◄ Carnets Geol. 22 (9) ►
![]()
Sommaire
[1. Introduction]
[2. Le Turonien du Massif d'Uchaux (Vaucluse)]
[3. Le Membre des Peyras aux Prés Morel ...]
[4. Les nouveautés dans la distribution verticale ... d'Uchaux]
[5. Paléontologie systématique ...]
[6. Microconches, macroconches et dimorphisme sexuel]
[Références bibliographiques]
et ...
[Planches]
26 rue de Nottingham, 62100 Calais (France) ;
Université de Bourgogne
- Franche-Comté, UMR 6282, CNRS Biogéosciences, 6 boulevard Gabriel, 21000 Dijon (France)
16 rue Frédéric Mistral, 26200 Montélimar (France)
Chemin du Pestrier, 06670 Levens (France)
33 chemin des Cargaules, 84420 Piolenc (France)
57 rue Desmortiers, 17100 Saintes (France) ;
Université de
Mons, Faculté Polytechnique, Département Mines-Géologie, 9 rue de Houdain, B. 7000
(Belgique)
Manuscrit en ligne depuis le 1 juillet 2022
DOI 10.2110/carnets.2022.2209
![]()
[Éditeur : Bruno R.C. Granier]
![]()
Le Massif d'Uchaux (Vaucluse, SE France) est un des rares lieux en Europe oů des récoltes d'ammonites peuvent ętre effectuées du sommet du Turonien moyen au Turonien supérieur. Ce qui a conduit Alcide d'Orbigny ŕ citer en 1852 Uchaux comme "type français de l'étage Turonien" en complément de Saumur, Tourtenay, Montrichard et Tours, localités nommées en 1842 lors de la création de l'étage. En 2014, la monographie de Robaszynski et al. a synthétisé et réactualisé les informations disponibles sur la litho- et la biostratigraphie du Turonien du Massif d'Uchaux. Plus récemment, un nouvel affleurement ŕ Mondragon, au lieu-dit les Prés Morel, a recoupé la totalité des siltites et grčs carbonatés ŕ Spongia du Membre des Peyras (UL3). Le matériel récolté comprend Lewesiceras peramplum (Mantell, 1822), Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, R. (Yubariceras) ornatissimum (Stoliczka, 1864), R. (Y.) kanei Jones, 1938, Collignoniceras woollgari regulare (Haas, 1946), C. carolinum (Orbigny, 1841), C. papale (Orbigny, 1841), C. turoniense (Orbigny, 1841) et une nouvelle espčce de Collignoniceras : C. rhodanicum Amédro, Baudouin & Delanoy sp. nov. Cette association, inédite pour la région, est caractéristique de la Zone ŕ Romaniceras ornatissimum qui est la troisičme des quatre zones d'ammonites du Turonien moyen (Tm 3). Au sommet de cette 3e zone, la surprise fut de retrouver le biohorizon ŕ Romaniceras marigniacum mis en évidence en 2020 dans la partie méridionale du Bassin de Paris au sommet du Tuffeau de Bourré et ŕ la base du Tuffeau Jaune de Touraine. De plus, nous pensons maintenant que le Membre des Peyras doit ętre attribué dans sa totalité ŕ la Zone ŕ Romaniceras ornatissimum. Enfin le matériel inclut l'une des plus riches populations de Collignoniceras carolinum connues au Monde (24 ex.), ce qui permet d'apprécier le spectre de variation morphologique de l'espčce et de mettre en évidence un dimorphisme avec l'existence de formes microconches et macroconches.
• sud-est de la France ;
• Massif d'Uchaux ;
• Turonien moyen ;
• ammonites ;
• Zone ŕ Romaniceras ornatissimum ;
• biohorizon ŕ Romaniceras marigniacum ;
• Collignoniceras carolinum ;
• Collignoniceras rhodanicum sp. nov.
Amédro F., Baudouin C., Delanoy G., Devalque Ch. & Robaszynski F. (2022).- Une association d'ammonites rare dans la Zone ŕ Romaniceras ornatissimum (Turonien moyen) du Massif d'Uchaux, Vaucluse, France.- Carnets Geol., Madrid, vol. 22, nº 9, p. 333-393.
A rare ammonite assemblage in the Romaniceras ornatissimum Zone (Middle Turonian) from the Uchaux Massif, Vaucluse, France.- The Uchaux Massif (Vaucluse, SE France) is one of the rare places in Europe where ammonites collecting can be led from a high middle Turonian through Late Turonian. That incited Alcide d'Orbigny in 1852 to quote Uchaux as "the French type of the Turonian stage" to complement the Saumur, Tourtenay, Montrichard and Tours localities mentionned in 1842 when the stage was created. In 2014, the monograph by Robaszynski et al. synthetized and actualized the informations available on the litho- and biostratigraphy of the Turonian from the Uchaux Massif. More recently, a new outcrop at Mondragon, in a place named Prés Morel, exposes the whole carbonate siltstones with Spongia of the Peyras Member (UL3). Were collected : Lewesiceras peramplum (Mantell, 1822), Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, R. (Yubariceras) ornatissimum (Stoliczka, 1864), R. (Y.) kanei Jones, 1938, Collignoniceras woollgari regulare (Haas, 1946), C. carolinum (Orbigny, 1841), C. papale (Orbigny, 1841), C. turoniense (Orbigny, 1841) and a new species of Collignoniceras : C. rhodanicum Amédro, Baudouin & Delanoy sp. nov. This assemblage, unprecedented in the studied area, is characteristic of the Romaniceras ornatissimum Zone which is the third of the four ammonite zones of the middle Turonian (Tm 3). At the top of this third zone, the surprise was to find the Romaniceras marigniacum bio-horizon defined in 2020 in the southern part of the Paris Basin at the top of the Tuffeau de Bourré and at the base of the Tuffeau Jaune de Touraine. Moreover, we now think that the Peyras Member has to be fully assigned to the Romaniceras ornatissimum Zone. Lastly, the assemblage includes a rich population of Collignoniceras carolinum (24 specimens), the most important known to-day over the world. All these ammonites allow to know the spectrum of their morphological variations and show a dimorphism through the existence of micro- and macroconchs.
• south-east France;
• Uchaux Massif;
• middle Turonian;
• ammonites;
• Romaniceras ornatissimum Zone;
• Romaniceras marigniacum biohorizon;
• Collignoniceras carolinum;
• Collignoniceras rhodanicum sp. nov.
Malgré son intéręt historique, le stratotype de l'étage Turonien, qui s'étend "depuis Saumur jusqu'ŕ Montrichard" (Orbigny, 1852), ne peut pas ętre considéré comme la référence unique ŕ l'échelle mondiale dans la mesure oů les récoltes paléontologiques réalisées dans le Saumurois et en Touraine ne couvrent pas la totalité du temps correspondant ŕ l'étage. Dčs la création de son étage en 1847, Alcide d'Orbigny a eu conscience de cette question et la liste des fossiles caractéristiques de l'étage Turonien, en particulier des ammonites, publiée dans le Prodrome (Orbigny, 1850) comprend un certain nombre d'espčces alors inconnues dans la région type de l'étage Turonien, mais bien représentées ailleurs en France. C'est ainsi qu'Orbigny mentionne en 1852, dans le Cours élémentaire de paléontologie et de géologie stratigraphique, le Massif d'Uchaux dans le Vaucluse au sud-est de la France comme "type français de l'étage Turonien" en complément de Saumur, Tourtenay, Montrichard et Tours, les grčs d'Uchaux ayant livré les "Ammonites peramplus Mantell, 1822, A. Papalis d'Orbigny, 1841, A. Deverianus d'Orbigny, 1841, A. Requienianus d'Orbigny, 1841, A. Bravaisianus d'Orbigny, 1841, Baculites undulatus d'Orbigny, 1850, Scaphites Rochatianus d'Orbigny, 1850 et Hamites gracilis d'Orbigny, 1841". Aujourd'hui encore, les index des deux premičres zones d'ammonites du Turonien supérieur : Romaniceras deverianum et Subprionocyclus bravaisianus ont leur type provenant du Massif d'Uchaux. Et ce Massif est l'un des rares gisements au monde, peut-ętre męme le seul, oů l'on trouve des faunes d'ammonites riches et diversifiées au sommet du Turonien moyen et ŕ la base du Turonien supérieur, avec en particulier la succession des zones d'ammonites ŕ Romaniceras ornatissimum, ŕ Romaniceras mexicanum, ŕ Romaniceras deverianum et ŕ Subprionocyclus bravaisianus. De ce fait, l'intéręt du Massif d'Uchaux est considérable pour la zonation biostratigraphique internationale.
La Figure 1 ![]() présente la succession des zones d'ammonites actuellement en usage dans le Turonien en Europe (Amédro et al., coords. 2018). Dix zones sont distinguées le long de la bordure méridionale du Bassin anglo-parisien (au sein des Tuffeaux de Touraine et du Saumurois), dans le Bassin aquitain, dans le Sud-Est de la France, dans les Corbičres et en Espagne. Toutes ces aires géographiques sont localisées sur la marge nord de la Téthys oů les Euomphaloceratinae (Kamerunoceras puis Romaniceras) abondent dans les parties moyenne et supérieure de l'étage Turonien avec plusieurs populations qui se suivent et se relayent. La situation est en revanche différente dans les facičs crayeux du Bassin anglo-parisien (nord de la France et sud-est de l'Angleterre), dans le nord-ouest de l'Allemagne, ainsi que sur la plate-forme russe jusqu'aux abords de la Mer Caspienne. Ici, les Euomphaloceratinae ŕ affinités téthysiennes sont rares, voire absents, remplacés par des Collignoniceratinae (Lecointriceras, Collignoniceras, puis Subprionocyclus) ŕ caractčre boréal. C'est la raison pour laquelle en Europe du Nord, l'usage au sein du Turonien moyen des zones successives ŕ Kamerunoceras turoniense (Tm 1), ŕ Romaniceras kallesi (Tm 2), ŕ R. ornatissimum (Tm 3) et enfin ŕ R. mexicanum (Tm 4) est abandonné au profit d'une vaste
Zone ŕ Collignoniceras woollgari dans laquelle on peut néanmoins reconnaître deux divisions en fonction du développement des sous-espčces woollgari puis regulare.
présente la succession des zones d'ammonites actuellement en usage dans le Turonien en Europe (Amédro et al., coords. 2018). Dix zones sont distinguées le long de la bordure méridionale du Bassin anglo-parisien (au sein des Tuffeaux de Touraine et du Saumurois), dans le Bassin aquitain, dans le Sud-Est de la France, dans les Corbičres et en Espagne. Toutes ces aires géographiques sont localisées sur la marge nord de la Téthys oů les Euomphaloceratinae (Kamerunoceras puis Romaniceras) abondent dans les parties moyenne et supérieure de l'étage Turonien avec plusieurs populations qui se suivent et se relayent. La situation est en revanche différente dans les facičs crayeux du Bassin anglo-parisien (nord de la France et sud-est de l'Angleterre), dans le nord-ouest de l'Allemagne, ainsi que sur la plate-forme russe jusqu'aux abords de la Mer Caspienne. Ici, les Euomphaloceratinae ŕ affinités téthysiennes sont rares, voire absents, remplacés par des Collignoniceratinae (Lecointriceras, Collignoniceras, puis Subprionocyclus) ŕ caractčre boréal. C'est la raison pour laquelle en Europe du Nord, l'usage au sein du Turonien moyen des zones successives ŕ Kamerunoceras turoniense (Tm 1), ŕ Romaniceras kallesi (Tm 2), ŕ R. ornatissimum (Tm 3) et enfin ŕ R. mexicanum (Tm 4) est abandonné au profit d'une vaste
Zone ŕ Collignoniceras woollgari dans laquelle on peut néanmoins reconnaître deux divisions en fonction du développement des sous-espčces woollgari puis regulare.
|
|
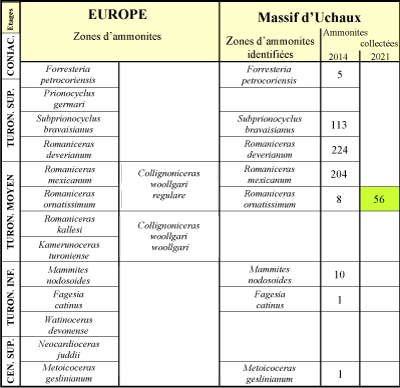
Figure 1 : Les zones d'ammonites en usage dans le Turonien en Europe et celles identifiées dans le massif d'Uchaux
(Vaucluse) avec, en regard, l'inventaire des récoltes d'ammonites réalisées par l'un d'entre nous (C.D.) en 2014 et les nouvelles récoltes d'un autre d'entre nous
(C.B.) en 2019. |
Les zones identifiées dans le Massif d'Uchaux (Amédro & Devalque, 2014) sont reportées sur la
Figure 1 ![]() avec, en regard, le nombre de spécimens trouvés dans chacune d'entre elles par l'un d'entre nous (C.D.) jusqu'en 2014. Son examen montre une importante disparité entre, d'une part les récoltes réalisées dans le Turonien inférieur et le Turonien moyen jusqu'ŕ la
Zone ŕ Romaniceras ornatissimum incluse (19 exemplaires) et d'autre part celles effectuées au sommet du Turonien moyen dans la
Zone ŕ Romaniceras mexicanum et dans le Turonien supérieur (541 spécimens). Comment expliquer cette différence ?
avec, en regard, le nombre de spécimens trouvés dans chacune d'entre elles par l'un d'entre nous (C.D.) jusqu'en 2014. Son examen montre une importante disparité entre, d'une part les récoltes réalisées dans le Turonien inférieur et le Turonien moyen jusqu'ŕ la
Zone ŕ Romaniceras ornatissimum incluse (19 exemplaires) et d'autre part celles effectuées au sommet du Turonien moyen dans la
Zone ŕ Romaniceras mexicanum et dans le Turonien supérieur (541 spécimens). Comment expliquer cette différence ?
Le Turonien inférieur et les deux premičres zones d'ammonites du Turonien moyen, successivement ŕ Kamerunoceras turoniense et ŕ Romaniceras kallesi, sont représentés dans le Massif d'Uchaux par des grčs calcaires glauconieux ŕ graviers noirs surmontés de grčs calcaires blancs ŕ Epiaster (le Membre des Vachčres de Robaszynski et al., 2014). Ces niveaux affleurent mal en raison du couvert végétal et les facičs gréseux ne sont pas favorables ŕ la préservation des macrofossiles. La troisičme zone d'ammonites du Turonien moyen ŕ Romaniceras ornatissimum correspond quant ŕ elle essentiellement ŕ des alternances de siltites gréso-carbonatées et de grčs gris ŕ Spongia (Membre des Peyras). Les "marnes ŕ Spongia" suivant l'appellation de Mennessier (1950) sont trčs sensibles ŕ l'altération et les fossiles (en dehors des éponges qui forment des boules centimétriques) se désagrčgent trčs vite lorsqu'ils sont soumis ŕ l'action des agents atmosphériques. C'est la raison pour laquelle les fossiles y semblent apparemment si rares sauf en cas de coupes récentes, comme ŕ l'occasion des travaux de terrassement réalisés en 2019-2020 sur le territoire de la commune de Mondragon au lieu-dit les Prés Morel qui ont fourni 48 nouvelles ammonites, ŕ comparer aux 8 découvertes précédemment aprčs quatre décennies de recherches.
Au-dessus, la derničre zone d'ammonites du Turonien moyen ŕ Romaniceras mexicanum et les deux premičres zones du Turonien supérieur, ŕ Romaniceras deverianum puis ŕ Subprionocyclus bravaisianus, sont préservées dans des sédiments toujours gréseux, mais plus carbonatés et beaucoup moins sensibles ŕ l'altération. Les riches populations d'ammonites qui ont fait la célébrité du Massif d'Uchaux dans le Monde entier depuis les travaux fondateurs d'Alcide d'Orbigny au XIXe sičcle proviennent de ces "Grčs d'Uchaux", nommés Formation de Massillan par Robaszynski et al. (2014).
Cet article a pour objet la description des ammonites découvertes par le second auteur (C.B.) depuis 2019 au sein de la Zone ŕ Romaniceras ornatissimum dans le gisement des Prés Morel. Malgré le nombre a priori modeste de spécimens recueillis (une cinquantaine), leur intéręt est considérable pour quatre raisons.
Le matériel inclut une riche population de Collignoniceras carolinum (24 exemplaires), la plus importante actuellement connue dans le monde, permettant d'apprécier pour la premičre fois le spectre de variation morphologique de l'espčce et de mettre en évidence un dimorphisme avec l'existence de formes microconches et macroconches.
Le type d'Ammonites papalis (Orbigny, 1841, p. 354, Pl. 109, figs. 1-3), une ammonite caractéristique de la Zone ŕ Romaniceras ornatissimum, provient du massif d'Uchaux. Ce spécimen, issu de la collection Requien, n'a pas été retrouvé au Musée d'Avignon qui héberge cette collection et les seuls autres représentants de l'espčce connus jusqu'ŕ présent proviennent de la partie méridionale du Bassin de Paris et plus précisément du Tuffeau de Saumur dans le Maine-et-Loire (1 exemplaire), du Tuffeau de Poncé dans la Sarthe (1 ex.) et du Tuffeau de Bourré dans le Loir-et-Cher (14 ex.). Le moule interne décrit ici des "Prés Morel" est l'unique spécimen actuellement préservé de la localité type.
Une nouvelle espčce est décrite : Collignoniceras rhodanicum sp. nov. connue par trois spécimens.
Enfin, Romaniceras (Yubariceras) kanei Jones, 1938, un taxon originaire du Golfe du Mexique, est identifié pour la premičre fois dans le nord-ouest de l'Europe.
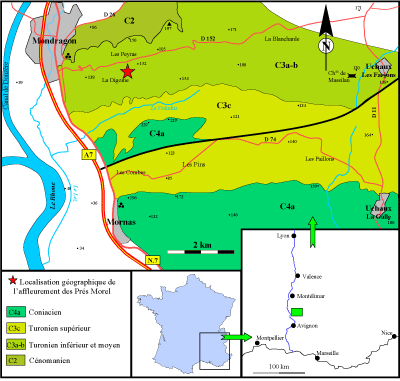
Le Massif d'Uchaux est situé dans le sud-est de la France, sur la rive gauche du Rhône ŕ une douzaine de kilomčtres au nord d'Orange, ŕ la latitude de Mondragon dans le département du Vaucluse. Il tire son nom de la commune d'Uchaux qui s'étend sur prčs de 4 km entre les hameaux d'Uchaux-Les Farjons au nord et d'Uchaux-La Galle au sud
(Fig. 2 ![]() ). Les formations visibles dans le Massif d'Uchaux appartiennent ŕ la partie moyenne du Crétacé et couvrent un intervalle allant du Cénomanien au Coniacien.
). Les formations visibles dans le Massif d'Uchaux appartiennent ŕ la partie moyenne du Crétacé et couvrent un intervalle allant du Cénomanien au Coniacien.
|
|
Figure 2 : Localisation géographique du massif d'Uchaux au sud-est de la France. |
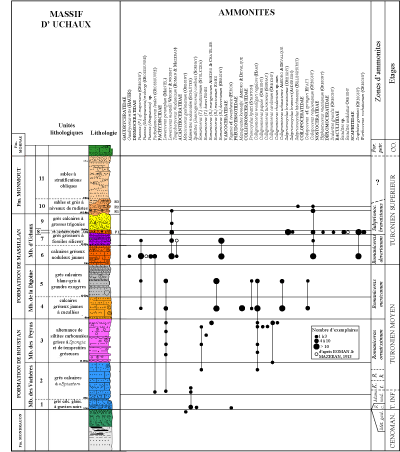
De nombreuses études ont été consacrées aux formations turoniennes du Massif d'Uchaux et ŕ leur contenu paléontologique, en particulier ammonitologique, depuis le XIXe sičcle, parmi lesquelles les plus significatives sont celles d'Orbigny (1840-1842, 1850, 1852), Hébert et Toucas (1875), Roman et Mazeran (1913), Mennessier (1950), Sornay (1950) et plus récemment Robaszynski et al. (2014) qui ont repris l'historique complet des travaux antérieurs.
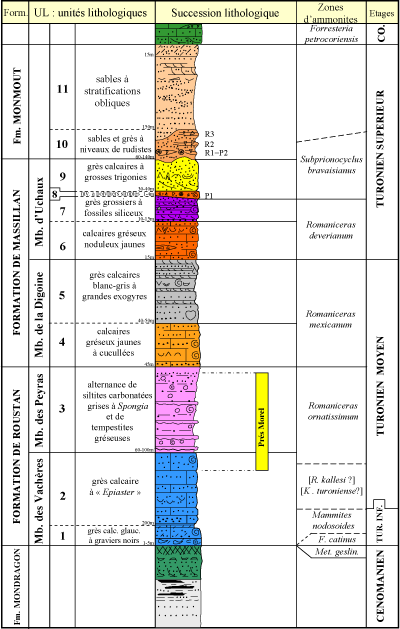
La succession lithologique a été décrite dans le détail par Robaszynski et al. (2014) ŕ partir de douze coupes partielles échelonnées sur une distance de 7 km entre la commune de Mondragon ŕ l'ouest et le hameau d'Uchaux-Les-Farjons ŕ l'est. Trois formations sont identifiées, du bas vers le haut : la Formation de Roustan (Turonien inférieur et moyen pro parte), ensemble de couches gréso-marneuses trčs peu fossilifčres de 250 ŕ 300 m d'épaisseur ; la Formation de Massillan (Turonien moyen p. p.et supérieur p. p.), ensemble gréso-calcaire de 150 ŕ 180 m d'épaisseur et enfin la Formation de Montmout (partie élevée du Turonien supérieur), couches sablo-gréseuses de 200 ŕ 300 m d'épaisseur. Plusieurs membres sont reconnus au sein des deux premičres formations, ces membres étant eux-męmes divisés en unités lithologiques de rang inférieur, numérotées UL1 ŕ 11
(Fig. 3 ![]() ).
).
|
|
Figure 3 : Les unités lithologiques reconnues dans le Turonien du massif d'Uchaux
(d'aprčs Robaszynski et al., 2014). L'épaisseur des unités lithologiques varie considérablement d'une coupe ŕ l'autre et les mesures indiquées sont des valeurs
moyennes. |
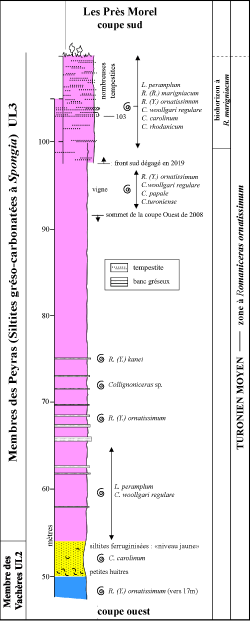
Les ammonites recueillies dans l'affleurement des Prés Morel proviennent de la moitié supérieure de la Formation de Roustan, et plus précisément du Membre des Peyras constitué de siltites gréso-carbonatées et de grčs calcaires fins riches en éponges sphériques, centimétriques, rapportées ŕ l'espčce Spongia (Hippalimus) pilula Michelin, 1843. Il s'agit des "Marnes ŕ Spongia" de Mennessier (1950). L'épaisseur du membre est estimée ŕ une centaine de mčtres au lieu-dit les Peyras. Mais le Membre des Peyras s'amincit rapidement vers l'ouest et aux Prés Morel, pourtant distants de moins d'un kilomčtre, sa puissance ne semble pas dépasser 60 mčtres.
Le Membre des Peyras : des siltites gréso-carbonatées et non des marnes sableuses
Le terme de "Marnes ŕ Spongia" a été créé par Mennessier (1950, p. 12) pour les niveaux riches en éponges sphériques et centimétriques (Spongia pilula) qui sont intercalés entre les calcaires blancs ŕ Epiaster en dessous et les Grčs ŕ Cucullées au-dessus. Le souci de mettre en conformité la terminologie des divisions lithologiques reconnues dans le Turonien du Massif d'Uchaux avec les rčgles actuelles de nomenclature stratigraphique a conduit récemment Robaszynski et al. (2014) ŕ remplacer le terme de "Marnes ŕ Spongia" par celui de "Membre des Peyras", en considérant cependant toujours qu'il s'agit d'une unité en grande partie marneuse. Les analyses pétrographiques (calcimétries et lames minces) réalisées ŕ l'Université de Dijon par notre collčgue et ami - le professeur Jean-François Deconinck - révčlent que la nature de la roche est en réalité différente. Les niveaux "tendres" sont constitués d'une siltite gréso-carbonatée contenant 39% de CaCO3. Les bancs décimétriques plus cohérents contiennent 71% de CaCO3 et correspondent ŕ un grčs trčs fin dont les grains de quartz sont cimentés par de la calcite.
Ŕ l'affleurement, la dissolution partielle des carbonates entraîne une désagrégation de la roche, ce qui explique pourquoi le Membre des Peyras apparaît toujours en creux dans le paysage, intercalé entre deux unités lithologiques plus résistantes ŕ l'altération et ŕ l'érosion. Mais il s'agit bien d'une alternance de siltites gréso-carbonatées et de grčs calcaires trčs fins et non de marnes sableuses comme on le pensait jusqu'ŕ présent - puisque la teneur en minéraux argileux est extręmement faible.
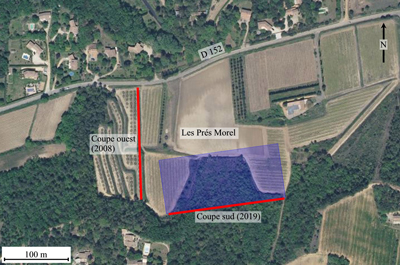
L'affleurement des Prés Morel est situé sur le territoire de la commune de Mondragon, ŕ 1 km ŕ l'est de la localité et au sud de la D 152, entre les lieux-dits les Peyras et la Digoine. Il comprend deux coupes, indexées ouest et sud
(Fig. 4 ![]() ). La coupe ouest a été révélée en 2008 ŕ l'occasion du défonçage du flanc d'une petite colline pour y planter un verger d'oliviers. Les travaux ont alors mis au jour la partie élevée du Membre des grčs blancs ŕ Epiaster des Vachčres (UL2), observée sur une épaisseur de 54 m et, au-dessus, les deux tiers inférieurs (vus sur 37,50 m) des siltites gréso-carbonatées et grčs calcaires ŕ Spongia des Peyras (UL3) (Robaszynski et al., 2014). Cette premičre coupe, toujours visible, débute prčs de la route départementale 152 et suit sensiblement un axe nord-sud.
). La coupe ouest a été révélée en 2008 ŕ l'occasion du défonçage du flanc d'une petite colline pour y planter un verger d'oliviers. Les travaux ont alors mis au jour la partie élevée du Membre des grčs blancs ŕ Epiaster des Vachčres (UL2), observée sur une épaisseur de 54 m et, au-dessus, les deux tiers inférieurs (vus sur 37,50 m) des siltites gréso-carbonatées et grčs calcaires ŕ Spongia des Peyras (UL3) (Robaszynski et al., 2014). Cette premičre coupe, toujours visible, débute prčs de la route départementale 152 et suit sensiblement un axe nord-sud.
|
|
Figure 4 : Vue aérienne de l'affleurement des Prés Morel localisé ŕ 1 km ŕ l'est de
Mondragon, au sud de la route départementale 152 (www.geoportail.gouv.fr). Les terrasses supportant les plantations d'oliviers visibles sur la partie gauche de la photo correspondent ŕ la coupe
ouest, dégagée en 2008 mais levée en 2014 (trait rouge vertical). L'aire bleutée représente la partie défoncée en 2019 et le trait rouge horizontal le front de taille d'oů proviennent les ammonites décrites ici (coupe
sud). Ce nouvel affleurement a été métré et décrit sur le plan de la lithologie en mai 2021. |
La coupe sud, dégagée en 2019, résulte de l'entaille d'une autre petite colline jouxtant la précédente un peu plus au sud-ouest. L'escarpement dégagé ŕ la faveur des travaux de terrassement recoupe la partie supérieure des siltites gréso-carbonatées et grčs calcaires ŕ Spongia des Peyras (UL3) sur une hauteur de 12,50 m. La coupe s'étend sur une longueur d'environ 200 mčtres et suit une direction est-ouest, parallčle ŕ la route départementale 152.
En 2014, le levé réalisé ŕ l'ouest des Prés Morel, le long des terrasses supportant les plantations actuelles d'oliviers (bien visibles sur la
Fig. 4 ![]() ), s'était arręté ŕ l'orée d'un bois de chęnes verts et de pins d'Alep, au niveau 91,50 m. Les terrassements effectués en 2019 ont dégagé l'affleurement sur une hauteur supplémentaire de 13,50 m et la coupe ouest des Prés Morel se termine maintenant au niveau 105 m. La corrélation entre les coupes ouest et sud des Prés Morel est réalisée grâce ŕ un niveau repčre commun dont la base est tracée ŕ 103 m. Il s'agit d'un banc gréseux pluridécimétrique onduleux, bien exprimé sur le plan de la lithologie, correspondant ŕ une tempestite et au-dessus duquel on observe de trčs nombreux bancs de grčs calcaires décimétriques en forme de fuseau. La genčse de ces bancs en fuseau résulte de rides de vagues qui remanient en fin de tempęte les couches de sable ŕ lamination plane et les rendent discontinues (information communiquée par Serge Ferry). En corrélant les deux affleurements ŕ l'aide de ce niveau repčre que l'on peut suivre latéralement, la coupe sud des Prés Morel débute au niveau 97,50 m et monte jusqu'au niveau 110 m
(Fig. 5
), s'était arręté ŕ l'orée d'un bois de chęnes verts et de pins d'Alep, au niveau 91,50 m. Les terrassements effectués en 2019 ont dégagé l'affleurement sur une hauteur supplémentaire de 13,50 m et la coupe ouest des Prés Morel se termine maintenant au niveau 105 m. La corrélation entre les coupes ouest et sud des Prés Morel est réalisée grâce ŕ un niveau repčre commun dont la base est tracée ŕ 103 m. Il s'agit d'un banc gréseux pluridécimétrique onduleux, bien exprimé sur le plan de la lithologie, correspondant ŕ une tempestite et au-dessus duquel on observe de trčs nombreux bancs de grčs calcaires décimétriques en forme de fuseau. La genčse de ces bancs en fuseau résulte de rides de vagues qui remanient en fin de tempęte les couches de sable ŕ lamination plane et les rendent discontinues (information communiquée par Serge Ferry). En corrélant les deux affleurements ŕ l'aide de ce niveau repčre que l'on peut suivre latéralement, la coupe sud des Prés Morel débute au niveau 97,50 m et monte jusqu'au niveau 110 m
(Fig. 5 ![]() ).
).
|
|
Figure 5 : Vue du flanc sud de l'affleurement des Prés Morel
(Turonien moyen) photographié en mai 2021. |
Dans la partie inférieure de l'affleurement des Prés Morel, un niveau jaune de siltite ferruginisée, identifié entre les niveaux 50 m et 54 m, marque la limite entre le Membre des grčs blancs ŕ Epiaster des Vachčres (UL2) et le Membre des siltites carbonatées et grčs calcaires ŕ Spongia des Peyras (UL3). C'est ici que débute la coupe levée en mai 2021, pour laquelle nous avons gardé le métrage de 2014. Le pendage, mesuré ŕ plusieurs endroits au toit de bancs gréseux, augmente progressivement en s'éloignant de la D 152, passant de 20° sud prčs de la route ŕ 30° sud au sommet de la colline. Du bas vers le haut, la succession lithologique composite réunissant les deux affleurements est la suivante
(Fig. 6 ![]() ) :
) :
Sommet des Grčs blancs ŕ Epiaster du Membre des Vachčres (UL2)
45 ŕ 50 m : bancs pluridécimétriques de grčs calcaires blanc-gris, plus ou moins bioturbés, contenant de fréquents oursins irréguliers (Epiaster), séparés par des niveaux décimétriques de siltites carbonatées dans lesquels on observe quelques rares éponges (Spongia) présentes sous la forme de petites sphérules centimétriques isolées. Un exemplaire de l'ammonite Romaniceras (Yubariceras) ornatissimum (Stoliczka, 1864), illustré par Amédro et Devalque (2014, Pl. 16, fig. 1) a été récolté au sommet des Grčs ŕ Epiaster dans le gisement des Prés Morel (vers le niveau 17 m, sous l'intervalle décrit ici) ;
50 m ŕ 54 m : niveau jaune constitué de siltites et incluant dans sa partie inférieure des petites huîtres dont la plupart appartiennent ŕ l'espčce Rhynchostreon suborbiculatum (Lamarck, 1801) [= Exogyra columba Lamarck, 1819]. Les Spongia deviennent abondantes. La teinte jaune du lit est liée ŕ la présence d'hydroxydes de fer. Une ammonite appartenant ŕ l'espčce Collignoniceras carolinum (Orbigny, 1841) provient de ce lit.
En toute honnęteté, il n'existe pas de limite franche entre les Grčs ŕ Epiaster (UL2) et les siltites gréso-carbonatées et grčs calcaires ŕ Spongia du Membre des Peyras (UL3). Des Spongia sont déjŕ présentes au sommet des Grčs ŕ Epiaster tandis que des Epiaster subsistent dans la partie inférieure des niveaux ŕ Spongia. De la męme façon, il n'y a pas de passage brutal des grčs aux siltites carbonatées dans la mesure oů des intercalations silteuses apparaissent dans la partie élevée des Grčs ŕ Epiaster. La limite retenue par Robaszynski et al. (2014) au sommet du "niveau jaune" est purement conventionnelle ;
Siltites gréso-carbonatées et grčs calcaires ŕ Spongia du Membre des Peyras (UL3)
54 m ŕ 75 m : succession de siltites gréso-carbonatées blanc-gris ŕ Spongia, entrecoupée de bancs décimétriques ŕ pluridécimétriques de grčs calcaires, en particulier ŕ 65,50 m, 67 m, 68,50 m, 69,80 m, 71,50 m, 73 m et 75 m. Les surfaces inférieure et supérieure des bancs gréseux sont planes et nous n'avons pas observé de granoclassement au sein des bancs ce qui suggčre qu'il ne s'agit pas de dépôts de tempętes. Les ammonites recueillies dans l'unité lithologique sont les suivantes :
Lewesiceras peramplum (Mantell, 1822) (4 exemplaires) et Collignoniceras woollgari regulare (Haas, 1946) (2 ex.) dans l'intervalle 54 m-68 m,
R. (Y.) ornatissimum (1 ex.) ŕ 68 m,
Collignoniceras sp. (1 ex.) ŕ 72 m,
Romaniceras (Yubariceras) kanei Jones, 1938 (1 ex.) ŕ 75 m ;
La macrofaune accompagnante inclut quelques nautiles, des Epiaster et des bivalves (Panopea, Cardium, Cucullaea) ;
75 m ŕ 98 m : siltites carbonatées plus ou moins micacées, avec nombreuses Spongia et parfois des bioturbations bien exposées (Thalassinoides). Les quelques ammonites trouvées ici proviennent de la partie supérieure de l'intervalle, entre les niveaux 92 m et 98 m :
Romaniceras (Y.) ornatissimum (1 exemplaire),
Collignoniceras woollgari regulare (2 ex.),
C. papale (Orbigny, 1841) (1 ex.),
C. turoniense (Sornay, 1951) (1 ex.) ;
98 m ŕ 103 m : siltites gréso-carbonatées tenant en paroi subverticale, dans lesquelles s'intercalent quelques fines couches de grčs calcaires décimétriques, ondulées ou en forme de fuseau, formant des dalles non jointives dont la taille peut atteindre plusieurs mčtres. Ces fines couches gréseuses sont caractéristiques de dépôts de tempętes. Les Spongia abondent toujours. Le niveau 102 m a fourni un fragment de Lewesiceras peramplum ;
103 m ŕ 110 m (fin de la coupe) : alternance de niveaux décimétriques de siltites gréso-carbonatées et micacées grises riches en Spongia et de trčs nombreux bancs centimétriques ŕ décimétriques onduleux de grčs calcaires. Les bancs gréseux, plus résistants ŕ l'altération et en relief, se présentent sous l'aspect de fines couches ŕ litage en mamelons et creux associés, soit suivant la formulation anglo-saxonne, en HCS (hummocky cross stratification). Il s'agit de rides de tempętes ou tempestites mises en place sous une faible tranche d'eau, dans un environnement de shoreface (quelques mčtres ŕ une ou deux dizaines de mčtres) ou d'offshore supérieur (quelques dizaines de mčtres). Des bioturbations de type Thalassinoides sont visibles dans certains bancs. Des paillettes de muscovite et quelques grains de glauconie sont associés aux grains de quartz, le tout cimenté par de la calcite d'oů les teneurs relativement élevées en CaCO3. On trouve aussi quelques bioclastes et des foraminifčres. L'examen des lames minces réalisé par J.-F. Deconinck montre qu'au sein des bancs gréseux, les grains de quartz sont fins ŕ trčs fins, bien classés et majoritairement anguleux ŕ subanguleux.
De façon générale, l'alimentation en détritique des zones ŕ sédimentation fine se fait par décantation. La finesse des grčs et silts explique l'angularité des grains car ils subissent plus difficilement l'usure que les grains de plus grande taille. Dans le cas d'une tempęte, les courants de retour entraînent vers le large des sédiments fins qui se décantent et donnent de la lamination plane. Lorsque la tempęte se calme, la couche laminée plane n'est pas remaniée en domaine d'offshore supérieur dans la mesure oů l'on se trouve sous la limite d'action des vagues. En revanche, en domaine de shoreface, l'oscillation des vagues de fin de tempęte ou męme de beau temps peut prendre le relai et remanier la couche qui vient d'ętre déposée en rides ou fuseaux.
La grande quantité de bancs gréseux confčre ŕ l'intervalle 103 m-110 m une teinte gris-blanc, plus claire qu'en dessous oů les siltites carbonatées apparaissent jaune pâle. Dans cette partie du gisement des Prés Morel, les ammonites ont été trouvées non en place au pied de l'escarpement de la coupe sud. C'est la raison pour laquelle nous n'avons pas pu différencier le matériel provenant de l'intervalle qui contient les premičres tempestites (98-103 m) de celui oů elles abondent (103-110 m). Les 41 spécimens provenant des douze mčtres de hauteur de cette coupe partielle sont les suivants :
Lewesiceras peramplum (5 exemplaires),
Romaniceras (Romaniceras) marigniacum (4 ex.),
Romaniceras (Yubariceras) ornatissimum (1 ex.),
Collignoniceras woollgari regulare (4 ex.),
C. carolinum (23 ex.),
C. rhodanicum Amédro, Baudouin & Delanoy sp. nov. (3 ex.)
C. sp. (1 ex.).
Le reste de la macrofaune trouvée dans l'intervalle 98 m-110 m, c'est-ŕ-dire dans les couches de tempętes, comprend quelques callianasses, des nautiles dont un exemplaire d'Angulithes sowerbyanus (Orbigny, 1840) illustré Pl. 9, fig. 1, des trigonies, de fréquentes cucullées et des fragments de bois flotté.
Ŕ 3 ou 4 m au-dessus du sommet de l'escarpement, soit vers 113-114 m, apparaissent les premiers bancs de calcaires gréseux ŕ cucullées du Membre de la Digoine (UL4) constituant la partie inférieure de la Formation de Massillan. En tenant compte de ces observations, l'épaisseur du Membre des siltites et marnes sableuses ŕ Spongia des Peyras peut raisonnablement ętre estimée ŕ une soixantaine de mčtres aux Prés Morel.
En 2014, nous avons attribué la partie supérieure des grčs blancs ŕ Epiaster du Membre des Vachčres (UL2), ainsi que les deux tiers inférieurs des siltites gréso-carbonatées ŕ Spongia du Membre des Peyras (UL3), ŕ la Zone ŕ Romaniceras (Y.) ornatissimum en nous appuyant sur la découverte de deux exemplaires de l'espčce index dans le gisement des Prés Morel, respectivement vers 17 m et ŕ 68 m (Robaszynski et al., 2014). Au-dessus, la présence ŕ 75 m d'un Romaniceras mal préservé, mais présentant certaines affinités avec Romaniceras (Romaniceras) mexicanum Jones, 1938 nous avait conduit ŕ placer ici la base de la zone d'ammonites suivante ŕ R. mexicanum. Le dégagement de la gangue des tours internes de ce spécimen montre qu'il possčde en réalité 11 rangées de tubercules sur les côtes longues. Cette observation conduit ŕ l'attribuer maintenant au sous-genre Yubariceras et plus précisément ŕ l'espčce Romaniceras (Yubariceras) kanei Jones, 1938, un taxon connu jusqu'ŕ présent uniquement dans le Golfe du Mexique.
La détermination de la cinquantaine de spécimens collectés depuis 2019 dans la coupe des Prés Morel par l'un d'entre nous (Cyril Baudouin) et la révision du matériel réuni en 2008-2009 par un autre d'entre nous (Christian Devalque) montrent en définitive qu'aucun R. (R.) mexicanum n'est présent au sein des siltites gréso-carbonatées ŕ Spongia et que la totalité du Membre des Peyras doit ętre attribué ŕ la
Zone ŕ R. ornatissimum (Fig. 6 ![]() ). La seconde nouveauté concerne le contenu paléontologique de la
Zone ŕ R. ornatissimum dans le Massif d'Uchaux. L'association recueillie dans la partie supérieure du Membre des Vachčres (UL2) et dans la plus grande partie du Membre des Peyras (UL3), jusqu'au niveau 98 m de la coupe des Prés Morel, est identique ŕ celle connue le long de la vallée du Cher en Touraine dans le Tuffeau de Bourré. Elle comprend : Lewesiceras peramplum, Romaniceras (Yubariceras) ornatissimum, R. (Y.) kanei, Collignoniceras woollgari regulare, C. papale, C. turoniense et C. carolinum. En revanche dans l'intervalle 98 m-110 m, c'est-ŕ-dire dans les dix derniers mčtres du Membre des Peyras oů apparaissent de nombreux niveaux de tempętes, le contenu de l'association évolue sensiblement avec la présence de nombreux Collignoniceras carolinum, Romaniceras (Y.) ornatissimum et Romaniceras (R.) marigniacum ainsi que l'apparition d'une nouvelle espčce de Collignoniceras : C. rhodanicum. Il s'agit de l'horizon ŕ Romaniceras marigniacum, situé au sommet de la
Zone ŕ R. ornatissimum et identifié pour la premičre fois dans les environs de Richelieu en Touraine méridionale par Amédro et al. (2020a, 2020b).
). La seconde nouveauté concerne le contenu paléontologique de la
Zone ŕ R. ornatissimum dans le Massif d'Uchaux. L'association recueillie dans la partie supérieure du Membre des Vachčres (UL2) et dans la plus grande partie du Membre des Peyras (UL3), jusqu'au niveau 98 m de la coupe des Prés Morel, est identique ŕ celle connue le long de la vallée du Cher en Touraine dans le Tuffeau de Bourré. Elle comprend : Lewesiceras peramplum, Romaniceras (Yubariceras) ornatissimum, R. (Y.) kanei, Collignoniceras woollgari regulare, C. papale, C. turoniense et C. carolinum. En revanche dans l'intervalle 98 m-110 m, c'est-ŕ-dire dans les dix derniers mčtres du Membre des Peyras oů apparaissent de nombreux niveaux de tempętes, le contenu de l'association évolue sensiblement avec la présence de nombreux Collignoniceras carolinum, Romaniceras (Y.) ornatissimum et Romaniceras (R.) marigniacum ainsi que l'apparition d'une nouvelle espčce de Collignoniceras : C. rhodanicum. Il s'agit de l'horizon ŕ Romaniceras marigniacum, situé au sommet de la
Zone ŕ R. ornatissimum et identifié pour la premičre fois dans les environs de Richelieu en Touraine méridionale par Amédro et al. (2020a, 2020b).
|
|
Figure 6 :
Les "Marnes ŕ Spongia" (Membre des Peyras) aux Prés Morel : Lithologie et ammonites. |
Curieusement, en Touraine comme dans le Massif d'Uchaux, c'est-ŕ-dire le long des bordures méridionale du Bassin anglo-parisien et occidentale du Bassin vocontien, la partie supérieure de la Zone ŕ Romaniceras ornatissimum correspond ŕ des couches de tempętes. En męme temps, on observe une augmentation de fréquence momentanée des ammonites ŕ caractčre boréal, celles-ci représentant 76 % de la population dans l'intervalle 98 m-110 m (jusqu'ŕ 96 % au sommet du Tuffeau de Bourré en Touraine ; cf. Amédro et al., 2020b). Une migration vers le sud d'ammonites et d'échinides ŕ caractčre boréal, associée ŕ une variation du rapport isotopique de l'oxygčne ou δ18O, a été mise en évidence ŕ ce niveau dans quatre domaines de sédimentation (sud-est de l'Angleterre, nord-ouest de l'Allemagne, Italie et nord de l'Espagne) par Voigt et Wiese (2000). Selon les auteurs, il y aurait eu au cours de cette phase un refroidissement d'environ 2°C des eaux marines parallčlement ŕ une baisse du niveau marin. Ce double événement pourrait ętre la conséquence de l'arrivée momentanée de masses d'eaux plus froides en relation, soit avec un changement des circulations océaniques, soit avec un événement eustatique et/ou tectonique.
En intégrant les nouvelles récoltes d'ammonites réalisées dans le Turonien du Massif d'Uchaux depuis 2019, la répartition verticale actualisée des espčces est présentée dans la
Figure 7 ![]() . Celle-ci révčle l'extręme richesse et la diversité des associations successives ainsi que la coexistence, au Turonien moyen, d'ammonites ŕ caractčre téthysien (Puzosia, Romaniceras, Masiaposites) et ŕ caractčre boréal (Collignoniceras).
. Celle-ci révčle l'extręme richesse et la diversité des associations successives ainsi que la coexistence, au Turonien moyen, d'ammonites ŕ caractčre téthysien (Puzosia, Romaniceras, Masiaposites) et ŕ caractčre boréal (Collignoniceras).
|
|
Figure 7 :
Distribution verticale des ammonites dans le Turonien du massif d'Uchaux. |
Seules les ammonites recueillies dans les siltites gréso-carbonatées et calcaires gréseux ŕ Spongia du Membre des Peyras (UL3) aux Prés Morel sont décrites dans la présente publication. La majorité des spécimens, indexés PM 1 ŕ 39, proviennent de la collection de Cyril Baudouin (C.B.) et ont été récoltés en 2019 et 2020. Le type de Collignoniceras rhodanicum ainsi que des moulages d'un paratype et d'un grand nombre de figurés (ŕ l'exception des plus volumineux) sont déposés dans les collections du Musée Requien ŕ Avignon oů des numéros additionnels leur ont été attribués. Les originaux y seront également remis ultérieurement.
Ŕ ce matériel s'ajoutent quelques moules internes recueillis dans le męme affleurement en 2008 par Christian Devalque (C.D.) et Bertrand Matrion (B.M.). Ces spécimens, dont certains ont déjŕ été décrits et illustrés par Amédro et Devalque (2014), sont simplement cités dans la liste du matériel se rapportant ŕ chaque espčce. La collection Devalque se trouve actuellement dans un musée sis au 30 avenue Charles de Gaulle ŕ Piolenc (Vaucluse). La collection Matrion rejoindra dans le futur celles de l'Université de Bourgogne ŕ Dijon.
La classification utilisée est celle du Treatise on Invertebrate Paleontology (C.W. Wright, 1996).
Ordre Ammonoidea Zittel, 1884
Sous-ordre Ammonitina Hyatt, 1889
Superfamille Desmoceratoidea Zittel, 1895
Famille Pachydiscidae Spath, 1922
Genre Lewesiceras Spath, 1939
Espčce-type : Ammonites peramplus Mantell, 1822, par désignation originale.
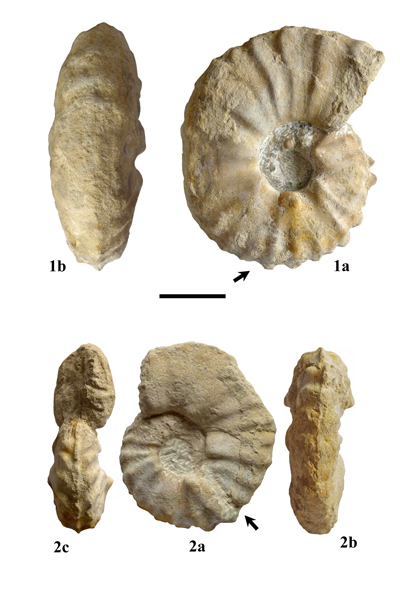
Lewesiceras peramplum (Mantell, 1822)
Pl. 1 ![]() , figs. 1-2 ;
Pl. 2
, figs. 1-2 ;
Pl. 2 ![]() , fig. 1
, fig. 1
1822 Ammonites peramplus Mantell, p. 200.
1981 Lewesiceras peramplum (Mantell) ; C.W. Wright & Kennedy, p. 29, Figs. 9-12 ; Pl. 2, figs. 1-3 ; Pl. 3 (avec synonymie).
1981 Lewesiceras peramplum (Mantell) ; Kennedy & C.W. Wright, p. 495, Figs. 1, 2 A-B ; Pl. 74 ; Pl. 75, figs. 1-7.
2014 Lewesiceras peramplum (Mantell) ; Amédro & Devalque, p. 133, Pl. 12, figs. 1-2 ; Pl. 16, fig. 2 (avec synonymie additionnelle).
2018 Lewesiceras peramplum (Mantell) ; Amédro & Matrion, p. 192, Figs. 128 A, 131 A.
2019 Lewesiceras peramplum (Mantell) ; Kennedy & Kaplan, p. 35, Pl. 7, figs. 3, 9-10, 17 ; Pl. 8-11 ; Pl. 13.
Type : le lectotype, désigné par Amédro in Robaszynski & Amédro coords. (1980, p. 254), est un des syntypes de Mantell (1822). Ce spécimen, n° BMNH 8108, est préservé dans les collections du Natural History Museum ŕ Londres (U.K.). Il provient du Middle Chalk ŕ Lewes (Sussex, U.K.).
Matériel : 9 spécimens collectés sur toute la hauteur de l'affleurement et répartis de la façon suivante : 4 exemplaires coll. C.B. (PM 12, 25, 29 et 39) et 5 exemplaires non numérotés coll. C.D.
Tableau 1 : Dimensions de quelques Lewesiceras peramplum (Mantell,
1822) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 1: Dimensions of some specimens of Lewesiceras peramplum (Mantell,
1822) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
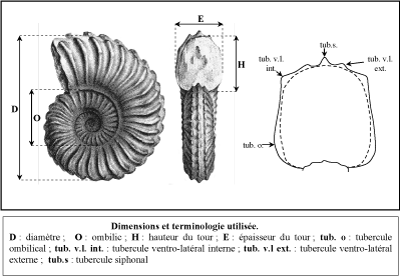
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM12 | non figuré | 179 | 56 | - | - | 0,31 | - | - | - | - |
| PM25 |
Pl. 2 |
128 | 38 | 57 | 52 | 0,30 | 0,45 | 0,41 | 0,91 | 1,10 |
| PM29 |
Pl. 1 |
302 | 85 | 123 | - | 0,28 | 0,41 | - | - | - |
| PM39 |
Pl. 1 |
34 | 7 | 17 | 11 | 0,21 | 0,50 | 0,32 | 0,65 | 1,55 |
Discussion : Les moules internes en notre possession ont un diamčtre compris entre 34 et 302 mm. La section du tour est ogivale, légčrement comprimée (rapport H/E de la hauteur sur l'épaisseur du tour de 1,10). La coquille est ornée de faibles ondulations visibles le long de la bordure ombilicale et qui correspondent ŕ des tubercules ombilicaux mieux développés sur les tours internes du phragmocône. Certains spécimens montrent également la présence sur les tours internes de côtes primaires incurvées vers l'avant, séparées par une ŕ trois courtes côtes intercalaires. Ces ammonites sont identiques aux spécimens figurés par Kennedy et C.W. Wright (1981), C.W. Wright et Kennedy (1981) et Amédro et Matrion (2018) des craies et tuffeaux du Bassin anglo-parisien, ainsi que ceux déjŕ décrits du Massif d'Uchaux par Amédro et Devalque (2014).
Distribution : Turonien inférieur (Zone ŕ Mammites nodosoides) et moyen (Zone ŕ Kamerunoceras turoniense ŕ Zone ŕ Romaniceras mexicanum) ; Formation de Roustan et moitié inférieure de la Formation de Massillan (unités 4 et 5). Distribution en dehors du Massif d'Uchaux : France, Angleterre, Belgique, Allemagne, Pologne, République tchčque, Russie européenne, Crimée, Espagne, Tunisie et Maroc.
Superfamille Acanthoceratoidea Grossouvre, 1894
Famille Acanthoceratidae Grossouvre, 1894
Sous-famille Euomphaloceratinae Cooper, 1978
Genre et sous-genre Romaniceras Spath, 1923
Espčce-type : Ammonites deverianus Orbigny, 1841, par désignation originale.
Discussion : deux sous-genres sont reconnus en Europe au sein du genre Romaniceras, différenciés en fonction du nombre de rangées de tubercules portés par les côtes longues : R. (Romaniceras) Spath, 1923, ŕ 9 rangées et R. (Yubariceras) Matsumoto et al., 1957, ŕ 11 rangées (Kennedy, C.W. Wright & Hancock, 1980a).
Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020
Pl. 13 ![]() , fig. 1 ; Pl. 14
, fig. 1 ; Pl. 14 ![]() ,
fig. 1
,
fig. 1
2020a Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, p. 51, Fig. 9
![]() ; Pl. 1
; Pl. 1 ![]() , fig. 2 ; Pl.
2
, fig. 2 ; Pl.
2 ![]() , figs. 1-2, 4 ; Pl.
4
, figs. 1-2, 4 ; Pl.
4 ![]() , figs. 1-2, 4 ; Pl.
5
, figs. 1-2, 4 ; Pl.
5 ![]() , figs. 1-2.
, figs. 1-2.
2020b Romaniceras (Romaniceras) marigniacum Amédro & Châtelier ; Amédro et al., Fig. 9 ; Pl. 1, fig. 2 a-b ; Pl. 2, figs. 1, 3, 5 ; Pl. 4, figs. 1-2, 4 ; Pl. 5, figs. 1-2.
Types : l'holotype, préservé dans les collections de Géologie de l'Université de Rennes en France sous le n° IGR-94970, provient du sommet du Tuffeau de Bourré dans la carričre "des Bruns de Marigny-Marmande" (Indre-et-Loire, France). Le paratype, IGR-94962, provient de la base du Tuffeau Jaune de Touraine dans la męme carričre.
Matériel : 4 exemplaires, coll. C.D. et coll. C.B. (PM 09, 10, 11) provenant de l'intervalle 98 m-110 m.
Tableau 2 : Dimensions de quelques Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 2: Dimensions of specimens of Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM09 |
Pl. 14 |
119 | 37 | 45 | 39 | 0,31 | 0,38 | 0,33 | 0,87 | 1,15 |
| PM10 | non figuré | 189 | 58 | 81 | 75 | 0,31 | 0,43 | 0,40 | 0,93 | 1,08 |
| PM11 |
Pl. 13 |
190 | 57 | 84 | 74 | 0,30 | 0,44 | 0,39 | 0,88 | 1,14 |
Discussion : la réalité de cette espčce a été démontrée ŕ partir de récoltes récentes réalisées en Touraine méridionale dans des niveaux attribués au sommet de la Zone ŕ R. ornatissimum et dans lesquels on trouve Romaniceras (R.) marigniacum associé ŕ l'index de zone (Amédro et al., 2020a, 2020b). Ŕ l'exception du nombre de rangées de tubercules, R. (R.) marigniacum Amédro & Châtelier, 2020 [ŕ 9 rangées de tubercules sur les côtes longues] et R. (Y.) ornatissimum (Stoliczka, 1864) [ŕ 11 rangées] sont deux espčces trčs proches morphologiquement, toutes deux caractérisées par une section du tour épaisse et le développement de côtes fortes, le plus souvent alternativement longues et courtes. La premičre et principale différence entre les deux taxons est l'absence de tubercules latéraux externes chez R. (R.) marigniacum. La seconde est le caractčre souvent plus comprimé de la coquille chez R. (R.) marigniacum oů le rapport H/E de la hauteur sur l'épaisseur du tour varie de 1,15 ŕ 0,77, ŕ comparer ŕ R. (Y.) ornatissimum oů celui-ci est compris entre 0,97 et 0,70.
Le spécimen PM 09 illustré Pl. 14 ![]() ,
fig. 1, est un adulte de 119 mm de diamčtre qui correspond ŕ un variant comprimé de l'espčce (rapport H/E = 1,15). La section du tour est subtrapézoďdale avec une épaisseur maximale au niveau des tubercules latéraux situés au tiers interne du flanc. La région ventrale, surélevée et arrondie, porte cinq rangées de tubercules arrondis : siphonal, ventro-latéraux externes et internes ŕ peu prčs équidistants les uns des autres. Les côtes, fortes et radiales, sont alternativement longues et courtes. Au total, on compte 9 rangées de tubercules, ombilicaux pincés radialement, latéraux (internes), ventro-latéraux internes et externes et siphonal. Quinze côtes sont présentes sur le dernier tour de spire. Aux abords du péristome, les côtes s'atténuent fortement et la tuberculation disparaît.
,
fig. 1, est un adulte de 119 mm de diamčtre qui correspond ŕ un variant comprimé de l'espčce (rapport H/E = 1,15). La section du tour est subtrapézoďdale avec une épaisseur maximale au niveau des tubercules latéraux situés au tiers interne du flanc. La région ventrale, surélevée et arrondie, porte cinq rangées de tubercules arrondis : siphonal, ventro-latéraux externes et internes ŕ peu prčs équidistants les uns des autres. Les côtes, fortes et radiales, sont alternativement longues et courtes. Au total, on compte 9 rangées de tubercules, ombilicaux pincés radialement, latéraux (internes), ventro-latéraux internes et externes et siphonal. Quinze côtes sont présentes sur le dernier tour de spire. Aux abords du péristome, les côtes s'atténuent fortement et la tuberculation disparaît.
Le spécimen PM 11, photographié Pl. 13 ![]() ,
fig. 1, est le moule interne d'un phragmocône de 190 mm de diamčtre, un peu plus robuste que le précédent. Les côtes longues portent 9 rangées de tubercules avec un développement plus important des tubercules latéraux. Entre les côtes longues, on observe une, parfois deux côtes courtes qui naissent ŕ des hauteurs variables sur le flanc. On compte 14 côtes sur le dernier tour de spire préservé.
,
fig. 1, est le moule interne d'un phragmocône de 190 mm de diamčtre, un peu plus robuste que le précédent. Les côtes longues portent 9 rangées de tubercules avec un développement plus important des tubercules latéraux. Entre les côtes longues, on observe une, parfois deux côtes courtes qui naissent ŕ des hauteurs variables sur le flanc. On compte 14 côtes sur le dernier tour de spire préservé.
Les deux autres moules internes recueillis aux Prés Morel sont moins bien préservés, mais présentent le męme type d'ornementation avec 9 rangées de tubercules et une alternance assez réguličre de côtes longues et courtes.
Distribution : Turonien moyen, Zone ŕ Romaniceras ornatissimum ; moitié supérieure de la Formation de Roustan (unité 3). Distribution en dehors du Massif d'Uchaux : sommet du Tuffeau de Bourré en Indre-et-Loire.
Sous-genre Romaniceras (Yubariceras) Matsumoto et al., 1957
Espčce-type : Yubariceras yubarense Matsumoto et al., 1957 par désignation originale (= Ammonites ornatissimum Stoliczka, 1864)
Romaniceras (Yubariceras) ornatissimum (Stoliczka, 1864)
Pl. 15 ![]() , fig. 1 ; Pl.
16
, fig. 1 ; Pl.
16 ![]() , fig. 2
, fig. 2
1864 Ammonites ornatissimum Stoliczka, p. 75, Pl. 40.
1889 Ammonites Deverioides Grossouvre, p. 524, Pl. 12, figs. 1-2 (incluant les variétés inermis et armata).
1980b Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Kennedy et al., p. 348, Figs. 3E, 7-8 ; Pl. 39, figs. 1-6 ; Pl. 40, figs. 1, 3-5 ; Pl. 45, figs. 1 ; Pl. 48, figs. 1-4 ; Pl. 50, figs. 1-4 (avec synonymie).
1998 Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Zonova & Yazykova, Pl. 7, figs. 1-2.
2014 Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Amédro & Devalque, p. 141, Pl. 16, fig. 1 ; Pl. 17, fig. 1 ; Pl. 18, fig. 1 (avec synonymie additionnelle).
2015 Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Kennedy et al., p. 462, Figs. 15 C-F, I, J, L ; 18 A, B.
2018 Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Amédro & Matrion, p. 194, Figs. 145 A, B ; 150 A, B.
2020a Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Amédro et al., p. 55, Pl.
1 ![]() , fig. 1 ; Pl.
3
, fig. 1 ; Pl.
3 ![]() , figs. 1-2 ; Pl.
5
, figs. 1-2 ; Pl.
5 ![]() , figs. 3-4.
, figs. 3-4.
2020b Romaniceras (Yubariceras) ornatissimum (Stoliczka) ; Amédro et al., Pl. 1, fig. 1 ; Pl. 3, figs. 3, 5.
Type : l'holotype, par monotypie, est l'original de Stoliczka (1864), p. 75; Pl. 40, fig. 1 a-e. Le specimen, préservé dans les collections de l'Indian Geological Survey ŕ Calcutta (Inde) sous le n° 174, provient de l'Uttatur Group ŕ Odium, dans le Sud de l'Inde.
Matériel : Trois exemplaires. Le premier, coll. C.D., collecté au niveau 68 m, a été illustré par Amédro et Devalque (2014, Pls. 17-18). Les deux autres, coll. C.B., proviennent respectivement des intervalles 92 m-97 m (PM 16) et 98 m-110 m (PM 26).
Tableau 3 : Dimensions de quelques Romaniceras (Yubariceras) ornatissimum (Stoliczka,
1864) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 3: Dimensions of specimens of Romaniceras (Yubariceras) ornatissimum (Stoliczka,
1864) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM16 |
Pl.
16 |
86 | 27 | 35 | 32 | 0,31 | 0,41 | 0,37 | 0,91 | 1,09 |
| PM26 |
Pl. 15 |
180 | 63 | 73 | 84 | 0,35 | 0,41 | 0,47 | 1,15 | 0,87 |
Discussion : Romaniceras (Yubariceras) ornatissimum (Stoliczka, 1864) est une espčce bien connue en Europe, en particulier dans le stratotype de l'étage Turonien, caractérisée par la présence de 11 rangées de tubercules sur les côtes longues : ombilicaux, latéraux internes et externes, ventro-latéraux internes et externes et enfin siphonal. La coquille présente le plus souvent une section du tour épaisse et des côtes fortes, alternativement longues et courtes. Chez certains variants, les tubercules restent bien marqués jusqu'au dernier tour de spire, tandis que chez d'autres, les tubercules disparaissent totalement sur la chambre d'habitation. L'enroulement est modérément évolute, avec un ombilic représentant en moyenne 30 ŕ 32 % du diamčtre.
Kennedy et al. (1980a) ont distingué trois formes dans les stades moyen et adulte de croissance :
forme A (= R. deverioides Grossouvre, 1889) : la section du tour est déprimée, les côtes alternativement longues et courtes, les côtes longues portent 11 rangées de tubercules, les courtes seulement 7 ;
forme B (= R. ornatissimum typique) : avec une section du tour plus déprimée, un ventre moins arrondi, presque plat et des tubercules latéraux internes tendant parfois ŕ devenir épineux ;
forme C (R. deverioides var. inermis Grossouvre, 1889 = bizeti Grossouvre, 1901) : avec une section du tour qui tend ŕ devenir subrectangulaire, un ventre plat et surtout une atténuation de la tuberculation au profit de la costulation, les tubercules tendant ŕ disparaître au-delŕ de 100 mm de diamčtre.
Le spécimen (coll. C.D.) du niveau 68 m, illustré par Amédro et Devalque (2014, Pl. 17, fig. 1, et Pl. 18, fig. 1) et non refiguré pour cette raison ici, est un adulte de 220 mm de diamčtre proche de la forme C. Le spécimen PM 16, photographié
Pl.
16 ![]() ,
fig. 2, possčde un diamčtre de 86 mm. Il s'agit d'un moule interne assez mal préservé, mais qui montre néanmoins la présence de 11 rangées de tubercules sur les côtes longues. Il s'agit d'une forme relativement comprimée de R. (Y.) ornatissimum. Le troisičme spécimen (PM 26), illustré
Pl. 15
,
fig. 2, possčde un diamčtre de 86 mm. Il s'agit d'un moule interne assez mal préservé, mais qui montre néanmoins la présence de 11 rangées de tubercules sur les côtes longues. Il s'agit d'une forme relativement comprimée de R. (Y.) ornatissimum. Le troisičme spécimen (PM 26), illustré
Pl. 15 ![]() ,
fig. 1, est mieux conservé. Il s'agit d'un individu adulte de 180 mm de diamčtre. La coquille est massive avec une section du tour épaisse, plus large que haute (rapport H/E de la hauteur sur l'épaisseur du tour = 0,87) et un ventre faiblement arrondi. Les côtes sont alternativement longues et courtes, avec une exception vers le péristome oů l'on observe deux côtes intercalaires successives. Sur le dernier tour de spire, on compte 13 côtes par demi-tour. Toutes les côtes longues portent 11 rangées de tubercules, avec des tubercules latéraux internes plus développés et saillants. Par l'ensemble de ces caractčres, cet exemplaire correspond ŕ la forme B, c'est-ŕ-dire ŕ un R. (Y.) ornatissimum typique.
,
fig. 1, est mieux conservé. Il s'agit d'un individu adulte de 180 mm de diamčtre. La coquille est massive avec une section du tour épaisse, plus large que haute (rapport H/E de la hauteur sur l'épaisseur du tour = 0,87) et un ventre faiblement arrondi. Les côtes sont alternativement longues et courtes, avec une exception vers le péristome oů l'on observe deux côtes intercalaires successives. Sur le dernier tour de spire, on compte 13 côtes par demi-tour. Toutes les côtes longues portent 11 rangées de tubercules, avec des tubercules latéraux internes plus développés et saillants. Par l'ensemble de ces caractčres, cet exemplaire correspond ŕ la forme B, c'est-ŕ-dire ŕ un R. (Y.) ornatissimum typique.
Distribution : Romaniceras ornatissimum est l'index de la troisičme zone d'ammonites du Turonien moyen (Tm 3) ; moitié supérieure de la Formation de Roustan (sommet de l'unité 2 et unité 3). Distribution en dehors du Massif d'Uchaux : Vienne, Sarthe, Aube et Corbičres en France, sud de l'Angleterre, Espagne, République tchčque, Tunisie, Israël, Liban, Madagascar, sud de l'Inde, Japon, Russie asiatique (Sakhaline), Californie, Texas et nord du Mexique.
Romaniceras (Yubariceras) kanei Jones, 1938
Pl. 17 ![]() , fig. 1
, fig. 1
1938 Romaniceras kanei Jones, p. 120, Pl. 8, figs. 2, 7-8 ; Pl. 9, fig. 6.
1980a Romaniceras (Yubariceras) kanei Jones ; Kennedy et al., p. 342, Fig. 9.
1988 Romaniceras (Yubariceras) kanei Jones ; Kennedy & Cobban, p. 601, Figs. 4, 7-8, Table 2.
Type : l'holotype est le spécimen figuré par Jones (1938), p. 120, Pl. 8, figs. 7-8. Ce moule interne, dont l'horizon et la localité sont inconnus, provient de l'état du Coahuila au Mexique. Il est préservé dans les collections de l'Université du Michigan (USA) sous le n° 16819.
Matériel : un exemplaire (coll. C.D.) provenant du niveau 75 m.
Tableau 4 : Dimensions du Romaniceras (Yubariceras) kanei Jones, 1938, du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 4: Dimensions of Romaniceras (Yubariceras) kanei Jones, 1938, from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| coll. C.D., ex. non numéroté | Pl. 17, fig. 1 | 128 | 45 | 46 | c44 | 0,35 | 0,36 | 0,34 | 0,96 | 1,05 |
Description : le moule interne illustré Pl. 17 ![]() est un individu adulte de 128 mm de diamčtre montrant la totalité du phragmocône et un demi-tour de la chambre d'habitation. L'enroulement est évolute (O = 35 % du diamčtre). La section du tour est aussi haute que large (H/E = 1,05), avec une épaisseur maximale observée au niveau des tubercules latéraux internes. Sur le phragmocône, la section du tour prise au niveau des côtes est polygonale. Sur la chambre d'habitation, elle devient subquadratique en gardant toutefois une région ventrale arrondie. Cette modification résulte de l'atténuation des tubercules latéraux internes, les flancs devenant plus plats et subparallčles. Le mur ombilical est plat et la bordure ombilicale étroitement arrondie. On compte 16 côtes par demi-tour. La plupart sont des côtes longues, droites ou légčrement sinueuses, surtout sur la chambre d'habitation, qui naissent au niveau de la bordure ombilicale et traversent la région ventrale sans interruption. Quelques côtes courtes, le plus souvent seules, s'intercalent parfois entre les côtes longues. Celles-ci portent 11 rangées de tubercules qui sont surtout bien visibles sur le phragmocône : ombilicaux en forme de bulle, latéraux internes arrondis et saillants, latéraux externes situés au tiers supérieur du flanc, également coniques mais moins développés et plus discrets, ventro-latéraux internes d'abord arrondis, puis qui tendent ŕ se pincer dans le sens de l'enroulement, ventro-latéraux externes pincés dans le sens de l'enroulement et enfin siphonal plus conique. Sur la chambre d'habitation, les bulles ombilicales s'allongent radialement tandis que les tubercules latéraux s'atténuent.
est un individu adulte de 128 mm de diamčtre montrant la totalité du phragmocône et un demi-tour de la chambre d'habitation. L'enroulement est évolute (O = 35 % du diamčtre). La section du tour est aussi haute que large (H/E = 1,05), avec une épaisseur maximale observée au niveau des tubercules latéraux internes. Sur le phragmocône, la section du tour prise au niveau des côtes est polygonale. Sur la chambre d'habitation, elle devient subquadratique en gardant toutefois une région ventrale arrondie. Cette modification résulte de l'atténuation des tubercules latéraux internes, les flancs devenant plus plats et subparallčles. Le mur ombilical est plat et la bordure ombilicale étroitement arrondie. On compte 16 côtes par demi-tour. La plupart sont des côtes longues, droites ou légčrement sinueuses, surtout sur la chambre d'habitation, qui naissent au niveau de la bordure ombilicale et traversent la région ventrale sans interruption. Quelques côtes courtes, le plus souvent seules, s'intercalent parfois entre les côtes longues. Celles-ci portent 11 rangées de tubercules qui sont surtout bien visibles sur le phragmocône : ombilicaux en forme de bulle, latéraux internes arrondis et saillants, latéraux externes situés au tiers supérieur du flanc, également coniques mais moins développés et plus discrets, ventro-latéraux internes d'abord arrondis, puis qui tendent ŕ se pincer dans le sens de l'enroulement, ventro-latéraux externes pincés dans le sens de l'enroulement et enfin siphonal plus conique. Sur la chambre d'habitation, les bulles ombilicales s'allongent radialement tandis que les tubercules latéraux s'atténuent.
Discussion : jusqu'ŕ présent, la seule espčce rapportée au sous-genre Yubariceras connue en Europe était Romaniceras (Yubariceras) ornatissimum (Stoliczka, 1864), caractérisée au stade moyen de croissance par un enroulement évolute, avec un ombilic représentant 30 ŕ 32 % du diamčtre, une section du tour déprimée, nettement plus large que haute (H/E variant de 0,97 ŕ 0,70) et des côtes alternativement longues et courtes.
Le spécimen décrit ici présente une morphologie assez différente. L'ombilic est plus ouvert (O = 35 % du diamčtre). Il n'y a pas d'alternance réguličre de côtes longues et courtes, mais une majorité de côtes primaires. La section du tour n'est pas déprimée, mais aussi haute que large (H/E = 1,05). Enfin, la section du tour prise au niveau des côtes primaires est polygonale sur le phragmocône. Tous ces caractčres rappellent ceux des Romaniceras décrits dans le nord du Mexique par Jones (1938, Pl. 8, figs. 7-8) et Kennedy et Cobban (1988, Figs. 4, 7-8) sous le nom de Romaniceras (Yubariceras) kanei Jones, 1938 (détermination confirmée par le Dr W.J. Kennedy, Oxford, UK).
Cet exemplaire isolé d'origine nord-américaine correspond probablement ŕ une coquille vide, flottée post-mortem et prise en charge par les circulations océaniques de surface ŕ travers le Proto-Atlantique depuis le Golfe du Mexique jusqu'en Europe. Cette migration est ŕ mettre en relation avec la courbe de variation du niveau marin qui, selon Gale (1996), présente un maximum de hauteur au tiers inférieur du Turonien moyen, ŕ la limite entre les zones d'ammonites ŕ Romaniceras kallesi (Tm 2) et ŕ R. ornatissimum (Tm 3). Męme si la Zone ŕ R. ornatissimum est interprétée comme le début d'une phase régressive, le niveau marin était encore trčs haut.
Distribution : Turonien moyen, Zone ŕ Romaniceras ornatissimum (Tm 3) ; moitié supérieure de la Formation de Roustan (Membre des Peyras ŕ Spongia ou unité 3). Distribution en dehors du Massif d'Uchaux : nord du Mexique.
Famille Collignoniceratidae C.W. Wright & E.V. Wright, 1951
Sous-famille Collignoniceratinae C.W. Wright & E.V. Wright, 1951
Genre Collignoniceras Breistroffer, 1947
Espčce-type : Ammonites Woollgari Mantell, 1822, par désignation originale de Meek, 1876 (pour le genre Prionotropis Meek, 1876, non Fieber, 1853, pour lequel Breistroffer, 1947, a proposé le nouveau nom de Collignoniceras). Opinion ICZN 861, 1968.
Collignoniceras woollgari regulare (Haas, 1946)
Pl. 8 ![]() ,
fig. 2 ;
Pl. 11
,
fig. 2 ;
Pl. 11 ![]() ,
fig. 2 ;
Pl. 12
,
fig. 2 ;
Pl. 12 ![]() ,
fig. 1
,
fig. 1
1946 Prionotropis woollgari var. regularis Haas, p. 154, Figs. 10-12, 59-74, 78, 80, 81, 83 ; Pl. 16, figs. 1-17.
1979 Collignoniceras woollgari regulare (Haas) ; Cobban & Hook, p. 22, Pl. 3, figs. 1-14 ; Pl. 12, fig. 3 (avec synonymie).
2001 Collignoniceras woollgari regulare (Haas) ; Kennedy et al., p. 45, Figs. 17-33, 49 (avec synonymie additionnelle).
2014 Collignoniceras woollgari regulare (Haas) ; Amédro & Devalque, p. 150, Pl. 19, fig. 1 ; Pl. 25, fig. 1 ; Pl. 26, fig. 1.
2018 Collignoniceras woollgari regulare (Haas) ; Amédro & Matrion, p. 197, Figs. 146 A, 147 A, B.
2019 Collignoniceras woollgari regulare (Haas) ; Kennedy & Kaplan, p. 63, Pl. 34.
2020a Collignoniceras woollgari regulare (Haas) ; Amédro et al., p. 57, Pl.
6 ![]() , figs. 1, 4 ; Pl.
9
, figs. 1, 4 ; Pl.
9 ![]() , figs. 1-2, 4.
, figs. 1-2, 4.
2020b Collignoniceras woollgari regulare (Haas) ; Amédro et al., Pl. 3, fig. 4, Pl. 4, figs. 1-2.
Type : l'holotype est, par désignation originale, le spécimen illustré par Haas (1946), Pl. 16, figs. 14, 16, d'un horizon et d'une localité inconnus dans la région des Black Hills (Dakota du Sud, USA). Il est préservé au sein des collections de la South Dakota State School of Mines sous le n° 1470.
Matériel : Matériel : 8 moules internes, dont 3 non numérotés coll. C.D (collectés respectivement dans les intervalles 54 m-65 m, 92 m-97 m et 98 m-110 m) et 5 coll. C.B. (PM 15 du niveau 97 m, PM 17 et 24 de l'intervalle 92 m-97 m, PM 33 et 36 de l'intervalle 98 m-110 m). Un moulage du spécimen PM 33 est déposé dans les collections du Musée Requien ŕ Avignon (n° 2021.08.10).
Tableau 5 : Dimensions de quelques Collignoniceras woollgari regulare (Haas,
1946) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 5: Dimensions of specimens of Collignoniceras woollgari regulare (Haas,
1946) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM15 | non figuré | 103 | 35 | 41 | 35 | 0,34 | 0,40 | 0,34 | 0,85 | 1,17 |
| PM17 |
Pl. 12 |
124 | 50 | 44 | 38 | 0,40 | 0,35 | 0,31 | 0,86 | 1,16 |
| PM24 |
Pl. 11 |
- | - | 57 | - | - | - | - | - | - |
| PM33 |
Pl. 8 |
94 | 34 | 36 | 28 | 0,36 | 0,38 | 0,30 | 0,78 | 1,29 |
| PM36 | non figuré | - | - | 43 | 37 | - | - | - | 0,86 | 1,16 |
Discussion : le spécimen (PM 24) illustré Pl. 11 ![]() ,
fig. 2, est un moule interne fragmentaire d'un individu adulte, avec un diamčtre estimé entre 14 et 15 cm, montrant un tiers de tour de spire avec le début de la chambre d'habitation. L'enroulement est modérément évolute et la section du tour ŕ peine plus haute que large. L'ornementation est constituée de côtes droites, étroites, faiblement inclinées vers l'avant. Toutes sont des côtes longues qui portent un tubercule latéral situé au tiers interne du flanc et un tubercule ventro-latéral épineux. La ligne siphonale est surélevée, dominée par une rangée de tubercules siphonaux pincés dans le sens de l'enroulement, en nombre identique aux tubercules ventro-latéraux, mais légčrement décalés vers l'avant.
,
fig. 2, est un moule interne fragmentaire d'un individu adulte, avec un diamčtre estimé entre 14 et 15 cm, montrant un tiers de tour de spire avec le début de la chambre d'habitation. L'enroulement est modérément évolute et la section du tour ŕ peine plus haute que large. L'ornementation est constituée de côtes droites, étroites, faiblement inclinées vers l'avant. Toutes sont des côtes longues qui portent un tubercule latéral situé au tiers interne du flanc et un tubercule ventro-latéral épineux. La ligne siphonale est surélevée, dominée par une rangée de tubercules siphonaux pincés dans le sens de l'enroulement, en nombre identique aux tubercules ventro-latéraux, mais légčrement décalés vers l'avant.
Le second moule interne illustré Pl. 8 ![]() ,
fig. 2 (PM 33), est un phragmocône de 94 mm de diamčtre. On compte 13 côtes par demi-tour. Le plus souvent, il s'agit de côtes longues, légčrement concaves et inclinées vers l'avant, mais quelques côtes courtes apparaissent ŕ mi-flanc ou au quart interne du flanc. Les côtes longues naissent au niveau de petits tubercules ombilicaux pincés radialement et qui restent bien individualisé ŕ tous les stades de croissance. La région ventrale porte quant ŕ elle cinq rangées de tubercules saillants : ventro-latéraux internes et externes et siphonal, ce dernier étant encore plus proéminent, pincé dans le sens de l'enroulement et décalé vers l'avant. Ce spécimen, rapporté aux formes comprimées de Collignoniceras woollgari regulare (Haas,
1946), présente une ornementation assez comparable ŕ celle de Collignoniceras carolinum (Orbigny, 1841) ŕ ce stade de croissance du phragmocône. En l'absence de la chambre d'habitation, les critčres de détermination retenus dans le cas présent sont, premičrement la croissance moins rapide de la hauteur du tour chez C. carolinum par rapport ŕ notre spécimen, deuxičmement la quasi absence de bulles ombilicales également chez C. carolinum.
,
fig. 2 (PM 33), est un phragmocône de 94 mm de diamčtre. On compte 13 côtes par demi-tour. Le plus souvent, il s'agit de côtes longues, légčrement concaves et inclinées vers l'avant, mais quelques côtes courtes apparaissent ŕ mi-flanc ou au quart interne du flanc. Les côtes longues naissent au niveau de petits tubercules ombilicaux pincés radialement et qui restent bien individualisé ŕ tous les stades de croissance. La région ventrale porte quant ŕ elle cinq rangées de tubercules saillants : ventro-latéraux internes et externes et siphonal, ce dernier étant encore plus proéminent, pincé dans le sens de l'enroulement et décalé vers l'avant. Ce spécimen, rapporté aux formes comprimées de Collignoniceras woollgari regulare (Haas,
1946), présente une ornementation assez comparable ŕ celle de Collignoniceras carolinum (Orbigny, 1841) ŕ ce stade de croissance du phragmocône. En l'absence de la chambre d'habitation, les critčres de détermination retenus dans le cas présent sont, premičrement la croissance moins rapide de la hauteur du tour chez C. carolinum par rapport ŕ notre spécimen, deuxičmement la quasi absence de bulles ombilicales également chez C. carolinum.
Les autres exemplaires déterminés comme C. woollgari regulare sont fragmentaires ou usés, mais présentent la męme morphologie que les spécimens mieux préservés provenant des Tuffeaux de Poncé (Sarthe) et de Bourré (Loir-et-Cher) illustrés par Kennedy et al. (1980b, 2001) et Amédro et Matrion
(2018). Ŕ la différence de Collignoniceras woollgari woollgari qui la précčde dans le temps, la sous-espčce Collignoniceras woollgari regulare garde toujours des côtes simples et ne présente pas une multiplication des tubercules siphonaux ni de côtes bouclées sur la région ventrale du dernier tour de spire du phragmocône et sur la chambre d'habitation. Il s'agit de la premičre citation de la sous-espčce dans le Membre des Peyras ŕ Spongia du Massif d'Uchaux. Ŕ noter que le spécimen PM 17, illustré
Pl. 12 ![]() ,
fig. 1, montre une réminiscence de quelques côtes bouclées sur la région ventrale, rappelant la sous-espčce antérieure C. woollgari woollgari.
,
fig. 1, montre une réminiscence de quelques côtes bouclées sur la région ventrale, rappelant la sous-espčce antérieure C. woollgari woollgari.
Distribution : Turonien moyen, Zone ŕ Romaniceras ornatissimum et Zone ŕ Romaniceras mexicanum ; moitié supérieure de la Formation de Roustan (sommet de l'unité UL2 et unité UL3) et moitié inférieure de la Formation de Massillan (unités UL4 et UL5). Distribution en dehors du Massif d'Uchaux : Indre-et-Loire, Loir-et-Cher, Sarthe et Gard en France, sud de l'Angleterre, Belgique, Allemagne, Iran, Western Interior des USA et du Canada, Californie, Oregon et nord du Mexique.
Collignoniceras carolinum (Orbigny, 1841)
Fig. 8 ![]() ; Pl. 2
; Pl. 2 ![]() ,
fig. 2 ; Pl. 3
,
fig. 2 ; Pl. 3 ![]() , figs. 1-2 ;
Pl. 4
, figs. 1-2 ;
Pl. 4 ![]() , figs. 1 ;
Pl. 5
, figs. 1 ;
Pl. 5 ![]() , figs. 1-2 ;
, figs. 1-2 ;
Pl. 6 ![]() , figs. 1-2 ;
Pl. 7
, figs. 1-2 ;
Pl. 7 ![]() , fig. 1 ;
Pl. 9
, fig. 1 ;
Pl. 9 ![]() , fig. 2 ;
Pl. 16
, fig. 2 ;
Pl. 16 ![]() ,
fig. 1
,
fig. 1
1841 Ammonites Carolinus d'Orbigny, p. 310, Pl. 91, figs. 5-6.
1980b Collignoniceras carolinum (d'Orbigny) ; Kennedy et al., p. 574, Figs. 1B, 5 ; Pl. 68, figs. 1-11 ; Pl. 76, figs. 1-2 (avec synonymie).
1981 Collignoniceras carolinum (d'Orbigny) ; C.W. Wright & Kennedy, p. 106, Pl. 30, figs. 4-5.
1982 Collignoniceras carolinum (d'Orbigny) ; Amédro & Badillet in Robaszynski coord. et al., p. 196, Pl. 5, fig. 4.
1988 Collignoniceras carolinum (d'Orbigny) ; Kaplan, p. 14, Pl. 6, fig. 3.
non 1996 Collignoniceras n. sp. ? aff. C. carolinum (d'Orbigny) ; Kirkland, p. 98, Pl. 45, figs. G, J ; Pl. 46, fig. A [= ? Collignoniceras woollgari regulare (Haas, 1946)].
2006 Collignoniceras carolinum (d'Orbigny) ; Kennedy & Juignet in Gauthier, p. 113, Pl. 60, fig. 2 a, b.
2014 Collignoniceras carolinum (d'Orbigny) ; Wilmsen & Nagm, p. 220, Pl. 12 d.
2016 Collignoniceras carolinum (d'Orbigny) ; Kennedy & Gale, p. 291, Fig. 20 C-F.
2019 Collignoniceras sp. aff. canthus (d'Orbigny) ; Kennedy & Kaplan, p. 65, Pl. 35, figs. 3-5 (uniquement).
Type : le lectotype est, par désignation subséquente de Kennedy et al. (1980b), le spécimen n° 6778 a de la collection Orbigny sise au Muséum national d'Histoire naturelle ŕ Paris. Il provient du Calcaire ŕ Céphalopodes de Martrou prčs de Rochefort (Charente-Maritime).
Matériel : 24 exemplaires répartis comme suit : 20 ex. coll. C. B. (PM 01, 02, 05, 06, 07, 08, 18, 19, 20, 21, 22, 23, 28, 30, 31, 32a et 32b, 34, 37 et 38, tous de l'intervalle 98 m-110 m), 3 ex. non numérotés coll. C.D. du męme intervalle et 1 ex. coll. B. M., récolté en place au sein du "niveau jaune" situé ŕ la limite supérieure de l'unité UL2, vers 53 m. Des moulages des spécimens PM 01, 02, 05, 06 et 21 sont déposés dans les collections du Musée Requien (portant respectivement les n° 2021.08.01, 2021.08.02, 2021.08.05, 2021.08.06 et 2021.08.09).
Tableau 6 : Dimensions de quelques Collignoniceras carolinum (Orbigny, 1841) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 6: Dimensions of specimens of Collignoniceras carolinum (Orbigny, 1841) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM01 |
Pl. 5 |
114 | 39 | 43 | 30 | 0,34 | 0,38 | 0,26 | 0,70 | 1,43 |
| PM02 |
Pl. 3 |
141 | 49 | 53 | 41 | 0,35 | 0,38 | 0,29 | 0,77 | 1,29 |
| PM05 |
Pl. 6 |
147 | 54 | 55 | 49 | 0,37 | 0,37 | 0,33 | 0,89 | 1,12 |
| PM06 |
Pl. 4 |
180 | 69 | 64 | 48 | 0,38 | 0,36 | 0,27 | 0,75 | 1,33 |
| PM07 |
Pl. 5 |
127 | 40 | 48 | - | 0,31 | 0,38 | - | - | - |
| PM08 | non figuré | 100 | 32 | 40 | 36 | 0,32 | 0,40 | 0,36 | 0,90 | 1,11 |
| PM18 | non figuré | 124 | 44 | 46 | 36 | 0,35 | 0,37 | 0,29 | 0,78 | 1,28 |
| PM19 | non figuré | 122 | 42 | 45 | - | 0,34 | 0,37 | - | - | - |
| PM20 | non figuré | 172 | 69 | 57 | c48 | 0,40 | 0,33 | 0,28 | 0,84 | 1,19 |
| PM21 |
Pl. 7 |
173 | 71 | 58 | 49 | 0,41 | 0,34 | 0,28 | 0,84 | 1,18 |
| PM22 |
Pl. 3 |
103 | 31 | 42 | 30 | 0,30 | 0,41 | 0,29 | 0,71 | 1,40 |
| PM23 |
Pl.
16 |
c172 | 72 | 57 | 42 | 0,42 | 0,33 | 0,24 | 0,74 | 1,36 |
| PM28 |
Pl. 2 |
99 | 26 | 42 | 26 | 0,26 | 0,42 | 0,26 | 0,62 | 1,62 |
| PM30 |
Pl. 6 |
113 | 36 | 45 | - | 0,32 | 0,40 | - | - | - |
| 77 | 23 | 32 | 25 | 0,30 | 0,42 | 0,32 | 0,78 | 1,28 | ||
| PM31 | non figuré | 99 | 33 | 38 | 33 | 0,33 | 0,38 | 0,33 | 0,87 | 1,15 |
| PM32a |
Pl. 9 |
90 | 26 | 32 | - | 0,29 | 0,36 | - | - | - |
| PM32b | non figuré | 61 | 37 | 26 | - | 0,61 | 0,43 | - | - | - |
| PM34 | non figuré | 118 | 37 | 43 | c25 | 0,31 | 0,36 | 0,21 | 0,58 | 1,72 |
| PM37 | non figuré | - | - | 42 | 37 | - | - | - | 0,88 | 1,14 |
| PM38 | non figuré | 127 | 36 | 49 | 30 | 0,28 | 0,39 | 0,24 | 0,61 | 1,63 |
Description : tous les spécimens recueillis dans le Membre des Peyras ŕ Spongia aux Prés Morel sont des moules internes correspondant ŕ des individus adultes dont le diamčtre varie de 61 ŕ 180 mm. L'enroulement est modérément évolute avec un ombilic assez large (26 ŕ 41 % du diamčtre). La coquille est comprimée (rapport H/E de la hauteur sur l'épaisseur du tour variant de 1,72 ŕ 1,11), avec une bordure ombilicale arrondie, des flancs plats, légčrement convergents et une région ventrale tectiforme. L'ornementation est constituée de nombreuses côtes fines et simples (une trentaine par tour), inclinées vers l'avant, la plupart du temps longues et qui naissent prčs de la bordure ombilicale. De petites bulles ombilicales allongées radialement sont visibles sur les tours internes des variants robustes. Elles disparaissent en revanche sur la chambre d'habitation et ne sont pas perceptibles sur les variants les plus comprimés. Le plus souvent, les côtes sont légčrement flexueuses, mais elles peuvent aussi ętre droites. Plus rarement, les côtes peuvent naître par paire au niveau de la bordure ombilicale ou ętre intercalaires. Sur le phragmocône, les côtes portent un petit tubercule ventro-latéral interne et un tubercule ventro-latéral externe bien marqué, pincé dans le sens de l'enroulement en un clavus. La ligne siphonale est dominée par une ligne de tubercules siphonaux, en męme nombre que les côtes, mais décalés vers l'avant et pincés dans le sens de l'enroulement en formant des clavi. Sur le dernier tour du phragmocône et au niveau de la chambre d'habitation, les tubercules ventro-latéraux internes disparaissent et seuls subsistent les tubercules ventro-latéraux externes qui restent toujours bien marqués et tendent ŕ ętre légčrement pincés dans le sens de l'enroulement en clavi et obliques par rapport ŕ la ligne siphonale.
|
|
Figure 8 : Collignoniceras carolinum. Copie de la figure originale d'Orbigny (1841, Pl. 91, figs. 5-6) et termes descriptifs
utilisés. |
Discussion : le lectotype de Collignoniceras carolinum (Orbigny, 1841) est un spécimen juvénile de 46 mm de diamčtre montrant la chambre d'habitation et une partie du phragmocône. Quatre autres spécimens adultes, d'un diamčtre compris entre 100 et 120 mm et provenant des formations turoniennes du Bassin anglo-parisien, ont permis ŕ Kennedy et al. (1980b) de préciser les caractčres spécifiques.
Jusqu'ŕ présent, l'espčce Collignoniceras carolinum (Orbigny, 1841) était connue par une dizaine de spécimens : deux exemplaires, dont le lectotype, du Calcaire ŕ Céphalopodes en Charente-Maritime, deux exemplaire du Tuffeau de Saumur en Anjou, deux exemplaires du Tuffeau de Bourré en Touraine, deux exemplaires des craies du sud de l'Angleterre et trois moules internes des calcaires de Westphalie et de Saxe en Allemagne. Avec vingt quatre nouveaux individus, la population récoltée dans le Massif d'Uchaux enrichit considérablement la connaissance de l'espčce, permettant en particulier de mieux apprécier son spectre de variation morphologique. Celui-ci est plus large qu'estimé par Kennedy et al. (1980b) et s'étend depuis des formes graciles ŕ côtes fines et denses jusqu'ŕ des formes robustes ŕ faible densité costale et ŕ côtes saillantes portant des tubercules proéminents.
Le moule interne figuré Pl. 2 ![]() ,
fig. 2 (PM 28), est le morphotype le plus comprimé recueilli aux Prés Morel (H/E = 1,62) et le plus involute (O/D = 0,26). Les côtes sont fines et denses (31 par tour) et tendent ŕ s'atténuer vers le péristome. Le spécimen illustré
Pl. 5
,
fig. 2 (PM 28), est le morphotype le plus comprimé recueilli aux Prés Morel (H/E = 1,62) et le plus involute (O/D = 0,26). Les côtes sont fines et denses (31 par tour) et tendent ŕ s'atténuer vers le péristome. Le spécimen illustré
Pl. 5 ![]() ,
fig. 2 (PM 01), est un autre variant comprimé (H/E = 1,43), ŕ enroulement modérément évolute (O/D = 34 %) et ŕ costulation fine et dense (30 côtes par tour). L'exemplaire photographié
Pl. 3
,
fig. 2 (PM 01), est un autre variant comprimé (H/E = 1,43), ŕ enroulement modérément évolute (O/D = 34 %) et ŕ costulation fine et dense (30 côtes par tour). L'exemplaire photographié
Pl. 3 ![]() ,
fig. 2 (PM 22), est trčs comparable au précédent, mais montre de façon distincte la présence de petits tubercules ventro-latéraux internes jusqu'ŕ un stade avancé du phragmocône. Le spécimen illustré
Pl. 6
,
fig. 2 (PM 22), est trčs comparable au précédent, mais montre de façon distincte la présence de petits tubercules ventro-latéraux internes jusqu'ŕ un stade avancé du phragmocône. Le spécimen illustré
Pl. 6 ![]() ,
fig. 2 (PM 30), est encore une forme comprimée (H/E = 1,28) avec une ornementation constituée de côtes le plus souvent longues, légčrement sinueuses et inclinées vers l'avant, entre lesquelles on observe parfois une ou deux côtes intercalaires qui naissent assez bas sur le flanc. Les côtes longues naissent au niveau de petites bulles ombilicales pincées radialement. On compte 28 côtes sur le dernier tour de spire qui inclut la terminaison du phragmocône et la chambre d'habitation. Les spécimens photographiés
Pl. 5
,
fig. 2 (PM 30), est encore une forme comprimée (H/E = 1,28) avec une ornementation constituée de côtes le plus souvent longues, légčrement sinueuses et inclinées vers l'avant, entre lesquelles on observe parfois une ou deux côtes intercalaires qui naissent assez bas sur le flanc. Les côtes longues naissent au niveau de petites bulles ombilicales pincées radialement. On compte 28 côtes sur le dernier tour de spire qui inclut la terminaison du phragmocône et la chambre d'habitation. Les spécimens photographiés
Pl. 5 ![]() ,
fig. 1, et Pl. 9
,
fig. 1, et Pl. 9 ![]() ,
fig. 2 (PM 07 et PM 32a), sont d'autres variants comprimés. Les moules internes illustrés
Pl. 4
,
fig. 2 (PM 07 et PM 32a), sont d'autres variants comprimés. Les moules internes illustrés
Pl. 4 ![]() ,
fig. 1 (PM 06), et Pl. 3
,
fig. 1 (PM 06), et Pl. 3 ![]() ,
fig. 1 (PM 02), correspondent ŕ des formes moyennes dans le spectre de variation morphologique intraspécifique (H/E voisin de 1,30 ; O/D compris entre 0,35 et 0,38). L'exemplaire illustré
Pl. 6
,
fig. 1 (PM 02), correspondent ŕ des formes moyennes dans le spectre de variation morphologique intraspécifique (H/E voisin de 1,30 ; O/D compris entre 0,35 et 0,38). L'exemplaire illustré
Pl. 6 ![]() ,
fig. 1 (PM 05), est plus épais (H/E = 1,12). Enfin le moule interne photographié
Pl. 7
,
fig. 1 (PM 05), est plus épais (H/E = 1,12). Enfin le moule interne photographié
Pl. 7 ![]() ,
fig. 1 (PM 21), correspond ŕ l'individu le plus robuste présent au sein de notre collection (H/E = 1,18 ; O = 41 % du diamčtre, 23 côtes par tour).
,
fig. 1 (PM 21), correspond ŕ l'individu le plus robuste présent au sein de notre collection (H/E = 1,18 ; O = 41 % du diamčtre, 23 côtes par tour).
Les populations recueillies en Charente-Maritime, en Touraine et dans le Vaucluse sont homogčnes, malgré leur localisation dans des bassins sédimentaires différents. Le spécimen d'Uchaux illustré ici
Pl. 3 ![]() ,
fig. 2 (PM 22), est ainsi trčs comparable ŕ celui du Tuffeau de Bourré figuré par Kennedy et al. (1980b, Pl. 76, figs. 1-2). De la męme façon, les exemplaires figurés ici
Pl. 6
,
fig. 2 (PM 22), est ainsi trčs comparable ŕ celui du Tuffeau de Bourré figuré par Kennedy et al. (1980b, Pl. 76, figs. 1-2). De la męme façon, les exemplaires figurés ici
Pl. 6 ![]() ,
fig. 1 (PM 05), et in Kennedy et al. (1980b, Pl. 68, fig. 11) sont quasi identiques.
,
fig. 1 (PM 05), et in Kennedy et al. (1980b, Pl. 68, fig. 11) sont quasi identiques.
Les deux moules internes d'une dizaine de centimčtres de diamčtre provenant de la Zone ŕ Collignoniceras woollgari de Dortmund en Rhénanie-Westphalie (Allemagne), déterminés avec doute par Kennedy et Kaplan (2019, Pl. 35, figs. 3-5) comme Collignoniceras sp. aff. canthus (Orbigny), semblent ętre également des Collignoniceras carolinum. Le spécimen illustré Pl. 35, fig. 3, est un variant robuste de l'espčce et le spécimen illustré Pl. 35, fig. 4, un variant gracile.
Distribution : Turonien moyen, avec une premičre occurrence dans la Zone ŕ Romaniceras kallesi (deux exemplaires connus dans le Tuffeau de Saumur), mais l'espčce est surtout connue dans la zone suivante ŕ Romaniceras ornatissimum ; moitié supérieure de la Formation de Roustan dans le Massif d'Uchaux (unité UL3). Distribution en dehors du Massif d'Uchaux : Aquitaine, Anjou et Touraine en France, sud de l'Angleterre (Surrey) et Allemagne (Rhénanie-Westphalie et Saxe).
Collignoniceras papale (Orbigny, 1841)
Pl. 11 ![]() ,
fig. 1
,
fig. 1
1841 Ammonites Papalis d'Orbigny, p. 354, Pl. 109, figs. 1-3.
1875 Ammonites Papalis d'Orbigny ; Hébert & Toucas, p. 43.
1980b Collignoniceras papale (d'Orbigny) ; Kennedy et al., p. 578, Figs. 1C, 6, 7 ; Pl. 69, figs. 1-2 ; Pl. 70, figs. 3-5 (avec synonymie).
1984 Collignoniceras papale (d'Orbigny) ; Kennedy et al., p. 40, Figs. 4a-c, 5.
2018 Collignoniceras papale (d'Orbigny) ; Amédro & Matrion, p. 197, Figs. 148 a, b ; 149 a.
2019 Collignoniceras cf. papale (d'Orbigny) ; Kennedy & Kaplan, p. 64, Pl. 29, fig. 1.
Type : l'holotype est, par monotypie, le moule interne décrit et figuré par Alcide d'Orbigny (1841), p. 356, Pl. 109, figs. 1-3, provenant de la "Craie chloritée du département du Vaucluse". Selon Orbigny, le spécimen serait déposé dans les collections du Musée Requien ŕ Avignon (Vaucluse, France), mais nous ne l'avons pas retrouvé.
Matériel : un spécimen, coll. C.B. (PM 14), collecté dans l'intervalle 92 m-97 m. Un moulage est déposé dans les collections du Musée Requien (n° 2021.08.08).
Tableau 7 : Dimensions du Collignoniceras papale (Orbigny, 1841) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 7: Dimensions of Collignoniceras papale (Orbigny, 1841) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM14 |
Pl. 11 |
95 | 29 | 40 | 47 | 0,31 | 0,42 | 0,49 | 1,18 | 0,85 |
Discussion : l'unique spécimen recueilli aux Prés Morel est un moule interne de 95 mm de diamčtre montrant la fin du phragmocône et le début de la chambre d'habitation. L'enroulement est modérément évolute. La section du tour intercostale est arrondie, la section costale polygonale avec une épaisseur maximale au niveau des tubercules ventro-latéraux proéminents. L'ornementation est constituée de côtes primaires droites, faiblement inclinées vers l'avant, qui naissent au niveau de la bordure ombilicale arrondie, se renflent au niveau de tubercules ombilicaux allongés radialement, ces derniers étant situés au tiers interne du flanc et rejoignent des tubercules ventro-latéraux développés en cornes. Sur la région ventrale, on observe une multiplication des tubercules siphonaux de forme arrondie. Certains sont connectés aux cornes ventro-latérales par des côtes bouclées. Les autres correspondent ŕ de courtes intercalaires présentes uniquement sur la région ventrale.
Le type d'Ammonites papalis (Orbigny, 1841, p. 354) provient du Massif d'Uchaux. Ce spécimen, issu de la collection Requien, n'a pas été retrouvé au Musée d'Avignon oů la collection est conservée. Le seul autre représentant de l'espčce signalé jusqu'ŕ présent du Massif d'Uchaux est celui cité par Hébert et Toucas (1875, p. 43) dans les "Marnes ŕ Spongia", mais cet exemplaire a également disparu. Le moule interne recueilli aux Prés Morel est en définitive le seul Collignoniceras papale disponible de la localité type, d'oů son intéręt. La récolte du spécimen dans la partie supérieure du Membre des siltites gréso-carbonatées et calcaires gréseux ŕ Spongia des Peyras (UL3) permet de préciser le niveau d'occurrence de l'espčce dans la suite stratigraphique du Massif d'Uchaux.
La figure publiée par Orbigny (1841, Pl. 109, figs. 1-2) est probablement idéalisée, en particulier en ce qui concerne la présence systématique et réguličre de côtes bouclées sur la région ventrale. La série de spécimens provenant du Tuffeau de Bourré (Loir-et-Cher) illustrée par Kennedy et al. (1980b ; 1984) et Amédro et Matrion (2018) permet d'apprécier la morphologie de la coquille ŕ différents stades de croissance. Au stade adulte qui correspond ŕ notre spécimen, Collignoniceras papale est caractérisé par l'atténuation, puis la disparition des tubercules ombilicaux tandis que les tubercules ventro-latéraux internes et externes fusionnent en une petite corne. Parallčlement, des côtes relient par paires sur la région ventrale les cornes ventro-latérales aux tubercules siphonaux en formant des boucles. De courtes intercalaires limitées ŕ la région ventrale, dont le nombre varie de une (le plus fréquemment) ŕ deux (plus rarement), se développent parfois entre les côtes bouclées.
Notre spécimen du Massif d'Uchaux est assez comparable morphologiquement ŕ la population tourangelle, mais correspond ŕ un variant sur lequel on observe une multiplication des tubercules siphonaux en raison de la présence systématique sur la région ventrale de deux courtes côtes intercalaires entre les côtes bouclées.
Distribution : Turonien moyen, Zone ŕ Romaniceras ornatissimum ; moitié supérieure de la Formation de Roustan dans le Massif d'Uchaux (unité UL3). Distribution en dehors du Massif d'Uchaux : Loir-et-Cher en France, Allemagne et République tchčque.
Collignoniceras turoniense (Sornay, 1951)
Pl. 8 ![]() , fig. 1
, fig. 1
1951 Prionotropis turoniense Sornay, p. 630, Pl. 21, figs. 1-3.
1980a Collignoniceras turoniense (Sornay) ; Kennedy, C.W. Wright & Hancock, p. 584, Pl. 71, figs. 4-5 ; Pl. 72, figs. 1-3.
2018 Collignoniceras turoniense (Sornay) ; Amédro & Matrion, p. 198, fig. 149 B.
2020a Collignoniceras turoniense (Sornay) ; Amédro et al., p. 58, Pl.
6 ![]() , fig. 3 ; Pl.
9
, fig. 3 ; Pl.
9 ![]() , figs. 3, 5 ; Pl.
10
, figs. 3, 5 ; Pl.
10 ![]() , figs. 1-2 ; Pl.
11
, figs. 1-2 ; Pl.
11 ![]() , fig. 2.
, fig. 2.
2020b Collignoniceras turoniense (Sornay) ; Amédro et al., Pl. 2, figs. 4, 6 ; Pl. 4, figs. 3, 6.
Type : l'holotype est, par monotypie, le spécimen décrit et illustré par Sornay (1951), p. 630, Pl. 21, figs. 1-3. Cette ammonite, de la collection Péron, est préservée dans les collections du Muséum national d'Histoire naturelle ŕ Paris, sous le numéro R 52555. Il provient du Tuffeau de Bourré dans sa localité type (Loir-et-Cher, France).
Matériel : un spécimen, coll. C.B. (PM 13), collecté dans l'intervalle 92 m-97 m. Un moulage est déposé dans les collections du Musée Requien ŕ Avignon (n° 2021.08.07).
Tableau 8 : Dimensions du Collignoniceras turoniense (Sornay, 1951) du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 8: Dimensions of Collignoniceras turoniense (Sornay, 1951) from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM13 |
Pl. 8 |
103 | 31 | 44 | 39 | 0,30 | 0,43 | 0,38 | 0,89 | 1,13 |
Discussion : Ŕ la différence de l'espčce précédente, Collignoniceras turoniense (Sornay, 1951) est caractérisé par le développement de tubercules ombilicaux coniques et massifs sur la totalité du phragmocône et la naissance de la chambre d'habitation. Elle diffčre en cela de Collignoniceras canthus (Orbigny, 1856) oů la quasi disparition de l'ornementation sur la chambre d'habitation la rend presque lisse. La multiplication des tubercules siphonaux sur le dernier quart de tour du phragmocône et sur la chambre d'habitation permet également de différencier aisément C. turoniense d'une autre espčce voisine : C. rhodanicum sp. nov, décrite ci-dessous, qui garde un nombre identique de tubercules ventro-latéraux et siphonaux jusqu'au péristome.
Le spécimen PM 13, illustré Pl. 8 ![]() ,
fig. 1, est un adulte de 103 mm de diamčtre montrant le phragmocône et l'intégralité de la chambre d'habitation. La section du tour, assez épaisse, est subtrapézoďdale sur le phragmocône avec une épaisseur maximale au niveau des tubercules ombilicaux saillants et une ligne siphonale surélevée, puis devient ovale sur la chambre suite ŕ l'atténuation des tubercules ombilicaux. L'ornementation est constituée de gros tubercules ombilicaux arrondis, au nombre de cinq par tour sur le phragmocône, ŕ partir desquels naissent des faisceaux de 2 ou 3 côtes longues, droites, entre lesquels on observe fréquemment une ou deux côtes courtes naissant assez bas sur le flanc. La région ventrale est malheureusement mal préservée sur le phragmocône. La chambre d'habitation est caractérisée par une multiplication des tubercules siphonaux qui deviennent plus arrondis et sont portés par de fines côtes bouclées reliant les tubercules ventro-latéraux. La principale différence par rapport aux exemplaires tourangeaux illustrés par Kennedy et al. (1980b), Amédro et Matrion
(2018) et Amédro et Châtelier (2020) est l'atténuation des tubercules ombilicaux au niveau de la chambre d'habitation et l'apparition sur celle-ci de côtes essentiellement longues, simples et flexueuses. Ce caractčre morphologique rapproche notre spécimen de Collignoniceras rhodanicum, en revanche la multiplication des tubercules siphonaux l'en sépare. Faut-il créer une nouvelle espčce pour cette ammonite ? Nous ne le pensons pas dans la mesure oů les Collignoniceratinae possčdent une trčs grande variabilité morphologique et montrent fréquemment une modification de l'ornementation sur la chambre d'habitation. Selon nous, ce spécimen représente une forme gracile de C. turoniense.
,
fig. 1, est un adulte de 103 mm de diamčtre montrant le phragmocône et l'intégralité de la chambre d'habitation. La section du tour, assez épaisse, est subtrapézoďdale sur le phragmocône avec une épaisseur maximale au niveau des tubercules ombilicaux saillants et une ligne siphonale surélevée, puis devient ovale sur la chambre suite ŕ l'atténuation des tubercules ombilicaux. L'ornementation est constituée de gros tubercules ombilicaux arrondis, au nombre de cinq par tour sur le phragmocône, ŕ partir desquels naissent des faisceaux de 2 ou 3 côtes longues, droites, entre lesquels on observe fréquemment une ou deux côtes courtes naissant assez bas sur le flanc. La région ventrale est malheureusement mal préservée sur le phragmocône. La chambre d'habitation est caractérisée par une multiplication des tubercules siphonaux qui deviennent plus arrondis et sont portés par de fines côtes bouclées reliant les tubercules ventro-latéraux. La principale différence par rapport aux exemplaires tourangeaux illustrés par Kennedy et al. (1980b), Amédro et Matrion
(2018) et Amédro et Châtelier (2020) est l'atténuation des tubercules ombilicaux au niveau de la chambre d'habitation et l'apparition sur celle-ci de côtes essentiellement longues, simples et flexueuses. Ce caractčre morphologique rapproche notre spécimen de Collignoniceras rhodanicum, en revanche la multiplication des tubercules siphonaux l'en sépare. Faut-il créer une nouvelle espčce pour cette ammonite ? Nous ne le pensons pas dans la mesure oů les Collignoniceratinae possčdent une trčs grande variabilité morphologique et montrent fréquemment une modification de l'ornementation sur la chambre d'habitation. Selon nous, ce spécimen représente une forme gracile de C. turoniense.
Distribution : Turonien moyen, sommet de la Zone ŕ Romaniceras kallesi (Tm 2) et Zone ŕ Romaniceras ornatissimum (Tm 3) ; moitié supérieure de la Formation de Roustan dans le Massif d'Uchaux (unité UL3). Distribution en dehors du Massif d'Uchaux : Vienne, Loir-et-Cher et Indre-et-Loire en France.
Collignoniceras rhodanicum Amédro, Baudouin & Delanoy sp. nov.
Pl. 10 ![]() ,
figs. 1-2
,
figs. 1-2
Type : l'holotype, PM 03 (coll. Cyril Baudouin), provient du Membre des siltites gréso-carbonatées et calcaires gréseux ŕ Spongia des Peyras (UL3) dans le gisement des Prés Morel, dans l'intervalle 98 m-110 m. Ce spécimen est déposé dans les collections du Musée Requien (n° 2021.08.03).
Autre matériel : deux exemplaires, coll. C.B. (PM 04 et 35), du męme niveau et de la męme localité. Un moulage du spécimen PM 04 est déposé dans les collections du Musée Requien (n° 2021.08.04).
Origine du nom : de rhodanicus, "qui navigue sur le Rhône" en latin (Gaffiot, 1934).
Tableau 9 : Dimensions de trois Collignoniceras rhodanicum Amédro, Baudouin & Delanoy sp. nov. du Turonien moyen
(Zone ŕ Romaniceras ornatissimum) du Massif d'Uchaux (Vaucluse).
Table 9: Dimensions of Collignoniceras rhodanicum Amédro, Baudouin & Delanoy sp.
nov. from middle Turonian (Romaniceras ornatissimum
Zone) of the Massif d'Uchaux (Vaucluse).
| Dimensions (mm) | Rapports | |||||||||
| n° | D | O | H | E | O/D | H/D | E/D | E/H | H/E | |
| PM03 |
Pl. 10 |
106 | 32 | 39 | - | 0,30 | 0,37 | - | - | - |
| 77 | 20 | 34 | 29 | 0,26 | 0,44 | 0,38 | 0,85 | 1,17 | ||
| PM04 |
Pl. 10 |
79 | 24 | 33 | - | 0,30 | 0,42 | - | - | - |
| 55 | 16 | 22 | 20 | 0,29 | 0,40 | 0,36 | 0,91 | 1,10 | ||
| PM35 | non figuré | 89 | 17 | 43 | 35 | 0,19 | 0,48 | 0,39 | 0,81 | 1,23 |
Diagnose : Collignoniceras comparable ŕ C. turoniense (Sornay, 1951) sur les tours internes du phragmocône, mais avec une ornementation plus robuste et surtout caractérisé par l'atténuation puis la disparition des tubercules ombilicaux sur la chambre d'habitation, ainsi que par la non multiplication des tubercules siphonaux sur le dernier tour de spire, ces derniers restant en nombre identique ŕ celui des tubercules ventro-latéraux.
Description : L'holotype, figuré Pl. 10 ![]() ,
fig. 1, est un moule interne de 106 mm de diamčtre, complet, avec la totalité du phragmocône et de la chambre d'habitation. Sur le phragmocône, la section du tour est légčrement plus haute que large (H/E = 1,17), subtrapézoďdale avec une région ventrale tectiforme et une épaisseur maximale au niveau des tubercules ombilicaux saillants. L'ornementation est constituée de gros tubercules ombilicaux coniques saillants au nombre de 6 ŕ 7 par tour, ŕ partir desquels naissent des faisceaux de deux, plus rarement trois, côtes longues, larges et droites. Une côte courte s'intercale parfois entre les côtes longues. Toutes les côtes portent un petit tubercule ventro-latéral interne arrondi, surtout visible sur les tours internes du phragmocône, et un tubercule ventro-latéral externe plus marqué, situé sur l'épaule ventro-latérale et pincé dans le sens de l'enroulement. La ligne siphonale, surélevée, porte une rangée de tubercules siphonaux fortement pincés dans le sens de l'enroulement, en nombre identique aux tubercules ventro-latéraux, mais légčrement décalés vers l'avant. Quatorze tubercules siphonaux sont visibles sur le dernier demi-tour de spire du phragmocône. Sur la chambre d'habitation, l'ornementation change. Les tubercules ombilicaux disparaissent, rendant la coquille plus comprimée, tandis que toutes les côtes deviennent longues, simples et légčrement flexueuses.
,
fig. 1, est un moule interne de 106 mm de diamčtre, complet, avec la totalité du phragmocône et de la chambre d'habitation. Sur le phragmocône, la section du tour est légčrement plus haute que large (H/E = 1,17), subtrapézoďdale avec une région ventrale tectiforme et une épaisseur maximale au niveau des tubercules ombilicaux saillants. L'ornementation est constituée de gros tubercules ombilicaux coniques saillants au nombre de 6 ŕ 7 par tour, ŕ partir desquels naissent des faisceaux de deux, plus rarement trois, côtes longues, larges et droites. Une côte courte s'intercale parfois entre les côtes longues. Toutes les côtes portent un petit tubercule ventro-latéral interne arrondi, surtout visible sur les tours internes du phragmocône, et un tubercule ventro-latéral externe plus marqué, situé sur l'épaule ventro-latérale et pincé dans le sens de l'enroulement. La ligne siphonale, surélevée, porte une rangée de tubercules siphonaux fortement pincés dans le sens de l'enroulement, en nombre identique aux tubercules ventro-latéraux, mais légčrement décalés vers l'avant. Quatorze tubercules siphonaux sont visibles sur le dernier demi-tour de spire du phragmocône. Sur la chambre d'habitation, l'ornementation change. Les tubercules ombilicaux disparaissent, rendant la coquille plus comprimée, tandis que toutes les côtes deviennent longues, simples et légčrement flexueuses.
Le second spécimen illustré Pl. 10 ![]() ,
fig. 2 (PM 04), possčde un diamčtre de 79 mm. Il s'agit également d'une coquille entičre, mais dont la chambre d'habitation est moins bien préservée. La morphologie générale de la coquille est comparable ŕ celle de l'holotype. En revanche la costulation est ici majoritairement constituée de côtes longues et simples, męme sur le phragmocône et non de côtes naissant par faisceaux de deux ŕ partir des tubercules ombilicaux. De tels faisceaux sont uniquement observés ŕ deux reprises sur le dernier tour de spire.
,
fig. 2 (PM 04), possčde un diamčtre de 79 mm. Il s'agit également d'une coquille entičre, mais dont la chambre d'habitation est moins bien préservée. La morphologie générale de la coquille est comparable ŕ celle de l'holotype. En revanche la costulation est ici majoritairement constituée de côtes longues et simples, męme sur le phragmocône et non de côtes naissant par faisceaux de deux ŕ partir des tubercules ombilicaux. De tels faisceaux sont uniquement observés ŕ deux reprises sur le dernier tour de spire.
Discussion : Collignoniceras rhodanicum Amédro, Baudouin & Delanoy sp. nov. fait clairement partie du męme groupe d'espčces que C. canthus (Orbigny, 1856) et C. turoniense (Sornay, 1951) par l'aspect tectiforme de la région ventrale. Elle en diffčre cependant par la costulation du tour externe lorsque l'on se rapproche du péristome, avec la présence de côtes longues, simples et flexueuses. Chez Collignoniceras canthus, il y a quasi disparition de l'ornementation sur la chambre d'habitation, les tubercules siphonaux et les fines côtes restant ŕ peine visibles sur une coquille devenue presque lisse. Les variants robustes de Collignoniceras turoniense (cf. Amédro & Châtelier, 2020, Pl.
9 ![]() , fig. 3) sont assez comparables ŕ C. rhodanicum, tout au moins en ce qui concerne le phragmocône. Mais, que ce soit chez les formes comprimées, moyennes ou épaisses de C. turoniense, la chambre d'habitation est toujours caractérisée par, premičrement, la présence de tubercules ombilicaux coniques et massifs et, deuxičmement, une multiplication des tubercules siphonaux devenus arrondis et qui sont portés par de fines côtes bouclées naissant par paires au niveau des tubercules ventro-latéraux (Kennedy et al.,
1980b ; Amédro & Matrion,
2018 ; Amédro & Châtelier, 2020) alors que chez C. rhodanicum, les tubercules siphonaux et ventro-latéraux restent en nombre identique.
, fig. 3) sont assez comparables ŕ C. rhodanicum, tout au moins en ce qui concerne le phragmocône. Mais, que ce soit chez les formes comprimées, moyennes ou épaisses de C. turoniense, la chambre d'habitation est toujours caractérisée par, premičrement, la présence de tubercules ombilicaux coniques et massifs et, deuxičmement, une multiplication des tubercules siphonaux devenus arrondis et qui sont portés par de fines côtes bouclées naissant par paires au niveau des tubercules ventro-latéraux (Kennedy et al.,
1980b ; Amédro & Matrion,
2018 ; Amédro & Châtelier, 2020) alors que chez C. rhodanicum, les tubercules siphonaux et ventro-latéraux restent en nombre identique.
Distribution : Turonien moyen, Zone ŕ Romaniceras ornatissimum ; moitié supérieure de la Formation de Roustan dans le Massif d'Uchaux (unité UL3).
Les sexes sont séparés chez les céphalopodes actuels et de nombreuses espčces sont dimorphiques, avec des différences plus ou moins marquées. La possibilité d'un dimorphisme d'origine sexuel chez les ammonites a été perçue dčs le XIXe sičcle par de Blainville (1840). Les publications d'Arkell et al. (1957), Makowski (1962) et Callomon (1963) ont été par la suite déterminantes dans l'acceptation de cette hypothčse. Depuis, de trčs nombreux travaux ont envisagé ou démontré l'existence de couples dimorphiques au sein d'un nombre important de genres d'ammonites jurassiques et crétacées. Le dimorphisme peut se manifester sous diverses maničres et les deux morphes (les microconches qui pourraient correspondre ŕ des mâles et les macroconches qui pourraient représenter des femelles), peuvent diverger par la taille, la morphologie du péristome, de la coquille ou l'ornementation de certaines parties de celle-ci. Nous renvoyons au travail de Klug et al. (2015) pour l'important inventaire, avec références, des ammonites jurassiques et crétacées chez lesquelles un dimorphisme d'ordre sexuel est envisagé.
En ce qui concerne la sous-famille des Collignoniceratinae, Kennedy et al. (1984) ont mis en évidence un dimorphisme se rapportant ŕ la taille des individus au sein de l'espčce Collignoniceras papale (Orbigny, 1841). Les macroconches sont adultes ŕ 150 mm, les microconches ŕ 75 % de ce diamčtre. Dans les deux cas, la terminaison de la chambre d'habitation est marquée par un déclin brutal de l'ornementation qui est réduite ŕ de faibles côtes sur la partie externe du flanc et le ventre.
Plus récemment, un dimorphisme a également été observé chez Collignoniceras woollgari regulare (Haas,
1946) par Kennedy et al. (2001). Sur la plus grande partie de la chambre d'habitation, les côtes, en nombre quasi identique ŕ celui des tubercules siphonaux, restent simples et les tubercules ventro-latéraux se développent en cornes proéminentes. Dans un premier groupe, les individus, interprétés comme microconches, deviennent adultes vers 200 mm de diamčtre. Il s'agit le plus souvent de formes graciles oů la partie terminale de la chambre d'habitation avant l'ouverture tend ŕ devenir lisse et tubulaire. Dans un second groupe, les individus interprétés comme macroconches, ont des chambres d'habitation qui atteignent jusqu'ŕ 360 mm de diamčtre. Leur morphologie correspond ŕ des formes robustes. Le spécimen PM 17, illustré
Pl. 12 ![]() , fig. 1, possčde un diamčtre maximal de 119 mm, celui du phragmocône atteignant 98 mm. Sur la premičre moitié de la chambre d'habitation se trouvent des côtes saillantes supportant des tubercules ombilicaux pincés radialement et des tubercules ventro-latéraux bien marqués. Cette ornementation change au diamčtre de 106 mm et, sur la seconde partie de la chambre, l'ornementation n'est plus constituée que de côtes simples de plus en plus inclinées vers l'avant et qui s'atténuent. Il s'agit trčs probablement d'une forme microconche.
, fig. 1, possčde un diamčtre maximal de 119 mm, celui du phragmocône atteignant 98 mm. Sur la premičre moitié de la chambre d'habitation se trouvent des côtes saillantes supportant des tubercules ombilicaux pincés radialement et des tubercules ventro-latéraux bien marqués. Cette ornementation change au diamčtre de 106 mm et, sur la seconde partie de la chambre, l'ornementation n'est plus constituée que de côtes simples de plus en plus inclinées vers l'avant et qui s'atténuent. Il s'agit trčs probablement d'une forme microconche.
Notre matériel permet aujourd'hui de décrire un dimorphisme au sein de Collignoniceras carolinum (Orbigny, 1841). Le diamčtre des adultes révčle l'existence de deux groupes d'individus interprétés comme des microconches et des macroconches. De façon anecdotique, tous les représentants ŕ peu prčs complets de l'espčce figurés dans la littérature sont des microconches (Kennedy et al., 1980b, Pl. 68, figs. 1-3 ; Pl. 76, figs. 1-2 ; Kennedy & Kaplan, 2019, Pl. 35, fig. 3). Les caractčres distinctifs des deux formes apparaissent comme suit.
Formes microconches : la taille adulte maximale se situe aux alentours de 120 mm, le diamčtre atteint ŕ la fin du phragmocône étant compris entre 70 et 100 mm. Un changement d'ornementation est observé ŕ la fin du phragmocône/début de la chambre d'habitation, caractérisé par :
l'atténuation plus ou moins prononcée, voire la quasi disparition de la costulation sur les flancs ;
la disparition des tubercules ombilicaux et des tubercules ventro-latéraux internes ;
la transformation des tubercules ventro-latéraux externes en clavi ;
l'affaiblissement de la carčne ;
et enfin le développement de fines côtes vers le péristome, accompagné d'une forte projection vers l'avant des clavi.
Les spécimens PM 01 (Pl. 5 ![]() , fig. 2), 07
(Pl. 5
, fig. 2), 07
(Pl. 5 ![]() , fig. 1), 08, 13, 19, 22
(Pl. 3
, fig. 1), 08, 13, 19, 22
(Pl. 3 ![]() ,
fig. 2), 28 (Pl. 2
,
fig. 2), 28 (Pl. 2 ![]() , fig. 2), 30
(Pl. 6
, fig. 2), 30
(Pl. 6 ![]() , fig. 2), 31, 32a
(Pl. 9
, fig. 2), 31, 32a
(Pl. 9 ![]() , fig. 2), 32b, 36 et 37 sont considérés comme des microconches.
, fig. 2), 32b, 36 et 37 sont considérés comme des microconches.
Formes macroconches : la taille adulte maximale est légčrement supérieure ŕ 180 mm. Ŕ la différence des microconches, l'ornementation reste vigoureuse sur la chambre d'habitation, mais les côtes, quelque peu flexueuses et inclinées vers l'avant, ne portent plus que les tubercules ventro-latéraux externes. Ces tubercules sont maintenant pincés dans le sens de l'enroulement en clavi et obliques par rapport ŕ la ligne siphonale. La région prépéristomale est marquée par un affaiblissement plus ou moins important de la costulation. Les spécimens PM 02
(Pl. 3 ![]() fig. 1), 05 (Pl. 6
fig. 1), 05 (Pl. 6 ![]() , fig. 1), 06
(Pl. 4
, fig. 1), 06
(Pl. 4 ![]() , fig. 1), 18, 20, 21
(Pl. 7
, fig. 1), 18, 20, 21
(Pl. 7 ![]() , fig. 1) et 23
(Pl. 16
, fig. 1) et 23
(Pl. 16 ![]() , fig. 1) sont interprétés comme des macroconches.
, fig. 1) sont interprétés comme des macroconches.
Si les microconches présentent une certaine homogénéité ornementale et morphologique et correspondent le plus souvent des formes graciles de façon comparable ŕ ce que l'on observe chez C. woollgari regulare, les formes macroconches montrent une variabilité plus importante qui s'exprime principalement ŕ travers l'ornementation généralement plus robuste et la section du tour plus massive.
Un dimorphisme existe également au sein d'autres familles ou sous-familles d'ammonites du Crétacé supérieur, en particulier chez les Euomphaloceratinae. Kennedy et al. (1980a) ont ainsi suggéré que l'espčce Romaniceras (Yubariceras) ornatissimum (Stoliczka, 1864) pourrait ętre dimorphique, les microconches étant représentés par la forme inerme Grossouvre, 1889 qui perd toute tuberculation au profit de la costulation au-delŕ d'un diamčtre de 100 mm, tandis que les macroconches restent toujours tuberculés (forme ornatissimum typique et forme deverioides Grossouvre, 1889).
L'espčce Romaniceras (Romaniceras) marigniacum décrite par Amédro et Châtelier (2020) en Touraine méridionale et dont quatre exemplaires viennent d'ętre récoltés dans le Massif d'Uchaux est probablement aussi dimorphique. Les spécimens tourangeaux atteignent leur taille adulte ŕ un diamčtre compris entre 200 et 230 mm (Amédro et al.,
2020a, Pl.
2 ![]() , figs. 1-2 ; Pl.
4
, figs. 1-2 ; Pl.
4 ![]() , fig. 1 ; Pl.
5
, fig. 1 ; Pl.
5 ![]() , fig. 1). L'ornementation est constituée sur la chambre d'habitation de côtes fortes, droites et radiales, alternativement longues et courtes. Les moules internes PM 10 et PM 11 (ce spécimen est illustré
Pl. 13
, fig. 1). L'ornementation est constituée sur la chambre d'habitation de côtes fortes, droites et radiales, alternativement longues et courtes. Les moules internes PM 10 et PM 11 (ce spécimen est illustré
Pl. 13 ![]() ,
fig. 1), collectés aux Prés Morel, sont des spécimens presque complets et de dimension adulte sensiblement équivalente aux formes de Touraine. Ces exemplaires sont interprétés comme des macroconches.
,
fig. 1), collectés aux Prés Morel, sont des spécimens presque complets et de dimension adulte sensiblement équivalente aux formes de Touraine. Ces exemplaires sont interprétés comme des macroconches.
Ŕ l'opposé, chez le spécimen PM 09 des Prés Morel (Pl. 14 ![]() , fig. 1), la taille adulte est atteinte au diamčtre de 119 mm. La section du tour est plus comprimée que dans les spécimens précédents et on observe la disparition brutale de la costulation dans la région péripéristomale oů celle-ci est remplacée par des bourrelets de faible relief et de fines stries. Par ces caractéristiques morphologiques et ornementales, ce spécimen peut vraisemblablement ętre considéré comme représentant la forme microconche de R. (R.) marigniacum. Un réexamen des exemplaires de taille réduite de la population de Touraine (Amédro et al.,
2020a, Pl. 1
, fig. 1), la taille adulte est atteinte au diamčtre de 119 mm. La section du tour est plus comprimée que dans les spécimens précédents et on observe la disparition brutale de la costulation dans la région péripéristomale oů celle-ci est remplacée par des bourrelets de faible relief et de fines stries. Par ces caractéristiques morphologiques et ornementales, ce spécimen peut vraisemblablement ętre considéré comme représentant la forme microconche de R. (R.) marigniacum. Un réexamen des exemplaires de taille réduite de la population de Touraine (Amédro et al.,
2020a, Pl. 1 ![]() , fig. 2) serait nécessaire pour mettre en évidence les formes microconches au sein de cette population.
, fig. 2) serait nécessaire pour mettre en évidence les formes microconches au sein de cette population.
Monsieur Jean-François Deconinck (Dijon) a analysé deux prélčvements provenant des "Marnes ŕ Spongia" (calcimétrie, fraction argileuse et lames minces). Le Dr William James Kennedy (Oxford, UK) nous a donné son avis sur la détermination de plusieurs Collignoniceras et Romaniceras. Monsieur Hervé Châtelier (Rennes) nous a communiqué de précieuses informations sur la population de Romaniceras (R.) marigniacum recueillie par ses soins en Touraine méridionale. Monsieur Bertrand Matrion (Troyes) nous a transmis des moulages des types de Collignoniceras carolinum. Messieurs Serge Ferry (Briançon) et William James Kennedy ont bien voulu relire cet article en y apportant de nombreuses remarques constructives. Que tous en soient chaleureusement remerciés.
Amédro F. & Châtelier H. (2020).- Paléontologie systématique : Description des ammonites. In : Amédro F., Robaszynski F., Châtelier H., Ferchaud P. & Matrion B., Identification d'un biohorizon d'ammonites ŕ Romaniceras (Romaniceras) marigniacum sp. nov. (Turonien moyen) ŕ la base du Tuffeau Jaune de Touraine (France).- Carnets Geol., Madrid, vol. 20, n° 4, p. 48-63 (11 Pls.). DOI : 10.4267/2042/70720
Amédro F. & Devalque C. (2014).- Les ammonites in Robaszynski F., Amédro F., Devalque C. & Matrion B.- Le Turonien des massifs d'Uchaux et de la Cčze (S.E. France). Migration globale d'ammonites et conséquences sur la zonation internationale, rudistes et corrélations entre les massifs.- Académie Royale de Belgique, Mémoires de la Classe des Sciences, Bruxelles, 2, 197 p. (48 Pls.).
Amédro F. & Matrion B. (2018).- Les ammonites in Amédro F., Matrion B. & Robaszynski F. ( coord.), Stratotype Turonien.- Patrimoine géologique, Paris, Mčze, vol. 8, p. 183-244.
Amédro F., Matrion B. & Robaszynski F. (coord., 2018).- Stratotype Turonien.- Patrimoine géologique, Paris, Mčze, vol. 8, 416 p.
Amédro F., Robaszynski F., Châtelier H., Ferchaud P. & Matrion B. (2020a).- Identification d'un biohorizon d'ammonites ŕ Romaniceras (Romaniceras) marigniacum sp. nov. (Turonien moyen) ŕ la base du Tuffeau Jaune de Touraine (France).- Carnets Geol., Madrid, vol. 20, n° 4, p. 37-89 (11 Pls.). DOI : 10.4267/2042/70720
Amédro F., Robaszynski F., Châtelier H., Ferchaud P., Matrion B. & Bréhéret J.-G. (2020b).- Des nouveautés sur les tuffeaux du Turonien moyen et sur leurs faunes d'ammonites en Touraine méridionale.- Bulletin d'Information des Géologues du Bassin de Paris, vol. 57, n° 2, p. 3-33 (4 Pls.).
Arkell W.J., Kummel B. & Wright C.W. (1957).- Mesozoic Ammonoidea. In : Moore R.C. (ed.), Treatise on Invertebrate Paleontology. Part L, Mollusca 4, Cephalopoda, Ammonoidea.- Geological Society of America, Boulder - CO, 489 p.
Blainville M.H. de (1840).- Prodrome d'une monographie des ammonites. In : Supplément du Dictionnaire des Sciences naturelles.- Bertrand, Paris, 131 p.
Breistroffer M. (1947).- Sur les zones d'ammonites de l'Albien de France et d'Angleterre.- Travaux du Laboratoire de Géologie de l'Univrsité de Grenoble, vol. 26, p. 1-88.
Callomon J.H. (1963).- Sexual dimorphism in Jurassic ammonites.- Transactions of the Leicester Literary and Philosophical Society, vol. 57, p. 21-56.
Cobban W.A. & Hook S.C. (1979).- Collignoniceras woollgari woollgari (Mantell) ammonite faunas from the Upper Cretaceous of Western Interior, United States.- New Mexico Bureau of Mines and Mineral Resources Memoir, Albuquerque - NM, n° 37, 51 p. (12 Pls.).
Cooper M.R. (1978).- Uppermost Cenomanian-basal Turonian ammonites from Salinas, Angola.- Annals of the South African Museum, Cape Town, vol. 75, p. 51-152.
Fieber I. (1853).- Synopsis der europäischen Orthopteren mit besonderer Rücksicht der böhmischen Arten.- Lotos, 3, p. 90-261.
Gaffiot F. (1934).- Dictionnaire illustré Latin-Français.- Hachette, Paris, 1720 p.
Gale A.S (1996).- Turonian correlation and sequence stratigraphy of the chalk in southern England. In : Hesselbo S.P. & Parkinson D.N. (ed.), Sequence stratigraphy in British geology.- Geological Society, London, Special Publications, vol. 103, p. 177-195.
Gauthier H. (2006).- Révision critique de la Paléontologie française d'Alcide d'Orbigny. Volume IV - Céphalopodes Crétacés.- Backhuys, Leiden, 292 p. (65 Pls.).
Grossouvre A. de (1889).- Sur le terrain crétacé dans le sud-ouest du Bassin de Paris.- Bulletin de la Société géologique de France (3e Série), Paris, tome 17, p. 475-525 (Pl. 12). URL : https://patrimoine.sorbonne-universite.fr/idurl/1/2701
Grossouvre A. de (1894).- Recherches sur la Craie supérieure. Deuxičme partie. Paléontologie. Les ammonites de la Craie supérieure.- Mémoires du Service de la Carte géologique détaillée de la France, Paris, 264 p. (39 Pls.). URL : https://gallica.bnf.fr/ark:/12148/bpt6k6302080k
Haas O. (1946).- Intraspecific variation in, and ontogeny of Prionotropis woollgari and Prionocyclus wyomingensis.- American Museum of Natural History Bulletin, n° 86, p. 141-224 (Pls. 11-24).
Hébert E. & Toucas A. (1875).- Description du Bassin d'Uchaux.- Annales des Sciences Géologiques, Paris, tome VI, 112 p. (3 Pls.).
Hyatt A. (1889).- Genesis of the Arietidae.- Smithsonian Contributions to Knowledge, Washington - DC, vol. 26, 238 p. (14 Pls.).
Kaplan U. (1988).- Die Ammoniten-Subfamilie Collignoniceratinae Wright & Wright, 1951 aus dem Turon (Ober-Kreide) von Westfalen und Niedersachsen (NW-Deutschland).- Geologie und Paläontologie in Westfalen, Münster, 12, p. 5-45 (7 Pls.).
Kennedy W.J., Amédro F., Badillet G., Hancock J.M. & Wright C.W. (1984).- Notes on Late Cenomanian and Turonian ammonites from Touraine, Western France.- Cretaceous Research, vol. 5, p. 29-45.
Kennedy W.J., Bilotte M. & Melchior P. (2015).- Turonian ammonite faunas from the southern Corbičres, Aude, France.- Acta Geologica Polonica, Varsaw, vol. 65, p. 437-494. URL : https://geojournals.pgi.gov.pl/agp/article/view/25693
Kennedy W.J. & Cobban W.A. (1976).- Aspects of Ammonite biology, biogeography and biostratigraphy.- Palaeontological Association London, Special papers in Palaeontology, n° 17, p. 1-94 (11 Pls.). URL : https://www.palass.org/publications/special-papers-palaeontology/archive/17/article_pp1-94
Kennedy W.J., Cobban W.A. & Landman N.H. (2001).- A revision of the Turonian members of the ammonite subfamily Collignoniceratinae from the United States Western Interior and Gulf Coast.- Bulletin of the American Museum of Natural History, New York - NY, 148 p.
Kennedy W.J. & Gale A.S. (2016).- Turonian ammonites from northwestern Aquitaine, France.- Cretaceous Research, vol. 58, p. 265-296.
Kennedy W.J. & Kaplan U. (2019).- Ammoniten aus dem Turonium des Münsterländer Kreidebeckens.- Geologie und Paläontologie in Westfalen, Münster, 92, p. 3-223 (52 Pls.).
Kennedy W.J. & Wright C.W. (1981).- Desmoceratacean ammonites from the type Turonian.- Palaeontology, London, vol. 24, p. 493-506 (Pls. 74-76).
Kennedy W.J., Wright C.W. & Hancock J.M. (1980a).- The European species of the Cretaceous ammonite Romaniceras with a revision of the genus.- Palaeontology, London, vol. 23, p. 325-362 (Pls. 39-50).
Kennedy W.J., Wright C.W. & Hancock J.M. (1980b).- Collignoniceratid ammonites from the Mid-Turonian of England and northern France.- Palaeontology, London, vol. 23, p. 557-603 (Pls. 62-77).
Kirkland J.I. (1996).- Paleontology of the Greenhorn Cyclothem (Cretaceous: Late Cenomanian to Middle Turonian) at Black Mesa, Northeastern Arizona.- New Mexico Museum of Natural History and Science Bulletin, n° 9, 131 p.
Klug C., Zaton M., Parent H., Hostettler B. & Tajika A. (2015).- Mature modifications and sexual dimorphism. In : LKlug C., Korn D., De Baets D., Kruta K. & Mapes R.H. (eds), Ammonoid paleobiology: From anatomy to ecology.- Topics in Geobiology, vol. 43, p. 253-320.
Makowski H. (1962).- Problem of sexual dimorphism in ammonites.- Palaeontologica Polonia, Varszawa, n° 12, p. 1-92. URL : http://www.palaeontologia.pan.pl/Archive/1962-12.pdf
Mantell G. (1822).- The fossils of the South Downs; or illustrations of the Geology of Sussex.- Rolfe, London, 327 p. (42 Pls.).
Matsumoto T., Saito R. & Fukada A. (1957).- Some Acanthoceratids from Hokkaido.- Memoirs of the Faculty of Science, Kyushu University (D, Geology), vol. 6, 45 p. (18 Pls.).
Meek F.B. (1876).- A report on the invertebrate Cretaceous and Tertiary fossils of the upper Missouri country.- Report of the United States Geological Survey of the Territories, Washington - DC, vol. IX, lxiv + 629 p. (45 Pls.).
Mennessier G. (1950).- Monographie géologique du massif d'Uchaux.- Bulletin de la Carte géologique de la France et des Topographies souterraines, Paris, n° 227, 50 p. (2 Pls.).
Orbigny A. d' (1840-1842).- Paléontologie française. Terrains Crétacés. Tome I : Céphalopodes.- Masson, Paris, p. 1-120 (1840) ; p. 121-430 (1841) ; p. 431-662 (1842) (148 Pls.).
Orbigny A. d' (1847-1851).- Paléontologie française. Terrains Crétacés. Tome IV : Brachiopodes.- Masson, Paris, p. 1-32 (1847) ; p. 33-104 (1849) ; p. 105-390 (1851) (Pls. 513-599).
Orbigny A. d' (1849-1852).- Cours élémentaire de paléontologie et de géologie stratigraphiques.- Masson, Paris, 3 volumes : tome I (1849), 299 p. ; tome II, fasc. I (1851), 382 p. ; tome II, fasc. II (1852), 464 p.
Orbigny A. d' (1850).- Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson, Paris, tome II, 428 p.
Robaszynski F. (coord.), Alcaydé G., Amédro F., Badillet G., Damotte R., Foucher J.-C., Jardiné S., Legoux O., Manivit H., Monciardini C. & Sornay J. (1982).- Le Turonien de la région-type : Saumurois et Touraine. Stratigraphie, biozonations, sédimentologie.- Bulletin des Centres de Recherche, Exploration-Production d'elf-Aquitaine, Pau, volume 6, n° 1, p. 119-225 (18 Pls.).
Robaszynski F., Amédro F., Devalque C. & Matrion B. (2014).- Le Turonien des massifs d'Uchaux et de la Cčze (SE France). Migration globale d'ammonites et conséquences sur la zonation internationale, rudistes et corrélations entre les massifs.- Académie royale de Belgique, Mémoire de la Classe des Sciences (IVe Série), Bruxelles, tome 2, 197 p. (48 Pls.).
Robaszynski F., Amédro F. (coord.), Foucher J.-C., Gaspard D., Magniez-Jannin F., Manivit H. & Sornay J. (1980).- Synthčse biostratigraphique de l'Aptien au Santonien du Boulonnais ŕ partir de sept groupes paléontologiques : Foraminifčres, nannoplancton, dinoflagellés et macrofaunes. Zonations micropaléontologiques intégrées dans le cadre du Crétacé boréal nord-européen.- Revue de Micropaléontologie, Paris, vol. 22, n° 4, p. 195-321 (20 Pls.).
Roman F. & Mazeran P. (1913).- Monographie paléontologique de la faune du Turonien du bassin d'Uchaux et de ses dépendances.- Archives du Muséum d'Histoire naturelle de Lyon, tome 12, p. 1-160 (11 Pls.). URL: https://www.persee.fr/doc/mhnly_0374-5465_1920_num_12_1_969
Sornay J. (1950).- Étude stratigraphique sur le Crétacé supérieur de la vallée du Rhône entre Valence et Avignon et des régions voisines.- Thčse, Allier, Grenoble, 254 p. ; Travaux du Laboratoire de Géologie de la Faculté des Sciences de l'Université de Grenoble, vol. 27, p. 35-278.
Sornay J. (1951).- Sur deux espčces d'ammonites inédites de d'Orbigny et sur une nouvelle espčce du Tuffeau de Touraine.- Bulletin de la Société Géologique de France (6e Série), Paris, tome 1, n° 8, p. 627-631 (Pl. 21).
Spath L.F. (1922).- On the Senonian ammonite faunas from Pondoland.- Transactions of the Royal Society of South Africa, Cape Town, vol. 10, p. 113-147 (Pls. 5-9).
Spath L.F. (1923).- On the ammonite horizons of the Gault and contiguous deposits.- Summary of Progress of the Geological Survey, London, p. 139-149.
Spath L.F. (1939).- Problems of ammonite nomenclature (6). The genus Pachydiscus Zittel.- Geological Magazine, Cambridge - MA, vol. 76, p. 293-296.
Stoliczka F. (1863-1866).- The fossil cephalopoda of the Cretaceous rocks of southern India: Ammonitidae, with revision of the Nautilidae.- Memoirs of the Geological Survey of India, Palæontologica Indica, Calcutta, 216 p. (94 Pls.).
Voigt S. & Wiese F. (2000).- Evidence for Late Cretaceous (Late Turonian) climate cooling from oxygen-isotope variations and palaeobiogeographic changes in Western and Central Europe.- Journal of the Geological Society, London, vol. 157, p. 737-743.
Wilmsen M. & Nagm E. (2014).- Die Kreide in Sachsen. 7 - Ammoniten.- Geologica Saxonica, Dresden, vol. 60, n° 1, p. 201-240 (15 Pls.).
Wright C.W. (1996).- Cretaceous Ammonoidea (with contributions by Callomon J.H. and Howarth M.K.). In : Kaesler R.L., Treatise on Invertebrate Paleontology. Part L, Mollusca 4 Revised.- Geological Society of America, Boulder - CO, 362 p.
Wright C.W. & Kennedy W.J. (1981).- The Ammonoidea of the Plenus Marls and the Middle Chalk.- Monographs of the Palaeontographical Society, London, vol. 134, n° 2, 148 p. (32 Pls.).
Wright C.W. & Wright E.V. (1951).- A survey of the fossil Cephalopoda of the chalk of Great Britain.- Monographs of the Palaeontographical Society, London, vol. 105, 40 p.
Zittel K.A. (1884).- Handbuch der Palaeontologie, Cephalopoda.- Oldenbourg, München & Leipzig, p. 329-522.
Zittel K.A. (1895).- Grundzüge der Palaeontologie.- Oldenbourg, München & Leipzig, 972 p.
Zanova T.D. & Yazykova E.A. (1998).- Biostratigraphy and correlation of the Turonian-Coniacian succession and the Turonian-Coniacian boundary problem in the Far East Russia based on ammonites and inoceramids.- Acta Geologica Polonica, Warszawa, vol. 48, n° 4, p. 483-494 (14 Pls.).
Planche
1 :
Figs. 1-2 :
Lewesiceras peramplum (Mantell,
1822), 1 : PM 29 (coll.
C.B.) ; 2 : PM 39 (coll. C.B.), les deux spécimens proviennent du gisement des Prés Morel ŕ Mondragon
(Vaucluse), Formation de Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
2 :
Fig. 1 :
Lewesiceras peramplum (Mantell,
1822), PM 25 (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
3 :
Figs. 1-2 : Collignoniceras carolinum (Orbigny, 1841), 1 : PM 02 (coll.
C.B.), spécimen macroconche ; 2 : PM 22 (coll. C.B.), spécimen
microconche. Les deux proviennent du gisement des Prés Morel ŕ Mondragon (Vaucluse) dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
4 :
Fig. 1 : Collignoniceras carolinum (Orbigny, 1841), PM 06 (coll.
C.B.), spécimen macroconche, du gisement des Prés Morel ŕ Mondragon (Vaucluse), récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
5 :
Figs. 1-2 : Collignoniceras carolinum (Orbigny, 1841), 1: PM 07 (coll.
C.B.) ; 2: PM 01. Les deux spécimens sont des microconches qui proviennent du gisement des Prés Morel ŕ Mondragon
(Vaucluse) dans l'intervalle 98 m-110 m, Formation de Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
6 :
Figs. 1-2 : Collignoniceras carolinum (Orbigny, 1841), 1 : PM 05 (coll.
C.B.), spécimen macroconche ; 2 : PM 30 (coll. C.B.), spécimen
microconche. Les deux spécimens proviennent du gisement des Prés Morel ŕ Mondragon
(Vaucluse) dans l'intervalle 98 m-110 m, Formation de Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
7 :
Fig. 1 : Collignoniceras carolinum (Orbigny, 1841), PM 21 (coll.
C.B.), spécimen macroconche, du gisement des Prés Morel ŕ Mondragon (Vaucluse), récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
8 :
Fig. 1 : Collignoniceras turoniense (Sornay, 1951), PM 13 (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), spécimen récolté dans la vigne située en contrebas de la terrasse réalisée en 2019, dans l'intervalle 92 m-97 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
9 :
Fig. 1 : Angulithes sowerbyanus (Orbigny, 1840), PM 32c (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), nautile récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
10 :
Figs. 1-2
: Collignoniceras rhodanicum Amédro, Baudouin & Delanoy sp.
nov., 1 : PM 03 : holotype (coll. C.B.) ; 2 : PM 04 (coll.
C.B.). Les deux spécimens proviennent du gisement des Prés Morel ŕ Mondragon
(Vaucluse), dans l'intervalle 98 m-110 m, Formation de Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
11 :
Fig. 1: Collignoniceras papale (Orbigny, 1841), PM 14 (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), spécimen récolté dans la vigne située en contrebas de la terrasse réalisée en 2019, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
12 :
Fig. 1:
Collignoniceras woollgari regulare (Haas,
1946), PM 17 (coll.
C.B.), spécimen microconche, du gisement des Prés Morel ŕ Mondragon (Vaucluse), récolté dans la vigne située en contrebas de la terrasse réalisée en 2019, dans l'intervalle 92 m-97 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
13 :
Fig. 1 :
Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, PM 11 (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), spécimen récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
14 :
Fig. 1 : Romaniceras (Romaniceras) marigniacum Amédro & Châtelier, 2020, PM 09 (coll.
C.B.), spécimen microconche, du gisement des Prés Morel ŕ Mondragon (Vaucluse), récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
15 :
Fig. 1 : Romaniceras (Yubariceras) ornatissimum (Stoliczka,
1864), PM 26 (coll.
C.B.), du gisement des Prés Morel ŕ Mondragon (Vaucluse), spécimen récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
16 :
Fig. 1 : Collignoniceras carolinum (Orbigny, 1841), PM 23 (coll.
C.B.), spécimen macroconche, du gisement des Prés Morel ŕ Mondragon (Vaucluse), récolté dans l'intervalle 98 m-110 m, Formation de
Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
Planche
17 :
Fig. 1 :
Romaniceras (Yubariceras) kanei Jones, 1938, spécimen non numéroté (coll. C.D.), du gisement des Prés Morel ŕ Mondragon
(Vaucluse), récolté au niveau 75 m, Formation de Roustan, Membre des siltites et marnes sableuses ŕ Spongia des Peyras (UL3), Turonien
moyen, Zone ŕ Romaniceras ornatissimum. |
|
| Nomenclatural note: Life Sciences Identifier (LSID) https://zoobank.org/References/FF665394-ABB4-4698-B5F1-F2248FCDCCC8 Species Group • Collignoniceras rhodanicum Amédro, Baudouin & Delanoy in Amédro et al., 2022 https://zoobank.org/NomenclaturalActs/80FA0172-E0D8-4A2D-8CEE-F4E3A844B14C |