◄ Carnets Geol. 22 (7) ►
![]()
Outline:
[1. Introduction]
[2. Material ]
[3.History of investigations ...]
[4. Discussion and results]
[5. Systematics]
[6. Paleoecology]
[7. Stratigraphy]
[8. Geographical distribution]
[9. Conclusions]
[Bibliographic references] [Plates] and ...
[Appendices]
Croatian Geological Survey, Sachsova 2, HR-10000 Zagreb (Croatia)
Published online in final form (pdf) on June 4, 2022
DOI 10.2110/carnets.2022.2207
![]()
[Editor: Igor Vlahović; language editor: Simon F. Mitchell; technical editor: Bruno Granier]
![]()
The taxonomic classification and mutual distinction of the genera Physoporella Steinmann and Oligoporella Pia (Dasycladales, green algae) have proven problematic and that has impeded their use in stratigraphy and other studies. The presence of single whorls of piriform laterals in Physoporella and double whorls in Oligoporella is considered as the main characteristic that separates these genera. According to that, diagnoses of genera and all species and varieties are changed or supplemented, and detailed descriptions are given. For this purpose the type-material from Pia's collection was re-examined and its redocumentation and reinterpretation performed. Additional information was obtained from abundant material from Croatia (Ivan�čica Mt, Medvednica Mt, �umberak Mt, Lika, Dalmatia). Only a few taxa were examined based on literature data. For all species and varieties reconstructions are given. Taxa from the Permian of East Asia are only briefly described. Among the investigated taxa, some of them have a structure that significantly differs from that present in the type species of the genera Physoporella and Oligoporella. They are separated into two new genera Ardeiporella and Neophysoporella. The revised genera Physoporella and Oligoporella, together with the newly established ones, give a clearer picture of phylogenetic relations.
� Triassic;
� Permian;
� calcareous algae;
� Dasycladales;
� taxonomy;
� phylogeny
Grgasović T. (2022).- Taxonomy of the fossil calcareous algae: Revision of genera Physoporella Steinmann and Oligoporella Pia (Dasycladales).- Carnets Geol., Madrid, vol. 22, no. 7, p. 171-310.
Taxinomie des algues calcaires fossiles : R�vision des genres Physoporella Steinmann et Oligoporella Pia (Dasycladales).- La classification taxinomique et la distinction entre les genres Physoporella Steinmann et Oligoporella Pia (Dasycladales, algues vertes) se sont av�r�es probl�matiques et ont entrav� leur utilisation dans la stratigraphie et d'autres �tudes. La pr�sence de verticilles simples de piriformes lat�rales chez Physoporella et de doubles verticilles chez Oligoporella est consid�r�e comme la principale caract�ristique qui s�pare ces genres. Sur cette base, les diagnoses des genres, mais aussi de toutes les esp�ces et vari�t�s, sont modifi�es ou compl�t�es, et des descriptions d�taill�es sont donn�es. � cette fin, les types de la collection de Pia ont �t� r�examin�s et leur redocumentation et r�interpr�tation ont �t� effectu�es. Des informations suppl�mentaires ont �t� obtenues � partir d'un mat�riel abondant provenant de gisements en Croatie (Mont Ivan�cica, Mont Medvednica, Mont �umberak, Lika, Dalmatie). Seuls quelques taxons ont �t� examin�s sur la base des donn�es de la litt�rature. Pour toutes les esp�ces et vari�t�s, des reconstructions sont donn�es. Les taxons du Permien d'Asie de l'Est ne sont que bri�vement d�crits. Parmi les taxons �tudi�s, certains pr�sentent une structure significativement diff�rente de celle pr�sente dans les esp�ces types des genres Physoporella et Oligoporella. Deux nouveaux genres, Ardeiporella et Neophysoporella, sont introduits pour les distinguer. Les genres Physoporella et Oligoporella, r�vis�s, ainsi que les genres nouvellement �tablis donnent une image plus claire des relations phylog�n�tiques.
� Trias ;
� Permien ;
� algues calcaires ;
� Dasycladales ;
� taxinomie ;
� phylog�nie
Dasycladalean algae are a group of green calcareous benthic algae that are important guide fossils in carbonate successions and in some levels of the Permian, Middle Triassic, Upper Jurassic, Lower Cretaceous and Paleocene are important lithogenetic fossils. Due to their relatively large dimensions and thus easier visibility, they are a great help in geological mapping. Despite the frequency of fossil algae in carbonate successions and their generally accepted importance in sedimentology, paleoecology and biostratigraphy of carbonates, the possibilities of their application in geological research have not yet been sufficiently exploited. One of the reasons is the inconsistency of taxonomic criteria in the description of new taxa, as well as the fragmentation of the description of algae in numerous papers, which makes it difficult for non-experts to make a taxonomic determination. Monographic papers on individual groups of these algae are generally rare.
The aim of this paper is to investigate in detail a group of these algae from the Triassic (partly Permian) that have proved to have a problematic taxonomomy, yet are widely distributed through many carbonate successions. These are the genera Physoporella Steinmann and Oligoporella Pia. I tried to explore this group on the basis of uniform taxonomic criteria and jointly present and redefine their taxonomic position. Each taxon (genus, species and variety) will be described in detail, a reconstruction of the algal structure will be given, and differences and similarities with related taxa will be described. For most taxa, a new or supplemented diagnosis will be given, since many taxa were described a long time ago and either, do not have appropriate diagnoses, or only have a shorter or longer description without diagnoses presented in uniform ways. Efforts will also be made to give a precise stratigraphic position for each taxon or group of taxa. Synonymies will be given only in a shorted form because they have been presented in detail by Granier and Grgasović (2000).
I hope that this paper will deepen the knowledge about this really interesting fossil group and facilitate the determination of, and further research on individual taxa from the Permian and Triassic.
In this paper all 52 species and 14 varieties of the genera Physoporella Steinmann and Oligoporella Pia that were previously known are described.
Revisions of certain species, especially the type ones, are based as much as possible on the original type material. G�mbel's material is unfortunately lost (Pia, 1912, p. 26, 1935a, p. 223, Ott, 1974, p. 31-32, Steininger in Barattolo et al., 1993, p. 28), so it was necessary to select neotypes among the material of Julius von Pia. I studied in detail the type material from Pia's collection which is kept in the Natural History Museum in Vienna (App. 1), which was necessary to make a correct and comprehensive analysis of the studied genera. The catalogue of the collection was very helpful for this work and is based on Pia's notes (partly published in the work of the Pia, 1919) and prepared by Dr. E. Gasche, the long-time head of the Geological Department of the Natural History Museum in Basel, where the collection was housed until his death. This collection has so far been used in taxonomic studies by Herak (1950), Herak in Kochansky and Herak (1960), Ott (1963), and Zorn (1972, 1974). The rock samples from which the thin-sections were made are kept mainly at the Federal Geological Institute in Vienna, while some are at the Geological Institute of the University of Vienna. Their partial catalogue was prepared by Hofmann and Granier (1994). In addition to this collection, I also studied the collection of Prof. Milan Herak, which is kept in the Geological and Paleontological Institute of the Faculty of Science in Zagreb. Part of the material described by Pia and Herak remained inaccessible to me (lost or possibly kept elsewhere), as well as material of taxa described by other authors: Bystrick�, Fl�gel and Mu, G�mbel, Hurka, Kochansky-Devid�, Kotański, Pastwa-Leszczyńska and Śliwiński, Praturlon, and Zanin Buri. Work on this material was based on the illustrations from the original works.
The investigation of the type material was supplemented by research on extensive material that I collected in north-western Croatia (Ivan�čica Mt, Medvednica Mt and �umberak Mts). These successions are described in the works of Goričan et al. (2005), Grgasović (1997), and Slovenec et al. (2019). I also used material from the Outer Dinarides of Croatia (mostly from Lika and the vicinity of Trilj in the Dalmatian hinterland), which was collected by academician Branko Sokač.
The species described by Endo (and also Ishijima, Ozaki & Nakamura) from the Permian of Japan are based mainly on poorly preserved and recrystallized material, and the author has often combined very heterogeneous material within one species or genus. For a complete revision of these taxa, it will be necessary to review the type material again, but due to the importance of Permian taxa, I give an overview of these forms.
It was necessary to literally transcribe the original diagnoses of all taxa, and in almost all of them the term "branch" is used for lateral extensions of the stem cell. This term is outdated and is not used any more, having beeing replaced with the term "lateral" (Berger & Kaever, 1992; De Castro, 1997).
Detailed statistical analysis of dimensions has been performed and dimensions of all taxa are given in the Table 1.
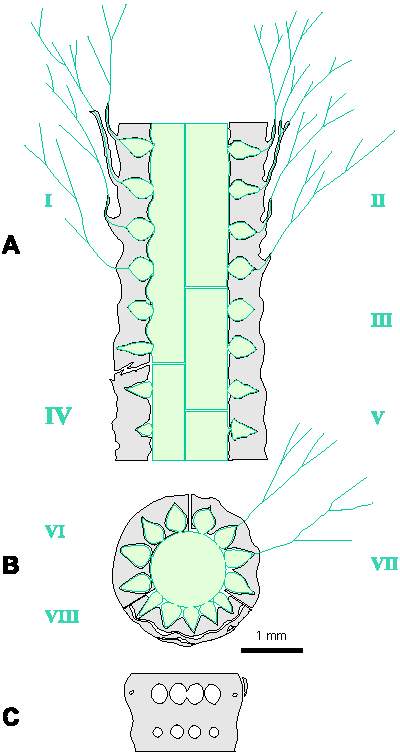
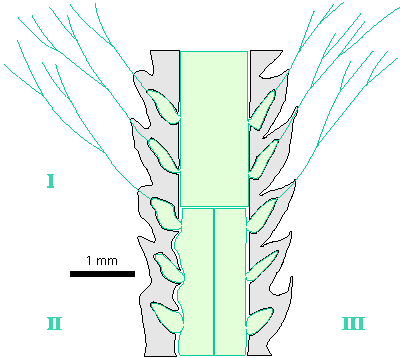
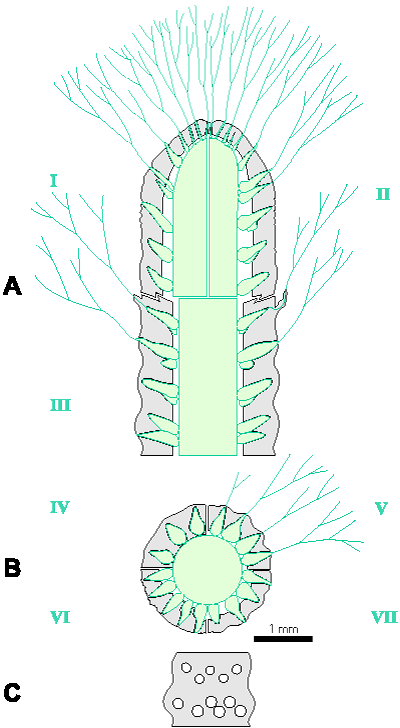
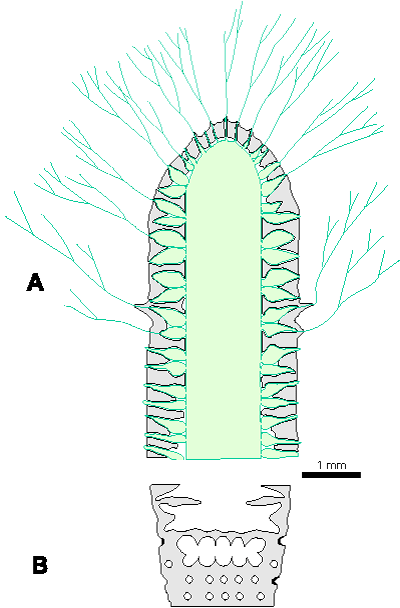
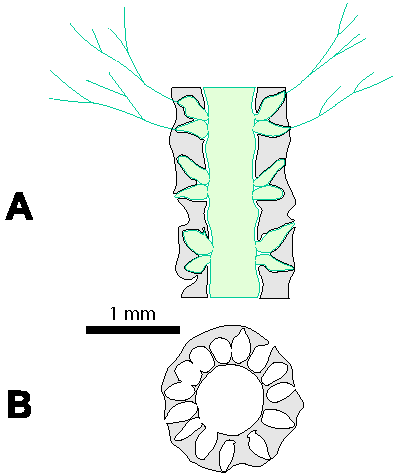
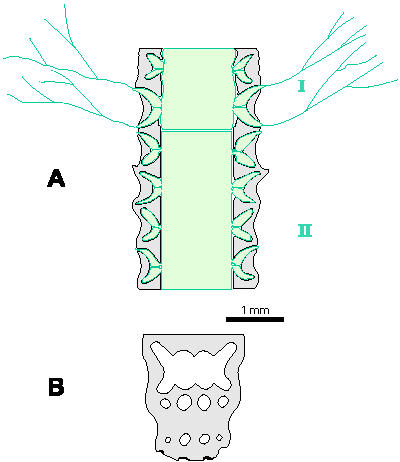
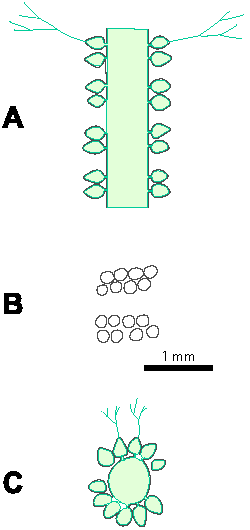
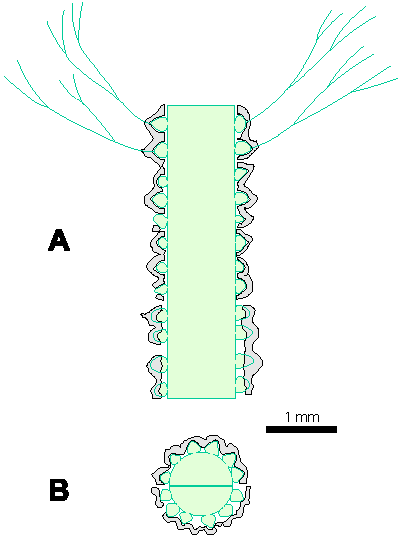
Reconstructions are not based on singular specimens, but they represent compilations of all investigated material. That is why reconstructions are divided in several parts to illustrate variations in morphology and preservation of particular taxon. A scale bar (1 mm) is added to each reconstruction, but should only be used as an approximative scale to compare the general dimensions of these algae.
Table 1: Dimensions of investigated algae (in mm). In each box, the first row of numbers represents the most common values (= mean � standard deviation), the second row of numbers the mean � standard deviation in%, the third row of numbers the total range of values, and the fourth row the number of measurements. Symbols for dimensions: Lmax - maximum thallus length, D - outer thallus diameter, d - diameter of the central cavity, d1 - diameter in its widest part, d2 - diameter in its narrowest part, d/D - inner/outer diameter ratio, h - distance between whorls, h/D - whorls distance/outer diameter ratio, h/d - whorls distance/inner diameter ratio, p - maximum thickness of primary laterals, α - inclination of laterals to the main axis, β - inclination of the lower row of laterals to the main axis, β-α - divergence of two rows of laterals, w - number of laterals per whorl.
| Lmax | D | d (d1) | d2 | d/D | d2/D | h | h/D | h/d | p | α | β | β - α | w | |
| Acroporella? prisca | 23.3 | 1.44-2.24 1.84�22% 1.32-2.57 12 |
0.51-0.99 0.75�33% 0.39-1.15 12 |
0.33-0.47 0.40�16% 0.29-0.54 12 |
0.28-1.52 0.4�31% 0.30-0.64 7 |
0.19-0.33 0.25�33% 0.39-1.1 6 |
0.44-0.72 0.58�25% 0.41-0.73 6 |
0.11-0.17 0.14�22% 0.10-0.20 13 |
≈ 52� 47-55� |
13-17 15 |
||||
| Ardeiporella karrerioidea | 5 | 1.75-2.9 2.16 4 |
0.27-0.55 0.42 4 |
0.15-0.22 0.19 4 |
0.38-0.48 0.43 2 |
0.22-0.23 0.23 2 |
1.14-1.41 0.28 2 |
0.12-0.20 0.15 4 |
≈ 60� | 16-22 19 |
||||
| Euteutloporella chia | 9.3 | 1.33 | 0.70 | 0.53 | 0.37 | 0.28 | 0.53 | 0.04-0.05 | 40� | 16 | ||||
| Neophysoporella lotharingica | 15 | 0.4-3.1 1.25 10 |
0.25-2.5 0.93 11 |
0.55-0.81 0.71 10 |
0.20-0.68 0.47 4 |
0.27-0.38 0.33 4 |
0.48-0.68 0.58 5 |
0.06-0.30 0.20 5 |
90� | 10-17 | ||||
| Neophysoporella jomdaensis | 11.7 | 1.50-1.75 | 0.8-1.3 | 0.63-0.77 | 0.12-0.25 | ≈ 0.11 | ≈ 0.18 | 0.15-0.22 | 90� | ?15 | ||||
| Neophysoporella jomdaensis from Croatia | 4.9 | 0.85-1.06 0.96 3 |
0.47-0.68 0.55 3 |
0.49-0.64 0.57 3 |
0.23-0.25 0.24 2 |
≈ 0.25 | ≈ 0.44 | 0.11 3 |
90� | 14 1 |
||||
| Neophysoporella n. sp. from Spain |
4.6 | 0.82-1.16 0.99�17% 0.67-1.25 8 |
0.41-0.75 0.58�29% 0.37-0.92 8 |
0.49-0.69 0.59�17% 0.47-0.74 8 |
0.28-0.88 0.58�52% 0.27-1.01 7 |
0.27-0.75 0.50�50% 0.27-0.81 6 |
0.55-1.09 0.82�33% 0.50-1.11 6 |
0.12-0.20 0.16�25% 0.12-0.23 8 |
90� | ? 9-10 ? 8-11 |
||||
| Neophysoporella zamparelliae | 7 | 0.88-1.38 1.13�22% 0.55-1.55 24 |
0.31-0.55 0.43�28% 0.20-0.70 24 |
0.33-0.43 0.38�13% 0.29-0.56 24 |
0.19-0.29 0.24�21% 0.13-0.30 14 |
≈ 0.21 | ≈ 0.56 | 0.13-0.23 0.18�28% 0.08-0.25 13 |
90� | 8-12 10�20% 7-14 16 |
||||
| Oligoporella cornuta | 7.2 | 1.84-2.32 2.08�12% 1.66-2.44 13 |
1.06-1.54 1.30�19% 1.0-1.66 13 |
0.68-1.06 0.87�22% 0.66-1.16 13 |
≈ 0.63 | 0.36-0.48 0.42�15% 0.36-0.52 13 |
0.55-1.0 0.71 4 |
0.30-0.46 0.38 4 |
≈ 0.82 | 0.22-0.33 0.25 12 |
47-66� 56.3��16% 43-75� 15 |
108-127� 117.3��8% 105-139� 15 |
45-77� 61��27% 33-83� 15 |
≈ 2x 13 |
| Oligoporella dissita | 12 | 1.65-2.12 1.88�12% 1.42-2.33 22 |
0.71-1.17 0.94�24% 0.55-1.40 24 |
0.38-0.54 0.46�17% 0.34-0.68 24 |
0.30-1.22 0.83 4 |
≈ 0.44 | ≈ 0.88 | ≈ 0.25 | 73-89� 81��10% 68-90� 14 |
76-98� 87.2��13% 70-99� 14 |
0-14� 6.7��100% 0-20� 14 |
≈ 2x 15 | ||
| Oligoporella fluctuosa | 5.12 | 1.56 | 0.64 | 0.41 | 0.76 | 0.49 | 1.19 | 0.10 | 55� (40-70�) | 58� (40-70�) | ≈ 3� | 2x ?18-?20 | ||
| Oligoporella intusannulata | 10.3 | 0.93-1.37 1.15�19% 0.62-1.72 598 |
0.49-0.71 0.60�18% 0.31-0.84 130 |
0.39-0.63 0.51�24% 0.12-0.81 259 |
≈ 0.52 | ≈ 0.44 | 0.59-0.81 0.70�16% 0.37-0.94 455 |
≈ 0.5-0.7 0.61 |
≈ 1.16 | 0.12-0.20 0.16�25% 0.06-0.31 1199 |
37-57� 47��21% 27-79� 181 |
83-123� 103��26% 28-144� 181 |
≈ 56� | 2x 13-15 |
| Oligoporella laevis | 4.5 | 0.96-1.48 1.22�21% 0.9-1.6 10 |
0.47-0.79 0.63�25% 0.4-0.9 9 |
0.46-0.56 0.51�10% 0.44-0.57 9 |
0.5-0.7 0.65�14% 7 |
0.45-0.67 0.54�15% 6 |
0.83-1.5 1.11�21% 6 |
0.18-0.24 0.21�14% 0.17-0.25 10 |
≈ 90� | ≈ 90� | ≈ 0� | 2x 10-13 | ||
| Oligoporella minutula | 7 | 1.30-2.90 1.85 7 |
0.50-2.00 0.90 7 |
0.35-0.66 0.45 7 |
0.47-2.00 0.80 3 |
≈ 0.43 | ≈ 0.89 | ≈ 0.19 | 68.7��15% 52-88� 10 |
74.7� �12% 58-90� 9 |
2-10� 6.1� �60% 2-14� 9 |
≈ 2x 15 | ||
| Oligoporella pilosa var. intusannulata |
11.5 | 2.05-3.07 2.56�20% 1.72-3.22 27 |
1.33-2.23 1.78�25% 1.05-2.44 27 |
0.92-1.72 1.32�30% 0.72-2.00 27 |
0.62-0.76 0.69�10% 0.61-0.83 27 |
≈ 0.52 | 0.53-0.81 0.67�21% 0.5-1.0 14 |
0.27 0.20-0.42 14 |
≈ 0.38 | 0.2-0.4 0.30�33% 0.13-0.50 27 |
79-85� 82��40% 75-88� 24 |
85-92� 93��9% 78-111� 24 |
3-20� 11��45% 3-36� 24 |
2x 14-15 |
| Oligoporella pilosa var. pilosa |
35 | 1.87-2.59 2.23�16% 1.4-3.0 33 |
0.91-1.49 1.20�24% 0.8-2.0 30 |
1.06-1.52 1.29�18% 1.0-1.7 13 |
0.46-0.62 0.54�14% 0.40-0.72 30 |
≈ 0.58 | 0.44-0.84 0.61 7 |
0.19-0.33 0.27 7 |
≈ 0.51 | 0.11-0.19 0.15�27% 0.11-0.33 31 |
74-86� 80��7% 74-89� 14 |
83-90� 87��4% 80-91� 14 |
4-10� 7��45% 1-11� 14 |
2x 15-18 |
| Oligoporella praealpina var. bosniaca |
12.4 | 2.7-3.1 2.9 |
1.1-2.0 1.7 |
0.38-0.74 0.56 |
0.83-0.90 0.87 |
0.29-0.31 0.30 |
0.45-0.75 0.60 |
≈ 0.23 | ≈ 77� | ≈ 97� | ≈ 20� | ?2x 16-18 | ||
| Oligoporella praealpina var. praealpina |
7.5 | 2.0-2.6 2.7 |
1.0-1.6 1.3 |
0.49-0.62 0.56 |
0.52-0.87 0.7 |
0.26-0.33 0.29 |
0.52-0.54 0.53 |
0.16-0.33 0.27 |
72-83� 77��7% 68-87� 23 |
73-88� 81��9% 70-91� 23 |
0-5� 3��86% 0-10� 23 |
2x 11 | ||
| Oligoporella varicans | 15 | 2.03-3.34 2.70�24% 1.6-3.9 10 |
0.93-1.91 1.42�35% 0.45-2.20 10 |
0.41-0.61 0.51�20% 0.28-0.61 10 |
0.86-1.40 1.13�24% 0.75-1.50 6 |
0.43-0.53 0.48�20% 0.43-0.56 6 |
≈ 0.80 | 0.24-0.40 0.32�25% 0.2-0.5 11 |
46-60� 49��21% 33-62� 10 |
117-133� 125��7% 115-141� 10 |
65-96� 80��19% 64-98� 10 |
12-16 14�14% 11-17 7 |
||
| Physoporella chichibuensis | 1.16-1.60 1.4 3 |
0.60-0.84 0.72 3 |
0.52-0.53 0.52 3 |
? | 0.15-0.21 0.18 3 |
?90� | ≈ 10 | |||||||
| Physoporella croatica | 16 | 1.16-2.68 1.92�40% 0.64-3.42 56 |
0.27-0.74 0.50�47% 0.09-1.67 56 |
0.15-0.40 ≈ 0.26 56 |
0.34-0.73 0.53�37% 0.24-1.07 13 |
≈ 0.28 | ≈ 1.06 | 0.25-0.42 0.34�25% 0.16-0.58 46 |
90� | pvert 0.20-0.80 0.46 15 |
7-9 7.5 5-9 13 |
|||
| Physoporella? elegans | 6.6 | 0.73-1.03 0.84 3 |
0.34-0.62 0.45 3 |
0.47-0.63 0.54 3 |
0.20-0.24 0.22 3 |
0.24-0.31 0.28 2 |
0.39-0.67 0.53 2 |
0.09 3 |
≈ 70� | ? 13 | ||||
| Physoporella? heraki var. heraki |
7.04 | 2.44-3.86 3.15�23% 2.2-5.3 61 |
1.51-2.63 2.07�27% 1.3-3.9 61 |
0.59-0.71 0.65�9% 0.50-0.77 61 |
0.22-0.32 0.27�20% 0.17-0.39 60 |
90� | 15-29 22�30% 14-38 10 |
|||||||
| Physoporella? heraki var. tenuipora |
2.06-4.78 3.69 6 |
1.33-3.50 2.52 6 |
0.65-0.73 0.68 6 |
0.17-0.22 0.17 6 |
90� | |||||||||
| Physoporella kitakamiensis | 1.42-1.90 1.66 6 |
0.94-1.38 1.08 6 |
0.60-0.75 0.68 6 |
0.20-0.29 0.25 3 |
0.11-0.18 0.15 3 |
0.21-0.24 0.23 3 |
0.14-0.23 0.19 6 |
60-90� 90� 4 |
14-18 16 3 |
|||||
| Physoporella lativentrusa | 1.46 | 0.96 | 0.66 | 0.18 | 0.12 | 0.19 | 0.12 | 90� | 19 | |||||
| Physoporella leptotheca | 4 | 0.76-1.12 0.94�19% 0.5-1.29 12 |
0.42-0.62 0.52�18% 0.4-0.68 12 |
0.50-0.62 0.56�11% 0.47-0.64 12 |
0.48-0.66 0.57�16% 0.40-0.69 10 |
0.48-0.68 0.58�17% 0.40-0.66 10 |
0.85-1.25 1.05�19% 0.70-1.39 10 |
0.08-0.14 0.11�16% 9 |
90� | ≈ 20 | ||||
| Physoporella longipora | 4 | 1.40-1.85 1.62 3 |
0.37-0.52 0.46 3 |
0.26-0.30 0.28 3 |
0.53 | 0.38 | 1.89 | 0.11?-0.16 0.15 3 |
40� | 12-14 | ||||
| Physoporella nipponica | 4.1 | 1.26 | 0.80 | 0.63 | 0.37 | 0.29 | 0.46 | 0.16 | 80-90� | 14 | ||||
| Physoporella nyugawensis | 1.20 | 0.70 | 0.58 | ? | 0.15 | ?90� | ≈ 14 | |||||||
| Physoporella pauciforata var. gemerica |
10.9 | 2.32-2.90 2.61�11% 1.99-3.25 17 |
1.10-1.60 1.35�19% 0.84-1.72 17 |
0.43-0.59 0.51�14% 0.31-0.60 17 |
0.41-0.61 0.51�20% 0.37-0.69 11 |
0.16-0.22 0.19�16% 0.13-0.24 10 |
0.28-0.42 0.35�21% 0.22-0.44 10 |
0.37-0.47 0.42�12% 0.35-050 16 |
90� 80-90� |
? 10-15 | ||||
| Physoporella pauciforata var. pauciforata |
13.5 | 1.71-2.85 2.28�25% 1.07-3.70 47 |
0.83-1.58 1.21�31% 0.5-2.0 46 |
0.45-0.59 0.52�14% 0.35-0.71 46 |
0.52-0.82 0.67�23% 0.45-0.95 26 |
0.21-0.35 0.28�24% 0.18-0.49 26 |
0.21-0.35 0.28�24% 0.35-1.20 24 |
0.24-0.40 0.32�24% 0.11-0.43 34 |
76-90� 84��10% 70-90� 27 |
12-16 14�11% 10-16 8 |
||||
| Physoporella pauciforata var. sulcata |
10.3 | 1.29-2.29 1.79�28% 0.84-2.76 17 |
0.54-1.12 0.83�35% 0.33-1.34 17 |
0.40-0.52 0.46�13% 0.38-0.57 17 |
0.45-0.77 0.61�26% 0.24-0.91 15 |
0.26-0.46 0.36�28% 0.19-0.47 15 |
0.55-1.09 0.82�33% 0.38-1.20 15 |
0.17-0.32 0.25�32% 0.13-0.36 17 |
52-70� 61��15% 50-80� 17 |
≈ 11 ? 10-13 |
||||
| Physoporella pauciforata var. undulata |
2.05-3.15 2.6�21% 0.71-3.66 39 |
0.90-1.68 1.29�30% 0.18-2.08 39 |
0.45-0.59 0.52�14% 0.41-0.62 14 |
0.67-0.95 0.81�17% 0.45-1.00 24 |
0.26-0.32 0.29�11% 0.24-0.33 7 |
0.23-0.37 0.30�24% 0.11-0.50 36 |
≈ 90� | 15-19 17�13% 14-20 6 |
In his work on the new species Tetraploporella Reme�i Steinmann (1903, p. 50), he also established a new genus Physoporella. In the chapter "Comparison", after the part of the text where he discusses the formation of spores in the stem cell, he writes the following: "The same applies to the Triassic (and Permian) representatives of the genus Diplopora, whose verticillated branches are more or less filamentous and unramified. In addition to such forms, however, Diplopora also include others, in which verticillated branches are very wide, often almost spherically shaped as in D. pauciforata Gue., macropora Gue. and others. The cavities of these forms, corresponding to the branches, seem to have been closed to the outside when completely preserved; and only with erosion do they get an outward opening. Due to this feature they are similar to the genus Gyroporella, in which it has undoubtedly been established that there are cavities of branches closed outwards. I have always suspected that these forms with pear-shaped to spherical branches that do not perforate the calcareous shell were fertile, which on the other hand indicates that they thus represent the starting point of other species� I name these forms Physoporella."
(In German: "Das gleiche gilt f�r die triadischen (und permischen) Vertreter der Gattung Diplopora, deren Wirtelaste � fadenf�rmig und unverzweigt sind. Ausser solchen Formen werden aber als Diplopora auch andere bezeichnet, bei denen die Wirtelaste sehr weit, oft fast kugelig gestaltet sind, wie D. pauciforata Gue., macropora Gue. u. A. Die den Wirtelasten entsprechenden H�hlungen dieser Formen scheinen bei vollst�ndiger Erhaltung auch gegen Au�en geschlossen gewesen zu sein; erst durch Abrollung erhalten sie eine M�ndung nach Au�en. Durch diese Beschaffenheit n�hern sie sich der Gattung Gyroporella, bei welcher zweifellos stets nach Au�en geschlossene Zweighohlen vorhanden sind. Ich habe von jeher vermuthet, dass diese Forme mit birnf�rmigen bis kugelf�rmigen, die Kalk h�lle nicht perforierenden Zweigen fertil gewesen sind, und werde an anderer Stelle zeigen, dass daf�r auch concrete Anhaltspunkte anderer Art vorliegen� Ich nenne diese formen Physoporella.")
Two errors were made in the mentioned diagnosis, which were also noticed by Pia (1912, p. 75): The species macropora does not exist in G�mbel's original work, its name is similar to Gyroporella macrostoma (Pia, 1920, transferred it to Diplopora annulata); and the last sentence ("I name..."), probably due to a typographic error, is placed on the same page one paragraph below.
Hence, after Steinmann (1903), the genus Physoporella includes all forms with broad pear-shaped to spherical laterals that were enclosed in a calcareous sheath.
The origin of the genus name probably comes from the Greek "" ("fysa" = bellows, bladder, bubble), which refers to the wide pores of the laterals.
Steinmann (1907, p. 19-20) gives another description of this genus in his palaeontology textbook: "Like Diplopora, often articulated. Outer surface with spaced vesicular protrusions, into which the sack-shaped cavities (verticillated branches), which exit the stem cell, are closed, and that open to the outside only when the outer surface is eroded. Spores probably in these sac-shaped verticillated branches." In the attached illustration of Physoporella pauciforata (Steinmann, 1907, Fig. 9.A-B) the laterals are vesiculifer, so Pia (1912, p. 76) warns that this does not match with his observations.
(In German: "Wie vorige oft gegliedert. Oberflache mit entfernt stehenden, blasigen Erh�hungen (w), in welche sackf�rmige, geschlossene H�hlungen (s) (Wirtelzweige) von der Stammzelle hineinsetzen, die sich nur gegen Au�en �ffnen, wenn die Oberflasche abgerieben ist. Sporen wahrscheinlich in diesen sackf�rmigen Wirtelasten.")
Pia (1912, p. 43; see also Granier & Sander, 2013, p. 32) gives a modified description of the genus Physoporella: "The type of verticillated branches I have defined as pyriferous is decisive in an assignment to this genus, that is, the pores terminate blindly, but differ from the vesiculiferous type in that the calcareous skeleton shows no distal widening. As a rule the basal part of the branches is the thickest. All species known to date have pore series, and on phylogenetic grounds it is probable that this arrangement is typical of the entire genus for presumably it was derived from the already euverticillate Oligoporella. In our genus the occurrence of closely spaced biserial verticils is common (apparently in the more specialized forms). The type of segmentation of the skeleton that we have learned to call "bulge" reaches an extreme development in some forms assigned here." As for the type species of the genus, Pia (1912, p. 44) names the species Gyroporella pauciforata G�mbel.
Hence, after Pia (1912), the genus Physoporella would include all forms with piriform laterals located in whorls and enclosed in a calcareous sheath.
In the same paper Pia (1912, p. 41-42; see also Granier & Sander, 2013, p. 28) established the genus Oligoporella: "A small number of relatively thick pores taper outwards more or less strongly, but this applies strictly only to the upper part of the plant. In primitive species the basal verticils could be of the phloiophorous type (Pia refers to the species Oligoporella prisca, see chapter 5.5.2. - comment T.G.). To date all known species are euverticillate. Spore development probably took place in the swollen distal part of the verticillated branches. Undoubtedly this genus has much in common with the genus Teutloporella. Originally I considered both of them as subgenera of a single genus. However, their habitus is so different and so easily recognizable that for practical reasons complete separation is to be recommended. Also the importance of the distinguishing characteristics compiled in the following table must not be underestimated, so nominating two discrete genera seems fully justified at this time. During the work on the phylogenetic section I became convinced that the two genera are quite distinct from each other phylogetically.
Oligoporella: The number of branches in a verticil is 10-20. Only euverticillate forms are known. Verticils are separated by distinct intervals. Verticils are commonly closely packed. Almost certainly derived from Macroporella.
Teutloporella: The number of branches in a cross-section (in normal individuals), always over 30, up to 60. The majority of forms are proverticillate. Verticils, when present, are set densely, touching each other. Verticils, when present, are always very simple. Origin unknown, if derived from Macroporella, independent for sure."
Hence, after Pia (1912), the genus Oligoporella include forms with slightly thicker trichoform laterals arranged in whorls.
As for the type species of the genus Pia (1912, p. 42) designated Oligoporella pilosa n. sp. The origin of the genus name probably comes from the Greek "" ("oligos" = small, little), which refers to the relatively small number of laterals in the whorl.
The separation of the genera Physoporella and Oligoporella on the basis of very small differences in the shape of laterals and especially their closure or openness proved to be problematic at the beginning. This is because one species and even one individual specimen may have open and closed pores. Therefore, in his next work, Pia (1920, p. 50) complements his previous diagnosis of the genus Physoporella: "Rod-shaped diplopores with pyriferous undivided branches, arranged in whorls. During the determination, it is less important whether all pores are closed outwards, but it is much more important whether reliably closed pores occur at all, and whether their shape can be assigned to the piriferous type, due to the rounding of the distal end. Whorls are often compacted to doubled. The shell is segmented by annulation or fissuration, and also undulation can be present."
(In German: "Stabf�rmige Diploporen mit piriferen, in Wirteln gestellten, unverzweigten Asten. Bei der Bestimmung ist weniger darauf Gewicht zu legen, dass alle Poren gegen Au�en geschlossen sind, als vielmehr darauf, dass �berhaupt sicher geschlossene Poren auftreten und dass die Form derselben sich durch die Zurundung des distalen Endes dem piriferen Typus anschliesst. die Wirtel sind h�ufig gedr�ngt bis zweizeilig. An der Schale ist eine Gliederung durch Annullation oder Fissuration h�ufig entwickelt oder es macht sich Undulation bemerkbar.")
Hence, after Pia (1920), the genus Physoporella would include forms with piriform laterals arranged in whorls, at least some of which are enclosed in a calcareous sheath.
It should be noted that for both genera Oligoporella (Pia, 1912) and Physoporella (Pia, 1912, 1920) Pia notes the frequent occurrence of double whorls, but neither gives this phenomenon, nor skeletal segmentation greater importance; but uses shape and closedness of pores as the diagnostic criteria. Note, however, that in the diagnosis of the type species P. pauciforata Pia (1912, p. 44) explicitly states single whorls. The problems were not solved, and Pia himself has on several occasions expressed doubts about the distinction between the genera Physoporella and Oligoporella, and the need to re-examine the type material, with the possibility of unification of these two genera. Thus Pia (1925, p. 331-332) states: "... On the contrary, it is necessary to re-examine the question of the validity of the genus Oligoporella and the accuracy of its present reconstruction. It is clear, however, that a significant change in the development tree may result, if, for example, it turns out that there were no euspondyl trichophorous Dasycladaceae in the Triassic other than Teutloporella. Furthermore, it seems possible that the name Physoporella pauciforata has so far included several related but different species." His conclusion in this paper (Pia, 1925, p. 331) is very important: "The whorls in Olig. pilosa are always compressed to double-rowed, and in Physop. pauciforata as a rule are purely single-rowed" - this will be commented on further in this text. Pia (1927, p. 71) wrote similarly: "Physoporella Steinmann was certainly derived from the genus Oligoporella. Moreover, it is doubtful whether these two genera will not have to be reunited later." Pia (1940, p. 5) described a new variety of Oligoporella pilosa forma physoporelloidea, which he established as a transitional form because of his inability to distinguish open or closed pores of the two genera, and wrote: "Physoporella can most likely be derived from Oligoporella." Furthermore Pia (1935a, p. 221-223) suggests: "The best and safest way to further separate these genera would surely be to go through all my material on Physoporellas again. However, it seems to me that this will not be possible for me in the foreseeable future. Therefore, I have to give preference to a less favourable but more practical approach, so that in each of my papers on diplopores in a particular area, I systematically examine Physoporellas in some detail. This, of course, will not be possible without a later proper revision of previous conclusions. Yet I must hold this lesser risk than no longer working at all on this genus." So despite the acceptance of the need to revise the genera Physoporella and Oligoporella and to re-examine the type material Pia never did it, but obviously left it for future generations. From previous quotations it seems that he was contemplating the rejection of the genus Oligoporella and the incorporation of all species with piriform laterals into the genus Physoporella. He considered the single-rowed and double-rowed whorls important only for distinguishing species.
Endo (1958, p. 266) gave a "re-emendation" of the genus Physoporella and established the new genus Clavaphysoporella: "Physoporella is proposed to include the species congeneric with Physoporella pauciforata G�mbel which has pirifer type pores. While I choose Physoporella minutula G�mbel as the genotype of Clavaphysoporella, established as a new genus in this paper. Its diagnosis is as follows: The thallus is cylindrical, relatively straight and consisting of fine annulations. It sometimes shows slight innerannulations. The pores are relatively slender, and usually somewhat expanded toward the exterior, and enclosed in the calcareous body, that is, they show all the characteristics of the so-called phloiophor type, and they are moderately ascending toward exterior. The pores are arranged as definite whorls which are assembled into two or three gathered lines and make cluster-like appearance." This revision, and in fact only the naming of a new genus, cannot be considered valid, as P. minutula, like all other species of the genus Physoporella, has laterals that taper outwards, and are in no case phloioform. The genus Clavaphysoporella is thus a junior synonym of the genus Physoporella, as first recognized by Kochansky and Herak (1960, p. 86), nullifying the so-called "re-emendation" of the genus Physoporella and the establishment of the new genus Clavaphysoporella, within which very heterogeneous taxa were united. Forms, with phloioform laterals arranged in diverging whorls, described by Endo, have subsequently been united within the new genus Clavaporella Kochansky & Herak with type species Clavaporella caliciformis Kochansky & Herak. This opinion was accepted by all later authors (Praturlon, 1963, p. 127; Vachard, 1980, p. 363-367; Granier & Deloffre, 1995, p. 56, 58; Granier & Grgasović, 2000, p. 22-23).
Botteron (1961, p. 59), in his establishment of the genus Anisoporella discusses the arrangement of laterals in "double whorls, as in Oligoporella", but without further comment.
Praturlon (1963, p. 127) makes a significant step forward, and for the first time clearly emphasizes the existence of two separate groups within the genus Physoporella: one with simple, more or less gathered whorls, and the other, with biseriate, alternating whorls.
Some other authors also distinguish the group with single whorls and the group with double whorls within both genera, and even discuss their different paleoecology (Bechst�dt & Brandner, 1970, p. 59-62; Zorn, 1974; Bystrick�, 1986, p. 295; Kotański, 2013, p. 42) and different stratigraphic range (Ott, 1972a, 1974), which will be commented upon later. Kotański and Čatalov (1973) introduced names for two groups within the genus Physoporella: the "Physoporella pauciforata group" (with single whorls) and the "Physoporella praealpina group" (with double whorls). Species described within the genus Oligoporella were not found in their material, so they are not commented on.
Many later authors, as Pia did earlier, repeatedly emphasized the problems of distinguishing the two genera, but no one engaged in a taxonomic revision. Thus Hurka (1967, p. 73-75) extensively discussed the great difficulties in distinguishing the genera Physoporella and Oligoporella. After analysing previous works on this topic, he again concludes that "the only criterion for distinguishing these two genera remains the answer to the question: Are all pores open (Oligoporella) or are all or many of them closed (Physoporella)?" The author announces further difficulties after such a conclusion, e.g., if we find 5 closed out of 100 pores, it seems inappropriate to add this specimen to the genus Physoporella, and suggests that the boundary between these genera should be quantitatively defined (but he did not suggest how). Hurka (1967, p. 73) also stated the existence of two morphologically well-separated groups within the both genera: a) pores in single whorls and b) pores in packed (double) whorls. An important work on the genera Physoporella and Oligoporella is his next work (Hurka, 1969) on "Tendencies to transform shapes of laterals in Physoporella-Oligoporella populations from the Anisian of Pragser Dolomites (Italy)". Pointing out the difficulties in distinguishing these genera, Hurka (1969) cites several authors who do not warn of these problems, but illustrate and describe Physoporellas that can also be easily attributed to the genus Oligoporella. This author believed that many forms labelled Physoporella are actually the result of a thin-section being too thin, such that the point of exit of the pore from the calcareous wall is simply not "captured" in the thin-section. In his paper, the author deals exclusively with the forms of laterals. He distinguishes six types of laterals and presents a scheme according to which one type of lateral would develop from another type, going from the trichoform through the piriform to the vesiculiform type. Hurka (1969) shows that there is continuous transition between Physoporella-type and Oligoporella-type pores. However, he does not engage in a revision of the genera, citing (on p. 107) that it is not the meaning and purpose of his paper to engage in a taxonomic study of the justification or unjustification of both genera. The exact taxonomic affiliation of all illustrated specimens is difficult to determine, as these illustratitions are drawings (Hurka, 1969, Figs. 1-6), so it is impossible to verify the actual appearance of the algae. Furthermore, several photographs (Hurka, 1969, Fig. 7) represent only individual laterals without the illustration of entire specimens, which is not sufficient for accurate species determination. It should be noted, however, that all these specimens in the accompanying illustrations represent forms with single-row whorls that most likely belong to the species P. pauciforata, although some specimens might also belong to the species P. croatica, which makes this whole analysis dubious. The basic problem is that the author studied exclusively the shapes of the pores, as if they were separate entities, not taking into account that they are only parts of the algae, which also has other characteristics.
The next paper dealing with this topic is Bucur et al. (1994), where in the chapter "Remarks on the Physoporella-Oligoporella group" they again repeat the conclusions from previous works, and emphasize that the principle of separation of the genera Physoporella and Oligoporella according to closed or open pores "may be equated to the degree of calcification of the thallus, a principle that, if thoroughly considered, can hardly be accepted to substantiate a generic differentiation." Citing the conclusions of Hurka (1969) on the existence of a complete transition between Physoporella and Oligoporella pores, the authors comment: "Due to his detailed analysis Hurka (1969) actually would have been entailed to unify the two genera under a single name (in which case Physoporella should have had priority)". Then they state that: "The arrangement of laterals in one-row verticils (i.e., laterals having the same inclination to the longitudinal axis within the verticil) or two-row verticils (i.e., laterals having inclination arranged in two diverging directions) may form a more adequate criterion for separation of species than the form of laterals which may vary considerably, making assignment to a particular type (i.e., trichophorous or piriferous) difficult." Even these authors do not dare to revise these genera, so in the end they conclude: "As Hurka (1969) was reluctant to unify the two genera, it seems that the solution may only be found by a rigorous revision of the type materials and reinterpretation of the taxonomic criteria"; a similar conclusion to Pia (1925, p. 331-332, see above) seventy years earlier.
Grgasović (1995) in the Abstract Book from the 6th International Symposium on Fossil Algae and Carbonate Platforms published his preliminary research results. Schlagintweit et al. (2003) first cited that paper as an emendation of the genera Oligoporella and Physoporella. Subsequently, Bucur in Rychliński et al. (2013) stated the modified names of Oligoporella cf. dissita and Oligoporella cf. minutula, and Bucur in Gawlick et al. (2021) cited the modified names Oligoporella dissita and Oligoporella praealpina according to the afore mentioned work. This emendation of the genus was also mentioned by Bucur et al. (2020) and Bucur and Matysik (2020). Later authors used this to synonymize the species O. elegans and O. prisca, with the latter species cited as a senior synonym and assigned it to the genus Physoporella. All these combinations, which are based on Grgasović (1995), are not valid, because that paper is not validly published according Art. 29.1. of the ICN (Turland et al., 2018); that is, it is not distributed and widely available to the general public.
Piros (2002, p. 135) also briefly commented on the problem of the separation of the genera Physoporella and Oligoporella: "Due to their almost identical size and pore shape a close relationship, possible even identity, of the two algal groups may also be assumed. This potential identity is confirmed by the fact that, between certain species of the Physoporella and Oligoporella groups (Oligoporella pilosa physoporelloidea - Physoporella pauciforata, Oligoporella pilosa varicans - Physoporella varicans), the only difference is in the wide aperture (characteristic of Oligoporella) and narrow aperture (characteristic of Physoporella) of the pores, respectively." Piros (2003) in her descriptions always states closeness of pores.
Granier and Sander (2013; Granier et al., 2013) translated and re-published the important work of Pia (1912) and had all his reconstructions redrawn, somewhat reinterpreted, and transformed to 3D animations.
In a recent paper, Kotański (2013) states that in both the genus Physoporella and the genus Oligoporella there are species with single-row and double-row whorls that he classifies into separate groups: Oligoporella prisca group (with single whorls), Oligoporella pilosa group (with double whorls), Physoporella pauciforata group (with single whorls) and Physoporella praealpina group (with double whorls). He suggests that these groups could be regarded as subgenera within Physoporella and Oligoporella, respectively.
From the above synthesis, there is a clear need for the (repeatedly requested) re-examination of the entire material of these two genera and to the solution of the problems of their differentiation.
4.1. Discussion
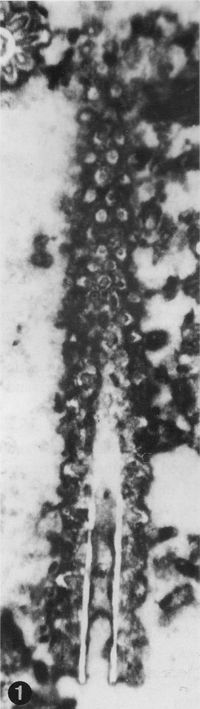
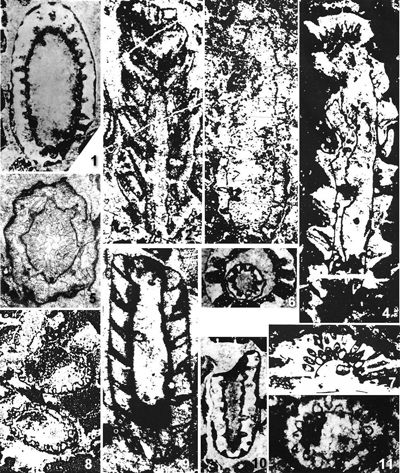
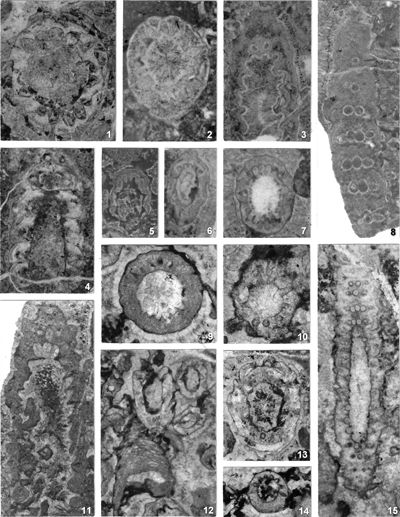
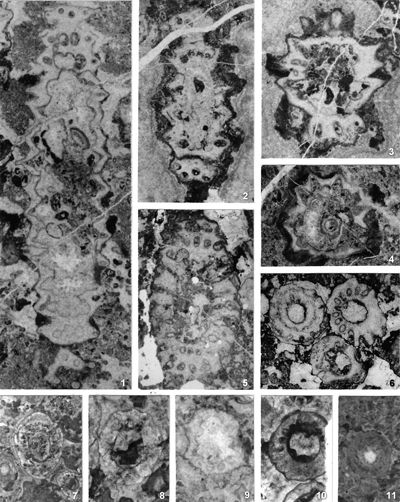
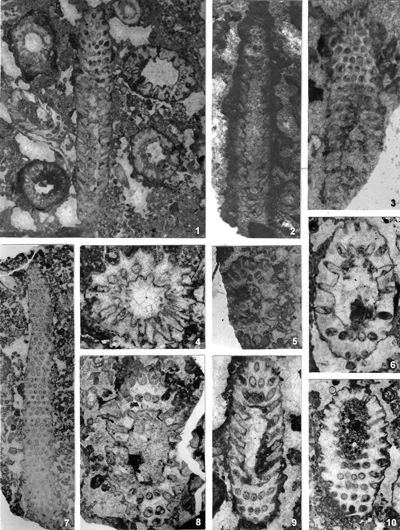
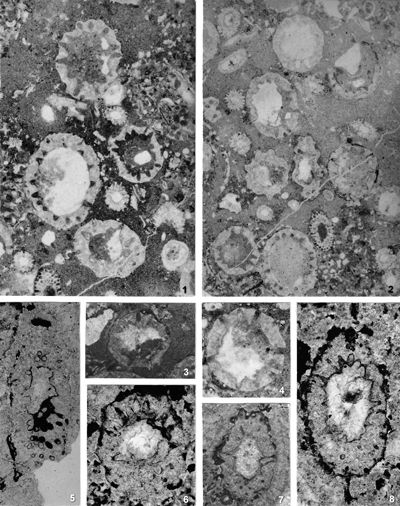
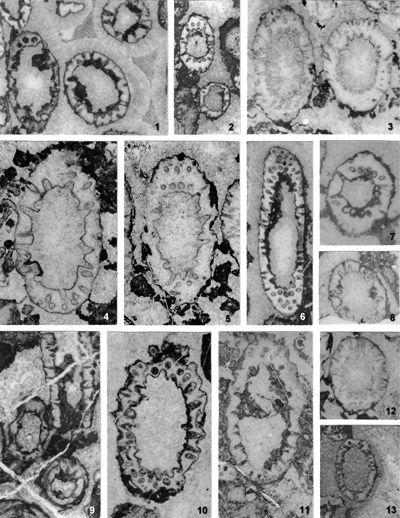
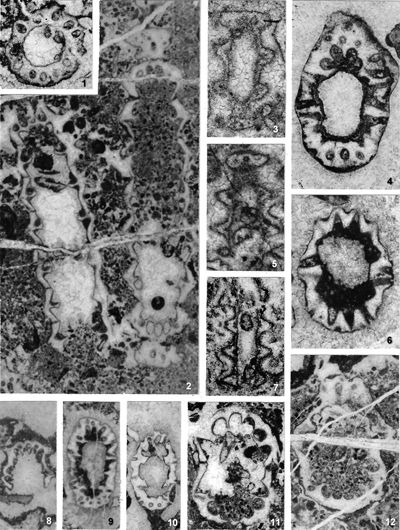
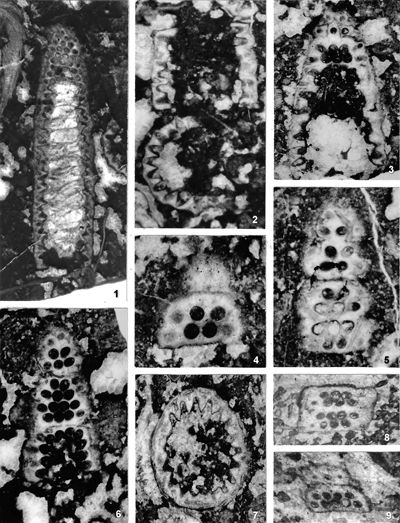
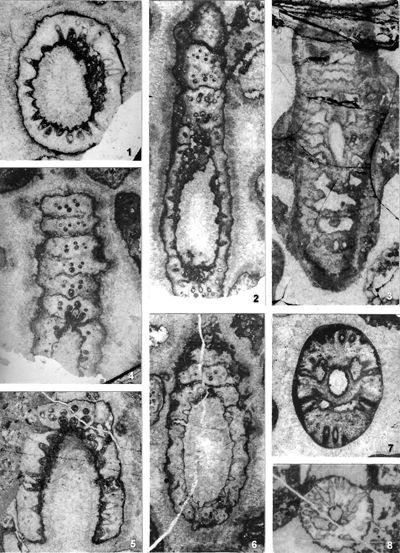
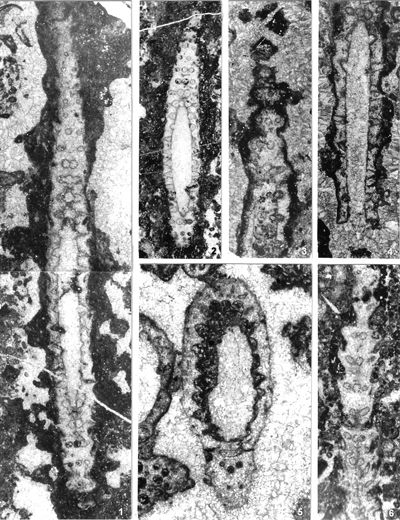
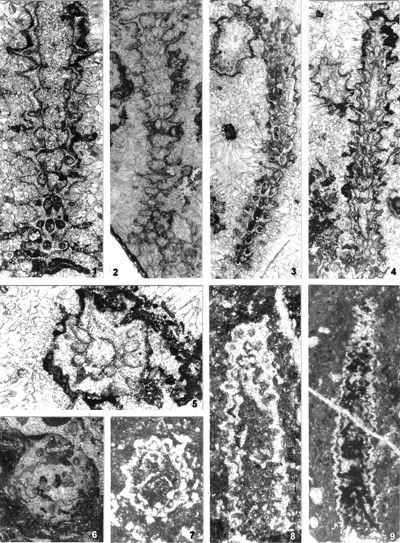
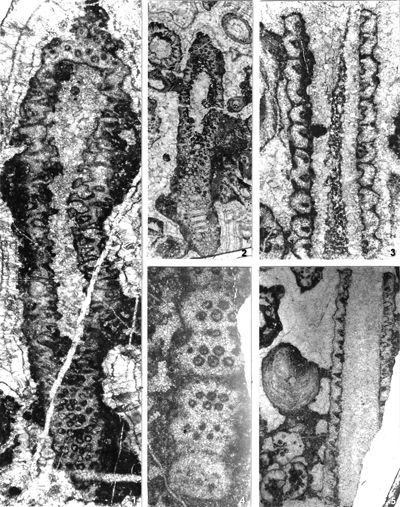
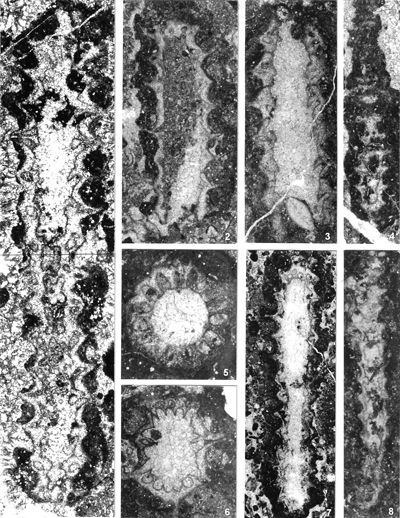
The criterion based on closedness and openness of pores of laterals has proved to be problematic in the establishment of these two genera. This becomes clear if we look at the type material of each genus as an example. In the case of type species Oligoporella pilosa with all specimens originating from the same sample (Pl. 5 ![]() , figs.
1-6, 8-11; Pl. 6
, figs.
1-6, 8-11; Pl. 6 ![]() , figs. 1-4) we see that in some specimens the pores are open (e.g., Pl. 5
, figs. 1-4) we see that in some specimens the pores are open (e.g., Pl. 5 ![]() , fig. 6), in some cases they are closed (e.g., Pl. 6
, fig. 6), in some cases they are closed (e.g., Pl. 6 ![]() , fig. 1 middle right), and in most specimens we find both open and closed pores. In the type species Physoporella pauciforata, also with all specimens from one sample, but less well preserved (Pl. 1
, fig. 1 middle right), and in most specimens we find both open and closed pores. In the type species Physoporella pauciforata, also with all specimens from one sample, but less well preserved (Pl. 1 ![]() , figs. 1-8), we see the same phenomenon of both open and closed pores. We find this in a more or less pronounced form in all (sic!) other species of these two genera.
, figs. 1-8), we see the same phenomenon of both open and closed pores. We find this in a more or less pronounced form in all (sic!) other species of these two genera.
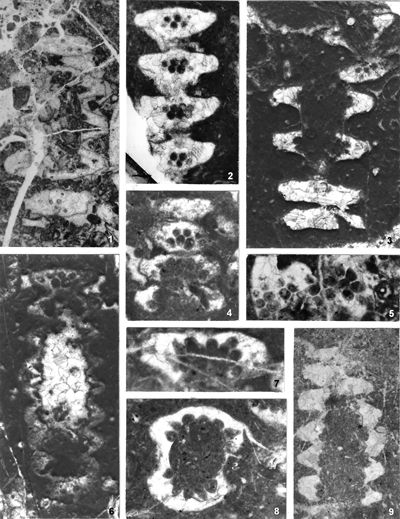
From the above examples, it is clear that this taxonomic criterion is not suitable for separating the two genera, or even the two species. The openness or closedness of pores is influenced by two factors: the degree of algae calcification and the degree of post-mortem and diagenetic change. The total extent of calcification depends on environmental factors: sea water saturation with CaCO3, temperature, light intensity and water turbulence, and possibly some biological factors. Although more detailed data on the impact of the environment on calcification are lacking, many authors cite their interrelationship (Valet, 1969, p. 552; Berger & Kaever, 1992, p. 23; De Castro, 1997, p. 108, 179; Berger et al., 1997; Granier, 2012). Particularly great variability in the extent of calcification can be seen in the well-preserved type material of O. pilosa (Pl. 6 ![]() , figs. 1-2) where in the same thin-section (sic!) we can observe laterals that are calcified only at the distal end (e.g., Pl. 6
, figs. 1-2) where in the same thin-section (sic!) we can observe laterals that are calcified only at the distal end (e.g., Pl. 6 ![]() , fig. 1
middle left) and those that are calcified all the way to the stem cell (e.g., Pl. 6
, fig. 1
middle left) and those that are calcified all the way to the stem cell (e.g., Pl. 6 ![]() , fig. 1 below). This variability is also visible, but to a lesser extent, in P. pauciforata; e.g., if we compare almost completely calcified laterals in the type material (Pl. 1
, fig. 1 below). This variability is also visible, but to a lesser extent, in P. pauciforata; e.g., if we compare almost completely calcified laterals in the type material (Pl. 1 ![]() , figs. 1, 15) with those only distally calcified from Herak (1965, Pl. XIII, fig. 5) and Bucur et al. (1994, Pl. 11, fig. 9). Postmortem and diagenetic changes can significantly alter the original morphology of the calcareous thallus in the following ways: by erosion of the outer surface during the sedimentation process, by dissolving of the calcareous shell in the early phase (by meteoric water) or in the late phase of diagenesis (by pore water), as well as by micritization. The intensity of all these processes is very variable, so it is no wonder that this taxonomic criterion proved to be unsuitable.
, figs. 1, 15) with those only distally calcified from Herak (1965, Pl. XIII, fig. 5) and Bucur et al. (1994, Pl. 11, fig. 9). Postmortem and diagenetic changes can significantly alter the original morphology of the calcareous thallus in the following ways: by erosion of the outer surface during the sedimentation process, by dissolving of the calcareous shell in the early phase (by meteoric water) or in the late phase of diagenesis (by pore water), as well as by micritization. The intensity of all these processes is very variable, so it is no wonder that this taxonomic criterion proved to be unsuitable.
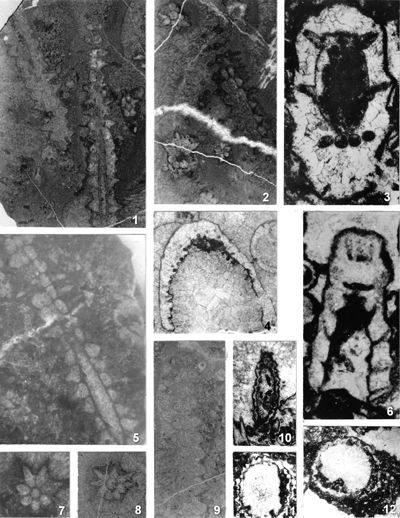
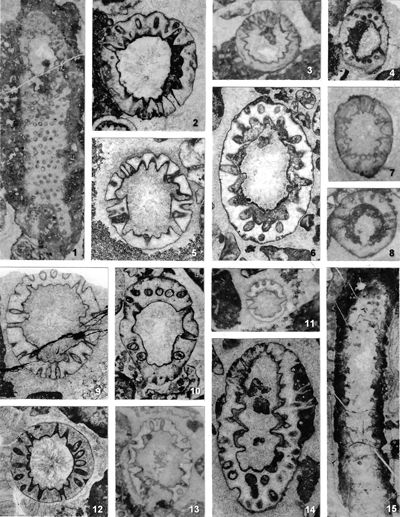
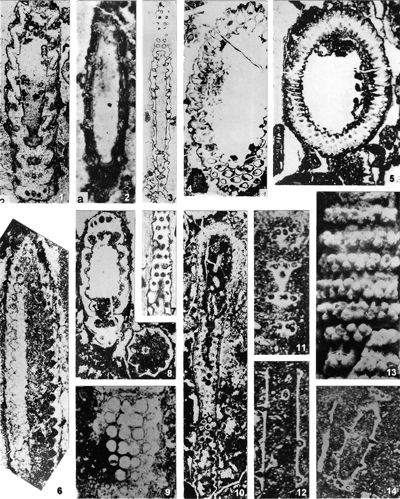
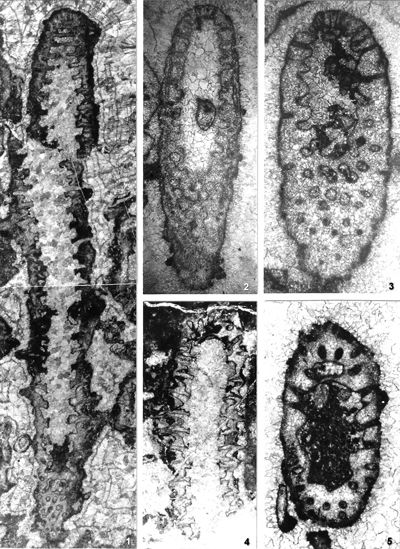
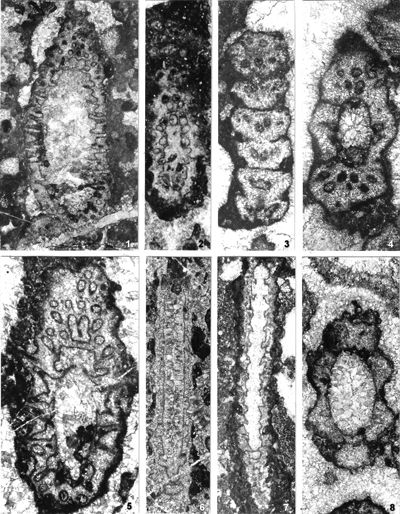
The shape of laterals in investigated genera is considered different in respective original descriptions (see the former chapter): Physoporella should have piriform laterals, and Oligoporella should have trichoform laterals. After investigation of the type material, it is clear that the shape of the laterals in the type species of both genera is very similar. If we compare the type material of P. pauciforata shown in
Pl. 1 ![]() and that of O. pilosa shown in Pls. 5
and that of O. pilosa shown in Pls. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() (as well as other species) we can see that the type of laterals is identical: a wider rounded proximal part and a gradual distal narrowing. They are similar to the extent that in cross-sections we usually cannot say which genus a specimen belongs to. There are some minor differences, so the laterals of Oligoporella are on average somewhat longer and thinner, but they are not trichoform types of laterals. Typical trichoform laterals can be found in Teutloporella herculea, Euteutloporella triasina, Diplopora nodosa, etc. and are significantly thinner than those in the genus Oligoporella. Due to their narrow diameter, they cannot be assumed to have served as gametangia, in contrary to wider piriform laterals. It is not probable that the two genera, which are obviously very closely related, had different types of reproductive organs, so it is justified to consider that the laterals of Oligoporella are of the same type, i.e., piriform.
(as well as other species) we can see that the type of laterals is identical: a wider rounded proximal part and a gradual distal narrowing. They are similar to the extent that in cross-sections we usually cannot say which genus a specimen belongs to. There are some minor differences, so the laterals of Oligoporella are on average somewhat longer and thinner, but they are not trichoform types of laterals. Typical trichoform laterals can be found in Teutloporella herculea, Euteutloporella triasina, Diplopora nodosa, etc. and are significantly thinner than those in the genus Oligoporella. Due to their narrow diameter, they cannot be assumed to have served as gametangia, in contrary to wider piriform laterals. It is not probable that the two genera, which are obviously very closely related, had different types of reproductive organs, so it is justified to consider that the laterals of Oligoporella are of the same type, i.e., piriform.
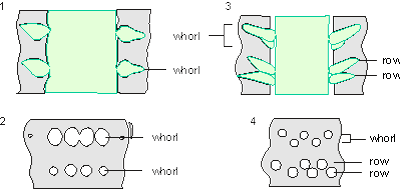
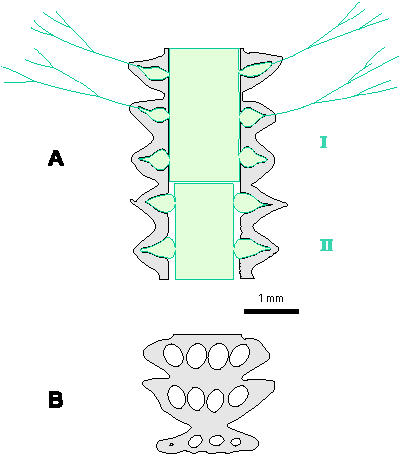
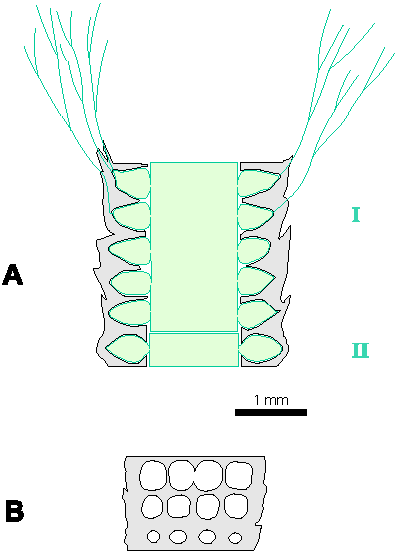
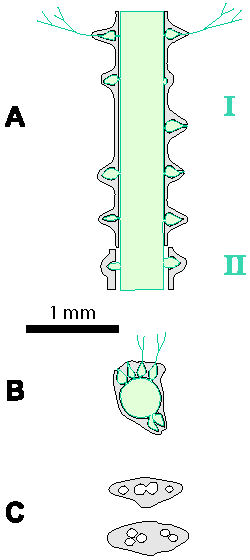
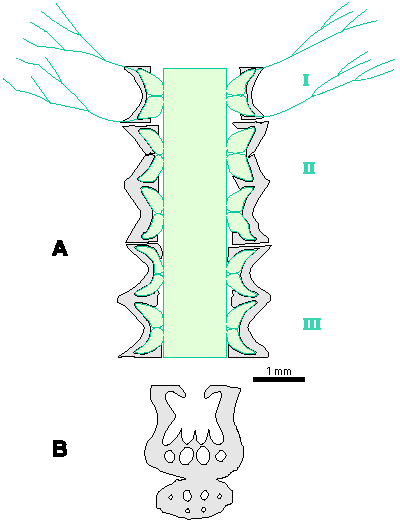
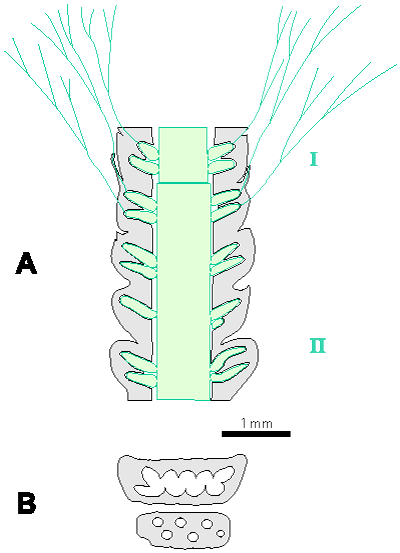
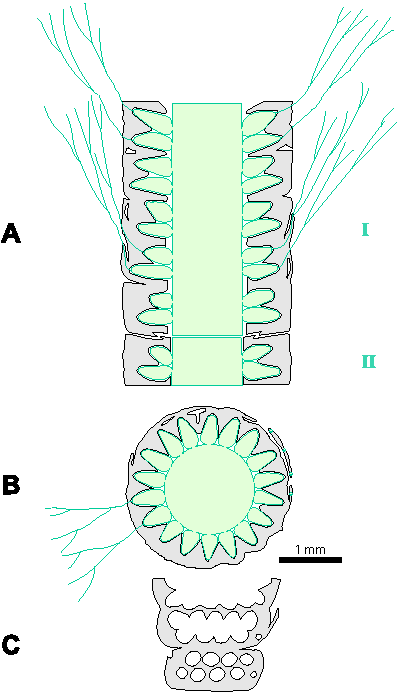
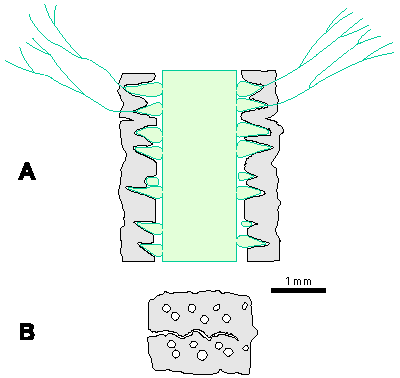
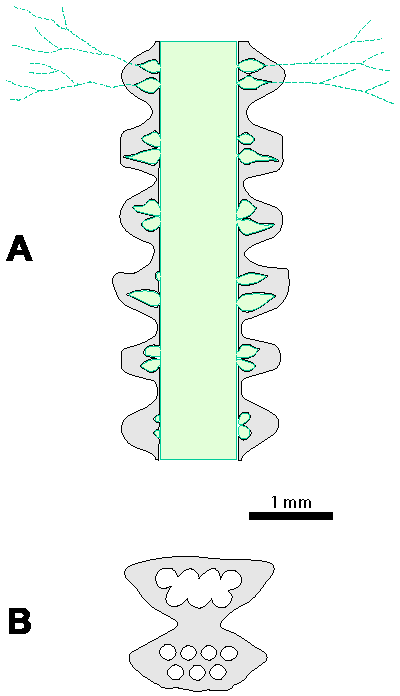
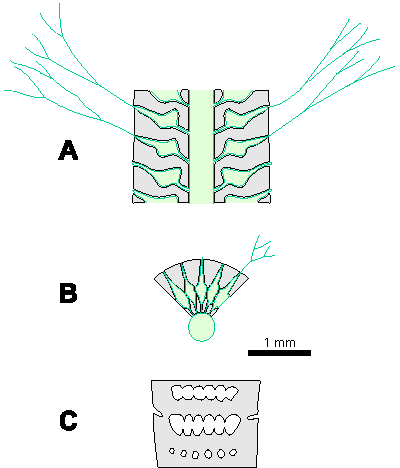
Which criterion, then, can be used to separate the genera and why? In my opinion, the presence of single or double whorls is the main characteristic that separates the genera Physoporella and Oligoporella (Fig. 1 ![]() ), primarily because of its biological importance, while openness and closedness of pores, as we have seen, are associated with environmental conditions, preservation and diagenesis. We must strive to make the taxonomic relationships among fossil algae as close as possible to actual biological relationships, and therefore to give more importance to those taxonomic criteria that have biological significance, than those that are genetically defined. The application of the above criterion, as we shall see later, facilitates a taxonomic classification and gives much clearer interrelationships among taxa. Luckily, the type species of the genus Physoporella has single whorls, and the type species of the genus Oligoporella has double whorls, so it is only necessary to change the diagnoses of the genera according to the criterion accepted above.
), primarily because of its biological importance, while openness and closedness of pores, as we have seen, are associated with environmental conditions, preservation and diagenesis. We must strive to make the taxonomic relationships among fossil algae as close as possible to actual biological relationships, and therefore to give more importance to those taxonomic criteria that have biological significance, than those that are genetically defined. The application of the above criterion, as we shall see later, facilitates a taxonomic classification and gives much clearer interrelationships among taxa. Luckily, the type species of the genus Physoporella has single whorls, and the type species of the genus Oligoporella has double whorls, so it is only necessary to change the diagnoses of the genera according to the criterion accepted above.
There can be a question of the validity of such a criterion at the generic level, since it was originally used mostly at the specific level. Unfortunately, living species do not have an euspondyl arrangement of laterals, neither do they have only the first order of laterals, so it is impossible to test this criteria using genetic tools. Researchers of fossil taxa are forced to use a diverse approach, so more pronounced differences are used on a generic level and less pronounced differences on a specific level, also trying to include evolutionary trends. What is important here is that the presence of both single and double whorls, or a transition form between these two types, has not been recorded in any investigated taxon, so these genera are clearly distinguished. This means that the presence of double whorls can be considered important at the genus level. The asumed presence of two close but distinct whorls, oposite to the possibilities of existence of one whorl with different inclined laterals (heteroclinal) or of one whorl with "crowded" pores seemingly arranged in double whorls, is well proved in many illustrations of type species and other species of investigated genera, which will be pointed out in the respective chapters.
|
|
Figure 1: Distinguishing between the genera Physoporella and Oligoporella. 1-2. Physoporella; 3-4. Oligoporella. 1, 3 - longitudinal sections; 2, 4 - tangential sections. |
4.2. Emendation of genera
Physoporella Steinmann, 1903, emend.
Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in single whorls.
Type species: Physoporella pauciforata (G�mbel, 1872), Steinmann, 1903.
Oligoporella Pia, 1912, emend.
Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in double whorls. Each double whorl consists of two very close rows of laterals.
Type species: Oligoporella pilosa Pia, 1912.
4.3. Taxonomy of species and varieties
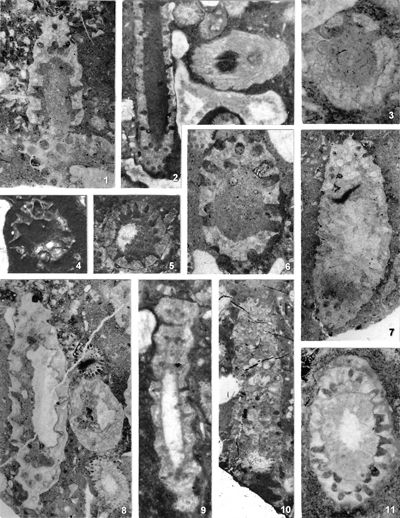
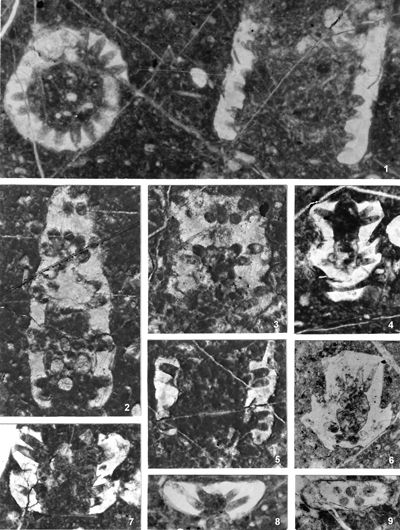
In accordance with the changed diagnoses of the genera, it is necessary to further re-examine the taxonomic affiliation of all species and varieties described so far. I believe that only such a complete analysis can solve the problem of these two genera, because by processing only some taxa, the problem would again remain for some future researchers. The criteria for the taxonomic classification of species and varieties, however, have not been uniform and there are significant differences from case to case. The differences in the previous criteria are evident if, for example, we compare almost identical specimens in Pl. 3 ![]() , fig. 2, described as Physoporella pauciforata (Pia, 1912) with that in Pl. 3
, fig. 2, described as Physoporella pauciforata (Pia, 1912) with that in Pl. 3 ![]() , fig. 1, described as Oligoporella prisca (Pia, 1912). Differences exists also in the quality of described material, if we compare Oligoporella pilosa var. physoporelloidea (Pl.
17
, fig. 1, described as Oligoporella prisca (Pia, 1912). Differences exists also in the quality of described material, if we compare Oligoporella pilosa var. physoporelloidea (Pl.
17 ![]() , fig. 9), described in only one cross-section, with Oligoporella pilosa var. pilosa (Pl. 5
, fig. 9), described in only one cross-section, with Oligoporella pilosa var. pilosa (Pl. 5 ![]() , figs. 1-6,
8-11; Pl. 6
, figs. 1-6,
8-11; Pl. 6 ![]() , figs. 1-4), described on more than, 20 specimens. Or, if we compare variety O. pilosa var. pilosa with O. pilosa var. intusannulata which is very similar, or with variety O. pilosa var. varicans whose shape of laterals differs significantly (compare reconstructions in Pia, 1935a, Figs. 15 and
28; this work, Figs. 9
, figs. 1-4), described on more than, 20 specimens. Or, if we compare variety O. pilosa var. pilosa with O. pilosa var. intusannulata which is very similar, or with variety O. pilosa var. varicans whose shape of laterals differs significantly (compare reconstructions in Pia, 1935a, Figs. 15 and
28; this work, Figs. 9 ![]() ,
12
,
12 ![]() ). In this revision, I have tried to adhere to uniform taxonomic criteria as far as possible and to avoid the dilemmas previously illustrated, and to simplify the determination of fossil material, even to those for whom fossil algae are not of primary interest.
). In this revision, I have tried to adhere to uniform taxonomic criteria as far as possible and to avoid the dilemmas previously illustrated, and to simplify the determination of fossil material, even to those for whom fossil algae are not of primary interest.
Another issue that arose during the work was the maintaining or abandonment of varieties established within some species. In today's systematics of Dasycladales, varieties are no longer used, and even in very similar forms, species are being established. It could be said that varieties are "no longer fashionable". In this paper, some varieties with clearly different characteristics were converted into species, while others still retained the same rank. I was guided in my decision-making by the important fact that there is a continuous series of forms between individual varieties within the same species, and the named varieties represent only the extremely pronounced respective forms that most probably belonged to the same biological species. By possibly rising varieties to the rank of species, forms with characteristics between the two extremes could not be taxonomically defined, while in this case a clear definition of species affiliation is possible. Additional weight is given by the fact that we find different varieties together in the same thin-section, and sometimes we can even observe the whole transition series. More about that in the chapters on individual species.
4.4. Comparison with other genera
The genera Physoporella and Oligoporella have a unique piriform shape to laterals and can be easily distinguished from other genera. Several genera with euspondyl arrangement of unramified laterals have some similarities with the genus Physoporella. The genus Uragiella Pia, 1924 has laterals somewhat similar to the piriform ones, but with a characteristic asymmetrical extension of the central part of the lateral. The Upper Permian genus Nipponophysoporella Endo, 1959 has similar laterals widened in the base, although it may present the proximal part of later vesiculifer laterals, which cannot be distinguished due to the poor preservation of the material (only one species N. elegans is known). It differs in the claviform thallus with "neck" and "head", which is probably only an article of the moniliform thallus, similar to the widespread Permian genus Mizzia, and may be its synonym. The genus Poikiloporella (Pia, 1920), 1943, also has proximally dilated laterals, but they are also distally dilated into a bubble, so they look like bowling cones. The genus Ardeiporella n. gen. differs in a substantially different form of laterals with a narrow proximal part, a pronounced extension in the middle and an elongated distal part. It also differs by having a very thick calcareous thallus (see Chapter 5.4.). Other genera, which also have an euspondyl arrangement of laterals of the first order, differ in the shape of the laterals: the genus Euteutloporella De Castro, 1993, has thin trichoform laterals; Salpingoporella Pia, 1918, emend. Conrad et al., 1973, has a phloioform shape of laterals; Griphoporella Pia, 1915, emend. Barattolo et al., 1993, and Gyroporella G�mbel, 1872, emend. Benecke, 1876, have vesiculifer laterals. The genus Anisoporella Botteron, 1961, is similar to Oligoporella in having double whorls, but with vesiculifer types of laterals.
4.5. Suprageneric position
The genera Physoporella and Oligoporella show a number of common characteristics, especially the shape of the laterals, which indicates that they are undoubted related, and they must certainly be classified in the same suprageneric taxon. This was already done by the Pia (1912 and, 1920) by including them in the subtribe Macroporellinae, which is characterized by aspondyl or euspondyl laterals of the first order, as opposed to the metaspondyl Diploporinae within the same tribe Diploporeae. Bassoullet et al. (1979) established a new tribe Salpingoporelleae characterized by a cylindrical unbranched thallus and first-order laterals. Within it, the subtribe Salpingoporellinae with phloioform laterals and the subtribe Oligoporellinae with trichoform and piriform laterals are separated. Both genera were included in the subtribe Oligoporellinae by Deloffre (1988) and Berger and Kaever (1992) which I agree with. In contrary, Granier and Deloffre (1995, p. 71) included the genus Physoporella in Salpingoporellinae, which does not seem justified.
Here we come to the question of belonging to one of the families. Dividing the order Dasycladales into families, Deloffre (1987, 1988) within the family Dasycladaceae combines all euspondyl forms with the same type of laterals along its thallus. For reproduction, he states that it takes place either in the stem cell or in modified laterals. Modified laterals also include laterals that have been modified only by function (cladospore type) and laterals that have been modified both by the function and by the form (choristospore type) (Bassoullet et al., 1975, 1977). This taxonomic approach caused very heterogeneous forms to be found in the same family. In contrast, Berger and Kaever (1992) consider the above two types of laterals to be phylogenetically and taxonomically important, and based on that they separate these into two families: Dasycladaceae (choristospore) and Triploporellaceae (cladospore), included tribe Salpingoporelleae. Granier and Bucur (in Granier et al., 2013, p. 218) changed the diagnosis of family Triploporellaceae that now included endospore and cladospore forms.
4.6. Phylogeny
Insufficient knowledge of the phylogeny of Dasycladalean algae is often a consequence of inconsistent criteria in their systematics, which can be seen, for example, in the very diverse ideas about the phylogeny of investigated genera. The leading problem is the fact that some living Dasycladalean algae do not calcify at all (Berger & Kaever, 1992) and it was probably the same with some extinct forms; so certain links in the evolutionary chain remain unknown to us.
Following the ideas of Pia (1920), Physoporella and Oligoporella are considered to have evolved from the genus Macroporella during Anisian, and consequently they developed in one lineage to Actinoporella, and in another to Triploporella, and further from that into Cymopolia and Neomeris, and Bornetella and Acetabularia, respectively. Following Kamptner (1958), the Diploporeae are interpreted to have developed from the early Palaeozoic Rhabdoporella. The subtribes Diploporinae and Macroporellinae separated very early on. Within the Macroporellinae, Physoporella forms a blind branch, and Oligoporella develops along one line into Poikiloporella, and on another into Neogyroporella, Uragiella, Clypeina and Actinoporella. After Herak et al. (1977), during the Carboniferous, aspondyl forms (represented by Rhabdoporella), give rise to the evolutionary line of euspondyl forms with acroform and trichoform laterals represented by Nanopora, Beresella, Dvinella (with the latter two subsequently excluded from the Dasycladales), Teutloporella and Oligoporella. The genus Uragiella, characterized by pirifer, vesiculiform and phloioform laterals, developed from the Oligoporella line in the late Triassic. In the Cretaceous, forms with several orders of laterals separated from it, and are typified by the genus Neomeris. Following Berger and Kaever (1992), the Salpingoporelleae tribe separated from the Dasyporelleae tribe during the Silurian, with the Coniporelleae, Dissocladelleae, Triploporellaleae and Clypeinae developed from these later. The Oligoporellinae and Mizziineae were derived from Salpingoporellineae (Deloffre & Granier, 1991). They originate from Macroporellinea. Recent Dasycladaceae have their origins in the early Palaeozoic Primicorallineae, while the Acetabulariaceae have theirs in the Diploporaceae, formed from the Salpinoporellineae tribe. With Oligoporellinae and Mizziineae are blind branches.
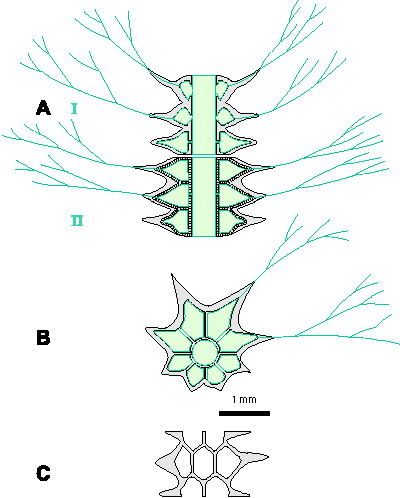
If we take into account the conclusions about the importance of the shape of the laterals from the previous chapter, we get a clearer picture of the evolutionary development of the studied genera (Fig. 2 ![]() ). Forms of the Oligoporellinae subtribe probably originated from aspondyl Teutloporellinae. There has thus been a simple expansion of the laterals and their arrangement into whorls, which is a clear evolutionary tendency in Dasycladales. This separation probably occurred somewhere in the late Palaeozoic, as evidenced by the finding of Physoporella? sp. from the Upper Carboniferous of Japan (Endo & Horiguchi, 1957, p. 172, Pl. XV, fig. 6). Looking evolutionary forward, the Oligoporellinae probably evolved in the direction of the family Dasycladaceae. This theory is supported by the fact that the early juvenile stage of the recent Neomeris annulata with its piriform laterals strongly resembles the fossil Physoporella (Cramer, 1891, Pl. I, fig. 2, copied in Pia, 1912, Pl. VIII, fig. 9, and Valet, 1968, Pl. 6, fig. 4.i), which was already observed by Pia (1912, p. 29, 41). This coincidence between ontogeny and phylogeny indicates a possible wider significance of the studied genera in the evolution of the Dasycladalean algae. The evolutionary sequence can be assumed from the genus Physoporella, through the genus Poikiloporella, which has distally extended laterals, and further in terms of creating second-order laterals into the Triploporelleae tribe and finally into the tribe Dasycladeae, i.e., the family Dasycladaceae (Fig. 2
). Forms of the Oligoporellinae subtribe probably originated from aspondyl Teutloporellinae. There has thus been a simple expansion of the laterals and their arrangement into whorls, which is a clear evolutionary tendency in Dasycladales. This separation probably occurred somewhere in the late Palaeozoic, as evidenced by the finding of Physoporella? sp. from the Upper Carboniferous of Japan (Endo & Horiguchi, 1957, p. 172, Pl. XV, fig. 6). Looking evolutionary forward, the Oligoporellinae probably evolved in the direction of the family Dasycladaceae. This theory is supported by the fact that the early juvenile stage of the recent Neomeris annulata with its piriform laterals strongly resembles the fossil Physoporella (Cramer, 1891, Pl. I, fig. 2, copied in Pia, 1912, Pl. VIII, fig. 9, and Valet, 1968, Pl. 6, fig. 4.i), which was already observed by Pia (1912, p. 29, 41). This coincidence between ontogeny and phylogeny indicates a possible wider significance of the studied genera in the evolution of the Dasycladalean algae. The evolutionary sequence can be assumed from the genus Physoporella, through the genus Poikiloporella, which has distally extended laterals, and further in terms of creating second-order laterals into the Triploporelleae tribe and finally into the tribe Dasycladeae, i.e., the family Dasycladaceae (Fig. 2 ![]() ) with choristospore reproductive organs. Polyphysaceae developed possibly from Dasycladaceae, i.e., by the widening and specialization of gametangia, that originated from second-order laterals, is obvious by studying the morphogenesis of recent species (Valet, 1968), and has not originated from the Salpingoporelleae tribe by further spreading and by specialization of first-order phloioform laterals.
) with choristospore reproductive organs. Polyphysaceae developed possibly from Dasycladaceae, i.e., by the widening and specialization of gametangia, that originated from second-order laterals, is obvious by studying the morphogenesis of recent species (Valet, 1968), and has not originated from the Salpingoporelleae tribe by further spreading and by specialization of first-order phloioform laterals.
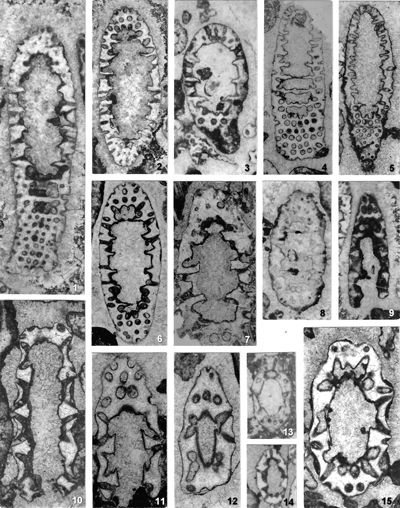
As for the phylogenetic development within the tribe Oligoporelleae, which includes the studied genera, it can be traced from the Early Permian, as the first certain Physoporella (P. chichibuensis Ishijima et al., P. kitakamiensis Endo, P. konishii Endo) and Oligoporella (O. fluctuosa (Endo)) have been found in the Asselian, and P. nipponica (Endo) in the Artinskian, while the species P. lativentrusa Endo, P. longipora (Endo) and P. nyugawensis Endo are found in the lower and middle Permian. Former species are quite similar to the Anisian ones, especially P. pauciforata, so it is very likely that it is a direct successor in the phylogenetic sense. The species O. fluctuosa differs somewhat from the typical species of the genus, but due to the insufficient material available does not allow a more detailed phylogenetic analysis: Is it the ancestor of later Oligoporellas, or did they later develop separately from Physoporellas by converging whorls into the double rows? Oligoporella laevis (Praturlon, 1963) appears in the uppermost layers of the Permian Bellerophon Formation in the Italian Dolomites. There are no finds of algae in the Lower Triassic, so it is not possible to follow the development of the studied genera, but they certainly survived in a protected environment that allowed Dasycladalean algae to live. From Physoporella, a peculiar branch of algae develops in the Illyrian that has slightly smaller and more rounded piriform laterals that are included in the new genus Neophysoporella (N. lotharingica, N. jomdaensis, N. zamparelliae and Neophysoporella n. sp.). The main "branch" of the genus Physoporella gave rise to two more "shoots". In the Fassanian P. leptotheca appears with typical piriform laterals, but sometimes located in groups of two or three. The genus Poikiloporella also probably evolved from the genus Physoporella in the early late Triassic by modifying the shape of the laterals, and the same is true for the genus Uragiella.
|
|
Figure 2: Phylogenetic development of the genera Physoporella, Oligoporella and related taxa: 1. Teutloporellinae (aspondyl trichoform laterals); 2. Oligoporella with double whorls of piriform laterals (O. fluctuosa, O. laevis, O. pilosa, O. intusannulata, O. cornuta, O. varicans, O. minutula, O. praealpina, O. dissita, O. chrzanowensis, O. polonoandalusica); 3. Typical Physoporella with single whorls of piriform laterals (P. chichibuensis, P. kitakamiensis, P. konishii, P. lativentrusa, P. longipora, P. nipponica, P. nyugawensis, P. pauciforata, P. croatica); 4. Ardeiporella n. gen. with laterals shaped like a heron's head; 5. P. leptotheca with grouped piriform laterals; 6. Neophysoporella n. gen. with single whorls of small rounded piriform laterals (N. lotharingica, N. zamparelliae, N. jomdaensis, Neophysoporella n. sp.); 7. Uragiella with asymmetrically widened laterals; 8. Poikiloporella with piriform laterals distally expanded in the vesicle; 9-10. Triploporelleae: 9.
Palaeodasycladus; 10. Triploporella; 11. Dasycladaceae (Cymopolia, Neomeris); 12) Polyphysaceae (Halicoryne). |
5.1. Genus Physoporella Steinmann, 1903, emend.
Phylum Chlorophyta
Class Ulvophyceae Mattox & Stewart, 1978
Order Dasycladales Pascher, 1931
Family Triploporellaceae Berger & Kaever, 1992, emend. Granier & Bucur in Granier et al., 2013
Tribe Salpingoporelleae Bassoullet et al., 1979
Subtribe Oligoporellinae Bassoullet et al., 1979
Genus Physoporella Steinmann, 1903, emend.
5.1.1. Physoporella pauciforata (G�mbel, 1872), Pia, 1912
(= Gyroporella pauciforata G�mbel, 1872)
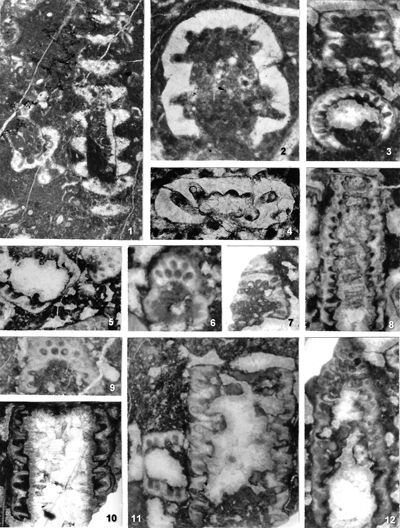
Pl. 1 ![]() ; Pl. 2
; Pl. 2 ![]() , figs. 1-4, 7-11; Pl. 4
, figs. 1-4, 7-11; Pl. 4 ![]() , fig. 3; Pl. 16
, fig. 3; Pl. 16 ![]() , figs. 1, 6, 8-9; Pl. 18
, figs. 1, 6, 8-9; Pl. 18
Origin of the name: Probably after relatively small number of pores (Latin "paucus" = scarce, few, rare; and "foramen" = hole).
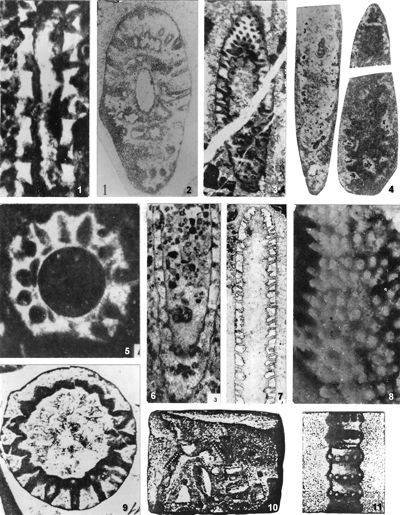
Studied material: G�mbel (1872, p. 274-275) cites "black 'Reiflinger Kalk' from Reisalp (collection Geologische Reichsanstalt, Wien) and black limestone from Pertisau in Tyrol (shipment from Professor Pichler)" as typical strata and localities for G. pauciforata, but does not state which specimen is from which locality (G�mbel, 1872, Pl. D.III, fig. 2.a-f). G�mbel's material has been lost (Pia, 1912, p. 26; 1935a, p. 223). Lein (1993) states that the material used by G�mbel from Reisalp (east of T�rnitz, southwest of Vienna) was collected by the "Geologische Reichsanstalt" during detailed research related to coal prospecting in the, 1850s. Since the exact location of G. pauciforata G�mbel, 1872 (also G. minutula G�mbel, 1872 and O. prisca Pia, 1912 are from the same type area) is unknown, Lein (1993) cites the quarry SSE of Kandlhof, north of Furthof with a dark Anisian (? Upper Pelsonian) "Gutenstein Limestone" as the best algae-bearing outcrop in the type area of Reisalp. The samples I collected there during the field trip "Alpine Algae '93" contain P. pauciforata pauciforata, P. pauciforata sulcata and O. pilosa intusannulata. Although algae appear abundant at this locality, they are rather poorly preserved.
Kotański (2013, p. 68) chose as "holotype" of P. pauciforata the specimen figured by G�mbel (1872, Pl. D.III, fig. 2b). It is not valid according to rules of botanical nomenclature after Art. 9.3. of ICN (Turland et al., 2018) since a subsequent author can only chose lectotype among the original material, not holotype, and also after Art. 40.4 (see also Ex. 5), since the type of a name of a new species on or after 1 January, 2007 cannot be an illustration; the type must be a specimen. G�mbel's figure is an illustration, it is not certain that it represents a real specimen completely, the number in the collection is unidentified, and it is known that his collection has been lost (Pia, 1912, p. 26; 1935a, p. 223). Also, the specimen from G�mbel (1872, Pl. D.III, fig. 2.e) is not the only syntype, since according to Art. 9.6. a syntype is any specimen cited in the protologue when there is no holotype.
The material of P. pauciforata described by the Pia (1912, p. 44), originates from several localities: samples XXI from N Brandm�uer, W T�rnitz (Austria), XXII from Ne�linger Wand, SW Kitzb�hel (Austria), XXVIII from Schwarzenbach an der Pielach, W T�rnitz (Austria) and LIV from a secondary sample from Sarlkofel (Monte Serla) in the Dolomites (Italy). Since Pia (1935a, p. 223) identified specimens from Schwarzenberg as type material (Pia, 1912, Pl. V, figs. 13-16), among them I choose one from the Pia (1912, Pl. V, fig. 13 = this work, Pl. 1 ![]() , fig. 1) and state it as a neotype, since it has the best visible piriform laterals and is the best preserved specimen. According to Lein (1993), the Schwarzenberg site forms a part of some steep cliffs between the peaks of Schwarzenberg (1096 m) and Schl�gelberg (1057 m, see also Piros et al., 1994, p. 345) and was built from the Anisian Steinalmkalk, and not from Reiflingkalk, as originally thought by Pia (1912). In Pia's collection, I could not find thin-section 1036 with P. pauciforata as described in Pia (1920, p. 51-52; Pl. III, figs. 10, 15), which originates from a secondary sample of Mendoladolomites from the Puster Valley (Italy).
, fig. 1) and state it as a neotype, since it has the best visible piriform laterals and is the best preserved specimen. According to Lein (1993), the Schwarzenberg site forms a part of some steep cliffs between the peaks of Schwarzenberg (1096 m) and Schl�gelberg (1057 m, see also Piros et al., 1994, p. 345) and was built from the Anisian Steinalmkalk, and not from Reiflingkalk, as originally thought by Pia (1912). In Pia's collection, I could not find thin-section 1036 with P. pauciforata as described in Pia (1920, p. 51-52; Pl. III, figs. 10, 15), which originates from a secondary sample of Mendoladolomites from the Puster Valley (Italy).
Material from Croatia includes specimens from the Anisian limestone from Belski dol quarry, Ivan�čica Mt (Pl. 18 ![]() , figs. 1-4; Pl. 19
, figs. 1-4; Pl. 19 ![]() , fig.
6).
, fig.
6).
History of investigations: G�mbel (1872, p. 274) establishes a new species with the following diagnosis: "Gyroporella pauciforata n. sp. The shell is cylindrical-tubular in shape, often slightly curved, small in size with distinct ring articles and two rows of less numerous (10-12) and very wide canals in each of them (Pl. D III, fig. 2.a, 2.c) The surface is clearly annular due to a slightly protruding swelling and covered with pits due to the openings of the canals. Tubes have a diameter of 2-2.5 mm; and a ring height of 0.6-0.7 mm. So far only specimens are known to be enclosed in the dense limestone." As can be seen, G�mbel (1872) based his description primarily on dimensions (number of canals and rings), and did not pay attention to the structure of the whorls and the shape of the laterals, which is not surprising when it is known that he considered these fossils foraminifers.
(In German: "Gehause cylindrisch-rohrenformig, oft etwas gebogen, von geringer Grosse mit deutlichen Ringgliederen und zwei Reihen wenig zahlreicher und ser weiter Kanalchen in jedem derselben. Die Oberflache ist durch etwas vorstehende Wulste deutlich geringelt und durch die Ausmundungsoffnungen der Kanalchen mit Grubchen bedeckt. Im Durchmesser haben die Rohrchen 2-2.5 mm, in der Ringhohe 0.6-0.7 mm. Bisher sind nur Exemplare im dichten Kalk eingeschlossen bekant.")
Pia (1912, p. 44; see also Granier & Sander, 2013, p. 33) gives a revision of this species: "Physoporella pauciforata G�mbel spec. The calcareous skeleton is cylindrical and linearly prolate without any marked segmentation. The lateral branches occur in true, simple and often very regular verticils. They are egg- or pear-shaped, more or less elongated and taper outward. Mostly they are completely enclosed in lime. Occasionally, however, a tapered pore perforates the skeleton (see Pl. V, figs. 11, 14). Since this occurs only exceptionally and, it appears, involves only single pores of a plant, I do not consider that much importance should be attached to it. The openings can scarcely be considered as passageways for the filaments, but only as accidental openings that would disappear if the calcareous layer were thicker. The angle of the verticillated branches and the distance between the verticils is subject to great variation. As a whole our species displays the essential attributes of the genus Physoporella without specific complications. Thus it may pass for the type of the genus." He comments on the shortcomings of G�mbel's (1872) description, which is based primarily on the dimensions and external morphology of the shell (Pia, 1912, p. 69): "G�mbel's mistakes are in large part understandable when one looks at the thin sections that he must have studied. Their poor quality obviously made it impossible for him to examine the form of the single pores, to which, in my opinion, so much importance must be given." Pia (1935a, p. 223) also writes about this: "The starting point of the whole research should naturally be the knowledge of the type of Physoporella pauciforata. Unfortunately, from G�mbel's descriptions and paintings (1872, p. 274, tab. D III, fig. 2.a-e), as usual, it is not possible to understand what his missing originals actually looked like." From the quoted parts of Pia's text, it is evident that in his work he repeatedly faces the problems of recognizing heterogeneous and poorly described G�mbel material, especially because his material is lost, which Pia witnessed after his trip to Munich especially for this purpose. As the already more experienced Pia researcher (1931, p. 272) states: "I now regret that I have reintroduced some of these names into scientific papers on the basis of insufficient support." As one can see, drawings submitted by G�mbel do not provide insight into the structure of algae and successfully differentiate between species, and they also seem to contain a lot of "interpretation," unlike Pia's drawings which show almost no deviation from the photographs (compare Pia, 1912, 1935a with this work). In many cases, we cannot be sure that all the specimens shown on the plate belong to the same species, especially because they come from two different localities, and it is not stated which specimen is from which locality. Since G�mbel's material has disappeared, there are no nomenclatural problems to define Pia's material as type, i.e., neotype material, and thus Pia's description becomes the only valid one.
Pia (1920, p. 51) completes the description of this species: "There is good new material of this stratigraphically important species, which allows the description to be supplemented on a few points. The whorls are not always very simple, but sometimes greatly packed. In the samples from the Steiner Alps, the calcification often extends to the stem cell, as is undoubtedly evident from the very even and smooth inner surface of the skeleton. It was therefore possible to obtain data for the diameter of the central cell. Their thickness is 31 to 44% of D. The shape of the verticiled branches is quite variable, sometimes they are noticeably slender, as in of my previous work (Pia, 1912, Pl. 5, fig. 9). The shell of Physoporella pauciforata lacks a real, externally recognizable, articulation. However, it can often be observed that the calcareous tubes break through on strikingly smooth surfaces. Occasionally, rather sharply delimited, dark transverse stripes run through the skeleton between the individual whorls, which in any case correspond to partially open fissures. We will get to know this fissuration in the next species in an even better way. Some cross-sections in the thin-section from the Mendola dolomite from the Sarlkofel show very strange holes in the shell. The outer part of the skeleton is completely filled with open fissures which cut diagonally at an angle of about 30� to the stem cell. As a result, the skeleton dissolves towards the outside into a series of interlocking, irregular cones, occasionally connected by calcareous bridges. It seems that these cross-sections belong to several specimens. The sculpture described is to be understood as a deformity with no functional meaning (cf. Pia, 1912, Pl. 3, figs. 10, 15)."
(In German: "Von dieser stratigraphisch wichtigen Arte liegt gutes, neues Material vor, das gestattet, die Beschreibung in einigen Punkten zu erg�nzen. Die Wirtel sind nicht immer ganz einfach, sondern gelegentlich massig gedr�ngt. Die Verkalkung reicht in den Handstucken aus den Steiner Alpen �fter bis an die Stammzelle, wie sich aus der ganz ebenen und glatten Innenflache des Skelettes unzweifelhaft ergibt. Es konnten daher f�r den Durchmesser der Zentralzelle Daten gewonnen werden. Ihre Dicke betr�gt 31 bis 44 % von D. Die Form der Wirtelaste ist ziemlich variabel, manchmal sind sie auffallend schlank, wie schon auf der Abbildung Pl. 5, fig. 9 meiner fr�heren Arbeit zu sehen ist. Eine echte, ausserlich wahrnehmbare Gliederung fehlt der Schale von Physoporella pauciforata. Es ist aber h�ufig zu beobachten, dass die Kalkrohrchen an auffallend glatten Fl�chen streng quer durchbrechen. gelegentlich verlaufen durch das Skelett zwischen den einzelnen Wirteln ziemlich scharf begrenzte, dunkle Querstreifen, die jedenfalls teilweise offenen Spalten entsprechen. Wir werden diese Fissuration bei der n�chsten Art in noch besserer Entwicklung kennen lernen. Sehr merkw�rdige L�cken in der Schale zeigen einige Schnitte in einem Schliff von Mendola-Dolomit des Sarlkofels. der �u�ere Teil des Skelettes ist ganz erf�llt von offenen Spalten, die sehr schrag, unter einem Winkel von etwa 30o zur Stammzelle, einschneiden. Das Skelett l�st sich dadurch gegen au�en in eine Serie ineinander steckender, unregelm��iger, hie und da durch Kalkbr�cken verbundener D�ten auf. es scheint, dass die betreffenden Schnitte zu mehreren Exemplaren geh�ren. Die beschriebene Skulptur ist wohl als eine Missbildung ohne funktionelle Bedeutung aufzufassen (vgl. Pl. 3, figs. 10, 15).")
Schlagintweit et al. (2003) publised findings of the species P. pauciforata from the Guttenstein Limestone of Salzkammergut, Austria. Kotański (2013, p. 66) gives a diagnosis on the base of isolated specimens from the Upper Silesia (Poland): "Cylindrical thallus without segmentation. Only very little undulation may be observable. The branches (tubercles) are distributed in single whorls. They are pyriform, mostly closed within the calcareous wall (if present), oriented perpendicularly to the longitudinal axis. Whorls are rather closely spaced. It is a model representative of Physoporella pauciforata group (one row of pores per one whorl)." It is not easy to compare his material with that from the thin-section, but the illustrated specimens, judging by the shape and arrangement of the branches, as well as by other characteristics, indicate positively that they belong to the species described.
Bucur and Matysik (2020) described specimens from thin sections, found also in the Upper Silesia. They are very similar to the typical specimens of P. pauciforata, although of smaller dimensions, and in my opinion belong to this species.
Physoporella pauciforata var. pauciforata Pia ex Bystrick�, 1964
(= Physoporella pauciforata var. simplex Pia, 1935a, nom. nud.)
Pl. 1 ![]() , figs. 1-10, 13, 15; Pl. 16
, figs. 1-10, 13, 15; Pl. 16 ![]() , fig. 8; Pl. 18
, fig. 8; Pl. 18 ![]() , figs. 1-4
, figs. 1-4
1872 - Gyroporella pauciforata n. sp. - G�mbel, p. 274-275, Pl. DIII, figs. 2a-f.
1903 - Physoporella pauciforata n. comb. - Steinmann, p. 50, no figure.
1912 - Physoporella pauciforata - Pia, p. 44, 51-53, Pl. V, figs. 9-11, 13-17, 19.
1920 - Physoporella pauciforata - Pia, p. 51-52, Pl. III, figs. 10, 15.
1935a (nom. nud.) - Physoporella pauciforata var. simplex n. var. - Pia, p. 221-223, no figure.
1957 (nom. nud.) - Physoporella pauciforata simplex - Bystrick�, p. 232, Pl. VI, fig. 1-2.
1958 - Physoporella pauciforata - Herak, p. 59, 61, Pl. 13, figs. 1-5.
1964 - Physoporella pauciforata var. pauciforata n. var.- Bystrick�, p. 118-121, Pl. XIII, figs. 1-6; Pl. XIV, figs. 3-6; Pl. XV, fig. 6; Pl. XXXIV, fig. 2 pars.
1965 - Physoporella pauciforata - Herak, p. 16-17, Pl. IX, fig. 1.b.
1965 (nom. nud.) - Physoporella pauciforata var. simplex - Herak, Pl. XIII, fig. 5.
1989 - Physoporella pauciforata var. pauciforata - Buček, p. 134, Pl. II, fig. 1.
1993 - Physoporella pauciforata pauciforata - Budai et al., Pl. V, fig. 2.
1994 - Physoporella pauciforata - Bucur et al., Pl. 11, figs. 8-9.
2003 - Physoporella pauciforata var. pauciforata - Piros, p. 54-55, Pl. D-II, figs. 2-4; Pl. D-IV, fig. 2.
2003 - Physoporella pauciforata.- Schlagintweit et al., p. 607-608, Pl. 1, figs. 1-10
2013 - Physoporella pauciforata var. pauciforata - Kotański, p. 66-69, Pl. XI, figs. 1-25; Pl. XV, figs. 22-23; Pl. XVIII, figs. 4, 68.b; Pl. XIX, fig. 6; Pl. XX, figs. 5, 7, 11, 13.b; Pl. XXXII, figs. 2.b, 3.a, 4.a; Pl. XXXIII, figs. 1.b, 2.b, 3 pars, 4.a; Pl. XXXIV, figs. 1.a, 2 pars, 3.b, 4.b.
2020 - Physoporella cf. pauciforata - Bucur & Matysik, p. 10, Fig. 6H-O.
2021 - Physoporella pauciforata - Gawlick et al., Pl. 5, figs. b-g, k, o.
2021 - Physoporella cf. prisca - Gawlick et al., Pl. 5, fig. h.
2022 - Physoporella pauciforata - Karádi et al., Fig. 8B.
Pia (1935a, p. 221-223) introduces different varieties within the species Physoporella pauciforata and names Physoporella pauciforata simplex: "I have already indicated on an earlier occasion (Pia, 1925, p. 331-332) that the species Physoporella pauciforata is very broad. It will be useful to divide it into varieties and to separate some forms entirely from it. The first step has already been made with the distinction between the var. lotharingica and the separation of Physoporella praealpina... As a starting point for further discussions, I choose specimens from Schwarzenberg west of T�rnitz, adjacent to Reisalp, on which my first description of the species is based (Pia, 1912, Pl. V, figs. 13-16). They usually have strictly single-row whorls and a cylindrical, not noticeably articulated shell. The pores are covered on the outside by a rather thick calcareous layer. The diameter of the shell is about 3 mm. I will refer to this form as Physoporella pauciforata simplex."
(In German: "Ich habe schon bei fr�herer Gelegenheit (Pia, 1925, p. 331-332) angedeutet, dass die Art Physoporella pauciforata sehr weit gefasst ist. es wird zweckmassig sein, sie in Variet�ten zu teilen und manche Formen ganz von ihr abzutrennen. Ein Anfang dazu ist durch die Unterscheidung der var. lotharingica und durch die Abtrennung der praealpina schon gemacht� Als Ausgangspunkt der weiteren Er�rterungen w�hle ich aber die St�cke von dem der Reisalpe, benachbarten Schwarzenberg westlich T�rnitz, die meiner ersten Beschreibung der Art zugrunde liegen (Pia). Sie haben in der Regel streng einreihige Wirtel und eine zylindrische, nicht merklich gegliederte Schale. die Poren sind aussen durch eine recht dicke Kalklage �berdeckt. Der Durchmesser der Schale betr�gt etwa 3 mm. Ich werde diese Form als Physoporella pauciforata simplex bezeichnen.")
Bystrick� (1964, p. 118) gives a new name to the type variety, in accordance with the rules of botanical nomenclature (Art. 26.1 ICN, Turland et al., 2018) and revises the diagnosis of Physoporella pauciforata var. pauciforata nom. nov.: "The tubular thallus with a smooth or moderately wavy outer surface is divided into differently high articles by narrow cracks (fissures) which are laid perpendicular to the longitudinal axis. The pores are pear-shaped; widest is the basal part and thinning towards the outer surface such that the end of the pore does not protrude from the shell (the pores are closed). The pores are arranged in single-rowed whorls at regular distances, and also perpendicular to the longitudinal axis."
(In Slovak: "Trubickovit� schr�nka s rovn�m, alebo mierne zvlnen�m vonkaj��m povrchom, je rozdelen� na r�zne vysok� cl�nky iba �zkymi �trbinkami (fisuratio), prebiehaj�cimi kolmo na pozdl�nu os stielky. P�ry maj� hru�kovit� tvar; ich najhrub�ia baz�lna cast sa k vonkaj�iemu povrchu schr�nky stencuje, av�ak dist�lny koniec p�rov nevycnieva zo schr�nky (p�ry zatvoren�). P�ry s� zoraden� do jednoradov�ch praslenov, takmer kolm�ch na pozdl�nu os stielky, v pravideln�ch vzdialenostiach.")
Description: Thallus has a cylindrical shape, mostly straight or only very slightly curved (Pl. 1 ![]() , fig. 15; Pl. 18
, fig. 15; Pl. 18 ![]() , figs. 1-2, 4). The inner surface of the calcareous thallus is usually even (Pl. 18
, figs. 1-2, 4). The inner surface of the calcareous thallus is usually even (Pl. 18 ![]() , figs. 1-4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, figs. 1-2; Schlagintweit et al., 2003, Pl. 1, fig. 5; Gawlick et al., 2021, Pl. 5, figs. e-f), and may be less (Pl. 1
, figs. 1-4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, figs. 1-2; Schlagintweit et al., 2003, Pl. 1, fig. 5; Gawlick et al., 2021, Pl. 5, figs. e-f), and may be less (Pl. 1 ![]() , fig. 15) or more wrinkled (Bystrick�, 1964, Tab. XIII, figs. 5-6; Schlagintweit et al., 2003, Pl. 1, fig. 2), which is a reflection of the wrinkling of the stem cell itself. Calcification generally does not reach the stem cell itself (except for rare exceptions, such as in Pl. 19
, fig. 15) or more wrinkled (Bystrick�, 1964, Tab. XIII, figs. 5-6; Schlagintweit et al., 2003, Pl. 1, fig. 2), which is a reflection of the wrinkling of the stem cell itself. Calcification generally does not reach the stem cell itself (except for rare exceptions, such as in Pl. 19 ![]() , fig. 6), but to a very short distance from it, which suggests incompleteness of the lateral pores, e.g., the basal part is missing (Pl. 1
, fig. 6), but to a very short distance from it, which suggests incompleteness of the lateral pores, e.g., the basal part is missing (Pl. 1 ![]() , figs. 1-2, 15; Pl. 18
, figs. 1-2, 15; Pl. 18 ![]() ,
figs. 1-4). Such an interpretation is also given by Pia (1912, fig. 16; Pia in Diener, 1925, Pl. XXVII, fig. 2.a-b) in the reconstruction of P. pauciforata. Rarely calcification affects only more distal parts of the laterals (Pl. 1
,
figs. 1-4). Such an interpretation is also given by Pia (1912, fig. 16; Pia in Diener, 1925, Pl. XXVII, fig. 2.a-b) in the reconstruction of P. pauciforata. Rarely calcification affects only more distal parts of the laterals (Pl. 1 ![]() , fig. 5; Bystrick�, 1964, Pl. XIV, fig. 3; Herak, 1965, Pl. XIII, fig. 5; Bucur et al., 1994, Pl. 11, figs. 9, 11). Where there has been a subsequent dissolution of the shell, its inner surface is uneven (Pl. 1
, fig. 5; Bystrick�, 1964, Pl. XIV, fig. 3; Herak, 1965, Pl. XIII, fig. 5; Bucur et al., 1994, Pl. 11, figs. 9, 11). Where there has been a subsequent dissolution of the shell, its inner surface is uneven (Pl. 1 ![]() , fig. 3; Pl. 2
, fig. 3; Pl. 2 ![]() , fig. 9; Bystrick�, 1964, Pl. XIII, fig. 3; Herak, 1958, Pl. 13, figs. 1-5). The outer surface of the calcareous thallus is never completely flat, as shown in the reconstruction by Pia in Diener (1925, Pl. XXVII, fig. 2.a-c), but is usually more or less uneven. It is represented mainly by a slight undulation, so that the protruding parts are at the level of the whorls, and the depressions between them (Pl. 1
, fig. 9; Bystrick�, 1964, Pl. XIII, fig. 3; Herak, 1958, Pl. 13, figs. 1-5). The outer surface of the calcareous thallus is never completely flat, as shown in the reconstruction by Pia in Diener (1925, Pl. XXVII, fig. 2.a-c), but is usually more or less uneven. It is represented mainly by a slight undulation, so that the protruding parts are at the level of the whorls, and the depressions between them (Pl. 1 ![]() , figs. 4-5; Pl. 18
, figs. 4-5; Pl. 18 ![]() , figs. 1-4; Bystrick�, 1957, Pl. VI, fig. 1, 1964, Pl. XIII, figs. 1-5; Bucur et al., 1994, Pl. 11, figs. 9, 11; Schlagintweit et al., 2003, Pl. 1, fig. 2). In the specimens illustrated by Pia (1920, Pl. III, figs. 10, 15), narrow cavities are observed laid obliquely to the longitudinal axis and arranged concentrically around the stem cell. Pia (1920, p. 51, 165) considers this phenomenon as "abnormal". Bystrick� (1964, p. 119; Pl. XIV, figs. 3-4) finds also in his material such an alteration of calcareous lamellae separated by cracks, interpreting them as changes in skeletal secretion caused by periodic changes in the environment. These "cracks" are not an exceptional occurrence in P. pauciforata, as the mentioned authors write, but they occur regularly and are a reflection of the biological structure of the alga (Fig. 3
, figs. 1-4; Bystrick�, 1957, Pl. VI, fig. 1, 1964, Pl. XIII, figs. 1-5; Bucur et al., 1994, Pl. 11, figs. 9, 11; Schlagintweit et al., 2003, Pl. 1, fig. 2). In the specimens illustrated by Pia (1920, Pl. III, figs. 10, 15), narrow cavities are observed laid obliquely to the longitudinal axis and arranged concentrically around the stem cell. Pia (1920, p. 51, 165) considers this phenomenon as "abnormal". Bystrick� (1964, p. 119; Pl. XIV, figs. 3-4) finds also in his material such an alteration of calcareous lamellae separated by cracks, interpreting them as changes in skeletal secretion caused by periodic changes in the environment. These "cracks" are not an exceptional occurrence in P. pauciforata, as the mentioned authors write, but they occur regularly and are a reflection of the biological structure of the alga (Fig. 3 ![]() ). These represent calcified trichoblasts that continue from the apex of piriform
laterals. Pia (1912), despite the similarity of shape, considered the pores of the laterals of the genus Physoporella to be closed and those of the genus Oligoporella to be open, and assumed that only the branch of Oligoporella continued into a thin trichoblast, based on comparison with the laterals of the recent Neomeris annulata (Pia, 1912, p. 29, 41; Pl. VIII, fig. 9). However, in the type species of the genus Physoporella these trichoblasts are often fossillized. Each lateral carries a trichoblast directed upwards at its apex, i.e., towards the light. There is a possibility that trichoblasts occurred in clusters of two or perhaps more, as suggested by illustration in Hurka (1969, fig. 7.6). Depending on the intensity of calcification, as well as the subsequent erosion of the skeleton, calcified trichoblasts are recognized as longer or shorter upward-pointing spikes (Pl. 1
). These represent calcified trichoblasts that continue from the apex of piriform
laterals. Pia (1912), despite the similarity of shape, considered the pores of the laterals of the genus Physoporella to be closed and those of the genus Oligoporella to be open, and assumed that only the branch of Oligoporella continued into a thin trichoblast, based on comparison with the laterals of the recent Neomeris annulata (Pia, 1912, p. 29, 41; Pl. VIII, fig. 9). However, in the type species of the genus Physoporella these trichoblasts are often fossillized. Each lateral carries a trichoblast directed upwards at its apex, i.e., towards the light. There is a possibility that trichoblasts occurred in clusters of two or perhaps more, as suggested by illustration in Hurka (1969, fig. 7.6). Depending on the intensity of calcification, as well as the subsequent erosion of the skeleton, calcified trichoblasts are recognized as longer or shorter upward-pointing spikes (Pl. 1 ![]() , figs. 13, 15 right; Pl. 2
, figs. 13, 15 right; Pl. 2 ![]() , fig. 7; Pl. 18
, fig. 7; Pl. 18 ![]() , figs. 1, 2 above; Bystrick�, 1964, Pl. XIII, fig. 4 = this paper, Pl. 16
, figs. 1, 2 above; Bystrick�, 1964, Pl. XIII, fig. 4 = this paper, Pl. 16 ![]() , fig. 8; Schlagintweit et al., 2003, Pl. 1, figs. 2, 10). In the case of more intensive calcification, much longer parts of the trichoblasts are fossilized, so that in longitudinal and longitudinal-oblique sections they are recognized as long curved spikes (Pl. 1
, fig. 8; Schlagintweit et al., 2003, Pl. 1, figs. 2, 10). In the case of more intensive calcification, much longer parts of the trichoblasts are fossilized, so that in longitudinal and longitudinal-oblique sections they are recognized as long curved spikes (Pl. 1 ![]() , fig. 15; Pl. 18
, fig. 15; Pl. 18 ![]() , fig. 1; Buček, 1989, Pl. II, fig. 1), and in transverse and transverse-oblique cross-sections as concentric calcified lamellae (Pl. 1
, fig. 1; Buček, 1989, Pl. II, fig. 1), and in transverse and transverse-oblique cross-sections as concentric calcified lamellae (Pl. 1 ![]() , figs. 1, 4-6; Pl. 2
, figs. 1, 4-6; Pl. 2 ![]() , fig. 7; Pia, 1920, Pl. III, figs. 10, 15; Herak, 1958, Pl. 13, fig. 5; Bystrick�, 1964, Pl. XIV, figs. 3-4; Herak, 1965, Pl. XIII, fig. 5; Schlagintweit et al., 2003, Pl. 1, figs. 1, 4, 6, 8-9). These lamellae were formed by calcification of the proximal parts of close trichoblasts and their fusion. Similarly, close and very thin laterals of the genus Cambelliella calcify, forming funnel-shaped articles (De Castro, 1993). In the case of intensive calcification and good preservation, in cross or oblique sections, several lamellae can be seen, which represents calcified trichoblasts of several lower whorls (Fig. 3
, fig. 7; Pia, 1920, Pl. III, figs. 10, 15; Herak, 1958, Pl. 13, fig. 5; Bystrick�, 1964, Pl. XIV, figs. 3-4; Herak, 1965, Pl. XIII, fig. 5; Schlagintweit et al., 2003, Pl. 1, figs. 1, 4, 6, 8-9). These lamellae were formed by calcification of the proximal parts of close trichoblasts and their fusion. Similarly, close and very thin laterals of the genus Cambelliella calcify, forming funnel-shaped articles (De Castro, 1993). In the case of intensive calcification and good preservation, in cross or oblique sections, several lamellae can be seen, which represents calcified trichoblasts of several lower whorls (Fig. 3 ![]() ; Pia, 1920, Pl. III, figs. 10, 15; Bystrick�, 1964, Pl. XIV, figs. 3-4), while in the case of weaker calcification and/or poorer preservation, only one lamella is seen (Pl. 1
; Pia, 1920, Pl. III, figs. 10, 15; Bystrick�, 1964, Pl. XIV, figs. 3-4), while in the case of weaker calcification and/or poorer preservation, only one lamella is seen (Pl. 1 ![]() , figs. 1, 4-6; Pl. 2
, figs. 1, 4-6; Pl. 2 ![]() , fig. 7). Some specimens also show dichotomous
branching (Pl. 1
, fig. 7). Some specimens also show dichotomous
branching (Pl. 1 ![]() , fig. 15 above right; Bystrick�, 1964, Pl. XIV, fig.
3), characteristic of trichoblasts of recent species (Berger & Kaever, 1992). In many specimens, either because the calcareous spikes are eroded or because the trichoblasts have fallen off earlier, the outer surface of the thallus has no spikes, but is only slightly undulated. In recent species of Dasycladales after the vegetative phase, and even earlier, trichoblasts generally fall off before calcification, which is the most likely the case with the fossil species. The calcification of trichoblasts described here can therefore be considered exceptional. Besides already mentioned slight undulation, thin cracks often appear between the whorls (Pl. 1
, fig. 15 above right; Bystrick�, 1964, Pl. XIV, fig.
3), characteristic of trichoblasts of recent species (Berger & Kaever, 1992). In many specimens, either because the calcareous spikes are eroded or because the trichoblasts have fallen off earlier, the outer surface of the thallus has no spikes, but is only slightly undulated. In recent species of Dasycladales after the vegetative phase, and even earlier, trichoblasts generally fall off before calcification, which is the most likely the case with the fossil species. The calcification of trichoblasts described here can therefore be considered exceptional. Besides already mentioned slight undulation, thin cracks often appear between the whorls (Pl. 1 ![]() , fig. 4; Bystrick�, 1964, Pl. XIII, figs. 3, 4 left, 6), which Pia (1920, p. 176) named "Fissuration". He identified P. pauciforata, P. praealpina and Diplopora hexaster as species for which fissuration is typical. Since calcification begins firstly around the laterals (to protect the gametes in cladospore species), the parts between the whorls are less calcified and mechanically weaker, so here the calcareous thallus cracks more easily. This phenomenon is consequently dependent on the intensity of calcification and the mechanical forces acting on the thallus, and therefore has no taxonomic significance. The already described cracks located between the lamellae of calcified trichoblasts do not represent fissuration in the sense as defined by Pia (1920, p. 166).
, fig. 4; Bystrick�, 1964, Pl. XIII, figs. 3, 4 left, 6), which Pia (1920, p. 176) named "Fissuration". He identified P. pauciforata, P. praealpina and Diplopora hexaster as species for which fissuration is typical. Since calcification begins firstly around the laterals (to protect the gametes in cladospore species), the parts between the whorls are less calcified and mechanically weaker, so here the calcareous thallus cracks more easily. This phenomenon is consequently dependent on the intensity of calcification and the mechanical forces acting on the thallus, and therefore has no taxonomic significance. The already described cracks located between the lamellae of calcified trichoblasts do not represent fissuration in the sense as defined by Pia (1920, p. 166).
The shape of the laterals in Physoporella pauciforata Steinmann (1903) marked as pear-shaped (German: birnformig), and Pia (1912) latinised this term in pirifer (Latin Pirum = pear). The laterals in reality rarely have a shape somewhat similar to a pear, i.e., with a rounded distal end (Pl. 18 ![]() , fig. 2 left; Bystrick�, 1964, Pl. XIV, fig. 5 bottom right; Bucur et al., 1994, Pl. 11, fig. 8). Because of the wide and round proximal and pointed distal part, the shape of laterals may resemble an onion, turnip, or the roofs of the Cathedral of St. Basil the Blessed in the Red Square in Moscow (Pl. 1
, fig. 2 left; Bystrick�, 1964, Pl. XIV, fig. 5 bottom right; Bucur et al., 1994, Pl. 11, fig. 8). Because of the wide and round proximal and pointed distal part, the shape of laterals may resemble an onion, turnip, or the roofs of the Cathedral of St. Basil the Blessed in the Red Square in Moscow (Pl. 1 ![]() , figs. 1-2, 7, 15; Pl. 18
, figs. 1-2, 7, 15; Pl. 18 ![]() , fig. 1 left; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, figs. 1-2, 4-6). Sometimes the laterals may be slightly elongated and thinner (Pl. 1
, fig. 1 left; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, figs. 1-2, 4-6). Sometimes the laterals may be slightly elongated and thinner (Pl. 1 ![]() , fig. 10; Pl. 2
, fig. 10; Pl. 2 ![]() , fig. 7). The very basal part of the lateral is not preserved in the fossils, but comparing the pore of the studied species with the pores of laterals of other Dasycladalean algae, including the recent ones, it is very likely that a relatively wide proximal part of the lateral was connected by a very narrow opening to the stem cell, which was also assumed by Pia (1912, p. 29). The shape of the laterals and stem cell was reconstructed in Fig. 3
, fig. 7). The very basal part of the lateral is not preserved in the fossils, but comparing the pore of the studied species with the pores of laterals of other Dasycladalean algae, including the recent ones, it is very likely that a relatively wide proximal part of the lateral was connected by a very narrow opening to the stem cell, which was also assumed by Pia (1912, p. 29). The shape of the laterals and stem cell was reconstructed in Fig. 3 ![]() . Since the openness and closure of the pores were rejected as a taxonomic criterion (see
Chapter 4), and it has just been proved that the laterals of the type species of the genus Physoporella, which "should" be completely enveloped in calcareous skeleton, regularly extended beyond the calcareous sheath, including in the neotype (Pl. 1
. Since the openness and closure of the pores were rejected as a taxonomic criterion (see
Chapter 4), and it has just been proved that the laterals of the type species of the genus Physoporella, which "should" be completely enveloped in calcareous skeleton, regularly extended beyond the calcareous sheath, including in the neotype (Pl. 1 ![]() , fig. 1), this criterion will no longer be discussed in further descriptions of individual taxa.
, fig. 1), this criterion will no longer be discussed in further descriptions of individual taxa.
The laterals are arranged in evenly spaced single whorls. In adjacent whorls, the laterals are positioned mostly above each other, and not alternately, which can be seen not only in the longitudinal-oblique and tangential sections (Pl. 1 ![]() , figs. 8, 15; Pl. 18
, figs. 8, 15; Pl. 18 ![]() , figs. 1-2; Bystrick�, 1964, Pl. XIII, figs. 2-3, 6; Gawlick et al., Pl. 5, fig. b), but also, after the symmetrical arrangement of pores, in longitudinal sections (Pl. 18
, figs. 1-2; Bystrick�, 1964, Pl. XIII, figs. 2-3, 6; Gawlick et al., Pl. 5, fig. b), but also, after the symmetrical arrangement of pores, in longitudinal sections (Pl. 18 ![]() , fig. 4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII , figs. 4-5; Herak, 1965, Pl. XIII, fig. 5). In the dilated proximal part, the laterals mainly touch each other within the same whorl (Pl. 1
, fig. 4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII , figs. 4-5; Herak, 1965, Pl. XIII, fig. 5). In the dilated proximal part, the laterals mainly touch each other within the same whorl (Pl. 1 ![]() , figs. 1-2, 7.9; Pl. 2
, figs. 1-2, 7.9; Pl. 2 ![]() , fig. 10; Pl. 18
, fig. 10; Pl. 18 ![]() , figs. 1-2; Bystrick�, 1964, Pl. XIII, figs. 2-4, 6; Pl. XIV, figs. 5-6), and occasionally they are also lateraly flattened (Pl. 1
, figs. 1-2; Bystrick�, 1964, Pl. XIII, figs. 2-4, 6; Pl. XIV, figs. 5-6), and occasionally they are also lateraly flattened (Pl. 1 ![]() , figs. 15). Very rarely they can be so compacted that their arrangement within the whorl is no longer symmetrical in one plane (Pl. 18
, figs. 15). Very rarely they can be so compacted that their arrangement within the whorl is no longer symmetrical in one plane (Pl. 18 ![]() , fig. 3). The laterals are generally perpendicular to the longitudinal axis (Pl. 1
, fig. 3). The laterals are generally perpendicular to the longitudinal axis (Pl. 1 ![]() , figs. 4, 15; Pl. 18
, figs. 4, 15; Pl. 18 ![]() , figs. 1-3; Bystrick�, 1964, Pl. XIII, figs. 3-6; Herak, 1965, Pl. XIII, fig. 5; Gawlick et al., Pl. 5, fig. b), and may be slightly inclined upwards (Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, fig. 2; Buček, 1989, Pl. II, fig. 1), approaching var. sulcata. A peculiar specimen is shown on Pl. 18
, figs. 1-3; Bystrick�, 1964, Pl. XIII, figs. 3-6; Herak, 1965, Pl. XIII, fig. 5; Gawlick et al., Pl. 5, fig. b), and may be slightly inclined upwards (Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, fig. 2; Buček, 1989, Pl. II, fig. 1), approaching var. sulcata. A peculiar specimen is shown on Pl. 18 ![]() , fig. 4, where the laterals in the lower part of the thallus are perpendicular to the axis, and in the upper part are oblique, but "exceptions confirm the rule" and it would be meaningless to name this specimen as a new variety or a species.
, fig. 4, where the laterals in the lower part of the thallus are perpendicular to the axis, and in the upper part are oblique, but "exceptions confirm the rule" and it would be meaningless to name this specimen as a new variety or a species.
Emended diagnosis: Cylindrical thallus with a smooth, flat or slightly wrinkled, inner surface that reflects the shape of the stem cell. The outer surface is slightly undulated and/or ornamented with calcareous lamellae and spikes that represent calcified trichoblasts. The shape of the laterals is piriform, characterized by a wide rounded proximal part and a gradual thinning towards the distal end, similar to the shape of an onion or a turnip. The distal end can also be rounded, so it resembles a pear to some extent (Lat. "Pirum" = pear). In its widest part, the laterals within one whorl mostly touch each other, and may distally continue into trichoblasts that are predominantly bent upwards. The laterals are arranged in evenly spaced single whorls and are mostly perpendicular to the longitudinal axis, and less frequently slightly inclined upwards. In adjacent whorls the laterals are located mostly one above the other.
|
|
Figure 3: Physoporella pauciforata var. pauciforata. Idealized longitudinal (A), transverse (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of calcareous skeleton. The longitudinal section (A) shows variants with wrinkled (I) and smooth inner surface (II-III, V), slightly undulated outer surface of the thallus (I, III, V), diagenetically dissolved inner and outer surface (IV), calcification very close to the stem cell
(I-II) and near to the stem cell (III, V), calcified proximal part of the trichoblast (I-II), and a rare form of a "conical" lateral (V). The cross section (B) shows variants with a smooth (VI) and undulated (VII) outer surface of the thallus, lamellae representing calcified trichoblasts (VIII), calcification that reaches very close to the stem cell (VI), near to the stem cell (VII) and only in the distal part of the laterals (VIII). The tangential section (C) shows the sections of the proximal (upper whorl) and distal parts of the laterals (lower whorl). |
Physoporella pauciforata var. sulcata Bystrick�, 1962
Pl. 1 ![]() , figs. 11, 12 pars; Pl. 4
, figs. 11, 12 pars; Pl. 4 ![]() , fig. 3; Pl. 16
, fig. 3; Pl. 16 ![]() , fig. 1, Pl. 18
, fig. 1, Pl. 18 ![]() , fig. 6
, fig. 6
1912 - Physoporella pauciforata - Pia, p. 44, Pl. V, fig. 12.
1962 - Physoporella pauciforata var. sulcata n. var. - Bystrick�, p. 232-233, Pl. IV, figs. 2-3, 6.
1964 - Physoporella pauciforata var. sulcata - Bystrick�, p. 121-122, Pl. XIV, figs. 1-2; Pl. XV, figs. 1-2, from Bystrick� (1962, Pl. IV, figs. 3 2,); Pl. XV, figs. 3, 5a.
1964 - Physoporella pauciforata var. pauciforata n. var. - Bystrick�, Pl. XV, figs. 4, 5.b.
1964 - Physoporella pauciforata var. sulcata - Dieni & Spagnulo, p. 8, Pl. II, figs. 1-2.
1967a - Physoporella minutuloidea n. sp. - Herak, p. 101, Pl. 9, fig. 1.
1970 - Physoporella pauciforata var. sulcate - Pantić, p. 380, Pl. X, figs. 1-4.
1986 - Physoporella pauciforata var. sulcate - Bystrick�, p. 295-301, Pl. I, fig. 10, from Bystrick� (1962, Pl. IV, fig. 3); Pl. II, fig. 5.
1989 - Physoporella pauciforata var. sulcata - Buček, p. 134, Pl. II, fig. 3.
1993 - Physoporella pauciforata var. sulcata - Budai et al., Pl. VI, fig. 3.
2003 - Physoporella pauciforata var. sulcata - Piros, p. 57, Pl. D-I, fig. 4.
2003 - Physoporella minutuloidea - Piros, p. 58, Pl. D-III, fig. 3.b; Pl. D-IV, fig. 1.
2003 - Physoporella pauciforata var. undulata - Piros, p. 56-57, Pl. D-III, figs. 1.b, 3-4.
2021 - Physoporella pauciforata - Gawlick et al., Pl. 5, fig. a.
Origin of the name: After the furrowed outer surface of the calcareous thallus (Latin "sulcus" = furrow, groove).
Studied material: Holotype of Physoporella pauciforata var. sulcata (Bystrick�, 1962, Pl. IV, fig. 2 = this work Tab. XIX, fig. 1) originates from excavation M-5513/59, taken from the Upper Anisian limestone from the Rakot� locality on the Silicka planina in Slovakia.
The specimen chosen by Herak (1967a, p. 101, Pl. 9, fig. 1 = this paper Pl. 4 ![]() , fig. 3) as a holotype of a new species Physoporella minutuloidea, which originates from the Anisian of the Greek island of Khios (specimen T219/4), by its characteristics also belongs to the variety P. pauciforata var. sulcata, and is therefore its junior synonym. According to the author, the sample is kept in the collection of K.-W. Tietze in "Geologisch-paleontologischen Institut der Universit�t Marburg".
, fig. 3) as a holotype of a new species Physoporella minutuloidea, which originates from the Anisian of the Greek island of Khios (specimen T219/4), by its characteristics also belongs to the variety P. pauciforata var. sulcata, and is therefore its junior synonym. According to the author, the sample is kept in the collection of K.-W. Tietze in "Geologisch-paleontologischen Institut der Universit�t Marburg".
In the material from Julius Pia, originally described as P. pauciforata, I determined as var. sulcata specimens from Tab. 1, figs. 11, 12 pars (thin-sections XXI2 and XXI3). One illustrated specimen (Pl. 18 ![]() , fig. 6) originates from the Anisian limestones of the Belski dol locality on Ivan�čica Mt.
, fig. 6) originates from the Anisian limestones of the Belski dol locality on Ivan�čica Mt.
History of investigations: Bystrick� (1962, p. 232, 239) give the diagnosis of Physoporella pauciforata var. sulcata var. nov.: "The outer surface of the shell is markedly furrowed with oblique furrows that are sharply cut into the shell. Each segment of the shell bounded by furrows has only one whorl of pores." It will be noticed that the focus is given to the morphology of the skeleton (furrows, segmentation), and not to the structure of the algae itself.
(In Slovak: "Vonkaj�i povrch schranky vyrazne ryhovany �ikmymi, ostro do schranky sa zarezevajucimi ryhami. Na ka�dy segment schranky obmedzeny ryhami pripada iba jeden praslen porov.")
Herak (1967a, p. 101) give the diagnosis of Physoporella minutuloidea n. sp. : "The thallus is cylindrical, the pores are rounded outwards. The outer shape of the calcareous shell shows an undulation rather than a real annulation. It is a shape that is very similar to Ph. minutula Pia in terms of dimensions and shape of the pores. It differs from it, however, in particular by distinctly single-row whorls, which are quite distant from each other." Although the specimens illustrated by Herak (1967a, Pl. 12, fig. 3), as well as those by Piros (2003, Pl. D-III, fig. 3.b; Pl. D-IV, fig. 1), are somewhat smaller, nevertheless they fall within the range of dimensions of P. p. sulcata, so I consider this species to be its synonym.
Description: The thallus is mostly cylindrical and straight (Pl. 1 ![]() , fig. 11; Bystrick�, 1962, Pl. IV, fig. 3; 1964, Pl. XIV, fig. 2; Pl. XV, fig. 3; Dieni & Spagnulo, 1964, Pl. II, fig. 1), and rarely very slightly claviform (Bystrick�, 1962, Pl. IV, fig. 2 = this paper Pl. 16
, fig. 11; Bystrick�, 1962, Pl. IV, fig. 3; 1964, Pl. XIV, fig. 2; Pl. XV, fig. 3; Dieni & Spagnulo, 1964, Pl. II, fig. 1), and rarely very slightly claviform (Bystrick�, 1962, Pl. IV, fig. 2 = this paper Pl. 16 ![]() , fig. 1). The inner surface of the thallus is smooth (Bystrick�, 1962, Pl. IV, figs. 2-3; Herak, 1967a, Pl. 9, fig. 1 = this work Pl. 4
, fig. 1). The inner surface of the thallus is smooth (Bystrick�, 1962, Pl. IV, figs. 2-3; Herak, 1967a, Pl. 9, fig. 1 = this work Pl. 4 ![]() , fig. 3), and exceptionally it can be wrinkled (Bystrick�, 1964, Pl. XIV, fig. 1 left; Dieni & Spagnulo, 1964, Pl. II, fig. 1), as described in var. pauciforata. The inner surface can be uneven when the proximal part of the laterals is not calcified (Pl. 1
, fig. 3), and exceptionally it can be wrinkled (Bystrick�, 1964, Pl. XIV, fig. 1 left; Dieni & Spagnulo, 1964, Pl. II, fig. 1), as described in var. pauciforata. The inner surface can be uneven when the proximal part of the laterals is not calcified (Pl. 1 ![]() , fig. 11 above right) or when there has been later erosion or dissolution (Pl. 1
, fig. 11 above right) or when there has been later erosion or dissolution (Pl. 1 ![]() , fig. 11 top left and bottom). The outer surface of the calcareous thallus is regularly furrowed with sharp and oblique furrows. These furrows are a direct consequence of the shape and position of the laterals (Fig. 4
, fig. 11 top left and bottom). The outer surface of the calcareous thallus is regularly furrowed with sharp and oblique furrows. These furrows are a direct consequence of the shape and position of the laterals (Fig. 4 ![]() ), and only in this sense they can have taxonomic importance. The pores are very similar in shape to var. pauciforata, only inclined upwards. The shape of the pores and the structure of this variety are best seen in the illustrations in Bystrick� (1962, Pl. IV, figs. 2-3; 1964, Pl. XIV, figs. 1-2; Pl. XV, fig. 3; 1986, Pl. II, fig. 5) and Dieni and Spagnulo (1964, Pl. II, fig. 1). The pores are often somewhat thinner and longer than in the type variety (Pl. 18
), and only in this sense they can have taxonomic importance. The pores are very similar in shape to var. pauciforata, only inclined upwards. The shape of the pores and the structure of this variety are best seen in the illustrations in Bystrick� (1962, Pl. IV, figs. 2-3; 1964, Pl. XIV, figs. 1-2; Pl. XV, fig. 3; 1986, Pl. II, fig. 5) and Dieni and Spagnulo (1964, Pl. II, fig. 1). The pores are often somewhat thinner and longer than in the type variety (Pl. 18 ![]() , fig. 6; Bystrick�, 1962, Pl. IV, figs. 2-3; 1964, Pl. XV, fig. 4; Pantić, 1970, Pl. X, figs. 1-4), which is a consequence of their oblique position. The laterals, as in var. pauciforata, had a trichoblast on the apex, which can be recognized as spikes on the outer wall of the thallus (Pl. 18
, fig. 6; Bystrick�, 1962, Pl. IV, figs. 2-3; 1964, Pl. XV, fig. 4; Pantić, 1970, Pl. X, figs. 1-4), which is a consequence of their oblique position. The laterals, as in var. pauciforata, had a trichoblast on the apex, which can be recognized as spikes on the outer wall of the thallus (Pl. 18 ![]() , fig. 6 right; Bystrick�, 1962, Pl. IV, fig. 2 right; 1964, Pl. XIV, fig. 2; 1986; Pl. I, fig. 10; Pl. II, fig. 5). Due to the oblique position of the laterals and trichoblasts that are bent upwards, the formation of a calcareous sheath uniformly around the laterals causes furrows between whorls that, in fact, represent a pronounced form of undulation (Fig. 4
, fig. 6 right; Bystrick�, 1962, Pl. IV, fig. 2 right; 1964, Pl. XIV, fig. 2; 1986; Pl. I, fig. 10; Pl. II, fig. 5). Due to the oblique position of the laterals and trichoblasts that are bent upwards, the formation of a calcareous sheath uniformly around the laterals causes furrows between whorls that, in fact, represent a pronounced form of undulation (Fig. 4 ![]() ). On the inner side of the thallus, calcification mainly reaches close to the stem cell (Bystrick�, 1962, Pl. IV, figs. 1-3; Herak, 1967a, Pl. 9, fig. 1; Pantić, 1970, Pl. X, figs. 1-4), and rarely immediately close to it (Dieni & Spagnulo, 1964, Pl. II, fig. 1; Bystrick�, 1986, Pl. II, fig. 5). The laterals are arranged in spaced whorls, and rarely the whorls are close to each other (Pl. 1
). On the inner side of the thallus, calcification mainly reaches close to the stem cell (Bystrick�, 1962, Pl. IV, figs. 1-3; Herak, 1967a, Pl. 9, fig. 1; Pantić, 1970, Pl. X, figs. 1-4), and rarely immediately close to it (Dieni & Spagnulo, 1964, Pl. II, fig. 1; Bystrick�, 1986, Pl. II, fig. 5). The laterals are arranged in spaced whorls, and rarely the whorls are close to each other (Pl. 1 ![]() , fig. 11; Bystrick�, 1964, Pl. XIV, fig. 1). The laterals in adjacent whorls are located mainly one above the other (Pl. 18
, fig. 11; Bystrick�, 1964, Pl. XIV, fig. 1). The laterals in adjacent whorls are located mainly one above the other (Pl. 18 ![]() , fig. 6; Bystrick�, 1962, Pl. IV, fig. 2; 1964, Pl. XIV, fig. 1; Pl. XV, fig. 3).
, fig. 6; Bystrick�, 1962, Pl. IV, fig. 2; 1964, Pl. XIV, fig. 1; Pl. XV, fig. 3).
That this is a variety, and not a separate species, is evident from a complete transitional series of forms whose laterals are perpendicular to the longitudinal axis (Pl. 1 ![]() , figs. 4, 15; Pl. 18
, figs. 4, 15; Pl. 18 ![]() , figs. 1-3; Bystrick�, 1964 , Pl. XIII, figs. 3-6) or are slightly oblique (Pl. 18
, figs. 1-3; Bystrick�, 1964 , Pl. XIII, figs. 3-6) or are slightly oblique (Pl. 18 ![]() , fig. 4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, fig. 2; Buček, 1989, Pl. II, fig. 1) and are included in var. pauciforata, over the less oblique ones (Pl. 1
, fig. 4; Bystrick�, 1957, Pl. VI, fig. 1; 1964, Pl. XIII, fig. 2; Buček, 1989, Pl. II, fig. 1) and are included in var. pauciforata, over the less oblique ones (Pl. 1 ![]() , fig. 11; Bystrick�, 1964, Pl. XIV, fig. 1; Pl. XV, figs. 3, 5; Herak, 1967a, Pl. 9, fig. 1), to very steep ones (Pl. 18
, fig. 11; Bystrick�, 1964, Pl. XIV, fig. 1; Pl. XV, figs. 3, 5; Herak, 1967a, Pl. 9, fig. 1), to very steep ones (Pl. 18 ![]() , fig. 6; Bystrick�, 1962, Pl. IV, figs. 2-3; Pantić, 1970, Pl. X, figs. 1 -4) that are arbitrarily included in var. sulcata. The greater the inclination of the laterals, the narrower and deeper are furrows in the shell.
, fig. 6; Bystrick�, 1962, Pl. IV, figs. 2-3; Pantić, 1970, Pl. X, figs. 1 -4) that are arbitrarily included in var. sulcata. The greater the inclination of the laterals, the narrower and deeper are furrows in the shell.
Emended diagnosis: Cylindrical, rarely slightly claviform, thallus with a smooth flat or slightly wrinkled inner surface that reflects shape of the stem cell. The outer surface of the calcareous thallus is characterized by oblique furrows that reflect the shape and the inclination of the laterals. There is a continuous transition from var. pauciforata: the greater the slope of the laterals, the narrower and deeper are furrows in the shell. The laterals are of the piriform type, carrying an upward-pointing trichoblast at their apex. They are located in evenly spaced single whorls and in adjacent whorls are located mostly one above the other.
|
|
Figure 4: Physoporella pauciforata var. sulcata. Idealized longitudinal section of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Variants with smooth (I, III) and wrinkled stem cell (II), and calcification very close to the stem cell (I) and near to the stem cell (II-III) are shown. Trichoblasts on the top of the laterals are shown on the upper three whorls. Intense undulation of the outer surface of the thallus with oblique grooves is the result of calcification of the steeply inclined laterals. |
Physoporella pauciforata var. undulata Pia, 1935a
Pl. 2 ![]() , figs. 1-4
, figs. 1-4
1920 - Physoporella pauciforata var. ? lotharingica n. stat. n. comb. - Pia, p. 52-53, Pl. III, figs. 12, 14.
1935a - Physoporella pauciforata var. undulata n. var. - Pia, p. 223-226, Figs. 33-34.
1957 - Physoporella pauciforata undulata.- Bystrick�, p. 233, Pl. VI, figs. 3-4.
1964 - Physoporella pauciforata var. undulata n. var. - Bystrick�, p. 124-125, Pl. XVII, figs. 1-6.
1983 - Physoporella pauciforata var. undulata - Bystrick�, p. 515-523, Pl. III, fig. 17; Pl. IV, figs. 21-22; Pl. V, fig. 23; Pl. IV, fig., 20, from Bystrick� (1957, Pl. VI, fig. 4).
1993 - Physoporella pauciforata undulata - Budai et al., Pl. V, fig. 3.
2003 - Physoporella pauciforata var. undulata - Piros, p. 56-57, Pl. D-IV, figs. 2-4.
Origin of the name: Probably after the wavy outer surface of the thallus (Latin "undosus" = wavy).
Studied material: The type material from Pia (1935a) originates from Trebević mountain south of Sarajevo (Bosnia and Herzegovina). Thin-section 561a/5 is from the Anisian limestone from site "h" (Pia, 1935a). The illustrated specimen (Pl. 2 ![]() , fig. 1 = Pia, 1935a, Fig. 34) was designated by Bystrick� (1983) as lectotype. Other thin sections are (localities are shown on plate descriptions): 555a (Pl. 2
, fig. 1 = Pia, 1935a, Fig. 34) was designated by Bystrick� (1983) as lectotype. Other thin sections are (localities are shown on plate descriptions): 555a (Pl. 2 ![]() , figs. 2-3 = Pia, 1935a, Figs. 33 and 68 (Pl. 2
, figs. 2-3 = Pia, 1935a, Figs. 33 and 68 (Pl. 2 ![]() , fig. 4).
, fig. 4).
History of investigations: Pia (1935a, p. 223-226) defines a new variety: "I found Physoporellas here quite often, whose calcareous shell between the whorls is very strongly constricted. Even the tips of the individual branches stand out as bulges on the outside (see Pia, 1935a, Figs. 33-34). The whorls are single-row, which can be seen especially in Pia (1935a, Fig. 34 = this paper, Pl. 3 ![]() , fig. 1), the pores strongly tapered towards the outside, but are almost always closed by a very thin calcareous layer. Otherwise they just had to be filled with sediment in the distal part. But here they only contain clear crystalline calcite, which is often difficult to distinguish from the shell (which is of course not easy to see from the schematic drawings). I name this alga Physoporella pauciforata var. undulata." Specimens previously described from Bosnia as Physoporella pauciforata var. ? lotharingica (Pia, 1920, Pl. III, figs. 12, 14) the author also adds to this variety.
, fig. 1), the pores strongly tapered towards the outside, but are almost always closed by a very thin calcareous layer. Otherwise they just had to be filled with sediment in the distal part. But here they only contain clear crystalline calcite, which is often difficult to distinguish from the shell (which is of course not easy to see from the schematic drawings). I name this alga Physoporella pauciforata var. undulata." Specimens previously described from Bosnia as Physoporella pauciforata var. ? lotharingica (Pia, 1920, Pl. III, figs. 12, 14) the author also adds to this variety.
(In German: "Unten den bosnischen diploporen scheint sie nicht vorzukommen. Dagegen fand ich hier nicht ganz selten Physoporellen, deren Kalkschale zwischen den Wirteln sehr stark eingeschn�rt ist. Ja sogar die Spitzen der einzelnen Zweige zeichnen sich auf der Aussenseite etwas als Erh�hungen ab. Die Wirtel sind, wie besonders fig. 34 zeigt, einreihig, die Poren gegen aussen stark verj�ngt, aber doch wohl fast immer durch eine ganz d�nne Kalklage verschlossen. Sonst mussten sie gerade im distalen Teil mit Sediment erf�llt sein. Sie enthalten hier aber (was aus den schematischen Zeichnungen freilich nicht gut zu ersehen ist) nur klaren, kristallinen Kalkspat, der sich oft schlecht von der Schale abhebt. Ich nenne diese Alge Undunata.")
Bystrick� (1964, p. 124) simplifies this diagnosis: "The thallus is distinctly wavy, the pores of pirifer type in single-rowed whorls, perpendicular to the longitudinal axis of the cell". Bystrick� (1983, p. 517-518) gives a statistical analysis of dimensions and a detailed description of this variety: "The thallus is a simple cylinder with a completely smooth inner surface, but a strongly undulated outer surface. The sleeve articulation is restricted to fissures (gaps); it is irregular and the height of the segments limited by them varies considerably even within one specimen. Branches are of the typical piriferous type, mostly with their distal ends closed by a fine layer of the calcareous sleeve. However, branches with their distal ends open also occur. It is not always possible to decide whether it is a primary phenomenon or only a secondary abrasion of the most external parts of the sleeve by transport or by other destruction. The cavities after branches could have been filled by sediment also from the inner side, through the central cavity. The transverse section through the branches is round or almost round, not only in their distal part, but also in the proximal one adhering to the central stem. The branches are arranged in one-rowed whorls, but not always exactly in the same plane. The whorls are almost perpendicular to the plant axis. Though the thallus inner cavity is completely smooth, it very probably does not completely correspond to the central stem diameter. A pore, connecting with the central stem has not been observed even in one case. The number of the branches in a whorl is 14�1." Again, there is also a futile discussion of the openness and closure of the pores, while the shapes and arrangement of the laterals are given relatively less importance.
Description: The thallus is generally cylindrical in shape (Pl. 2 ![]() , fig. 1; Bystrick�, 1964, Pl. XVII, figs. 1-3), and exceptionally, as with most other Dasycladales, a slightly curved thallus may also occur (Bystrick�, 1957, Pl. VI, fig. 3). The inner surface of the thallus is completely smooth (Pl. 2
, fig. 1; Bystrick�, 1964, Pl. XVII, figs. 1-3), and exceptionally, as with most other Dasycladales, a slightly curved thallus may also occur (Bystrick�, 1957, Pl. VI, fig. 3). The inner surface of the thallus is completely smooth (Pl. 2 ![]() , figs. 2-4; Pia, 1920, Pl. III, fig. 12; Bystrick�, 1957, Pl. VI, fig. 4; 1964, Pl. XVII, figs. 1- 2, 4-6), so no wrinkling of the stem cell has been observed. In some specimens, due to diagenetic changes (dissolution, erosion), the inner surface is uneven (Pl. 2
, figs. 2-4; Pia, 1920, Pl. III, fig. 12; Bystrick�, 1957, Pl. VI, fig. 4; 1964, Pl. XVII, figs. 1- 2, 4-6), so no wrinkling of the stem cell has been observed. In some specimens, due to diagenetic changes (dissolution, erosion), the inner surface is uneven (Pl. 2 ![]() , fig. 1; Bystrick�, 1964, Pl. XVII, fig. 3). As with other varieties, calcification reaches very close to the stem cell, but it does not calcify itself. Due to the relative spacing of the whorls and the fact that each lateral is surrounded by a relatively thin individual calcareous sheath, the outer surface of the thallus is markedly undulated (Fig. 5
, fig. 1; Bystrick�, 1964, Pl. XVII, fig. 3). As with other varieties, calcification reaches very close to the stem cell, but it does not calcify itself. Due to the relative spacing of the whorls and the fact that each lateral is surrounded by a relatively thin individual calcareous sheath, the outer surface of the thallus is markedly undulated (Fig. 5 ![]() ). Depending on the distance between the whorls and the intensity of calcification, undulation may be milder (Bystrick�, 1964, Pl. XVII, fig. 3; 1983, Pl. I, fig. 21), thus approaching the undulated forms of var. pauciforata, moderately pronounced, as in most specimens, or very pronounced as in the specimen from Bystrick� (1964, Pl. XVII, fig. 5).
). Depending on the distance between the whorls and the intensity of calcification, undulation may be milder (Bystrick�, 1964, Pl. XVII, fig. 3; 1983, Pl. I, fig. 21), thus approaching the undulated forms of var. pauciforata, moderately pronounced, as in most specimens, or very pronounced as in the specimen from Bystrick� (1964, Pl. XVII, fig. 5).
The laterals are of the piriform type, often distally rounded, and sometimes pointed. That these laterals also continued into trichoblasts is proven by spikes on the outer wall of the limestone shell, that are visible sometimes in longitudinal and oblique sections (Pl. 2 ![]() , figs. 1-3). In other published specimens, these spikes are not visible or are only slightly pronounced (Bystrick�, 1983, Pl. V, fig. 23). The reason is probably that the trichoblasts were not steeply bent upwards, as in var. pauciforata, but were (at least in the proximal part) more or less horizontal, so they came out of the mucous sheet in which calcification takes place.
, figs. 1-3). In other published specimens, these spikes are not visible or are only slightly pronounced (Bystrick�, 1983, Pl. V, fig. 23). The reason is probably that the trichoblasts were not steeply bent upwards, as in var. pauciforata, but were (at least in the proximal part) more or less horizontal, so they came out of the mucous sheet in which calcification takes place.
The laterals are arranged in regularly spaced single-rowed whorls. Due to the relative compactness of the laterals within a whorl, that is visible in the cross and cross-oblique sections (Pl. 2 ![]() , figs. 3-4; Pia, 1920, Pl. III, fig. 14), the laterals are generally not distributed within one plane but they protrude slightly upwards or downwards, is visible in oblique and tangential sections (Pl. 2
, figs. 3-4; Pia, 1920, Pl. III, fig. 14), the laterals are generally not distributed within one plane but they protrude slightly upwards or downwards, is visible in oblique and tangential sections (Pl. 2 ![]() , fig. 2; Bystrick�, 1983, Pl. IV, figs., 20-22), and was also mentioned by Bystrick� (1983). It is difficult to assess whether they are positioned alternately or one above the other in adjacent whorls, although I am more inclined to favour the second suggestion (Pl. 2
, fig. 2; Bystrick�, 1983, Pl. IV, figs., 20-22), and was also mentioned by Bystrick� (1983). It is difficult to assess whether they are positioned alternately or one above the other in adjacent whorls, although I am more inclined to favour the second suggestion (Pl. 2 ![]() , figs. 1-2; Bystrick�, 1983, Pl. IV, figs., 20-22). The laterals are mostly perpendicular to the longitudinal axis (Pl. 2
, figs. 1-2; Bystrick�, 1983, Pl. IV, figs., 20-22). The laterals are mostly perpendicular to the longitudinal axis (Pl. 2 ![]() , figs. 1-2; Bystrick�, 1964, Pl. XVII, figs. 1-2), and rarely slightly inclined upwards (Bystrick�, 1957, Pl. VI, fig. 4).
, figs. 1-2; Bystrick�, 1964, Pl. XVII, figs. 1-2), and rarely slightly inclined upwards (Bystrick�, 1957, Pl. VI, fig. 4).
Emended diagnosis: Cylindrical thallus with smooth inner surface. Due to the relative spacing of the whorls and the fact that each lateral is surrounded by an individual calcareous sheath, the outer surface of the thallus is markedly undulated. Depending on the distance between the whorls and the intensity of calcification, the intensity of undulation can vary. The laterals are of the piriform type, often distally rounded, sometimes pointed. Distally, they continue into the trichoblast, which was more or less horizontal in the basal part. The laterals are densely arranged in regularly spaced single whorls.
|
|
Figure 5: Physoporella pauciforata var. undulata. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows the variants with calcification very close to the stem cell (I) and near to the stem cell (II). Trichoblasts on the top of laterals are shown on the upper two whorls. The tangential section (B) shows the sections of the proximal (upper two whorls) and distal parts of the laterals (lower whorl). The undulation of the outer surface of the calcareous thallus is visible. |
Physoporella pauciforata var. gemerica Bystrick�, 1962
Pl. 16 ![]() , figs. 6, 9; Pl. 18
, figs. 6, 9; Pl. 18 ![]() , fig. 5
, fig. 5
1962 - Physoporella pauciforata var. gemerica n. var. - Bystrick�, p. 233, Pl. IV, figs. 4-5.
1964 - Physoporella pauciforata var. gemerica - Bystrick�, p. 122-123, Pl. XVI, figs. 4, 1, from Bystrick� (1962, Pl. IV, figs. 4-5); Pl. XVI, figs. 2-3, 5; Pl. XXXIV, fig. 2 pars.
1967b - Physoporella pauciforata var. gemerica - Herak, Pl. 1, fig. 7.
1989 - Physoporella pauciforata var. gemerica - Buček, p. 134, Pl. II, fig. 2.
2003 - Physoporella pauciforata var. gemerica - Piros, p. 58, Pl. D-IV, fig. 1.b.
Origin of the name: After the geographical area in which the type locality is: Gemer in Slovakia.
Studied material: The holotype of P. pauciforata var. gemerica (Bystrick�, 1962, Pl. IV, fig. 4 = this work Pl. 16 ![]() , fig. 6) is in thin-section Pč-109 that originates from Doln� Vrch, NE Torn�nadaska in the Slovak Karst or, since the site is on the Hungarian side of the border, in Aggteleki-karszt. The sample was taken from the light limestone of the Upper Anisian (Pelsonian - Lower Illyrian). The holotype is kept in Geologick�� prieskum, n.p. (now Progeo spol. s r.o.), �ilina, Slovakia. In addition to the holotype, I also illustrated a specimen from Bystrick� (1964, Pl. XVI, fig. 3 = this paper Pl. 16
, fig. 6) is in thin-section Pč-109 that originates from Doln� Vrch, NE Torn�nadaska in the Slovak Karst or, since the site is on the Hungarian side of the border, in Aggteleki-karszt. The sample was taken from the light limestone of the Upper Anisian (Pelsonian - Lower Illyrian). The holotype is kept in Geologick�� prieskum, n.p. (now Progeo spol. s r.o.), �ilina, Slovakia. In addition to the holotype, I also illustrated a specimen from Bystrick� (1964, Pl. XVI, fig. 3 = this paper Pl. 16 ![]() , fig. 9). The specimen illustrated on Pl. 18
, fig. 9). The specimen illustrated on Pl. 18 ![]() , fig. 5 is from the Anisian limestone of Hum �agudovac on the northern side of Medvednica Mt.
, fig. 5 is from the Anisian limestone of Hum �agudovac on the northern side of Medvednica Mt.
History of investigations: Bystrick� (1962, p. 233) gives a diagnosis of a new variety of Physoporella pauciforata var. gemerica: "The shell is unevenly grooved. The grooves that cut into the outer surface of the shell are sharp but very shallow. The pores are in single-row of close whorls, laid almost perpendicular to the longitudinal axis of the thallus."
(In Slovak: "Schranka neravnomerne ryhovana. Ryhy zarezavajuce sa do vonkaj�ieho povrchu schranky su ostre, ale velmi plytke. Pory v jednoradovych tesne k sebe priliehajucich praslenoch prebiehajucich takmer kolmo na pozdl�nu os stielky.")
In his next work, Bystrick� (1964, p. 122-123) repeats the previous conclusion and point out that "The pores touch each other, not only within a single whorl, but also in adjacent whorls, or are separated only by a thin strip of the skeleton."
Description: The thallus is cylindrical in shape. The main characteristic of this variety is a dense arrangement of the whorls along the thallus. Therefore, the proximal parts of the laterals are close or compressed together, so that there was not much space for calcifying mucus and calcification usually does not reach the stem cell. Therefore, the inner surface of the thallus, i.e., the outer surface of the stem cell is usually not visible. Due to a lack of information I interpreted the outer surface of the stem cell as smooth (Fig. 6 ![]() ), although it is possible that it was finely wrinkled, as is suggested on the left side of the holotype (Bystrick�, 1962, Pl. IV, fig. 4 = this paper Pl. 16
), although it is possible that it was finely wrinkled, as is suggested on the left side of the holotype (Bystrick�, 1962, Pl. IV, fig. 4 = this paper Pl. 16 ![]() , fig. 6). Maybe this fine folding of the stem cell caused a reduction in the distance between the whorls? I originally planned to elevate this variety to the rank of an independent species, as there seems to be very little variation in this alga and there are no transitional forms. However, if we compare, for example, the illustrations from Bystrick� (1964) that show var. undulata (Pl. XVII, fig. 3), var. sulcata (Pl. XIV, figs. 1-2) and var. gemerica (Pl. XVI, fig. 5) it is obvious that a transitional series of forms does exist.
, fig. 6). Maybe this fine folding of the stem cell caused a reduction in the distance between the whorls? I originally planned to elevate this variety to the rank of an independent species, as there seems to be very little variation in this alga and there are no transitional forms. However, if we compare, for example, the illustrations from Bystrick� (1964) that show var. undulata (Pl. XVII, fig. 3), var. sulcata (Pl. XIV, figs. 1-2) and var. gemerica (Pl. XVI, fig. 5) it is obvious that a transitional series of forms does exist.
The laterals are piriform, somewhat larger and with less pronounced thinning at the distal end. The laterals continue into trichoblasts that are relatively steeply bent upwards as in var. pauciforata and var. sulcata. They are visible in the longitudinal sections with spikes and horns pointing upwards, that represent the calcified proximal parts of the trichoblasts, which can be seen in almost all illustrated specimens (Bystrick�, 1962, Pl. IV, figs. 4-5; 1964, Pl. XVI, fig. 5; Herak, 1967b, Pl. 1, fig. 7; Buček, 1989, Pl. II, fig. 2), and on Pl. 18 ![]() , fig. 5 only as indications. As a consequence of calcified trichoblasts, the "grooves" mentioned by Bystrick� (1962, 1964) appear on the surface of the thallus. The laterals in the adjacent whorls are placed one above the other. Due to the dense arrangement and mutual "squeezing" of the laterals, they sometimes have a slightly square shape in cross section (Pl. 16
, fig. 5 only as indications. As a consequence of calcified trichoblasts, the "grooves" mentioned by Bystrick� (1962, 1964) appear on the surface of the thallus. The laterals in the adjacent whorls are placed one above the other. Due to the dense arrangement and mutual "squeezing" of the laterals, they sometimes have a slightly square shape in cross section (Pl. 16 ![]() , fig. 9 = Bystrick�, 1964, Pl. XVI, fig. 3). The laterals are mostly perpendicular to the longitudinal axis, but can also be slightly oblique to it (Pl. 16
, fig. 9 = Bystrick�, 1964, Pl. XVI, fig. 3). The laterals are mostly perpendicular to the longitudinal axis, but can also be slightly oblique to it (Pl. 16 ![]() , fig. 6 = Bystrick�, 1962, Pl. IV, fig. 4).
, fig. 6 = Bystrick�, 1962, Pl. IV, fig. 4).
Emended diagnosis: Cylindrical thallus with calcification that rarely reaches the stem cell. The laterals are densely arranged in very close single whorls, mostly perpendicular, and seldom slightly oblique to the longitudinal axis. In adjacent whorls laterals stand one above the other. The laterals are of the piriform type, and carry an upward-pointing trichoblast at their apex. Calcification of the laterals creates an uneven outer surface of the thallus with spikes and grooves.
|
|
Figure 6: Physoporella pauciforata var. gemerica. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows variants with calcification very close to the stem cell (II) and near to the stem cell (I). Trichoblasts on the top the laterals are shown on the upper two whorls. The tangential section (B) shows the sections of the proximal (upper two whorls) and distal parts of the laterals (lower whorl). |
New diagnosis of the species Physoporella pauciforata (G�mbel): The thallus is cylindrical, rarely slightly claviform, with a smooth or slightly wrinkled inner surface, that reflects the outline of the stem cell. Calcification usually reaches close to the stem cell (exceptionally next to it), except when the proximal parts of the laterals are compressed together. The shape of the laterals is piriform, and is characterized by a wide rounded proximal part and a gradual thinning towards the distal end, similar to the shape of onion or turnip. The distal end can also be rounded, so it resembles a pear to some extent. The laterals within one whorl mostly touch each other, and the laterals of adjacent whorls stand mostly one above the other, although it is possible that due to the compactness this regular arrangement is disturbed. The pronounced variability within this species that are interpreted as varieties is a reflection mainly of the variability in the inclination of the laterals and especially the trichoblasts, and the spacing of the whorls. To a lesser extent it is a reflection of the variability of the shape of the laterals, dimensions of the algae, and intensity of the calcification. These factors are mostly reflected in the ornamentation of the outer surface of the thallus that can be more or less undulated and furrowed.
The following table compares the individual varieties:
| variety | stem cell | laterals | trichoblasts | whorls | calcification |
| pauciforata | smooth or wrinkled | perp. to sl. oblique | oblique | spaced | stronger |
| sulcata | smooth or wrinkled | oblique | oblique | spaced | stronger |
| undulata | smooth | perp. (rarely sl. obl.) | perpendicular | spaced | weaker |
| gemerica | smooth? | perp. (rarely sl. obl.) | oblique | compressed | weaker |
Similarities and differences: Infraspecific variability has been described in previous chapters. Physoporella pauciforata is similar in the shape of laterals to Oligoporella pilosa, but the later has double whorls, so it is easy to distinguish from it in oblique and longitudinal sections. The problem is cross-sections in which differentiation is difficult or impossible, so such forms are most often referred to as Physoporella sp. (Pl. 1 ![]() , fig. 14; Pl. 19
, fig. 14; Pl. 19 ![]() , fig. 5) or Oligoporella sp. (Pl. 23
, fig. 5) or Oligoporella sp. (Pl. 23 ![]() , fig. 5). It differs from other species of the genus Physoporella by the characteristic shape of its laterals. P. pauciforata var. undulata is somewhat similar to P. croatica, from which it differs by the spaced whorls, the larger number of laterals in the whorl and their different appearance, as laterals are not so distinctly compressed together.
, fig. 5). It differs from other species of the genus Physoporella by the characteristic shape of its laterals. P. pauciforata var. undulata is somewhat similar to P. croatica, from which it differs by the spaced whorls, the larger number of laterals in the whorl and their different appearance, as laterals are not so distinctly compressed together.
5.1.2. Physoporella croatica Herak, 1958
Pl. 4 ![]() , figs. 1-2, 5, 7-9; Pl. 19
, figs. 1-2, 5, 7-9; Pl. 19 ![]() , figs. 1-3, 5
, figs. 1-3, 5
1958 (nom. nud.) - Physoporella croatica n. sp. - Herak, p. 61-67, Figs. 2, no. 1, 2 (= Pl. 14 ![]() , fig. 4 right), 3-5, 6 (= ? Pl. 14
, fig. 4 right), 3-5, 6 (= ? Pl. 14 ![]() , fig. 3 below left), 7 (= Pl. 14
, fig. 3 below left), 7 (= Pl. 14 ![]() , fig. 1), 8, 9 (= Pl. 14
, fig. 1), 8, 9 (= Pl. 14 ![]() , fig. 3 right), 10 (= Pl. 14
, fig. 3 right), 10 (= Pl. 14 ![]() , fig. 2 left), 11 (= Pl. 14
, fig. 2 left), 11 (= Pl. 14 ![]() , fig. 2 right), 12 (= Pl. 14
, fig. 2 right), 12 (= Pl. 14 ![]() , fig. 5); Pl. 14
, fig. 5); Pl. 14 ![]() , figs. 1-5.
, figs. 1-5.
1983 (nom. nud.) - Physoporella croatica - Bystrick�, p. 507-515, Pl. I, figs. 1-4; Pl. II, fig. 11, Pl. III, figs. 12-16; Pl. V, figs. 24-25.
1983 (nom. nud.) - Physoporella aff. croatica - Bystrick�, p. 523-531, Pl. II, figs. 5-10; Pl. III, fig., 18; Pl. V, figs. 26-27.
Origin of the name: After the country in which it was found - Croatia.
Studied material: The type material originates from the outcrops of the Anisian limestone near the hamlet of Miljevići, on the road between Bajići and Dobrijevići, W from Golubić, NW from Knin in Croatia. Thin sections are kept in the collection of Professor Herak
at the Faculty of Science in Zagreb. Specimens from the following thin-sections were analysed and illustrated: 61 (Pl. 4 ![]() , fig. 8 = Herak, 1958, Pl. 14, fig. 1), 66 (Pl. 4
, fig. 8 = Herak, 1958, Pl. 14, fig. 1), 66 (Pl. 4 ![]() , fig. 1 = Herak, 1958, Pl. 14, fig. 2), 70 (Pl. 4
, fig. 1 = Herak, 1958, Pl. 14, fig. 2), 70 (Pl. 4 ![]() , fig. 2 = Herak, 1958, Pl. 14, fig. 3), 73 (Pl. 4
, fig. 2 = Herak, 1958, Pl. 14, fig. 3), 73 (Pl. 4 ![]() , fig. 7 = Herak, 1958, Pl. 14
, fig. 7 = Herak, 1958, Pl. 14 ![]() , fig. 4), 74 (Pl. 4
, fig. 4), 74 (Pl. 4 ![]() , fig. 5 = Herak, 1958, Pl. 14, fig. 5). I was not able to identify the thin-section for the specimen from Pl. 4
, fig. 5 = Herak, 1958, Pl. 14, fig. 5). I was not able to identify the thin-section for the specimen from Pl. 4 ![]() , fig. 9 (= Herak, 1958, Fig. 2.8). The specimen from Pl. 4
, fig. 9 (= Herak, 1958, Fig. 2.8). The specimen from Pl. 4 ![]() , fig. 1 right (= Herak, 1958, Pl. 14, fig. 2 right) was identified as a lectotype by Herak (in Granier & Deloffre, 1995, p. 57). All photos (Pl. 4) are made from original films and glass photographic plates of Prof. Herak. The research on the type material was supplemented by specimens that Dr. Branko Sokač collected at the Jabuka site near Trilj in Dalmatia, Croatia (Pl. 19
, fig. 1 right (= Herak, 1958, Pl. 14, fig. 2 right) was identified as a lectotype by Herak (in Granier & Deloffre, 1995, p. 57). All photos (Pl. 4) are made from original films and glass photographic plates of Prof. Herak. The research on the type material was supplemented by specimens that Dr. Branko Sokač collected at the Jabuka site near Trilj in Dalmatia, Croatia (Pl. 19 ![]() , figs. 1-5).
, figs. 1-5).
History of investigations: Herak (1958, p. 61) gives a description of a new species: "The stem is elongated; in the cross section predominantly polygonal and generally isodiametrical. The branches are closed at their distal end. The calcareous wall is very thin. The proximal part of each branch is large and lies with a broad base on the stem. The branches are pressed together and therefore polygonal in the cross section of the whorl. The width of the proximal part of the branch, in the section perpendicular to the main stem, is narrower than in the section parallel to the main stem. This concerns especially its base. The distal cone-like part of the branch is tapered and covered with a thin calcareous wall. The branches are situated in whorls consisting of only one row. Some whorls become somewhat irregular. In most cases there are 7 branches in one whorl. One specimen of our material has only five branches (Herak, 1958, Fig. 2.5; mistyped 2.7 in the text). Most probably it is a section of an end portion of the stem. Two oblique, nearly cross sections show more than 7 branches, but not strictly in one plane (Herak, 1958, Fig. 2.6-7; mistyped 2.5-6 in the text)."
Bystrick� (1983, p. 514) gives a new diagnosis, i.e., a supplement to the description of this alga: "The thallus is inarticulated, morphologically considerably disserted, of an irregular transversal section. The calcareous sleeve consists of only a very thin layer, covering each branch individually. Branches of the pyripherous type are pressed in their proximal part and vertically prolonged, but their distal parts ending often in prolonged hair reaching to the outer surface of the sleeve are of a round section. The branches are arranged in one-rowed whorls perpendicular to the plant axis and they are separated from each other only by a very thin layer of the calcareous sleeve. The central stem is of a round section and in correlation with the outer diameter of the thallus of a small diameter."
In the same paper, Bystrick� (1983, p. 523-524, 530-531) describes specimens which he designates as Physoporella aff. croatica. According to the results of statistical analysis, these specimens are much closer to P. pauciforata var. undulata than to P. croatica, but due to similarities in morphology and structure it is designated as Physoporella aff. croatica (and not P. aff. pauciforata). In my opinion (studying photographs, and not statistical graphs), these are only slightly larger, but typical specimens of Physoporella croatica.
Description: This species is well investigated and there is not much to add. The thallus is cylindrical and mostly very long, straight (Pl. 4 ![]() , figs. 1-2, 5, 9; Pl. 19
, figs. 1-2, 5, 9; Pl. 19 ![]() , fig. 1) or more or less curved (Pl. 19
, fig. 1) or more or less curved (Pl. 19 ![]() , figs. 2-3). Due to the densely distributed large piriform laterals, the outer surface of the thallus is covered with large protrusions - horns (Fig. 7
, figs. 2-3). Due to the densely distributed large piriform laterals, the outer surface of the thallus is covered with large protrusions - horns (Fig. 7 ![]() ). In the cross section, the inner cavity is round (Pl. 4
). In the cross section, the inner cavity is round (Pl. 4 ![]() , fig. 7; Pl. 19
, fig. 7; Pl. 19 ![]() , fig. 5; Bystrick�, 1983), and sometimes barely polygonal (Pl. 4
, fig. 5; Bystrick�, 1983), and sometimes barely polygonal (Pl. 4 ![]() , fig. 8). Calcification generally extends all the way to the stem cell. The type of calcification in this alga is very peculiar. In the type material (Pl. 4
, fig. 8). Calcification generally extends all the way to the stem cell. The type of calcification in this alga is very peculiar. In the type material (Pl. 4 ![]() , figs. 1-2, 5, 7-9) a relatively thin dark sheath is seen around each lateral ("individual sheath" according to De Castro, 1997), both on the outside and on the inside next to the stem cell. At higher magnification, it can be seen that these are very thin prismatic calcite crystals arranged perpendicular to the edge. The uniform thickness and regular occurrence of this sheath indicate that it is of primary origin. On the outside of the laterals, another type of microsparite crystal shell has been developed, which is slightly lighter in shade and thicker than the first type. The material of Bystrick� (1983, Pl. I, Pl. II, fig. 11; Pl. III, figs. 12-16; Pl. V, figs. 24-25) is very similar, except that the sheath of this first type is somewhat thicker, and along the edges a typical micritization rim is observed. In slightly larger specimens (Physoporella aff. croatica, see above; Bystrick�, 1983, Pl. II, figs. 1-10; Pl. III, fig., 18; Pl. V, figs. 26-27), the inner side of the laterals is not calcified. The second type of sheath is partially visible only on Bystrick� (1983, Pl. V, fig. 25), but it is possible that it was developed on other specimens as well, and due to its small thickness and recrystallization it merged with the first envelope. The same is the case with specimens from the vicinity of Trilj (Pl. 19
, figs. 1-2, 5, 7-9) a relatively thin dark sheath is seen around each lateral ("individual sheath" according to De Castro, 1997), both on the outside and on the inside next to the stem cell. At higher magnification, it can be seen that these are very thin prismatic calcite crystals arranged perpendicular to the edge. The uniform thickness and regular occurrence of this sheath indicate that it is of primary origin. On the outside of the laterals, another type of microsparite crystal shell has been developed, which is slightly lighter in shade and thicker than the first type. The material of Bystrick� (1983, Pl. I, Pl. II, fig. 11; Pl. III, figs. 12-16; Pl. V, figs. 24-25) is very similar, except that the sheath of this first type is somewhat thicker, and along the edges a typical micritization rim is observed. In slightly larger specimens (Physoporella aff. croatica, see above; Bystrick�, 1983, Pl. II, figs. 1-10; Pl. III, fig., 18; Pl. V, figs. 26-27), the inner side of the laterals is not calcified. The second type of sheath is partially visible only on Bystrick� (1983, Pl. V, fig. 25), but it is possible that it was developed on other specimens as well, and due to its small thickness and recrystallization it merged with the first envelope. The same is the case with specimens from the vicinity of Trilj (Pl. 19 ![]() , figs. 1-5), where the calcareous thallus is built of a fine-crystalline mosaic of calcite with a micritization rim, formed by recrystallization of the primary calcareous envelopes. In these specimens, the inner parts of the laterals were only partially calcified.
, figs. 1-5), where the calcareous thallus is built of a fine-crystalline mosaic of calcite with a micritization rim, formed by recrystallization of the primary calcareous envelopes. In these specimens, the inner parts of the laterals were only partially calcified.
The laterals are large, of piriform type and arranged in compact whorls. The number of laterals in the whorl is relatively small (mostly 7-9). A distinct feature of this species is that the laterals in the proximal part are strongly compressed, both inside the whorl and between adjacent whorls. Therefore, the laterals in cross section (Pl. 4 ![]() , figs. 7-8) have a conical proximal part, while distally they taper uniformly towards the top. In adjacent whorls, the laterals usually stand alternately, so due to compaction; in deep tangential-longitudinal sections they have a slightly polygonal (hexagonal) appearance (Pl. 4
, figs. 7-8) have a conical proximal part, while distally they taper uniformly towards the top. In adjacent whorls, the laterals usually stand alternately, so due to compaction; in deep tangential-longitudinal sections they have a slightly polygonal (hexagonal) appearance (Pl. 4 ![]() , fig. 2; Pl. 19
, fig. 2; Pl. 19 ![]() , fig. 3; Bystrick�, 1983, Pl. I, fig. 2 below), but might also be one above the other (Pl. 19
, fig. 3; Bystrick�, 1983, Pl. I, fig. 2 below), but might also be one above the other (Pl. 19 ![]() , fig. 2 above). The laterals are arranged perpendicular to the thallus axis. The laterals are also longer than wide, and is clearly visible in longitudinal sections (Pl. 4
, fig. 2 above). The laterals are arranged perpendicular to the thallus axis. The laterals are also longer than wide, and is clearly visible in longitudinal sections (Pl. 4 ![]() , figs. 1-2, 5; Pl. 19
, figs. 1-2, 5; Pl. 19 ![]() , figs. 2-3). In the specimens from Trilj (Pl. 19
, figs. 2-3). In the specimens from Trilj (Pl. 19 ![]() , figs. 1-3), the laterals are slightly asymmetric, so that the distal thinning is not in the axis of the lateral, but slightly above it. The laterals were connected to the stem cell probably through a narrow pore that is not observed in the fossil material, either because it was very narrow, or it was closed during diagenesis. The laterals extend into the trichoblast (Pl. 19
, figs. 1-3), the laterals are slightly asymmetric, so that the distal thinning is not in the axis of the lateral, but slightly above it. The laterals were connected to the stem cell probably through a narrow pore that is not observed in the fossil material, either because it was very narrow, or it was closed during diagenesis. The laterals extend into the trichoblast (Pl. 19 ![]() , figs. 1-2), as in other species. Trichoblasts probably arose from the top of the lateral in tufts of two (or more?) (Pl. 19
, figs. 1-2), as in other species. Trichoblasts probably arose from the top of the lateral in tufts of two (or more?) (Pl. 19 ![]() , fig. 1 below).
, fig. 1 below).
The specimen in Pl. 19 ![]() , fig. 4 (determined as P. cf. croatica) somewhat differs from the others. The laterals are not of a typical piriform shape, but have a pronounced bulge on their underside. Since the other characteristics are identical to those of P. croatica, and it is found with it in the same sample, it does not seem likely that it is another (new) taxon, but rather a variation within the species.
, fig. 4 (determined as P. cf. croatica) somewhat differs from the others. The laterals are not of a typical piriform shape, but have a pronounced bulge on their underside. Since the other characteristics are identical to those of P. croatica, and it is found with it in the same sample, it does not seem likely that it is another (new) taxon, but rather a variation within the species.
Emended diagnosis: The thallus is cylindrical with a smooth inner surface and, since the alga has densely arranged large piriform laterals, a horned (prickly) outer surface. Calcareous thallus consists of two calcareous sheets. The first one is very thin and wraps individually each lateral on the outer side, and often on the inner side next to the stem cell. The second one is much thicker and envelops the alga on the outside. After diagenetic changes, these sheets can merge into one. The laterals are large piriform, compressed together within the whorl and between adjacent whorls, so that their tangential section is polygonal. They gradually taper out distally and carry trichoblasts on the top. The laterals are arranged perpendicular to the thallus axis and mostly alternately in adjacent whorls, although they may be one above the other.
Similarities and differences: Due to its peculiar appearance, similarity exists only with P. pauciforata var. undulata, from which it differs by its distinctly compressed laterals, which are higher than wide, dense whorls, a smaller number of laterals in the whorl, and a narrower central cavity.
|
|
Figure 7: Physoporella croatica Herak. Idealized longitudinal (A), transverse-very slightly oblique (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows variants with a single-layer (I) and a double-layer (II) calcareous sheet (the thickness of the inner layer is enlarged). Trichoblasts are shown on some laterals. |
5.1.3. Physoporella leptotheca Kochansky-Devid�, 1967
Pl. 16 ![]() , figs. 11-12, 14
, figs. 11-12, 14
1958 - "Physoporella lotharingica" - Ellenberger, p. 194, Fig. 11.
1967 - Physoporella leptotheca n. sp. - Kochansky-Devid�, p. 171-174, Pl. I, figs. 1-12.
1979 - Physoporella leptotheca.- Fois, p. 63-65, Pl. 3, figs. 1-5, 8; Pl. 5, figs. 1-13.
1983 - Physoporella leptotheca.- Fois & Jadoul, p. 13, 15, 20-22, Pl. 1, figs. c pars, d.2, f.1.
Origin of the name: After a thin calcareous shell (Greek "leptos" = thin, delicate; Latin "theca" = shell).
Studied material: The holotype of Physoporella leptotheca (Kochansky-Devid� (1967, Pl. I, fig. 1 above = this work, Pl. 16 ![]() , fig. 14) is in thin-section J 173. I also illustrate a specimen from sample J 178 (Kochansky-Devid� (1967, Pl. I, figs. 3-4 = this work, Pl. 16
, fig. 14) is in thin-section J 173. I also illustrate a specimen from sample J 178 (Kochansky-Devid� (1967, Pl. I, figs. 3-4 = this work, Pl. 16 ![]() , figs. 12, 11). Thin-section J 173 is kept at the Department of Geology and Paleontology, University of Ljubljana, with other thin-sections (J 174, 175, 176, 178 and 518) at the Technical Museum in Jesenice. The secondary sample,
from which the thin-sections were made, was found in Jermenov potok in the �eleznica valley, north of Gozd-Martuljek in the Karavanke (Slovenia). The
age of the material is unknown.
, figs. 12, 11). Thin-section J 173 is kept at the Department of Geology and Paleontology, University of Ljubljana, with other thin-sections (J 174, 175, 176, 178 and 518) at the Technical Museum in Jesenice. The secondary sample,
from which the thin-sections were made, was found in Jermenov potok in the �eleznica valley, north of Gozd-Martuljek in the Karavanke (Slovenia). The
age of the material is unknown.
History of investigations: Kochansky-Devid� (1967, p. 172) diagnoses a new species of Physoporella leptotheca: "A Physoporella with very thin walls, very swollen rings with deeply serrated edges, each with a relatively irregular circle of branches (pores) protruding at regular, widely spaced distances."
(In German: "Eine Physoporella mit sehr d�nnen Wanden. In regelmassigen, weit voneinander stehenden Abstanden ragen sehr hervorspringende Ringe mit tief gezacktem Rand und mit je einem verh�ltnism��ig unregelm��igen �stchenkreis hervor.")
In the description she adds: "In longitudinal section, the rings on each side of the tube are seen as teeth; they protrude quite abruptly above the wall of the tube. The teeth are of different heights depending on whether the cross-section hit the ring or between the rings. Exceptionally, the tooth can fall right in the place of the tube. From this we can conclude that the "rings" are very deeply serrated, even broken in places. The pirifer euspondile pores lie parallel to the transverse plane and obliquely to the radial plane, usually with the two together in a divergent position. That is why we often find obliquely cut pores in axial sections� In tangential sections it can be seen that the pores often stand rather irregularly; often pairs of pores stand slightly obliquely. A total of five examples of 3 pores in one bundle were observed in the available material."
Kochansky-Devid� and Gu�ić (1971, Fig. 5 pars) give a reconstruction of P. leptotheca, as an illustration of a "horned" alga.
Fois (1979, p. 63), on the basis of rich material (idem, Pl. 3, figs. 1-5, 8; Pl. 5, figs. 1-13) gives a new diagnosis of the species: "Thallus cylindrical, originally flexible, with very thin wall and whorled branches. Distances between the whorls are variable. Max. size: 7 mm long and 1 mm wide."
In description she adds: "Thallus: Long, thin, cylindrical, with low calcification and a very thin wall. The tubes are straight, rarely bending. In longitudinal section the skeleton shows many prominent rings corresponding to the whorls. Branches have a typical triangular section sharply connected to the wall. Generally whorls are well spaced (1 mm in average), however some specimens show very close whorls with a more irregular branch arrangement. Sometimes in the same specimen, the distance between the whorls may vary. The inner surface of the central stem is smooth and straight. Transversal sections are circular with a very low D/L ratio. Branches: Single whorled, metaspondyle, trichophorous or piriferous type; they are open through a thin canal which can be frequently occluded by recrystallization. Branches of a single whorl can be differently spaced. Branches insert perpendicularly on the central stem or are slightly bended upwards."
Specimens described by Braga (1986, p. 242, 244), probably prompted by the presumed late Triassic age, under the name of P. leptotheca do not belong to this species. The laterals have a more rounded shape, so this alga probably belongs to the genus Neophysoporella n. gen., and is described in this work as Neophysoporella n. sp. (see chapter 5.3.4).
Specimens described by Parente and Climaco (1999, p. 178-179, Pl. 40, figs. 9-10) from the Norian of Monte Rotonda as Physoporella aff. leptotheca probably do not belong to this species, since they have very thin walls and a roughly triangular shape to the laterals.
Description: The thallus is cylindrical and mostly straight (Pl. 16 ![]() , fig. 12; Kochansky-Devid�, 1967, Pl. I, fig. 2; Fois, 1979, Pl. 3, fig. 8; Pl. 5, figs. 1-6), and rarely slightly bent (Fois, 1979, Pl. 5, fig. 9). The inner wall of the thallus is completely flat and smooth (Pl. 16
, fig. 12; Kochansky-Devid�, 1967, Pl. I, fig. 2; Fois, 1979, Pl. 3, fig. 8; Pl. 5, figs. 1-6), and rarely slightly bent (Fois, 1979, Pl. 5, fig. 9). The inner wall of the thallus is completely flat and smooth (Pl. 16 ![]() , figs. 12, 14; Kochansky-Devid�, 1967, Pl. I, figs. 1-12; Fois, 1979, Pl. 3, figs. 2, 4; Pl. 5, figs. 11, 13). Calcification in the specimens illustrated by Kochansky-Devid� (1967, Pl. I) reached the stem cell or very close to it, as indicated by the completely preserved laterals. In the specimens illustrated by Fois (1979, Pl. 3, figs. 2-4) the calcification did not reach the stem cell, and in some specimens (Fois, 1979, Tab. 3, Figs. 5, 8) the inner wall was uneven due to dissolution during diagenesis. The calcareous wall is relatively thin and envelops the stem cell and laterals uniformly. Therefore, the outer surface of the thallus is extremely embossed, and it could be considered annulated, as the laterals protrude outwards (Fig. 8
, figs. 12, 14; Kochansky-Devid�, 1967, Pl. I, figs. 1-12; Fois, 1979, Pl. 3, figs. 2, 4; Pl. 5, figs. 11, 13). Calcification in the specimens illustrated by Kochansky-Devid� (1967, Pl. I) reached the stem cell or very close to it, as indicated by the completely preserved laterals. In the specimens illustrated by Fois (1979, Pl. 3, figs. 2-4) the calcification did not reach the stem cell, and in some specimens (Fois, 1979, Tab. 3, Figs. 5, 8) the inner wall was uneven due to dissolution during diagenesis. The calcareous wall is relatively thin and envelops the stem cell and laterals uniformly. Therefore, the outer surface of the thallus is extremely embossed, and it could be considered annulated, as the laterals protrude outwards (Fig. 8 ![]() ). The conclusion of Fois (1979, p. 63) that the thallus was originally flexible cannot be accepted, because a rigid calcareous wall enveloped the entire algae. Algae are flexible only before they are calcified, except in articulated algae (e.g., Mizzia velebitana, Diplopora nodosa, etc.).
). The conclusion of Fois (1979, p. 63) that the thallus was originally flexible cannot be accepted, because a rigid calcareous wall enveloped the entire algae. Algae are flexible only before they are calcified, except in articulated algae (e.g., Mizzia velebitana, Diplopora nodosa, etc.).
The laterals are typically piriform with a broad proximal part and gradually thin outwards, which is nicely seen on all illustrated specimens. Thanks to the calcification of the laterals all the way to the stem cell, it can be seen that they were connected to the stem cell by a very thin and short peduncle (Kochansky-Devid�, 1967, Pl. I, fig. 8 left). As with other species of this genus, they probably continued distally into a thin trichoblast, which most likely fell off during the life of the algae.
The laterals are arranged in spaced single-rowed whorls. The
main characteristic of this species is the irregular arrangement of laterals in the whorl and the uneven spacing of the whorls. Laterals most often occur in groups of two or three (Pl. 16 ![]() , figs. 11, 14; Kochansky-Devid�, 1967, Pl. I, figs. 5, 7, 12; Fois, 1979, Pl. 3, figs. 3-4). I do not consider this convergence of laterals to be metaspondility (as stated by Fois, 1979), as for example in Diplopora sinica (Bucur & Enos, 2001), where the laterals emerge from the stem cell almost from one point, i.e., their bases are very close to each other, unlike in Physoporella leptotheca, where laterals emerging points are clearly separated (Kochansky-Devid�, 1967, Pl. I, fig. 12). It has no affinity with the family Diploporaceae, but its association with other Physoporellae is obvious. This grouping of laterals into "twos" or "threes" just represents the relatively irregular structure of this alga, somewhat similar to Clypeina bucuri Barattolo & Romano, 2002 (see Sokač et al., 2012). The laterals are very often not symmetrically arranged within the whorl, but are located only on one side of the thallus (Kochansky-Devid�, 1967, Pl. I, fig. 6) or there are gaps between individual groups of laterals (Kochansky-Devid�, 1967, Pl. I, fig. 12). In longitudinal sections, this irregularity is expressed in the way that the laterals appear only on one side of the thallus, and not on the other (Pl. 16
, figs. 11, 14; Kochansky-Devid�, 1967, Pl. I, figs. 5, 7, 12; Fois, 1979, Pl. 3, figs. 3-4). I do not consider this convergence of laterals to be metaspondility (as stated by Fois, 1979), as for example in Diplopora sinica (Bucur & Enos, 2001), where the laterals emerge from the stem cell almost from one point, i.e., their bases are very close to each other, unlike in Physoporella leptotheca, where laterals emerging points are clearly separated (Kochansky-Devid�, 1967, Pl. I, fig. 12). It has no affinity with the family Diploporaceae, but its association with other Physoporellae is obvious. This grouping of laterals into "twos" or "threes" just represents the relatively irregular structure of this alga, somewhat similar to Clypeina bucuri Barattolo & Romano, 2002 (see Sokač et al., 2012). The laterals are very often not symmetrically arranged within the whorl, but are located only on one side of the thallus (Kochansky-Devid�, 1967, Pl. I, fig. 6) or there are gaps between individual groups of laterals (Kochansky-Devid�, 1967, Pl. I, fig. 12). In longitudinal sections, this irregularity is expressed in the way that the laterals appear only on one side of the thallus, and not on the other (Pl. 16 ![]() , fig. 12). The isolated silicified specimens illustrated by Fois (1979, Pl. 5) show that not all laterals are retained in groups but that there are specimens in which individual laterals predominate (idem, Pl. 5, figs. 1-4). In these specimens, a great variability in the spacing of the whorls is observed, and the arrangement of the laterals in the whorls is somewhat denser than in the material from Slovenia.
, fig. 12). The isolated silicified specimens illustrated by Fois (1979, Pl. 5) show that not all laterals are retained in groups but that there are specimens in which individual laterals predominate (idem, Pl. 5, figs. 1-4). In these specimens, a great variability in the spacing of the whorls is observed, and the arrangement of the laterals in the whorls is somewhat denser than in the material from Slovenia.
Emended diagnosis: Cylindrical thallus with smooth inner wall and relatively wide central cavity. Calcification in a relatively thin layer covers the stem cell and individual laterals, and consequently the outer surface of the thallus is extremely embossed. The laterals are typical piriform, located often in groups of two or three and usually irregularly distributed within the spaced whorls. They are laid perpendicular to the thallus axis, rarely slightly obliquely upwards. The number of laterals in the whorl and the distance between the whorls varies greatly.
Similarities and differences: It differs from other species of the genus Physoporella by the grouping of laterals into groups of two or three and by their irregular arrangement within the whorl. From the morphologically somewhat similar species of the genus Neophysoporella n. gen. it differs, aside from the mentioned features, by its elongated piriform laterals and more spaced whorls.
|
|
Figure 8: Physoporella leptotheca Kochansky-Devid�. Idealized longitudinal (A), transverse (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows variants with calcification very close to the stem cell (I) and near to the stem cell (II). Trichoblasts on the top the laterals are shown on some laterals. The cross-section (B) shows an irregular arrangement of laterals in the whorl. The tangential section (C) shows a grouping of laterals into groups of two or three. |
5.1.4. Physoporella? heraki Bystrick�, 1967
Pl. 16 ![]() , figs. 4-5
, figs. 4-5
1967 - Physoporella heraki n. sp. - Bystrick�, p. 295-298, Pl. IX, figs. 1-3; Pl. X, figs. 1-4; Pl. XI, figs. 3-4; Pl. XIV, fig. 5.
1967 - Physoporella heraki var. tenuipora n. var. - Bystrick�, p. 298-299, Pl. XI, figs. 1-2.
1982 - Physoporella heraki - Fl�gel & Mu, p. 61, Pl. 8, fig. 4 pars; Pl. 9, figs. 5-6.
1986 - Physoporella heraki var. heraki - Bystrick�, p. 311-312, Pl. V, fig. 8.
1999 - Physoporella heraki - Velledits et al., p. 580, Pl. II, fig. 2.
Origin of the name: Dedicated to Prof. Milan Herak, student of Julius von Pia and the founder of fossil algae research in Croatia.
Studied material: Bystrick� (1967) describes Physoporella heraki with the help of numerous illustrations (idem, Pl. IX, figs. 1-3; Pl. X, figs. 1-4; Pl. XI, figs. 3-4; Pl. XIV, fig. 5). As a holotype he chose a specimen from thin-section no. 2183 (Bystrick�, 1967, Pl. IX, fig. 1 = this paper, Pl. 16 ![]() , fig. 4). Bystrick� (1967, tab. XI, figs. 1-2) also described the variety Physoporella heraki var. tenuipora. As a holotype he chose a specimen from thin-section no. 2804 (idem, Pl. XI, fig. 2 = this paper, Pl. 16
, fig. 4). Bystrick� (1967, tab. XI, figs. 1-2) also described the variety Physoporella heraki var. tenuipora. As a holotype he chose a specimen from thin-section no. 2804 (idem, Pl. XI, fig. 2 = this paper, Pl. 16 ![]() , fig. 5). The type material is kept in the collection of the Geological Department of the Slovak Academy of Sciences in Bratislava. Since the original material was not available to me, I based the description on the published illustrations.
, fig. 5). The type material is kept in the collection of the Geological Department of the Slovak Academy of Sciences in Bratislava. Since the original material was not available to me, I based the description on the published illustrations.
History of investigations: Bystrick� (1967, p. 295) gave diagnosis of the new species Physoporella heraki: "Unsegmented thallus with a spacious inner cavity. The outer surface is covered with outgrowths of different lengths, which are in fact a fine covering of the distal parts of branches. The branches are of the piriform type and have the distal end somewhat stretched and widened in the shape of a club. The whorls are very close to each other."
(In German: "Ungegliederte Schale mit ger�umigem innerem Hohlraum. Aussen-oberflache mit Auslaufern von verschiedener Lange bedeckt, die eine feine H�lle der distalen Teile der Wirtel�ste bilden. Die �stchen von piriferen Typus haben das distale Ende etwas gedehnt und zur Form eines Keulchens erweitert. Wirteln zueinander sehr nahestehende.")
In the same paper, Bystrick� (1967, p. 298) describe the variety Physoporella heraki var. tenuipora with the following diagnosis: "Single undivided thallus with a spacious internal cavity. The whorled branches significantly thinner than in the type species and are round in cross section."
(In German: "Einfache ungegliederte Schale mit ger�umigem Innenhohlraum. Wirtel�ste wesentlich d�nner als bei der typischen Art und von kreisrunden Durchschnit.")
Ott (1972a, p. 104-105) comments on this species: "Described from the Lower Carnian it can certainly be classified as a Physoporella based on the sole criteria of the shape of the laterals. However, it differs so much from the Anisian species of Physoporella through the wide central cavity and densely packed whorls, rising from a gap, that a relationship to these is much less likely than to young Mesozoic algae, e.g., with the almost identical Teutloporella gallaeformis or with other Dasycladaceae with a large central cavity in Upper Triassic and Jura."
Fl�gel and Mu (1982, p. 61) describe specimens of this species from the Carnian of Tibet: "Thallus cylindrical, straight, curved or slightly constricted. Branches piriform, thickened at the base and tapering towards the periphery. The branches are very gently curved upwards and are densely spaced into verticils. The horn-like distal ends of the branches are covered by a thin calcareous sheath."
Description: Calcification individually covers the distal part of each lateral in a relatively uniform layer (Pl. 16 ![]() , fig. 4 below; Bystrick�, 1967, Pl. IX, fig. 2; Pl. X, figs. 1-2; Pl. XI , fig. 3), and in the proximal part these sheaths merge into a common calcareous thallus. In Physoporella? heraki var. tenuipora (Pl. 16
, fig. 4 below; Bystrick�, 1967, Pl. IX, fig. 2; Pl. X, figs. 1-2; Pl. XI , fig. 3), and in the proximal part these sheaths merge into a common calcareous thallus. In Physoporella? heraki var. tenuipora (Pl. 16 ![]() , fig. 5) calcification, due to the narrow laterals, is less regular, so that the outer surface of the thallus is jagged. The shape of the pores resembles piriform types, but since calcification is limited to the outer part of the algae, it is not possible to determine their actual shape. It is also possible, as indicated by the very spacious central cavity, that they might be second-order laterals. That is why I included this alga in the genus Physoporella with a question mark. The laterals are bent upwards in the distal part. It is not true that in the distal part the laterals expand again. This is only an impression, because the distal parts of the laterals from the lower whorls are also cut in cross section, so their pores are sometimes fused with the proximal parts of the pores from the upper whorl. To get to know the exact structure of this alga, we need to wait for better preserved specimens.
, fig. 5) calcification, due to the narrow laterals, is less regular, so that the outer surface of the thallus is jagged. The shape of the pores resembles piriform types, but since calcification is limited to the outer part of the algae, it is not possible to determine their actual shape. It is also possible, as indicated by the very spacious central cavity, that they might be second-order laterals. That is why I included this alga in the genus Physoporella with a question mark. The laterals are bent upwards in the distal part. It is not true that in the distal part the laterals expand again. This is only an impression, because the distal parts of the laterals from the lower whorls are also cut in cross section, so their pores are sometimes fused with the proximal parts of the pores from the upper whorl. To get to know the exact structure of this alga, we need to wait for better preserved specimens.
5.1.5. Physoporella? hydrae (Richter et al., 1999), n. comb.
(= Oligoporella hydrae Richter et al., 1999)
1999 - Oligoporella hydrae n. sp. - Richter et al., p. 141, Pl. 1, figs. 3-4, 5 pars, 6 pars, 7.
Origin of the name: From Hydra Island, the type locality.
Original diagnosis: "Thallus small, cylindrical, strongly calcified, crossed by a narrow central cavity. The ramifications are large, disposed in whorls, piriform inflated to the base and very narrow to the top. In the distal part, the ramifications present a very small, circular opening. The disposition of the ramifications in the whorls is regular having a diagonal-helicospiral pattern."
Discussion: Since the whorls are single-rowed, the species was transferred to the genus Physoporella. The question mark is left because it differs from typical representatives of this genus. The difference is that the laterals are not placed symmetrically in the whorl but are "shifted" so the whorls have a "diagonal-helicospiral pattern". When looking at the longitudinal section of the holotype (Richter et al., Pl. 1, fig. 3), the seemingly piriform laterals appear to be dense bundles of thin laterals, similar to the species Euteutloporella triassina (Schauroth). These possible bundles are slightly concavely bent. It is necessary to check this by examining additional material.
5.1.6. Physoporella? elegans (Assmann ex Pia, 1931), n. comb.
(= Oligoporella elegans Assmann ex Pia, 1931; = Diplopora elegans Assmann, 1926, nom. nud.)
Pl. 16 ![]() , figs. 3, 7
, figs. 3, 7
1926 (nom. nud.) - Diplopora elegans n. sp. - Assmann, p. 505-506, no figure. Type no figure.
1931 Oligoporella elegans n. comb. - Pia, p. 273-274, 275-276, Pl. XXI, figs. 3-6.
2013 - Oligoporella elegans - Kotański 52-57, Pl. V, figs. 1-37; Pl. VI, figs. 1-50; Pl. VII, figs. 1-29; Pl. XXIX, fig. 27; Pl. XXXIII, fig. 4.c; Pl. XXXVI, figs. 2.c, 4.c; Pl. XXXVII, figs. 1.c, 2.e, 4.c; Pl. XXXVIII, figs. 1.c, 4.b.
Origin of the name: Probably after its appearance (Latin "elegans" = elegant, graceful).
Studied material: Pia (1931) does not identify the samples or thin-sections he used to illustrate this species. In the catalogue of Pia's collection (see
Chapter 2), under the name of the species the following numbers are written: DXXX, DXXXI and DXXXIV. On a piece of paper in the box with thin-sections, the following is written for thin-sections DXXX1 and DXXXI2: "Zur�ck an Preuss. Geol. Landesamst.". According to Pia (1931), he borrowed the material from Assmann
and later returned it (Assmann worked at the "Preussischen Geologischen Landesanstalt"), so I assume that the specimens illustrated in the paper were in the box. In other thin-sections: DXXX2, DXXXI3, DXXXI4, DXXXI5 and DXXXIV1 I found highly recrystallized specimen of Diplopora annulatissima, one specimen of Macroporella alpina, and extremely poorly preserved specimens of algae that could belong to the species Physoporella? elegans, but due to poor preservation, determination is impossible. Illustrations from Pl. 16 ![]() , figs. 3, 7 were copied from Pia (1931, Pl. XXI, figs. 3-4). Samples from Pia (1931, Pl. XXI, figs. 3-6) originate from the vicinity of the settlement of Nowe Koszyce (in Pia's time this area was in Germany, so it is referred to as Neu-Kosch�tz), northwest of Strzelce Opolskie in Upper Silesia, Poland.
It is assigned to the Upper Anisian due to its occurrence with D. annulatissima. Kotański (2013) place Pia's finds in the "Diplopora dolomite" of Illyrian age. Since it is not certain where the type material is stored, no lectotype can be designated. Pia's samples are possibly not from the type locality, since Assmann (1926) states "south of Tarnowitz (in Polish: Tarnowske Gōry), and Nowe Koszyce is west from it. However, it is within the type area.
, figs. 3, 7 were copied from Pia (1931, Pl. XXI, figs. 3-4). Samples from Pia (1931, Pl. XXI, figs. 3-6) originate from the vicinity of the settlement of Nowe Koszyce (in Pia's time this area was in Germany, so it is referred to as Neu-Kosch�tz), northwest of Strzelce Opolskie in Upper Silesia, Poland.
It is assigned to the Upper Anisian due to its occurrence with D. annulatissima. Kotański (2013) place Pia's finds in the "Diplopora dolomite" of Illyrian age. Since it is not certain where the type material is stored, no lectotype can be designated. Pia's samples are possibly not from the type locality, since Assmann (1926) states "south of Tarnowitz (in Polish: Tarnowske Gōry), and Nowe Koszyce is west from it. However, it is within the type area.
History of investigations: Assmann (1926, p. 505-506) describes a new species Diplopora elegans: "In the Diplopora dolomites south of Tarnowitz there are some isolated small diploporas, which differ from Diplopora annulata Schaf. sp. quite significantly. Shell cylindrical-tubular, thick-walled, mostly slightly curved, with dotted pits covering the openings of the radial canals; Radial canals quite wide and apparently arranged in four rows on each ring article; Ring articles on the outer surface usually very weakly or not at all visible, occasionally protruding like bulges; Ring height 0.4-0.5 mm, ring width 0.8-1.0 mm. The most similar relationship is with Gyroporella triasina V. Schauroth sp. and G. multiserialis G�mbel, both of which occur in the Mendoladolomit, the former also in the Alpine Muschelkalk of Recoaro. All three match well with one another in the form of the ring articles and in the shape of the shell. But they differ mainly in size, in which the Alpine species exceed the Silesian form by many times. Furthermore, the pits on the outside of the shell in Diplopora elegans are much coarser than in the other two species, where they emerge as fine points."
(In German: "In den Diploporendolomiten s�dlich von Tarnowitz kommen vereinzelt kleine Diploporen vor, die sich von Diplopora annulata Schaf. sp. recht wesentlich unterscheiden. Geh�use zylindrisch-r�hrenf�rmig, dickwandig, meist etwas gekr�mmt, mit Punktgr�bchen, den Ausm�ndungen der Radialkan�lchen, vollkommen bedeckt; Radialkan�lchen ziemlich weit und an scheinend in 4 Reihen auf jedem Ringglied angeordnet; Ringglieder an der Au�enfl�che meist sehr schwach oder garnicht sichtbar, gelegentlich etwas wulstartig hervortretend; Ringh�he 0,4-.0,5 mm, Ringbreite 0,8-1,0 mm. Die meisten verwandtschaftlichen Beziehungen bestehen zu Gyroporella triasina V. Schauroth sp. und G. multiserialis G�mbel die beide im Mendoladolomit, die erstere auch noch im alpinen Muschelkalk von Recoaro vorkommen. Alle drei stimmen in der Ausbildung der Ringglieder und in der Form des Geh�uses gut miteinander �berein. Sie unterscheiden sich aber vor allem in der Gr��e, worin die alpinen Arten die schlesische Form um ein mehrfaches �bertreffen. Ferner sind die Gr�bchen auf der Au�enseite der Schale bei Diplopora elegans viel gr�ber als bei den beiden anderen Arten, an denen sie, mehr als feine Punkte hervortreten.")
Since Assmann (1926) did not attach any illustration or figure to his description, it is not valid (Art. 43.2. ICN, Turland et al., 2018).
Pia (1931) attaches illustrations of this species and thus makes it valid, changing the name of this species to Oligoporella elegans Assmann, and giving the following description (idem, p. 273-274): "From site 4 I have a favourably weathered specimen of the species determined by Assmann. You can see from the stone core that the pores are filled from the stem cell to the surrounding rock. The pores were therefore open to the outside, not closed, as in Physoporella lotharingica. The total diameter of the shell is about 0.8 mm. There are 4 whorls with a length of 0.7 mm. The average distance between the whorls is 0.175 mm. The specimens in thin-sections (Pia, 1931,
Pl. 21, figs. 3-6) agree well with this specimen. You can tell from them that the whorls are in one row. The number of branches in a whorl apparently changes quite a lot and cannot be taken with certainty from the small number of sufficiently well-preserved sections, but it should be around 12-20. The direction of the pores is usually clearly oblique. The shell is often indented a little between the individual whorls, but regular undulation only occurs in exceptional cases. The pores widen slightly towards the outside in some sections (Pia, 1931, Pl. 21 ![]() , fig. 6). Olig. elegans is obviously very close to Olig. prisca (Pia, 1912, p. 42). The alpine species is larger on average, so that its smallest specimens are roughly equal to the largest of the Silesian ones. The inner cavity in Olig. prisca is relatively narrower and the whorls are closer together. They are likely to be more numerous on average. Of course, these are not very important features. The occasional widening of the pores towards the outside is common to both types. For the time being, however, it should be better to keep them separate, at least as long as we do not know any sites with transitional specimens. Because Olig. elegans occurs in several places in the same sections with Diplop. annulatissima, we shall regard them as upper Anisian."
, fig. 6). Olig. elegans is obviously very close to Olig. prisca (Pia, 1912, p. 42). The alpine species is larger on average, so that its smallest specimens are roughly equal to the largest of the Silesian ones. The inner cavity in Olig. prisca is relatively narrower and the whorls are closer together. They are likely to be more numerous on average. Of course, these are not very important features. The occasional widening of the pores towards the outside is common to both types. For the time being, however, it should be better to keep them separate, at least as long as we do not know any sites with transitional specimens. Because Olig. elegans occurs in several places in the same sections with Diplop. annulatissima, we shall regard them as upper Anisian."
(In German: "Hieher vielleicht auch alle oder die meisten oberschlesischen St�cke, die beschrieben wurden als Gyroporella pauciforata, Gyr. minutula, Gyr. silesiaca, Cylindrella silesiaca. Von Fundort 4 liegt mir ein durch Assmann bestimmtes, ziemlich g�nstig ausgewittertes Exemplar der Art vor. Man sieht an dem Steinkern, da� die Ausf�llungen der Poren von der Stammzelle bis zum umh�llenden Gestein durchgehen. Die Poren waren also gegen au�en offen, nicht geschlossen, wie bei Physoporella lotharingica. Der Gesamtdurchmesser der Schale betr�gt etwa 0.8 mm. Es kommen 4 Wirtel auf eine L�nge von 0.7 mm. Der durchschnittliche Abstand der Wirtel ist also 0.175 mm. Mit diesem Befund stimmt der an den Schliffen (Taf. 21, Fig. 3-6) gut �berein. Man erkennt an ihnen, da� die Wirtel einreihig sind. Die Zahl der �ste in einem Wirtel wechselt offenbar ziemlich stark und ist bei der geringen Menge hinl�nglich gut erhaltener Schnitte nicht sicher zu entnehmen, d�rfte aber etwa 12-20 betragen. Die Richtung der Poren ist meist deutlich schr�g. �fter ist die Schale zwischen den einzelnen Wirteln etwas eingezogen, aber eine regelm��ige Wellung kommt nur ausnahmsweise zustande. Die Poren erweitern sich an manchen Schnitten gegen au�en deutlich etwas (Taf. 21, Fig. 6). Die Abmessungen der abgebildeten St�cke sind aus der folgenden Aufstellung zu entnehmen. Olig. elegans steht offensichtlich der Olig. prisca recht nahe (Pia, 1912, S. 42). Die alpine Art ist im Durchschnitt gr��er, so da� ihre kleinsten St�cke etwa den gr��ten der schlesischen gleichkommen. Der innere Hohlraum ist bei Olig. prisca verh�ltnism��ig enger und die Wirtel stehen n�her beisammen. Sie d�rften bei ihr im Durchschnitt wohl reicher sein. Das sind freilich durchwegs keine sehr wichtigen Merkmale. Die gelegentliche Erweiterung der Poren gegen au�en ist beiden Arten gemeinsam. Vorl�ufig d�rfte es aber doch besser sein, sie getrennt zu halten, wenigstens so lange wir keine Fundorte mit vermittelnden St�cken kennen. Da Olig. elegans an mehreren Stellen in denselben Schliffen wie Diplop. annulatissima auftritt, werden wir sie f�r oberanisisch ansehen.")
Kotański (2013, p. 53) gives a new diagnosis: "Long and narrow central stem with thin calcareous wall. Pores are perpendicular or slightly oblique to the wall and are open from the outside. They are arranged in single whorls. The distance between whorls is rather large. The number of pores in one whorl is about 12-20. The shape of the pores is generally trichoform (wedge-like), but one can observe the enlargement of pores in some thin sections." He illustrates quite an extensive material, both from thin sections (idem, Pls. V-VI) and from isolated specimens of moulds and imprints of algae with tubercles instead of pores (idem, Pl. VII; see Chapter 5.2.9.). Specimens from thin-sections are very poorly preserved (due to diagenetic processes described by Kotański, 2013) and the photographs are very small, so it is very difficult to compare them with Pia's illustrations, but we have to trust the author with such an experience in investigation of Upper Silesian algae. Some of them have phloioform laterals (idem, Pl. 5, figs. 6, 29) and belong to a different genus. Kotański (2013) probably include the specimens (also in diagnosis) following Pia's description of the similar alga O. prisca, supposedly having phloioform laterals in the basal part that has been disproved here (see Chapter 5.5.2.). Isolated specimens are better preserved, especially the specimen from Kotański (2013, Pl. VII, fig. 1) that resembles that from Pia (1931, Pl. XXI, fig. 3). The synonymy list of O. elegans also includes Cylindrella silesiaca G�mbel, 1872. This cannot be accepted since it is significantly larger than Physoporella? elegans and has a pronounced intusannulation.
Description: The thallus is cylindrical and straight, and relatively narrow. The inner wall is smooth, except in places where it is secondarily dissolved, while the outer surface appears uneven - serrated. The laterals are slightly oblique and are arranged in relatively dense whorls. It seems that in adjacent whorls they can be arranged alternately or one above the other (Pl. 16 ![]() , fig. 7). Due to the poor preservation of the material and the absence of photographs (Pia gave drawings), it is very difficult to reconstruct the shape of the laterals. They are round in cross section (Pl. 16
, fig. 7). Due to the poor preservation of the material and the absence of photographs (Pia gave drawings), it is very difficult to reconstruct the shape of the laterals. They are round in cross section (Pl. 16 ![]() , figs. 3, 7), so they probably have a uniform diameter. However, they taper distally and show some similarity with piriform the type, although they may be acroform as well. That is why this alga is included in the genus Physoporella with a question mark. Small protrusions on the outside of the thallus (Pl. 16
, figs. 3, 7), so they probably have a uniform diameter. However, they taper distally and show some similarity with piriform the type, although they may be acroform as well. That is why this alga is included in the genus Physoporella with a question mark. Small protrusions on the outside of the thallus (Pl. 16 ![]() , fig. 3; Pia, 1931, Pl. XXI, fig. 6) may represent extensions of the laterals outwards. It is very difficult to determine the generic affiliation of this species, especially due to the lack of sufficient material, and also due to its structure.
, fig. 3; Pia, 1931, Pl. XXI, fig. 6) may represent extensions of the laterals outwards. It is very difficult to determine the generic affiliation of this species, especially due to the lack of sufficient material, and also due to its structure.
Similarities and differences: This species looks similar to Acroporella? prisca (Pia, 1912), n. comb. (see discussion bellow), and it differs mostly in its larger dimensions.
5.2. Genus Oligoporella Pia, 1912, emend.
5.2.1. Oligoporella pilosa Pia, 1912
Pl. 5 ![]() , figs. 1-6, 8-11; Pl. 6
, figs. 1-6, 8-11; Pl. 6 ![]() , figs. 1-8; Pls. 7
, figs. 1-8; Pls. 7 ![]() - 8
- 8 ![]() ; Pl. 9
; Pl. 9 ![]() , figs. 1-8; Pl. 12
, figs. 1-8; Pl. 12 ![]() , ? fig. 2; Pl. 17
, ? fig. 2; Pl. 17 ![]() , figs. 3, 6-7, 9;
Pl. 20
, figs. 3, 6-7, 9;
Pl. 20 ![]() ; Pl. 21
; Pl. 21 ![]() , figs. 1-2
, figs. 1-2
Origin of the name: Probably after assumed appearance of living alga with trichoblasts in extension of laterals (Latin "pilosus" = hairy).
Studied material: The studied and illustrated material of Oligoporella pilosa, described by Pia (1912, p. 42), originates from two localities. Sample I originates west of the village of Lapčići north from Budva in Montenegro (then in the Austrian province of Dalmatia), while the age of the sample is marked as Muschelkalk (hence the Middle Triassic). Sample V originates from the vicinity of the village of Stani�ići, near the previous one, while the age is also marked as Muschelkalk.
Since the first description did not specify a type specimen, Bucur and Grgasović in Bucur (1997) chose the specimen illustrated in Pia (1912, Pl. IV, fig. 3 = Bucur, 1997, Pl. I, figs. 1, 16 = this paper, Pl. V, fig. 1 above) as lectotype. Other specimens from the same sample (thin-sections I1 - I11) are therefore isotypes.
In the same paper, Pia (1912, p. 42) also describes the species of Oligoporella serripora based on the following material: sample LIV from Sarlkofel (Monte Serla) southeast of Niederdorf (Villabassa) in the Dolomites of northern Italy. The age was first defined by Pia (1912) as Muschelkalk (i.e., Middle Triassic), only to later give more details stating that the material is from the Upper Sarldolomite of Pelsonian-early Illyrian age (Pia, 1937, p. 46).
I also investigated the material described as Oligoporella pilosa forma typica by Pia (1935a) from Bosnia and Herzegovina: thin-section 231 from Ograda (site "t") on the road Are�in brijeg - Grab (probably a part of the road between Bosansko Grahovo and Knin along which Triassic deposits are well exposed), 46a from the village of Studenkovići on Mt Trebević (site "e"), 557c from the western part of Trebević Mt (site "f"), 575b and 577a from the village of Haliluci, east of Sarajevo (site "r"). In the same paper, Pia (1935a, p. 42) also describes Oligoporella pilosa var. subvaricans based on the following material: thin-section 46a and 46b also come from the site "e" (see above), 231 from the site "t" (see above) and 557a from the site "f".
The material from Croatia, that improved the research of this species, includes specimens from the area of Hum-�agudovac on the northern slopes of Medvednica Mt (Pl. 20 ![]() , figs. 2-3) and the quarry Belski dol on Ivan�čica Mt (Pl. 21
, figs. 2-3) and the quarry Belski dol on Ivan�čica Mt (Pl. 21 ![]() , fig. 2), as well as the areas of Jabuka near Trilj in Dalmatia from the collection of Branko Sokač (Pl. 20
, fig. 2), as well as the areas of Jabuka near Trilj in Dalmatia from the collection of Branko Sokač (Pl. 20 ![]() , fig.
5) and Udbina in Lika (Pl. 20
, fig.
5) and Udbina in Lika (Pl. 20 ![]() , fig.
1; Pl. 21
, fig.
1; Pl. 21 ![]() , fig.
1).
, fig.
1).
The material of O. pilosa forma physoporelloidea from Pia (1940) and P. pauciforata var. variabilis from Zanin Buri (1965) is studied from the published illustrations.
History of investigations: Pia (1912, p. 42; see also Granier & Sander, 2013, p. 29) establishes a new species Oligoporella pilosa: "Occasionally the calcareous skeleton shows a slight curvature. The canaliculae that occur in relatively dense verticils taper outward only moderately but for the most part very clearly. As a rule the verticils are well separated from one another by pore-free spaces, but exceptionally they are quite close together (Pl. IV, fig. 8 = this paper, Pl. 5 ![]() , fig. 11). The rather thin-walled calcareous skeleton is mostly unsegmented. However, it occasionally develops a well-marked bulge (Pl. IV, fig. 6 = this paper, Pl. 5
, fig. 11). The rather thin-walled calcareous skeleton is mostly unsegmented. However, it occasionally develops a well-marked bulge (Pl. IV, fig. 6 = this paper, Pl. 5 ![]() , fig. 9)."
, fig. 9)."
(In German: "Das Geh�use zeigt gelegentlich eine leichte Kr�mmung. Die Kan�lchen, die in ziemlich gedr�ngten Wirteln stehen, sind gegen au�en nur m��ig, aber meist doch deutlich verj�ngt. In der Regel sind die Wirtel durch porenfreie R�ume gut voneinander getrennt, ausnahmsweise k�nnen sie einander aber ganz nahe r�cken (vergl. Taf. IV, Fig. 8). Die eher d�nnwandige Schale ist meist ungegliedert. Gelegentlich entwickelt sich jedoch eine deutliche Wulstung (Taf. IV, Fig. 6).")
In the same work, Pia (1912, p. 42; see also Granier & Sander, 2013, p. 30) also establishes a new species of Oligoporella serripora: "This species is closely related to Oligoporella pilosa. The average size is somewhat smaller; the verticils are more closely spaced so that they appear to be biserial. The inclination of the branches against the main axis is rather large. But the most striking difference is that the pores taper very sharply outward. This may be related, at least in part, to the greater thickness of the calcification. The inner cavity (and obviously the corresponding main axis) is proportionately narrow." The species O. serripora is synonymous with the species O. pilosa, due to their identical structure, especially the shape of the laterals and the arrangement into double whorls. The mentioned thinning of the pores to the outside is within the range of variations of the species O. pilosa. The identity of these two species is well illustrated in Pl. 6 ![]() , where the type material of O. serripora is illustrated together with the type material of O. pilosa.
, where the type material of O. serripora is illustrated together with the type material of O. pilosa.
(In German: "Diese Art ist mit Oligoporella pilosa enge verwandt. Die durchschnittliche Gr��e ist etwas geringer, die Wirtel sind noch dichter gedr�ngt, so da� sie zweizeilig erscheinen. Die Neigung der �ste gegen die Stammzelle ist ziemlich gro�. Der auffallendste Unterschied zeigt sich aber darin, da� die Puren sich gegen. au�en sehr stark verj�ngen. Dies mag wohl, wenigstens zum Teil, mit der gr��eren relativen Dicke der Verkalkung zusammenh�ngen. Der innere Hohlraum ( und dem entsprechend offenbar auch die Stammzelle) war verh�ltnism��ig d�nn.")
Herak (1965, p. 15) gives a new diagnosis of O. pilosa: "The thallus is cylindrical; the branches rare, relatively large, tapering towards the distal end; distributed in whorls. Variability is rather emphasized (intusannulation, different position of the branches, protuberances on the surface of the thallus, etc.). But all the mentioned features are infraspecific. Therefore several varieties can be distinguished."
Oligoporella pilosa var. pilosa Pia, 1935
Pl. 5 ![]() , figs. 1-6, 8-11; Pl. 6
, figs. 1-6, 8-11; Pl. 6 ![]() , figs. 1-8; Pl. 7
, figs. 1-8; Pl. 7 ![]() ; Pl. 8
; Pl. 8 ![]() , figs. ?1, 2-10, 12; Pl. 12
, figs. ?1, 2-10, 12; Pl. 12 ![]() ,
fig. ?2; Pl. 17
,
fig. ?2; Pl. 17 ![]() , figs. 6-7, 9; Pl. 20
, figs. 6-7, 9; Pl. 20 ![]() , figs. 2-3, 5
, figs. 2-3, 5
1912 - Oligoporella pilosa n. sp. - Pia, p. 42, Pl. IV, figs. 1-4, 6-8.
1912 - Oligoporella serripora n. sp. - Pia, p. 42, Pl. IV, figs. 9-11.
1935a (nom. nud.) - Oligoporella pilosa forma typica n. forma. - Pia, p. 205-211, Figs. 5-11, ?12.
1935a - Oligoporella pilosa var. subvaricans n. var. - Pia, p. 214-216, Fig. 20.
1940 - Oligoporella pilosa forma physoporelloidea n. forma.- Pia, p. 5, Pl. 1, fig. 11.
1957 (nom. nud.) - Oligoporella pilosa typica.- Bystrick�, p. 229-230, Pl. IV, figs. 1-2.
1964 - Oligoporella pilosa var. pilosa nom. nov.- Bystrick�, p. 109-111, Pl. X, figs. 2, 6; Pl. XI, figs. 2, 4; Pl. X, fig. 4, from Bystrick� (1957: Pl. IV, fig. 1).
1965 - Physoporella pauciforata var. variabilis n. var. - Zanin Buri, p. 476-478, Pl. 49, figs. 3, 6.b.
1989 - Oligoporella pilosa var. pilosa.- Buček, p. 134, Pl. I, fig. 4.
1994 - Oligoporella pilosa var. pilosa.- Bucur et al., p. 94, 96, Pl. 10, figs. 1 pars, 5 pars; Pl. 11, figs. 16-18, 19 pars; Pl. 12, figs. 1-10, 12, 14; Pl. 13, fig. 1 pars.
1997 - Oligoporella pilosa var. pilosa.- Bucur, p. 181-186, Pl. I, figs. 2, 8, 12; Pl. I, figs. 1, 16, from Pia (1912: Pl. IV, fig. 3); Pl. I, figs. 3-7, 9-11, 13-15, from Bucur et al. (1994).
2002 - Oligoporella pilosa pilosa - Piros, p. 126-127, Pl. I, fig 1; Pl. II, fig. 2.
2013 - Oligoporella pilosa pilosa - Rychliński et al., Pl. 2, figs. 11-12.
2021 - Oligoporella pilosa pilosa - Gawlick et al., Pl. 5, fig. n; Pl. 15, figs. l, n-p.
Pia (1935a, p. 205, 207) divides the species Oligoporella pilosa into varieties and described Oligoporella pilosa forma typica: "The branches of a single whorl are only slightly inclined upwards and downwards. As with many species, the calcification between the pores of the same whorl does not extend as far inward as in the pore-free zones between the whorls. This must be taken into account when taking measurements. Especial care should be taken that on the cross sections as it is not possible to reliably recognize whether we are in one whorl or between two... The described calcification arrangement creates small inwardly protruding calcareous bars between the whorls. But in the typical form I include only those specimens in which these bars are hardly noticeable."
(In German: "Die �ste eines Wirtels sind nur wenig gegen oben und unten geneigt. Wie bei vielen Arten reicht die Verkalkung zwischen den Poren desselben Wirtels weniger weit nach innen, als in den porenfreien Zonen zwischen den Wirteln. Darauf muss man bei den Messungen R�cksicht nehmen� Durch die beschriebene Anordnung des Kalkes entstehen zwischen den Wirteln kleine, gegen innen vorspringende Kalkstein. Zur typischen Form rechne ich aber nur solche Stucke, bei denen diese Leisten wenig auffallen.")
In the same paper, Pia (1935a, p. 214-216) establishes a variety of Oligoporella pilosa var. subvaricans: "In some specimens of Oligoporella pilosa, the pores of the upper and lower rows of a compressed whorl diverge more than in the specimens previously discussed as 'forma typica'. But even within this group of different specimens, the mentioned feature is developed to very different degrees. Since the specimens in which it is most pronounced also differ from the type in other respects, I consider it best to establish two varieties. I designate as var. subvaricans those specimens of Olig. pilosa in which the whorled branches were inclined only moderately upwards and downwards, and which in the other characters are still closely related to the 'forma typica'. According to the table of dimensions, the total size seems to be slightly smaller than that of the type. It is 2.01 � 0.088 mm. The difference is 0.13 � 0.106 mm, so it is of little importance. The distance between the whorls is relatively large, which is understandable with the divergent position of the branches." Divergence of laterals in the material described as O. pilosa var. subvaricans does not differ from the range observed in specimens described as forma typica, as well as from other published material of var. pilosa. This is best visible on Pl. 7-8 where the type material of both varieties is illustrated together. It is obvious that there is a continuous series of lateral divergence values, and I consider that O. pilosa var. subvaricans is a younger synonym of O. pilosa var. pilosa.
(In German: "Bei nicht wenigen Stucken von Oligoporella pilosa divergieren die Poren der oberen und unteren Reihe eines gedr�ngten Wirtels mehr als bei den oben als forma typica besprochenen Individuen. Aber auch innerhalb dieser Gruppe abweichender Stucke ist das erw�hnte Merkmal sehr verschieden stark entwickelt. Da die Exemplare, bei denen es am kr�ftigsten hervortritt, auch in anderen Hinsicht vom Typus abweichen, halte ich es f�r das Beste, zwei Variet�ten aufzustellen. Ich bezeichne als var. subvaricans jene Stucke von Oligoporella pilosa, bei denen die Wirtelaste nur massig gegen oben und unten geneigt waren und die sich in den �brigen Merkmalen noch eng an die forma typica anschlie�en. Die Gesamtgr�sse scheint nach der Tabelle etwas geringer zu sein, als beim Typus. Sie betr�gt 2.01�0.088 mm. Die Differenz ist 0.13�0.106 mm, also wenig gesichert. der Abstand der Wirtel ist verh�ltnism�ssig gross, was bei der gespreizten Stellung der Aeste ja begreiflich ist.")
Pia (1940, p. 5) establishes a new form (i.e., variety) Oligoporella pilosa forma physoporelloidea: "Many Oligoporellas are characterized by pores that are very thick in the inner part, and narrow outwards in the shape of a bottle neck. Occasionally, a thin calcareous lamella extends between the two parts of the pore, the colour of which completely corresponds to the remaining skeleton, so it was probably created during the life of the algae, and not only during fossilization (Pia, 1940, Fig. 11 right = this paper, Pl. 13 ![]() , fig. 9). These features are somewhat similar to the genus Physoporella, from which they differ, however, by pores that are always open to the outside. Physoporella is most likely derived from Oligoporella. It is probably a transitional form, in which the proximal parts of the branches are already formed as sporangia, while the distal ones have not yet fallen off." In the description of this variety, Pia was obviously guided by the still unresolved issue of openness and closedness of pores, i.e., the distinction between the two genera. Since this criterion has been rejected as taxonomically relevant
(see Chapter 3), the existence of this variety is not justified, and I consider it a junior synonym of O. pilosa var. pilosa. As for "a thin calcareous lamella" it is clearly part of the calcite cement. The same cement that fills the mold cavity formed by dissolution of the primary aragonite algal thallus.
, fig. 9). These features are somewhat similar to the genus Physoporella, from which they differ, however, by pores that are always open to the outside. Physoporella is most likely derived from Oligoporella. It is probably a transitional form, in which the proximal parts of the branches are already formed as sporangia, while the distal ones have not yet fallen off." In the description of this variety, Pia was obviously guided by the still unresolved issue of openness and closedness of pores, i.e., the distinction between the two genera. Since this criterion has been rejected as taxonomically relevant
(see Chapter 3), the existence of this variety is not justified, and I consider it a junior synonym of O. pilosa var. pilosa. As for "a thin calcareous lamella" it is clearly part of the calcite cement. The same cement that fills the mold cavity formed by dissolution of the primary aragonite algal thallus.
(In German: "Mehrere der vorliegenden Oligoporellen zeichnen sich dadurch aus, dass die Poren im inneren Teil sehr dick sind und sich gegen aussen flaschenhalsartig verengen. Gelegentlich l�uft zwischen den beiden Abschnitten der Pore eine d�nne Kalklamelle durch, die in der Farbe ganz mit dem �brigen Skelett �bereinstimmt, also wohl w�hrend des Lebens der Alge, nicht erst bei der Fossilisation gebildet wurde (Fig. 11, rechts). Diese Merkmale n�hern unsere Form etwas der Gattung Physoporella, von der sie sich aber durch die gegen aussen stets offenen Poren unterscheidet. Physoporella ist h�chstwahrscheinlich von Oligoporella abzuleiten. Vermutlich liegt uns jetzt also eine �bergangsrasse vor, bei der die proximalen Teile der Wirtel�ste schon deutlich als Sporangien ausgebildet waren, die distalen aber noch nicht fr�hzeitig abfielen.")
Zanin Buri (1965, p. 476-478) establishes Physoporella pauciforata var. variabilis: "Skeleton. The calcareous sleeve is of modest thickness and cylindrical in shape; it usually has a slightly wavy outer surface. In none of the sections, however oriented, was any trace of fissuration or cracks found, that Pia consider to be typical for the species. The shape of the axial cell is not, with very few exceptions, clearly distinguishable. The maximum length of the skeleton is about 2 cm. Branches. They have a verticillated arrangement (Zanin Buri, 1965, Pl. 49, fig. 6b); the verticils are spaced apart at intervals that are not always strictly constant; the average value of this distance is around 0.3-0.5 mm; each whorl can consist of one or two series of branches, even in the same individual, without any regular and systematic alternation being noticed (Zanin Buri, 1965, Pl. 49, fig. 3; below, two series of circular pores are observed for each whorl, while in the central and upper parts only one is observed). The typical pear-shaped appearance of the branches is clearly visible, which are always closed on the outside and are arranged perpendicular to the central cavity or slightly oblique towards the top. The branches, cross-sectioned, have an irregularly circular shape." The main characteristic that distinguishes this taxon from others, according to the author, is the existence of both single and double whorls on the same thallus. Although the two illustrated specimens are poorly preserved (especially the one in Zanin Buri, 1965, Pl. 49, fig. 6b), the holotype (Zanin Buri, 1965, Pl. 49, fig. 3 = this paper, Pl. 17 ![]() , fig. 6) clearly shows the existence of a double whorl in the lower part of the thallus, where it is cut longitudinally-tangentially. In the upper part of the thallus, the whorls are cut longitudinally, so due to the alternating arrangement of laterals in the whorl, only one lateral can be seen in the section of each whorl. This is a phenomenon that every experienced fossil algae researcher is familiar with. Therefore, there are no single whorls. Due to its structure, I consider that this alga is also a junior synonym of O. pilosa var. pilosa.
, fig. 6) clearly shows the existence of a double whorl in the lower part of the thallus, where it is cut longitudinally-tangentially. In the upper part of the thallus, the whorls are cut longitudinally, so due to the alternating arrangement of laterals in the whorl, only one lateral can be seen in the section of each whorl. This is a phenomenon that every experienced fossil algae researcher is familiar with. Therefore, there are no single whorls. Due to its structure, I consider that this alga is also a junior synonym of O. pilosa var. pilosa.
(In Italian: "Scheletro. Il manicotto calcareo e di spessore modesto e di forma cilindrica; di solito presenta una superficie esterna debolmente ondulata. In nessuna sezione, comunque orientata, si e rinvenuta traccia di fessurazioni o spaccature, come invece Pia sembra considerare tipico della specie. L'andamento della cellula assiale non e, tranne pochissime eccezioni, chiaramente distinguibile. La lunghezza massima dello scheletro e di circa 2 cm. Rami. Hanno disposizione verticillata (v. tav. 49, fig. 6b); i verticilli sono distanziati tra loro ad intervalli che non sono sempre rigorosamente constanti; il valore medio di tale distanza si aggira attorno a 0.3-0.5 mm; ciascun verticillo pu� essere costituito da una o da due serie di rami, anche in uno stesso individuo, senza che si possa notare nessuna regolare e sistematica alternanza (v. tav. 49; in basso si osservano due serie di pori circolari per ogni verticillo, mentre nelle parti centrale e superiore se ne osserva soltanto una). E' chiaramente visibile il tipico aspetto piriforme dei rami, i quali si presentano sempre chiusi all'esterno e sono disposti perpendicolarmente al canale centrale o debolmente obliqui verso l'alto. I rami, sezionati trasversalmente, presentano forma irregolarmente circolare.")
Bystrick� (1964, p. 109-111) gives the type variety a new name Oligoporella pilosa var. pilosa, in accordance with the rules of botanical nomenclature (Art. 26.1 ICN, Turland et al., 2018), and completes the diagnosis: "The thallus is made of a simple tube, without articulation. The pores are of trichophore type, very wide in the basal part, gradually thinning outwards and exit the thallus. They are arranged in alternating two-row whorls with moderately diverging pores. The calcareous sheath reaches between the whorls into the internal cavity farther than between the pores of the two-rowed whorl. The protrusions on the shell thus formed are small and indistinct (Pia, 1935a)."
Bucur (1997) publishes a synthesis of relevant and updated informations on the variety pilosa with a detailed description. He accepted my suggestion, so in the definition of the lectotype he states that it was chosen by Bucur and Grgasović. In the description of the algae he writes (translation from French): "The thallus of Oligoporella pilosa pilosa is cylindrical, non-articulated. The outer surface is generally smooth or has slight undulation. The laterals are trichophore, that is, they gradually narrow outwards, but their shape is variable. Some have laterals of which the basal part is very thick, narrowing very quickly, and whose shape is similar to that of the piriform one of Physoporella from which they differ, however, by a very sharp termination. In other specimens, the differences in diameter of the proximal part and the distal part of the lateral are less. This variability has also been underlined by Bystrick� (1964). The ramifications are arranged in "biseriate" whorls: more precisely, they are regularly alternating with divergent inclinations (up and down). This divergence is variable but the angle is generally low in this variety, compared to those found in the varieties subvaricans and varicans." He does not give a new diagnosis, but translate that of Bystrick� (1964).
Description: The thallus is cylindrical, straight (Pl. 5 ![]() , fig. 2; Pl. 6
, fig. 2; Pl. 6 ![]() , fig. 5; Pl. 17
, fig. 5; Pl. 17 ![]() , figs. 6-7; Pl. 20
, figs. 6-7; Pl. 20 ![]() , fig. 2) or slightly curved (Pl. 5
, fig. 2) or slightly curved (Pl. 5 ![]() , figs. 8-9). The inner surface of the calcareous thallus is smooth in well-preserved specimens (Pl. 5
, figs. 8-9). The inner surface of the calcareous thallus is smooth in well-preserved specimens (Pl. 5 ![]() , figs. 1-2, 6, 9; Pl. 6
, figs. 1-2, 6, 9; Pl. 6 ![]() , figs. 1 below, 2 middle; Bucur et al., 1994, Pl. 11, figs. 16-18), but it is usually, due to subsequent dissolution, more or less uneven. In the thallus of O. pilosa pilosa the wrinkling of the stem cell is not observed as in P. pauciforata. Calcification in this species is very variable. This is best seen in the lectotype material where in the same sample we find specimens in which the calcification reaches all the way to the stem cell (Pl. 5
, figs. 1 below, 2 middle; Bucur et al., 1994, Pl. 11, figs. 16-18), but it is usually, due to subsequent dissolution, more or less uneven. In the thallus of O. pilosa pilosa the wrinkling of the stem cell is not observed as in P. pauciforata. Calcification in this species is very variable. This is best seen in the lectotype material where in the same sample we find specimens in which the calcification reaches all the way to the stem cell (Pl. 5 ![]() , figs. 2, 4; Pl. 6
, figs. 2, 4; Pl. 6 ![]() , fig. 1 below), those in which the calcification covers only the distal parts of laterals (Pl. 6
, fig. 1 below), those in which the calcification covers only the distal parts of laterals (Pl. 6 ![]() , fig. 1 middle, 2 below), as well as all values in between (Pl. 5
, fig. 1 middle, 2 below), as well as all values in between (Pl. 5 ![]() , figs. 1, 3, 5-6, 8-10; Pl. 6, figs. 1-4). This also applies to specimens from other localities, often with uncalcified proximal parts of the laterals. The thickness of the calcareous shell also varies. The outer surface of the thallus is usually slightly undulated (Pl. 5
, figs. 1, 3, 5-6, 8-10; Pl. 6, figs. 1-4). This also applies to specimens from other localities, often with uncalcified proximal parts of the laterals. The thickness of the calcareous shell also varies. The outer surface of the thallus is usually slightly undulated (Pl. 5 ![]() , figs. 1, 8-9; Pl. 6
, figs. 1, 8-9; Pl. 6 ![]() , figs. 1 up and down, 2 middle; Pl. 7
, figs. 1 up and down, 2 middle; Pl. 7 ![]() , figs. 4, 10-11; Pl. 8
, figs. 4, 10-11; Pl. 8 ![]() , fig. 2; Buček, 1989, Pl. I, fig. 4; Bucur et al., 1994, Pl. 12, figs. 2, 6), and may be flat, although not quite smooth (Pl. 5
, fig. 2; Buček, 1989, Pl. I, fig. 4; Bucur et al., 1994, Pl. 12, figs. 2, 6), and may be flat, although not quite smooth (Pl. 5 ![]() , fig. 2; Pl. 6
, fig. 2; Pl. 6 ![]() , fig. 1 middle, 2 down, 5, 8; Pl. 7
, fig. 1 middle, 2 down, 5, 8; Pl. 7 ![]() , figs. 1-3; Pl. 8
, figs. 1-3; Pl. 8 ![]() , fig. 6; Pl. 17
, fig. 6; Pl. 17 ![]() , fig. 7; Pl. 20
, fig. 7; Pl. 20 ![]() , figs. 2-3; Bystrick�, 1957, Pl. IV, figs. 1-2; 1964, Pl. X, fig. 2; Zanin Buri, 1965, Pl. 49, fig. 3). As with most other Dasycladalean algae, the skeleton is composed of mosaic crystals of calcite formed by heteroaxial replacement of the primary aragonite skeleton, often with a pronounced micritic rim and micritic overgrowth. The exceptions are well-preserved specimens from the lectotype material, characterized by extremely thin micritic sheaths and a skeleton of yellowish mosaic calcite, formed probably by homoaxial replacement of the primary skeleton, as evidenced by the excellent preservation of these algae specimens.
, figs. 2-3; Bystrick�, 1957, Pl. IV, figs. 1-2; 1964, Pl. X, fig. 2; Zanin Buri, 1965, Pl. 49, fig. 3). As with most other Dasycladalean algae, the skeleton is composed of mosaic crystals of calcite formed by heteroaxial replacement of the primary aragonite skeleton, often with a pronounced micritic rim and micritic overgrowth. The exceptions are well-preserved specimens from the lectotype material, characterized by extremely thin micritic sheaths and a skeleton of yellowish mosaic calcite, formed probably by homoaxial replacement of the primary skeleton, as evidenced by the excellent preservation of these algae specimens.
The shape of the laterals in the genus Oligoporella, and thus in the typical species O. pilosa, where described by Pia (1912) as trichoform, having in mind primarily the openness of the pores, i.e., the uprise of laterals outside the calcareous thallus. This taxonomic criterion was rejected, and it is proved
(see Chapter 5.2.2.) that the laterals of Oligoporella are of the piriform type. The shape of the laterals somewhat varies: from quite large (Pl. 5 ![]() , figs. 1, 4, 8; Pl. 6
, figs. 1, 4, 8; Pl. 6 ![]() , figs. 1 pars, 2 pars; Pia, 1940, Pl. 1, fig. 11; Bystrick�, 1964, Pl. X, fig. 6; Bucur et al., 1994, Pl. 12, figs. 2-3), elongated ones (Pl. 5
, figs. 1 pars, 2 pars; Pia, 1940, Pl. 1, fig. 11; Bystrick�, 1964, Pl. X, fig. 6; Bucur et al., 1994, Pl. 12, figs. 2-3), elongated ones (Pl. 5 ![]() , fig. 11; Pl. 6
, fig. 11; Pl. 6 ![]() , figs. 1 pars, 2 pars, 5- 8; Pl. 7
, figs. 1 pars, 2 pars, 5- 8; Pl. 7 ![]() , figs. 1-4, 10; Pl. 8
, figs. 1-4, 10; Pl. 8 ![]() , figs. 2-10, 12; Bucur et al., 1994, Pl. 11, figs. 16, 19; Pl. 12, fig. 9), relatively small (Pl. 5
, figs. 2-10, 12; Bucur et al., 1994, Pl. 11, figs. 16, 19; Pl. 12, fig. 9), relatively small (Pl. 5 ![]() , figs. 2, 9; Pl. 17
, figs. 2, 9; Pl. 17 ![]() , figs. 2-3; Zanin Buri, 1965, Pl. 49, fig. 3), to quite thin (Pl. 7
, figs. 2-3; Zanin Buri, 1965, Pl. 49, fig. 3), to quite thin (Pl. 7 ![]() , figs. 6, 8, 12; Pl. 17
, figs. 6, 8, 12; Pl. 17 ![]() , fig. 7; Bucur et al., 1994, Pl. 12, figs. 7, 10), with all possible intermediates. In the example from Pl. 6
, fig. 7; Bucur et al., 1994, Pl. 12, figs. 7, 10), with all possible intermediates. In the example from Pl. 6 ![]() , fig. 1 below, in which the calcification extends all the way to the stem cell, a typical piriform type of laterals is nicely seen. The junction of the lateral and the stem cell was probably through a very narrow pore that is not preserved in the fossils. Trichoblasts at the top of the laterals (Fig. 8
, fig. 1 below, in which the calcification extends all the way to the stem cell, a typical piriform type of laterals is nicely seen. The junction of the lateral and the stem cell was probably through a very narrow pore that is not preserved in the fossils. Trichoblasts at the top of the laterals (Fig. 8 ![]() ), which are often found in P. pauciforata, are rarely observed in O. pilosa. Exceptions are the specimen on Pl. 7
), which are often found in P. pauciforata, are rarely observed in O. pilosa. Exceptions are the specimen on Pl. 7 ![]() , fig. 5 (described as O. pilosa var. subvaricans), in which "horns" on the outer surface represent calcified trichoblasts, and the specimen on Pl. 20
, fig. 5 (described as O. pilosa var. subvaricans), in which "horns" on the outer surface represent calcified trichoblasts, and the specimen on Pl. 20 ![]() , fig. 2, in which an outer "lamellae" is formed by calcification of the upward-pointing trichoblasts, identical to those of P. pauciforata
(Chapter 5.1.1.). Trichoblasts are rarely preserved, probably because they generally stood more perpendicular to the stem cell, similar to those in var. undulata
(Chapter 5.1.1.), so they were outside the mucous membrane of the algae. Interesting are the specimens on Pl. 17
, fig. 2, in which an outer "lamellae" is formed by calcification of the upward-pointing trichoblasts, identical to those of P. pauciforata
(Chapter 5.1.1.). Trichoblasts are rarely preserved, probably because they generally stood more perpendicular to the stem cell, similar to those in var. undulata
(Chapter 5.1.1.), so they were outside the mucous membrane of the algae. Interesting are the specimens on Pl. 17 ![]() , fig. 7 and Pl. 20
, fig. 7 and Pl. 20 ![]() , figs. 2-3, where the top parts of the thallus with very thin laterals are preserved. An identical phenomenon has been reported in other fossil and recent Dasycladales.
, figs. 2-3, where the top parts of the thallus with very thin laterals are preserved. An identical phenomenon has been reported in other fossil and recent Dasycladales.
The laterals are arranged in double whorls consisting of two close rows (essentially whorls) of laterals. The presence of double whorls is especially well-visible on the lectotype (Pl. 5 ![]() , fig. 1) and specimens on Pl. 5
, fig. 1) and specimens on Pl. 5 ![]() , fig. 8; Pl. 6
, fig. 8; Pl. 6 ![]() , figs. 5, 7; Pl. 7
, figs. 5, 7; Pl. 7 ![]() , fig. 5; Pl. 20
, fig. 5; Pl. 20 ![]() , figs. 3, 5. In these, the two rows of laterals are arranged alternately (Pl. 5
, figs. 3, 5. In these, the two rows of laterals are arranged alternately (Pl. 5 ![]() , figs. 1, 9; Pl. 6
, figs. 1, 9; Pl. 6 ![]() , fig. 5; Pl. 7
, fig. 5; Pl. 7 ![]() , figs. 3 left, 6; Pl. 20
, figs. 3 left, 6; Pl. 20 ![]() , fig. 2; Zanin Buri, 1965, Pl. 49, fig. 3; Bucur et al., Pl. 10, fig. 1 middle), but are sometimes less pronounced (Pl. 5
, fig. 2; Zanin Buri, 1965, Pl. 49, fig. 3; Bucur et al., Pl. 10, fig. 1 middle), but are sometimes less pronounced (Pl. 5 ![]() , figs. 2, 9). The double whorls are usually clearly spaced from each other (Pl. 5
, figs. 2, 9). The double whorls are usually clearly spaced from each other (Pl. 5 ![]() , figs. 1-2, 8-9; Pl. 6
, figs. 1-2, 8-9; Pl. 6 ![]() , fig. 5; Pl. 20
, fig. 5; Pl. 20 ![]() , fig. 5; Zanin Buri, 1965, Pl. 49, fig. 3; Bucur et al., 1994, Pl. 12, figs. 2, 8), but they can also be relatively close (Pl. 5
, fig. 5; Zanin Buri, 1965, Pl. 49, fig. 3; Bucur et al., 1994, Pl. 12, figs. 2, 8), but they can also be relatively close (Pl. 5 ![]() , fig. 10; Pl. 6
, fig. 10; Pl. 6 ![]() , fig. 8; Pl. 7
, fig. 8; Pl. 7 ![]() , figs. 3 left, 4-5, 10-11; Pl. 8
, figs. 3 left, 4-5, 10-11; Pl. 8 ![]() , fig. 6; Pl. 20
, fig. 6; Pl. 20 ![]() , figs. 2-3). The inclination of the laterals inside the whorl is not the same, but the laterals in the upper row are slightly more inclined towards the axis than those in the lower row (on average about 5�), so that the whole whorl is slightly inclined towards the axis (Pl. 5
, figs. 2-3). The inclination of the laterals inside the whorl is not the same, but the laterals in the upper row are slightly more inclined towards the axis than those in the lower row (on average about 5�), so that the whole whorl is slightly inclined towards the axis (Pl. 5 ![]() , figs. 1, 8; Pl. 7
, figs. 1, 8; Pl. 7 ![]() , fig. 5) as with most other Oligoporella species. Slightly oblique sections often cut through both rows of laterals (Pl. 5
, fig. 5) as with most other Oligoporella species. Slightly oblique sections often cut through both rows of laterals (Pl. 5 ![]() , fig. 5; Pl. 6
, fig. 5; Pl. 6 ![]() , figs. 1-2, 6; Pl. 8
, figs. 1-2, 6; Pl. 8 ![]() , figs. 2-3, 5, 9, 12), so that the number of pores in such cross-sections can be relatively large (32-36), which is a criterion for distinguishing, the otherwise very similar, genera Physoporella and Oligoporella if only such cross-sections are available. The mentioned illustrations also show that the laterals are quite close to each other, and the slightly thicker ones often touch each other.
, figs. 2-3, 5, 9, 12), so that the number of pores in such cross-sections can be relatively large (32-36), which is a criterion for distinguishing, the otherwise very similar, genera Physoporella and Oligoporella if only such cross-sections are available. The mentioned illustrations also show that the laterals are quite close to each other, and the slightly thicker ones often touch each other.
Emended diagnosis: Cylindrical thallus with a smooth inner and slightly undulated or more or less flat outer surface. The laterals are piriform with a wide rounded proximal part and a gradual thinning towards the distal end, on which they carried trichoblasts. They are arranged in double whorls consisting of two close rows of laterals. The laterals within the whorl are mostly alternately arranged. They are usually slightly inclined upwards. The thickness of the laterals and the distance between the whorls can vary.
|
|
Figure 9: Oligoporella pilosa var. pilosa Pia. Idealized longitudinal (A), transverse (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows variants with calcification close to the stem cell (III), away from the stem cell (I) only in the distal part of the laterals (II), and with relatively smooth (I-II) and slightly undulated (III) outer surface of the thallus. The top part of the thallus with slightly thinner laterals (I-II) is also shown. Section I shows a case where whorls can be cut in a way that gives a false sense of one-rowed whorls. Trichoblasts are shown on some laterals. The cross section (B) shows variants with a relatively smooth (VI) and less (IV, VII) or more (V) undulated outer surface of the thallus, and calcification that reaches very close to the stem cell (IV), near to the stem cell (V) and only in the distal part of the laterals (VI-VII). The tangential section (C) shows the sections of the proximal (lower whorl) and distal parts of the laterals (upper whorl). |
Oligoporella pilosa var. intusannulata Pia, 1935
Pl. 8 ![]() , figs. 11, 13, 15, not 14; Pl. 9
, figs. 11, 13, 15, not 14; Pl. 9 ![]() , figs. 1-8; Pl. 17
, figs. 1-8; Pl. 17 ![]() , fig. 3; Pl. 20
, fig. 3; Pl. 20 ![]() , figs. 1, 4; Pl. 21
, figs. 1, 4; Pl. 21 ![]() , fig. 1
, fig. 1
1935a - Oligoporella pilosa var. intusannulata n. var. - Pia, p. 212-214, Figs. 16-18.
1940 - Oligoporella pilosa forma intusannulata (sic) - Pia, p. 4, Pl. 1, fig. 7.
1957 - Oligoporella pilosa intusannulata - Bystrick�, p. 230, Pl. IV, figs. 3-6.
1964 - Oligoporella pilosa var. intusannulata - Bystrick�, p. 111-113, Pl. X, figs. 1, 3, 5, 8-9; Pl. XI, figs. 1, 3, 5, 7; Pl. XII, figs. 1, 3; Pl. XXXIV, fig. 1; Pl. XII, fig. 2.
1965 - Oligoporella pilosa var. intusannulata - Assereto et al., p. 819-820, Pl. 78.b; Pl. 79, figs. 2-4.
1965 - Oligoporella pilosa var. pilosa - Assereto et al., p. 818-819, Pl. 78.a & pars; Pl. 81, fig. 2.
1965 - Oligoporella pilosa var. varicans - Assereto et al., p. 821-822, Pl. 79, fig. 1; Pl. 81, figs. 6-7.
1965 - Oligoporella pilosa var. intusannulata - Herak, p. 16, Pl. XI, figs. 5, 7.
1986 - Oligoporella pilosa var. intusannulata - Bystrick�, p. 295-299, 301, Pl. I, figs. 1-2, 8.
1989 - Oligoporella pilosa var. intusannulata - Buček, p. 134, Pl. I, fig. 3.
2021 - Oligoporella pilosa intusannulata - Gawlick et al., Pl. 11, fig. a.
2021 - Oligoporella intusannulata - Gawlick et al., Pl. 11, figs. b-c, e, g, i.
Origin of the name: After the uneven inner surface of the calcareous thallus, which Pia (1920) called intusannulation (Latin: "intus" = on the inside; "annulus" = ring).
Studied material: The material described by Pia (1935a) originates from Bosnia and Herzegovina: thin-section 231 and 235a from Ograda (site "t") on the road Are�in brijeg - Grab, 555b NW from the village Blizanac on Trebević Mt. (site "d"), 557a from the western part of Trebević Mt (site "f"), 575a from the village of Haliluci, east of Sarajevo (site "r").
Specimen from Pl. 9 ![]() , fig. 1 (= Pia, 1935a, Fig. 17
, fig. 1 (= Pia, 1935a, Fig. 17 ![]() ) best represents all the characteristics of this variety, so I choose it as a lectotype. The other specimens from thin-section 575a (Pl. 8
) best represents all the characteristics of this variety, so I choose it as a lectotype. The other specimens from thin-section 575a (Pl. 8 ![]() , figs. 11, 13; Pl. 9
, figs. 11, 13; Pl. 9 ![]() ,
figs. 2, 4 = Pia, 1935a, Fig. 18) represent isotypes.
,
figs. 2, 4 = Pia, 1935a, Fig. 18) represent isotypes.
The specimen from thin-section 575a (Pl. 8 ![]() , fig. 14 = Pia, 1935a, Pl. II, fig. 8) does not belong to the genus Oligoporella, as it has two orders of laterals: rather wide first order laterals and smaller second order laterals carrying trichoblasts on the top. Based on only one specimen, it is not possible to define which genus it belongs to, but it is likely that it belongs to the tribe Triploporelleae.
, fig. 14 = Pia, 1935a, Pl. II, fig. 8) does not belong to the genus Oligoporella, as it has two orders of laterals: rather wide first order laterals and smaller second order laterals carrying trichoblasts on the top. Based on only one specimen, it is not possible to define which genus it belongs to, but it is likely that it belongs to the tribe Triploporelleae.
The material from Croatia, with which I supplemented the research of this species, includes specimens from the vicinity of Udbina in Lika from the collection of Branko Sokač (Pl. 20 ![]() , figs. 1, 4; Pl. 21
, figs. 1, 4; Pl. 21 ![]() , fig. 1).
, fig. 1).
History of investigations: Pia (1935a, p. 212-213) diagnoses the newly established variety Oligoporella pilosa var. intusannulata: "The only characteristic of this variety lies in the fact that inwardly protruding calcareous bars between the whorls are more developed, so that in sections (especially very oblique ones) they conspicuously stand out. I consider the systematic value of this characteristic quite low. On the other hand, it is sometimes very noticeable on the fossil skeletons and it is the real reason why I also express it in the nomenclature. Originally, I wanted to make another subdivision of the specimens included here according to the degree of development of the bars, but this now seems superfluous to me. It suffices to point out that all transitional forms, with regard to this feature, belong to the 'forma typica'."
(In German: "Das einzige Merkmal dieser Variet�t liegt darin, dass die schon auf p. 207 erw�hnten, gegen innen vorspringenden Kalk-leisten zwischen den Wirteln mehr entwickelt sind, so dass sie in de.r Schliffen - besonders in stark schr�gen - auffallend her-vortreten. Ich halte den systematischen Wert dieser Eigenschaft f�r recht gering. Dagegen f�llt sie an den fossilen Skeletten manchmal stark in die Augen und das ist der eigentliche Grund, warum ich sie auch in der Namengebung zum Ausdruck bringe. Urspr�nglich wollte ich unter den hier einzureihenden St�cken noch eine Unterteilung nach dem Grad der Entwicklung der Leisten machen. Dies scheint mir aber jetzt doch �berfl�ssig. Es gen�gt, darauf hinzuweisen, dass in dieser Beziehunp- alle Ueberg�nge zu der forma typica bestehen. Vergl. auch die Textabbildungen.")
Bystrick� (1964, p. 111) gives a short diagnosis: "The shape and size of the thallus as in var. pilosa. The calcareous bars extending between the whorls into the inner cavity are very pronounced and mostly adhere to the cell."
Description: The thallus can be cylindrical (Pl. 8 ![]() , fig. 15; Pl. 20
, fig. 15; Pl. 20 ![]() , figs. 1, 4) or very slightly claviform (Pl. 9
, figs. 1, 4) or very slightly claviform (Pl. 9 ![]() ,
fig. 1), straight (Bystrick�, 1964, Pl. X, fig. 1) or very slightly bent (Pl. 8
,
fig. 1), straight (Bystrick�, 1964, Pl. X, fig. 1) or very slightly bent (Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() , fig. 1; Pl. 20
, fig. 1; Pl. 20 ![]() , figs. 1, 4; Buček, 1989, Pl. I, fig. 3). The main characteristic of this variety is the rather more intense calcification. As Pia (1935a) has already pointed out, the taxonomic value of this is quite low. However, I decided to keep this variety because it is morphologically quite distinctive and easily recognizable. Calcification generally reaches almost to the stem cell, but this only applies to the intervals between the whorls. The laterals of the lower and upper rows of the same whorl touch each other, so these parts are not calcified. The calcified parts between the whorls in longitudinal and oblique sections have the shape of the letter "T" (Pl. 20
, figs. 1, 4; Buček, 1989, Pl. I, fig. 3). The main characteristic of this variety is the rather more intense calcification. As Pia (1935a) has already pointed out, the taxonomic value of this is quite low. However, I decided to keep this variety because it is morphologically quite distinctive and easily recognizable. Calcification generally reaches almost to the stem cell, but this only applies to the intervals between the whorls. The laterals of the lower and upper rows of the same whorl touch each other, so these parts are not calcified. The calcified parts between the whorls in longitudinal and oblique sections have the shape of the letter "T" (Pl. 20 ![]() , figs. 1, 4; Pia, 1940, Pl. 1, fig. 7; Bystrick�, 1957, Pl. IV, figs. 3-4; 1964, Pl. X, fig. 1; Pl. XI, figs. 3, 8; Pl. XII, figs. 1, 3; 1986, Pl. I, figs. 1-2; Herak, 1965, Pl. XI, fig. 5). This shape actually represents the outline of the space between the whorls and the stem cell (Fig. 10
, figs. 1, 4; Pia, 1940, Pl. 1, fig. 7; Bystrick�, 1957, Pl. IV, figs. 3-4; 1964, Pl. X, fig. 1; Pl. XI, figs. 3, 8; Pl. XII, figs. 1, 3; 1986, Pl. I, figs. 1-2; Herak, 1965, Pl. XI, fig. 5). This shape actually represents the outline of the space between the whorls and the stem cell (Fig. 10 ![]() ). In some specimens the calcification was slightly weaker, so the shape of the letter "T" is less pronounced (Pl. 8
). In some specimens the calcification was slightly weaker, so the shape of the letter "T" is less pronounced (Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() , figs. 2, 5-6; Bystrick�, 1957, Pl. IV, fig. 5; 1964, Pl. XI, fig. 5), or in the longitudinal and oblique sections between the whorls we observe "spikes", which shows that calcification did not reach the stem cell (Pl. 9
, figs. 2, 5-6; Bystrick�, 1957, Pl. IV, fig. 5; 1964, Pl. XI, fig. 5), or in the longitudinal and oblique sections between the whorls we observe "spikes", which shows that calcification did not reach the stem cell (Pl. 9 ![]() , figs. 4, 8; Pl. 20
, figs. 4, 8; Pl. 20 ![]() , fig. 3; Pl. 21
, fig. 3; Pl. 21 ![]() , fig. 1; Bystrick�, 1957, Pl. IV, fig. 6; 1964, Pl. X, fig. 9; Pl. XI, fig. 1), or there was a partial dissolution or erosion of the skeleton (Pl. 9
, fig. 1; Bystrick�, 1957, Pl. IV, fig. 6; 1964, Pl. X, fig. 9; Pl. XI, fig. 1), or there was a partial dissolution or erosion of the skeleton (Pl. 9 ![]() , figs. 1, 3; Bystrick�, 1964, Pl. X, fig. 3, Pl. XII, fig. 2; Assereto et al., 1965, Pl. 78.b; Pl. 79, figs. 2-4). In tangential sections, the calcified parts between the whorls are represented by "bars", while the whorls are represented by unconnected or connected cavities (Pl. 9
, figs. 1, 3; Bystrick�, 1964, Pl. X, fig. 3, Pl. XII, fig. 2; Assereto et al., 1965, Pl. 78.b; Pl. 79, figs. 2-4). In tangential sections, the calcified parts between the whorls are represented by "bars", while the whorls are represented by unconnected or connected cavities (Pl. 9 ![]() , figs. 1 below, 4 below, 8 above; Pl. 20
, figs. 1 below, 4 below, 8 above; Pl. 20 ![]() , fig. 1 top; Pl. 21
, fig. 1 top; Pl. 21 ![]() , fig. 1 top; Bystrick�, 1964, Pl. XI, fig. 3 bottom; Pl. XII, fig. 1 top; Assereto et al., 1965, Pl. 78.b; Pl. 79, figs. 2 down, 3 down). The presence of "bars" is not unique to var. intusannulata, although it is more pronounced in this variety, but also occurs in all species and varieties of both described genera, in which the whorls are relatively close, and calcification reaches the stem cell, for example, in P. pauciforata (Pl. 1, fig. 15 bottom left), O. cornuta (Pl. 9
, fig. 1 top; Bystrick�, 1964, Pl. XI, fig. 3 bottom; Pl. XII, fig. 1 top; Assereto et al., 1965, Pl. 78.b; Pl. 79, figs. 2 down, 3 down). The presence of "bars" is not unique to var. intusannulata, although it is more pronounced in this variety, but also occurs in all species and varieties of both described genera, in which the whorls are relatively close, and calcification reaches the stem cell, for example, in P. pauciforata (Pl. 1, fig. 15 bottom left), O. cornuta (Pl. 9 ![]() , fig. 11; Pl. 10
, fig. 11; Pl. 10 ![]() , fig. 9; Pl. 21
, fig. 9; Pl. 21 ![]() , fig. 5), O. praealpina (Pl. 13
, fig. 5), O. praealpina (Pl. 13 ![]() , fig. 8; Pl. 14
, fig. 8; Pl. 14 ![]() , fig. 1; Pl. 25, figs. 1-2), and in some specimens of O. pilosa pilosa (Pl. 7
, fig. 1; Pl. 25, figs. 1-2), and in some specimens of O. pilosa pilosa (Pl. 7 ![]() , fig. 3 left; Pl. 21
, fig. 3 left; Pl. 21 ![]() , fig. 2). The outer surface of the thallus is generally more or less smooth (Pl. 9
, fig. 2). The outer surface of the thallus is generally more or less smooth (Pl. 9 ![]() , figs. 1-6; Pl. 17
, figs. 1-6; Pl. 17 ![]() , fig. 3; Bystrick�, 1957, Pl. IV, fig. 4; 1964, Pl. X, figs. 1, 9), often slightly undulated, with protruding parts between the whorls and concave within the whorls, i.e., between two rows of laterals (Pl. 9
, fig. 3; Bystrick�, 1957, Pl. IV, fig. 4; 1964, Pl. X, figs. 1, 9), often slightly undulated, with protruding parts between the whorls and concave within the whorls, i.e., between two rows of laterals (Pl. 9 ![]() , fig. 5; Pia, 1940, Pl. 1, fig. 7; Bystrick�, 1957, Pl. IV, figs. 3, 5; 1964, Pl. XI, figs. 3, 5). This undulation is far more pronounced in O. cornuta and O. varicans. Specimens subjected to erosion or micritisation have an uneven outer surface (Pl. 8
, fig. 5; Pia, 1940, Pl. 1, fig. 7; Bystrick�, 1957, Pl. IV, figs. 3, 5; 1964, Pl. XI, figs. 3, 5). This undulation is far more pronounced in O. cornuta and O. varicans. Specimens subjected to erosion or micritisation have an uneven outer surface (Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() , figs. 7-8; Pl. 21
, figs. 7-8; Pl. 21 ![]() , fig. 1). The specimens on Pl. 20
, fig. 1). The specimens on Pl. 20 ![]() ,
figs. 1, 4, are peculiar due to the more intense calcification, and some laterals extends into the spikes, so that some parts of the thallus show pronounced undulation.
,
figs. 1, 4, are peculiar due to the more intense calcification, and some laterals extends into the spikes, so that some parts of the thallus show pronounced undulation.
It should be noted that, despite the name of this variety, this feature represents intusannulation only in its morphological sense, since it is the result of a difference in the extent of calcification within the whorl and between two adjacent whorls, not a reflection of the algal biological structure. True intusannulation is present, for example, in the species Diplopora phanerospora (Pia, 1920) and is a reflection of the stem cell shape, so it has also a biological sense.
The shape of the laterals is identical to that of var. pilosa and is designated as
piriform. The shape of the laterals in this variety also varies to some extent from slightly thicker (Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() , fig. 7; Bystrick�, 1957, Pl. IV,
figs. 3-5; 1964, Pl. X, fig. 1; Pl. XI, figs. 3, 5, 8; Pl. XII,
figs. 2-3), slightly thinner (Pl. 9
, fig. 7; Bystrick�, 1957, Pl. IV,
figs. 3-5; 1964, Pl. X, fig. 1; Pl. XI, figs. 3, 5, 8; Pl. XII,
figs. 2-3), slightly thinner (Pl. 9 ![]() ,
figs. 1-6, 8; Pl. 20
,
figs. 1-6, 8; Pl. 20 ![]() ,
figs. 1, 4; Bystrick�, 1957, Pl. IV, fig. 6; 1964, Pl. X,
figs. 3, 9; Pl. XI, fig. 1; 1966, Pl. VI, figs. 2-3; Assereto et al., Pl. 78.a) and relatively thin (Pl. 17
,
figs. 1, 4; Bystrick�, 1957, Pl. IV, fig. 6; 1964, Pl. X,
figs. 3, 9; Pl. XI, fig. 1; 1966, Pl. VI, figs. 2-3; Assereto et al., Pl. 78.a) and relatively thin (Pl. 17 ![]() , fig. 3; Pl. 21
, fig. 3; Pl. 21 ![]() , fig. 1; Assereto et al., Pl. 78.b; Pl. 79,
figs. 2-4). The laterals at the top part of the thallus are thinner, i.e., of the trichoform type, which is nicely seen in the upper parts of the photographs on Pl. 9
, fig. 1; Assereto et al., Pl. 78.b; Pl. 79,
figs. 2-4). The laterals at the top part of the thallus are thinner, i.e., of the trichoform type, which is nicely seen in the upper parts of the photographs on Pl. 9 ![]() ,
figs. 1, 5 and Bystrick� (1957, Pl. IV, fig. 6; 1964, Pl. XI, fig. 1; Pl. XII, fig. 3). The reason is that in that way the alga in the top part of the thallus
(the most exposed to light) has a large number of laterals on a small area, so it has a larger area for
photosynthesis. The same adaptation is present, for example, in the living Neomeris annulata
(Cramer, 1891; Berger & Kaever, 1992).
The laterals of this variety also carried trichoblasts on the top, but they are usually not preserved in the
fossils. The existence of trichoblasts is indicated by spikes in the continuation of certain laterals (Pl. 20
,
figs. 1, 5 and Bystrick� (1957, Pl. IV, fig. 6; 1964, Pl. XI, fig. 1; Pl. XII, fig. 3). The reason is that in that way the alga in the top part of the thallus
(the most exposed to light) has a large number of laterals on a small area, so it has a larger area for
photosynthesis. The same adaptation is present, for example, in the living Neomeris annulata
(Cramer, 1891; Berger & Kaever, 1992).
The laterals of this variety also carried trichoblasts on the top, but they are usually not preserved in the
fossils. The existence of trichoblasts is indicated by spikes in the continuation of certain laterals (Pl. 20 ![]() ,
figs. 1, 4), which represent their calcified proximal parts. The laterals, as in other species of the
genus, are arranged in double whorls, that is best seen on Pl. 8
,
figs. 1, 4), which represent their calcified proximal parts. The laterals, as in other species of the
genus, are arranged in double whorls, that is best seen on Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() ,
figs. 1-8; Pl. 20
,
figs. 1-8; Pl. 20 ![]() ,
figs. 1-4; Pl. 21
,
figs. 1-4; Pl. 21 ![]() , fig. 1. The whorls are somewhat closer to each other than in the type
variety, but there is a whole transitional sequence. The laterals in the two rows within the whorl diverge from each
other, and the bisector of the angle between the two rows is generally slightly inclined
upwards. The laterals are located mainly one above the other, which can be seen by the fact that the longitudinal sections evenly intersect the laterals in both rows within the
whorl, even those in the adjacent whorls (Pl. 8
, fig. 1. The whorls are somewhat closer to each other than in the type
variety, but there is a whole transitional sequence. The laterals in the two rows within the whorl diverge from each
other, and the bisector of the angle between the two rows is generally slightly inclined
upwards. The laterals are located mainly one above the other, which can be seen by the fact that the longitudinal sections evenly intersect the laterals in both rows within the
whorl, even those in the adjacent whorls (Pl. 8 ![]() , fig. 15; Pl. 9
, fig. 15; Pl. 9 ![]() , fig. 1; Pl. 10
, fig. 1; Pl. 10 ![]() , fig. 3; Pl. 20
, fig. 3; Pl. 20 ![]() ,
figs. 1, 4; Bystrick�, 1957, Pl. IV, fig. 5; 1964, Pl. X, fig. 1; Pl. XI, fig. 3; Pl. XII,
figs. 1, 3; 1986, Pl. I, fig. 1).
,
figs. 1, 4; Bystrick�, 1957, Pl. IV, fig. 5; 1964, Pl. X, fig. 1; Pl. XI, fig. 3; Pl. XII,
figs. 1, 3; 1986, Pl. I, fig. 1).
Emended diagnosis: Cylindrical thallus with a smooth inner surface and a more or less flat, rarely undulated outer surface. The laterals are of the piriform type, and on their appex they carried trichoblasts. The laterals are arranged in double whorls consisting of two very close rows of laterals. Adjacent whorls are relatively close, so the calcified parts between the whorls have a specific appearance in some sections: in longitudinal and oblique sections they have the appearance of "spikes", and when the calcification reaches to the stem cell they look like the letter "T", while in tangential sections they have the appearance of "bars" between the cavities of the laterals. The laterals touch each other inside the whorl in the proximal part and stand mostly one above the other. They are inclined very slightly upwards. In the apex of the thallus the laterals are trichoform.
|
|
Figure 10: Oligoporella pilosa var. intusannulata Pia. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous
skeleton. The longitudinal section (A) also shows the top part of the thallus with slightly thinner
laterals. Trichoblasts are shown on some laterals. In the lowest part of the longitudinal section (A), calcification is shown only in the distal part of the
laterals. The tangential section (C) shows the sections of the proximal (upper
whorl), central (middle whorl) and distal parts of the laterals (lower whorl). |
New diagnosis of the species Oligoporella pilosa Pia: Cylindrical thallus with a smooth inner and more or less smooth or slightly undulated outer surface. The laterals are piriform with a wide rounded proximal part and a gradual thinning towards the distal end, and on their apex they bear trichoblasts. They are arranged in double whorls consisting of two close rows of laterals. They are generally slightly inclined upwards. In the apex of the thallus the laterals are trichoform. The thickness of the laterals and the distance between the whorls can vary. The parts of the skeleton between the whorls in specimens with closer whorls have the appearance of "spikes" in longitudinal and oblique cross-sections or, when calcification reaches all the way to the stem cell, the letter "T" and belong to var. intusannulata.
The following table compares the two varieties:
| variety | stem cell | laterals | whorls | calcification |
| pilosa | smooth | alternately | spaced | weaker |
| intusannulata | smooth | above one another | rather close | stronger |
Similarities and differences: Oligoporella pilosa is most similar to other species of the genus Oligoporella. It differs from O. dissita, O. minutula and O. praealpina by the slightly thicker laterals and the absence of more pronounced undulation. It differs from O. intusannulata by a relatively smaller divergence of the laterals and a relatively symmetrical inclination of the laterals in the whorl. It differs from O. cornuta and O. varicans by a smaller divergence of laterals and their rather different shape. It differs from O. laevis because it has thinner laterals and a different type of calcification.
O. pilosa is somewhat similar in shape to P. pauciforata, but the latter has single
whorls, so it is easy to distinguish in oblique and longitudinal sections. The problem is in cross-sections, where differentiation is difficult or impossible, so such forms are most often referred to as Physoporella sp. (Pl. 1 ![]() , fig. 14; Pl. 19
, fig. 14; Pl. 19 ![]() , fig. 5) or Oligoporella sp. (Pl. 23
, fig. 5) or Oligoporella sp. (Pl. 23 ![]() , fig. 5). Due to the same stratigraphic range and the common occurrence of these two
species, this is of little importance.
, fig. 5). Due to the same stratigraphic range and the common occurrence of these two
species, this is of little importance.
5.2.2. Oligoporella intusannulata (Hurka, 1967), n. comb.
(= Physoporella intusannulata Hurka, 1967, nom. nud.)
Pl. 17 ![]() ,
figs. 1, 5
,
figs. 1, 5
1957 - Oligoporella pilosa varicans - Bystrick�, p. 230, Pl. V, fig. 2
1967 (nom. nud.) Physoporella intusannulata n. sp. - Hurka, p. 48-78, Figs. 3.a-g, 4, 6, 7.a-g, 9-10, 11.a-e; Pl. 6, figs. 1-5.
2021 non - Oligoporella intusannulata (= Oligoporella pilosa var. intusannulata) - Gawlick et al., Pl. 11, figs. b-c, e, g, i.
Origin of the name: After the uneven inner surface of the calcareous thallus, which Pia (1920) called intusannulation (Latin "intus" = inside; "annulus" = ring). This alga has double whorls, so it was transferred to the genus Oligoporella, in which there is already an alga of a very similar name Oligoporella pilosa var. intusannulata. Notwithstanding the possibility of confusion, both names must be retained, since they are legitimately established.
Studied material: The material of Oligoporella intusannulata described by Hurka (1967) originates from a ridge east of Bagni di
Piandim�ia (Bad Maistatt) near Vilabasse (Niederdorf) in the Puster Valley (Dolomites, NE
Italy). The author defined as types "thin-sections UG 1/1-UG 1/45, Nr. UG 1/16". This is not in accordance with the rules of botanical nomenclature, since only one specimen for fossil taxa should be chosen as the type (Art. 8.5
ICN, Turland et al., 2018), so as a lectotype I determine the specimen from Pl. 17 ![]() , fig. 1 (= Hurka, 1967, Pl. 6, fig. 2), since it best represents the typical characteristics of this
species. Hurka did not state in which thin-section this specimen was. The thin-sections are stored in the "Institut f�r spezielle Botanik der Universit�t T�bingen".
, fig. 1 (= Hurka, 1967, Pl. 6, fig. 2), since it best represents the typical characteristics of this
species. Hurka did not state in which thin-section this specimen was. The thin-sections are stored in the "Institut f�r spezielle Botanik der Universit�t T�bingen".
History of investigations: Hurka (1967, p. 77) gives a diagnosis of a new species: "External articulation with all transitions: unarticulated, undulation, annulation (one double whorl per segment); Tubes usually between 3 and 6 mm long and 1 mm wide; Pores pirifer, slender cone-shaped with an average diameter (p) of 0.16 mm, with excessive distribution, e.g., high constancy of p; Pores arranged in distinct double whorls, the branches of the lower row being disadvantaged in terms of length growth compared to those of the upper row; mostly 16 pores per double whorl, both rows equally occupied (8); upper row inclined an average of 47� to the thallus axis, lower row 77�, whorl distance h in the single individual more constant than in the population, where h = 0.7 mm (average); Stem cell undulated, widened between two double whorls, narrowed in the insertion area of the whorl (intusannulation with average st1 = 0.5 mm and st2 = 0.6 mm). Correlations between p and 1, D and k, st2 and k, st2 and D, st2 and h."
(In German: "�u�ere Gliederung mit allen �berg�ngen : ungegliedert Undulation Annulation (1 Doppelwirtel pro Gliedh�he); R�hrchen meist zwischen 3 und 6 mm lang und 1 mm breit; Poren pirifer, schlank kegelf�rmig mit einem p von durchschnittlich 0,16 mm, Verteilung exzessiv, d. h. hohe Konstanz von p; Poren in deutlichen Doppelwirteln angeordnet, dabei die �ste der unteren Reihe gegen�ber denen der oberen Reihe im L�ngenwachstum benachteiligt; meist 16 Poren pro Doppelwirtel, die beiden Reihen gleich besetzt (8); obere Reihe durchschnittlich 47� gegen die Stammachse geneigt, untere 77�, Wirtelabst�nde h am einzelnen Individuum konstanter als in der Population, dort h = 0,7 mm (Durch-schnitt); Stammzelle unduliert, zwischen je zwei Doppelwirteln erweitert, im Insertionsbereich der Wirtel verengt (Intusannulation mit durch-schnittlichen st1 = 0,5 mm und st2 = 0,6 mm); Korrelationen zwischen p und 1, D und k, st2 und k, st2 und D, st2 und h.")
Description: Although the author described this species in great detail and statistically analysed 600 specimens of this
alga, he illustrates only five of them (two longitudinal and one oblique, tangential and transverse sections), so it is not easy to confirm his
results. The thallus is cylindrical and straight. The outer surface of the thallus is slightly uneven (Pl. 17 ![]() , fig. 1). The part of that unevenness is certainly the result of diagenetic changes, but depressions or furrows between the whorls described by Hurka (1967) are occasionally
present. Narrow furrows between the whorls are more clearly seen on the tangential section (Hurka, 1967, Pl. 6, fig. 1). These furrows are probably the result of a large gap between the
whorls. Calcification begins around the laterals (to protect gametangia), so the part exactly in the middle between the whorls can sometimes remain with weaker calcification. Similar furrows or cracks can
occur, although less frequently, on the inner surface of the thallus. Although the author cites undulation and annulation in the diagnosis of the
species, these indentations, furrows and cracks, due to their irregularity, uneven appearance and
origin, could rather be classified as fissuration, which occurs in the genus Oligoporella and has little taxonomic
significance. The inner surface of the thallus is smooth with a pronounced intusannulation (or
better: "intusundulation"). This intusannulation is the result of the shape of the stem cell (Fig. 11
, fig. 1). The part of that unevenness is certainly the result of diagenetic changes, but depressions or furrows between the whorls described by Hurka (1967) are occasionally
present. Narrow furrows between the whorls are more clearly seen on the tangential section (Hurka, 1967, Pl. 6, fig. 1). These furrows are probably the result of a large gap between the
whorls. Calcification begins around the laterals (to protect gametangia), so the part exactly in the middle between the whorls can sometimes remain with weaker calcification. Similar furrows or cracks can
occur, although less frequently, on the inner surface of the thallus. Although the author cites undulation and annulation in the diagnosis of the
species, these indentations, furrows and cracks, due to their irregularity, uneven appearance and
origin, could rather be classified as fissuration, which occurs in the genus Oligoporella and has little taxonomic
significance. The inner surface of the thallus is smooth with a pronounced intusannulation (or
better: "intusundulation"). This intusannulation is the result of the shape of the stem cell (Fig. 11 ![]() ), which was slightly wider between the whorls (Fig. 11
), which was slightly wider between the whorls (Fig. 11 ![]() ; Pl. 17
; Pl. 17 ![]() , fig. 1). The calcification reached near to the stem
cell, not next to it, as indicated by the non-calcified basal parts of the
laterals.
, fig. 1). The calcification reached near to the stem
cell, not next to it, as indicated by the non-calcified basal parts of the
laterals.
The laterals are piriform in shape and, as in other species of the genus Oligoporella, are arranged in double
whorls. Trichoblasts are not observed on the illustrated material, but after comparison with other species of Oligoporella and Physoporella, they probably existed at the top of each
lateral. Hurka (1967) states a pronounced uniformity of pore diameter (p), which means that the laterals are of uniform shape. Some of the laterals illustrated by Hurka in his drawings (1967, Fig. 11) differ from typical piriform laterals by small protrusions ("protuberances") on the upper or lower side of the proximal part of the laterals (Fig. 11 ![]() A, first whorl on the
left). The existence of protuberances can also be seen in the cross section (Pl. 17
A, first whorl on the
left). The existence of protuberances can also be seen in the cross section (Pl. 17 ![]() , fig. 5) in the form of less calcified parts of the thallus at the level of
protuberances, although it can be due to a lack of calcification at the points of contact of the enlarged parts of the
laterals, as stated also by Hurka (1967). These protuberances possibly played a role in reproduction as
gametophores. Protuberances are not developed in all specimens, and that is why this is so difficult to
interpret; perhaps algae only created such structures under adverse conditions. A similar rationale may be for an unusual specimen of Physoporella aff. croatica (Pl. 19
, fig. 5) in the form of less calcified parts of the thallus at the level of
protuberances, although it can be due to a lack of calcification at the points of contact of the enlarged parts of the
laterals, as stated also by Hurka (1967). These protuberances possibly played a role in reproduction as
gametophores. Protuberances are not developed in all specimens, and that is why this is so difficult to
interpret; perhaps algae only created such structures under adverse conditions. A similar rationale may be for an unusual specimen of Physoporella aff. croatica (Pl. 19 ![]() , fig. 4; see
Chapter 5.1.2.). Hurka (1967) finds these protuberances
(he calls them "spurs") on 50% of the laterals, and within the same specimen there are laterals with and without
protuberances.
, fig. 4; see
Chapter 5.1.2.). Hurka (1967) finds these protuberances
(he calls them "spurs") on 50% of the laterals, and within the same specimen there are laterals with and without
protuberances.
The main characteristic of this species is the different inclination of the two rows of laterals in the whorl
(Pl. 17 ![]() ,
fig. 1). The lower row (β = 103�) diverges less from the whorl axis than the upper row (α = 47�). Laterals of the upper row are therefore slightly longer than those in the lower row. The laterals within one row and between two rows within the whorl mostly touch each other (Hurka, 1967, Fig. 6; Pl. 6, figs. 1, 5), and less often they are slightly spaced (Pl. 17
,
fig. 1). The lower row (β = 103�) diverges less from the whorl axis than the upper row (α = 47�). Laterals of the upper row are therefore slightly longer than those in the lower row. The laterals within one row and between two rows within the whorl mostly touch each other (Hurka, 1967, Fig. 6; Pl. 6, figs. 1, 5), and less often they are slightly spaced (Pl. 17 ![]() , fig. 5). The spacing of whorls is relatively large and fairly constant (h = 0.7 mm � 16%). The laterals of the two rows within the whorl stand one above the other (Hurka, 1967, Pl. 6, fig. 1), and probably also in the adjacent whorls, which can be recognised by the uniform shape of the laterals in longitudinal section (Pl. 17
, fig. 5). The spacing of whorls is relatively large and fairly constant (h = 0.7 mm � 16%). The laterals of the two rows within the whorl stand one above the other (Hurka, 1967, Pl. 6, fig. 1), and probably also in the adjacent whorls, which can be recognised by the uniform shape of the laterals in longitudinal section (Pl. 17 ![]() , fig. 1).
, fig. 1).
Emended diagnosis: Cylindrical thallus with slightly uneven or slightly undulated outer surface. The inner surface of the thallus is smooth and wavy (intusannulation), as a reflection of the shape of the stem cell, which was slightly narrower at the level of the whorls. The laterals are typical piriform and arranged in spaced double whorls. The laterals in the upper row diverge more from the whorl axis than those in the lower row and are therefore somewhat shorter. They mostly touch each other in the proximal part and stand one above the other. Occasionally, protuberances appear in the laterals.
Similarities and differences: Oligoporella intusannulata (Hurka), n. comb., is most similar to the species O. cornuta (Pia), nom. nov., from which it differs by the undulated inner surface of the thallus, and by the dissimilar divergence of the laterals. The dimensions of the thallus are also smaller (Table 1). It differs from O. pilosa mainly in the greater divergence of its laterals.
|
|
Figure 11: Oligoporella intusannulata (Hurka). Idealized longitudinal (A) and transverse (B) sections of the thallus copied from Hurka (1967, Figs. 1-2). Reconstruction of the organic part of the thallus is added. |
5.2.3. Oligoporella cornuta (Pia, 1935a), nom. nov.
(= Oligoporella pilosa var. varicans Pia, 1935a)
Pl. 9 ![]() , figs. 9-15; Pl. 10
, figs. 9-15; Pl. 10 ![]() , figs. 4, 6, 9-10; Pl. 21
, figs. 4, 6, 9-10; Pl. 21 ![]() , figs. 5-7
, figs. 5-7
1935a - Oligoporella pilosa var. varicans n. var. - Pia, p. 216-219, Figs. 21-27.
1940 - Oligoporella pilosa forma varicans (sic) - Pia, p. 4-5, Pl. 1, fig. 9.
1957 - Oligoporella pilosa varicans - Bystrick�, p. 230, Pl. IV, fig. 7; Pl. V, figs. 1, 3-5.
1964 - Oligoporella pilosa var. varicans.- Bystrick�, p. 114-115, Pl. XII, figs. 4-5.
1993 - Oligoporella pilosa varicans - Senowbari-Daryan et al., p. 200, 236, Pl. 52, fig. 7.2.; Pl. 56, figs. 1-5, 6.1, 7-9.
2003 - Physoporella varicans - Piros, p. 58, Pl. D-III, fig. 2.
Origin of the name: This species is primarily described as Oligoporella pilosa var. varicans (Pia, 1935a). I transfer this taxon to the rank of species (see below), but it is not possible to keep its name because the species Physoporella varicans (Pia, 1935a) is transferred into the genus Oligoporella (see below), and since it is primarily described at the rank of species, it has priority to keep the specific name (Art. 11.2 ICN, Turland et al., 2018). So I chose a new name for this species, according to the shape of the laterals arraged in double whorls (Fig. 12 ![]() ) that resemble horns (Latin "cornus" = horn).
) that resemble horns (Latin "cornus" = horn).
Studied material: The material described by Pia (1935a) originates from Bosnia and Herzegovina: thin-section 235b is from Ograda (site "t") on the road Are�in brijeg - Grab, 574 and 577a are from the village of Haliluci, east of Sarajevo (site "r"). The specimen cited by Pia (1935a, Fig. 26) is from site "r", but has no thin-section number, I have not found it in his collection.
The material from Croatia, with which I supplemented the research of this species, includes specimens from the Belski dol quarry on Ivan�čica Mt. (Pl. 21 ![]() , figs. 6-7), and from the surroundings of Udbina in Lika from the collection of Branko Sokač (Pl. 21
, figs. 6-7), and from the surroundings of Udbina in Lika from the collection of Branko Sokač (Pl. 21 ![]() , fig. 5).
, fig. 5).
Since Pia did not designate a type specimen, among them I choose the one from Pia (1935a, Fig. 25 (= this paper, Pl. 9 ![]() , fig. 10) as a lectotype, and the other specimens from the same sample 235b are isotypes (Pl. 9
, fig. 10) as a lectotype, and the other specimens from the same sample 235b are isotypes (Pl. 9 ![]() , figs. 13 -14).
, figs. 13 -14).
History of investigations: Pia (1935a, p. 216-219) described the new variety Oligoporella pilosa var. varicans: "In this variety I include the specimens of Oligoporella pilosa, in which the inclination of the whorled branches down and up is strongest. It is thus primarily characterized by closeness to the var. subvaricans. A closer examination shows, however, that it differs quite clearly from the type variety and from the just mentioned similar variety also in other characteristics, especially in the dimensions. Of all the subspecies of the Oligoporella pilosa, the var. varicans could most likely have been viewed as a separate species. To show this, let us first analyse the measurements again. Above all, they show that Oligoporella pilosa varicans is much smaller than Ol. p. typica. The difference between the mean values is 0.64 � 0.091 mm. It is therefore to be regarded as objective with a probability of 100%. In most cases, the distance between the whorls is greater than that of the other varieties of Oligoporella pilosa. This is easy to understand in view of the strong divergence of the branches belonging to the same whorl. The fact that the parts of the skeleton protruding towards the inside between the whorls are relatively wide is related to this greater distance between the whorls. It is not uncommon for them to be so straight and smooth on the inside and one has to conclude that they reached up to the stem cell. What is peculiar is the paired arrangement of pores in the (actually not very typical) specimen of Pia (1935a, Fig. 26), which do not belong to the same whorl, but to two consecutive ones. The sum of all these characteristics makes Oligoporella pilosa varicans quite easy to identify."
(In German: "Zu dieser Variet�t stelle ich die Exemplare von Oligoporella pilosa, bei denen die Neigung der Wirtel�ste gegen unten und oben am st�rksten ausgebildet ist. Sie wird also zun�chst durch eine Steigerung des Merkmales der var. subvaricans gekennzeichnet. Eine n�here Betrachtung ergibt aber, dass sie vom Typus und von der eben genannten, ihm n�her stehenden Variet�t auch in anderen Merkmalen, besonders in den Abmes-sungen, recht deutlich verschieden ist. Von allen Unterarten der Oligoporella pilosa h�tte die var. varicans jedenfalls am ehesten als eigene Art angesehen werden k�nnen. Um das zu zeigen, seien zun�chst wieder die Messungen zusammengestellt. Vor allem zeigt diese Tabelle, dass Oligoporella pilosa varicans wesentlich kleiner ist, als O. p. typica. Der Unterschied der Mittelwerte betr�gt. Differenz = 0.64 + 0.091 mm. Er ist also mit einer Wahrscheinlichkeit von 100% als objektiv anzusehen. Der Abstand der Wirtel ist in weitaus den meisten F�llen gr�sser, als bei den anderen Variet�ten von Oligoporella pilosa (vergl. die Messungstabellen). Das ist in anbetracht der starken Divergenz der zum selben Wirtel geh�rigen Aeste leicht ver-st�ndlich. Mit diesem gr�sseren Wirtelabstand h�ngt wieder zusammen, dass die zwischen den Wirteln gegen innen vorra-genden Teile des Skelettes verh�ltnism�ssig breit sind. Nicht selten sind sie innen so gerade und glatt begrenzt, dass man wohl schliessen muss, sie h�tten bis an die Stammzelle herangereicht. Eigent�mlich ist die an dem (�brigens nicht sehr typischen) Exemplar Fig. 26 ersichtliche, paarweise Zusammenordnung von Poren, die nicht zum selben Wirtel, sondern zu je zwei aufeinan-derfolgenden geh�ren. Die Summe aller dieser Merkmale macht Oligoporella pilosa varicans ziemlich leicht kenntlich.")
Bystrick� (1964, p. 114-115) gives a short diagnosis of this variety: "The pores in double-rowed whorls diverge strongly. The distances between the whorls are greater than in other varieties, therefore the calcareous strips between the whorls are quite high."
Description: Thallus of Oligoporella cornuta (Pia, 1935a), nom. nov., is cylindrical and straight (Pl. 9 ![]() , fig. 10; Pl. 21
, fig. 10; Pl. 21 ![]() , fig. 6), occasionally slightly curved (Pl. 21
, fig. 6), occasionally slightly curved (Pl. 21 ![]() , fig. 7; Pia, 1935a, Fig. 26). The outer side of the thallus is usually slightly undulated (Pl. 9
, fig. 7; Pia, 1935a, Fig. 26). The outer side of the thallus is usually slightly undulated (Pl. 9 ![]() , figs. 10-12, 15; Pia, 1935a, Fig. 26; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, figs. 3, 5; 1964, Pl. XII, fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1, 3-5, 9) so that the recessed (concave) parts are inside the whorls and the protruding (convex) parts are at the contact of the upper and lower row of adjacent whorls (Fig. 12
, figs. 10-12, 15; Pia, 1935a, Fig. 26; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, figs. 3, 5; 1964, Pl. XII, fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1, 3-5, 9) so that the recessed (concave) parts are inside the whorls and the protruding (convex) parts are at the contact of the upper and lower row of adjacent whorls (Fig. 12 ![]() ), thus following the morphology of the organic part of the thallus. In some cases, spikes may appear on the surface of the thallus (Pl. 21
), thus following the morphology of the organic part of the thallus. In some cases, spikes may appear on the surface of the thallus (Pl. 21 ![]() , figs. 5-6; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2), which is the result of calcification of more distal parts of the laterals. The outer surface of the specimen on Pl. 9
, figs. 5-6; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2), which is the result of calcification of more distal parts of the laterals. The outer surface of the specimen on Pl. 9 ![]() , fig. 9 is relatively flat due to slightly weaker calcification, while on the specimen on Pl. 21
, fig. 9 is relatively flat due to slightly weaker calcification, while on the specimen on Pl. 21 ![]() , fig. 7 it is relatively uneven due to diagenetic changes, but such phenomena are present in most of the algae described here. The inner side of the thallus is, in well-preserved specimens, mostly smooth and flat (Pl. 9
, fig. 7 it is relatively uneven due to diagenetic changes, but such phenomena are present in most of the algae described here. The inner side of the thallus is, in well-preserved specimens, mostly smooth and flat (Pl. 9 ![]() , figs. 11-12, 15; Pl. 21
, figs. 11-12, 15; Pl. 21 ![]() , fig. 5; Bystrick�, 1957, Pl. V, figs. 3, 5; 1964, Pl. XII, fig. 5; 1986, Pl. I, fig. 3). It is thus evident that the calcification reached very close to the stem cell. A very interesting specimen is shown by Bystrick� (1957, Pl. IV, fig. 7) on which a thin micritic layer is visible; it does not represent a micritisation envelope since it is of uniform thickness and completely flat and smooth, but probably represents the primary micritic envelope between the stem cell and calcareous thallus. This interpretation is also indicated by the basal pores of the laterals that break through this layer. This type of micritic envelope in the studied material has been found only in Physoporella croatica (see chapter 5.1.2), and is otherwise characteristic of some other genera and species, such as Salpingoporella dinarica (see discussion in Sokač, 1996). Some specimens of O. cornuta have a pronounced wrinkling of the inner side of the thallus of very small amplitude (Pl. 21
, fig. 5; Bystrick�, 1957, Pl. V, figs. 3, 5; 1964, Pl. XII, fig. 5; 1986, Pl. I, fig. 3). It is thus evident that the calcification reached very close to the stem cell. A very interesting specimen is shown by Bystrick� (1957, Pl. IV, fig. 7) on which a thin micritic layer is visible; it does not represent a micritisation envelope since it is of uniform thickness and completely flat and smooth, but probably represents the primary micritic envelope between the stem cell and calcareous thallus. This interpretation is also indicated by the basal pores of the laterals that break through this layer. This type of micritic envelope in the studied material has been found only in Physoporella croatica (see chapter 5.1.2), and is otherwise characteristic of some other genera and species, such as Salpingoporella dinarica (see discussion in Sokač, 1996). Some specimens of O. cornuta have a pronounced wrinkling of the inner side of the thallus of very small amplitude (Pl. 21 ![]() , fig.
7 above; Pia, 1935a, Fig. 26; Bystrick�, 1964, Pl. XII, fig. 7 Senowbari-Daryan et al., 1993, Pl. 56, fig. 1). This shallow intusannulation has already been described in Physoporella pauciforata (see
Chapter 5.1.1.) and was interpreted as a fine stem cell wrinkling, possibly due to environmental factors. Specimens with a wrinkled inner surface do not appear to have a smaller spacing between the whorls, as might be expected. Algae, thus, appear to arrange their laterals with optimal density regardless of whether the stem cell is smooth or wrinkled. Calcification between the whorls, that follows the shape of the laterals and the stem cell, similar to that of O. pilosa var. intusannulata, has the shape of the letter "T" in longitudinal sections, or "bars" in tangential sections (Pl. 9
, fig.
7 above; Pia, 1935a, Fig. 26; Bystrick�, 1964, Pl. XII, fig. 7 Senowbari-Daryan et al., 1993, Pl. 56, fig. 1). This shallow intusannulation has already been described in Physoporella pauciforata (see
Chapter 5.1.1.) and was interpreted as a fine stem cell wrinkling, possibly due to environmental factors. Specimens with a wrinkled inner surface do not appear to have a smaller spacing between the whorls, as might be expected. Algae, thus, appear to arrange their laterals with optimal density regardless of whether the stem cell is smooth or wrinkled. Calcification between the whorls, that follows the shape of the laterals and the stem cell, similar to that of O. pilosa var. intusannulata, has the shape of the letter "T" in longitudinal sections, or "bars" in tangential sections (Pl. 9 ![]() , fig. 11; Pl. 21
, fig. 11; Pl. 21 ![]() , fig. 5).
, fig. 5).
The shape of the laterals is piriform with wide a proximal part and a gradual thinning outwards. The laterals are evidently arranged in the double whorls. The shape of the laterals in the double whorls is best seen in longitudinal and oblique sections (Pl. 9 ![]() , figs. 10-11, 15; Pl. 21
, figs. 10-11, 15; Pl. 21 ![]() , fig. 5; Pia, 1935a, Fig. 26; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, figs. 3, 5; 1964, Pl. XII, figs. 5, 7; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1-3). In the proximal part, laterals in two rows inside the whorl are compressed together so there is no calcification between them. Distally laterals from two rows diverge and are slightly bent. The laterals from the lower and upper rows of adjacent whorls are close to each other, and sometimes touch in the distal part (e.g., Pl. 9
, fig. 5; Pia, 1935a, Fig. 26; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, figs. 3, 5; 1964, Pl. XII, figs. 5, 7; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1-3). In the proximal part, laterals in two rows inside the whorl are compressed together so there is no calcification between them. Distally laterals from two rows diverge and are slightly bent. The laterals from the lower and upper rows of adjacent whorls are close to each other, and sometimes touch in the distal part (e.g., Pl. 9 ![]() , fig. 10 left). The upper row of laterals may diverge more strongly from the whorl axis than the lower row (Pl. 21
, fig. 10 left). The upper row of laterals may diverge more strongly from the whorl axis than the lower row (Pl. 21 ![]() , figs. 5, 7; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1, 3) or may have approximately equal divergence (Pl. 9
, figs. 5, 7; Bystrick�, 1957, Pl. IV, fig. 7; Pl. V, fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, figs. 1, 3) or may have approximately equal divergence (Pl. 9 ![]() , figs. 9-10; Bystrick�, 1964, Pl. XII, fig. 7; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2). The laterals, as with other species from the studied genera, continued into trichoblasts, which can be seen as the already mentioned spikes on the surface of the thallus (Pl. 21
, figs. 9-10; Bystrick�, 1964, Pl. XII, fig. 7; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2). The laterals, as with other species from the studied genera, continued into trichoblasts, which can be seen as the already mentioned spikes on the surface of the thallus (Pl. 21 ![]() ,
fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2). The laterals inside the whorl always stand one above the other, and mainly in the adjacent whorls, that are visible in tangential (Senowbari-Daryan et al., 1993, Pl. 56, figs. 3-4) and longitudinal sections (Pl. 9
,
fig. 5; Senowbari-Daryan et al., 1993, Pl. 56, fig. 2). The laterals inside the whorl always stand one above the other, and mainly in the adjacent whorls, that are visible in tangential (Senowbari-Daryan et al., 1993, Pl. 56, figs. 3-4) and longitudinal sections (Pl. 9 ![]() , figs. 9-10; Pl. 21
, figs. 9-10; Pl. 21 ![]() , figs. 6-7; Bystrick�, 1964, Pl. XII, fig. 7).
, figs. 6-7; Bystrick�, 1964, Pl. XII, fig. 7).
Due to the described characteristic structure of this alga, and especially the shape of the laterals and their arrangement, which differ from those of other species of the genus Oligoporella, I changed the rank of this taxon from variety to species, as already suggested by Pia (1935a).
Specimens from the Slovak Karst (Pia, 1940, p. 4-5, Pl. 1, fig. 9; Bystrick�, 1957, Pl. V, fig. 2) which deviate from typical O. cornuta according to a different inclination of the laterals (also described by Pia, 1935a), and the appearance of protuberances below and above the whorl, may represent a transitional form to O. intusannulata (Hurka). The main difference between the two illustrated specimens to O. intusannulata is the small distance between the whorls and the absence of intusannulation, so they are probably closer to their primary determination.
Emended diagnosis: Cylindrical thallus, usually with a slightly undulated outer surface, with concave parts inside the whorls, following the external morphology of the organic part of the algae. The inner side of the thallus is mostly smooth and flat, and sporadically and finely undulated due to the wrinkling of the stem cell. The laterals are piriform, and on their top they carried trichoblasts. They are arranged in double whorls. The upper and lower rows of laterals within the whorl diverge from each other and are slightly bent. The laterals are arranged mostly one above the other, and due to divergence, the lower and upper rows of the laterals of adjacent whorls are very close or touching.
Similarities and differences: This species according to the divergence of laterals stands between O. pilosa Pia and O. varicans (Pia). So far, no transitional forms have been found between these three species, so they are defined as separate taxa. In longitudinal and tangential sections, they can be easily recognized because of the different divergence of laterals that also causes different manifestations in the morphology of the calcareous thallus (see descriptions of species and reconstructions). Other species in which the laterals do not diverge (O. dissita, O. minutula, O. praealpina) are easily distinguished from O. cornuta.
|
|
Figure 12: Oligoporella cornuta (Pia, 1935a), nom. nov. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts are shown on some laterals. The longitudinal section (A) shows the wrinkled (I) and smooth stem cell (II) variants. The tangential section (B) shows the sections of the proximal (upper whorl) and distal parts of the laterals (lower whorl). A slightly undulated outer surface of the thallus is visible. |
5.2.4. Oligoporella varicans (Pia, 1935a), n. comb.
(= Physoporella varicans Pia, 1935a)
Pl. 10 ![]() , figs. ?1, 2, 8, 11-12; Pl. 23
, figs. ?1, 2, 8, 11-12; Pl. 23 ![]() , figs. 1-4, 6-8
, figs. 1-4, 6-8
1935a - Physoporella varicans n. sp. - Pia, p. 229-232, Figs. 43-44.
1957 - Physoporella varicans - Bystrick�, p. 233, Pl. VI, fig. 5.
1964 - Physoporella varicans - Bystrick�, p. 125-126, Pl. XVIII, fig. 1, from Bystrick� (1957, Pl. VI, fig. 5); Pl. XVIII, figs. 2-6.
2021 - Oligoporella cf. varicans - Gawlick et al., Pl. 5, fig. m.
Origin of the name: Possibly after divergence of laterals within the whorl (Latin "varico" = straddle, stride).
Studied material: The material described by Pia (1935a) originates from Bosnia and Herzegovina: thin-section 551 is from the village of Blizanac on Trebević Mt. (site "c"), 235a is from Ograda (site "t") on the road from Are�in brijeg - Grab, 587 is also from Ograda (but site "v").
Pia did not designate a type specimen, so among his material I choose the one from Pia (1935a, Fig. 44 = this paper, Pl. 10 ![]() , fig. 2 right) as the lectotype, while other specimens from the same sample 551 represent isotypes (Pl. 10
, fig. 2 right) as the lectotype, while other specimens from the same sample 551 represent isotypes (Pl. 10 ![]() , figs. 2 left, 11-12).
, figs. 2 left, 11-12).
The material from Croatia, with which I supplemented the research of this species, includes specimens from the Belski dol quarry in Ivan�čica Mt. (Pl. 23 ![]() , figs. 1-4, 6-8).
, figs. 1-4, 6-8).
History of investigations: Pia (1935a, p. 229, 231) describes the new species Physoporella varicans: "This relatively common species shows the most pronounced characteristics we have known so far in Oligoporella pilosa varicans. The branches are arranged in two-row whorls. They are firmly close to each other at their base, so that calcite cannot be deposited there. Immediately from the base they incline strongly upwards and downwards, so that they form an angle of, 20-30� to the stem cell and for a considerable length almost lie next to it. One is reminiscent of the Characeae cortex cells because of these characteristics - and especially in cross-sections, as in Pia (1935a) Figure 45. The upward and downward inclined branches of the adjacent whorls have their tips very close to each other, never seem to touch, and are always separated by a thin calcite lamella. Clear cracks are often present in the thallus. It is significant that these do not lie, as in other species, between the whorls, but always through the middle of the whorl between the two corresponding rows of pores. Incidentally, the shell is rather tender, mostly clearly undulated. These broad, shallow constrictions also lie above the whorls. As far as I could see, the pores are always closed to the outside."
(In German: "Diese gar nicht seltene Art zeigt die h�chste bisher beobachtete Entwicklung des Merkmales, das wir schon bei Oligoporella pilosa varicans kennen gelernt haben. Die Aeste sind in zweizeiligen Wirteln angeordnet. An ihrer Basis schliessen sie fest aneinander, so dass hier kein Kalk eingelagert werden kann. Sogleich aber vom Ursprung an neigen sie sich sehr stark gegen oben und unten, so dass sie mit der Stammzelle nur einen Winkel von, 20-30� bilden und ihr �ber eine ziemliche Strecke fast anliegen. Man wird durch diese Verh�ltnisse - und besonders durch Querschnitte, wie Textfig 45 - eirnigermassen an die Rindenzellen der Characeen erinnert. Die gegen oben und unten geneigten Aeste benachbarter Wirtel kommen einander mit den Spitzen sehr nahe, scheinen einander aber doch nie zu ber�hren, sondern immer durch eine d�nne Kalkplatte getrennt zu sein. Oft sind in der Schale deutliche Fissuren vorhanden. Bezeichnender Weise liegen diese aber nicht, wie bei anderen Arten, zwischen den Wirteln, sondern immer �ber den Wirtel-mitten, zwischen den beiden zusammengeh�rigen Zeilen von Poren. Uebrigens ist die Schale eher zart, meist deutlich unduliert. Auch diese breiten, seichten Einschn�rungen liegen �ber den Wirteln. Die Poren sind, so viel ich sah, immer gegen aussen geschlossen.")
Bystrick� (1964, p. 125-26, 190) largely repeats Pia's conclusions. Herak (1965) gives a brief description of this species: "An elongated cylindrical thallus carries whorls consisting of two sets of branches joined together at their base and diverging and bending towards the surface of the calcareous wall. Particularly characteristic is the convergence of adjacent whorls at the distal ends of the branches, creating external convexities that cause longitudinal undulation of the thallus. It depends on the orientation of the cross-section."
Description: The thallus is cylindrical and straight, often relatively long (Pl. 10 ![]() , fig. 2; Pl. 23
, fig. 2; Pl. 23 ![]() , figs. 1-4, 7-8). The outer surface of the thallus is distinctly undulated, following the shape of the laterals in the whorl. In some specimens this undulation is milder (Pl. 10
, figs. 1-4, 7-8). The outer surface of the thallus is distinctly undulated, following the shape of the laterals in the whorl. In some specimens this undulation is milder (Pl. 10 ![]() , fig. 2; Gawlick et al., Pl. 5, fig. b), and in others more pronounced (Pl. 23
, fig. 2; Gawlick et al., Pl. 5, fig. b), and in others more pronounced (Pl. 23 ![]() , fig. 1), since the distal ends of the laterals protrude more strongly outwards (Fig. 13
, fig. 1), since the distal ends of the laterals protrude more strongly outwards (Fig. 13 ![]() ). Calcification occurred only on the outside of the laterals and between the whorls, where it only occasionally reached near to the stem cell (Pl. 10
). Calcification occurred only on the outside of the laterals and between the whorls, where it only occasionally reached near to the stem cell (Pl. 10 ![]() , fig. 2 left). Due to this, the central cavity is relatively wide. As the calcareous thallus is relatively thin and intensely undulated, the mechanically weakest point is in the depressions inside the whorl, so it often cracks, and thin cracks appear in the thallus, which was also noticed by Pia (1935a). These cracks are not a diagnostic criterion, but the result of the mechanical properties of the calcareous thallus.
, fig. 2 left). Due to this, the central cavity is relatively wide. As the calcareous thallus is relatively thin and intensely undulated, the mechanically weakest point is in the depressions inside the whorl, so it often cracks, and thin cracks appear in the thallus, which was also noticed by Pia (1935a). These cracks are not a diagnostic criterion, but the result of the mechanical properties of the calcareous thallus.
The laterals are of the piriform type, but more elongated. They are evidently arranged in double whorls (Pl. 10 ![]() , figs. 2, 8, 11; Pl. 23
, figs. 2, 8, 11; Pl. 23 ![]() , figs. 1-4, 7-8). In the proximal part, the laterals of the two rows within the whorl diverge greatly - one going up and the other down - and bending distally, not to overlap with the laterals from the adjacent whorl. In the proximal part the laterals are compressed, which is best seen on Pl. 23
, figs. 1-4, 7-8). In the proximal part, the laterals of the two rows within the whorl diverge greatly - one going up and the other down - and bending distally, not to overlap with the laterals from the adjacent whorl. In the proximal part the laterals are compressed, which is best seen on Pl. 23 ![]() , fig. 1. They always stand one above the other, both inside the whorl and in the adjacent whorls and is visible in tangential (Pl. 23
, fig. 1. They always stand one above the other, both inside the whorl and in the adjacent whorls and is visible in tangential (Pl. 23 ![]() , fig. 1) and longitudinal sections (Pl. 10
, fig. 1) and longitudinal sections (Pl. 10 ![]() , fig. 2). Adjacent whorls are as close as the distinctly divergent laterals allow. The laterals carried trichoblasts, which is revealed by the spikes into which the pores on the Pl. 23
, fig. 2). Adjacent whorls are as close as the distinctly divergent laterals allow. The laterals carried trichoblasts, which is revealed by the spikes into which the pores on the Pl. 23 ![]() , figs. 1-2, 6-7 extend.
, figs. 1-2, 6-7 extend.
Emended diagnosis: Cylindrical thallus with pronounced undulation that follows the shape of the laterals, so recessed (concave) parts are within the whorl. Calcification covers the outside of the laterals and partially the space between the whorls. The laterals are of piriform type, arranged in distinctly diverging double whorls. In the proximal part the laterals are compressed together, and in the distal part they are bent, not to overlap with the laterals of the adjacent whorl. The laterals stand one above the other.
Similarities and differences: This species is very similar to the species Oligoporella cornuta (Pia), from which it differs mostly by the stronger divergence of the laterals within the whorl and, as a consequence, by a thinner and strongly undulated calcareous thallus. For the same reasons, but to a greater extent, it also differs from the species Oligoporella intusannulata (Hurka) and Oligoporella pilosa Pia.
|
|
Figure 13: Oligoporella varicans (Pia, 1935a), n. comb. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts in extension of laterals are shown on the upper whorl (I). The longitudinal section (A) shows variants with calcification close to the stem cell (III), away from the stem cell (II) and only in the distal part of the laterals (I). The tangential section (C) shows the sections of the proximal (upper whorl) and distal parts of the laterals (lower whorl). An intensely undulated outer surface of the thallus is visible. |
5.2.5. Oligoporella minutula (G�mbel, 1872), n. comb.
(= Physoporella minutula (G�mbel, 1872), Pia, 1912; = Gyroporella minutula G�mbel, 1872)
Pl. 12 ![]() , figs. 1, ?2, 3-9; Pl. 13
, figs. 1, ?2, 3-9; Pl. 13 ![]() , figs. 2, 4; Pl. 17
, figs. 2, 4; Pl. 17 ![]() , fig. 8
, fig. 8
1872 - Gyroporella minutula n. sp. (Gyroporella minuta n. sp. after plate legend) - G�mbel, p. 275, Pl. DIII, fig. 4.a-b.
1892 - Gyroporella balinensis n. sp. - Raciborski, p. 7-8, no figure
1912 - Physoporella minutula n. comb. - Pia, p. 45, 52, 54, 59, 80, Pl. VI, figs. 5-10, 12.
1920 - Physoporella minutula - Pia, p. 56, 178, Pl. II, figs. 19-22.
1958 - Physoporella minutula - Ellenberger, p. 178, 190, Pl. 6, figs. 18-19.
1958 - Clavaphysoporella minutula n. gen., n. comb. - Endo, p. 266, no figure.
1960 - Oligoporella pilosa forma balinensis n. comb., n. stat. - Pastwa-Leszczyńska & Śliwiński, p. 689, Fig. 9; Pl. III, fig. 14 (= Fig. 9)
1960 - Diplopora cf. silesiaca - Pastwa-Leszczyńska & Śliwiński, p. 687, Fig. 7; Pl. III, fig. 13 (= Fig. 7).
1961 - Physoporella minutula - Botteron, p. 63, Pl. VI, fig. 4 pars.
1964 - Physoporella cf. minutula.- Bystrick�, p. 129-131, Pl. XX, fig. 4.
2013 - Physoporella minutula (Physoporella cf. minuta n. sp. after plate legend) - Kotański, p. 81, Pl. XXI, figs. 29-44.
2013 - Physoporella cf. minutula - Rychliński et al., Pl. 2, fig. 9.
2021 - Oligoporella minutula - Gawlick et al., Pl. 5, figs. i, l, r.
2021 non - Oligoporella minutula (= Oligoporella dissita) - Gawlick et al., Pl. 15, figs. h-j.
Origin of the name: Probably after the smaller dimensions of this alga (Latin "minutulus" = very small, tiny).
Studied material: G�mbel (1872, p. 275) cites "black 'Reiflinger Kalk' from Reisalp (collection of the Geologische Reichsanstalt, Wien) and black limestone from Pertisau in Tyrol (shipment from Professor Pichler)" as typical strata and localities for Gyroporella minutula, but does not state which specimen is from which locality (G�mbel, 1872, Pl. D.III, figs. 4.a-b). G�mbel's material has been lost (Pia, 1912, p. 26; 1935a, p. 223). For other comments see Chapter 5.1.1. on P. pauciforata described from the same sites.
The material of P. minutula described by Pia (1912, p. 45; Pl. VI, figs. 5-110 = this paper, Pl. 12 ![]() , figs. 1, 3-9) originates from the vicinity of Mythen in the Zweckenalp area (Canton de Schwyz, Switzerland) from the Wetterstein Limestone of presumed Middle Triassic age: thin-sections XVI1 and XVI3. The specimen from thin-section XVI4 (Pl. 12
, figs. 1, 3-9) originates from the vicinity of Mythen in the Zweckenalp area (Canton de Schwyz, Switzerland) from the Wetterstein Limestone of presumed Middle Triassic age: thin-sections XVI1 and XVI3. The specimen from thin-section XVI4 (Pl. 12 ![]() , fig. 2 = Pia, 1912, Pl. VI, fig. 11) has a questionable affiliation to this species, it is possibly Oligoporella pilosa. I did not find the specimen from Pia (1912) Pl. VI, fig. 10, in Pia's collection, but it is very likely that it comes from the same sample.
, fig. 2 = Pia, 1912, Pl. VI, fig. 11) has a questionable affiliation to this species, it is possibly Oligoporella pilosa. I did not find the specimen from Pia (1912) Pl. VI, fig. 10, in Pia's collection, but it is very likely that it comes from the same sample.
Since G�mbel's material is lost, among Pia's material, as the most representative specimen, I chose the one on Pl. 12 ![]() , fig. 7 = Pia, 1912, Pl. VI, fig. 7 as neotype. Other specimens from sample XVI become isoneotypes. As with P. pauciforata (see according text), the designation of a "holotype" by Kotański (2013, p. 80) as the specimen figured by G�mbel (1872, Pl. D.III, fig. 4.a) is not valid according to rules of botanical nomenclature after Art. 9.3. of ICN (Turland et al., 2018) since a later author can only chose a lectotype from the original material and not a holotype.
, fig. 7 = Pia, 1912, Pl. VI, fig. 7 as neotype. Other specimens from sample XVI become isoneotypes. As with P. pauciforata (see according text), the designation of a "holotype" by Kotański (2013, p. 80) as the specimen figured by G�mbel (1872, Pl. D.III, fig. 4.a) is not valid according to rules of botanical nomenclature after Art. 9.3. of ICN (Turland et al., 2018) since a later author can only chose a lectotype from the original material and not a holotype.
The material of Physoporella minutula described by Pia (1920, p. 56, Pl. II, figs. 19-22) originates also from the Swiss Zweckenalps, but the age is determined as ? Anisian. In Pia's collection I found only a specimen from Pia (1920, Pl. II, fig. 20 = this paper, Pl. 13 ![]() , fig. 4) in thin-section 49. I did not find thin-section 65, and from the thin-section 15 I photographed one specimen of O. minutula, which was not illustrated in the cited work (Pl. 13
, fig. 4) in thin-section 49. I did not find thin-section 65, and from the thin-section 15 I photographed one specimen of O. minutula, which was not illustrated in the cited work (Pl. 13 ![]() , fig. 2).
, fig. 2).
The specimen described by Pastwa-Leszczyńska and Śliwiński (1960, p. 689, fig. 9 = Pl. III, fig. 14 = this paper, Pl. 17 ![]() , fig. 8) as Oligoporella pilosa forma balinensis originates from the Anisian dolomites of the Chrzanow area in Upper Silesia, Poland. The material is kept in the "Katedrze Zł�ż Rud na AGH w Krakowie", Krak�w. This specimen belongs to the species Oligoporella minutula by its characteristics, and especially by the shape and arrangement of the laterals, and should be considered a junior synonym.
, fig. 8) as Oligoporella pilosa forma balinensis originates from the Anisian dolomites of the Chrzanow area in Upper Silesia, Poland. The material is kept in the "Katedrze Zł�ż Rud na AGH w Krakowie", Krak�w. This specimen belongs to the species Oligoporella minutula by its characteristics, and especially by the shape and arrangement of the laterals, and should be considered a junior synonym.
History of investigations: G�mbel (1872, p. 275) describes a new species (of foraminifera, according to him) Gyroporella minutula: "The shell is as in G. pauciforata, but half the size, also with comparatively wide, less numerous canals, which are usually so close together that the two rows are often difficult to distinguish from one another and it seems as if it is only one row on each ring. The tubes have a diameter of 1 mm and a ring height of 0.5 mm."
(In German: "Geh�use wie bei G. pauciforata, doch um die H�lfte kleiner, mit gleichfalls verh�ltnissm�ssig weiten , wenig zahlreichen Kan�lchen, welche meist so dicht beisammen stehen, dass beide Reihen oft schwierig voneinander zu unterscheiden sind und es das Ansehen hat, als ob nur eine Reihe derselben auf jedes Ringglied treffe. Im Durchmesser haben die R�hrchen 1 mm in der Ringh�he 0.5 mm.")
Pia (1912, p. 45) gives a new description and renames the species as Physoporella minutula: "The shell of this species shows all transitions from an almost smooth surface to a deeply segmented structure that almost approaches the species Physoporella dissita. Compare the transition series on Pia (1912, Pl. VI, figs. 5-8 = this paper, Pl. 12 ![]() , figs. 1 left, 5, 7, 4.). I find it quite clear that we are here dealing only with an advanced undulation. The branches are again in crowded, two-rowed whorls (see especially the individual article, Pl. VI, fig. 10). What differs from the species described so far is, however, their shape. They are not, or at least are only barely visibly, pointed outwards. Rather, their shape is tubular or, if you like, sausage-shaped, with a rounded distal end."
, figs. 1 left, 5, 7, 4.). I find it quite clear that we are here dealing only with an advanced undulation. The branches are again in crowded, two-rowed whorls (see especially the individual article, Pl. VI, fig. 10). What differs from the species described so far is, however, their shape. They are not, or at least are only barely visibly, pointed outwards. Rather, their shape is tubular or, if you like, sausage-shaped, with a rounded distal end."
(In German: "Die Schale dieser Art zeigt alle �berg�nge von einer fast glatten Oberfl�che bis zu einer tief eingreifenden Gliederung, die sich beinahe schon dem Zustand von Physoporella dissita n�hert. Vergl. die �bergangsreihe Taf. VI, Fig. 5 bis 8. Ich glaube, es ist hier vollst�ndig klar, da� wir es nur mit einer gesteigerten Undulation zu tun haben. Die �ste stehen wieder in gedr�ngten, zweizeiligen Wirteln (siehe besonders das einzelne Glied Taf. VI, Fig. 10). Abweichend von den bisher beschriebenen Arten ist jedoch ihre Form. Sie sind n�mlich gegen au�en nicht oder doch kaum merklich zugespitzt. Ihre Gestalt ist vielmehr eine schlauchoder, wenn man will wurstf�rmige, mit abgerundetem distalen Ende.")
Pia (1920, p. 56) gives a description of the new specimens: "The new material of this species which I have received does not give a basis to any special comments. The pores are relatively regularly open to the outside. Whether one should conclude from this that the species could be better included in Oligoporella seems to me uncertain. The whole shape of the pores does not suggest the idea of a hair-like extension in this species either. In one specimen it seems that the annular furrow behind every second whorl is considerably stronger and sharper than the intervening ones, so that may indicate articles with two whorls each. But this observation is not certain. The wall of the calcareous tube in some cases differs in thickness on the opposite sides of the same article. On a clearly measurable cross-section, the thickness of the wall on the left side was just twice as much as on the right side of the longitudinal axis. It is noteworthy that in the thin part of the wall the pores are wide open to the outside. In this specimen, and probably in others, it is probably erosion, which is one more reason why not to give any systematic importance to some open pores."
(In German: "Das neue Material, das ich von dieser Art erhalten habe, gibt zu keinen besonderen Bemerkungen Anla�. Die Poren sind relativ oft gegen au�en offen. Ob man daraus schlie�en soll, da� die Art besser bei Oligoporella unterzubringen w�re, scheint mir keineswegs sicher. Die ganze Form der Poren legt die Vorstellung einer haarf�rmigen Verl�ngerung der Kurztriebe auch bei dieser Art nicht nahe. Bei einem Exemplar scheint es, als wenn die Ringfurche hinter jedem zweiten Wirtel bedeutend st�rker und sch�rfer w�re als die zwischenliegenden, so da� eine Andeutung von Gliedern mit je zwei Wirteln entstehen w�rde. Doch ist die Beobachtung nicht sicher. Die Wandung der Kalkr�hre ist in einigen F�llen auf gegen�berliegenden Seiten desselben Gliedes sehr verschieden stark. An einem deutlich me�baren Schr�gschnitt betrug die Dicke der Wand links gerade doppelt soviel als rechts von der langen Achse. Es ist bemerkenswert, da� in dem d�nnen Skeletteil die Poren gegen au�en weit offen sind. Es wird sich also bei diesem und wohl auch bei anderen St�cken um Abrollung handeln, ein weiterer Grund, um einzelnen offenen Poren keine systematische Bedeutung beizulegen.")
Ellenberger (1958, p. 190) states the presence of transitional morphologies between P. minutula and P. praealpina in the same sample. He concludes that these two species are only one genetic (biological) species. P. praealpina would be the adult form, while in unfavourable living conditions P. minutula would predominate as the juvenile or atrophied form. Botteron (1961, p. 63-64) supports this idea, concluding that P. minutula would represent the juvenile member of the sequence, P. dissita the transitional member, and P. praealpina the adult member. These conclusions cannot be accepted since only adult algae calcify. Calcification begins only at the end of the vegetative phase when the algae has reached the adult stage (De Castro, 1997), because otherwise it could no longer grow, thus only adult specimens are preserved as fossils.
Raciborski (1892, p. 8) describes the new species of Gyroporella balinensis from the Triassic of Chrzanow in Poland: "In Balina below Chrzanow I found layers of dolomitic limestone full of fragments of Gyroporella, similar to G. cylindrica, but only 1-1.2 mm wide. On one very narrow ring there are two rows of canals. I name this, the smallest species known of the Triassic genus Gyroporella: G. balinensis nov. sp." No description (drawing, photograph) of the new species is attached to this description, but after Article 43.2. ICN (Turland et al., 2018), species described before January 1st, 1912 does not need to be accompanied by an illustration, so the species is valid. Pastwa-Leszczyńska and Śliwiński (1960, p. 689) used this already existing species name, but with a new combination and a new taxon rank Oligoporella pilosa forma balinensis, to name a specimen of algae found in the same area: "It is the smallest form we have found so far. The diameter of the preserved shell fragment is 1.2 mm. The branches are grouped in double whorls, clearly separated from each other. The shape of the branches is similar to the trichophorus type. The most common is the alternating arrangement of the branches within one double whorl. The height of the branches does not exceed 0.3 mm. According to the descriptions of Pia (1925, p. 101-102; 1940, p. 4) we identify this form as Oligoporella pilosa. Its small dimensions, already noted by M. Raciborski (1892, p. 532-533), encourage us to distinguish the variety Oligoporella pilosa f. balinensis. We took the name of the variety from M. Raciborski." As already mentioned, I consider this form as a younger synonym of Oligoporella minutula n. comb. To the same species belongs the specimen described as Diplopora cf. silesiaca (Pastwa-Leszczyńska & Śliwiński, 1960, p. 687, Fig. 7 = Pl. III, fig. 13).
(In Polish: "Jest to najmniejsza ze znalezionych przez nas dotychczas form. średnica zachowanego fragmentu wałka wynosi 1,2 mm. Guzki grupują się w podw�jnych ok�łkach, wyraźnie od siebie oddzielonych. Kształt guz-k�w zbliżony jest do typu trichophor. Najczęściej obserwuje się naprzemianległe ułożenie guzk�w w ramach jednego podw�jnego ok�łka. Wysokość guzk�w nie przekracza 0,3 mm. Stosownie do opis�w J. Pia (1925, p. 101-102; 1940, p. 4) identyfi-kujemy tę formę jako Oligoporella pilosa. Drobne jej wymiary, zauważone już przez M. Raciborskiego (1892, str. 532-533), skłaniają nas do wy-r�żnienia odmiany Oligoporella piłosa f. balinensis. Nazwę odmiany przyjęliśmy od M. Raciborskiego.")
Kotański (2013, p. 81) gave a description on the basis of isolated specimens from the Upper Silesia (Poland): "Most frequently algae with divided (undulated or even annulated) cylinder. Segments, if they exist, are sharper that in Ph. dissita and have a very characteristic shape (comp. G�mbel's holotype). This feature is so characteristic, that it is frequently used for species determination even when pores are not visible. The two rows of pores in one whorl are very well marked and the shape of the pores, although still piriform, is generally elongated, having a rather "sausage-like" character. Nevertheless, in some cases it is typically piriform. The contour of the inner tube of cylinder is straight, regular and very markedly separated from the wall. Dimensions are comparable with Ph. praealpina, or smaller."
Description: Species Oligoporella minutula (G�mbel), n. comb., is relatively poorly known and with few high-quality illustrations in the literature. The thallus is cylindrical, and a well-preserved specimen from Pia (1920, Pl. III, fig. 22) shows a slight curvature in the lower part of the thallus. The outer surface of the thallus is less (Pl. 12 ![]() , figs. 5, 7) or more undulated (Pl. 12
, figs. 5, 7) or more undulated (Pl. 12 ![]() , figs. 4, 6; Pia, 1920, Pl. III, fig. 22; Botteron, 1961, Pl. VI, fig. 4 pars; Bystrick�, 1964, Pl. XX, fig. 4). The protruding parts are at the level of the whorl, and due to the inclination of the whorl, the undulation is asymmetric. The inner wall of the thallus is, in the case of preserved specimens, smooth and flat, and the calcification reached near to the stem cell (Pl. 12
, figs. 4, 6; Pia, 1920, Pl. III, fig. 22; Botteron, 1961, Pl. VI, fig. 4 pars; Bystrick�, 1964, Pl. XX, fig. 4). The protruding parts are at the level of the whorl, and due to the inclination of the whorl, the undulation is asymmetric. The inner wall of the thallus is, in the case of preserved specimens, smooth and flat, and the calcification reached near to the stem cell (Pl. 12 ![]() , figs. 1 left, 4, 6-7; Pl. 13
, figs. 1 left, 4, 6-7; Pl. 13 ![]() , fig. 2; Botteron, 1961, Pl. VI, fig. 4 pars; Bystrick�, 1964, Pl. XX, fig. 4) or close to it (Pia, 1920, Pl. III, fig. 22).
, fig. 2; Botteron, 1961, Pl. VI, fig. 4 pars; Bystrick�, 1964, Pl. XX, fig. 4) or close to it (Pia, 1920, Pl. III, fig. 22).
The laterals, according to the obvious similarity with those of other species of Physoporella and Oligoporella, can be classified as of piriform type, but they have less pronounced proximal widening and distal narrowing (Fig. 14 ![]() ), so, as Pia (1912) stated, they have a more sausage-like appearance (Pl. 12
), so, as Pia (1912) stated, they have a more sausage-like appearance (Pl. 12 ![]() , figs. 3, 5, 7-8; Pl. 13
, figs. 3, 5, 7-8; Pl. 13 ![]() , figs. 2, 4; Bystrick�, 1964, Pl. XX, fig. 4). However, they are not always completely distally rounded, but sometimes show a thinning at their end (Pl. 12
, figs. 2, 4; Bystrick�, 1964, Pl. XX, fig. 4). However, they are not always completely distally rounded, but sometimes show a thinning at their end (Pl. 12 ![]() , figs. 1 left, 4, 6; Pl. 13
, figs. 1 left, 4, 6; Pl. 13 ![]() , figs. 2, 4; Pia, 1920, Pl. III, fig. 22), so there is some variability, as with other studied species. The existence of trichoblasts in the continuation of the laterals is indicated by "lamellae" (Pl. 12
, figs. 2, 4; Pia, 1920, Pl. III, fig. 22), so there is some variability, as with other studied species. The existence of trichoblasts in the continuation of the laterals is indicated by "lamellae" (Pl. 12 ![]() , fig. 1 right) and "spikes" (Pl. 12
, fig. 1 right) and "spikes" (Pl. 12 ![]() , fig. 4 right) on the outer surface of the thallus. The laterals are arranged in spaced double whorls (Pl. 12
, fig. 4 right) on the outer surface of the thallus. The laterals are arranged in spaced double whorls (Pl. 12 ![]() , figs. 1 right, 3-9; Pl. 13
, figs. 1 right, 3-9; Pl. 13 ![]() , fig. 2) and always laid obliquely on the longitudinal axis of the thallus. Within the whorl they are probably arranged alternately (Pl. 12
, fig. 2) and always laid obliquely on the longitudinal axis of the thallus. Within the whorl they are probably arranged alternately (Pl. 12 ![]() , figs. 3, 9; Botteron, 1961, Pl. VI, fig. 4 pars).
, figs. 3, 9; Botteron, 1961, Pl. VI, fig. 4 pars).
Emended diagnosis: Cylindrical thallus with more or less undulated outer and smooth inner surface. The laterals are of the piriform type, but with less pronounced proximal widening and distal narrowing, often of sausage-like appearance. Arranged in spaced double whorls and laid obliquely on the longitudinal axis of the thallus. Within the whorl they are arranged mostly alternately.
Similarities and differences: Oligoporella minutula is most similar to the species Oligoporella praealpina, from which it differs in its smaller dimensions, undulated outer surface of the thallus, oblique sausage-like laterals and more spaced whorls.
|
|
Figure 14: Oligoporella minutula (G�mbel, 1872), n. comb. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The upper two whorls (I) show trichoblasts in the extension of laterals. The longitudinal section (A) shows variants with calcification close to the stem cell (II) and away from the stem cell (I). Due to the alternating arrangement of laterals in the whorl, some laterals are not or are only partially cut in cross-section. The tangential section (B) shows sections of the proximal (upper whorl) and distal parts of the laterals (lower whorl). The undulation of the outer surface of the calcareous thallus is visible. |
5.2.6. Oligoporella praealpina (Pia, 1920), n. comb.
(= Physoporella praealpina Pia, 1920)
Pl. 13 ![]() , figs. 3, 5-12; Pl. 14
, figs. 3, 5-12; Pl. 14 ![]() , figs. 1-9; Pl. 15
, figs. 1-9; Pl. 15 ![]() , figs. 1-2, 4-6; Pl. 16
, figs. 1-2, 4-6; Pl. 16 ![]() , fig. 13; Pl. 17
, fig. 13; Pl. 17 ![]() , fig. 4; Pl. 22
, fig. 4; Pl. 22 ![]() , figs. 1-4, 5 pars
, figs. 1-4, 5 pars
Origin of the name: Probably after the type locality southeast of Bern that is located in the area of Praealps.
History of investigation: Pia (1920, p. 53-55) describes the new species Physoporella praealpina: "This splendid species has very large pores that are densely packed in pronounced two-row whorls. They are only very slightly inclined upwards. The whorled branches touch each other next to the stem cell, so that no calcareous mass could be deposited between them. This is why - as in other species with a similar arrangement of pores - the skeleton in the poreless zones between the whorls extends further inward than between the pores of one whorl. The articulation of the skeleton is interesting because it provides one of the best examples of what I call Fissuration. Firstly, it is noticeable that shells very often split regularly along transverse planes. There are also sections (Pia, 1920, Pl. 3. figs. 3, 6, 7, 9 = this work, Pl. 14 ![]() , figs. 5, 6, 4, 9) where there is a narrow crack, only partially closed by calcareous mass, between every two whorls. The resulted articles comprise one or two whorls. I have not observed any other numbers. Incidentally, both of these cases are not quite clearly separated, so I consider it possible that in one specimen with one-whorled articles the fissures are alternately more or less pronounced. In typical articles with two whorls, however, this form of metamerism is expressed in the structure of the whole plant, because both whorls that belong together are very close to each other, while those from the adjacent articles are separated through an approximately pore-wide gap (Pia, 1920, Pl. 3, fig. 6 = this paper, Pl. 16
, figs. 5, 6, 4, 9) where there is a narrow crack, only partially closed by calcareous mass, between every two whorls. The resulted articles comprise one or two whorls. I have not observed any other numbers. Incidentally, both of these cases are not quite clearly separated, so I consider it possible that in one specimen with one-whorled articles the fissures are alternately more or less pronounced. In typical articles with two whorls, however, this form of metamerism is expressed in the structure of the whole plant, because both whorls that belong together are very close to each other, while those from the adjacent articles are separated through an approximately pore-wide gap (Pia, 1920, Pl. 3, fig. 6 = this paper, Pl. 16 ![]() , fig. 6). Although these articles are poorly developed and the grooves that separate them are poorly noticeable, so far I have not had thin-sections in which I did not notice the articulation (of course, the cut has to be quite oblique to show the structure at all). However, it must be taken as a fact that there are quite a few (about one third of all examined) individuals of Physoporella praealpina who are completely inarticulate (e.g., Pia, 1920, Pl. 3, figs. 5, 2 = this paper, Pl. 14
, fig. 6). Although these articles are poorly developed and the grooves that separate them are poorly noticeable, so far I have not had thin-sections in which I did not notice the articulation (of course, the cut has to be quite oblique to show the structure at all). However, it must be taken as a fact that there are quite a few (about one third of all examined) individuals of Physoporella praealpina who are completely inarticulate (e.g., Pia, 1920, Pl. 3, figs. 5, 2 = this paper, Pl. 14 ![]() , fig. 1). In the same way, poorly closed cracks, such as those between the articles, separate the calcareous parts belonging to the individual whorled branches in several specimens in the most superficial part of the calcareous shell. It seems to me that in this case it is not a question of zones of contamination, but of really open cracks (Pl. 3
, fig. 1). In the same way, poorly closed cracks, such as those between the articles, separate the calcareous parts belonging to the individual whorled branches in several specimens in the most superficial part of the calcareous shell. It seems to me that in this case it is not a question of zones of contamination, but of really open cracks (Pl. 3 ![]() , fig. 1)."
, fig. 1)."
(In German: "Diese stattliche Art weist sehr grobe Poren auf, die dicht gedr�ngt in ausgesprochen zweizeiligen Wirteln stehen. Sie sind nur ganz schwach gegen oben geneigt. Die Wirtel�ste ber�hrten einander n�chst der Stammzelle, so da� hier zwischen ihnen keine Kalkmasse abgelagert werden konnte. Daher kommt es, da� - wie auch bei anderen Arten mit �hnlicher Anordnung der Poren - das Skelett in den porenfreien Zonen zwischen den Wirteln weiter gegen innen reicht als zwischen den Poren eines Wirtels. Interessant ist die Gliederung des Skelettes, weil sie eines der besten Beispiele f�r die Erscheinung liefert, die ich Fissuration nenne. Zun�chst f�llt schon auf, da� die Schalen sehr h�ufig in regelm��iger Weise entlang von Querebenen zerfallen sind. Bald findet man auch Schnitte (Taf. 3. Fig. 3, G. 7, 9). bei denen zwischen je zwei Wirteln ein schmaler, nur teilweise durch Kalkmasse geschlossener Spalt verl�uft. Die so gebildeten Glieder umfassen ein oder zwei Wirtel. Andere Zahlen habe ich nicht beobachtet. �brigens sind diese beiden F�lle vielleicht nicht ganz scharf getrennt, denn ich glaube, bemerkt zu haben, da� bei einem Exemplar mit einwirteligen Gliedern die Fissuren abwechselnd einmal st�rker und einmal weniger deutlich sind. Bei typischen Gliedern mit zwei Wirteln dr�ckt sich diese Form dar Metamerie allerdings im Bau der ganzen Pflanze aus, denn die beiden zusammengeh�rigen Wirtel sind einander dann stark gen�hert, w�hrend sie von denen der Nachbarglieder durch einen etwa porenbreiten Zwischenraum getrennt sind (vgl. Taf. 3, Fig. 6). Wie gering der Zusammenhang der Glieder trotz ihrer wenig auffallenden Scheidung ist, geht daraus hervor, da� ich keim, gegliedertes Exemplar beobachtet habe, das nicht innerhalb des Bereiches des Schnittes wenigstens einmal zerbrochen w�re. (Naturgem�� mu� der Schnitt ja ziemlich schr�g liegen, um die Gliederung �berhaupt zu zeigen). Es mu� jedoch als Tatsache angesehen werden, da� es ziemlich viele (etwa ein Drittel aller untersuchten) Individuen von Physoporella praealpina gibt, die vollst�ndig ungegliedert sind (z. B. Tab. 3, Fig. 2, 5). Ganz �hnlich schlecht geschlossene Fugen, wie sie zwischen den Gliedern liegen, trennen bei mehreren Exemplaren im oberfl�chlichsten Teil der Schale die zu den einzelnen Wirtel�sten geh�rigen Kalkpartien voneinander. Es scheint mir aus dem mikroskopischen Bild klar hervorzugehen, da� es sich in diesem Falle nicht um Zonen von Verunreinigung, sondern um wirklich offene Spalten handelt (vgl. Taf. 3, Fig. 1).")
Pia (1935a, p. 226-229) describes as Physoporella aff. praealpina specimens from western Bosnia and Herzegovina, but I believe that they should be included in a separate variety Physoporella praealpina var. bosniensis n. var., which will be explained below. By isolating a new variety, type variety become Physoporella praealpina var. praealpina n. var. (Art. 26.3. ICN, Turland et al., 2018).
Pastwa-Leszczyńska and Śliwiński (1960, p. 686-687, 1960, Fig. 6 = Pl. IV, fig. 15 = this work, Pl. 16 ![]() , fig. 13) establish a new form of Diplopora annulata forma physoporelloidea. This taxon Kotański in Kotański and Čatalov (1973, p. 203) considered a younger synonym of the species Physoporella praealpina. I agree with him as the illustrated specimens, according to the shape and euspondyl arrangement of laterals, belong to the species P. praealpina.
, fig. 13) establish a new form of Diplopora annulata forma physoporelloidea. This taxon Kotański in Kotański and Čatalov (1973, p. 203) considered a younger synonym of the species Physoporella praealpina. I agree with him as the illustrated specimens, according to the shape and euspondyl arrangement of laterals, belong to the species P. praealpina.
Bystrick� (1964, p. 126-127) describes specimens of this species under the name Physoporella cf. praealpina, not because he doubts the determination of the species, but because he did not notice a series of whorls separated by larger parts without pores that is stated in the diagnosis (1920). Here is his description: "The most characteristic feature of these specimens, which can be compared with the species Physoporella praealpina, is the calcareous shell with a flat or slightly wavy outer surface and the pores arranged in two-row whorls. Articulation of the thallus by fissures is very irregular, often missing; the height of the segments limited by them is therefore very variable. The inner surface is flat or grooved in the same way as in Ph. pauciforata var. pauciforata. The pores are pear-shaped, their distal ends are pointy and enclosed in a calcareous sheet. They form distinctive alternating two-row whorls, lying perpendicular to the thallus' longitudinal axis. The basal thickened portions of the pores in the whorl are close together; therefore, the shell above the whorl axis penetrates inside much less than between the whorls. The distances between the whorls are uniform but variable. In the specimens on Pl. XIX, figs. 1, 5, the whorls are very close, separated from each other only by fine calcareous lamellae; in others (e.g., Pl. XIX, figs. 2, 3, 6) the distances between the whorls are significantly larger. Only the individual on Pl. XIX, fig. 8 (longitudinal section) showed signs of grouping the whorls into a series, separated by a higher part of calcareous shell."
Kotański and Čatalov (1973, p. 191) named a new variety Physoporella praealpina var. sulcata: "Two specimens (Kotański & Čatalov, 1973, Pl. VII, figs. 18-19) form a new variety of Ph. praealpina. The presence of oblique fissures (idem, Pl. VII, fig. 18) and obliquely oriented pairs of pores (idem, Pl. VII, fig. 19) are features comparable only with Ph. pauciforata var. sulcata, but the existence of two rows of pores in each whorl suggests for their assignment to Ph. praealpina. Physoporella cf. praealpina Pia with branches obliquely oriented to the main stem and without calcification between their proximal portions was described by Herak (1965). These features are outside the range of Ph. praealpina var. praealpina. For the new variety is proposed the name Ph. praealpina var. sulcata - per analogiam to Ph. pauciforata var. sulcata Bystrick�." Considering the taxon is nomen nudum due to the lack of a defined holotype, but also according to poorly preserved specimens, the inclination of the pores, as well as other characteristics, does not differ from other specimens described as P. praealpina, this variety can be rejected.
Ott (1972b, p. 4-6) gives his description of this species: "The branch pores have a pirifer shape typical of the genus Physoporella. In the section they typically appear closed within the calcareous shell. From the cross-section in Ott (1972b, Fig. 1/16), however, it can be seen that there were also hair-like channels outwards. As a rule, in the two-row whorls the lower branch is also inclined diagonally upwards and does not diverge downwards, as in Ph. varicans. The calcareous skeleton shows two peculiarities, which were also mentioned in the first description by Pia. The first phenomenon was called "fissuration" by Pia. It consists of weak zones in the calcareous tube that are incised transversely to the longitudinal axis, along which the skeleton split into short tubular segments. Tubular segments of Ph. praealpina typically contain two whorls, like the ones illustrated by Ott (1972b, Fig. 1/17-18). However, there are also more or less unbroken specimens (idem, Fig. 1/1-4). If the fissures only divide articles from at the level of one whorl, the sections are reminiscent of Ph. dissita, in which the furrows are indented so far towards the stem cell after each whorl that Pia considers as true annulation. Another peculiarity is lamellar cavities parallel to the longitudinal axis in the outer layers of the skeleton, which in some places peel off the outer wall layers. In longitudinal sections you can see spurs parallel to the wall splitting off (idem, Fig. 1/1, 7, 17, 20), in cross sections you can see covered hollows or glacier-table-like plates, which are still connected to the skeletal tube with a thin base (especially beautiful in Ott (1972b, Fig. 1/5, 11). A functional meaning of these phenomena is not obvious."
Kotański (2013, p. 74) gave a diagnosis on the basis of isolated specimens from the Upper Silesia (Poland): "This species is characterized by a cylindrical, generally undivided thallus and by alternating vertical branches (pores, tubercles), forming two rows belonging to one whorl. Whorls are closely spaced and tubercles are very regularly arranged. If the calcareous (or dolomitic) wall is preserved, initial undulation or fissuration can be observed. The shape of the pores (tubercles) is piriform. Generally, they are perpendicular to the main axis, but in some cases they are slightly oblique to the wall."
Oligoporella praealpina var. praealpina n. var.
Pl. 13 ![]() , figs. 3, 5-12; Pl. 14
, figs. 3, 5-12; Pl. 14 ![]() , figs. 1-9; Pl. 16
, figs. 1-9; Pl. 16 ![]() , fig. 13; Pl. 17
, fig. 13; Pl. 17 ![]() , fig. 4; Pl. 22
, fig. 4; Pl. 22 ![]() , figs. 1-4, 5 pars
, figs. 1-4, 5 pars
1920 - Physoporella praealpina n. sp. - Pia, p. 53-55, Pl. III, figs. 1-9.
1960 - Diplopora annulata forma physoporelloidea n. forma - Pastwa-Leszczyńska & Śliwiński, p. 686-687, Fig. 6; Pl. IV, fig. 15 (= Fig. 6).
1964 - Physoporella cf. praealpina - Bystrick�, p. 126-127, Pl. XIX, figs. ?1, 2.-4, ?5, 6-9; Pl. XX, figs. 1-2, ?3.
1972d - Physoporella praealpina - Ott, p. 4-6, Fig. 1, no. 1-21.
1973 (nom. nud.) - Physoporella praealpina var. sulcata n. var.- Kotański & Čatalov, p. 191, Pl. VII, figs. 18-19, no type.
1975 - Physoporella praealpina - Pantić-Prodanović, p. 12, Pl. XXVI, fig. 1 pars; Pl. XXVII, fig. 1.
1975 - Oligoporella pilosa var. intusannulata - Pantić-Prodanović, p. 12, Pl. XXVI, fig. 1 pars.
1975 - Physoporelles - Pantić-Prodanović, Pl. XXVIII, fig. 1 pars.
2013 - Physoporella praealpina - Kotański, p. 73-78, Pl. XIII, figs. 1-17; Pl. XIV, figs. 1-23; Pl. XV, figs. 1-22; Pl. XVIII, figs. 1-3, 5, 8; Pl. XIX, figs. 1-5; Pl. XX, figs. 3.b, 4, 6, 9, 13-14; Pl. XXXII, figs. 1.b, 2.a, 3.b, 4b; Pl. XXXIII, figs. 1.a, 2.a, 3 pars, 4.b; Pl. XXXIV, figs. 1.b, 2 pars, 3.1, 4.a; Pl. XXXV, figs. 1.a, 2-a; Pl. XXXVI, figs. 1.b, 3.b, 4.a; Pl. XXXVII, figs. 1.b, 2.d, 3.b, 4.a; Pl. XXXVIII, figs. 1.a, 2.a, 3.a, 4.a.
2013 - Physoporella cf. praealpina - Rychliński et al., Pl. 2, fig. 3.
2021 - Oligoporella praealpina - Gawlick et al., Pl. 15, figs. m, q.
Studied material: The analysed and illustrated material, described by Pia (1920), comes from a secondary sample of Anisian limestone found in the vicinity of Horboden in the Diemtigtal, west of Spiez, southeast Bern in Switzerland: thin-sections CLVII1, CLVII2 and CLVII3. I could not find the specimen from Pia (1920, Pl. III, fig. 5). Pia (1920) did not choose a holotype, so as a lectotype I choose a specimen from Pl. 14 ![]() , fig. 1, as it best reveals the characteristics of this species. Other specimens from the same sample are isotypes. The material from Croatia includes specimens from the Anisian deposits near Udbina, Lika (Pl. 22
, fig. 1, as it best reveals the characteristics of this species. Other specimens from the same sample are isotypes. The material from Croatia includes specimens from the Anisian deposits near Udbina, Lika (Pl. 22 ![]() , figs. 1-2) and those from Hum-�agudovac on the northern side of Medvednica Mt (Pl. 22
, figs. 1-2) and those from Hum-�agudovac on the northern side of Medvednica Mt (Pl. 22 ![]() , figs. 3-5).
, figs. 3-5).
Description: This species is abundantly illustrated in the literature and is not difficult to describe. The thallus is cylindrical and straight, sometimes slightly curved (Pl. 14 ![]() , fig. 1; Pl. 22
, fig. 1; Pl. 22 ![]() , figs. 1-3, 5 pars; Kotański & Čatalov, 1973, Pl. VI, fig. 1; Pantić-Prodanović, 1975, Pls. XXVI-XXVIII). Long specimens are quite common. The outer surface of the thallus is usually uneven, i.e., rough (Pl. 13
, figs. 1-3, 5 pars; Kotański & Čatalov, 1973, Pl. VI, fig. 1; Pantić-Prodanović, 1975, Pls. XXVI-XXVIII). Long specimens are quite common. The outer surface of the thallus is usually uneven, i.e., rough (Pl. 13 ![]() , figs. 8, 12; Pl. 14
, figs. 8, 12; Pl. 14 ![]() , figs. 3, 5-6; Pl. 22; Bystrick�, 1964, Pl. XX, fig. 1). This is probably due to more irregular calcification in this species. This is also indicated by small, mostly triangular, cavities between the whorls in the lectotype (Pl. 14
, figs. 3, 5-6; Pl. 22; Bystrick�, 1964, Pl. XX, fig. 1). This is probably due to more irregular calcification in this species. This is also indicated by small, mostly triangular, cavities between the whorls in the lectotype (Pl. 14 ![]() , fig. 1). In other specimens the outer surface is relatively smooth (Pl. 14
, fig. 1). In other specimens the outer surface is relatively smooth (Pl. 14 ![]() , fig. 1), often slightly undulated with protruding parts at the level of the whorls (Pl. 13
, fig. 1), often slightly undulated with protruding parts at the level of the whorls (Pl. 13 ![]() , figs. 3, 10-11; Pl. 14
, figs. 3, 10-11; Pl. 14 ![]() , fig. 2; Bystrick�, 1964, Pl. XIX, figs. 7, 9; Pantić-Prodanović, 1975, Pl. XXVII left). Since calcification begins around the laterals, in order to protect them, parts of the thallus between the laterals are mechanically weaker, especially due to sporadic irregular calcification, so they are more susceptible to cracking (Pl. 13
, fig. 2; Bystrick�, 1964, Pl. XIX, figs. 7, 9; Pantić-Prodanović, 1975, Pl. XXVII left). Since calcification begins around the laterals, in order to protect them, parts of the thallus between the laterals are mechanically weaker, especially due to sporadic irregular calcification, so they are more susceptible to cracking (Pl. 13 ![]() , figs. 10-11; Pl. 14
, figs. 10-11; Pl. 14 ![]() , figs. 3-6, 8-9; Pl. 22
, figs. 3-6, 8-9; Pl. 22 ![]() , figs. 3-4). Therefore, it makes no sense to count the cracks and the distance between them as significant, as in some older works, because it is not in any way related to the living organism, but the result of physical processes. An identical phenomenon is found in P. pauciforata var. pauciforata (see chapter 5.1.1.1.). Particularly interesting is the appearance of "scales" and "horns" on the outer side of the thallus in well-preserved specimens (Pl. 13
, figs. 3-4). Therefore, it makes no sense to count the cracks and the distance between them as significant, as in some older works, because it is not in any way related to the living organism, but the result of physical processes. An identical phenomenon is found in P. pauciforata var. pauciforata (see chapter 5.1.1.1.). Particularly interesting is the appearance of "scales" and "horns" on the outer side of the thallus in well-preserved specimens (Pl. 13 ![]() , figs. 8, 10; Pl. 14
, figs. 8, 10; Pl. 14 ![]() , figs. 7-9; Pl. 22
, figs. 7-9; Pl. 22 ![]() , figs. 3- 4; Ott, 1972d, Fig. 1), which are identical to those described in Physoporella pauciforata var. pauciforata and have a similar interpretation. These are calcified parts of the trichoblasts, which were bent upwards (Fig. 15
, figs. 3- 4; Ott, 1972d, Fig. 1), which are identical to those described in Physoporella pauciforata var. pauciforata and have a similar interpretation. These are calcified parts of the trichoblasts, which were bent upwards (Fig. 15 ![]() ). As the trichoblasts were numerous, their calcareous envelopes merged into "scales", which in longitudinal sections look like horns, spikes or scales (see chapter 5.1.1.1). The same phenomenon was observed by Ott (1972d, p. 6), but he could not explain it. Since the whorls are quite dense, calcification could not reach the stem cell, and the parts of the calcareous sheath between the whorls in longitudinal and oblique sections look like thin bars or wedges (Pl. 13
). As the trichoblasts were numerous, their calcareous envelopes merged into "scales", which in longitudinal sections look like horns, spikes or scales (see chapter 5.1.1.1). The same phenomenon was observed by Ott (1972d, p. 6), but he could not explain it. Since the whorls are quite dense, calcification could not reach the stem cell, and the parts of the calcareous sheath between the whorls in longitudinal and oblique sections look like thin bars or wedges (Pl. 13 ![]() , figs. 8, 10; Pl. 14
, figs. 8, 10; Pl. 14 ![]() , fig. 1), and in tangential sections as bars between the whorls (Pl. 14
, fig. 1), and in tangential sections as bars between the whorls (Pl. 14 ![]() , fig. 6; Pl. 22
, fig. 6; Pl. 22 ![]() , fig. 2). Rare specimens on which calcification reached to the stem cell preserved in the fossils (Pl. 22
, fig. 2). Rare specimens on which calcification reached to the stem cell preserved in the fossils (Pl. 22 ![]() , fig. 1 right and partially 3 right and 5 right; Ott, 1972d, Fig. 1/10) show a smooth inner thallus wall.
, fig. 1 right and partially 3 right and 5 right; Ott, 1972d, Fig. 1/10) show a smooth inner thallus wall.
The shape of the laterals is certainly piriform, similar to those of the already described species of Physoporella and Oligoporella, but less pointed in the distal part and with a rounded end, which can be seen in almost all the illustrated specimens. The proximal part of the laterals, due to incomplete calcification, is generally not visible, but on the above-mentioned preserved specimens it can be seen that it was slightly enlarged. At their apex the laterals bear trichoblasts, as mentioned earlier. It is also clear from almost all the illustrated specimens that the laterals are arranged in dense double whorls. The laterals lie perpendicular to the stem cell
(Pl. 13 ![]() ,
figs. 7-9; Pl. 14
,
figs. 7-9; Pl. 14 ![]() ,
fig. 1; Pl. 15
,
fig. 1; Pl. 15 ![]() ,
figs. 1-2) or quite slightly sloping upwards (Pl. 13
,
figs. 1-2) or quite slightly sloping upwards (Pl. 13 ![]() , figs. 3, 10-12; Pl. 14
, figs. 3, 10-12; Pl. 14 ![]() , figs. 1-3; Pl. 22
, figs. 1-3; Pl. 22 ![]() , figs. 3, 5). Inside the whorls the laterals stood very close to each other. Due to their dense arrangement, the laterals stood mostly alternately both inside the whorl and in the adjacent whorls (Pl. 13
, figs. 3, 5). Inside the whorls the laterals stood very close to each other. Due to their dense arrangement, the laterals stood mostly alternately both inside the whorl and in the adjacent whorls (Pl. 13 ![]() , figs. 9, 12; Pl. 14
, figs. 9, 12; Pl. 14 ![]() , figs. 1, 3-6, 8-9; Pl. 22
, figs. 1, 3-6, 8-9; Pl. 22 ![]() , figs. 2, 4).
, figs. 2, 4).
Pia (1920, Fig. 11) on the basis of a sample from Pl. 14 ![]() , fig. 6, gives a reconstruction of the algae in which each of the two adjacent double whorls are close together and are separated from the other two double whorls, although in the text he mentions that this is not a regular occurrence (see above). Therefore, specimens in which this is not seen are named by Pia (1935a) Physoporella aff. praealpina, and for the same reason this is also done later by Bystrick� (1964), Patrulius (1970), Bucur (1997) and Bucur et al. (1994). The described characteristic can be seen only on the mentioned specimen, but it fits into the range of variations of each living and fossil species, so the diagnosis of the genus cannot be based on it alone. Let us recall the numerous phenomena of heteromorphosis in recent Dasycladales described by Valet (1968).
, fig. 6, gives a reconstruction of the algae in which each of the two adjacent double whorls are close together and are separated from the other two double whorls, although in the text he mentions that this is not a regular occurrence (see above). Therefore, specimens in which this is not seen are named by Pia (1935a) Physoporella aff. praealpina, and for the same reason this is also done later by Bystrick� (1964), Patrulius (1970), Bucur (1997) and Bucur et al. (1994). The described characteristic can be seen only on the mentioned specimen, but it fits into the range of variations of each living and fossil species, so the diagnosis of the genus cannot be based on it alone. Let us recall the numerous phenomena of heteromorphosis in recent Dasycladales described by Valet (1968).
Emended diagnosis of the species Oligoporella praealpina (Pia), n. comb.: Cylindrical thallus with a slightly uneven, slightly undulated or smooth outer surface. Calcification first envelops the laterals, so sometimes the parts between the whorls are less calcified and more susceptible to mechanical disintegration. Calcification generally does not reach the stem cell. The laterals are piriform, distally somewhat less tapered than in other species and with rounded ends. At the apex laterals bear trichoblasts bent upwards, and occasionally mutually calcified in the form of "scales" on the thallus surface. The laterals are laid vertically or slightly obliquely to the longitudinal axis and arranged in relatively dense double whorls. Inside the whorl and in the adjacent whorls the laterals are arranged mostly alternately.
Similarities and differences: Oligoporella praealpina is somewhat similar to the species Oligoporella pilosa, from which it differs in generally thinner laterals, although relatively thicker and more rounded in the distal part, and a relatively smaller distance between the whorls. It differs from the species Oligoporella dissita by its distally rounded laterals, relatively smaller distance between the whorls and the absence of annulation. It differs from the similar species Oligoporella minutula in its larger dimensions, weaker undulation of the thallus, less oblique laterals and more densely distributed whorls. It clearly differs from other species of this genus by a smaller divergence of laterals.
Oligoporella praealpina var. praealpina differs from the other variety O. praealpina var. bosniensis mainly by being somewhat thicker and the more regularly arranged laterals.
|
|
Figure 15: Oligoporella praealpina var. praealpina n. var. Idealized longitudinal (A), transverse (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. The longitudinal section (A) shows variants with calcification close to the stem cell (I) and adjacent to the stem cell (II). Trichoblasts are shown on some laterals. The tangential section (C) shows the sections of the proximal (upper whorl), central (middle whorl) and distal parts of the laterals (lower whorl). |
Oligoporella praealpina var. bosniensis n. var.
Pl. 15 ![]() , figs. 1-2, 4-6
, figs. 1-2, 4-6
1935a - Physoporella aff. praealpina - Pia, p. 226-229, Figs. 37-39,
Origin of the name: After the type locality of this variety in Bosnia.
Studied material: The material described by Pia (1935a) originates from Bosnia and Herzegovina from the Ograda site, northwest side of the peak (site "u") on the road from Are�in brijeg - Grab in Bosnia and Herzegovina (probably a part of the road between Bosanski Grahovo and Knin along which Triassic deposits crop out): Thin-sections 583a (Pl. 15 ![]() , fig. 1 = Pia, 1935a, Fig. 37; Pl. 15
, fig. 1 = Pia, 1935a, Fig. 37; Pl. 15 ![]() , figs. 5-6) and 583b (Pl. 15
, figs. 5-6) and 583b (Pl. 15 ![]() , figs. 2, 4 = Pia, 1935a, Figs. 38-39).
, figs. 2, 4 = Pia, 1935a, Figs. 38-39).
As holotype I select a specimen from Pl. 15 ![]() , fig. 2, while the other specimens become isotypes.
, fig. 2, while the other specimens become isotypes.
History of investigations: Pia (1935a, p. 226-229) describes Physoporella aff. praealpina (see previous chapter): "Some good sections from western Bosnia (site" u ") belong to the species Physoporella praealpina with distinctly two-row whorls and by the presence of cracks (fissures) in the shell, between which there are 1-2 whorls. They also match it in dimensions. However, among the few usable specimens, I have not found any in which larger and smaller distances between the whorls regularly alternate. The shape of the pores is also slightly different from that of the western Alpine specimens. They taper markedly but gradually, as can be seen from the decreasing size of the pores towards the edge in oblique sections of the shell (Pia, 1935a, Fig. 38; = this paper, Pl. 15 ![]() , fig. 2). In this respect they are closer to those of Physoporella pauciforata (see Pia, 1935a, Figs. 31, 35, 42). It appears to be a transitional form, the position of which can be accurately assessed only when physoporellas from other sites are described."
, fig. 2). In this respect they are closer to those of Physoporella pauciforata (see Pia, 1935a, Figs. 31, 35, 42). It appears to be a transitional form, the position of which can be accurately assessed only when physoporellas from other sites are described."
(In German: "Einige gute Schnitte aus Westbosnien (Fundort u) schliessen sich der Physoporella praealpina durch die ausgesprochen zwei-zeiligen Wirtel und durch das Vorhandensein deutlicher Fissuren in der Schale an, zwischen denen 1-2 Wirtel liegen. Auch in den Abmessungen passen sie zu ihr. Ich habe jedoch unter meinen wenigen brauchbaren St�cken keine gefunden, bei denen gr�ssere und kleinere Abst�nde zwischen den Wirteln regelm�ssig abwechseln. Auch die Gestalt der Poren ist etwas anders, als bei den westalpinen Exemplaren. Sie sind gegen aussen stark, aber allm�hlich verj�ngt, wie aus der abnehmenden Gr�sse der Porenquerschnitte gegen das Ende von Schr�gschnitten der ganzen Schale zu hervorgeht (Textfig. 38). Sie n�hern sich in dieser Beziehung also mehr denen von Physoporella pauciforata (vergl. Textfig. 31, 35, 42). Es scheint sich demnach um eine Uebergangsform zu handeln, deren Stellung erst genau wird beurteilt werden k�nnen, bis die Physoporellen anderer Fundorte beschrieben sind.")
Description: Specimens from Bosnia, although very similar to other specimens of Oligoporella praealpina, show some differences, as stated by the Pia (1935a), so I decided to separate them into a new variety. The thallus is cylindrical. Calcification is also more irregular in this variety than in other species of the genus Oligoporella. The inner side of the thallus, which appears smooth and flat, shows a slight unevenness, while the outer side is very rough and uneven (Pl. 15 ![]() , figs. 2, 5-6). Similar to the type variety, calcification begins around the laterals, so the parts between the laterals are mechanically weaker and often brake (Pl. 15
, figs. 2, 5-6). Similar to the type variety, calcification begins around the laterals, so the parts between the laterals are mechanically weaker and often brake (Pl. 15 ![]() , figs. 2, 4, 6).
, figs. 2, 4, 6).
The laterals are of the piriform type, but somewhat thinner and distally more taper than the type variety. Their arrangement within the whorl also seems to be more "untidy". Within the whorls, the laterals are generally arranged alternately, but they can also be one above the other (Pl. 15 ![]() , figs. 2, 4-6). The laterals are laid perpendicular to the longitudinal axis of the thallus (Pl. 15
, figs. 2, 4-6). The laterals are laid perpendicular to the longitudinal axis of the thallus (Pl. 15 ![]() , figs. 2, 5-6). Although this is not seen in the illustrated specimens, they surely had trichoblasts on the top, as did other taxa studied (Fig. 16
, figs. 2, 5-6). Although this is not seen in the illustrated specimens, they surely had trichoblasts on the top, as did other taxa studied (Fig. 16 ![]() ).
).
Diagnosis: Cylindrical thallus with relatively flat inner and uneven outer surface. The parts between the whorls are somewhat less calcified and were more susceptible to mechanical breaking. Calcification generally does not reach the stem cell. The laterals are of the piriform type, but somewhat thinner and taper distally more than in the type variety. They are laid perpendicular to the longitudinal axis of the thallus and arranged in relatively dense double whorls. Their arrangement within the whorl is somewhat untidy. They are usually in an alternating arrangement, and may be one above the other.
|
|
Figure 16: Oligoporella praealpina var. bosniensis n.var. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts are shown on some laterals. Due to the alternating arrangement of laterals in the whorls, some laterals are not or are only partially cut by cross-section. |
5.2.7. Oligoporella dissita (G�mbel, 1872), n. comb.
(= Physoporella dissita (G�mbel, 1872), Pia, 1912; = Gyroporella dissita G�mbel, 1872)
Pl. 11 ![]() , figs. 1-9; Pl. 13
, figs. 1-9; Pl. 13 ![]() , fig. 1; Pl. 21
, fig. 1; Pl. 21 ![]() , figs. 3-4, 8; ? Pl. 10
, figs. 3-4, 8; ? Pl. 10 ![]() , figs. 3, 5, 7
, figs. 3, 5, 7
1872 - Gyroporella dissita n. sp. - G�mbel, p. 271, Pl. D.III, fig. 1.
1912 - Physoporella dissita n. comb. - Pia, p. 45, Pl. VI, figs. 1-4.
1957 - Physoporella dissita - Bystrick�, p. 234, Pl. VI, figs. 6-7.
1964 - Physoporella dissita - Bystrick�, p. 127-129, Pl. XX, fig. 6, Pl. XXI, fig. 7.
1966 - Physoporella dissita - Bystrick�, p. 251, Pl. VI, fig. 5; Pl. VII, figs. 1, 2.b.
1989 - Physoporella dissita - Buček, p. 134, Pl. I, fig. 1.
1994 - Physoporella dissita - Bucur et al., p. 92, Pl. 11, fig. 2.
1994 - Physoporella dissita - Piros et al., p. 347, Pl. 1, figs. 9 pars, 11 pars, 12 pars.
2002 - Physoporella disita - Piros, p. 126-127, Pl. I, figs. 2-3.
2013 - Physoporella dissita - Kotański, p. 78-80, Pl. XVI, figs. 1-30, Pl. XX, figs. 3.a, 9; Pl. XXXII, figs. 1.a, 2.b, 3.b; Pl. XXXIII, fig. 1.c; Pl. XXXIV, fig. 4.c; Pl. XXXV, figs. 1.c, 2.c; Pl. & XXXVII, figs. 2.c, 3.a; Pl. XXXVIII, fig. 2.b
2013 - Physoporella cf. dissita - Rychliński et al., Pl. 2, figs. 6-8
2021 - Oligoporella dissita - Gawlick et al., Pl. 15, figs. k, r.
2021 - Oligoporella minutula - Gawlick et al., Pl. 15, figs. h-j.
Origin of the name: Perhaps after Latin "dissitus" = scattered, possibly referring to laterals.
Studied material: G�mbel (1872, p. 271) cites "Wetterstein" limestone from Zugspitzegruppe, and pale limestone from Schwarzau, southeast of T�rnitz in Lower Austria as type locality of Gyroporella dissita. The material of Physoporella dissita, described by Pia (1912, p. 45; Pl. VI, figs. 1-4; = this paper, Pl. 11 ![]() , figs. 1-3, 8) originates from the vicinity Saalfelden, Austria. Pia (1920, p. 55) stated that Tiefenbach was incorrectly defined as the original locality, and changed it to �fenbach north of Saalfelden. It belongs to thin sections LXII1, LXII3, LXII4 and LXII5.
, figs. 1-3, 8) originates from the vicinity Saalfelden, Austria. Pia (1920, p. 55) stated that Tiefenbach was incorrectly defined as the original locality, and changed it to �fenbach north of Saalfelden. It belongs to thin sections LXII1, LXII3, LXII4 and LXII5.
In thin-sections CDXXXIII1 (Pl. 10 ![]() , figs. 3, 5), CDXXXIII3 (Pl. 10
, figs. 3, 5), CDXXXIII3 (Pl. 10 ![]() , fig. 7) and CDXXXIII4, from Pia's collection I found cross-sections of algae which, with their structure (double whorls of piriform laterals and annulation), strongly resemble Oligoporella dissita, but are significantly smaller, so I determined it as Oligoporella cf. dissita. The material is from the Ladinian deposits, as indicated by the presence of Diplopora annulata in the mentioned thin-sections. The catalogue of Pia's collection for these thin-sections states: "Physoporella lotharingica, 1931". A specimen of this algae is illustrated by Pia (1931, Pl. XXI, Fig. 8) from possibly the Ladinian of Moselle, east France as an imprint of algae on the rock surface, but not from a thin-section. Due to poor preservation it cannot be determined with certainty. Unfortunately, there is no sample label in the work. I have no other information about the site from which the sample CDXXXIII was taken, but it is not impossible that this is the same material described by Pia (1931). Dimension fits to those stated by Pia (1931).
, fig. 7) and CDXXXIII4, from Pia's collection I found cross-sections of algae which, with their structure (double whorls of piriform laterals and annulation), strongly resemble Oligoporella dissita, but are significantly smaller, so I determined it as Oligoporella cf. dissita. The material is from the Ladinian deposits, as indicated by the presence of Diplopora annulata in the mentioned thin-sections. The catalogue of Pia's collection for these thin-sections states: "Physoporella lotharingica, 1931". A specimen of this algae is illustrated by Pia (1931, Pl. XXI, Fig. 8) from possibly the Ladinian of Moselle, east France as an imprint of algae on the rock surface, but not from a thin-section. Due to poor preservation it cannot be determined with certainty. Unfortunately, there is no sample label in the work. I have no other information about the site from which the sample CDXXXIII was taken, but it is not impossible that this is the same material described by Pia (1931). Dimension fits to those stated by Pia (1931).
G�mbel's material is lost and among Pia's material, as the most representative specimen, I choose the one from thin-section LXII2 illustrated on Pl. 11 ![]() , fig. 1 (= Pia, 1912, Pl. VI, fig. 1) as a neotype. Other specimens from the same sample become isotypes. The material from Croatia, with which I supplemented the research of this species, includes specimens from the Anisian deposits of the Hum-�agudovac area on the northern slopes of Medvednica Mt (Pl. 21
, fig. 1 (= Pia, 1912, Pl. VI, fig. 1) as a neotype. Other specimens from the same sample become isotypes. The material from Croatia, with which I supplemented the research of this species, includes specimens from the Anisian deposits of the Hum-�agudovac area on the northern slopes of Medvednica Mt (Pl. 21 ![]() , figs. 3-4, 8).
, figs. 3-4, 8).
History of investigations: G�mbel (1872, p. 271) describes a new species of Gyroporella dissita: "The shell as in previous species (G. cylindrica), the diameter of the tubes is 2 mm; but the annular articles are very high (1 mm), and have fewer but wider canals (G�mbel, 1872, Pl. DIII, fig. 1). So far, this species is known only from cross-sections and cuttings, enclosed in rock. However, these cross-sections show such deep incisions at the ring joints, and relatively very widely spaced bulges, such as have never been observed in normal G. annulata, so this feature was crucial for establishing the new species."
(In German: "Geh�use, wie bei den vorausgehenden Arten, der Durchmesser der R�hrchen betr�gt jedoch 2 mm dabei sind die Ringglieder sehr hoch (1 mm) und von weniger zahlreichen, aber weiteren Kan�lchen durchzogen (Taf. D III Fig. 1) Bis jetzt ist diese Art nur aus Durchschnitten und D�nnschliffen rings eingeschlossen im Gesteine bekannt. Diese Durchschnitte zeigen aber so tiefe Einschnitte an den Ringfugen und verh�ltnissm�ssig sehr, weit auseinander stehende W�lste, wie sie bei der normalen G. annulata nie beobachtet wurden, wesshalb dieser Eigenth�mlichkeit durch die Aufstellung einer besonderen Art Rechnung getragen wurde.")
Although, the diagnosis is focused on the morphology, the illustration (G�mbel, 1872, Pl. DIII, fig. 1) is relatively good, in relation to his other drawings, and it shows an annulled thallus and thin laterals in double whorls, which are the basic characteristics of this species.
Pia (1912, p. 45; see also Granier & Sander, 2013, p. 32) gives a new description and renames the species as Physoporella dissita: "The most conspicuous feature of this species is the segmentation of the calcareous skeleton. At first sight it appears that it is a typical annulate form. However, closer inspection shows that there is never more than one whorl to a segment. Therefore I believe we are more in accordance with the facts if we interpret the segmentation of our species as an extreme of bulge development, as an enhanced advancement of the behaviour displayed in some specimens of Oligoporella pilosa (Pl. IV, fig. 5 = this paper, Pl. 5 ![]() , fig. 7) and also in the same way in Physoporella minutula, described below. This is even more significant because true segmentation occurs neither in Physoporella nor in the related Oligoporella. The branches, always tapered distally, are arranged in tightly packed whorls most of which have a very regular biserial structure. The outer surface is always completely enclosed in calcite."
, fig. 7) and also in the same way in Physoporella minutula, described below. This is even more significant because true segmentation occurs neither in Physoporella nor in the related Oligoporella. The branches, always tapered distally, are arranged in tightly packed whorls most of which have a very regular biserial structure. The outer surface is always completely enclosed in calcite."
Bystrick� (1964, p. 127) summarizes Pia's diagnosis: "Thallus extremely undulated ("undulatio") or articulated ("annulatio"). The pores in two-row whorls, perpendicular to the longitudinal axis of the cell, pear-shaped and with closed distal ends. The article belongs to only one whorl."
Kotański (2013, p. 79) gave a diagnosis on the base of isolated specimens: "Markedly segmented or even annulated forms. Each segment is connected with one whorl, composed of two rows with alternating pores. This segmentation is very well developed and is an indispensable feature of this species. The tubercles are piriform, but not so remarkable pear-shaped, as in Ph. praealpina. Nevertheless, the pores never, or only very rarely, transect the wall of the cylinder, which is decisive in assigning these forms to the genus Physoporella. Dimensions are comparable to those of Ph. praealpina. If the wall is not preserved, the only feature allowing the distinction of these two species is the distances between whorls of the double-row tubercles, which are distinctly larger in Ph. dissita."
Description: The thallus is cylindrical with a smooth inner and annulated outer wall (Pl. 11 ![]() , figs. 1, 3; Pl. 13
, figs. 1, 3; Pl. 13 ![]() , fig. 1; Pl. 21
, fig. 1; Pl. 21 ![]() , figs. 4, 8; G�mbel, Pl. DIII, fig. 1; Bystrick�, 1957, Pl. VI, figs. 6-7; 1964, Pl. XXI, figs. 1-7; 1966, Pl. VI, figs. 1, 5; Buček, 1989, Pl. I, fig. 1; Gawlick et al., 2021, Pl. 15, figs. h-i). Annulation is the result of intense calcification at the level of the whorls and weak or very weak calcification between the whorls. It is pronounced because the whorls are clearly spaced (Fig. 17
, figs. 4, 8; G�mbel, Pl. DIII, fig. 1; Bystrick�, 1957, Pl. VI, figs. 6-7; 1964, Pl. XXI, figs. 1-7; 1966, Pl. VI, figs. 1, 5; Buček, 1989, Pl. I, fig. 1; Gawlick et al., 2021, Pl. 15, figs. h-i). Annulation is the result of intense calcification at the level of the whorls and weak or very weak calcification between the whorls. It is pronounced because the whorls are clearly spaced (Fig. 17 ![]() ). Calcification often reaches the stem cell, as indicated by the preserved basal parts of the laterals (Pl. 11
). Calcification often reaches the stem cell, as indicated by the preserved basal parts of the laterals (Pl. 11 ![]() , fig. 1; Pl. 21
, fig. 1; Pl. 21 ![]() , figs. 4, 8). In the less calcified specimens, part of the stem cell is covered only with a very thin calcareous layer (Pl. 11
, figs. 4, 8). In the less calcified specimens, part of the stem cell is covered only with a very thin calcareous layer (Pl. 11 ![]() , figs. 1, 3; Pl. 13
, figs. 1, 3; Pl. 13 ![]() , fig. 1), while in more heaverly calcified specimens this layer is thicker, but due to a mechanically weak spot it is sometimes susceptible to breakage (Pl. 21
, fig. 1), while in more heaverly calcified specimens this layer is thicker, but due to a mechanically weak spot it is sometimes susceptible to breakage (Pl. 21 ![]() , figs. 4, 8). In some specimens, calcification did not reach the stem cell, so the basal parts of the laterals are not preserved (Pl. 11
, figs. 4, 8). In some specimens, calcification did not reach the stem cell, so the basal parts of the laterals are not preserved (Pl. 11 ![]() , fig. 6). Annulation is the most characteristic feature of the species Oligoporella dissita and enables its differentiation from other species of the genus Oligoporella in tangential sections (Pl. 11
, fig. 6). Annulation is the most characteristic feature of the species Oligoporella dissita and enables its differentiation from other species of the genus Oligoporella in tangential sections (Pl. 11 ![]() , figs. 2, 4, 7; Pl. 21
, figs. 2, 4, 7; Pl. 21 ![]() , fig. 3; Bystrick�, 1966, Pl. VI, fig. VI. 2b; Rychliński et al., 2013, Pl. 2, fig. 6; Gawlick et al., 2021, Pl. 15, fig. k), and also in poorly preserved material.
, fig. 3; Bystrick�, 1966, Pl. VI, fig. VI. 2b; Rychliński et al., 2013, Pl. 2, fig. 6; Gawlick et al., 2021, Pl. 15, fig. k), and also in poorly preserved material.
The shape of the laterals is piriform, but with a more pronounced distal narrowing, and somewhat resembles the tip of a spear
(Pl. 11 ![]() ,
fig. 1; Pl. 21
,
fig. 1; Pl. 21 ![]() , fig. 8; Bystrick�, 1957, Pl. VI, fig. 7; 1964, Pl. XXI, figs. 1, 3; 1966, Pl. VI, figs. 1, 5), although there are also specimens in which this narrowing is not so pronounced (Pl. XI, figs. 3, 6, 8; Bystrick�, 1964, Pl. XXI, figs. 2, 5; 1986, Pl. III, fig. 2; Bucur et al., 1994, Pl. 11, fig. 2; Gawlick et al., 2021, Pl. 15, fig. i). The laterals, as with other species, had trichoblasts that probably fell off at the end of the vegetative phase, so they are rarely preserved as fossils. Calcified trichoblasts are visible on the right side of the specimen on
Pl. 11
, fig. 8; Bystrick�, 1957, Pl. VI, fig. 7; 1964, Pl. XXI, figs. 1, 3; 1966, Pl. VI, figs. 1, 5), although there are also specimens in which this narrowing is not so pronounced (Pl. XI, figs. 3, 6, 8; Bystrick�, 1964, Pl. XXI, figs. 2, 5; 1986, Pl. III, fig. 2; Bucur et al., 1994, Pl. 11, fig. 2; Gawlick et al., 2021, Pl. 15, fig. i). The laterals, as with other species, had trichoblasts that probably fell off at the end of the vegetative phase, so they are rarely preserved as fossils. Calcified trichoblasts are visible on the right side of the specimen on
Pl. 11 ![]() ,
fig. 8.
,
fig. 8.
The laterals are arranged in spaced double whorls, so that each annulled article corresponds to one whorl, and within the whorls laterals are arranged alternately, which is best seen in tangential sections
(Pl. 11 ![]() ,
figs. 2, 4-7). The laterals were densely arranged within the whorl and were separated in the proximal part only by thin calcareous lamellae (Pl. 11
,
figs. 2, 4-7). The laterals were densely arranged within the whorl and were separated in the proximal part only by thin calcareous lamellae (Pl. 11 ![]() , figs. 4-6). They were placed mostly perpendicular to the stem cell (Pl. 11
, figs. 4-6). They were placed mostly perpendicular to the stem cell (Pl. 11 ![]() , figs. 1, 3, 6, 9; Pl. 21
, figs. 1, 3, 6, 9; Pl. 21 ![]() , fig. 8; Bystrick�, 1957, Pl. VI, figs. 6-7; 1964, Pl. XXI, figs. 1, 5, 7; Bucur et al., 1994, Pl. 11, fig. 2), and rarely very slightly inclined upwards (Pl. 13
, fig. 8; Bystrick�, 1957, Pl. VI, figs. 6-7; 1964, Pl. XXI, figs. 1, 5, 7; Bucur et al., 1994, Pl. 11, fig. 2), and rarely very slightly inclined upwards (Pl. 13 ![]() , fig. 1; Pl. 21
, fig. 1; Pl. 21 ![]() , figs. 3-4; Bystrick�, 1964, Pl. XXI, figs. 2-4).
, figs. 3-4; Bystrick�, 1964, Pl. XXI, figs. 2-4).
Emended diagnosis: Cylindrical thallus with smooth inner and annulated outer surface. The shape of the laterals is piriform, often with a more pronounced distal narrowing, so it somewhat resembles the tip of a spear. The laterals are densely arranged in spaced double whorls, so that each annulated thallus article corresponds to a single whorl. Within the whorl laterals are arranged alternately. They are placed perpendicular, rarely slightly obliquely to the longitudinal axis of the thallus.
Similarities and differences: The species Oligoporella dissita differs from other species of this genus by the more pronounced thinning of the laterals and a distinct annulation of the thallus, and is relatively easily recognizable in thin-sections.
|
|
Figure 17: Oligoporella dissita (G�mbel, 1872), n. comb. Idealized longitudinal (A) and tangential (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts in laterals extension are shown on the upper whorl. The tangential section (B) shows the sections of the proximal (upper whorl) and distal parts of the laterals (lower whorl). The annulation of the outer surface of the calcareous thallus is visible. |
5.2.8. Oligoporella laevis (Praturlon, 1963), n. comb.
(= Physoporella laevis Praturlon, 1963)
1963 - Physoporella laevis n. sp. - Praturlon, p. 128-130, Pl. IV, figs. 1-7.
Origin of the name: after Latin "laevis" = light, referring to delicate calcareous envelope.
Studied material: Praturlon (1963, Pl. IV, figs. 1-7) illustrated seven specimens of this alga from several thin-sections from sample G1/1-8 from the area of Auronzo (Cadore region) in the Dolomites, Italy. The type strata are the uppermost levels of the Upper Permian "Bellerophon-zone," dark bituminous clayey micritic limestones, just a few inches below the light grey marls of the Lower Triassic with Myophoria and Claraia. It occurs together with Gymnocodium bellerophontis. The author chose a specimen from thin-section G 1/8 (illustrated on Pl. IV, fig. 2 = this paper, Fig. 18 ![]() ) as a holotype. Specimens from the same sample (Praturlon, 1963, Pl. IV, figs. 5, 7) are therefore isotypes.
) as a holotype. Specimens from the same sample (Praturlon, 1963, Pl. IV, figs. 5, 7) are therefore isotypes.
History of investigations: Praturlon (1963, p. 128) gives a rather long description of the new species Physoporella laevis: "Aspect in thin section - Owing perhaps to a feeble calcification, well-preserved specimens are few. The branches are filled with crystalline calcite, the central hollow is frequently marked by dark secondary detritus. Thin calcareous walls, reddish in colour, line axial hollow and single branches. Sections very oblique to the axis show an ellipse truncated at both extremities (Pl. IV, 4) by meeting of two consecutive whorls with the cutting plane. More elongated sections (Pl. IV, 2, 3) show besides, at regular intervals, lateral branches couples, to each of which corresponds a biseriate whorl. Easily recognizable, but nearly always incomplete, the cross-sections. Very rare the axial sections. Main cell and calcareous wall - Thallus cylindrical, lined with a very thin calcareous layer, which would correspond not to the usual calcareous wall but merely to the calcified cell-membrane. This is a rather rare character among the Dasycladaceae; it seems to be constant in this species, and not in connection with unfavourable lighting. Branches shape and disposition - Branches euspondyl, alternating in biseriate whorls arranged at regular intervals along the stem (Pl. IV, figs. 1-2). The branches depart from main cell perpendicular to the axis, but often they get inclined in opposite directions. In their axial sections they appear as egg-shaped bodies, fastened to the wall by means of a brief stalk, not always visible. They swell very quickly in proximal portion, then close to the exterior lengthening in a rather smooth point. We are in presence of the pirifer type, and it is justifiable to consider them as both assimilatory and reproductive organs."
Description: The thallus is cylindrical, but due to the specific style of calcification it has a different shape than in other species of this genus (Fig. 18 ![]() ). The calcareous envelope is made of a very thin (≈ 0.02 mm) micrite layer that envelops the stem cell and each lateral separately, the so-called "individual sheath" (De Castro, 1997). The cell wall can not calcify, as stated by the author of the species, because among Dasycladalean algae calcification take part in the mucus layer right next to the cell wall. This type of calcification is also present in Cylindroporella taurica and Salpingoporella sellii (see discussion in Conrad & Varol, 1990). This style of calcification may be a reaction to a very calm and protected environment where the protection by a thick skeleton was not required. The cavities of the laterals and stem cell were filled with sparite cement during diagenesis.
). The calcareous envelope is made of a very thin (≈ 0.02 mm) micrite layer that envelops the stem cell and each lateral separately, the so-called "individual sheath" (De Castro, 1997). The cell wall can not calcify, as stated by the author of the species, because among Dasycladalean algae calcification take part in the mucus layer right next to the cell wall. This type of calcification is also present in Cylindroporella taurica and Salpingoporella sellii (see discussion in Conrad & Varol, 1990). This style of calcification may be a reaction to a very calm and protected environment where the protection by a thick skeleton was not required. The cavities of the laterals and stem cell were filled with sparite cement during diagenesis.
The laterals can be classified as piriform, since they are wide in the proximal part and taper distally. This distal narrowing is less pronounced than in other species, so they sometimes look ovoid. Due to the peculiar calcification, the basal part of the lateral, i.e., the peduncle, has been preserved (Praturlon, 1963, Pl. IV, fig. 3). Trichoblasts have not been preserved, they probably fell off before calcification. The laterals are arranged in distinct double whorls (Figs. 18 ![]() - 19
- 19 ![]() ; Praturlon, 1963, Pl. IV, figs. 1-3, 6). The laterals in a whorl are next to each other, but between them is (in most cases) a micritic wall. Due to the dense arrangement of laterals within the whorl, they generally stand alternately. The two rows of laterals only slightly diverge, i.e., they are almost parallel. The reason for this is in the pronounced distance of their basal pores, which is very small in other species of the genus Oligoporella. The whorls are clearly spaced.
; Praturlon, 1963, Pl. IV, figs. 1-3, 6). The laterals in a whorl are next to each other, but between them is (in most cases) a micritic wall. Due to the dense arrangement of laterals within the whorl, they generally stand alternately. The two rows of laterals only slightly diverge, i.e., they are almost parallel. The reason for this is in the pronounced distance of their basal pores, which is very small in other species of the genus Oligoporella. The whorls are clearly spaced.
Emended diagnosis: Cylindrical thallus with a distinctive dark micritic sheath that envelops the stem cell and each lateral separately. The laterals are of the piriform type, more ovoid and arranged in spaced double whorls. In the basal part they have a thin and a short peduncle. The divergence of the rows within the whorl is not pronounced. The laterals stand inside the whorl mostly alternately.
Similarities and differences: Due to the distinctive calcification and less elongated laterals, there is no possibility of confusion with any other species of the genera Oligoporella and Physoporella.
|
|
Figure 18: Original illustration of the holotype of Oligoporella laevis (Praturlon, 1963), n. comb. (from Praturlon, 1963, Pl. IV, fig.
2). Scale bar = 1 mm. |
|
|
Figure 19: Oligoporella laevis (Praturlon, 1963), n. comb. Idealized longitudinal (A), tangential (B) and transverse (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts are shown on some laterals. |
5.2.9. Species from the Middle Triassic of Upper Silesia
Kotański (2013) published a monograph on Anisian Dasycladales from Upper Silesia and adjacent regions of southern Poland. In contrary to the "classical" specimens of Dasycladales that are preserved as calcareous tubes with pores of laterals, specimens from Upper Silesia are from dolomite and are preserved as natural moulds and imprints, with tubercles instead of pores. Although the material is well described and extensively illustrated, it is very difficult to compare it with the "classical" material. Material from thin-sections is usually very poorly preserved and that from isolated samples has, due to its different preservation, a different appearance. This is why I simply reproduce the original diagnosis of this author's species and give only a short comment with the new generic attributions according to conclusions from this work. Previously known species are commented on in the corresponding chapters.
Kotański (2013, p. 58-61) established Oligoporella silesiaca (G�mbel, 1872), n. comb., based on the species Gyroporella silesiaca G�mbel, 1872. Later this species has been considered as a synonym partly of Diplopora annulata and partly of Diplopora nodosa by Pia (1920, p. 28). Analysing the original illustrations, I consider G. silesiaca a junior synonym of D. nodosa only. Kotański (2013) in the synonymy list also included two citations. Gyroporella infundibuliformis G�mbel, 1872 is also considered as synonym partly of D. annulata and partly of D. nodosa by Pia (1920, p. 28). De Castro (1979) included it in the synonymy of D. nodosa which is accepted here. Diplopora cf. silesiaca was described by Pastwa-Leszczyńska and Śliwiński (1960, p. 687, fig. 7 = Pl. III, fig. 13), that I already included in the synonymy list of Oligoporella minutula (Granier & Grgasović, 2000). It was used by Kotański (2013) as a neotype of O. silesiaca. I keep my previous decision, not only because the mentioned specimen strongly resembles those of O. minutula, but it clearly differs by having thicker laterals that the original material of G�mbel (1872, Pl. DIII, fig. 6).
Kotański (2013, p. 61-63) established Oligoporella balinensis (Raciborski ex Pastwa-Leszczyńska & Śliwiński, 1960), n. stat., based on Oligoporella pilosa forma balinensis described by Pastwa-Leszczyńska and Śliwiński (1960, p. 689, fig. 9 = Pl. III, fig. 14: = this paper, Pl. 17 ![]() , fig. 8) with only one specimen. I consider it as a junior synonym of Oligoporella minutula (G�mbel, 1872), n. comb. (see
Chapter 5.2.5.).
, fig. 8) with only one specimen. I consider it as a junior synonym of Oligoporella minutula (G�mbel, 1872), n. comb. (see
Chapter 5.2.5.).
Oligoporella chrzanowensis Kotański, 2013
1960 - Gyroporella multiserialis G�mbel - Pastwa-Leszczyńska & Śliwiński, Pl. II, fig. 11.
2013 - Oligoporella chrzanowensis n. sp. - Kotański, p. 64-66, Pl. X, figs. 1-24, Pl. XXXVI, fig. 1.a.
Holotype: Kotański (2013, Pl. X, fig. 1), paratypes: idem, Fig. 15; Pl. X, 2, 5, 14). Repository: Polish Geological Institute, Warsaw, Geological Institute, coll. Z. Kotański no.MuzPIG 1653.II.
Origin of the name: After the type area Chrzan�w: locality Kąty Chrzanowskie (Rosowa G�ra), no. 48, and Balin, no. 50, in the Cracow-Chrzan�w region, Upper Silesia, Poland.
Type horizon: Upper part of the Diplopora Dolomite, Upper Anisian.
Original diagnosis: "Thallus without annulation or undulation, with amassment of short, trichophore oblique branches (tubercles) arranged in indistinct, dense whorls (euspondyl arrangement) or irregularly aspondyl arrangement)."
Description: Laterals are typical piriform, not trichoform, if we follow conclusions from this work. Analysing illustrations it seems to me that the whorls are double-rowed, although very dense, so it is not easily to observe. This is clearly visible on the upper part of the holotype (Pastwa-Leszczyńska & Śliwiński, Pl. II, fig. 11) and in Kotański (2013, Pl. X, fig. 7). An aspondyl arrangement is just an illusion based on the very dense arrangement of the laterals. There is no aspondyl piriform species known yet.
Emended diagnosis: Cylindrical thallus with piriform laterals arranged alternately in very dense double whorls.
Oligoporella polonoandalusica (Kotański, 2013), n. comb.
1935b - Physoporella aff. lotharingica - Pia, Pl. II, fig. 2.
2013 - Oligoporella polonoandalusica n. comb. - Kotański, p. 83-86, Pl. XVII, figs. 1-20.
Holotype: Kotański (2013, Pl. XVII, fig. 6); paratypes: idem, Pl. XVII, figs. 1, 5.
The type locality: Przełajka, Upper Silesia, Poland.
Repository: Polish Geological Institute, Warsaw, Geological Institute, coll. Z. Kotański no. MuzPIG 1653.II.
Origin of the name: After a palaeogeographical province in the Middle Triassic, stretching from Poland to Andalusia.
Original diagnosis: Cylindrical thallus with a very small diameter (1-2 mm), very long and frequently bent. The calcareous wall is rather thin and slightly undulated. The pores (tubercles) are piriform and are distributed in very widely spaced double whorls. This species belongs to the Physoporella praealpina group (two rows of alternate pores (tubercles) per one whorl).
Description: the species is easily distinguished in having typical piriform laterals arranged in quite widely spaced double whorls. The author put Diplopora rauffi Ahlburg in the synonymy list, although with question mark; this is not acceptable since Ahlburg's specimens have only one row of laterals.
5.3. Genus Neophysoporella n. gen.
Previous authors, as described in Chapter 4.6. based the generic determination mainly on the openness and closedness of the pores, not taking sufficient account on the structure of the alga itself. In this paper, priority is given to biological characteristics, i.e., the actual appearance and structure of the algae. Thus, it was shown in the investigated material that Ladinan and late Triassic species attributed to the genus Physoporella have a different structure compared to those from the Permian and Anisian. This primarily refers to the shape of the laterals that are relatively shorter, and do not have such a pronounced tapering towards their distal end, but are more rounded. In addition, the whorls are more densely packed, and the calcareous sheath is relatively thinner. These differences are clear when comparing the illustrations on the attached figures. Therefore, I decided to combine these species into a new genus with the following diagnosis:
Neophysoporella n. gen.: Cylindrical thallus with a relatively thin calcareous sheath. Laterals are of the piriform type, but relatively short, more rounded and with less pronounced tapering towards the distal end, and arranged in whorls.
The type species of the genus is Diplopora lotharingica Benecke, 1898.
The new genus most likely evolved from the genus Physoporella, as explained in Chapter 4.6. In addition to the type, the new genus includes the species Physoporella jomdaensis Fl�gel & Mu, 1982, and Physoporella zamparelliae Parente & Climaco, 1999, as well as specimens described as Physoporella leptotheca from the Norian of Betic Cordilleras by Braga (1986).
5.3.1. Neophysoporella lotharingica (Benecke, 1898), n. comb.
(= Physoporella lotharingica (Benecke, 1898), Pia, 1931;
= Physoporella pauciforata var.? lotharingica (Benecke, 1898), Pia, 1920;
= Diplopora lotharingica Benecke, 1898)
Pl. 4 ![]() , figs. 10-12; Pl. 17
, figs. 10-12; Pl. 17 ![]() , figs. 10-11
, figs. 10-11
1898 - Diplopora lotharingica n. sp. - Benecke, p. 280-281, Fig. 1.
1920 - Physoporella pauciforata var. ? lotharingica n. stat. n. comb. - Pia, p. 52-53, Pl. III, figs. 11, 13.
1931 - Physoporella lotharingica n. comb. - Pia, p. 266-267, Pl. XXI, fig. 8.
1965 - Physoporella lotharingica - Herak, p. 21-22, Pl. XIV, figs. 4-6.
1979 - Dasicladacee - De Castro, Pl. XVII, figs. 3, 5-10.
1997 - Physoporella lotharingica - Pugliese, p. 74, Pl. 1, figs. 10-13.
2001 - ?Physoporella lotharingica - Senowbari-Daryan & Di Stefano, p. 99-100, Pl. III, figs. 1-5.
2013 - Physoporella lotharingica - Kotański, p. 69-73, Pl. XII, figs. 1-20.
Origin of name: After the area where it was found - Lothringien in then western part of Germany, and after the First World War Moselle in France.
Studied material: Benecke (1898) in his description of the new species illustrated a rock sample with two specimens of this alga (idem, Fig. 1; = this paper; Pl. 17 ![]() , fig. 10). The other figure (idem, Fig. 2) is a reconstruction of the alga based on both specimens. As a type locality Benecke (1898, p. 279) cites the village of G�nglingen, about 6 km northwest of Falkenberg, today Faulquemonta is in the Moselle area of eastern France. The new alga is found in yellowish dolomites with an oolithic fabric, a few meters below the Trochiten-limestone. He defines the age as middle Muschelkalk. The author did not stated where his material is kept, but Pia (1931, p. 265) stated "Benecke, 1898. Nat. Mus., Alg. Nr. 1082 und 1083". Since I do not have any information about those samples, and the illustration is poor (magnification is 1x), I gave up defining a lectotype. It would be good to find an illustrated sample or, if the sample is lost, to re-sample.
, fig. 10). The other figure (idem, Fig. 2) is a reconstruction of the alga based on both specimens. As a type locality Benecke (1898, p. 279) cites the village of G�nglingen, about 6 km northwest of Falkenberg, today Faulquemonta is in the Moselle area of eastern France. The new alga is found in yellowish dolomites with an oolithic fabric, a few meters below the Trochiten-limestone. He defines the age as middle Muschelkalk. The author did not stated where his material is kept, but Pia (1931, p. 265) stated "Benecke, 1898. Nat. Mus., Alg. Nr. 1082 und 1083". Since I do not have any information about those samples, and the illustration is poor (magnification is 1x), I gave up defining a lectotype. It would be good to find an illustrated sample or, if the sample is lost, to re-sample.
From the same G�nglingen locality (but samples obtained from a certain director E. de Margerie), Pia (1931, Pl. XXI, fig. 8) illustrates one specimen of this species (see Chapter 5.2.7.), and changed the stratigraphic determination from Benecke (1898) to Lingula-Dolomite.
Pia (1920) also illustrates this alga. Specimens from the vicinity of Sarajevo (Pia, 1920, Pl. II, figs. 12, 14) were transferred to Physoporella pauciforata var. undulata by Pia (1935a). Specimens from Pia (1920, Pl. III, figs. 11, 13) are, according to the catalogue of Pia's collection, from thin-section 1041, which I could not find. I found only thin-section 1041a, which is completely sterile. As a site, Pia (1920) denotes Furthof, east of T�rnitz in Lower Austria, and assigns the age as Anisian Reiflingerkalk. This age deviates from the probable lower range for this species, and this statement needs to be verified.
Illustrations of Neophysoporella lotharingica on Pl. 4 ![]() , figs. 10-12 are made from the original films of Prof. Herak and correspond to the illustrations from Herak (1965, Pl. XIV, figs. 4-6). His material originates from the Ladinian deposits of Debelo brdo northwest of Knin (Pl. 4
, figs. 10-12 are made from the original films of Prof. Herak and correspond to the illustrations from Herak (1965, Pl. XIV, figs. 4-6). His material originates from the Ladinian deposits of Debelo brdo northwest of Knin (Pl. 4 ![]() , fig. 10), together with D. annulata and D. nodosa, Lower Ladinian of Gradina near Sv. Rok (Pl. 4
, fig. 10), together with D. annulata and D. nodosa, Lower Ladinian of Gradina near Sv. Rok (Pl. 4 ![]() , fig. 11), together with D. annulata and D. annulatissima, and from an undefined level of the Middle Triassic from Sekulin vrh, northwest of Lička Kaldrma (Pl. 4
, fig. 11), together with D. annulata and D. annulatissima, and from an undefined level of the Middle Triassic from Sekulin vrh, northwest of Lička Kaldrma (Pl. 4 ![]() , fig. 12).
, fig. 12).
History of investigations: Benecke (1898, p. 280-281) describes a new species Diplopora lotharingica: "Preservation is one of the most common in these organisms. All the limestone made by the plant is dissolved, and the one remaining has only to do with the cylindrical filling of the cell tube or with the imprint of the calcareous sheath. The piece of rock shown here (Fig. 1; = this paper, Pl. 17 ![]() , fig. 10) shows on the left the impression of the outer side, and on the right the tentaculite-like filling. The dimensions of the numerous available specimens are always small. The diameter does not reach 1 mm. Only fragments up to 15 mm in length were found. The apical part is not known. It was necessary to make a drawing combined from the two specimens shown on the rock sample (Benecke, 1898, Fig. 2 = this paper, Pl. 17
, fig. 10) shows on the left the impression of the outer side, and on the right the tentaculite-like filling. The dimensions of the numerous available specimens are always small. The diameter does not reach 1 mm. Only fragments up to 15 mm in length were found. The apical part is not known. It was necessary to make a drawing combined from the two specimens shown on the rock sample (Benecke, 1898, Fig. 2 = this paper, Pl. 17 ![]() , fig. 11) to show the inner structure as far as it can be recognized. There is only one whorl of lateral branches on the each ring, the number of which is very small, usually 10. If the lateral branches ended uncalcified on the outside, one should assume that one would see openings (Fig. 2 above) at the level of the outer margin. But since these openings appear to be absorbed, it is not impossible that the branch ends that protruded outwards on the older parts of the plant were also calcified and covered with a hump-like layer of limestone."
, fig. 11) to show the inner structure as far as it can be recognized. There is only one whorl of lateral branches on the each ring, the number of which is very small, usually 10. If the lateral branches ended uncalcified on the outside, one should assume that one would see openings (Fig. 2 above) at the level of the outer margin. But since these openings appear to be absorbed, it is not impossible that the branch ends that protruded outwards on the older parts of the plant were also calcified and covered with a hump-like layer of limestone."
(In German: "Die Erhaltung ist eine bei diesen Organismen h�ufig vor-kommende. Aller von der Pflanze abgeschiedene Kalk ist aufgel�st und man hat es nur mit der cylindrischen Ausf�llung des Zell-schlauches oder mit dem Abdruck der kalkigen H�lle zu thun. Das hier abgebildete Gesteinsst�ck (Fig. 1) zeigt links den Abdruck der Aussenseite, rechts die tentaculiten�hnliche Ausf�llung. Die Dimensionen zahlreich vorliegender Exemplare sind stets gering. Der Durchmesser erreicht noch nicht 1 mm. Es sind nur Fragmente bis zu 15 mm L�nge gefunden. Der Scheitel ist nicht bekannt. Uni den inneren Bau so weit derselbe erkannt werden kann - zu zeigen, wurde eine aus den beiden auf dem Gesteins-st�ck dargestellten Exemplaren combinirte Zeichnung entworfen (Fig. 2). Auf jeden Ring kommt nur ein Wirtel seitlicher Zweige, deren Zahl sehr gering, in der Regel 10, ist. Wenn die seitlichen Zweige aussen urverkalkt endigten, so sollte man vermuthen, dass man im Abdruck (Fig. 2 oben) im Niveau der �usseren Begrenzung liegende Oeffnungen zu sehen bek�me. Da aber diese Oeffnungen grubig vertieft erscheinen, so ist es nicht unm�glich, dass die nach aussen hinaustretenden Zweigenden an den �lteren Theilen der Pflanze ebenfalls verkalkten und mit einer buckelartig aufgetriebenen Kalklage �berzogen wurden.")
Pia (1920, p. 52-53) changes the name and rank of this species to Physoporella pauciforata var. ? lotharingica, and illustrates specimens from the Reisalp and the vicinity of Sarajevo which, as already mentioned, were later added by Pia (1935a) to the species Physoporella pauciforata var. undulata.
Pia (1931, p. 266) again changed the name and rank of the species to Physoporella lotharingica, with the following comments: "The description from Benecke applies in all essential points. The pores were apparently closed to the outside. The calcification was weak, the shell was deeply constricted between the whorls. Benecke's figure is pretty good. As a supplement, I bring a photo (Pia, 1931, Pl. XXI, fig. 8) that is similar to Benecke's Figure 2 and shows the stone core at the bottom and the impression at the top. As I have repeatedly pointed out, there is every reason to believe that we are dealing with a Physoporella. A clear difference to certain stunted forms of the Physopor. pauciforata does not exist."
Ellenberger (1958, p. 194), comparing washed stone algal cores from Ladino Vanoise (Savoie, France), considers Physoporella lotharingica to be an atrophied variety of Diplopora uniserialis Pia. This, of course, does not make sense because the mentioned forms belong to different families of Dasycladalean algae.
Another description of this alga was given by Pugliese (1997, p. 74): "The thallus is simple, with a cylindrical shape. The calcareous wall is thin, due to calcification that involves only a small part of the branches, in particular the inner portion. For this reason, it is quite difficult to evaluate the branch distribution, even if they seem to be organised in single verticils. The shape of the pores is tricophorous, in some cases piripherous and they are generally closed at the distal end. Reproductive organs were not found." It is noticeable that even in the recent works, openness and closedness of pores are considered as an important taxonomic criterion (see Chapter 4).
The most recent diagnosis is by Kotański (2013, p. 69): "The thallus is cylindrical with a very small diameter (1-2 mm). The calcareous wall is rather thin. Pores (tubercles) are piriform (closed) and distributed in single whorls. The spaces between the whorls are so large that particular whorls of tubercles are distinctly separated. This species belongs to the Physoporella pauciforata group (one row of pores per one whorl)."
Description: The structure of these algae is very difficult to reconstruct on the basis of stone cores or thallus impressions (Benecke, 1898; Pia, 1931; Ellenberger, 1958; Kotański, 2013), especially since they are mostly drawings and not photographs, so I base my description primarily on specimens from thin-sections (Pia, 1920; Herak, 1965; De Castro, 1979; Pugliese, 1997; Senowbari-Daryan & Di Stefano, 2001).
The thallus is cylindrical, and the preserved thallus is generally quite short. The diameter of the thallus and thus the number of laterals in a whorl is very variable (compare Pl. 4 ![]() , fig. 10 and figs. 11-12). Calcification is very peculiar in this species. The calcareous sheath is relatively thin, but very irregular. It covers the outer surface of the laterals as an irregular layer of uneven thickness with a rough outer surface. Due to the irregular and uneven calcification, the thallus was more susceptible to diagenetic changes and more difficult to preserve. Cracks are also very common. In better preserved specimens (Pl. 4
, fig. 10 and figs. 11-12). Calcification is very peculiar in this species. The calcareous sheath is relatively thin, but very irregular. It covers the outer surface of the laterals as an irregular layer of uneven thickness with a rough outer surface. Due to the irregular and uneven calcification, the thallus was more susceptible to diagenetic changes and more difficult to preserve. Cracks are also very common. In better preserved specimens (Pl. 4 ![]() , figs. 10, 12 above; Pia, 1920, Pl. III, fig. 11; De Castro, 1979, Pl. XVII, figs. 3, 6), the calcareous sheath covers the laterals in an uneven layer, so that their shape is largely preserved. In other specimens (Pl. 4
, figs. 10, 12 above; Pia, 1920, Pl. III, fig. 11; De Castro, 1979, Pl. XVII, figs. 3, 6), the calcareous sheath covers the laterals in an uneven layer, so that their shape is largely preserved. In other specimens (Pl. 4 ![]() , figs. 11, 12 below; De Castro, 1979, Pl. XVII, figs. 5, 7-10; Senowbari-Daryan & Di Stefano, 2001, Pl. III, figs. 1-4), it appears that the calcareous sheath does not completely cover the laterals, but only covered, or, perhaps only touched, their distal parts. Due to this type of calcification, it is more difficult to reconstruct the form of this alga (Fig. 20
, figs. 11, 12 below; De Castro, 1979, Pl. XVII, figs. 5, 7-10; Senowbari-Daryan & Di Stefano, 2001, Pl. III, figs. 1-4), it appears that the calcareous sheath does not completely cover the laterals, but only covered, or, perhaps only touched, their distal parts. Due to this type of calcification, it is more difficult to reconstruct the form of this alga (Fig. 20 ![]() ), but on the other hand it simplifies the determination of this alga in thin-sections.
), but on the other hand it simplifies the determination of this alga in thin-sections.
The shape of the laterals is piriform with a wide proximal part and a distal narrowing, yet this narrowing is not pronounced and the laterals are shorter and more rounded. The basal part of the laterals and their contact with the stem cell are not preserved. The shape of the laterals is best seen in Pl. 4 ![]() , fig. 10; Pia (1920, Pl. III, fig. 11) and in De Castro (1979, Pl. XVII, figs. 3, 6). The shape of the laterals is not trichoform, as stated by Pugliese (1997), who probably focused on the openness and closedness of the pores. Like other laterals, those in N. lotharingica probably carried trichoblasts. The laterals were arranged in relatively dense whorls, as can be seen from the symmetrical position of the laterals on both sides of the thallus in longitudinally-oblique sections (Pl. 4
, fig. 10; Pia (1920, Pl. III, fig. 11) and in De Castro (1979, Pl. XVII, figs. 3, 6). The shape of the laterals is not trichoform, as stated by Pugliese (1997), who probably focused on the openness and closedness of the pores. Like other laterals, those in N. lotharingica probably carried trichoblasts. The laterals were arranged in relatively dense whorls, as can be seen from the symmetrical position of the laterals on both sides of the thallus in longitudinally-oblique sections (Pl. 4 ![]() , fig. 10; De Castro, 1979, Pl. XVII, fig. 3), uniformly crossed laterals in cross section (De Castro, 1979, Pl. XVII, fig. 6) and in tangential sections (Pugliese, 1997, Pl. 1, fig. 13). The laterals are placed perpendicular to the central stem. In the whorl they probably touched in the proximal part or were very close. Adjacent whorls were very close, but probably did not touch, or very rarely did so. Although there is no doubt about the euspondyl arrangement of the laterals, they appear to have been arranged a little more "untidy" than in the previously described species of Physoporella and Oligoporella.
, fig. 10; De Castro, 1979, Pl. XVII, fig. 3), uniformly crossed laterals in cross section (De Castro, 1979, Pl. XVII, fig. 6) and in tangential sections (Pugliese, 1997, Pl. 1, fig. 13). The laterals are placed perpendicular to the central stem. In the whorl they probably touched in the proximal part or were very close. Adjacent whorls were very close, but probably did not touch, or very rarely did so. Although there is no doubt about the euspondyl arrangement of the laterals, they appear to have been arranged a little more "untidy" than in the previously described species of Physoporella and Oligoporella.
Emended diagnosis: Cylindrical thallus, quite variable in diameter. The calcareous sheath is thin and irregular, of uneven thickness and with rough outer surface. It covers the outer surfaces of the laterals or only their distal parts. The laterals are of the piriform type, but somewhat shorter and more rounded. They are placed perpendicular to the stem cell and distributed in relatively dense whorls.
Similarities and differences: Neophysoporella lotharingica n. comb., is very similar to other species of the genus Neophysoporella n. gen., from which it differs by the irregular calcification and somewhat larger dimensions.
|
|
Figure 20: Neophysoporella lotharingica (Benecke, 1898), n. comb. Idealized longitudinal (A) and cross (B) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous skeleton. Trichoblasts are shown on some laterals. Irregular calcification is visible. |
5.3.2. Neophysoporella jomdaensis Fl�gel & Mu, 1982, n. comb.
(= Physoporella jomdaensis Fl�gel & Mu, 1982)
Pl. 16 ![]() , fig. 10; Pl. 19
, fig. 10; Pl. 19 ![]() , figs. 7-9
, figs. 7-9
1982 - Physoporella jomdaensis n. sp. - Fl�gel & Mu, p. 61-62, Pl. 8 ![]() , fig. 5 pars; Pl. 9
, fig. 5 pars; Pl. 9 ![]() , figs. 3-4.
, figs. 3-4.
1997 - Physoporella jomdaensis - Grgasović, p. 203, 207, Pl. II, figs. 10, 14-15.
2020 - Physoporella jomdaensis - Bucur et al., p. 517-518, Pl. 8J.
Origin of the name: After the town of Jomda in eastern Tibet which is next to the type locality.
Studied material: Fl�gel and Mu (1982) illustrate material from thin-sections 76 CJ 12 (Pl. 8 ![]() , fig. 5 pars), 76 CJ 23 (Pl. 8
, fig. 5 pars), 76 CJ 23 (Pl. 8 ![]() , fig. 4) and 76 CJ 25 (Pl. 8
, fig. 4) and 76 CJ 25 (Pl. 8 ![]() , fig. 3, holotype = this paper, Pl. 16
, fig. 3, holotype = this paper, Pl. 16 ![]() , fig. 10). The sample is from the Carnian Jiapeila Formation in the vicinity of the town of Jomda in the Qambdo district of eastern Tibet. The thin-sections are stored at the Nanjing Institute of Geology and Paleontology, Nanjing (China).
, fig. 10). The sample is from the Carnian Jiapeila Formation in the vicinity of the town of Jomda in the Qambdo district of eastern Tibet. The thin-sections are stored at the Nanjing Institute of Geology and Paleontology, Nanjing (China).
I also based the description on the material from the Norian of �umberak Mt (Grgasović, 1997, Pl. II, figs. 10, 14-15 = this work, Pl. 19 ![]() , figs. 7-9).
, figs. 7-9).
History of investigation: Fl�gel and Mu (1982, p. 61-62) gave a diagnosis of a new species Physoporella jomdaensis: "Thallus thin, cylindrical in shape, slightly undulated up to 11.70 mm in length. Branches piriform; thickest at the proximal parts, tapering to the distal ones and ending with distinct rounded terminations. The branches are arranged perpendicularly to the stem cell and organized into spiral rows and verticils. Calcification is rather weak; the distal part of the thallus is covered by a thin calcareous sheet (00035-0.070 mm thick), which also protects the horn-like protuberances on the surface of the thallus."
Grgasović (1997, p. 207) describes specimens of this species from the Norian Main Dolomite of �umberak Mt: "In their morphological characteristics the �umberak specimens fully agree with those from the original description or this species. There are some differences in dimensions, however they are consistent with the limits of variations known in the other species of this genus. The thallus of Physoporella jomdaensis is cylindrical, straight (Grgasović, 1997, Pl. II, fig. 15 = this work, Pl. 19 ![]() , fig. 8), very slightly bent (idem, Pl. II, fig. 14 = this work, Pl. 19
, fig. 8), very slightly bent (idem, Pl. II, fig. 14 = this work, Pl. 19 ![]() , fig. 9) or slightly clavate (Fl�gel & Mu, 1982, Pl. 9, fig. 3). Branches are of the typical pear like - pirifer type and disposed in whorls, which are the principal characteristics of the genus Physoporella. Internal branch parts, as well as the surface of the main axis were most likely not calcified. Calcification affects separately the external side of each branch and is complete, so that the external thallus surface is undulated. The calcareous envelope is about 0.06 mm thick. The proximal parts of branch envelopes are fused (idem, Pl. II, fig. 15, top = this work, Pl. 19
, fig. 9) or slightly clavate (Fl�gel & Mu, 1982, Pl. 9, fig. 3). Branches are of the typical pear like - pirifer type and disposed in whorls, which are the principal characteristics of the genus Physoporella. Internal branch parts, as well as the surface of the main axis were most likely not calcified. Calcification affects separately the external side of each branch and is complete, so that the external thallus surface is undulated. The calcareous envelope is about 0.06 mm thick. The proximal parts of branch envelopes are fused (idem, Pl. II, fig. 15, top = this work, Pl. 19 ![]() , fig. 8, top; Fl�gel & Mu, 1982, Pl. 9, fig. 3, middle part). Branches in individual and neighbouring whorls are densely disposed as far as their calcareous envelopes allow it. The original thallus structure was most probably aragonite, like in recent Dasycladales, but, in the early diagenetic phases it was replaced by tiny mosaic dolomite crystals, as are other skeletal particles in this dolomite. The phenomenon described by Fl�gel and Mu (1982) that "... the distal part of the thallus is covered by a thin calcareous sheet (0.035 - 0.070 mm thick)..." has not been recorded in the original material. This might be due to the diagenetic origin of this envelope in the type specimens or possibly in the form of the early diagenetic acicular calcite."
, fig. 8, top; Fl�gel & Mu, 1982, Pl. 9, fig. 3, middle part). Branches in individual and neighbouring whorls are densely disposed as far as their calcareous envelopes allow it. The original thallus structure was most probably aragonite, like in recent Dasycladales, but, in the early diagenetic phases it was replaced by tiny mosaic dolomite crystals, as are other skeletal particles in this dolomite. The phenomenon described by Fl�gel and Mu (1982) that "... the distal part of the thallus is covered by a thin calcareous sheet (0.035 - 0.070 mm thick)..." has not been recorded in the original material. This might be due to the diagenetic origin of this envelope in the type specimens or possibly in the form of the early diagenetic acicular calcite."
Bucur et al. (2020, p. 517-518) described one well-preserved specimen from the Upper Triassic of Shikoku Island, Japan, that fits in dimensions of the specimens from Croatia.
Description: I have little to add to my earlier description, except that the arrangement of the laterals in the spiral rows cited by Fl�gel and Mu (1982, p. 61) does not exist, but the laterals in the adjacent whorls are arranged alternately. In any accumulation of alternately stacked objects (balls, wooden logs, snowballs, etc.) we can notice a spiral arrangement, but it is only an illusion. According to the shape of the laterals, which are relatively shorter and not so distinctly distally tapered, their dense arrangement, and the style of calcification, I attribute this species to the genus Neophysoporella n. gen.
Similarities and differences: Neophysoporella zamparelliae (Parente & Climaco, 1999) is very similar to this alga, but this species has an inner wall formed by sparite crystals and alveoli in the lateral envelopes.
5.3.3. Neophysoporella zamparelliae Parente & Climaco, 1999, n. comb.
(= Physoporella zamparelliae Parente & Climaco, 1999)
1999 - Physoporella zamparelliae n. sp. - Parente & Climaco, p. 177-178, Pl. 37, figs. 1-10; Pl. 38, figs. 6, 9-10; Pl. 39, fig. 1.
Origin of the name: Species is dedicated to Prof. Valeria Zamparelli (Naples) for her precious contribution to the knowledge of stratigraphy, facies distribution and palaeontology of the Triassic successions of the Calabria-Lucania border.
Studied material: The authors based their description (Parente & Climaco, 1999, Pl. 37, figs. 1-4, 6-10; Pl. 38, figs. 6, 9-10; Pl. 39, fig. 1) on quite rich material from samples AC 554, AC 561, and AC 565. As a holotype they name the specimen from thin-section AC 554.15 (idem, Pl. 37, fig. 5 = this paper, Fig. 21 ![]() ), while the other specimens are isotypes. The material is kept in the Zamparelli collection in the "Dipartimento di Scienze della Terra", Universit� degli studi di Napoli "Federico II" in Naples (Italy).
), while the other specimens are isotypes. The material is kept in the Zamparelli collection in the "Dipartimento di Scienze della Terra", Universit� degli studi di Napoli "Federico II" in Naples (Italy).
The type locality is Mt. Rotunda in the southern part of the Apennines, on the border of the Italian provinces of Calabria and Lucania. Algae were found in the Norian dolomites.
History of investigations: Parente and Climaco (1999, p. 177) give a diagnosis of a new species: "Calcareous skeleton simple, undulated, made by partly welded thin individual sheaths enclosing the laterals. The median and distal outer surface of the sheaths are pitted by many small alveoles. Some individual sheaths end with a spine-like apophysis opening outward with a thin pore. Central cavity bordered by a thin "inner wall", sometimes finely perforated. Laterals perpendicular to the central stem axis, arranged in closely spaced simple whorls, alternating in subsequent whorls. Laterals piriferous, vertically compressed, roughly triangular both in vertical and in verticillar section. Reproductive organs unknown, probably cladospore and located in the swollen, proximal part of laterals."
Description: Parente and Climaco (1999) gave a detailed description of the algae, and I will only mention that "spine-like apophysis" represented trichoblasts on the top of laterals, as already described in numerous species. The tiny pores (alveoli) seen in the calcareous sheaths of the laterals indicate that trichoblasts (or trichoblast-like hairs) may have emerged from the entire lateral, thus increasing the assimilation surface of the algae. After the relatively short and rounded piriform laterals, arranged in dense whorls, and the relatively thin calcareous sheath, I include this species in the genus Neophysoporella n. gen. It is unusual that the authors do not mention similar N. lotharingica or very similar N. jomdaensis when comparing other species.
Similarities and differences: Other algae of the genus Neophysoporella n. gen. are similar to this alga. Missidentification with N. lotharingica is not possible, but N. jomdaensis is quite similar to it. An important difference is the existence of an inner sparite wall and alveoli in laterals, although this second characteristic may be lost due to diagenesis.
|
|
Figure 21: Neophysoporella zamparelliae (Parente & Climaco, 1999), n. comb. (from Parente & Climaco, 1999, Pl. 37, fig. 5,
holotype). Scale bar = 1 mm. |
5.3.4. Neophysoporella n. sp. (Braga, 1986)
Pl. 16 ![]() , fig. 2
, fig. 2
1986 - Physoporella leptotheca - Braga, p. 242, 244, 2.a-2.g, 2.o.
Studied material: The material illustrated by Braga (1986, Fig. 2.b-2.g, 2.o; Fig.2.a = this paper, Pl. 16 ![]() , fig. 2) originates from the Norian dolomites of Sierra de Baza, Alpuj�rride complex, Betic Cordillera, SE Spain. The illustrated specimens are from samples SB-105 and SB-108. From the same dolomites and the same locality the following taxa were described (Braga & Mart�n, 1987): Gyroporella plumosa Zanin Buri, Griphoporella curvata (G�mbel), Spinaporella andalusica Fl�gel & Fl�gel-Kahler, and Spinaporella? granadiensis Fl�gel & Fl�gel-Kahler.
, fig. 2) originates from the Norian dolomites of Sierra de Baza, Alpuj�rride complex, Betic Cordillera, SE Spain. The illustrated specimens are from samples SB-105 and SB-108. From the same dolomites and the same locality the following taxa were described (Braga & Mart�n, 1987): Gyroporella plumosa Zanin Buri, Griphoporella curvata (G�mbel), Spinaporella andalusica Fl�gel & Fl�gel-Kahler, and Spinaporella? granadiensis Fl�gel & Fl�gel-Kahler.
History of investigations: Braga (1986, p. 242, 244) described specimens of this alga as Physoporella leptotheca Kochansky-Devid�: "The thalli are long and cylindrical with a thin wall, whose inner surface is always well defined, although its thickness varies according to the amount of calcareous material accumulated. The branches are arranged in whorls. As in the examples of Ph. leptotheca described by Fois (1979), they are generally well spaced, but even in the same individual, there may also be closely or irregularly spaced whorls. The arrangement of the branches varies from approximately metaspondylus to euspondylus. They are piriferous, opened by fine canals, perpendicular or slightly oblique to the internal face and irregularly scattered in the whorl."
Description: Calcification is very peculiar in this alga. The calcareous wall is not thin, as stated by Braga (1986), but is relatively thick, e.g., "double". The structure of the calcareous wall is best seen in a specimen from Braga (Fig. 2.a = this work, Pl. 16 ![]() , fig.
2). A very thin white (transparent) wall is placed next to the stem cell and around the laterals. However, another such thin wall is visible slightly below the level of the tops of the laterals, so the calcareous wall appears "hollow". This "cavity" is filled with micrite, which may be part of the primary thallus of algae. In other specimens (idem, Fig. 2.b-d) the "cavity" is filled with microsparite, possibly resulting from recrystallization of the micrite filling. The laterals protrude slightly beyond the described calcareous thallus. The laterals are very similar to those of other species of the genus Neophysoporella n. gen., being somewhat more rounded, almost bubble-shaped, but with visible tapering toward distal end. They are arranged in whorls, whose spacing highly varies. The illustrations do not show the metaspondyl arrangement or irregular scattering mentioned by Braga (1986), perhaps prompted by other descriptions of P. leptotheca, to which he ascribed his specimens. The laterals do not show a grouping of two or three, as in the mentioned species, but are normally arranged in the whorls.
, fig.
2). A very thin white (transparent) wall is placed next to the stem cell and around the laterals. However, another such thin wall is visible slightly below the level of the tops of the laterals, so the calcareous wall appears "hollow". This "cavity" is filled with micrite, which may be part of the primary thallus of algae. In other specimens (idem, Fig. 2.b-d) the "cavity" is filled with microsparite, possibly resulting from recrystallization of the micrite filling. The laterals protrude slightly beyond the described calcareous thallus. The laterals are very similar to those of other species of the genus Neophysoporella n. gen., being somewhat more rounded, almost bubble-shaped, but with visible tapering toward distal end. They are arranged in whorls, whose spacing highly varies. The illustrations do not show the metaspondyl arrangement or irregular scattering mentioned by Braga (1986), perhaps prompted by other descriptions of P. leptotheca, to which he ascribed his specimens. The laterals do not show a grouping of two or three, as in the mentioned species, but are normally arranged in the whorls.
Due to the characteristic shape of the whorled laterals, as well as their arrangement, this alga is not Physoporella leptotheca (compare Pl. 16 ![]() , fig. 2 and Pl. 16
, fig. 2 and Pl. 16 ![]() , figs. 11-12, 14), but is a new species that belongs to the genus Neophysoporella n. gen. Since the original material was not available to me, I did not dare to name a new species, but left it under open nomenclature. I suggest re-examination of the original material of this very interesting species.
, figs. 11-12, 14), but is a new species that belongs to the genus Neophysoporella n. gen. Since the original material was not available to me, I did not dare to name a new species, but left it under open nomenclature. I suggest re-examination of the original material of this very interesting species.
5.4. Genus Euteutloporella De Castro, 1993
The genus Euteutloporella was established by De Castro (1993) on the basis of the type species Chaetetes? triasinus Schauroth, 1859 (later Teutloporella triasina (Schauroth, 1859) Pia, 1912) for euspondyl algae with trichoform laterals. Later Euteutloporella peniculiformis (Ott, 1963) Grgasović & Sokač ex Piros & Preto, 2008 was included in this genus. The following species can also be added:
5.4.1. Euteutloporella chia (Herak, 1967a), n. comb.
(= Oligoporella chia Herak, 1967a)
Pl. 4 ![]() , fig. 6
, fig. 6
1967a - Oligoporella chia n. sp. - Herak, p. 100, Pl. 7, figs. 5-7; Pl. 8, figs. 1-3.
1972b - Oligoporella chia - Ott, p. 266-270, Pl. II.
Origin of the name: After the Greek island of Chios in the Aegean Sea where it was found.
Studied material: The species is described from Ladinian deposits of the Ligaridi site on the Greek island of Kh�os, i.e., Chios in the Aegean Sea. Among the illustrated material Herak
(1967a, Pl. 7, figs. 5-7; Pl. 8, figs. 1-3) chose as holotype the specimen from Pl. 7, fig. 7 = this paper, Pl. 4 ![]() , fig. 6) from sample T359, thin-section c. The type material is kept in the collection of K.-W. Tietze, Geologisch-pal�ontologischen Institut der Universit�t Marburg Lahn (Germany).
, fig. 6) from sample T359, thin-section c. The type material is kept in the collection of K.-W. Tietze, Geologisch-pal�ontologischen Institut der Universit�t Marburg Lahn (Germany).
History of investigations: Herak (1967a, p. 100) gave a diagnosis for the new species Oligoporella chia: "The stem cell is cylindrical; the branches are slender, slightly tapered towards the outside and set obliquely to the stem cell. They are arranged in a clear, distinctly single-row in regular whorls. The distance between the whorls is relatively large. The surface of the calcareous cylinder is variable. Most of the specimens show a true regular undulation. But there are also sections where this feature cannot be determined or where only traces of it are present. At first I suspected that on the basis of this characteristic one could establish different species or varieties. But an overview of all the material has led me to the conviction that it is a question of individual variability, and not in the organization of the organism, but in the dimensions and the external shape of the calcareous shell. You can see all possible transitions. I therefore think it is justified to provide a somewhat broader framework for the species. This species is most similar to the O. pilosa. The differences relate primarily to the distance between the individual whorls, their clear single rows and the undulation of the calcareous envelope, which can be seen in O. chia."
(In German: Die Stammzelle ist zylindrisch; die Aste sind schlank, gegen au�en etwas verj�ngt und schr�g zur Stammzelle gestellt. Sie sind in deutlichen, ausgesprochen einzeiligen und regelm��igen Wirteln angeordnet. Die Entfernung der Wirtel voneinander ist relativ gro�. Die Oberfl�che des Kalkzylinders ist variabel. Die meisten Exemplare zeigen eine echte regelm��ige Undulation. Es gibt aber auch solche Schnitte, wo man dieses Merkmal nicht feststellen kann oder wo nur Spuren davon vorhanden sind. Zuerst vermutete ich, da� man auf Grund dieses Merkmals verschiedene Arten oder Variet�ten aufstellen k�nnte. Aber eine �bersicht des ganzen Materials hat mich zur �berzeugung gef�hrt, da� es sich um individuelle Variabilit�t handelt, und zwar nicht in der Organisation des Organismus, sondern in den Dimensionen und in der Au�enform der Kalkh�lle. Man kann alle m�glichen Oberg�nge feststellen. Ich halte es daher f�r gerecht-fertigt, einen etwas breiteren Rahmen der Art vorzusehen. Diese Art ist am �hnlichsten der O. pilosa. Die Unterschiede beziehen sich vor allem auf die Entfernung zwischen den einzelnen Wirteln, ihre deutliche Einreihigkeit und die Undulation der Kalkh�lle, welche man bei O. chia feststellen kann.")
Ott (1972a, p. 106) comments on this species: "It bears whorls arranged at distinct intervals. The thin branches are more reminiscent of Teutloporella than Oligoporella. For the most part, they do not thin at all inside the wall, but are unchanged. The author's observations on the rich material of this species from Karaburun have shown that a phloiophorous funnel follows from the outside, after a sudden narrowing, of which in many cases only the beginning has been preserved. This relationship is reminiscent of the genus Favoporella described by Sokač. In general, Oligoporella chia has similarities with the Anisian Physoporella-Oligoporella flora through clearly spaced whorls, through sausage-like pores (which also occur in Ph. minutula) and through probably phloiophorous branch extensions (Hurka, 1969, Fig. 6)." Later Ott (1972b, Pl. II) published specimens from Karaburun (Turkey), without description, but on two illustrated specimens no "phloiophorous extensions" are visible, but only trichoform laterals. These may be secondary (diagenetically) enlarged pore openings.
Description: Laterals of this species are typical for the trichoform type: thin, and further thinning to the top, quite similar to those of the species Teutloporella herculea (Stoppani), which is clearly seen in the published illustrations and the holotype (Herak, 1967a, Pl. 7, fig. 7; = this work, Pl. 4 ![]() , fig. 6). That is why I add this species to the genus Euteutloporella.
, fig. 6). That is why I add this species to the genus Euteutloporella.
Emended diagnosis: Cylindrical thallus with smooth inner and slightly undulated, rarely flat, outer wall. The laterals are trichoform, oblique and slightly concavely bent, arranged in spaced whorls.
5.5. Genus Ardeiporella n. gen.
The algae described by Pia (1935a) under the name Oligoporella karrerioidea has a structure, and this is especially true of laterals, which differs significantly from that of the genera Physoporella and Oligoporella, as well as other known genera. Therefore, based on this species, I establish a new genus Ardeiporella. Since the new genus, for now, contains only one species, I give a joint diagnosis of genus and species (descriptio generico-specifica, Art. 38.5. ICN, Turland et al., 2018):
Ardeiporella n. gen.: Cylindrical thallus with a relatively thick calcareous wall and a relatively narrow stem cell. The laterals consist of a narrow proximal part ("neck"), a clearly widened and flattened central part ("head") and an elongated distal part tapering towards the end ("beak"). The laterals are oblique and arranged in whorls.
The name of the genus is derived from the shape of the laterals, which resembles the head of a heron (Latin "ardea" = heron).
5.5.1. Ardeiporella karrerioidea (Pia, 1935a), n. comb.
(= Oligoporella karrerioidea Pia, 1935a)
Pl. 15 ![]() , figs. 3, 7-8; Pl. 17
, figs. 3, 7-8; Pl. 17 ![]() , fig. 2
, fig. 2
1935a - Oligoporella karrerioidea n. sp. - Pia, p. 219-221, Pl. III, figs. 1-3.
Origin of the name: Unknown, perhaps from the German "karren", meaning cart, and the laterals in cross-section somewhat resemble a cart, i.e., wheelbarrow with a "head" as a wheel and "beak" as handles, but this is only a guess.
Studied material: The material described by Pia (1935a) originates from Bosnia and Herzegovina: thin-section 235a from Ograda (site "t") on the road Are�in brijeg - Grab, 575b from the village of Haliluci, east of Sarajevo (site "r"). For the specimen from Pia (1935a, Pl. III, fig. 1 = this work, Pl. 17 ![]() , fig. 2) the author does not mentioned the locality, and I could not find it in Pia's collection.
, fig. 2) the author does not mentioned the locality, and I could not find it in Pia's collection.
Pia (1935a) as a type specimen (i.e., holotype) determined the specimen on Pl. XV, fig. 3. Specimen from Pl. XV, fig. 7 is thus an isotype.
History of investigations: Pia (1935a, p. 219-221) describes the new species Oligoporella karrerioidea: "This is one of the unusual, but also one of the most difficult forms in the flora. Already in the type of preservation, it noticeably differs from the others. As a rule, the shells of the Bosnian diplopores - and especially the Oligoporellas - are composed of very pure, whitish translucent calcite, so that they form the lightest part in a thin section. In contrast, the shells of Oligoporella karrerioidea appear cloudy and murky in cross section. Apparently they were relatively loose during life and rich in organic components. The middle part of the shell is occupied by a very narrow cavity, which certainly only offered space for the stem cell. It is surrounded by a thin, almost closed cylinder. The passage points of the whorled branches can only rarely be seen in it (Pia, 1935a, Pl. 3, fig. 3 = this work, Pl. 15 ![]() , fig. 3). I assume that these were usually blocked by calcite towards the end of the life of alga. There is a zone which contains ring-shaped cavities that stand close to each other and surround the stem cell. As can be seen from their shape, especially in the more oblique sections (idem, Pl. III, figs. 1, 3; = this paper, Pl. 17
, fig. 3). I assume that these were usually blocked by calcite towards the end of the life of alga. There is a zone which contains ring-shaped cavities that stand close to each other and surround the stem cell. As can be seen from their shape, especially in the more oblique sections (idem, Pl. III, figs. 1, 3; = this paper, Pl. 17 ![]() , fig. 2; Pl. 15
, fig. 2; Pl. 15 ![]() , fig. 3), they correspond to the proximal parts of the branches that are close to one another or perhaps that have grown together. As a result of the lack of space, these were rather narrow, but high, and apparently often irregularly turned up and down (idem, Pl. 3, fig. 3 = this work, Pl. 15
, fig. 3), they correspond to the proximal parts of the branches that are close to one another or perhaps that have grown together. As a result of the lack of space, these were rather narrow, but high, and apparently often irregularly turned up and down (idem, Pl. 3, fig. 3 = this work, Pl. 15 ![]() , fig. 3). They probably contained the sporangia, for which there was hardly any space in the thin stem cell. Finally, in the outermost part of the shell, the branches separate from each other and are somewhat arched upwards. They are only slightly or not at all thinned towards the exit point."
, fig. 3). They probably contained the sporangia, for which there was hardly any space in the thin stem cell. Finally, in the outermost part of the shell, the branches separate from each other and are somewhat arched upwards. They are only slightly or not at all thinned towards the exit point."
(In German: "Dies ist eine der merkw�rdigsten, aber auch de schwierigsten Formen der ganzen Flora. Schon in der Art der ErhaltungErhaltung weicht sie von den anderen auffallend ab. Vergl. die Tafel. In der Regel sind die Schalen der bosnischen Diploporen - und gerade der Oligoporellen - aus sehr reinem, weisslich durchscheinendem Kalkspat zusammengesetzt, so dass sie in den D�nnschliffen den hellsten Teil bilden. Dagegen erscheinen die Skelette von Oligoporella karrerioidea im Schliff tr�b und schmutzig. Offenbar waren sie im Leben verh�ltnism�ssig locker und reich an organischen Beimengungen. Der mittlere Teil der Schale wird von einem sehr engen Hohlraum eingenommen, der sicher nur f�r die Stammzelle Platz bot. Er wird umgeben von einem d�nnen, fast geschlossenen Zylinder. Nur selten erkennt man in ihm die Durchtritt-stellen der Wirtel�ste (Taf. 3, Fig. 3). Ich m�chte vermuten, dass diese gegen das Lebensende der Alge zu h�ufig durch Kalk verschlossen wurden. An diesen d�nnen Hohlzylinder schliesst sich aussen eine Zone, die nahe �ber einander folgende, die Stammzelle umgreifende, ringf�rmige Hohlr�ume enth�lt. Wie aus ihrer Form, besonders in den st�rker schr�gen Schnitten 1 und 3, hervorgeht, entsprechen sie den dicht an einander schlies-senden, oder vielleicht mit einander verwachsenen proximalen Teilen der Wirter�ste. Diese waren hier infolge Raummangels ziemlich schmal, aber hoch, scheinbar oft sogar unregelm�ssig gegen oben und unten ausgest�lpt (Taf. 3, Fig. 3). Wahrscheinlich enthielten sie die Sporangien, f�r die ja in der d�nnen Stammzelle kaum Platz war. Im �ussersten Teil der Schale endlich l�sen sich die Aeste von einander ab und verlaufen etwas bogenf�rmig gegen oben. Sie sind gegen ihre Austrittsstelle zu nur wenig oder gar nicht verj�ngt.")
Description: The thallus is cylindrical with a thick wall, which envelops all the laterals and the stem cell together
(the so-called collective sheath after De Castro, 1997), and a narrow central
cavity. The inner wall is smooth, and such was probably the outer, but it was later damaged by diagenetic processes
(micricitization, dissolution). On the illustrated material (Pl. 15 ![]() ,
figs. 3, 7-8; Pl. 17
,
figs. 3, 7-8; Pl. 17 ![]() , fig. 2) it is not possible to observe a different mineral structure of the
thallus, which is stated by Pia (1935a). It is a usual a finely-crystalline calcite thallus with an irregular micrite
envelope.
, fig. 2) it is not possible to observe a different mineral structure of the
thallus, which is stated by Pia (1935a). It is a usual a finely-crystalline calcite thallus with an irregular micrite
envelope.
The shape of the laterals is best visible on the holotype (Pl.15, fig. 3), although some pores are secondarily expanded by dissolution. The proximal part of the lateral pore ("neck") is represented by a horizontally flattened tube (compare Pl. 15 ![]() ,
figs. 3, 8). The larger central part of the lateral ("head") is also horizontally
flattened, but it is thicker than the proximal and distal part, so the adjacent laterals touch each
other, which is nicely seen in the cross (Pl. 15
,
figs. 3, 8). The larger central part of the lateral ("head") is also horizontally
flattened, but it is thicker than the proximal and distal part, so the adjacent laterals touch each
other, which is nicely seen in the cross (Pl. 15 ![]() , fig. 8) and tangential section (Pl. 15
, fig. 8) and tangential section (Pl. 15 ![]() , fig. 3
above), and form a cavity in the calcareous thallus. The distal part of the lateral ("beak") tapers slightly towards the end. Although it is not seen on the fossil
material, it is to be expected that these laterals also carried trichoblasts in the vegetative phase (Fig. 22
, fig. 3
above), and form a cavity in the calcareous thallus. The distal part of the lateral ("beak") tapers slightly towards the end. Although it is not seen on the fossil
material, it is to be expected that these laterals also carried trichoblasts in the vegetative phase (Fig. 22 ![]() ). The extended middle part of the lateral probably had the function of
gametangia, as stated by Pia (1935a). Laterals from adjacent whorls do not touch each
other.
). The extended middle part of the lateral probably had the function of
gametangia, as stated by Pia (1935a). Laterals from adjacent whorls do not touch each
other.
|
|
Figure 22: Ardeiporella karrerioidea (Pia, 1935a), n.
comb. Idealized longitudinal (A), transverse-oblique (B) and tangential (C) sections of the thallus showing examples of possible shapes of laterals and morphology of the calcareous
skeleton. Trichoblasts are shown on some laterals. |
5.6. Genus Acroporella Praturlon, 1964
The genus Acroporella was established by Praturlon (1964, p. 177) with type species Acroporella radoicicae as "not segmented Dasycladaceae having simple, long, not ramified, acrophorous laterals". Based on the new well-preserved material Praturlon and Radoičić (1974) changed the diagnosis to: "cylindrical, not segmented Dasycladaceae having whorls of long primary laterals distally ramified in clusters of button-like secondary twigs" Sokač (1996) accepted A. radoicicae as it was originally described, and advocated that the well preserved material used by Praturlon and Radoičić (1974) does not belong to the same species. In more recent work Mancinelli (2006) supports the emendation of Praturlon and Radoičić (1974) and disagrees with Sokač (1996).
Tribe Triploporelleae Pia, 1920, emend. Bassoullet et al., 1978
Genus Acroporella Praturlon, 1964
5.6.1. Acroporella? prisca (Pia, 1912), n. comb.
(= Oligoporella prisca Pia, 1912)
Pl. 2 ![]() , figs. 5-6; Pl. 3
, figs. 5-6; Pl. 3 ![]() , figs. 1-10
, figs. 1-10
1912 - Oligoporella prisca n. sp. - Pia, p. 42-43, Pl. V, figs. 1-2.
1912 - Physoporella pauciforata - Pia, p. 44, Pl. V, fig. 18.
2020 - Physoporella prisca - Bucur & Matysik, p. 8, Fig. 6A-G
2021 non - Physoporella cf. prisca (= Phsoporella cf. pauciforata) - Gawlick et al., Pl. 5, fig. h.
Origin of the name: Probably from Latin "priscus" = former, primaeval, antique. Probably Pia gave this name thinking that the laterals in the basal part represent an ontogenetic repetition of the phylogenetically older stage (see below).
Studied material: The material of Oligoporella prisca described by Pia (1912) originates from several localities: sample XXXIV from Schlegelberg (Sandkogel) near Schwarzenbach an der Pielach, W T�rnitz, sample LVI from Schwarzenberg near Schwarzenbach an der Pielach, W T�rnitz, sample LXXIX SW Kleinzel, Reisalpe, I T�rnitz, Austria. The specimen from sample XXXIV that Pia (1912, Pl. V, fig. 18) originally described as Physoporella pauciforata, but according to all characteristics it belongs to the species Acroporella? prisca, together with all other specimens from the same sample. According to Lein (1993), the Schwarzenberg site is a part of steep cliffs between the peaks of Schwarzenberg (1096 m) and Schl�gelberg (1057 m, see also Piros et al., 1994, p. 345) and was built from the Anisian Steinalmkalk, and not from Reiflingkalk, as originally thought by Pia (1912).
Pia (1912) did not define a holotype, so I choose a specimen from Pia (1912, Pl. V, fig. 4 = this paper, Pl. 3 ![]() , fig. 3) as a lectotype, as it best shows the characteristics of this species. Other specimens from the same sample (Pl. 3
, fig. 3) as a lectotype, as it best shows the characteristics of this species. Other specimens from the same sample (Pl. 3 ![]() , figs. 1-2, 4-10) are isotypes.
, figs. 1-2, 4-10) are isotypes.
Pia's specimens are the only specimens of this species published so far. The specimens illustrated by Bystrick� (1964, Pl. IX, figs. 4-6) as Oligoporella cf. prisca show some similarities, but do not belong to this species. The specimen illustrated by Popa and Dragastan (1973, Pl. VIII, fig. 31) does not belong to this species (probably Ph. pauciforata). Kotański (2013, p. 52) stated that it is relatively frequent in several localities of the Diplopora Dolomite in Upper Silesia but did not give any illustrations of this alga in his well-illustrated monograph. He stated that it is difficult to distinguish it from Oligoporella elegans, which is true, so I believe that his material belongs to the former species.
History of investigations: Pia (1912, p. 42-43) gives a description of a new species Oligoporella prisca: "This species has very remarkable and phylogenetically important characteristics. Originally I had assigned the various sections I had of this species not only to two different species, but even to different genera. If one considers, for example, the figures in Pia (1912, Pl. V, fig. 7 and Pl. V, fig. 8 = this work, Pl. 3 ![]() , figs. 6, 8), one will probably not hesitate to define the former as Oligoporella and the latter as euverticillate Macroporella. Although both forms always occur together in the same rock and individual thin-sections, one such as Pia (1912, Pl. V, fig. 6 = this work, Pl. 3
, figs. 6, 8), one will probably not hesitate to define the former as Oligoporella and the latter as euverticillate Macroporella. Although both forms always occur together in the same rock and individual thin-sections, one such as Pia (1912, Pl. V, fig. 6 = this work, Pl. 3 ![]() , fig. 10) seems to show a transition. I would probably not have recognized the error of such a view if I had not come into possession of the unusually long and conveniently located tangential section (idem, Pl. V, fig. 1 = this work, Pl. 3
, fig. 10) seems to show a transition. I would probably not have recognized the error of such a view if I had not come into possession of the unusually long and conveniently located tangential section (idem, Pl. V, fig. 1 = this work, Pl. 3 ![]() , fig. 7). This shows at one end (judging by the inclination of the whorl branches it is the lower one) undoubtedly an expansion of the pores towards the outside, while at the other end they narrow somewhat in the distal direction. The same specimen thus combines the characteristics of Macroporella and Oligoporella in itself. At first glance, this seems to question the basis of our whole system. On closer inspection, however, we recognize in the relationships described a phylogenetically highly instructive analogy with recent algae. We know for example from Neomeris that their most basal whorled branches have a much more primitive structure than the upper ones and is described as a youth stage. These of youth stage, as I shall have the opportunity to explain further, form a repetition of phylogenetically older stages. On the other hand, we have good reason to believe that Oligoporella evolved from Macroporella (see the section on phylogeny). I therefore consider it a perfectly plausible explanation of the observations if we assume that the lower, phloiophore whorls of Oligoporella prisca are the repetition of an ancestral form. They may at the same time give us a picture of what the youth stages of not only this but also many other trichoform species. In Pia (1912, Fig. 15) I tried to show how we can imagine the transition between the phloiophore and trichoform whorls. Perhaps similar conditions existed as in the present species, only to a much lesser extent, also in Oligoporella pilosa, since the degree of tapering of the pores also changes with it. The basal part of Oligoporella prisca is not easy to miss, since it is the only known combination of phloiophore form with euverticillate arrangement of the whorl branches. The upper section of the plant is reminiscent of Oligoporella serripora due to the narrowness of the inner cavity, but it clearly differs from it by the always only slight tapering of the whorled branches, which in most cases is even weaker than in Oligoporella pilosa. The pores are in pure, simple whorls in contrast to the other two species of the genus. In the outer part of the shell this regularity is of course blurred, as on Pia (1912, Pl. V, fig. 1 = this work, Pl. 3
, fig. 7). This shows at one end (judging by the inclination of the whorl branches it is the lower one) undoubtedly an expansion of the pores towards the outside, while at the other end they narrow somewhat in the distal direction. The same specimen thus combines the characteristics of Macroporella and Oligoporella in itself. At first glance, this seems to question the basis of our whole system. On closer inspection, however, we recognize in the relationships described a phylogenetically highly instructive analogy with recent algae. We know for example from Neomeris that their most basal whorled branches have a much more primitive structure than the upper ones and is described as a youth stage. These of youth stage, as I shall have the opportunity to explain further, form a repetition of phylogenetically older stages. On the other hand, we have good reason to believe that Oligoporella evolved from Macroporella (see the section on phylogeny). I therefore consider it a perfectly plausible explanation of the observations if we assume that the lower, phloiophore whorls of Oligoporella prisca are the repetition of an ancestral form. They may at the same time give us a picture of what the youth stages of not only this but also many other trichoform species. In Pia (1912, Fig. 15) I tried to show how we can imagine the transition between the phloiophore and trichoform whorls. Perhaps similar conditions existed as in the present species, only to a much lesser extent, also in Oligoporella pilosa, since the degree of tapering of the pores also changes with it. The basal part of Oligoporella prisca is not easy to miss, since it is the only known combination of phloiophore form with euverticillate arrangement of the whorl branches. The upper section of the plant is reminiscent of Oligoporella serripora due to the narrowness of the inner cavity, but it clearly differs from it by the always only slight tapering of the whorled branches, which in most cases is even weaker than in Oligoporella pilosa. The pores are in pure, simple whorls in contrast to the other two species of the genus. In the outer part of the shell this regularity is of course blurred, as on Pia (1912, Pl. V, fig. 1 = this work, Pl. 3 ![]() , fig. 7) shows, on which not all of the originally present pores are visible."
, fig. 7) shows, on which not all of the originally present pores are visible."
(In German: "Diese Art weist sehr bemerkenswerte und phylogenetisch wichtige Eigenheiten auf. Urspr�nglich hatte ich die verschiedenen Durchschnitte, die mir von St�cken dieser Spezies vorlagen, nicht nur zwei verschiedenen Arten, sondern sogar verschiedenen Gattungen zugeteilt. Betrachtet man etwa die Figuren Taf. V, Fig. 7 und Taf. V, Fig. 8, so wird man wohl nicht z�gern, die erstere als Oligoporella, die letztere als euvertizillate Macroporella zu bestimmen. Das Irrt�mliche einer solchen Auffassung h�tte ich, trotzdem beide Formen stets zusammen im selben Gestein vorkommen und gewisse Schliffe, wie Taf. V, Fig. 6 einen �bergang zu vermitteln scheinen, wohl nicht erkannt, wenn ich nicht durch einen gl�cklichen Zufall in den Besitz des ungew�hnlich langen und g�nstig gelegenen Tangentialschliffes Taf. V, Fig. 1 gekommen w�re. Dieser zeigt an seinem einen Ende (nach der Neigung der Wirtel�ste zu urteilen ist es das untere) zweifellos eine Erweiterung der Poren gegen au�en, w�hrend sie sich am anderen Ende eher schon etwas in distaler Richtung verengern. Es vereinigt also dasselbe Exemplar die Merkmale von Macroporella und Oligoporella in sich. Auf den ersten Blick scheint dies die Grundlage unserer ganzen Systematik in Frage zu stellen. Bei n�herem Zusehen jedoch erkennen wir in den geschilderten Verh�ltnissen eine phylogenetisch h�chst lehrreiche Analogie mit rezenten vertizillierten Siphoneen. Wir wissen n�mlich z. B. von Neomeris, da� ihre basalsten Wirtel�ste einen viel primitiveren Bau als die oberen haben und sich an die Jugendstadien anschlie�en. Diese Jugendstadien wiederum bilden, wie ich weiterhin auszuf�hren Gelegenheit haben werde, eine Wiederholung phylogenetisch �lterer Zust�nde. Anderseits haben wir guten Grund zu der Annahme, da� Oligoporella aus Macroporella hervorgegangen ist (siehe den Abschnitt �ber Phylogenie, Seite 53). Ich halte es daher f�r eine durchaus plausible Erkl�rung der Beobachtungen, wenn wir annehmen, da� die unteren, phloiophor gebauten Wirtel von Oligoporella prisca die Wiederholung einer Ahnenform sind. Sie m�gen uns zugleich ein beil�ufiges Bild davon geben, wie die Jugendstadien nicht nur dieser, sondern auch vieler anderer trichophorer Arten beschaffen waren. In Fig. 15 habe ich darzustellen versucht, wie wir uns den �bergang zwischen den phloiophoren und trichophoren Wirteln etwa denken k�nnen. Vielleicht bestanden �hnliche Verh�ltnisse wie bei der vorliegenden Art, nur in viel geringerem Ausma�e, auch bei Oligoporella pilosa, da auch bei ihr das Ma� der Verj�ngung der Poren wechselt. Der basale Teil von Oligoporella prisca ist nicht leicht zu verkennen, da er die einzige bisher sicher bekannte Kombination phloiophorer Form mit euvertizillater Anordnung der Wirtel�ste darstellt. Der obere Abschnitt der Pflanze erinnert durch die Enge des inneren Hohlraumes an Oligoporella serripora, von der er sich aber sofort durch die stets nur geringe Verj�ngung der Wirtel�ste unterscheidet, die in den meisten F�llen sogar noch schwacher als bei Oligoporella pilosa ist. Die Poren stehen in reinen, einfachen Wirteln im Gegensatz zu den beiden anderen Arten der Gattung. Im �u�eren Teile der Schale ist diese Regelm��igkeit freilich verwischt, wie Taf. V, Fig. 1 zeigt, an der allerdings auch nicht alle urspr�nglich vorhandenen Poren mehr sichtbar sein d�rften.")
Bucur and Matysik (2020) followed Grgasović (1995) and synonymized the species O. elegans and O. prisca, with the latter species as senior synonym being assigned to the genus Physoporella. They did not give a new description.
Description: The thallus is cylindrical, relatively slender, mostly straight (Pl. 3 ![]() , figs. 1-3; Bucur & Matysik, 2020, Fig. 6.A-D), rarely slightly curved (Pl. 3
, figs. 1-3; Bucur & Matysik, 2020, Fig. 6.A-D), rarely slightly curved (Pl. 3 ![]() , fig. 7). The inner wall of the thallus is smooth (Pl. 2
, fig. 7). The inner wall of the thallus is smooth (Pl. 2 ![]() , fig. 6; Pl. 3
, fig. 6; Pl. 3 ![]() , figs. 1 pars, 6, 8-9; Bucur & Matysik, 2020, Fig. 6.A-B), except when damaged by diagenetic processes. Calcification probably did not reach the stem cell, since the pores are wide in their proximal part, i.e., there is no narrowing that usually occurs at the base of the laterals. The outer wall of the thallus may be relatively smooth (Pl. 3
, figs. 1 pars, 6, 8-9; Bucur & Matysik, 2020, Fig. 6.A-B), except when damaged by diagenetic processes. Calcification probably did not reach the stem cell, since the pores are wide in their proximal part, i.e., there is no narrowing that usually occurs at the base of the laterals. The outer wall of the thallus may be relatively smooth (Pl. 3 ![]() , fig. 6), but usually is slightly serrated with short thorns (Pl. 2
, fig. 6), but usually is slightly serrated with short thorns (Pl. 2 ![]() , fig. 5; Pl. 3
, fig. 5; Pl. 3 ![]() , figs. 1-3, 9-10; Bucur & Matysik, 2020, Fig. 6.A-B), which are possibly the result of calcification of the distal parts of laterals (see below).
, figs. 1-3, 9-10; Bucur & Matysik, 2020, Fig. 6.A-B), which are possibly the result of calcification of the distal parts of laterals (see below).
The shape of the laterals, despite the relatively rich and well-preserved material, is difficult to determine. It is clear that it is not a piriform type, since there is no proximal thickening or the distal thinning. From the tangential and oblique cross-sections (Pl. 3 ![]() , figs. 1, 3, 7, 9-10) it is clear that the pores have a round cross-section of relatively uniform diameter, indicating an acroform type of laterals, while their distal end is rounded. It is almost certain that the laterals continued out of the calcareous thallus, but it is not certain whether they are perhaps second-order phloioform laterals or only a bundle of trichoblasts. Indication for the trichoblasts is due to the shape of some thorns (Pl. 3
, figs. 1, 3, 7, 9-10) it is clear that the pores have a round cross-section of relatively uniform diameter, indicating an acroform type of laterals, while their distal end is rounded. It is almost certain that the laterals continued out of the calcareous thallus, but it is not certain whether they are perhaps second-order phloioform laterals or only a bundle of trichoblasts. Indication for the trichoblasts is due to the shape of some thorns (Pl. 3 ![]() , figs. 4, 10). Second order laterals are identified on the left side of the specimen from Pl. 3
, figs. 4, 10). Second order laterals are identified on the left side of the specimen from Pl. 3 ![]() , fig. 9, and especially the upper part of specimen from Pl. 3
, fig. 9, and especially the upper part of specimen from Pl. 3 ![]() , fig. 3, where bundles of the second-order laterals appear to grow on the primary ones. All this is not completely certain, so I gave generic determination with a question mark. This question mark also matches if we take into account previously mentioned disagreements on the diagnosis of the type species of the genus Acroporella, regardless of whether we accept the view of Praturlon and Radoičić (1974) or Sokač (1996). Photographs that I made from Pia's thin-sections are not of sufficient quality, so for the better determination of this material more research is needed.
, fig. 3, where bundles of the second-order laterals appear to grow on the primary ones. All this is not completely certain, so I gave generic determination with a question mark. This question mark also matches if we take into account previously mentioned disagreements on the diagnosis of the type species of the genus Acroporella, regardless of whether we accept the view of Praturlon and Radoičić (1974) or Sokač (1996). Photographs that I made from Pia's thin-sections are not of sufficient quality, so for the better determination of this material more research is needed.
As for the phloioform laterals in the lower part of the thallus, which Pia discusses extensively, they are not visible on the specimen on Pl. 3 ![]() , fig. 7 (= Pia, 1912, Pl. V, fig. 1). On the contrary, cross-sections of the pores in the lower part of the thallus are exactly the same as in the rest.
, fig. 7 (= Pia, 1912, Pl. V, fig. 1). On the contrary, cross-sections of the pores in the lower part of the thallus are exactly the same as in the rest.
The laterals are uniformly arranged in regularly spaced, relatively dense, whorls. The laterals are inclined with respect to the longitudinal axis of the thallus at an angle of about 50�. Within the whorl and in adjacent whorls laterals are clearly separated from each other and only exceptionally touch each other (Pl. 3 ![]() , figs. 6, 9). In adjacent whorls, the laterals stand approximately one above the other (Pl. 3
, figs. 6, 9). In adjacent whorls, the laterals stand approximately one above the other (Pl. 3 ![]() , figs. 1, 10) or alternately (Pl. 3
, figs. 1, 10) or alternately (Pl. 3 ![]() , figs. 1, 3, 7). Specimens from Bucur and Matysik (2020, Fig. 6.A-G) are smaller and have a relatively larger distance between whorls.
, figs. 1, 3, 7). Specimens from Bucur and Matysik (2020, Fig. 6.A-G) are smaller and have a relatively larger distance between whorls.
Similarities and differences: The similarity exists only with the species Physoporella? elegans, which is also little known, and besides supposedly two orders of laterals, it differs mostly in its smaller dimensions. There is a possibility that they are synonyms, but there is a need to reinvestigate the type material.
5.7. Genus Teutloporella Pia, 1912
Family Seletonellaceae Korde, 1950, emend. Berger & Kaever, 1992
Tribe unknown
Genus Teutloporella Pia, 1912
5.7.1. Teutloporella herculea (Stoppani, 1857), Pia, 1912
Pl. 4 ![]() , fig. 4
, fig. 4
1857 - Gastrochaena herculea n. sp. - Stoppani, p. 376-377, no figure.
1858 - Gastrochaena herculea - Stoppani, Pl. 16 ![]() , figs. 11-12.
, figs. 11-12.
1912 - Teutloporella herculea n. comb. - Pia, Pl. II, fig. 27; Pl. III, figs. 1-2.
1965 (nom. nud.) - Physoporella likana n. sp. - Herak, p. 20-21, Pl. XIV, figs. 1-3.
To this species, as a junior synonym, should be ascribed one described by Herak (1965, p. 20-21) as Physoporella likana: "The thallus is cylindrical but probably not very elongated; the main stem is relatively large in comparison to the tiny branches and calcareous wall. The shape of the pores is piriferous, generally closed at the distal end. There are traces which show that some of them penetrate through the calcareous wall. There are no visible indications of forming clear whorls."
A typical locality is near Sv. Rok in Lika, Croatia. It was found together with Teutloporella herculea and Macroporella sp. All illustrated specimens (Herak, 1965, Pl. XIV, figs. 1-3) come from the thin-section Alg.86, which is kept in the Geological and Paleontological Institute of the Faculty of Science in the collection of Prof. Herak. Since the author originally defined the syntypes, Herak (in Granier & Deloffre, 1995) chooses among them the specimen from Fig. 2 (= this paper, Pl. 4 ![]() , fig. 4) as a lectotype.
, fig. 4) as a lectotype.
The illustrated specimens, according to their aspondyl arranged and numerous typical trichoform (not piriform) laterals undoubtedly belong to the Ladinian species Teutloporella herculea (Stoppani).
At the end of this chapter, I would like to remind readers that the species Oligoporella duplicata Pia, 1920, was transferred to the genus Poikiloporella by Pia (1943), as its type-species.
I would also like to mention here the erroneous determination of the Early Cretaceous species Acroporella radoicicae Praturlon as Oligoporella radoicicae in Raviv and Lorch (1992, p. 36, 38), and which Bucur et al. (2000, p. 450, 454; see also Bucur et al., 2021) consider it to be a synonym of Similiclypeina aff. somalica.
5.8. Species from the Permian of East Asia
Algae from the studied genera appeared already in the early Permian, and they were described mainly by the Japanese author Riuji Endo, who investigated Dasycladalean algae thoroughly during his career, and in his 21 works established as many as 9 new genera and 92 new species of Dasycladalean algae. Among them, 22 species are from the genera Physoporella and Oligoporella. Most of the algae are from Japan, and a few are from Thailand. In addition to Endo, the Permian algae of Japan were described by Ishijima et al. (1971), establishing as many as, 18 new species, of which only two were found once more, by the same author (Nakamura, 1981). Only one Permian species Physoporella longipora (Endo), n. comb., nom. nov. has been later found by another author outside Asia (Fl�gel, 1990). The species mentioned, and especially those described by Endo, are based mainly on poorly preserved and recrystallized material, so it is rarely possible to reconstruct the actual structure of the algae. In addition, the author has often united very heterogeneous material within one species, and especially within the genus. An illustrative example is the species Physoporella classiforata Endo, 1969, whose "holotype" (i.e., one of the syntypes; Endo, 1969, Pl. XXIV, fig. 7) is almost identical to the specimen defined by the author in the same paper as Gyroporella microporosa (Endo, 1969, Pl. XIX, fig. 4). Many species represent nomina nuda because no holotype has been named or multiple specimens (syntypes) have been named. As will be seen below, Endo's diagnoses of the species are very similar to each other, many of them almost identical, differing only in whether the pores have rounded ends (closed) or narrow outward passages (open), and whether they are vertical on an axis or oblique.
Due to all the above, it is not possible to give a more detailed description of the mentioned species or a new diagnosis. For a complete revision of Endo taxa, it is necessary to review the type material again. For now, based on his illustrations, some species can be safely excluded from the genera Physoporella and Oligoporella, while some can be more or less safely left in one of these two genera. Some species probably had vesiculifer laterals, so they can be included in the genus Gyroporella, while some possibly belongs to the genus Mizzia, as the specimens shown probably represent moniliform thallus articles rather than the very short thallus as described by Endo.
Due to the importance of the appearance of the studied genera in the Permian, I still consider it necessary to give a brief overview of them.
Genus Physoporella Steinmann, 1903, emend.
Physoporella chichibuensis Ishijima et al., 1971
Fig. 23 ![]() /5
/5
1971 - Physoporella chichibuensis n. sp. - Ishijima et al., p. 126, Pl. 14, figs. 1-5.
Original diagnosis: "Thallus is short, broadly cylindrical, rather ovate and slightly undulating. Central stem is relatively broad. Pores are given off from the central stem at relatively broad basements and expand to the broadest width at midway, then diminish to the pointed terminations, and they are closed in the calcareous body and arranged in regularly spaced whorls."
Discussion: Typical Physoporella, very similar to the type species P. pauciforata both in the typical piriform shape of the laterals and in slight external undulation (Fig. 23 ![]() /5, holotype). The inner wall of the thallus is smooth. The illustrated specimens are quite small, so the possibility that it has a moniliform thallus is not completely ruled out, but it is unlikely.
/5, holotype). The inner wall of the thallus is smooth. The illustrated specimens are quite small, so the possibility that it has a moniliform thallus is not completely ruled out, but it is unlikely.
The species is described from the "lower" Permian (? Asselian -? Kungurian) of the island of Honsh� in Japan.
Physoporella kitakamiensis Endo, 1956, emend.
Fig. 23 ![]() /8
/8
1956 - Physoporella kitakamiensis n. sp. - Endo, p. 236, Pl. 30, figs. 5-6; Pl. 31, figs. 1-4.
1957 - Physoporella kitakamiensis - Endo & Horiguchi, p. 174-175, Pl. XIII, fig. 1.
Original diagnosis: "The thallus is rather short and broadly cylindrical. The longest specimen is estimated to be 6.8 mm, tapering gradually towards the rounded end. Pores broadly rounded, usually closing to the exterior. The pores are arranged in closely and regularly spaced whorls, about, 20 pores in a whorl at the middle part. They appear to have developed approximately perpendicular to the main stem or slightly ascending. No reproductive organs were found."
Discussion: The material, although only having a small number of specimens, is relatively well preserved with several oblique and longitudinal-oblique cross-sections, which allows insight into the structure of this alga. The thallus is cylindrical with piriform laterals arranged in whorls perpendicular or slightly oblique to the thallus axis. The whorls are relatively dense, but do not touch each other. The laterals in adjacent whorls are placed alternately. Since the author named several specimens as syntypes (Endo, 1956, Pl. 30, fig. 6; Pl. 31, figs. 1-4), I determine the specimen from Endo (1956, Pl. 31, fig. 3 middle, left = this work, Fig. 23 ![]() /8) as a lectotype, since the characteristics of the mentioned species are best seen on it. Thin-section number is 212 or 213. It is possibly kept in Department of Earth Sciences, Saitama University, Urawa, Japan. The species was first described in the Lower Permian (Artinskian-Kungurian) of
Honsh� Island, Japan (Endo, 1956), and was found on the same island once again (Endo & Horiguchi, 1957), also in the Lower Permian (? Asselian).
/8) as a lectotype, since the characteristics of the mentioned species are best seen on it. Thin-section number is 212 or 213. It is possibly kept in Department of Earth Sciences, Saitama University, Urawa, Japan. The species was first described in the Lower Permian (Artinskian-Kungurian) of
Honsh� Island, Japan (Endo, 1956), and was found on the same island once again (Endo & Horiguchi, 1957), also in the Lower Permian (? Asselian).
Physoporella konishii Endo, 1959, emend.
Fig. 23 ![]() /6, 10
/6, 10
1959 (nom. nud.) - Physoporella konishii n. sp. - Endo, p. 195, Pl. 42, fig. 4; Pl. 42, fig. 5 centre (= Pl. 41, fig. 7 centre).
Original diagnosis: "The thallus is cylindrical and moderately undulating. It shows also slight innerannulation. The pores are given off from the central stem as rather broad expansions which gradually diminish into rounded terminations toward the exterior. The pores are often nearly perpendicular to the exterior or slightly ascending. No reproductive organs are found."
Discussion: Relatively well-preserved longitudinal-oblique and cross-section. The thallus is relatively large with pronounced undulation, and less pronounced intusannulation as in P. pauciforata. The laterals are slightly thinner, piriform, arranged in whorls and perpendicular to the longitudinal axis. Laterals touch each other in the same whorl. The whorls are close, but do not touch each other.
When establishing the species, the author named two "holotypes" (Endo, 1959, p. 195, Pl. 42, figs. 4-5 = this work, Figs. 23/10, 6), which is a nomen nudum (Art. 8.5. and 40.1 of ICN, Turland et al., 2018). In order for the species to become valid, I name the specimen in Endo (1959, Pl. 42, fig. 4 = this work, Fig. 23 ![]() /10) as a lectotype.
/10) as a lectotype.
The species is described from the Lower Permian (? Asselian) island of Honsh� in Japan.
Physoporella lativentrusa Endo, 1959, emend.
Fig. 23 ![]() /1
/1
1959 (nom. nud.) - Physoporella lativentrusa n. sp. - Endo, p. 196, Pl. 42, figs. 8-9.
1971 (nom. nud.) - Physoporella lativentrusa - Ishijima et al., p. 125, Pl. 13 ![]() , fig. 4.
, fig. 4.
Original diagnosis: "The thallus is broadly cylindrical and slightly undulating. The pores are given off from the central stem as relatively broad expansions which gradually diminish into rounded terminations toward the exterior. The pores are often perpendicular to the exterior or slightly ascending."
Discussion: On a well-preserved oblique-cross section (Endo, 1959, Pl. 42, fig. 8 = this paper, Fig. 23 ![]() /1) one can see somewhat more rounded piriform laterals arranged in dense whorls, perpendicular on the thallus axis. Each whorl carries, 18 laterals. The thallus is cylindrical with a smooth inner wall, while the outer wall is very slightly undulated.
/1) one can see somewhat more rounded piriform laterals arranged in dense whorls, perpendicular on the thallus axis. Each whorl carries, 18 laterals. The thallus is cylindrical with a smooth inner wall, while the outer wall is very slightly undulated.
The author names two holotypes (Endo, 1959, p. 196, Pl. 42, figs. 8-9), which represents a nomen nudum (Art. 8.5. and 40.1 of ICN, Turland et al., 1907), and therefore, in order for the species to become valid, I name the specimen in Endo (1959, Pl. 42, fig. 8 = this paper, Fig. 23 ![]() /1) from thin-section no. 611 as a lectotype.
/1) from thin-section no. 611 as a lectotype.
The species has been described from Lower-Middle Permian deposits from the island of Honsh� in Japan (Endo, 1959), and has also been described by Ishijima et al. (1971) from the "Lower" Permian.
Physoporella longipora (Endo, 1969), n. comb., nom. nov.
(= Oligoporella elegans Endo, 1969, nom. nud.)
Fig. 23 ![]() /2, 9
/2, 9
1969 (nom. nud.) - Oligoporella elegans n. sp. - Endo, p. 73, Pl. XXXIV, figs. 1-5.
1990 (nom. nud.) Teutloporella ? sp. (cf. Oligoporella elegans) - Fl�gel, p. 112, Pl. 3, fig. 1 pars.
Original diagnosis: "The thallus is long, cylindrical and slightly undulating. The longest specimen on hand is estimated to be 3.5 mm. The pores are remarkably long; they are given off from the central stem at its rather large, round basements and gradually diminish in width into slender passages toward the exterior. The pores ascend moderately toward the exterior; the angles between the pores and central stem are estimated to be 30� to 80�. The pores form fine regularly spaced whorls and the length of each whorl is estimated to be 0.365 to 0.5333 mm."
Discussion: Relatively well-preserved material. The holotype is from thin-section no. 13077 (Endo, 1969, Pl. XXXIV, fig. 1 = this paper, Fig. 23 ![]() /9). The thallus is cylindrical and usually annulated by oblique grooves between the whorls (Fig. 23
/9). The thallus is cylindrical and usually annulated by oblique grooves between the whorls (Fig. 23 ![]() /2). The inner wall of the thallus is smooth and calcification reaches close to the stem cell. The laterals are long piriform, distinctly oblique and very slightly concave bent. The laterals calcified along their entire length (Endo, 1969, Pl. XXXIV, fig. 5) or the top parts remained uncalcified (Endo, 1969, Pl. XXXIV, figs. 2, 4). The number of laterals in the whorl is 12-14.
/2). The inner wall of the thallus is smooth and calcification reaches close to the stem cell. The laterals are long piriform, distinctly oblique and very slightly concave bent. The laterals calcified along their entire length (Endo, 1969, Pl. XXXIV, fig. 5) or the top parts remained uncalcified (Endo, 1969, Pl. XXXIV, figs. 2, 4). The number of laterals in the whorl is 12-14.
The name of the species Oligoporella elegans Endo, 1969 is nomen nudum (Art. 53.1. ICN; Turland et al., 1907), as the later homonym of the species Oligoporella elegans Assmann ex Pia, 1931, having the same name based on another holotype. In addition, due to the arrangement of the piriform laterals in single-rowed whorls, this species belongs to the genus Physoporella. Therefore, I give this species a new name: Physoporella longipora Endo, 1969 n. comb., nom. nov., because it has long pores (Latin "longus" = long).
The species was first described from the Middle Permian (Kuberghandian) Rat Buri limestone from Thailand (Endo, 1969). It was also illustrated by Fl�gel (1990) from the Lower Permian (Bolorien) of Anatolia in Turkey as Teutloporella? sp. (cf. Oligoporella elegans). Although somewhat less well preserved and with slightly thinner laterals, it is the same species.
Physoporella nipponica (Endo, 1956), n. comb.
(= Oligoporella nipponica Endo, 1956)
Fig. 23 ![]() /3
/3
1956 - Oligoporella nipponica n. sp. - Endo, p. 235-236, Pl. 24, figs. 10-12; Pl. 25, figs. 1-2.
Original diagnosis: "Thallus broadly long, cylindrical and slightly undulating; circular in cross section. Pores start from the central axis as rather broad expansions which gradually diminish into slender passages toward the exterior. Pores are arranged perpendicularly to both inner and outer surfaces of the calcareous wall; however, they sometimes climb slightly outwards."
Discussion: A well-preserved specimen in Endo (1956, Pl. 24, fig. 11 = this paper, Fig. 23 ![]() /3) shows typical piriform laterals, positioned perpendicular to the thallus axis. The whorls are spaced, and the laterals within the whorl are quite close to each other, although they rarely touch. One whorl has about 14 laterals. The thallus is slightly undulated on the outside and very slightly intusannulated on the inside, which are also characteristics of the genus Physoporella, so I rename the species Physoporella nipponica (Endo, 1956), n. comb. I name the specimen in Endo (1956, Pl. 24, fig. 11 = this paper, Fig. 23
/3) shows typical piriform laterals, positioned perpendicular to the thallus axis. The whorls are spaced, and the laterals within the whorl are quite close to each other, although they rarely touch. One whorl has about 14 laterals. The thallus is slightly undulated on the outside and very slightly intusannulated on the inside, which are also characteristics of the genus Physoporella, so I rename the species Physoporella nipponica (Endo, 1956), n. comb. I name the specimen in Endo (1956, Pl. 24, fig. 11 = this paper, Fig. 23 ![]() /3) from thin-section no. 302 as a lectotype. The species was found in the Artinskian (Lower Permian) of Honsh� Island, Japan.
/3) from thin-section no. 302 as a lectotype. The species was found in the Artinskian (Lower Permian) of Honsh� Island, Japan.
Physoporella nyugawensis Endo, 1959
Fig. 23 ![]() /11
/11
1959 - Physoporella nyugawensis n. sp. - Endo, p. 193-194, Pl. 35, fig. 9.
Original diagnosis: "The thallus is short, and broadly cylindrical. The pores start from the central stem at ball-like expansions which rather slightly diminish toward the exterior. From the fact that the expanded part of the pore contains well-defined spores it is presumed that this part functioned as a sporangium."
Discussion: The holotype (Endo, 1959, Pl. 35, fig. 9 = this paper, Fig. 23 ![]() /11) from thin-section no. 21126 is a somewhat less well-preserved oblique section, but with a visible whorls of densely distributed piriform laterals. Grooves appear on the outer surface of the thallus that probably represents calcified trichoblasts. The spores mentioned by the author are not visible, but only irregular micrite particles that each limestone thin-section has. Due to the existence of only one specimen, we cannot be completely sure of the generic affiliation of this species. The two cross-sections (Endo, 1959, Pl. 35, fig. 10; Pl. 40, fig. 2) have vesicular laterals, so they do not belong to Physoporella.
/11) from thin-section no. 21126 is a somewhat less well-preserved oblique section, but with a visible whorls of densely distributed piriform laterals. Grooves appear on the outer surface of the thallus that probably represents calcified trichoblasts. The spores mentioned by the author are not visible, but only irregular micrite particles that each limestone thin-section has. Due to the existence of only one specimen, we cannot be completely sure of the generic affiliation of this species. The two cross-sections (Endo, 1959, Pl. 35, fig. 10; Pl. 40, fig. 2) have vesicular laterals, so they do not belong to Physoporella.
The only specimen of this species was found in the Permian deposits of the island of Honsh� in Japan, presumably of the early middle Permian range.
The following are species whose affiliation to the genus Physoporella is questionable.
Physoporella? hidaensis (Endo in Endo & Kanuma, 1954), n. comb.
(= Oligoporella hidaensis Endo in Endo & Kanuma, 1954)
1954 - Oligoporella hidaensis n. sp. - Endo in Endo & Kanuma, p. 198-199, Pl. XV, figs. 4-6.
Original diagnosis: "Thallus is relatively short, and cylindrical pores are given off from the central stem as egg-like expansions which gradually diminish into very slender passages toward the exterior. Pores are arranged as definite whorls and they are slightly ascending toward the exterior. As will be seen in Pl. III, fig. 5, pores may sometimes cluster."
Discussion: The poorly preserved material has the shape of laterals not clearly discernible, only their arrangement in whorls. Due to the absence of double whorls, I rename this species Physoporella? hidaensis (Endo and Endo & Kanuma, 1954), n. comb.
The species is described from the early Permian (? Sakmarian -? Artinskian) of Honsh� Island, Japan.
Physoporella? japonica Endo, 1957
1957 - Physoporella japonica n. sp. - Endo, p. 291, Pl. 40, figs. 7-9; Pl. 40, figs. 10-11; Pl. 41, fig. 1.
1959 - Physoporella japonica - Endo, p. 194, Pl. 36, figs. 1-4.
1961b - Physoporella japonica - Endo, p. 131, Pl. 5, figs. 3-4
Original diagnosis: "Thallus is relatively short and broadly cylindrical. The longest specimen at hand is estimated to be 4.5 mm, tapering gradually to a rounded end. Pores given off from the stem cell at relatively narrow basement which gradually expand into its broadest width at the middle part and narrow again towards exterior as rounded terminations; they usually closed in the calcareous body. Pores appear to have been developed approximately perpendicular to the main stem in the middle portion. However, they gradually become inclined toward the upper and lower ends of the thallus. The pores are arranged in regularly spaced whorls, about, 20 in a whorl in the middle part. The central stem is relatively wide in proportion to the entire thallus."
Discussion: The presented material is poorly preserved and quite heterogeneous. Specimens from Endo (1957, Pl. 40, fig. 9) have piriform laterals; specimens from Pl. 40, figs. 7-8 are questionable, while the specimen from Pl. 40, figs. 10-11; Pl. 41, fig. 1 have vesiculifer laterals and does not belong to the genus Physoporella.
The species was first described from the "Lower" Permian (? Artinskian - ? Kungurian) deposits of the Honsh� Island, Japan (Endo, 1957), and was identified in the Lower and Middle Permian (? Artinskian -? Murghabian) by Endo (1959), and in the Lower Permian (? Asselian) by Endo (1961b). Specimens from Ishijima et al. (1971) certainly do not belong to the genus Physoporella having relatively large oval laterals.
Physoporella? kanmerai Endo, 1969
1969 - Physoporella kanmerai n. sp. - Endo, p. 64, Pl. XXVI, figs. 6-10; Pl. XXVII, figs. 1-3.
Original diagnosis: "The longest specimen on hand is estimated to be 3.6 mm long. The thallus is broadly cylindrical and slightly undulating. The pores are given off from the stem cell at the broader basements and diminish gradually in width toward the rounded ends. They are usually enclosed in the calcareous body. The pores appear to have developed ascending diagonally toward the exterior. One series of complete pores can be seen in the inner side and another series of incomplete open pores in the outer part in cross - section, as in Pl. 27 (not 23 as in Endo), fig. 2."
Discussion: In this case, too, the material is recrystallized and heterogeneous. The specimens from Endo (1969, Pl. XXVI, fig. 6 (holotype), 8, Pl. XXVII, fig. 1) possibly belongs to the genus Physoporella having piriform laterals and an aspondyl arrangement. Some cross-sections show a very small thallus (Endo, 1969, Pl. XXVI, figs. 3-5).
The species is described from Rat Buri limestones of Middle Permian age (? Kuberghandian -? Murghabian) from Thailand.
Physoporella? ovata Endo, 1961a
1961a - Physoporella ovata n. sp. - Endo, p. 99, Pl. 15 ![]() , fig. 1; Pl. 16
, fig. 1; Pl. 16 ![]() , fig. 1; Pl. 17
, fig. 1; Pl. 17 ![]() , figs. 5-6.
, figs. 5-6.
1969 - Physoporella ovata - Endo, p. 65, Pl. XXIV, figs. 8-10; Pl. XXV, figs. 7-9.
1971 - Physoporella ovata - Ishijima et al., p. 125-126, Pl. 13, figs. 2-3.
Original diagnosis: "Thallus relatively short and broadly cylindrical. The rays given off from the stem cell at relatively narrow basements which expand into broadest width at the middle parts and narrow again toward exterior as rounded terminations; they usually closed in the calcareous body. The rays appear to have been developed approximately perpendicular to the main stem in the middle portion and gradually become inclined towards the upper and lower ends of thallus. The central stem is much wider in proportion to the entire thallus."
Discussion: Poorly preserved material. The laterals are possibly piriform, laid perpendicular to the longitudinal axis. They are arranged, most likely, in dense whorls.
The species was first described from the deposits of the Honsh� Island (Japan) in the Lower - Middle Permian (Endo, 1961a), and is also mentioned by Endo (1969) from the Middle Permian (? Kuberghandian) in Thailand, and by Ishijima et al. (1971) from the "Lower" Permian, also from Honsh� Island.
Physoporella? skawadai (Endo, 1952), n. comb.
(= Oligoporella s-kawadai Endo, 1952)
1952 - Oligoporella s-kawadai n. sp. - Endo, p. 245-246, Pl. 23, figs. 9-10
1956 - Oligoporella s-kawadai - Endo, p. 234-235, Pl. 24, figs. 5-9.
Original diagnosis: "Thallus 6 mm long, cylindrical; nearly circular in cross-section. Pores given off from the central stem as ball-like expansions which gradually diminish into very slender passages toward the exterior. Pores are arranged perpendicularly to both the inner and outer surfaces of the calcareous wall; however, they are sometimes slightly ascending towards exterior. Pores are arranged as definite whorls. The basal, spherical part of pores seems to be a sporangia."
Discussion: A relatively well-preserved specimen is in Endo (1952, Pl. 23, fig. 9), which shows the unusual structure of this alga. The laterals are not typically piriform, but look like a laboratory flask with a spherical base and a narrow neck, so placement in the genus Physoporella is questionable. Their euspondyl arrangement is clearly visible. Morphologically it is quite reminiscent of the genus Acicullela, but the later has only round cavities, which represent gametangia, but no laterals. Other illustrated material (Endo, 1952, Pl. 23, fig. 10; 1956; 1961a) does not certainly belong to the same species as this specific shape of laterals is not visible.
Since the author named both illustrated specimens as "holotypes" (i.e., syntypes), I designate the above-mentioned specimen from Endo (1952, Pl. 23, fig. 9) as the lectotype and simplify the species name (deleting the hyphen) in skawadai (Granier & Deloffre, 1995, p. 70).
The species is described from the "middle" Permian of Honsh� Island in Japan (Endo, 1952), and was registered on the same island in Lower Permian (Artinian) deposits (Endo, 1956).
Genus Oligoporella Pia, 1912, emend.
Only one species from the Japanese Permian can be attributed to the genus Oligoporella:
Oligoporella fluctuosa (Endo, 1958), n. comb.
(= Likanella fluctuosa (Endo, 1958), Vachard, 1980, nom. nud.; = Physoporella? fluctuosa (Endo, 1958), Kochansky & Herak, 1960, nom. nud.; = Clavaphysoporella fluctuosa Endo, 1958, nom. nud.)
Fig. 23 ![]() /4, 7
/4, 7
1958 (nom. nud.) - Clavaphysoporella fluctuosa n. sp. - Endo, p. 267-268, Pl. 39, figs. 8-9.
1959 (nom. nud.) - Clavaphysoporella fluctuosa - Endo, p. 198-199, Pl. 38, fig. 12.
1963 (nom. nud.) - Physoporella ? fluctuosa n. comb. - Kochansky & Herak, p. 86-87, no figure.
1980 (nom. nud.) Likanella fluctuosa n. comb. - Vachard, p. 363, no figure.
Original diagnosis: "The thallus long, broadly cylindrical and consisting of very fine outer annulations as well as slight innerannulations. The outer annulations show wide and deep furrows which sometimes reach the central stem. The pores are 0.675 mm long and given off from the central stem at the slightly broader basement which gradually diminish into rather pointed terminations. The pores are moderately ascending toward the exterior and constitute a fine cluster. One whorl of clustering pores may be contained in each annulation. No reproductive organs described."
Discussion: As already described in Chapter 3. Endo (1958, p. 265-266) "revises" the genus Physoporella by extracting the species P. minutula and naming a new genus Clavaphysoporella, characterized by phloioform pores arranged in whorls, although the said species does not show these characteristics. He also includes C. fluctuosa in the newly established genus. Kochansky and Herak (1960, p. 86) nullify the "revision" of the genus Physoporella and the establishment of a new genus Clavaphysoporella, and combine the forms with phloioform laterals arranged in the diverging whorls into a new genus Clavaporella, and the species P. minutula and "probably" C. fluctuosa are considered to belong to the genus Physoporella. Vachard (1980, p. 363) transfers this species to the genus Likanella without any explanation. The genus Likanella is characterized by specific spindle-shaped (fusiform) laterals arranged in triple whorls, and the mentioned species certainly does not belong to this genus in terms of its structure.
The thallus is cylindrical, markedly annulated with oblique furrows, rounded at the base. The annulation is the result of calcification of the whorls which are constructed of two very close rows of oblique piriform laterals (Fig. 23 ![]() /4). The laterals are distinctly inflated at the base and gradually thin distally. They are relatively long, and the inclination towards the longitudinal axis is about 60�. Two rows of laterals, i.e., double whorls are clearly visible in longitudinal section (Endo, 1958, Pl. 39, fig. 8; = this, work, Fig. 23
/4). The laterals are distinctly inflated at the base and gradually thin distally. They are relatively long, and the inclination towards the longitudinal axis is about 60�. Two rows of laterals, i.e., double whorls are clearly visible in longitudinal section (Endo, 1958, Pl. 39, fig. 8; = this, work, Fig. 23 ![]() /4), and also in cross-section (Endo, 1958, Pl. 39, fig. 9; = this, work, Fig. 23
/4), and also in cross-section (Endo, 1958, Pl. 39, fig. 9; = this, work, Fig. 23 ![]() /7) where one can see alternately arranged laterals of the lower and upper row. In this section, as well as in the upper part of Fig. 23
/7) where one can see alternately arranged laterals of the lower and upper row. In this section, as well as in the upper part of Fig. 23 ![]() /4, it is clear that there is no possibility of clusters of laterals, i.e., metaspondyl arrangement. The inner wall of the thallus is smooth, and the calcification reached the very base of the laterals, i.e., the stem cell. The intusannulation reported by Endo (1958) is visible only on the left side of the specimen, and may be some kind of deformation.
/4, it is clear that there is no possibility of clusters of laterals, i.e., metaspondyl arrangement. The inner wall of the thallus is smooth, and the calcification reached the very base of the laterals, i.e., the stem cell. The intusannulation reported by Endo (1958) is visible only on the left side of the specimen, and may be some kind of deformation.
According to its structure (double whorl of piriform laterals) the species certainly belongs to the genus Oligoporella, so it gets a new name Oligoporella fluctuosa (Endo, 1958), n. comb. Given that the author named two "holotypes", i.e., syntypes (Endo, 1958, Pl. 39, figs. 8-9), the species is nomen nudum. In order for the species to become valid, I name the specimen from thin-section no. 21044 (Endo, 1958, Pl. 39, fig. 8; = this work, Fig. 23 ![]() /4) as a lectotype.
/4) as a lectotype.
The species is described from the lower Permian (? Asselian) of Honsh� Island in Japan, and is illustrated from the same locality by Endo (1959).
|
|
Figure 23: Species from the Permian of East Asia: 1) Physoporella lativentrusa Endo, emend., from Endo (1959, Pl. 42, fig. 8); 2, 9) Physoporella longipora (Endo), n. comb., nom. nov., from Endo (1969, Pl. XXXIV, figs. 2, 1); 3) Physoporella nipponica (Endo), n. comb., from Endo (1956, Pl. 24, fig. 11); 4, 7) Oligoporella fluctuosa (Endo), n. comb., from Endo (1958, Pl. 39, figs. 8-9); 5) Physoporella chichibuensis Ishijima et al., from Ishijima et al. (1971, Pl. 14, fig. 1); 6, 10) Physoporella konishii Endo, emend., from Endo (1959, Pl. 42, figs. 5 middle, 4); 8) Physoporella kitakamiensis Endo, emend., from Endo (1956, Pl. 31, fig. 3); 11) Physoporella nyugawensis Endo, from Endo (1959, Pl. 35, fig. 9). Magnification x25, except fig. 8 (x
10). All scale bars = 1 mm. |
Genus Teutloporella Pia, 1912
Teutloporella? himurensis (Ishijima et al., 1971), n. comb.
(= Oligoporella himurensis Ishijima et al., 1971)
1971 - Oligoporella himurensis n. sp. - Ishijima et al., p. 123-124, Pl. 7, fig. 5; Pl. 12, fig. 1.A.
Original diagnosis: "The thallus is large, broadly cylindrical, round in cross section. The largest specimen available is about 11 mm long and 5.6 mm wide. The central tree is particularly wide and the calcareous wall is very thin, more precisely the tree is 13 times pores are long and thin, curved downwards, they protrude from the central tree at round bases and gradually thin and curve into pointed ends outwards and are usually open towards the surface of the calcareous wall. The pores are arranged diagonally climbing outwards and they form properly spaced whorls. The basal spherical parts of the pores are possible sporangia."
Discussion: As the authors state in the diagnosis, the central cavity is extremely wide and the calcareous envelope is very thin. The inner surface of the wall is uneven, suggesting that it has not calcified all the way to the stem cell, so it is possible that the proximal parts of the laterals, and perhaps even lower-order laterals, are not visible. The laterals are narrow and pointed at the top, i.e., of the trichoform rather than the piriform type. They are bent upwards, that is, towards the sunlight, and not downwards, as the authors state, and they are densely distributed. The illustrated cross-sections (Ishijima et al., 1971, Pl. 7, fig. 5; Pl. 12, fig. 1A), however, do not prove the presence of whorls, so I rename this alga Teutloporella? himurensis (Ishijima et al., 1971), n. comb., because of the aspondylly arranged trichoform laterals.
The species is described from the "Lower" Permian island of Honsh� in Japan.
Genus Gyroporella G�mbel, 1872, emend. Benecke, 1876
The following are species, also based on very poor material, which certainly do not have piriform laterals, and therefore do not belong to the genera Physoporella or Oligoporella. In general, vesiculifer laterals are recognizable in slightly better preserved specimens, so all these algae are included in the genus Gyroporella (with a question mark). Judging by the very small thallus, some of them could belong to the genus Mizzia, but for any conclusion it is necessary to study the type material. Their descriptions are also almost identical. These species are therefore only listed:
Gyroporella? cauliculus (Endo, 1961a, nom. nud.), n. comb.
[= Oligoporella cauliculus Endo, 1961a, nom. nud. (syntypes)]
Gyroporella? classiforata (Endo, 1969, nom. nud.), n. comb.
[= Physoporella classiforata Endo, 1969, nom. nud. (two holotypes designated)]
Gyroporella? expansa Endo, 1959, nom. nud., n. comb.
[= Oligoporella expansa Endo, 1959, nom. nud. (syntypes)]
Gyroporella? horiguchii (Endo, 1957), n. comb.
(= Oligoporella horiguchii Endo, 1957)
Gyroporella? isomii (Endo, 1961a), n. comb.
(= Oligoporella isomii Endo, 1961a)
Gyroporella? omiensis (Endo, 1952), n. comb.
(= Oligoporella �miensis Endo, 1952)
Gyroporella? propilosa (Endo, 1969), n. comb.
(= Oligoporella propilosa Endo, 1969)
Gyroporella? rotunda (Endo, 1969, nom. nud.), n. comb.
[= Physoporella? rotunda Endo, 1969, nom. nud. (type not designated)]
Gyroporella? tenuiaxis (Endo, 1959, nom. nud.), n. comb.
[= Physoporella tenuiaxis Endo, 1959 (syntypes)]
I would like to mention here that Physoporella sp. described by Johnson and Danner (1966, p. 430-431, Pl. 56, fig. 5) from the Permian Porcupine Creek, N Vancouver, British Columbia (Canada) does not belong to this genus, nor does it belong to any Dasycladalean algae.
The species Oligoporella sola Endo, 1969, from the middle Permian of Thailand was transferred by Vachard (1980, p. 354) to the genus Uragiellopsis due to the characteristic claviform shape of the large laterals.
5.9. Key to identification of investigated taxa
To facilitate the identification of the investigated taxa, a key for recognizing genera, species and varieties is attached (Table 2). The species Acroporella? prisca (Pia) and Physoporella? elegans (Pia, 1912), n. comb., are not included due to their uncertain systematic position (see Chapter 5.6.). Species with uncertain generic attribution (with question mark) and species from the Permian of East Asia (see Chapter 5.8.) are also not included.
Table 2: Key for the identification of the investigated taxa.
| Subtribe Oligoporellinae Bassoullet et al., 1979 Cylindrical thallus with euspondyl laterals of the first order expanded in the proximal part and thinning distally (i.e., trichoform or piriform type). | ||
| 1. | a) Laterals only slightly expanded in the proximal part (trichoform type) | Euteutloporella |
| b) Laterals proximally thin, extended in the middle part and distally tapper | Ardeiporella | |
| Ard. karrerioidea | ||
| c) Laterals markedly expanded in the proximal part | → 2 | |
| 2. | a) Laterals expanded in the proximal part, with gradual thinning distally (piriform type) | → 3 |
| b) Laterals with asymmetrically expanded proximal part | Uragiella | |
| c) Laterals expanded in the proximal part, thinned in the central part, and spherically expanded in the distal part | Poikiloporella | |
| 3. | a) Typical piriform laterals arranged in single whorls | Physoporella |
| b) Short and rounded piriform laterals, arranged in dense single whorls | Neophysoporella | |
| c) Typical piriform laterals arranged in double whorls | Oligoporella | |
| Physoporella Steinmann, 1903, emend. Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in single whorls. | ||
| 1. | Piriform laterals arranged in close or spaced single whorls | Phys. pauciforata |
| a) Laterals horizontal or slightly oblique, whorls spaced, stronger calcification | var. pauciforata | |
| b) Laterals oblique, whorls spaced, stronger calcification | var. sulcata | |
| c) Laterals horizontal, whorls spaced, weaker calcification | var. undulata | |
| d) Laterals horizontal, whorls close, weaker calcification | var. gemerica | |
| 2. | Large piriform laterals arranged in compacted single whorls | Phys. croatica |
| 3. | Piriform laterals arranged in spaced whorls and irregularly arranged in groups of two or three | Phys. leptotheca |
| Oligoporella Pia, 1912, emend. Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in double whorls. Each double whorl consists of two very close rows of laterals. | ||
| 1. | Piriform laterals arranged in close or spaced double whorls, slightly oblique and slightly divergent | Olig. pilosa |
| a) Whorls spaced apart, weaker calcification | var. pilosa | |
| b) Whorls closer, stronger calcification | var. intusannulata | |
| 2. | Piriform laterals arranged in spaced double whorls, divergent and slightly bent | |
| a) Both rows of laterals equally divergent | Olig. cornuta | |
| b) The upper row of laterals is more divergent than the lower, protrusions on the laterals | Olig. intusannulata | |
| 3. | Piriform laterals arranged in spaced double whorls, markedly divergent and bent | Olig. varicans |
| 4. | Piriform laterals with less pronounced proximal expansion and distal narrowing, arranged in spaced double whorls, oblique and slightly divergent | Olig. minutula |
| 5. | Piriform laterals with less pronounced distal narrowing, arranged in dense double whorls, horizontal to slightly oblique and slightly divergent | Olig. praealpina |
| a) Slightly wider laterals, more evenly distributed | var. praealpina | |
| b) Slightly narrower laterals, more irregularly distributed | var. bosniensis | |
| 6. | Piriform laterals with more pronounced distal narrowing, arranged in spaced double whorls, horizontal to slightly oblique and slightly divergent; calcification stronger at the level of the whorls | Olig. dissita |
| 7. | Piriform laterals, slightly elongated, with a short basal peduncle, arranged in clearly spaced double whorls, horizontal and slightly divergent; calcification by a thin micritic sheath | Olig. laevis |
| Neophysoporella n. gen. Cylindrical thallus with a relatively thin calcareous sheath. Laterals of the piriform type, but relatively shorter, more rounded and with less pronounced thinning towards the distal end, arranged in relatively dense whorls. | ||
| 1. | Distinctly uneven calcification, variable central cavity, slightly larger thallus | Neophys. lotharingica |
| 2. | More regular calcification, slightly smaller thallus | Neophys. jomdaensis |
| 3. | Calcareous wall with tinny pores | Neophys. zamparelliae |
The microfacies of the investigated material can be grouped in two basic types: with micritic and with sparite matrix. Microfacies with a micritic matrix are represented mainly by skeletal packstones or biomicrites (Pls. 5 ![]() - 6
- 6 ![]() ,
11
,
11 ![]() -
12
-
12 ![]() ). Skeletal particles are exclusively fragments and thalluses of Dasycladalean algae, and of other allochemes only tiny peloids occur. These microfacies indicate protected lagoon environments which are also the most common habitat of living Dasycladales (Berger & Kaever, 1992). Part of the microfacies from this group is represented by poorly washed biomicrites (Pl. 3
). Skeletal particles are exclusively fragments and thalluses of Dasycladalean algae, and of other allochemes only tiny peloids occur. These microfacies indicate protected lagoon environments which are also the most common habitat of living Dasycladales (Berger & Kaever, 1992). Part of the microfacies from this group is represented by poorly washed biomicrites (Pl. 3 ![]() ) with sparite cement precipitated beside the micrites. Sparite cement also precipitated in thallus cavities of algae that were not filled with micritic sediment. The sparite matrix microfacies is represented mainly by skeletal grainstones or biosparites (Pls. 7
) with sparite cement precipitated beside the micrites. Sparite cement also precipitated in thallus cavities of algae that were not filled with micritic sediment. The sparite matrix microfacies is represented mainly by skeletal grainstones or biosparites (Pls. 7 ![]() -
8
-
8 ![]() -
9
-
9 ![]() , 15
, 15 ![]() , 18
, 18 ![]() -
19
-
19 ![]() -
20
-
20 ![]() -
21
-
21 ![]() -
22
-
22 ![]() -
23
-
23 ![]() ). In this type also, almost all bioclasts are fragments and thalluses of algae, but in the most thin-sections we find a small number of fragments of mollusc. Of the other allochems, intraclasts are common, mainly in the form of aggregated grains. These are typical sand bars and shoals microfacies. Dasycladalean algae that lived above storm wave base were, along with other particles, deposited on carbonate sand bars and shoals. Thinner or thicker irregular micritic envelopes around the thallus of algae can be seen in most of the photographs. These envelopes are overgrowths by epilithic microorganisms, most likely cyanobacteria. Micritic envelopes protect the thallus of Dasycladales from dissolving in the vadose diagenetic environment of the shoal, and allow their preservation.
). In this type also, almost all bioclasts are fragments and thalluses of algae, but in the most thin-sections we find a small number of fragments of mollusc. Of the other allochems, intraclasts are common, mainly in the form of aggregated grains. These are typical sand bars and shoals microfacies. Dasycladalean algae that lived above storm wave base were, along with other particles, deposited on carbonate sand bars and shoals. Thinner or thicker irregular micritic envelopes around the thallus of algae can be seen in most of the photographs. These envelopes are overgrowths by epilithic microorganisms, most likely cyanobacteria. Micritic envelopes protect the thallus of Dasycladales from dissolving in the vadose diagenetic environment of the shoal, and allow their preservation.
The investigated fossil material confirms that most Triassic Dasycladalean algae lived in similar environments to recent species: in shallow warm seas of the tropical and subtropical zone, normal salinity and protected from strong waves, in subtidal areas of open but protected lagoons and bays. There are, as with recent algae, exceptions to "standard" living environments, which are often overlooked. Some algae could tolerate turbid water, i.e., an increased proportion of clay; some tolerated and some even preferred more turbulent reef environments. Hence Praturlon (1963) describes Physoporella laevis from the uppermost Permian black clayey micritic limestones, very rich in organic matter, together with Mizzia velebitana. These algae appear to have tolerated a more protected environment with poor seawater circulation and some water turbidity (clayey and organic component). Adaptation to these conditions is evident in the very thin micritic thallus (see Chapter 5.2.8.), as it did not need stronger protection in such a quiet environment, and possible due to poor sunlight it could not even develop a thicker calcareous sheath. Also, an interesting species is Scinderella scopuliformis that has been described from the Anisian reef limestones on Ivan�čica Mt. (Grgasović et al., 2002). It was adapted to a high-energy environment so that it was attached to the reef rock. It was found together with O. cornuta, O. pilosa, O. praealpina, O. varicans, P. croatica and P. pauciforata, which means that they also could tolerate high energy environments.
Bechst�dt and Brandner (1970), as reported by Zorn (1976), provide an analysis of the distribution of algal groups in the Anisian facies of the Pragser Dolomite. From west to east they identify conglomerate and sandstone of the coastal area ("Peresschichten"), clayey nodular limestone with algae ("Algenwellenkalk") in the zone slightly distal from the coast, marls of the intraplatform trough, and pure platform limestones with algae ("Hauptdiploporenk"). The intraplatform trough, according to the authors, was used to purify the terrigenous material, so that the sea above the platform was clear. Within the "Algenwellenkalk" in the near-coast zone the Poncetella hexaster group dominated, and a little further from the coast the Physoporella-Oligoporella group with double whorls (i.e., the revised Oligoporella). Within the "Hauptdiploporenkalk", in the area closer to the trough the Macroporella alpina-Euteutloporella peniculiformis group dominated, and in the platform interior the group Physoporella-Oligoporella with single whorls (i.e., the revised Physoporella). Observing the columns with data on the percentage of individual algae (Bechst�dt & Brandner, 1970, App. 3), the dependence on lithofacies does not seem to be as high as the authors state. Subsequent research by Senowbari-Daryan et al. (1993) found that the mentioned deposits represent a carbonate ramp system into which an braided river flows, thus with a differently interpreted and more complex arrangement of facies than shown by previous authors. The aforementioned "Peresschichten" (now the Voltago Formation) represent fluvial and marginal-marine clastic sediments; "Algenwellenkalk" (now the Gracilis Formation) represents mixed clastic-carbonate sediments of the upper part of the ramp; marls correspond to the basin sediments of the Dont Formation; while the "Hauptdiploporenkalk" (now Upper Serla Formation) represents the carbonates of the upper part of the ramp. Senowbari-Daryan et al. (1993) nowhere mention reflections on the facies dependence of individual algae, although they share the same co-author (T. B.). Kotański (2013) however accepts the conclusions of Bechst�dt and Brandner (1970) and Zorn (1976), using them for his own interpretation of the dependence of algae on the environment in the Anisian of Upper Silesia. Thus, the Physoporella-Oligoporella group with single whorls is placed in the inner part of the platform, protected by shallow banks, the group with double whorls is placed on a muddy bottom far from the shore with poor water circulation and silt spread from the shore, Salpingoporella krupkaensis in an extremely closed environment, Diplopora annulatissima group in shallow banks with "diplopore meadows," and the Diplopora annulata group in a low turbulence environment. Still, this author states that is impossible to precisely locate the described assemblages since "the diplopores are rarely found in situ".
However, according to research on very abundant material from Croatia, as well as the Pia's collection, species of the genera Physoporella and Oligoporella regularly appear together in the same thin section (see App. 2) and there are no indications of their environmental dependence as already stated by Bystrick� (1986). The same is true with the genus Poncetella and with the species Euteutloporella peniculiformis. This dependence on the environment has not been mentioned also by other authors, such as Bassi and Fugagnoli (2005), and Pantić-Prodanović (1975).
Dasycladalean algae of the investigated genera have also been found in resedimented intercalations in deeper-water limestones of Late Bithynian to Early Pelsonian age in the Eastern Alps (Gawlick et al., 2021).
Most information on the stratigraphic ranges of the studied species can be obtained from the work of Granier and Grgasović (2000). Finds of Permian species are relatively rare, so their stratigraphic ranges are given in the relevant chapters and will not be further discussed.
The first biozonation of the Triassic on the basis of Dasycladalean algae originates from the book "Fundamentals of Stratigraphy" (Pia, 1930). In all his works Pia emphasized the stratigraphic importance of algae. The next author to renew interest in the application of Dasycladalean algae in stratigraphy was Ernst Ott, who in several of his contributions (Ott, 1972a, 1972c, 1974) gives a detailed account of this issue based on the investigations in the Northern Calcareous Alps. The contribution of Jan Bystrick� is particularly significant (Bystrick�, 1986). On the basis of rich fossil material, collected by very systematic research of the wide area of Slovak Karst and the West Carpathians, he showed the stratigraphic value of Dasycladalean algae and established a biostratigraphic zoning, on which I largely based my research. In the same area, but within the Hungarian territory in the Aggtelek karst, algae-based biostratigraphy is further elaborated by Piros (2002). The same author also analyses algal stratigraphy in the eastern part of the Northern Calcareous Alps (Piros et al., 1994, 2001; Piros & Preto, 2008). Important stratigraphic works are also Bassi and Fugagnoli (2005) and Kotański (2013). Let us also mention the table of stratigraphic ranges of the Permian and Triassic Dasycladales given by Granier and Deloffre (1995), which is transferred in its entirety by Bucur (1999). There is no discussion on stratigraphic ranges of particular species, so they cannot be commented upon, just to mention that some ranges of the studied species are not accurately presented: O. dissita (Anisian-Ladinian), O. minutula (Anisian-Ladinian), and Ph. pauciforata (Anisian-Ladinian).
As far as the Croatian Dinarides are concerned, historically the most significant work is Herak et al. (1967), not only because this paper presents for the first time a table of stratigraphic ranges of Dasycladalean algae, but also because it synthesizes the earliest phycological and stratigraphic research of Milan Herak (Herak, 1950, 1958, 1965), and also research related to the sections of the Basic Geological Map in the Velebit Mt area, which was conducted by Branko Sokač and his colleagues from the Institute of Geology. The work points out the stratigraphic importance of algae, especially in the separation of the Anisian and Ladinian. Discoveries on the ranges of Triassic algae in Croatia, including the studied genera, were synthesized in the work of Grgasović and Sokač (2003).
There is no evidence that the studied genera appear from the base of Anisian (see below), so the zone from the base of Anisian to their first appearance is defined by Grgasović and Sokač (2003) as the Poncetella cellulata Interval Zone. In addition to the leading species, Poncetella cellulata (Hurka), Poncetella hexaster (Pia), Poncetella helvetica (Pia) and Julpiaella subtilis (Pia) also occur. Algae of the studied genera are mainly present through the Physoporella pauciforata Range Zone (Grgasović & Sokač, 2003), which Bystrick� (1986) defined as the O. pilosa - P. pauciforata Assemblage Zone. In this fossil community we find relatively numerous specimens of algae from the investigated genera (Physoporella-Oligoporella group in further text): Oligoporella cornuta nom. nov. (= O. pilosa var. varicans Pia), O. dissita n. comb. (= P. dissita (G�mbel), Pia), O. minutula n. comb. (= P. minutula (G�mbel), Pia), O. pilosa Pia with varieties, O. praealpina n. comb. (= P. praealpina Pia) with varieties, O. varicans n. comb. (= P. varicans Pia), and Physoporella pauciforata (G�mbel), Steinmann with varieties. We can include here also the species Acroporella? prisca (Pia), n. comb., that is known only from the original description of Pia (1912). In thin-section LVI1 I found it together with P. pauciforata, and according to Lein (1993) it is a sample from the Anisian Steinalmkalk, hence there is no doubt about its Anisian age. Some rare species also belong to this zone, although their stratigraphic position cannot be precisely defined: O. intusannulata n. comb (= P. intusannulata Hurka), P. croatica Herak and Ardeiporella karrerioidea n. comb. (= O. karrerioidea Pia). In addition to the above, we also find species from the lower zone, as well as the species Macroporella alpina Pia, Euteutloporella peniculiformis (Ott), Scinderella scopuliformis Grgasović & Sokač, and Teutloporella tabulata Pia, and also the foraminifera Meandrospira dinarica Kochansky-Devid� & Pantić, and Pilammina densa Pantić. This fossil community is widespread in the Anisian Carbonates in the Outer Dinarides. Due to their relatively large dimensions, these algae (at the genus level) can be easily recognized in the field, and are very good guide fossils.
In his first works, Julius Pia defined the range of these algae as Anisian (Pia, 1920, 1925, 1927; Pia in Diener, 1925). In his work on the Anisian algae of Bosnia, Pia (1935a) for the first time gave age determinations for some localities at the sub-stage level, relying on the work of Kittl (1904) on the geology of the Sarajevo area. The vicinity of Sarajevo is a typical area for "Bosnische Stuffe" (Mojsisovics et al., 1895), which Pia (1930) renamed as "Illyrian", and its typical deposits are the "Hanbulog Limestone". The deposits in the bedrock of the Hanbulog Limestone are thus defined as "Pelsonian" (localities a, c, f-i, m, r), renamed by Pia (1930) from "Balatonische Stuffe", and those between the Lower Triassic and Pelsonian as "Hydaspian" (localities f, t). In locality "d" (see chapter O. p. var. intusannulata) Pia was not sure whether the light limestones from that locality were on the same level with the Hanbulog Limestone, so he marked the age as "Pelsonian, ? Illyrian". Works of Kittl (1904) and Pia (1930, 1935a) are now outdated due to the huge improvement in Triassic stratigraphy (Lucas, 2010) and should be used carefully. The biostratigraphy of the same area was later explored in detail by Sudar (1986), based mainly on conodonts, but also on foraminifera and dasyclads. Although he unfortunately does not use Pia's results in his work, it can be reconstructed that all the described Dasycladalean algae probably originate from the "Trebević Limestone" and "Dedović Carbonates" of Pelsonian age (in terms of the ammonite zone Balatonicus). As for the mentioned locality "d", it probably belongs to the "Dedović carbonates", while it is less probable that it belongs to the lower part of the Hanbulog Limestone, that has Upper Pelsonian - Illyrian range, so its lower part would correspond to the Upper Pelsonian or Basal Illyrian.
Pia (1937) in his work on the stratigraphy and tectonics of the Pragser Dolomites, based on the relationship of individual lithostratigraphic units, and the findings of different fossil groups and comparisons with other areas, gives ranges of individual algae at the sub-stage level. Thus the range of Physoporella and Oligoporella is defined as Hydaspian (early Anisian) - early Illyrian, which is later taken over by Ott (1972a) in his stratigraphic table. Extensive research in the same area has been conducted by Bechst�dt and Brandner (1970) and Senowbari-Daryan et al. (1993) (see Chapter 6). The Dolomites region is important for the studied genera because this is where the stratigraphically lowest published findings of the Physoporella-Oligoporella group originate from the "Lower Serla Dolomite" (= "Unterer Sarldolomit"). Pia (1930), in his first biostratigraphic zoning of the Middle Triassic, defines the Lower Hydaspian as Physoporella pauciforata Zone. For his table, unfortunately, he does not give further comments, so we can only assume that he did it based of the aforementioned findings in Lower Serla Dolomite. From this formation Pia (1937, p. 25) cites findings of P. pauciforata, Macroporella cf. dinarica and Oligoporella sp. These findings are very rare and there are no indications in the text that they refer to the lowest part of the Middle Triassic, and the table and zoning from the previous paper are not mentioned in the text. Bechst�dt and Brandner (1970, p. 51) from the Lower Serla Dolomites cite the findings of P. pauciforata, O. pilosa and M. alpina, unfortunately without illustrations. According to the attached columns (idem, App. 3) it can be indirectly concluded that the algae finds are from the upper part of this formation. The Lower Serla Dolomite is of Aegean Age, after Senowbari-Daryan et al. (1993) and Gianolla et al. (1998). In the Slovak karst, the first algae finds originate from the upper part of the Gutenstein Limestone, the lowest Anisian formation: P. pauciforata, O. dissita and O. pilosa var. intusannulata (Koll�rov�-Andrusovov� & Bystrick�, 1974; Bystrick�, 1986). In the neighbouring Aggtelek karst, the Gutenstein limestone is completely sterile (Piros, 2002), and the same is the case with the eastern part of the Northern Calcareous Alps (Piros et al., 1994, 2001; Gawlick et al., 2021). In Croatia, the lowest finding of investigated algae is from the Kremen hill in Lika, where an undoubted specimen of Physoporella sp., together with Macroporella alpina and the foraminifera Meandrospira dinarica, was found about 10 m above the Lower-Middle Triassic boundary.
We can therefore conclude that the studied algae in the wider Alpine region do not appear from the base of the Anisian, but slightly above, so the lower limit of the Physoporella-Oligoporella group's range, due to the impossibility of correlation, is roughly approximated as late Aegean.
The upper limit of the Physoporella-Oligoporella group's range is discussed in a number of papers. Almost all authors consider the upper limit to be the early Illyrian (Pia, 1937; Bechst�dt & Brandner, 1970; Ott, 1972a, 1972c; Bystrick�, 1986; Piros et al., 1994; Grgasović & Sokač, 2003). Piros et al. (2001), based on research of the Hochschwab massif, divide the Middle and Upper Triassic into eight algal-based zones, two of which are Anisian: 1) Physoporella spp. and P. hexaster (Pelsonian), 2) D. annulatissima (Illyrian), but there is an unsampled interval between them. Piros (2002), based on research in the Aggtelek Mts., which are an extension of the Slovak Karst to the Hungarian side, instead of one zone singles out three: I) Poncetella hexaster, Physoporella pauciforata, Oligoporella pilosa; II) Oligoporella pilosa, Euteutloporella peniculiformis; III) Oligoporella pilosa, Euteutloporella peniculiformis, Oligoporella dissita, Oligoporella varicans, Diplopora annulatissima. He places the boundary of Pelsonian and Illyrian within the third zone. As I have already pointed out, the different stratigraphic occurrences of certain species of the Physoporella - Oligoporella group do not correspond with the material from the Pia's collection and from Croatia. Certainly the most significant work is Bystrick� (1986) because it is based on the most extensive material and the most extensive research. The author names the Physoporella pauciforata - Oligoporella pilosa Assemblage Zone as the lowest Anisian zone. He determined the upper boundary as early Illyrian, which he discusses in detail. We can therefore conclude that the upper boundary of the studied algae in the wider Alpine region is the early Illyrian.
What is very important is that there is an interval between the last appearance of algae from the Physoporella-Oligoporella group and the first appearance of the Diplopora annulata - Kantia dolomitica group, which is also emphasized by many authors (Pia, 1930, 1937; Herak et al., 1967; Bystrick�, 1986; Piros et al., 1994; Piros, 2002; Grgasović & Sokač, 2003). It is also the case in the Germanic basin in Upper Silesia (Kotański, 2013). Bystrick� (1986) named this interval the Diplopora annulatissima Partial Range Zone, and Grgasović and Sokač (2003) the Diplopora annulatissima Interval Zone by its dominant species. Otherwise, the species D. annulatissima has the range Illyrian-Fassanian (Grgasović & Sokač, 2003), and very rare finds are from the Upper Pelsonian (Senowbari-Daryan et al., 1993). It is very rarely found in a sample together with the studied genera.
I would also like to mention that the former well-defined Anisian-Ladinan boundary in shallow water deposits was the first mass occurrence of algae from the Diplopora annulata - Kantia dolomitica group that roughly corresponded to the traditional Ladinian base in the base of the Reitzi ammonite zone (Kov�cs et al., 1994). By moving the base of Ladinian much higher to the base of the Curionii ammonite zone (Brack et al., 2005) it has become impossible to recognize.
Some species of the studied genera have a different range. As for the species Physoporella leptotheca Kochansky-Devid� (see Chapter 5.1.3.), its stratigraphic range has been problematic so far. It is described by Kochansky-Devid� (1967) from a sample from the �eleznica Valley in the Karavanke Mts. that the author received from another person without knowing whether it was a primary or secondary sample, and the only information about this locality is that the terrain is mostly covered with Lower Triassic deposits, and the sample is lithologically reminiscent of the Permian Trogkofel limestone. The Triassic age is suggested by the findings of Đurđanović from the Upper Triassic (Lower Norian) deposits near the village of Borovnica NW of Ljubljana, but the author does not illustrate these poorly preserved specimens. Fois (1979, p. 64) cites oral information from Ramov� who believes that the age of P. leptotheca is more likely Upper Anisian, perhaps close to the Anisian-Ladinian boundary, but without any arguments to confirm this allegation. We cannot accept all the above "evidences" because they are not substantiated. Fois (1979) illustrates the rich finds of these algae from the M. Poppera "algal limestones" in the Dolomites, along with Diplopora comelicana and Diplopora annulata dolomitica. Due to the findings of D. annulata, and based on the work of Ott (1972a), the age is defined as Late Illyrian. This should be discussed. In his works Ott (1972a, 1972c) for the first time lowered the lower limit of D. annulata in the late Illyrian, which was followed by some later authors (Fois, 1979; Fois & Jadoul, 1983; Senowbari-Daryan et al., 1993; Granier & Deloffre, 1995; Bucur, 1999). As the author himself says, he did so not because of the findings of these algae at a lower level, but because of changes in ammonite biostratigraphy (Rieber, 1967; Assereto, 1969). It later emerged that these changes were not substantiated and that this alga did not occur below the traditional Reitzi ammonite zone (Brack et al., 2005; Kov�cs et al., 1994). Gaetani et al. (1981) correctly place these findings in the Lower Latemar limestone. Due to all the above, the findings of P. leptotheca can be safely placed in the early Ladinian.
The species Euteutloporella chia (Herak), n. comb., is found only on the island of Chios in the Aegean Sea (Herak, 1967a) and the nearby Karaburun Peninsula in Turkey (Ott, 1972b) along with D. annulata, while findings from Romania are dubious (Popa & Dragastan, 1973; Dragastan, 1981). According to the geological profiles from Ott (1972b), it can be seen that in both areas it occurs in the upper part of the Ladinian, below the first appearance of Poikiloporella duplicata, so its age is Longobardian.
The species Physoporella? heraki Bystrick� is found often together with the species Poikiloporella duplicata which is typical for Carnian and is the main species of the range zone of the same name (Bystrick�, 1986). The species Poikiloporella brezovica Bystrick�, Salpingoporella humilis Bystrick�, S. spectabilis Bystrick�, S. sturi Bystrick�, Teutloporella herculea (Stoppani) and Uragiella supratriasica Bystrick� also occur in the same zone. Physoporella? heraki has been found in the Carnian not only in Slovakia, but also in the neighbouring area of northern Hungary (Velledits et al., 1999; Piros, 2002) and in Tibet (Fl�gel & Mu, 1982).
The species Neophysoporella jomdaensis (Fl�gel & Mu), n. comb., is described from the Carnian of Tibet, but it was also found in the western part of the Tethys in the Norian Main Dolomite of �umberak Mt. (Grgasović, 1997) and in the Upper Triassic of Shikoku Island, Japan (Bucur et al., 2020) in the Panthalassian realm. Neophysoporella zamparelliae (Parente & Climaco), n. comb., from the Apennines, as well as Neophysoporella n. sp. found in the Betic Cordilleras of Spain (Braga, 1986), are also of the Norian age.
Algae from the Triassic of the Germanic Basin are especially interesting. The so-called Germanic Triassic development represents an epicontinental development in an indented basin that was separated from the western part of the Tethys by raised Hercynian blocks. They were breached by tectonic controlled depressions (gates), which allowed communication with Tethys. The German basin embraced a large territory of the present-day Germany and Poland, reaching through France and Spain as far as to North Africa. Dasycladalean algae were found in three areas of the Germanic Basin: 1) Upper Silesia, southern Poland (G�mbel, 1872; Pia, 1931; Pastwa-Leszczyńska & Śliwiński, 1960; Kotański, 1994, 2013; Bucur & Matysik, 2020); 2) Moselle, NE France (Benecke, 1898) and 3) Piedmont, Italy (M�gard-Galli, 1968) - French Alps, Vanoise, France (Ellenberger, 1958) - Vaud, Switzerland (Botteron, 1961). The studied algae were also found in the Germanic Triassic in Upper Silesia in Poland. Some species are found in both Germanic and Alpine development: Anisian Physoporella pauciforata (G�mbel) Steinmann, Oligoporella minutula (G�mbel), n. comb., Oligoporella praealpina (Pia), n. comb., Oligoporella dissita (G�mbel), n. comb., and Ladinian Neophysoporella lotharingica (Benecke), n. comb. Other species are found only in the Germanic Triassic: Physoporella? elegans (Assmann ex Pia), n. comb., Oligoporella chrzanowensis Kotański and Oligoporella polonoandalusica (Kotański), n. comb. Kotański (2013) describes in detail the stratigraphic ranges of species from Upper Silesia and singles out six horizons with algae: I. very rare P. pauciforata, O. minutula (Upper Gogolin Fm., G�rażdże Fm. and Karchowice Fm.) of ?Bithynian - Pelsonian age; II. Rare P. pauciforata, O. minutula, O. praealpina, very rare P. ? elegans (at the bottom of the Diplopora Beds (Jemielnica Fm.)) of Pelsonian age; III. Abundant Physoporellae and Oligoporellae (lower part of Diplopora Dolomite) of Pelsonian age; IV. Diversified and abundant Physoporellae and Oligoporellae and Diplopora annulatissima (the middle part of the Diplopora Dolomite) of the early Illyrian age; V. Predominance of Diplopora annulatissima, subordinate P. pauciforata, P. ? elegans , O. minutula, N. lotharingica, D. nodosa (the upper part of Diplopora Dolomite) of the late Illyrian age; VI. Diplopora annulata and Kantia dolomitica, subordinate P. ? elegans , O. minutula, N. lotharingica, D. annulatissima, D. nodosa (the uppermost part of the Diplopora Dolomite) of late Illyrian (former Ladinian) age.
An important work is that of Nawrocki and Szulc (2000) that synthesizes all previous knowledge about the stratigraphy of the Germanic Triassic of Poland, including that of algae based on the work of Kotański (1994). A precise correlation was made between the German (Pery-Tethys) and Alpine (Tethys) developments based on conodonts and magnetostratigraphy which, along with ammonites, are the foundations of modern chronostratigraphy. The Pelsonian and Illyrian boundary is in the middle of Karchowice Fm., and the Illyrian and Fassanian boundary, based on the old boundary at the base of the Reitzi ammonite zone, is well above the Diplopora Beds, in the top of the Tarnowice Beds. The modern boundary at the base of the Curionii ammonite zone is not even present in Upper Silesia. This means that Diplopora annulata is present in the German Basin already in the late Illyrian. Diplopora annulata is also present in the late Anisian of China (Bucur & Enos, 2001). For Physoporella? elegans, which occurs only in Upper Silesia we can conclude an Illyrian age.
The species Neophysoporella lotharingica (Benecke), n. comb., occurs in the Germanic Triassic from the Illyrian, and at the top of its range is found together with Diplopora annulata. In the Dolomites, this species occurs from the base of Ladinian along with D. annulata and T. herculea (Gaetani et al., 1981). Pugliese (1997) also found it in Ladinian together with D. annulata and D. annulatissima. In Croatia, this alga was found in the vicinity of Knin, in the vicinity of Lička Kaldrma, and near Sv. Rok (Herak, 1965) together with D. annulata and D. annulatissima, suggesting an early Ladinian age. By reviewing the synonymy in Granier & Grgasović (2000), we can conclude that this alga is most common in the Lower Ladinian, and occurs from the Illyrian.
Most information on the geographical distribution of the studied species can be obtained from the work Granier and Grgasović (2000). As noted in Chapter 7, most of the taxa studied (Physoporella-Oligoporella group) come together at the same stratigraphic level, but also at the same locality, even in the same thin-section (App. 2). Algae from this group occur in Anisian deposits throughout the Alpine-Mediterranean area: Austria (G�mbel, 1872; Pia, 1912, 1920; Herak, 1967b; Ott, 1972d, 1974; Lein, 1993; Piros et al., 1994; Gawlick et al., 2021), Bosnia and Herzegovina (Pia, 1935a; Sudar, 1986), Bulgaria (Kotański & Čatalov, 1973), Croatia (Herak, 1965; Herak et al., 1967; Grgasović & Sokač, 2003; this work), Greece (Herak, 1967a), Hungary (Piros, 2002, 2003), Italy (Pia, 1912, 1920, 1930, 1937; Dieni & Spagnulo, 1964; Assereto et al., 1965; Zanin Buri, 1965; Hurka, 1969; Bechst�dt & Brandner, 1970; Senowbari-Daryan et al., 1993; Bassi & Fugagnoli, 2005), Montenegro (Pia, 1912, 1920; Pantić-Prodanović, 1975), Tatra Mts, Poland (Kotański, 1986; Rychliński et al., 2013), Romania (Patrulius, 1970; Popa & Dragastan, 1973; Dragastan, 1981; Bucur et al., 1994; Bucur, 1997), Serbia (Pantić, 1970), Slovakia (Pia, 1940; Bystrick�, 1957, 1962, 1964, 1967, 1983, 1986; Koll�rov�-Andrusovov� & Bystrick�, 1974; Buček, 1989), Slovenia (Herak, 1958; Fl�gel et al., 1994) and Switzerland (Pia, 1912, 1920; Zorn, 1971). Some of them (see Chapter 7) are also present in the epicontinental deposits of Poland (G�mbel, 1872; Pia, 1931; Pastwa-Leszczyńska & Śliwiński, 1960; Kotański, 1994, 2013; Bucur & Matysik, 2020), Italy (M�gard-Galli, 1968), France (Ellenberger, 1958) and Switzerland (Botteron, 1961). Very interesting is the finding of Oligoporella sp. by Gaździcki and Smit (1977) from NW Malaysia in the Middle Triassic Kodiang Limestone Formation.
Acroporella? prisca (Pia, 1912), n. comb., has been described only from Austria (Pia, 1912). Ardeiporella karrerioidea (Pia), n. comb., has been described only from Bosnia and Herzegovina (Pia, 1935a). Oligoporella intusannulata (Hurka), n. comb has been described from Italy (Hurka, 1967) and Slovakia (Bystrick�, 1957). Physoporella croatica Herak has been described from Croatia (Herak, 1958; this work) and Slovakia (Bystrick�, 1983, 1986). Physoporella leptotheca Kochansky-Devid� has been described from Slovenia (Kochansky-Devid�, 1967), Italy (Fois, 1979; Fois & Jadoul, 1983), and Switzerland (Ellenberger, 1958). Euteutloporella chia (Herak), n. comb., has been described from the island Chios, Greece (Herak, 1967a) and Turkey (Ott, 1972b), possibly (?) also from Romania (Popa & Dragastan, 1973). Physoporella? heraki Bystrick� has been described from Slovakia (Bystrick�, 1967, 1986), Tibet, China (Fl�gel & Mu, 1982) and Hungary (Piros, 2002). Neophysoporella jomdaensis (Fl�gel & Mu), n. comb., has been described from Tibet, China (Fl�gel & Mu, 1982), Croatia (Grgasović, 1997; this work) and Shikoku Island, Japan (Bucur et al., 2020). Neophysoporella zamparelliae (Parente & Climaco), n. comb., has been described only from Italy (Parente & Climaco, 1999).
Neophysoporella lotharingica (Benecke, 1898), n. comb., has been found in the Alpine Triassic of Austria (Pia, 1920), Croatia (Herak, 1965; Herak et al., 1967) and Italy (De Castro, 1979; Pugliese, 1997) and in the Germanic Triassic of France (Benecke, 1898; Pia, 1931), Poland (Kotański, 2013), and possibly (?) Sicily, Italy (Senowbari-Daryan & Di Stefano, 2001).
The species Physoporella? elegans (Assmann ex Pia, 1931), n. comb., Oligoporella chrzanowensis Kotański, 2013, and Oligoporella polonoandalusica (Kotański, 2013), n. comb., have been found only in the Germanic Triassic of Upper Silesia, Poland.
Taxonomic classification and differentiation of the genera Physoporella and Oligoporella has proved problematic since their establishment (Pia, 1912), while many later authors only reaffirmed the existence of the problem, but were unwilling to solve it. These algae are very common in Anisian deposits, and occur from the Lower Permian to the Norian. The research presented here is based primarily on the original type material from the collection of Julius von Pia, supplemented with material from Croatia. A small part of the taxa are described based on the literature.
The genus Physoporella was established by Steinmann (1903) on the basis of material and description by G�mbel (1872), encompassing all species with pear-shaped to spherical laterals enclosed in a calcareous sheath. Julius Pia in his first work (Pia, 1912) takes lateral closure as an important taxonomic characteristic, and slightly corrects the diagnosis of the genus, which now encompasses all species with piriform (pear-shaped) laterals, arranged in whorls and enclosed in a calcareous sheath. In the same work he establishes a new genus Oligoporella. Although the laterals of Oligoporella pilosa, the type species of this genus, are almost identical to those of Physoporella, Pia believes that they were not completely encased in calcareous sheet and protruded beyond the thallus, and adds them to the trichoform type. Due to this decision, the separation of these two genera also created problems for Pia himself, so in a series of his works (Pia, 1920, 1925, 1927, 1935a, 1935b) he discussed the issue of open and closed pores of laterals, doubting whether it matters whether there are closed pores or it may be better, however, to unite these two genera, and he expresses similar doubts in the description of each species. Numerous later authors (Hurka, 1967, 1969; Bechst�dt & Brandner, 1970; Ott, 1972a, 1974; Kotański & Čatalov, 1973; Zorn, 1974; Bucur et al., 1994; Piros, 2002) discuss the openness and closeness of pores and difficulties in separating these two genera, and regularly emphasize the need to solve this problem, but also stop there. On the other hand, Praturlon (1963) already noticed the existence of "two separate groups" with single and double whorls, followed later by other authors (see above), but do not give this feature enough importance and use it, like Pia, in defining species.
The taxonomic classification should be based as much as possible on those morpological characteristics of a calcareous skeleton that, as much as possible, reflect biologically significant properties. Algal calcification is variable and depends mostly on environmental factors. The variability of calcification is nicely seen on the submitted plates. It is also seen that the shape of the pores of these two genera is almost identical, and is designated as piriform. The biologically significant difference is the presence of single whorls in Physoporella pauciforata and double whorls in Oligoporella pilosa. Considering that, these genera are unambiguously different. In this sense, the diagnoses of the genera have been changed.
Physoporella Steinmann, 1903, emend.: Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in single whorls.
Oligoporella Pia, 1912, emend.: Cylindrical, sometimes slightly claviform thallus with piriform laterals arranged in double whorls. Each double whorl consists of two very close rows of laterals.
Although calcification is also influenced by environmental factors, so the calcareous shell can have different thicknesses, the shape of the outer wall of the thallus is completely related to the shape and arrangement of the laterals:
Physoporella pauciforata var. pauciforata has horizontal laterals, quite spaced, so that there was enough space between the laterals to fill, and it calcified fully. The laterals are relatively large, so the outer surface of the thallus is slightly undulated with protruding parts at the level of the whorls.
Physoporella pauciforata var. sulcata has oblique laterals, and since calcification follows the shape of the laterals, on the outer surface of the thallus has furrows between the whorls, and protruding parts at the level of the whorls.
Physoporella pauciforata var. undulata has relatively spaced horizontal whorls, so the space between the whorls is not completely calcified, but each lateral is surrounded by a relatively thin calcareous sheath, and the outer surface of the thallus is distinctly undulated.
Physoporella pauciforata var. gemerica has densely distributed horizontal laterals, so there was no space between the laterals for calcifying mucus and calcification generally does not reach the stem cell. Calcification on the outside follows the laterals and trichoblasts, so the outer surface is uneven.
Physoporella croatica has very large and densely distributed horizontal laterals, and the calcareous sheath is relatively thin, so the outer surface of the thallus is covered with conical protrusions - horns. This species has a characteristic calcification with a thin dark sheath around each lateral (both on the outside and on the inside next to the stem cell), while on the outside of the laterals a thicker sheath of microsparite crystals is developed
Physoporella leptotheca has spaced horizontal or slightly oblique whorls and laterals, so that a relatively thin calcareous wall envelops them evenly, resulting in "horns" and "rings" with laterals protruding from the calcareous cylinder.
Oligoporella pilosa var. pilosa has a thallus very similar to that of Physoporella pauciforata var. pauciforata with calcified space between the whorls and a slightly undulated outer surface. The difference is that the protruding parts are at the level of the double whorl, not the single ones, while the spacing of the whorls is similar. The variability of calcification in different specimens is characteristic.
Oligoporella pilosa var. intusannulata has relatively close double whorls. Calcification extends between the whorls to the stem cell, and since the laterals in adjacent whorls are closer in their central than in the basal part, the calcified parts between the whorls have the shape of the letter "T" in longitudinal and oblique sections, and the appearance of "bars" between the cavities of laterals. The outer surface of the thallus is flat or slightly undulated.
Oligoporella cornuta has divergent laterals. Calcification follows the outer surfaces of the laterals, so the outer surface of the thallus is usually slightly undulated, so that the recessed parts are inside the whorls, and protruding at the contact of the upper and lower rows of adjacent whorls. Calcification between the whorls that accompanies the shape of the laterals and the stem cell, identical to that of Oligoporella pilosa var. intusannulata, has the shape of the letter "T" in longitudinal sections, or "bars" in tangential sections.
Oligoporella intusannulata has calcification similar to the previous species, and differs mainly by the undulated inner wall, which follows the outline of the stem cell.
Oligoporella varicans has extremely divergent laterals, so the outer surface of the thallus is extremely undulated, because it follows the outer surface of the laterals. The concave parts are inside the double whorls, and the protruding ones in between. Due to the proximity of the distal portions of the laterals of adjacent whorls, calcification very rarely reaches the stem cell.
Oligoporella minutula has relatively spaced slightly oblique double whorls, so calcification covers the space between and above the whorls, and the outer surface is slightly undulated. The protruding parts are at the level of the whorls, and due to the inclination of the whorls, the undulation is asymmetric.
Oligoporella praealpina has a slightly different calcification. Usually calcification begins around the laterals, but in this species the parts between the laterals are often unevenly calcified, so the outer surface of the thallus is often uneven, i.e., rough, but can also be flat or slightly undulated. Due to the relatively dense whorls, calcification generally does not reach the stem cell.
Oligoporella dissita has relatively widely spaced double whorls, so calcification is mainly concentrated at the level of the whorls protruding above the less calcified part between the whorls. The outer surface of the thallus of this species is therefore annulated.
Oligoporella laevis from the Permian has a completely different mode of calcification with a thin micritic sheath that envelops the stem cell and each lateral separately, and the outer surface of the thallus is very embossed with each lateral recognizable.
Also characteristic of the thallus of the studied genera were calcified trichoblasts located on the tops of the laterals. In almost all species (P. pauciforata var. sulcata, P. p. var. undulata, P. p. var. gemerica, Oligoporella pilosa var. pilosa, O. cornuta, O. varicans) the constricted distal parts of the trichoblast are represented by shorter or longer spikes on the surface of the thallus, while in some specimens of Physoporella pauciforata var. pauciforata and Oligoporella praealpina var. praealpina larger parts of trichoblasts from adjacent laterals calcified together into more or less continuous shells.
Some species need to be further explored so their genus determinations are questionable. These are: Physoporella? heraki Bystrick�, Physoporella? hydrae (Richter et al., 1999), n. comb., and Physoporella? elegans (Assmann ex Pia), n. comb. Among the studied species, there were those whose structure differed significantly from those of the mentioned type species, so they were separated into other genera: Oligoporella karrerioidea Pia is placed in a new genus Ardeiporella, due to the characteristic shape of oblique laterals with a thin proximal part (neck), with an widened oval central part (head) and a long distal part (beak). The species Physoporella lotharingica Benecke has become a type species of a new genus Neophysoporella, characterized by slightly shorter and more rounded piriform laterals arranged in relatively dense whorls. In addition to the type species, Physoporella jomdaensis Fl�gel & Mu, and Physoporella zamparelliae Parente & Climaco are also included, as well as specimens from Braga (1986) that are described here as Neophysoporella n. sp. The species Oligoporella chia Herak was transferred to the genus Euteutloporella because it has trichoform laterals arranged in the whorls. Due to the shape of the laterals, the species Oligoporella prisca Pia does not belong to the studied genera, so it was transferred (with question mark) to the genus Acroporella, but further research on this problem is needed. Permian species have also been studied, but not in such detail.
I would like to thank everyone who helped me finish this work. I would like to thank my colleagues from the Croatian Geological Survey Josip Halamić, �eljka Glovacki Jernej, and Georg Koch for their joint field work, technicians Otilia Branković, Đurđica Novoselec, and Damir Galović for quality microscopic thin-sections, Nenad Kurtanjek for computer processing of the drawings, and late former superiors Josip Bukovac and Đuro Benček for support. From the time before the Google Translate, I thank the late Daniela Kabelač and Mihovil Juri�a for translating works from German, French, and Italian. I thank the professors from the Faculty of Science in Zagreb, Ivan Gu�ić, Jasenka Sremac, Vlasta Čosović, and the late Milan Herak, for their efforts in connection with my long time ago defence of PhD. I would like to thank Prof. Bruno Granier from Brest for our joint work in compiling the list of synonyms, and for the trust expressed. I thank Sigrid Berger from Heidelberg for introducing me to recent Dasycladales which has opened up completely different views of these wonderful organisms to me. I enhanced these views visting Bahamas with Prof. Antun Husinec from St. Lawrence University, USA. I thank Prof. Werner Piller, in that time from the Department of Paleontology, Vienna University, for the opportunity to study the collection of Julius von Pia that was stored there. Finally, I would like to express my special and sincere gratitude to my late mentor Branko Sokač, who passed to me a part of his exceptional knowledge about fossil algae, encouraged me and helped me in my work on a daily basis, and gave me many tips that improved this work.
I especially express my gratitude to the reviewers Prof. Ioan Bucur from Cluj-Napoca and Prof. Filippo Barattolo from Naples for the effort, useful corrections and suggestions which have contributed to this work. Thanks also to the language editor Prof. Simon F. Mitchell.
This study was supported by the Ministry of Science, Education and Sports of the Republic of Croatia under the projects, 181-1811096-1093, 181-1951126-1134 and by the Croatian Science Foundation under the project GOST IP-2019-04-3824. It is also a part of the PETRALGA project.
Assereto R. (1969).- Sul signiicato stratigrafico della "Zona ad Avisianus" del Trias Medio delle Alpi.- Bolletino della Societ� Geologica Italiana, Roma, vol. 88, p. 123-145 (2 Pls.).
Assereto R., Casati P. & Zanin Buri C. (1965).- Il Trias in Lombardia (Studi geologici e paleontologici). XIV. Sulla presenza di una scogliera anisica nella bassa Val Camonica.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 71, no. 3, p. 805-836 (4 Figs., 5 Pls.).
Assmann P. (1926).- Die Fauna der Wirbellosen und die Diploren der oberschlesischen Trias mit Ausnahme der Brachiopoden, Lamellibranchiaten, Gasteropoden und Korallen.- Jahrbuch Preu�ischen Geologischden Landesanstalt, Berlin, vol. 46 (1925), p. 504-527 (1 Fig., 2 Pls.).
Barattolo F., De Castro P. & Parente M. (1993).- Some remarks on Griphoporella curvata (G�mbel, 1872) Pia, 1915, dasycladacean green alga from the Upper Triassic. In: Barattolo F., De Castro P. & Parente M. (eds.), Studies on fossil benthic algae.- Bollettino della Societ� Paleontologica Italiana, Special Volume no. 1, Modena, p. 23-45 (7 Figs., 1 Table, 7 Pls.).
Barattolo F. & Romano R. (2002).- Clypeina bucuri n. sp. and Clypeina lucana n. sp. (green algae, Dasycladales) from the uppermost Paleocene(?)-Lower Eocene of Trentinara Formation (Southern Italy). In: Bucur I.I. & Filipescu S. (eds.), Research advances in calcareous algae and microbial carbonates.- Proceedings of the 4th IFAA Regional Meeting, Cluj-Napoca, August 29-September 5, 2001, p. 41-69.
Bassi D. & Fugagnoli A. (2005).- Triassic Dasycladalean algae from the Dolomites (northern Italy): Stratigraphic assessment.- Revista Espa�ola de Micropaleontolog�a, Madrid, vol. 37, no. 1, p. 95-103 (2 Figs.).
Bassoullet J.-P., Bernier P., Deloffre R., G�not P., Jaffrezo M., Poignant A.F. & Segonzac G. (1975).- R�flexions sur la syst�matique des Dasycladales fossiles. �tude critique de la terminologie et importance relative des crit�res de classification.- G�obios, Lyon, vol. 8, no. 4, p. 259-290 (6 Figs.).
Bassoullet J.-P., Bernier P., Deloffre R., G�not P., Jaffrezo M., Poignant A.F. & Segonzac G. (1977).- Classification criteria of fossil Dasycladales. In: Fl�gel E. (ed.), Fossil algae: Recent results and developments.- Springer-Verlag, p. 154-166.
Bassoullet J.-P., Bernier P., Deloffre R., G�not P., Jaffrezo M. & Vachard D. (1979).- Essai de classification des Dasycladales en tribus. In: 2�me Symposium international sur les Algues fossiles (Paris, 23-26 avril, 1979).- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 3, no. 2, p. 429-442 (2 Tables).
Bechst�dt T. & Brandner R. (1970).- Das Anis zwischen St. Vigil und dem H�hlensteintal (Pragser-und Olanger Dolomiten, S�dtirol).- Festband des Geologischen Institutes, 300-Jahr-Feier Universit�t Innsbruck, p. 9-103 (4 Figs., 18 Pls., 3 Suppls.).
Benecke E.W. (1876).- Ueber einige Versteinerungen, insbesondere aus den Umgebungen von Esino. In: �ber die Umgebungen von Esino in der Lombardei.- Geognostisch-pal�ontologische Beitr�ge, M�nchen, vol. 2, no. 3, p. 296-317 (4 Pls.).
Benecke E.W. (1898).- Diplopora und einige andere Versteinerungen im elsa�-lothringischen Muschelkalk.- Mittheilungen der Geologischen Landesanstalt von Elsa�-Lothringen, Strassburg, Band IV, p. 277-285 (2 Figs.).
Berger S. & Kaever M.J. (1992).- Dasycladales: An illustrated monograph of a fascinating algal order.- Thieme, Stuttgart, 247 p. (146 Figs., 20 Tables.).
Berger S., Kaever M.J. & Reichel B. (1997).- Influence of ecological conditions on magnesium and strontium content and ultrastructure of the calcareous skeleton of Bornetella sphaerica (Zarnadini), Dasycladales.- Pal�ontologische Zeitschrift, Stuttgart, vol. 71, no. 3-4, 189-195 (1 Fig., 3 Tables.).
Botteron G. (1961).- �tude g�ologique de la r�gion du Mont d'Or (Pr�alpes romandes).- Eclogae Geologicae Helvetiae, Basel, vol. 54, no. 1, p. 29-106 (16 Figs., 12 Pls.).
Brack P., Rieber H. & Nicora A. (2005).- The Global Boundary Stratotype Section and Point (GSSP) of the Ladinian Stage (Middle Triassic) at Bagolino (Southern Alps, Northern Italy) and its implications for the Triassic time scale.- Episodes, Bengalore, vol. 28, no. 4, p. 233-244. DOI: 10.18814/epiiugs/2005/v28i4/001
Braga J.C. (1986).- Upper Triassic Dasycladaceans in the Sierra de Baza (Alpuj�rride complex, Betic Cordillera, SE Spain).- Neues Jahrbuch f�r Geologie und Pal�ontologie, Monatshefte, Stuttgart, Jahrgang 1986, no. 4, p. 239-247 (1 Fig., 1 Table).
Braga J.C., & Martin J.M. (1987).- Distribucion de las Algas Dasycladaceas en el Trias Alpuj�rride.- Cuadernos de Geolog�a Ib�rica, Madrid, vol. 11, p. 475-489 (2 Figs., 1 Pl.).
Bucur I.I. (1997).- R�vision de la vari�t� Pilosa Pia ex Bystrick�, 1964, de l'esp�ce Oligoporella pilosa Pia, 1912, une algue dasycladac�e de l'Anisien.- Revue de Pal�obiologie, Gen�ve, vol. 16, no. 1 (1997), p. 181-186 (1 Table, 1 Pl.).
Bucur I.I. (1999).- Stratigraphic significance of some skeletal algae (Dasycladales, Caulerpales) of the Phanerozoic. In: Farinacci A. & Lord A.R. (eds.), Depositional episodes and bioevents.- Palaeopelagos Special Publication, Roma, no. 2, p. 53-104 (5 Figs., 18 Tables).
Bucur I.I. & Enos P. (2001).- Middle Triassic dasyclad algae from Guizhou, China.- Micropaleontology, Flushing � NY, vol. 47, no. 4, p. 317-338 (5 Figs., 7 Pls.).
Bucur I.I., Jovanović D., Radoičić R, Sudar M. & Mircescu C.V. (2021).- Lower Cretaceous carbonate deposits from the Derezna borehole (Carpatho-Balkkanides, Eastern Serbia) and remarks on some Dasycladalean Algae.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 17, no. 1, p. 3-14. DOI: 10.35463/j.apr.2021.01.01
Bucur I.I., Koch R., Kirmaci Z.M. & Tasli K. (2000).- Les algues Dasycladales du Cr�tac� inf�rieur (Calcaire de Berdiga) de Kircaova (r�gion de Kale-G�m�shane, NE Turquie).- Revue de Pal�obiologie, Gen�ve, vol. 19, no. 2, p. 435-463 (4 Figs., 9 Pls., 18 Tables).
Bucur I.I. & Matysik M. (2020).- Dasycladalean green algae and associated foraminifers in Middle Triassic (Lower and Middle Muschelkalk) carbonates of the south-eastern Germanic basin (Upper Silesia, Poland).- Annales Societatis Geologorum Poloniae, Krak�w, vol. 90, p. 391-407. DOI: 10.14241/asgp.2020.18
Bucur I.I., Rigaud S., Del Piero N., Fucelli A., Heerwagen E., Peybernes C., Peyrotti G., Verard C., Chablais J. & Martini R. (2020).- Upper Triassic calcareous algae from the Panthalassa ocean.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), Milano, vol. 126, no. 2, p. 499-540 (6 Figs., 16 Pls., 3 Tables). DOI: 10.13130/2039-4942/13681
Bucur I.I., Strutinski C. & Pop-Stratila D. (1994).- Middle Triassic Carbonate Deposits and Calcareous Algae from the Sasca Zone (Southern Carpathians, Romania).- Facies, Erlangen, 30, p. 85-100 (1 Fig., 6 Pls.).
Buček S. (1989).- V�sledky v�skumu dasyklad�lnych rias v triasov�ch sekvenci�ch Mal�ch Karp�t (Results of research of dasycladalean algae in triassic sequences of the Mal� Karpaty Mts.).- Zborn�k z paleontologickej konferencie: S�časn� probl�my a trendy v československej paleontol�gii, Geologick� �stav Dion�za �t�ra, Bratislava, p. 133-138 (3 Pls.).
Budai T., Lelkes G. & Piros O. (1993).- Evolution of Middle Triassic shallow marine carbonates in the Balaton Highland (Hungary).- Acta Geologica Hungarica, Budapest, vol. 36, no. 1, p. 145-165 (4 Figs., 6 Pls.).
Bystrick� J. (1957).- Pr�spevok k poznaniu diplop�r triasu Gemer�d.- Geologick� Sborn�k, Bratislava, vol. 8, no. 2, p. 226-241 (6 Pls.).
Bystrick� J. (1962).- Nov� Dasycladaceae triasu Slovensk�ho krasu (New Dasycladaceae of the Triassic in the Slovak Karst).- Geologick� Sborn�k, Bratislava, vol. 13, no. 2, p. 227-240 (2 Pls.).
Bystrick� J. (1964).- Slovensk� kras: Stratigrafia a Dasycladaceae mezozoika Slovensk�ho krasu (Stratigraphie und Dasycladaceen des Gebirges Slovensk� kras).- �stredn� �stav geologick�, Bratislava, 204 p. (38 Pls.).
Bystrick� J. (1967).- Die obertriadischen Dasycladazeen der Westkarpaten.- Geologick� Sborn�k - Geologica carpathica, Bratislava, vol. 18, no. 2, p. 285-309 (3 Figs., 10 Tables, 18 Pls.).
Bystrick� J. (1983).- Physoporella croatica Herak, 1958 of the Slovak Karst Anisian (Slovakia, the West Carpathians Mts.).- Geologick� Zborn�k - Geologica carpathica, Bratislava, vol. 34, no. 5, p. 507-533 (5 Figs., 17 Tables, 5 Pls.).
Bystrick� J. (1986).- Stratigraphic Ranging and Zonation of Dasycladal Algae in the West Carpathians Mts., Triassic.- Mineralia Slovaca, Bratislava, vol. 18 (1986), no. 4, p. 289-321 (3 Figs., 11 Columns, 1 Table, 6 Pls.).
Conrad M.A., Praturlon A. & Radoičić R. (1973).- Reinstatement of the genus Salpingoporella Pia (Dasycladaceae), followed by Salpingoporella steinhauseri n. sp.- Compte Rendu des S�ances de la Soci�t� de Physique et d'Histoire naturelle de Gen�ve, NS, vol. 7 (1972), no. 2-3, p. 103-111 (1 Fig., 1 Pl.).
Conrad M.A. & Varol B. (1990).- Cylindroporella taurica, n.sp., urges to review different patterns of calcification in the Mesozoic Dasycladales (Green Algae).- Archives des Sciences, Gen�ve, vol. 43, no. 1, p 193-214 (12 Figs.).
Cramer C. (1891).- �ber die verticillirten Siphoneen besonders Neomeris und Bornetella.- Neue Denkschriften der allgemeinen Schweizerischen Gesellschaft f�r die gesammten Naturwissenschaften, Z�rich, vol. 32, 48 p. (4 Pls.).
De Castro P. (1979).- Osservazioni su Diplopora nodosa Schafh�utl, 1863 (alga verde Dasicladacea) delle scogliere triassiche della Lucania (Appennino meridionale).- Bollettino della Societ� dei Naturalisti in Napoli, vol. 88, p. 1-51 (12 Figs., 1 Table, 20 Pls.).
De Castro P. (1993).- Observations on Campbelliella Radoičić 1959 and Neoteutloporella Bassoullet et al. 1978 (green algae, Dasycladales). In: Barattolo F., De Castro P. & Parente M. (eds.), Studies on fossil benthic algae.- Bollettino della Societ� Paleontologica Italiana, Modena, Special Volume no. 1, p. 121-184 (14 Figs., 3 Tables, 21 Pls.).
De Castro P. (1997).- Introduzione allo studio in sezione sottile delle Dasicladali fossili (An approach to thin-section study of fossil Dasycladales).- Quaderni dell'Accademia Pontaniana, Napoli, no. 22, 261 p. (39 Figs., 30 Pls.).
Deloffre R. (1987).- Nouvelle classification des algues Dasycladales fossiles.- Compte Rendu de l'Acad�mie des Sciences, Paris (Serie 2), no. 305, p. 1017-1020.
Deloffre R. (1988).- Nouvelle taxonomie des algues Dasycladales.- Bulletin des Centres Recherches Exploration-Production elf-Aquitaine, Pau, vol. 12, no. 1, p. 165-217 (9 Tables, 13 Pls.).
Deloffre R. & Granier B. (1991).- Hypoth�se phylog�nique des Algues Dasycladales.- Compte Rendu de l'Acad�mie des Sciences (s�rie II), Paris, vol. 312, p. 1673-1676 (2 Figs.).�
Diener C. (1925).- Leitfossilien der Trias. Wirbellose Tiere und Kalkalgen. In: G�rich G. (ed.), Leitfossilien, Lief. 4.- Verlag von Gebr�der Borntraeger, Berlin, 118 p. (28 Pls.).
Dieni I. & Spagnulo G. (1964).- Alghe calcaree nell'Anisico del Dosso dei Morti (Adamello sud-orientale).- Memorie della Accademia Patavina di SS.LL.AA., Classe di Scienze Matematiche e Naturali, Padova, vol. LXXVI (1963-1964), p. 1-13 (1 Fig., 2 Pls.).
Dragastan O. (1981).- Mesozoic Dasycladaceae from Romania: Distribution and biostratigraphical importance.- Facies, Erlangen, vol. 4, p. 165-196 (8 Figs., 1 Pl.).
Ellenberger F. (1958).- �tude g�ologique du pays de Vanoise.- M�moires pour servir � l'explication de la Carte g�ologique d�taill�e de la France, Paris, 561 p. (111 Figs., 42 Pls., 8 Panoramas).
Endo R. (1952).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, IV. Notes on the calcareous algae of the Omi limestone.- Transactions and Proceedings of the Paleontological Society of Japan, N.S., Tokyo, no. 8, p. 241-248 (1 Pl.).
Endo R. (1956).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, X. Fossil algae from the Kwant� and Kitakami Mountains.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, vol. 2, no. 2, p. 221-248 (2 Figs., 10 Pls.).
Endo R. (1957).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XI. Fossil algae from the Taishaku district, Hiroshima-Ken, and Kitami-no-kuni, Hokka�do.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, vol. 2, no. 3, p. 279-305 (8 Pls.).
Endo R. (1958).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XIII. A restudy of the genus Physoporella.- Transactions and Proceedings of the Paleontological Society of Japan, N.S., Tokyo, no. 31, p. 265-269 (1 Fig., 1 Pl.).
Endo R. (1959).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XIV. Fossil algae from the Nyugawa Valley in the Hida massif.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, vol. 3, no. 2, p. 177-207 (13 Pls.).
Endo R. (1961a).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XVI. Fossil algae from the Ominagahama district.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, Commemorative Volume dedicated to Professor Riuji Endo, p. 77-118 (19 Pls.).
Endo R. (1961b).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XVII. Fossil Algae from the Akiyoshi Limestone Group.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, Commemorative Volume dedicated to Professor Riuji Endo, p. 119-142 (7 Pls.).
Endo R. (1969).- Fossil algae from the Khao Phlong Phrab district in Tha�land. Contributions to the Geology and Paleontology of Southeast Asia, LXXIV. In: Kobayashi T. & Toriyama R. (eds.), Geology and paleontology of Southeast Asia, vol. 7.- University of Tokyo Press, p. 33-85 (38 Pls.).
Endo R. & Horiguchi M. (1957).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, XII. Fossil algae from the Fukuji District in the Hida Massif.- Japanese Journal of Geology and Geography, Tokyo, vol. 28, no. 4, p. 169-177 (3 Pls.).
Endo R. & Kanuma M. (1954).- Stratigraphical and paleontological studies of the later Paleozoic calcareous algae in Japan, VII. Geology of the Mino Mountain Land and southern part of Hida Plateau, with descriptions of the algal remains found in those discrictc.- The Science Reports of the Saitama University, series B (Biology and Earth Sciences), Urawa, vol. 1, no. 3, p. 177-208 (2 Figs., 5 Pls.).
Fl�gel E. (1990).- "Einschnitte" in der Entwicklung permischer Kalkalgen.- Mitteilungen des Naturwissenschaftlichen Vereines f�r Steiermark, Graz, vol. 190, p. 99-124 (2 Figs., 4 Pls.).
Fl�gel E., Ramov� A. & Bucur I.I. (1994).- Middle Triassic (Anisian) Limestones from Bled, Northwestern Slovenia: Microfacies and Microfossils.- Geologija, Ljubljana, vol. 36 (1993), p. 157-181 (2 Figs., 7 Pls.).
Fl�gel E. & Mu X. (1982).- Upper Triassic Dasycladaceae from Eastern Tibet.- Facies, Erlangen, vol. 6, p. 59-74 (3 Pls.).
Fois E. (1979).- A new Dasycladaceans (Calcareous Algae) assemblage from Triassic of M. Popera (Belluno, Italy).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 85, no. 1, p. 57-84 (3 Figs., 6 Pls.).
Fois E. & Jadoul F. (1983).- La Dorsale Paleocarnica anisica di Pontebba.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 89, no. 1, p. 3-30 (9 Figs., 1 Table, 1 Pl.).
Gaetani M., Fois E., Jadoul F. & Nicora A. (1981).- Nature and evolution of Middle Triassic carbonate buildups in the Dolomites (Italy). In: Cita M.B. & Ryan W.B.F. (eds.), Carbonate platforms of the passive-type continental margins, present and past.- Marine Geology, vol. 44, p. 25-57 (15 Figs., 3 Tables).
Gawlick H.-J., Lein R. & Bucur I.I. (2021).- Precursor extension to final Neo-Tethys break-up: Flooding events and their significance for the correlation of shallow-water and deep-marine organisms (Anisian, Eastern Alps, Austria).- International Journal of Earth Sciences, vol. 110, p. 419-446. DOI: 10.1007/s00531-020-01959-w
Gaździcki A. & Smit O.E. (1977).- Triasic foraminifers from the Malay Peninsula.- Acta Geologica Polonica, Warszawa, vol. 27, no. 3, p. 319-332 (4 Figs., 10 Pls.).
Gianolla P., De Zanche V. & Mietto P. (1998).- Triassic sequence stratigraphy in the Southern Alps (northern Italy): Definition of sequences and basin evolution. In: Graciansky P.-C. de, Hardenbol J., Jacquin J. & Vail P.R. (eds.), Mesozoic and Cenozoic sequence stratigraphy of European basins.- SEPM Special Publication, Tulsa - OK, vol. 60, p. 719-747 (14 Figs.).
Goričan �., Halamić J., Grgasović T. & Kolar-Jurkov�ek T. (2005).- Stratigraphic evolution of Triassic arc-backarc system in northwestern Croatia.- Bulletin de la Soci�t� g�ologique de France, vol. 176, p. 3-22 (5 Figs., 3 Pls.).
Granier B. (2012).- The contribution of calcareous green algae to the production of limestones: A review.- Geodiversitas, vol. 34, no. 1, p. 35-60 (10 Figs., 1 Table, 1 App.). DOI: 10.5252/g2012n1a3
Granier B. & Deloffre R. (1995).- Inventaire critique des Algues Dasycladales fossiles. IIIe partie - Les Algues Dasycladales du Permien et du Trias.- Revue de Pal�obiologie, Gen�ve, vol. 14, no. 1 (1994), p. 49-84 (1 Table).
Granier B., Dias-Brito D., Bucur I.I. & Tibana P. (2013).- Brasiliporella, a new mid-Cretaceous dasycladacean genus: The earliest record of the Tribe Batophoreae.- Facies, Erlangen, vol. 59, p. 207-220 (9 Figs.).
Granier B.R.C. & Grgasović T. (2000).- Les Algues Dasycladales du Permien et du Trias. Nouvelle tentative d'inventaire bibliographique, g�ographique et stratigraphique (Permian and Triassic Dasyclad algae. Bibliographic, geographic, and stratigraphic reappraisal).- Geologia Croatica, Zagreb, vol. 53, no. 1, p. 1-197 (27 Figs., 1 Table).
Granier B., Lethiers A. & Sander N.J. (2013).- The XXIst Century Edition of the "New studies on Triassic Siphoneae verticillatae, by Julius von Pia".- Jahrbuch der Geologischen Bundesanstalt, Wien, Band 153, Heft 1-4, p. 239-300 (24 Figs., 8 Tables, 7 Pls.).
Granier B. & Sander N.J. (2013).- The XXIst Century (the 100th Anniversary) Edition of the "New studies on Triassic Siphoneae verticillatae" by Julius von Pia.- Carnets de G�ologie (Notebooks on Geology), Madrid, Book, 2013/01 (CG2013_B01), 108 p. (7 Pls., 56 videos). DOI: 10.4267/2042/48735
Grgasović T. (1995).- Revision of genera Physoporella Steinmann and Oligoporella Pia (Dasycladales, Green Algae).- 6th International Symposium on Fossil Algae and Carbonate Platforms, Abstracts, Ankara, p. 15.
Grgasović T. (1997).- Upper Triassic biostratigraphy and algae from �umberak (Croatia).- Geologia Croatica, Zagreb, vol. 50, no. 2, p. 201-214 (1 Fig., 3 Tables, 2 Pls.).
Grgasović T. & Sokač B. (2003).- Review on fossil dasycladalean associations in the Triassic of Croatia.- Acta Micropalaeontologica Sinica, Nanjing, vol. 20, no. 1, p. 75-79 (1 Fig.).
Grgasović T., Sokač B. & Halamić J. (2002).-Scinderella scopuliformis nov. gen., nov. sp. (Dasycladales, green algae) from the Middle Triassic of Croatia.- Geobios, vol. 35, p. 303-312 (5 Figs.).
G�mbel C.W. (1872).- Die sogenannten Nulliporen (Lithothamnium und Dactylopora) und ihre Betheiligung an der Zusammensetzung der Kalkgesteine. Zweiter Theil: Die Nulliporen des Thierreichs (Dactyloporideae) nebst Nachtrag zum ersten Theile.- Abhandlungen der II. Classe der k�n. Akademie der Wissenschaften, M�nchen, vol. 11, no. 1, p. 231-290 (4 Pls.).
Herak M. (1950).- Ladiničke Dasycladaceae Jugoslavije i njihovo stratigrafsko značenje.- Rad Jugoslavenske akademije znanosti i umjetnosti, Zagreb, vol. 280, p. 115-141 (4 Pls.).
Herak M. (1958).- The Dasyclad genus Physoporella in the Anisian of Yugoslavia.- Journal of the Palaeontological Society of India, Lucknow, Birbal Sahni Memorial Number, vol. 3, p. 59-63 (2 Figs., 2 Pls.).
Herak M. (1965).- Comparative study of some Triassic Dasycladaceae in Yugoslavia (Komparativni studij nekih trijaskih dasikladaceja u Jugoslaviji).- Geolo�ki vjesnik, Zagreb, vol. 18, no. 1 (1964), p. 3-34 (1 Fig., 15 Pls.).
Herak M. (1967a).- Mesozoische Kalkalgen der Insel Chios (Griechenland).- Neues Jahrbuch f�r Geologie und Pal�ontologie, Abhandlungen, Stuttgart, vol. 129, no. 1, p. 97-104 (1 Table, 5 Pls.).
Herak M. (1967b).- Anisische Dasycladaceen vom Gosaukamm (Dachsteingebiet, �sterreich).- Mitteilungen der geologischen Gesellschaft in Wien, vol. 59 (1966), no. 2, p. 213-217 (3 Pls.).
Herak M., Kochansky V. & Gu�ić I. (1977).- The Development of the Dasyclad algae through the Ages. In: Fl�gel E. (ed.), Fossil algae: Recent results and sevelopments.- Springer-Verlag, p. 143-153 (2 Figs.).
Herak M., Sokač B. & �ćavničar B. (1967).- Correlation of the Triassic in SW Lika, Paklenica and Gorski Kotar (Croatia).- Geologick� Sborn�k - Geologica carpathica, Bratislava, vol. 18, no. 2, p. 189-202 (5 Figs., 1 Table).
Hurka H. (1967).- Variationsstatistische Untersuchungen an anisischen Dasycladaceen aus den Pragser Dolomiten in S�dtirol.- Neues Jahrbuch f�r Geologie und Pal�ontologie, Abhandlungen, Stuttgart, vol. 128, no. 1, p. 41-100 (19 Figs., 3 Tables, 2 Pls.).
Hurka H. (1969).- Umbildungstendenzen der Astformen in Physoporella-Oligoporella-Populationen (Dasycladaceen) aus dem Anis der Pragser Dolomiten (Italien).- Neues Jahrbuch f�r Geologie und Pal�ontologie, Monatshefte, Stuttgart, Jahrgang, 1969, no. 2, p. 104-120 (7 Figs.).
Hofmann T. & Granier B. (1994).- A guide to the collection of Julius Pia. 1st Part: "Neue Studien �ber triadische Siphonae verticilatae" Julius v. Pia, 1912.- Beitr�ge zur Pal�ontologie, vol. 19, p. 149-151.
Ishijima W., Ozaki H. & Nakamura M. (1971).- Upper Paleozoic calcareous algae from the limestone at Sakaishimachibun, Sa�tama-ken.- Bulletin of the National Science Museum, Tokyo, vol. 14, no. 1, p. 97-136 (16 Figs., 16 Pls.).
Kamptner E. (1958).- �ber das System und die Stammesgeschichte der Dasycladaceen (Siphoneae verticillatae).- Annalen des naturhistorischen Museums in Wien, 62 (1958), p. 95-102 (1 Table).
Kar�di V., Budai T., Haas J., V�r�s A., Piros O., Dunkl I. & T�th E. (2022).- Change from shallow to deep-water environment on an isolated carbonate platform in the Middle Triassic of the Transdanubian Range (Hungary).- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 587, no. 110793, p. 1-16 (14 Figs.).
Kittl E. (1904).- Geologie der Umgebung von Sarajevo.- Jahrbuch der Geologischen Reichanstalt, Wien, vol. 53 (1903), p. 515-748.
Kochansky V. & Herak M. (1960).- On the Carboniferous and Permian Dasycladaceae of Yugoslavia (O karbonskim i permskim dasikladacejama Jugoslavije).- Geolo�ki vjesnik, Zagreb, vol. 13 (1959), p. 65-94 (7 Figs., 9 Pls.).
Kochansky-Devid� V. (1967).- Eine interessante neue Physoporella-Art (Dasycladaceae) aus der Trias von Slowenien (Interesantna nova vrsta roda Physoporella (Dasycladaceae) iz trijasa Slovenije).- Geolo�ki vjesnik, Zagreb, vol. 20 (1966), p. 171-174 (1 Pl.).
Kochansky-Devid� V. & Gu�ić I. (1971).- Evolutions-Tendenzen der Dasycladaceen mit besonderer Ber�cksichtigung neuer Funde in Jugoslawien.- Pal�ontologische Zeitschrift, Stuttgart, vol. 45, no. 1/2, p. 82-91 (7 Figs.).
Koll�rov�-Andrusovov� V. & Bystrick� J. (1974).- �bersicht �ber den gegenw�rtigen Stand der Biostratigraphie der Trias der Westkarpaten. In: Zapfe H. (ed.), Die Stratigraphie der alpin-mediterranen Trias (The Stratigraphy of the Alpine-Mediterranen Triassic), Symposium (Wien, Mai, 1973).- �sterreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissionen, Wien (Springer Verlag), vol. 2, p. 125-136 (2 Tables).
Kord� K.B. (1950).- Ostatki vodorosley iz kembriya Kazakhstana.- Doklady Akademii Nauk SSSR, Moskow, vol. 73, no. 4, p. 809�812.
Kotański Z. (1986).- Macroflora. Class Chlorophyta. In: Gaździcki A., Fugelwicz R., Karczewski L., Kotański Z., Marcinkiewicz, T., Morycowa E., Orlowska-Zwolinska T., Piotrowski A., Reymanowna M., Roniewich E., Senkowiczowa H., Styk O., Trammer J. & Zawidzka K., Geology of Poland. Volume III. Atlas of guide and characteristic fossils. Part 2a. Mesozoic. Triassic, 1986 English edition (Budowa Geologiczna Polski, Tom III).- Instytut Geologiczny, Wydawnictwa Geologiczne, Warszawa, p. 177-184 (1 Table, 4 Pls.).
Kotański Z. (1994).- Middle Triassic Dasycladacea of Upper Silesian-Cracow Region and their stratigraphical and paleogeological significance. In: Third international meeting of Peri-Tethyan epicratonic basins. Excursion Guidebook.- Polish Geological Institute, Warszawa, p. 59-66.
Kotański Z. (2013).- Anisian Dasycladales from Upper Silesia and adjacent regions.- Carnets de G�ologie (Notebooks on Geology), Madrid, Book, 2013/02 (CG2013_B02), 205 p. (25 Figs., 39 Pls.). DOI: 10.4267/2042/53036
Kotański Z. & Čatalov G.A. (1973).- Triassic Dasycladaceae from central Balkan and fore-Balkan, Bulgaria.- Acta palaeontologica polonica, Warszawa, vol. 18, no. 2, p. 187-210 (2 Figs., 8 Pls.).
Kov�cs S., Doszt�ly L., G�cz�n F., Oravecz-Scheffer A. & Budai T. (1994).- The Anisian/Ladinian boundary in the Balaton Highland, Hungary - A complex microbiostratigraphic approach.- Albertiana, M�nster, vol. 14, p. 53-64 (7 Figs.).
Lein R. (1993).- B 5. Middle Triassic dasyclads from type localities of G�mbel and Pia; Lower Austria. In: H�fling R., Moussavian E. & Piller W.E. (eds.), Facial development of Algae-bearing carbonates sequences in the Eastern Alps. Field Trip Guidebook, Alpine Algae '93, M�nchen - Wien, 5 p. (2 Figs.).
Lucas S.G. (2010).- The Triassic Timescale.- Geological Society, London, Special Publication, vol. 334, 514 p.
Mancinelli A. (2006).- Acroporella cairensis n. sp. (Dasycladales) from the Barremian of Monte Cairo (southern Latium, Italy).- Facies, Erlangen, vol. 52, p. 411-416 (1 Table, 2 Figs.). DOI: 10.1007/s10347-006-0047-3
M�gard-Galli J. (1968).- Étude stratigraphique et tectonique du Monte Boulliagna (Haut Val d'Acceglio, Italie).- G�ologie alpine, Grenoble, vol. 44, p. 281-322 (8 Figs., 6 Panoramas).
Mojsisovics E.M. von, Waagen W. & Diener C. (1895).- Entwurf iner Gliederung der pelagischen Sedimente des Trias-Systems.- Sitzungsberichte, �sterreichische Akademie der Wissenschaften, Mathematisch-naturwissenschaftlische Klasse, Wien, vol. 104, no. 1, p. 1271-1302.
Nakamura M. (1981).- Fossil algae from the Ishiyama Limestone, Gifu Prefecture.- Bulletin of the National Science Museum, Series C (Geology and Paleontology), vol. 7, no. 2, p. 27-60 (10 Pls.).
Nawrocki J. & Szulc J. (2000).- The Middle Triassic magnetostratigraphy from the Peri-Tethys basin in Poland.- Earth and Planetary Science Letters, vol. 182, p. 77-92 (8 Figs., 2 Tables). DOI: 10.1016/S0012-821X(00)00230-2
Ott E. (1963).- Untersuchungen an ladinishen Dasycladaceen aus den N�rdlichen Kalkalpen.- Inaugural-Dissertation, Doktors der Naturwissenschaften, Matematisch-naturwissenschaftlichen Fakult�t der Eberhard-Karls Universit�t, T�bingen, 43 p. (47 Figs.).
Ott E. (1972a).- Die Kalkalgen-Chronologie der alpinen Mitteltrias in Angleichung an die Ammoniten-Chronologie.- Neues Jahrbuch f�r Geologie und Pal�ontologie, Abhandlungen, Stuttgart, vol. 141, no. 1, p. 81-115 (2 Figs., 1 Table).
Ott E. (1972b).- Mitteltriadische Riffe der N�rdlichen Kalkalpen und altersgleiche Bildungen auf Karaburun und Chios (�g�is).- Mitteilungen der Gesellschaft der Geologie- und Bergbaustudenten in �sterreich, vol. 21, no. 1, Symposium Innsbruck, 20.-23. M�rz, 1972: Mikrofazies und Mikrofauna der Alpinen Trias und deren Nachbargebiete, Innsbruck, p. 251-276 (7 Figs., 1 Suppl., 2 Pls.).
Ott E. (1972c).- Zur Kalkalgen-Stratigraphie der Alpinen Trias.- Mitteilungen der Gesellschaft der Geologie- und Bergbaustudenten in �sterreich, 21/1, Symposium Innsbruck, 20.-23. M�rz, 1972: Mikrofazies und Mikrofauna der Alpinen Trias und deren Nachbargebiete, Innsbruck , p. 455-463 (1 Suppl.).
Ott E. (1972d).- Neufunde anisischer Dasycladaceen (Kalkalgen) in �sterreich.- Anzeiger der mathematisch-naturwissenschaftlichen Klasse der �sterreichischen Akademie der Wissenschaften, Wien, Jahrgang, 1972, no. 9, p. 187-200 (4 Figs.).
Ott E. (1974).- Algae (Dasycladaceae). In: Zapfe H. (ed.), Catalogus Fossilium Austriae. Eine systematisches Verzeichnis aller auf osterreichischen Gebiet festgestellten Fossilien, Heft XVIIb.- �sterreichischen Akademie der Wissenschaften, Wien (Springer-Verlag, Wien), 64 p. (4 Tables).
Pantić S. (1970).- Mikropaleontolo�ke karakteristike trijaskog stuba antiklinale �drela (istočna Srbija) (Charact�ristiques micropal�ontologiques de la colonne triasique de l'anticlinal de �drelo (Serbie oriental)).- Zavod za geolo�ka i geofizička istra�ivanja, Vesnik (Serija A), Beograd, vol. 28, p. 377-386 (1 Fig., 1 Suppl., 13 Pls.).
Pantić-Prodanović S. (1975).- Trijaske mikrofacije Dinarida. Crna Gora, istočna Bosna i Hercegovina i zapadna Srbija (Les microfaci�s triasiques des Dinarides. Le Montenegro, la Bosnie orientale et l'Herzegovine et la Serbie occidentale).- Dru�tvo za nauku i umjetnost Crne Gore, Posebna izdanja, Titograd (Podgorica), vol. 4, 56 p. (1 Fig., 1 Suppl., 100 Pls.).
Parente M. & Climaco A. (1999).- Dasycladalean Green Algae from the Upper Triassic of Mt. Rotonda (Verbicaro Unit, Calabria-Lucania Border, Southern Italy).- Facies, Erlangen, vol. 41, p. 159-182 (2 Figs., 5 Tables, 7 Pls.).
Pastwa-Leszczyńska C. & Śliwiński S. (1960).- Występowanie glon�w (Dasycladaceae) w dolomitach kruszconośnych okolic Chrzanowa (Algae (Dasycladaceae) from orebearing dolomites in the vicinity of Chrzan�w).- Kwartalnik Geologiczny, Warszawa, vol. 4, no. 3, p. 679-699 (10 Figs., 2 Tables, 4 Pls.).
Patrulius D. (1970).- Inventar sumar al algelor Dasycladaceae triasice din Carpaţii rom�neşti (Inventaire sommaire des Algues Dasycladac�es triasiques des Carpathes Roumaines).- Dări de seamă ale şedinţelor, Bucureşti, vol. 55 (1967-1968), 3. Pal�ontologie, p. 187-196 (5 Pls.).
Pia J. (1912).- Neue Studien �ber die triadischen Siphoneae verticillatae.- Beitr�ge zur Pal�ontologie und Geologie �sterreich-Ungarns und des Orients, Wien und Leipzig, vol. 25, p. 25-81 (24 Figs., 7 Pls.).
Pia J. (1915).- Griphoporella curvata G�mb. sp. In: Spitz A. & Dyhrenfurth G. (eds.), Monographie der Engadiner Dolomiten zwischen Schuls, Scanfs und dem Stilfserjoch.- Beitr�ge zur geologischen Karte der Schweiz, Bern, N.F., vol. 44, p. 62 (1 Pl.).
Pia J. (1918).- Dasycladaceae. In: Trauth F. (ed.). Das Eoz�nvorkommen bei Radstadt im Pongau und seine Beziehungen zu den Gleichalterigen Ablagerungen bei Kirchberg am Wechsel und Wimpassing am Leithagebirge.- Denkschriften der Kaiserlichen Akademie der Wissenschaften Wien, Mathematisch-Naturwissenschaftliche Classe, vol. 95, p. 171-278.
Pia J. (1919).- Katalog der Diploporensammlung des naturhistorischen Museums in Wien.- Annalen des naturhistorischen Museums in Wien, Wien, vol. 33, no. 1, Notizen, p. 1-16.
Pia J. (1920).- Die Siphoneae verticillatae vom Karbon bis zur Kreide.- Abhandlungen der zoologisch-botanischen Gesellschaft in Wien, Wien, vol. 11, no. 2, 263 p. (8 Pls.).
Pia J. (1925).- Die Diploporen der Trias von S�ddalmatien.- Sitzungsberichte der Akademie der Wissenschaften in Wien, Mathematisch-naturwissenschaftliche Klasse, Abteilung I, vol. 133 (1924), nos. 7-8, p. 329-346 (1 Figs., 1 Table, 1 Pl.).
Pia J. (1927).- 1. Abteilung: Thallophyta. In: Hirmer M. (ed.), Handbuch der Pal�obotanik.- Oldenbourg, M�nchen und Berlin, vol. 1, p. 31-136 (Figs. 14-95).
Pia J. (1930).- Grundbegriffe der Stratigraphie mit ausf�hrlicher Anwendung auf die europ�ische Mitteltrias.- Verlag von Franz Deuticke, Leipzig und Wien, 252 p. (3 Figs.).
Pia J. (1931).- Die Dasycladaceen der germanischen Trias.- Annalen des naturhistorischen Museums in Wien, vol. 45, p. 265-279 (1 Pl.).
Pia J. (1935a).- Die Diploporen der anisischen Stufe Bosniens.- Geolo�ki anali Balkanskog poluostrva (Annales g�ologiques de la P�ninsule balkanique), Beograd, vol. 12, no. 2, p. 190-246 (55 Figs., 5 Pls.).
Pia J. (1935b).- Algen und Pseudoalgen aus der spanischen Trias. In: Schmid M. (ed.), Fossillien der spanischen Trias.- Abhandlungen der Heidelberger Akademie der Wissenschaften, Stifung H. Lang, M.-n. Kl., Heidelberg, Band 22, p. 9-17 (2 Pls.).
Pia J. (1937).- Stratigraphie und Tektonik der Pragser Dolomiten in S�dtirol.- Self-published, Wien, 15+248 p. (17 Figs., 14 Pls.).
Pia J. (1940).- Wirtelalgen (Dasycladaceen) aus den anisischen Kalken des Szilicei fenns�k in Nordungarn (A Szilicei fenns�k anisusi m�szk�veinek Dasycladacea-alg�i).- Abhandlungen aus dem mineralogisch-geologischen Institut der St. Tisza-Universit�t in Debrecen (K�zlem�nyek a Debreceni Tisza Istv�n-Tudom�nyegyetem �sv�ny - �s F�ldtani Intezetebol), vol. 4, no. 18, p. 1-7 (1 Pl.).
Pia J. (1943).- Geologische Untersuchungen in der Salmgruppe (Oberdonau).- Annalen des Naturhistorischen Museum in Wien, vol. 53, no. 1, p. 5-155 (9 Figs., 7 Pls.).
Piros O. (2002).- Anisian to Carnian carbonate platform facies and dasycladacean biostratigraphy of the Aggtelek Mts, Northeastern Hungary.- Acta Geologica Hungarica, Budapest, vol. 45, no. 2, p. 119-151 (2 Figs., 1 Table, 9 Pls.). DOI: 10.1556/ageol.45.2002.2.1
Piros O. (2003).- The Pelsonian Dasycladales of the Balaton Highland.- Geologica Hungarica (series Palaeontologica), Budapest, vol. 55, p. 53-69 (1 Fig., 4 Pls.).
Piros O., Mandl G.W., Lein R., Pavlik W, B�rczi-Makk A., Siblik M. & Lobitzer H. (1994).- Dasycladaceen-Assoziationen aus triadischen Seichtwasserkarbonaten des Ostabschnittes der N�rdlichen Kalkalpen. In: Lobitzer H. & Cs�sz�r G. (eds.).- Jubil�umsschrift, 20 Jahre Geologische Zussamenarbeit �sterreich-Ungarn (Geologischen Bundesanstalt), Wien, no. 2, p. 343-362 (4 Figs., 3 Pls.).
Piros O., Pavlik W., Moser M. & Bryda G. (2001).- Vorl�ufige Ergebnisse zur Kalkalgen-Stratigraphie der alpinen Mitteltrias aus dem Hochschwabmassiv (M�rzalpen-Decke, Steiermark).- Geologischen Bundesanstalt Arbeitstagung, 2001 - Neuberg an der M�rz Beitr�ge, p. 95-103 (2 Figs., 1 Table).
Piros O. & Preto N. (2008).- Dasycladalean algae distribution in ammonoid-bearing Middle Triassic platforms (Dolomites, Italy).- Facies, Erlangen, vol. 54, p. 581-595 (4 Figs.). DOI: 10.1007/s10347-008-0154-4
Popa E. & Dragastan O. (1973).- Alge si foraminifere triasice (Anisian - Ladinian) din estul Padurii Craiului (Muntii Apuseni).- Studii si Cercetari de Geologie, Geofizica, Geografie (Seria Geologie), Bucuresti, vol. 18, no. 2, p. 425-442 (1 Fig., 22 Pls.).
Praturlon A. (1963).- Dasycladaceae from Upper Permian of the Dolomites (Italy).- Geologica Romana, vol. 2 (1963), p. 119-137 (3 Figs., 5 Pls.).
Praturlon A. (1964).- Calcareous algae from Jurassic-Cretacoeus limestones of Central Apennines (southern Latium-Abruzzi).- Geologica Romana, vol. 3, p. 171-202 (34 Figs., 1 Table).
Praturlon A & Radoičić R. (1974).- Emendation of genus Acroporella (Dasycladaceae).- Geologica Romana, vol. 13, p. 17-20 (3 Figs.).
Pugliese A. (1997).- Middle-Late Triassic Dasycladales (green algae) from Brenta Dolomites (Giudicarie Alps, Italy).- Rivista Italiana de Paleontologia e Stratigrafia, Milano, vol. 103, no. 1, p. 71-80 (1 Fig., 1 Table, 2 Pls.).
Raciborski M. (1892).- Zapiski paleobotaniczne.- Odbitka z Kosmosu, Lw�w, vol. 11-12, 8 p.
Raviv V. & Lorch J. (1992).- Mesozoic Dasycladaceae (Chlorophyceae) from Israel.- Israel Journal of Botany, Tel Aviv, vol. 41, no. 1, p. 1-54 (12 Figs., 24 Tables, 18 Pls.).
Richter D.K., Dragastan O., Kube B., Beck T. & Zuhl H. (1999).- Fazies und Biostratigraphie des finalen Stadiums der obertriadischen Karbonattplatform von Hydra (Kap Kastello/Westrand des Pelagonikums/Griechenland).- Bochumer geologische und geotechnische Arbeiten, vol. 53, p. 127-148 (3 Figs., 1 Table, 6 Pls.).
Rieber H. (1967).- �ber die Grenze Anis-Ladin in den S�dalpen.- Eclogae Geologicae Helvetiae, Basel, vol. 60, p. 611-614.
Rychliński T., Ivanova D.K., Jaglarz P. & Bucur I.I. (2013).- Benthic foraminifera and calcareous algae from the Anisian-Norian succession in the Tatras (Poland and Slovakia): New data from High-Tatric and Kr�na units.- Studia UBB Geologia, Cluj Napoca, vol. 58, no. 1, p. 21-43 (5 Figs., 2 Pls.). DOI: 10.1007/s00531-020-01959-w
Schlagintweit F., R�nstler H., Gawlick H.J. (2003).- �ber ein Vorkommen von Physoporella pauciforata (G�mbel, 1872) Steinmann, 1903 (Dasycladaceae) aus Gutensteiner Kalken (Anis) des Rabenkogel (Steirisches Salzkammergut, �sterreich).- Jahrbuch der Geologischen Bundesanstalt, Wien, vol. 143, no. 4, p. 605-612 (2 Figs., 1 Pl., 1 Table).
Senowbari-Daryan B. & Di Stefano P. (2001).- Middle Triassic Dasycladales in Sicily: Evidence of an Anisian?-Ladinian carbonate platform.- Acta Geologica Hungarica, Budapest, vol. 44, no. 1, p. 95-109 (2 Tables, 4 Pls.).
Senowbari-Daryan B., Z�hlke R., Bechst�dt T. & Fl�gel E. (1993).- Anisian (Middle Triassic) buildups of the Northern Dolomites (Italy): The recovery of reef communities after the Permian/Triassic crisis.- Facies, Erlangen, vol. 28, p. 181-256 (17 Figs., 26 Pls.).
Slovenec D., �egvić B., Halamić J., Goričan �. & Zanoni G. (2019).- An ensialic volcanic arc along the northwestern edge of Palaeotethys - Insights from the Mid-Triassic volcano-sedimentary succession of Ivan�čica Mt. (northwestern Croatia).- Geological Journal, vol. 55, no. 6, p. 4324-4351 (15 Figs., 5 Tables).
Sokač B. (1996).- Taxonomic review of some Barremian and Aptian calcareous algae (Dasycladales) from the Dinaric and Adriatic karst regions of Croatia.- Geologia Croatica, Zagreb, vol. 49, no. 1, p. 1-35 (1-79) (3 Figs., 10 Tables, 22 Pls.).
Sokač B., Velić I., Grgasović T., Ćosović V. & Vlahović I. (2012).- Taxonomy and stratigraphy of an algal assemblage in Palaeogene deposits of the northern foothills of Mt. Biokovo (Southern Croatia).- Geologia Croatica, Zagreb, vol. 65, no. 2, p. 161-205 (2 Figs., 19 Pls.).
Steinmann G. (1903).- "Tetraporella Reme�i", eine neue Dasycladacea aus dem Tithon vom Stramberg.- Beitr�ge zur Pal�ontologie und Geologie �sterreich-Ungarns und des Orients, Wien, vol. 15, no. 2-3, p. 45-54 (11 Figs.).
Steinmann G. (1907).- Einf�rung in die Pal�ontologie.- Wilhem Engelmann Verlag, Leipzig, 542 p.
Stoppani A. (1857).- Studii geologici e paleontologici sulla Lombardia.- C. Turati, Milano, 461 p.
Stoppani A. (1858).- Monographie des Ac�phales des environs d'Esino, pr�s du lac de C�me et de Lenna, en Val-Brembana appartenant au d�p�t des p�trifications d�Esino. In: Les p�trifications d'Esino ou Description des fossiles appartenant au d�p�t Triasique sup�rieur des environs d'Esino en Lombardie. Pal�ontologie lombarde ou Description des fossiles de Lombardie (1e S�rie).- J. Bernardoni, Milano, p. 73-107 (8 pls.).
Sudar M. (1986).- Mikrofosili i biostratigrafija trijasa unutra�njih Dinarida Jugoslavije između Gučeva i Ljubi�nje (Triassic microfossils and biostratigraphy of the Inner Dinarides between Gučevo and Ljubi�nja mts., Yugoslavija).- Geolo�ki anali Balkanskoga poluostrva (Annales g�ologiques de la P�ninsule balkanique), Beograd, vol. 50, p. 151-394 (66 Figs., 30 Pls.).
Turland N.J., Wiersema J.H., Barrie F.R., Greuter W., Hawksworth D.L., Herendeen P.S., Knapp S., Kusber W.-H., Li D.-Z., Marhold K., May T.W., Mcneill J., Monro A.M., Prado J., Price M.J. & Smith G.F. (eds., 2018).- International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July, 2017. Regnum Vegetabile 159. Glash�tten.- Koeltz Botanical Books. DOI: 10.12705/Code.2018
Vachard D. (1980).- T�thys et Gondwana au Pal�ozo�que sup�rieur : Les donn�es afghanes ; biostratigraphie, micropal�ontologie, pal�og�ographie.- Th�se de Doctorat d'�tat, Documents et Travaux IGAL, Paris, no. 2, 2 vols., 463 p. (75 Figs., 50 Tables, 35 Pls.).
Valet G. (1968).- Contribution � l'�tude des Dasycladales : 1. Morphogen�se.- Nova Hedwigia, Lehre, vol. 16, p. 21-82 (23 Pls.).
Valet G. (1969).- Contribution � l'�tude des Dasycladales : 2. Cytologie et reproduction. 3. R�vision syst�matique.- Nova Hedwigia, Lehre, vol. 17, p. 552-644 (30 Pls.).
Velledits F., B�rczin�-Makk A. & Piros O. (1999).- A Kisfenns�ki M�szko (B�kk hegys�g) f�ciese �s kora (with English abstract: Facies and age of the Kisfenns�k Limestone (B�kk Mts.)).- F�ldtani K�zl�ny, Budapest, vol. 129, no. 4, p. 573-592 (4 Pls.).
Zanin Buri C. (1965).- Il Trias in Lombardia (Studi geologici e paleontologici). XIII. Le Alghe calcaree delle Prealpi Lombarde.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 71, no. 2, p. 449-554 (1 Fig., 1 Table, 23 Pls.).
Zorn H. (1971).- Pal�ontologische, stratigraphische und sedimentologische Untersuchungen des Salvatoredolomits (Mitteltrias) der Tessiner Kalkalpen. Unter besonderer Ber�cksichtigung der Mikrofazies, Diagenese und Taxonomie der Lamellibranchiata. In: Kuhn-Schnyder E. & Peyer B. (eds.), Der Triasfauna der Tessiner Kalkalpen.- Schweizerische Pal�ontologische Abhandlungen, Basel, vol. 91, p. 1-90 (14 Figs., 5 Tables, 17 Pls.).
Zorn H. (1972).- Bau und Funktion der Wirtel�ste einiger triassischer Dasycladaceen (Kalkalgen).- Neues Jahrbuch f�r Geologie und Pal�ontologie, Monatshefte, Stuttgart, Jahrgang 1972, no. 12, p. 727-740 (3 Figs.).
Zorn H. (1974).- Variationsstatistische Untersuchungen an Diplopora annulata (Schafh.) und D. philosophi (Pia), Kalkalgen der alpinen Mitteltrias.- Eclogae Geologicae Helvetiae, Basel, vol. 67, no. 1, p. 1-30 (18 Figs., 5 Tables, 2 Pls.).
Zorn H. (1976).- �ber den Lebensraum fossiler Wirtelalgen in der Trias der Alpen.- Naturwissenschaften, Berlin, vol. 63, p. 426-429 (2 Figs.).
Plate 1:
Physoporella pauciforata (G�mbel, 1872) Pia, 1912. 1-10, 13, 15 Physoporella pauciforata var. pauciforata Pia ex Bystrick�,
1964. 1. Slightly oblique cross section, Pia (1912, V, 13), neotype, XXVIII1, x 14.8; 2. Slightly oblique cross section, Pia (1912, V, 14), XXVIII2, x 18.5; 3. Slightly oblique longitudinal section, Pia (1912, V, 15), XXVIII2, x 11.6; 4. Longitudinal-oblique section, XXVIII1, x 11.6; 5-6. Oblique section, XXVIII2, x 7.3; 7. Oblique section, XXVIII2, x 10; 8. Tangential section, Pia (1912, V, 16), XXVIII2, x 12.7; 9. Oblique section, Pia (1912, V, 10), XXI3, x 18.5; 10. Slightly oblique cross section, Pia (1912, V, 9), XXI1, x 29.6; 13. Oblique section, Pia (1912, V, 11), XXI2, x 14.8; 15. Longitudinal-oblique section, Pia (1912, V, 19), LIV1, x 7.9. 11, 12 pars Physoporella pauciforata var. sulcata Bystrick�, 1962. 11. Longitudinal-oblique section, Pia (1912, V, 12), XXI2, x 12.7; 12 middle up. Oblique section, XXI3, x 12.7. 12 pars, 14 Physoporella sp. 12 right up. Oblique sections, XXI3, x 12.7; 14. Cross section, XXI3, x 14.8. 12 pars. Euteutloporella peniculiformis (Ott,
1963). 12 down. Oblique section, XXI3, x
12.7. All scale bars = 1 mm. |
|
Plate 2:
1-4 Physoporella pauciforata var. undulata Pia, 1935a. 1. Slightly oblique longitudinal section, Pia (1935a, 34), lectotype (Bystrick�, 1983), 561a, x 10; 2. Longitudinal-oblique section, 555a, x 12.7; 3. Oblique section, Pia (1935a, 33), 555a, x 12.7; 4. Cross-oblique section, (Pia, 1935a, ? No. 4 from the table of dimensions), 68, x 10. 5-6 Acroporella? prisca (Pia, 1912), n. comb. 5. Oblique section, LXXIX1, x 18.5; 6. Cross sections, LXXIX1,
x 18.5. 7-11 Physoporella pauciforata (G�mbel, 1872) Pia, 1912. 7. Oblique section, XXI2, x 10.1; 8. Oblique section, XXI3, x 12.7; 9. Oblique section, XXI1, x 13.8; 10. Slightly oblique cross section, XXI3, x 14.8; 11. Cross section, XXVIII2, x 10. All scale bars = 1 mm. |
|
Plate 3: 1-10 Acroporella? prisca (Pia, 1912), n.
comb. 1 middle. Slightly oblique longitudinal section, Pia (1912, V, 3), XXXIV1, x 10.1; 1 left and right. Oblique and cross sections, XXXIV2, x 10.1; 2. Slightly oblique longitudinal section, Pia (1912, V, 18), originally described as P. pauciforata, XXXIV1, x 7.9; 3. Longitudinal-oblique section, Pia (1912, V, 4), XXXIV1, x 12.7; 4. Cross section, Pia (1912, V, 2), LXXIX1, x 29.6; 5. Oblique section, LXXIX1, x 12.7; 6. Cross section, Pia (1912, V, 7), XXXIV2, x 18.5; 7. Tangential section, Pia (1912, V, 1), LXXIX1, x 7.3; 8. Oblique section, Pia (1912, V, 8), LVI1, x 14.8; 9. Oblique section, Pia (1912, V, 5), XXXIV2, x 18.5; 10. Oblique section, Pia (1912, V,6), XXXIV2, x
14.8. All scale bars = 1 mm. |
|
Plate 4: 1-2, 5, 7-9 Physoporella croatica Herak, 1958. 1 up left. Slightly oblique longitudinal section, Herak (1958, 14, 2), 66, x 7.2; 1 right. Slightly oblique longitudinal section, Herak (1958, 14, 2), lectotype (Herak in Granier & Deloffre,
1995, p. 57), 66, x 7.2; 1 far right. Cross section, Herak
(1958, 14, 2), 66, x 7.2; 2. Diverse sections, Herak (1958, 14, 3), 70, x 7.2; 5. Slightly oblique longitudinal section, Herak (1958, 14, 5), 74, x 11; 7. Cross section, Herak (1958, 14, 4), 73, x 12; 8. Cross section, Herak
(1958, 14, 1), 61, x 7.2; 9. Slightly oblique longitudinal section, Herak (1958, Fig. 2.8), x 7.2. 3 Physoporella pauciforata var. sulcata Bystrick�, 1962, originally described as Physoporella minutuloidea Herak,
1967a. 3. Oblique section, Herak (1967a, 9, 1),
holotype, T219/4, x 21. 4 Teutloporella herculea (Stoppani, 1857) Pia, 1912, originally described as Physoporella likana Herak, 1965. 4. Oblique section, Herak (1965, XIV, 2), lectotype (Herak in
Granier & Deloffre, 1995, p. 57), 86, x 15. 6 Euteutloporella chia (Herak,
1967a), n.
comb. 6. Longitudinal-oblique section, Herak (1967a, 7, 7),
holotype, T359.c, x 27. 10-12. Neophysoporella lotharingica (Benecke, 1898), n.
comb. 10. Oblique section, Herak (1965, XIV, 4), x 10: 11. Slightly oblique cross section, Herak (1965, XIV, 5), x 12; 12. Cross section, Herak (1965, XIV, 6), x
12. All scale bars = 1 mm. |
|
Plate 5: 1-6, 8-11 Oligoporella pilosa Pia, 1912. Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964. 1 up. Longitudinal-oblique section, Pia (1912, IV, 3), lectotype (Bucur & Grgasović in Bucur,
1997), I1, x 14.8; 1 down. Oblique section, I1, x 14.8; 2 left. Longitudinal section, Pia (1912, IV, 1), I11, x 12.7; 3. Cross section, Pia (1912, IV, 2), I9, x 29.6; 4. Cross section, I11, x 15.8; 5. Cross section, I1, x 12.7; 6. Oblique section, Pia (1912, IV, 4), I1, x 23.2; 8 pars. Longitudinal-oblique
(left) and oblique section (right), I1, x 12.7; 9. Longitudinal-oblique section, Pia (1912, IV, 6), I11, x 15.8; 10. Longitudinal-oblique section, Pia (1912, IV, 7), I6, x 12.7; 11. Oblique section, Pia (1912, IV, 8), V1, x 14.8. 2 pars Euteutloporella peniculiformis (Ott,
1963). 2 right. Oblique section, I11, x 12.7. 7 Teutloporella tenuis Pia, 1912, originally described as Oligoporella pilosa Pia, 1912. 7. Oblique section, Pia (1912, IV, 5), I3, x 14.8; 8 pars Macroporella dinarica Pia, 1912. 8 up right. Cross section, Pia (1912, I, 4), I1, x 12.7; 8 down right. Oblique section, I1, x
12.7. All scale bars = 1 mm. |
|
Plate 6: 1-8 Oligoporella pilosa Pia, 1912. Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964, originally described as Oligoporella pilosa Pia, 1912. 1 pars. Oblique sections, I1, x 12.7; 2 pars. Oblique and cross sections, I7, x 10.1; 3. Slightly oblique cross section, I1, x 12.7; 4. Cross section, I1, x 15.8; 5-8 originally described as Oligoporella serripora Pia, 1912. 5. Oblique section, Pia (1912, IV, 11), LIV8, x 14.8; 6. Slightly oblique cross section, Pia (1912, IV, 9), LIV2, x 18.5; 7. Oblique section, LIV3, x 10.1; 8. Oblique section, Pia (1912, IV, 10), LIV3, x 18.5. 1-2 Macroporella dinarica Pia, 1912. 1 pars. Cross sections and part of oblique section down
left, Pia (, 1912, II, 3), I1, x12.7; 2 pars. Cross and oblique sections, I7, x
10.1. All scale bars = 1 mm. |
|
Plate 7: Oligoporella pilosa Pia, 1912. 1-13 Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964. 1-2, 3 pars, 4, ?6, 8, 9 pars, 10-12 originally described as Oligoporella pilosa forma typica Pia, 1935a. 1 middle. Oblique section, Pia (1935a, ? No. 2 from the table of dimensions), 235b, x 10.8; 1 left and up. Oblique sections, 235b, x 10.8; 2. Oblique (up) and cross-oblique sections (down), 235b, x 7.9; 3 left. Oblique section, 231, x 10; 4. Oblique section, Pia (1935a, 8), 231, x 18.5; ? 6. Longitudinal-oblique section, 235a, x 10; 8. Oblique section, Pia (1935a, No. 4 from the table of dimensions), 235b, x 10.8; 9 pars. Longitudinal (up) and cross section, 46a, x10.1; 10. Oblique section, Pia (1935a, 10), 46a, x 18.5; 11. Oblique section, Pia (1935a, 11), 236b, x 11.6; 12. Oblique section, 231, x 10. 5, 7, 9 pars,
13 originally described as Oligoporella pilosa var. subvaricans Pia, 1935a. 5. Oblique section, Pia (1935a, 20), 231, x 14.8;
7. Oblique-cross section, Pia (1935a, ? No. 13 from the table of dimensions), 235a, x 10; 9 pars. Oblique section
(left), Pia (1935a, No. 2 from the table of dimensions), 46a, x 10.1; 13.
Oblique section, Pia (1935a, No. 9 from the table of dimensions), 232, x 10.8. 3 pars Teutloporella tabulata Pia, 1935a. 3 right. Cross-oblique section 231, x
10. All scale bars = 1 mm. |
|
Plate 8: Oligoporella pilosa Pia, 1912. ?1, 2-10, 12 Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964. ?1, 2-3, 5-9, 12 originally described as Oligoporella pilosa forma typica Pia, 1935a. ? 1. Longitudinal-tangential section, Pia (1935a, 12), 46a, x 10.1; 2. Slightly oblique cross section, Pia (1935a, 6), 46b, x 18.5; 3. Cross section, Pia (1935a, No. 7 from the table of dimensions), 46b, x 10.8; 5. Slightly oblique cross section, 577a, x 18.5; 6. Oblique section, Pia (1935a, 9), 577a, x 18.5; 7. Oblique section, Pia (1935a, No. 5 from the table of dimensions), 46b, x 10.8; 8. Cross section, Pia (1935a, No. 11 from the table of dimensions), 235b, x 10.8; 9. Oblique section, Pia (1935a, 5), 575b, x 18.5; 12. Slightly oblique cross section, Pia (1935a, 7), 557c, x 14.8. 4, 10 originally described as Oligoporella pilosa var. subvaricans Pia, 1935a. 4. Oblique-cross section, Pia (1935a, No. 6 from the table of dimensions), 46b, x 10.1; 10. Oblique section, Pia (1935a, No. 5 from the table of dimensions), 557a, x 18.5. 11, 13, 15, not 14 Oligoporella pilosa var. intusannulata Pia, 1935a. 11. Cross section, 575a, x 13.8; 13. Oblique section, 575a, x 10.8; 15. Longitudinal section, 555b, x 7.3; not 14. Oblique section,
gen. et sp. indet., Pia (1935a, II, 8), 576b, x 18.5. All scale bars =
1 mm. |
|
Plate 9: 1-8 Oligoporella pilosa Pia, 1912.
1-8 Oligoporella pilosa var. intusannulata Pia, 1935a. 1. Longitudinal-oblique section, Pia (1935a, 17),
lectotype, 575a, x 15.8; 2. Oblique section, Pia (1935a, No. 10 from the table of dimensions), 575a, x 11.6; 3. Oblique section, Pia (1935a, No. 8 from the table of dimensions), 574, x 12.7; 4. Longitudinal-oblique section, Pia (1935a, 18), 575a, x 11.6; 5. Longitudinal-oblique section, Pia (1935a, 19), 235a, x 11.6; 6. Oblique section, Pia (1935a, 16), 557a, x 14.8; 7. Oblique section, Pia (1935a, No. 2 from the table of dimensions), 573, x 18.5; 8. Oblique section, Pia (1935a, No. 7 from the table of dimensions), 231, x 10. 9-15 Oligoporella cornuta (Pia, 1935a), nom. nov., originally described as Oligoporella pilosa var. varicans Pia, 1935a. 9. Longitudinal-oblique section, 575a, x 10.1; 10. Longitudinal section, Pia (1935a, 25),
lectotype, 235b, x 18.5; 11. Longitudinal-oblique section, Pia (1935a, 24), 574, x 23.2; 12. Oblique section, Pia (1935a, 23), 575b, x 23.2; 13. Oblique section, Pia (1935a, 22), 235b, x 10,8; 14. Oblique section, 235b, x 10.8; 15. Oblique section, Pia (1935a, 21), 577a, x
23.2. All scale bars = 1 mm. |
|
Plate 10:
Oligoporella varicans (Pia, 1935a), n. comb. ?1. Originally described as ? Physoporella varicans Pia, 1935a. ? 1. Cross section, Pia (1935a, 45), 587, x 23.2. 2, 8, 11-12
Originally described as Physoporella varicans Pia, 1935a. 2 right. Slightly oblique longitudinal section, Pia (1935a, 44),
lectotype, 551, x 10.8; 2 left. Slightly oblique longitudinal section, 551, x 10.8; 8. Oblique section, Pia (1935a, No. 3 from the table of dimensions), 235a, x 10; 11. Oblique section, Pia (1935a, 43), 551, x 14.8; 12. Oblique section, 551, x 14.8. 4, 6, 9-10 Oligoporella cornuta (Pia, 1935a), n.
comb. Originally described as Oligoporella pilosa var. varicans Pia, 1935a. 4. Oblique section, Pia (1935a, N.F., No. 7 from the table of dimensions), 575b, x 23.2; 6. Oblique section, 574, x 29.6; 9. Oblique section, 575a, x 10.8; 10. Oblique section, Pia (1935a, No. 6 from the table of dimensions), 574, x 12.7. 3, 5, 7 Oligoporella cf. dissita (G�mbel, 1872), n.
comb. 3. Oblique section, CDXXXIII1, x 29.6; 5. Slightly oblique longitudinal section, CDXXXIII1, x 25.3; 7. Longitudinal section, CDXXXIII3, x
23.2. All scale bars = 1 mm. |
|
Plate 11: 1-9 Oligoporella dissita (G�mbel, 1872), n.
comb, originally described as Physoporella dissita (G�mbel, 1872) Pia, 1912. 1. Oblique section, Pia (1912, VI, 1),
neotype, LXII2, x 15.8; 2. Tangential section, Pia (1912, VI, 2), LXII4, x 18.5; 3. Oblique section, Pia (1912, VI, 3), LXII4, x 15.8; 4. Oblique section, LXII4, x 14.8; 5. Tangential section, LXII1, x 14.8; 6. Longitudinal-oblique section, LXII1, x 12.7; 7. Tangential section, LXII1, x 23.2; 8. Oblique section, Pia (1912, VI, 4), LXII4, x 18.5; 9. Longitudinal-oblique section, LXII5, x 14.8.
All scale bars = 1 mm. |
|
Plate 12:
1, ?2, 3-9 Oligoporella minutula (G�mbel, 1872), n. comb., originally described as Physoporella minutula (G�mbel, 1872) Pia, 1912. 1 left. Cross section, Pia (1912, VI, 5), XVI3, x 24.4; 1 right. Longitudinal section, Pia (1912, VI, 12), XVI3, x 24.4; 2. Longitudinal-oblique section, Pia (1912, VI, 11), XVI4, x 21.6; 3. Oblique-tangential section, XVI1, x 25.3; 4. Oblique section, Pia (1912, VI, 8), XVI3, x 23.2; 5. Longitudinal section, Pia (1912, VI, 6), XVI3, x 20.3; 6. Oblique section, Pia (1912, VI, 9), XVI1, x 29.6; 7. Longitudinal section, Pia (1912, VI, 7),
neotype, XVI3, x 23.2; 8. Slightly oblique longitudinal section, XVI3, x 23.2; 9. Tangential section, XVI1, x
29.6. All scale bars = 1 mm. |
|
Plate 13:
1 Oligoporella dissita (G�mbel, 1872), n. comb, originally described as Physoporella dissita (G�mbel, 1872) Pia, 1912. 1. Oblique
(left) and longitudinal-oblique (right) sections, LXII4, x 12.7. 2, 4 Oligoporella minutula
(G�mbel, 1872), n.
comb., originally described as Physoporella minutula (G�mbel, 1872) Pia, 1912. 2. Oblique section, 15, x 25.3; 4. Tangential section, Pia (1920, II, 20), 49, x 29.6. 3, 5-12 Oligoporella praealpina (Pia, 1920), n.
comb. Oligoporella praealpina var. praealpina n. var., originally described as Physoporella praealpina Pia, 1920. 3. Longitudinal-tangential (up) and cross section (down), CLVII3, x 10.8; 5. Longitudinal-oblique
(left) and tangential section (right), CLVII1, x 10.1; 6. Oblique section, CLVII3, x 10; 7. Oblique section, CLVII1, x 10.1; 8. Longitudinal section, CLVII3, x 10.8; 9. Oblique section, CLVII3, x 10.8; 10. Longitudinal section, CLVII3, x 10.8; 11. Longitudinal sections, CLVII3, x 10.8; 12. Slightly oblique longitudinal section, CLVII3, x
10. All scale bars = 1 mm. |
|
Plate 14: 1-9 Oligoporella praealpina (Pia, 1920), n.
comb. Oligoporella praealpina var. praealpina n. var., originally described as Physoporella praealpina Pia, 1920. 1. Slightly oblique longitudinal section, Pia (1920, III, 2),
lectotype, CLVII3, x 7.9; 2. Slightly oblique longitudinal (up) and cross section (down), CLVII1, x 12.7; 3. Oblique section, Pia (1920, III, 1), CLVII3, x 11.6; 4. Tangential section, Pia (1920, III, 7), CLVII3, x 18.5; 5. Tangential section, Pia (1920, III, 3), CLVII3, x 14.8; 6. Tangential section, Pia (1920, III, 6), CLVII3, x 11.6; 7. Slightly oblique cross section, Pia (1920, III, 4), CLVII2, x 14.8; 8. Tangential section, Pia (1920, III, 8), CLVII2, x 15.8; 9. Tangential section, Pia (1920, III, 9), CLVII2, x
15.8. All scale bars = 1 mm. |
|
Plate 15: 1-2, 4-6 Oligoporella praealpina (Pia, 1920), n.
comb. Oligoporella praealpina var. bosniensis n. var. Originally
described as Physoporella praealpina Pia, 1920. 1. Oblique section, Pia (1935a, 37), 583a, x 14.8; 2. Longitudinal-oblique section, Pia (1935a, 38),
holotype, 583b, x 14.8; 4. Longitudinal-tangential section, Pia (1935a, 39), 583b, x 11.6; 5. Oblique section, 583 a, x 14,8; 6. Oblique section, 583a, x 10.1; 3, 7-8 Ardeiporella karreiroidea (Pia, 1935a), n.
gen, n. comb. Originally described as Oligoporella karreiroidea Pia, 1935a. 3. Longitudinal-oblique section, Pia (1935a, III, 3),
holotype, 575b, x 20.3; 7. Oblique section, Pia (1935a, III, 2), 575b, x 18.5; 8. Cross section, 235a, x
10. All scale bars = 1 mm. |
|
Plate 16: Figures transferred from other
works. 1. Physoporella pauciforata var. sulcata Bystrick�, 1962, longitudinal-oblique section (Bystrick�, 1962, IV, 2),
holotype, x 11.2; 2. Neophysoporella n. sp., longitudinal-oblique section (Braga,
1986, 2.a), x 19.4; 3. Physoporella? elegans (Pia, 1912), n.
comb, slightly oblique longitudinal section (Pia, 1931, XXI, 3), originally described as Oligoporella elegans
n.sp., x 13; 4. Physoporella? heraki var. heraki Bystrick�, 1967, oblique section (Bystrick�, 1967, IX, 1),
holotype, x 11.8; 5. Physoporella? heraki var. tenuipora Bystrick�, 1967, oblique section (Bystrick�, 1967, XI, 2),
holotype, x 11.2; 6. Physoporella pauciforata var. gemerica Bystrick�, 1962, longitudinal section (Bystrick�,
1964, XVI, 4),
holotype, x 9.8; 7. Physoporella? elegans (Pia, 1912), n.
comb., tangential section (Pia, 1931, XXI, 4 pars), originally described as Oligoporella elegans
n.sp., x 18; 8. Physoporella pauciforata var. pauciforata Pia ex Bystrick�, 1962, longitudinal-oblique
(left) and cross section (right) (Bystrick�, 1964, PCIII, 4), x 9.6; 9. Physoporella pauciforata var. gemerica Bystrick�, 1962, tangential section (Bystrick�,
1964, XVI, 3), x 12.8; 10. Neophysoporella jomdaensis (Fl�gel & Mu,
1982), n.
comb., longitudinal section (Fl�gel & Mu, 1982, 9, 3),
holotype, x 9.6; 11. Physoporella leptotheca Kochansky-Devid�, 1967, tangential section (Kochansky-Devid�, 1967, I, 4), x 23.8; 12. Physoporella leptotheca, Kochansky-Devid�, 1967, longitudinal section (Kochansky-Devid�, 1967, I, 3), x 23.8; 13. Oligoporella praealpina var. praealpina n. var, isolated specimen (Pastwa-Leszczyńska & Śliwiński, 1960, IV, 15 = Fig. 6), originally described as Diplopora annulata forma physoporelloidea n. forma, x13; 14. Physoporella leptotheca Kochansky-Devid�, 1967, oblique section (Kochansky-Devid�, 1967, I, 1 up),
holotype, x 20. All scale bars = 1 mm. |
|
Plate 17:
Figures transferred from other works. 1. Oligoporella intusannulata (Hurka, 1967), n.
comb., longitudinal section (Hurka, 1967, 6, 2), lectotype, originally described as Physoporella intusannulata n.
sp, x 28.7; 2. Ardeiporella karrerioidea (Pia, 1935a), n.
gen., n. comb., oblique section (Pia , 1935a, III, 1), originally described as Oligoporella karrerioidea n. sp., x 18.9; 3. Oligoporella pilosa var. intusannulata Pia, 1935a, longitudinal-oblique section (Assereto et al., 1965, 79, 1), originally described as Oligoporella pilosa var. varicans, x 10.1; 4. Oligoporella praealpina var. praealpina n. var., longitudinal-oblique section (Kotański & Čatalov,
1973, VII, 13, 18-19), types, originally described as Physoporella praealpina var. sulcata n. var., x 11.7; 5. Oligoporella intusannulata (Hurka, 1967), n.
comb., cross section, (Hurka, 1967, 6, 4), originally described as Physoporella intusannulata n.
sp, x 45.3; 6. Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964, slightly oblique longitudinal section (Zanin Buri, 1965, 49, 3), originally described as Physoporella pauciforata var. variabilis n. var, x 18.8; 7. Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964, longitudinal section (Bystrick�,
1964, XI, 4), 245, x10; 8. Oligoporella minutula (G�mbel, 1872), n.
comb., izolirani primjerak (Pastwa-Leszczyńska & Śliwiński, 1960, III, 14 = Fig. 9), originally described as Oligoporella pilosa forma balinensis n. forma, x 44.3; 9. Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964, cross section (Pia, 1940, I, 11), originally described as Oligoporella pilosa var. physoporelloidea n. var., x 21.7; 10. Neophysoporella lotharingica (Benecke, 1898), n.
comb., rock surface (Benecke, 1898, 1), types, originally described as Diplopora lotharingica n. sp., x 1.2; 11. Neophysoporella lotharingica (Benecke, 1898), n.
comb, reconstruction (Benecke, 1898, 2), originally described as Diplopora lotharingica n. sp., x
12.1. All scale bars = 1 mm. |
|
Plate 18: Physoporella pauciforata (G�mbel, 1872) Pia, 1912. 1-4 Physoporella pauciforata var. pauciforata Pia ex Bystrick�, 1962. 1. Longitudinal-oblique section, BD I/21/3, x 11; 2. Longitudinal-oblique section, BD I/31, x 7.3; 3. Longitudinal-oblique section, BD I/24/5, x 11; 4. Longitudinal section, BD I/26/4, x 14.6; 5 Physoporella pauciforata var. gemerica Bystrick�, 1962. 5. Longitudinal-oblique section, VS 1108-B/9, x 14.6. 6 Physoporella pauciforata var. sulcata Bystrick�, 1962. 6. Longitudinal-tangential section, BD I/23/3, x 14.6. 1-4,
6 Belski dol, Ivan�čica,
Croatia: Anisian, Pelsonian - Lower Illyrian; 5 Hum �agudovec, Medvednica Mt,
Croatia: Anisian. All scale bars = 1 mm. |
|
Plate 19:
1-3, 5 Physoporella croatica Herak, 1958. 1. Slightly oblique longitudinal section, OM-1491 B, x 11; 2.
Longitudinal-tangential, OM-1491 A, x 7.3; 3. Longitudinal-tangential section, OM-1491, x 7.3;
5. Cross section, OM-1491 O, x 18.3. 4 Physoporella cf. croatica Herak, 1958. 4. Slightly oblique longitudinal section, OM-1491 P, x 7.3. 6 Physoporella sp. 6. Cross section, BD I/23/5; x 14.6. 7-9 Neophysoporella jomdaensis (Fl�gel & Mu,
1982), n.
comb. 7. Cross slightly oblique section, Grgasović (1997, II, 10);
SL, 19-15/IV; x 22; 8. Longitudinal section, Grgasović (1997, II, 15);
SL, 19-15/V; x 22; 9. Longitudinal section, Grgasović (1997, 11, 14), SL 17-9, x 22. 1-5
Jabuka, Trilj, Croatia: Anisian; 6 Belski dol, Ivan�čica, Croatia:
Anisian, Pelsonian - Lower Illyrian; 7-9 �umberak, NE Kra�ić, Croatia: Norian
(Hauptdolomit, Interval-zone Aulotorlus sinuosus). All scale bars = 1
mm. |
|
Plate 20: Oligoporella pilosa Pia, 1912. 1, 4 Oligoporella pilosa var. intusannulata Pia, 1935a. 1. Longitudinal section, U-5233, x 11; 4. Longitudinal section, U-5026, x 11; 2-3, 5 Oligoporella pilosa var. pilosa Pia ex Bystrick�,
1964. 2. Longitudinal-oblique section, VS 1108 B/1, x 19.1; 3. Oblique section, VS 1108 B/4, x 30.6; 5. Oblique section, OM 1495 C, x 22. 1, 4
Udbina, Croatia: Anisian; 2-3 Hum �agudovec, Medvednica Mt, Croatia: Anisian; 5
Jabuka, Trilj, Croatia: Anisian. All scale bars = 1 mm. |
|
Plate 21: 2 Oligoporella pilosa Pia, 1912. 2. Oblique section, BD I/23/8, x 14.6. 1 Oligoporella pilosa var. intusannulata Pia, 1935a. 1. Oblique section, U-5026 A, x 11. 5-7 Oligoporella cornuta (Pia, 1935a), nom. nov. 5. Longitudinal-oblique section, U-5233/2, x 22; 6. Longitudinal section, BD I/28/1, x 14.6; 7. Longitudinal section, BD I/22/7, x 12. 3-4, 8 Oligoporella dissita (G�mbel, 1872), n.
comb. 3. Tangential section, VS 1108 B/4, x 22; 4. Oblique section, VS 1108 B/3, x 30.6; 8. Oblique section, VS 1108 B/2, x 18.3. 1, 5
Udbina, Croatia: Anisian; 2, 6-7 Belski dol, Ivan�čica, Croatia: Anisian, Pelsonian - Lower
Illyrian; 3-4, 8 Hum �agudovec, Medvednica Mt, Croatia: Anisian. All scale
bars = 1 mm. |
|
Plate 22: Oligoporella praealpina (Pia, 1920), n.
comb. 1-4, 5 pars Oligoporella praealpina var. praealpina n. var. 1. Slightly oblique longitudinal section, U-5233, x 18.3; 2. Slightly oblique longitudinal section, U-5233, x 5.5; 3. Longitudinal section, VS 1108 B/2, x 14.6; 4. Tangential section, VS 1108 B/2, x 24; 5 right. Longitudinal section, VS 1108 B/6, x 9.8. 5 pars Euteutloporella peniculiformis (Ott,
1963). 5 left. Slightly oblique cross section, VS 1108 B/6, x 9.8. 1-2
Udbina, Croatia: Anisian; 3-5 Hum �agudovec, Medvednica Mt, Croatia: Anisian.
All scale bars = 1 mm. |
|
Plate 23:
1-4, 6-8 Oligoporella varicans (Pia, 1935a), n. comb. 1. Longitudinal section, BD I/24/10, x 22; 2. Longitudinal section, BD I/24/6, x 14.6; 3. Longitudinal section, BD I/21/5, x 14.6; 4. Slightly oblique longitudinal section, BD I/25/3, x 14.6; 6. Oblique section, BD I/21/1, x 14.6; 7. Longitudinal section, BD I/23/5, x 7.8; 8. Longitudinal section, BD I/22/1, x 11. 5 Oligoporella sp. 5. Cross section, BD I/25/2, x 22. 1-8 Belski dol, Ivan�čica, Croatia: Anisian, Pelsonian - Lower
Illyrian. All scale bars = 1 mm. |
|
Appendix 1: List of investigated thin-sections from the Julius Pia collection. Name of the taxon is followed by the year of Pia's publication, plate and plate number. The names of the taxa are given according to the original statements. The names in parentheses are according to my investigations. Tag "N.F." means that it is a specimen that Pia did not illustrate, but cited his finding in the thin-section. Sometimes it was possible to recognize a specimen from the table of dimensions, so this was marked as "Num".
| THIN-SECTION | ALGAE |
| I1 | Macroporella dinarica, 1912, II, 3, 4 Oligoporella pilosa, 1912, IV, 3, 4 (Ol. pilosa, M. dinarica, D. hexaster) |
| I3 | Ol. pilosa, 1912, IV, 5 (= T. tenuis) M. dinarica, 1912, II, 1, 2, 5, 6 Teutloporella? tenuis, 1912, III, 9, 10 Diplopora hexaster, 1912, VI, 13 |
| I4 | (Ol. pilosa, Teutloporella? tenuis) |
| I5 | (Macroporella dinarica) (Oligoporella sp.) |
| I6 | Oligoporella pilosa, 1912, IV, 7 Teutloporella tenuis, 1912, III, 7 (Macroporella dinarica) |
| I7 | (Oligoporella pilosa) (Macroporella dinarica) |
| I8 | (Macroporella dinarica) |
| I9 | Oligoporella pilosa, 1912, IV, 2 (Euteutloporella peniculiformis) (Macroporella dinarica) |
| I10 | Oligoporella pilosa, 1912, IV, 6 Teutloporella tenuis, 1912, III, 8 (M. dinarica, Oligoporela sp.) |
| I11 | Oligoporella pilosa, 1912, IV, 1 (Ol. pilosa, E. peniculiformis) (M. dinarica, D. hexaster) |
| I12 | (Ol. pilosa, Macroporella dinarica) |
| III1 | Teutloporella triasina, 1912, IV, 12, 13, 14 |
| V1 | Oligoporella pilosa, 1912, IV, 8 (Ol. pilosa, M. dinarica) |
| VI1 | T. triasina, 1912, IV, 17 (T. triasina) |
| XVI1 | Ph. minutula, 1912, VI, 9, 10 Diplopora helvetica, 1912, II, 17 (D. helvetica, E. peniculiformis) |
| XVI2 | (Physoporella minutula) |
| XVI3 | P. minutula, 1912, VI, 5, 6, 7, 8, 12 Diplopora helvetica, 1912, II, 16 (P. minutula, D. helvetica) |
| XVI4 | Physoporella minutula, 1912, VI, 11 (= ? Ol. pilosa pilosa) |
| XVI5 | (Physoporella minutula) |
| XXI1 | Ph. pauciforata, 1912, V, 9 (Physoporella pauciforata) |
| XXI2 | Physoporella pauciforata, 1912, V, 11, 12 (P. pauciforata sulcata) (Euteutloporella peniculiformis) |
| XXI3 | Ph. pauciforata, 1912, V, 10 (Euteutloporella peniculiformis) (Physoporella pauciforata) (P. pauciforata sulcata) |
| XXVIII1 | Physop. pauciforata, 1912, V, 13 Physoporella pauciforata |
| XXVIII2 | P. pauciforata, 1912, V, 14, 15, 16 (Physoporella pauciforata) |
| XXXIV1 | Physoporella pauciforata, 1912, V, 18 (= O. prisca) Oligoporella prisca, 1912, V, 4 |
| XXXIV2 | Ol. prisca, 1912, V, 3, 5, 6, 7 |
| XXXIV3 | (Oligoporella prisca) |
| XXXIV4 | (Oligoporella prisca) |
| XLI2 | Teutloporella vicentina, 1912, III, 14 |
| LIV1 | (Euteutloporella peniculiformis) Ph. pauciforata, 1912, V, 19 |
| LIV2 | Ol. serripora, 1912, IV, 9 |
| LIV3 | Ol. serripora, 1912, IV, 10 |
| LIV4-7 | (sterile) |
| LIV8 | Ol. serripora, 1912, IV, 11 |
| LVI1 | Oligoporella prisca, 1912, V, 8 (Physoporella pauciforata) |
| LVII1 | Macroporella alpina, 1912, II, 13, 14 (M. alpina, Diplopora helvetica) |
| LVII2 | Macroporella alpina, 1912, II, 15 (M. alpina, D. helvetica) |
| LVIII1 | Teutloporella gigantea, 1912, III, 5 |
| LVIII2 | Teutloporella gigantea, 1912, III, 6 (T. gigantea) |
| LXII1 | (Physoporella dissita) (Euteutloporella peniculiformis) |
| LXII2 | Physoporella dissita, 1912, VI, 1 (Physoporella dissita) (Scinderella scopuliformis) |
| LXII3 | (Physoporella dissita) (Euteutloporella peniculiformis) |
| LXII4 | Physoporella dissita, 1912, VI, 2, 3, 4 (Physoporella dissita) |
| LXII5 | (Physoporella dissita) (E. peniculiformis, Diplopora sp.) |
| LXXIX1 | Oligoporella prisca, 1912, V, 1, 2 (Oligoporella prisca) |
| LXXX1 | T. gigantea, 1912, III, 4 (Teutloporella gigantea) |
| LXXX2 | (Teutloporella gigantea) |
| LXXX5 | (E. peniculiformis, T. gigantea) |
| LXXX6 | T. gigantea, 1912, III, 3 |
| CLVII1 | (Physoporella praealpina) |
| CLVII2 | Ph. praealpina, 1920, III, 4, 8, 9 (Physoporella praealpina) |
| CLVII3 | Ph. praealpina, 1920, III, 1, 2, 3, 6, 7 (Physoporella praealpina) (Euteutloporella peniculiformis) |
| CDXXXIII1 | (Oligoporella cf. dissita) (Diplopora annulata) |
| CDXXXIII3 | (Oligoporella cf. dissita) (Diplopora annulata) |
| CDXXXIII4 | (Oligoporella cf. dissita) (Diplopora annulata) |
| DXXX2 | Oligoporella elegans, 1931 (Diplopora annulatissima) |
| DXXXI3 | Oligoporella elegans, 1931 (Macroporella alpina) |
| DXXXI4 | Oligoporella elegans, 1931 (Diplopora annulatissima) |
| DXXXI5 | (Diplopora annulatissima) |
| DXXXIV1 | Oligoporella elegans, 1931 |
| 15 | Ph. minutula (sample from Pia, 1920) |
| 46a | Ol. pilosa typica, 1935a, 10, 12 Macroporella alpina, 1935a, N.F.(Num11) Diplopora hexaster, 1935a, N.F. (7x) Oligoporella pilosa subvaricans, 1935a, N.F. (Num 2& 6) |
| 46b | Ol. pilosa typica, 1935a, 6 Ol. pilosa subvaricans, 1935a, N.F. (1x) Oligoporella pilosa typica, 1935a, N.F. (Num 5, 7) (E. cf. peniculiformis) Diplopora hexaster, 1935a, IV, 2 D. hexaster, 1935a, N.F. (4x) |
| 49 | Physoporella minutula, 1920, II, 20 |
| 68 | Physoporella pauciforata undulata, 1935a, N.F. (2x) |
| 231 | Ol. pilosa typica, 1935a, 8 Ol. pilosa subvaricans, 1935a, 20 Oligoporella pilosa subvaricans, 1935a, N.F. (Num 3 pars) Ol. pilosa intusannulata, 1935a, N.F. (Num 7) Teutloporella tabulata, 1935a, II, 6 T. tabulata, 1935a, N.F. (Num 6?) |
| 232 | Oligoporella pilosa subvaricans, 1935a, N.F. (Num 9) (Oligoporella pilosa) |
| 234d | (Oligoporella sp.) (Euteutloporella peniculiformis) |
| 235a | Ol. pilosa intusannulata, 1935a, 19 Oligoporella pilosa subvaricans, 1935a, N.F. (1x, Num 13?) Physoporella varicans, 1935a, N.F. (1x, Num 3) (E. peniculiformis, T. tabulata) (Oligoporella karrerioidea) (Oligoporella pilosa typica) |
| 235b | Ol. pilosa varicans, 1935a, 22, 25 Ol. pilosa subvaricans, 1935a, N.F. (1x) Oligoporella pilosa typica, 1935a, N.F. (4x, Num ?2, 4, ?6, ?11) Macroporella alpina, 1935a, I, 1 D. subtilis typica, 1935a, N.F. Teutloporella hirsuta, 1935a, N.F. |
| 236b | Ol. pilosa typica, 1935a, 11 (Ol. pilosa, M. alpina, Ph. varicans) |
| 386b | Holosporella siamensis, 1930, II |
| 551 | Physoporella varicans, 1935a, 43, 44 (Physoporella varicans) |
| 552 | Teutloporella hirsuta, 1935a, III, 7 |
| 555a | Ph. p. undulata, 1935a, 33 Ph. p. undulata, 1935a, N.F. (1x) |
| 555b | Diplopora serialis, 1935a, IV, 7 (O. pilosa intusannulata) (Diplopora sp.) |
| 555c | Teutloporella hirsuta, 1935a, N.F. |
| 555d | Teutloporella tabulata, 1935a, II, 5 |
| 557a | Ol. pilosa intusannulata, 1935a, 16 Oligoporella pilosa subvaricans, 1935a, N.F. (1x, Num 5) Diplopora hexaster, 1935a, IV, 1 Diplopora proba, 1935a, V, 1 Diplopora subtilis dissocladelloidea, 1935a, N.F. (1x) |
| 557c | Oligop. pilosa typica, 1935a, 7 Diplopora hexaster, 1935a, IV, 3 Diplopora hexaster, 1935a, N.F. |
| 561a | Ph. pauciforata undulata, 1935a, 34 Physoporella pauciforata undulata, 1935a, N.F. (1x) |
| 573 | Oligoporella pilosa intusannulata, 1935a, N.F. (1x, Num 2) D. subtilis typica, 1935a, N.F.(3x) D. subtilis calicina, 1935a, N.F.(1x) |
| 574 | Ol. pilosa varicans, 1935a, 24 Oligoporella pilosa varicans, 1935a, N.F. (1x, Num 6) M. alpina, 1935a, N.F. (Num 6) Diplopora subtilis dissocladelloidea, 1935a, IV, 5 Ol. pilosa intusannulata, 1935a, N.F. (1x, Num 8) Diplopora subtilis typica, 1935a, V, 12, N.F.(4x) |
| 575a | Ol. pilosa intusannulata, 1935a, 17, 18 Ol. pilosa intusannulata, 1935a, N.F. (Num 10) Macroporella alpina, 1935a, I, 5 D. subtilis typica, 1935a, N.F.(2x) (Oligoporella pilosa varicans) |
| 575b | Ol. pilosa varicans, 1935a, 23 Oligoporella pilosa typica, 1935a, 5 O. karreroidea, 1935a, III, 2, 3 Oligoporella pilosa varicans, 1935a, N.F. (1x, Num 7) Ol. pilosa subvaricans, 1935a, ?N.F.(1x) M. alpina, 1935a, N.F. (Num 7) D. subtilis typica, 1935a, N.F.(2x) (Ph. pauciforata sulcata) |
| 576a | Macroporella alpina, 1935a, II, 2 Diplopora subtilis typica, 1935a, V, 11, N.F.(2x) |
| 576b | non Ol. pilosa intusannulata, 1935a, II, 8 D. subtilis typica, 1935a, N.F.(3x) Diplopora subtilis dissocladelloidea, 1935a, IV, 6 |
| 576c | Diplopora subtilis typica, 1935a, V, 8, N.F. |
| 577a | Ol. pilosa typica, 1935a, 9 Ol. pilosa varicans, 1935a, 21 T. tabulata, 1935a, II, 4 D. subtilis calicina, 1935a, V, 6 D. subtilis typica, 1935a, V, 9 (Macroporella alpina) |
| 583a | Ph. praealpina, 1935a, 37 Teutloporella tabulata, 1935a, II, 3 Macroporella alpina, 1935a, I, 3 Diplopora subtilis calicina, 1935a, N.F. (1x) (Physoporella pauciforata) |
| 583b | Physoporella praealpina, 1935a, 38, 39 (Physoporella varicans) |
| 587 | ? Physoporella varicans, 1935a, 45 |
Appendix 2: Joint occurrence of investigated algae in the same thin-section. The table is based on materials from Croatia and materials from Pia's collection. Findings of the foraminifera Meandrospira dinarica are also added.
| Acroporella? prisca | P. pauciforata |
| Ardeiporella karrerioidea | D. subtilis var. subtilis, E. peniculiformis, M. alpina, O. cornuta, O. pilosa var. intusannulata, Ol. pilosa var. pilosa, O. varicans, P. pauciforata sulcata, T. tabulata. |
| Neophysoporella jomdaensis | ? |
| Neophysoporella lotharingica | D. annulata, D. annulatissima, M. beneckei, K. dolomitica, E. peniculiformis |
| Oligoporella cornuta | A. karrerioidea, D. subtilis var. subtilis, D. subtilis var. dissocladeoidea, E. peniculiformis, M. alpina, O. pilosa var. intusannulata, O. pilosa var. pilosa, O. varicans, P. pauciforata sulcata, S. scopuliformis, T. hirsuta, T. tabulata, Meandrospira dinarica |
| Oligoporella dissita | E. peniculiformis, O. ? minutula, O. pilosa var. pilosa, O. praealpina, O. varicans, P. pauciforata gemerica, P. pauciforata pauciforata |
| Oligoporella pilosa var. intusannulata | A. karrerioidea, D. proba, D. serialis, D. subtilis var. subtilis, D. subtilis var. calicina, D. subtilis var. dissocladeoidea, E. peniculiformis, M. alpina, O. cornuta, O. pilosa var. pilosa, O. varicans, P. hexaster, T. tabulata, S. scopuliformis, Meandrospira dinarica |
| Oligoporella pilosa var. pilosa | A. karrerioidea, D. proba, D. subtilis var. subtilis, D. subtilis var. dissocladeoidea, E. peniculiformis, M. alpina, M. dinarica, O. cornuta, O. dissita, O. pilosa var. intusannulata, O. praealpina, O. varicans, P. hexaster, P. pauciforata gemerica, P. pauciforata pauciforata, P. pauciforata sulcata, S. scopuliformis, T. hirsuta, T. tabulata, ?T. tenuis, Meandrospira dinarica |
| Oligoporella praealpina var. bosniaca | D. subtilis var. calicina, M. alpina, ?O. pilosa var. pilosa, P. pauciforata var.gemerica, S. scopuliformis, T. tabulata |
| Oligoporella praealpina var. praealpina | E. peniculiformis |
| Oligoporella varicans | P. cellulata, O. cornuta, O. pilosa var. pilosa, P. pauciforata sulcata, S. scopuliformis, E. peniculiformis, Meandrospira dinarica |
| Physoporella croatica | S. scopuliformis, E. peniculiformis, Meandrospira dinarica |
| Physoporella pauciforata var. gemerica | E. peniculiformis, O. dissita, ?O. minutula, O. pilosa var. pilosa, O. praealpina var. bosniaca |
| Physoporella pauciforata var. pauciforata | D. subtilis var. calicina, E. peniculiformis, M. alpina, O. cornuta, O. dissita, O. pilosa var. pilosa, O. praealpina var. bosniaca, O. varicans, P. pauciforata sulcata, S. scopuliformis, T. tabulata, Meandrospira dinarica |
| Physoporella pauciforata var. sulcata | A. karrerioidea, D. subtilis var. subtilis, E. peniculiformis, M. alpina,O. cornuta, O. pilosa var. pilosa, D. subtilis var. subtilis, P. pauciforata pauciforata, S. scopuliformis, Meandrospira dinarica |
| Physoporella pauciforata var. undulata | ? |