◄ Carnets Geol. 22 (8) ►
![]()
Outline:
[1. Introduction]
[2. Material and methods]
[3. Petrographic description of the material studied]
[4. Paragenetic sequence]
[5. Distorted ooids or collapsed molds]
[6. Conclusion]
[Bibliographic references]
[Appendix]
and ...
[Plates]
Dépt. STU, Fac. Sci. Tech., UBO, 6 avenue Le Gorgeu, CS 93837, F-29238 Brest (France)
Department of Earth and Ocean Sciences, University of South Carolina, Columbia (U.S.A.)
1 rue des tilleuls, F-59152 Gruson (France)
Published online in final form (pdf) on June 2, 2022
DOI 10.2110/carnets.2022.2208
![]()
[Editor: Michel
Moullade; technical editor: Bruno R.C. Granier]
![]()
The subsurface upper Spraberry Formation in NW Texas is dominantly composed of calcareous turbidites and debris-flows. The petrographic analysis of its porous grain-supported fabrics revealed the presence of controversial superimposed compaction fabrics, sometimes referred to as "distorted ooids". Based on the paragenetic sequence, it is demonstrated that these allochems were not distorted. In fact, the related zigzag and silcrow (§) patterns result from the collapse of molds, either oomolds or biomolds, in response to mechanical compaction of the fragile framework made up of isopachous Low-Mg Calcite cement surrounding these empty molds. As a side finding, this stratigraphic unit, which was said to be late Kungurian in age, is proved to be nearly 7 millions of year older.
NW Texas;
Permian;
ooids;
oomolds;
compaction
Granier B.R.C., Kendall Ch.G.St.C. & Vachard D. (2022).- The Kalkowsky Project - Chapter IV. Case study of the Happy Spraberry oil reservoirs in NW Texas (with a micropaleontologic and biostratigraphic supplement): Collapsed molds should not be treated as a category of distorted ooids.- Carnets Geol., Madrid, vol. 22, no. 8, p. 311-332.
Le projet Kalkowsky - Chapitre IV. Étude de cas des réservoirs pétroliers de Happy Spraberry dans le nord-ouest du Texas (avec un supplément micropaléontologique et biostratigraphique) : Les cavités de dissolution effondrées ne doivent pas être traitées comme une catégorie d'ooïdes déformés.- La Formation Spraberry supérieur reconnue dans des gisements pétroliers du nord-ouest du Texas correspond pour l'essentiel à des turbidites calcaires et coulées de débris. L'analyse pétrographique de ses faciès poreux à grains autoportés a révélé la présence de structures superposées de compaction, structures controversées parfois appelées "ooïdes déformés". Sur la base de la séquence paragénétique, il est démontré que ces allochems n'ont pas été significativement déformés. En fait, les motifs associés en zigzag et ceux en § (deux S combinés, symbole typographique de paragraphe) résultent de l'effondrement de moules de dissolution, soit d'ooïdes, soit de bioclastes, en réponse à la compaction mécanique de la structure fragile constituée par le ciment isopaque en calcite peu magnésienne délimitant ces cavités. Par ailleurs, cette unité stratigraphique, à laquelle on attribuait un âge Kungurien supérieur, s'avère être près de 7 millions dannées plus ancienne.
nord-ouest du Texas ;
Permien ;
ooïdes ;
moules de dissolution d'ooïdes ;
compaction
This is the fourth contribution to the Kalkowsky Project (for details see earlier contributions: Granier & Lapointe, 2021, 2022a, 2022b) and a reminder of the importance of careful petrography. During the preparatory work for a manuscript discussing a paper recently published in the journal Sedimentology[*], the first author (BRCG) received a temporary loan of a set of 40 thin sections from 7 wells of the Happy Spraberry oil field in the Garza County (NW Texas, USA). The cored intervals comprise both siliciclastic and calcareous turbidites, as well as debris-flows, from the Leonardian upper Spraberry Formation (e.g., Handford, 1981), late Kungurian in age according to Ahr and Hammel (1999, and followers: Clayton & Kerans, 2013) but probably older, i.e., latest Sakmarian to early Artinskian as established by one of us (D.V.): See § Appendix: Micropaleontologic and biostratigraphic supplement. Where the rock has retained a certain amount of both residual primary intergranular and secondary moldic porosity, the best flow units correspond to calcareous grainy turbidites with up to 33% porosity and variable permeability reaching 124 mD. The petrographic study of the thin sections revealed that many molds are not forming separate vug pores sensu Lucia (1983), but touching vug pores sensu Lucia (1983), contributing to improving their connectivity, hence their permeability. The corresponding fabric is quite unique: Should these pores have been filled with LMC (Low-Mg Calcite) cement they would have been identified either as "pseudooliths" or "verwitterte Kalksteinkörner" [corroded limestone allochems] (Bornemann,1886, Pl. VII, fig. 1), "zerbrochene und auseinander gezogene Oolithkörner" [broken and pulled apart ooids] (Frantzen, 1888, Pl. III, figs. 2-3), "oolithes déformées" [distorted ooids] (Cayeux, 1935, Pl. XV, fig. 55), "distorted oolites and pseudoolites" (Carozzi, 1961, Fig. 6), "deformed recrystallized pisoliths" in chains or in clusters (Kettenbrink & Manger, 1971, Fig. 3.B, 3.D-F), "distorted ooids" (Radwański & Birkenmajer, 1977, Fig. 16.d-e; Richter, 1983, Fig. 3.H; Cantrell & Walker, 1985, Fig. 9.C-D), "distorted pisomolds" (Conley, 1977, Fig. 5.A-D), "spastolithic (deformed) and elephantine (trunk-to-tail) connected ooids" (Wilkinson et al., 1984, Fig. 6.F; Chatalov, 2003, Fig. 1.a-b), or "spastoliths" (Scholle & Ulmer-Scholle, 2003, p. 240 middle-upper, p. 354 middle). Contrary to our predecessors who were dealing with surface samples, hence fully cemented, tight limestones, we have subsurface material sampled from cores from a depth of about 1500 m TVD (true vertical depth). Because these rocks lack ultimate stages of cementation reported from most surface sampling, the interpretation of the paragenetic sequence and the related resulting fabrics is significantly easier.
The first author (BRCG) used a Wild Heerbrugg M5A binocular microscope to study the 40 thin sections. The latter were probably made from horizontal or vertical plugs taken from cores of 7 wells (Table 1) in the Happy Spraberry Field, located some 90 km SE of Lubbock, NW Texas. The core plug chips were impregnated with blue epoxy prior to the final cut, which helps visualizing the porous network. On each thin section a notch indicates the top. One third to half of each thin section surface is stained with Alizarin Red-S, which however proved to be useless considering the limited amount of dolomite in the rock samples. Photomicrographs of the material (i.e., microfacies and microfossils) were taken in both transmitted and reflected lights with a Nikon D3100 camera mounted on the microscope.
Table 1 : List and location of the Happy Spraberry wells (Garza County, NW Texas, USA) operated by Citation Oil & Gas Corp. with studied samples:
| Well | GSP | completion year |
| Lott 19#2 | 33°00'27.2"N 101°21'25.0"W | 1989 |
| Lott 19#3 | 33°00'12.1"N 101°21'25.5"W | 2012 |
| Lott 19#4 | 33°00'12.3"N 101°21'41.1"W | 1990 |
| Lott 19#5 | 32°59'59.2"N 101°21'41.3"W | 2002 |
| Lott 19#6 | 32°59'59.1"N 101°21'27.0"W | 2016 |
| Lott 19#10 | 32°59'52.4"N 101°21'54.5"W | 1992 |
| Lott 19#11 | 33°00'18.7"N 101°21'33.2"W | 1992 |
Five out of forty thin sections studied, of which two (i.e., Lott 19#3 4911' and Lott 19#4 4930') also include areas with the grainy turbidite facies, correspond to the debris-flow facies, i.e., to lithoclastic floatstones with grainstone matrices. The extraclasts are mostly either silty mudstones or bryozoan-Tubyphites boundstones.
Thirty-seven thin sections display a grainstone facies. The allochems are mostly bioclasts and ooids. A significant number of the allochems were leached and remain as molds, whereas some others were cemented and a few recrystallized sensu lato (recrystallized sensu stricto to form a mosaic of subequant LMC crystals or partly leached and cemented by drusy LMC cement). Common bioclasts are Bivalvia shells and crinoid ossicles, each category forming up to 10% of the grains. Bivalvia shells, which originally had an aragonite mineralogy, are commonly found as empty molds and micritic envelopes, and sometimes as partly or fully cemented molds and micritic envelopes; similar observations are made for the previously aragonite layers of those Bivalvia shells that once had a dual calcite and aragonite composition. It is noteworthy to mention that syntaxial overgrowths on HMC (High-Mg Calcite) crinoidal remains have surprisingly a rather limited extension. Two rare occurrences of ammonoid shells (? goniatites) are reported here from Lott 19#4 4958' and 4961.7'. Other bioclasts include echinoid spines, gastropod shells, brachiopod remains (including some spines), ostracods, and foraminifers. Among the foraminifers the tests of the primitive Miliolata that were made of HMC are commonly leached (see § Appendix: Micropaleontologic and biostratigraphic supplement) whereas the larger pseudomonocrystalline echinodermal remains were not. Contrary to the primary nature of the angular molds (mostly the remains of former aragonite molluscan shells) there is always some ambiguity regarding the origins of the many rounded molds. The latter should not necessarily be described as oomolds because, although some retained a nucleus or few cortical layers, others were definitely both bioclasts and probable peloids. Some ooids were not leached but partly or fully recrystallized sensu lato. There is no evidence either that these ooids were ever micritized because micritization would have erased the concentric structures that remain visible in most recrystallized ooids. Besides the secondary moldic porosity after ooids or bioclasts the porous network comprises a significant volume of intergranular porosity, which is merely a residual primary porosity. Intergranular porosity is commonly found in the majority of the thin sections studied; it is lacking only when the centripetal drusy LMC cementation occluded these pores or when late dolomite or celestine cements occurred. Stylolites are rare, except where the debris-flow facies are tightly cemented or have contrasting lithologies. However, solution seams are common in the grainstone facies. They are often observed in collapsed moldic pores, next to residual intergranular pores.
Any study
on cement stratigraphy implies that cementation and leaching cannot be coeval
because both processes require fluids with different discrete pH. For example,
two commonly observed sequences of events involving originally aragonite ooids
(Fig. 1.A ![]() ) and bioclasts
(Fig. 1.B
) and bioclasts
(Fig. 1.B ![]() ) are presented in
Figure 1
) are presented in
Figure 1 ![]() , redrawn after a
sketch by the second author (Kendall, 2005).
, redrawn after a
sketch by the second author (Kendall, 2005).
|
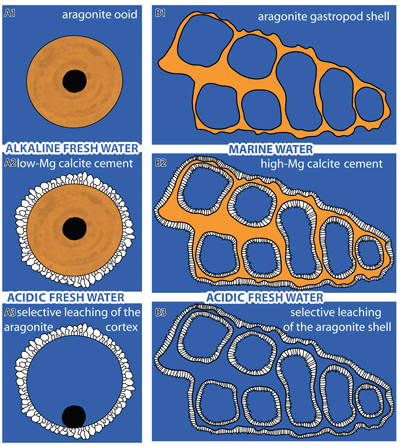
Figure 1:
A1 to A3) Aragonite
ooid cemented by Low-Mg calcite (LMC) in alkaline fresh water, followed by
leaching of the cortical aragonite in acidic fresh water. B1 to B3) Aragonite
gastropod shell cemented by High-Mg calcite (HMC) in marine water, followed by
selective leaching of the gastropod shell aragonite in acidic fresh water,
possibly due to the presence of acid from organic proteins. |
With regard to the grainstone reservoir facies of the Plattsburg Limestone (uppermost Carboniferous) of Kansas, it is possible to reconstruct the paragenetic sequence on the basis of the above petrological observations.
First, peripheral micritization probably affected some bioclasts in their original shallow-water setting, hence predating their resedimentation in turbidites. Later, provided that the formerly aragonite or LMC bioclasts did not suffer any recrystallization sensu stricto, their leaching would form molds that correspond to a secondary, moldic porosity. Oolitic cortices are also eligible to recrystallization and/or leaching. Although it is commonly stated that leached ooids or recrystallized ooids were primarily aragonite ooids (e.g., Buczynski & Wilkinson, 1982; Tucker, 1984), it cannot be excluded that they could have been primarily HMC ooids (e.g., Granier & Lapointe, 2022b). The nature of the organic framework of these ooid cortices (as well as of some bioclasts), its amount, its degree of oxidation and the timing of the later are probably the keys to understand why some were selectively leached whereas others were recrystallized (e.g., Skipwith & Shearman, 1965).
Second, the resedimentation of the various calcareous allochems in grainy turbidites led to a good grain sorting within the graded beds. There is no obvious evidence for mechanical compaction prior to cementation in the primary intergranular and intragranular pores of this calcareous sands.
Third,
the intragranular (organic, e.g., foraminifer chambers) pores, especially
the smallest, were fully filled with a LMC cement while intergranular pores, as
well as the larger intragranular pores, were incompletely filled with an
isopachous cement made of small (i.e., ~20 µm in width) more or less
equidimensional (i.e., "equant", or "granular" sensu
Flügel, 2004) LMC crystals, and thus leaving some residual primary
intergranular porosity (Pls. 1 ![]() - 2
- 2 ![]() - 3
- 3 ![]() ,
figs. c, f, i, l, o, r, u). Considering
the crystal habit, hence their LMC nature (Folk, 1974), such cementation
could have taken place either in the meteoric phreatic zone or in the subsurface
burial zone; the former setting should be excluded when considering both the
depositional environment (turbidites) and the regional burial history. It is
noteworthy that the common but volumetrically minor syntaxial
overgrowths on crinoidal ossicles also contributed in reducing the intergranular
porosity. Finally, this LMC cementation stopped quite early in the paragenetic
sequence of most grainstone samples studied whereas the few tight grainstone
samples (e.g., Lott 19#3 4915'; Lott19#4
4919.3', 4926.9', 4937.5'; Lott 19#6 4948') were almost fully filled with a
drusy LMC cement.
,
figs. c, f, i, l, o, r, u). Considering
the crystal habit, hence their LMC nature (Folk, 1974), such cementation
could have taken place either in the meteoric phreatic zone or in the subsurface
burial zone; the former setting should be excluded when considering both the
depositional environment (turbidites) and the regional burial history. It is
noteworthy that the common but volumetrically minor syntaxial
overgrowths on crinoidal ossicles also contributed in reducing the intergranular
porosity. Finally, this LMC cementation stopped quite early in the paragenetic
sequence of most grainstone samples studied whereas the few tight grainstone
samples (e.g., Lott 19#3 4915'; Lott19#4
4919.3', 4926.9', 4937.5'; Lott 19#6 4948') were almost fully filled with a
drusy LMC cement.
Fourth, the drusy LMC cement in the tight grainstone samples results from the competitive overgrowth of the originally equant LMC crystals in the former intergranular pores. Besides molds and micritic envelopes that are also affected by LMC drusy cementation, these samples are also characterized by the occurrence of recrystallized oolitic cortices (e.g., Lott 19#3 4915'; Lott 19#4 4911.5'). It is assumed here that recrystallization probably occurred before leaching, preventing these oolitic cortices being leached. However a lack of oxidation of the organic framework of the oolitic cortices at the time of leaching would also have limited the dissolution efficiency. Therefore the hypothesis that the recrystallisation could have occurred after the leaching cannot be definitely ruled out.
Fifth, in the few tight grainstone samples studied, LMC cement occurs in some molds (e.g., bioclastic molds). However, in most grainstone samples, dissolution molds (both bioclastic and oolitic) are empty indicating that cementation predated their leaching (e.g., Shearman, unpublished; Conley, 1977; Scholle & Ulmer-Scholle, 2003; Flügel, 2004; Kendall, 2005).
Sixth, as seen
above, there was no obvious evidence for mechanical
compaction in the early stages. However, the leaching of aragonite and HMC of
oolitic cortices and bioclasts to form a secondary moldic porosity, combined
with the weak isopachous "granular" cementation, hence the presence of
a residual primary intergranular porosity, all these features favoured the
collapse of some of the largest pores. This late mechanical compaction implies a
stratiform arrangement similar to that of the stratiform stylolites (Pls. 1 ![]() - 2
- 2 ![]() - 3
- 3 ![]() ,
figs. a, d, g, j, m, p, s). However, as reported by Conley
(1977),
"linear series of elegant
curves" that form "long zigzag chains" are not only arranged
perpendicular to compaction stress (i.e., stratiform), but also at an
angle (Pls. 1
,
figs. a, d, g, j, m, p, s). However, as reported by Conley
(1977),
"linear series of elegant
curves" that form "long zigzag chains" are not only arranged
perpendicular to compaction stress (i.e., stratiform), but also at an
angle (Pls. 1 ![]() - 2
- 2 ![]() - 3
- 3 ![]() ,
figs. b, e, h, k, n, q, t) delimitating breccia
fragments. The brecciation was probably favored by differences in both size and
shape existing between rounded moldic pores (mostly after
ooids) and/or angular moldic pores (mostly after bioclasts).
,
figs. b, e, h, k, n, q, t) delimitating breccia
fragments. The brecciation was probably favored by differences in both size and
shape existing between rounded moldic pores (mostly after
ooids) and/or angular moldic pores (mostly after bioclasts).
Seventh, chemical compaction in the form of non-sutured pressure solution features (e.g., Lott 19#2 4934'; Lott 19#4 4911.5', 4918') developed at a later stage when mechanical compaction could not accomodate the exceeding compaction stress.
Eighth, flushing of the brine out the reservoir rock with its replacement by oil brought an end to the paragenetic sequence.
This paragenetic sequence could be enhanced considering that 1) dissolution of the aragonite of the bioclasts [or the aragonite oolitic cortices] was not coeval to that of the HMC of the bioclasts [or the HMC oolitic cortices], 2) timings of recrystallisation of aragonite bioclasts and of aragonite or HMC oolitic cortices could be better constrained, 3) dissolution of the HMC of the porcellaneous foraminifer tests, 4) syntaxial overgrowths on echinodermal remains were not analysed, etc. However, on one hand, it is outside of the scope of this paper and, on the other hand, we were short in resource (no access to cathodoluminescence microscopy) and time to push forward our investigation.
In the opinions of Bornemann (1886), Frantzen (1888), Carozzi (1961), or Kettenbrink and Manger (1971), the plastic deformation of their ooids was caused by compaction and preceded both lithification and cementation whereas according to 1) Radwański and Birkenmajer (1977), the distortion of the ooids is related to "boudinage"; 2) Wilkinson et al. (1984), it was caused by compaction following cementation and "during and/or following aragonite dissolution"; 3) Cantrell and Walker (1985), it is due to pressure-solution; and 4) Chatalov (2003), compaction initiated "early deformation" that "may have continued during the late diagenesis, e.g. through pressure-solution". However, in our case study, none of the above assumptions is valid because 'what were distorted' were indeed the molds, not the allochems (i.e., neither bioclasts, nor ooids).
Although Conley
(1977) quoted 48 times "distorted" grains for only 3 times "distorted
pisomolds" (plus 2 times "undistorted pisomolds"), he was the
first to correctly point out that he was not dealing with distorted ooids but
distorted oomolds (Fig. 2 ![]() ). He pointed out that "Solution of some pisoliths
followed precipitation of first generation cement and preceded compaction"
(op.cit., p. 561). Furthermore, he noted that the "cement framework"
between the "pisomolds" could be "destroyed in varying degrees by
compaction" and that it resulted in variety of unique fabrics. The first
steps of his paragenetic sequence are rather similar to those of the paragenetic
sequence we describe above for the upper Spraberry oolitic grainstone reservoir
facies.
). He pointed out that "Solution of some pisoliths
followed precipitation of first generation cement and preceded compaction"
(op.cit., p. 561). Furthermore, he noted that the "cement framework"
between the "pisomolds" could be "destroyed in varying degrees by
compaction" and that it resulted in variety of unique fabrics. The first
steps of his paragenetic sequence are rather similar to those of the paragenetic
sequence we describe above for the upper Spraberry oolitic grainstone reservoir
facies.
Figure 2 ![]() ,
redrawn and adapted from Conley's (1977) Figure 8, illustrates the
striking similarity between both paragenetic sequences. Our interpretation of
the paragenetic sequence and that of the related structures are significantly
easier because our subsurface samples lack the ultimate stages of cementation
reported by Conley. Figure 3
,
redrawn and adapted from Conley's (1977) Figure 8, illustrates the
striking similarity between both paragenetic sequences. Our interpretation of
the paragenetic sequence and that of the related structures are significantly
easier because our subsurface samples lack the ultimate stages of cementation
reported by Conley. Figure 3 ![]() is expanding Conley's model to
include models for both the zig-zag pattern (Fig. 3
is expanding Conley's model to
include models for both the zig-zag pattern (Fig. 3 ![]() , left column) sensu Carozzi
(1961) and the typographic symbol of silcrow (§) (Fig. 3
, left column) sensu Carozzi
(1961) and the typographic symbol of silcrow (§) (Fig. 3 ![]() , right column) sensu
Carozzi (1961).
, right column) sensu
Carozzi (1961).
The only
minor flaw in Conley's interpretation is that he refers to "blocky"
intergranular cement, which implies the absence of residual primary porosity,
whereas we refer to isopachous LMC cement that did not fully filled the
intergranular pores. According to Conley (1977, p. 561), the "cement
framework failed at points of least strength and partially collapsed under
compactional forces". However, he was reasoning in 2 dimensions only (Conley,
1977, Figs. 8-9), not in 3 dimensions. Besides the grain nature, their external
morphology, their sizes and their arrangement are variable, compaction with
"blocky" (sensu Conley) intergranular cement would have
only produced stratiform seams and stylolites, never oblique chains of oomolds (Fig.
2 ![]() ), nor the silcrow fabric
(Fig. 3
), nor the silcrow fabric
(Fig. 3 ![]() , right column).
, right column).
In the
studied material, most collapsed molds are empty whereas others contain some
material that is probably the remains of partly dissolved oolitic cortices (Pl. 1 ![]() , figs. a, d, g-h, j-k, m-o;
Pl. 2
, figs. a, d, g-h, j-k, m-o;
Pl. 2 ![]() ,
figs. a-b, d-f, j-k, s-u) and oolitic
nuclei, if any. In the latter case, similarity with the tight oolitic limestones
documented by our predecessors (Bornemann, 1886, Pl. VII, fig. 1; Frantzen,
1888, Pl. III, figs. 2-3; Cayeux,
1935, Pl. XV, fig. 55; Carozzi, 1961, Fig. 6; Kettenbrink & Manger,
1971, Fig. 3.B, 3.D-F; Conley,
1977, Fig. 5.A-D; Radwański & Birkenmajer,
1977, Fig.
16.d-e; Richter, 1983, Fig. 3.H; Wilkinson et al.,
1984,
Fig. 6.F; Cantrell & Walker,
1985, Fig. 9.C-D; Chatalov, 2003, Fig. 1.a-b; Scholle & Ulmer-Scholle,
2003, p. 240
middle-upper, p. 354 middle) are striking. The only difference is that our
subsurface material (it is a producing oil reservoir) lacks final stages of LMC
cements found in most specimens picked in outcrops.
,
figs. a-b, d-f, j-k, s-u) and oolitic
nuclei, if any. In the latter case, similarity with the tight oolitic limestones
documented by our predecessors (Bornemann, 1886, Pl. VII, fig. 1; Frantzen,
1888, Pl. III, figs. 2-3; Cayeux,
1935, Pl. XV, fig. 55; Carozzi, 1961, Fig. 6; Kettenbrink & Manger,
1971, Fig. 3.B, 3.D-F; Conley,
1977, Fig. 5.A-D; Radwański & Birkenmajer,
1977, Fig.
16.d-e; Richter, 1983, Fig. 3.H; Wilkinson et al.,
1984,
Fig. 6.F; Cantrell & Walker,
1985, Fig. 9.C-D; Chatalov, 2003, Fig. 1.a-b; Scholle & Ulmer-Scholle,
2003, p. 240
middle-upper, p. 354 middle) are striking. The only difference is that our
subsurface material (it is a producing oil reservoir) lacks final stages of LMC
cements found in most specimens picked in outcrops.
|
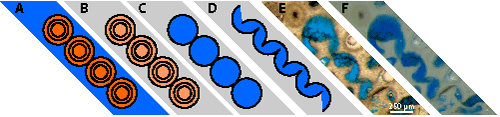
Figure 2:
Schematic paragenetic sequence from aragonite or
HMC ooids to a chain of distorted oomolds (redrawn and adapted from Conley,
1977, Fig. 8): A) undistorted ooids; B) cemented slightly oxided ooids; C)
leached ooids, i.e., oomolds; D) collapsed oomolds; E) transmitted light;
F) reflected light. E-F: Lott 19#4 4918' (Scale bar = 250 µm). Orange: oolitic
cortices; pale orange: oolitic cortices after oxidation; grey: "blocky"
cement; blue: pores. |
|
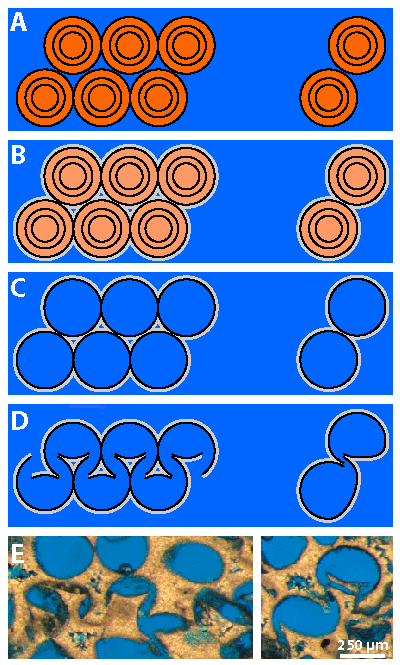
Figure 3: Schematic paragenetic sequence from aragonite or HMC ooids to
a chain of distorted oomolds: A)
undistorted ooids; B) weakly cemented slightly oxided ooids; C) leached ooids, i.e.,
oomolds; D) collapsed oomolds forming a zig-zag pattern to the left and the
typographic symbol of silcrow to the right; E) to the left, photomicrograph of a
zig-zag pattern sensu Carozzi (1961), Lott 19#5 4958'; to the
right, photomicrograph of the
typographic symbol of silcrow (§) pattern sensu Carozzi
(1961),
Lott 19#2 4934' (Scale bar = 250 µm). Orange: oolitic cortices; pale orange:
oolitic cortices after oxidation; grey: isopachous cement; blue: pores. |
The uppermost Sakmarian to lowermost Artinskian calcareous turbidites and associated debris-flows of the upper Spraberry Formation in NW Texas comprise some oolitic grainstones that display peculiar compactional fabrics known in the past literature by many names including so-called "distorted ooids". A simplified paragenetic sequence drawn from the petrographic analysis starts with an isopachous LMC cementation filling in part intergranular pores. It is followed by oxidation of organic matter framework of many allochems, then the leaching of aragonite and HMC allochems (bioclasts, oolitic cortices), with the notable exception of the HMC echinodermal remains. It ends with compaction that led to the collapse of some moldic pores and the local limited development of solution seams. The ooids were not distorted but their molds did collapse. This interpretation better matches Conley's (1977), who presented a paragenetic model to explain the occurrence of distorted molds (op.cit., Figs. 8-9). The only minor flaw in his model was to consider that the grain-supported fabric was fully cemented before leaching and collapsing. Our study of the Texan Permian material demonstrates that collapse would not have been possible without the presence of some residual intergranular porosity, hence the presence of a limited LMC cementation of these primary pores. The converse of this conclusion is also true: The occurrence of collapsed molds would imply the presence of a residual intergranular porosity at the time of collapse.
The first author (BRCG) would like to thank Citation Oil & Gas Corp., Houston - TX, for having granted him permission to use their material in this publication. He is very grateful to Jay Portwood, Vice President - Reservoir, and Jorge L. Romero, Senior Reservoir Engineer, who acted as facilitator. He also wishes to thank the Association "Carnets de Géologie" and donators, namely D.M. Basso, J. Charollais, C. Collom, T. Grgasović, R. Manni, D.M. Martill, R. Martini, M.V. Pardo Alonso, F. Sciuto, M. Septfontaine, and R.W. Scott, for financial support. The authors acknowledge the help of Matt Gentry, "Robert" Handford, Felicitas Kaiser, Charles Kerans, John Layman II, "Bob" Loucks, Vincent Mazingue-Desailly, and Robert W. Scott when tracking material from the Spraberry Formation of the Happy Field (Garza County, Texas). By proxy of J.C. Laya P.K. Pufahl has arranged the loan of material that was previously studied by the late Professor Wayne M. Ahr (1938-2011) and a group of MSc students (Matt Gentry, John Layman II, Vincent Mazingue-Desailly, ), and that is currently stored at the Texas A&M University, College Station - TX. Finally, thanks are due to Jan Golonka, Brian Pratt, and Andreas "André" Strasser for their careful review and useful suggestions (even if we did not follow up on all of these). The authors are solely responsible for the interpretations (and opinions) expressed here.
Ahr W.M. & Hammel B.S. (1999).- Identification and mapping of flow units in carbonate reservoirs: An example from the Happy Spraberry (Permian) Field Garza County, Texas USA.- Energy Exploration & Exploitation, London, vol. 17, nos. 3-4, p. 311-334.
Baryshnikov V.V., Zolotova V.P. & Koshaleva V.F. (1982).- Novye vidy foraminifer Artinskogo yarusa Permskogo Priuralya [New species of foraminifers from Artinskian stage of the Pre-Ural].- Akademiya Nauk SSSR, Uralskii Nauchnyi Tsentr, Institut Geologii i Geokhimii, preprint, 54 p. [in Russian]
Bensh F.P. (1987).- Reviziya sistematiki psevdofuzulinid roda Pseudofusulina Dunbar et Skinner i blizkikh rodov (Revision of the systematics of the genus Pseudofusulina Dunbar et Skinner and related genera).- Voprosy Mikropaleontologii, Moscow, vol. 29, p. 20-53 [in Russian].
Bornemann J.G. (1886).- Beiträge zur Kentniss des Muschelkalks, insbesondere der Schlehtenfolge und der Gesteine des Unteren Muschelkalkes in Thüringen.- Jahrbuch der Königlich Preussischen Geologischen Landesanstalt und Bergakademie zu Berlin, Bd. 6, Jahr 1885, p. 268-321 (Pls. VII-XIV).
Buczynski C. & Wilkinson B.H. (1982).- Two-phase (aragonite-calcite) ooids from the Pennsylvanian Plattsburg limestone of southeastern Kansas.- Abstracts of Papers, Eleventh International Congress on Sedimentology, Hamilton, Ontario, p. 115.
Cantrell D.L. & Walker K.R. (1985).- Depositional and diagenetic patterns, ancient oolite, Middle Ordovician, eastern Tennessee.- Journal of Sedimentary Petrology, Tulsa - OK, vol. 55, no. 4, p. 518-531.
Carozzi A.V. (1961).- Distorted oolites and pseudoolites.- Journal of Sedimentary Petrology, Tulsa - OK, vol. 31, no. 2, p. 262-274.
Cayeux L. (1935).- Les roches sédimentaires de France. Roches carbonatées.- Masson & Cie, Paris, 447 p.
Chatalov A. (2003).- On the origin of distorted ooids in the Triassic limestones from northwestern Bulgaria.- Comptes rendus de l'Académie bulgare des Sciences (Sciences mathématiques et naturelles), Sofia, t. 56, no. 10, p. 63-68.
Clayton J.L. & Kerans C. (2013).- Reservoir compartmentalization of a deep-water ooid fan, Happy Field, Permian Basin. In: Verwer K., Playton T.E. & Harris P.M. (eds.), Deposits, architecture, and controls of carbonate margin, slope and basinal settings.- SEPM Special Publication, Tulsa - OK, vol. 105, p. 359-383.
Conley C.D. (1977).- Origin of distorted ooliths and pisoliths.- Journal of Sedimentary Petrology, Tulsa - OK, vol. 47, no. 2, p. 554-564.
Cushman J.A. & Waters J.A. (1928).- Some Foraminifera from the Pennsylvanian and Permian of Texas.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon - MA, vol. 4, p. 31-55.
Davydov V.I., Krainer K. & Chernykh V. (2013).- Fusulinid biostratigraphy of the Lower Permian Zweikofel Formation (Rattendorf Group; Carnic Alps, Austria) and Lower Permian Tethyan chronostratigraphy.- Geological Journal, vol. 48, p. 57-100.
Folk R.L. (1974).- The natural history of crystalline calcium carbonate: Effect of magnesium content and salinity.- Journal of Sedimentary Petrology, Tulsa - OK, vol. 44, no. 1, p. 40-53.
Flügel E. (2004).- Microfacies of carbonate rocks. Analysis, interpretation and application.- Springer Verlag, Berlin, Heidelberg, New York, xx + 976 p.
Frantzen W. (1888).- Untersuchungen über die Gliederung des unteren Muschelkalks in einem Theile von Thüringen und Hessen und über die Natur des Oolithkörner in diesen Gebirgsschichten.- Jahrbuch der Königlich Preussischen Geologischen Landesanstalt und Bergakademie zu Berlin, Bd. 8, Jahr 1887, p. 1-321 (Pls. I-III).
Granier B.R.C. & Lapointe Ph. (2021).- The Kalkowsky Project - Chapter I. Ooid - stromatoid relationship in a stromatolite from the Maiz Gordo Fm (Argentina).- Carnets Geol., Madrid, vol. 21, no. 9, p. 193-201. DOI: 10.2110/carnets.2021.2109
Granier B.R.C. & Lapointe Ph. (2022a).- The Kalkowsky Project - Chapter II. Wobbly ooids in a stromatolite from the Yacoraite Formation (Argentina).- Carnets Geol., Madrid, vol. 22, no. 3, p. 111-117. DOI: 10.2110/carnets.2022.2203
Granier B.R.C. & Lapointe Ph. (2022b).- The Kalkowsky Project - Chapter III. Significance of primary radial fabrics associated with ancient partly leached or recrystallized calcareous ooids.- Carnets Geol., Madrid, vol. 22, no. 5, p. 149-160. DOI: 10.2110/carnets.2022.2205
Handford C.R. (1981).- Sedimentology and genetic stratigraphy of Dean and Spraberry formations (Permian), Midland Basin, Texas.- AAPG Bulletin, Tulsa - OK, vol. 65, p. 1602-1616.
Henderson C.M., Davydov V.I. & Wardlaw B.R. (2012).- The Permian Period (Chapter 24). In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 653-679.
Kendall C.G.St.C. (2005).- Lecture 6: Cementation & diagenesis.- Geol 751: Carbonate petrography, University of South Carolina, Columbia - SC, 118 slides. URL: http://www.sepmstrata.org/CMS_Files/CarbonateDiagenesis.ppt
Kettenbrink E.C. & Manger W.L. (1971).- A deformed marine pisolite from the Plattsburg Limestone (Upper Pennsylvanian) of southeastern Kansas.- Journal of Sedimentary Petrology, Tulsa - OK, vol. 41, no. 2, p. 435-443.
Krainer K., Vachard D. & Schaffhauser M. (2019).- Early Permian (Yakhtashian; Artinskian-early Kungurian) foraminifers and microproblematica from the Carnic Alps (Austria/Italy).- Abhandlungen der Geologischen Bundesanstalt, Wien, vol. 73, 247 p.
Lipina O.A. (1949).- Melkie foraminifery pogrebennykh massivov Bashkirii [Smaller foraminifers from the buried massifs of Bashkiria].- Trudy Instituta Geologicheskikh Nauk (Geol. Ser.), Akademiya Nauk SSSR, Moscow, vol. 105, no. 35, p. 198-235 [in Russian; French translation BRGM, no. 791].
Lucas S.G., Krainer K. & Vachard D. (2015).- The Lower Permian Hueco Group, Robledo Mountains, New Mexico (U.S.A.). In: Lucas S.G. & DiMichele W.A. (eds.), Carboniferous-Permian Transition in the Robledo Mountains, Southern New Mexico.- New Mexico Museum of Natural History and Science Bulletin, Albuquerque - NM, vol. 65, p. 43-95.
Lucia F.J. (1983).- Petrophysical parameters estimated from visual description of carbonate rocks: A field classification of carbonate pore space.- Journal of Petroleum Technology, Richardson - TX, vol. 35, no. 3, p. 626-637.
Maslov V.P. (1956).- Iskopaemye izvestkovye vodorosli SSSR (Calcareous algae from USSR).- Trudy Instituta Geologicheskikh Nauk (Geol. Ser.), Akademiya Nauk SSSR, Moscow, vol. 160, 297 p. [in Russian; French translation BRGM, no. 351, 382 p.].
Miklukho-Maklay K.V. (1954).- Permian foraminifers from North Caucasus.- Trudy VSEGEI, Gosgeoltekhizdat, Moscow, vol. 1, 163 p. [in Russian; French translation BRGM no. 2683, 216 p.].
Radwański A. & Birkenmajer K. (1977).- Oolitic/pisolitic dolostones from the Late Precambrian of south Spitsbergen: Their sedimentary environment and diagenesis.- Acta Geologica Polonica, Warsaw, vol. 27, no. 1, p. 1-40.
Rauzer-Chernousova D.M. (1949).- Некоторые псевдофузулины и парафузулины Башкирского Приуралья (Some Pseudofusulina and Parafusulina from the Bashkirian Pre-Urals).- Trudy Instituta Geologicheskikh Nauk (Geol. Ser.), Akademiya Nauk SSSR, Moscow, vol. 105, no. 35, p. 118-162.
Richter D.K. (1983).- Calcareous ooids: A synopsis. In: Peryt T.M. (ed.), Coated grains.- Springer, Berlin, Heidelberg, p. 71-99.
Ross C.A. & Ross J.R.P. (1960).- Fusulinids from the Hess Member of the Leonard Formation, Leonard Series (Permian), Glass Mountains, Texas.- Contributions from the Cushman Foundation for Foraminiferal Research, Sharon - MA, vol. 11, no. 4, p. 116-133. URL: https://cedar.wwu.edu/geology_facpubs/57/
Scholle P.A. & Ulmer-Scholle D.S. (2003).- A color guide to the petrography of carbonate rocks: Grains, textures, porosity, diagenesis.- AAPG Memoir, Tulsa - OK, vol. 77, 474 p.
Shearman D.J. (circa 1977, unpublished).- Laboratory handbook of carbonate petrology.- Imperial College, London, 55 p.
Skipwith P.A.d'E. & Shearman D.J. (1965).- Organic matter in Recent and Ancient limestones and its role in their diagenesis.- Nature, London, vol. 208, p. 1310-1311.
Tucker M.E. (1984).- Calcitic, aragonitic and mixed calcitic-aragonitic ooids from the mid-Proterozoic Belt Supergroup, Montana.- Sedimentology, vol. 31, no. 5, p. 627-644.
Vachard D., Krainer K. & Lucas S.G. (2015).- Late Early Permian (late Leonardian; Kungurian) algae, microproblematica, and smaller foraminifers from the Yeso Group and San Andres Formation (New Mexico; USA).- Palaeontologia Electronica 18.1.21A, 77 p. URL: https://doi.org/10.26879/433
Wilkinson B.H., Buczynski C. & Owen R.M. (1984).- Chemical control of carbonate phases: Implications from Upper Pennsylvanian calcite-aragonite ooids of southeastern Kansas.- Journal of Sedimentary Research, Tulsa - OK, vol. 54, no. 3, p. 932-947.
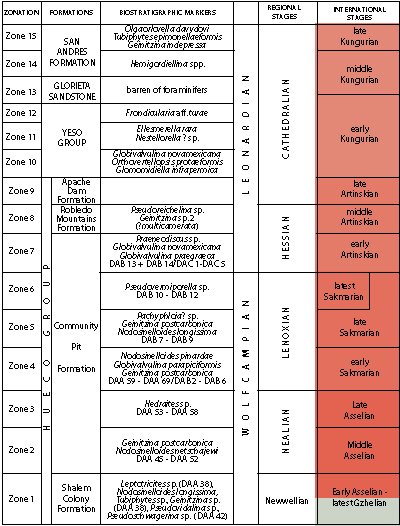
In the Lower Permian sedimentary series recently studied in New Mexico (Lucas
et al., 2015), the investigated levels
correspond to the latest Sakmarian and early Artinskian biozones 6 and 7 (Fig. 4 ![]() ) , on the basis of the occurrence of Pseudovermiporella,
Praeneodiscus, Glomomidiella infrapermica, Grovesella, and Schubertella,
as well as the absence of Pseudoreichelina.
) , on the basis of the occurrence of Pseudovermiporella,
Praeneodiscus, Glomomidiella infrapermica, Grovesella, and Schubertella,
as well as the absence of Pseudoreichelina.
When compared with the Russian sequences, the studied samples seem to also correlate with the lower Artinskian beds of the Pre-Urals of Perm (Baryshnikov et al., 1982), on the basis of the occurrence of Grovesella chomatifera (Zolotova in Baryshnikov et al., 1982) and its nodosariates.
Classically, these units correspond to the Tastubella pedissequa Zone of larger fusulinids of Russia, Praeskinnerella crassitectoria-P. guembeli of the USA, and Sweetognathus "whitei" of the conodonts (Rauzer-Chernousova, 1949; C.A. Ross & J.R.P. Ross, 1960; Bensh, 1987; Henderson et al., 2012).
The observed assemblage comprises algae (Gymnocodiaceae, Gyroporella? sp.), foraminifers with microgranular tests (Abadehellopsis cf. protea, Climacammina sp., Endothyra sp., Globivalvulina spp., Grovesella chomatifera, Grovesella spp., Neoendothyra sp., Palaeotextularia sp., Schubertella sp., Spireitlina sp., Tetrataxis sp.), foraminifers with porcelaneous tests (Ammovertella sp., Cornuspira sp., Glomomidiella infrapermica, Glomodiella spp., Hemigordiellina sp., Palaeonubecularia graiferi, Palaeonubecularia sp., Paraellesmerella sp., Planiinvoluta heathi, Praeneodiscus spp., Pseudovermiporella sp., "Tolypammina" sp., Tubiphytes obscurus), and foraminifers with hyaline tests (Nodosinelloides bella, N. shikhanica, Pachyphloia sp., Praerectoglandulina cf. turae, P. pusilla).
|
Figure
4:
Comparison with regional and international stratigraphic scales in New
Mexico (according to Lucas et al., 2015). |
Gyroporella? sp.
Pl. 4 ![]() ,
fig. 7
,
fig. 7
Description: A tangential section of a Dasycladale apparently aspondyl.
Occurrence: Lott 19#3 4915'.
Spireitlina sp.
Pl. 5 ![]() ,
fig. 42
,
fig. 42
Description: A transverse section of an immature (i.e., only coiled) specimen.
Occurrence: Lott 19#3 4957'.
Endothyra sp.
Pl. 4 ![]() ,
fig. 6
,
fig. 6
Description: Subaxial section.
Occurrence: Lott 19#3 4960'.
Neoendothyra sp.
Pl. 5 ![]() ,
fig. 59
,
fig. 59
Description: An axial section of a small, compressed species with well carinated last whorl.
Occurrence: Lott 19#11 4891'.
Palaeotextularia sp.
Pl. 4 ![]() ,
fig. 7
,
fig. 7
Description: Several axial sections of a typical Palaeotextularia which exhibits 4-8 pairs of chambers.
Occurrence: Lott 19#3 4911'; Lott 19#4 4972.9'.
Climacammina sp.
Pl. 4 ![]() ,
fig. 4
,
fig. 4
Description: Several subaxial sections seem to be more assignable to Climacammina than typical Palaeotextularia.
Occurrence: Lott 19#2 4930'.
Tetrataxis sp.
Pl. 5 ![]() , fig. 60
, fig. 60
Description: Several subaxial sections of a small Tetrataxis.
Occurrence: Lott 19#4 4923.8', 4937.5', 4960'; Lott 19#11 4891', 4905'.
Abadehellopsis
cf. protea
(Cushman & Waters,
1928)
Pl. 5 ![]() ,
fig. 41
,
fig. 41
*1928 Patellina protea Cushman & Waters, p. 54-55, Pl. 7, figs. 8-10.
Description: Abadehellopsis was recently described in the Carnic Alps (Austria). However, one representative was known in Texas for a longer time period under the name of Patellina protea. It was recently refound in New Mexico (Lucas et al., 2015). One oblique section was found in our material.
Occurrence: Lott 19#4 4957.5'.
Globivalvulina spp.
Pl. 4 ![]() ,
figs. 11, 15, 19
,
figs. 11, 15, 19
Description: Several subtransverse and subaxial sections of two or three small species.
Occurrence: Lott 19#2 4926'; Lott 19#3 4925', 4928'; Lott 19#4 4923.8', 4940.8', 4944.9', 4956', 4958', 4960'; Lott 19#5 4950'.
Grovesella
chomatifera
(Zolotova in Baryshnikov et
al., 1982)
Pl. 4 ![]() ,
fig. 9
,
fig. 9
*1982 Schubertella sphaerica chomatifera Zolotova in Baryshnikov et al., p. 21, Pl. 6, figs. 5, 8.
Description: A typical axial section of a small, nautiloid test with three whorls, the two first whorls are deviated at 90°. Proloculus spherical relatively large; chomata low and triangular with a tunnel poorly defined.
Occurrence: Lott 19#3 4915'5.
Grovesella spp.
Pl. 4 ![]() ,
figs. 18, 20, 29; Pl. 5
,
figs. 18, 20, 29; Pl. 5 ![]() , fig. 43
, fig. 43
Description: Several oblique sections.
Occurrence: Lott 19#4 4911.5', 4923.8', 4940.8', 4944.9', 4960'.
Schubertella sp.
Pl. 4 ![]() ,
fig. 13; Pl. 5
,
fig. 13; Pl. 5 ![]() ,
fig. 52
,
fig. 52
Description: Several oblique sections.
Occurrence: Lott 19#3 4928'; Lott 19#5 4958'.
Planiinvoluta
heathi
(Cushman
& Waters, 1928)
Pl. 4 ![]() ,
figs. 2, 5, 26b
,
figs. 2, 5, 26b
Description: Several subaxial sections of this abundant species with one series of an oscillating deuteroloculus, regularly increasing in height and width.
Occurrence: Lott 19#2 4930', 4940'; Lott 19#3 4911', 4928'; Lott 19#4 4933.1', 4944.9', 4960'; Lott 19#5 4942', 4945', 4950'; Lott 19#11 4891'.
Tolypammina sp.
Pl. 4 ![]() ,
figs. 22, 25
,
figs. 22, 25
Description: Several subaxial sections.
Occurrence: Lott 19#2 4926'; Lott 19#3 4915'; Lott 19#4 4926.9', 4933.1', 4950', 4957.5'.
Ammovertella sp.
Pl. 5 ![]() ,
fig. 56
,
fig. 56
Description: Rare subaxial sections.
Occurrence: Lott 19#4 4926.9'; Lott 19#6 4958'.
Orthovertella sp.
Pl. 4 ![]() ,
fig. 32
,
fig. 32
Description: Rare subaxial sections.
Occurrence: Lott 19#3 4909'; 19#4 4944.9'.
Palaeonubecularia
graiferi
(Baryshnikov
in Baryshnikov et al., 1982)
Pl. 4 ![]() ,
fig. 23
,
fig. 23
*1982 Tolypammina graiferi Baryshnikov in Baryshnikov et al., p. 11, Pl. 1, figs. 8, 12-13.
?2019 Pseudovermiporella graiferi (Baryshnikov in Baryshnikov et al., 1982) - Krainer et al., p. 94, Pl. 36, figs. 4-8; Pl. 37, fig. 2 (center)
Description: This species is characterized by concentric rows of chambers and a thick wall. The wall is homogeneous and the perforated wall described by Krainer et al. (2019) belongs probably to another species and perhaps genus.
Occurrence: Lott 19#4 4930'.
Palaeonubecularia sp.
Pl. 4 ![]() ,
fig. 12
,
fig. 12
Description: An elongated test composed of two superposed series of chambers.
Occurrence: Lott 19#3 4925'.
Pseudovermiporella sp.
Pl. 5 ![]() ,
fig. 39b
,
fig. 39b
Description: A tangential section with numerous cylindrical pits. Wall neosparitized
Occurrence: Lott 19#4 4956'.
Paraellesmerella sp.
Pl. 4 ![]() ,
fig. 26a
,
fig. 26a
Description: Some sections of this recently discovered taxon in the Carnic Alps (Vachard in Krainer et al., 2019)
Occurrence: Lott 19#4 4933.1', 4972.9'.
Tubiphytes
obscurus
Maslov,
1956
Pl. 5 ![]() ,
fig. 47
,
fig. 47
Description: Rare typical bioconstructions.
Occurrence: Lott 19#4 4980.7'.
Hemigordiellina sp.
Pl. 4 ![]() ,
figs. 3, 14, 24, 39a
,
figs. 3, 14, 24, 39a
Description: Several small glomospirally coiled miliolates.
Occurrence: Lott 19#2 4930'; Lott 19#3 4928'; Lott 19#4 4926.9', 4933.1', 4944.9', 4961.7'.
Cornuspira sp.
Pl. 4 ![]() ,
figs. 4, 27, 28
,
figs. 4, 27, 28
Description : Rare, planispirally coiled, small foraminifers.
Occurrence: Lott 19#2 4930'; Lott 19#3 4915'; Lott 19#4 4937.5', 4940.8', 4944.9'.
Hemigordius sp.
Pl. 4 ![]() ,
fig. 1
,
fig. 1
Description: A discoid to subrectangular species with few planispirally coiled, evolute, last whorls.
Occurrence: Lott 19#2 4926'; Lott 19#3 4909', 4911'; Lott 19#4 4911.5'; Lott 19#5 4950'.
Glomomidiella
infrapermica
Vachard et al., 2015
Pl. 4 ![]() ,
fig. 39a
,
fig. 39a
*2015 Glomomidiella infrapermica Vachard et al., p. 54 , Figs. 22.1, 22.3, 23.1, 23.3, 23.7, 23.8, 35.9, 35.11, 35.14, 36.1-36.12, 36.16.
Description: Large species, streptospirally coiled, with small pseudo-septa and a neosparitized wall.
Occurrence: Lott 19#4 4956'.
Glomomidiella spp.
Pl. 4 ![]() ,
figs. 16, 21; Pl. 5
,
figs. 16, 21; Pl. 5 ![]() ,
fig. 40
,
fig. 40
Description: Several subaxial sections.
Occurrence: Lott 19#3 4928'; Lott 19#4 4919.3', 4926.9', 4956', 4957.5', 4958', 4960'; Lott 19#5 4950'.
Praeneodiscus spp.
Pl. 5 ![]() ,
figs. 49, 58
,
figs. 49, 58
Description: Several subaxial sections of this recently described genus of miliolates (Krainer et al., 2019).
Occurrence: Lott 19#2 4930'; Lott 19#3 4911', 4925', 4928'; Lott 19#4 4933.1', 4956', 4957.5', 4960'; Lott 19#5 4945', 4950'; Lott 19#11 4891'.
Nodosinelloides
bella
(Lipina, 1949)
Pl. 4 ![]() ,
fig. 9; Pl. 5
,
fig. 9; Pl. 5 ![]() ,
fig. 53
,
fig. 53
*1949 Nodosaria bella Lipina, p. 216, Pl. 4, fig. 9; Pl. 6, fig. 4.
Description: Small species with less than 10 globular chambers
Occurrence: Lott 19#3 4915'; Lott 19#4 4944.9'; Lott 19#5 4968'.
Nodosinelloides
shikhanica
(Lipina, 1949)
Pl. 5 ![]() ,
fig. 54
,
fig. 54
*1949 Nodosaria shikhanica Lipina, p. 217-218, Pl. 4, figs. 7-8; Pl. 6, figs. 3, 9.
Description: Small species, very compressed and with numerous chambers.
Occurrence: Lott 19#6 4946'.
Nodosinelloides sp.
Pl. 5 ![]() ,
fig. 55
,
fig. 55
Description: Small species with irregular chambers a curved axis of growth.
Occurrence: Lott 19#6 4946'.
Praerectoglandulina cf. turae (Baryshnikov in Baryshnikov et al., 1982)
Pl. 4 ![]() ,
fig. 38; Pl. 5
,
fig. 38; Pl. 5 ![]() ,
fig. 57
,
fig. 57
*1982 Frondicularia turae Baryshnikov in Baryshnikov et al., p. 33-34, Pl. 9, figs. 2-3.
Description: Rare subaxial sections.
Occurrence: Lott 19#4 4956'; Lott 19#11 4891'.
Praerectoglandulina
pusilla
(Miklukho-Maklay, 1954)
Pl. 4 ![]() ,
fig. 36; Pl. 5
,
fig. 36; Pl. 5 ![]() ,
figs. 46, 48
,
figs. 46, 48
*1954 Geinitzina pusilla Grozd. (in litt.) Miklukho-Maklay, p. 34, Pl. 3, fig. 7.
Description: Several subaxial sections of this small species
Occurrence: Lott 19#2 4934'; Lott 19#4 4944.9', 4950'; Lott 19#5 4945'; Lott 19#11 4891'.
Pachyphloia sp.
Pl. 4 ![]() ,
figs. 10, 13-14, 16
,
figs. 10, 13-14, 16
Description: Several subaxial sections of a small, primitive species.
Occurrence: Lott 19#3 4911'; Lott 19#4 4944.9', 4956'.
Plate 1: Column 1 (A): a, d, g, j, m, p, s, "touching vugs" subparallel to bedding; column 2 (B): b, e, h, k, n, q, t, "touching vugs" oblique to bedding; column 3 (C): c, f, i, l, o, r, u, residual primary intergranular pores. Row 1: a-c, Lott 19#10 4939'; row 2: d-f, Lott 19#11 4891'; row 3: g-i, Lott 19#11 4905'; row 4: j-l, Lott 19#2 4926'; rows 5-6: m-r, Lott 19#2 4934'; row 7: s-u, Lott 19#2 4940'. |
|
Plate 2: Column 1 (A): a, d, g, j, m, p, s, "touching vugs" subparallel to bedding; column 2 (B): b, e, h, k, n, q, t, "touching vugs" oblique to bedding; column 3 (C): c, f, i, l, o, r, u, residual primary intergranular pores. Rows 1-2: a-f, Lott 19#4 4918'; row 3: g-i,, Lott 19#4 4926.9'; row 4: j-l, Lott 19#4 4937.5'; row 5: m-o, Lott 19#4 4940.8'; row 6: p-r, Lott 19#4 4957.5'; row 7: s-u, Lott 19#5 4958'. |
|
Plate 3: Column 1 (A): a, d, g, j, m, p, s, "touching vugs" subparallel to bedding; column 2 (B): b, e, h, k, n, q, t, "touching vugs" oblique to bedding; column 3 (C): c, f, i, l, o, r, u, residual primary intergranular pores. Row 1: a-c, Lott 19#5 4968'; row 2: d-f, Lott 19#6 4946'; row 3: g-i, Lott 19#6 4948'; row 4: j-l, Lott 19#6 4952'; row 5: m-o, Lott 19#6 4955'; rows 6-7: p-u, Lott 19#6 4958'. |
|
Plate
4: 1. Hemigordius sp. Axial section. Lott 19#2 4926'; 2. Planiinvoluta heathi (Cushman & Waters,
1928). Subaxial section. Lott 19#2 4930'; 3. Hemigordiellina sp. Axial
section. Lott 19#2 4930'; 4. Cornuspira
sp. (subaxial section, left) and Climacammina sp. (subaxial section,
right). Lott 19#2 4930'; 5. Planiinvoluta
heathi (Cushman & Waters,
1928). Axial section. Lott 19#2
4940'; 6. Endothyra sp.
Subaxial section. Lott 19#2 4940'; 7.
Palaeotextularia sp. Axial sections. Lott 19#3 4911'; 8. Gyroporella? sp. Tangential section. Lott 19#3 4915';
9. Grovesella chomatifera (Zolotova in Baryshnikov et
al., 1982). Axial section. Lott 19#3 4915'; 10. Praeneodiscus sp. Oblique section. Lott 19#3 4925';
11. Globivalvulina sp. Axial section. Lott 19#3 4925'; 12. Praeneodiscus sp. (oblique section, top
left) and Palaeonubecularia
sp. (with two superposed series of chambers, bottom). Lott 19#3 4925';
13.
Schubertella sp. Oblique section. Lott 19#3 4928'; 14. Hemigordiellina sp. Axial section. Lott 19#3 4928';
15. Globivalvulina sp. Transverse section. Lott 19#3 4928'; 16. Glomomidiella sp. Axial section. Lott 19#3 4928';
17. Hemigordius sp. Axial section. Lott 19#4 4911.5'; 18. Grovesella sp. Oblique section. Lott 19#4 4911.5';
19. Globivalvulina sp. Transverse section. Lott 19#4 4923.8'; 20. Grovesella sp. Subtransverse section. Lott 19#4 4923.8';
21. Glomomidiella sp. Axial section. Lott 19#4 4926.9'; 22. "Tolypammina" sp. Subaxial section. Lott 19#4
4926.9'; 23. Palaeonubecularia graiferi (Baryshnikov in
Baryshnikov et al., 1982) with
concentric rows of chambers and thick wall. Lott 19#4 4930'; 24. Hemigordiellina sp. Subaxial section. Lott 19#4 4933.1'; 25.
"Tolypammina" sp. Axial section. Lott 19#4 4933.1'; 26a. Paraellesmerella sp. attached on an
ooid. Axial section. Lott
19#4 4933.1'; 26b. Planiinvoluta
heathi (Cushman & Waters,
1928). Subaxial section. Lott
19#4 4933.1'; 27.
Hemigordiellina sp. (subaxial section, left) and Cornuspira sp.
(axial section, top right). Lott 19#4 4937.5'; 28. Cornuspira sp. Axial section. Lott 19#4 4940.8';
29. Grovesella sp. Subaxial section. Lott 19#4 4940.8'; 30. Nodosinelloides bella (Lipina,
1949). Axial section.
Lott 19#4 4944.9'; 31. Pachyphloia
sp. Subaxial section. Lott 19#4 4944.9'; 32. Globivalvulina sp. (subaxial section,
left) and Orthovertella
sp. (subaxial section, right). Lott 19#4 4944.9'; 33. Globivalvulina sp. Transverse section. Lott 19#4 4944.9';
34. Pachyphloia sp. Transverse section. Lott 19#4 4944.9'; 35. Pachyphloia sp. Subaxial section. Lott 19#4 4944.9';
36. Praerectoglandulina pusilla (Miklukho-Maklay,
1954).
Axial section. Lott 19#4 4950'; 37.
Pachyphloia sp. Subaxial section. Lott 19#4 4956'; 38. Praerectoglandulina cf. turae (Baryshnikov in
Baryshnikov et al., 1982).
Axial section. Lott 19#4 4956'; 39a.
Glomomidiella infrapermica Vachard et al.,
2015. Transverse
section. Lott 19#4 4956'. |
|
Plate
5: 39b. Pseudovermiporella sp. Tangential section. Lott 19#4 4956';
40. Glomomidiella sp. Subaxial section. Lott 19#4 4957.5'; 41. Abadehellopsis cf. protea (Cushman & Waters,
1928). Oblique section. Lott 19#4 4957.5'; 42. Spireitlina sp. Transverse section. Lott 19#3 4957';
43. Grovesella sp. Subtransverse section. Lott 19#4 4960'; 44. Spireitlina?
sp. (left) and Glomomidiella sp. (right). Lott 19#4 4960'; 45. Globivalvulina sp. Oblique section. Lott 19#4 4960';
46. Praerectoglandulina pusilla (Miklukho-Maklay,
1954) (subaxial
section, left) and Hemigordiellina sp. (axial section, right). Lott 19#4
4961.7'; 47. Tubiphytes obscurus
Maslov, 1956. Typical bioconstruction. Lott 19#4 4980.7'; 48. Praerectoglandulina pusilla (Miklukho-Maklay,
1954).
Axial section. Lott 19#5 4945'; 49.
Praeneodiscus sp. Axial section. Lott 19#5 4950'; 50. Globivalvulina sp. Subaxial section. Lott 19#5 4950';
51. Glomomidiella sp. Axial section. Lott 19#5 4950'; 52. Schubertella sp. Subtransverse section. Lott 19#5 4958';
53. Nodosinelloides bella (Lipina, 1949). Axial section.
Lott 19#5 4968'; 54.
Nodosinelloides sikhanica (Lipina, 1949). Subaxial section. Lott 19#6
4946'; 55. Nodosinelloides
sp. Axial section. Lott 19#6 4946'; 56.
Ammovertella sp. Subaxial section. Lott 19#6 4958'; 57. Praerectoglandulina cf. turae (Baryshnikov in
Baryshnikov et al., 1982).
Subaxial section. Lott 19#11 4891'; 58.
Praeneodiscus sp. Axial section. Lott 19#11 4891'; 59. Neoendothyra sp.
Axial section. Lott 19#11 4891'; 60.
Tetrataxis sp. Subaxial section. Lott 19#11 4891'; 61. Brachiopod shell. Lott 19#3 4825'. |
|