0 mm, H = 0.9-1.2 mm, D/H 0.55-0.75
Occurrences: Common in the Kamenica section, rare in the Prekonozi section.
Gen. et sp. indet 1 (no rafters)
(Pl. 13 ![]() , figs. 1-6)
, figs. 1-6)
Remarks: This common taxon is characterized by medium- to high-conical tests, with adult chambers often reduced in width, and an almost plane base (e.g., Pl. 13, fig. 1). There are no rafters in the marginal zone, and the vertical main partitions are alternating between subsequent chambers; in axial sections the chambers are subdivided by numerous vertical pillar-like elements, their arrangement in the central zone remains unclear due to the lack of adequate transverse sections (Pl. 13, figs. 3, 5). Similar specimens have been illustrated by Polavder (2014) from the Kamenica section as belonging to Paleodictyoconus beckerae Clavel et al., 2009b. This species has, like any other species of the genus, a prominent early coil in the megalospheric form; the initial part of our specimens is unknown so that a preference is made herein for an open nomenclature.
Dimensions: D up to 0.85 mm, H up to 1.1 mm.
Occurrences: rare to common in the Kamenica section.
Gen. et sp. indet 2 (no rafters)
(Pl. 13 ![]() , figs. 7,
9-10)
, figs. 7,
9-10)
Remarks: This rare taxon is characterized by a coarse test structure, comparably high chambers and pillar-like elements in the central zone often triangular in shape (= the acute end pointing towards the apex = "pointe en haut"). In oblique sections, these elements do not regularly reach the septum of the chamber (= hemi-pillars; e.g., Clavel in Granier et al., 2013a, p. 153; 2017, p. 133) thus leaving a free space at its base. The poor material does not allow for a better characterization. The test habitus shows affinities to Paracoskinolina hispanica Peybernčs, 1976, a species that has been described from the Barremian of the Spanish Pyreneans, later recorded from the Upper Hauterivian - lower Barremian of various localities in southeastern France (Clavel et al., 2010, 2014; Clavel in Granier et al., 2013a, 2017). For comparison purpose one oblique section of Paracoskinolina? hispanica from southeastern France is re-illustrated herein (Clavel et al., 2014, Pl. 6, specimen EB H 27.4). In any case, the revision of this taxon is still pending; it cannot belong to the genus Paracoskinolina as the vertical main partitions are alternating between subsequent chambers (e.g., Peybernčs, 1976, Pl. 1, figs. 2 and 8, each in the upper test part).
Dimensions: D up to 0.85 mm, H up to 1.1 mm; noting a poor database.
Occurrences: Rare in the Kamenica section.
Gen. et sp. indet 3 (with rafters)
(Pl. 14 ![]() ,
figs. 6-11)
,
figs. 6-11)
Remarks: This rare and rather large-sized taxon is characterized by high-conical tests with up to 35 uniserial chambers; the initial part is unknown. The marginal zone is subdivided by one rafter per chamber; the number of intercalary beams (at least one up to ?) is unknown. The main partitions and vertical elements (pillars?) of the central zone are alternating between subsequent chambers. These structural features exclude the assignment to Cribellopsis on the one hand and Paracoskinolina on the other hand. More sections including the initial part and transverse sections showing the structure of the central zone and the vertical subdivision of the marginal zone are needed.
Dimensions: H up to 1.8 mm, D up to 0.75 mm, D/H = 0.5, number of chambers per last mm = 12-15
Occurrences: Lacking in the Kamenica section, gen. et sp. indet 3 has been observed only in the upper part of the Jerma section within high-energey deposits such as grainstones with ooids.
Subfamily DICTYORBITOLININAE Schroeder in Schroeder et al., 1990
Remarks: The three genera Paracoskinolina Moullade, Praedictyorbitolina Schroeder, and Dictyorbitolina Cherchi & Schroeder are attributed to the subfamily Dictyorbitolininae in the Lower Cretaceous. According to Schroeder in Schroeder et al. (1990, p. 196, translated), it is defined as follows: "The genera assigned to this subfamily are characterized by the presence of marginal foramina, situated at the periphery of the central zone forming a circular row. These apertures are arranged along vertical lines from the apex to the base. The primary septules show the same arrangement as the apertures". In the central zone, pilars may alternate (Dictyorbitolina, Praedictyorbitolina), or are aligned (Paracoskinolina).
Genus Paracoskinolina Moullade, 1965, emend. Arnaud-Vanneau, 1980
Remarks: The genus Paracoskinolina was originally established as a subgenus of Meyendorffina of the subfamily Dictyoconinae defined by a distinct intial trochospire, pillars in the central zone and the lack of horizontal partitions (Moullade, 1965, p. 4033). Paracoskinolina was emended by Arnaud-Vanneau (1980, p. 602) defined by the alignment of both the main partitions of the marginal zone and the pillars of the central zone (from apex to base). It is generally accepted that horizontal partitions are present in some species of the genus, e.g., P. arcuata (Arnaud-Vanneau) or P. reicheli (Guillaume). In the 70s and 80s Paracoskinolina became a wastebasket for orbitolinids with differing internal structures, and in any case incompatible with the emended diagnosis: P. fleuryi Decrouez & Moullade, 1974, P. pfenderae Canérot & Moullade, 1971 (see this work), P. prestati Peybernčs & Delmas, 1981, P. querolensis Canérot & Peybernčs, 1981, and P. hispanica Peybernčs, 1976.
Paracoskinolina aff. arcuata (Arnaud-Vanneau, 1976)
(Pl. 14 ![]() , figs. 1-4, 5?)
, figs. 1-4, 5?)
1976 Dictyoconus maynci arcuatus n. ssp. - Arnaud-Vanneau, p. 9; Pl. 1, figs. 4-8; Pl. 2, figs. 6-9.
1980 Paracoskinolina arcuata (Arnaud-Vanneau) - Arnaud-Vanneau, p. 609; Pl. 61, fig. 12; Pl. 99, figs. 11-13
2010 Paracoskinolina arcuata (Arnaud-Vanneau) - Clavel et al.; Pl. 1, specimen 136.2-4c; Pl. 14, specimen 407.2-1a; Pl. 24, specimen 6922a1
2014 Paracoskinolina arcuata (Arnaud-Vanneau) - Clavel et al.; Pl. 9, specimen MM373a; Pl. 13, specimen 6901-1a; Pl. 17, specimen 500.5-1a; Pl. 21, specimen 407.2-1a,
2019 Paracoskinolina arcuata (Arnaud-Vanneau) - Bonvallet et al.; Figs. 10/42, 10/51, 10/53, 10/59.
Remarks: The present rare oblique sections belong to medium- to high-conical tests displaying one rafter per chamber and aligned skeletal elements (Pl. 14, fig. 2). The latter feature is not always well constrained due to the obliqueness of the section plane. The specimens might belong to the high-conical to cylindro-conical P. arcuata. The presence of several orders of intercalary beams in the oblique transverse section (Pl. 14, fig. 5) however points to Paracoskinolina reicheli Guillaume, 1956, at least for this specimen rather than P. arcuata that has just one intercalary beam (Arnaud-Vanneau, 1976).
Dimensions: no data are provided to the rare oblique sections.
Occurrences: Rare in the upper parts of the Kamenica and Jerma sections.
The range of the family Orbitolinidae is indicated as Middle Jurassic to Oligocene in the fundamental work of Loeblich and Tappan (1987). The Middle Jurassic "record" of these taxa refers to Gutnicella (former Lucasella) (Gutnig & Moullade, 1981; Moullade et al., 1981) including also "Dictyoconus" cayeuxi (Lucas, 1938), and Meyendorffina Auroze & Bizon (see Moullade, 1965; Loeblich & Tappan, 1987, p. 159-160). Gutnicella, the oldest "foraminifčre orbitolinomorphe" (Septfontaine et al., 1991, p. 263), has been assigned to the Hauraniidae as well as Meyendorffina (Septfontaine, 1988, fig. 3; Kaminski, 2014). Ataxella Peybernčs, 1988 (type-species Paracoskinolina occitanica Peybernčs, 1974), from the Middle Jurassic of France, previously assigned to the Dictyoconinae has not been included in the classification of Kaminski (2014). It was assigned to the genus Gutnicella by BouDagher-Fadel (2008), later retained in the genus Ataxella Peybernčs, 1988 (not Bassoullet & Lorenz, 1988, as indicated in BouDagher-Fadel, 2018), and assigned to the Hauraniidae. Due to the rather homogeneous and massive columella, Ataxella is in our opinion better placed within the Pfenderinidae. The Upper Jurassic "record" refers to the taxa Kilianina Pfender, 1933, respectively the type-species K. blancheti Pfender, 1933, and K. rahonensis Foury & Vincent, 1967 (= Neokilianina acc. to Septfontaine, 1988), and Parurgonina Cuvillier et al., 1968. According to the classification of Kaminski (2014), Kilianina belongs to the Paravalvulinidae Banner et al. whereas Parurgonina belongs to the Parurgoninidae Septfontaine. Summarizing, all Middle and Upper Jurassic species previously assigned to the Orbitolinidae have meanwhile been included in other families. The first records of the Orbitolinidae date back to the upper Berriasian (Darsac, 1983; Granier, 2019; Bucur, et al., 2020; Schlagintweit & Yazdi-Moghadam, 2020: Cribellopsis delicatula). Another form, Valdanchella miliani (Schroeder), considered by many workers as a Valanginian marker taxon (Azéma et al., 1977; Martín-Closas & Salas, 1998; Cherchi & Schroeder in Schroeder et al., 2000) has also been reported from upper Berriasian strata (Bádenas et al., 2004; Granier et al., 2014, Fig. 2). The reported assemblage herein from eastern Serbia further manifests the late Berriasian age of the first representatives of the family. The outstanding species enrichment from the upper Berriasian of eastern Serbia with 18 taxa is highlighted by the contrast to just three different taxa recognized by Darsac (1983) from the upper Berriasian of the French Jura Mountains.
The discussion of phylogenetic relationships and models for any fossil group requires detailed knowledge on the stratigraphic ranges of the taxa involved. This prerequisite is even more important for a group such as the Orbitolinidae that lack any modern representatives, and thus exclude the application of molecular data analyses (e.g., Pawlowski et al., 2013). Otherwise, the assumption of different stratigraphic ranges must necessarily result in contradictionary views on the phylogeny of the group. In the literature there are different views on the phylogeny of the Orbitolininidae (or the genus Orbitolina) of which the most important ones have been summarized by Henson (1948, p. 73). Henson already recognized the most important "variation(s) from older to younger communities", namely increase of (i) test size, (ii) complexity of the marginal zone ("sub-epidermal layer"), (iii) the size and complexity of the megalospheric embryo along with its (iv) shift from a spiral-eccentric towards a central (apical) position. Note that the increasing complexity in the marginal zone is a prominent criterion for the subfamily Dictyoconinae, while in the Orbitolininae it is the increasing complexity of the embryo. It is worth mentioning that the principles of Henson are still the base for the widely accepted temporary concept of different lineages within the three subfamilies Dictyoconinae, Orbitolininae, and Dictyorbitolininae (e.g., Schroeder, 1975; Schroeder et al., 2002, 2010). In his review, Henson (1948, p. 75), distinguished the four structurally different groups of Dictyoconus, Dictyoconella, Orbitolinopsis, and Orbitolina. Direct relationships (e.g., ancestor-descendant) between different genera were not refined certainly also due to the lack of knowledge on the accuracy of stratigraphic ranges. Numerous genera that might have functioned as missing links, have also been described later. A phylogeny, that took "into account not only the morphologic stage of development but also the stratigraphic occurrence" was proposed almost 20 years later by Douglass (1960, p. 249). Douglass regarded Coskinolinoides texanus Keijzer, a "primitive" form without horizontal partitions (= rafters) as the ancestor from which other genera evolved in "several directions", namely Orbitolina (= Mesorbitolina) texana, Simplorbitolina manasi Ciry & Rat, Iraqia simplex Henson, and two branches with the Eocene Dictyoconus (= Coskinon) floridanus (Cole) and the Albian Dictyoconus walnutensis (Carsey), and another one with the Eocene D. indicus Davies (Douglass, 1960, Fig. 3). This scheme is not commented in detail. We only want to stress that the stratigraphic distribution of these genera does not fit such a reconstruction: the Albian C. texanus cannot be the ancestor of the older forms S. manasi or I. simplex.
Several
(if not all) authors however agree that the Orbitolinidae represent a
morphologically rather heterogeneous group composed of convergent evolutionary
lines (e.g., Pokorný, 1963).
The most recent contribution to this topic was provided by BouDagher-Fadel
& Price (2019, Fig. 4) with a phylogenetic evolution scheme where
"gradual morphological changes" occurred through time. Among several points
that would be worth mentioning, we just name two. First, the stratigraphic
ranges indicated for the genera following each other (ancestor-descendant) are,
independently from the new data from Serbia (Bucur et
al., 2020), not accurate. The range of Orbitolinopsis
for example is indicated as Aptian-Albian to stress its descendance from Paleodictyoconus
with an inferred range of late Valanginian to Aptian. It is already widely known
that the oldest record of both Orbitolinopsis
and Paleodictyoconus, previously
known, dates back to the Late Hauterivian (Charollais et al.,
2010; Clavel et
al., 2010, 2014; Granier et
al., 2013a, 2017). In addition, the assumed evolution from forms with
hemi-pillars (Paleodictyoconus; e.g.,
Schroeder in Schroeder & Charollais,
1966, p. 109-110)
to forms with cupules in the central zone (Orbitolinopsis;
e.g., Arnaud-Vanneau, 1980)
appears quite arbitrarly and has not been further discussed. Last but not least,
the presentation may create the appearance of facts rather than of one
hypothesis. Anyway, the new data give evidence that numerous genera display a
near-simultaneous appearance (or FAD) in a parallel manner, not one after the
other. Independently from the evolutionary first appearance, the lower part of
the Kamenica section appears unsuitable from a facies point of view (e.g.,
fenestral limestones) for the proliferation of the group (see Fig. 2 ![]() ). Higher up
in the stratigraphic section the establishment of inner platform conditions
allowed the occupation and colonization of available or new ecospaces, e.g.,
muddy lagoonal facies (wackestones) and external shoal facies (grainstones). In
the latter, the preference of some taxa is evident (Orbitolinopsis debelmasi, Urgonina
alpillensis, Paracoskinolina aff. arcuata,
gen. et sp. indet 3). Thus, the early history of the Orbitolinidae can be
interpreted as an example of adaptive radiation where a clade displays a rapid
diversification (18 taxa recorded herein!) and ecological exploitation (Simpson,
1944). This kind of evolution is also often referred to as "early bursts" (e.g., Puttick,
2018). The major part of the genera that later
lead to the "Urgonian" diversification (e.g., Arnaud-Vanneau, 1980; Clavel et
al., 2014) were already present at the very beginning of the group.
Therefore, the new data provide a solid data base for future further refinements
of phylogenetic traits within the Orbitolinidae. The
explosive radiation of the Orbitolinidae in the late Berriasian is accompanied
by the FAD of several other large benthic foraminifera including mostly
agglutinating (e.g., Ammocycloloculina Maync, Choffatella
Schlumberger, Drevennia Arnaud-Vanneau,
Eclusia Septfontaine, Moulladella
Bucur & Schlagintweit, Pfenderina
Henson, Pseudotextulariella Barnard)
but also complex porcelaneous taxa (Pavlovcevina
Loeblich & Tappan) (e.g.,
Granier, 2019; Bucur et al.,
2020) providing evidence for a thus far unknown bioevent in this time period
that exceeds the number of taxa originating in the previous (Tithonian) and the
following stage (Valanginian) (e.g., Kaminski
et al., 2008, Fig. 1). Last but not
least, the Berriasian also witnessed the appearance of new genera of the
aragonitic Robertinida described only recently by Rigaud et
al. (2021).
). Higher up
in the stratigraphic section the establishment of inner platform conditions
allowed the occupation and colonization of available or new ecospaces, e.g.,
muddy lagoonal facies (wackestones) and external shoal facies (grainstones). In
the latter, the preference of some taxa is evident (Orbitolinopsis debelmasi, Urgonina
alpillensis, Paracoskinolina aff. arcuata,
gen. et sp. indet 3). Thus, the early history of the Orbitolinidae can be
interpreted as an example of adaptive radiation where a clade displays a rapid
diversification (18 taxa recorded herein!) and ecological exploitation (Simpson,
1944). This kind of evolution is also often referred to as "early bursts" (e.g., Puttick,
2018). The major part of the genera that later
lead to the "Urgonian" diversification (e.g., Arnaud-Vanneau, 1980; Clavel et
al., 2014) were already present at the very beginning of the group.
Therefore, the new data provide a solid data base for future further refinements
of phylogenetic traits within the Orbitolinidae. The
explosive radiation of the Orbitolinidae in the late Berriasian is accompanied
by the FAD of several other large benthic foraminifera including mostly
agglutinating (e.g., Ammocycloloculina Maync, Choffatella
Schlumberger, Drevennia Arnaud-Vanneau,
Eclusia Septfontaine, Moulladella
Bucur & Schlagintweit, Pfenderina
Henson, Pseudotextulariella Barnard)
but also complex porcelaneous taxa (Pavlovcevina
Loeblich & Tappan) (e.g.,
Granier, 2019; Bucur et al.,
2020) providing evidence for a thus far unknown bioevent in this time period
that exceeds the number of taxa originating in the previous (Tithonian) and the
following stage (Valanginian) (e.g., Kaminski
et al., 2008, Fig. 1). Last but not
least, the Berriasian also witnessed the appearance of new genera of the
aragonitic Robertinida described only recently by Rigaud et
al. (2021).
A "remarkable stratigraphic identity" between the Urgonian of southeastern France and the southern Pyreneans (France-Spain) has been stated by Becker (1999) and Schroeder et al. (2000, p. 34). This cross reference is provided here, due to species common to both this area and eastern Serbia. The orbitolinid microfauna of southeastern France also shows many species in common with Sardinia, Italy (e.g., Dieni et al., 1963; Cherchi & Schroeder, 1978, 1999). This accordance is not really astonishing bearing in mind that during the Lower Cretaceous the Corsica-Sardinia microplate was in a position proximal to the European continent (here: Provence area of southeastern France) (e.g., Gattacceca et al., 2007). The diversity of Lower Cretaceous Orbitolinidae from these northern Tethysian occurrences is distinctly greater than the one recorded from its southern margin, e.g., northern Africa (e.g., Peybernčs, 1982). There, the records are also younger lacking any late Berriasian taxa. It is worth mentioning that later, for example during the Cenomanian, the diversity of the southern Tethysian margin (with special reference to the Arabian Plate) exceeds the one observed from the northern Neotethysian margin (Schroeder & Neumann, 1985; Yazdi-Moghadam & Schlagintweit, 2021).
The upper Berriasian orbitolinid microfauna of eastern Serbia is characterized by very high diversity including at least 18 taxa belonging to at least seven genera. One species is described as the new species Cribellopsis sudari. Morphologically, high-conical forms with predominantly simple exoskeleton (= lack of rafters) are dominating. The observed taxa belong to the Dictyorbitolininae (genus Paracoskinolina) and Dictyoconinae (all other genera obsereved); Orbitolininae are absent.
With presumably inconvenient environmental conditions in the lower Berriasian (e.g., fenestral limestones), the appearing of inner platform facies favoured the rapid (explosive) adaptive radiation of the group. The sudden and more or less time-equivalent appearance of all these genera, contrasts evolutionary concepts with "gradual morphological changes" and ancestor-descendant relationships. This observation does not question lineages among the Dictyoconinae (e.g., with increasing complexity of the marginal zone) and the Orbitolininae (e.g., increasing complexity and size of the embryo).
Almost all taxa observed herein are known from the Lower Cretaceous (Upper Hauterivian-lower Aptian) of southeastern France. With the distinct extension of the stratigraphic ranges, the biostratigraphic value of several species is distinctly lowered.
The rich assemblages of the Orbitolinidae of eastern Serbia (upper Berriasian) and southeastern France (Upper Hauterivian-lower Aptian) belong to the former northern Neotethysian margin contrasting an impoverished assemblage from the southern Neotethysian margin, where upper Berriasian taxa have so far not been recorded.
Remarks of reviewers Michel Septfontaine (Froideville), Danielle Decrouez (Genčve) and editor Daniela Basso (Milano) are kindly acknowledged. We are grateful to the memory of Bernard Clavel (Charollais & Granier, 2020) for providing us some pictures from his personal material from southeastern France for comparison purposes. Thanks to Milan Sudar, Divna Jovanovic and Svetlana Polavder for assistance in the field. The samples have been collected within the framework of the bilateral cooperation between the Serbian Academy of Science and Arts and the Romanian Academy, 2016.
Albrich S., Bernaus J.M., Boix C., Caus E., Martín-Closas C., Salas R., Vicedo V. & Villalonga R. (2006).- Caracterizacíon bioestratigráfica y paleoambiental del Cretácico inferior (Berriasiense-Barremiense) del Macizo de Garraf (Cadena Costera Catalana).- Revista Espańola de Micropaleontología, Madrid, vol. 38, nos. 2-3, p. 429-452.
Anđelković J., Krstić B., Bogdanović P., Jadranin D., Milošaković R., Milenković P., Fotić V. & Jovanović L. (1975).- Basic Geological Map of SFRY, 1:100,000, Sheet Breznik (K 34-46).- Savezni geološki zavod, Beograd (Zavod za geološka i geofizička istraživanja, Beograd, 1964-1968).
Anđelković J., Krstić B., Bogdanović P., Jadranin D., Milenković P., Milošaković R., Urošević D., Dimitrijević M.N., Dolić D., Rakić M.O., Jovanović L., Maslarević L., Marković B., Divljan M. & Đorđević M. (1977).- Explanatory text: Geology of the Sheets Pirot (K 34-34) and Breznik (K 34-46).- Savezni geološki zavod, Beograd (Zavod za geološka i geofizička istraživanja, Beograd, 1969), 67 p. [in Serbian, English and Russian summaries].
Arnaud-Vanneau A. (1976).- Étude de l'évolution Barrémo-Bédoulienne d'un Orbitolinidé Urgonien Dictyoconus maynci (Chevalier) et description d'une nouvelle sous-espčce : Dictyoconus maynci arcuatus.- Revista Espańola de Micropaleontología, Madrid, vol. 3, no. 1, p. 5-14.
Arnaud-Vanneau A. (1980).- L'Urgonien du Vercors septentrional et de la Chartreuse.- Géologie Alpine Mémoire, Grenoble, vol. 11 (3 volumes), 874 p.
Arnaud-Vanneau A. & Schroeder R. (1976).- Paleodictyoconus actinostoma n. sp. Orbitolinidé nouveau des "couches ŕ orbitolines" intra-urgoniennes du Vercors (France).- Géobios, Villeurbanne, vol. 9, no. 3, p. 279-289.
Arnaud-Vanneau A. & Thieuloy J.-P. (1972).- Orbitolinopsis buccifer n. sp. nouveau représentant du genre Orbitolinopsis Silvestri dans les calcaires urgoniens du Vercors (France).- Revue de Micropaléontologie, Paris, vol. 14, no. 4, p. 248-258.
Azéma J., Chabrier G., Fourcade É. & Jaffrezo M. (1977).- Nouvelles données micropaléontologiques, stratigraphiques et paléogéographiques sur le Portlandien et le Néocomien de Sardaigne.- Revue de Micropaléontologie, Paris, vol. 20, no. 3, p. 125-139.
Azéma J., Chabrier G., Chauve P. & Fourcade É. (1979).- Nouvelles données stratigraphiques sur le Jurassique et le Crétacé du Nord-Ouest d'Ibiza (Baléares, Espagne).- Geologica Romana, vol. 8, p. 1-12.
Bádenas B., Salas R. & Aurell M. (2004).- Three orders of regional sea-level changes control facies and stacking pattern of shallow platform carbonates in the Maestrat Basin (Tithonian-Berriasian, NE Spain).- International Journal of Earth Sciences (Geologische Rundschau), vol. 93, no. 1, p. 144-162.
Becker E. (1999).- Orbitoliniden-Biostratigraphie der Unterkreide (Hauterive - Barręme) in den spanischen Pyrenäen (Profil Organyŕ, Prov. Lérida).- Revue de Paléobiologie, Genčve, vol. 18, no. 2, p. 359-489.
Benzaggagh M. (2020).- Discussion on the calpionellid biozones and proposal of a homogeneous calpionellid zonation for the Tethyan Realm.- Cretaceous Research, vol. 114, article 104184, 24 p.
Bernaus J.M. (2000).- L'Urgonien du Bassin d'Organyŕ (NE de l'Espagne) : Micropaléontologie, sédimentologie et stratigraphie séquentielle.- Géologie Alpine Mémoire, Grenoble, no. 33, 138 p.
Bernaus J.M., Arnaud-Vanneau A. & Caus E. (2002).- Stratigraphic distribution of Valanginian-Early Aptian shallow-water benthic foraminifera and algae, depositional sequences of a carbonate carbonate platform in a tectonically-controlled basin: The Organyŕ Basin, Pyrenees, Spain.- Cretaceous Research, vol. 23, no. 1, p. 25-36.
Blanc É. (1996).- Transect plate-forme-bassin dans les séries carbonatées du Berriasien supérieur et du Valanginien inférieur (Domaines jurassien et du nord-Vocontien) - Chronostratigraphie et transferts des sédiments.- Géologie Alpine Mémoire H.S., Grenoble, no. 25, 312 p.
Blau J. & Grün B. (1997).- Late Jurassic/Early Cretaceous revised calpionellid zonal and subzonal division and correlation with ammonite and absolute time scales.- Mineralia Slovaca, Bratislava, vol. 29, p. 297-300.
Boisseau T. (1987).- La plate-forme jurassienne et sa bordure subalpine au Berriasien-Valanginien (Chartreuse-Vercors). Analyse et corrélation avec les séries de bassin.- Thčse Université Grenoble, 413 p.
Bonvallet L. (2015).- Evolution of the Helvetic shelf (Switzerland) during the Barremian-Early Aptian: Paleoenvironmental, paleogeographic and paleooceanic controlling factors.- Thčse Université Lausanne, 333 p., URL: https://serval.unil.ch/resource/serval:BIP_8DF48AF8B7D5.P001/REF.pdf
Bonvallet L., Arnaud-Vanneau A., Arnaud H., Adatte T., Spangenberg J.E., Stein M., Godet A. & Föllmi K.D. (2019).- Evolution of the Urgonian shallow-water carbonate platform on the Helvetic shelf during the late Early Cretaceous.- Sedimentary Geology, vol. 387, p. 18-56.
BouDagher-Fadel M.K. (2008).- Evolution and geological significance of Larger Benthic Foraminifera (First edition).- Developments in Palaeontology and Stratigraphy, 544 p.
BouDagher-Fadel M.K. (2018).- Evolution and geological significance of Larger Benthic Foraminifera (Second edition).- UCL Press London, 693 p.
BouDagher-Fadel M.K. & Price G.D. (2019).- Global evolution and paleogeographic distribution of mid-Cretaceous orbitolinids.- UCL Open: Environment, London, URL: https://dx.doi.org/10.14324/111.444/ucloe.000001
Bucur I.I., Arnaud-Vanneau A. & Arnaud H. (2007).- Remarks on Triploporella praturlonii Barattolo, 1982 from the Lower Barremian of Southern Vercors (SE France).- Neues Jahrbuch Geologie und Paläontologie Monatshefte, Stuttgart, vol. 244, no. 2, p. 133-141.
Bucur I.I. & Cociuba I. (1988).- La plate-forme carbonatée du Crétacé inférieur des Monts Pădurea Craiului (Monts Apuseni, Roumanie). Biostratigraphie et configuration.- Studia Universitatis Babeş-Bolyai, Geologia, Cluj-Napoca, vol. 43, no. 2, p. 89-100.
Bucur I.I. & Schlagintweit F. (2018).- Moulladella jourdanensis (Foury & Moullade, 1966) n. gen., n. comb.: Valanginian-early late Barremian larger benthic foraminifera from the Northern Neotethyan Margin.- Acta Palaeontologica Romaniae, Bucharest, vol. 14, no. 2, p. 45-59.
Bucur I.I., Conrad M.A. & Radoičić R. (1995).- Foraminifers and calcareous algae from Valanginian limestones in the Jerma River Canyon, eastern Serbia.- Revue de Paléobiologie, Genčve, vol. 14, no. 2, p. 349-377.
Bucur I.I., Sudar M., Schlagintweit F., Pleş G., Săsăran E., Jovanović D., Polavder S. & Radoičić R. (2020).- Lowermost Cretaceous limestones from the Kučaj zone (Carpatho-Balkanides, Eastern Serbia): New data on their age assignment.- Cretaceous Research, vol. 116, article 104575, 20 p.
Canérot J. (1974).- Recherches géologiques aux confins des chaînes ibérique et catalane (Espagne).- Thčse Université Paul Sabatier Toulouse, 517 p.
Canérot J. (1984).- Fluctuations marines et évolution biologique : Exemple du Néocomien des Ibérides Orientales (Espagne).- Benthos '83, 2nd International Symposium on Benthic Foraminifera (Pau, April 1983), Pau and Bordeaux, p. 131-139.
Canérot J. & Moullade M. (1971).- Le Valanginien ŕ facičs marin dans le Maestrazgo (Provincias de Castéllon, Tarragona, Teruel - Espagne). Étude particuličre des Orbitolinidées Valdanchella n. gen, Paracoskinolina pfendere n. sp.- Archives des Sciences, Genčve, vol. 24, no. 2, p. 207-218.
Canérot J. & Peybernčs B. (1981).- Paracoskinolina querolensis n. sp., orbitolinidé nouveau de l'Hauterivien calcaire des Ibérides sud-orientales (Espagne).- Archives des Sciences et Compte Rendu des Séances de la Société, Genčve, vol. 34, p. 191-200.
Castro J.M., Company M., De Gea G.A. & Aguado R. (2001).- Biostratigraphy of the Aptian-Middle Cenomanian platform to basin domain in the Prebetic Zone of Alicante, SE Spain: Calibration between shallow-water benthonic and pelagic scales.- Cretaceous Research, vol. 22, p. 145-156.
Charollais J., Clavel B., Busnardo R., Conrad M., Müller A. & Decrouez D. (2009).- Olistolithes et coulées bioclastiques : Prémices de l'installation de la plate-forme urgonienne aux confins des Bornes et des Aravis (Haute-Savoie, France).- Archives des Sciences, Genčve, vol. 62, p. 35-70.
Charollais J. & Granier B. (2020).- Nécrologie Bernard Clavel.- Archives des Sciences, Genčve, vol. 71, p. 95-100.
Cherchi A. & Schroeder R. (1978).- Osservazioni sul gen. Orbitolinopsis Silvestri (Foraminiferida) e sua presenza nel Barremiano della Sardegna.- Bollettino della Societŕ Sarda di Scienze Naturali, Sassari, Anno XI, vol. 17, p. 159-167.
Cherchi A. & Schroeder R. (1999).- Montseciella, a new orbitolinid genus (Foraminiferida) from the Uppermost Hauterivian - Early Barremian of SW Europe.- Treballs del Museu de Geologia de Barcelona, vol. 8, p. 5-23.
Chiocchini M. (1989).- Cribellopsis arnaudae n. sp. (Foraminiferida, Orbitolinidae) nel Cretaceo inferior del Lazio meridionale e dell' Abruzzo (Italia central).- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 28, no. 1, p. 39-48.
Clavel B., Busnardo R., Charollais J., Conrad M. & Granier B. (2009a).- Nouvelles données sur la répartition biostratigraphique des orbitolinidés ŕ l'Hauterivien supérieur, au Barrémien et ŕ l'Aptien inférieur.- Archives des Sciences, Genčve, vol. 62, p. 125-146.
Clavel B., Busnardo R., Charollais J., Conrad M. & Granier B. (2010).- Répartition biostratigraphique des orbitolinidés dans la biozonation ŕ amonites (plate-forme urgonienne du Sud-Est de la France). Partie 1: Hauterivien supérieur-Barrémian basal).- Carnets Geol., Madrid, vol. 10, no. A06 (CG2010_A06), 53 p., DOI: 10.4267/2042/33369.
Clavel B., Charollais J., Busnardo R., Granier B., Conrad M.A., Desjaques P. & Metzger J. (2014).- La plate-forme carbonatée urgonienne (Hauterivien supérieur-Aptien inférieur) dans le Sud-Est de la France et en Suisse : Synthčse.- Archives des Sciences, Genčve, vol. 67, p. 1-100.
Clavel B., Decrouez D., Charollais J. & Busnardo R. (2009b).- Paleodictyoconus beckerae n. sp., un orbitolinidé (Foraminifčre) nouveau de l'Hauterivien supérieur (Crétacé) ŕ facičs urgonien (SE France, Jura franco-suisse, Pyrénées espagnoles).- Archives des Sciences, Genčve, vol. 62, p. 23-34.
Cruz-Abad E. (2018).- Textura y arquitectura de los Orbitolinoideos (Superfamilia Orbitolinoidea): Revisión y caracterización.- PhD Thesis University of Barcelona, 154 p., URL: https://www.tesisenred.net/handle/10803/665998#page=1
Darsac C. (1983).- La plate-forme berriaso-valanginienne du Jura méridional aux massifs subalpins (Ain, Savoie). Sédimentologie, minéralogie, stratigraphie, paléogéographie, micropaléontologie.- Thčse de 3e cycle, Université de Grenoble, 319 p.
Decrouez D. & Moullade M. (1974).- Orbitolinidés nouveaux de l'Albo-Cénomanien de Grčce.- Archives des Sciences, Genčve, vol. 27, no. 1, p. 75-92.
Dieni L., Massari F. & Moullade M. (1963).- Sur quelques Orbitolinidae des calcaires ŕ facies "urgonien" du Crétacé inférieur des environs d'Orosei (Sardaigne).- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 2, no. 2, p. 3-8.
Douglass R.C. (1960).- Revision of the family Orbitolinidae.- Micropaleontology, New York, vol. 6, p. 249-270.
Fourcade É. (1970).- Microfacičs du Jurassique et du Crétacé aux confins des Chaînes Bétiques et Ibériques (Sud-Est de l'Espagne).- Thčse Université de Paris, 427 p.
Foury G. (1963).- Deux nouvelles espčces d'Orbitolinidae du facičs Urgonien des Alpilles (Bouches du Rhône).- Revue de Micropaléontologie, Paris, vol. 6, no. 1, p. 3-12.
Foury G. (1968).- Le Crétacé inférieur des Alpilles. Contribution ŕ l'étude stratigraphique et micropaléontologique.- Géobios, Villeurbanne, vol. 1, p. 119-164.
Foury G. & Moullade M. (1966).- Orbitolinidae nouveaux du Barrémien (Facičs Urgonien) des Alpilles (Bouches-du-Rhône).- Revue de Micropaléontologie, Paris, vol. 8, p. 249-257.
García-Hernández M. (1978).- El Jurasico terminal y el Cretacico inferior en las Sierras de Carzola y del Segura (Zona Prebetica).- PhD Thesis University of Granada, 344 p., URL: https://digibug.ugr.es/handle/10481/53663?show=full
Gattacceca J., Deino A., Rizzo R., Jones D.S., Henry B., Beaudoin B. & Vadeboin F. (2007).- Miocene rotation of Sardinia: New paleomagnetic and geochronological constraints and geodynamic implications.- Earth and Planetary Science Letters, vol. 258, p. 359-377.
Ghafor I.M & Mohialdeen I.M.J (2018).- Early cretaceous microfosiils associations (foraminifera, ostracoda, calcareous algae, and coral) from the Garagu Formation, Duhok Area, Kurdistan Region, Northern Iraq.- Arabian Journal of Geosciences, vol. 11, no. 15, article 407, 1-17.
Granier B. (2019).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition.- Cretaceous Research, vol. 93, p. 245-274.
Granier B. (2020).- Discussion of the paper by Imad M. Ghafor and Ibrahim M.J. Mohialdeen, 2018, entitled "Early Cretaceous microfosiils associations (foraminifera, ostracoda, calcareous algae, and coral) from the Garagu Formation, Duhok Area, Kurdistan Region, Northern Iraq" (Arabian Journal of Geosciences, 11:407).- Arabian Journal of Geosciences, vol. 13, article 60, 10 p.
Granier B. & Clavel B. (2019).- Revised dating of the major earliest Cretaceous transgression in S Aquitaine (SW France).- Cretaceous Research, vol. 99, p. 246-254.
Granier B., Clavel B., Charollais J. & Weidmann M. (2014).- Latest Jurassic-Early Cretaceous dasycladalean algae (Chlorophyta) from the Morand drilling at Montricher (Canton of Vaud, Switzerland).- Acta Palaeontologica Romaniae, Bucharest, vol. 10, no. 1-2, p. 25-38.
Granier B., Clavel B., Moullade M., Busnardo R., Charollais J., Tronchetti G. & Desjaques P. (2013a).- L'Estellon (Baronnies, France), a "Rosetta Stone" for the Urgonian biostratigraphy.- Carnets Geol., Madrid, vol. 13, no. A04 (CG 2013_A04), p. 139-183.
Granier B., Clavel B., Moullade M., Busnardo R., Charollais J., Tronchetti G. & Desjaques P. (2013b).- Erratum to the article 2013/04 (CG 2013_A04), entitled "L'Estellon (Baronnies, France), a "Rosetta Stone" for the Urgonian biostratigraphy".- Carnets Geol., Madrid, vol. 13, no. A04 - Erratum (CG 2013_A04 - Erratum), p. 302.
Granier B., Clavel B., Moullade M., Busnardo R., Charollais J., Tronchetti G. & Desjaques P. (2017).- L'Estellon (Baronnies, France), a "Rosetta Stone" for the Urgonian biostratigraphy. In: Granier B. (ed.), Some key Lower Cretaceous sites in Drôme (SE France).- Carnets de Géologie, Madrid, Book 2017/01 (CG2017_B01), ISBN13: 978-2-916733-13-5, p. 117-163.
Guex J. (2020).- The controversial Cope's, Haeckel's and Dollo's evolutionary rules: The role of evolutionary retrogradation. In: Guex J., Torday J.S. & Miller W.B. Jr (eds.), Morphogenesis, environmental stress and reverse evolution.- Springer International Publishing, p. 13-22.
Guex J., Galster F. & Hammer Ř. (2016).- Discrete biochronological time scales.- Springer, Cham, 166 p.
Gutnig M. & Moullade M. (1981).- Données nouvelles sur le Jurassique et le Crétacé inférieur du Barla Dağ au Sud de Senırkent (Taurus de Pisidie, Turquie).- Bulletin of the Mineral Research Exploration Institute of Turkey, Foreign Edition, Ankara, vol. 69, p. 60-78.
Guillaume H. (1956).- Une nouvelle espčce crétacée du genre Dictyoconus Blanckenhorn.- Eclogae Geologicae Helvetiae, Basel, vol. 49, p. 141-146.
Haas J., Kovács S., Gawlick H.J., Grădinaru E., Karamata S., Sudar M., Péró C., Mello J., Polák M., Ogorelec B. & Buser S. (2011).- Jurassic evolution of the tectonostratigraphic units of the circum-Pannonian region.- Jahrbuch der Geologischen Bundesanstalt, Vienna, vol. 151, no. 3-4, p. 281-354.
Henson F.R.S. (1948).- Larger imperforate Foraminifera of south-western Asia. Families Lituolidae, Orbitolinidae and Meandropsinidae.- Monograph British Museum (Natural History), London, 127 p.
Holland S.M. & Patzkowsky M.E. (2002).- Stratigraphic variation in the timing of First and Last Occurrences.- Palaios, Lawrence - KS, vol. 17, p. 134-146.
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, no. M02 (CG2006_M02), 126 p.
Hottinger L. & Drobne K. (1980).- Early Tertiary conical imperforate foraminifera.- Razprave IV. razr. SAZU, Ljubljana, vol. 22, p. 188-276.
Hughes G.W (2000).- Bioecostratigraphy of the Shu'aiba Formation, Shaybah field, Saudi Arabia.- GeoArabia, vol. 5, p. 545-578.
Ivanova D. & Koleva-Rekalova E. (2004).- Agglutinated foraminifers in the framework of Southwestern Bulgarian palaeoenvironmental evolution during the Late Jurassic and Early Cretaceous. In: Bubík M. & Kaminski M.A. (eds.).- Proceedings of the 6th International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 8, p. 217-227.
Jankičević J. (1978).- Barrémien et Aptien des parties moyennes des Carpatho-Balkanides dans la Serbie orientale au point de vue du développement d'Urgonien.- Annales Géologiques de la Péninsule Balkanique, Belgrade, vol. 42, p. 103-194 [in Serbian, French summary].
Jankičević J. (1996).- Urgonian of the Carpatho-Balkanides of Serbia in the light of formation analysis.- Annales Géologiques de la Péninsule Balkanique, Belgrade, vol. 60, no. 2, p. 29-40 [in Serbian and English].
Kaminski M.A. (2014).- The year 2010 classification of the agglutinated foraminifera.- Micropaleontology, Paris, vol. 60, no. 1, p. 89-108.
Kaminski M.A., Setoyama E. & Cetean C.G. (2008).- Revised stratigraphic ranges and the Phanerozoic diversity of Agglutinated Foraminiferal Genera. In: Kaminski M.A. & Coccioni R. (eds.), Proceedings of the Seventh International Workshop on Agglutinated Foraminifera.- Gryzybowski Foundation Special Publication, Krakow, vol. 13, p. 79-106.
Krstić B., Rakić B., Veselinović M., Dolić D., Rakić M., Anđelković J. & Banković V. (1978).- Geological Map of RSF Yougoslavia, 1:10000, Sheet K 34-20 (Aleksinac).- Geological Institute, Belgrade.
Krstić B., Veselinović M., Divljan M. & Rakić M. (1980).- Explanation to the map of RSF Yougoslavia, 1:10000, sheet K 34-20 (Aleksinac).- Geological Institute, Belgrade, 59 p. [in Serbian].
Lakova I. & Petrova S. (2013).- Towards a standard Tithonian to Valanginian calpionellid zonation of the Tethyan Realm.- Acta Geologica Polonica, Warsaw, vol. 63, no. 2, p. 201-221.
Léonide P., Borgomano J., Masse J.-P. & Doublet S. (2012).- Relation between stratigraphic architecture and multi-scale heterogeneities in carbonate platforms: The Barremian-Lower Aptian of the Monts de Vaucluse, SE France.- Sedimentary Geology, vol. 265, p. 87-109.
Loeblich A.R. Jr & Tappan H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, 2 vol., 970 p. (847 Pls.).
Martín-Closas C. & Salas R. (1998).- Lower Cretaceous Charophyte biozonation in the Maestrat Basin (Iberian Ranges, Spain). A reply to P.O. Mojon [1996].- Géologie Alpine, Grenoble, vol. 74, p. 97-110.
Masse J.-P. & Fenerci-Masse M. (2013).- Stratigraphic updating and correlation of Late Barremian-Early Aptian Urgonian successions and their marly cover, in their type region (Orgon-Apt, SE France).- Cretaceous Research, vol. 39, p. 17-28.
Mathieu R., Bellier J.-P. & Granier B. (2011).- Manuel de micropaléontologie.- Carnets de Géologie, Madrid, Book 2011/02 (CG2011_B02), ISBN13: 978-2-916733-04-3, 123 p.
Moullade M. (1960).- Les Orbitolinidae des microfacičs barrémiens de la Drôme.- Revue de Micropaléontologique, Paris, vol. 3, no. 3, p. 188-198.
Moullade M. (1965).- Contribution au problčme de la classification des Orbitolinidae (Foraminiferida, Lituolacea).- Comptes rendus de l'Académie des Sciences, Paris, vol. 260, p. 4031-4034.
Moullade M. (1966).- Étude stratigraphique et micropaléontologique du Crétacé inférieur de la "fosse vocontienne".- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, Villeurbanne, vol. 15, 369 p. URL: https://www.persee.fr/doc/geoly_0076-1672_1966_mon_15_1
Moullade M., Haman D. & Huddleston R.W. (1981).- Gutnicella, a new name for Lucasella Gutnic and Moullade, 1967 (Foraminiferida) non Stewart, 1936 (Ostracoda).- Journal of Paleontology, vol. 55, p. 484.
Moullade M. & Thieuloy J.-P. (1965).- Précisions sur Orbitolina? kiliani, espčce-type d'Orbitolinopsis Silvestri.- Comptes Rendus de l'Académie des Sciences (groupe 9), Paris, t. 261, no. 11, p. 4175-4178.
Nichols G. (2009).- Sedimentology and stratigraphy. Second Edition.- Wiley-Blackwell, Chichester, 419 p.
Pawlowski J., Holzmann M. & Tyszka J. (2013).- New supraordinal classification of Foraminifera: Molecules meet morphology.- Marine Micropaleontology, vol. 100, p. 1-10.
Petković V. (1935).- Geology of Eastern Serbia.- Posebno izdanje Srpske kraljevske akademije, 105, Prirodnjački i matematički spisi, vol. 28, 218 p. [in Serbian].
Peybernčs B. (1974).- Paracoskinolina occitanica n. sp., Orbitolinidé nouveau du Bathonien supérieur des Pyrénées françaises.- Géologie méditerranéenne, Marseille, vol. 1, no. 1, p. 27-34.
Peybernčs B. (1976).- Paracoskinolina hispanica n. sp. : Orbitolinidé nouveau du Barrémien des Pyrénées espagnoles.- Archives des Sciences, Genčve, vol. 29, no. 3, p. 259-265.
Peybernčs B. (1982).- Les Orbitolinidés crétacés d'Afrique : Essai de synthčse.- Cahiers de Micropaléontologie, Paris, vol. 2, p. 13-28.
Peybernčs B. (1988).- Description d'Ataxella n. gen. (genero-type : Paracoskinolina occitanica Peybernčs, 1974) nouveau genre d'Orbitolinidé du Bathonien supérieur ouest-téthysien.- Comptes Rendus de l'Académie des Sciences (série II), Paris, vol. 307, p. 1097-1100.
Peybernčs B. & Delmas J. (1981).- Description de Paracoskinolina prestati nov. sp., Orbitolinidé nouveau du Crétacé moyen de la République de Somalie.- Bulletin de la Société d'Histoire naturelle de Toulouse, vol. 117, p. 80-86.
Pokorný V. (1963).- Principles of zoological micropaleontology.- International Series of Monographs of Earth Sciences, vol. 1, 668 p.
Polavder S. (2014).- Contribution to the biostratigraphy of the Lower Cretaceous in eastern Serbia: Upper Hauterivian Orbitolinidae from the Kamenica 1 section, Kurilovo anticline.- Cretaceous Research, vol. 48, p. 12-22.
Polavder S. & Radulović B. (2005).- Microfossils and brachiopods from the Lower Barremian at Prekonozi, eastern Serbia.- Annales Géologiques de la Péninsule Balkanique, Belgrade, vol. 66 (2004-2005), p. 55-61.
Pop G. (1994).- Calpionellid evolutive events and their use in biostratigraphy.- Romanian Journal of Stratigraphy, Bucharest, vol. 76, p. 7-24.
Puttick M.N. (2018).- Mixed evidence for early bursts of morphological evolution in extant clades.- Journal of Evolutionary Biology, Hoboken, vol. 31, no. 4, p. 502-515.
Radoičić R. (1978).- On distribution of Dasycladales in the Lower Cretaceous of Kurilovo, East Serbia.- Bulletin du Museum d'Histoire naturelle Beograd (Series A), vol. 33, p. 183-197 [in Serbian, with English summary].
Rehakova D. & Michalik J. (1997).- Evolution and distribution of calpionellids - the most characteristic constituents of Lower Cretaceous Tethyan mikroplankton.- Cretaceous Research, vol. 18, p. 493-504.
Remane J., Bakalova-Ivanova D., Borza K., Knauer J., Nagy I., Pop G. & Tardi-Filácz E. (1986).- Agreement on the subdivision of the standard calpionellid zones defined at IInd Planktonic Conference, Roma 1970.- Acta Geologica Hungarica, Budapest, vol. 29, no. 1-2, p. 5-14.
Rigaud S., Granier B. & Masse J.-P. (2021).- Aragonitic foraminifers: An unsuspected wall diversity.- Journal of Systematic Paleontology, vol. 19, no. 6, 27 p.
Schlagintweit F. (2020).- Taxonomic revision of Dictyoconus? vercorii Arnaud-Vanneau, 1980 (Foraminifera, Orbitolinidae) from the Lower Cretaceous (lowermost Barremian-lower Aptian) of southeastern France, Switzerland and Spain.- Cretaceous Research, vol. 110, article 104349, 6 p.
Schlagintweit F., Bucur I.I. & Sudar M.N. (2019a).- Bispiraloconulus serbiacus gen. et sp. nov., a giant arborescent benthic foraminifer from the Berriasian of Serbia.- Cretaceous Research, vol. 93, p. 98-106.
Schlagintweit F., Bucur I.I. & Sudar M.N. (2019b).- Corrigendum to "Bispiraloconulus serbiacus gen et sp. nov., a giant arborescent benthic foraminifer from the Berriasian of Serbia".- Cretaceous Research, vol. 94, p. 259.
Schlagintweit F. & Yazdi-Moghadam M. (2020).- Cribellopsis delicatula n. sp., an early Orbitolinidae from the upper Berriasian-lower Valanginian of SE France, Switzerland and Iran.- Micropaleontology, vol. 66, no. 4, p. 339-349.
Schroeder R. (1975).- General evolutionary trends in orbitolinas.- Revista Espańola de Micropaleontologia, Num. esp. 1975, Madrid, p. 117-128.
Schroeder R. (1993).- Évolution du genre Valserina Schroeder et al. 1968 et l'origine des Palorbitolines [Foraminiferida].- Paläontologische Zeitschrift, vol. 67, nos. 3-4, 245-251.
Schroeder R. & Cherchi A. (2002).- Hallazgo de Paracoskinolina pertenuis Foury (Orbitolinidae) en el Barremiense inferior del Pirineo catalán: Reflexiones sobre su posición sistemática y filogenética.- Treballs del Museu de Geologia de Barcelona, vol. 11, p. 97-103.
Schroeder R. & Charollais J. (1966).- Quatričme note sur les Foraminifčres du Crétacé inférieur de la région genevoise. Sur quelques Orbitolinidae des facičs urgoniens.- Archives des Sciences, Genčve, vol. 19, no. 1, p. 93-114.
Schroeder R. & Conrad M.A. (1968).- Huitičme note sur les Foraminifčres du Crétacé inférieur de la région genevoise. Eopalorbitolina charollaisi n.gen., n.sp., un Orbitolinidé nouveau du Barrémien ŕ facičs urgonien.- Compte rendu des Sciences de la Société de Physique et d'Histoire naturelle de Genčve (Nouvelle Série), [1967], p. 145-162.
Schroeder R. & Neumann M. (coords.) (1985).- Les grands Foraminifčres du Crétacé moyen de la région méditerranénne.- Geobios Mémoire Spécial, Villeurbanne, no. 7, 160 p.
Schroeder R., Altenbach A., Brinnel P., Cherchi A. & Wallrabe H.-J. (1982).- El Barremiense marino de la Sierra de Montsec (Prov. Lerida-Huesca).- Cuadernos Geología Ibérica, Madrid, vol. 8, p. 915-933.
Schroeder R., Charollais J. & Conrad M.A. (1969).- Neunter Beitrag über die Foraminiferen der Unterkreide der Gegend von Genf. Weitere Studien an Orbitoliniden des Urgons.- Archives des Sciences, Genčve, vol. 22, p. 91-104.
Schroeder R., Clavel B., Cherchi A. & Charollais J. (1999).- Praedictyorbitolina busnardoi n. sp. (Foraminiferida) et évolution de la lignée Praedicytorbitolina - Dictyorbitolina (Hauterivien supérieur - Barrémien basal).- Paläontologische Zeitschrift, vol. 73, nos. 3-4, p. 203-215.
Schroeder R., Clavel B., Cherchi A., Busnardo R., Charollais J. & Decrouez D. (2002).- Lignées phylétiques d'Orbitolinidés dans Hauterivien supérieur - Aptien inférieur du Sud-Ouest de l'Europe et leur importance stratigraphique.- Revue de Paléobiologie, Genčve, vol. 21, no. 2, p. 853-863.
Schroeder R., Clavel B., Conrad M.A., Zaninetti L., Busnardo R., Charollais J. & Cherchi A. (2000).- Corrélations biostratigraphiques entre la coupe d'Organyŕ (Pyrénées Catalanes, NE de l'Espagne) et le Sud-Est de la France pour l'intervalle Valanginian-Aptien.- Treballs del Museu de Geologia de Barcelona, vol. 9, p. 5-41.
Schroeder R., Conrad M.A. & Charollais J. (1967).- Sixičme note sur les Foraminifčres du Crétacé inférieur de la région genevoise. Contribution ŕ l'étude des Orbitolinidae: Valserina brönnimanni Schroeder & Conrad, n. gen., n. sp.; Paleodictyoconus barremianus (Moullade) et Paleodictyoconus cuvillieri (Foury).- Archives des Sciences, Genčve, vol. 20, no. 2, p. 199-222.
Schroeder R., Buchem F.S.P. van, Cherchi A., Baghbani D., Vincent B., Immenhauser A. & Granier B. (2010).- Revised orbitolinid biostratigraphic zonation for the Barremian - Aptian of the eastern Arabian Plate and implications for regional stratigraphic correlations.- GeoArabia Special Publication, Manama, vol. 4, no. 1, p. 49-96.
Schroeder R., Clavel B. & Charollais J. (1990).- Praedictyorbitolinina carthusiana n. gen., n. sp., Orbitolinidé (Foraminiferida) de la limite Hauterivien-Barrémien des Alpes occidentales.- Paläontologische Zeitschrift, vol. 64, no. 3-4, p. 193-202.
Schubert R. (1912).- Über Lituonella und Coskinolina liburnica Stache sowie deren Beziehungen zu den anderen Dictyoconinen.- Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, Vienna, vol. 62, p. 195-208.
Septfontaine M. (1988).- Vers une classification évolutive des Lituolidés (foraminifčres) jurassiques en milieu de plate-forme carbonatée.- Revue de Paléobiologie, Genčve, vol. spec. 2 (Benthos '86), p. 229-256.
Septfontaine M., Arnaud-Vanneau A., Bassoullet J-P., Gusič Y., Ramalho M.M. & Velić I. (1991).- Les foraminifčres imperforés des plates-formes carbonatées jurassiques : État des connaissances et perspectives d'avenir.- Bulletin de la Société vaudoise des Sciences naturelles, Lausanne, vol. 80, no. 3, p. 255-277.
Simpson G.G. (1944).- The tempo and mode in evolution.- Columbia University Press, New York, 217 p.
Strasser A., Charollais J., Conrad M.A., Clavel B., Pictet A. & Mastrangelo B. (2016).- The Cretaceous of the Swiss Jura Mountains: An improved lithostratigraphic scheme.- Swiss Journal of Geosciences, Basel, vol. 109, p. 201-220.
Sudar M., Jovanović D., Maran A. & Polavder S. (2008).- Late Barremian-Early Aptian Urgonian limestones from the south-eastern Kućaj Mountains (Carpatho-Balkanides, eastern Serbia).- Annales Géologiques de la Péninsule Balkanique, Belgrade, vol. 69, p. 13-30.
Ungureanu R., Săsăran E., Bucur I.I., Mircescu C.V., Ungur C.G. & Ungureanu A. (2017).- The Cretaceous conglomerates from Piatra Craiului syncline (South Carpathians, Romania): Searching for the source area.- Facies, Erlangen, vol. 63, no. 4, article 30, 30 p.
Virgone A. (1997, unpublished).- Stratigraphie, sédimentologie et dynamique d'une plate-forme carbonatée : Le Berriasien supérieur-Valanginien basal de Basse Provence occidentale (S.E. France).- PhD Thesis University of Provence Aix-Marseille, 196 p.
Yazdi-Moghadam M. & Schlagintweit F. (2021).- Cenomanian "orbitoliniform" foraminifera - State of the art and description of Ebrahimiella dercourti (Decrouez and Moullade, 1974) gen. et comb. nov. (family Coskinolinidae) from the Sarvak Formation (SW Iran, Zagros Zone).- Cretaceous Research, vol. 126, article 104885, 12 p.
Plate
1: |
|
|
Plate
2: |
|
|
Plate
3: |
|
|
Plate
4: |
|
|
Plate
5: |
|
|
Plate
6: |
|
|
Plate
7: |
|
|
Plate
8: |
|
|
Plate
9: |
|
|
Plate
10: |
|
|
Plate
11: |
|
|
Plate
12: |
|
|
Plate
13: |
|
|
Plate
14: |
|
| Nomenclatural note: Life Sciences Identifier (LSID) http://zoobank.org/References/E00D3DCB-77DD-4FF9-9692-826E174333D0 • Cribellopsis sudari Schlagintweit & Bucur, 2021 http://zoobank.org/NomenclaturalActs/40A5B99E-4DBC-4F4E-9328-AF3CE767CA85 |