◄ Carnets Geol. 21 (17) ►
![]()
Outline:
[1. Introduction and setting]
[2. materials and methods]
[3. Systematic ichnology] [4. Description of the substrate and the trace fossils]
[5. Remarks] [6. Concluding remarks]
and ... [Bibliographic references]
Department of Paleontology, Calvert Marine Museum, P.O. Box
97, Solomons, Maryland, 20688 (U.S.A.);
Research Associate, Department of Paleobiology, National Museum of Natural History, Smithsonian
Institution, Washington, DC, 20013-7012 (U.S.A.)
Department of Paleontology, Calvert Marine Museum, P.O. Box
97, Solomons, Maryland, 20688 (U.S.A.);
Current Address: 2421 N Utah St, Arlington, VA, 22207 (U.S.A.)
Published online in final form (pdf) on October 24, 2021
DOI 10.2110/carnets.2021.2117
![]()
[Editor: Alberto Collareta; technical editor: Bruno
R.C. Granier]
![]()
An isolated Miocene baleen whale left radius was marked repeatedly by shark bite-shake traces. The radius probably derives from the Plum Point Member of the Calvert Formation, Calvert Cliffs, Calvert County, Maryland, U.S.A. At least three successive bite-shake traces marking the radius, made by multiple teeth, are attributed to the trace fossil Linichnus bromleyi. These bite-shake traces consisting of shallow, thin arching gouges on a radius, likely indicate scavenging rather than active predation. The most likely means of producing the bundle of L. bromleyi within each of the three sets of traces would be through repeated biting as the shark re-positioned the prey in its mouth or, perhaps, by a shark species with multiple functional teeth within its tooth row. If the bite traces were produced by a non-serrated tooth (as they appear to have been), then the most likely candidate would be Carcharodon hastalis.
bite traces;
trophic interaction;
shark;
Mysticeti;
Miocene;
Calvert Cliffs;
Maryland
Godfrey S.J. & Lowry A.J. (2021).- The ichnospecies Linichnus bromleyi on a Miocene baleen whale radius preserving multiple shark bite-shake traces suggests scavenging.- Carnets Geol., Madrid, vol. 21, no. 17, p. 391-398.
L'ichnoespèce Linichnus bromleyi sur un radius de baleine à fanons miocène comportant de multiples traces de morsure-secouage de requin suggère son charognage.- Un radius gauche isolé de baleine à fanons miocène a enregistré à plusieurs reprises des traces de morsure-secouage de requin. Le radius provient probablement du Plum Point Member de la Calvert Formation des Calvert Cliffs (Comté de Calvert, Maryland, États-Unis d'Amérique). Au moins trois ensembles de morsure-secouage successifs marquant ce radius et provenant de plusieurs dents sont attribués à la trace fossile Linichnus bromleyi. Ces traces de morsure-secouage se composant de gouges peu profondes, fines et arquées sur le radius indiquent vraisemblablement du charognage plutôt qu'une prédation active. L'origine la plus probable de ce regroupement de L. bromleyi au sein de chacun de ces trois ensembles de traces serait par le biais de morsures répétées alors que le requin repositionne sa proie dans sa gueule ou, autre possibilité, de morsures d'une espèce de requin dotée de plusieurs dents fonctionnelles au sein même de sa rangée dentaire. Si les traces de morsure sont produites par des dents non-crantées (comme cela semble être le cas), alors le candidat le plus probable serait Carcharodon hastalis.
traces de morsures ;
interactions trophiques ;
requin ;
Mysticètes ;
Miocène ;
Calvert Cliffs;
Maryland
The Calvert Cliffs are one of the most fossiliferous regions on the east coast of the continental United States (Godfrey, 2018; Vogt et al., 2018). During part of the Miocene epoch (from approximately 22-8 Ma), the Chesapeake area (i.e., the Salisbury Embayment) was intermittently flooded by the Atlantic Ocean. The shallow marine sediments that now comprise the cliffs abound with marine fossils, including the tests of planktonic organisms, shelled invertebrates, shark teeth, teleosts, and marine mammals (Godfrey, 2018, and the references therein). In addition to innumerable body fossils, Calvert Cliffs preserve trace fossils including invertebrate burrows (Thalassinoides, Gyrolithes and Ophiomorpha Kidwell et al., 2015), coprolites (Wetmore, 1943; Godfrey & Smith, 2010; Kent, 2018; Weems, 2018; Godfrey et al., accepted), and shark bite traces, usually on cetacean bone (Godfrey, 2003; Godfrey et al., 2018).
There are now many examples of shark bite traces on bone, either from active predation or scavenging (Deméré & Cerutti, 1982; Cigala Fulgosi, 1990; Purdy, 1996; Schwimmer et al., 1997; Bianucci et al., 2000, 2009, 2010, 2018; Purdy et al., 2001; Hanks & Shimada, 2002; Renz, 2002; Godfrey, 2003; Corral et al., 2004; Godfrey & Altman, 2005; Noriega et al., 2007; Aguilera et al., 2008; Diedrich, 2008; Cicimurri & Knight, 2009; Ehret et al., 2009; Bianucci & Gingerich, 2011; Kallal et al., 2012; Govender & Chinsamy, 2013; Takakuwa, 2014; Govender, 2015, 2021; Carrillo-Briceño et al., 2016; Collareta et al., 2017, 2019; Godfrey et al., 2018, 2021; Kent, 2018; Cortés et al., 2019; Mierzwiak & Godfrey, 2019; Muñiz et al., 2020; Bosio et al., 2021; Feichtinger et al., 2021; Perez et al., accepted). The authors of some of these case studies have been able to determine the species of shark that left the bite trace, either by finding shark teeth imbedded in the fossil bone, or by comparing the morphology of the tooth traces to shark teeth found in the same formation (Everhart et al., 1995; Schwimmer et al., 1997; Shimada, 1997; Shimada et al., 2002; Corral et al., 2004; Shimada & Everhart, 2004; Shimada & Hooks, 2004; Rothschild et al., 2005; Noriega et al., 2007; Bianucci et al., 2009; Cicimurri & Knight, 2009; Ehret et al., 2009; Govender & Chinsamy, 2013; Govender, 2015; Godfrey et al., 2018, 2021; Kent, 2018; Cortés et al., 2019; Feichtinger et al., 2021).
Location and nature of bite traces can indicate the context of the interaction, as deep bites on bones near vital organs might be indicative of predation by a large shark (Kent, 2018). In contrast, small, thin tooth traces on appendages suggest scavenging, as smaller sharks will feed leisurely on a whale carcass at the water line (Dudley et al., 2000; Govender & Chinsamy, 2013; Bisconti et al., 2020).
Here we describe an isolated Miocene baleen whale radius from Calvert Cliffs that preserves signs of a unique trophic interaction with a single shark. The bite traces are exceptional in that they preserve three successive shark bite-shake behaviors that left arching traces on both the dorsal (external) and ventral (internal) sides of the bone. Although the species of the shark and whale are yet undetermined, these bite traces are superb examples of the recently named trace fossil Linichnus bromleyi (Muñiz et al., 2020) and worthy of being described here.
CMM-V = the fossil vertebrate collection at the Calvert Marine Museum in Solomons, Maryland, U.S.A.
USNM PAL, Department of Paleobiology collections, National Museum of Natural History, Smithsonian Institution, Washington, DC, U.S.A.
To measure the depth and width of the shark tooth traces on the radius (CMM-V-3979), a silicone rubber mold was prepared of both upper and lower sides of the bone. Depth and width of the bite traces were measured from the positive mold using digital calipers.
In
its natural state, CMM-V-3979 is very dark, so to improve contrast and
visibility in Figure 1 ![]() , the bone was
lightly dusted with sublimed ammonium chloride, i.e., a whitening
technique described by Cooper (1935) and Feldman
(1989). After the specimen was
photographed with a Nikon CoolPix P510 camera under fluorescent light, the
ammonium chloride was removed by holding CMM-V-3979 under running water (Shelburne & Thompson,
2016).
, the bone was
lightly dusted with sublimed ammonium chloride, i.e., a whitening
technique described by Cooper (1935) and Feldman
(1989). After the specimen was
photographed with a Nikon CoolPix P510 camera under fluorescent light, the
ammonium chloride was removed by holding CMM-V-3979 under running water (Shelburne & Thompson,
2016).
Linichnus Jacobsen & Bromley, 2009
Linichnus bromleyi Muñiz et al., 2020
Horizon
and locality: CMM-V-3979, Figure 1 ![]() , was collected by William (Douggie) Douglass, as float on the
beach south of Parkers Creek along Calvert Cliffs in Calvert County, Maryland,
U.S.A. When found, the specimen was devoid of entombing sediment. Even though it
was not found in situ, there is no
reason to think that it was not locally derived from the adjacent cliffs at
Warrior's Rest Wildlife Sanctuary. At the point where CMM-V-3979 was found,
Shattuck Zones 11-19 are exposed, which includes the upper portion of the Plum
Point Member (Shattuck Zones 11-16A) of the Calvert Formation along with the
Drumcliff (Shattuck Zones 16B-17), St. Leonard (Shattuck Zone 18), and Boston
Cliff (Shattuck Zone 19) members of the Choptank Formation (Kidwell
et al., 2015). The sediments
comprising this section of the cliffs range in age from approximately 16.5-12.5
Ma (Perez et al., 2019, Fig.
1).
, was collected by William (Douggie) Douglass, as float on the
beach south of Parkers Creek along Calvert Cliffs in Calvert County, Maryland,
U.S.A. When found, the specimen was devoid of entombing sediment. Even though it
was not found in situ, there is no
reason to think that it was not locally derived from the adjacent cliffs at
Warrior's Rest Wildlife Sanctuary. At the point where CMM-V-3979 was found,
Shattuck Zones 11-19 are exposed, which includes the upper portion of the Plum
Point Member (Shattuck Zones 11-16A) of the Calvert Formation along with the
Drumcliff (Shattuck Zones 16B-17), St. Leonard (Shattuck Zone 18), and Boston
Cliff (Shattuck Zone 19) members of the Choptank Formation (Kidwell
et al., 2015). The sediments
comprising this section of the cliffs range in age from approximately 16.5-12.5
Ma (Perez et al., 2019, Fig.
1).
|
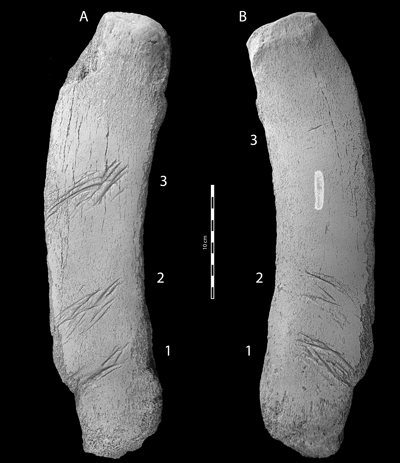
Figure 1:
CMM-V-3979, a Miocene baleen whale radius preserving several specimens of the
ichnospecies Linichnus bromleyi. A.
Dorsal (external) view. B. Ventral
(internal) view. Top of the image is proximal. Numbers 1-3 mark the locations
where the shark teeth raked the surface of the radius. Numbers 1-1, 2-2, 3-3
correspond to bite traces made on the radius by teeth in opposing jaws. To
improve contrast and highlight detail, CMM-V-3979 was whitened with sublimed
ammonium chloride. |
From its size and gently curving shape, CMM-V-3979 can only be a
mysticete radius (Fig. 1 ![]() ). CMM-V-3979
measures 275 mm in length and 68 mm at its widest point (i.e., at approximately
53 mm along the shaft of the bone from its proximal articular surface). The
maximum dorsoventral thickness of the bone is 45 mm through the proximal
articular condyle. Both epiphyses are fully fused to the diaphysis and articular
surfaces are well defined, indicating that the radius came from a mature
individual. The proximal condyle is gently concave anteriorly. The distal
condyle is bifurcated into two concave surfaces that articulated with the
carpals. It is not known what damaged both the proximal and distal ends of the
bone; if this was the result of shark-feeding damage or modern damage from the
time the bone fell out of the cliffs until it was found. Furthermore, the
anterior and posterior faces of the bone are also missing the outer smooth
compact cortical bony layer.
). CMM-V-3979
measures 275 mm in length and 68 mm at its widest point (i.e., at approximately
53 mm along the shaft of the bone from its proximal articular surface). The
maximum dorsoventral thickness of the bone is 45 mm through the proximal
articular condyle. Both epiphyses are fully fused to the diaphysis and articular
surfaces are well defined, indicating that the radius came from a mature
individual. The proximal condyle is gently concave anteriorly. The distal
condyle is bifurcated into two concave surfaces that articulated with the
carpals. It is not known what damaged both the proximal and distal ends of the
bone; if this was the result of shark-feeding damage or modern damage from the
time the bone fell out of the cliffs until it was found. Furthermore, the
anterior and posterior faces of the bone are also missing the outer smooth
compact cortical bony layer.
Both
dorsal (external) and ventral (internal) surfaces show three repeating patterns
of bite-shake traces lying at nearly right angles to the curved longitudinal
axis of the bone (Fig. 1 ![]() ). The bite traces reach a maximum depth of 1.47 mm (although
most are less than 1 mm deep) and a maximum width of 1.33 mm (although these measurements did not coincide
on the same trace).
No serration marks were observed along the edges of the gouges, but because only
the tips of the teeth cut into the bone, it is not surprising that serration
traces are absent (if the originating teeth were serrated at all). Furthermore,
the cut bone is coarsely textured, further obscuring any possibility of
preserving serration traces.
). The bite traces reach a maximum depth of 1.47 mm (although
most are less than 1 mm deep) and a maximum width of 1.33 mm (although these measurements did not coincide
on the same trace).
No serration marks were observed along the edges of the gouges, but because only
the tips of the teeth cut into the bone, it is not surprising that serration
traces are absent (if the originating teeth were serrated at all). Furthermore,
the cut bone is coarsely textured, further obscuring any possibility of
preserving serration traces.
There are a number of basal thalassotherians (Mysticeti) known from the Calvert and Choptank formations along Calvert Cliffs for which their radius is also known. The radius of the Calvert Formation mysticete Pelocetus calvertensis (USNM PAL 11976) is 397 mm long (Kellogg, 1965), 122 mm longer than CMM-V-3979. The radius in "Eschrichtius" pusillus was even longer than in Pelocetus (Kellogg, 1968). The radius of the Choptank Formation mysticete Thinocetus arthritis (USNM PAL 23794) is 385 mm long (Kellogg, 1969), 110 mm longer than CMM-V-3979, and it is not even from a fully mature individual. The best match is an isolated radius (USNM PAL 23019, Kellogg, 1968, Fig. 73), questionably assigned to the Calvert Formation mysticete, Diorocetus hiatus, which is exactly the same length as the radius of CMM-V-3979. Although they are not a perfect match morphologically, they are very close. Both the type specimen of Diorocetus (USNM PAL 16738) and CMM-V-3979 were found just south of Parkers Creek, along Calvert Cliffs.
From the sharks that are known from along Calvert Cliffs (Visaggi & Godfrey, 2010; Kent, 2018), we think that the bite traces on CMM-V-3979 could have been made by any one of the following sharks (listed here alphabetically: Alopias grandis, Alopias palatasi, Carcharhinus spp., Carcharodon hastalis, Galeocerdo aduncus, Hemipristis serra, a juvenile Otodus megalodon, Physogaleus contortus, or Sphyrna laevissima. The most likely means of producing the bundle of L. bromleyi specimens within each of the three sets of traces would be through repeated biting as the shark re-positioned the prey in its mouth or, perhaps, by a shark species with multiple functional teeth within their tooth row. A similar interpretation in a different situation was discussed by Bianucci et al. (2018, Fig. 13B-E & G). If the bite traces were produced by a non-serrated tooth (as they appear to have been), then the most likely candidate would be Carcharodon hastalis (V. Perez, pers. comm. July 26, 2021).
|
Figure 2:
One possible view of the origin of the bitten Miocene whale radius. The whale
could also have been bloated and floating belly up at the time the shark bite
traces were made. Furthermore, the distal limb elements below the radius and
ulna may already have been missing and/or eaten. Original artwork by Tim Scheirer
and Clarence (Shoe) Shumaker. © Calvert Marine Museum. |
When a whale dies, it inverts and floats at the surface of the water due to the buildup of abdominal gases from decomposition. Scavenging sharks habitually feed at the water line, occasionally lifting their heads out of the water (Dudley et al., 2000; Tucker et al., 2019). The flippers of the whale lie at the water line and extend outward from the body, providing an easy target for scavengers. Indeed, of previous literature documenting shark-bitten cetacean flippers, all posit scavenging as the context of feeding (Noriega et al., 2007; Cicimurri & Knight, 2009; Govender & Chinsamy, 2013). The shark tooth markings on Pliocene balaenopterid flippers bones from Panama described by Cortés et al. (2019, Fig. 3) are here attributed to Linichnus bromleyi.
From the similarity of the three bite traces on the dorsal side of the radius, we think that they were made by the same teeth during successive bite-shake events. Teeth in the same position in different files (rows of teeth) of the shark's mouth marked both the upper and lower surface of the radius. The shark would have clenched down on the flipper firmly and then shaken its head vigorously in an attempt to cut through the bone (unsuccessfully) or to simply remove flesh.
The existence of aligned bite-shake traces on both the upper and lower surface of the radius is due to the shark teeth marking the bone with upper and lower teeth simultaneously. It is not known which teeth in the dental arcade created the bite traces. Neither do we know if the whale was inverted during the scavenging event.
Furthermore, we do not know
the order in which the three bite traces were made, distal to proximal - proximal to distal
- or some other order. We assume distal to proximal because
the most distal bite (number 1 in Fig. 1 ![]() ) would likely have resulted in the
removal of some flesh covering the flipper. Following that, the shark might have
re-bitten the radius at site 2, shaken its head and successfully removed more
flesh before proceeding to biting and head shaking at site 3. Notice that for
site number three, there are fewer bite traces on the ventral surface of the
radius. Presumably, those teeth only just barely penetrated the soft tissue to
contact the bone. Notice also that those bite traces are not aligned as in bite
traces 1 and 2. Perhaps those shallow grooves on the ventral (internal) surface
of the radius represent markings made during another head-thrashing event.
) would likely have resulted in the
removal of some flesh covering the flipper. Following that, the shark might have
re-bitten the radius at site 2, shaken its head and successfully removed more
flesh before proceeding to biting and head shaking at site 3. Notice that for
site number three, there are fewer bite traces on the ventral surface of the
radius. Presumably, those teeth only just barely penetrated the soft tissue to
contact the bone. Notice also that those bite traces are not aligned as in bite
traces 1 and 2. Perhaps those shallow grooves on the ventral (internal) surface
of the radius represent markings made during another head-thrashing event.
There is no evidence of healing (i.e., no bony callus formations as a result of the originating trauma). The whale (if it was alive at the time of the trophic encounter, which is doubtful) did not survive the biting by the shark.
Amongst extant sharks, head thrashing is employed when removing chunks from a large food source (Curtis et al., 2006). Apparently, tiger sharks use a method of head movement much like head shaking during feeding bouts; however, movement is a slower side-to-side motion (Curtis et al., 2006). Saw-biting is the term coined by Lea et al. (2018) to describe this behavior. It seems to be specific to tiger sharks whose serrated teeth have evolved to saw through prey too large to swallow whole (Frazzetta, 1988; Curtis et al., 2006).
Linichnus bromleyi describes vertebrate tooth bite traces raking the surface of biogenic material, in straight lines or arching curves. Head shaking/thrashing, or saw-biting behavior would account for the formation of this kind of trace fossil. Linichnus bromleyi can occur either as a result of exploration, predation, or scavenging. However, in CMM-V-3979, the shallow thin arching gouges on the radius indicate scavenging rather than active predation.
We would like to thank William
(Douggie) Douglass for having collected the radius described herein, and
for having donated it to the Calvert Marine Museum. CMM artists Tim Scheirer
and Clarence (Shoe) Shumaker very skillfully created Figure 2 ![]() . CMM
Paleontology Collections Manager, John R. Nance is also gratefully
acknowledged for encouraging the study of specimens under his care. Victor Perez
(CMM) graciously reviewed an early version of this paper. We are very grateful
for the constructive reviews by Bretton W. Kent and one anonymous reviewer. Alberto Collareta
edited this paper for the journal, many thanks! This research was funded in part
by the citizens of Calvert County Maryland, the County Board of Calvert County
Commissioners, and the Clarissa and Lincoln Dryden Endowment for
paleontology at the Calvert Marine Museum.
. CMM
Paleontology Collections Manager, John R. Nance is also gratefully
acknowledged for encouraging the study of specimens under his care. Victor Perez
(CMM) graciously reviewed an early version of this paper. We are very grateful
for the constructive reviews by Bretton W. Kent and one anonymous reviewer. Alberto Collareta
edited this paper for the journal, many thanks! This research was funded in part
by the citizens of Calvert County Maryland, the County Board of Calvert County
Commissioners, and the Clarissa and Lincoln Dryden Endowment for
paleontology at the Calvert Marine Museum.
Aguilera O.A., García L. & Cozzuol M.A. (2008).- Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation.- Paläontologische Zeitschrift, Stuttgart, vol. 82, p. 204-208.
Bianucci G., Bisconti M., Landini W., Storai T., Zuffa M., Giuliani S. & Mojetta A. (2000).- Trophic interactions between white sharks (Carcharodon carcharias) and cetaceans: A comparison between Pliocene and recent data. In: Vacchi M., La Mesa G., Serena F. & Séret B. (eds.), Proceedings of the 4th Meeting of the European Elasmobranch Association, Livorno (Italy), 27-30 September 2000.- European Elasmobranch Association, imprimerie F. Paillart, Abbeville, p. 33-48.
Bianucci G., Collareta A., Bosio G., Landini W., Gariboldi K., Gioncada A., Lambert O., Malinverno E., Muizon C. de, Varas-Malca R., Villa I.M, Coletti G., Urbina M. & Di Celma C.- (2018).- Taphonomy and palaeoecology of the lower Miocene marine vertebrate assemblage of Ullujaya (Chilcatay Formation, East Pisco Basin, southern Peru).- Palæogeography, Palæoclimatology, Palæoecology, vol. 511, p. 256-279.
Bianucci G. & Gingerich P.D. (2011).- Aegyptocetus tarfa, n. gen. et sp. (Mammalia, Cetacea), from the middle Eocene of Egypt: Clinorhynchy, olfaction, and hearing in a protocetid whale.- Journal of Vertebrate Paleontology, Lawrence - KS, vol. 31, p. 1173-1188.
Bianucci G., Sorce B., Storai T. & Landini W. (2010).- Killing in the Pliocene: Shark attack on a dolphin from Italy.- Palaeontology, London, vol. 5, p. 457-470.
Bianucci G., Vaiani S.C. & Casati S. (2009).- A new delphinid record (Odontoceti, Cetacea) from the Early Pliocene of Tuscany (Central Italy): Systematics and biostratigraphical considerations.- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 254, p. 275-292.
Bisconti M., Damarco P., Pavia M., Sorce B. & Carnevale G. (2020).- Marzanoptera tersillae, a new balaenopterid genus and species from the Pliocene of Piedmont, north-west Italy.- Zoological Journal of the Linnean Society, London, vol. 20, p. 1-40.
Bosio G., Collareta A., Di Celma C., Lambert O., Marx F.G., Muizon C. de, Gioncada A., Gariboldi K, Malinverno E., Varas-Malca R., Urbina M. & Bianucci G. (2021).- Taphonomy of marine vertebrates of the Pisco Formation (Miocene, Peru): Insights into the origin of an outstanding Fossil-Lagerstätte.- Plos One, vol. 16, no. 7, e0254395, 53 p. DOI: https://doi.org/10.1371/journal.pone.0254395
Carrillo-Briceño J.D., Aguilera O.A., De Gracia C., Aguirre-Fernández G., Kindlimann R. & Sánchez-Villagra M.R. (2016).- An early Neogene elasmobranch fauna from the southern Caribbean (western Venezuela).- Palaeontologia Electronica, vol. 19, no. 2, 32 p. DOI: https://doi.org/10.26879/664
Cicimurri D.J. & Knight J.L. (2009).- Two shark-bitten whale skeletons from Coastal Plain deposits of South Carolina.- Southeastern Naturalist, Steuben - ME, vol. 8, no. 1, p. 71-82.
Cigala Fulgosi F. (1990).- Predation (or possible scavenging) by a great white shark on an extinct species of bottlenosed dolphin in the Italian Pliocene.- Tertiary Research, Orpington, Kent, vol. 12, p. 17-36.
Collareta A., Fulgosi F.C. & Bianucci G. (2019).- A new kogiid sperm whale from northern Italy supports psychrospheric conditions in the early Pliocene Mediterranean Sea.- Acta Palaeontologica Polonica, Warsaw, vol. 64, no. 3, p. 609-626.
Collareta A., Lambert O., Landini W., Di Celma C., Malinverno E., Varas-Malca R., Urbina M. & Bianucci G. (2017).- Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru.- Palæogeography, Palæoclimatology, Palæoecology, vol. 469, p. 84-91.
Cortés D., De Gracia C., Carrillo-Briceño J.D., Aguirre-Fernández G., Jaramillo C., Benites-Palomino A. & Atencio-Araúz J.E. (2019).- Shark-cetacean trophic interactions during the late Pliocene in the Central Eastern Pacific (Panama).Palaeontologia Electronica, vol. 22.2.49A, 13 p. DOI: https://doi.org/10.26879/953
Cooper C.L. (1935).- Ammonium chloride sublimate apparatus.- Journal of Paleontology, Cambridge, vol. 9, no. 4, p. 357-359.
Corral J.C., Pereda Suberbiola X. & Bardet N. (2004).- Shark-bite marks in a mosasaur vertebra from the Late Cretaceaous of Álava (Basque-Cantabrian Region).- Revista Española de Paleontología, Madrid, vol. 19, no. 1, p. 23-32.
Curtis T.H., Kelly J.T. Menard K.L., Laroche R.K., Jones R.E. & Klimley A.P. (2006).- Observations on the behavior of white sharks scavenging from a whale carcass at Point Reyes, California.- California Fish and Game, Sacramento - CA, vol. 92, no. 3, p. 113-124.
Deméré T.A. & Cerutti R.A. (1982).- A Pliocene shark attack on a cethotheriid whale.- Journal of Paleontology, Cambridge, vol. 56, p. 1480-1482.
Diedrich C. G. (2008).- The food of the miosiren Anomotherium langenwieschei (Siegfried)indirect proof of seaweed or seagrass by xenomorphic oyster fixation structures in the Upper Oligocene (Neogene) of the Doberg, Bünde (NW Germany) and comparisons to modern Dugong dugon (Müller) feeding strategies.- Senckenbergiana Maritima, Frankfurt am Main, vol. 38, no. 1, p. 59-73.
Dudley S.F.J., Anderson-Reade M.D., Thompson G.S. & McMullen P.B. (2000).- Concurrent scavenging off a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier.- Fishery Bulletin, Seattle - WA, vol. 98, p. 646-649.
Ehret D.J., MacFadden B.J., Jones D.S., DeVries T.J. & Salas-Gismondi R. (2009).- Caught in the act: Trophic interactions between a 4-million-year-old white shark (Carcharodon) and mysticete whale from Peru.- Palaios, Lawrence - KS, vol. 24, p. 329-333.
Everhart M.J., Everhart P.A. & Shimada K. (1995).- A new specimen of shark bitten mosasaur vertebrae from the Smoky Hill Chalk (Upper Cretaceous) in western Kansas.- Abstracts, 127th Annual Meeting, Kansas Academy of Science, Pittsburg - KS, 14, p. 19.
Feichtinger I., Fritz I. & Göhlich U.B. (2021).- Tiger shark feeding on sirenian - first fossil evidence from the middle Miocene of the Styrian Basin (Austria).- Historical Biology, no numbering, 8 p. DOI: https://doi.org/10.1080/08912963.2021.1906665
Feldman R.M. (1989).- Whitening fossils for photographic purposes. In: Feldman R.M. (ed.), Paleotechniques.- The Paleontological Society Special Publication, Cambridge, vol. 4, p. 342-346.
Frazzetta T. (1988).- The mechanics of cutting and the form of shark teeth (Chondrichthyes, Elasmobranchii).- Zoomorphology, Berlin, vol. 108, no. 2, p. 93-107.
Godfrey S.J. (2003).- Miocene sharks and smoking guns - Bugeye Times (Quarterly Newsletter of the Calvert Marine Museum), Solomons Island - MD, vol. 28, no. 1, p. 1, 5-7.
Godfrey S.J. (ed.). (2018).- The geology and Vertebrate paleontology of Calvert Cliffs.- Smithsonian Contributions to Paleobiology, Washington, D.C., vol. 100, 265 p. DOI: https://doi.org/10.5479/si.1943-6688.100
Godfrey S.J. & Altman, J. (2005).- A Miocene cetacean vertebra showing a partially healed compression fracture, the result of convulsions or failed predation by the Giant White Shark, Carcharodon megalodon.- Jeffersoniana, Martinsville - VA, vol. 16, p. 1-12.
Godfrey S.J., Collareta A. & Nance J.R. (accepted).- Coprolites from Calvert Cliffs: Miocene fecal pellets and burrowed crocodilian droppings from the Chesapeake Group of Maryland, U.S.A.- Rivista Italiana di Paleontologia e Stratigrafia, Milano.
Godfrey S.J., Ellwood M., Groff S. & Verdin M.S. (2018).- Carcharocles-bitten odontocete caudal vertebrae from the Coastal Eastern United States.- Acta Palaeontologica Polonica, Warsaw, vol. 63, no. 3, p. 463-468.
Godfrey S.J., Nance J.R. & Riker N.L. (2021).- Otodus-bitten sperm whale tooth from the Coastal Eastern United States.- Acta Palaeontologica Polonica, Warsaw, vol. 66, 5 p. DOI: https://doi.org/10.4202/app.00820.2020
Godfrey S.J. & Smith J. (2010).- Shark-bitten vertebrate coprolites from the Miocene of Maryland.- Naturwissenschaften, Cham, vol. 97, no. 5, p. 461-467.
Govender R. (2015).- Shark-cetacean trophic interaction, Duinefontein, Koeberg, (5 Ma), South Africa.- South African Journal of Science, Pretoria, vol. 111, no. 11/12, art. #2014-0453, 7 p. DOI: https://doi.org/10.17159/ sajs.2015/20140453
Govender, R. (2021).- Shark-seal interaction off South Africa's west coast during the early Pliocene (5 Ma).- South African Journal of Science, Pretoria, vol. 117, no. 3/4, p. 123-129. DOI: https://doi.org/10.17159/sajs.2021/8837
Govender R. & Chinsamy A. (2013).- Early Pliocene (5 MA) shark-cetacean trophic interaction from Langebaanweg, Western Coast of South Africa.- Palaios, Lawrence - KS, vol. 28, no. 5, p. 270-277.
Hanks H.D. & Shimada K. (2002).- Vertebrate fossils, including non-avian dinosaur remains and the first shark-bitten bird bone, from a Late Cretaceous (Turonian) marine deposit of northeastern South Dakota.- Journal of Vertebrate Paleontology, vol. 22 (Supplement to Number 3), p. 62A.
Kallal R.J., Godfrey S.J. & Ortner D.J. (2012).- Bone reactions on a Pliocene cetacean rib indicate short-term survival of predation event.- International Journal of Osteoarchaeology, Hoboken - NJ, vol. 22, p. 253-260.
Kellogg R. (1965).- Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 1. A new whalebone whale from the Miocene Calvert Formation.- United States National Museum Bulletin, Washington, D.C., vol. 247, p. 1-45.
Kellogg R. (1968).- Fossil marine mammals from the Miocene Calvert Formation of Maryland and Virginia. Part 8. Supplement to description of Parietobalaena palmeri.- United States National Museum Bulletin, Washington, D.C., vol. 247, p. 175-197.
Kellogg R. (1969).- Cetothere skeletons from the Miocene Choptank Formation of Maryland and Virginia.- United States National Museum Bulletin, Washington, D.C., vol. 294, p. 1-40.
Kent B.W. (2018).- 2. The cartilaginous fishes (chimaeras, sharks and rays) of Calvert Cliffs, Maryland, USA. In: Godfrey S.J. (ed.), The geology and Vertebrate paleontology of Calvert Cliffs.- Smithsonian Contributions to Paleobiology, Washington, D.C., vol. 100, p. 45-160.
Kidwell S.M., Powars D.S., Edwards L.E., & Vogt P.R. (2015).- Miocene stratigraphy and paleoenvironments of the Calvert Cliffs, Maryland.- The Geological Society of America Field Guide, Boulder - CO, vol. 40, p. 231-279.
Lea J., Daly R., Leon C., Daly C. & Clarke C. (2018).- Life after death: Behaviour of multiple shark species scavenging a whale carcass.- Marine and Freshwater Research, Clayton, vol. 70, no. 2, p. 302-306.
Mierzwiak J.S. & Godfrey S.J. (2019).- Megalodon-bitten whale rib from South Carolina.- The Ecphora, Solomons, vol. 34, no. 2, p. 15-20.
Mikulá R., Kadlecová E., Fejfar O. & Dvořák Z. (2006).- Three new ichnogenera of biting and gnawing traces on reptilian and mammalian bones: A case study from the Miocene of the Czech Republic.- Ichnos, London, vol. 13, no. 3, p. 113-127.
Muñiz F., Belaústegui Z., Toscano A., Ramirez-Cruzado S. & Gámez Vintaned J.A. (2020).- New ichnospecies of Linichnus Jacobsen & Bromley, 2009.- Ichnos, vol. 27, no. 3, p. 344-351.
Noriega J.I., Cione A.L. & Aceñolaza F.G. (2007).- Shark tooth marks on Miocene balaenopterid cetacean bones from Argentina.- Neues Jahrbuch für Geologie und Paläontologie Abh., Stuttgart, vol. 245, p. 185-192.
Perez V.J., Godfrey S.J., & Chapman P.F. (accepted).- Rare evidence of shark-on-shark trophic interactions in the fossil record.- Acta Palaeontologica Polonica, Warsaw.
Perez V.J., Godfrey S.J., Kent B.W., Weems R.E. & Nance J.R. (2019).- The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): Lateral cusplet loss through time.- Journal of Vertebrate Paleontology, Abingdon, vol. 38, no. 6, article e1546732, 14 p.
Purdy R.W. (1996).- Paleoecology of fossil white sharks. In: Klimley A.P. & Ainley D.G. (eds.), Chapter 8. Great white sharks: The biology of Carcharodon carcharias.- Academic Press, San Diego, p. 67-78.
Purdy R.W., Schneider V.P., Applegate S.P., McLellan J.H., Meyer R.L. & Slaughter B.H. (2001).- The Neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. In: Ray C.E. & Bohaska D.J. (eds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, III.- Smithsonian Contributions to Paleobiology, Washington, D.C., vol. 90, p. 71-202.
Rothschild B.M., Martin L.D. & Schulp A.S. (2005).- Sharks eating mosasaurs, dead or alive?- Netherlands Journal of Geosciences, Cambridge, vol. 84, no. 3, p. 335-340.
Renz M. (2002).- Megalodon, hunting the hunter.- PaleoPress, Lehigh Acres - FL, 159 p.
Schwimmer D.R., Stewart J.D. & Williams G.D. (1997).- Scavenging by sharks of the genus Squalicorax in the late Cretaceous of North America.- Palaios, Lawrence - KS, vol. 12, p. 71-83.
Shelburne E.C.H. & Thompson A.C. (2016).- Specimen whitening: An assessment of methods of ammonium chloride smoke removal.- Collection Forum, Lawrence - KS, vol. 30, no. 1, p. 63-72.
Shimada K. (1997).- Dentition of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli, from the Niobrara Chalk of Kansas.- Journal of Vertebrate Paleontology, Abingdon, vol. 17, no. 2, p. 269-279.
Shimada K. & Everhart M.J. (2004).- Shark-bitten Xiphactinus audax (Teleostei: Ichthyodectiformes) from the Niobrara Chalk (Upper Cretaceous) of Kansas.- The Mosasaur, Plymouth Meeting, vol. 7, p. 35-39.
Shimada K., Everhart M.J. & Hooks G.E. III (2002).- Ichthyodectid fish and protostegid turtle bitten by the Late Cretaceous lamniform shark, Cretoxyrhina mantelli.- Journal of Vertebrate Paleontology, Abingdon, vol. 22 (Supplement to Number 3), p. 106A.
Shimada K. & Hooks G.E. III (2004).- Shark-bitten protostegid turtles from the Upper Cretaceous Mooreville Chalk, Alabama.- Journal of Paleontology, Cambridge, vol. 7, p. 205-210.
Takakuwa Y. (2014).- A dense occurrence of teeth of fossil "mako" shark ("Isurus" hastalis: Chondrichthyes, Lamniformes), associated with a balaenopterid-whale skeleton of the Late Miocene Pisco Formation, Peru, South America.- Bulletin of the Gunma Museum of Natural History, Tucker, vol. 18, p. 77-86.
Tucker J.P., Vercoe B., Santos I.R., Dujmovic M. & Butcher P.A. (2019).- Whale carcass scavenging by sharks.- Global Ecology and Conservation, vol. 19, article e00655, 11 p.
Visaggi C.C. & Godfrey S.J. (2010).- Variation in composition and abundance of Miocene shark teeth from Calvert Cliffs, Maryland.- Journal of Vertebrate Paleontology, Abingdon, vol. 30, p. 26-35.
Vogt P.R., Eshelman R.E. & Godfrey S.J. (2018).- 1. Calvert cliffs: Eroding mural escarpment, fossil dispensary, and paleoenvironmental archive in space and time. In: Godfrey S.J. (ed.), The geology and Vertebrate paleontology of Calvert Cliffs.- Smithsonian Contributions to Paleobiology, Washington, D.C., vol. 100, p. 3-44.
Weems R.E. (2018).- 4. Crocodilians of the Calvert Cliffs. In: Godfrey S.J. (ed.), The geology and Vertebrate paleontology of Calvert Cliffs.- Smithsonian Contributions to Paleobiology, Washington, D.C., vol. 100, p. 213-240.
Wetmore A. (1943).- The occurrence of feather impressions in the Miocene deposits of Maryland.- The Auk, Boston - MA, vol. 60, p. 440-441.