◄ Carnets Geol. 22 (13) ►
![]()
Outline:
[1. Introduction]
[2. Material and methods]
[3. Data]
[4. Results]
[5. A revised Berriasian-Hauterivian Time Scale]
[6. Conclusions] [Bibliographic references]
and ...
[Appendices]
Precision Stratigraphy Associates, 149 West Ridge Road, Cleveland Oklahoma 74040
(U.S.A.)
Published online in final form (pdf) on August 1, 2022
DOI
10.2110/carnets.2022.2213
![]()
[Editor: Bruno R.C. Granier]
![]()
New radioisotopic dates of Tithonian-Hauterivian strata in the Neuquén Basin significantly recalibrate Early Cretaceous numerical ages. In order to evaluate the implications of these revised ages, a graphic correlation experiment of twenty-three Andean Tithonian to Hauterivian sections integrated the ranges of 254 species, sequence boundaries, polarity chrons, and radioisotopic ages that compose the ANDESCS DB. This database accurately reproduces the order of Andean ammonite zones and places them in a relative metric scale of a composite reference section. The ranges in the ANDESCS DB were correlated with the LOK2016 DB that comprises Tithonian-Albian ammonites, calpionellids, nannofossils, and polarity chrons in Mediterranean-Caucasian Subrealm stage reference sections. In 2017 these ranges were calibrated to GTS2016 mega-annums (MA). Although most Andean ammonoids were endemic to the Indo-Pacific Subrealm, nannofossils, calpionellids and polarity chrons were present in both areas.
This stratigraphic experiment correlates base Berriasian as defined in France within the Substeueroceras koeneni Zone. In Andean sections this boundary is correlated with the Crassicolaria/Calpionella zone boundary dated at about 141 Ma. The base of the Valanginian defined by Calpionellites darderi correlates with the Neocomites wichmanni Zone of the Neuquén Basin (NB) recalibrated at 139.50 Ma, which is confirmed by multiple dates in Argentina, Mexico, Tibet, and elsewhere. The base Hauterivian correlates with base of Holcoptychites neuquensis Zone in the NB recalibrated at 131 Ma. Top of Hauterivian is in the Sabaudiella riverorum Zone in the NB and is dated at 127 Ma below an unconformity.
Previous cyclostratigraphic astrochronologic cycles are averaged and calibrate the duration of the Tithonian at 5.67 myr, the Berriasian at 5.27 myr, the Valanginian at 5.30 myr, and the Hauterivian at 5.60 myr. The age of each stage is recalibrated by adding revised durations to the most common age of base Valanginian of 139.5 Ma. These ages revise the Berriasian to Hauterivian stages time scale, and the ages of stage boundaries are on average 2.8 myr longer than proposed by the new Neuquén Basin radioisotopic dates.
• Early Cretaceous numerical dates;
• Tithonian;
• Berriasian;
• Valanginian;
• Hauterivian;
• biostratigraphy;
• Indo-Pacific Subrealm
Scott R.W. (2022).- Tithonian-Hauterivian chronostratigraphy (latest Jurassic-Early Cretaceous), Mediterranean-Caucasian Subrealm and southern Andes: A stratigraphic experiment and Time Scale.- Carnets Geol., Madrid, vol. 22, no. 13, p. 619-660.
• Indo-Pacific Subrealm new high quality
radioisotopic ages of the Tithonian-Hauterivian stages in the Neuquén Basin,
Indo-Pacific Subrealm, propose significant changes to the Early Cretaceous
numerical time scale.
• A chronostratigraphic database of of ammonites, calpionellids, nannofossils,
dinoflagellates, and polarity chrons spans uppermost Tithonian to Albian stages
from outcrops and drill cores on five continents, the LOK2016 DB, serves as a
chronostratigraphic reference data set.
• Stratigraphic range data of Andean taxa and polarity chrons, the ANDESCS DB,
integrates stratigraphic events into a common metric scale.
• Correlation of Andean ammonite zones with the global database projects
European stage boundaries into Andean sections about as predicted.
• The new U/Pd zircon dates would shorten the durations of stages, ammonite
zones and depositional cycles.
• New radioisotopic dates together with stage durations measured in Tethys
sections suggest that age of base Valanginian is close to 139.5 Ma and ages of
other stage boundaries may be calibrated by cyclostratigraphy.
Chronostratigraphie du Tithonien-Hauterivien (Jurassique terminal-Crétacé inférieur), du sous-domaine méditerranéen-caucasien et des Andes méridionales : Un exercice stratigraphique et l'échelle de temps.- De nouvelles datations radio-isotopiques des strates de l'intervalle Tithonien-Hauterivien du Bassin de Neuquén contribuent ŕ significativement recalibrer les âges numériques du Crétacé inférieur. Afin d'évaluer les implications de la révision de ces âges, un exercice de corrélation graphique incluant vingt-trois coupes andines de l'intervalle Tithonien-Hauterivien a été réalisé. Il intčgre les distributions de 254 espčces, les limites de séquence, les chrons de polarité et les âges radio-isotopiques qui composent la base de données ANDESCS. Cette base de données reproduit fidčlement l'ordre des zones d'ammonites andines et les replace sur l'échelle métrique relative d'une coupe composite de référence. Les éléments de la base de données ANDESCS ont été corrélés avec la base de données LOK2016 qui restitue les distributions des ammonites, calpionelles et nannofossiles ainsi que des chrons de polarité pour l'intervalle Tithonien-Albien pour des coupes de référence d'étages du sous-domaine méditerranéo-caucasien. En 2017, ces distributions furent calibrées sur les millions d'années de la GTS2016. Bien que la plupart des ammonoďdes andins soient endémiques du sous-domaine indo-pacifique, des zones de nannofossiles et de calpionelles ainsi que des chrons de polarité ont été reconnus dans les deux sous-domaines.
Cet exercice stratigraphique permet de placer la base du Berriasien telle que définie en France au sein de la Zone ŕ Substeueroceras koeneni. Dans les coupes andines, cette limite est corrélée avec celle des zones ŕ Crassicolaria et ŕ Calpionella datée d'environ 141 Ma. La base du Valanginien définie par Calpionellites darderi se corrčle avec la Zone ŕ Neocomites wichmanni du Bassin de Neuquén recalibrée ŕ 139,50 Ma, ce qui est confirmé par de multiples datations en Argentine, au Mexique, au Tibet et en d'autres régions. La base de l'Hauterivien est corrélée avec la base de la Zone ŕ Holcoptychites neuquensis du Bassin de Neuquén recalibrée ŕ 131 Ma. Le sommet de l'Hauterivien se trouve dans la Zone ŕ Sabaudiella riverorum du Bassin de Neuquén et est daté de 127 Ma sous une discordance.
Les cycles astrochronologiques cyclostratigraphiques précédents ont fait l'objet de calculs de moyennes qui attribuent au Tithonien une durée de 5,67 myr, 5,27 myr au Berriasien, 5,30 myr au Valanginien, et 5,60 myr ŕ l'Hauterivien. L'âge de chaque étage est alors recalculé en soustrayant ou ajoutant les durées révisées ŕ l'âge le plus couramment attribué ŕ la base du Valanginien soit 139,5 Ma. Ces âges constituent une révision de l'échelle de temps des étages Berriasien ŕ Hauterivien. Les âges des limites des étages sont ainsi en moyenne 2,8 myr plus longs que ceux proposés suite aux derničres datations radio-isotopiques du Bassin de Neuquén.
• datations numériques du Crétacé inférieur
;
• Tithonien ;
• Berriasien ;
• Valanginien ;
• Hauterivien ;
• biostratigraphie ;
• sous-domaine indo-pacifique
Numerical-age calibration of the Cretaceous Period has evolved over more than sixty years as radioisotopic measurements have been acquired and revised. Numerical ages were first estimated by radioisotopic ages, then by rates of sea-floor spreading and most recently by astrochronology and strontium isotopes. In 1959 numerical ages of the beginning and end of the Cretaceous Period were dated from 135±5 to 70±2 Ma (Holmes in Hinte, 1976) (Table 1). Since 1976 this time scale has been revised at least nineteen times. Beginning in 1995 a series of frequent updates adjusted the Cretaceous time scale as new data and methods were acquired (Ogg et al., 2004, 2012, 2016; Huang, 2018; Walker et al., 2018; Gale et al., 2020; Cohen et al., 2021). The most recent update, GTS2020 (Gale et al., 2020), resulted in more precise dates based on improved isotopic procedures and techniques. The development of cyclostratigraphy and astrochronology provide more accurate stage durations. In addition, biostratigraphic correlation of stages in the Mediterranean-Caucasian Subrealm of the Tethys Realm, where most type localities lie, with other provinces has become reliably demonstrated.
Table 1. Evolution of Cretaceous Period time scale. Ages from Hinte (1976), Gradstein et al. (1995), Remane et al. (2002), Ogg et al. (2004, 2012, 2016), and Gale et al. (2020). Andean ages as recalibrated from radioisotopic dates herein.
| Evolution of the Cretaceous Time Scale - Ma of Bases | |||||||||
| AGES | 1976 | 1995 | 2002 | 2004 | 2012 | 2016 | 2020 | 2021-ICS | Andean |
| Paleogene | 65 | 65 | 65.5 | 65.5 | 66 | 66 | 66.04 | 66 | |
| Maastrichtian | 70 | 71.3 | 71.3 | 70.6 | 72.1 | 72.1 | 72.17 | 72.1 ±0.2 | |
| Campanian | 78 | 83.5 | 83.5 | 83.5 | 83.6 | 89.2 | 83.65 | 83.6 ±0.2 | |
| Santonian | 82 | 85.8 | 85.8 | 85.8 | 86.3 | 86.5 | 85.7 | 86.3 ±0.5 | |
| Coniacian | 86 | 89 | 89 | 89.3 | 89.8 | 89.8 | 89.39 | 89.8 ±0.3 | |
| Turonian | 92 | 93.5 | 93.5 | 93.5 | 93.9 | 93.9 | 93.9 | 93.9 | |
| Cenomanian | 100 | 98.9 | 98.9 | 99.6 | 100.5 | 100.5 | 100.5 | 100.5 | |
| Albian | 108 | 112.2 | 112.2 | 112 | 113 | 113.1 | 113.7 | ~113 | |
| Aptian | 115 | 121 | 121 | 125 | 126.3 | 126.3 | 121.4 | ~125 | |
| Barremian | 121 | 127 | 127 | 130 | 130.8 | 130.8 | 126.5 | ~ 129.4 | 127 |
| Hauterivian | 126 | 132 | 132 | 136.4 | 133.9 | 134.7 | 132.6 | ~ 132.6 | 131 |
| Valanginian | 131 | 137 | 136.5 | 140.2 | 139.4 | 139.4 | 137.7 | ~ 139.8 | 139 |
| Berriasian | 135 | 144.2 | 142 | 145.5 | 145 | 145 | 143.1 | ~145 | 141 |
The use of "absolute" as an adjective for geological ages carries the connotation that the date will never change, is complete, is true, or is unlimited. A review of the Cretaceous time scale demonstrates that numerical ages of stage boundaries have changed as new data and technical methods have evolved and been applied (Table 1).
Along the eastern Pacific convergent margin of South America Upper Jurassic and Lower Cretaceous strata extend from Chile to southern Argentina. Andean retroarc basins were deformed during Middle Jurassic-Early Cretaceous time (Naipauer et al., 2012; Horton et al., 2016; Kietzmann et al., 2020, 2021a). This thick succession was deposited in a series of basins from the Abanico and Cura-Mallin basins in central Chile to the Neuquén Basin in west-central Argentina. The Lower Cretaceous strata are an essential source of chronostratigraphic data that enable correlation between the Tethys-Caucasian-Himalayan Province (sensu Page 2008 for the Tithonian) and the Andean area of the Indo-Pacific Subrealm.
New high-quality radioisotopic dates of the Tithonian-Hauterivian stages in the Neuquén Basin of the Indo-Pacific Subrealm propose important changes to the numerical age calibration of that time interval (Table 2) (Vennari et al., 2014; Aguirre-Urreta et al., 2017, 2019; Lena et al., 2019). These measurements would shift the age of the base Berriasian by 2-4 million years and less so the bases of the Valanginian, Hauterivian and Barremian. The result would be major recalibration of the ages of all the Tithonian-Hauterivian biozones (Ogg et al., 2016; Aguirre-Urreta et al., 2017; Reboulet et al., 2018; Gale et al., 2020; Kietzmann et al., 2020) and potentially affects ages and durations of subjacent and suprajacent stages.
Table 2. Important radioisotopic dates of uppermost Jurassic-Lower Cretaceous strata.
| Early Cretaceous radioisotopic dates | ||||||||
| Authors | Method | Location | Biostratigraphy | Stage | ANDESCS Dates Mu | Radioisotope Dates (Ma) | GTS2016 FAD taxa | LOK2016DB FAD taxa |
| Bralower et al., 1990 | Tuff, zircon, U/Pb | Grindstone Creek, California | Grantarhabdus meddii | Valanginian | 137.1±0.6 | 139 | ||
| Wan et al., 2011 | SHRIMP of rhyolite | Gyangze, southern Tibet | Calcicalathina oblongata | Valanginian | 136±3 | 139,4 | 139.6 | |
| Lopez-Martínez et al., 2017 | Tuff, zircon, U/Pb | Tlatlauquitepec, Puebla | Calpionellites major Zone | Valanginian | 134.0 ± 0.5 | 139.4 | ||
| Lopez-Martínez et al., 2017 | LAMC-ICPMS 87Sr86 | Tlatlauquitepec, Puebla | Calpionellites darderi Zone | Valanginian | 139.85 | 139.4 | 139.5 | |
| Lopez-Martínez et al., 2015 | Zircon LA-ICPMS U/Pb | Tamazunchale, San Luis Potosí | Calpionella elliptica overlies tuff above Crassicolaria | upper Berriasian | 139.1±2.6 | NA | 139.2 LAD 139.8 LAD | |
| Lena et al., 2019 | Zircon TIMS U/Pb | Puebla State, Mexico | N. steinmanni minor | Berriasian | 140.51±0.03 | 145.5 | 145.9 | |
| Lena et al., 2019 | Sediment rate | Puebla State, Mexico | N. steinmanni minor | Berriasian | 140.7 | 145.5 | 145.9 | |
| Lena et al., 2019 | Sediment rate | Puebla State, Mexico | Calpionella alpina | upper Tithonian | 140.9 | 145.7 | 146.9 | |
| Liu et al., 2013 | Zircon SIM U/Pb | Nagarze, southern Tibet | Manivitella pemmatoidea | Berriasian | 141-140 | 146.2 | ||
| Aguirre-Urreta et al., 2019 | Zircon TIMS U/Pb | El Portón, Argentina | S. riverorum | Upper Hauterivian | 1290 Mu | 126.97±0.15 | 131.3 | |
| Aguirre-Urreta et al., 2015 | Zircon TIMS U/Pb | Neuquén Basin Argentina | P. groeberi | Upper Hauterivian | 1281 Mu | 127.42±0.15 | NA | 131.8 |
| Aguirre-Urreta et al., 2015 | Zircon TIMS U/Pb | Mina San Eduardo, Argentina | S. riccardii | Upper Hauterivian | 1085 Mu | 129.09±0.16 | NA | 132.9 |
| Aguirre-Urreta et al., 2008 | Zircon SHRIMP U/Pb | Caepe Malal, Argentina | S. riccardii | Upper Hauterivian | 132.5±1.3 | NA | 132.9 | |
| Aguirre-Urreta et al., 2017, 2019 | Zircon LA-ICPMS U/Pb | El Portón, Argentina | H. agrioensis | Lower Hauterivian | 810 Mu | 130.39±0.16 | NA | 134.5 |
| Schwartz et al., 2016 | Zircon SHRIMP U/Pb | Neuquén Basin Argentina | H. neuquenensis | Lower Hauterivian | 130.0±0.80 | NA | 134.7 | |
| Vennari et al., 2014 | Zircon TIMS | Las Loicas, Argentina | A. noduliferum | Berriasian | 160 Mu | 139.55±0.18 | NA | 143.9 |
| Lena et al., 2019, Fig. 2 | Zircon TIMS U/Pb | Las Loicas, Argentina | A. noduliferum | Berriasian | 153 Mu | 139.24±0.05 | NA | 143.9 |
| Lena et al., 2019 | Zircon TIMS U/Pb | Las Loicas, Argentina | A. noduliferum | Berriasian | 130 Mu | 139.96±0.06 | NA | 143.9 |
| Lena et al., 2019 | Bayesian age-depth | Las Loicas, Argentina | N. winteri | Berriasian | 140.22±0.13 | 145.5 | 145.9 | |
| Lena et al., 2019 | Zircon TIMS U/Pb | Las Loicas, Argentina | R. asper | Tithonian | 112 Mu | 140.34±0.08 | 145.5 | 145.9 |
| Lena et al., 2019 | Bayesian age-depth | Las Loicas, Argentina | R. asper | Tithonian | 140.54±0.34 | 145.5 | 145,9 | |
| Lena et al., 2019 | Bayesian age-depth | Las Loicas, Argentina | R. asper | Tithonian | 140.6±0.4 | 145.5 | 145.9 | |
| Lena et al., 2019 | Bayesian age-depth | Las Loicas, Argentina | U. granulosa | Tithonian | 141.31±0.56 | |||
| Lena et al., 2019 | Zircon TIMS U/Pb | Las Loicas, Argentina | Crassicolaria Zone | Tithonian | 60 Mu | 142.04±0.06 | 147.7 | |
| Aguirre-Urreta et al., 2014; Lena et al., 2019 | Zircon CA-ID-TIMS | La Yesera, Argentina | Tordillo Fm. 1.5m below V. andesensis | Tithonian | 147.11±0.08 | |||
| Naipauer et al., 2015b | Zircon LA-ICPMS U/Pb | Las Loicas, Argentina | Tordillo Formation | Tithonian | 144 | |||
| Horton et al., 2016 | Zircon LA-ICPMS U/Pb | Neuquén Basin, Argentina | Tordillo Formation | Tithonian | 143.0±1.0 -149.5±1.2 | |||
| Naipauer et al., 2015c | Tordillo Formation | Tithonian | 144 | |||||
| Lena et al., 2019 | Tordillo Formation | Kimmeridgian | 147.11±0.078 | |||||
| Naipauer et al., 2012 | Zircon U/Pb | Tordillo Formation | Kimmeridgian | 152 | ||||
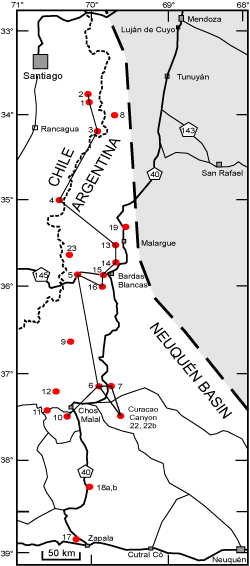
In order to evaluate the effects of these recent numerical dates, a stratigraphic experiment was conducted to integrate new Andean biostratigraphic taxa into a relative metric numerical database. From among the many well documented outcrop stratigraphic sections twenty-three were selected to represent the Andean
Tithonian-Hauterivian stages (Fig. 1 ![]() ). The second objective was to correlate the Andean zonal database with fossil zones in the Mediterranean Tethys in order to correlate the positions of stage boundaries with Andean zones. The
Tethys and Andean range databases were combined and then the new radioisotope dates were projected into the database. The relative ages of first and last occurrences of nearly 250 stratigraphic events were recalibrated to new dates. These Andean stage ages are compared with GTS2020 ages. The recalibration of numerical ages of Andean stratigraphic markers has significant implications on durations of stages and zones as well as sedimentary rates and durations. Ages of Berriasian-Hauterivian stage boundaries recalibrated by different methods are compared.
). The second objective was to correlate the Andean zonal database with fossil zones in the Mediterranean Tethys in order to correlate the positions of stage boundaries with Andean zones. The
Tethys and Andean range databases were combined and then the new radioisotope dates were projected into the database. The relative ages of first and last occurrences of nearly 250 stratigraphic events were recalibrated to new dates. These Andean stage ages are compared with GTS2020 ages. The recalibration of numerical ages of Andean stratigraphic markers has significant implications on durations of stages and zones as well as sedimentary rates and durations. Ages of Berriasian-Hauterivian stage boundaries recalibrated by different methods are compared.
|
|
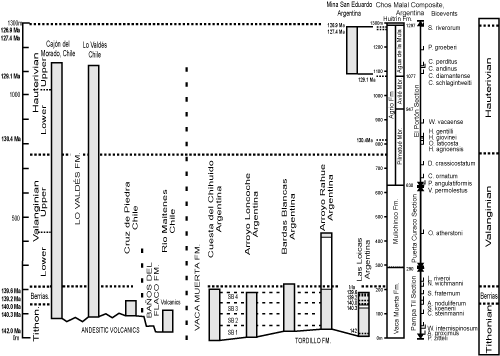
Figure 1: Location of outcrop measured sections of upper Tithonian-Hauterivan stages in Chile and Argentina that compose the ANDESCS Database. 1-Lo
Valdés, Chile; 2-Cajón del Morado, Chile; 3-Cruz de Piedra, Chile; 4-Rio
Maitenes, Chile; and Argentinian sections 5-Las Loicas; 6-Pampa Tril; 7-El
Portón; 8-Real de las Coloradas; 9-Cerro Domuyo; 10-Mina San Eduardo; 11-Arroyo
Truquico; 12-Cerro La Parva; 13-Arroyo Loncoche; 14-Cuesta del Chihuido; 15-Bardas Blancas; 16-Arroyo
Rahue; 17-Los Catuto; 18a Bajada Viejo; 18b Bajada del Agrio; 19 Arroyo
Cieneguita; 22b Puerta Curaco Section; and 23 Las Tapaderas Section. The composited section of Pampa Tril (6), Puerta Curaco (22b) and El Portón (7) is indicated by the triangle. |
Abbreviations: CA-ID-TIMS - Chemical Abrasion Isotope-Dilution Thermal Ionization Mass Spectrometry; CLS - correlation line of synchroneity; DB - database; FO/LO - first and last occurrence datums in a given section; FAD/LAD - first and last appearance datums in all database sections; GSSP - global stratotype section and point; Ma - mega-annums; MU - metric units; myr - million years duration; RS - reference section; SAR - sediment accumulation rate; U-Pb - uranium-lead.
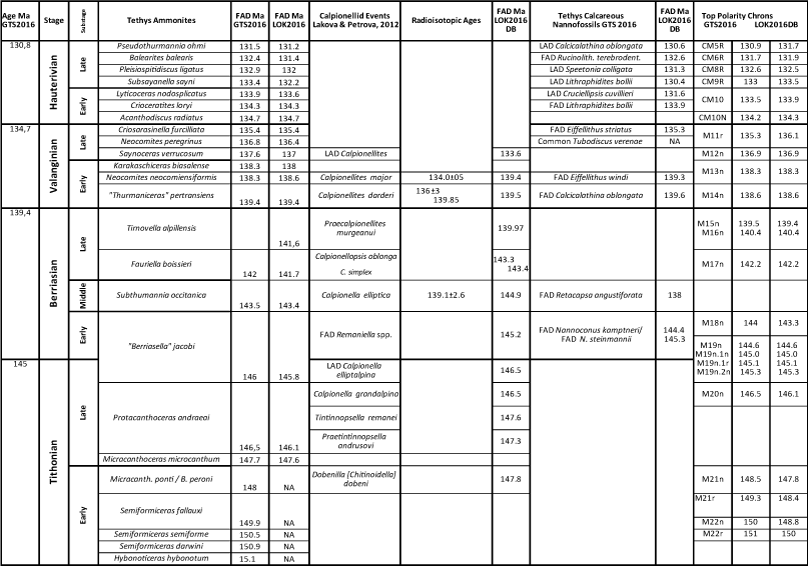
A comprehensive chronostratigraphic database of the first and last appearance datums (FAD/LAD) of ammonites, calpionellids, nannofossils, dinoflagellates, and polarity chrons in uppermost Tithonian to Maastrichtian stages from numerous public documents of outcrops and drill cores on five continents was compiled (Scott, 2014, 2019a). A subset of this database is composed of 70 Lower Cretaceous reference sections in France, Spain, Italy, Eastern Europe, North Africa, Iran, Tibet, the Atlantic basin, North and South America (Appendix 1). Included are GSSP or candidate reference sections of Berriasian to Barremian stages. This data set also includes polarity chrons M16n through M20r from nine sections in Spain, Italy and Poland and DSDP 534 core in the western Atlantic. Beginning in 2017 fossil ranges were integrated into a single database, LOK16CS DB, scaled to what then was the most recent time scale GTS2016 (Ogg et al., 2016) using the graphic correlation technique (Carney & Pierce, 1995) and the GraphCor software (Hood, 1995) (Appendix 2). GTS2020 (Gale et al., 2020) was published after this project was completed.
Bioevents and polarity chrons in the Mediterranean-Caucasian (Westernman, 2000) sections in meters/feet were cross-plotted on the Y-axis with the GTS2016 geologic time scale in mega-annums on the X-axis to create hypotheses of synchroneity between section pairs. The correlation line of synchroneity (CLS) extended the first and last species occurrences in each section (FOs, LOs) relative to ranges in other sections combining ranges in all sections, in which each taxon was present. The composited range extensions in all sections approximated first and last appearance datums (FADs, LADs) calibrated to numerical ages (Ma) (Table 3) of the 2016 Geologic Time Scale (Ogg et al., 2016). This stratigraphic experiment placed the calpionellid, nannofossil, dinoflagellate, and ammonite FADs in the predicted order relative to polarity chrons M16n through M22r (Wimbledon, 2017; Reboulet et al., 2018). The numeric ages of all taxa calibrated by this method are within less than 0.1% of the ages predicted by GTS2016.
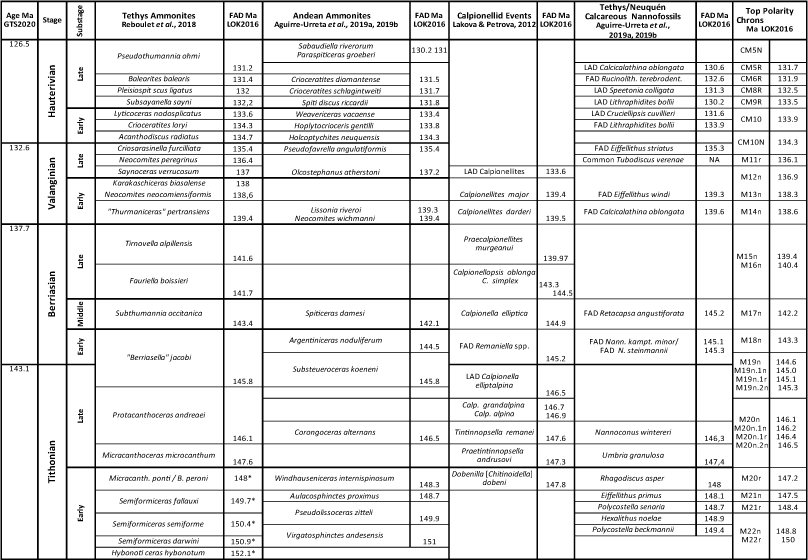
Table 3. Numerical mega-annum ages calibrated to GTS2016/2020 of Tethys ammonites, calpionellids and calcareous nannofossils correlated with Andean ammonites and polarity chrons.
|
|
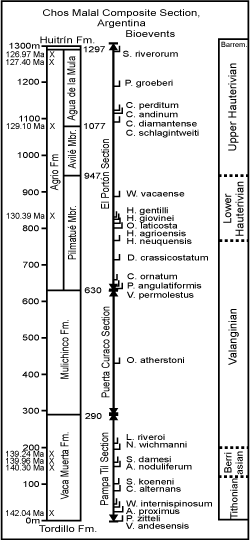
In order to construct a quantitative database of Andean Tithonian-Hauterivian biostratigraphy twenty-three stratigraphic outcrop sections were selected from among the many excellent published data. Experienced professional geologists have measured, described, sampled, and analyzed these sections for ammonites, and where possible calpionellids, dinoflagellates, nannofossils, and polarity chrons (Appendix 3). The Andes Chronostratigraphic Database, ANDESCS DB, comprises bioevent data scaled to metric units (MUs) of the Chos Malal composite reference section (Table 4).
Because no single stratigraphic section is known in the Andes that spans the upper
Tithonian-Hauterivian stages, a composited reference section was necessary in order to scale taxon ranges relative to each other. The Chos Malal reference section represents the Mendoza Group in the Neuquén Basin and was assembled by combining the Pampa Tril section at the base (Parent et
al., 2015) with the overlying Puerta Curaco section (Schwarz et al.,
2006;
Keitzmann et al., 2021a) at the contact of the Vaca Muerta and Mulichinco formations; then the El Portón section was added above at the base of the Agrio Formation (Aguirre-Urreta et al., 2015, 2017)
(Fig. 2 ![]() ). These sections are within 50 km of each other, two of which were studied by the same team and the third by a most experienced team. The Pampa Tri section exposes the Vaca Muerta shale with diverse ammonites (Parent et al., 2015; Vennari, 2016). The Puerta Curaco section spans the Vaca Muerta-Quintuco and Mulinchinco formations. The nearby El Portón section spans the upper
Valanginian-upper Hauterivian Agrio Formation, which yields ammonites, nannofossils and a succession of radioisotope ages (Aguirre-Urreta et al., 2017,
2019). These three sections document detailed biostratigraphy that correlates with
Tethys stages (Aguirre-Urreta & Rawson, 2010).
). These sections are within 50 km of each other, two of which were studied by the same team and the third by a most experienced team. The Pampa Tri section exposes the Vaca Muerta shale with diverse ammonites (Parent et al., 2015; Vennari, 2016). The Puerta Curaco section spans the Vaca Muerta-Quintuco and Mulinchinco formations. The nearby El Portón section spans the upper
Valanginian-upper Hauterivian Agrio Formation, which yields ammonites, nannofossils and a succession of radioisotope ages (Aguirre-Urreta et al., 2017,
2019). These three sections document detailed biostratigraphy that correlates with
Tethys stages (Aguirre-Urreta & Rawson, 2010).
|
|
Figure 2: Chos Malal composite section composed of three stratigraphic sections stacked at common lithostratigraphic contacts: Vaca
Muerta/Mulichinco and Mulichinco/Agrio formations to form a single reference section calibrated in meters. |
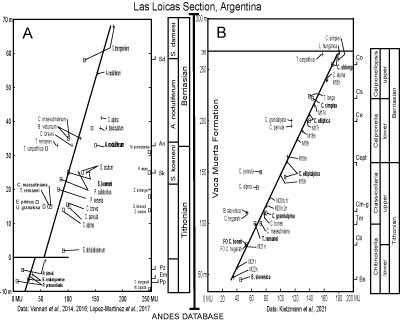
As successive sections were plotted to the reference section the metric positions of
FO/LOs were extended by the correlation line of synchroneity (CLS), which was positioned by the stratigraphic interpreter to align with known bioevents
(Fig. 3.A-B ![]() ). For example, the Las Loicas section was plotted to the Andes database and the CLS was constrained by ammonite and nannofossil bioevents
(Fig. 3.A
). For example, the Las Loicas section was plotted to the Andes database and the CLS was constrained by ammonite and nannofossil bioevents
(Fig. 3.A ![]() ). The offset in the lower part of section is an artifact of stacking
separately measured sections, the lower Tithonian section (Vennari et al.,
2016) with the upper Tithonian-Berriasian interval (Vennari et al., 2014). The FOs of many other nannofossils were previously calibrated in the Agrio Formation and they range lower in the Vaca Muerta Formation and their ranges were extended and recalibrated at lower metric positions. Several LOs (plus signs) are left of the CLS and were extended higher-younger in the database. The calpionellid bioevents and zones were integrated from the nearby Las Loicas outcrop
(Fig. 3.B
). The offset in the lower part of section is an artifact of stacking
separately measured sections, the lower Tithonian section (Vennari et al.,
2016) with the upper Tithonian-Berriasian interval (Vennari et al., 2014). The FOs of many other nannofossils were previously calibrated in the Agrio Formation and they range lower in the Vaca Muerta Formation and their ranges were extended and recalibrated at lower metric positions. Several LOs (plus signs) are left of the CLS and were extended higher-younger in the database. The calpionellid bioevents and zones were integrated from the nearby Las Loicas outcrop
(Fig. 3.B ![]() )
(Kietzmann et al., 2021b) and the Arroyo Loncoche and Cuesta del Chihuido sections (Kietzmann et al.,
2020). The composited ranges compose the ANDESCS Database
(Table 4). The stage boundaries previously have been correlated by ammonites and nannofossils (Aguirre-Urreta et al.,
2005,
2017, 2019; Vennari et al., 2014). The vertical spacing and scaling of the zones are in meters of the thickness of the reference section
(MUs) and do not measure zone durations.
)
(Kietzmann et al., 2021b) and the Arroyo Loncoche and Cuesta del Chihuido sections (Kietzmann et al.,
2020). The composited ranges compose the ANDESCS Database
(Table 4). The stage boundaries previously have been correlated by ammonites and nannofossils (Aguirre-Urreta et al.,
2005,
2017, 2019; Vennari et al., 2014). The vertical spacing and scaling of the zones are in meters of the thickness of the reference section
(MUs) and do not measure zone durations.
|
|
Figure 3: Stratigraphic correlation plots of two data sets of the Las Loicas section with the ANDESCS DB based on the Chos Malal Composite reference section (SRS) (□ signs are
FOs, + signs are LOs). Sloping correlation lines (CLS) are constrained by ammonite and nannofossil
bioevents. A. Nannofossil FO bioevents right of the CLS in the Agrio Formation will be extended into the Vaca Muerta Formation and their ranges will be recalibrated in meters of the reference section. B. Calpionellid and polarity chrons tightly constrain the
CLS. |
Table 4. Chronostratigraphic classification of ammonite zones and polarity chrons in the ANDESCS DB. Scale is metric units (MUs) in the Chos Malal composite section (SRS). Early-middle Tithonian zones after Vennari (2016); late Tithonian to Berriasian zones after Kietzmann et al. (2018); Valanginian-Hauterivian zones after Aguirre-Urreta et al. (2015, 2017, 2019); central Chilean zones after Salazar et al. (2020).
|
|
Stratigraphy of the Mediterranean-Caucasian Subrealms. The uppermost Jurassic Tithonian Stage and the Berriasian, Valanginian and Hauterivian stages of the Lower Cretaceous System time scale were initially defined in southern France, and as of this writing only the Hauterivian at La Charce, Drôme, southern France, has been designated a Global Section Stratotype Points (GSSP) (Ogg et al., 2016; Gale et al., 2020; Mutterlose et al., 2020). Reference sections of Tithonian-Hauterivian stages, substages, ammonite, calpionellid, and nannofossil zones were calibrated to GTS2016 mega-annums in the LOK2016 DB (Table 3). Tithonian-Berriasian polarity chrons were correlated with biostratigraphic zones in 23 European sections (Grabowski & Pszczólkowski, 2006; Grabowski, 2011; Grabowski et al., 2018), nine of which are in our database.
The upper Tithonian Stage is represented in part by the LOK2016 DB by the FADs of Micracanthoceras microcanthum at 147.6 Ma, Protacanthoceras andraeai at 146.1 Ma and "Berriasella" jacobi at 145.8 Ma. Two Tithonian chitinodellid calpionellid species are Dobinella [Chitinoidella] dobeni at 147.83 Ma and Bonetilla [Chitinoidella] boneti at 147.73 Ma (systematics revised by Benzaggagh, 2021). Calcareous nannofossil events span the Tithonian-Berriasian boundary as documented by Casellato and Erba (2021). The absence of lower Tithonian ammonite zones in the LOK2016 DB indicates that this interval of the database is incomplete, because no older sections are in the DB.
The Berriasian Stage is represented in southeastern France by marine carbonates and siliciclastics with ammonite, calpionellid and calcareous nannofossil zones (Wimbledon, 2017; Reboulet et al., 2018; Wimbledon et al., 2020). The base of the Berriasian has been defined by the base of the Calpionella Zone, which was defined as the "...abrupt increase in the abundance of Calpionella alpina ... (and) ... becomes the predominate element of the fauna" (Alleman et al., 1971). Wimbledon et al. defined the C. alpina Zone more precisely as the "...the turnover from Crassicollaria and large Calpionella to small orbicular Calpionella alpina (together with Crassicollaria parvula and Tintinopsella carpathica..." (2017, p. 182). These definitions differ from the FAD of Calpionella alpina (Gale et al., 2020, p. 1025), which is diachronous (Scott, 2019a). This transition is in polarity Chron M19n.2n. The commonly used ammonite species, "Berriasella" jacobi, has been revised, most of its records challenged, and the species reassigned to Strambergella (Frau et al., 2016). These authors rejected use of the "Jacobi" Zone to define base Berriasian. The most recent revision of late Tithonian-early Berriasian ammonite biostratigraphy in the Mediterranean region replaced the former "Jacobi" Zone with a refined zonation (Szives & Főzy, 2022). Most ammonite species used to subdivide the stage are endemic to the Mediterranean region so that global substage correlation is problematic (Wimbledon, 2017). Calcareous nannofossils define effective secondary biomarkers. Candidate GSSP sections considered by the former Berriasian Working Group (BWG) at Tré Maroua and Le Chouet in France, and Puerto Escańo and Rio Argos in Spain are in LOK2016 DB. Definition of the Berriasian Stage as base of Cretaceous is reviewed by Énay (2020) and Granier et al. (2020), who proposed to define base Cretaceous at base Valanginian following Oppel. A new BWG II is discussing the issue and will officially propose the base of the Berriasian Stage, its GSSP and its role in defining (or not) the J/K boundary.
The Valanginian Stage as first defined in southern France is subdivided by ammonite zones (Bulot et al., 1993; Reboulet & Atrops, 1999; Reboulet et al., 2018 and references therein; Kenjo et al., 2021). The FAD of the ammonite "Thurmaniceras" pertransiens defines base Valanginian (Martinez et al., 2013; Reboulet et al., 2018; Kenjo et al., 2021; Szives & Fözy, 2022). Closely associated is the FAD of the calpionellid Calpionellites darderi, which is proposed as the primary marker (Ogg et al., 2016; Gale et al., 2020). The Rio Argos reference section, Caravaca, Spain, yields calpionellids, ammonites, planktic foraminifera, dinoflagellates, and polarity chrons (Hoedemaker & Leereveld, 1995; Hoedemaker et al., 2016). Other reference sections in France are the Barret-le-Bas and the Angles sections with ammonites, calpionellids and cycles (Ogg et al., 2016). Marker species in each of these sections are incorporated in LOK2016 DB. Detailed ammonite and nannofossil biostratigraphy of the Vergol section, France, is proposed as the candidate GSSP (Kenjo et al., 2021).
The Hauterivian Stage GSSP is in southeastern France where the FO of the ammonite Acanthodiscus radiatus is used as the primary marker (Ogg et al., 2004, 2016; Gale et al., 2020). The La Charce outcrop section is accepted as the GSSP with detailed ammonite zones, carbon isotope chemozones and depositional cycles (Bulot et al., 1993; Gale et al., 2020; Mutterlose et al., 2020). The base of the Barremian is defined by the FAD of the ammonite Taveraidiscus hughi in the basinal Rio Argos section (Ogg et al., 2004, 2016; Gale et al., 2020). Each of these sections is in LOK2016 DB. On the carbonate shelf the Barremian is represented by benthic foraminifers and calcareous algae (Clavel et al., 2010, in the HA-BA set of sections in LOK2016 DB).
Andean Lithostratigraphy: Uppermost Jurassic and Lower Cretaceous Mendoza Group of the Neuquén Basin is composed of the lower Tithonian-Valanginian-Hauterivian Vaca
Muerta, Quintuco, Mulichinco or Chachao, and Agrio formations (Aguirre-Urreta,
2001; Leanza et al., 2011; Schwarz et al.,
2006; Kietzmann et al., 2020,
2021a). At its base the non-marine clastic Tordillo Formation disconformably overlies older Jurassic strata and conformably underlies the Tithonian-Valanginian Vaca Muerta Formation
(Schwarz et al., 2006; Naipauer et al., 2015a, 2015b;
Horton et al., 2016)
(Fig. 4 ![]() ). Lower but not lowermost Tithonian ammonites are in the basal part of the Vaca Muerta (Vennari, 2014,
2016; Kietzmann et al., 2021a). This stratigraphic succession comprises three long-term cycles of paralic sandstone to flooding organic-rich marine shale to shoaling-up marl, limestone, and sandstone (Schwarz et al.,
2006; Kietzmann et al., 2015, 2020).
). Lower but not lowermost Tithonian ammonites are in the basal part of the Vaca Muerta (Vennari, 2014,
2016; Kietzmann et al., 2021a). This stratigraphic succession comprises three long-term cycles of paralic sandstone to flooding organic-rich marine shale to shoaling-up marl, limestone, and sandstone (Schwarz et al.,
2006; Kietzmann et al., 2015, 2020).
In central Chile the Tithonian-Hauterivian Lo Valdés Formation correlates with the Mendoza Group. At its type locality near the village of Lo
Valdés, Chile, the Lo Valdés overlies Jurassic andesite and is composed of four lithological subunits, a lower interval of andesite overlain by a lower sandstone interval 73 m thick, a middle siltstone interval 214 m thick, and an upper limestone interval 252 m thick (Salazar
Soto, 2012; Salazar & Stinnesbeck,
2015, 2016; Salazar et al., 2020). The top of the Lo Valdés is unconformably overlain by volcanic breccia with limestone clasts. It is laterally equivalent in part with the marine Bańos del Flaco Formation, which overlies Kimmeridgian continental strata
(Fig. 4 ![]() ).
).
A 600 km north-south stratigraphic correlation cross section depicts the lithostratigraphic relations among the various formations
(Fig. 4 ![]() ). This transect is approximately subparallel with the north-south Malargüe and Agrio fold and thrust belts
(Horton et al., 2016; Lena et al.,
2019), which is the trend of the eastern proto-Pacific Ocean shoreline.
). This transect is approximately subparallel with the north-south Malargüe and Agrio fold and thrust belts
(Horton et al., 2016; Lena et al.,
2019), which is the trend of the eastern proto-Pacific Ocean shoreline.
The Tithonian to upper Berriasian Vaca Muerta Formation is composed of bituminous shale, calcareous shale, and sandstone (Leanza et al., 2011; Parent et al., 2011, 2015, 2017). Its thickness ranges from 100 to 1200 m. Regularly interbedded limestone and marlstone cycles approximate 21
ky, 90-120 ky and 400 ky frequencies (Kietzmann et al., 2018,
2020). Cyclostratigraphy and biostratigraphy suggest that the Tithonian duration was 5.67 myr and the Berriasian duration was 5.27 myr (Kietzmann et al., 2018). Four transgressive-regressive composite depositional sequences are composed of bundles of limestone and marlstone bounded by sequence boundaries SB 1-4. A basin-to-ramp succession extends from Cuesta del
Chihuido, Arroyo Loncoche, Bardas Blancas, and Arroyo Rahue (Fig. 4 ![]() ) (Kietzmann et al., 2018,
2020). The 280 m-thick Arroyo Loncoche section integrates ammonite biostratigraphy and polarity chrons (Iglesia Llanos & Kietzmann,
2020). At the Pampa Tril section farther south, the Vaca Muerta contains diverse ammonite faunas and is subdivided into ammonite zones, subzones and biohorizons (Parent et al., 2015;
Kietzmann et al., 2016; Vennari, 2016).
) (Kietzmann et al., 2018,
2020). The 280 m-thick Arroyo Loncoche section integrates ammonite biostratigraphy and polarity chrons (Iglesia Llanos & Kietzmann,
2020). At the Pampa Tril section farther south, the Vaca Muerta contains diverse ammonite faunas and is subdivided into ammonite zones, subzones and biohorizons (Parent et al., 2015;
Kietzmann et al., 2016; Vennari, 2016).
The Berriasian-lower Valanginian Quintuco Formation gradationally overlies the Vaca Muerta Formation and is up to 300 m-thick marine claystone, sandstone and limestone comprising several transgressive-regressive sequences (Schwarz et al., 2006; Leanza et al., 2011; Kietzmann et al., 2016; Garrido & Parent, 2017). It is overlain conformably to disconformably by the Valanginian Mulichinco Formation, which is composed mainly of paralic terrigenous clastic units and the upper member is composed of mixed siliciclastic-carbonate transgressive-regressive sequences (Schwarz et al., 2006, 2013; Garrido & Parent, 2017).
The Mulichinco is overlain conformably by the upper Valanginian to Hauterivian Agrio Formation. The Agrio is up to 540 m thick and is disconformably overlain by the regressive Barremian Huitrín Formation. The Agrio is composed of three members from lower to upper: the Pilmatué, Avilé and Agua de la Mula members (Aguirre-Urreta et al., 2017). The Pilmatué was deposited in a mixed siliciclastic-carbonate ramp setting and is composed of limestone/marl cycles suggestive of climatic control (Kietzmann & Paulin, 2019). The Avilé is a regressive-transgressive sandstone 25 to 40 m thick that disconformably overlies marine shale and grades up into a marine unit (Schwarz et al., 2016).
|
|
Figure 4: Stratigraphic cross section of Andean sections. Biozone numeric scale in metric units
(MUs) of the Chos Malal composite section. Sequence contacts (SB) in Kietzmann et al. (2018); dated ash beds in Las Loicas and El Portón sections (dotted lines) from Vennari et al. (2014) and Aguirre-Urreta et al. (2017). Stratigraphic data from Aguirre-Urreta et al., 2005, 2007, 2015, 2017;
Salazar, 2012; Vennari et al., 2014,
2016; Salazar and
Stinnesbeck, 2015; Parent et al., 2015; Kietzmann et al., 2018; Kohan Martínez et al.,
2018. |
Andean Biostratigraphy: Andean assemblage and interval biozones are based on ammonites, calpionellids, calcareous nannofossils, and calcareous dinoflagellates that are correlated with Tithonian-upper Hauterivian stages in the Mediterranean-Caucasian Subrealm (Table 3) (Aguirre-Urreta et al., 2005, 2007, 2015, 2017, 2019; Kietzmann et al., 2011, 2015; Lazo et al., 2009; Soto, 2012; Vennari et al., 2014, 2017; Parent et al., 2015, 2017; Salazar & Stinnesbeck, 2015, 2016; Vennari, 2016; Ivanova & Kietzmann, 2017; Kietzmann, 2017; Salazar et al., 2020; Iglesia Llanos and Kietzmann, 2020). In this stratigraphic experiment zones are defined by the FO of nominal species rather than basing zones on genera or assemblages.
In the Neuquén Basin five ammonite FO events in the Pampa Tril and Arroyo Loncoche sections and six other sections are correlated with the Tethys Tithonian Stage (Vennari, 2016; Parent et al., 2017; Kietzmann et al., 2018). Different correlation hypotheses correlate base Berriasian in the Mediterranean sections with the Vaca Muerta Formation. One interpretation correlates base of the Substeueroceras koeneni Zone at 101 MU with base Berriasian (Salazar & Stinnesbeck, 2016; Iglesia Llanos & Kietzmann, 2020). Alternative correlations of base Berriasian are either within the S. koeneni Zone (Vennari et al., 2014; Kietzmann et al., 2020, 2021a) or with the base of the Argentiniceras noduliferum Zone (Parent et al., 2015) at 115 Mu. The basal part of the Vaca Muerta Formation records polarity chrons M22r to M15r (Iglesia Llanos et al., 2017; Kohan Martinez et al., 2018). The top of polarity chron M19n at 112 MU in the ANDESCS DB is correlated above the Tithonian-Berriasian boundary and the FO of Calpionella alpina below at 81 Mu (Table 3). The FADs of several calcareous nannofossils that span the Tithonian-Berriasian boundary (Casellato & Erba, 2021) are slightly above the FAD of S. koeneni.
The FO of Neocomites wichmanni at 180 MU is correlated with base Valanginian (Aguirre-Urreta, 2001; Parent et al., 2015; Riccardi, 2015). It occurs together with Calpionellites darderi in the Cuesta del Chihuido and Puerta Curaco sections (Kietzmann et al., 2020). The early-late Valanginian boundary is correlated within the Olcostephanus atherstoni Zone (Aguirre-Urreta, 2001), which spans 429-472 MU (Table 4).
Base Hauterivian is correlated with the FO of Holcoptychites neuquensis at MU 772 in the Bajada Viejo section (Aguirre-Urreta et al., 2015, 2017), which is slightly above the FO of the nannofossil Retacapsa surirella at 765 MU in the El Portón section. The lower-upper boundary is at the base of the Spitidiscus riccardii Zone (Lazo et al., 2009; Aguirre-Urreta et al., 2019). The Hauterivian/Barremian boundary is correlated in the midst of the Sabaudiella riverorum Zone (Aguirre-Urreta et al., 2019; Table 3).
The first and last occurrences (FO, LO) of calcareous nannofossils have been integrated with ammonite zones of the Neuquén Basin because they support correlation with the Tethys zones (Aguirre-Urreta et al., 2005, 2007, 2019; Riccardi, 2015); they are also calibrated in the ANDESCS DB (Table 3). However, ranges of some important species are not yet fully extended in the ANDESCS DB because they are reported in single sections. A succession of upper Hauterivian nannofossils, Lithraphidites bollii, C. cuvillieri, E. striatus, and Nannoconus liguis (Aguirre-Urreta et al., 2019) is represented in the ANDESCS DB with minor changes in the order (Table 3). Ages of nannofossils in the LOK2016 DB support the correlation of the Pilmatué Member of the Agrio Formation spanning upper Valanginian to lower Hauterivian.
In central Chile the Tithonian-lower Valanginian zones are different (Salazar et al., 2020) (Table 4). At the base of the Tithonian is the Virgatosphinctes mexicanus / Pseudolissoceras zitteli Zone, and the upper Tithonian zones are the Windhauseniceras internispinosum and Micracanthoceras microcanthum / Corongoceras alternans Zone. Base Berriasian is marked by the Berriasiella jacobi Zone; middle-upper Berriasian is the Groebericeras roccardi Zone. The lower Valanginian zone is the Thurmanniceras thurmanni / Argentiniceras fasciculatus Zone.
Paleobiogeography: A brief summary of Early Cretaceous ammonite biogeography frames the different zonal schemes used in the Mediterranean and Andean regions. The biogeographic distribution of Early Cretaceous ammonoids was influenced by a complex of interrelated factors including climate, ocean temperatures and oceanic circulation (Énay, 1972; Cecca, 1998; Westermann, 2000; Page, 2008; Lehmann et al., 2015). Endemism resulted in distinct geographic ammonite assemblages although the calpionellids and calcareous nannofossils were distributed widely (López-Martínez et al., 2017a). During the Berriasian through Hauterivian ages, the Mediterranean-Caucasian Subrealm of the Tethys Realm hosted a biota distinct from the Andean Indo-Pacific Subrealm (Westermann, 2000; Page 2008; Lehmann et al., 2015). However, the Berriasian "Berriasiella", Grobericeras, Spiticeras, and some Olcostephanid ammonites occupied both subrealms (Salazar et al., 2020), although, many genera were endemic to the Andes: Andiceras, Argentiniceras, Frenguelliceras, Hemispiticeras, Cuyaniceras, and Pseudoblanfordia (Riccardi, 1988; Aguirre-Urreta et al., 2007; Parent et al., 2011; Vennari et al., 2012). During the Valanginian Age Olcostephanids were widely distributed from Mediterranean-Caucasian, Pacific to Andean basins including Neocomites, Kilianella, Sarasinella, and Thurmanniceras (Aguirre-Urreta, 1998; Aguirre-Urreta & Rawson, 1999; Rawson, 2007; Aguirre-Urreta et al., 2008a). Endemism increased in Andean basins during the latest Valanginian when common Neocomitidae genera were Pseudofavrella, Chacantuceras and Decliveites (Aguirre-Urreta & Rawson, 2003, 2010). During the Hauterivian Age Tethys Indo-Pacific genera in the Andean basins were Holcoptychites, Favrella, Jeannoticeras, and Plesiospitidiscus. The characteristic early Hauterivan genera differ from the late Hauterivian genera (Lehmann et al., 2015). These genera comprise the basis of Andean Berriasian-Hauterivian biostratigraphy (Aguirre-Urreta & Rawson, 2003, 2010).
Andean Magnetostratigraphy: Polarity chrons are key to correlating Andean biozones with Tethys Mediterranean stages. The Tithonian-Berriasian polarity sequence in the Neuquén Basin is defined in the Vaca Muerta Formation at Arroyo Loncoche (Kietzmann et al., 2018b; Iglesia Llanos & Kietzmann, 2020) and at the Los Catutos section (Kohan Martínez et al., 2018). The Tithonian-Berriasian boundary has been consistently correlated in the middle of polarity chron M19n.2n (Ogg et al., 2016; Wimbledon et al., 2020). In the Neuquén Basin this unit correlates with the lower part of the Substeueroceras koeneni Zone in the Vaca Muerta Formation (Kietzmann et al., 2018b). The lower-upper Tithonian boundary has been correlated with polarity chron M20n (Ogg et al., 2016) and at the base of the Windhauseniceras internispinosum Zone (Kietzmann et al., 2018b; Kohan Martínez et al., 2018) (Table 3). The new radioisotpic age of 140.3 Ma projects at 112 MUs in the Vaca Muerta Formation, which correlates with polarity chron M19n (Table 3).
Tithonian-Hauterivian Radioisotope Dates: In the past thirteen years numerous new radioisotopic dates spanning the Tithonian-Hauterivian stages have been added to previous ages in GTSS2016 and GTS2020 (Table 2). Prior to that date only five numerical ages had been published and numerical ages in the Geologic Time Scale were estimated by the polarity time scale (Ogg et al., 2012, 2016). However, the new Argentinian dates would significantly alter the Early Cretaceous time scale by 1-4 myr. Most new dates are based on euhedral zircon crystals extracted from ash beds. Such dates are quite precise because selected crystals were apparently deposited penecontemporaneously and were not reworked or displaced down-section or altered (Aguirre-Urreta et al., 2015, 2017, 2019; Lena et al., 2019). Each date was related to a bioevent and its correlative stage (Table 2) (Aguirre-Urreta et al., 2019; Lena et al., 2019).
Radioisotopic U/Pb dates of detrital zircon crystals in the Tordillo Formation underlying the Tithonian Vaca Muerta Formation range in age from 275 Ma to 144 Ma and indicate that the Jurassic Andean arc was the primary sediment source and that older igneous sources contributed minor amounts (Naipauer et al., 2015c). Dates from the basal interval of the Tordillo of 149.5±1.2 Ma and from a higher bed of 143.0±1.0 Ma (Horton et al., 2016; Naipauer et al., 2015a, 2015b) suggest that the Tithonian Stage may be younger than 152.1 Ma as in GTS2016.
Five volcanic tuff beds in the Tithonian-Berriasian Vaca Muerta Formation at the Las Loicas section (Vennari et al., 2014; Lena et al., 2019) are dated by U-Pb zircons or the Bayesian age-depth model. A date of 142.04±0.17 Ma is in the Crassicolaria Zone (Table 2). Four dates are associated with uppermost Tithonian nannofossil FADs: 140.6±0.4 Ma, 140.54±0.34 Ma, 140.34±0.18 and 140.22±0.13 Ma (Table 2). These dates are interpolated into the ANDESCS DB database by their co-occurrence with ammonites, calpionellids and nannofossils (Table 3).
The base of the uppermost Tithonian-lower Berriasian Substeueroceras koeneni Zone in the Las Loicas section underlies an ash bed dated at 140.34±0.08 Ma, which is within polarity Chron M19n. The middle Berriasian Argentiniceras noduliferum Zone and the FO of Nannoconus kamptneri minor are bracketed by the dates of 140.34±0.08 Ma and 139.96±0.06 Ma. The Spiticeras damesi Zone is dated at 139.24±0.05 Ma. The FO of basal Valanginian C. darderi and N. wichmanni are projected directly above these dates. These ages are significantly younger than calibrated in GTS2016 and in LOK2016 DB (Scott, 2019a). Based on these radioisotopic dates, Vennari et al. (2014) and Lena et al. (2019) proposed that the numerical age of the base Berriasian should be 141.0 Ma, which is four myr younger than in GTS2016 (Ogg et al., 2016) and about two myr younger than 143.1 Ma in GTS2020.
In the Valanginian-Hauterivian Agrio Formation, zircons from four ash beds date Hauterivian biozones
(Fig. 4 ![]() ) (Aguirre-Urreta et al., 2015, 2017,
2019; Kohan Martínez et al., 2017; Rawson et al., 2017). The Olcostephanus laticosta Zone in the middle of the Pilmatué Member is dated at 130.39±0.16 Ma (Aguirre-Urreta et al., 2015). The tuff bed in the Agua de la Mula Member about 7 m above the top of the Avilé Member in the Spitidiscus riccardii Zone was first dated at 132.5±1.3 Ma by SHRIMP U-Pb on zircons (Aguirre-Urreta et al., 2015) and subsequently a CA-ID TIMS date at 129.09±0.04 Ma. The upper part of the Paraspiticeras groeberi Zone was dated at 127.42±0.03 Ma (Aguirre-Urreta et al., 2017) and the Sabaudiella riverorum zone that spans the Hauterivian-Barremian boundary was dated by CA-ID-TIMS at 126.97 +/- 0.04 Ma (Aguirre-Urreta et al.,
2019).
) (Aguirre-Urreta et al., 2015, 2017,
2019; Kohan Martínez et al., 2017; Rawson et al., 2017). The Olcostephanus laticosta Zone in the middle of the Pilmatué Member is dated at 130.39±0.16 Ma (Aguirre-Urreta et al., 2015). The tuff bed in the Agua de la Mula Member about 7 m above the top of the Avilé Member in the Spitidiscus riccardii Zone was first dated at 132.5±1.3 Ma by SHRIMP U-Pb on zircons (Aguirre-Urreta et al., 2015) and subsequently a CA-ID TIMS date at 129.09±0.04 Ma. The upper part of the Paraspiticeras groeberi Zone was dated at 127.42±0.03 Ma (Aguirre-Urreta et al., 2017) and the Sabaudiella riverorum zone that spans the Hauterivian-Barremian boundary was dated by CA-ID-TIMS at 126.97 +/- 0.04 Ma (Aguirre-Urreta et al.,
2019).
Numerical ages in the LOK2016 DB are constrained by nine radioisotopic dates (Table 2).
Dates of Valanginian Stage calpionellids and calcareous nannofossils in Mexico, California and Tibet range between 139.85 and 134.0 Ma.
The upper lower Berriasian Calpionella elliptica Zone in the Lower Tamaulipas Formation in Morelos, Mexico, is dated at 140.512±0.031 Ma by U-Pb zircon CA-ID-TIMS from an ash bed (Lena et al., 2019). This date suggests that the base of the Berriasian Stage must be older than proposed by Lena et al. (2019).
The Berriasian/Valanginian boundary in the Lower Tamaulipas Formation in eastern Mexico is dated at 139.85 Ma by 87Sr/86Sr of a limestone 0.4 m above the FO of Calpionellites darderi (López-Martínez et al., 2017b).
The overlying upper Valanginian Calpionellites major Subzone is dated at 134.0±0.5 Ma by U-Pb of zircons from a felsic tuff. Thus, the duration of the Valanginian is at least 5.85 myr compared to 5.1 myr in GTS2020 (Gale et al., 2020).
An uppermost Berriasian-lowermost Valanginian calpionellid assemblage in the Pimienta Formation near San Luis Potosí overlies a bentonite bed, from which zircons were dated by U-Pb at 139.1±2 Ma (López-Martínez et al., 2015, Table 1), although, the average age of 20 "best ages" is 141.17 Ma.
In the California Coastal Range in the Great Valley Sequence, zircons from two tuff beds 64.6 m apart were dated by U-Pb at 137.1±1.6/-0.6 Ma (Bralower et al., 1990). The lower tuff bed directly underlies the Valanginian assemblage of Cretarhabdus angustiforatus and a few meters above are the FOs of Micrantholithus hoschulzii and Rhagodiscus nebulosus.
In southern Tibet the uppermost Tithonian-Berriasian-Valanginian succession was recognized by ammonite and calcareous nannofossil assemblages (Liu et al., 2013; Wan et al., 2011). Ash beds yielded zircons dated from 140.0±1.3 to 141.8±1.2 Ma by SIMS U-Pb.
An ash bed dated at 141±1 is bracketed by the FOs of three upper Tithonian-Valanginian calcareous nannofossils.
In a separate Tibetan section, an ash bed overlying the C. oblongata Zone is dated at 136 Ma (Wan et al., 2011).
The age of the base Albian Stage is constrained by a date of 113.1±0.3 Ma by 206Pb/238U of zircons from an ash bed in the Gault Formation, Vöhrum, Germany (Selby et al., 2009), which in GTS2020 is 113.2 (Gale et al., 2020).
Correlation of Andean zones with Mediterranean Tithonian-Hauterivian Stages: Standard European stage boundaries can be correlated with the Andean sections by means of nannofossils, calpionellids and polarity chrons that are in both the LOK2016 DB and the ANDESCS DB. Stratigraphic positions in the former database are in mega-annums and in the latter database positions are scaled in meters (MUs) of the Chos Malal reference section.
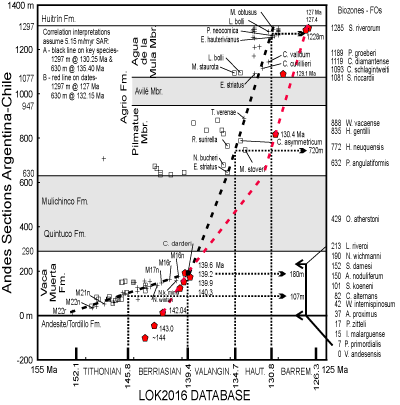
The two data sets were correlated by plotting the LOK2016 DB in MA on the X-axis, and the ANDESCS DB on the Y-axis in MUs
(Fig. 5 ![]() ). The black correlation lines (CLS) on the X/Y plots project stage boundaries defined in European reference sections with the Andean standard ammonite zones. On the right side of the plot are the FOs of Andean ammonites and their numerical ages in Ma are derived by projecting to the Mediterranean data by the CLS.
). The black correlation lines (CLS) on the X/Y plots project stage boundaries defined in European reference sections with the Andean standard ammonite zones. On the right side of the plot are the FOs of Andean ammonites and their numerical ages in Ma are derived by projecting to the Mediterranean data by the CLS.
The first correlation hypothesis in the Vaca Muerta Formation (black, bold, dashed CLS) is constrained by tops of polarity chrons and the Agrio Formation is correlated by fossil FADs or LADs
(Fig. 5 ![]() ). The plot has three segments at two bends, and the contact between the Quintuco and Mulichinco formations separates segments three and four. The lower segment in the Vaca Muerta Formation is constrained by the tops of polarity chrons M22n through M16n (plus signs). Nannofossil and calpionellid FOs (squares) also constrain the line including the FO of Calpionellites darderi
(Fig. 5
). The plot has three segments at two bends, and the contact between the Quintuco and Mulichinco formations separates segments three and four. The lower segment in the Vaca Muerta Formation is constrained by the tops of polarity chrons M22n through M16n (plus signs). Nannofossil and calpionellid FOs (squares) also constrain the line including the FO of Calpionellites darderi
(Fig. 5 ![]() ). Several Andean FO bioevents are above and left of the line because they have not been recorded lower in the Vaca Muerta; conversely several FOs to the right of the line (not shown) will be extended older in the LOK2016 DB, which incorporates few Tithonian sections and species. The second CLS segment spans the upper part of the Vaca Muerta and the Quintuco and Mulichinco formations; it is unconstrained by bioevents because none of the fossiliferous sections of the Quintuco and Mulichinco formations are in the ANDES Database, although a few Berriasian and lower Valanginian ammonites are in the Quintuco (Schwarz et al.,
2006; Garrido & Parent,
2017). The Mulichinco overlies the L. riveroi Zone and underlies the P. angulatiformis Zone and in its uppermost intervals O. atherstoni and O. permolestus are present where the Mulichinco grades into the Agrio (Schwarz et al., 2006).
). Several Andean FO bioevents are above and left of the line because they have not been recorded lower in the Vaca Muerta; conversely several FOs to the right of the line (not shown) will be extended older in the LOK2016 DB, which incorporates few Tithonian sections and species. The second CLS segment spans the upper part of the Vaca Muerta and the Quintuco and Mulichinco formations; it is unconstrained by bioevents because none of the fossiliferous sections of the Quintuco and Mulichinco formations are in the ANDES Database, although a few Berriasian and lower Valanginian ammonites are in the Quintuco (Schwarz et al.,
2006; Garrido & Parent,
2017). The Mulichinco overlies the L. riveroi Zone and underlies the P. angulatiformis Zone and in its uppermost intervals O. atherstoni and O. permolestus are present where the Mulichinco grades into the Agrio (Schwarz et al., 2006).
In the Agrio Formation two correlation interpretations are reasonable. The black CLS A is constrained by nannofossil FOs and LOs, and it will extend fewer LOs than alternate CLSs. At its base is the FO of Eiffellithus striatus, which is a well-established upper Valanginian bioevent in both databases and GTS2016 (Bown et al., 1998). The FOs of a number of other nannofossils are left of the CLS and would be projected lower in Andean strata but have yet to documented there. The upper part of the black CLS A is tightly constrained by the LOs of the nannofossils Tubodiscus verenae, Cruciellipsis cuvillieri, and Lithraphidites bolli. To the right of the CLS is a stack of numerous other nannofossils that have longer ranges. To the left side of CLS A is a smaller group of LOs that are younger in the Andes than in sections in the LOK2016 DB. Their range ages will be extended by the Andean data set.
The second correlation interpretation places the red CLS through the nine
radioisotopic ages (red polygons,
Fig. 5 ![]() ). The correlation line in the lower part of the Vaca Muerta Formation would date it much younger than ages of polarity chrons in GTS2016 and GTS2020. Also, the Agrio Formation would be younger than projected by the LOK2016 DB.
). The correlation line in the lower part of the Vaca Muerta Formation would date it much younger than ages of polarity chrons in GTS2016 and GTS2020. Also, the Agrio Formation would be younger than projected by the LOK2016 DB.
|
|
Figure 5: Correlation plot of LOK2016 DB (X-axis) in
mega-annums (Ma) with ANDESCS DB (Y-axis) in metric units (MUs). Berriasian,
Valanginian, Hauterivian, and Barremian stage boundaries defined at Tethys reference sections. Polygons are radioisotopic dates of Andean ash beds. Black dashed correlation lines are tied to polarity chrons M22n to M16n and calcareous
nannofossils. Dotted lines project standard stage boundaries into Andean sections. |
The slopes of the CLSs represent sediment accumulation rates (SARs), not sedimentation rates because these rocks have been compacted and lithified. The SAR of the Vaca Muerta Formation increases from 0.050 mm/kyr to 0.344 mm/kyr. The SAR of the combined Mulichinco and Agrio formations is estimated at 0.633 mm/kyr by CLS A and 0.851 mm/kyr. Because the base of the Mulichinco varies from conformable to unconformable across the basin (Schwarz et al., 2006), the duration of the hiatus in
Figure 5 ![]() is not estimated.
is not estimated.
The black CLSs of this stratigraphic experiment correlates base Berriasian at MU 107 above the FO of S. koeneni at MU 101. This is consistent with correlations that place base Berriasian within the S. koeneni Zone (Leanza et al., 2011; Salazar Soto, 2012; Salazar & Stinnesbeck, 2015; Kietzmann et al., 2018, 2021a). Base Valanginian projects at MU 180 at the base of the N. wichmanni Zone and FO of C. darderi, which is consistent with projections by Leanza et al. (2011) and Kietzmann et al. (2018) among others. Base of the Hauterivian Stage is projected at MU 720 below the base of the Holcoptychites neuquensis Zone at 772 Mu, which is lower than previous correlations (Aguirre-Urreta et al., 2017, 2019). Base of Barremian Stage as defined in the Mediterranean sections, projects into the S. riverorum Zone below the unconformable contact between the Agrio and Huitrín formations consistent with previous correlations (Aguirre-Urreta et al., 2017).
The base of the Mulichinco Formation, base of the Middle Mendoza Subgroup, is bracketed in the middle part of the Valanginian Stage (Kietzmann et al., 2020) and may correlate with the 136.4 Ma sea-level event of Haq (2014) and the intra-Valanginian unconformity in the Texas Gulf Coast (Scott, 2019b). The unconformity at base of Avilé Member of the Agrio Formation, base of the Upper Mendoza Subgroup, may correlate with the 132.8 Ma or the 131.8 Ma mid Hauterivian sea-level event (Haq, 2014).
The X/Y plot of the Andean data with the LOK2016 data correlates Andean biozones with Tethys biozones and interpolates numerical ages for the FOs (Table 4). These correlations generally reproduce those of the Tithonian-Berriasian (Riccardi, 2015). The calpionellid zonal schemes of Riccardi differ in some details from that of Lakova and Petrova (2013), so the correlations differ. However, both schemes correlate similarly with polarity chrons. The calcareous nannofossil zones in both regions are based on FAD/LADs and reproduce those of Aguirre-Urreta et al. (2019).
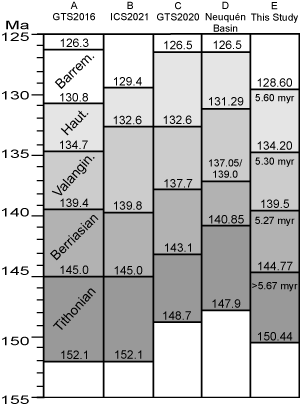
Stage and Ammonite Zone Durations: Calculating durations of Tithonian and Lower Cretaceous stages and associated ammonite zones by different methods tests numerical ages of stage boundaries
(Fig. 6 ![]() ). Cyclostratigraphic astrochronologic calibration of stage durations is an important tool to calibrate numerical ages of stage boundaries. However, the durations vary depending on the stratigraphic sections, the boundary criteria and the methods. The GTS2020 time scale measures the duration of base Berriasian to top Hauterivian at 16.5 myr
(Fig. 6.C
). Cyclostratigraphic astrochronologic calibration of stage durations is an important tool to calibrate numerical ages of stage boundaries. However, the durations vary depending on the stratigraphic sections, the boundary criteria and the methods. The GTS2020 time scale measures the duration of base Berriasian to top Hauterivian at 16.5 myr
(Fig. 6.C ![]() ). New dates from the Neuquén Basin measure the duration of this interval at 14.35 Myr
(Fig. 6.D
). New dates from the Neuquén Basin measure the duration of this interval at 14.35 Myr
(Fig. 6.D ![]() ). This stratigraphic experiment estimates the duration of the upper part of the Tithonian Stage into the lower part of the Valanginian Stage in the Vaca Muerta and Quintuco formations to be about 15 myr
(Fig. 5
). This stratigraphic experiment estimates the duration of the upper part of the Tithonian Stage into the lower part of the Valanginian Stage in the Vaca Muerta and Quintuco formations to be about 15 myr
(Fig. 5 ![]() ).
).
Cyclostratigraphic astrochronology of the Vaca Muerta Formation calculated the durations of the Tithonian Stage at least 5.67 myr and the Berriasian Stage at 5.27 myr
(Fig. 6.E ![]() ) (Kietzmann et al., 2018,
2020) compared with durations of 6.1 and 5.4 myr in GTS2020
(Fig. 6.C
) (Kietzmann et al., 2018,
2020) compared with durations of 6.1 and 5.4 myr in GTS2020
(Fig. 6.C ![]() ; Gale et al., 2020; Hesselbo et al.,
2020).
; Gale et al., 2020; Hesselbo et al.,
2020).
The duration of the Valanginian Stage may have been up to 6 myr duration based on the Sr isotope date of 139.85 Ma and the U-Pb zircon date of 134.0 Ma in eastern Mexico
(Table 2). However, radio-astrochronology calibrated the duration at 5.08 myr in French and Spanish reference sections using the FO of Tirnovella pertransiens as the base Valanginian (Martinez et al., 2013, 2015). A slightly shorter duration of 4.74 myr was measured in the Angles section, France, where base Valanginian is defined at FO Calpionellites darderi and its top at FO of Acanthodiscus radiatus (data from Busnardo et al., 1979,
Fig. 8E). The mid-point duration of this range is 5.3 myr, which is used here to calculate stage boundary ages
(Fig. 6.E ![]() ).
).
The duration of the Hauterivian Stage ranges from 5.96 to 5.21 myr. In French reference sections four cyclostratigraphic astrochronologic studies measured the duration from 5.3 myr to 5.93±0.41 myr (Martinez et al., 2015). In the Neuquén Basin at El Portón in the Agrio Formation precise CA-ID TIMS U-Pb radioisotopic dates, biostratigraphy and astrochronology of bedding cycles calculated the duration of the Hauterivian at 5.21±0.08 myr (Aguirre-Urreta et al., 2019). Low-frequency eccentricity cycles of the Agrio Formation at Arroyo Loncoche calculated the duration of the Hauterivian at 5.96 myr (Kietzmann et al., 2020). The mid-point age of 5.585 myr is rounded to 5.6 myr as Hauterivian duration.
The graphic correlation experiment of the Neuquén Basin data presents two possible interpretations of the Hauterivian duration
(Fig. 5 ![]() ). Correlation line A estimates the duration at 5.15 myr, close to the proposed 5.21 myr duration. It is constrained at base of the Agrio Formation by the FO of Eiffellithus striatus with an age of 135.30 Ma
(Appendix 2). The FAD of E. striatus is uppermost Valanginian (GTS2016, Ogg et al.,
2016). The top of correlation line A at 130 Ma is constrained by a cluster of late Hauterivian nannofossil LADs (Bown et al., 1998): Cruciellipsis cuvillieri, Eiffellithus striatus and Lithraphidites bollii (Fig. 5
). Correlation line A estimates the duration at 5.15 myr, close to the proposed 5.21 myr duration. It is constrained at base of the Agrio Formation by the FO of Eiffellithus striatus with an age of 135.30 Ma
(Appendix 2). The FAD of E. striatus is uppermost Valanginian (GTS2016, Ogg et al.,
2016). The top of correlation line A at 130 Ma is constrained by a cluster of late Hauterivian nannofossil LADs (Bown et al., 1998): Cruciellipsis cuvillieri, Eiffellithus striatus and Lithraphidites bollii (Fig. 5 ![]() ).
).
Correlation hypothesis B calibrates the duration at 5.18 myr. Correlation line B is constrained by the set of four radioisotopic dates with an age of 132.15 Ma at the base of the Agrio and 126.97 Ma at its top. These durations assume a uniform rate of accumulation of the Avilé Member with no significant hiatus, which assumptions need to be fully evaluated. Longer durations could be represented between these two hypotheses by correlation lines with lower slopes. The range of durations calculated by astrochronology from 5.21 myr to 5.93 myr suggests that the assumptions should also be reevaluated as noted by Martinez et al. (2015).
Kietzmann et al. (2018, 2020) estimated the durations of Tithonian-Berriasian ammonite zones in the Vaca Muerta Formation by the number of 405 myr-period depositional cycles. Durations calculated by this method are compared to durations measured between the FADs of each zonal species in LOK2016 DB (Table 5, Appendix 2). Most zonal durations estimated by the graphic method are longer than those measured by cyclostratigraphy because the graphic method relates the zonal ages to ages of polarity chrons in GTS2016. In contrast, zonal durations measured using the Andean radioisotopic dates are much shorter.
Table 5. Comparison of durations of Andean ammonite zones with revisions by new radioisotopic dates.
| Zone | Durations kyr | |||
| Kietzmann, 2018 | LOK2016DB | Revised Age Ma | Duration | |
| N. wichmanni | 139.51 | |||
| S. damesi | 1.62 | 2.09 | 140.09 | 0.58 |
| A. noduliferum | 0.81 | 1.61 | 140.14 | 0.05 |
| S. koeneni | 2.43 | 0.95 | 140.7 | 0.56 |
| C. alternans | 1.21 | 2.11 | 141.28 | 0.58 |
| W. interspinosum | 1.21 | 1.63 | 141.52 | 0.24 |
| A. proximus | 0.61 | 0.65 | 141.59 | 0.07 |
| P. zitteli | 0.61 | 1.38 | 141.74 | 0.15 |
| V. andesensis | 0.81 | 1.56 | 141.91 | 0.17 |
Since 2016 four studies have revised the Berriasian-Hauterivian time scale
(Fig. 6.A-D ![]() ). New radioisotopic dates and cyclostratigraphic
astrochronologic durations from the Neuquén Basin revise the ages of these stages. If the age of one stage is dated consistently be several methods, it can anchor ages of other stages by adding or subtracting stage durations. Using this method, the ages of other stage boundaries are proposed
(Fig. 6.E
). New radioisotopic dates and cyclostratigraphic
astrochronologic durations from the Neuquén Basin revise the ages of these stages. If the age of one stage is dated consistently be several methods, it can anchor ages of other stages by adding or subtracting stage durations. Using this method, the ages of other stage boundaries are proposed
(Fig. 6.E ![]() ).
).
The base of the Valanginian Stage in Mediterranean sections is
consistently defined by the FAD of "Thurmanniceras" pertransiens and alternatively by Calpionellites darderi (Reboulet et al., 2018) and secondarily by calcareous nannofossil biomarkers. The dates range between 140 Ma and 139 Ma in Argentina, Mexico, California, and Tibet
(Table 2). The mid-point date of 139.5 Ma is used here as the numerical age of the Berriasian/Valanginian boundary. In the Neuquén Basin base Valanginian correlates with the base of the Neocomites wichmanni Zone, which is projected to range in age from 139.45 to 139.16 Ma by the graphical plot
(Fig. 5 ![]() ).
).
By adding the astrochronologically derived durations of the Berriasian (5.27 myr) and Tithonian (5.67 myr) to 139.50, a recalibrated age of base Berriasian is 144.77 Ma and Tithonian is 150.44 Ma
(Fig. 6.E ![]() ). New radioisotope dates of the two lowermost Berriasian zones that span Tithonian-Berriasian, Nannoconnus steinmanni minor and Argentiniceras noduliferum, are younger ranging from 140.7 Ma to 137.9 Ma
(Table 2). Lena et al.
(2019,
Fig. 4) estimated the age of this boundary between 141.0 and 140.7 Ma below an ash bed dated at 140.34±0.18 Ma
(Table 2). The age of 152.1 Ma at base of the Tithonian Stage
(Fig. 6.A
). New radioisotope dates of the two lowermost Berriasian zones that span Tithonian-Berriasian, Nannoconnus steinmanni minor and Argentiniceras noduliferum, are younger ranging from 140.7 Ma to 137.9 Ma
(Table 2). Lena et al.
(2019,
Fig. 4) estimated the age of this boundary between 141.0 and 140.7 Ma below an ash bed dated at 140.34±0.18 Ma
(Table 2). The age of 152.1 Ma at base of the Tithonian Stage
(Fig. 6.A ![]() ; Ogg et al.,
2016) was supported by astronomical calibration (Huang, 2018). GTS2020 revised the age to 149.24 Ma (Hesselbo et al.,
2020). A new radioisotopic date of 147.112±0.078 Ma in the Tordillo Formation 1.5 m below the lower but not lowest Tithonian Virgatosphinctes andesensis Zone in the Vaca Muerta Formation is consistent with the GTS2020 age
(Table 2; Lena et al., 2019, p. 10).
; Ogg et al.,
2016) was supported by astronomical calibration (Huang, 2018). GTS2020 revised the age to 149.24 Ma (Hesselbo et al.,
2020). A new radioisotopic date of 147.112±0.078 Ma in the Tordillo Formation 1.5 m below the lower but not lowest Tithonian Virgatosphinctes andesensis Zone in the Vaca Muerta Formation is consistent with the GTS2020 age
(Table 2; Lena et al., 2019, p. 10).
The base of the Hauterivian Stage is recalibrated at 134.20 Ma by subtracting the Valanginian duration of 5.30 myr from 139.50 Ma. However, a range of radioisotopic dates between 132 to 130.5 Ma near the stage base
(Table 2) is younger even than the GTS2020 age of 132.6 Ma
(Fig. 6.C ![]() ). In the Neuquén Basin base Hauterivian is correlated with the base of the Holcoptychites neuquensis Zone (Aguirre-Urreta, 2001); the FO of this taxon is dated at 131.16 Ma (Aguirre-Urreta et al.,
2019). An alternative age of 131.96±1.0 Ma was derived from a radioisotopically dated tuff bed in the Neuquén Basin constrained biostratigraphically (Martinez et al., 2015). The graphic plot to the new radioisotopic date of 130.40 Ma
(Fig. 5
). In the Neuquén Basin base Hauterivian is correlated with the base of the Holcoptychites neuquensis Zone (Aguirre-Urreta, 2001); the FO of this taxon is dated at 131.16 Ma (Aguirre-Urreta et al.,
2019). An alternative age of 131.96±1.0 Ma was derived from a radioisotopically dated tuff bed in the Neuquén Basin constrained biostratigraphically (Martinez et al., 2015). The graphic plot to the new radioisotopic date of 130.40 Ma
(Fig. 5 ![]() ) would recalibrate the base age at 130.49 Ma. A new date of 130.39±0.16 Ma in the middle of the N. neuquensis Zone in the Pilmatúe Member of the Agrio Formation
(Table 2) is consistent with these ages. This new data suggests an age of 132 to 131 Ma for base Hauterivian, thus the duration of the Valanginian would be longer than calculated. The incompatibility between Hauterivian ages derived by stage durations and radioisotope dates is yet to be resolved.
) would recalibrate the base age at 130.49 Ma. A new date of 130.39±0.16 Ma in the middle of the N. neuquensis Zone in the Pilmatúe Member of the Agrio Formation
(Table 2) is consistent with these ages. This new data suggests an age of 132 to 131 Ma for base Hauterivian, thus the duration of the Valanginian would be longer than calculated. The incompatibility between Hauterivian ages derived by stage durations and radioisotope dates is yet to be resolved.
The top of the Hauterivian is recalibrated at 128.60 Ma by subtracting the mean duration of 5.60 myr from the recalibrated age of 134.20 Ma
(Fig. 6.E ![]() ). In the Neuquén Basin top Hauterivian correlates approximately within the Sabaudiella riverorum Zone at the top of the Agua de la Mula Member of the Agrio Formation. The FO of S. riverorum coincides with a new radioisotopic date of 126.97 Ma. Martinez et al.
(2012, 2015) dated top Hauterivian at 126.02±1.0 Ma by cyclostratigraphy.
). In the Neuquén Basin top Hauterivian correlates approximately within the Sabaudiella riverorum Zone at the top of the Agua de la Mula Member of the Agrio Formation. The FO of S. riverorum coincides with a new radioisotopic date of 126.97 Ma. Martinez et al.
(2012, 2015) dated top Hauterivian at 126.02±1.0 Ma by cyclostratigraphy.
|
|
Figure 6: Comparison of five recent time scales of Tithonian to Barremian stages. Column A, Ogg et al.
(2016); B, International Commission on Stratigraphy (Cohen et al.,
2021); C, Gale et al. (2020); D, composited time scale of
Martinez et al. (2013, 2015), Kietzmann et al. (2018),
Aguirre-Urreta et al. (2019), and
Lena et al. (2019); E, alternative time scale based on stage durations anchored on base Valanginian dated at 139.50Ma. |
The graphic correlation experiment of twenty-three sections in the Andean part of the Indo-Pacific Subrealm span middle Tithonian to Hauterivian stages and integrates ranges of 254 species, sequence boundaries, polarity chrons, and radioisotopic ages that compose the ANDESCS DB. This database accurately reproduces the order of the Andean ammonite zones and places them in a relative metric scale of the Chos Malal reference section. This composite of three measured sections represents continuous deposition throughout this time interval in the Neuquén Basin. This achievement demonstrates that the ANDESCS DB is reliable so that correlation with standard reference sections in the Mediterranean-Caucasian Subrealm will produce meaningful results. A larger database of 70 sections and 877 stratigraphic markers primarily in the Mediterranean-Caucasus Subrealm compose the LOK2016 DB and prior to publication of GTS2020 was calibrated to GTS2016. This database contains the standard reference sections of the Berriasian, Valanginian and Barremian stages and the Hauterivian GSSP.
The X/Y plot of the LOK2016 DB to ANDESCS DB projects boundaries of the Berriasian, Hauterivian and Barremian stages as defined in the Mediterranean region into the ANDESCS DB. This stratigraphic experiment confirms the approximate correlation of stages defined by endemic ammonites and cosmopolitan calcareous nannofossils. The FO of Substeueroceras koeneni is latest Tithonian. The base of the Valanginian correlates with the FOs of Neocomites wichmanni and Calpionellites darderi. These two bioevents are younger than three new upper Berriasian dates that average 139.58 Ma, which is consistent with an age of 139.50 Ma at base Valanginian. This age of the base Valanginian defined by Calpionellites darderi is confidently confirmed by multiple dates in Argentina, Mexico, Tibet, and California. The base of the Hauterivian projects between the FOs of Pseudofavrella angulatiformis and Holcoptychites neuquensis. Top of the Hauterivian Stage is projected into the uppermost part of the Agrio Formation in the Sabaudiella riverorum Zone.
A revised time scale of the Tithonian to Hauterivian stages is recalibrated by adding or subtracting stage durations from the age of base Valanginian Stage, which is dated consistently by various methods in widely separate sections. Durations of the have been measured by different methods in both subrealms, so they are reliable. The age of the Tithonian base is proposed at 150.40 Ma, base Berriasian Stage at 144.77 Ma, base Valanginian at 139.50 Ma, base Hauterivian at 134.20 Ma, and top Hauterivian at 128.60 Ma.
The new radioisotopic ages in the Neuquén Basin would result in several significant differences if adopted. The ages of most of the biostratigraphic and magnetostratigraphic events would be recalibrated much younger. The duration of the Jurassic would increase by up to 4 myr. The rates of sediment accumulation would increase dramatically. The durations of ammonite zones would be reduced to unreasonable numbers. The precise ages of Tithonian to Barremian stage boundaries will continue to evolve as new data become available from other localities.
This stratigraphic correlation experiment was possible only because of superb research by the international community of Cretaceous stratigraphers, biostratigraphers, magnetostratigraphers, and radioisotopic geochemists, who have documented an enormous amount of high quality data during many decades. I am particularly grateful to the late Luc G. Bulot for sharing his original ammonite data from the French sections and encouraging the compilation of the Cretaceous database. The methodology of quantitative stratigraphy by graphic correlation was pioneered by Alan Shaw and the software by Kenneth Hood. The former Berriasian Working Group (International Subcommission on Cretaceous Stratigraphy) under the leadership of W.A.P. Wimbledon has produced a large volume of reliable and essential stratigraphic data spanning the Tithonian-Berriasian stage boundary in a global set of sections (Wimbledon, 2017; Wimbledon et al., 2020). Granier et al. (2020) have thoughtfully reviewed the history and issues surrounding definition of the Jurassic-Cretaceous boundary. Helpful reviews by Dr. Beatriz Aguirre-Urreta, Dr. Diego A. Kietzmann, Dr. Ottilia Szives, Dr. Jacek Grabowski, Dr. Christian A. Salazar Soto, and an anonymous reviewer materially improved the accuracy of this stratigraphic experiment, but this author is responsible for all interpretations.
This research received no specific grants from funding agencies in the public, commercial, or not-for-profit sectors. It has been self-funded by Precision Stratigraphy Associates.
Aguado R., Company M. & Tavera J.M. (2000).- The Berriasian/Valanginian boundary in the Mediterranean region: New data from the Caravaca and Cehegín sections, SE Spain.- Cretaceous Research, vol. 21, p. 1-21.
Aguirre-Urreta M.B. (1998).- The ammonites Karakaschiceras and Neohoploceras (Valanginian Neocomitidae) from the Neuquen Basin, West-Central Argentina.- Journal of Paleontology, Cambridge, UK, vol. 72, p. 39-59.
Aguirre-Urreta M.B. (2001).- Marine Upper Jurassic-Lower Cretaceous stratigraphy and biostratigraphy of the Aconcagua-Neuquén Basin, Argentina and Chile.- Journal of Iberian Geology, Madrid, vol. 27, p. 71-90.
Aguirre-Urreta M.B., Casadío S., Cichowolski M., Lazo D.G. & Rodriguez D.L. (2008a).- Afinidades paleobiogeográficas de los invertebrados cretácicos de la Cuenca Neuquina.- Ameghiniana, Buenos Aires, vol. 45, p. 593-613.
Aguirre-Urreta B., Lescano M., Schmitz M.D., Tunik M., Concheyro A., Rawson P.F. & Ramos V.A. (2015).- Filling the gap: New precise Early Cretaceous radioisotopic ages from the Andes.- Geological Magazine, Cambridge (UK), vol. 152, p. 557-564.
Aguirre-Urreta B., Martinez M., Schmitz M., Lescano M., Omarini J., Tunik M., Kuhnert H., Concheyro A., Rawson P.F., Ramos V.A., Reboulet S., Noclin N., Frederichs T., Nickl A.-L. & Pälike H. (2019).- Interhemispheric radio-astrochronological calibration of the time scales from the Andean and the Tethyan areas in the Valanginian-Hauterivian (Early Cretaceous).- Gondwana Research, vol. 70, p. 104-132.
Aguirre-Urreta M.B., Mourgues A.F., Rawson P.F., Bulot L.G. & Jaillard E. (2007).- The Lower Cretaceous Chańarcillo and Neuquén Andean basins: Ammonoid biostratigraphy and correlations.- Geological Journal, vol. 42, p. 143-173.
Aguirre-Urreta M.B., Pazos P.J., Lazo D.G., Fanning C.M. & Litvak V.D. (2008b).- First U-Pb SHRIMP age of the Hauterivian stage, Neuquén Basin, Argentina.- Journal of South American Earth Sciences, vol. 26, p. 91-99.
Aguirre-Urreta M.B. & Rawson P.F. (1999).- Stratigraphic position of Valanginites, Lissonia, and Acantholissonia in the Lower Valanginian (Lower Cretaceous) ammonite sequence of Neuquén Basin, Argentina. In: Olóriz F. & Rodríguez-Tovar F.J. (eds.), Advancing research on living and fossil Cephalopods.- Kluwer/Plenum, New York, p. 521-529.
Aguirre-Urreta M.B. & Rawson P.F. (2003).- Lower Cretaceous ammonites from the Neuquén Basin, Argentina: The Hauterivian genus Holcoptychites.- Cretaceous Research, vol. 24, p. 589-613.
Aguirre-Urreta M.B. & Rawson P.F. (2010).- Lower Cretaceous ammonites from the Neuquén Basin, Argentina: The neocomitids of the Pseudofavrella angulatiformis Zone (upper Valanginian).- Cretaceous Research, vol. 31, p. 321-343.
Aguirre-Urreta M.B., Rawson P.F., Concheyro G.A., Bown P.R. & Otton E.G. (2005).- Lower Cretaceous (Berriasian-Aptian) biostratigraphy of the Neuquén Basin. In: Veiga G., Spalletti L., Howell J.A. & Schwarz E. (eds.), The Neuquén Basin: A case study in sequence stratigraphy and basin dynamics.- Geological Society, London, Special Publications, vol. 25, p. 57-81.
Aguirre-Urreta M.B., Schmitz M.D., Lescano M., Tunik M., Rawson P.F., Concheyro A., Buhler M. & Ramos V.A. (2017).- A high precision U-Pb radioisotopic age for the Agrio Formation, Neuquén Basin, Argentina: Implications for the chronology of the Hauterivian Stage.- Cretaceous Research, vol. 75, p. 193-204.
Alleman F., Catalano R., Farés F. & Remane J. (1971).- Standard Calpionellid zonation (Upper Tithonian-Valanginian) of the Western Mediterranean Province. In: Farininaci A. (ed.), Proceedings of the II Planktonic Conference, Roma, 1970.- Edizioni Tecnoscienza, Rome, p. 1337-1340.
Andreini G., Caracuel J. & Parisi G. (2007).- Calpionellid biostratigraphy of the Upper Tithonian-Upper Valanginian interval in Western Sicily (Italy).- Swiss Journal of Geosciences, 100, p. 179-198. DOI: https://doi.org/10.1007/s00015-007-1227-z.
Applegate J.L. & Bergen J.A. (1988).- Cretaceous calcareous nannofossil biostratigraphy of sediments recovered from the Galicia Margin, ODP Leg 103.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 103, p. 293-348. DOI: https://doi.org/10.2973/odp.proc.sr.103.144.1988
Arkadiev V., Guzhikov A.U., Baraboshkin E., Savelieva Y., Feodorva A., Shurekova O., Egor P. (2018).- Biostratigraphy and magnetostratigraphy of the upper Tithonian-Berriasian of the Crimean Mountains.- Cretaceous Research, vol. 87, p. 5-41.
Arnaud H., Arnaud-Vanneau A., Blanc-Alétru M.-C., Adatte T., Argot M., Delanoy G., Thieuloy J.-P., Vermeulen J., Virgone A., Virlouvet B. & Wermeille S. (1998).- Répartition stratigraphique des orbitolinidés de la plate-forme urgonienne subalpine et jurassienne (SE de la France).- Géologie Alpine, Grenoble, vol. 74, p. 3-89.
Arnaud-Vanneau A. & Masse J.-P. (1989).- Les foraminifčres benthiques des formations carbonatées de l’Hauterivien-Barrémien pro parte du Jura vaudois et neuchâtelois (Suisse).- Mémoires de la Société neuchâteloise des Sciences naturelles, vol. 11, p. 257-276.
Atasoy S.G. (unpublished, 2017).- Foraminiferal and calpionellid biostratigraphy, microfacies analyses and tectonic implications of the Upper Jurassic - Lower Cretaceous carbonate platform to slope successions in Sivrihisar Region (Eskişehir, NW Turkey).- MSc. Thesis, Middle East Technical University, Ankara, 254 p.
Atasoy S.G., Altiner D., Okay A.I. (2018).- Reconstruction of a Late Jurassic - Early Cretaceous carbonate platform margin with composite biostratigraphy and microfacies analysis (western Sakarya Zone, Turkey): Paleogeographic and tectonic implications.- Cretaceous Research, vol. 92, p.66-93.
Benzaggagh M. (2021).- Systematic revision and evolution of the Tithonian family Chitinoidellidae Trejo, 1975.- Carnets Geol., vol. 21, no. 2, p. 27-53. DOI: 10.2110/carnets.2021.2102
Blauser W.H. & McNulty C.L. (1980).- Calpionellids and nannoconids of the Taraises Formation (Early Cretaceous) in Santa Rosa Canyon, Sierra de Santa Rosa, Nuevo Leon, Mexico.- Gulf Coast Association of Geological Societies, Transactions, Lafayette - LA, vol. 30, p. 263-272.
Bown P.R., Rutledge D.C., Crux J.A. & Gallagher L.T. (1998).- Early Cretaceous. In: Bown P.R. (ed.), Calcareous nannofossil biostratigraphy.- British Micropalaeontological Society Publication Series, Chapman & Hall, London, p. 86-131.
Bralower T.J., Ludwig K.R., Obradovich J.D. & Jones D.L. (1990).- Berriasian (Early Cretaceous) radiometric ages from the Grindstone Creek section, Sacramento Valley, California.- Earth and Planetary Science Letters, vol. 98, p. 62-73.
Bralower T.J., Monechi S. & Thierstein H.R. (1989).- Calcareous nannofossil zonation of the Jurassic-Cretaceous boundary interval and correlation with the geomagnetic polarity timescale.- Marine Micropaleontology, vol. 14, p. 153-235.
Buffler R.T., Schlager W. & Pisciotto K.A. (1984).- 1. Introduction and Explanatory Notes.- Initial Reports DSDP, College Station - TX, vol. 77, p. 5-22. DOI: 10.2973/dsdp.proc.77.101.1984.
Bujtor L., Krische O. & Gawlick H.-J. (2013).- Late Berriasian ammonite assemblage and biostratigraphy of the Leube quarry near Salzburg (Northern Calcareous Alps, Austria).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 267, no. 3, p. 273-295.
Bulot L. (unpublished, 1995).- Les formations ŕ ammonites du Crétacé inférieur dans le Sud-Est de la France (Berriasien ŕ Hauterivien) : Biostratigraphie, paléontologie et cycles sédimentaires.- Ph.D. thesis, Muséum National d’Histoire Naturelle, Paris, 398 p.
Bulot L. Arnaud-Vanneau A. & Blanc É. (1992).- Basin type succession: Biostratigraphic tools, stratigraphic gaps and reworked sediments [excursion guide].- First day: Saturday May 9, International Symposium "Platform Margins", 6-8 May, Chichilianne, IAS & ASF, p. 25-40.
Bulot L.G., Thieuloy J.-P., Blanc É. & Klein J. (1993).- Le cadre stratigraphique du Valanginien supérieur et de l'Hauterivien du Sud-Est de la France : Définition des biochronozones et caractérisation de nouveaux biohorizons.- Géologie Alpine, Grenoble, vol. 68, p. 13-56.
Busnardo R., Thieuloy J.-P. & Moullade M. (1979).- Hypostratotype Mesogéen de l'étage Valanginien.- Les stratotypes Francais, vol. 6, Centre National de la Recherche Scientifique, Paris, 143 p.
Busnardo R., Clavel B., Charollais J. & Schroeder R. (1991).- Hauterivian-Barremian boundary at Mont Aiguille (Vercors, France): Biostratigraphy and sequence stratigraphy.- Revue de Paléobiologie, Genčve, vol. 10, p. 359-364.
Caracuel J., Monaco P. & Olóriz F. (2000).- Taphonomic tools to evaluate sedimentation rates and stratigraphic completeness in Rosso Ammonitica facies (epioceanic Tethyan Jurassic).- Rivista Italiana di Paleontologia e Stratigraphia, Milano, vol. 106, p. 353-368. DOI: https://doi.org/10.13130/2039-4942/6150
Carney J.L. & Pierce R.W. (1995).- Graphic correlation and composite standard databases as tools for the exploration biostratigrapher. In: Mann K.O. & Lane H.R. (eds.), Graphic correlation.- SEPM (Society for Sedimentary Geology) Special Publication, Tulsa - OK, vol. 53, p. 23-43.
Casellato C.E. & Erba E. (2021).- Reliability of calcareous nannofossil events in the Tithonian-early Berriasian time interval: Implications for a revised high resolution zonation.- Cretaceous Research, vol. 117, article 104611, 30 p.
Cecca F. (1998).- Early Cretaceous (pre-Aptian) ammonites of the Mediterranean Tethys: Palaeoecology and palaeobiogeography.- Palæogeography Palæoclimatology Palæoecology, vol. 138, p. 305-323.
Celestino R., Wohlwend S., Rehakova D. & Weissert H. (2017).- Carbon isotope stratigraphy, biostratigraphy and sedimentology of the Upper Jurassic - Lower Cretaceous Rayda Formation, central Oman Mountains.- Newsletters on Stratigraphy, Stuttgart, vol. 50, p. 91-109.
Channell J.E.T., Cecca F. & Erba E. (1995).- Correlations of Hauterivian and Barremian (Early Cretaceous) stage boundaries to polarity chrons.- Earth & Planetary Science Letters, vol. 134, p. 125-140.
Channell J.E.T. & Erba E. (1992).- Early Cretaceous polarity chrons CM0 to CM11 recorded in northern Italian sections near Brescia.- Earth Planetary Science Letters, vol. 108, p. 161-179.
Clavel B., Busnardo R., Charollais J., Conrad M. & Granier B. (2010).- Répartition biostratigraphique des orbitolinidés dans la biozonation ŕ ammonites (plate-forme urgonienne du Sud-Est de la France) Partie 1: Hauterivien supérieur-Barrémien basal.- Carnets Geol., Madrid, vol. 10, no. A06 (CG2010_A06), 53 p. DOI: 10.4267/2042/33369
Coccioni R. & Premoli-Silva I. (1994).- Planktonic foraminifera from the Lower Cretaceous of Rio Argos sections (southern Spain) and biostratigraphic implications.- Cretaceous Research, vol. 15, p. 645-687.
Cohen K.M., Finney S.C., Gibbard P.L. & Fan J.-X. (2021).- The ICS International Chronostratigraphic Chart (originally published 2013, 2021).- Episodes, vol. 36, p. 199-204. URL: http://www.stratigraphy.org/ICSchart/ChronostratChart2021-05.pdf
Di Lucia M., Trecalli A., Mutti M. & Parente M. (2012).- Bio-chemostratigraphy of the Barremian-Aptian shallow-water carbonates of the southern Apennines (Italy): Pinpointing the OAE1a in a Tethyan carbonate platform.- Solid Earth, vol. 3, p. 1-28.
Edgell H.S. (1971).- Calpionellid stratigraphy and the Jurassic-Cretaceous boundary in south-east Iran. In: Colloque de Jurassique, Luxembourg 1967.- Mémoires du B.R.G.M., Orléans, no. 75, p. 213-247.
Énay R. (2020).- The Jurassic/Cretaceous System Boundary is at an impasse. Why not go back to Oppel's 1865 original and historic definition of the Tithonian?.- Cretaceous Research, vol. 106, article 104241, 20 p.
Erba E. & Quadrio B. (1987).- Biostratigrafia a nannofossili calcarei calpionellidi e foraminiferi planctonici della Maiolica (Titoniano superiore-Aptiano) nelle Prealpi bresciane (Italia settentrionale).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 93, no. 1, p. 3-108. URL: https://riviste.unimi.it/index.php/RIPS/article/view/13228
Frau C., Bulot L.G., Reháková D., Wimbledon W.A.P. & Ifrim C. (2016).- Revision of the ammonite index species Berriasella jacobi Mazenot, 1939 and its consequences for the biostratigraphy of the Berriasian Stage.- Cretaceous Research, vol. 66, p. 94-114.
Frau C., Bulot L.G. & Wimbledon W.A.P. (2015).- Upper Tithonian Himalayitidae Spath, 1925 (Perisphinctoidea, Ammonitina) from Le Chouet (Drôme, France): Implications for the systematics.- Geologica Carpathica, Bratislava, vol. 66, p. 117-132.
Frau C., Bulot L., Wimbledon W.A.P. & Ifrim C. (2016).- Systematic palaeontology of the Perisphinctoidea in the Jurassic/Cretaceous boundary interval at Le Chouet (Drôme, France), and its implications for biostratigraphy.- Acta Geologica Polonica, Warsaw, vol. 66, p. 175-204.
Gale A.S., Mutterlose J., Battenberg S., Gradstein F.M., Agterberg F.P., Ogg J.G. & Petrizzo M.R. (2020).- Chapter 27. The Cretaceous Period. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale 2020.- Elsevier, Amsterdam, vol. 2, p. 1023-1086.
Galbrun B., Rasplus L. & Le Hégarat G. (1986).- Données nouvelles sur le stratotype du Berriasien : Corrélations entre magnétostratigraphie et biostratigraphie.- Bulletin de la Société géologique de France, Paris, vol. 4, p. 575-584.
Garrido A.C. & Parent H. (2017).- Lithofacies and age of the Quintuco Formation (Lower Cretaceous) in the Mallín Quemado area (Sierra de la Vaca Muerta, Neuquén Basin, Argentina). Stratigraphic and depositional implications.- Boletín del Instituto de Fisiografía y Geología, Rosario, vol. 87, p. 1-16.
Grabowski J. (2011).- Magnetostratigraphy of the Jurassic/Cretaceous boundary interval in the Western Tethys and its correlations with other regions: A review.- Volumina Jurassica, Warsaw, vol. IX, p. 105-128.
Grabowski J., Lodowski D.G., Schnyder J., Sobień K., Chadimova L., Pszczólkowski A., Krzemiński L. & Schnabl P. (2018).- 17. Magnetic stratigraphy, stable isotopes and chemostratigraphy in the upper Berriasian of Rówienka section (Western Tatra Mts., Fatric succession, Poland): Towards a consistent model of late Berriasian paleoenvironmental changes in the Western Tethys.- JK2018 (December 5-7, 2018), Muséum d'Histoire naturelle de Genčve, p. 30-33.
Grabowski J. & Pszczólkowski A. (2006).- Magneto- and biostratigraphy of the Tithonian-Berriasian pelagic sediments in the Tatra Mountains (central Western Carpathians, Poland): Sedimentary and rock magnetic changes at the Jurassic/Cretaceous boundary.- Cretaceous Research, vol. 27, p. 398-417.
Gradstein F.M., Ogg J.G., Smith A.G., Bleeker W. & Lourens L.J. (2004).- A new Geologic Time Scale with special reference to Precambrian and Neogene.- Episodes, Bangalore, vol. 27, p. 83-100.
Granier B., Énay R. & Charollais J. (2020).- Discussion of the paper by Wimbledon et al., 2020b, entitled "The proposal of a GSSP for the Berriasian Stage (Cretaceous System): Part 1" [Volumina Jurassica, XVIII (1)].- Volumina Jurassica, Warsaw, vol. XVIII, no. 2, p. 111-124. URL: https://vjs.pgi.gov.pl/article/view/28578/20272
Haq B.U. (2014).- Cretaceous eustasy revisited.- Global and Planetary Change, vol. 113, p. 44-58.
Hesselbo S.P., Ogg J.G. & Ruhl M. (2020).- The Jurassic Period, Chap. 26. In: Gradstein F.M, Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale 2020.- Elsevier, Amsterdam, vol. 2, p. 955-1022.
Hinte J.E. van (1976).- A Cretaceous time scale.- American Association of Petroleum Geologists Bulletin, vol. 60, p. 269-287.
Hoedemaeker P. (2013).- Genus Pseudothurmannia Spath, 1923 and related subgenera Crioceratites (Balearites) Sarkar, 1954 and C. (Binelliceras) Sarkar, 1977 (Lower Cretaceous Ammonoidea).- Revue de Paléobiologie, Genčve, vol. 32, no. 1, p. 1-209.
Hoedemaeker P., Janssen N., Casellato C., Gardin S., Reháková D. & Jamrichová M. (2016).- Jurassic/Cretaceous boundary in the Río Argos succession (Caravaca, SE Spain).- Revue de Paléobiologie, Genčve, vol. 35, p. 111-247.
Hoedemaeker P.J. & Leereveld H. (1995).- Biostratigraphy and sequence stratigraphy of the Berriasian-lowest Aptian (Lower Cretaceous) of the Rio Argos succession.- Cretaceous Research, vol. 16, p. 195-230.
Hood K.C. (1995).- Evaluating the use of average composite sections and derived correlations in the graphic correlation technique. In: Mann K.O. & Lane H.R. (eds.), Graphic correlation.- SEPM (Society for Sedimentary Geology) Special Publication, Tulsa - OK, vol. 53, p. 83-93.
Horton B.K., Fuentes F., Boll A., Starck D. Ramirez S.G. & Stockli D.F. (2016).- Andean stratigraphic record of the transition from backarc extension to orogenic shortening: A case study from the northern Neuquén Basin, Argentina.- Journal of South American Earth Sciences, vol. 71, p. 17-40.
Housa V., Krsa M., Mana O., Prunera P., Venhodova D., Cecca F., Nardic G. & Piscitello M. (2004).- Combined magnetostratigraphic, palaeomagnetic and calpionellid investigations across Jurassic/Cretaceous boundary strata in the Bosso Valley, Umbria, central Italy.- Cretaceous Research, vol. 25, p. 771-785.
Huang C. (2018).- Chapter 2. Astronomical Time Scale for the Mesozoic.- Stratigraphy & Timescales, vol. 3, p. 81-150.
Ice R.G. & McNulty CL. (1980).- Foraminifers and calcispheres from the Cuesta del Cura and lower Agua Nueva(?) formations (Cretaceous) in east-central Mexico.- Association of Geological Societies, Transactions, Lafayette - LA, vol. 30, p. 403-425.
Iglesia Llanos M.P. & Kietzmann D.A. (2020).- Magnetostratigraphy of the Jurassic through Lower Cretaceous in the Neuquén Basin. In: Kietzmann D.A. & Folguera A. (eds.), Opening and Closure of the Neuquén Basin in the Southern Andes.- Springer Earth System Sciences, New York - NY, p. 175-210.
Iglesia Llanos M.P., Kietzmann D.A., Martinez M.K. & Palma R.M. (2017).- Magnetostratigraphy of the Upper Jurassic-Lower Cretaceous from Argentina: Implications for the J-K boundary in the Neuquén Basin.- Cretaceous Research, vol. 70, p. 189-208.
Ivanova D.K. & Kietzmann D.A. (2017).- Calcareous dinoflagellate cysts from the Tithonian-Valanginian Vaca Muerta Formation in the southern Mendoza area of the Neuquén Basin, Argentina.- Journal of South American Earth Sciences, vol. 77, p. 150-169.
Kedzierski M. & Ochabska A. (2012).- Calcareous nannofossil biostratigraphy and sedimentary environment of Valanginian-Hauterivian rhythmites (Silesian Nappe, Polish Carpathians).- Annales Societatis Geologorum Poloniae, Kraków, vol. 82, p. 225-237.
Kenjo S., Reboulet S., Mattioli E. & Ma'louleh K. (2021).- The Berriasian-Valanginian boundary in the Mediterranean Province of the Tethyan Realm: Ammonite and calcareous nannofossil biostratigraphy of the Vergol section (Montbrun-les-Bains, SE France), candidate for the Valanginian GSSP.- Cretaceous Research, vol. 121, article 104738.
Kietzmann D.A. (2017).- Chitinoidellids from the Early Tithonian-Early Valanginian Vaca Muerta Formation in the northern Neuquén basin, Argentina.- Journal of South American Earth Science, vol. 76, p. 152-164.
Kietzmann D.A., Ambrosio A.L., Suriano J., Alonso M.S., González Tomassini F., Depine G. & Repol D. (2016).- The Vaca Muerta-Quintuco system (Tithonian-Valanginian) in the Neuquén Basin, Argentina: A view from the outcrops in the Chos Malal fold and thrust belt.- American Association of Petroleum Geologists Bulletin, Tulsa - OK, vol. 100, p. 743-771. DOI: 10.1306/02101615121
Kietzmann D.A., Iglesia Llanos M.P. & Kohan Martínez M. (2018).- Ch. 8, Astronomical calibration of the Tithonian-Berriasian in the Neuquén Basin, Argentina: Contribution from the Southern Hemisphere to the Geological Time Scale.- Stratigraphy & Timescales, vol. 3, p. 327-351.
Kietzmann D.A., Iglesia Llanos M.P. & Kohan Martínez M. (2020).- Orbital Controls and high-resolution cyclostratigraphy of Late Jurassic-Early Cretaceous in the Neuquén Basin. In: Kietzmann D.A. & Folguera A. (eds.), Opening and closure of the Neuquén Basin in the Southern Andes.- Springer Earth System Sciences, New York, p. 211-235.
Kietzmann D.A., Iglesia Llanos M.P., Palacio J.P. & Sturlesi M.A. (2021a).- Facies analysis and stratigraphy across the Jurassic-Cretaceous boundary in a new basinal Tithonian-Berriasian section of the Vaca Muerta Formation, Las Tapaderas, Southern Mendoza Andes, Argentina.- Journal of South American Earth Sciences, vol. 109, article 103267, 22 p.
Kietzmann D.A., Iglesia Llanos M.P., González Tomassini F., Lanusse Noguera I., Vallejo D. & Reijenstein H. (2021b).- Upper Jurassic-Lower Cretaceous calpionellid zones in the Neuquén Basin (Southern Andes, Argentina): Correlation with ammonite zones and biostratigraphic synthesis.- Cretaceous Research, vol. 127, article 104950, 28 p.
Kietzmann D.A., Martín-Chivelet J., Palma R.M., López-Gómez J., Lescano M. & Concheyro A. (2011).- Evidence of precessional and eccentricity orbital cycles in a Tithonian source rock: The mid-outer carbonate ramp of the Vaca Muerta Formation, Northern Neuquén Basin, Argentina.- American Association of Petroleum Geologists Bulletin, Tulsa - OK, vol. 95, p. 1459-1474.
Kietzmann D.A., Palma R.M. & Iglesia Llanos M.P. (2015).- Cyclostratigraphy of an orbitally-driven Tithonian-Valanginian carbonate ramp succession, Southern Mendoza, Argentina: Implications for the Jurassic-Cretaceous boundary in the Neuquén Basin.- Sedimentary Geology, vol. 315, p. 29-346.
Kietzmann D. & Paulin S. (2019).- Cyclostratigraphy of an upper Valanginian - lower Hauterivian mixed siliciclastic-carbonate ramp succession (Pilmatué Member of the Agrio Formation), Loma La Torre Section, Northern Neuquén Basin, Argentina.- Cretaceous Research, vol. 98, p. 26-46.
Kohan Martínez M., Aguirre-Urreta B.M., Marina L., Julieta O., Maisa T. Thomas F., Anna-Leah N. & Heiko P. (2017).- Radio-astrochronology of the Agrio Formation (Neuquén Basin, Argentina) to reduce the uncertainties of the geological time scale in Early Cretaceous times.- Geophysical Research Abstracts, vol. 19, article EGU2017-12075, 1 p. (abstract).
Kohan Martínez M., Kietzmann D.A. & Iglesia Llanos M.P. (2018).- Magnetostratigraphy and cyclostratigraphy of the Tithonian interval from the Vaca Muerta Formation, southern Neuquén Basin, Argentina.- Journal of South American Earth Science, vol. 85, p. 209-228.
Lakova I. & Petrova S. (2013).- Towards a standard Tithonian to Valanginian calpionellid zonation of the Tethyan Realm. Acta Geologica Polonica, Warsaw, vol. 63, p. 201-221.
Lakova I., Stoykova K. & Ivanova D. (1997).- Tithonian to Valanginian bioevents and integrated zonation on calpionellids, calcareous nannofossils and calcareous dinocycts from the Western the Western Balkanides, Bulgaria.- Mineralia Slovaca, Dionýz Štúr, vol. 29, p. 301-303.
Lazo D.G., Concheyro A., Ottone G., Guler M.V. & Aguirre-Urreta B. (2009).- Bioestratigrapfía integrada de la Formación Agrio en su localidad tipo, Cretácico temprano de Cuenca Neuquina.- Revista de la Asociación Geológica Argentina, Buenos Aires, vol. 65, p. 322-341.
Leanza H.A., Sattler F., Martinez R.S. & Carbone O. (2011).- La Formación Vaca Muerta y equivalentes (Jurásico Tardío-Cretácico Temprano) en la Cuenca Neuquina.- Relatorio del XVIII Congreso Geológico Argentino, Neuquén, 2011, p. 113-129.
Le Hégarat G. (1973).- Le Berriasien du Sud-Est de la France.- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, Villeurbanne, no. 43, 575 p.
Le Hégarat G. & Remane J. (1968).- Tithonique supérieur et Berriasien de l'Ardčche et de l'Hérault. Corrélation des ammonites et des calpionelles.- Geobios, Villeurbanne, vol. 1, no. 1, p. 7-70.
Lehmann J., Ifrim C., Bulot L. & Frau C. (2015).- Paleobiogeography of Early Cretaceous Ammonoids. In: Klug C., Korn D., De Baets K., Kruta I. & Mapes R.H. (eds.), Ammonoid paleobiology: From macroevolution to paleogeography.- Topics in Geobiology, Dordrecht, vol. 44, p. 229-257.
Lena L., López-Martínez R., Lescano M., Aguirre-Urreta B., Concheyro A., Vennari V., Naipauer M., Samankassou E., Pimentel M., Ramos V. & Schaltegger U. (2019a).- High precision U-Pb ages in the early Tithonian to early Berriasian and implications for the numerical age of the Jurassic/Cretaceous boundary.- Solid Earth, vol. 10, p. 1-14. DOI: 10.5194/se-10-1-2019
Liu Yong-Qing, Ji Qiang, Jiang Xiao-Jun, Kuang Hong-Wei, Ji Shu'an, Gao Lian-Feng, Zhang Zhen-Guo, Peng Nan Yuan, Chong-Xi Wang, Xu Ri & Xu Huan (2013).- U-Pb Zircon Ages of Early Cretaceous Volcanic Rocks in the Tethyan Himalaya at Yangzuoyong Co Lake, Nagarze, Southern Tibet, and Implications for the Jurassic/Cretaceous Boundary.- Cretaceous Research, vol. 40, p. 90-101.
López-Martínez R., Aguirre-Urreta B., Lescano M., Concheyro A., Vennari V. & Ramos V.A. (2017a).- Tethyan calpionellids in the Neuquén Basin (Argentine Andes), their significance in defining the Jurassic/Cretaceous boundary and pathways for Tethyan-Eastern Pacific connections.- Journal of South American Earth Sciences, vol. 78, p. 116-125.
López-Martínez R., Barragán R., Bernal J.P., Reháková D., Gómez-Tuena A., Martini M. & Ortega C. (2017b).- Integrated stratigraphy and isotopic ages at the Berriasian-Valanginian boundary at Tlatlauquitepec (Puebla, Mexico).- Journal of South American Earth Sciences, vol. 85, p. 209-228.
López-Martínez R., Barragán R., Rehákova D. & Cobiella-Reguera J.L. (2013).- Calpionellid distribution and microfacies across the Jurassic/Cretaceous boundary in western Cuba (Sierra de los Órganos).- Geologica Carpathica, Bratislava, vol. 64, p. 195-208.
López-Martínez R., Barragán R., Reháková D., Martini M. & Eguiluz de Antuńano S. (2015).- Calpionellid biostratigraphy, U-Pb geochronology and microfacies of the Upper Jurassic-Lower Cretaceous Pimienta Formation (Tamazunchale, San Luis Potosí, central-eastern Mexico).- Boletín de la Sociedad Geológica Mexicana, Mexico City - D.C., vol. 67, p. 75-86.
Lowrie W. & Channell J.E.T. (1984).- Magnetostratigraphy of the Jurassic-Cretaceous boundary in the Maiolica limestone (Umbria, Italy).- Geology, Boulder - CO, vol. 12, p. 44-47.
Lukenender A., Halásova E., Kroh A., Mayrhofer S., Pruner D., Rehákova D., Schnabl P., Sprovieri M. & Wagreich M. (2010).- High resolution stratigraphy of the Jurassic-Cretaceous boundary interval in the Gresten Klippenbelt (Austria).- Geologica Carpathica, Bratislava, vol. 61, p. 365-381.
Maaloui K. & Zargouni F. (2016).- The lower and middle Berriasian in Central Tunisia: Integrated ammonite and calpionellid biostratigraphy of the Sidi Kralif Formation.- Acta Geologica Polonica, Warsaw, vol. 66, p. 43-58.
Maaloui K. & Zargouni F. (2017).- Biostratigraphical study around the Jurassic/Cretaceous boundary in Central Tunisia zonal schemes and correlation.- Journal of African Earth Sciences, vol. 125, p. 166-176.
Martinez M., Aguirre-Urreta B.M., Marina L., Julieta O., Maisa T., Thomas F., Anna-Leah N. & Heiko P. (2017).- Radio-astrochronology of the Agrio Formation (Neuquén Basin, Argentina) to reduce the uncertainties of the geological time scale in Early Cretaceous times.- Geophysical Research Abstracts, vol. 19, article EGU2017-12075, 1 p. (abstract).
Martinez M., Deconinck J.-F., Pellenard P., Reboulet S. & Riquier L. (2013).- Astrochronology of the Valanginian Stage from reference sections (Vocontian Basin, France) and palaeoenvironmental implications for the Weissert Event.- Palæogeography Palæoclimatology Palæoecology, vol. 376, p. 91-102.
Martinez M., Deconinck J-F., Pellenard P., Riquier L., Company M., Reboulet S. & Moiroud M. (2015).- Astrochronology of the Valanginian-Hauterivian stages (Early Cretaceous): Chronological relationships between the Paraná-Etendeka large igneous province and the Weissert and the Faraoni events.- Global and Planetary Change, vol. 131, p. 158-17.
Martinez M., Pellenard P., Deconinck J.-F., Monna F., Riquier L., Boulila S., Moiroud M. & Company M. (2012).- An orbital floating time scale of the Hauterivian/Barremian GSSP from a magnetic susceptibility signal (Río Argos, Spain).- Cretaceous Research, vol. 36, p. 106-115.
Masure E. (1988).- 25. Berriasian to Aptian dinoflagellate cysts from the Galicia margin, Offshore Spain, Sites 638 and 639, ODP Leg 103.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 103, p. 433-438.
Mutterlose J., Rawson P.F., Reboulet S., Baudin F, Bulot L., Emmanuel L., Gardin S., Martinez M. & Renard M. (2020).- The Global Boundary Stratotype Section and Point (GSSP) for the base of the Hauterivian Stage (Lower Cretaceous), La Charce, southeast France.- Episodes, Beijing, vol. 44, p. 129-150. DOI: 10.18814/epiiugs/2020/020072
Naipauer M., García Morabito E., Marques J.C., Tunik V., Rojas Vera E., Vujovich G.I., Pimentel M.P. & Ramos V.A. (2012).- Intraplate Late Jurassic deformation and exhumation in western central Argentina: Constraints from surface data and U-Pb detrital zircon ages.- Tectonophysics, vol. 524-525, p. 59-75.
Naipauer M., Lescano M., Zavala C., Aguirre-Urreta B., Pimente M.M. & Ramos V.A. (2015a).- Estudio multidisciplinario en la Formación Vaca Muerta: Ambiente de sedimentación, bioestratigrafía, edades U-Pb en circones y áreas de proveniencia del Miembro Huncal.- XIV Congresso Geológica Chilean, La Serena, Resumenes Expandidos Electrónicos, p. 1-4.
Naipauer M., Tapi F., Mescu J., Faria M., Pimente M.M. & Ramos V.A. (2015b).- Detrital and volcanic zircon U-Pb ages from southern Mendoza (Argentina): An insight on the source regions in the northern part of the Neuquén Basin.- Journal of South American Earth Sciences, vol. 64, p. 434-451.
Naipauer M., Tunik M., Marques J.C., Rojas-Vera E.A., Vujovich G.I., Pimentel M.M. & Ramos V.A. (2015c).- U-Pb detrital zircon ages of Upper Jurassic continental successions: Implications for the provenance and absolute age of the Jurassic-Cretaceous boundary in the Neuquén Basin. In: Sepúlveda S.A., Giambiagi L.B., Moreiras S.M., Pinto L., Tunik M., Hoke G.D. & Farías M. (eds)., Geodynamic Processes in the Andes of central Chile and Argentina.- Geological Society, London, Special Publications, vol. 399, p. 131-154.
Ogg J.G., Agterberg F.P. & Gradstein F.M. (2004).- The Cretaceous Period. In: Gradstein F.M., Ogg J.G. & Smith A.G. (eds.), A Geologic Time Scale, 2004.- Cambridge University Press, Cambridge - MA, p. 344-383.
Ogg J.G., Hinnov L.A. & Huang C. (2012).- Cretaceous. In: Gradstein F.M, Ogg J.G., Schmitz M.D.& Ogg G.M. (eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 793-853.
Ogg J.G., Ogg G.M. & Gradstein F.M. (2016).- A Concise Geologic Time Scale 2016.- Elsevier, Amsterdam, 234 p.
Olóriz F., Villaseńor A.B., Gonzalez-Arreola C. & Westermann G.E.G. (1999).- Ammonite biostratigraphy and correlations in the Upper Jurassic-lowermost Cretaceous La Caja Formation of North-Central Mexico (Sierra de Catorce, San Luis Potosí). In: Olóriz F. & Rodríguez-Tovar F.J. (eds.), Advancing research on living and fossil Cephalopods.- Kluwer/Plenum, New York, p. 463-491.
Parent H., Garrido A.C., Scherzinger A., Schweigert G. & Fözy I. (2015).- The Tithonian-Lower Valanginian stratigraphy and ammonite fauna of the Vaca Muerta Formation in Pampa Tril, Neuquén Basin, Argentina.- Boletín del Instituto de Fisiografía y Geología, Rosario, vol. 86, p. 1-96.
Parent H., Scherzinger A. & Schweigert G. (2011).- The Tithonian-Berriasian ammonite fauna and stratigraphy of Arroyo Cieneguita, Neuquén-Mendoza Basin, Argentina.- Boletín del Instituto de Fisiografía y Geología, Rosario, vol. 79-81, p. 21-94
Parent H., Scherzinger A., Schweigert G. & Garrido A.C. (2017).- Additional Tithonian and Berriasian ammonites from the Vaca Muerta Formation in Pampa Tril, Neuquén Basin, Argentina.- Volumina Jurassica, Varsaw, vol. XV, p. 139-154.
Petrova S., Koleva-Rekalova E., Ivanova D. & Lakova I. (2019).- Biostratigraphy and microfacies of the pelagic carbonate formations in the Yavorets section (Tithonian-Berriasian), Western Balkan Mts, Bulgaria.- Geologica Balcanica, Sofia, vol. 48, p. 51-73.
Pruner P., Houša V.F., Oloriz F., Košak M.M., Krs M., Man O., Schnabl P., Venhodová D., Tavera J.M. & Mazuch M. (2010).- High-resolution magnetostratigraphy and biostratigraphic zonation of the Jurassic/Cretaceous boundary strata in the Puerto Escańo section (southern Spain).- Cretaceous Research, vol. 31, p. 192-206.
Pszczólkowski A. & Myczynski R. (2010).- Tithonian-Early Valanginian evolution of deposition along the proto-Caribbean margin of North America recorded in Guaniguanico successions (western Cuba).- Journal of South American Earth Science, vol. 29, p. 225-253.
Rawson P.F. (2007).- Global relationships of Argentine (Neuquén Basin) Early Cretaceous ammonite faunas.- Geological Journal, vol. 42, p. 175-183.
Rawson P.F., Concheyro A., Buhler M. & Ramos V.A. (2017).- A high precision U-Pb radioisotopic age for the Agrio Formation, Neuquén Basin, Argentina: Implications for the chronology of the Hauterivian Stage.- Cretaceous Research, vol. 75, p. 193-204.
Reboulet S. (2008).- The Global Boundary Stratotype Sections and Points (GSSP) of the Hauterivian: La Charce section (Drôme, France, Vocontian basin). In: Mattioli E., Gardin S., Giraud F., Olivero F., Pittet B. & Reboulet S. (Eds.), Guidebook for the post-congress fieldtrip in the Vocontian Basin, SE France (September 11-13 2008).- Carnets de Géologie, Madrid, Book 2008/01 (CG2008_B01), p. 7-10. DOI: 10.4267/2042/18137
Reboulet S. & Atrops F. (1999).- Comments and proposals about the Valanginian-Lower Hauterivian ammonite zonation of south-eastern France.- Eclogae Geologicae Helvetiae, Basel, vol. 92, no. 2, p. 183-198.
Reboulet S., Szives O., Aguirre-Urreta B., Barragán R., Company M., Frau C., Kakabadze M.V., Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S.N., Vašíček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Rehakova D., Halásova E. & Lukeneder A. (2009).- The Jurassic-Cretaceous boundary in the Gresten Klippenbelt (Nutzhof, Lower Austria): Implications for micro- and nannofacies analysis.- Annalen des Naturhistorischen Museums in Wien, vol. 110A, p. 345-381.
Remane J., Faure-Muret A., Odin G.S., Cita M.B., Dercourt J., Bouysse P. & Repetto F. (2002).- International stratigraphic chart.- International Commission on Stratigraphy, International Union of Geological Sciences, Paris, 1 chart.
Riccardi A.C. (1988).- The Cretaceous system of southern South America. In: Klug C., Korn D., De Baets K., Kruta I. & Mapes R.H. (eds.), Ammonoid paleobiology: From macroevolution to paleogeography.- Geological Society of America, Memoir, Boulder - CO, vol. 186, p. 116-143.
Riccardi A.C. (2015).- Remarks on the Tithonian-Berriasian ammonite biostratigraphy of west central Argentina.- Volumina Jurassica, Varsaw, vol. XIII, no. 2, p. 23-52.
Salazar Soto C.A. (unpublished, 2012).- The Jurassic-Cretaceous boundary (Tithonian-Hauterivian) in the Andean Basin of central Chile: Ammonites, bio- and sequence stratigraphy and palaeobiogeography.- Unpublished Dissertation, Rupecht-Karls-Universität, Heidelberg, 388 p.
Salazar C. & Stinnesbeck W. (2015).- Redefinition, stratigraphy and facies of the Lo Valdes Formation (Upper Jurassic-Lower Cretaceous) in central Chile.- Boletín del Museo Nacional de Historia Natural, Santiago, vol. 64, p. 41-68.
Salazar C. & Stinnesbeck W. (2016).- Tithonian-Berriasian ammonites from the Bańos del Flaco Formation, central Chile.- Journal of Systematic Palaeontology, vol. 14, no. 2, p. 149-182.
Salazar C., Stinnesbeck W. & Álvarez M. (2020).- Ammonite biostratigraphy and bioevents in the Jurassic - Cretaceous boundary of central Chile.- Cretaceous Research, vol. 107, article 104282, 18 p.
Schwarz E., Álvarez-Trentini G. & Valenzuel M.E. (2013).- Ciclos mixtos carbonáticos/silicoclásticos en el Miembro Superior de la Formación Mulichinco (Yacimiento Cańadón Amarillo, Cuenca Neuquina Central, Argentina): Implicancias secuenciales y para caracterización de reservorios.- Latin American Journal of Sedimentology and Basin Analysis, Buenos Aires, vol. 20, p. 21-49.
Schwarz E., Spalletti L.A. & Howell J.A. (2006).- Sedimentary response to a tectonically induced sea-level fall in a shallow back-arc basin: The Mulichinco Formation (Lower Cretaceous), Neuquén Basin, Argentina.- Sedimentology, vol. 53, p. 55-81.
Schwarz E., Spalletti L.A., Veiga G.D. & Fanning C.M. (2016).- First U-Pb SHRIMP age for the Pilmatué Member (Agrio Formation) of the Neuquén Basin, Argentina: Implications for the Hauterivian lower boundary Hauterivian lower boundary.- Cretaceous Research, vol. 58, p. 223-233.
Scott R.W. (1990).- Models and stratigraphy of Mid-Cretaceous reef communities, Gulf of Mexico.- SEPM (Society for Sedimentary Geology), Concepts in Sedimentology and Paleontology, Tulsa - OK, vol. 2, 102 p.
Scott R.W. (2014).- Cretaceous chronostratigraphic database: Construction and applications.- Carnets Geol., Madrid, vol. 14, no. 2, p. 1-13. DOI: 10.4267/2042/53522
Scott R.W. (2019a).- Jurassic-Cretaceous boundary bioevents and magnetochrons: A stratigraphic experiment.- Cretaceous Research, vol. 100, p. 97-104.
Scott R.W. (2019b).- Valanginian Knowles Limestone, East Texas, A potential reservoir.- Gulf Coast Association of Geological Societies, Houston - TX, vol. 8, p. 79-88.
Selby D., Mutterlose J. & Condon D.J. (2009).- U-Pb and Re-Os geochronology of the Aptian/Albian and Cenomanian/Turonian stage boundaries: Implications for timescale calibration, osmium isotope seawater composition and Re-Os systematics in organic-rich sediments.- Chemical Geology, vol. 265, p. 394-409.
Sheridan R.E. & Gradstein F.M. (1983).- Site 534: Blake-Bahama Basin (Shipboard Scientific Party).- Initial Reports of the Deep Sea Drilling Project, College Station - TX, vol. 76, p. 141-340. DOI: 10.2973/dsdp.proc.76.101.1983
Simmons M.D. & Hart M.B. (1987).- The biostratigraphy and microfacies of the Early to mid-Cretaceous carbonates of Wadi Mi’aidin, Central Oman Mountains. In: Hart M.B. (ed.), Micropalaeontology of carbonate environments.- Ellis Horwood, Totnes, p. 176-207.
Skupien P. & Doupovcová P. (2019).- Dinoflagellates and calpionellids of the Jurassic-Cretaceous boundary, Outer Western Carpathians (Czech Republic).- Cretaceous Research, vol. 99, p. 209-228.
Speranza F., Satolli S., Mattioli E. & Calamita F. (2005).- Magnetic stratigraphy of Kimmeridgian-Aptian sections from Umbria-Marche (Italy): New details on the M polarity sequence.- Journal of Geophysical Research, vol. 110, B12109. DOI: https://agupubs.onlinelibrary.wiley.com/doi/epdf/10.1029/2005JB003884.
Szives O. & Főzy I. (2022).- Towards the ammonite zonation of the Jurassic/Cretaceous transition: New data from ammonitico rosso/biancone sections of the Transdanubian Range (Hungary).- Newsletters on Stratigraphy, Stuttgart, 42 p. DOI: 10.1127/nos/2022/0679
Vennari V.V., Lescano M., Naipauer M., Aguirre-Urreta B., Concheyro A., Schalteggerb U., Armstrong R., Pimentel M. & Ramos V.A. (2014).- New constraints on the Jurassic-Cretaceous boundary in the High Andes using high-precision U-Pb data.- Gondwana Research, vol. 26, p. 374-385.
Vennari V.V. (2016).- Tithonian ammonoids (Cephalopoda, Ammonoidea) from the Vaca Muerta Formation, Neuquén Basin, West-Central Argentina.- Palaeontographica Abteilung A, Stuttgart, Band 306, Lieferung 1-6, p. 85-165.
Vennari V.V. & Pujana I. (2017).- Finding of two new radiolarian associations calibrated with ammonoids in the Vaca Muerta Formation (Late Jurassic-Early Cretaceous), Neuquén Basin, Argentina.- Journal of South American Earth Sciences, vol. 75, p. 35-50.
Walker J.D., Geissman J.W., Bowring S.A. & Babcock L.E. (compilers, 2018).- Geologic Time Scale v. 5.0.- Geological Society of America, Boulder - CO. URL: https://www.geosociety.org/documents/gsa/timescale/timescl.pdf
Wan Xiaoqiao, Scott R.W., Chen Wen, Gao Lianfeng & Zhang Yiyi (2011a).- Early Cretaceous stratigraphy and SHRIMP U-Pb age constrain the Valanginian-Hauterivian Boundary in Southern Tibet.- Lethaia, Oslo, vol. 43, p. 231-244. DOI: 10.1111/j.1502-3931.2010.00238.x
Wan X., Scott R., Chen W., Gao L. & Yiyi Z. (2011b).- Early Cretaceous stratigraphy and SHRIMP U-Pb age constrain the Valanginian-Hauterivian Boundary in Southern Tibet.- Lethaia, Oslo, vol. 44, p. 231-244.
Westermann G.E.G. (2000).- Biochore classification and nomenclature in paleobiogeography: An attempt at order.- Palæogeography Palæoclimatology Palæoecology, vol. 158, p. 1-13.
Wimbledon W.A.P., Reháková D., Pszczółkowski A., Cassellato C.E., Halásová E., Frau C., Bulot L.G., Grabowski J., Sobień K., Pruner Schnabl P. & Čížková K. (2013).- A preliminary account of the bio- and magnetostratigraphy of the upper Tithonian-lower Berriasian interval at Le Chouet, Drôme (SE France).- Geologica Carpathica, Bratislava, vol. 64, p. 437-460.
Wimbledon W.A.P. (2017).- Developments with fixing a Tithonian/Berriasian (J/K) boundary.- Volumina Jurassica, Warsaw, vol. XV, p. 181-186.
Wimbledon W.A.P., Reháková D.A., Svobodová A., Schnabl P., Pruner P., Elbra T., Šifnerová K., Kdýr š., Frau C., Schnyder J. & Galbrun B. (2020).- Fixing a J/K boundary: A comparative account of key Tithonian-Berriasian profiles in the departments of Drôme and Hautes-Alpes, France.- Geologica Carpathica, Bratislava, vol. 71, p. 24-46.
Wippich M.G.E. (2003).- Valanginian (Early Cretaceous) ammonite faunas from the western High Atlas, Morocco, and the recognition of western Mediterranean 'standard' zones.- Cretaceous Research, vol. 24, p. 357-374.
Appendix 1: List of sections and sources of biostratigraphic and lithostratigraphic data in LOK2016DB. Catalog numbers are used by GraphCor software.
| Catalog #-Name: | Reference |
| CRET.16 Standard Reference Section-2016 Geologic Time Scale: | Ogg et al., 2012, Figs. 13.4, 27.6, Table 27.2-3, Appendix 2 |
| LOK.1 DSDP 534 Blake Plateau: | Sheridan et al., 1983 |
| LOK.2 DSDP 535 Blake Plateau: | Buffler et al., 1984 |
| LOK.3 Rio Argos section, Spain, Barremian Candidate GSSP: | Coccioni & Premoli-Silva, 1994; Hoedemaeker & Leereveld, 1995 |
| LOK.5 Santa Rosa Canyon, Mexico: | Blauser & McNulty, 1980; Ice & McNulty, 1980 |
| LOK.6 Berrias Section, France (Galbrun et al.): | Galbrun et al., 1986, Fig. 2 |
| LOK.6B Berrias Section, France (Le Hegarat): | Le Hégarat & Remane, 1968, Table VII, p. 45; Le Hégarat, 1971, Table 7 |
| LOK.7 ODP 638B & C, Offshore Spain: | Applegate & Bergen, 1988; Masure, 1988, Figs. 2-3 |
| LOK.8 Bosso Valley, Italy: | Housa et al., 2004 |
| LOK.10 Barret-le-Bas Section, France: | Busnardo et al., 1979, p. 44, Fig. 13; p. 90, Fig. 28; p. 105, Fig. 30; Bulot, 1995, Figs. 3, 6-7 |
| LOK.11 Angles Section, France: | Busnardo et al., 1979 |
| LOK.12 Angles Section (Bulot 1993): | Bulot et al., 1993, Table VI, p. 27, VII, p. 30, Table VIII, p. 35, XII, p. 44, XIV. Bulot, 1995 |
| LOK.13 La Charce Section, France 1995, Hauterivian GSSP: | Bulot et al., 1993, Table VI, p. 27, VII, p. 30, Table VIII, p. 35, XII, p. 44, XIV. Bulot, 1995 |
| LOK.13b La Charce Section, France 2008, Hauterivian GSSP: | Reboulet, 2008, Figs. 2.1, 3.1 |
| LOK.14 Curnier Section, France: | Bulot et al., 1993, Table III, p. 22 |
| LOK.15 Moriez (St-Firmin) Section, France: | Bulot et al., 1993, Table IV, p. 24 |
| LOK.16 Baumugne Section, France: | Bulot et al., 1993, Table V, p. 25 |
| LOK.17 La Charce Combe Reboul, France: | Bulot et al., 1993; Bulot, 1995, this is a key reference section for 2 Upper Hauterivian zones |
| LOK.18 Chamaloc-Col du Rousset, France: | Bulot et al., 1992, Fig. 1.7, p. 40; Hoedemaeker, 2013 |
| LOK.19 Mont Aiguille I, Vercors, France: | Busnardo et al., 1991 |
| LOK.20 Mont Aiguille II, Vercors, France: | Busnardo et al., 1991 |
| LOK.21 Miravetes Section, Spain: | Aguado et al., 2000, Fig. 3 |
| LOK.22 Canada Lengua Section, Spain: | Aguado et al., 2000, Fig. 4 |
| LOK.23 Canada Lengua-2 Section, Spain, Valanginian Candidate GSSP: | Aguado et al., 2000, Fig. 5 |
| LOK.24 Barlya Section, Bulgaria: | Lakova et al., 1997, Fig. 1 |
| LOK.25 Gyangze Section, Tibet : | Wan et al., 2010. |
| LOK.26 Fiume Bosso Section, Italy: | Lowrie & Channell, 1984, magnetostratigraphic data; Bralower et al., 1989, Fig. 3, p. 162 |
| LOK.27 Fonte Giordano Section, Italy: | Bralower et al., 1989, Fig. 4, p. 163 |
| LOK.28 Puerto Escańo Section, South Spain, base Berriasian: | Caracuel et al., 2000; Pruner et al., 2010 |
| LOK.29 Grindstone Creek, California: | Bralower et al., 1990 |
| LOK.30 Miravetes-1 Rio Argos, Spain: | Aguado et al., 2000, Fig. 3 |
| LOK.31 Canada Luenga-2 Rio Argos, Spain: | Aguado et al., 2000, Fig. 4 |
| LOK.32 Canada Luenga-3 Rio Argos, Spain: | Aguado et al., 2000, Fig. 5 |
| LOK.34 Tang-E Asbu, Kuh-E Ginau, Iran: | Edgell, 1967, Fig. 8 |
| LOK.35 Le Chouet, Drome, SE France: | Wimbledon et al., 2013; Frau et al., 2015, Fig. 1, p. 118; Frau et al., 2016, Fig. 1 |
| LOK.36 Crimea West: | Arkadiev et al., 2018 |
| LOK.37 Crimea East: | Arkadiev et al., 2018 |
| LOK.38 Leube Quarry, Salzburg, Austria: | Bujtor et al., 2013 |
| LOK.39 Guidaloca Section, NW Sicily: | Andreini et al., 2007, Figs. 2, 4 |
| LOK.40 Dieni I & II Section, W Sicily: | Andreini et al., 2007, Fig. 3 |
| LOK.41 Polaveno Italy: | Channell & Erba, 1992; Channell et al., 1995, Fig. 12, Table 2 |
| LOK.42 Jebel Rheouis Section, Tunisia: | Maaloui & Zargouni, 2016 |
| LOK.43 Jebel Meloussi Section, Tunisia: | Maaloui & Zargouni, 2016 |
| LOK.44 Nara section, Tunisia: | Maaloui & Zargouni, 2016 |
| LOK.45 Sidi Kralif section, Tunisia: | Maaloui & Zargouni, 2016 |
| LOK.49 Nutzhof Section 2009, Austria [2009+2010]: | Rehakova et al., 2009; Lukenender et al., 2010, Figs. 2, 6 |
| LOK.52 Bruzovice Section, West Carpathians: | Skupien & Doupovcova, 2019 |
| LOK.56 SGT Section, Turkey: | Atasoy, S.G., 2017, ranges from Appendices A-B; Atasoy et al., 2018 |
| LOK.57A Wadi Mi'iadin, Oman 1987: | Simmons & Hart 1987, Fig. 10.7-8, .10-11. |
| LOK.57B Wadi Mi'iadin, Oman 1990: | Scott, 1990 |
| LOK.57C Wadi Mi'iadin, Oman 2016: | Celestino et al., 2016 |
| LOK.60 Tamazunchale, San Luis Potosi, Mexico: | Lopez-Martinez et al., 2015, Fig. 8 |
| LOK.61 Poznachowice Dolne, Poland: | Kedzierski & Ochabska, 2012, Fig. 3, Table 1 |
| LOK.62 Tre Maroua, Drome, SE France | Wimbledon et al., 2020 |
| LOK.63 Rancho San Vicente, SW Cuba: | Pszczółkowski & Myczyński, 2010; López-Martínez et al., 2013 |
| LOK.64 Ain Hammouch, Morocco Val-Haut: | Wippich, 2003, Fig. 5 |
| LOK.65 65 Torre de Busi section Italy: | Erba & Quadrio, 1987; Andreini et al., 2007; Casellato & Erba, 2020, Fig. 5 |
| LOK.66 66 Mt. Pernice section Italy: | Erba & Quadrio, 1987; Casellato & Erba, 2020, Fig. 6 |
| LOK.69 69 Yavorets section, Bulgaria: | Petrova et al., 2019, Fig. 3 |
| LOK.70 Vergol section, France, Vocontian Basin, France: | Kenjo et al.,2021 |
| HA-BACS.1 Catalog File of Hauterivian-Barremian Carbonates: | HA-BA.1 Pont de Laval, France; HA-BA. 2 Pas de l'Essaure, France; HA-BA. 3 Mont Aiguille, France; HA-BA. 4 Grands Goulets, France; HA-BA. 5 Combe de Bella Cha, France; HA-BA. 6 Pic de l'Oeillette, Chartreuse, France; HA-BA. 7 Chames Vivarais, France; HA-BA. 8 Arredons Vivarais, France. Urgo.1 Col de Rousset, SE France, Arnaud et al., 1998, Fig. 21. Urgo.2 Gorges du Nant, Arnaud et al., 1998, Fig. 27. Urgo.3 Gorges du Frou, Arnaud et al., 1998, Fig. 28. Urgo.4 Rocher de Cluses, Arnaud et al., 1998, Fig. 29. MIDK.102 La Russille, Switzerland; MIDK 103 Vaulion, Switzerland; MIDK 104 La Sarraz Eclepens, Switzerland; Gorges de l'Orbe, Switzerland: Arnaud-Vanneau & Masse, 1989; MIDK.103 Vaulion, Switzerland; Arnaud-Vanneau & Masse, 1989; MIDK.104 La Sarraz Éclepens, Switzerland; MIDK.105 Gorges de l'Orbe, Switzerland; MIDK.106 Gellin-Rochejean, Switzerland. BACI.1 Mt. Croce Section, Italy; BACI.2 Mt. Motola Section, Italy; BACI.3 Mt. Coccovello Section, Italy; BACI.4 Mt. Raggeto Section, Italy; BACI.5 Mt. Tobenna Section, Italy: Di Lucia et al., 2012, Fig. 8. |
| Tatra.1 Posrednie III Section, Poland; Tatra.2 Posrednie II Section, Poland; Tatra.3 Rowienka Section, Poland: | Grabowski & Pszczolkowski, 2006, Figs. 4-6 |
| ANDESCS.1 Andes Sections Argentina-Chile–ANDES DB, see Appendix 3. |
Appendix 2: LOK2016 DB 09/17/2021: Bioevents, Polarity Chrons, Radioisotopic Ages, and Sequences (SB) in Mega-annums calibrated to GTS2016. Asterisks = no data. Negative sign an artifact of orientation of X/Y plot.
| Acaenolithus vimineus | -130.7224 | -130.2981 |
| Acanthodiscus radiatus | -134.7294 | -134.1235 |
| Acanthodiscus rebouli | -134.7887 | -134.1235 |
| Achomosphaera neptuni | -139.7813 | -130.4596 |
| Acrioceras pulcherrinum | -131.5870 | -131.4780 |
| Acrioceras puzosianum | -130.8221 | -130.8221 |
| Acrioceras seringuei | -131.3472 | -131.3472 |
| Acrioceras tabarelli | -130.8640 | -130.8539 |
| Agrio tuff bed 126.97 | -130.1558 | *** |
| Agrio tuff bed 127.42 | -130.2259 | *** |
| Agrio tuff bed 129.09 | -131.7887 | *** |
| Agrio tuff bed 130.40 | -133.9840 | *** |
| Aldorfia dictyotum | -135.1896 | -135.1896 |
| Alvellodinium falsificum | -136.8908 | -136.5085 |
| Amphizygus brooksii | -132.8665 | -130.6650 |
| Amphizygus infracretacea | -134.9580 | -133.5160 |
| Amphorellina lanceolata | -140.4337 | *** |
| Amphorula delicata | -140.0724 | -138.8577 |
| Amphorula metaelliptica | -142.6067 | -137.7134 |
| Ancyloceras vandenheckii | -129.1604 | -129.1604 |
| Andes J-K SB 1 | -147.7527 | *** |
| Andes J-K SB 2 | -145.9279 | *** |
| Andes J-K SB 3 | -142.6842 | *** |
| Andes J-K SB 4 | -140.5289 | *** |
| Andiceras acuticostum | -148.1857 | -146.5407 |
| Aprobolocysta eilema | -131.9987 | -131.2840 |
| Apteodinium maculatum | -136.2827 | -133.9153 |
| Ardesciella rhodanica | -145.8887 | -145.8887 |
| Argentiniceras fasciculatum | -143.8925 | -141.6715 |
| Argentiniceras noduliferum | -144.4639 | -141.9289 |
| Aspidoceras depressus | -139.4232 | -139.4232 |
| Aspidoceras euomphalum | -148.2864 | -144.9288 |
| Aspidoceras quinchaoi | -146.7752 | -146.3036 |
| Aspidoceras rogoznicensis | -145.9821 | -144.0502 |
| Assipetra infracretacea | -144.8638 | -132.4596 |
| Assipetra terebrodentarius | -130.3153 | -130.2755 |
| Aulacosphinctes proximus | -148.6696 | -143.7945 |
| Aulacosphinctes sulcatus | -147.8308 | -146.3750 |
| Avramidiscus kiliani | -130.7834 | -130.4925 |
| Axopodorhabdus dietzmannii | -139.0080 | -130.0161 |
| Balearites balearis | -131.3690 | -131.0289 |
| Baronnites hirsutus | -137.5468 | -137.1511 |
| Barremites spp. | -131.1221 | -125.8838 |
| Base Albian | -113.1400 | *** |
| Base Aptian | -126.3000 | *** |
| Base Barremian | -130.8600 | *** |
| Base Berriasian | -145.7000 | 145.0000 |
| Base Hauterivian | -134.7000 | *** |
| Base Valanginian | -139.4000 | *** |
| Batioladinium gochtii | -136.8908 | -134.8651 |
| Batioladinium varigranosa | -137.7466 | -135.1885 |
| Berriasella bebrovensis | -142.6661 | -140.2646 |
| Berriasella callisto | -141.5873 | -139.4634 |
| Berriasella chomeracensis | -145.7500 | -144.0591 |
| Berriasella jacobi | -145.7583 | -143.7890 |
| Berriasella malbosi | -144.6594 | -140.2568 |
| Berriasella paramacillenta | -144.9717 | -143.8667 |
| Berriasella picteti | -142.6421 | -139.2625 |
| Berriasella privasensis | -143.8669 | -142.0898 |
| Berriasella subcallisto | -145.7340 | -143.6672 |
| Berriasella subprivasensis | -144.1210 | -144.1210 |
| Berriasella tithonica | -146.3000 | -145.8000 |
| Biorbifera johnewingii | -142.8262 | -135.4659 |
| Biscutum constans | -145.9889 | -130.0000 |
| Blanfordiceras vetustum | -145.7957 | -143.9674 |
| Bochianites neocomiensis | -138.8470 | -134.6804 |
| Bochianites oosteri | -134.7000 | -134.6256 |
| Borzaiella atava | -142.1924 | -141.0966 |
| Borzaiella slovenica | -148.9733 | -146.4902 |
| Boughdiriella chouetensis | -146.1110 | -145.8887 |
| Bourkidinium granulatum | -133.4430 | -130.9993 |
| Braarudosphaera bigelowii | -140.0507 | -134.3880 |
| Braarudosphaera discula | -134.9864 | -133.5160 |
| Braarudosphaera regularis | -134.1125 | -132.2382 |
| Breistrofferella castellanensis | -134.7282 | -134.0627 |
| Breistrofferella varappensis | -134.6804 | -134.4245 |
| Buchia pacifica | -137.6250 | -135.5000 |
| Buchia uncitoides | -139.0000 | -138.0750 |
| Bukrylithus ambiguus | -139.0080 | -130.0161 |
| Burckhardticeras peroni | -147.8308 | -147.8308 |
| Busnardoiceras busnardoi | -146.0433 | -145.8887 |
| Busnardoites campylotoxum | -138.5141 | -135.6475 |
| Busnardoites desori | -136.8564 | -135.6475 |
| Busnardoites subcampylotoxum | -136.795 | -136.795 |
| Caddasphaera halosa | -138.0278 | -134.8203 |
| Cadosina fusca | -146.8745 | -141.4428 |
| Cadosina fusca cieszynica | -145.2419 | -145.2419 |
| Cadosina semiradiata | -149.3071 | -143.8031 |
| Calcicalathina oblongata | -139.5554 | -130.5848 |
| Calcicalathina praeoblongata | -141.3878 | -138.5064 |
| Callaiosphaeridium asymmetricum | -134.153 | -130.00 |
| Calpionella alpina | -146.9167 | -134.0660 |
| Calpionella elliptalpina | -146.4940 | -145.0320 |
| Calpionella elliptica | -144.9150 | -139.2050 |
| Calpionella grandalpina | -146.7081 | -143.2117 |
| Calpionella minuta | -142.1674 | *** |
| Calpionellites caravacaensis | -139.4586 | -138.4756 |
| Calpionellites coronata | -139.3957 | -136.5576 |
| Calpionellites darderi | -139.4672 | -133.5600 |
| Calpionellites major | -139.3957 | -133.5600 |
| Calpionellopsis oblonga | -143.3071 | -133.5600 |
| Calpionellopsis simplex | -144.4744 | -133.5600 |
| Canninginopsis colliveri | -134.0444 | -132.7406 |
| Carbon peak Valanginian OAEb | -138.963 | -138.769 |
| Carbon peak Valanginian OAEc | -136.381 | *** |
| Carbon peak Valanginian OAEd | -134.1475 | *** |
| Carpistomiosphaera borzai | -150.8000 | -149.1181 |
| Carpistomiosphaera tithonica | -149.1700 | -148.8213 |
| Carpistomiosphaera valanginiana | -139.160 | -138.830 |
| Cassiculosphaeridia reticulat | -134.1811 | -132.5330 |
| Catutosphinctes americanensis | -148.380 | -146.817 |
| Catutosphinctes guenenokenensis | -150.87 | -149.51 |
| Catutosphinctes inflatus | -146.4850 | -145.6913 |
| Catutosphinctes proximus | -148.5300 | -146.4675 |
| Cerbia tabulata | -132.7406 | -132.5330 |
| Chacantuceras ornatum | -135.1895 | -134.9810 |
| Cheloniceras spp. | -125.6695 | -125.6695 |
| Chiastozygus bilamellus | -137.2383 | -134.2975 |
| Chiastozygus litterarius | -133.5523 | -132.5309 |
| Chiastozygus platyrhethus | -133.5035 | -132.5414 |
| Chiastozygus striatus | -135.0461 | -130.5160 |
| Chiastozygus tenuis | -136.8009 | -133.5160 |
| Chigaroceras loteroense | -146.7041 | -144.3597 |
| Chitinoidella boneti | -147.7338 | -145.4263 |
| Chitinoidella dobeni | -147.8319 | -147.0225 |
| Chitinoidella elongata | -147.5023 | -147.0316 |
| Chitinoidella hegarati | -147.5443 | -146.7081 |
| Chitinoidella slovenica | -147.8308 | -146.2811 |
| Chlamydophorella huguoniotii | -135.695 | -133.805 |
| Chlamydophorella membranoidea | -133.738 | -133.76 |
| Chlamydophorella nyei | -135.2687 | -130.000 |
| Choicensisphinctes burckhardti | -150.68 | -149.227 |
| Choicensisphinctes choicensis | -150.150 | -148.304 |
| Choicensisphinctes erinoides | -150.221 | -147.295 |
| Choicensisphinctes platyconus | -150.873 | -149.748 |
| Choicensisphinctes striolatus | -145.378 | -143.203 |
| Choicensisphinctes windhauseni | -149.244 | -148.639 |
| Cieneguiticeras falculatum | -149.670 | -147.1275 |
| Cieneguiticeras perlaevis | -149.9339 | -148.6956 |
| Circulodinium distinctum | -142.2500 | -130.0000 |
| Clavihedbergella eocretacea | -130.2629 | -125.608 |
| Clavihedbergella semielongata | -130.263 | -125.608 |
| Clavithurmannia foraticostata | -140.076 | -139.782 |
| Clepsilithus maculosus | -135.0204 | -130.2356 |
| Colchidites sp. | -127.1700 | -126.0369 |
| Colomisphaera carpathica | -147.4556 | -141.6868 |
| Colomisphaera cieszynica | -147.8319 | -143.8745 |
| Colomisphaera conferta | -139.8093 | -138.6887 |
| Colomisphaera fortis | -147.5243 | -143.6067 |
| Colomisphaera helicosphaera | -139.255 | -138.7358 |
| Colomisphaera lapidosa | -147.4556 | -139.3957 |
| Colomisphaera lucida | -145.1938 | -143.8944 |
| Colomisphaera nagyi | -150.8000 | -147.0225 |
| Colomisphaera pieniniensis | -150.8000 | -147.0225 |
| Colomisphaera radiata | -147.4556 | -147.4556 |
| Colomisphaera tenuis | -148.1071 | -143.3798 |
| Colomisphaera volgeri | -139.1604 | -138.6887 |
| Cometodinium whitei | -138.6217 | -130.1032 |
| Cometodinium? whitei | -135.9912 | -132.7406 |
| Comittosphaera sublapidosa | -147.4556 | -146.4450 |
| Conusphaera maledicto | -150.0800 | *** |
| Conusphaera mexicana | -149.6262 | -130.0000 |
| Conusphaera mexicana minor | -149.636 | -140.6229 |
| Corollithion acutum | -136.7605 | -132.5927 |
| Corollithion ellipticum | -139.0080 | -132.4596 |
| Corollithion geometricum | -134.3456 | -130.5618 |
| Corollithion signum | -135.3959 | -135.3959 |
| Corongoceras alternans | -146.4757 | -145.9821 |
| Corongoceras evolutum | -146.4386 | -146.2102 |
| Corongoceras involutum | -146.4014 | -146.2968 |
| Corongoceras koellikeri | -145.2441 | -143.5279 |
| Corongoceras koeni | -146.4014 | -146.1786 |
| Corongoceras lotenoense | -147.5985 | -145.9506 |
| Corongoceras mendozanum | -147.1025 | -144.0502 |
| Corongoceras multimum | -146.4386 | -146.4386 |
| Corongoceras praecursor | -148.5164 | -146.8825 |
| Corongoceras steinmanni | -145.7957 | -145.7957 |
| Coronifera oceanica | -132.6670 | -130.0000 |
| Crassicollaria brevis | -146.6856 | -144.4639 |
| Crassicollaria colomi | -146.4554 | -144.5822 |
| Crassicollaria intermedia | -147.0800 | -144.9200 |
| Crassicollaria massutiniana | -147.6800 | -143.8944 |
| Crassicollaria parvula | -147.6800 | -139.8093 |
| Crassiculosphaeridia reticulata | -130.573 | -130.000 |
| Cretarhabdus angustiforatus | -141.900 | -135.500 |
| Cretarhabdus conicus | -142.8262 | -130.0000 |
| Cretarhabdus crenulatus | -142.0705 | -134.3880 |
| Cretarhabdus loriei | -135.6338 | -132.5414 |
| Cretarhabdus octofenestratus | -145.004 | -140.979 |
| Cretarhabdus striatus | -134.8215 | -130.4350 |
| Cretarhabdus surirellus | -146.3025 | -130.1558 |
| Cribellopsis elongata | -131.4775 | -130.5234 |
| Cribellopsis neoelongata | -131.0552 | -130.4745 |
| Cribellopsis schroederi | -130.8380 | -130.7193 |
| Cribellopsis thieuloyi | -130.8380 | -130.7193 |
| Cribroperidinum sepimentum | -133.002 | -130.000 |
| Cribrosphaerella ehrenbergii | -135.3959 | -135.3959 |
| Crioceratites andinum | -131.5133 | -131.4798 |
| Crioceratites basseae | -131.3690 | -131.3690 |
| Crioceratites diamantense | -131.5196 | -131.4798 |
| Crioceratites duvalii | -132.3500 | -131.8057 |
| Crioceratites fabreae | -131.3254 | -131.2818 |
| Crioceratites gr. quenstedti | -133.4112 | -132.1908 |
| Crioceratites loryi | -134.2881 | -133.8625 |
| Crioceratites majorisensis | -131.6088 | -131.4344 |
| Crioceratites matsumotoi | -132.0532 | -131.9672 |
| Crioceratites nolani | -134.4682 | -131.2448 |
| Crioceratites perditum | -131.5053 | -131.4239 |
| Crioceratites remanei | -131.3690 | -131.3690 |
| Crioceratites schlagintweiti | -131.7270 | -131.7110 |
| Criohimantoceras gigas | -136.0119 | -134.9787 |
| Criosarasinella furcillata | -135.4300 | -135.0247 |
| Criosarasinella heterocostata | -135.430 | -135.193 |
| Criosarasinella mandovi | -135.2400 | -134.9506 |
| Cruasiceras cruasense | -132.2300 | -132.1683 |
| Crucibiscutum nequenensis | -131.7509 | -131.2963 |
| Crucibiscutum salebrosum | -134.9864 | -133.5160 |
| Cruciellipsis chiasta | -140.0507 | -134.5708 |
| Cruciellipsis cuvillieri | -147.0599 | -131.5509 |
| Cruciplacolithus furtivus | -132.8665 | -131.4172 |
| Cruciplacolithus salebrosus | -140.5103 | -135.6505 |
| Ctenidodinium elegantulum | -135.4659 | -134.6907 |
| Ctenidodinium scissum | -135.8210 | -135.8210 |
| Cuyaniceras raripartitum | -141.7864 | -140.9119 |
| Cuyaniceras transgrediens | -143.5279 | -139.4232 |
| Cyclagelosphaera deflandrei | -146.8181 | -130.7779 |
| Cyclagelosphaera margerelii | -148.6707 | -130.0000 |
| Cyclagelosphaera tubulata | -131.5944 | -131.5944 |
| Cyclonephelium distinctum | -134.4564 | -132.7406 |
| Cyclonephelium hystrix | -139.7813 | -133.6636 |
| Cymatiosphaera delicatula | -134.2732 | -133.5532 |
| Cymososphaeridium validum | -136.3043 | -131.0296 |
| Dalmasiceras biplanum | -144.9607 | -144.9607 |
| Dalmasiceras crassicostatum | -145.4730 | -145.4730 |
| Dalmasiceras dalmasi | -142.8000 | -142.6902 |
| Dalmasiceras djanelidzei | -144.8350 | -144.8350 |
| Dalmasiceras punctatum | -142.8000 | -141.8496 |
| Dalmasiceras subloevis | -145.2603 | -144.8780 |
| Dapsilidinium warrenii | -139.5724 | -130.3853 |
| Decliveites agrioensis | -134.4865 | -134.3429 |
| Decliveites crassicostatum | -134.7178 | -134.7178 |
| Deshayesites oglanlensis | -126.3000 | -126.3000 |
| Deshayesites weissi | -126.0369 | -125.8991 |
| Diadorhombus rectus | -139.0080 | -131.7612 |
| Diazomatolithus lehmanii | -146.6772 | -130.0000 |
| Dicanthum hollisteri | -142.8262 | -130.0000 |
| Dichotomites petschi | -136.2850 | -135.9287 |
| Dichotomites vergunnorum | -135.4482 | -135.3385 |
| Diloma placinum | -133.6829 | -133.6118 |
| Dingodinium albertii | -137.3749 | -133.3539 |
| Dingodinium cerviculum | -138.9044 | -130.0000 |
| Dingodinium europaeum | -129.0379 | -127.2312 |
| Discorhabdus biradiatus | -133.7892 | -133.6447 |
| Discorhabdus ignotus | -142.8098 | -130.0000 |
| Discorhabdus rotatorius | -142.5769 | -133.5160 |
| Discorsia nanna | -133.8054 | -133.8054 |
| Djurjuriceras catutosense | -146.4084 | -146.3036 |
| Dobeniella bermudezi | -147.5489 | -146.9398 |
| Dobeniella cubensis | -147.5489 | -146.9398 |
| Druggidium apicopaucicum | -138.4730 | -131.9987 |
| Druggidium deflandrei | -135.6038 | -127.1240 |
| Druggidium rhabdoreticulatum | -136.8969 | -128.9920 |
| Durangites acanthicus | -146.3000 | -145.8000 |
| Durangites astillerensis | -146.3000 | -145.8000 |
| Durangites vulgaris | -146.3000 | -145.8000 |
| Eiffellithus primus | -148.0991 | -137.0000 |
| Eiffellithus striatus | -135.3000 | -131.7509 |
| Eiffellithus windi | -139.2677 | -132.5450 |
| Elenaelia cularensis | -145.7340 | -145.6760 |
| Eleniceras nikolovi | -134.7718 | -134.4611 |
| Eleniceras tchechitevi | -135.4683 | -134.6804 |
| Eleniceras transsylvanicum | -134.9075 | -134.7650 |
| Emericiceras emerici | -130.8640 | -130.7513 |
| Endoscrinium campanula | -142.2500 | -130.0000 |
| Endoscrinium glabra | -136.5227 | -136.5085 |
| Eopalorbitolina charollaisi | -130.8100 | -130.7437 |
| Eopalorbitolina pertenuis | -130.8221 | -130.7437 |
| Eprolithus floralis | -131.2166 | -131.0969 |
| Erdenella paquieri | -141.6455 | -138.6995 |
| Erdenella zianidia | -141.7535 | -141.0331 |
| Escharisphaeridia pocockii | -142.8262 | -142.8262 |
| Ethmorhabdus gallicus | -145.9784 | -133.5160 |
| Ethmorhabdus hauterivianus | -139.7364 | -130.4350 |
| Euptychoceras meyrati | -133.4112 | -132.1908 |
| Euvirgalithacoceras malarguense | -150.3983 | -148.6394 |
| Exiguisphaera phragma | -137.7466 | -135.2687 |
| Falsurgonina pileola | -131.4775 | -130.7193 |
| Falsurgonina vanneauae | -131.5768 | -130.4745 |
| Fauriella boissieri | -141.6945 | -139.3891 |
| Fauriella donzei | -140.0000 | -139.8813 |
| Fauriella gallica | -142.6902 | -140.8890 |
| Fauriella kiliani | -140.0965 | -138.1346 |
| Fauriella rarefurcata | -143.2876 | -139.9524 |
| Faviconus multicolumnatus | -150.7160 | -144.4940 |
| Flabellites oblonga | -133.5933 | -132.6252 |
| Foucheria modesta | -140.0724 | -136.6659 |
| Frenguelliceras magister | -143.1385 | -141.6715 |
| Fromea amphora | -135.0502 | -135.0502 |
| Gaarderella granulifera | -130.3600 | -130.0161 |
| Glob'oides algeriana | -126.4917 | -126.4028 |
| Glob'oides aptiense | -127.1700 | -126.7106 |
| Glob'oides blowi | -127.9355 | -125.6082 |
| Glob'oides gottisi | -135.9654 | -125.6082 |
| Glob'oides maridalensis | -125.6082 | -122.5459 |
| Globochaete alpina | -144.6595 | *** |
| Globuligerina hoterivica | -138.3380 | -125.6695 |
| Gonyaulacysta helicoidea | -145.7073 | -130.0000 |
| Gonyaulacysta kostromiensis | -136.5085 | -133.1871 |
| Grantarhabdus meddii | -139.0080 | -132.4492 |
| Groebericeras bifrons | -143.8400 | -141.9926 |
| Groebericeras rocardi | -143.1393 | -143.1007 |
| Gubkinella graysonensis | -135.1336 | -125.6695 |
| Haploceras staszycii | -149.6700 | -149.2267 |
| Haplophylloceras strigile | -144.6797 | *** |
| Haqius circumradiatus | -142.9085 | -130.0000 |
| Hayesites atlanticus | -135.0461 | -133.6447 |
| Hayesites radiatus | -133.9498 | -130.0711 |
| Hedb aptiana | -130.2629 | -125.6082 |
| Hedb aptica | -138.3380 | -125.6082 |
| Hedb delrioensis | -135.5587 | -125.6082 |
| Hedb excelsa | -125.6695 | -125.6695 |
| Hedb kuznetsovae | -130.2629 | -125.6695 |
| Hedb sigali | -138.3380 | -125.6695 |
| Hedb similis | -130.4925 | -125.6082 |
| Heinzia provincialis | -129.1604 | -129.1604 |
| Heinzia sartousi | -127.4149 | -127.3078 |
| Helenea chiastia | -148.6707 | -130.0000 |
| Heterosphaeridium? galliciae | -135.9912 | -132.7406 |
| Hexalithus geometricus | -146.7013 | -144.6900 |
| Hexalithus magharensis | -140.8365 | -140.8365 |
| Hexalithus noelae | -148.7000 | -143.3986 |
| Himalayites treubi | -142.9481 | -142.9481 |
| Himantoceras trinodosum | -136.0119 | -134.6937 |
| Holcodiscus caillaudianus | -130.0791 | -129.6351 |
| Holcophylloceras calypso | -144.8463 | -137.1906 |
| Holcoptychites agrioensis | -134.0638 | -134.0638 |
| Holcoptychites magdalensae | -134.3031 | -134.1356 |
| Holcoptychites neuquensis | -134.2861 | *** |
| Hoplytocrioceris gentilli | -133.7847 | -133.6252 |
| Hoplytocrioceris giovinei | -133.8644 | -133.8405 |
| Hypophylloceras courchonense | -135.5761 | -135.3385 |
| Hypophylloceras perlobatum | -137.7457 | -136.6546 |
| Hystrichodinium furcatum | -136.8969 | -133.1871 |
| Hystrichodinium pulchrum | -138.8068 | -131.1072 |
| Hystrichodinium voigtii | -142.2500 | -130.3853 |
| Indansites malarguensis | -150.3983 | -149.9233 |
| Inoceramus everesti | -144.0392 | *** |
| Jeanthieuloyites quinquestriatus | -135.4447 | -134.6433 |
| Karakaschiceras attenuatum | -137.0024 | -136.0532 |
| Karakaschiceras biasalense | -138.0339 | -136.4899 |
| Karakaschiceras heteroptychum | -136.7607 | -136.7734 |
| Karakaschiceras neumayri | -137.0780 | -136.8407 |
| Karakaschiceras pronecostatum | -136.7125 | -135.3691 |
| Kilianella busnardoi | -141.9457 | -141.7535 |
| Kilianella gr.chamalocensis | -141.3182 | -138.9727 |
| Kilianella lucensis | -139.7691 | -136.2014 |
| Kilianella pexiptycha | -139.9406 | -138.9333 |
| Kilianella retrocostata | -140.0965 | -137.6375 |
| Kilianella roubaudi | -139.0340 | -136.2014 |
| Kilianella roubaudiana | -139.1867 | -137.0405 |
| Kilianella superba | -137.0660 | -137.0405 |
| Kiokansium polypes | -137.7466 | -130.0000 |
| Kleithriasphaeridium corrugatum | -137.3749 | -137.3749 |
| Kleithriasphaeridium eoinodes | -133.7790 | -132.5487 |
| Kleithriasphaeridium fasciatum | -137.3302 | -132.5487 |
| Kleithriasphaeridium simplicispinum | -137.2264 | -136.5227 |
| Krantziceras azulense | -145.3783 | -145.3783 |
| Krantziceras compressum | -141.9926 | -141.9289 |
| Krantziceras disputabile | -147.1025 | -147.1025 |
| Krantziceras planulatum | -142.5022 | -142.3111 |
| Kutekiceras pseudocolubrinus | -147.8308 | -147.8308 |
| Laeviaptychus crassissimus | -149.9176 | -147.9057 |
| Laeviaptychus latus | -147.9692 | -146.5505 |
| Leopoldia buxtorfi | -134.4345 | -134.0750 |
| Leopoldia leopoldina | -134.5275 | -134.4087 |
| Leptoceras studeri | -141.1600 | -138.6778 |
| Leupoldina cabri | -125.6082 | -125.6082 |
| Leupoldina pustulans | -130.2629 | -125.6082 |
| Leymeriella schrammeni anterior | -113.1000 | -113.1000 |
| Lissonia riveroi | -139.2741 | -138.5129 |
| Lithastrinus septentrionalis | -132.8665 | -130.0000 |
| Lithoceras picunleufuense | -150.8733 | -149.7480 |
| Lithraphidites bollii | -133.9498 | -130.2356 |
| Lithraphidites carniolensis | -147.7160 | -130.0000 |
| Lorenziella hungarica | -143.9394 | -137.2302 |
| Lorenziella plicata | -145.3436 | -133.8360 |
| Luppovella superba | -137.5627 | -136.9573 |
| Lyticoceras nodosoplicatum | -133.5537 | -133.3875 |
| Lyticoceras subfimbriatum | -134.0600 | -134.0600 |
| Lytoceras juileti | -137.7457 | -136.3597 |
| Lytoceras montanum | -144.9609 | -144.9609 |
| Lytohoplites burckhardti | -146.3387 | -146.2804 |
| Lytohoplites rauloi | -146.4386 | -145.3387 |
| Lytohoplites vareloe | -146.2968 | -145.6480 |
| Lytohoplites zambranoi | -146.4757 | -145.6480 |
| Magnetochron CM0R | -126.3000 | -126.0000 |
| Magnetochron CM10 | -133.9313 | -133.6694 |
| Magnetochron CM10N | -135.3408 | -134.3082 |
| Magnetochron CM1R | *** | -127.5912 |
| Magnetochron CM2 | *** | -128.0208 |
| Magnetochron CM3R | -130.8011 | -128.6813 |
| Magnetochron CM5R | -131.7071 | -131.4307 |
| Magnetochron CM6R | -131.8776 | -131.7973 |
| Magnetochron M6 | *** | -132.0222 |
| Magnetochron M7 | *** | -132.1807 |
| Magnetochron CM7R | -132.3155 | -131.9912 |
| Magnetochron CM8R | -132.7334 | -132.5194 |
| Magnetochron M8 | *** | -133.0312 |
| Magnetochron CM9R | -133.4655 | -133.0008 |
| Magnetochron M9 | *** | -133.6578 |
| Magnetochron M11r | *** | -136.0997 |
| Magnetochron M12n | *** | -136.9000 |
| Magnetochron M12r | *** | -137.7535 |
| Magnetochron M13n | *** | -138.3000 |
| Magnetochron M13r | *** | -138.4059 |
| Magnetochron M14n | *** | -138.6000 |
| Magnetochron M14r | *** | -139.2348 |
| Magnetochron M15n | *** | -139.4462 |
| Magnetochron M15r | *** | -139.9000 |
| Magnetochron M16n | *** | -140.4000 |
| Magnetochron M16r | *** | -141.2865 |
| Magnetochron M17n | *** | -142.1538 |
| Magnetochron M17r | *** | -142.5645 |
| Magnetochron M18n | *** | -143.3173 |
| Magnetochron M18r | *** | -143.7134 |
| Magnetochron M19n | *** | -144.6496 |
| Magnetochron M19n.1n | *** | -145.0000 |
| Magnetochron M19n.1r | *** | -145.1333 |
| Magnetochron M19n.2n | *** | -145.3000 |
| Magnetochron M19r | *** | -145.8432 |
| Magnetochron M20n | *** | -146.1054 |
| Magnetochron M20n.1n | *** | -146.1689 |
| Magnetochron M20n.1r | *** | -146.3727 |
| Magnetochron M20n.2n | *** | -146.5132 |
| Magnetochron M20r | *** | -147.1557 |
| Magnetochron M21n | *** | -147.5443 |
| Magnetochron M21r | -149.2386 | -148.4143 |
| Magnetochron M22A | *** | -150.5000 |
| Magnetochron M22n | *** | -148.7717 |
| Magnetochron M22r | *** | -150.0481 |
| Malbosiceras malbosi | -144.0857 | -143.7945 |
| Manivitella pemmatoidea | -146.1787 | -130.0000 |
| Markalius circumradiatus | -142.5769 | -134.3880 |
| Markalius inversus | -130.6982 | -130.6982 |
| Marker bed 134.0 U-Pbf | *** | -134.0660 |
| Marker bed 138.45 | -138.0000 | -136.0000 |
| Marker bed 139.85 Sr | -139.8500 | -139.8500 |
| Marker bed 305 | -136.9518 | *** |
| Marker bed 321 | -136.5449 | *** |
| Marker bed A 137.1 | -138.0625 | -138.0525 |
| Marker bed Ap SB SL 1 | -126.5805 | -126.5805 |
| Marker bed B 137.1 | -136.4375 | -136.4275 |
| Marker bed Ba1 | -130.8446 | *** |
| Marker bed Ba2 | -130.7936 | *** |
| Marker bed Faraoni bed | -131.0776 | -131.0539 |
| Marker bed Ha6 | -131.5340 | *** |
| Marker bed Ha7 | -131.1679 | *** |
| Marker bed SbB3 | -131.4278 | *** |
| Marker bed Tuff 136 | -133.1500 | *** |
| Mazatepites arredondense | -149.6700 | -147.8187 |
| Mazenoticeras paramimounum | -142.2667 | -140.8890 |
| Mazenoticeras tarini | -145.7823 | -145.6760 |
| Megacrioceras doublieri | -131.4126 | -131.4126 |
| Meiourogonyaulax pertusa pertusa | -137.1198 | -131.9987 |
| Meiourogonyaulax stoveri | -134.4863 | -127.0475 |
| Micracanthoceras lamberti | -147.1504 | -146.5470 |
| Micracanthoceras microcanthum | -147.5538 | -145.5449 |
| Micracanthoceras spinulosum | -146.2157 | -143.9568 |
| Micracanthoceras vetustum | -144.5726 | -144.0502 |
| Micrantholithus hoschulzii | -142.2385 | -130.1000 |
| Micrantholithus obtusus | -141.1637 | -130.1399 |
| Micrantholithus speetonensis | -142.8851 | -138.6887 |
| Microhedbergella renilaevis | -113.1400 | -113.1400 |
| Microstaurus chiastius | -148.8920 | -135.5000 |
| Microstaurus quadratus | -146.0433 | -133.5160 |
| Micula infracretacea | -141.1637 | -134.3880 |
| Montseciella alguerensis | -131.0252 | -130.7279 |
| Montseciella glanensis | - 131.4458 | -130.7279 |
| Moravisphinctes fischeri | -146.5267 | -146.1400 |
| Moravisphinctes moravicus | -147.5538 | -146.3750 |
| Muderongia brachialis | -131.7188 | -131.7188 |
| Muderongia extensiva | -136.0875 | -136.0163 |
| Muderongia perforata | -135.2687 | -134.0444 |
| Muderongia simplex | -142.6012 | -134.2183 |
| Muderongia simplex microperforata | -137.7466 | *** |
| Muderongia staurota | -134.5668 | -128.5939 |
| Nannoconus bermudezii | -143.3213 | -130.0000 |
| Nannoconus boletus | -132.7014 | -132.7014 |
| Nannoconus bonetii | -133.6187 | -133.6187 |
| Nannoconus broennimannii | -145.2244 | -133.3458 |
| Nannoconus bucheri | -135.6983 | -130.0000 |
| Nannoconus circularis | -135.0293 | -130.2356 |
| Nannoconus colomii | -145.8403 | -130.0000 |
| Nannoconus compressus | -148.0278 | -145.3562 |
| Nannoconus cornutus | -145.9014 | -133.2976 |
| Nannoconus dolomiticus | -144.6766 | -141.0086 |
| Nannoconus elongatus | -135.6983 | -130.6982 |
| Nannoconus erbae | -146.6348 | -143.9735 |
| Nannoconus globulus | -146.2516 | -130.0000 |
| Nannoconus globulus globulus | -145.7047 | -130.1399 |
| Nannoconus globulus minor | -147.0800 | -133.2344 |
| Nannoconus infans | -148.1960 | -141.0538 |
| Nannoconus kamptneri | -144.3913 | -130.1399 |
| Nannoconus kamptneri minor | -145.0573 | -140.5965 |
| Nannoconus ligius | -131.2963 | -130.2259 |
| Nannoconus puer | -147.7160 | *** |
| Nannoconus quadratus | -143.9989 | -138.8302 |
| Nannoconus steinmannii | -145.2957 | -127.7118 |
| Nannoconus steinmannii minor | -145.9020 | -134.4311 |
| Nannoconus truitti | -141.0538 | -130.4589 |
| Nannoconus wassallii | -134.9132 | -126.3093 |
| Nannoconus wintereri | -146.3026 | -143.3310 |
| Neocomiceramus curacoensis | -131.6698 | -131.1772 |
| Neocomites callidiscus | -134.9787 | -134.9075 |
| Neocomites crassicostatum | -141.5930 | -141.4653 |
| Neocomites flucticulus | -134.8125 | -134.4087 |
| Neocomites neocomiensiformis | -138.5501 | -135.1636 |
| Neocomites neocomiensis | -139.4961 | -135.2914 |
| Neocomites pachydicranus | -135.4300 | -134.0600 |
| Neocomites peregrinus | -136.4329 | -135.9287 |
| Neocomites platycostatus | -136.9518 | -135.2871 |
| Neocomites polygonius | -135.0975 | -135.0975 |
| Neocomites premolicus | -139.3702 | -137.7446 |
| Neocomites subquadratus | -138.5865 | -136.7086 |
| Neocomites subtenuis | -137.1650 | -135.2914 |
| Neocomites teschenensis | -137.1927 | -135.8337 |
| Neocomites wichmanni | -139.4472 | -139.1555 |
| Neocosmoceras malbosiforme | -139.4232 | -139.3429 |
| Neocosmoceras sayni | -143.8189 | -143.1007 |
| Neohoploceras arnoldi | -137.0780 | -136.8407 |
| Neohoploceras depereti | -136.7125 | -136.1425 |
| Neohoploceras provinciale | -137.3214 | -135.5331 |
| Neohoploceras submartini | -136.9518 | -136.4901 |
| Neolisoceras salinarium | -137.8633 | -135.9640 |
| Neolissoceras aberrans | -137.4128 | -137.4128 |
| Neolissoceras grasianum | -141.6945 | -131.8057 |
| Occisucysta tentorium | -138.5607 | -130.2437 |
| Octopodorhabdus decussatus | -137.1751 | -135.6505 |
| Octopodorhabdus polytretus | -132.4927 | -131.3415 |
| Octopodorhabdus reinhardtii | -134.2539 | -133.4788 |
| Odontochitina operculata | -133.0018 | -127.0475 |
| Olcostephanus atherstoni | -137.2291 | -136.8407 |
| Olcostephanus balestrai | -136.1859 | -135.2162 |
| Olcostephanus densicostatus | -135.4116 | -134.0627 |
| Olcostephanus drumensis | -140.2899 | -137.4281 |
| Olcostephanus guebhardi | -137.0660 | -135.8612 |
| Olcostephanus hispanicus | -134.6758 | -134.6758 |
| Olcostephanus jeannoti | -134.3124 | -133.4523 |
| Olcostephanus josephinus | -137.5468 | -137.1511 |
| Olcostephanus laticosta | -133.9681 | -133.9202 |
| Olcostephanus nicklesi | -136.0119 | -135.3385 |
| Olcostephanus sayni | -133.9547 | -133.7912 |
| Olcostephanus stephanophorous | -137.6945 | -135.7266 |
| Olcostephanus tenuituberculatus | -138.5141 | -133.2827 |
| Olcostephanus thieuloyi | -135.3706 | -135.2471 |
| Olcostephanus variegatus | -133.5775 | -133.4587 |
| Oligosphaeridium complex | -137.4847 | -130.0000 |
| Oligosphaeridium dividuum | -134.6907 | -133.4430 |
| Oligosphaeridium pulcherrimum | -135.8581 | -130.2078 |
| Oligosphaeridium verrucosum | -132.5487 | *** |
| Oloriziceras magnum | -147.7615 | -147.6231 |
| Oloriziceras salariensis | -147.7615 | -147.6231 |
| Oosterella cultrata | -135.3113 | -134.4682 |
| Oosterella cultrataeformis | -135.5013 | -134.4245 |
| Oosterella fascigera | -136.4038 | -135.8337 |
| Oosterella garciae | -135.9202 | -134.9787 |
| Oosterella stevenini | -135.8575 | -135.3113 |
| Orbitolinopsis cuvillieri | -130.8282 | -130.7437 |
| Orbitolinopsis debelmasi | -130.8315 | -130.7193 |
| Orbitolinopsis flandrini | -130.7071 | -130.6993 |
| Orbitolinopsis subkiliani | -130.7071 | -130.5260 |
| Paleodictyoconus actinostoma | -130.8282 | -130.7437 |
| Paleodictyoconus beckerae | -131.5967 | -130.9483 |
| Paleodictyoconus cuvillieri | -131.6762 | -130.4745 |
| Paracoskinolina arcuata | -131.5768 | -130.7437 |
| Paracoskinolina hispanica | -130.8539 | -130.4745 |
| Paracoskinolina jourdanensis | -131.4079 | -130.4745 |
| Paracoskinolina maynci | -131.5570 | -130.4745 |
| Paracoskinolina praereicheli | -130.8315 | -130.7361 |
| Paracoskinolina querolensis | -131.4458 | -130.5357 |
| Paracoskinolina reicheli | -130.8446 | -130.7437 |
| Paradontoceras calistoides | -146.3225 | -144.1928 |
| Paraspiticeras groeberi | -130.9613 | -130.2259 |
| Paraspiticeras precrassispinum | -131.5216 | -131.5216 |
| Parastomiosphaera malmica | -149.3071 | -146.9757 |
| Parathurmannia sarasini | -131.1211 | -131.0581 |
| Paraulacosphinctes senoides | -146.5267 | -146.1400 |
| Paraulacosphinctes transitorius | -147.5538 | -146.1274 |
| Pareodinia ceratophora | -144.7305 | -135.1885 |
| Parhabdolithus achlyostaurion | -140.1490 | -137.2383 |
| Parhabdolithus asper | -145.1543 | -134.3880 |
| Parhabdolithus embergeri | -148.0337 | -134.3880 |
| Parhabdolithus infinitus | -136.9295 | -130.0000 |
| Parhabdolithus judithae | -138.8304 | -137.3639 |
| Parhabdolithus splendens | -142.2385 | -134.3880 |
| Parhabdolithus swinnertonii | -137.2383 | -132.9078 |
| Pasottia andina | -149.6700 | -148.8710 |
| Percivalia fenestrata | -140.8846 | -130.2356 |
| Percivalia nebulosa | -141.5873 | -138.9182 |
| Phoberocysta neocomica | -145.7073 | -130.3853 |
| Phoberocysta tabulata | -137.8081 | -137.8081 |
| Phylloceras tethys | -137.8994 | -134.9912 |
| Phyllopachyceras winckleri | -134.7170 | -134.6987 |
| Pickelhaube furtiva | -144.0957 | -133.5160 |
| Piriferella paucicalcarea | -131.5967 | -130.4745 |
| Platylenticeras cardioceroides | -137.5468 | -137.4281 |
| Platylenticeras occidentale | -138.1799 | -137.3094 |
| Plesiospitidiscus ligatus | -132.0230 | -130.9333 |
| Plesiospitidiscus subdifficilis | -131.2234 | -131.0135 |
| Podorhabdus dietzmanni | -138.1799 | -134.6804 |
| Polycostella beckmannii | -149.3960 | -144.3560 |
| Polycostella senaria | -148.8680 | -131.4000 |
| Polygonifera evittii | -144.7305 | -144.7305 |
| Polypodorhabus madingleyensis | -145.5646 | -132.4950 |
| Praecalpionellites murgeanui | -139.9667 | -138.4912 |
| Praedictyorbitolina busnardoi | -131.6762 | -131.6762 |
| Praedictyorbitolina carthusiana | -131.4775 | -130.5260 |
| Praedictyorbitolina claveli | -131.6762 | -130.7437 |
| Praetintinnopsella andrusovi | -147.2500 | -145.4263 |
| Praturlonella danilovae | -131.4079 | -130.4745 |
| Prediscosphaera columnata | -113.1000 | -113.1000 |
| Protacanthodiscus andreaei | -146.1400 | -145.8887 |
| Protacanthodiscus berriasensis | -145.7500 | -144.6359 |
| Protacanthodiscus heterocosmus | -145.7500 | -144.6359 |
| Protacrioceras ornatum | -131.4344 | -131.3908 |
| Protacrioceras puzosianum | -133.5537 | -133.5300 |
| Protancyloceras punicum | -140.0000 | -139.9209 |
| Protetragonites quadrisulcatus | -139.2250 | -135.3385 |
| Protoellipsodinium seghire | -134.0444 | -133.6636 |
| Protoellipsodinium touile | -134.5384 | -133.4430 |
| Pseudhimalayites subpretiosus | -148.3400 | -147.8187 |
| Pseudinvoluticeras douvillei | -150.3033 | -148.0435 |
| Pseudinvoluticeras primordialis | -150.5670 | -149.9339 |
| Pseudoacanthodiscus hexagonus | -145.8887 | -145.8887 |
| Pseudoceratium anaphrissum | -128.9001 | -127.2006 |
| Pseudoceratium pelliferum | -142.6067 | -127.2312 |
| Pseudocyclammina lituus | -138.8436 | -138.5807 |
| Pseudofavrella angulatiformis | -135.4160 | -135.3982 |
| Pseudofavrella australe | -135.1485 | -135.1485 |
| Pseudofavrella garatei | -135.4160 | -135.3892 |
| Pseudolissoceras zitteli | -149.9233 | -147.2950 |
| Pseudomoutoniceras annulare | -131.8486 | -131.8486 |
| Pseudosaynella termieri | -125.6695 | -125.6695 |
| Pseudosubplanites euxinus | -144.9717 | -143.9896 |
| Pseudosubplanites lorioli | -145.0197 | -143.3660 |
| Pseudosubplanites ponticus | -145.3079 | -144.2326 |
| Pseudothurmannia angulicostata | -131.2138 | -130.9150 |
| Pseudothurmannia catulloi | -131.1457 | -130.8447 |
| Pseudothurmannia ohmi | -131.1915 | -130.8141 |
| Pseudothurmannia picteti | -131.5833 | -131.0050 |
| Pseudothurmannia pseudomalbosi | -131.4967 | -131.1491 |
| Pterolytoceras exoticum | -144.3114 | -144.3114 |
| Pterospermella aureolata | -138.1600 | -133.1871 |
| Pterospermella australiensis | -137.7146 | -137.5051 |
| Ptychophylloceras diphyllum | -136.8009 | -135.3385 |
| Ptychophylloceras semisulcatum | -143.9243 | -131.8057 |
| Raimondiceras alexandrense | -141.3556 | -141.3556 |
| Reinhardtites elegans | -135.0461 | -133.4800 |
| Reinhardtites fenestratus | -141.5000 | -130.0000 |
| Remaniella borzai | -144.2732 | -138.8622 |
| Remaniella cadischiana | -145.0677 | -138.4756 |
| Remaniella catalanoi | -144.7714 | -139.6111 |
| Remaniella colomi | -144.5900 | -141.7265 |
| Remaniella dadayi | -139.4672 | -137.2662 |
| Remaniella duranddelgai | -144.9500 | -136.3200 |
| Remaniella ferasini | -145.2167 | -143.1667 |
| Remaniella filipescui | -143.3071 | -136.6650 |
| Remaniella murgeanui | -139.8458 | -137.2662 |
| Retecapsa angustiforata | -145.2134 | -130.0000 |
| Retecapsa levis | -130.3715 | -130.2500 |
| Retecapsa neocomiana | -143.0732 | -133.2059 |
| Retecapsa octofenestratus | -143.8907 | -130.2356 |
| Retecapsa surirella | -139.6485 | -130.1558 |
| Rhabdolekiskus parallelus | -132.8665 | -130.3715 |
| Rhabdolithus rectus | -139.7230 | -135.0540 |
| Rhagodiscus angustus | -133.9498 | -133.9498 |
| Rhagodiscus asper | -148.0337 | -130.0000 |
| Rhagodiscus eboracensis | -134.5589 | -134.5589 |
| Rhagodiscus nebulosus | -140.7146 | -136.0000 |
| Rhagodiscus reightonensis | -135.2495 | -133.4800 |
| Rhagodiscus splendens | -142.9579 | -130.0000 |
| Rhynchodiniopsis aptiana | -133.6636 | -132.5487 |
| Rhynchodiniopsis fimbriata | -137.7466 | *** |
| Rodighieroites rutimeyeri | -136.3800 | -136.1425 |
| Rotelapillus laffittei | -144.9670 | -130.0000 |
| Rotelapillus radians | -143.5726 | -143.5726 |
| Rucinolithus irregularis | -130.7224 | -130.1995 |
| Rucinolithus terebrodentarius | -132.6331 | -130.7224 |
| Rucinolithus wisei | -141.4818 | -134.8773 |
| Sabaudiella riverorum | -130.1957 | -130.1957 |
| Salpingoporella genevensis | -131.4458 | -130.4745 |
| Sarasinella biformis | -136.9518 | -136.5449 |
| Sarasinella eucyrta | -139.0444 | -137.4384 |
| Sarasinella hirticula | -136.1425 | -135.7165 |
| Sarasinella longi | -137.8546 | -137.7274 |
| Sarasinella trezanensis | -138.7673 | -137.4384 |
| Sarasinella uhligi | -137.5366 | -137.5366 |
| Saynella clypeiformis | -133.5867 | -133.4587 |
| Saynoceras verrucosum | -136.9525 | -135.2914 |
| Scriniodinium attadalense | -135.1873 | -130.0000 |
| Sirmiodinium grossii | -134.7146 | -134.7146 |
| Sollasites horticus | -141.1526 | -132.4596 |
| Sollasites lowei | -130.5160 | -130.5160 |
| Sornayites gp. simionescui | -131.5833 | -131.1877 |
| Speetonia colligata | -141.5541 | -131.3415 |
| Spiniferites dentatus | -132.6670 | -130.0000 |
| Spiniferites ramosus | -137.4847 | -130.0000 |
| Spiniferites ramosus multibrevis | -138.2895 | -133.4430 |
| Spiticeras acutum | -145.8110 | -143.2667 |
| Spiticeras damesi | -142.1348 | -139.3786 |
| Spiticeras fraternum | -143.8400 | -139.4232 |
| Spiticeras gr. multiforme | -141.6945 | -140.0091 |
| Spiticeras pricei | -142.6245 | -142.6245 |
| Spiticeras pseudogroteanum | -145.4730 | -144.9703 |
| Spiticeras spitiense | -143.1259 | -142.4513 |
| Spiticeras tripartitum | -144.1585 | -141.4983 |
| Spitidiscus fasciger | -133.7912 | -132.4590 |
| Spitidiscus gr. lorioli | -134.5757 | -134.0627 |
| Spitidiscus gr. pavlowi | -133.9575 | -133.5062 |
| Spitidiscus hugii | -130.8141 | -130.5385 |
| Spitidiscus kilapiae | -131.8159 | -131.8159 |
| Spitidiscus riccardii | -131.8227 | -131.7819 |
| Staurolithites crux | -139.3556 | -130.4350 |
| Staurolithites mutterlosei | -138.7112 | -132.4500 |
| Stenosemellopsis hispanica | -139.5829 | -138.7121 |
| Stephanolithion laffittei | -140.1019 | -132.4596 |
| Stomiosphaera acculeata | -145.7712 | -145.7712 |
| Stomiosphaera echinata | -146.1865 | -138.6887 |
| Stomiosphaera proxima | -146.6375 | -141.8957 |
| Stomiosphaera wanneri | -143.8625 | -138.8302 |
| Stradneria crenulata | -143.3366 | -130.0000 |
| Sturiella oblonga | -141.1871 | -141.1871 |
| Subaspinoceras mulsanti | -131.3908 | -131.3908 |
| Suboosterella heliaca | -133.6962 | -133.6962 |
| Subpulchellia nicklesi | -130.5538 | -130.1863 |
| Subsaynella mimica | -132.1538 | -132.1102 |
| Subsaynella sayni | -132.2424 | -131.8014 |
| Substeueroceras calistoide | -146.2968 | -144.0198 |
| Substeueroceras ellipsostomum | -147.3397 | -146.7469 |
| Substeueroceras koeneni | -145.8110 | -142.9481 |
| Substeueroceras striolatissimum | -147.2261 | -143.1007 |
| Substreblites callomoni | -141.1798 | -141.1798 |
| Substreblites zonarius | -136.8009 | -136.5815 |
| Subthurmannia boissieri | -141.3556 | -139.1912 |
| Subthurmannia clareti | -142.8583 | -142.8583 |
| Subthurmannia floquinensis | -144.1311 | -143.7469 |
| Subthurmannia occitanica | -143.3864 | -141.2972 |
| Subthurmannia patruliusi | -142.8102 | -142.0657 |
| Subthurmannia subalpina | -143.9390 | -143.6000 |
| Subtilisphaera perlucida | -132.8252 | -127.0475 |
| Subtilisphaera senegalensis | -130.5538 | -127.0475 |
| Subtilisphaera terrula | -133.0018 | -130.0000 |
| Systematophora areolata | -139.7813 | -137.6920 |
| Systematophora fasciculigera | -136.5093 | -136.5093 |
| Systematophora palmula | -142.1618 | -135.5812 |
| Systematophora silybum | -137.1198 | -133.4430 |
| Tanyosphaeridium boletus | -138.8068 | -130.2437 |
| Tanyosphaeridium magneticum | -140.0724 | -127.2312 |
| Tanyosphaeridium salpinx | -143.6494 | -130.0000 |
| Tanyosphaeridium variecalamus | -132.8252 | -130.0000 |
| Taveraidiscus intermedius | -130.8609 | -130.5522 |
| Taveridiscus oosteri | -130.8600 | -130.6916 |
| Tegumentum stradneri | -135.9912 | -130.2156 |
| Teschenites callidiscus | -134.9787 | -134.7782 |
| Teschenites castellanensiformis | -134.7377 | -134.5275 |
| Teschenites flucticulus | -134.8125 | -134.0898 |
| Teschenites neocomiensiformis | -135.9202 | -134.6758 |
| Teschenites pachydicranus | -135.4482 | -134.0898 |
| Teschenites subflucticulus | -135.0247 | -134.7406 |
| Teschenites subpachydicranus | -134.9224 | -134.7310 |
| Tetrapodorhabdus coptensis | -139.0080 | -130.2156 |
| Tetrapodorhabdus decorus | -139.0080 | -135.2414 |
| Thurmanniceras gratianopolitense | -139.4667 | -138.5064 |
| Thurmanniceras otopeta | -140.2899 | -138.3606 |
| Thurmanniceras perisphinctoides | -139.9483 | -139.8458 |
| Thurmanniceras pertransiens | -139.4356 | -137.9424 |
| Thurmanniceras salientum | -139.9739 | -139.9483 |
| Thurmanniceras thurmanni | -142.6296 | -137.9309 |
| Tintinnopsella carpathica | -147.0565 | -133.5600 |
| Tintinnopsella dacica | -138.5053 | *** |
| Tintinnopsella doliphormis | -145.1938 | -142.1260 |
| Tintinnopsella longa | -144.1758 | -133.5600 |
| Tintinnopsella remanei | -147.6200 | -144.4639 |
| Tintinnopsella subacuta | -140.4337 | -139.2093 |
| Tirnovella alpillensis | -141.5873 | -139.2418 |
| Tirnovella occitanica | -145.7340 | -145.6760 |
| Tirnovella pertransiens | -139.4000 | -137.7731 |
| Tirnovella romani | -139.7667 | -139.7640 |
| Top Mulichino Fm | -137.3348 | -135.3357 |
| Toulisphinctes rafaeli | -148.3400 | -147.1025 |
| Toxaster retusus | -131.6265 | -131.1899 |
| Toxaster seynensis | -131.2138 | -130.5724 |
| Tranolithus gabalus | -139.0080 | -130.0138 |
| Tranolithus salillium | -138.8495 | -134.1125 |
| Trichodinium castanea | -136.6366 | -133.7586 |
| Tubodiscus jurapelagicus | -141.3878 | -130.7939 |
| Tubodiscus verenae | -140.7182 | -133.3460 |
| Andean tuff beds | ||
| Tuff bed 139.24 | -142.0638 | *** |
| Tuff bed 139.55 | -141.6066 | *** |
| Tuff bed 139.96 | -143.5496 | *** |
| Tuff bed 140.34 | -144.6925 | *** |
| Tuff bed 142.04 | -147.2261 | *** |
| Turnovella kayseri | -144.5726 | -144.3114 |
| Umbria granulosa | -147.4496 | -139.1604 |
| Umbria granulosa minor | -147.8161 | *** |
| Urgonina alpillensis | -131.4775 | -130.4745 |
| Vagalapilla compacta | -140.0507 | -135.0643 |
| Vagalapilla stradneri | -144.5798 | -130.0000 |
| Valanginites bachelardi | -138.3092 | -135.2914 |
| Valanginites nucleus | -136.9518 | -135.4237 |
| Valserina broennimanni | -131.2138 | -130.5260 |
| Valserina primitiva | -131.4775 | -130.8539 |
| Valserina turbinata | -130.7437 | -130.5234 |
| Varlheideites peregrinus | -136.3800 | -135.0957 |
| Vekshinella angusta | -133.7586 | -130.9746 |
| Vekshinella stradneri | -138.8495 | -133.3458 |
| Viluceras permolestus | -135.4249 | -135.4249 |
| Virgatosphinctes andesensis | -151.0000 | -148.2974 |
| Virgatosphinctes mendozanum | -149.7544 | -148.2974 |
| Virgatosphinctes scythicus | -149.3303 | -148.9848 |
| Wallodinium cylindrica | -138.8068 | -130.0000 |
| Wallodinium krutzschii | -142.2500 | -130.0000 |
| Wallodinium lunua | -134.1410 | -133.1769 |
| Watznaueria barnesae | -148.6707 | -130.0000 |
| Watznaueria biporta | -148.6707 | -130.0161 |
| Watznaueria britannica | -148.6707 | -130.0000 |
| Watznaueria communis | -148.6707 | -130.0000 |
| Watznaueria fossacincta | -148.6707 | -130.1000 |
| Watznaueria manivitiae | -148.6707 | -131.1926 |
| Watznaueria oblonga | -134.7194 | -130.1032 |
| Watznaueria ovata | -148.3400 | -130.1558 |
| Watznaueria supraretacea | -133.6164 | -130.0390 |
| Weavericeras vacaense | -133.3620 | -132.9153 |
| Windhauseniceras internispinosum | -148.3400 | -146.3036 |
| Windhauseniceras windhauseni | -147.6203 | -147.6203 |
| Wollemanniceras keilhacki anterior | -113.1000 | -113.1000 |
| Wollemanniceras keilhacki keilhacki | -113.1000 | -113.1000 |
| Zeugrhabdotus diplogrammus | -138.4768 | -130.2117 |
| Zeugrhabdotus embergeri | -150.3728 | -130.0000 |
| Zeugrhabdotus erectus | -146.5460 | -130.1558 |
| Zeugrhabdotus fluxus | -146.5460 | -144.7867 |
| Zeugrhabdotus pseudoangustus | -139.0080 | -132.5414 |
| Zeugrhabdotus trivectis | -134.9864 | -133.5160 |
| Zygodiscus bicrescenticus | -134.9580 | -133.6510 |
| Zygodiscus diplogrammus | -137.7970 | -130.0000 |
| Zygodiscus elegans | -141.0456 | -130.0000 |
| Zygodiscus erectus | -146.5460 | -131.1695 |
Appendix 3: Sections and data (including taxa) that compose ANDESCS DB. Appendix 3. Andes.# are GraphCor catalog IDs. Preceding * means item not used.
| Datum | Taxa/morph | base (m) | top (m) |
| Andes.1 Lo Valdés, Chile [33°52'25.7"S 70°02'54.6"W]: Salazar Soto, 2012, Base Valanginian at 275 m @ FO T. thurmanni. Type section of Lo Valdés Formation 0-539 m; Tithonian-Berriasian boundary at base of sill 75 m. | |||
| Argentiniceras fasciculatum | /am | 280 | 320 |
| Aulacosphinctes proximus | /am | 125 | 200 |
| Berriasella jacobi | /am | 105 | 200 |
| Corongoceras alternans | /am | 10 | 50 |
| Corongoceras evolutum | /am | 15 | 45 |
| Corongoceras involutum | /am | 20 | 20 |
| Corongoceras koeni | /am | 20 | 50 |
| Corongoceras koellikeri | /am | 115 | 115 |
| Corongoceras lotenoense | /am | 15 | 45 |
| Corongoceras multimum | /am | 15 | 15 |
| Corongoceras mendozanum | /am | 20 | 20 |
| Crioceratites andinum | /am | 525 | 525 |
| Crioceratites diamantense | /am | 525 | 530 |
| Crioceratites perditum | /am | 526 | 526 |
| Cuyaniceras transgrediens | /am | 270 | 270 |
| Frenguelliceras magister | /am | 280 | 345 |
| Groebericeras rocardi | /am | 245 | 245 |
| Lytohoplites vareloe | /am | 35 | 65 |
| Lytohoplites vouloi | /am | 15 | 45 |
| Lytohoplites zambranoi | /am | 10 | 70 |
| Malbosiceras malbosi | /am | 180 | 200 |
| Micracanthoceras microcanthum | /am | 20 | 45 |
| Micracanthoceras spinulosum | /am | 45 | 160 |
| Spiticeras acutum | /am | 205 | 205 |
| Spiticeras spitiense | /am | 285 | 285 |
| Spiticeras tripartitum | /am | 175 | 315 |
| Substeueroceras calistoide | /am | 35 | 65 |
| Substeueroceras koeneni | /am | 100 | 245 |
| Thurmanniceras thurmanni | /am | 280 | 320 |
| Andes.2 Cajón del Morado, Chile [33°48'06.1"S 70°04'12.4"W]: Salazar Soto, 2012. Lo Valdés Formation 0-580 m; Tithonian-Berriasian boundary at ~160-165 m; Base Valanginian at 290 m FO T. thurmanni; base Hauterivian between 370-540 at FO C. diamantense. | |||
| Argentiniceras fasciculatum | /am | 290 | 345 |
| Aspidoceras rogoznicensis | /am | 75 | 75 |
| Aulacosphinctes proximus | /am | 173 | 305 |
| Berriasella jacobi | /am | 300 | 300 |
| Chigaroceras loteroense | /am | 115 | 173 |
| Corongoceras alternans | /am | 45 | 75 |
| Corongoceras involutum | /am | 26 | 26 |
| Corongoceras koellikeri | /am | 165 | 165 |
| Crioceratites andinum | /am | 540 | 540 |
| Crioceratites diamantense | /am | 540 | 540 |
| Crioceratites perditum | /am | 550 | 550 |
| Frenguelliceras magister | /am | 270 | 345 |
| Groebericeras rocardi | /am | 270 | 273 |
| Lytohoplites vareloe | /am | 26 | 100 |
| Lytohoplites vouloi | /am | 26 | 115 |
| Lytohoplites zambranoi | /am | 100 | 100 |
| Micracanthoceras microcanthum | /am | 105 | 105 |
| Micracanthoceras spinulosum | /am | 300 | 305 |
| Neocosmoceras sayni | /am | 271 | 273 |
| Pseudofavrella angulatiformis | /am | 290 | 300 |
| Spiticeras pricei | /am | 290 | 290 |
| Spiticeras spitiense | /am | 271 | 300 |
| Spiticeras tripartitum | /am | 240 | 355 |
| Substeueroceras calistoide | /am | 26 | 300 |
| Substeueroceras koeneni | /am | 225 | 255 |
| Substeueroceras striolatissimum | /am | 273 | 273 |
| Thurmanniceras thurmanni | /am | 290 | 370 |
| Andes.3 Cruz de Piedra, Chile [34°13'50.5"S 69°56'33.0"W]: Salazar Soto, 2012. Lo Valdés Formation 0-150 m; Base of section @ contact of Lo Valdes Fm.; Tithonian-Berriasian contact ~85-95m. | |||
| Aspidoceras rogoznicensis | /am | 120 | 120 |
| Aulacosphinctes proximus | /am | 90 | 120 |
| Berriasella jacobi | /am | 90 | 130 |
| Corongoceras koellikeri | /am | 140 | 140 |
| Corongoceras mendozanum | /am | 50 | 120 |
| Cuyaniceras transgrediens | /am | 140 | 150 |
| Micracanthoceras microcanthum | /am | 50 | 50 |
| Micracanthoceras spinulosum | /am | 50 | 90 |
| Micracanthoceras vetustum | /am | 100 | 120 |
| Pterolytoceras exoticum | /am | 110 | 110 |
| Spiticeras acutum | /am | 150 | 150 |
| Substeueroceras calistoide | /am | 50 | 110 |
| Substeueroceras koeneni *Occurrence at 70 is too low | /am | 90 | 130 |
| Substeueroceras striolatissimum | /am | 100 | 150 |
| Turnovella kayseri | /am | 100 | 110 |
| Andes.4 Rio Maitenes, Chile [35°00'25.6"S 70°23'18.2"W]: Salazar Soto, 2012. Lo Valdés Formation 0-535 m. Tithonian-Berriasian boundary at 400 m. | |||
| Aulacosphinctes proximus | /am | 165 | 200 |
| Catutosphinctes cf. americanensis | /am | 180 | 180 |
| Choicensisphinctes windhauseni | /am | 130 | 165 |
| Corongoceras alternans | /am | 360 | 360 |
| Corongoceras evolutum | /am | 360 | 360 |
| Euvirgalithacoceras malarguense | /am | 130 | 165 |
| Micracanthoceras microcanthum | /am | 360 | 360 |
| Micracanthoceras spinulosum | /am | 360 | 360 |
| Pseudolissoceras cf. zitteli | /am | 165 | 165 |
| Substeueroceras koeneni | /am | 430 | 430 |
| Virgatosphinctes scythicus *= V. mexicanus | /am | 125 | 145 |
| Windhauseniceras internispinosum | /am | 190 | 205 |
| Andes.5 Las Loicas, Argentina [35°46'59.9"S 70°09'W]: Vennari et al., 2014. CA-ID-TIMS age 139.6+/-0.09/0.18 Ma" at about 58 m. Vennari, 2016. Vaca Muerta Fm. overlies Tordillo Fm., Figs. 4, 20. Lena et al., 2019, Fig. 2; dated U-Pb zircons in 4 ash beds by CA-ID-TIMS 206Pb/238U: beds LL3-139.238+/-0.0.49Ma, LL9-139.956+/-0.063Ma, LL10-140.338+/-0.083Ma, LL13-142.039+/-0.058Ma. Lopez-Martinez et al., 2017a, Fig.1. | |||
| *Lena et al., 2019, Fig. 2 | |||
| Tuff bed 139.24 | /mb | -54 | *** |
| Tuff bed 139.96 | /mb | -41 | *** |
| Tuff bed 140.34 | /mb | -31 | *** |
| Tuff bed 142.04 | /mb | -2 | *** |
| *Vennari, 2016 | |||
| Argentiniceras cf. fasciculatum | /am | -38 | -38 |
| Argentiniceras noduliferum | /am | -53 | -66? |
| Berriasella subprivasensis | /am | -36 | -36 |
| Blanfordiceras sp. vetustum | /am | -35 | -35 |
| Cuyaniceras transgrediens | /am | -58? | -68 |
| Paradontoceras calistoides | /am | -23 | -25 |
| Spiticeras acutum | /am | -25 | -25 |
| Substeueroceras ellipsostomum | /am | -0? | -2 |
| Substeueroceras striolatissimum | /am | -2 | -2 |
| Euvirgalithacoceras malarguense | /am | -3 | 4.5 |
| Indansites malarguensis | /am | -3 | -4.5 |
| Pseudinvoluticeras primoridalis | /am | -3 | -4.5 |
| Pseudolissoceras zitteli | /am | -7 | -10 |
| *Lopez-Martinez et al., 2017a, Fig. 1 | |||
| Argentiniceras noduliferum | /am | -33 | -54 |
| *Berriasella sp. | /am | -25 | -25 |
| Berriasella subprivasensis | /am | -36 | -36 |
| Blanfordiceras sp. vetustum | /am | -35 | -35 |
| Cuyaniceras transgrediens | /am | -58 | -68 |
| Lytohoplites burckhardti | /am | -17 | -18 |
| *Neocosmoceras sp. | /am | -60 | -60 |
| Paradontoceras calistoides | /am | -23 | -25 |
| Spiticeras acutum | /am | -25 | -25 |
| Substeueroceras ellipsostomum | /am | -1 | -10 |
| Substeueroceras koeneni | /am | -25 | -25 |
| Substeueroceras striolatissimum | /am | -2 | -2 |
| Calpionella alpina | /ca | -14 | -41 |
| Crassicollaria brevis | /ca | -15.5 | -33 |
| Crassicollaria colomi | /ca | -15 | -15 |
| Crassicollaria parvula | /ca | -15 | -15 |
| Crassicollaria massutiniana | /ca | -15 | -36 |
| Tintinnopsella carpathica | /ca | -32 | -33 |
| Tintinnopsella remanei | /ca | -15 | -33 |
| *Vennari et al., 2014, Fig. 3: | |||
| Biscutum constans | /nn | -23 | -44 |
| Cyclagelosphaera deflandrei | /nn | -32.5 | 32.5 |
| Cyclagelosphaera margerelii | /nn | 0 | -48 |
| Diazomatolithus lehmanii | /nn | -15 | -15 |
| Eiffellithus primus | /nn | -17 | -36 |
| Manivitella pemmatoidea | /nn | 32.5 | -35.5 |
| Nannoconus cornutus | /nn | -24.5 | -35.5 |
| Nannoconus kamptneri minor | /nn | -33 | -37 |
| Nannoconus steinmannii minor | /nn | -35.5 | -35.5 |
| Nannoconus wintereri | /nn | -32.5 | -35.5 |
| Polycostella senaria | /nn | -20 | -37 |
| Rhagodiscus asper | /nn | -26 | -38 |
| Umbria granulosa | /nn | -15 | -36 |
| Watznaueria barnesae | /nn | 0 | -58 |
| Watznaueria biporta | /nn | 0 | -47.5 |
| Watznaueria britannica | /nn | -2 | -48 |
| Watznaueria fossacincta | /nn | 0 | -66 |
| Watznaueria manivitiae | /nn | -15 | -34 |
| Watznaueria ovata | /nn | 0 | -32 |
| Zeugrhabdotus embergeri | /nn | -17.5 | -48 |
| Zeugrhabdotus erectus | /nn | -15 | -34 |
| Watznaueria barnesae | /nn | 0 | -58 |
| Andes.6 Pampa Tril, Argentina [37°13'59.9"S 69°49'00.1"W]: Parent et al., 2015. Vaca Muerta Fm., Tithonian-lower Valanginian, Figs. 2, 5, 421.6 m thick; Overlies Tordillo Fm., underlies Quintuco Fm. | |||
| Argentiniceras noduliferum | /am | 155 | 155 |
| Aspidoceras depressus ID as cf. | /am | 183 | 183 |
| Aulacosphinctes proximus ID as Catutosphinctes ? | /am | 39 | 42 |
| Blanfordiceras vetustum | /am | 101 | 123 |
| Catutosphinctes guenenokenensis | /am | 2 | 6 |
| Catutosphinctes inflatus | /am | 82 | 102 |
| Catutosphinctes proximus | /am | 39 | 42 |
| Choicensisphinctes burckhardti | /am | 5 | 28 |
| Choicensisphinctes erinoides | /am | 17 | 28 |
| Choicensisphinctes platyconus | /am | 2 | 6 |
| Choicensisphinctes striolatus | /am | 105 | 135 |
| Cieneguiticeras falculatum | /am | 21 | 28 |
| Cieneguiticeras cf. perlaevis | /am | 17 | 21 |
| Corongoceras mendozanum | /am | 63? | 101 |
| Corongoceras steinmanni | /am | 101 | 101 |
| Cuyaniceras transgrediens | /am | 183 | 183 |
| Groebericeras bifrons | /am | 154 | 154 |
| Haploceras staszycii | /am | 21 | 28 |
| Himalayites cf. treubi | /am | 139 | 139 |
| Krantziceras azulense | /am | 105 | 105 |
| Krantziceras compressum | /am | 154 | 155 |
| Krantziceras disputabile | /am | 63 | 63 |
| Krantziceras planulatum | /am | 146 | 149 |
| Lissonia riveroi | /am | 231 | 285 |
| Lithoceras picunleufuense | /am | 2 | 6 |
| Lytoceras montanum | /am | 109 | 109 |
| Mazatepites arredondense | /am | 21 | 21 |
| Neocosmoceras malbosiforme | /am | 183 | 192 |
| Neocomites wichmanni | /am | 211 | 213 |
| Paradontoceras calistoides | /am | 87 | 109 |
| Pasottia andina | /am | 21 | 28 |
| Pseudhimalayites subpretiosus | /am | 42 | 42 |
| Pseudolissoceras zitteli | /am | 17 | 28 |
| Raimondiceras alexandrense | /am | 164 | 164 |
| Spiticeras fraternum | /am | 183 | 183 |
| Substeueroceras koeneni | /am | 137 | 139 |
| Subthurmannia boissieri | /am | 164 | 209 |
| Toulisphinctes cf. rafaeli | /am | 42 | 63 |
| Windhauseniceras internispinosum | /am | 42 | 43 |
| Andes.7 El Portón, Argentina [37°11'52.1"S 69°41'03.1"W]: Aguirre-Urreta et al., 2017, Fig. 3; Aguirre-Urreta et al., 2019, Figs. 3-4. | |||
| Top Mulichino | /mb | 0 | 0 |
| Agrio tuff bed 126.97 | /mb | -660 | *** |
| Agrio tuff bed 130.40 | /mb | -180 | *** |
| Assipetra terebrodentarius | /nn | -640 | -645 |
| Biscutum constans | /nn | -527 | -527 |
| Bukrylithus ambiguus | /nn | -580 | -580 |
| Clepsilithus maculosus | /nn | -470 | -650 |
| Cretarhabdus conicus | /nn | -43 | -580 |
| Cretarhabdus striatus | /nn | -75 | -625 |
| Cretarhabdus surirellus ID in Retacapsa | /nn | -135 | -660 |
| Crucibiscutum nequenensis | /nn | -460 | -517 |
| Cruciellipsis cuvillieri | /nn | -55 | -465 |
| Cyclagelosphaera deflandrei | /nn | -502 | -582 |
| Cyclagelosphaera margerelii | /nn | -5 | -650 |
| Diazomatolithus lehmanii | /nn | -127 | -470 |
| Eiffellithus striatus | /nn | -15 | -460 |
| Eiffellithus windi | /nn | -15 | -175 |
| Eprolithus floralis | /nn | -527 | -542 |
| Ethmorhabdus hauterivianus | /nn | -517 | -625 |
| Helenea chiastia | /nn | -165 | -625 |
| Lithraphidites bollii | /nn | -465 | -650 |
| Lithraphidites carniolensis | /nn | -185 | -655 |
| Manivitella pemmatoidea | /nn | -125 | -617 |
| Markalius inversus | /nn | -592 | -592 |
| Micrantholithus hoschulzii | /nn | -5 | -667 |
| Micrantholithus obtusus | /nn | -5 | -662 |
| Nannoconus bucheri | /nn | -185 | -655 |
| Nannoconus circularis | /nn | -95 | -650 |
| Nannoconus elongatus | /nn | -190 | -592 |
| Nannoconus globulus globulus | /nn | -43 | -662 |
| Nannoconus globulus minor | /nn | -43 | -274 |
| Nannoconus kamptneri | /nn | -43 | -662 |
| Nannoconus ligius | /nn | -517 | -622 |
| Nannoconus steinmannii | /nn | -147 | -662 |
| Nannoconus truitti | /nn | -260 | -622 |
| Percivalia fenestrata | /nn | -205 | -650 |
| Retecapsa angustiforata | /nn | -185 | -640 |
| Retecapsa octofenestratus | /nn | -475 | -650 |
| Retecapsa surirella | /nn | -135 | -660 |
| Rhagodiscus asper | /nn | -33 | -655 |
| Staurolithites crux | /nn | -140 | -625 |
| Tubodiscus jurapelagicus | /nn | -267 | -580 |
| Tubodiscus verenae | /nn | -260 | -260 |
| Watznaueria barnesae | /nn | -5 | -662 |
| Watznaueria biporta | /nn | -15 | -662 |
| Watznaueria fossacincta | /nn | -5 | -667 |
| Watznaueria manivitiae | /nn | -502 | -530 |
| Watznaueria ovata | /nn | -490 | -660 |
| Zeugrhabdotus embergeri | /nn | -5 | -635 |
| Zeugrhabdotus erectus | /nn | -157 | -660 |
| Zeugrhabdotus diplogrammus | /nn | -160 | -653 |
| Chacantuceras ornatum | /am | -35 | -50 |
| Crioceratites andinum | /am | -493 | -494 |
| Crioceratites diamantense | /am | -489 | -494 |
| Crioceratites perditum | /am | -498 | -501 |
| Crioceratites schlagintweiti | /am | -463 | -465 |
| Decliveites agrioensis | /am | -117 | -135 |
| Decliveites crassicostatum | /am | -88 | -88 |
| Holcoptychites agrioensis | /am | -170 | -178 |
| Holcoptychites magdalensae | /am | -140 | -161 |
| Hoplytocrioceris gentilli | /am | -205 | -225 |
| Hoplytocrioceris giovinei | /am | -195 | -198 |
| Olcostephanus laticosta | /am | -182 | -188 |
| Paraspiticeras groeberi | /am | -459 | -618 |
| Pseudofavrella australe | /am | -34 | -34 |
| Pseudofavrella garatei | /am | -2 | -5 |
| Sabaudiella riverorum | /am | -655 | -655 |
| Viluceras permolestus | /am | -1 | -1 |
| Weavericeras vacaense | /am | -258 | -314 |
| Andes.7b El Portón, Argentina [37°11'52.1"S 69°41'03.1"W]: Aguirre-Urreta et al., 2019. Pilmatue Mbr. 0-317 m; Avilé Mbr. Agua de la Mula Mbr. 447-700 m. | |||
| Marker tuff bed 126.97 | /mb | -660 | *** |
| Marker tuff bed 130.40 | /mb | -170 | *** |
| Assipetra terebrodentarius | /nn | -640 | -645 |
| Biscutum constans | /nn | -527 | -527 |
| Bukrylithus ambiguus | /nn | -580 | -580 |
| Clepsilithus maculosus | /nn | -470 | -650 |
| Cretarhabdus conicus | /nn | -43 | -580 |
| Cretarhabdus striatus | /nn | -75 | -625 |
| Cretarhabdus surirellus ID as Retacapsa | /nn | -135 | -660 |
| Crucibiscutum nequenensis | /nn | -460 | -517 |
| Cruciellipsis cuvillieri | /nn | -55 | -465 |
| Cyclagelosphaera deflandrei | /nn | -502 | -582 |
| Cyclagelosphaera margerelii | /nn | -5 | -650 |
| Diazomatolithus lehmanii | /nn | -127 | -470 |
| Eiffellithus striatus | /nn | -15 | -460 |
| Eiffellithus windi | /nn | -15 | -175 |
| Eprolithus floralis | /nn | -527 | -542 |
| Ethmorhabdus hauterivianus | /nn | -517 | -625 |
| Helenea chiastia | /nn | -165 | -625 |
| Lithraphidites bollii | /nn | -465 | -650 |
| Lithraphidites carniolensis | /nn | -185 | -655 |
| Manivitella pemmatoidea | /nn | -125 | -617 |
| Markalius inversus | /nn | -592 | -592 |
| Micrantholithus hoschulzii | /nn | -5 | -667 |
| Micrantholithus obtusus | /nn | -5 | -662 |
| Nannoconus bucheri | /nn | -185 | -655 |
| Nannoconus circularis | /nn | -95 | -650 |
| Nannoconus elongatus | /nn | -190 | -592 |
| Nannoconus globulus globulus | /nn | -43 | -662 |
| Nannoconus globulus minor | /nn | -43 | -274 |
| Nannoconus kamptneri | /nn | -43 | -662 |
| Nannoconus lignus | /nn | -517 | -622 |
| Nannoconus steinmannii | /nn | -147 | -662 |
| Nannoconus truitti | /nn | -260 | -622 |
| Percivalia fenestrata | /nn | -205 | -650 |
| Retecapsa angustiforata | /nn | -185 | -640 |
| Retecapsa octofenestratus | /nn | -475 | -650 |
| Retecapsa surirella | /nn | -135 | -660 |
| Rhagodiscus asper | /nn | -33 | -655 |
| Staurolithites crux | /nn | -140 | -625 |
| Tubodiscus jurapelagicus | /nn | -267 | -580 |
| Tubodiscus verenae | /nn | -260 | -260 |
| Watznaueria barnesae | /NN | -5 | -662 |
| Watznaueria biporta | /NN | -15 | -662 |
| Watznaueria fossacincta | /NN | -5 | -667 |
| Watznaueria manivitiae | /NN | -502 | -530 |
| Watznaueria ovata | /NN | -490 | -660 |
| Zeugrhabdotus embergeri | /NN | -5 | -635 |
| Zeugrhabdotus erectus | /NN | -157 | -660 |
| Zeugrhabdotus diplogrammus | /nn | -160 | -653 |
| Chacantuceras ornatum | /am | -35 | -50 |
| Crioceratites andinum | /am | -493 | -494 |
| Crioceratites diamantense | /am | -489 | -494 |
| Crioceratites perditum | /am | -498 | -501 |
| Crioceratites schlagintweiti | /am | -463 | -465 |
| Decliveites agrioensis | /am | -117 | -135 |
| Decliveites crassicostatum | /am | -88 | -88 |
| Holcoptychites agrioensis | /am | -170 | -178 |
| Holcoptychites magdalensae | /am | -140 | -161 |
| Hoplytocrioceris gentilli | /am | -205 | -225 |
| Hoplytocrioceris giovinei | /am | -195 | -198 |
| Olcostephanus laticosta | /am | -182 | -188 |
| Paraspiticeras groeberi | /am | -459 | -618 |
| Pseudofavrella australe | /am | -34 | -34 |
| Pseudofavrella garatei | /am | -2 | -5 |
| Sabaudiella riverorum | /am | -655 | -655 |
| Viluceras permolestus | /am | -1 | -1 |
| Weavericeras vacaense | /am | -258 | -314 |
| Andes.8 Real de las Coloradas, Argentina [34°01'59.9"S 69°43'59.9"W]: Vennari, 2016. Vaca Muerta Fm. overlies Tordillo Fm. Revises Parent's taxonomy(2011). | |||
| Choicensisphinctes choicensis = C. platyconus Parent | /am | 8 | 10 |
| Choicensisphinctes platyconus | /am | 1 | 10 |
| Cieneguiticeras perlaevis | /am | 8 | 18 |
| Pseudinvoluticeras douvillei = C. lotenoensis Parent | /am | 9 | 12 |
| Pseudinvoluticeras primoridalis = C. platyconus Parent | /am | 1 | 8 |
| Pseudolissoceras zitteli | /am | 10 | 18 |
| Virgatosphinctes andesensis | /am | 10 | 12 |
| Virgatosphinctes mendozanum | /am | 10 | 12 |
| Andes.9 Cerro Domuyo, Argentina [36°40'59.9"S 70°25'59.9"W]: Vennari, 2016. Vaca Muerta Fm. overlies Tordillo Fm. Revises Parent's taxonomy (2011). | |||
| Choicensisphinctes choicensis = C. platyconus Parent | /am | 3 | 5 |
| Choicensisphinctes platyconus | /am | 3 | 5 |
| Choicensisphinctes erinoides | /am | 26 | 40 |
| Pseudinvoluticeras douvillei | /am | 3 | 3 |
| Pseudolissoceras zitteli | /am | 26 | 40 |
| Virgatosphinctes andesensis | /am | 0 | 0 |
| Andes.10 Mina San Eduardo Composite Section, Argentina [~37°32'25.1"S 70°22'00.1"W]:Aguirre-Urreta et al., 2015, Fig. 2; 0-505 m Agrio Fm. Agua de la Mula Mbr. overlies Avilé Mbr., underlies Huitrín Fm. Inoceramid from Lazo, 2006. | |||
| Agrio tuff bed 127.42 | /mb | -470 | *** |
| Agrio tuff bed 129.09 | /mb | -10 | *** |
| Crioceratites diamantense | /am | -90 | *** |
| Crioceratites schlagintweiti | /am | -40 | *** |
| Paraspiticeras groeberi | /am | -410 | -470 |
| Sabaudiella riverorum | /am | -480 | *** |
| Spitidiscus kilapiae | /am | -2 | -2 |
| Spitidiscus riccardii | /am | 0 | -12 |
| Clepsilithus maculosus | /nn | -15 | -410 |
| Cruciellipsis cuvillieri | /nn | -27 | -80 |
| Lithraphidites bollii | /nn | -65 | -440 |
| Nannoconus ligius | /nn | -365 | -470 |
| Neocomiceramus curacoensis | /bi | -45 | -190 |
| Andes.11 Arroyo Truquico, Neuquén, Argentina [37°26'06.0"S 70°37'53.4"W]: Aguirre-Urreta., 1998, Fig. 2. Lower member Agrio Fm. overlies Mulichino Fm. at 0 m; | |||
| Top Mulichino | /mb | 18 | 18 |
| Karakaschiceras attenuatus | /am | 40 | 70 |
| Karakaschiceras neumayri | /am | 35 | 35 |
| Neohoploceras arnoldi | /am | 35 | 40 |
| Olcostephanus atherstoni | /am | 25 | 35 |
| Pseudofavrella angulatiformis | /am | 145 | 145 |
| Pseudofavrella garatei | /am | 145 | 145 |
| Andes.12 Cerro La Parva, Neuquén, Argentina [37°15'46.1"S 70°30'47.9"W]: Aguirre-Urreta, 1998, Fig. 2. Lower member Agrio Fm. overlyies Mulichino Fm., Valanginian. | |||
| Top Mulichino | /mb | -10 | -10 |
| Karakaschiceras attenuatus | /am | 30 | 73 |
| Karakaschiceras neumayri | /am | 20 | 22 |
| Neohoploceras arnoldi | /am | 20 | 22 |
| Olcostephanus atherstoni | /am | 5 | 22 |
| Pseudofavrella angulatiformis | /am | 115 | 115 |
| Andes.13 Arroyo Loncoche, Argentina [35°31'40.4"S 69°39'07.9"W]: Kietzmann et al., 2018, Figs. 3, 6; Iglesia Llanos et al., 2017. | |||
| Andes J-K SB 4 | /MB | 270 | *** |
| Andes J-K SB 3 | /MB | 215 | *** |
| Andes J-K SB 2 | /MB | 149 | *** |
| Andes J-K SB 1 | /MB | 75 | *** |
| Andiceras acuticostum ID cf. | /am | 65 | 70 |
| Argentiniceras noduliferum | /am | 220 | 225 |
| Aulacosphinctes proximus | /am | 60 | 70 |
| Blanfordiceras vetustum ID cf. | /am | 150 | 153 |
| Catutosphinctes americanensis | /am | 105 | 105 |
| Choicensisphinctes cf. erinoides | /am | 18 | 35 |
| Corongoceras lotenoense | /am | 140 | 145 |
| Corongoceras cf. mendozanum | /am | 135 | 135 |
| Cuyaniceras raripartitum | /am | 225 | 230 |
| Cuyaniceras transgrediens | /am | 240 | 255 |
| Laeviaptychus crassissimus | /am | 25 | 50 |
| Laeviaptychus latus | /am | 70 | 117 |
| Micracanthoceras lamberti | /am | 90 | 90 |
| Pseudinvoluticeras sp. primoridalis | /am | 10 | 12 |
| Pseudolissoceras zitteli | /am | 25 | 60 |
| Spiticeras damesi | /am | 220 | 275 |
| Substeueroceras koeneni | /am | 183 | 185 |
| Virgatosphinctes andesensis | /am | 0 | 5 |
| Windhauseniceras internispinosum | /am | 75 | 90 |
| Data from Kietzmann et al., 2011, Fig. 3 | |||
| Eiffellithus primus | /nn | 67 | *** |
| Polycostella beckmannii | /nn | 62 | *** |
| Polycostella senaria | /nn | 121 | *** |
| Umbria granulosa | /nn | 82 | *** |
| Data from Iglesia Llanos et al., 2017 | |||
| Magnetochron M15r | /MA | *** | |
| Magnetochron M16n | /MA | *** | 260 |
| Magnetochron M16r | /MA | *** | 240 |
| Magnetochron M17n | /MA | *** | 215 |
| Magnetochron M17r | /MA | *** | 200 |
| Magnetochron M18n | /MA | *** | 190 |
| Magnetochron M18r | /MA | *** | 180 |
| Magnetochron M19n | /MA | *** | 160 |
| *Magnetochron M19n.1r | /MA | *** | |
| *Magnetochron M19n.2n | /MA | *** | |
| Magnetochron M19r | /MA | *** | 147 |
| Magnetochron M20n | /MA | *** | 135 |
| Magnetochron M20n.1r | /MA | *** | 125 |
| Magnetochron M20n.2n | /MA | *** | 118 |
| *Magnetochron M20r | /MA | *** | |
| Magnetochron M21n | /MA | *** | 75 |
| Magnetochron M21r | /MA | *** | 58 |
| Andes.14 Cuesta del Chihuido, Argentina [35°45'39.6"S 69°42'35.3"W]: *Kietzmann et al., 2018, Fig. 3; Iglesia Llanos et al., 2017. Vaca Muerta Fm., Tithonian-Berriasian. Reported section thickness 185 m. *Kietzmann et al., 2021a [35°44'49.6"S 69°34'37.2"W] | |||
| Andes J-K SB 4 | /MB | 195 | * |
| Andes J-K SB 3 | /MB | 164 | * |
| Andes J-K SB 2 | /MB | 113 | * |
| Andes J-K SB 1 | /MB | 58 | * |
| Argentiniceras bituberculatum cf.? | /am | 165 | 168 |
| Aulacosphinctes proximus | /am | 72 | 75 |
| Blanfordiceras vetustum | /am | 114 | 117 |
| Choicensisphinctes erinoides cf.? | /am | 32 | 72 |
| Corongoceras mendozanum cf.? | /am | 80 | 110 |
| Laeviaptychus crassissimus | /am | 10 | 58 |
| Laeviaptychus latus | /am | 65 | 80 |
| Neocomites wichmanni cf.? | /am | 200 | 200 |
| Pseudinvoluticeras primordialis sp.? | /am | 17 | 32 |
| Pseudolissoceras zitteli | /am | 30 | 45 |
| Spiticeras damesi cf.? | /am | 170 | 175 |
| Substeueroceras koeneni sp.? | /am | 120 | 152 |
| *Virgatosphinctes andesensis sp.? * defines base of V. andesensis Zone | /am | 0 | 32 |
| Virgatosphinctes mendozanum | /am | 0 | 32 |
| Windhauseniceras internispinosum | /am | 60 | 78 |
| Windhauseniceras windhauseni | /am | 60 | 75 |
| *Kietzman et al., 2021a | |||
| Borzaiella slovenica | /dn | 50 | 61 |
| Chitinoidella boneti | /dn | 59 | 87 |
| Chitinoidella hegarati | /dn | 59 | 59 |
| Calpionella alpina | /ca | 110 | 155 |
| Calpionella elliptalpina | /ca | 118 | 121 |
| Calpionella elliptica | /ca | 155 | 155 |
| Calpionella grandalpina | /ca | 89 | 128 |
| Calpionellites darderi | /ca | 200 | 200 |
| Calpionellopsis oblonga | /ca | 190 | 200 |
| Calpionellopsis simplex | /ca | 160 | 200 |
| Crassicollaria brevis | /ca | 110 | 119 |
| Crassicollaria massutiniana | /ca | 89 | 100 |
| Crassicollaria parvula | /ca | 114 | 128 |
| Lorenziella hungarica | /ca | 160 | 200 |
| Tintinnopsella carpathica | /ca | 72 | 175 |
| Tintinnopsella doliphormis | /ca | 195 | 200 |
| Tintinnopsella longa | /ca | 155 | 185 |
| Tintinnopsella remanei | /ca | 72 | 89 |
| Andes.15 Bardas Blancas, Argentina [35°52'40.8"S 69°54'32.0"W]: Kietzmann et al., 2018, Fig. 3. Vaca Muerta Fm. | |||
| Andes J-K SB 4 | /MB | 235 | *** |
| Andes J-K SB 3 | /MB | 180 | *** |
| Andes J-K SB 2 | /MB | 110 | *** |
| Andes J-K SB 1 | /MB | 50 | *** |
| Andiceras cf. acuticostum | /am | 80 | 80 |
| Berriasella sp. | /am | 108 | 119 |
| Chigaroceras loteroense | /am | 72 | 72 |
| Choicensisphinctes choicensis | /am | 5 | 20 |
| Cuyaniceras raripartitum | /am | 205 | 205 |
| Laeviaptychus crassissimus | /am | 26 | 30 |
| Lissonia riveroi | /am | 270 | 270 |
| Neocomites cf. wichmanni | /am | 240 | 242 |
| Spiticeras acutum | /am | 160 | 160 |
| Substeueroceras koeneni | /am | 113 | 158 |
| Substeueroceras striolatissimum | /am | 100 | 100 |
| Virgatosphinctes andesensis | /am | 18 | 18 |
| Virgatosphinctes mendozanum | /am | 3 | 3 |
| Windhauseniceras internispinosum | /am | 50 | 70 |
| Andes.16 Arroyo Rahue, Argentina [35°59'56.8"S 69°56'35.9"W]: Kietzmann et al., 2018, Fig. 3. Vaca Muerta Fm. | |||
| Andes J-K SB 4 | /MB | 205 | *** |
| Andes J-K SB 3 | /MB | 158 | *** |
| Andes J-K SB 2 | /MB | 91 | *** |
| Andes J-K SB 1 | /MB | 25 | *** |
| Argentiniceras noduliferum | /am | 160 | 160 |
| Blanfordiceras vetustum | /am | 100 | 100 |
| Corongoceras lotenoense | /am | 45 | 50 |
| ?Neocomites crassicostatum | /am | 180 | 183 |
| Laeviaptychus latus | /am | 20 | 30 |
| Micracanthoceras lamberti | /am | 60 | 60 |
| Pseudinvoluticeras douvillei | /am | 10 | 10 |
| Pseudolissoceras zitteli | /am | 7 | 15 |
| Spiticeras damesi | /am | 190 | 195 |
| Substeueroceras cf. striolatissimum | /am | 92 | 105 |
| Virgatosphinctes andesensis | /am | 4 | 4 |
| Virgatosphinctes mendozanum | /am | 4 | 4 |
| Windhauseniceras internispinosum | /am | 25 | 30 |
| Windhauseniceras windhauseni | /am | 20 | 20 |
| Andes.17 Los Catutos, Argentina [38°49'12.0"S 70°10'12.0"W]: López-Martínez et al., 2017b, Fig. 6. Vaca Muerta Fm., Los Catutos Mbr. overlies Tordillo Fm. | |||
| Aspidoceras aff. euomphalum | /am | 81 | 81 |
| Aspidoceras quinchaoi | /am | 77 | 95 |
| Aulacosphinctes proximus | /am | 32 | 52 |
| Choicensisphinctes erinoides | /am | 3 | 16 |
| Choicensisphinctes choicensis | /am | 3 | 6 |
| Corongoceras cf. praecursor | /am | 35 | 35 |
| Djurjuriceras catutosense | /am | 91 | 95 |
| Pseudinvoluticeras douvillei | /am | 0 | 6 |
| Pseudinvoluticeras primoridalis | /am | 0 | 6 |
| Pseudolissoceras zitteli | /am | 18 | 22 |
| Toulisphinctes rafaeli | /am | 57 | 60 |
| *ID as Catutosphinctes | /am | ||
| Windhauseniceras internispinosum | /am | 61 | 95 |
| Magnetochron M20n.2n | /ma | * | 87 |
| Magnetochron M20r | /ma | * | 61 |
| Magnetochron M21n | /ma | * | 51 |
| Magnetochron M21r | /ma | * | 37 |
| Magnetochron M22n | /ma | * | 30 |
| Magnetochron M22r | /ma | * | 5 |
| Andes.18a Bajada Viejo, Neuquén Basin, Argentina [38°22'S 70°W]: Lazo et al., 2009, Fig. 2a. The Agrio Formation age @ 235m 136.4 Ma by biostratigraphic correlation. Valanginian-Hauterivian Stage boundary at 235 m at FAD Holcoptychites neuquensis. | |||
| Top Mulichino | /mb | 0 | 0 |
| Clepsilithus maculosus | /nn | 70 | *** |
| Eiffellithus striatus | /nn | 8 | *** |
| Nannoconus bucheri | /nn | 68 | *** |
| Nannoconus circularis | /nn | 68 | *** |
| Chacantuceras ornatum | /am | 32 | *** |
| Holcoptychites agrioensis | /am | 315 | *** |
| Holcoptychites neuquensis | /am | 235 | *** |
| Hoplytocrioceris gentilli | /am | 412 | *** |
| Hoplytocrioceris giovinei | /am | 408 | *** |
| Neocomites crassicostatum | /am | 79 | *** |
| Neocomites wichmanni | /am | 79 | *** |
| Olcostephanus laticosta | /am | 388 | *** |
| Pseudofavrella angulatiformis | /am | 3 | *** |
| Weavericeras vacaense | /am | 446 | *** |
| Andes.18b Bajada del Agrio, Neuquén Basin, Argentina [38°22'S 70°W]: Lazo et al., 2009, Fig. 2b. The Agrio Formation ages @ 5m 133.8 Ma & @ 470m = 130.0 Ma by biostratigraphic correlation. Hauterivian-Barremian Stage boundary at 470 m in Paraspiticeras groeberi Zone. | |||
| Top Avilé Member | /mb | 0 | 0 |
| Clepsilithus maculosus | /nn | *** | 432 |
| Eiffellithus striatus | /nn | 8 | *** |
| Nannoconus bucheri | /nn | *** | 390 |
| Neocomiceramus curacoensis | /bi | 138 | *** |
| Crioceratites andinum | /am | 78 | *** |
| Crioceratites diamantense | /am | 78 | *** |
| Paraspiticeras groeberi | /am | 417 | *** |
| Spitidiscus riccardii | /am | 5 | *** |
| Andes.19 Arroyo Cieneguita, Neuquén Basin, Argentina [35°33'S 69°24'W]: Parent et al., 2011. Vaca Muerta Fm., Tithonian-lower Valanginian, Fig. 2, 143 m thick. Overlies Upper Jurassic Tordillo Fm. sandstone. | |||
| Aspidoceras cf. euomphalum | /am | 28 | 100 |
| Blanfordiceras vetustum | /am | 95 | 110 |
| Catutosphinctes guenenokenensis | am | 0 | 7 |
| Catutosphinctes inflatus | /am | 89 | 10 |
| Catutosphinctes proximus | /am | 28 | 71 |
| Choicensisphinctes platyconus | /am | 0 | 3 |
| Choicensisphinctes striolatus | /am | 110 | 111 |
| Cieneguiticeras falculatum | /am | 28 | 49 |
| Cieneguiticeras cf. perlaevis | /am | 0 | 21 |
| Corongoceras mendozanum | /am | 89 | 100 |
| Cuyaniceras transgrediens | /am | 123 | 130 |
| Groebericeras bifrons | /am | 117 | 123 |
| Lithoceras picunleufuense | /am | 0 | 3 |
| Mazatepites arredondense | /am | 21 | 36 |
| Paradontoceras calistoides | /am | 95 | 111 |
| Pasottia andina | /am | 11 | 18 |
| Pseudhimalayites subpretiosus | /am | 28 | 36 |
| Pseudolissoceras zitteli | /am | 11 | 21 |
| Spiticeras fraternum | /am | 117 | 130 |
| Substeueroceras koeneni | /am | 110 | 111 |
| Toulisphinctes rafaeli ID as cf. | /am | 28 | 49 |
| Windhauseniceras internispinosum | /am | 49 | 71 |
| Andes.22 Composite Chos Malal Section, Argentina
[37°28'08.4"S
69°58'46.6"W]: Composed of Andes 6 Pampa Tril, Argentina, Parent et al., 2015. Vaca Muerta Fm., Tithonian-lower Valanginian, Figs. 2, 5, 421.6 m thick; Overlies Upper Jurassic Tordillo Fm. sandstone and underlies lower Valanginian Quintuco Fm. claystone. Puerta Curaco section, Schwarz et al., 2006, Fig. 11, Mulichinco Fm. 340m, base on Vaca Muerta, top at Agrio; add 630 m to all below: Andes.7 El Porton Section, Argentina [37°11'52.1"S 69°41'03.1"W]: Aguirre-Urreta et al., 2017, Fig. 3; Aguirre-Urreta et al., 2019, Figs. 3-4. Radiometric date @ 660m 126.97+/-0.04 Ma = 1290 Radiometric date @ 180m 130.39+/-0.16 Ma = 810 Mulichinco Fm. overlain by Agrio Fm., Pilmatue Mbr. base at 0m; Avile Mbr. continental deposits 317-437 m; Hauterivian Stage duration 5.21+/-0.08 myr from 131.29 to 126.08+/-0.19 Ma. Andes.22b, Puerta Curaco, Argentina [37°22'26.0"S 69°56'17.2"W]: Kietzman et al., 2021a (37°22'26.2"S 69°56'17.2"W); ss V. andesensis to S. damesi zones. *Kietzmann et al., 2018. |
|||
| *Zones on chart Fig. 6. | |||
| *Neocomites wichmanni | /am | 265 | 360 |
| *Spiticeras damesi | /am | 160 | 265 |
| *Argenticeras noduliferum | /am | 140 | 160 |
| *Substeueroceras koeneni | /am | 85 | 140 |
| *Corongoceras alternans | /am | 65 | 85 |
| *Windhauseniceras internispinosum | /am | 40 | 65 |
| *Aulacosphinctes proximus | /am | 30 | 40 |
| *Pseudolissoceras zitteli | /am | 10 | 30 |
| *Virgatosphinctes andesensis | /am | 0 | 10 |
| *Kietzmann et al., 2021a, Fig. 6. | |||
| Borzaiella slovenica | /dn | 20 | 92 |
| Chitinoidella boneti | /ca | 40 | 45 |
| Calpionella alpina | /ca | 92 | 265 |
| Calpionella elliptalpina | /ca | 92 | 92 |
| Calpionella elliptica | /ca | 195 | 265 |
| Calpionella grandalpina | /ca | 80 | 120 |
| Calpionellites darderi | /ca | 310 | 310 |
| Calpionellopsis oblonga | /ca | 225 | 360 |
| Calpionellopsis simplex | /ca | 165 | 300 |
| Crassicollaria brevis | /ca | 165 | 165 |
| Crassicollaria massutiniana | /ca | 60 | 15 |
| Crassicollaria parvula | /ca | 70 | 70 |
| Lorenziella hungarica | /ca | 195 | 290 |
| Tintinnopsella carpathica | /ca | 55 | 360 |
| Tintinnopsella remanei | /ca | 55 | 80 |
| Andes.23 Las Tapaderas, Argentina [estimated: 35°24'S 70°18'W]: *Kietzmann et al., 2021b, Fig. 3. base conformable above Tordillo Fm.; top overlain by Pleistocene volcanics | |||
| Aulacosphinctes proximus | /am | 0 | 2 |
| Corongoceras alternans | /am | 35 | 38 |
| Corongoceras lotenoense ID ? | /am | 8 | 8 |
| Corongoceras praecursor | /am | 20 | 20 |
| Substeueroceras koeneni | /am | 61 | 61 |
| Chitinoidella boneti | /ca | 8 | 42 |
| Calpionella alpina | /ca | 35 | 64 |
| Calpionella elliptica | /ca | 64 | 64 |
| Calpionella grandalpina | /ca | 25 | 25 |
| Crassicollaria brevis | /ca | 35 | 40 |
| *Crassicollaria colomi ID ? too high | /ca | 59 | 61 |
| Crassicollaria massutiniana | /ca | ||
| Crassicollaria parvula | /ca | 35 | 61 |
| Lorenziella hungarica | /ca | ||
| Tintinnopsella carpathica | /ca | 16 | 64 |
| Tintinnopsella doliphormis | /ca | ||
| Tintinnopsella longa | /ca | ||
| Tintinnopsella remanei | /ca | 16 | 16 |
| Cadosina fusca | /dn | 61 | 64 |
| Colomisphaera fortis | /dn | 25 | 64 |
| Colomisphaera tenuis | /dn | 2 | 64 |
| Stomiosphaera echinata | /dn | 36 | 61 |
| Stomiosphaera proxima | /dn | 40 | 64 |
| Stomiosphaera wanneri | /dn | 61 | 64 |