◄ Carnets Geol. 22 (2) ►
![]()
Outline:
[1. Introduction]
[2. Roloboceratinae as biostratigraphic index]
[3. Systematic study]
[4. Discussion] [5. Conclusions]
[Bibliographic references] and ... [Plates]
284
Chemin du Pestrier, 06670 Levens (France);
Centre d'Études Méditerranéennes, c/o
Madoux E., Rue Haute, 04330 Barrême (France)
16, rue Frédéric Mistral, 26200 Montélimar (France);
Centre d'Études Méditerranéennes, c/o Madoux E., Rue Haute, 04330 Barrême (France)
Musée Cantonal de Géologie, Quartier
UNIL-Dorigny, Bâtiment Anthropole, 1015 Lausanne (Switzerland)
Instituto de
Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria,
Coyoacán, 04510, Ciudad de Mexico (Mexico)
Groupement d'Intérêt Paléontologique, Science et Exposition, 60 bd Georges
Richard, 83000 Toulon (France)
1ter rue du Pont, 10450 Bréviandes (France);
Université de Bourgogne-Franche-Comté, UMR 6282, CNRS, Biogéosciences,
6 boulevard Gabriel, 21000 Dijon (France)
Published online in final form (pdf) on February 17, 2022
DOI 10.2110/carnets.2022.2202
![]()
[Editor: Bruno
R.C. Granier; language editor: Nigel D. Chapman]
![]()
The Lower Aptian marine deposits from southern Ardèche (South-East of France) yield a rich ammonite fauna of the sub-family Roloboceratinae Casey, 1961, which dates the upper Deshayesites forbesi Zone (Roloboceras hambrovi Subzone pro parte ?). The systematic study of this Roloboceratinae fauna has improved our knowledge of Megatyloceras coronatum Rouchadzé, 1933, type species of Megatyloceras Humphrey, 1949, as well as a better understanding of the genus thanks to the examination of the species from the Anglo-Paris Basin. A new species, M. leteilensis sp. nov. is introduced, and a description of M. transiens Casey, 1961, provided, with clarifications on its possible stratigraphic position. This study also establishes that the "species" Megatyloceras ricordeanum (Orbigny, 1850) represents only a growth stage shared by several species of Megatyloceras which is doubtfully a true taxon and must, therefore, be considered as a nomen dubium. The hypothesis of sexual dimorphism in Roloboceras Casey, 1954, and Megatyloceras is advanced. The study of this Roloboceratinae fauna highlights their heterogeneous distribution throughout Lower Aptian deposits of Ardèche, showing a direct correlation with bathymetric range. The biostratigraphical distribution of both genera on the Languedoc platform shows that Roloboceras is present from the uppermost C2 segment to the C4 segment of the stable carbon-isotope record, while Megatyloceras is limited to the middle C4 segment up to the lowermost C6 segment. Both genera coexist during the C4 segment.
• Ammonoidea;
• Roloboceratinae;
• Lower Cretaceous;
• Lower Aptian;
• Ardèche;
• South-East France;
• Paris Basin
Delanoy G., Baudouin C., Pictet A., Moreno-Bedmar J., Frau C. & Matrion B. (2022).- The genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949 (Ammonoidea, Ancyloceratina, Douvilleiceratidae), from the Lower Aptian of Ardèche (SE France) - Taxonomic and biostratigraphic implications.- Carnets Geol., Madrid, vol. 22, no. 2, p. 7-109.
Les genres Roloboceras Casey, 1954, et Megatyloceras Humphrey, 1949 (Ammonoidea, Ancyloceratina, Douvilleiceratidae), dans l'Aptien inférieur de l'Ardèche (Sud-Est de la France) - Conséquences taxinomiques et biostratigraphiques.- Les dépôts marins de l'Aptien inférieur (zone à Deshayesites forbesi) de la bordure ardéchoise (Sud-Est de la France) ont livré une riche faune d'ammonites de la sous-famille des Roloboceratinae Casey, 1961, qui date ces dépôts de la Zone à Deshayesites forbesi (Sous-zone à Roloboceras hambrovi pro parte?). L'étude systématique de cette faune de Roloboceratinae a permis de parfaire la connaissance de Megatyloceras coronatum Rouchadzé, 1933, espèce-type de Megatyloceras Humphrey, 1949, ainsi qu'une meilleure compréhension du genre grâce à l'examen des espèces décrites dans le Bassin anglo-parisien. Une espèce nouvelle, M. leteilensis sp. nov., est créée et une description de M. transiens Casey, 1961, est faite ; des éclaircissements sont donnés sur sa possible position stratigraphique. Cette étude a également permis d'établir que l'espèce Megatyloceras ricordeanum (Orbigny, 1850) ne représente qu'un stade de croissance commun à plusieurs espèces de Megatyloceras plutôt qu'à une espèce à part entière et devrait être considérée comme un nomen dubium. L'hypothèse d'un dimorphisme sexuel chez Roloboceras Casey, 1954, et Megatyloceras est proposée. L'étude paléoécologique de cette faune de Roloboceratinae met en évidence leur répartition hétérogène au sein des dépôts de la platforme ardéchoise, montrant que celle-ci est clairement corrélée à une tranche bathymétrique. La distribution biostratigraphique des deux genres sur la plateforme du Languedoc montre une distribution étagée avec Roloboceras couvrant la partie sommitale du segment C2 jusqu'au segment C4 de l'enregistrement des isotopes stables du carbone, tandis que Megatyloceras est limité au milieu du segment C4 jusqu'à la partie basale du segment C6. Les deux genres coexistent pendant le segment C4.
• Ammonoidea ;
• Roloboceratinae ;
• Crétacé inférieur ;
• Aptien inférieur ;
• Ardèche ;
• Sud-Est France ;
• Bassin de Paris
This
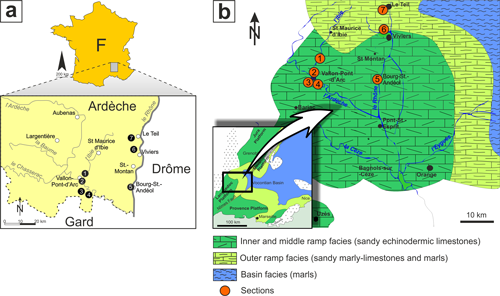
study covers the SE region of the Ardèche department, the so-called Bas-Vivarais, along the right bank of the Rhone valley in south-eastern France.
The limits of the study area are to the north of the Frayol stream near Le Teil,
southerly from the southern end of the Ardèche department, eastwards from the
Rhone river, and to the west of the Ardèche river (Fig. 1 ![]() ).
).
|
Figure 1:
Geographical and geological map of the studied area. Modified after
Pictet et al. (2015). |
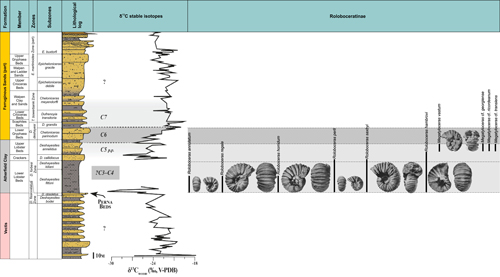
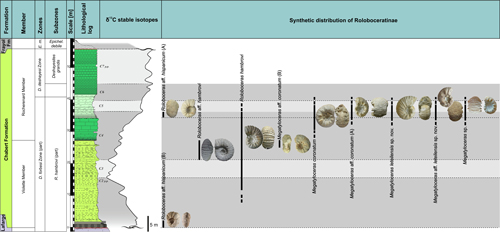
During the Early Cretaceous, shallow-water carbonate platforms developed around the Vocontian Basin: the Dauphiné-Jura platform to the north, the Provence platform to the south and the Languedoc platform to the west. In the Bas-Vivarais, the platform settles during the early Barremian on hemipelagic marl and marly-limestone deposited during the Valanginian-Barremian time interval (Lafarge, 1978; Pascal et al., 1989). The growing of the rudist-bearing carbonate platform is interrupted in the latest Barremian and replaced by oobioclastic echinodermic limestone deposits during the earliest Aptian, except along the Cevenol margin where the rudist platform continues to undergo subaerial exposure untill the onset of deposition of the OAE1a marls (Frau et al., 2018).
Following a long-term emersion phase documented by Pictet et al. (2015), shallow hemipelagic series subjected to detrital inputs with sandy-glauconitic oyster and cephalopod marls were deposited during the early Aptian drowning phase (Pictet, 2011; Pictet et al., 2015; Pictet & Delanoy, 2017). These deposits were first decribed by Carez (1882) who observed this basal sedimentary "unit 15", up to 25 m thick, which he called "Marly limestones with Ostrea aquila and cephalopods". These deposits are overlain by black marls with belemnites and ammonites indicating deepening and siltation (Pictet, 2011; Pictet et al., 2015; Pictet & Delanoy, 2017, with literature).
Kilian (1907-1913) and Kilian and Reboul (1915) reported the presence of Douvilleiceras hambrowii (sic) (Forbes, 1845) at Bourg-St-Andéol from a specimen housed in the collections of the Faculté des Sciences de Lyon. In the same work Kilian and Reboul reported, on the right bank of the Rhone, between Le Teil and Lafarge, a complex of siliceous limestones identical to those of l'Homme d'Armes. These limestones, which were exploited for cement production and hydraulic limes, were assigned to the lower Bedoulian series and would have yielded Douvilleiceras hambrowii (sic). At the Violette river, this limestone complex is followed by a nodular marl-limestone formation about 100 metres thick that Kilian and Reboul assigned to the upper Bedoulian series.
In 1958, Sornay (p. 170) described a section in the vicinity of
Viviers,
immediately west of the road from Paurière to Mélas. Above the siliceous and
cherty Lafarge limestones crops out a thick series of marls overlain by a sandy
limestone cliff with large-sized ammonites, including Cheloniceras
cf. hambrovii (Pl. 10 ![]() , fig. 2) and Procheloniceras
cf. pachystephanum (Uhlig, 1883) (= teratologic specimen of Megatyloceras
sp., Fig. 2.A-C
, fig. 2) and Procheloniceras
cf. pachystephanum (Uhlig, 1883) (= teratologic specimen of Megatyloceras
sp., Fig. 2.A-C ![]() ).
).
|
Figure 2:
Megatyloceras sp.,
specimen MNHN.F. A72532 coll. Sornay, "Bédoulien, quartier de
Paurière"
Viviers (Ardèche). Teratologic specimen cited as Procheloniceras cf. pachystephanum
(Uhlig, 1883) in Sornay
(1958, p. 170). D: Pseudosaynella
aff. raresulcata Orbigny, 1841: coll. R. Jaffré,
niveau condensé de la Sous-zone à Grandis, Jaulges, Yonne. |
Contensuzas (1980) described these upper Bedoulian series in a neighbouring section visible to the NNW of the Hellys hamlet and to the north of Rocherenard, which yield several specimens of Roloboceras sp. both in the marl-limestones series topping directly the siliceous limestones, and in the sandy limestone cliff above.
In 2013, Clavel et al. figured six sections of supra-Urgonian marly limestones in which two occurrences of Roloboceras Casey, 1954, are reported. The first one was found in the thick bar of echinoderm limestones in the Picourel section near Labastide-de-Virac, and the second in the same stratigraphic levels of the Chabert Farm section, NW of Viviers.
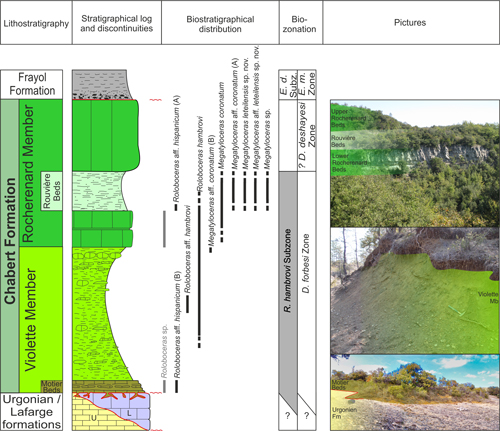
The
lower Bedoulian siliceous limestones were renamed as the Lafarge Formation,
while the "Marly limestones with Ostrea
aquila and cephalopods" or "upper Bedoulian" have recently been
revised and attributed to the Chabert Formation (Pictet et
al., 2015; Pictet & Delanoy,
2017) (Fig. 3 ![]() ). The latter
consists of a predominantly marl-dominated lower series, the Violette Member,
comprising at its base a shallow marly limestone unit, the Motier Beds. The
Violette Member grades upward to an upper series composed of echinodermic
limestones, the Rocherenard Member, within which a marly episode is inserted,
designated the Rouvière Marls (Pictet et
al., 2015; Pictet & Delanoy,
2017). The Chabert Formation
is topped by a thick marly series, the Frayol Formation, dated from the upper
Aptian. A systematic collection of ammonite faunas was carried out in different
lithological units of the Chabert Formation during the last 20 years, allowing a
detailed biostratigraphic and systematic study of the subfamily Roloboceratinae Casey,
1961.
). The latter
consists of a predominantly marl-dominated lower series, the Violette Member,
comprising at its base a shallow marly limestone unit, the Motier Beds. The
Violette Member grades upward to an upper series composed of echinodermic
limestones, the Rocherenard Member, within which a marly episode is inserted,
designated the Rouvière Marls (Pictet et
al., 2015; Pictet & Delanoy,
2017). The Chabert Formation
is topped by a thick marly series, the Frayol Formation, dated from the upper
Aptian. A systematic collection of ammonite faunas was carried out in different
lithological units of the Chabert Formation during the last 20 years, allowing a
detailed biostratigraphic and systematic study of the subfamily Roloboceratinae Casey,
1961.
|
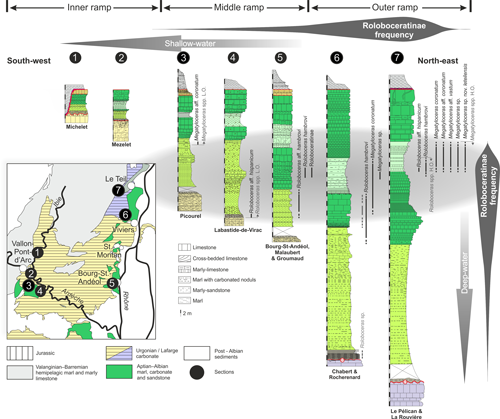
Figure 3:
Distribution of Roloboceratinae Casey, 1961, in the synthetic litho-log
of the Lower Aptian of the Languedoc platform. The stratigraphic ranges in light
grey are specimens cited in Contensuzas (1980) and Clavel et
al. (2013). |
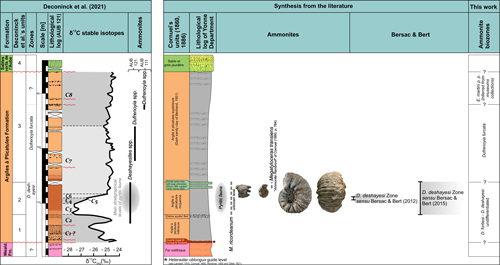
In France the genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949, have long been recognised as "Bedoulian" or Aptian, without any further precision, except for indicating either the "lower" or "upper" moiety (Orbigny, 1850b; Kilian, 1907-1913; Kilian & Reboul, 1915; Corroy, 1925; Roch, 1927). Subsequently, Megatyloceras ricordeanum (Orbigny, 1850) benefited from a more precise stratigraphic situation since it has been reported in the Parahoplites deshayesi Zone of the Paris Basin (Corroy, 1925), Deshayesites deshayesi Zone (Casey, 1961; Amédro & Matrion, 2004a, 2004b; Bersac & Bert, 2012, 2015; Amédro et al., 2017) or in the upper part of the Deshayesites forbesi Zone (Moreno et al., 2014; Bulot et al., 2018). Both Roloboceras and Megatyloceras are common components of the ammonite fauna of the mid-lower Aptian series.
In 1984, Busnardo made an important precision by recognizing a Roloboceras hambrovi Zone within the Lower Aptian (former "Bédoulien"/Bedoulian) unit-stratotype of Cassis-Roquefort-la-Bédoule, in which the presence of the index-species was known since the work of Roch (1927). More recent work undertaken on the same section has shown important divergences in the taxonomic interpretation of the species of Deshayesites Kazansky, 1914, which led to controversy over the stratigraphic position of the Roloboceratinae within either the Deshayesites forbesi Zone (Casey, 1961; Casey et al., 1998; Pictet, 2011; Najarro et al., 2011; Ivanov & Idakieva, 2013; Frau et al., 2015; Pictet et al., 2015) or perhaps the Deshayesites deshayesi Zone (Moullade et al., 2000; Ropolo et al., 2006, 2008a, 2009a, 2009b; Bersac & Bert, 2015). Because of this controversy, a significant stratigraphic shift was suggested between the biostratigraphic range of the Roloboceratinae of the unit-stratotype of Cassis-Roquefort-la-Bédoule and the distribution of these same faunas in southern England (Isle of Wight) where they characterise the upper part of the Deshayesites forbesi Zone (Casey, 1961; Casey et al., 1998). In fact, Ropolo et al. (2008b) explained this discrepancy by a delayed appearance of the Roloboceratinae in the South Provence intrashelf basin because of an ecological barrier.
In the meantime, Moreno et al. (2007, 2009) demonstrated that, in Spain, Roloboceras and Megatyloceras coexisted with a fauna containing Deshayesites, some elements of which were conspecific or had great affinities with forms of the English Lower Greensand Deshayesites forbesi Zone (Deshayesites forbesi Casey 1961, D. fittoni Casey, 1961, D. euglyphus Casey, 1964, D. gr. spathi Casey, 1964). Other works in Spain (Najarro et al., 2011), in Bulgaria (Ivanov & Idakieva, 2013) and in south-eastern France (Pictet, 2011; Frau et al., 2015; Pictet et al., 2015) support the positioning of Roloboceratinae levels of the Tethys regions in the Deshayesites forbesi Zone (ex D. weissi Zone), based on both micro- and macropaleontological, sedimentologic and chemostratigraphic data, still in conjunction with the Ocean Anoxic Event 1a (OAE1a).
Bersac and Bert (2012, 2015) have different conceptions of species of Deshayesites present in this stratigraphic interval. They indeed consider D. annelidus Casey, 1961, as a minor synonym of D. deshayesi (Orbigny, 1841), leading to a new conception of the Deshayesites deshayesi Zone, which then includes de facto the Deshayesites annelidus Subzone sensu Casey et al. (1998), and thus containing a large part of the biostratigraphic range of Megatyloceras.
The IUGS Lower Cretaceous ammonite Working Group, the Kilian Group, retained a Roloboceras hambrovi Subzone in the upper part of the Deshayesites forbesi Zone of the Standard Mediterranean Ammonite Scale (standard zonation of the Mediterranean domain of Reboulet et al., 2011, 2014, 2018) (Table 1), which serves as a stratigraphic framework for this work.
Table 1: Biostratigraphic scheme of the Aptian sensu Reboulet et al., 2011, 2014, 2018 (1), and Moullade et al., 2011, 2017 (2).
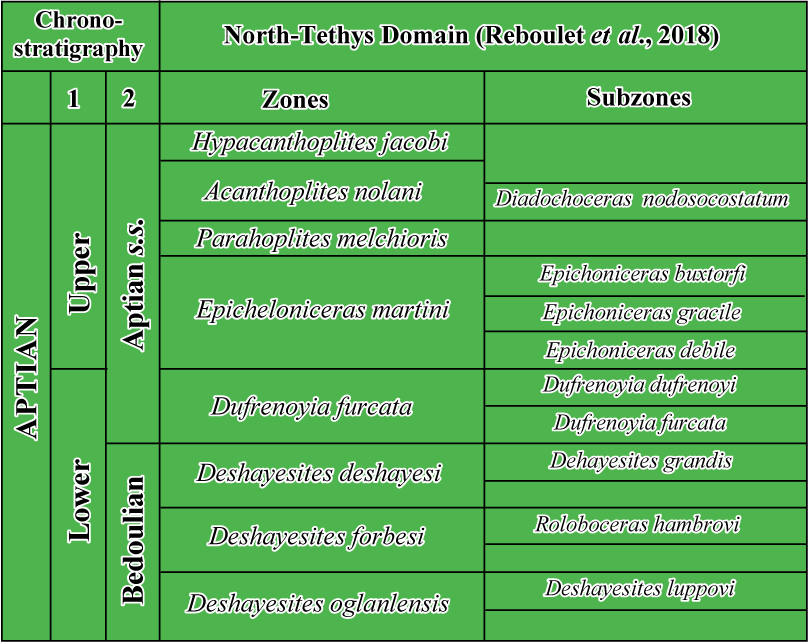 |
The subfamily Roloboceratinae was introduced by Casey (1961) for the two Lower Aptian ammonite genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949, but Casey also included the Hauterivian-Barremian genus Paraspiticeras Kilian, 1910. The latter was later reassigned to the Emericiceratidae Vermeulen, 2004. Roloboceras and Megatyloceras include about twenty taxa of ammonites with depressed to more or less cadicone shell morphologies. They also possess a wide latero-ventral region rounded at all growth stages and a single row of very strong tubercles along the umbilical margin. As pointed out by Casey (1961, p. 176) this peculiar morphology and ornamentation allows facile discrimination from other Douvilleiceratidae. At present, the origin of Roloboceratinae is uncertain. According to Pictet (2012) this origin would be among the group of Cheloniceras cornuelianum (Orbigny, 1841) and would be the result of a neotenic process from the Royerianum stage. More recently, Bersac and Bert (2017) debated the possibility that Roloboceratinae is a neotenic offshot of Procheloniceras Spath, 1923.
Although the origin of the Roloboceratinae is still unclear, the rich fauna of this sub-family collected in the marls and glauconitic sandy limestones of the Chabert Formation of the southern Ardèche improves our knowledge on systematics, biostratigraphy and facies distribution.
Conventions and repositories of specimens
GSM: British Geological Survey, Nottingham, UK
IWCMS: Dinosaur Isle Museum, Sandown, Isle of Wight, UK
UCB-FSL: Faculté des Sciences de Lyon, France
MGL: Musée Géologique de Lausanne, Suisse
MHNG: Muséum d'Histoire naturelle de Genève, Suisse
MNHN: Muséum National d'Histoire naturelle, France
GEPI: Département de Géologie et de Paléontologie, Genève, Suisse
UJF-ID: Université Joseph-Fourier, Grenoble, France
tl, chb, brg: Baudouin coll., except for tl059 (Gonnet coll.), tl 060, 061, 070 (Boselli coll.), tl071 and chb008 (Delanoy coll.)
VI: Boselli coll.
RG: Gonnet coll.
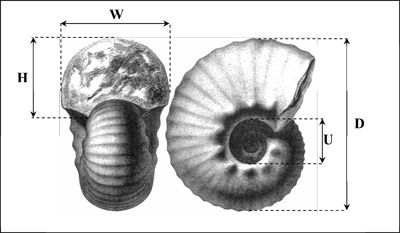
Dimensions
are given in millimetres: D = diameter; W
= whorl breadth; H
= whorl height; U = diameter
of umbilicus; Nc/2 = number of ribs per half whorl; Nt/2 = number
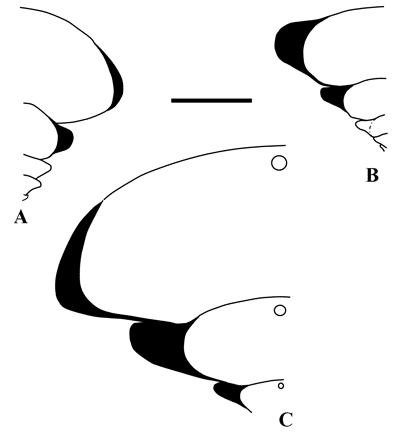
of tubercles per half whorl (Fig. 4 ![]() ).
).
|
Figure 4:
Diagram showing the
measurements made on the specimens in this study. |
Super-family Douvilleiceratoidea Parona & Bonarelli, 1897
Family Douvilleiceratidae Parona & Bonarelli, 1897
Subfamily Roloboceratinae Casey, 1961
Genus Roloboceras Casey, 1954
In 1954, Casey (p. 114) introduced Roloboceras for Douvilleiceratidae with a subcircular and depressed whorl section, a high and smooth umbilical wall, with an ornamentation of thick, blunt ventral ribs arising from large periumbilical nodular tubercles and, finally, a simplified suture line. He designates as type-species Ammonites hambrovi Forbes, 1845, a taxon of the English Lower Greensand whose generic attribution has been interpreted in various ways in the litterature, including Douvilleiceras Grossouvre, 1894, Cheloniceras Hyatt, 1903, and Megatyloceras Humphrey, 1949. This last genus is restricted (Casey, 1954, p. 114) to species showing a disproportionate development of periumbilical tubercles and affine to the type-species Douvilleiceras coronatum Rouchadzé, 1933, designated by Humphrey (1949).
Type-species: Ammonites hambrovi Forbes, 1845, by original designation of Casey (1954, p. 114).
Diagnosis: Ammonites, usually middle to large-sized, with a semi-circular, depressed cross-section. Ventral region rounded, very broad, merging with the flanks. Deep umbilicus with a high, sub-vertical, almost smooth umbilical wall. Ornamentation consisting of thick, low, rounded ribs, simple or arising in pairs, more rarely by three from large to very large obtuse or nodule-like periumbilical tubercles, prominent, becoming bulliform on the body chamber. Few intercalatory ribs arising near the tubercles. Simplified suture line with large external saddle.
Roloboceras Casey, 1954, presents five ontogenic stages: 1) the embryonic stage; 2) from 2-3 mm in diameter, a juvenile stage, resembling the Royerianum-stage of Cheloniceras Hyatt, 1903; 3) between 6 and 10 mm, the appearance of a more pronounced ribbing, differentiated by main and intercalatory ribs and by the presence of more or less thorn-like peri-umbilical tubercles (perli-stage of Spath, 1930); 4) an ornamentation typical of Roloboceras, appearing towards 20-25 mm in diameter; 5) senile stage with large simple radial ribs which can be erased in certain adult specimens.
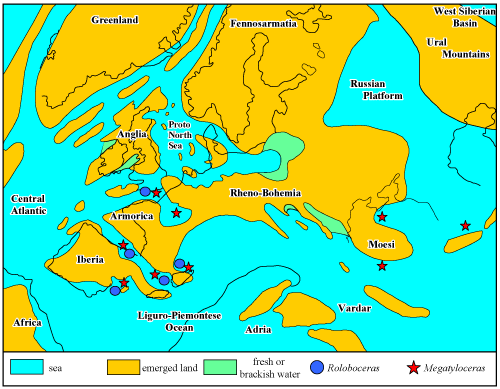
Geographic and stratigraphic distribution: Roloboceras Casey, 1954, was widely cited along the northern Tethys and Central Atlantic margins. Records from Venezuela (Renz, 1982) and Colombia (Etayo-Serna et al., 1994; Patarroyo, 2002; Sharikadze et al., 2004) are doubtful though. These are either based on specimens which are too incomplete, or on determinations which appear to be erroneous, or on imprecise data on the stratigraphic position of collected specimens, or deviating from the generally accepted distribution of the genus [see further discussion about Roloboceras hambrovi (Forbes, 1845)].
Thus,
the genus is known with certainty from France, England and Spain; a Bulgaria
record is unclear (Fig. 5 ![]() ). Its stratigraphic distribution is restricted to the
Lower Aptian, Deshayesites forbesi
Zone, Roloboceras hambrovi Subzone of
the Tethys regions, and Deshayesites
forbesi Zone, D. kiliani, D.
callidiscus and D. annelidus subzones in England.
). Its stratigraphic distribution is restricted to the
Lower Aptian, Deshayesites forbesi
Zone, Roloboceras hambrovi Subzone of
the Tethys regions, and Deshayesites
forbesi Zone, D. kiliani, D.
callidiscus and D. annelidus subzones in England.
Figure 5:
Paleogeographic distribution of the genera Roloboceras
Casey, 1954, and Megatyloceras Humphrey,
1949. |
Systematic of the genus: Casey (1954) placed Ammonites hambrovi (Forbes, 1845), Cheloniceras hambrovi var. horrida Spath, 1930, and C. perli (Spath, 1930) into Roloboceras. Later, other species were described by Casey (1961) from material collected from the Lower Greensand and by Sornay & Marin (1972) in Spain.
In the most recent Fossilum Catalogus, Klein and Bogdanova (2013) retained ten species or subspecies, respectively:
Roloboceras annulatum Casey, 1961
Roloboceras arnaudi (Coquand, 1866)
Roloboceras hambrovi hambrovi (Forbes, 1845)
Roloboceras hambrovi subnodosum Casey, 1961
Roloboceras hispanicum hispanicum Sornay, 1972
Roloboceras hispanicum rotundatum Sornay, 1972
Roloboceras horridum (Spath, 1930)
Roloboceras perli (Spath, 1930)
Roloboceras regale Casey, 1961
Roloboceras saxbyi Casey, 1961
A review of the reported type-specimens of these species, allows discernment of two main groups:
large-sized forms with a maximum diameter in excess of 200 mm including the species Roloboceras arnaudi, R. hambrovi, R. regale, R. horridum and R. hispanicum; R. saxbyi seems to have a special place in this group because of its smaller size and the early loss of the strong periumbilical tubercules;
small-sized forms with a maximum diameter of about 40 mm as Roloboceras annulatum and R. perli. To this group should be added the Iberian individuals figured by Moreno et al. (2007, Pl. I, figs. 4-5; 2010, electronic appendix, Pl. IV, fig. 1) and Najarro et al. (2011, Fig. 8T, W) under Roloboceras sp. microconch. The relationships between these two groups would undoubtedly merit further investigation, but unfortunately the material studied in this work does not permit this, as any form of the second group has been collected in the Ardèche sections.
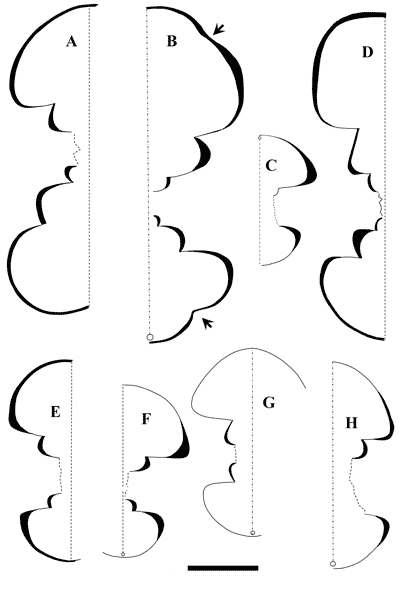
Roloboceras hambrovi (Forbes, 1845)
(Figs. 6H-I ![]() , 7A-C, E-F, H
, 7A-C, E-F, H ![]() ;
Pl. 1
;
Pl. 1 ![]() , figs.
1a-b, 3a-d;
Pl. 2
, figs.
1a-b, 3a-d;
Pl. 2 ![]() , figs. 1a-c, 2a-c;
, figs. 1a-c, 2a-c;
Pl. 3 ![]() , figs. 1, 2a-c;
Pl. 4
, figs. 1, 2a-c;
Pl. 4 ![]() , fig.
1a-b; Pl. 7
, fig.
1a-b; Pl. 7 ![]() , figs. 2a-b, 3a-c;
, figs. 2a-b, 3a-c;
Pl. 8 ![]() , fig. 2a-b;
Pl. 9
, fig. 2a-b;
Pl. 9 ![]() , figs. 1a-b, 3a-b;
Pl. 11
, figs. 1a-b, 3a-b;
Pl. 11 ![]() , figs. 1, 2a-b, 3a-b; Table
2)
, figs. 1, 2a-b, 3a-b; Table
2)
Synonymy:
1845. Ammonites Hambrovii sp. nov. - Forbes, p. 354, Pl. V, fig. 4.
? 1866. Ammonites Arnaudi sp. nov. - Coquand, p. 238, Pl. II, figs. 1-2.
? 1882. Ammonites Arnaudi Coquand - Mallada, p. 19, Pl. 5, figs. 1-2.
1921. Cheloniceras hambrovii (Forbes) - Spath, p. 317, Pl. XXVI, fig. 3.
non 1927. Douvilleiceras cf. hambrovi (Forbes) - Roch, p. 22, Pl. I, fig. 3 (= Megatyloceras cf. coronatum (Rouchadzé, 1933).
1930. Cheloniceras hambrovi (Forbes) var. horrida - Spath, p. 444.
? 1950. Ammonites Arnaudi Coquand - Bataller, p. 376, Fig. p. 377.
1954. Roloboceras hambrovi (Forbes) - Casey, p. 114.
1957. Roloboceras hambrovi (Forbes) - Arkell et al., p. L384, Fig. 501: 9a-b.
1961. Roloboceras hambrovi (Forbes) - Casey, p. 179, Pl. XXIX, fig. 5a-b; Pl. XXX, fig. 8; Pl. XXXI, fig. 3a-b; Pl. XXXII, fig. 5a-b; Figs. 54a-l, 55a-b, 57a-b.
? 1961. Ammonites Arnaudi Coquand - Casey, Fig. 55c-d.
1961. Roloboceras hambrovi subnodosum nov. var. - Casey, p. 182, Pl. XXIX, fig. 6a-b; Pl. XXX, fig. 7a-b; Fig. 55a (left specimen).
1961. Roloboceras horridum Casey - Casey, p. 185; Pl. XXXII, figs. 1a-b, 2-4.
?1961. Roloboceras horridum Casey - Casey, p. 185; Pl. XXXI, fig. 5a-b.
pars 1961. Roloboceras regale sp. nov. - Casey, p. 186, Pl. XXX,? fig. 9a-c; Pl. XXXI, fig. 1a-c (only).
? 1961. Roloboceras aff. regale Casey - Casey, p. 186, Pl. XXXI, fig. 2a-b, Pl. XXXII, fig. 6.
1966. Roloboceras hambrovi (Forbes) - Schindewolf, Fig. 412a-g.
? 1967. Roloboceras hambrovi (Forbes) - Cox et al., Pl. 64, fig. 4.
? 1972. Roloboceras sp. ex gr. arnaudi Coquand - Sornay in Sornay & Marin, p. 108(10), Fig. 7, Pl. B, fig. 4.
? 1976. Roloboceras regale Casey - Peybernès, Pl. XXV, fig. 8.
? 1982. Roloboceras hambrovi (Forbes) - Renz, p. 19, Pl. 2, fig. 25a-b.
? 1983. Roloboceras sp. gr. hambrovi (Forbes) - arnaudi Coquand - Murat, p. 59, Pl. VIII, fig. 3.
1996. Roloboceras hambrovi (Forbes) - Wright et al., p. 267, Fig. 207: 2a-b.
non 2006. Roloboceras hambrovi (Forbes) - Ropolo et al., Pl. 11, fig. 2 (= Cheloniceras sp.).
2007. Roloboceras hambrovi (Forbes) - Moreno-Bedmar et al., Pl. 1, fig. 1.
2008. Roloboceras hambrovi (Forbes) - Salas & Moreno-Bedmar, Pl. 1, figs. A-B.
2008b. Roloboceras hambrovi (Forbes) - Moreno-Bedmar et al., Fig. 6.
non 2008a. Roloboceras hambrovi (Forbes) - Ropolo et al., p. 26, Pl. 6, fig. 2 (= Cheloniceras sp.).
non 2008a. Roloboceras horridum Casey - Ropolo et al., p. 27, Pl. 12, fig. 3 (= Cheloniceras sp.).
2009. Roloboceras hambrovi (Forbes) - Moreno-Bedmar et al., Fig. 10E.
non 2009a. Roloboceras hambrovi (Forbes) - Ropolo et al., Pl. 11, fig. 2 (= Cheloniceras sp.).
non 2009b. Roloboceras hambrovi (Forbes) - Ropolo et al., p. 147, Pl. 6, fig. 2 (= Cheloniceras sp.).
non 2009b. Roloboceras horridum Casey - Ropolo et al., p. 149, Pl. 12, fig. 3 (= Cheloniceras sp.).
? 2010. Roloboceras hambrovi (Forbes) - Moreno-Bedmar et al., electronic appendix, Fig. IB.
2010. Roloboceras hambrovi (Forbes) - Moreno-Bedmar et al., electronic appendix, Fig. IC, D, IIA.
? 2011. Roloboceras hambrovi (Forbes) - Najarro et al., Fig. 8X.
? 2011. Roloboceras cf. hambrovi (Forbes) - Raisossadat, p. 169, Fig. 5G.
non 2013. Roloboceras aff. hambrovi (Forbes) - Ivanov & Idakieva, Fig. 14I-J (= Megatyloceras sp.).
2015. Roloboceras hambrovi (Forbes) - Pictet et al., p. 111, Fig. 9b-c.
Lectotype:
The lectotype is specimen GSM 2295 designated by Casey (1961, p. 179)
from the Lower Lobster Bed at Atherfield (Isle of Wight, UK), and it is housed
in the collections of the British Geological Survey Museum (Nottingham, UK):
Pl. 1 ![]() ,
fig. 1a-b. It
was first illustrated by Forbes (1845, Pl. V, fig. 4; right-hand specimen)
and then re-illustrated by Casey (1961, p. 179, Pl.
XXXII, fig. 5).
,
fig. 1a-b. It
was first illustrated by Forbes (1845, Pl. V, fig. 4; right-hand specimen)
and then re-illustrated by Casey (1961, p. 179, Pl.
XXXII, fig. 5).
Studied material (N = 15): Lectotype GSM 2295, Lower Aptian, Deshayesites forbesi Zone, Atherfield (Isle of Wight, UK); specimens tl003, tl004, tl005, tl006, tl008, tl012, tl081, tl105 coll. Baudouin, tl059 coll. Gonnet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche); chb003 coll. Baudouin, chb008 coll. Delanoy, VI004 coll. Boselli, MGL.104624, MGL.104622 coll. A. Pictet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Rocherenard, Viviers (Ardèche).
Measurements (in mm):
Table 2: Measurements of Roloboceras hambrovi (Forbes, 1845).
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl008 | c239 | 90 | c71 | c108 | 18 | 0 | 0.38 | 0.30 | 0.45 | 1.52 |
| tl012 | 258 | 87 | 105 | 134 | 11 | 0 | 0.34 | 0.41 | 0.52 | 1.28 |
| MGL.104624 | 216 | 76 | 79 | 113 | 18 | 5 | 0.35 | 0.37 | 0.52 | 1.43 |
| chb003 | 208 | 74 | 84 | 109 | 15 | 0 | 0.36 | 0.40 | 0.52 | 1.30 |
| tl003 | 203 | c73 | 79 | 118 | - | - | 0.36 | 0.39 | 0.58 | 1.49 |
| tl059 | 163 | 52 | 56 | c91 | 6 | 0.32 | 0.34 | 0.56 | 1.63 | |
| tl081 | 158 | 59 | 63 | 93 | 15 | 5 | 0.37 | 0.40 | 0.59 | 1.48 |
| 118 | 46 | 46 | 81 | 19 | 5 | 0.39 | 0.39 | 0.69 | 1.76 | |
| tl105 | 156 | 54 | 61 | c74 | - | 5 | 0.35 | 0.39 | 0.47 | 1.21 |
| 105 | 38 | 48 | 60 | - | 4 | 0.36 | 0.46 | 0.57 | 1.25 | |
| tl004 | 148 | 52 | 61 | 82 | - | - | 0.35 | 0.41 | 0.55 | 1.34 |
| c102 | 38 | 43 | 75 | - | 6 | 0.37 | 0.42 | 0.74 | 1.74 | |
| tl006 | 133 | 42 | 61 | - | - | 5 | 0.32 | 0.460 | - | - |
| 94 | 30 | 37 | 61 | - | 5 | 0.32 | 0.39 | 0.65 | 1.65 | |
| VI004 | 132 | 46 | 50 | 75 | 0.34 | 0.37 | 0.56 | 1.50 | ||
| MGL.104622 | 113 | 40 | 39 | - | - | 6 | 0.35 | 0.35 | - | - |
| chb008 | 110 | 39 | c45 | c61 | 10 | - | 0.35 | 0.41 | 0.55 | 1.36 |
| tl005 | 83 | 28 | - | c50 | - | 6 | 0.34 | - | 0.60 | - |
| c57 | c22 | c21 | 35 | - | 6 | 0.39 | 0.37 | 0.61 | 1.67 | |
| GSM2295 | C75 | 23 | 30 | 52 | - | - | 0.31 | 0.41 | 0.70 | 1.73 |
Description: Probably a dimorphic Roloboceras of medium to large size, whose body chamber represents approximately ¾ of the last whorl. The coiling is evolute, and the whorls overlap in the upper third of the flanks, with a notable opening of the umbilicus on the second half of the adult body chamber. The whorl section is depressed, with a subvertical to slightly convex umbilical wall. The flanks and the venter are merged into a broad, domed lateroventral region. The section becomes more subcircular on the body chamber of adult specimens.
Ornamentation
consists in the inner whorls of large and strong periumbilical tubercles,
decreasing in strength during ontogenic development; these are gradually
replaced by large periumbilical bullae towards about H
= 50-70 mm. There are 6-10 tubercles per whorl on the innermost observed whorls
(D = about 55-60 mm), later replaced
by large and rounded, nodule-like tubercles, sometimes more or less laterally
pinched (specimen chb008, Pl. 7 ![]() , fig. 3). The number of tubercles increases
during the growth to 10 or even 16 on the following whorls (specimen tl008, Pl. 11
, fig. 3). The number of tubercles increases
during the growth to 10 or even 16 on the following whorls (specimen tl008, Pl. 11 ![]() , fig. 1); they are weaker and give rise to 2, sometimes 3, rounded ribs. A
few intermediary ribs arise isolated close to the umbilical shoulder, sometimes
thinner and weaker in the ventral area. On the last whorl, the tubercles are
replaced by about 10-12 bullae, from which one, or more rarely two, thick,
rounded ribs originate. In the second part of the body chamber the bullae
disappear and the ribs, which tend to decrease in number, are simple, gradually
thickening towards the ventral area. The peristome could not be observed on any
of the specimens but the apertural margin seems characterized by a weakening of
the ribbing (tl008, Pl. 11
, fig. 1); they are weaker and give rise to 2, sometimes 3, rounded ribs. A
few intermediary ribs arise isolated close to the umbilical shoulder, sometimes
thinner and weaker in the ventral area. On the last whorl, the tubercles are
replaced by about 10-12 bullae, from which one, or more rarely two, thick,
rounded ribs originate. In the second part of the body chamber the bullae
disappear and the ribs, which tend to decrease in number, are simple, gradually
thickening towards the ventral area. The peristome could not be observed on any
of the specimens but the apertural margin seems characterized by a weakening of
the ribbing (tl008, Pl. 11 ![]() , fig. 1).
, fig. 1).
Poor preservation inhibited detailed analysis of the suture pattern.
|
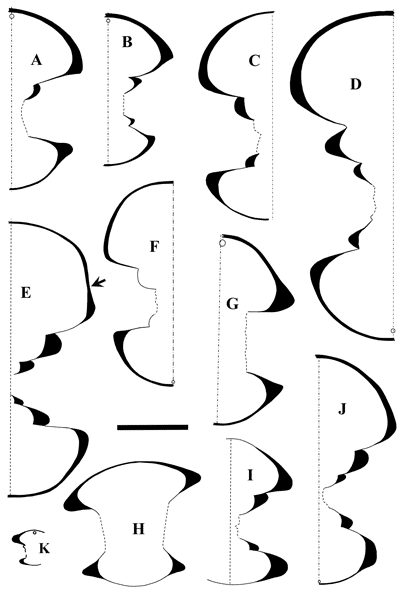
Figure 6:
Juvenile whorls (nuclei) of some English and Spanish specimens of Roloboceras
Casey, 1954, according to Casey, 1961: A-C:
Roloboceras regale Casey, 1961, GSM. 99287 (Casey, 1961, Pl.
XXX,
fig. 9a-c); D: Roloboceras hispanicum Sornay,
1972, PUAB 48.332; E-G:
Roloboceras horridum Casey,
1961, SM. B27128 (Casey,
1961, Pl. XXXI, fig. 5a-b); H-I: R.
hambrovi (Forbes, 1845), GSM 30952 (Casey, 1961, Pl.
XXXI,
fig. 3a-b). A-C, E-I
from Atherfield (UK); D: Tejeria de Josa (Spain). Scale
bar is 10 mm. |
Variability: The variability observed in the Ardèche population of Roloboceras hambrovi (Forbes, 1845) mainly concerns the size of the specimens preserved with body chamber, the strength of the peri-umbilical tubercles, their number and the consequences this has on ribbing. The variability in size seems to allow the recognition of two groups which do not appear to be linked by intermediate forms:
a group composed of large-sized specimens whose maximum diameter (D) is between 250 and 300 mm, the diameter at the end of the phragmocone is between 160 and 180 mm, and the height of the whorl (h) at the last suture being about 70 mm. The ornamentation is characteristic of the genus with large periumbilical tubercles on the innermost whorls which gradually change irregularly into more or less pinched tubercles and then into bullae on the last whorl, almost disappearing at the end of the body chamber where the ornamentation tends to weaken in the apertural margin;
a medium-sized group where D is
between 150 and 170 mm and the diameter of the phragmocone is between 80 and 110
mm, h being then about 40 mm. The ornamentation is strong for inner
whorls but seems to weaken on the body chamber even if some erosion is
discernible (tl004, Pl. 3 ![]() , fig. 2). Other observations show that in some
specimens the whorl section is weakly compressed at the end of the body chamber
(tl004), so that strong and spaced single ribs (VI004, Pl. 7
, fig. 2). Other observations show that in some
specimens the whorl section is weakly compressed at the end of the body chamber
(tl004), so that strong and spaced single ribs (VI004, Pl. 7 ![]() ,
fig. 2) may be
present at the end of the body chamber and, finally, that wider and flatter
tubercles may be present on the inner whorls (tl105, Pl. 8
,
fig. 2) may be
present at the end of the body chamber and, finally, that wider and flatter
tubercles may be present on the inner whorls (tl105, Pl. 8
![]() , fig. 2), an
ornamental feature found in R. hispanicum
Sornay, 1972, of which it represents one of the main characteristics.
These tubercles of the "hispanicum" morphology can be observed, but arranged
randomly, in large specimens (? = macroconchs; Pl. 11
, fig. 2), an
ornamental feature found in R. hispanicum
Sornay, 1972, of which it represents one of the main characteristics.
These tubercles of the "hispanicum" morphology can be observed, but arranged
randomly, in large specimens (? = macroconchs; Pl. 11 ![]() ,
fig. 1) and also for the
lectotype of R. hambrovi (Pl. 1
,
fig. 1) and also for the
lectotype of R. hambrovi (Pl. 1 ![]() , fig.
3a-d) which is a smaller specimen with onset of body-chamber. Such tubercles are
observable on some specimens cited and figured as R. horridum (Spath,
1930) "showing
early loss of gross tubercles" (Pl. 12
, fig.
3a-d) which is a smaller specimen with onset of body-chamber. Such tubercles are
observable on some specimens cited and figured as R. horridum (Spath,
1930) "showing
early loss of gross tubercles" (Pl. 12 ![]() , fig. 3a-c;
Pl. 30
, fig. 3a-c;
Pl. 30 ![]() , fig. 2a-c). It is
clear that the phylogenetic and stratigraphic relationships between these two
species requires further clarification.
, fig. 2a-c). It is
clear that the phylogenetic and stratigraphic relationships between these two
species requires further clarification.
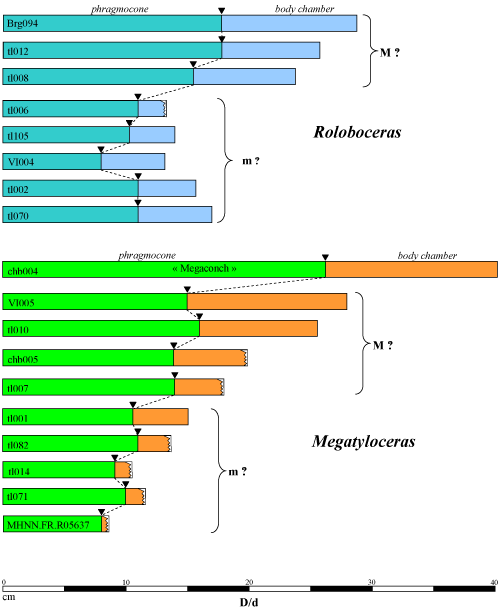
The apparent existence of these two groups, which are morphologically and ornamentally very similar but differ only in size, suggests the possibility of a dimorphism probably of sexual origin in which the medium-sized forms would represent microconchs and the large forms macroconchs. This hypothesis, which is based on a fairly small population, nevertheless needs to be reinforced by a larger study.
|
Figure 7:
Section of some Roloboceras Casey,
1954. The arrows indicate sequelae left by an injury. Scale bar is 5 cm. |
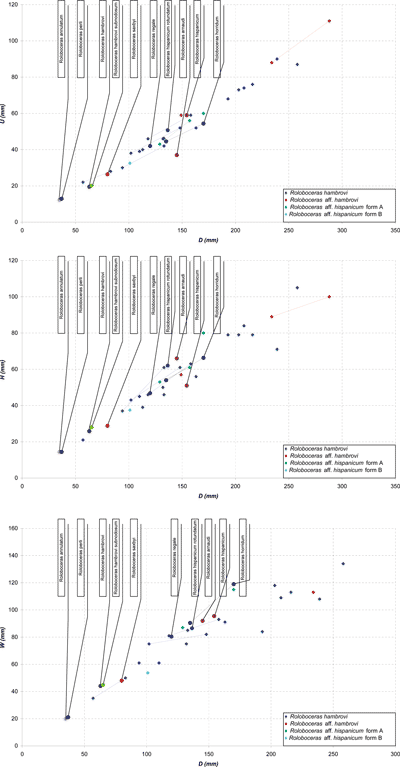
Discussion: Roloboceras hambrovi subnodosum Casey, 1961, was created for Roloboceras morphologies showing a prolonged juvenile stage with smaller and more numerous tubercles. Founded on small, incomplete specimens, this "variety" whose adult stage would not be distinguishable from the typical form of R. hambrovi (Forbes, 1845) (Casey, 1961, p. 182) must be regarded as an expression of the intraspecific variability of the species.
According
to Casey (1961, p. 186), R.
hambrovi can be distinguished from Roloboceras
regale Casey, 1961 (Fig.
6A-C ![]() ;
Pl. 1
;
Pl. 1 ![]() , fig. 2a-c) by its ornamental
features (strength and number of periumbilical tubercles, number of ventral ribs)
and morphological characters (narrower umbilicus). Although the specimens
studied here form a relatively homogeneous population, they nevertheless show
some variability (Fig. 8
, fig. 2a-c) by its ornamental
features (strength and number of periumbilical tubercles, number of ventral ribs)
and morphological characters (narrower umbilicus). Although the specimens
studied here form a relatively homogeneous population, they nevertheless show
some variability (Fig. 8 ![]() ). Their dimensional parameters correspond both to those
of Roloboceras hambrovi and to those of R. regale, without the possibility of distinguishing between the two
groups by using these characters. Thus, and by comparison with the values given
by Casey (1961), the U/D ratio
shows that 28% of specimens have values corresponding to R.
hambrovi, 22% to R. regale, 35% have values common to both taxa and finally 15% have
values corresponding to neither species. For the H/D ratio there are 0%, 28%, 35%, and 35% respectively. Finally, for
the W/D ratio the percentages are 7%,
14%, 14% and 64% respectively. Moreover, these values are randomly distributed
among the specimens. Thus, for example, tl006 has at D
= 94 mm, U/D and W/D ratios of R.
hambrovi and an H/D ratio of R. regale.
Among the studied material, only the specimen tl003 displays, for all 3 ratios,
values that are specific to R. regale.
). Their dimensional parameters correspond both to those
of Roloboceras hambrovi and to those of R. regale, without the possibility of distinguishing between the two
groups by using these characters. Thus, and by comparison with the values given
by Casey (1961), the U/D ratio
shows that 28% of specimens have values corresponding to R.
hambrovi, 22% to R. regale, 35% have values common to both taxa and finally 15% have
values corresponding to neither species. For the H/D ratio there are 0%, 28%, 35%, and 35% respectively. Finally, for
the W/D ratio the percentages are 7%,
14%, 14% and 64% respectively. Moreover, these values are randomly distributed
among the specimens. Thus, for example, tl006 has at D
= 94 mm, U/D and W/D ratios of R.
hambrovi and an H/D ratio of R. regale.
Among the studied material, only the specimen tl003 displays, for all 3 ratios,
values that are specific to R. regale.
|
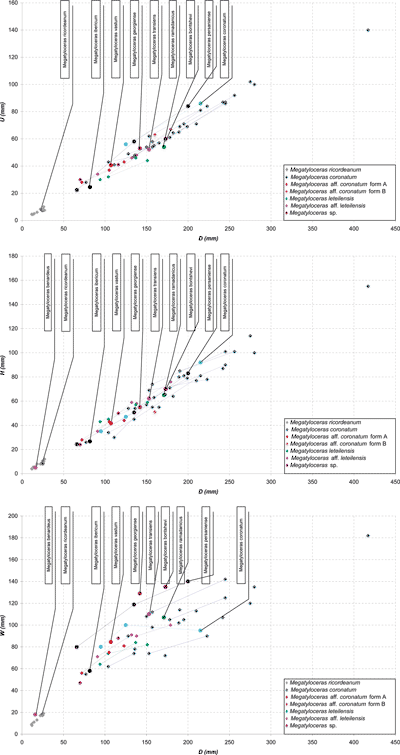
Figure 8:
Roloboceras
Casey, 1954: diameter variation of the umbilicus (U),
width (W) and height of the whorls (H) as a function of
the diameter (D)
of the population studied and the types of the literature. |
Ornamental characteristics also inhibit differentiation within the study sample. According to Casey (1961), the number of umbilical tubercles would be 14-15 at D = 140-150 mm in R. hambrovi and 12 on the holotype of R. regale at D = 120 mm. At the same diameters the number of ventral ribs would be 24 and 32 respectively, these differences in rib density are explained by a greater number of secondary and/or intercalatory ribs in R. regale.
On
the basis of the above morphological and ornamental criteria alone, it would be
tempting to consider that the criteria used by Casey (1961) to
differentiate R. hambrovi from R. regale
better reflect intraspecific variability. Nevertheless, examination of the
holotype of R. regale (No. GM99283; Pl. 1 ![]() , fig. 2a-c) shows that tubercles on the inner whorls are bulliform,
laterally pinched, and slightly prorsiradiate. On the last preserved whorl, the
tubercles, to the number of 12, become stronger and larger and they are still
quite strongly pinched radially. They give rise to 2 secondary ribs, between
which an intermediate rib is regularly present.
, fig. 2a-c) shows that tubercles on the inner whorls are bulliform,
laterally pinched, and slightly prorsiradiate. On the last preserved whorl, the
tubercles, to the number of 12, become stronger and larger and they are still
quite strongly pinched radially. They give rise to 2 secondary ribs, between
which an intermediate rib is regularly present.
The holotype of R. regale originates from the Deshayesites forbesi Zone, Deshayesites kiliani Subzone, and the distribution of the species is thought to be limited to this biostratigraphic unit (Casey, 1961; Casey et al., 1998; Simpson, 1985; Simpson, personal communication) within which it coexists with R. hambrovi. On the other hand, syntype BM46588 (Casey, 1961, Pl. XXXI, fig. 1a-c), whose stratigraphic origin is supposed to mirror that of the holotype (Casey, 1961, p. 187), shows stronger ornamentation comparable to that of R. hambrovi both at the level of tubercles and ribbing. This specimen is considered in this work as R. hambrovi. In conclusion, it appears that the dimensional criteria used by Casey (1961, p. 186) to differentiate R. hambrovi from R. regale cannot be taken into consideration. Because of the ornamental characters observed on the holotype of R. regale and its stratigraphic distribution, we keep R. hambrovi and R. regale (restricted to the holotype) separate.
Roloboceras horridum
(Spath, 1930) was first introduced by Spath as a variety of R.
hambrovi with a particularly more robust ornamentation than R.
hambrovi and based on a small (D =
54 mm) specimen showing body chamber. Furthermore, according to Spath
(1930, p. 444), the number of peri-umbilical tubercles is lower (5 at D
= 54 mm). Casey (1961) failed to find the corresponding specimen in the Spath
collection, and he thus proposed a neotype (Pl. 1 ![]() , fig. 1a-b). This specimen is
of larger size (170 mm), and shows large periumbilical tubercles, nodule-like on
the inner whorls, 8-9 in number on the last whorl. The first half of the
external whorl is poorly preserved, but 4-5 very large tubercles can be
recognized in the periumbilical region. The last whorl bears very thick strong
ribs and very large periumbilical bullae. The dimensional characters of the
neotype of R. horridum
given by Casey (1961, p. 185) are exactly the same as those given to R.
hambrovi. According to Casey (1961, p. 185), the persistence of the
very large tubercles stage is highly variable and could occur very early in
ontogenic development, as shown by the syntype SM.B27102 (Pl. 12
, fig. 1a-b). This specimen is
of larger size (170 mm), and shows large periumbilical tubercles, nodule-like on
the inner whorls, 8-9 in number on the last whorl. The first half of the
external whorl is poorly preserved, but 4-5 very large tubercles can be
recognized in the periumbilical region. The last whorl bears very thick strong
ribs and very large periumbilical bullae. The dimensional characters of the
neotype of R. horridum
given by Casey (1961, p. 185) are exactly the same as those given to R.
hambrovi. According to Casey (1961, p. 185), the persistence of the
very large tubercles stage is highly variable and could occur very early in
ontogenic development, as shown by the syntype SM.B27102 (Pl. 12 ![]() , fig. 3a-c)
which, moreover, has particularly strong ornamental affinities with the
lectotype of R. hambrovi. In the Lower Greensand, R. hambrovi and R.
horridum have the same stratigraphic distribution in the Deshayesites
forbesi Zone, Deshayesites kiliani
and D. callidiscus subzones and at the
base of the D. annelidus Subzone. This
leads us to consider R. horridum as a
particularly robust morphological pole of R. hambrovi. The nucleus of R.
horridum, figured by Casey, 1961 Pl. XXXI, fig. 5a-b shows very
pronounced Cheloniceras-like
characters by its simple primary ribs on the last whorl and its strong
periumbilical tubercles, and seems rather similar to a nucleus of a Cheloniceras
sp. (Fig. 6E-G
, fig. 3a-c)
which, moreover, has particularly strong ornamental affinities with the
lectotype of R. hambrovi. In the Lower Greensand, R. hambrovi and R.
horridum have the same stratigraphic distribution in the Deshayesites
forbesi Zone, Deshayesites kiliani
and D. callidiscus subzones and at the
base of the D. annelidus Subzone. This
leads us to consider R. horridum as a
particularly robust morphological pole of R. hambrovi. The nucleus of R.
horridum, figured by Casey, 1961 Pl. XXXI, fig. 5a-b shows very
pronounced Cheloniceras-like
characters by its simple primary ribs on the last whorl and its strong
periumbilical tubercles, and seems rather similar to a nucleus of a Cheloniceras
sp. (Fig. 6E-G ![]() ).
).
Roloboceras
hispanicum Sornay, 1972 (Pl. 5 ![]() , figs. 1a-b, 2a-b) can be distinguished from R.
hambrovi by its smaller adult size, broad, flat-rounded peri-umbilical
tubercles and its low weaker ribs which become evanescent when D
is about 95 mm. On adult specimens the ventro-lateral area is nearly smooth, the
peri-umbilical tubercles of the body-chamber are smaller, and more bulliform. On
juvenile specimens the ribs are mostly present on the venter, but are discreet,
sometimes missing, on the flanks (Fig. 6D
, figs. 1a-b, 2a-b) can be distinguished from R.
hambrovi by its smaller adult size, broad, flat-rounded peri-umbilical
tubercles and its low weaker ribs which become evanescent when D
is about 95 mm. On adult specimens the ventro-lateral area is nearly smooth, the
peri-umbilical tubercles of the body-chamber are smaller, and more bulliform. On
juvenile specimens the ribs are mostly present on the venter, but are discreet,
sometimes missing, on the flanks (Fig. 6D ![]() ). R.
hispanicum also shows a fairly high morphological and ornamental variability
affecting cross-section, umbilical dimensions, and number and strength of
tubercles (Sornay & Marin,
1972; Moreno-Bedmar & Bover,
2007; Moreno-Bedmar et al.,
2007, 2009, 2010). But the near absence of ventral ribbing is one of the most
significant characteristics for the species. As pointed out above, the presence
in the Rouvière Marls of specimens of R.
hambrovi showing great ornamental affinities with R.
hispanicum justify that the relationship between the two species must be
clarified.
). R.
hispanicum also shows a fairly high morphological and ornamental variability
affecting cross-section, umbilical dimensions, and number and strength of
tubercles (Sornay & Marin,
1972; Moreno-Bedmar & Bover,
2007; Moreno-Bedmar et al.,
2007, 2009, 2010). But the near absence of ventral ribbing is one of the most
significant characteristics for the species. As pointed out above, the presence
in the Rouvière Marls of specimens of R.
hambrovi showing great ornamental affinities with R.
hispanicum justify that the relationship between the two species must be
clarified.
Roloboceras arnaudi (Coquand, 1866) is only known from the original description and figurations which present morphological and ornamental analogies with R. hambrovi. Casey (1961, p. 182) discussed this taxon, pointing out the discrepancies with the Forbes species, considering the figures of Coquand (1866) to be idealized representations and considering, with uncertainty, the synonymy of R. arnaudi with R. hambrovi. The presence of regularly arranged, nodule-like tubercles, giving rise to a well-marked ventral ribbing, also excludes the consideration of R. arnaudi as conspecific with R. hispanicum. Recent research in the Coquand collection in Budapest has not located the type of Ammonites arnaudi (Ottilia Szives, personal communication). For these reasons it seems that the Coquand's species must be considered as a nomen dubium.
The
specimen of Roloboceras hambrovi cited by Kilian
(1907-1913) and Kilian
and Reboul (1915), still preserved in the FSL collections, shows a
particular ornamentation of the body chamber consisting of relatively thin and
spaced ribs. In addition, the internal whorls are not visible, the specimen is
relatively compressed and its precise stratigraphic level and location are
unknown. Nevertheless, the dark marly matrix suggests that this could belong to
the Violette Member. According to one of us (JMB), specimens with similar body
chamber ornamentation have been collected in Spain. Pending a better knowledge
of these forms, this specimen is considered here as Roloboceras
sp. (Pl. 8 ![]() , fig. 1a-c).
, fig. 1a-c).
Stratigraphic and geographical distribution: The specimen MGL.104622 was collected at the top of the Violette Member (Violette Member in Pictet & Delanoy, 2017), the specimens chb003, chb008, VI004 and MGL.104624 in the lower part of the Rocherenard Member (Rocherenard Member in Pictet & Delanoy, 2017), and the specimens tl003, tl004, tl005, tl006, tl008, tl012, tl059, tl070, tl081 and tl105 originate from the first marl-limestone levels of the Rouvière Marls (Rouvière beds in Pictet & Delanoy, 2017). All these levels are dated from the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone.
Roloboceras hambrovi (Forbes, 1845) is a species present in the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone in Spain and south-eastern France, and in the Deshayesites forbesi Zone, D. kiliani and D. callidiscus subzones in England (Casey, 1961; Casey et al., 1998).
Its occurrence in Venezuela (Renz, 1982) and Colombia (Etayo-Serna et al., 1994) remains highly uncertain. Indeed, the specimen described and figured by Renz (1982) seems much too fragmentary to be determined precisely at the generic level. In addition, the Roloboceras nov. sp. aff. regale (Casey, 1961) described and represented by Etayo-Serna et al. (1994), although presenting morphological similarities with Roloboceras hambrovi, comes from levels which have yielded Epicheloniceras bambucaense (Etayo-Serna et al., 1994) and Acanthoplites bigouretiformis (Etayo-Serna, 1979), species that indicate an Upper Aptian age.
Roloboceras aff. hambrovi (Forbes, 1845)
(Fig. 7D ![]() ;
Pl. 6
;
Pl. 6 ![]() , fig. 1;
Pl. 7
, fig. 1;
Pl. 7 ![]() , fig. 1;
Pl. 8
, fig. 1;
Pl. 8 ![]() , fig. 3; Table
3)
, fig. 3; Table
3)
Studied material (N = 2): Specimen brg094 coll. Baudouin, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, bed no. 7 of the Malaubert section, Pierrelatte (Drôme); specimen RG2003 coll. Gonnet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, bed no. 10 of the Malaubert section, Pierrelatte (Drôme).
Measurements (in mm):
Table 3: Measurements of Roloboceras aff. hambrovi (Forbes, 1845).
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| brg094 | 288 | 111 | 100 | - | 19 | 0 | 0.39 | 0.35 | - | - |
| 234 | 88 | 89 | 113 | c22 | c5 | 0.38 | 0.38 | 0.48 | 1.27 | |
| RG2003 | 149 | 59 | 57 | - | 16 | 7 | 0.40 | 0.38 | - | - |
Description: Ammonites of medium to large size, with a body chamber that represents approximately the last ¾ of the whorl. The coiling is evolute, the whorls overlapping in the upper third of the flanks and uncoiling slightly at the end of the body chamber. The whorl cross section is slightly wider than high, with a rather curved umbilical wall in the inner whorls, becoming more vertical with a rounded edge. At the end of the body chamber the umbilical wall becomes rounded and the section of the whorls becomes subcircular.
In
the innermost observable whorls, the ornamentation, clearly visible on specimen
brg094 (Pl. 6 ![]() , fig. 1), is composed of 8 large peri-umbilical tubercles per
whorl. From a whorl-height of 21 mm, the tubercles become prominent, gradually
taking on a triangular shape and somewhat overhanging the umbilical wall. They
are then separated by relatively wide inter-tubercular spaces. This type of
ornamentation continues on a whorl until the end of the phragmocone (H
= 72 mm on specimen brg094); there are 11 tubercles on this whorl but a decrease
in tubercles strength is noted on the last quarter. On the first part of the
body chamber, the tubercles are replaced by large peri-umbilical bullae, which
regularly give rise to 2 ribs with rounded sections, thickening in the ventral
region. There are a few rare isolated intercalatory ribs emerging at various
heights on the flanks. On the last third of the body chamber, only simple, thick
ribs with rounded sections are observed. These cross the venter in their maximum
thickness.
, fig. 1), is composed of 8 large peri-umbilical tubercles per
whorl. From a whorl-height of 21 mm, the tubercles become prominent, gradually
taking on a triangular shape and somewhat overhanging the umbilical wall. They
are then separated by relatively wide inter-tubercular spaces. This type of
ornamentation continues on a whorl until the end of the phragmocone (H
= 72 mm on specimen brg094); there are 11 tubercles on this whorl but a decrease
in tubercles strength is noted on the last quarter. On the first part of the
body chamber, the tubercles are replaced by large peri-umbilical bullae, which
regularly give rise to 2 ribs with rounded sections, thickening in the ventral
region. There are a few rare isolated intercalatory ribs emerging at various
heights on the flanks. On the last third of the body chamber, only simple, thick
ribs with rounded sections are observed. These cross the venter in their maximum
thickness.
Poor preservation inhibited detailed analysis of the suture pattern.
Discussion: These two specimens are distinguishable from the specimens of Roloboceras hambrovi (Forbes, 1845) described above by their very prominent tubercles, overhanging the umbilical wall, which are separated by wide inter-tubercular spaces on the inner whorls, a higher ribbing density, and both a more evolute shell and compressed whorl section. In addition, these specimens were collected in beds 7 and 10 of Malaubert section, which are located in the upper half of the Violette Member. They thus occupy an older stratigraphic position in the Roloboceras hambrovi Subzone than the material collected in Le Teil and Viviers sections from the Rocherenard Member (Pictet & Delanoy, 2017).
Stratigraphic and geographical distribution: Roloboceras aff. hambrovi (Forbes, 1845) was collected in the upper half of the Violette Member, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone of the Malaubert section, Pierrelatte (Drôme, S-E of France).
Roloboceras aff. hispanicum Sornay, 1972, form A
(Pl. 4 ![]() , fig. 2a-b;
Pl. 5
, fig. 2a-b;
Pl. 5 ![]() ,
fig. 3a-b; Table 4)
,
fig. 3a-b; Table 4)
Material studied (N = 2): specimens tl002 coll. Baudouin, tl070 coll. Boselli, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche).
Measurements (in mm):
Table 4: Dimensions of Roloboceras aff. hispanicum Sornay, 1972, form A.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl070 | 170 | 60 | 80 | 115 | - | - | 0.35 | 0.47 | 0.68 | 1.44 |
| tl002 | 157 | 56 | 61 | - | - | - | 0.36 | 0.39 | - | - |
| 129 | 43 | 53 | 87 | - | 4 | 0.33 | 0.41 | 0.67 | 1.64 |
Description: Medium-sized
ammonites, showing almost two whorls of spire, the body chamber occupying about
¾ of the last whorl. The whorl section is strongly domed to subogival with a
wide latero-ventral region. The umbilical wall is subvertical and smooth. In the
preserved internal whorls, corresponding to the last whorl of the phragmocone,
the tubercles are large, low, more or less wide, rounded, from 6 to 9 in number,
separated by narrow intertubercular spaces. This type of ornamentation continues
over the first half of the body chamber and then changes suddenly; the tubercles
are then diminished in both width and thickness and seem to stretch radially
into bullae. The latero-ventral ribbing is hardly visible on both specimens due
to poor preservation. Indistinct bundles of 2-3 ribs, rounded, broad and low,
arising from the umbilical tubercles are visible on some parts of the last half
whorl of specimen tl002 (Pl. 4 ![]() , fig. 2a-b). On specimen tl070
(Pl. 5
, fig. 2a-b). On specimen tl070
(Pl. 5 ![]() , fig.
3a-b), the end of the body chamber shows thick, simple primary ribs and
intercalatory ribs which appear on the lower third of the flanks. All these ribs
seem to cross the ventral region.
, fig.
3a-b), the end of the body chamber shows thick, simple primary ribs and
intercalatory ribs which appear on the lower third of the flanks. All these ribs
seem to cross the ventral region.
Poor preservation inhibited detailed analysis of the suture pattern.
Discussion: These two specimens are comparable to the medium-sized specimens studied in the population of Roloboceras hambrovi (Forbes, 1845) and described above (almost complete specimens, of medium size with a phragmocone diameter of around 100 mm). However, they differ from R. hambrovi by their very wide and flattened tubercles on the phragmocone and at the beginning of the body chamber.
Morphological
and ornamental affinities with R.
hispanicum Sornay, 1972, are certain, as shown by comparison with the
holotype of the Spanish species (Pl. 5 ![]() , figs. 1a-b, 2a-b). They are
characterised by the width and low relief of the tubercles on the internal
whorls, with a sudden decrease in the strength of the tubercles from the lower
third of the body chamber. The weakness of the latero-ventral ornamentation,
another characteristic of R. hispanicum, is visibly exaggerated by an erosion of this part of
the shell on both specimens.
, figs. 1a-b, 2a-b). They are
characterised by the width and low relief of the tubercles on the internal
whorls, with a sudden decrease in the strength of the tubercles from the lower
third of the body chamber. The weakness of the latero-ventral ornamentation,
another characteristic of R. hispanicum, is visibly exaggerated by an erosion of this part of
the shell on both specimens.
In view of the observations noted on the population of R. hambrovi described above, in particular those concerning the presence of "hispanicum" type tubercles in some specimens, the absence of the innermost whorls and the poor preservation of the latero-ventral region in R. aff. hispanicum form A, it is undesirable to consider these two specimens as original representatives of R. hispanicum. They could be extreme morphotypes of the medium-sized population of R. hambrovi in which tuberculation of the "hispanicum" type is highly expressed. It therefore appears increasingly important to clarify relationships between these two species.
The
specimen cited as Cheloniceras aff. hambrovi
by Sornay (1958, p. 170) (Pl. 10 ![]() , fig. 2a-b) can probably be related to Roloboceras
aff. hispanicum.
, fig. 2a-b) can probably be related to Roloboceras
aff. hispanicum.
Stratigraphic and geographical distribution: Roloboceras aff. hispanicum Sornay, 1972, form A comes from the yellow marly limestone (Rouvière beds in Pictet & Delanoy, 2017) of Le Teil (Ardèche), reported from the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone. Specimen tl002 was collected from the base of these marly limestones.
Roloboceras aff. hispanicum Sornay, 1972, form B
Studied material (N = 1): Specimen MGL.104623 coll. A. Pictet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Labastide-de-Virac section (Ardèche).
Measurements (in mm):
Table 5: Measurements of Roloboceras aff. hispanicum Sornay, 1972, form B.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| MGL. 104623 | 101 | 32.5 | 37.5 | 53.7 | - | - | 0.32 | 0.37 | 0.53 | 1.43 |
Description: Small-sized and poorly preserved ammonite. The section of the whorls is subogival with a high, rounded umbilical wall.
The internal whorls are very poorly preserved. On the last whorl, there are large and wide peri-umbilical tubercles, 7 in number, separated by deep intervals. The ribbing is almost indistinguishable, except for the last quarter whorl where it seems to be composed of very wide and thick ribs.
Discussion: This specimen shows some affinities with Roloboceras hispanicum Sornay, 1972, in particular with the specimen CPT-3234 from the section of Barranco de las Calzadas figured by Moreno-Bedmar and Bover (2007) and Moreno-Bedmar et al. (2009, Fig. 10F; 2010, Fig. IIIA) by the strength and size of its tubercles. It also closely resembles R. horridum (Spath, 1930) by its large periumbilical tubercles, but the latter differs in its very wide and rounded ventral region and particularly strong ribbing.
Stratigraphic and geographical distribution: Roloboceras aff. hispanicum Sornay, 1972, form B was collected from the Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone of the Labastide-de-Virac section (Ardèche). It is sourced from the Motier Beds, in the basal part of the Violette Member and is in fact the oldest known representative of Roloboceras Casey, 1954, in the sections of the Languedoc platform.
Genus Megatyloceras Humphrey, 1949
Megatyloceras was introduced by Humphrey in 1949 (p. 149) for ammonites of the group of Ammonites hambrovi Forbes, 1845, characterized by a very low coronatiform section, a deep umbilicus and strongly prominent lateral tubercles from which 2-3 strong ribs arise.
Type-species: Douvilleiceras coronatum Rouchadzé, 1933, by original designation of Humphrey, 1949, p. 149.
Diagnosis: Ammonites attaining a large size characterized by an extremely wide and strongly depressed coronate section in the internal whorls, a very deep umbilicus with a high, very broad and rounded ventral region, a subvertical and smooth umbilical wall, and an ornamentation made of strong primary and secondary ribs (Humphrey, 1949). Intercalatory ribs are few, irregularly arranged, and arise close to the tubercles. Simplified suture line with a large external saddle.
The ontogenic development of Megatyloceras Humphrey, 1949, differs from that of Roloboceras Casey, 1954, by a significantly reduced Royerianum stage. The succession of ontogenic stages is as follows: 1) Embryonic stage; 2) juvenile stage with a coronatiform morphology with low whorls and a wide, smooth latero-ventral region bordered by numerous small periumbilical tubercles which can be observed very early (D = 3 mm); 3) stage with the discrete appearance of ventral ribs, which originate singly and then by 2 (3?) from tubercles; all the ornamentation become gradually stronger. The episodic presence at this stage of a constriction between two ribs recalls the Royerianum stage observed in Roloboceras; 4) stage with a very coronatiform section and a pronounced wide ventro-lateral region, with considerable development of more or less conical spinous peri-umbilical tubercles and presence of a ventral ribbing; 5) stage with strong periumbilical tubercles more or less pinched and with strong ventral ribbing; 6) adult stage with peri-umbilical bullae disappearing on the body-chamber, which is similar to that of Roloboceras.
Geographic and
stratigraphic distribution: The
genus occurs in France, England, Spain, Bulgaria, Romania and Georgia (Fig. 5 ![]() ).
Its stratigraphic distribution seems to be restricted to the Lower Aptian, Deshayesites
forbesi Zone, upper part of the Roloboceras
hambrovi Subzone
).
Its stratigraphic distribution seems to be restricted to the Lower Aptian, Deshayesites
forbesi Zone, upper part of the Roloboceras
hambrovi Subzone
Systematic of the genus: Humphrey (1949) assigned to this new genus the Cheloniceratidae Spath, 1923, Megatyloceras casei Humphrey, 1949, Ammonites ricordeanus Orbigny, 1850, Cheloniceras hambrovi var. horrida Spath, 1930, ?C. crassum Spath, 1930, and Douvilleiceras coronatum Rouchadzé, 1933. In 1961, in his monograph on Lower Greensand ammonites, Casey erected the subfamily Roloboceratinae for grouping the two genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949, within family Douvilleiceratidae Parona & Bonarelli, 1897. He also introduced a new species of Megatyloceras, M. vastum sp. nov. Casey (p. 190) questions the occurrence of the genus in the Anglo-Paris Basin and Georgia and its apparent absence in the Tethys regions. However, later work will show that the genus Megatyloceras is well represented in these regions, whereas new taxa were described in Bulgaria (Dimitrova, 1967), Romania (Chiriac, 1981) and Spain (Sornay & Marin, 1972). Casey (1961, p. 190) expressed doubt about the generic assignment of the late Aptian species Megatyloceras casei, later reconsidered as belonging to Epicheloniceras Casey, 1954, by Moreno-Bedmar and Delanoy, 2013.
After a thorough literature survey, we consider that Megatyloceras is composed of the following taxa:
? Megatyloceras benardeus (Buvignier, 1852)
Megatyloceras bonsthevi Dimitrova, 1967
Megatyloceras coronatum (Rouchadzé, 1933)
Megatyloceras georgiense Casey, 1954
Megatyloceras ibericum Sornay, 1972
Megatyloceras persianense Pauliuc & Grădinaru, 1970
Megatyloceras ramadanicum (Chiriac, 1981)
Megatyloceras ricordeanum (Orbigny, 1850)
Megatyloceras transiens (Casey, 1961)
Megatyloceras vastum Casey, 1961
To which we add:
Megatyloceras leteilensis sp. nov. (this work).
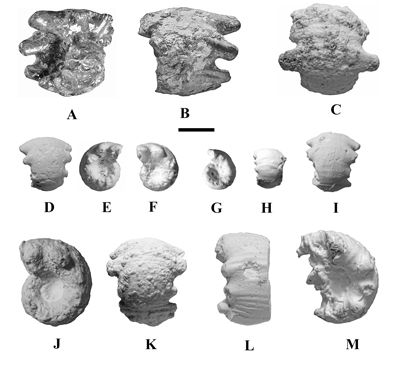
Megatyloceras ricordeanum (Orbigny, 1850)
(Figs. 9A-M ![]() ,
10K
,
10K ![]() ;
Pl. 14
;
Pl. 14 ![]() , fig. 3a-d; Table 6)
, fig. 3a-d; Table 6)
Synonymy:
1850a. Ammonites Ricordeanus sp. nov. - Orbigny, p. 113, no. 21.
1850b. Ammonites Ricordeanus d'Orbigny - Orbigny, p. 199, Pl. 8, figs. 5-8.
? 1852. Ammonites benardeus sp. nov. - Buvignier, p. 46, Pl. XXXI, figs. 10-12.
1949. Megatyloceras ricordeanum d'Orbigny - Humphrey, p. 149.
1961. Megatyloceras ricordeanum (d'Orbigny) - Casey, p. 192, Fig. 58f-g.
1970. Megatyloceras ricordeanum (d'Orbigny) - Pauliuc & Grădinaru, p. 23, Fig. 1A.
1996. Megatyloceras ricordeanum (d'Orbigny) - Wright et al., Fig. 207: 1b.
2004b. Megatyloceras ricordeanum (d'Orbigny) - Amédro & Matrion, p. 79, Pl. 1, fig. 8a-b.
non 2008a. Megatyloceras ricordeanum (d'Orbigny) - Ropolo et al., p. 28, Pl. 8, fig. 3 (= Megatyloceras aff. coronatum form A, see Discussion), Pl. 9, fig. 4 (= Megatyloceras cf. coronatum), Pl. 11, fig. 3 (= Cheloniceras sp.).
non 2009b. Megatyloceras ricordeanum (d'Orbigny) - Ropolo et al., p. 150, Pl. 8, fig. 3 (= Cheloniceras gr. cornuelianum Orbigny), Pl. 9, fig. 4 (= Megatyloceras cf. coronatum), Pl. 11, fig. 3 (= Cheloniceras sp.).
? 2013. Megatyloceras ricordeanum (d'Orbigny) - Ivanov & Idakieva, Fig. 14F.
2015. Megatyloceras ricordeanum (d'Orbigny) - Bersac & Bert, Pl. 5, fig. 9.
2018. Megatyloceras ricordeanum (d'Orbigny) - Bulot et al., Fig. 3 (1).
Lectotype:
We designate as lectotype of Ammonites
ricordeanum Orbigny, 1850, the syntype MNHNF.A25561 (coll. Orbigny),
from Gurgy (Yonne, France) (Fig. 9A-C ![]() ;
Pl. 14
;
Pl. 14 ![]() , fig. 3a-d) and stored in the collections of the Museum of Natural History in
Paris.
, fig. 3a-d) and stored in the collections of the Museum of Natural History in
Paris.
|
Figure 9:
Juvenile whorls (nuclei) of some specimens of Megatyloceras
ricordeanum (Orbigny, 1850) from the Argiles à Plicatules Formation: A-B: lectotype, MNHNF.A25561; C: MHNG GEPI
17455b; D-F: MHNG GEPI 17957; G-H:
unregistered specimen coll. AGA; I:
unregistered specimen coll. AGA; J-K:
MHNG GEPI 17453a; L-M: MHNG GEPI
17455a. Specimens C, D-F, G-H, I, J-K and L-M are plaster casts. See text for
more details. Scale bar is 1 cm. |
Studied material (N = 15): Lectotype MNHN.F.A25561 coll. Orbigny, casts of topotypes MHNG 17453a-c and 17454b-c coll. F.J. Pictet, topotypes 91-1091 (4 specimens) coll. Muséum d'Histoire Naturelle d'Auxerre, Lower Aptian, Argiles à Plicatules, Gurgy (Yonne); casts of specimens MHNG 17455a-c coll. F.J. Pictet, Lower Aptian, Argiles à Plicatules, Montiéramey (Aube); cast of specimen MHNG 17457 coll. F. J. Pictet, Lower Aptian, Argiles à Plicatules, Ervy (Aube); specimen 69-1091, coll. Muséum d'Histoire Naturelle d'Auxerre, Lower Aptian, Argiles à Plicatules, locality unspecified.
Measurements (in mm):
Table 6: Measurements of Megatyloceras ricordeanum (Orbigny, 1850).
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| MHNG 17455a | 27 | c7.5 | c12 | - | - | - | 0.27 | 0.44 | - | - |
| MHNG 17455b | 26 | c10 | c10 | C19 | - | - | 0.38 | 0.38 | 0.76 | 1.9 |
| Lectotype MNHN.F.A25561 | 24.6 | 7.6 | 8.5 | 17.7 | - | - | 0.30 | 0.34 | 0.71 | 2.0 |
| MHNG 17453a | 24.5 | 8 | 10.5 | 17 | - | - | 0.32 | 0.42 | 0.69 | 1.61 |
| MHNG 17454b | 22 | 8.3 | 8 | c17 | - | - | 0.37 | 0.36 | 0.77 | 2.12 |
| MHNG 17454c | 18 | 6 | 8 | 13 | - | - | 0.33 | 0.44 | 0.72 | 1.62 |
| MHNG 17457 | 14.5 | 5 | 5.5 | 11 | 0.34 | 0.37 | 0.75 | 2 | ||
| MHNG 17455c | 13 | 4 | 5.8 | c10.5 | 030 | 0.44 | 0.80 | 1.81 | ||
| MHNG 17453c | 12 | 4.5 | 4 | c8 | - | - | 0.37 | 0.33 | 0.66 | 2 |
| MHNG 17453b | 11.5 | 4.5 | 4 | c9 | - | - | 0.39 | 0.34 | 0.78 | 2.25 |
Description: The lectotype is a fairly well-preserved pyrite nucleus. It has undergone a resin treatment that slightly hinders observational analysis. The shell is coronatiform with a very low whorl section; the flanks and the ventral region are no longer separable, forming a large lateroventral area. Up to H = 4.5 mm, the ornamentation consists of simple ribs, visible on the umbilical wall, which bear a relatively long conical tubercle at the edge of the periumbilical region. There are then nine ribs per half whorl. The ventral ribs figured by Orbigny (1850b), derived in bundles from periumbilical tubercles, and are not visible probably because of the preservation of the specimen. From H = 4.5 mm, the tubercles develop very strongly. They are then proportionally very long (7 mm), somewhat flattened in the coiling direction and have a wide base, 6 to 7 mm on the last tubercles. Their tips are rounded. They give rise to 2-3 ribs of very low relief crossing the extensively wide lateroventral area, which is slightly convex. On the last half whorl, there are four tubercles arising from a barely visible bulge on the umbilical wall, which at this stage appears completely smooth.
Discussion: In addition to the lectotype of Megatyloceras ricordeanum (Orbigny, 1850), we were able to examine a small population only represented by more or less well-preserved pyritic nuclei depending on the state of degradation of the pyrite. The majority of these nuclei are fairly consistent with the description of Orbigny (1850b). Nevertheless, the best-preserved specimens show discrete ribbing on the ventral area. Some show that tubercle strength increases quite abruptly from D = 10 mm when H = 4-4.5 mm. Specimen MHNG 17455a reveals the presence of ventral ribs and some clearly visible intercalatory ribs on the umbilical wall.
The taxon was defined from a pyritic nucleus 24.7 mm in diameter characterized by exaggerated periumbilical tubercles in regard to the corresponding diameter and undoubtedly representing the innermost whorls of a large ammonite whose adult stage remains hitherto unknown. Although partially preserved, this particular ornamental stage has also been detected in the type material of Megatyloceras vastum Casey, 1961, and M. transiens (Casey, 1961) (see infra) and well-observed in M. coronatum (Rouchadzé, 1933), taxa which are distinguished from each other by different ontogenetic sequences at later growth stages. It appears, therefore, that the only ornamental stage characterizing M. ricordeanum cannot be considered as representative of the species. Due to the scarcity of complete material both from the type locality and environs, we consider M. ricordeanum as the expression of an ontogenic stage common to several species of Megatyloceras Humphrey, 1949, which we refer to here as the Ricordeanum stage. For this reason, we consider M. ricordeanum as nomen dubium.
Ammonites benardeus Buvignier, 1852, is founded on a 16 mm diameter nucleus collected in the Argiles à Plicatules of Sermaize (Marne). Buvignier, who refers only to the description without figuration of the Prodrome (Orbigny, 1850a), relates this new species closer to M. ricordeanum from which it would differ by the presence of well individualized ribs on the ventral region, which, however, are present on illustrated specimens of M. ricordeanum. In the absence of type-material and in view of the uncertainties concerning the morphology and ornamentation of this species, it seems reasonable to consider also A. benardeus as a nomen dubium.
In the literature, the citations and figurations of M. ricordeanum are always associated with specimens whose determinations are doubtful or erroneous and cannot be taken into consideration to represent the taxon ricordeanum.
Thus, M. ricordeanum was reported from the Lower Greensand, near Atherfield Point, in the Deshayesites forbesi Zone, D. annelidus Subzone, represented by a body chamber and by pyritic nuclei of uncertain specific determination (Casey et al., 1998, p. 516 and 532).
The presence of M. ricordeanum in the Lower Aptian series of Bulgaria (Ivanov & Idakieva, 2013) is based on a fragmentary and distorted specimen. Ropolo et al. (2008a, 2009b) have described and figured three specimens from La Bédoule as Megatyloceras ricordeanum but none of them can be assigned to the Orbigny's species. The specimen of Pl. 8, fig. 3 in Ropolo et al. (2008a) is Megatyloceras aff. coronatum form A, specimen tl014, described in this work but figured by mistake in Ropolo et al. (2008a) (see Ropolo et al., 2008a - erratum). In Ropolo et al. (2009b) the same Pl. 8, fig. 3, PRA1424 corresponds to a nucleus of Cheloniceras gr. cornuelianum (Orbigny, 1841) with an erroneous scale; PRA1425, Pl. 11, fig. 3 is also a Cheloniceras Hyatt, 1903, which is also refigured under this name Pl. 16, fig. 1. Finally, only one specimen (C.865, Pl. 9, fig. 4) corresponds to a Megatyloceras showing some affinities with M. coronatum. In the same publications, Ropolo et al. (2008a, 2009b) considered the Bulgarian species M. bontchevi Dimitrova, 1967, as a synonym of M. ricordeanum, which cannot be retained in view of the conclusions of this study.
Stratigraphic distribution: The lectotype of Megatyloceras ricordeanum (Orbigny, 1850) was collected in the Lower Aptian of the Argiles à Plicatules of the Paris Basin without any biostratigraphic precision. According to Bersac and Bert (2012, 2015), M. ricordeanum would be present in the Deshayesites deshayesi zone, D. deshayesi Subzone of the Paris Basin. However, it should be pointed out that the authors consider Deshayesites annelidus Casey, 1961, as a minor synonym of D. deshayesi (Orbigny, 1841), leading to a new conception of the Deshayesites deshayesi Zone which then includes de facto the Deshayesites annelidus Subzone sensu Casey et al. (1998).
|
Figure 10:
Sections of some Megatyloceras Humphrey,
1949. The arrow indicates sequelae left by an injury. Scale bar is 5 cm. |
Megatyloceras coronatum (Rouchadzé, 1933)
(Figs. 10D-E, J ![]() ,
11A
,
11A ![]() ;
Pl. 12
;
Pl. 12 ![]() , fig. 2;
Pl. 13
, fig. 2;
Pl. 13 ![]() , figs. 1a-b, 2a-c;
Pl. 14
, figs. 1a-b, 2a-c;
Pl. 14 ![]() , figs. 1a-c, 2a-c;
, figs. 1a-c, 2a-c;
Pl. 17 ![]() , figs.
1a-b, 2a-b; Pl. 18
, figs.
1a-b, 2a-b; Pl. 18 ![]() , fig. 1a-b;
Pl. 19
, fig. 1a-b;
Pl. 19 ![]() , fig. 1a-b;
Pl. 20
, fig. 1a-b;
Pl. 20 ![]() , fig. 1a-b;
, fig. 1a-b;
Pl. 21 ![]() ,
fig. 1a-c; Pl. 25
,
fig. 1a-c; Pl. 25 ![]() , fig. 1a-b;
Pl. 26
, fig. 1a-b;
Pl. 26 ![]() , fig. 2a-b;
Pl. 27
, fig. 2a-b;
Pl. 27 ![]() , fig. 1a-c;
, fig. 1a-c;
Pl. 28 ![]() , fig.
1; Pl. 29
, fig.
1; Pl. 29 ![]() , fig. 1; Table 7)
, fig. 1; Table 7)
Synonymy:
? 1927. Douvilleiceras cf. hambrovi (Forbes) - Roch, p. 22, Pl. I, fig. 3.
1933. Douvilleiceras coronatum sp. nov. - Rouchadzé, p. 195, Pl. III, fig. 4, Fig. 12, 13.
1938. Douvilleiceras coronatum Rouchadzé - Rouchadzé, p.178, Pl. I, fig. 7, Pl. II, fig. 1, Figs. 13-14.
1954. Megatyloceras georgiense nom. nov. - Casey, p. 114.
1961. Megatyloceras georgiense Casey - Casey, p.192, Fig. 58d-e.
1961. Megatyloceras coronatum (Rouchadzé) - Casey, p. 192, Fig. 58c.
? 1967. Megatyloceras bontshevi sp. nov. - Dimitrova, p. 167, Pl. LXXXV, fig. 1, Pl. LXXXVI, fig. 3.
1970. Megatyloceras coronatum (Rouchadzé) - Pauliuc & Grădinaru, p. 23, Fig. 1b, 2A.
1970. Megatyloceras georgiense Casey - Pauliuc & Grădinaru, p. 23, Fig.1c, 2B.
1970. Megatyloceras persaniense sp. nov. - Pauliuc & Grădinaru, p. 32, Pls. I-V, Fig. 1e, 2Da-b.
non 1972. Megatyloceras coronatum ibericum nov. ssp. - Sornay in Sornay & Marin, p. 109(11), Pl. B, figs. 1-2, Pl. D, fig. 4; Fig. 8 [= Megatyloceras ibericum Sornay, 1972 (see infra)].
? 1983. Megatyloceras coronatum ibericum Sornay - Murat, p. 58, Pl. VIII, fig. 2.
1996. Megatyloceras coronatum (Rouchadzé) - Wright et al., p. 267, Fig. 207: 1a.
? 2005. Megatyloceras coronatum (Rouchadzé) - Sharikadze & Kvantaliani in Kotetishvili et al., p. 386, Pl. 95, fig. 3a-б.
? 2007. Megatyloceras coronatum (Rouchadzé) - Moreno-Bedmar et al., Pl. 1, fig. 7.
? 2009. Megatyloceras coronatum (Rouchadzé) - Moreno-Bedmar et al., Fig. 10B-D, G.
? 2010. Megatyloceras coronatum (Rouchadzé) - Moreno-Bedmar et al., electronic appendix, Fig. IVB-D.
non 2012. Megatyloceras coronatum (Rouchadzé) - Moreno-Bedmar et al., Fig. 7H (= ? Megatyloceras sp.).
Lectotype:
When designating Douvilleiceras coronatum Rouchadzé, 1933, as type-species of Megatyloceras,
Humphrey (1949, p. 149) referred to the species described and figured by Rouchadzé
in 1933 (p. 195, Pl. III, fig. 4; Figs. 12-13) in a monograph on Aptian
ammonites of western Georgia. The only specimen figured in this work by Rouchadzé
came from the Bayern collection and had been collected in Kharagoüli, Horecha,
Georgia. It is a very incomplete specimen, the internal whorls being only very
partially preserved, and in a poor state of preservation (Pl. 13 ![]() , fig. 1a-b).
However, Rouchadzé specified that the description of the new species was
also based on two other specimens from the Gamkrélidzé collection, also
collected in Horécha, but then considered as lost (Rouchadzé, 1933,
footnote, p. 197). In addition, Rouchadzé did not designate a holotype
of the species coronatum.
, fig. 1a-b).
However, Rouchadzé specified that the description of the new species was
also based on two other specimens from the Gamkrélidzé collection, also
collected in Horécha, but then considered as lost (Rouchadzé, 1933,
footnote, p. 197). In addition, Rouchadzé did not designate a holotype
of the species coronatum.
A
few years later, in a complementary work, Rouchadzé (1938, p. 178)
indicated that specimens of Gamkrélidzé coll. had been recovered and he
figured in Pl. I, fig. 7 and Pl. II, fig. 1 the larger of the two specimens (Pl. 14 ![]() , fig. 1a-c). The latter is an ammonite with a diameter of about 200 mm in
good state of preservation which perfectly illustrates the succession of
ornamental stages described by Rouchadzé in 1933 in the original
description of the species, from the Ricordeanum growth stage to the stage with
large radially pinched tubercles, present at the end of the preserved whorls.
These same pinched tubercles are present at the beginning of the whorls of the
specimen illustrated in 1933, which effectively confers conspecificity on these
two specimens.
, fig. 1a-c). The latter is an ammonite with a diameter of about 200 mm in
good state of preservation which perfectly illustrates the succession of
ornamental stages described by Rouchadzé in 1933 in the original
description of the species, from the Ricordeanum growth stage to the stage with
large radially pinched tubercles, present at the end of the preserved whorls.
These same pinched tubercles are present at the beginning of the whorls of the
specimen illustrated in 1933, which effectively confers conspecificity on these
two specimens.
However,
in 1954, Casey (p. 114) considered that the specimen figured by Rouchadzé
in 1938 represented a new taxon which he named Megatyloceras
georgiense nom. nov. (Fig. 11B ![]() ). A few years later, Casey (1961, p.
192) had considered also the specimen figured by Rouchadzé in 1933, Pl.
3, fig. 4, as the "holotype" of the species M.
coronatum, thus conferring on this specimen the status of lectotype (Art.
74.5, ICZN Code, lectotype designation before 2000): Pl. 13
). A few years later, Casey (1961, p.
192) had considered also the specimen figured by Rouchadzé in 1933, Pl.
3, fig. 4, as the "holotype" of the species M.
coronatum, thus conferring on this specimen the status of lectotype (Art.
74.5, ICZN Code, lectotype designation before 2000): Pl. 13 ![]() ,
fig. 1a-b.
,
fig. 1a-b.
Note: Klein and Bogdanova (2013, p. 6) erroneously indicated that the specimen figured by Rouchadzé in 1933 (Pl. III, fig. 4) had been designated as the lectotype of the species by Humphrey in 1949 (Klein, personal communication).
Studied material (N = 20): Specimens tl009, tl010, tl011, tl013, tl016, tl058, tl062, tl068 coll. Baudouin, tl060, tl061 coll. Boselli, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche); specimens chb004, chb005 coll. Baudouin, VI002, VI003, VI005, VI007, VI008 coll. Boselli, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Rocherenard, Viviers (Ardèche); specimen MHNG GEPI 82461 coll. A. Pictet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Bourg-Saint-Andéol (Ardèche); specimen FSL 89026 coll. Busnardo, "Bédoulien supérieur", Rocherenard, Viviers (Ardèche).
Measurements (in mm):
Table 7: Measurements of Megatyloceras coronatum (Rouchadzé, 1933).
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| chb004 | 417 | 140 | 155 | 182 | - | 0 | 0.34 | 0.37 | 0.44 | 1.17 |
| VI005 | 280 | 100 | 100 | 135 | 16 | 0 | 0.36 | 0.36 | 0.48 | 1.35 |
| VI002 | 275 | 102 | 114 | 120 | - | 0 | 0.37 | 0.41 | 0.44 | 1.05 |
| tl010 | 256 | 92 | 101 | - | 20 | 1 | 0.36 | 0.39 | - | - |
| 190 | 69 | 85 | 114 | 21 | 3 | 0.36 | 0.45 | 0.60 | 1.34 | |
| tl011 | 245 | 86 | 101 | 142 | 17 | 0 | 0.35 | 0.41 | 0.58 | 1.41 |
| c157 | 54 | 74 | 112 | - | 4 | 0.34 | 0.47 | 0.71 | 1.51 | |
| tl058 | 245 | 87 | 90 | c125 | 18 | 3 | 0.36 | 0.37 | 0.51 | 1.39 |
| 178 | 61 | 71 | 105 | 20 | 5 | 0.34 | 0.40 | 0.59 | 1.48 | |
| tl009 | 223 | 84 | 78 | 90 | 18 | 0 | 0.38 | 0.35 | 0.40 | 1.15 |
| 153 | c62 | 69 | 74 | - | 4 | 0.41 | 0.45 | 0.48 | 1.07 | |
| tl062 | c215 | 81 | 81 | - | - | - | 0.38 | 0.38 | - | - |
| 135 | 47 | 45 | 74 | 17 | 6 | 0.35 | 0.33 | 0.55 | 1.64 | |
| 104 | 43 | 34 | 62 | 17 | 5 | 0.41 | 0.33 | 0.60 | 1.82 | |
| VI003 | 210 | 71 | 77 | 113 | 17 | 0 | 0.34 | 0.37 | 0.54 | 1.47 |
| chb005 | 199 | 69 | 79 | - | 23 | 4 | 0.35 | 0.40 | - | - |
| 136 | 48 | 54 | 78 | - | 4 | 0.35 | 0.40 | 0.57 | 1.44 | |
| tl060 | 189 | 65 | 80 | 102 | 23 | 1 | 0.34 | 0.42 | 0.54 | 1.28 |
| VI008 | 195 | 71 | 81 | 105 | - | 1 | 0.36 | 0.42 | 0.54 | 1.30 |
| MHNGGEPI 82461 | 182 | 64.5 | 64 | - | - | - | 0.35 | 0.35 | - | - |
| FSL 89026 | 172.6 | 59.4 | 65.5 | c72 | - | - | 0.34 | 0.38 | 0.42 | 1.10 |
| tl068 | 165 | 57 | 55 | - | 15 | 7 | 0.35 | 0.33 | - | - |
| 128 | 49 | 54 | 90 | 19 | 6 | 0.38 | 0.42 | 0.70 | 1.67 | |
| VI007 | 159 | 55 | 63 | - | - | - | 0.35 | 0.40 | - | - |
| tl061 | 157 | 58 | c55 | 98 | 16 | 0 | 0.37 | 0.35 | 0.62 | 1.78 |
| tl016 | 150 | 54 | 57 | c108 | - | - | 0.36 | 0.38 | 0.72 | 1.89 |
| 111 | 41 | 30 | - | 23 | 5 | 0.37 | 0.27 | - | - | |
| tl013 | 77 | c28 | c26 | c55 | 21 | 5 | 0.36 | 0.34 | 0.71 | 2.12 |
Description: The
species reaches a large to very large size (VI005, Pl. 20 ![]() , fig. 1, D
= 280 mm; chb004, Pl. 28
, fig. 1, D
= 280 mm; chb004, Pl. 28 ![]() , D = 417 mm).
The coiling is evolute, with whorls clearly wider than high, depressed in the
inner whorls, with a subcircular section during the adult stage. The umbilical
wall is high, subvertical in the inner whorls and rounded on the body chamber.
Four ontogenic stages can be distinguished, passing gradually from one to the
other:
, D = 417 mm).
The coiling is evolute, with whorls clearly wider than high, depressed in the
inner whorls, with a subcircular section during the adult stage. The umbilical
wall is high, subvertical in the inner whorls and rounded on the body chamber.
Four ontogenic stages can be distinguished, passing gradually from one to the
other:
a first Ricordeanum stage, up to about D = 35 mm, where the whorl section is very low and where the ornamentation consists of very strong peri-umbilical tubercles that are high, sub-conical, flattened on their umbilical side, attached to the umbilical wall of the next whorl, in the number of 7 to 10 per whorl;
a stage with highly conical, more or less developed tubercles,
approximately present from D = 35 mm
to D = 70-80 mm. Depending on the
specimens, the strength and number of tubercles is very variable, and even they
may be highly developed (VI002, Pl. 19 ![]() , fig. 2);
, fig. 2);
a more or less expressed stage, with radially pinched tubercles, of variable strength. On these first three stages, the tubercles give rise to groups of 3, more rarely 4, rounded broad ribs, which cross the latero-ventral region with their maximum thickness. During the pinched tubercle stage, a few intercalatory ribs are also observed emerging on or near the umbilical margin;
a stage where the tubercles develop into periumbilical bullae and then disappear
more or less distinctly and rapidly to give way to simple or bifurcated ribs in
the periumbilical area. This stage corresponds to the adult stage. The number of
ribs from the periumbilical bullae gradually decreases to two, then one rib at
the end of the adult body chamber. The presence of irregularly arranged
intercalatory ribs is observed, one to three per interval depending on the
specimens and appearing at varying heights on the flanks. They cross the ventral
region, identical to the main ribs. Their frequency decreases at the end of the
body chamber of adult specimens. On the largest specimen collected (chb004, D = 417 mm,
Pl. 28 ![]() ), the ribbing becomes evanescent on the body
chamber.
), the ribbing becomes evanescent on the body
chamber.
Poor preservation inhibited detailed analysis of the suture pattern.
|
Figure 11:
Whorl section of: A - Megatyloceras
coronatum (Rouchadzé, 1933); B
- M. georgiense Casey, 1954, and C - M. persaniense Pauliuc
& Grădinaru, 1970. According Rouchadzé, 1933 (A),
1938 (B), Casey (1961) (A-B),
Pauliuc & Grădinaru
(1970) (A-C).
Scale bar is 5 cm. |
Variability: The
population studied here is well individualized by a characteristic succession of
four ornamental stages. It nevertheless shows a relatively high variability (Fig. 12 ![]() ), especially in the duration and the morphological expression of these stages
as in the strength of the conical or/and the pinched tubercules which can be
very developed, with the exception of the Ricordeanum stage which is relatively
constant in its characteristics (duration and number of tubercles).
), especially in the duration and the morphological expression of these stages
as in the strength of the conical or/and the pinched tubercules which can be
very developed, with the exception of the Ricordeanum stage which is relatively
constant in its characteristics (duration and number of tubercles).
Nevertheless, a small number of the specimens originally studied show an early appearance of the adult stage. In fact, with the exception of the Ricordeanum stage, the ornamental stages recognized among M. coronatum (Rouchadzé, 1933) are much shorter and the adult size appears smaller. They are described as M. aff. coronatum form A and considered as possible microconchs forms (see infra).
|
Figure 12:
Megatyloceras Humphrey,
1949: diameter variation of the
umbilicus (U),
width (W) and height of the whorls (H) as a function of
the diameter (D)
of the population studied and the types of the literature. |
Discussion: Specimens from Ardèche perfectly show the succession of the ornamental stages (Ricordeanum stage, conical tubercles stage and finally the laterally pinched tubercles stage) observable on the lectotype of M. coronatum (Rouchadzé, 1933). The Ricordeanum stage is clearly visible within the internal whorls of some of these specimens and shows great morphological and ornamental similarities with the lectotype of M. ricordeanum (Orbigny, 1850) which is only a nucleus of 24.7 mm in diameter whose complete ontogenetic development remains unknown. Synonymy of M. coronatum with M. ricordeanum based solely on this single similarity of the comparable internal whorls cannot be envisaged because a similar stage to the Ricordeanum stage is partially observable during the internal whorls of M. transiens (Casey, 1961) and M. vastum Casey, 1961, and the complete ontogenetic development of M. ricordeanum remains unknown.
Examination
of the specimen illustrated by Roch (1927, Pl. I, fig. 3) under Douvilleiceras
cf. hambrovi (Forbes, 1845)
suggests that it represents a Megatyloceras
Humphrey, 1949, most probably conspecific with M.
coronatum as evidenced by the presence of large radially pinched tubercles
at the beginning of the last whorl (Pl. 30 ![]() , fig. 1a-b).
, fig. 1a-b).
M. persaniense Pauliuc & Grădinaru,
1970 (Fig. 11C ![]() ), shows the same succession of ontogenic stages as those
observed in M. coronatum, including a
Ricordeanum stage and a stage with clearly visible laterally pinched tubercles.
The poor preservation of the holotype restricts clear observation of the
intermediate conical tubercles, but this seems to be confirmed in the original
description (Pauliuc & Grădinaru,
1970, p. 33). We
therefore consider it a synonym of M.
coronatum.
), shows the same succession of ontogenic stages as those
observed in M. coronatum, including a
Ricordeanum stage and a stage with clearly visible laterally pinched tubercles.
The poor preservation of the holotype restricts clear observation of the
intermediate conical tubercles, but this seems to be confirmed in the original
description (Pauliuc & Grădinaru,
1970, p. 33). We
therefore consider it a synonym of M.
coronatum.
M. vastum
(Fig. 10B ![]() ;
Pl. 15
;
Pl. 15 ![]() , fig. 1a-e)
is clearly distinguished from M. coronatum
by the presence in the innermost whorls, and after the Ricordeanum stage, of
alternating relatively thick single ribs (two per interval) and relatively
spaced strong, tall tubercles of more or less oval cross-section in the
direction of coiling. This ornamentation disappears towards H
= 6 mm and it then gives way to tubercles pinched in the direction of coiling,
to the number of four per whorl, which tend to become conical at the end of the
last preserved whorl.
, fig. 1a-e)
is clearly distinguished from M. coronatum
by the presence in the innermost whorls, and after the Ricordeanum stage, of
alternating relatively thick single ribs (two per interval) and relatively
spaced strong, tall tubercles of more or less oval cross-section in the
direction of coiling. This ornamentation disappears towards H
= 6 mm and it then gives way to tubercles pinched in the direction of coiling,
to the number of four per whorl, which tend to become conical at the end of the
last preserved whorl.
M. transiens
(Fig. 10A ![]() ;
Pl. 16
;
Pl. 16 ![]() , fig. 1a-d) shows internal whorls similar to those of M.
vastum, from which it differs by the appearance of a stage with big, radially
oriented and relatively pinched periumbilical tubercles.
, fig. 1a-d) shows internal whorls similar to those of M.
vastum, from which it differs by the appearance of a stage with big, radially
oriented and relatively pinched periumbilical tubercles.
M. ramadanicum
(Chiriac, 1981) (Fig. 10G ![]() ;
Pl. 12
;
Pl. 12 ![]() , fig. 1a-c)
shows some ornamental and morphological affinities with M. coronatum but the holotype (by
monotypy) of the Romanian taxon
shows notable ornamental differences and it is too incomplete to be considered
synonymous with the Rouchadzé species. The internal whorls of the
specimen described by Chiriac (1981), a fully septate
phragmocone, are
not visible. The last preserved whorl first shows conical tubercles, more or
less pinched in the direction of coiling to about 110 mm diameter (H
= 36 mm). This is soon followed by a slightly conical to nodule-like tubercles
stage, somewhat rursiradiate, rapidly decreasing in strength, which seems to
announce the adult stage. The stage with radially pinched and very developed
tubercles, well individualized in M.
coronatum, is here absent.
, fig. 1a-c)
shows some ornamental and morphological affinities with M. coronatum but the holotype (by
monotypy) of the Romanian taxon
shows notable ornamental differences and it is too incomplete to be considered
synonymous with the Rouchadzé species. The internal whorls of the
specimen described by Chiriac (1981), a fully septate
phragmocone, are
not visible. The last preserved whorl first shows conical tubercles, more or
less pinched in the direction of coiling to about 110 mm diameter (H
= 36 mm). This is soon followed by a slightly conical to nodule-like tubercles
stage, somewhat rursiradiate, rapidly decreasing in strength, which seems to
announce the adult stage. The stage with radially pinched and very developed
tubercles, well individualized in M.
coronatum, is here absent.
M. coronatum ibericum Sornay, 1972
(Pl. 24 ![]() , fig. 3a-c), should be considered as a separate species (M.
ibericum), characterized by its small adult size and its tubercles pinched
obliquely to the direction of coiling. Unfortunately, the holotype of this taxon
is also incomplete, represented by a single whorl including the beginning of the
adult body chamber, and whose internal whorls are not preserved.
, fig. 3a-c), should be considered as a separate species (M.
ibericum), characterized by its small adult size and its tubercles pinched
obliquely to the direction of coiling. Unfortunately, the holotype of this taxon
is also incomplete, represented by a single whorl including the beginning of the
adult body chamber, and whose internal whorls are not preserved.
M. bontshevi Dimitrova,
1967 (Pl. 25 ![]() , fig. 2a-c), is characterized by its ornamentation consisting of
"highly conical" tubercles (Dimitrova,
1967, p. 230). The holotype of
this species corresponds to a phragmocone whose innermost whorls are not
preserved and whose ornamentation of strong conical tubercles which extend over
the lateroventral region in bundles of three thick ribs, shows on little more
than one and a half whorls. The stage with large radially pinched tubercles,
present in M. coronatum, is not
observable, which prevents consideration of the Bulgarian
species as a possible synonym for M. coronatum. As pointed out previously
(see supra), synonymy of M.
bontshevi with M. ricordeanum as
proposed by Ropolo et al.
(2008a, 2009b) is implausible.
, fig. 2a-c), is characterized by its ornamentation consisting of
"highly conical" tubercles (Dimitrova,
1967, p. 230). The holotype of
this species corresponds to a phragmocone whose innermost whorls are not
preserved and whose ornamentation of strong conical tubercles which extend over
the lateroventral region in bundles of three thick ribs, shows on little more
than one and a half whorls. The stage with large radially pinched tubercles,
present in M. coronatum, is not
observable, which prevents consideration of the Bulgarian
species as a possible synonym for M. coronatum. As pointed out previously
(see supra), synonymy of M.
bontshevi with M. ricordeanum as
proposed by Ropolo et al.
(2008a, 2009b) is implausible.
Stratigraphic and geographical distribution: Megatyloceras coronatum (Rouchadzé, 1933) has a fairly wide geographical distribution within the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone of south-eastern France (Ardèche), Spain (unpublished data from J.A. Moreno-Bedmar), Bulgaria, and in the "Lower Aptian" of Romania and Georgia.
Casey et al. (1998, p. 516) reported the presence of forms close to M. georgiense Casey, 1954 (= M. coronatum herein) in the Upper Lobster Beds of the Isle of Wight, but the absence of figuration prevents further confirmation.
Ropolo et al. (2008a, p. 28; 2009b, p. 107) also reveal the presence of Megatyloceras aff. coronatum in the Deshayesites deshayesi Zone of the section of the historical stratotype of Cassis - Roquefort-La Bédoule but the description, without figuration, of a single incomplete specimen does not confirm the presence of the species in this area.
Most of the specimens from Ardèche come from the Rouvière Marls, reported from the Lower Aptian, Deshayesites forbesi Zone, top of the Roloboceras hambrovi Subzone from La Rouvière, Le Teil (Ardèche) and from the locality of Rocherenard, Viviers (Ardèche).
Megatyloceras aff. coronatum (Rouchadzé, 1933) form A
[? = M. coronatum (Rouchadzé, 1933) microconch form]
(Fig. 10I ![]() ;
Pl. 24
;
Pl. 24 ![]() , figs. 1a-c, 2a-c; Table 8)
, figs. 1a-c, 2a-c; Table 8)
Studied material (N = 2): Specimens tl014, tl015 coll. Baudouin, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche).
Measurements (in mm):
Table 8: Measurements of Megatyloceras aff. coronatum (Rouchadzé, 1933) form A.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl015 | 123 | 43 | c44 | c81 | 19 | 5 | 0.35 | 0.36 | 0.66 | 1.84 |
| tl014 | 105 | 37 | 43 | 75 | 21 | 6 | 0.35 | 0.41 | 0.71 | 1.74 |
| 72 | 28 | 28 | 56 | 20 | 4 | 0.39 | 0.39 | 0.78 | 2.00 |
Description: The description is mainly based on the best-preserved specimen tl014. It is a small-sized ammonite of good preservation, showing two whorls of the spire. The innermost whorls are not observable. The whorl section is low, very depressed, with a subvertical, slightly convex umbilical wall. The ornamentation is strong. In the preserved inner whorls, strong periumbilical tubercles, four in number, are first observed, pinched in the coiling direction and stuck to the next whorl, characteristic of the Ricordeanum stage. From about H = 9 mm, these tubercles become stronger and tend to become conical. There are eight tubercles in the observed inner whorl. On the outer whorl, the periumbilical tubercles are long, very prominent, pinched in the coiling direction, almost in the extension of the lateroventral region which is very wide and slightly convex. From these tubercles, arise bundles of three to four rounded ribs, crossing the lateroventral region. Single ribs are irregularly present between the tubercles, with one or two ribs per interval. At the end of the external whorl, the tubercles become more massive and thicker, less prominent, pinched radially and originating from the base of the umbilical wall. These tubercles, which appear to announce the adult stage, give rise to two or three thick ribs, rounded in cross-section and crossing the lateroventral region. There are nine tubercles on the last whorl.
Poor preservation inhibited detailed analysis of the suture pattern.
Discussion: These ammonites show a succession of ornamental stages quite similar to that observed in M. coronatum (Rouchadzé, 1933), viz. the Ricordeanum stage, the conical tubercle stage, the pinched tubercle stage and finally the bullae stage. The difference with the previously described forms of M. coronatum lies in the fact that, with the exception of the Ricordeanum stage, these stages are here much shorter and the adult size appears smaller.
Morphological
and ornamental similarities exist with Megatyloceras
ibericum Sornay, 1972, although the holotype of this species is incomplete and the internal
whorls are not observable (Pl. 24 ![]() , fig. 3a-c). It has, on the preserved whorl,
a tuberculation similar to that of specimen tl014, which consists in seven
large, well-individualized tubercles (nine in the original description) pinched
subradially, with slightly prorsiradiate projection. From these tubercles appear
bundles of three or four thick rounded ribs, which radially cross the wide
lateroventral region. These ribs are only visible on the first quarter of the
preserved whorl where the lateroventral region is smooth. Between each of the
first three tubercles, a single rib is inserted, which originates on the
umbilical wall and crosses the latero-ventral region. Examination of the
holotype also revealed the presence of body chamber, very partially preserved at
the end of the preserved whorl. The sutures lines, clearly visible, show a clear
approximation of the last septa, thus testifying to the adult age of this ammonite. The
type-specimen of this taxon is nevertheless too incomplete (absence of internal
whorls) to allows conspecificity with the forms described just above.
, fig. 3a-c). It has, on the preserved whorl,
a tuberculation similar to that of specimen tl014, which consists in seven
large, well-individualized tubercles (nine in the original description) pinched
subradially, with slightly prorsiradiate projection. From these tubercles appear
bundles of three or four thick rounded ribs, which radially cross the wide
lateroventral region. These ribs are only visible on the first quarter of the
preserved whorl where the lateroventral region is smooth. Between each of the
first three tubercles, a single rib is inserted, which originates on the
umbilical wall and crosses the latero-ventral region. Examination of the
holotype also revealed the presence of body chamber, very partially preserved at
the end of the preserved whorl. The sutures lines, clearly visible, show a clear
approximation of the last septa, thus testifying to the adult age of this ammonite. The
type-specimen of this taxon is nevertheless too incomplete (absence of internal
whorls) to allows conspecificity with the forms described just above.
These ammonites present similar morphologies probably corresponding to relatively small adult forms as shown by the close approximation of the sutures in M. ibericum and by the appearance of the adult ornamental stage in specimen tl014 whose sutures are unfortunately not discernible. This same earlier adult-stage appearance is also observed in Megatyloceras sp. and M. leteilensis sp. nov. (see infra). The existence of these morphologies can be interpreted either as the simple expression of a very strong variability in adult size, or as the expression of a dimorphism, probably of a sexual nature, within Megatyloceras Humphrey, 1949. The similarities observed in the ornamental stages and their succession between the specimen tl014 and the larger forms attributed to M. coronatum and the similar observations made within Roloboceras hambrovi (Forbes, 1845) rather argue in favour of the latter hypothesis; however, in Megatyloceras the change of ornamentation is more brutal and therefore more noticeable.
Finally,
it should be noted that one of the syntypes of M.
coronatum, the specimen 21/1221 figured by Sharikadze and Kvantaliani
(in Kotetishvili et
al., 2005, Pl. 95, fig. 3a-b; figured here Pl. 13 ![]() , fig. 2a-c) shows
ornamental and morphological similarities with the two specimens described and
discussed above. However, only a visual examination could confirm the
conspecificity of this specimen with M.
aff. coronatum form A.
, fig. 2a-c) shows
ornamental and morphological similarities with the two specimens described and
discussed above. However, only a visual examination could confirm the
conspecificity of this specimen with M.
aff. coronatum form A.
Stratigraphic and geographical distribution: Megatyloceras aff. coronatum (Rouchadzé, 1933) form A was collected in the Rouvière Marls, reported from the Lower Aptian, Deshayesites forbesi Zone, top of the Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche).
Megatyloceras aff. coronatum (Rouchadzé, 1933) form B
Studied material (N = 1): Specimen MGL.104620 coll. A. Pictet, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Picourel section, Vagnas (Ardèche).
Measurements (in mm):
Table 9: Measurement of Megatyloceras aff. coronatum (Rouchadzé, 1933) form B.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| MGL.104620 | 160 | 63 | 51 | - | - | - | 0.39 | 0.32 | - | - |
Description: Medium-sized ammonite of rather poor preservation, represented by two whorls of spire. The innermost whorls are not preserved. At the beginning of the last whorl, the section appears relatively depressed, and then gradually changes to become subcircular on the body chamber.
In the preserved inner whorl, three to four tubercles more or less pinched in the coiling direction can be observed, probably stuck to the umbilical wall of the next whorls and corresponding to the Ricordeanum stage. This stage is quickly replaced by a stage with six to seven very large tubercles, at first rather nodular to slightly conical, round-based, then radially pinched, very thick and oval-based. These tubercles are still present at the beginning of the last whorl where they give rise to three thick ribs which cross the wide ventral region in their maximum thickness. On the last half whorl, probably partly corresponding to the body chamber, the tubercles change into large periumbilical bullae and then disappear.
Discussion: Megatyloceras aff. coronatum (Rouchadzé, 1933) form B shows ornamental and morphological affinities with the Georgian species, especially by the succession of ornamental stages which seem however to follow one another more rapidly. The absence of the most internal whorls, the state of preservation of the specimen and its presence in older stratigraphic levels (upper part of the Violette Member) than those of the other Megatyloceras Humphrey, 1949, studied in this work prevent us from including it with certainty in M. coronatum.
Stratigraphic and geographical distribution: Megatyloceras aff. coronatum (Rouchadzé, 1933) form B is only known in the extreme summit of the Violette Member, reported from the Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Picourel (Ardèche).
Megatyloceras transiens (Casey, 1961)
(Fig. 10A ![]() ;
Pl. 16
;
Pl. 16 ![]() , fig. 1a-d; Table 10)
, fig. 1a-d; Table 10)
Synonymy:
1961. Roloboceras (?) transiens sp. nov. - Casey, p. 182, Fig. 56.
non 1972. Roloboceras sp. ex gr. transiens Casey - Sornay in Sornay & Marin, p. 108(10), Pl. C, fig. 4 (=? Megatyloceras sp.).
non 1975. Roloboceras sp. gr. transiens Casey - Conte, p. 106, Figs. 2-3.(= Roloboceras sp.)
1998. Megatyloceras transiens (Casey) - Casey et al., p. 531.
? 2007. Roloboceras transiens Casey - Moreno-Bedmar et al., Pl. 1, figs. 14-15.
non 2008a. Roloboceras sp. gr. transiens Casey - Ropolo et al., p. 27, Pl. 8, fig. 2.
non 2009b. Roloboceras sp. gr. transiens Casey - Ropolo et al., p. 148, Pl. 8, fig. 2.
? 2013. Roloboceras transiens Casey - Ivanov & Idakieva, Fig. 14G-H.
2015. Megatyloceras transiens (Casey) - Bersac & Bert, p. 25, Pl. 5, fig. 10.
Holotype:
By original designation, the specimen MNHNP.F.A25552 (ex 5596a) coll. Orbigny,
Aptian, Gurgy (Yonne), stored in the collection of the Museum of Natural History
in Paris:
Pl. 16 ![]() , fig.
1a-d.
, fig.
1a-d.
Measurements (in mm):
Table 10: Measurement of Megatyloceras transiens (Casey, 1961).
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| MNHNP.F.A25552 | 153 | 52.2 | 62.2 | C110 | - | - | 0.34 | 0.39 | 1.71 | 1.76 |
Description: The specimen corresponds to a fully septate phragmocone of almost two whorls. However, there is a slight approximation of the suture lines at the end of the last whorl. The general shape of the shell is globular. The section is depressed, with a very wide lateroventral region. The umbilical wall is subvertical.
In the innermost whorls, the ornamentation is incompletely preserved. It is partially observable from a whorl height of about 7 mm. One then observes spaced tubercles (two visible tubercles but up to a number of four tubercles per half-whorl can be envisaged) which appear strongly pinched in the direction of coiling with a rounded top, more or less clearly separated from the umbilical wall of the next whorl. The ornamentation between these tubercles is not visible. Nevertheless, this short ornamental sequence suggests the presence of a Ricordeanum stage, of which only the very last developments would be observable. Between H = 9 mm and H = 12 mm the tubercles thicken strongly, become sub-conical while stretching radially. Between these tubercles, intermediary ribs appear, subangular in section, two per interval, elevating slightly on the flanks. Between H = 12 mm and H = 17 mm the specimen is badly preserved and the ornamentation, although not easily discernible, appears more uniform, probably consisting of ribs and/or tubercles that are less differentiated than in the previous ornamental sequence.
On the first half of the last whorl, the section is strongly depressed with a deep umbilicus and a smooth wall between the rather discrete umbilical ribs. The periumbilical tubercles, originating from the discrete umbilical ribs are very strong, raised and pinched radially on the umbilical margin, giving rise to two or three thick rounded secondaries, crossing the wide ventrolateral region. The central rib of these bundles is often the thickest. Thinner intermediate ribs are more or less evenly arranged between these rib bundles; they arise approximately at the division points of the periumbilical tubercles. In the second half of the last whorl, the tubercles tend to develop into large, thick, wedge-shaped bullae, some of which are prominently raised, announcing the adult stage.
The sutures lines are clearly visible on the last whorl and tend to get closer at the end of this whorl, indicating that this specimen has reached adult development.
Discussion: Although Megatyloceras transiens (Casey, 1961) is absent from the Ardèche deposits, it seemed essential to describe this taxon because it remains poorly described in the literature and, like M. vastum Casey, 1961, it has ornamental features that are important for the understanding of Megatyloceras Humphrey, 1949. The species was introduced without description by Casey (1961, p. 182, Fig. 56) who subsequently considered it as a Roloboceras Casey, 1954, forming the transition toward Megatyloceras. It was only much later that Casey et al. (1998) recognized it as a representative of Megatyloceras. The ornamentation of the internal whorls and of the beginning of the last whorl, as well as the section of the same part of the shell on the holotype, confirm this last systematic option.
Although the holotype of Megatyloceras transiens shows very partially the end of a Ricordeanum stage and originates from the same locality (Gurgy, Yonne) as M. ricordeanum (Orbigny, 1850), it can hardly be interpreted as representing the later growth stage of the species of Orbigny (and therefore a synonym of this one), for several reasons:
M. ricordeanum is only known from numerous pyritic nuclei collected at various locations in the Paris Basin (Gurgy, Ervy, Montieramey, see supra) and the largest specimen studied reaches 27 mm in diameter. Casey (1961, p. 192) writes that he observed a topotype of about 40 mm diameter in the Sorbonne collections; unfortunately it was not possible to find this ammonite in the collections now held at the Université Pierre et Marie Curie (Paris, France);
the Ricordeanum stage is present in the internal whorls of several taxa of Megatyloceras as M. transiens, M. coronatum (Rouchadzé, 1933), M. vastum, M. persaniense Pauliuc & Grădinaru, 1970;
M. transiens, on the other hand, is only known from a single specimen preserved as a calcareous phosphatized mould whose Ricordeanum stage, incompletely preserved, has a maximum estimated diameter of about 35 mm. The conservation of M. transiens is moreover quite similar to that of the holotype and several topotypes of Heminautilus lallierianus (Orbigny, 1841) recently studied (Baudouin et al., 2016), which, like M. transiens, all came from Gurgy. These very different preservations suggest the existence of a distinct lithological level of origin in this locality (see "Distribution of Roloboceratinae and palaeoenvironment" paragraph).
Megatyloceras transiens can be distinguished from M. coronatum mainly by the presence in the inner whorls of an alternation of tuberculate ribs and non tuberculate ribs, by the absence of the conical tuberculate stage, by a stage with very radially pinched tubercles, with a tendency to become cuneiform as the adult stage approaches. A Ricordeanum stage, or similar, is present in the whorls as evidenced by the morphology of the first tubercles visible on the specimen.
Megatyloceras vastum
(Pl. 15 ![]() , fig. 1a-e) shows internal whorls with an ornamentation quite similar to
those of M. transiens. A stage similar to the Ricordeanum stage is observed
up to about H = 4-5 mm with tubercles
pinched in the direction of coiling. This stage is then followed by an
alternation of less pinched tubercles and relatively thick, angular-sectioned
intercalatory ribs. Towards H = 6-7 mm
the tubercles become very large, they are pinched and stretched in the coiling
direction, spaced, with an oval basal section; there are two per half whorl. The
intercalory ribs have disappeared.
, fig. 1a-e) shows internal whorls with an ornamentation quite similar to
those of M. transiens. A stage similar to the Ricordeanum stage is observed
up to about H = 4-5 mm with tubercles
pinched in the direction of coiling. This stage is then followed by an
alternation of less pinched tubercles and relatively thick, angular-sectioned
intercalatory ribs. Towards H = 6-7 mm
the tubercles become very large, they are pinched and stretched in the coiling
direction, spaced, with an oval basal section; there are two per half whorl. The
intercalory ribs have disappeared.
Stratigraphic and geographical distribution: Megatyloceras transiens (Casey, 1961) is only formally known in the formation des Argiles à Plicatules of Gurgy (Yonne, France) by its holotype. The specimens from Spain and Bulgaria are too incomplete for positive identification. Casey et al. (1998, p. 516) report its occurrence in the Upper Lobster Beds of the section located east of Atherfield Point (Isle of Wight, UK) corresponding to the Deshayesites forbesi Zone, D. annelidus Subzone. However, in the same publication Casey et al. (1998, p. 531) specified that this specimen is represented by in fact an eroded whorl fragment, which may also belong to a sister species. Bersac and Bert (2015) located M. transiens in the Deshayesites deshayesi Zone which, according to their interpretation (Bersac & Bert, 2012, 2015) would be equivalent to the Casey's D. annelidus Subzone (see M. ricordeanum (Orbigny, 1850), supra).
Megatyloceras leteilensis sp. nov.
(Fig. 10F ![]() ;
Pl. 23
;
Pl. 23 ![]() , figs. 1a-c, 2a-d; Table 11)
, figs. 1a-c, 2a-d; Table 11)
Holotype: Specimen
tl001 coll. Baudouin, Lower Aptian, Deshayesites
forbesi Zone, Roloboceras hambrovi
Subzone, la Rouvière, Le Teil (Ardèche):
Pl. 23 ![]() ,
fig. 1a-c.
,
fig. 1a-c.
Locus typicus: La Rouvière, Le Teil (Ardèche).
Stratum typicum: Rouvière Beds, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone.
Derivation nominis: From the town of Le Teil in Ardèche.
Studied material (N = 2): Specimens tl001, tl082 coll. Baudouin, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche). Cast of the holotype and the paratype are stored in the Requien Museum (Avignon, France) with the no.2021.11.1 and 2021.11.2. The original specimens will also be deposited at a later date in the same institution.
Measurements (in mm):
Table 11: Measurements of Megatyloceras leteilensis sp. nov.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl001 | 151 | 44 | 59 | c82 | 24 | 5 | 0.29 | 0.39 | 0.54 | 1.39 |
| 104 | 32 | 45 | 74 | 26 | 4 | 0.31 | 0.43 | 0.71 | 1.64 | |
| c70 | - | 29 | 58 | - | - | - | 0.41 | 0.83 | 2.00 | |
| tl082 | 137 | 46 | 57 | 84 | 20 | 3 | 0.34 | 0.42 | 0.61 | 1.47 |
| 94 | c30 | 43 | 64 | - | 4 | 0.32 | 0.46 | 0.68 | 1.49 | |
| - | - | 25 | 56 | - | - | - | - | - | 2.24 |
Diagnosis: Medium-sized Megatyloceras Humphrey, 1949, whose phragmocone ornamentation is composed of spaced, conical and relatively small tubercles, which changes abruptly on the body chamber, showing ornamented, dense ribbing, arising from small umbilical bullae.
Description: Medium-sized ammonites, showing a moderately evolute coiling. The whorls overlap at mid-flank on the phragmocone and on the first part of the body chamber, which uncoil slightly towards the aperture on the holotype. The section of the whorls is depressed on the phragmocone (W/H = 2 on the holotype at D = about 70 mm), wider than high with a smooth, slightly convex subvertical umbilical wall and a wide rounded lateroventral region.
On the first preserved whorl, which corresponds to the phragmocone, the ornamentation consists of large, round and slightly conical, spaced, peri-umbilical tubercles, eight in number, with relatively wide and smooth inter-tubercular spaces. From these tubercles emanate bundles of four or, more rarely, three rounded ribs of low relief, crossing the ventral region, more or less curved proversly; an intercalatory rib is sometimes observed between two ribs bundles. At the end of this whorl (H = 45 mm, D = 104 mm on the holotype) the ornamentation changes rather abruptly, announcing the adult body chamber: the periumbilical tubercles change into radially stretched periumbilical bullae, quite close together. These somewhat elevated bullae give rise to a thick rounded rib that frequently divides towards the lower third of the flanks to give rise to a somewhat thinner secondary rib. Sometimes, a third rib of variable strength appears between the bullae. This type of ornamentation is particularly visible on the holotype. All these ribs cross the ventral region in their maximum thickness. This type of ornamentation continues over the body chamber.
Poor preservation inhibited detailed analysis of the suture pattern.
Discussion: Megatyloceras leteilensis sp. nov. differs from M. coronatum (Rouchadzé, 1933) by its fewer and much less developed tubercles on inner whorls, the lack of the radially pinched tubercles stage and a much smaller adult size.
Megatyloceras leteilensis sp. nov. shows affinities with M. vastum Casey, 1961, by its low number of tubercles, with wide intertubercular spaces. It differs from that species by a very different shape of its tubercles: in Casey's species the tubercles are fewer, very wide, large and stretched in the coiling direction, whereas they are less developed and more conical in M. leteilensis sp. nov. The ribbing is also much more vigorous in M. vastum, a species known only by an incomplete phragmocone.
Megatyloceras leteilensis sp. nov. is, by its internal whorls with spaced tubercles, close to the specimens of M. aff. leteilensis sp. nov. studied in this work. It differs by a very fast change in ornamentation accompanied by the disappearance of tubercles on the last quarter whorl indicating the adult body-chamber. In M. aff. leteilensis sp. nov. the tubercles are larger on the internal whorls, more pinched in the direction of coiling, the adult size is larger and shows different body-chamber ornamentation, which is vigorously ornamented showing thick ribs which cross the venter in their maximum thickness.
Stratigraphic and geographical distribution: Megatyloceras leteilensis sp. nov. is sourced uniquely from the Rouvière Beds, reported from the Lower Aptian, Deshayesites forbesi Zone, top of the Roloboceras hambrovi Subzone from la Rouvière, Le Teil (Ardèche).
Megatyloceras aff. leteilensis sp. nov.
[? = M. leteilensis sp. nov. macroconch form]
(Fig. 10C ![]() ;
Pl. 21
;
Pl. 21 ![]() , fig. 2a-b;
Pl. 22
, fig. 2a-b;
Pl. 22 ![]() , fig. 1a-b; Table 12)
, fig. 1a-b; Table 12)
Studied material (N = 2): Specimens tl007, tl083 coll. Baudouin, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche).
Measurements (in mm):
Table 12: Measurements of Megatyloceras aff. leteilensis sp. nov.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl007 | 179 | 67 | 76 | 100 | - | - | 0.37 | 0.42 | 0.56 | 1.32 |
| 138 | 48 | 58 | 90 | - | 5 | 0.35 | 0.42 | 0.65 | 1.55 | |
| tl083 | 132 | 46 | 59 | c91 | - | 4 | 0.35 | 0.45 | 0.69 | 1.54 |
| 91 | 34 | 35 | 71 | - | 5 | 0.37 | 0.38 | 0.78 | 2.03 |
Description: Large ammonites with a depressed whorl section, a very wide and convex lateroventral region, a deep umbilicus and a slightly convex, subvertical to gently sloping, smooth umbilical wall. On the largest specimen (tl007) the incomplete body chamber occupies the last quarter of whorl from a diameter of about 150 mm.
In the innermost preserved whorls, the ornamentation comprises spaced peri-umbilical tubercles, at first slightly stretched in the coiling direction, leaning against the umbilical wall of the next whorl, and then departing from it, five or six per whorl and separated by wide intertubercular spaces. Around 70 mm in diameter tubercles gradually become more conical, the umbilical face of the tubercles is flattened, lying in line with the umbilical wall. Then, towards a diameter of about 130 mm they become pinched radially and gradually turn into a bullae at the end of the preserved whorl. On the last whorl, the peri-umbilical tubercles give rise to bundles of 3 thick rounded ribs, which cross the flanks and the venter in their maximum strength.
Discussion: These two specimens show affinities with Megatyloceras vastum Casey, 1961, by the presence of a stage with relatively low tubercles somewhat stretched in the coiling direction, with wide intertubercular spaces. They are distinguished by slightly more numerous tubercles and much less vigorous ribbing. Because the internal whorls of the Ardèche specimens are not preserved, it is not possible to go any further with this comparison.
Megatyloceras aff. leteilensis sp. nov. can be distinguished from M. coronatum (Rouchadzé, 1933) by the more widely spaced, weaker and fewer internal tubercles, which are more stretched in the direction of coiling and by the absence of the radially pinched tubercles stage. In addition, the innermost whorls have not been observed in M. aff. leteilensis sp. nov. due to the poor preservation of the specimens.
M. aff. leteilensis sp. nov. shows some phragmoconic ornamental affinities with M. leteilensis sp. nov., with few, widely spaced conical tubercles, however differentiated by showing greater stretching in the direction of coiling. A larger adult size is also definitive, certainly approaching, or perhaps, exceeding 250 mm, contrasting with an estimated 100-?160 mm adult size for M. leteilensis sp. nov. (holotype shows initiation of body chamber at approximately 100 mm). Perhaps the morphological and ornamental affinities of the visible internal whorls of Megatyloceras aff. leteilensis sp. nov. and M. leteilensis sp. nov. as well as the size difference in the appearance of their adult stages could be the expression of a dimorphism of sexual origin. Thus M. aff. leteilensis sp. nov. would be the macroconch form, whereas M. leteilensis sp. nov., the suspected microconch. Other small micromorphs such as Megatyloceras aff. coronatum form A, Megatyloceras sp. and M. ibericum Sornay, 1972, are other suspected microconch megatyloceratid candidates.
Stratigraphic and geographical distribution: Megatyloceras aff. leteilensis sp. nov. is only known in the Rouvière Beds, reported from the Lower Aptian, Deshayesites forbesi Zone, top of the Roloboceras hambrovi Subzone, from la Rouvière, Le Teil (Ardèche).
Megatyloceras sp.
(Fig. 10H ![]() ;
Pl. 22
;
Pl. 22 ![]() , figs. 2a-c, 3a-b; Table 13)
, figs. 2a-c, 3a-b; Table 13)
Studied material (N = 2): Specimens tl071 coll. Delanoy, tl073 coll. Baudouin, Lower Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, la Rouvière, Le Teil (Ardèche).
Measurements (in mm):
Table 13: Measurements of Megatyloceras sp.
| No. | D | U | H | W | Nc/2 | Nt/2 | U/D | H/D | W/D | W/H |
| tl071 | 116 | 41 | 50 | 88 | 17 | 7 | 0.35 | 0.43 | 0.76 | 1.76 |
| - | - | 18 | 51 | - | - | - | - | - | 2.83 | |
| tl073 | c73 | c30 | 24 | 47 | 9 | 6 | 0.41 | 0.33 | 0.64 | 1.96 |
| - | - | 16 | 42 | - | - | - | - | - | 2.63 |
Description: The best-preserved specimen (tl071) is a small ammonite, whose internal whorls are not preserved. The last preserved whorl shows a depressed section, with a high umbilical wall, first slightly inclined and then subvertical and weakly convex, and a very wide and rounded lateroventral region.
On specimen tl071, ornamentation visible on the last whorl, is first composed of five conical tubercles, thorn-like and high, with an oval base. These tubercles then become distinctly more radially pinched and yet remain strong until the last preserved quarter whorl where the tuberculation loses its vigour and appears to announce the adult stage. It is not excluded that the last quarter whorl may be the beginning of the body chamber. All tubercles originate from a low relief bulge that runs through the umbilical wall. There are 12 tubercles on the preserved whorl. All these tubercles give rise to bundles of ribs in irregular numbers, from between two to four. These ribs are rounded and cross the wide lateroventral region in their maximum thickness.
Specimen tl073, which is very fragmentary, shows an ornamentation consisting of very spinous and high tubercles, slightly pinched radially, originating at the base of the umbilical wall and continuing over the ventral region in what appears to be a single broad, low rib. It cannot be excluded that the poor preservation of the ventral region does not allow the distinction of thinner secondary ribs.
Poor preservation inhibited detailed analysis of the suture pattern.
Discussion: The most complete specimen (tl071) appears to be a relatively small Megatyloceras Humphrey, 1949, with a strongly depressed section and showing the early appearance of very pinched tubercles indicating an early adult stage. By these characteristics it shows affinities with M. aff. coronatum (Rouchadzé, 1933) form A, M. ibericum Sornay, 1972, and M. leteilensis sp. nov. (see supra) for which the hypothesis that they could either represent a group of small species or perhaps microconch forms related to sexual dimorphism has been considered (see M. aff. coronatum form A, supra). Megatyloceras sp. thus appears as an additional argument in favour of this hypothesis. However, the absence of the internal whorls prohibits a comparison with one of the known species of Megatyloceras.
Megatyloceras sp. differs, however, from M. aff. coronatum form A and M. ibericum by its more numerous tubercles on the last whorl and its strongly spinous tubercles on (all ?) the phragmocone.
Stratigraphic and geographical distribution: Megatyloceras sp. is only known in the Rouvière Beds, reported from the Lower Aptian, Deshayesites forbesi Zone, top of the Roloboceras hambrovi Subzone, from la Rouvière, Le Teil (Ardèche).
Distribution of Roloboceratinae and palaeoenvironment: Roloboceratinae are characterized by coarsely ribbed coronate shells, quite similar to the Jurassic genus Gravesia Salfeld, 1913. Both genera flourished on the slope of carbonate platforms. This similarity in lifestyle induced identical environmental constraints, which can explain the similarity in the strongly ornamented coronate shape of the shell in both genera.
From
a stratigraphic and palaeoenvironmental point of view, the rich fauna of
Roloboceratinae collected in the Chabert Formation of the Ardèche border is
mainly distributed over five sections, all located on the median to external
ramp (Fig. 13 ![]() ). Within these five sections, the stratigraphic distribution of
Roloboceratinae shows a heterogeneity within the Deshayesites forbesi
Zone with a progressive shift of their occurrence from the Violette Member for
the more coastal sections of Picourel and Labastide de Virac, at the top of the
Violette Member and the base of the Lower Rocherenard Beds for the deeper sector
of Bourg-St-Andéol, in the Lower Rocherenard Beds and the Rouvière Beds of the
distal sector of Chabert, and finally exclusively in the Rouvière Beds of the
circalittoral section of La Rouvière. The absence of Roloboceratinae on the
innermost ramp (Michelet and Mezelet sections) as well as in the bathyal facies
of the outer ramp (i.e. Violette Member except Motier Beds, Chabert and La Rouvière
sections), and the progressive shift in the occurrence of Roloboceratinae within
the lithostratigraphic units of the Chabert Formation is remarkable and clearly
controlled by the bathymetric interval (Fig. 13
). Within these five sections, the stratigraphic distribution of
Roloboceratinae shows a heterogeneity within the Deshayesites forbesi
Zone with a progressive shift of their occurrence from the Violette Member for
the more coastal sections of Picourel and Labastide de Virac, at the top of the
Violette Member and the base of the Lower Rocherenard Beds for the deeper sector
of Bourg-St-Andéol, in the Lower Rocherenard Beds and the Rouvière Beds of the
distal sector of Chabert, and finally exclusively in the Rouvière Beds of the
circalittoral section of La Rouvière. The absence of Roloboceratinae on the
innermost ramp (Michelet and Mezelet sections) as well as in the bathyal facies
of the outer ramp (i.e. Violette Member except Motier Beds, Chabert and La Rouvière
sections), and the progressive shift in the occurrence of Roloboceratinae within
the lithostratigraphic units of the Chabert Formation is remarkable and clearly
controlled by the bathymetric interval (Fig. 13 ![]() ). Roloboceratinae thus seems to
favor a specific facies zone, or bathymetric interval, as suggested by Moreno-Bedmar
et al. (2009), neither too shallow nor
too deep. This specific facies zone seems to be characterised by fine-grained
marly-calcareous to calcareous facies whose fauna seems to be dominated by the
presence of numerous sponges, irregular sea urchins (Toxasteridae) and small
benthic foraminifera and, conversely, by the absence of neritic fauna such as
regular sea urchins, crinoid meadows, and bryozoans or pelagic microfauna. This
distribution thus seems to be reduced to microfacies F1 to F3 of the Arnaud-Vanneau
classification (Arnaud-Vanneau, 1980,
2005), corresponding to the aphotic
zone of the slopes. The possible dependence of Roloboceratinae on certain
biotopes was mentioned by Ropolo et
al. (2008b) to explain the apparent diachronism of Roloboceratinae from
Cassis - La Bédoule with those from the Ardèche and Spain (Reboulet et
al., 2011).
). Roloboceratinae thus seems to
favor a specific facies zone, or bathymetric interval, as suggested by Moreno-Bedmar
et al. (2009), neither too shallow nor
too deep. This specific facies zone seems to be characterised by fine-grained
marly-calcareous to calcareous facies whose fauna seems to be dominated by the
presence of numerous sponges, irregular sea urchins (Toxasteridae) and small
benthic foraminifera and, conversely, by the absence of neritic fauna such as
regular sea urchins, crinoid meadows, and bryozoans or pelagic microfauna. This
distribution thus seems to be reduced to microfacies F1 to F3 of the Arnaud-Vanneau
classification (Arnaud-Vanneau, 1980,
2005), corresponding to the aphotic
zone of the slopes. The possible dependence of Roloboceratinae on certain
biotopes was mentioned by Ropolo et
al. (2008b) to explain the apparent diachronism of Roloboceratinae from
Cassis - La Bédoule with those from the Ardèche and Spain (Reboulet et
al., 2011).
|
Figure 13:
Biostratigraphic occurrence of the ammonites of the subfamily Roloboceratinae Casey,
1961, in the studied sections. |
From
a biostratigraphic point of view, Roloboceras
Casey, 1954, and Megatyloceras Humphrey,
1949, are present on almost all of the dated deposits in the upper part of the D.
forbesi Zone within the Chabert
Formation (Pictet, 2011; Pictet et
al., 2015; Pictet & Delanoy,
2017) and coexist on a good
quarter of them (Table 14). Roloboceras
is present from the base of the Chabert Formation, in the Violette Marls (R. aff. hispanicum Sornay,
1972, form B) and Megatyloceras aff. coronatum
(Rouchadzé, 1933) from the topmost part of these marls. On the other
hand, both genera are abundant in the "Lower Rocherenard Beds", and in the
Rouvière Marls (Fig. 3 ![]() ). In these marls, well exposed in the vicinity of Le
Teil, the Roloboceratinae represent 30% of the cephalopod fauna, dominated by Pseudohaploceras
Hyatt, 1900 (49%). Within the Roloboceratinae, Megatyloceras quantitatively dominates (18%) over Roloboceras
(12%) (Fig. 14
). In these marls, well exposed in the vicinity of Le
Teil, the Roloboceratinae represent 30% of the cephalopod fauna, dominated by Pseudohaploceras
Hyatt, 1900 (49%). Within the Roloboceratinae, Megatyloceras quantitatively dominates (18%) over Roloboceras
(12%) (Fig. 14 ![]() ).
).
|
Figure 14:
Percentage distribution of the ammonites genera and nautiloids in
the Rouvière Marls. |
These
data differ somewhat from the observations made in the Lower Greensand of
southern England by Casey (1961) and Casey et
al. (1998). While both genera occur in the Deshayesites
forbesi Zone, their distribution within this zone diverges. Indeed, Roloboceras
is known to occur in the upper part of the D.
fittoni Zone, in the D. kiliani
and D. callidiscus zones and at the
base of the D. annelidus Zone. Megatyloceras
occurs only in the D. annelidus Zone
but above the levels that delivered the last Roloboceras
(Casey, 1961; Casey et al.,
1998) (Table 14; Fig. 17 ![]() ).
).
Table 14: Biostratigraphic occurrence of Roloboceras Casey, 1954 (in green) and Megatyloceras Humphrey, 1949 (in red) in Southern England, Anglo-Paris Basin and Languedoc platform regarding the zonal scheme used in these geographic regions [1) Aptian sensu Reboulet et al., 2011, 2014, 2018, 2) Moullade et al., 2011, 2017].
 |
In
the Paris Basin, the genus Roloboceras
was reported in 19th century literature as Ammonites
Hambrovii (Forbes, 1845) (Cornuel,
1860, p. 764) and in the
20th century as Douvilleiceras Hambrovii
by Corroy (1925, p. 440). Describing the Argiles à Plicatules Formation
from the Yonne Department, Lambert (1916) located the stratigraphic
occurrence of "Ammonites Hombronii"
(sic) in the more indurated sandy beds toping the exploited clays from Gurgy,
the same indurated beds that delivered the large ammonite fauna (Fig. 15 ![]() ; Péron,
1905; Lambert, 1916; Corroy, 1925). Cornuel
(1860, p. 762)
specifies that these indurated levels are situated in his Argiles à Plicatules
moyennes unit. The only known specimen that could correspond to Ammonites Hambrovii in view of the knowledge of that time is the
holotype of Megatyloceras transiens (Casey,
1961) that belongs to Orbigny's collection (MNHNP.F.A25552) and comes
from Gurgy outcrops. The specimen presents a large phosphatic shell of which one
of the faces is strongly corroded and encrusted by serpulids and oysters. This
specimen was first recognized with uncertainty as a Roloboceras by Casey (1961) and later revised by Casey
et al. (1998) as a representative of
the genus Megatyloceras. For its part,
Megatyloceras ricordeanum (Orbigny, 1850) is a species introduced from pyritic material also originating from Gurgy
outcrops. Still following Cornuel (1860,
1886), the pyritic fauna is
situated in the Argiles à Plicatules moyennes (Fig. 15
; Péron,
1905; Lambert, 1916; Corroy, 1925). Cornuel
(1860, p. 762)
specifies that these indurated levels are situated in his Argiles à Plicatules
moyennes unit. The only known specimen that could correspond to Ammonites Hambrovii in view of the knowledge of that time is the
holotype of Megatyloceras transiens (Casey,
1961) that belongs to Orbigny's collection (MNHNP.F.A25552) and comes
from Gurgy outcrops. The specimen presents a large phosphatic shell of which one
of the faces is strongly corroded and encrusted by serpulids and oysters. This
specimen was first recognized with uncertainty as a Roloboceras by Casey (1961) and later revised by Casey
et al. (1998) as a representative of
the genus Megatyloceras. For its part,
Megatyloceras ricordeanum (Orbigny, 1850) is a species introduced from pyritic material also originating from Gurgy
outcrops. Still following Cornuel (1860,
1886), the pyritic fauna is
situated in the Argiles à Plicatules moyennes (Fig. 15 ![]() ). The exact
stratigraphic level of M. ricordeanum
inside this unit is unfortunately unknown. Studying the rich pyritic fauna of
Deshayesites Kazansky, 1914, from the Paris Basin, Bersac and Bert
(2015, p. 282) suggested that Megatyloceras
could be a co-index of the Deshayesites
deshayesi Zone, D. deshayesi
Subzone sensu Bersac & Bert
(2012) (= Deshayesites
forbesi Zone, D. annelidus Subzone
sensu Casey et al., 1998)
(Table 14). Moreover, Bersac and Bert (2015) note the total
absence of the Roloboceras against the
presence of the Megatyloceras as an
argument to consider the whole Argiles à Plicatules Formation as a monozonal
formation belonging to the Deshayesites
deshayesi Subzone sensu Bersac & Bert (2015) (see
Fig. 15
). The exact
stratigraphic level of M. ricordeanum
inside this unit is unfortunately unknown. Studying the rich pyritic fauna of
Deshayesites Kazansky, 1914, from the Paris Basin, Bersac and Bert
(2015, p. 282) suggested that Megatyloceras
could be a co-index of the Deshayesites
deshayesi Zone, D. deshayesi
Subzone sensu Bersac & Bert
(2012) (= Deshayesites
forbesi Zone, D. annelidus Subzone
sensu Casey et al., 1998)
(Table 14). Moreover, Bersac and Bert (2015) note the total
absence of the Roloboceras against the
presence of the Megatyloceras as an
argument to consider the whole Argiles à Plicatules Formation as a monozonal
formation belonging to the Deshayesites
deshayesi Subzone sensu Bersac & Bert (2015) (see
Fig. 15 ![]() ). A recent sedimentary study was carried out by Amédro et
al. (2017) on several boreholes in the Brienne-le-Château sector and more
especially on the Juzanvigny drilling cores (core AUB 111 and AUB 121) in the
Aube Department for the ANDRA (Agence Nationale pour la Gestion des Déchets
Radioactifs). This study highlighted that the Argiles à Plicatules Formation is
composed of several sedimentary sequences separated by discontinuity surfaces
associated with iron-rich and/or phosphatic conglomerates, confirming the
assumptions of Amédro and Matrion
(2004b). Ammonites collected in
the boreholes date at least two Lower Aptian ammonite zones, the D.
deshayesi and the D. furcata
zones. Based on museum collections, Bulot et
al. (2018) support the interpretation that the Argiles à Plicatules is a
polyzonal formation that at least contains the upper parts of Deshayesites forbesi Zone, Deshayesites
deshayesi Zone, Dufrenoyia furcata
Zone and the lowermost part of Epicheloniceras
martini Zone. More recently, Deconinck et al.
(2021) performed stable isotope analyses on the Argiles à
Plicatules Formation and recognized the negative carbon-isotopes excursion which
precedes on a global scale the oceanic anoxic event 1a (OAE1a) inside their unit
2 (Fig. 15
). A recent sedimentary study was carried out by Amédro et
al. (2017) on several boreholes in the Brienne-le-Château sector and more
especially on the Juzanvigny drilling cores (core AUB 111 and AUB 121) in the
Aube Department for the ANDRA (Agence Nationale pour la Gestion des Déchets
Radioactifs). This study highlighted that the Argiles à Plicatules Formation is
composed of several sedimentary sequences separated by discontinuity surfaces
associated with iron-rich and/or phosphatic conglomerates, confirming the
assumptions of Amédro and Matrion
(2004b). Ammonites collected in
the boreholes date at least two Lower Aptian ammonite zones, the D.
deshayesi and the D. furcata
zones. Based on museum collections, Bulot et
al. (2018) support the interpretation that the Argiles à Plicatules is a
polyzonal formation that at least contains the upper parts of Deshayesites forbesi Zone, Deshayesites
deshayesi Zone, Dufrenoyia furcata
Zone and the lowermost part of Epicheloniceras
martini Zone. More recently, Deconinck et al.
(2021) performed stable isotope analyses on the Argiles à
Plicatules Formation and recognized the negative carbon-isotopes excursion which
precedes on a global scale the oceanic anoxic event 1a (OAE1a) inside their unit
2 (Fig. 15 ![]() ). This negative carbon-isotopes excursion is a particularly reliable
chemostratigraphic marker for the D.
forbesi Zone, R. hambrovi Subzone
in the Tethys Domain, highligthing the presence of time-equivalent series in the
Paris-Bassin despite the absence of the genus Roloboceras.
). This negative carbon-isotopes excursion is a particularly reliable
chemostratigraphic marker for the D.
forbesi Zone, R. hambrovi Subzone
in the Tethys Domain, highligthing the presence of time-equivalent series in the
Paris-Bassin despite the absence of the genus Roloboceras.
|
Figure 15:
Lithostratigraphy, ammonite biostratigraphy and lithologic log modified from Amédro
et al. (2017), and δ13C data with OAE 1a isotopic segments
modified from Deconinck et al.
(2021) from the Juzanvigny (AUB 121) drilling core, Aube Department, France.
Stratigraphic ranges of the Roloboceratinae taxa are discussed in the text and
photographs are from Orbigny's specimens (Orbigny,
1850). |
Further afield, if Roloboceras and Megatyloceras coexist in the Deshayesites forbesi Zone of the Mediterranean regions as in Spain and in the south-east of France, the presence of Roloboceras in Bulgaria remains to be demonstrated and would appear absent in the Caucasian regions. Faced with the divergences and/or difficulties often observed in the interpretation of Deshayesites Kazansky, 1914, the predominant index forms of the Lower Aptian, Roloboceratinae could become a good candidate for the role of index or co-index forms for outer platform environments, as it is already the case for Roloboceras hambrovi (Forbes, 1845). However, nominating Megatyloceras requires a prerequisite thorough knowledge of the species making up the genus (species definition, geographical and stratigraphic distribution, and ecological conditions).
Taxonomy of Roloboceratinae and presumed dimorphism: The important material collected on the Languedoc platform has made possible revision of M. coronatum (Rouchadzé, 1933), the type species of Megatyloceras Humphrey, 1949, to define its variability and to specify stratigraphic position. The necessary comparison with the types of taxa of the subboreal domain, collected in the south of England, and in the Paris Basin [Megatyloceras vastum Casey, 1961, M. transiens (Casey, 1961) and M. ricordeanum (Orbigny, 1850)] reveal great ornamental similarities in the juvenile whorls of all these taxa, with the systematic presence of a Ricordeanum stage. Moreover, the type of M. ricordeanum, consisting of a nucleus showing only this single stage, makes it difficult to nominate this species as viable taxa. A better definition of the Orbigny taxon ricordeanum can only be made with the help of additional material, more complete and stratigraphically well identified.
The hypothesis of a dimorphism within Roloboceratinae has been advanced by Najarro et al. (2011) who recognized microconchs of Roloboceras Casey, 1954 (Najarro et al., 2011, Fig. 8T, W) in very small, but fragmentary, forms. Similar micromorphic forms have been figured in the Tejera de Josa section (Moreno et al., 2010, Fig. 4A). Nevertheless, this hypothesis was founded on specimens whose geographical distribution appears to be restricted to the Basque-Cantabrian and Maestrat Basins in Spain. A further study of these micromorphic forms is necessary to precise their relationships with other Roloboceratinae.
In
the Ardèche, the study of Roloboceras
and Megatyloceras tends to show that, within these two genera, two
groups seem to coexist with great ornamental affinities shown on the internal
whorls, but differing in adult size (Table 15 and Fig. 16 ![]() ). In the smaller
specimens, which are supposed to represent microconch forms, the adult
ornamental stage with periumbilical bullae appears soon after the last suture
line. The end of the phragmocone is about 100-110 mm in Megatyloceras, and 80-110 m in Roloboceras.
The whorl height at the last septa is between 30-60 mm in Megatyloceras, 40 mm in Roloboceras.
The measurements of the diameters and whorl heights in some large forms (M?) and
small forms (m?) are presented in Table 15. Schematic
comparative ontogenetic development between the supposed dimorphs of both genera
is illustrated in Figure 17
). In the smaller
specimens, which are supposed to represent microconch forms, the adult
ornamental stage with periumbilical bullae appears soon after the last suture
line. The end of the phragmocone is about 100-110 mm in Megatyloceras, and 80-110 m in Roloboceras.
The whorl height at the last septa is between 30-60 mm in Megatyloceras, 40 mm in Roloboceras.
The measurements of the diameters and whorl heights in some large forms (M?) and
small forms (m?) are presented in Table 15. Schematic
comparative ontogenetic development between the supposed dimorphs of both genera
is illustrated in Figure 17 ![]() .
.
Table 15: Dimensions in mm of D and H (largest diameter and whorl height in supposed macroconchs (M?) and in supposed microconchs (m?) as in Fig. 2; Dimensions of d and h (largest diameter and whorl height at the end of the phragmocone in supposed macroconchs (M?) and microconchs (m?) within Megatyloceras Humphrey, 1949, and Roloboceras Casey, 1954.
| Genus | D M? | H M? | d M? | h M? | D m? | H m? | d m? | h m? |
| Megatyloceras | 240-300 (417) | 80-100 (150) | 140-160 (260) | 60-70 (115) | 150 | 60 | 100-110 | 30-60 |
| Roloboceras | 250-300 | 100-110 | 160-180 | 70 | 150-170 | 60-80 | 80-110 | 40 |
|
Figure 16:
Representation of the supposed dimorphism in Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949. |
|
Figure 17:
Schematic ontogenetic development of the phragmocone/body chamber in Roloboceras
Casey, 1954, and Megatyloceras Humphrey, 1949. |
In
this work, small-sized adult forms of Roloboceras
hambrovi (Forbes, 1845) and R.
aff. hispanicum Sornay, 1972,
and some Megatyloceras such as M. aff.
coronatum form A, M. ibericum Sornay, 1972, M. leteilensis sp. nov. and Megatyloceras
sp. are considered as possible microconchs forms (Fig. 16 ![]() ). Casey (1961)
reported and included in R. horridum (Spath, 1930) specimens showing rapid loss of strong tuberculation (Casey, 1961,
Pl. XXXII, figs. 2 and 3a-b; this work, Pl. 12
). Casey (1961)
reported and included in R. horridum (Spath, 1930) specimens showing rapid loss of strong tuberculation (Casey, 1961,
Pl. XXXII, figs. 2 and 3a-b; this work, Pl. 12 ![]() , fig. 3a-c;
Pl. 30
, fig. 3a-c;
Pl. 30 ![]() , fig. 2a-c).
This ornamental feature is similar to the observations mentioned above in some Roloboceras
and Megatyloceras of this study and
could be an additional proof of the existence of a dimensional dimorphism within
these genera.
, fig. 2a-c).
This ornamental feature is similar to the observations mentioned above in some Roloboceras
and Megatyloceras of this study and
could be an additional proof of the existence of a dimensional dimorphism within
these genera.
OAE 1a chemostratigraphy and the calibration of the Roloboceratinae: Faced with the difficulty of using Deshayesites Kazansky, 1914, the carbon isotope chemostratigraphy has imposed itself for calibrating the Lower Aptian marine deposits (Menegatti et al., 1998). It has been widely documented that the Lower Aptian records major disturbances in the global carbon cycle, associated with black shales of the Oceanic Anoxic Event (OAE) 1a in deep basins of the Tethys and Boreal realms (Arthur et al., 1990). The carbon signatures of the OAE 1a expose a stepped negative, then positive, excursion in the isotope profile that have been divided into eight successive segments (Menegatti et al., 1998). The massive deposition of OAE 1a-related black shales most commonly occurs between the C4 and C6 segments (Li et al., 2016, and references therein).
Gröcke
et al. (1999) first noticed that the
Roloboceratinae spread during the disturbances of OAE 1a in southern UK (Fig. 18 ![]() ). The C-isotope profile documented by those authors is based on the
composition of fossil driftwood fragments, collected sporadically throughout the
Lower Greensand succession. The profile, although imperfect, allow recognition
of most of the carbon segments. Subsequent work by Moreno-Bedmar et
al. (2008a, 2009, 2012) and Najarro et
al. (2011) on the Iberian ammonoid faunas have confirmed the observations of
Gröcke et al. (1999).
Furthermore, the Iberian Roloboceratinae are exclusively known by isolated
occurrences between the C2 and C5 segments through the Maestrat, Prebetic and
Basque-Cantabrian basins. Furthermore, the Iberian Roloboceratinae assemblages
are most often exclusive: Roloboceras Casey, 1954, and Megatyloceras
Humphrey, 1949, rarely co-occur together except in certain condensed
deposits (for example, Tejeria de Josa and Barranco Emilia section of the Oliete
sub-basin, Maestrat). Elsewhere (Paris Basin, Georgia and Bulgaria), there is no
chemostratigraphic calibration of the Roloboceratinae as yet.
). The C-isotope profile documented by those authors is based on the
composition of fossil driftwood fragments, collected sporadically throughout the
Lower Greensand succession. The profile, although imperfect, allow recognition
of most of the carbon segments. Subsequent work by Moreno-Bedmar et
al. (2008a, 2009, 2012) and Najarro et
al. (2011) on the Iberian ammonoid faunas have confirmed the observations of
Gröcke et al. (1999).
Furthermore, the Iberian Roloboceratinae are exclusively known by isolated
occurrences between the C2 and C5 segments through the Maestrat, Prebetic and
Basque-Cantabrian basins. Furthermore, the Iberian Roloboceratinae assemblages
are most often exclusive: Roloboceras Casey, 1954, and Megatyloceras
Humphrey, 1949, rarely co-occur together except in certain condensed
deposits (for example, Tejeria de Josa and Barranco Emilia section of the Oliete
sub-basin, Maestrat). Elsewhere (Paris Basin, Georgia and Bulgaria), there is no
chemostratigraphic calibration of the Roloboceratinae as yet.
|
Figure 18:
Lithostratigraphy, ammonite biostratigraphy, lithologic log and raw δ13C
wood data with OAE 1a isotopic segments from the Atherfield section, Isle of
Wight, southern United Kingdom (modified from Gröcke et
al., 1999, and Frau, 2020). Stratigraphic ranges of the English
Roloboceratinae taxa are discussed in the text and photographs are from Casey's
specimens (Casey, 1961; Casey et
al., 1998). |
In Ardèche, Pictet
et al. (2015)
documented the carbon and oxygen isotope signals from the Chabert Formation at
the Michelet, Picourel, Bourg-Saint-Andéol, and Chabert sections. Isotopic
values have proved to reflect a primary signal and the best-developed record is
found at the Chabert section. There, a long negative δ13C
excursion is followed by a positive shift and a short plateau, indicative of the
C3 to C7 segments according to the authors (Fig. 19 ![]() ). The overlying Frayol
Formation would mark the beginning of the C8 segment, as documented at Picourel.
Based on the recent works of one of us (Frau et
al., 2017, 2018; Tendil et al.,
2019), we here provide a re-assessment of the extension of the isotopic segments
at Chabert. Our main correction concerns the extension of the C2 segment, up to
the middle part of the Violette Marls Member, and thus the ensuing reduction of
the C3 segment (Fig. 19
). The overlying Frayol
Formation would mark the beginning of the C8 segment, as documented at Picourel.
Based on the recent works of one of us (Frau et
al., 2017, 2018; Tendil et al.,
2019), we here provide a re-assessment of the extension of the isotopic segments
at Chabert. Our main correction concerns the extension of the C2 segment, up to
the middle part of the Violette Marls Member, and thus the ensuing reduction of
the C3 segment (Fig. 19 ![]() ). Following these modifications, the stratigraphic range
of Roloboceras covers the uppermost C2 segment to the C4 segment, while
those of Megatyloceras is limited to the middle C4 segment up to the
lowermost C6 segment. The overlap between the two genera is, therefore, mostly
limited to part of the C4 segment. As a conclusion, the inception of Roloboceras
shortly predates the onset of the OAE 1a black shales contained between the C4
to C6 segments (op. cit.). Both Roloboceras and Megatyloceras co-occur
during the culmination of the anoxic event and decline together in its aftermath.
). Following these modifications, the stratigraphic range
of Roloboceras covers the uppermost C2 segment to the C4 segment, while
those of Megatyloceras is limited to the middle C4 segment up to the
lowermost C6 segment. The overlap between the two genera is, therefore, mostly
limited to part of the C4 segment. As a conclusion, the inception of Roloboceras
shortly predates the onset of the OAE 1a black shales contained between the C4
to C6 segments (op. cit.). Both Roloboceras and Megatyloceras co-occur
during the culmination of the anoxic event and decline together in its aftermath.
|
Figure 19:
Lithostratigraphy, ammonite biostratigraphy, lithologic log, and δ13C data
with OAE 1a isotopic segments from the Chabert section, Ardèche Department,
southern France (data from Pictet et
al., 2015, revised by Frau, 2020). Stratigraphic ranges of the French
Roloboceratinae taxa are discussed in the text. |
The stratigraphic distribution of Roloboceratinae on the Bas-Vivarais shows that they are present on the lower two thirds of the deposits of the Chabert Formation dated to the upper part of the D. forbesi Zone. The genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949, cohabit on a quarter of the D. forbesi Zone. Roloboceras is dominant in the Violette Member and the lower Rocherenard Beds while Megatyloceras predominates in the Rouvière Beds. These data differ somewhat from observations made in the Lower Greensand where the genera Roloboceras and Megatyloceras do not coexist. The study of the distribution of Roloboceratinae on a transect from the very inner ramp to the very outer facies shows an absence of this subfamily on the inner ramp, and a gradual shift of their occurrence towards the higher lithostratigraphic units in the seaward direction. This heterogeneous distribution is remarkable and clearly correlated to a bathymetric interval, or zone of optimal comfort, neither too superficial nor too deep, in the aphotic zone of the outer slope.
The rich collected fauna of Roloboceratinae has made possible revision of M. coronatum (Rouchadzé, 1933), the type species of Megatyloceras, to define its variability and to specify stratigraphic position. Comparison of the types of Megatyloceras reveal great ornamental similarities in the juvenile whorls of all taxa, with the systematic presence of a Ricordeanum stage. The existence of a sexual dimorphism within the genera Roloboceras and Megatyloceras seems very probable but requires confirmation.
We are very grateful to Roland Gonnet, Patrick and Marc Boselli for their invaluable help in the field.
We warmly thank Fabienne Giraud (OSUG, Saint-Martin d'Hères, France), Jean Michel Pacaud (Muséum National d'Histoire Naturelle, Paris, France), Jaap Klein (Naturalis Biodiversity Center, Leiden, Netherlands), Gilles Pavy and Gaëlle Labonne (Musée d'Histoire Naturelle d'Auxerre, France), Abel Prieur and Emmanuel Robert (Université Lyon 1, Villeurbanne, France), Mikheil V. Kakabadze (A. Janelidze Institute of Geology of Georgia, Tbilisi, Georgia), Alex Peaker (Dinosaur Isle, Sandown, Isle of Wight, UK), Matt Riley (Sedgwick Museum, Cambridge, UK), Simon Harris, Paul Shepherd, Neep Louise Frances and Mike Howe (British Geological Survey, Nottingham, UK), Eugen Grădinaru (University of Bucharest, Romania), Martin Simpson (Southampton, UK), Ottilia Szives (Hungarian Natural History Museum, Budapest, Hungary), Pablo Aragonés Suárez (Mexico) and Nigel D. Chapman (Bornemouth, UK) who, through their various aids, have greatly contributed to the preparation of this work. We also warmly thank Francis Amédro (Université de Bourgogne - Franche-Comté, Dijon, France) and an anonymous reviewer for their constructive remarks.
This work benefited from a grant from the French state managed by the Agence nationale de la Recherche via the programme "Investissements d'avenir" (ANR-11-INBS-0004-RECOLNAT).
Amédro F. & Matrion B. (2004a).- L'Aptien-Albien du bassin de Paris : Un nouveau regard à l'aube du XXIe siècle.- Bulletin d'Information des Géologues du Bassin de Paris, vol. 41, no. 2, p. 3-23.
Amédro F. & Matrion B. (2004b).- Les ammonites aptiennes des Argiles à plicatules de la bordure orientale du Bassin de Paris : Un aperçu.- Bulletin Annuel de l'Association Géologique Auboise, Sainte-Savine, no. 24-25, p. 75-89.
Amédro F., Matrion B., Deconinck J.-F., Huret E. & Landrein P. (2017).- Les forages de Juzanvigny (Aube, France) : Litho-biostratigraphie des formations du Barrémien à l'Albien moyen dans l'est du bassin de Paris et datations par les ammonites.- Geodiversitas, Paris, vol. 39, no. 2, p. 185-212.
Arkell W.J., Kummel B. & Wright C.W. (1957).- Mesozoic Ammonoidea. In: Moore R.C. (ed.), Treatise on Invertebrate Paleontology. Part L, Mollusca 4, Cephalopoda, Ammonoidea.- The Geological Society of America, Boulder - CO; The University of Kansas, Lawrence - KS, 490 p.
Arnaud-Vanneau A. (1980).- Micropaléontologie, paléoécologie et sédimentologie d'une plate-forme carbonatée de la marge passive de la Téthys : L'Urgonien du Vercors septentrional et de la Chartreuse (Alpes occidentales).- Géologie Alpine, Mémoire, Grenoble, no. 11, 874 p.
Arnaud-Vanneau A. (2005).- Environmental controls on tropical carbonate platform. In: Adatte T., Arnaud-Vanneau A., Arnaud H., Blanc-Aletru M.-C., Bodin S., Carrio-Schaffhauser E., Föllmi K.B., Godet A., Raddadi M.C. & Vermeulen J. (Eds.), The Hauterivian-Lower Aptian sequence stratigraphy from Jura Platform to Vocontian Basin: A multidisciplinary approach.- Géologie Alpine, Série spéciale "Colloques Excursions", Grenoble, no. 7, p. 29-78.
Arthur M.A., Brumsack H.J., Jenkyns H.C., Schlanger S.O., Ginsburg R.N. & Beaudoin B. (1990).- Stratigraphy, geochemistry, and paleoceanography of organic carbon-rich Cretaceous sequences. In: Ginsburg R.N. & Beaudoin B. (eds.), Cretaceous resources, events and rhythms: Background and plans for research.- Kluwer Academic Publishers & NATO Scientific Affairs Division, Dordrecht, p. 75-119.
Bataller J.R. (1950).- Sinopsis de las especies nuevas del Cretacico de España. Pars VIII - Mollusca, 3 Cephalopoda.- Anales de la Escuela de Peritos Agrícolas y de Especialidades Agropecuarias y de los Servicios Técnicos de Agricultura, Barcelona, vol. 9, p. 63-165.
Baudouin C., Delanoy G., Moreno-Bedmar J.A., Pictet A., Vermeulen J., Conte G., Gonnet R., Boselli P. & Boselli M. (2016).- Revision of the Early Cretaceous genera Heminautilus Spath, 1927, and Josanautilus Martínez & Grauges, 2006 (Nautilida, Cenoceratidae).- Carnets Geol., Madrid, vol. 16, no. 5, p. 61-212.
Bersac S. & Bert D. (2012).- Ontogenesis, variability and evolution of the Lower Greensand Deshayesitidae (Ammonoidea, Lower Cretaceous, Southern England): Reinterpretation of literature data; taxonomic and biostratigraphic implications.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXVII, p. 197-270.
Bersac S. & Bert D. (2015).- Two ammonite species under the same name: Revision of Deshayesites deshayesi (d'Orbigny, 1841) based on topotype material (Lower Aptian, Lower Cretaceous, Northeast of France).- Annales de Paléontologie, Paris, vol. 101, no. 4, p. 265-294.
Bersac S. & Bert D. (2017).- Revision of the lower Aptian (Lower Cretaceous) ammonite species Cheloniceras cornuelianum (d'Orbigny, 1841).- Annales de Paléontologie, Paris, vol. 104, no. 1, p. 45-70.
Bersac S. & Bert D. (2020).- The heteromorph ammonite genus Ancyloceras (Ancyloceratidae) in the Paris Basin (lower Aptian, Lower Cretaceous, NE France).- Annales de Paléontologie, Paris, vol. 106, no. 2, p. 1-15.
Bulot L.G., Frau C. & Pictet A. (2018).- Revision of Toxoceratoides royeri (d'Orbigny, 1842) and its bearing on the systematics of the Aptian Acrioceratidae Vermeulen, 2004 (Ammonoidea, Ancyloceratina, Ancyloceratoidea).- Cretaceous Research, vol. 88, p. 187-196.
Busnardo R. (1984).- Crétacé inférieur: 1.3.1. - Ammonites. In: Debrand-Passard S. (ed.), Synthèse géologique du sud-est de la France. I: Stratigraphie et paléogéographie.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, no. 125, p. 292-294.
Buvignier A. (1852).- Statistique géologique, minéralogique, minérallurgique et paléontologique du département de la Meuse.- Baillière, Paris, 694 p.
Carez L. (1882).- Sur l'Aptien et le Gault dans les départements du Gard et de l'Ardèche.- Bulletin de la Société Géologique de France (3e série), Paris, t. 11, p. 100-102.
Casey R. (1954).- New genera and sub genera of Lower Cretaceous ammonites.- Journal of the Washington Academy of Sciences, vol. 44, no. 4, p. 106-115.
Casey R. (1961).- A monograph of the Ammonoidea of the Lower Greensand. Part III.- Palaeontographical Society, London, p. 119-216.
R. (1964).- A monograph of the Ammonoidea of the Lower Greensand: Part V.- Palaeontographical Society, London, vol. 117 (1963), p. 289-398.
Casey R., Bayliss H.M. & Simpson M.I. (1998).- Observations on the lithostratigraphy and ammonite succession of the Aptian (Lower Cretaceous) Lower Greensand of Chale Bay, Isle of Wight, UK).- Cretaceous Research, vol. 19, p. 511-535.
Chiriac M. (1981).- Amoniţi Cretacici din Dobrogea de Sud, Studiu biostratigrafic.- Editura Academiei Republicii Socialiste România, Bucureşti, 138 p.
Clavel B., Conrad M.A., Busnardo R., Charollais J. & Granier B. (2013).- Mapping the rise and demise of Urgonian platforms (Late Hauterivian - Early Aptian) in southeastern France and the Swiss Jura.- Cretaceous Research, vol. 39, p. 29-46.
Conte G. (1975).- Une Ammonite peu commune, Roloboceras sp. dans l'Aptien de la Bédoule, Bouches-du-Rhône.- Géologie Méditerranéenne, Marseille, t. 2, no. 3, p. 105-110.
Contensuzas C. (1980).- Le Barrémien-Bédoulien entre Viviers-sur-Rhône et Cruas, Ardèche : Stratigraphie, analyse et répartition des faciès, cartographie.- Thèse, Université Claude Bernard, Lyon, 172 p.
Coquand H. (1866).- Monographie paléontologique de l'étage Aptien de l'Espagne.- Mémoires de la Société d'Émulation, Marseille, 221 p.
Cornuel J. (1860).- Sur le groupe du grès vert inférieur du bassin de la Seine, sur sa division d'après les oscillations du sol et les caractères géologiques, et sur ses rapports, assise par assise, avec les diverses parties du groupe Wealdien et du Lower Greensand d'Angleterre.- Bulletin de la Société Géologique de France (2e série), Paris, t. 17, p. 736-788.
Cornuel J. (1886).- Liste des fossiles du terrain Crétacé inférieur de la Haute-Marne.- Bulletin de la Société Géologique de France (3e série), Paris, t. 14, p. 312-323.
Corroy G. (1925).- Le Néocomien de la bordure orientale du Bassin de Paris.- Imprimerie J. Coubé & Fils, Nancy, 334 p.
Cox L.R., Castell C.P. & Muir-Wood H.M. (1967).- British Mesozoic Fossils. Third Edition.- Natural History Museum, London, 207 p.
Deconinck J.-F., Boué D., Amédro F., Baudin F., Bruneau L., Huret E., Landrein P., Moreau J.-D. & Santoni A.L. (2021).- First record of early Aptian Oceanic Anoxic event1a from the Paris Basin (France) - climatic signals on a terrigenous shelf.- Cretaceous Research, vol. 125, article 104846, 13 p.
Dimitrova N. (1967).- Les fossiles de Bulgarie, IV, Crétacé inférieur, Cephalopoda (Nautiloidea et Ammonoidea).- Académie Bulgare des Sciences, Sofia, 424 p.
Etayo-Serna F. (1979).- Zonation of the Cretaceous of central Colombia by ammonites.- Publicaciones Geológicas Especiales del Ingeominas, Bogotá, no. 2, 186 p.
Etayo-Serna F., Moreno Sanchez M. & Llinas Rivera R.D. (1994).- Estratigrafia de las capas basales de la Formacion El Ocal, Quebrada Bambuca (Aipe), valle superior del Magdalena, Colombia. In: Etayo-Serna F. (ed.), Estudios geológicos del valle superior del Magdalena.- Universidad Nacional de Colombia, Bogotá, p. 1-14.
Forbes E. (1845).- Catalogue of Lower Greensand fossils, in the Museum of the Geological Society, with notices of species new to Britain, contained in other collections.- The Quarterly Journal of the Geological Society of London, vol. 1, p. 237-250, 345-355.
Frau C. (2020).- Stable carbon-isotope chemostratigraphy versus ammonite biostratigraphy: Data from around the Barremian/Aptian boundary (Lower Cretaceous).- Strata (2e série), Gaillac, vol. 56, p. 1-31.
Frau C., Delanoy G. & Hourqueig E. (2015).- Le genre Macroscaphites Meek, 1876 (Ammonoidea) dans l'Aptien inférieur de Cassis-La Bédoule (Bouches-du-Rhône, France). Proposition d'un nouveau schéma zonal pour la série stratotypique.- Revue de Paléobiologie, Genève, vol. 34, no. 1, p. 45-57.
Frau C., Pictet A., Spangenberg J., Masse J.-P., Tendil A.J.-B. & Lanteaume C. (2017).- New insights on the age of the post-Urgonian marly cover of the Apt region (Vaucluse, SE France) and its implications on the demise of the North Provence carbonate platform.- Sedimentary Geology, Tulsa - OK, vol. 359, p. 44-61.
Frau C., Tendil A.J.-B., Lanteaume C., Masse J.-P., Pictet A., Bulot L.G., Luber T., Redfern J., Borgomano J., Léonide P., Fournier F. & Massonnat G. (2018).- Late Barremian-early Aptian ammonite bioevents from the Urgonian-type series of Provence (Southeast France): Regional stratigraphic correlations and implications for the peri-Vocontian carbonate platforms.- Cretaceous Research, vol. 90, p. 222-253.
Gröcke D.R., Hesselbo S.P. & Jenkyns H.C. (1999).- Carbon-isotope composition of lower Cretaceous fossil wood: ocean-atmosphere chemistry and relation to sea-level change.- Geology, Boulder - CO, vol. 27, no. 2, p. 155-158.
Humphrey W.E. (1949).- Geology of the Sierra de los Muertos Area, Mexico (with descriptions of Aptian Cephalopods from the la Peña Formation).- Bulletin of the Geological Society of America, Boulder - CO, vol. 60, p. 89-176.
A. (1903).- Pseudoceratites of the Cretaceous.- Monographs of the United States Geological Survey, Washington DC, vol. 54, 351 p.
Ivanov M. & Idakieva V. (2013).- Lower Aptian ammonite biostratigraphy and potential for further studies of OAE 1 a in Bulgaria.- Cretaceous Research, vol. 39, p. 47-69.
Kazansky P.A. (1914).- Opisanie kollektsii golovonogikh iz melovykh otlozheniin Dagestana (Description d'une collection de céphalopodes des terrains crétacés du Daghestan).- Izvestiia Tomskogo Teckhnolo-gicheskoga Instituta, vol. 32, no. 4, 127 p.
Kilian W. (1907-1913).- Erste Abteilung: Unterkreide (Palaeocretacicum). In: Frech F. (ed.), Lethaea Geognostica. II, Das Mesozoicum, Band 3 (Kreide).- Schweizerbart, Stuttgart, 398 p.
Kilian W. & Reboul P. (1915).- Contribution à l'étude des faunes paléocrétacés du Sud-Est de la France. Première partie. La faune de l'Aptien inférieur des environs de Montélimar (Drôme) [Carrière de l'Homme-d'Armes].- Mémoires pour Servir à l'Explication de la Carte géologique détaillée de la France, Paris, no. 14, 221 p.
Klein J. & Bogdanova T. (2013).- Fossilium Catalogus I: Animalia. Pars 151. Lower Cretaceous Ammonites VI Douvilleiceratoidea & Deshayesitoidea.- Backhuys Publishers, Leiden, 299 p.
Kotetishvili E.V., Kvantaliani I.V., Kakabadze M.V. & Tsirekidze L.R. (2005).- Atlas of Early Cretaceous fauna of Georgia.- Georgian Academy of Sciences, Tbilissi, 788 p.
Lafarge D. (1978, unpublished).- Étude géologique du plateau de Saint-Remèze, Ardèche. Stratigraphie, cartographie, sédimentologie, tectonique.- Thèse de 3e Cycle, Université Claude Bernard I, 119 p.
Lambert J. (1916).- Le Néocomien dans le bassin de Paris.- Bulletin de la Société Géologique de France (4e série), Paris, t. 16, p. 219-225.
Li J., Hu X., Zhao K., Cai Y. & Sun T. (2016).- Paleoceanographic evolution and chronostratigraphy of the Aptian Oceanic Anoxic Event 1a (OAE1a) to oceanic red bed 1 (ORB1) in the Gorgo a Cerbara section (central Italy).- Cretaceous Research, vol. 66, p. 115-128.
Mallada L. (1882).- Sinopsis paleontológica de España. Láminas.- Boletin de la Comisión del Mapa Geológico de España, Madrid, t. IX, láminas 3-12 (Cretáceo inferior).
Menegatti A.P., Weissert H., Brown R.S., Tyson R.V., Farrimond P., Strasser A. & Caron M. (1998).- High resolution δ13C stratigraphy through the early Aptian "Livello Selli" of the Alpine Tethys.- Paleoceanography and Paleoclimatology, vol. 13, no. 5, p. 530-545.
Moreno-Bedmar J.A. & Bover T. (2007).- Precisiones sobre la edad, mediante ammonoideos, de la Fm. Margas del Forcall, Aptiense inferior, en la subcuenca de Galve (Teruel, España).- XXIII Jornadas de la Sociedad Española de Paleontologia, Libro de Resumenes, Granada, p. 151-152.
Moreno-Bedmar J.A. & Delanoy G. (2013).- About the generic attribution of Megatyloceras casei Humphrey, 1949 (Ammonoidea, Ancyloceratina), from the Aptian of Mexico.- Carnets Geol., Madrid, vol. 13, no. L06 (CG2013_L06), p. 315-323.
Moreno-Bedmar J.A., Company M., Delanoy G., Grauges A., Martinez R. & Salas R. (2007).- Precisiones sobre la edad, mediante ammonoideos y nautiloideos, de la Fm. Margas del Forcall en la subcuenca de Oliete (Cadena Ibérica, España).- Geogaceta, Salamanca, vol. 42, p. 75-78.
Moreno-Bedmar J.A., Bover-Arnal T., Salas R. & Company M. (2008a).- The early Aptian oceanic anoxic event in the Maestrat Basin (NE Iberian Chain).- Geo-Temas, Salamanca, no. 10, p. 159-162.
Moreno-Bedmar J.A., Martin-Closas C. & Salas R. (2008b).- El registre fòssil marí (porífers, ammonítids, nautílids i equinoïdeus) i continental (caròfits) del cretaci inferior (barremià superior - aptià) del Garraf.- V Trobada d'Estudiosos del Garraf, Diputació de Barcelona, p. 205-211.
Moreno-Bedmar J.A., Company M., Bover-Arnal T., Salas R., Delanoy G., Martinez R. & Grauges A. (2009).- Biostratigraphic characterization by means of ammonoids of the lower Aptian Oceanic Anoxic Event (OAE 1a) in the eastern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, vol. 30, p. 864-872.
Moreno-Bedmar J.A., Company M., Bover-Arnal T., Salas R., Delanoy G., Maurasse F.J.-M.R., Grauges A. & Martínez R. (2010).- Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence.- Geologica Acta, Barcelona, vol. 8, no. 3, p. 281-299.
Moreno-Bedmar J.A., Company M., Sandoval J., Tavera J.M., Bover-Arnal T., Salas R., Delanoy G., Maurasse F.J.-M.R. & Martínez R. (2012).- Lower Aptian ammonite and carbon isotope stratigraphy in the eastern Prebetic Domain (Betic Cordillera, southeastern Spain).- Geologica Acta, Barcelona, vol. 10, no. 4, p. 333-350.
Moreno-Bedmar J.A., Barragán R., Delanoy G., Company M. & Salas R. (2014).- Review of the early Aptian (Early Cretaceous) ammonoid species Deshayesites deshayesi (d'Orbigny, 1841).- Cretaceous Research, vol. 51, p. 341-360.
Moullade M., Granier B. & Tronchetti G. (2011).- The Aptian stage: Back to fundamentals.- Episodes, Bangalore, vol. 34, no. 3, p. 148-156.
Moullade M., Tronchetti G., Balme C., Dutour Y., Ropolo P., Price G., Martinez M. & Granier B. (2017).- Proposition pour l'établissement du GSSP de l'Aptien supérieur dans le Bassin d'Apt (Vaucluse, SE France) : Synthèse des données stratigraphiques.- Carnets Geol., Madrid, vol. 17, no. 9, p. 161-189.
Moullade M., Tronchetti G. & Masse J.P. (eds.) (2000).- Le stratotype historique de l'Aptien inférieur (Bédoulien) dans la région de Cassis-La Bédoule (S.E. France).- Géologie méditerranéenne, Marseille, t. XXV, no. 3-4 (1998), 298 p.
Murat B. (1983).- Contribution à l'étude stratigraphique, sédimentologique et tectonique du bassin éocrétacé d'Oliete (Prov. de Teruel, Espagne).- Thèse Université Paul Sabatier, Toulouse, 263 p.
Najarro M., Rosales I., Moreno-Bedmar J.A., de Gea G.A., Barrón E., Company M. & Delanoy G. (2011).- High-resolution chemo- and biostratigraphic records of the Early Aptian oceanic anoxic event in Cantabria (N Spain): Palaeoceanographic and palaeoclimatic implications.- Palæogeography, Palæoclimatology, Palæoecology, vol. 299, p. 137-158.
Orbigny A. d' (1840-1842) - Paléontologie française, description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France. Terrains Crétacés, Vol. I. Céphalopodes.- Masson, Paris, 662 p.
Orbigny A. d' (1850a).- Prodrome de Paléontologie stratigraphique universelle des animaux mollusques & rayonnés faisant suite au cours élémentaire de paléontologie et de géologie stratigraphiques. Vol. 2.- Masson, Paris, 427 p.
Orbigny A. d' (1850b).- Notes sur quelques nouvelles espèces remarquables d'Ammonites des étages Néocomien et Aptien de la France.- Journal de Conchyliologie, Paris, t. 1, p. 196-201.
Parona C.F. & Bonarelli G. (1897).- Fossili albiani d'Escragnolles, del Nizzardo e della Liguria occidentale.- Paleontographia Italica, Pisa, vol. 2, p. 53-112.
Pascal M., Lafarge D., Chedhomme J. & Glintzboeckel C. (1989).- Bourg-Saint-Andéol.- Carte géologique de la France, Notice, 889, 67 p.
Patarroyo P. (2002).- Sobre las sucesiones marinas del Aptiano en cercanias de Ortega y Ataco (Tolima - Colombia).- Geologia Colombiana, Bogotá, no. 27, p. 87-93.
Pauliuc S. & Grădinaru E. (1970).- Sur la présence de l'Aptien inférieur dans le secteur du Sud-Ouest des Monts Persani, Megatyloceras persaniense nov. sp.- Analele Universităţii Bucureşti, Géologie, vol. 19, p. 21-41.
Péron A. (1905).- Note stratigraphique sur l'étage Aptien dans l'Est du bassin parisien.- Bulletin de la Société Géologique de France (4e série), Paris, t. 5, p. 359-378.
Peybernès B. (1976).- Le Jurassique et le Crétacé inférieur des Pyrénées Franco-Espagnoles entre la Garonne et la Méditerranée.- Thèse Université Paul Sabatier, Toulouse, 458 p.
Pictet A. (2011, unpublished).- Les Marnes à Exogyra aquila (Aptien inférieur) sur la plate-forme du Vivarais (Ardèche, France): Sédimentologie, stratigraphie, et biostratigraphie.- Mémoire de l'Université de Genève, 237 p.
Pictet A. (2012).- Evolution of the Cheloniceratinae: An alternative to Deshayesitidae for the Lower Aptian biozonation? Preliminary observations. In: Bert D. & Bersac S. (eds.): First meeting of the research group for paleobiology and biostratigraphy of the ammonites.- Boletín del Instituto de Fisiografia y Geologia, Rosario, vol. 82, p. 34-36.
Pictet A. & Delanoy G. (2017).- The Chabert Formation a newly defined stratigraphic unit of late early Aptian age in the southern Ardèche, SE France.- Archives des Sciences, Genève, vol. 69, p. 3-28.
Pictet A., Delanoy G., Adatte T., Spangenberg J.E., Baudouin C., Boselli P., Boselli M., Kindler P. & Föllmi K.B. (2015).- Three successive phases of platform demise during the early Aptian and their association with the oceanic anoxic Selli episode (Ardèche, France).- Palæogeography, Palæoclimatology, Palæoecology, vol. 418, p. 101-125.
Raisossadat S.N. (2011).- The Early Aptian (Early Cretaceous) ammonites from the Aralar Mountains, Basque-Cantabrian Basin, Northern Spain.- Carnets Geol., Madrid, vol. 11, no. M02 (CG2011_M02), p.163-201.
Reboulet S., Rawson P.F., Moreno-Bedmar J.A., Aguirre-Urreta M.B., Barragán R., Bogomolov Y., Company M., González-Arreola C., Idakieva Stoyanova V., Lukeneder A., Matrion B., Mitta V., Randrianaly H., Vašíček Z., Baraboshkin E.J., Bert D., Bersac S., Bogdanova T.N., Bulot L.G., Latil J.-L., Mikhailova I.A., Ropolo P. & Szives O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the "Kilian Group" (Dijon, France, 30th August 2010).- Cretaceous Research, vol. 32, p. 786-793.
Reboulet S., Szives O., Aguirre-Urreta B., Barragán R., Company M., Idakieva V., Ivanov M., Kakabadze M.V., Moreno-Bedmar J.A., Sandoval J., Baraboshkin E.J., Çağlar M.K., Főzy I., González-Arreola C., Kenjo S., Lukeneder A., Raisossadat S.N., Rawson P.F. & Tavera J.M. (2014).- Report of the 5th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Ankara, Turkey, 31st August 2013).- Cretaceous Research, vol. 50, p. 126-137.
Reboulet S., Szives O., Aguirre-Urreta B., Barragán R., Company M., Frau C., Kakabadze M.V, Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S.N., Vašíček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Renz O. (1982).- The Cretaceous ammonites of Venezuela.- Maraven Editor, Caracas, 132 p.
Roch E. (1927).- Etude stratigraphique et paléontologique de l'Aptien inférieur de la Bédoule (près Cassis) (Bouches-du-Rhône).- Mémoires de la Société Géologique de France, Paris, t. 4, fasc. 1, no. 8, p. 1-37.
Ropolo P., Moullade M., Gonnet R., Conte G. & Tronchetti G. (2006).- The Deshayesitidae Stoyanov, 1949 (Ammonoidea) of the Aptian historical stratotype region at Cassis-La Bédoule (SE France).- Carnets Geol., Madrid, vol. 6, no. M01 (CG2006_M01), 46 p.
Ropolo P., Conte G., Moullade M., Tronchetti G. & Gonnet R. (2008a).- The Douvilleiceratidae (Ammonoidea) of the Lower Aptian historical stratotype at Cassis-La Bédoule (SE France).- Carnets Geol., Madrid, vol. 8, no. M03 (CG2008_M03 + erratum), 60 p.
Ropolo P., Moullade M., Conte G. & Tronchetti G. (2008b).- About the stratigraphic position of the Lower Aptian Roloboceras hambrovi (Ammonoidea) level.- Carnets Geol., Madrid, vol. 8, no. L03 (CG2008_L03), 7 p.
Ropolo P., Moullade M., Gonnet R., Conte G. & Tronchetti G. (2009a).- The Deshayesitidae Stoyanov, 1949 (Ammonoidea) of the Aptian historical stratotype region at Cassis-La Bédoule (SE France).- Annales du Muséum d'Histoire Naturelle de Nice, t. XXIV, p. 43-104.
Ropolo P., Conte G., Moullade M., Tronchetti G. & Gonnet R. (2009b).- The Douvilleiceratidae (Ammonoidea) of the Lower Aptian historical stratotype at Cassis-La Bédoule (SE France).- Annales du Muséum d'Histoire Naturelle de Nice, t. XXIV, p. 115-181.
Rouchadzé I. (1933).- Les ammonites aptiennes de la Géorgie occidentale.- Bulletin de l'Institut Géologique de Géorgie, Tbilissi, vol. 1, fasc. 3, p. 165-273.
Rouchadzé I. (1938).- Céphalopodes nouveaux ou peu connus de l'Aptien de la Géorgie.- Bulletin de l'Institut Géologique de Géorgie, Tbilissi, vol. 2, fasc. 2, p. 97-190.
Salas R. & Moreno-Bedmar J.A. (2008).- Revisó de la Geologia i altres treballs complementaris de quatre Fulls del MGC25M a l'àrea del Massís de Garraf. Fulls Nos.: 420-1-1, 420-1-2, 448-1-1 i 448-1-2. Part I. Estratigratia dels materials aptians.- Institut Geològic de Catalunya, Barcelona, 41 p.
Schindewolf O.H. (1966).- Studien zur Stammesgeschichte der Ammoniten. Lieferung VI.- Abhandlungen der Mathematisch-Naturwissenschaftlichen Klasse, Akademie der Wissenschaften und der Literatur in Mainz, Wiesbaden, no. 8, p. 643-730.
Sharikadze M.Z., Kakabadze M.V. & Hoedemaeker P.J. (2004).- Early Albian Douvilleiceratidae, Acanthohoplitidae and Parahoplitidae of Colombia.- Scripta Geologica, Leiden, vol. 128, p. 313-514.
Simpson M.I. (1985).- The stratigraphy of the Atherfield Clay Formation (Lower Aptian, Lower Cretaceous) at the type and other localities in southern England.- Proceedings of the Geologists' Association, London, vol. 96, no. 1, p. 23-45.
Sornay J. (1958).- Observations sur le Tithonique supérieur de la région du Pouzin et sur le Cénomanien et l'Aptien des environs de Viviers-sur-Rhône.- Bulletin de la Carte géologique de la France, Paris, t. 56, no. 257, p. 167-171.
Sornay J. & Marin P. (1972).- Sur la faune d'ammonites aptiennes de la Tejeria de Josa (Teruel, Espagne).- Annales de Paléontologie (Invertébrés), Paris, t. 58, fasc. 1, p. 101-114.
Spath L.F. (1921).- On Cretaceous Cephalopoda from Zululand.- Annals of the South African Museum, Cape Town, vol. 12, p. 217-321.
L.F. (1923).- A monograph of the Ammonoidea of the Gault, part 1.- Monograph of the Paleontographical Society, London, (1921), vol. 75, 72 p.
Spath L.F. (1930).- On some Ammonoidea from the Lower Greensand.- The Annals and Magazine of Natural History (tenth series), London, vol. 5, no. 29, p. 417-464.
Tendil A.J.-B., Frau C., Léonide P., Fournier F., Borgomano J.R., Lanteaume C., Masse J.-P. & Rolando J.-P. (2019).- Chapter five: Stable-isotope chemostratigraphy of Urgonian-type platform carbonates: Time to be cautious? In: Montenari M. (ed.), Case Studies in Isotope Stratigraphy.- Stratigraphy & Timescales, vol. 4, p. 165-216.
Wright C.W., Callomon J.H. & Howarth M.K. (1996).- Cretaceous Ammonoidea. Treatise on Invertebrate Paleontology. Part L, Mollusca 4 Revised.- The Geological Society of America, Boulder - CO; The University of Kansas, Lawrence - KS, 362 p.
Plate
1: |
|
|
Plate
2: |
|
|
Plate
3: |
|
|
Plate
4: |
|
|
Plate
5: |
|
|
Plate
6: |
|
|
Plate
7: |
|
|
Plate
8: |
|
|
Plate
9: |
|
|
Plate
10: |
|
|
Plate
11: |
|
|
Plate
12: |
|
|
Plate
13: |
|
|
Plate
14: |
|
|
Plate
15: |
|
|
Plate
16: |
|
|
Plate
17: |
|
|
Plate
18: |
|
|
Plate
19: |
|
|
Plate
20: |
|
|
Plate
21: |
|
|
Plate
22: |
|
|
Plate
23: |
|
|
Plate
24: |
|
|
Plate
25: |
|
|
Plate
26: |
|
|
Plate
27: |
|
|
Plate
28: |
|
|
Plate
29: |
|
|
Plate
30: |
|
| Nomenclatural note: Life Sciences Identifier (LSID) http://zoobank.org/References/8C46DDA4-47B6-4493-9777-973A064AF0DE Species Group • Megatyloceras leteilensis Delanoy et al., 2022 http://zoobank.org/NomenclaturalActs/CB242214-1670-4C03-87EC-094364ADBB03 |