◄ Carnets Geol. 22 (12) ►
![]()
Outline:
[1. Introduction]
[2. Material and methods]
[3. Morpho-anatomical diagnostic characteristics of coralline algae]
[4. Geological settings]
[5. Results: Systematic palaeontology]
[6. Discussion]
[7. Conclusions]
[Bibliographic references]
[Supplement] and ...
[Appendix: Identification key for encrusting coralline algal genera]
Earth Science Institute of the Slovak Academy of Sciences, D�bravsk� cesta 9, Bratislava (Slovakia)
Earth Science Institute of the Slovak Academy of Sciences, D�bravsk� cesta 9, Bratislava (Slovakia)
Published online in final form (pdf) on August 1, 2022
DOI
10.2110/carnets.2022.2212
![]()
[Editor: Daniela Basso; language editor:
Stephen Carey; technical editor: Bruno R.C. Granier]
![]()
Encrusting coralline algae are important rock-building organisms of the lower Priabonian limestones from the Central Carpathian Paleogene Basin. Despite the effort of early palaeontologists, former classifications lacked many diagnostic characteristics and modern taxonomic concepts, which hinders their use for modern palaeontological interpretations. The situation is further complicated as recent DNA analyses of extant coralline algae highlight the limits of the morpho-anatomical classification and identify many coralline algal genera which can also be recognized in the fossil record. Because palaeontology deals exclusively with the morpho-anatomical characteristics, a synthesis of the new discoveries in molecular phylogenetic analyses with morphology-based palaeontological classifications is necessary. Our palaeontological study based on published investigations on coralline molecular genetics enabled: 1) the identification of seventeen coralline algal morphological species grouped in six genera: Sporolithon lugeonii, Sporolithon nummuliticum, Sporolithon sp. 1, Sporolithon sp. 2, Lithothamnion camarasae, Lithothamnion cf. corallioides, Lithothamnion prascoi, Lithothamnion cf. ramosissimum, Lithothamnion roveretoi, Lithothamnion sp., Phymatolithon sp., Mesophyllum fructiferum, Mesophyllum cf. engelhartii, Mesophyllum sp., Chamberlainium lemoinei, Lithoporella melobesioides, and Lithoporella cf. minus ; 2) the description of Chamberlainium lemoinei (Miranda) comb. nov. known from the Bartonian, and 3) the identification of a unique encrusting coralline alga tentatively assigned to the genus Lithothamnion.
� coralline algae;
� taxonomy;
� Priabonian;
� Central Carpathian Paleogene Basin
Hrabovsk� J. & Starek D. (2022).- Priabonian non-geniculate coralline algae from the Central Carpathian Paleogene Basin.- Carnets Geol., Madrid, vol. 22, no. 12, p. 567-617.
Les algues corallines encro�tantes priaboniennes du bassin pal�og�ne des Carpates centrales.- Les algues corallines encro�tantes constituent un groupe d'organismes important pour la formation des calcaires du Priabonien inf�rieur du bassin pal�og�ne des Carpates centrales. En d�pit des efforts des pre�miers pal�ontologues, aucune classification ancienne ne tenait compte ni de l'�ventail des carac�t�ris�tiques diagnostiques, ni des concepts taxinomiques modernes, ce qui compromettait leur utilisation � des fins d'interpr�tations pal�ontologiques modernes. La situation a �t� rendue encore plus complexe lors�que les analyses g�n�tiques des algues corallines actuelles ont mis en �vidence les limites de la clas�sification morpho-anatomique et l'existence d'un plus grand nombre de genres, lesquels peuvent aussi �tre identifi�s dans le registre fossile. Parce que la pal�ontologie utilise exclusivement des caract�res morpho-ana�to�miques, il est n�cessaire de faire une synth�se des approches phylog�n�tiques mol�culaires et de la classification pal�ontologique. Nos �tudes pal�ontologiques bas�es sur les r�sultats publi�s de la g�n�tique mol�culaire des algues corallines ont permis : 1) l'identification de dix-sept esp�ces mor�pho�lo�gi�ques d'algues corallines regroup�es en six genres : Sporolithon lugeoni, Sporolithon nummuliticum, Spo�rolithon sp. 1, Sporolithon sp. 2, Lithothamnion camarasae, Lithothamnion cf. corallioides, Litho�tham�nion prascoi, Lithothamnion cf. ramosissimum, Lithothamnion roveretoi, Lithothamnion sp., Phy�ma�tolithon sp., Mesophyllum fructiferum, Mesophyllum cf. engelhartii, Mesophyllum sp., Cham�ber�lai�nium lemoinei, Lithoporella melobesioides et Lithoporella cf. minus ; 2) la description de Chamberlainium lemoinei (Miranda) comb. nov. connue depuis le Bartonien et 3) l'identification d'une algue co�ral�line encro�tante unique provisoirement attribu�e au genre Lithothamnion.
� algues corallines ;
� taxinomie ;
� Priabonien ;
� bassin pal�og�ne des Carpates centrales
Geniculate and non-geniculate coralline algae (Corallinophycidae, Rhodophyta) are a major constituent of Paleogene bioclastic limestone of the Central Carpathian Paleogene Basin (Schalekov�, 1962, 1964; Filo et al., 2009; Buček et al., 2013; Hrabovsk� et al., 2022). Despite the effort of past researchers (Lemoine, 1934; Andrusov, 1937; Schalekov�, 1962, 1964) and the subsequent revision of the major coralline algal subfamilies, genera and species (e.g., Woelkerling, 1988; Woelkerling & Harvey, 1992; Braga et al., 1993; Braga & Aguirre, 1995; Basso et al., 1997; Athanasiadis & Ballantine, 2014; Athanasiadis, 2017) the systematic position of many species is still uncertain.
Recent progress in molecular taxonomy reveals significant cryptic diversity and polyphyly of coralline algal taxa (Bailey et al., 2004; Kato et al., 2011, 2013; Richards et al., 2016; Melbourne et al., 2017; Pezzolesi et al., 2019; Kato & Baba, 2019; Caragnano et al., 2020; Puckree-Padua et al., 2021). This has led to changes in the morpho-anatomical diagnostic characters used by botanists and palaeontologists to identify certain subfamilies or genera, and further questioning of their diagnostic value (Hind et al., 2016; Caragnano et al., 2018; Nelson et al., 2021). However, palaeontology mostly deals with morphologically defined species (e.g., De Queiroz, 2007) because genetic material is not normally preserved in specimens older than sub-fossil remains (e.g., Teichert et al., 2019). Palaeontologists have, in fact, two options to use the outcomes of molecular taxonomy. First, given the lack of DNA sequences in fossils, morphologically similar genera could be kept taxonomically separate, as in the cases of extant Adeylithon and extinct Aethesolithon (Pe�a et al., 2019). Second, DNA-based studies may reveal morpho-anatomical characteristics that would be useful in distinguishing fossil material, e.g., within the five orders of the Corallinophycidae (Jeong et al., 2021); this approach permitted the emended diagnosis of coralline algal subfamilies and genera (Athanasiadis & Ballantine, 2014; R�sler et al., 2016; Caragnano et al., 2018). The two approaches outlined have a significant effect on the identification of fossil taxa (Hrabovsk� et al., 2019; Coletti et al., 2020). However, only a very few specimens from the genera Mesophyllum, Chamberlainium and Spongites have as yet been examined in these ways (Hrabovsk� et al., 2019; Coletti et al., 2020). Nevertheless, the results so far are of great use in palaeoecology because descendants of these genera are known from Recent temperate climatic zones (Athanasiadis & Ballantine, 2014; Caragnano et al., 2018).
Therefore, description, classification and assessment of new specimens based on application of the accepted vegetative and reproductive characteristics, as well as re-description of species occurrences in the light of the new approaches is required prior to palaeoecological reconstructions of the coralline-algal-dominated limestones. For this purpose, we have analysed 1) new samples collected from the Tich� Dolina Valley - Ježov vrch and Čaplovka sections, and 2) samples from available historical collections. We also provide a key for the identification of encrusting coralline algae which summarizes published morphological characteristics in known taxa of encrusting coralline algae. This key has facilitated our classification of the Cenozoic coralline algae from the Central Carpathian Paleogene Basin.
Coralline algae were studied in 46 thin sections, 19 from Ježov vrch and 27 from Čaplovka (including seven large thin sections, 5x5 cm, the remainder being of standard size, 2x3.5 cm). Specimens from historical collections were also studied and we focused on the work of Schalekov� (1962) who identified 43 species among geniculate and non-geniculate coralline algae. Her historical collection is conserved at the Faculty of Sciences, Comenius University in Bratislava. The Paleogene collection includes four boxes, I-IV, with 199 thin sections. Because of the large amount of material, we restricted our study to selected specimens, namely those that were classified under the same name as species found in the new samples. Re-description of other species is beyond the scope of the present research. For the observations on coralline algae, we have used an AXIOZEISS scope A1 light microscope equipped with an AXIOCAM 105 Color camera and a Leica MZ6 stereomicroscope equipped with a Leica EC3 camera.
Identification of coralline algae at the rank of genus is based on morpho-anatomical characteristics discussed in numerous papers dealing with the taxonomy of both extinct and extant coralline algae (e.g., Johansen, 1981; Woelkerling, 1988; Woelkerling & Harvey, 1992; Braga et al., 1993; Braga & Aguirre, 1995; Basso et al., 1997; Rasser & Piller, 1999; Athanasiadis, 2001; Athanasiadis & Ballantine, 2014; Hrabovsk� et al., 2016; Athanasiadis, 2017). Characteristics used by palaeontologists are vegetative and reproductive (e.g., Braga et al., 1993; Basso et al., 1996; Hrabovsk� et al., 2019; Coletti et al., 2020) or they are simply related to growth forms (Woelkerling et al., 1993; Athanasiadis & Ballantine, 2014).
Vegetative characteristics which can be used by palaeontologists are associated with the internal organization of the thallus (e.g., Johansen, 1981; Caragnano et al., 2018), like the arrangement of ventral core filaments (e.g., Lemoine, 1928; Aguirre & Braga, 1998), stratification of peripheral filaments (e.g., Basso et al., 1998; Basso et al., 1997), branching patterns of the filaments (Athanasiadis, 2017), morphology of the cells (e.g., Braga & Aguirre, 1995; Basso et al., 1998), secondary and primary cell connections (e.g., Braga et al., 1993), morphology of epithallial cells (e.g., Braga et al., 1993; Rasser & Piller, 1999), length of meristematic cells (e.g., Aguirre et al., 1996) and trichocyte arrangement and types (e.g., Basso et al., 2014; Rōsler et al., 2015; Caragnano et al., 2018).
Reproductive anatomy includes characteristics associated with both the sporophytes and gametophytes (Basso et al., 1996; Coletti et al., 2020; Hrabovsk� et al., 2019). Reproductive characteristics of sporophytes include types of development of conceptacles, the manner of formation of the conceptacle roof, its chamber and its exit pore(s) (Johansen, 1981), roof morphology, e.g., presence or absence of peripheral rim and sunken pore plate or degree of conceptacle projection, and anatomy, e.g., length of the roof filaments, number of cells in roof filaments and their shape, pore canal lining filaments and shape and number of pore canal lining cells, morphology of pore canals (e.g., Basso et al., 1997; Coletti et al., 2016, 2018, 2020; Hrabovsk� et al., 2019), dimensions of conceptacles (Fravega et al., 1993; Coletti et al., 2020), and rarely features of bi/tetraspores (e.g., Teichert et al., 2019; Aguirre & Braga, 1998). Reproductive characteristics of gametophytes include morphology of carposporangial conceptacles - i.e., presence of a central columella in Mesophyllum and its absence in Melyvonnea or Leptophytum (Athanasiadis & Ballantine, 2014; Athanasiadis & Adey, 2006), monoecy/dioecy of gametophytes (Basso et al., 1996; Athanasiadis & Ballantine, 2014; Hrabovsk� et al., 2016, Coletti et al., 2020) and rarely carposporophyte characteristics (Woelkerling et al., 2012). Diagnostic features distinctive of Sporolithales are based on the occurrence within calcified compartments of tetrasporangia that undergo cruciate cleavage (Le Gall et al., 2010). In contrast, in Hapalidiales and Corallinapetrales, zonate sporangia are borne in conceptacles with multiporate roof and each sporangium possesses an apical plug, while in Corallinales the zonate sporangia are borne in uniporate conceptacles (Harvey et al., 2003; Nelson et al., 2015; Athanasiadis, 2016; Jeong et al., 2021). In the Order Sporolithales sporangia are borne either in sporangial complexes consisting of stalk cell and involucre developed within the calcified compartment (Heydrichia and Roseapetra) or within calcified sporangial compartments lacking involucre (Sporolithon). While sporangial complexes may produce few generation of spores resulting in multiple stalk cell structure, sporangial compartments, e.g., those of the genus Sporolithon produce only one spore so that chambers may contain relics of the single stalk cell (e.g., Vannucci et al., 2000; Chelaru & Bucur, 2016; Nelson et al., 2021). However, Heydrichia and Roseapetra, both bearing sporangial complexes, are morphologically indistinguishable from each other (Nelson et al., 2021).
Gametophytes are commonly used by palaeontologists in the species description, e.g., Mastrorilli (1968); Basso et al. (1996); Quaranta et al. (2007). However, gametophytic characteristics of high diagnostic value at the rank of the genus were identified only recently for sub-fossil and fossil coralline algae (Woelkerling et al., 2012; Hrabovsk� et al., 2019; Coletti et al., 2020).
Identification of gametophytes requires consideration of anatomical characteristics of both proposed gametophytes, and sporophytes (Basso et al., 1996). This, however, may be problematic since 1) reproductive structures such as spermatangial filaments, carpogonial filaments, carposporangia, gonimoblast filaments, carpospores are rarely preserved in fossil plants, and 2) matching the proposed gametophytes with sporophytes can be done using features which are of low diagnostic value (e.g., growth form, dimensions of cells in vegetative filaments, arrangement of filaments in ventral core, rarely shape of epithallial cells and meristematic cells or trichocytes (Braga & Aguirre, 1995; Hrabovsk�, 2019; Hrabovsk� et al., 2019; Coletti et al., 2020). On the other hand, identification of fossil gametophytes is facilitated where two types of conceptacles occur in a single plant. This suggests one of 1) fusion of the sporophyte with the gametophyte that is common in Recent species (Verheij, 1993), 2) monoecious gametophytic life cycle phases where both female and male conceptacles are borne on the same plant, or 3) a dioecious carpogonial-carposporangial plant with preserved developmental stages of the conceptacles from small non-fertilized carpogonial to large carposporangial conceptacles as documented, e.g., in extant Mesophyllum (Athanasiadis et al., 2004) and fossils (Hrabovsk� et al., 2022).
The proposed identification key for the Cenozoic coralline algae from the Carpathian Paleogene Basin is based on morpho-anatomical characteristics (Supplement - Appendix: Identification key).
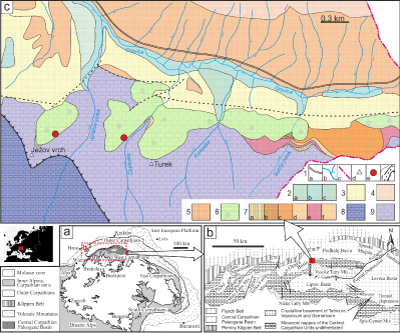
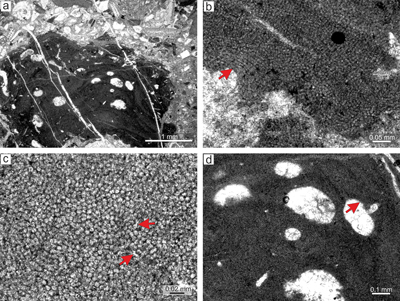
The Central Carpathian Paleogene Basin (CCPB) lies inside the Western Carpathian Mountain chain
(Fig. 1a ![]() ). It belongs to the basinal system of the Peri- and
Paratethys seas. The CCPB opening and evolution is probably related to crustal
thinning as a result of either subcrustal erosion (K�zm�r et al.,
2003) or extensional collapse of the overthickened central Western Carpathian crust and the pull of the external Western Carpathian oceanic lithosphere
during retreating subduction (Kov�č et al.,
2016).
). It belongs to the basinal system of the Peri- and
Paratethys seas. The CCPB opening and evolution is probably related to crustal
thinning as a result of either subcrustal erosion (K�zm�r et al.,
2003) or extensional collapse of the overthickened central Western Carpathian crust and the pull of the external Western Carpathian oceanic lithosphere
during retreating subduction (Kov�č et al.,
2016).
The basin covers a large part of the central Western Carpathian area (Fig. 1a-b ![]() ) and is filled mainly with marine deposits which overlap older nappe units.
The marine deposits range from Middle Eocene (Samuel & Fus�n,
1992; Gross et al., 1980) to
Late Oligocene (Olszewska & Wieczorek, 1998; Gedl, 2000; Sot�k et al., 2001; Garecka, 2005). The CCPB sediments are preserved in many structural sub-basins
(Fig. 1b
) and is filled mainly with marine deposits which overlap older nappe units.
The marine deposits range from Middle Eocene (Samuel & Fus�n,
1992; Gross et al., 1980) to
Late Oligocene (Olszewska & Wieczorek, 1998; Gedl, 2000; Sot�k et al., 2001; Garecka, 2005). The CCPB sediments are preserved in many structural sub-basins
(Fig. 1b ![]() ), located in the �ilina, Rajec, Turiec, Orava, Podhale, Liptov, Poprad and Horn�d regions as well as in the Spi�sk� Magura, Levočsk� vrchy and �ari�sk� vrchovina
Mountains.
), located in the �ilina, Rajec, Turiec, Orava, Podhale, Liptov, Poprad and Horn�d regions as well as in the Spi�sk� Magura, Levočsk� vrchy and �ari�sk� vrchovina
Mountains.
The CCPB deposits ("Podtatransk� skupina Group" according to Gross et al., 1984; Gross, 2008) are divided into the following formations (Fm):
At the base is the Borov� Fm, including the Horn�d Member (Mb), the Chrastianske Mb and the Tom�ovce Mb, according to Filo & Sir�ňov�, 1996, 1998). It consists of breccia, conglomerates, lithic sandstones to siltstones, marlstones, and organodetrital and organogenic limestones. It represents terrestrial, fluvial-deltaic and shallow marine transgressive deposits (Marshalko, 1970; Kulka, 1985; Gross et al., 1993; Bar�th & Kov�č, 1995; Filo & Sir�ňov�, 1996, 1998; �urka et al., 2012). The ages of the marine deposits range from the Late Lutetian to the late Priabonian. The latter is documented only in the Tom�ovce Mb. However, the age of the predominantly continental Horn�d Mb was recently established as Paleocene to Middle Eocene (Marshalko, 1970; Filo & Sir�ňov�, 1996).
The Borov� Fm is overlain by the Huty Fm, which includes mainly mud-rich shelf, slope to deep-marine deposits (Janočko & Jacko, 1999; Sot�k et al., 2001; Starek et al., 2004) intercalated with sandstone megabeds (Starek et al., 2013). The Zuberec Fm and Biely Potok Fm (including the Kežmarok Mb) compose the CCPB up-section, and predominantly consist of rhythmically bedded turbidites and massive sandstones, representing various sand-rich submarine-fan facies associations (Westwalewicz-Mogilska, 1986; Wieczorek, 1989; Sot�k, 1998; Starek et al., 2000; Sliva, 2005; Starek & Fuksi, 2017a, 2017b; Starek et al., 2019). The age of the Huty Fm ranges from the late Priabonian to Early Oligocene and the age of the Zuberec and Biely Potok formations Oligocene (Olszewska & Wieczorek, 1998; Gedl, 2000; Starek et al., 2000; Sot�k et al., 2001; Garecka, 2005; Filipek et al., 2017).
The study area is situated near the border of the Orava and Podhale depressions (Fig. 1b ![]() ),
south-east of the village of Oravice. The Paleogene deposits of the Podtatransk� skupina Group are bounded on the south by Mesozoic Central Carpathian units (Fig. 1c
),
south-east of the village of Oravice. The Paleogene deposits of the Podtatransk� skupina Group are bounded on the south by Mesozoic Central Carpathian units (Fig. 1c ![]() ). The studied deposits are exposed along cliffs of the Čaplovka valley and Ježov vrch hill (1086 m) (Fig. 1c
). The studied deposits are exposed along cliffs of the Čaplovka valley and Ježov vrch hill (1086 m) (Fig. 1c ![]() ). They are a part of the Borov�
Fm, which is locally represented by carbonate breccia, polymictic conglomerates and organogenic-biodetrital limestones (Fig. 1c
). They are a part of the Borov�
Fm, which is locally represented by carbonate breccia, polymictic conglomerates and organogenic-biodetrital limestones (Fig. 1c ![]() ). The bedded to massive limestones are originally micritic (often
recrystallized) and are formed predominantly by algae, large benthic foraminifers,
and bryozoans, with associated rare small benthic foraminifers, crinoids, molluscs and echinoderms as well as granule-to pebble-sized limestone clasts. Based on associated large benthic
foraminifers their age is regarded as early Priabonian (SBZ 19) (Filo et al., 2009).
). The bedded to massive limestones are originally micritic (often
recrystallized) and are formed predominantly by algae, large benthic foraminifers,
and bryozoans, with associated rare small benthic foraminifers, crinoids, molluscs and echinoderms as well as granule-to pebble-sized limestone clasts. Based on associated large benthic
foraminifers their age is regarded as early Priabonian (SBZ 19) (Filo et al., 2009).
|
|
Figure 1: a) Location of study area within the Alpine-Carpathian orogeny. b) The Central Carpathian Paleogene Basin system depicting structural sub-basins, basement massifs and surrounding units. c) Simplified geological sketch of part of the Orava Basin (Geological Map of Slovakia M 1:50,000 [online] �G�D�, 2013. |
Phylum Rhodophyta Wettstein, 1901
Subphylum Eurhodophytina Saunders & Hommersand, 2004
Class Florideophyceae Cronquist, 1960
Subclass Corallinophycidae Le Gall & Saunders, 2007
Order Sporolithales Le Gall & Saunders in Le Gall et al., 2010
Family Sporolithaceae Verheij, 1993
Genus Sporolithon Heydrich, 1897
Type species: Sporolithon ptychoides Heydrich, 1897, El Tor, Sinai Peninsula, Egypt, Recent.
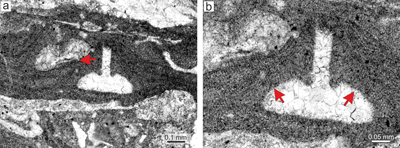
Sporolithon lugeonii (Pfender) Gosh & Maithy, 1996
(Fig. 2 ![]() )
)
Selected specimens examined. Ježov vrch: specimens from thin sections 28097, 28091, 28092, 28079, 28081, 28085, 28080.
Material. Specimen from thin section 28097. Proposed carpogonial gametophyte is described based on the specimen from thin section 28092.
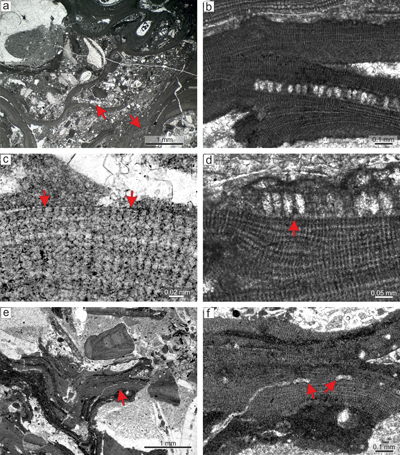
Sporolithon lugeonii is the dominant species in this thin section. This plant has an encrusting-layered growth habit with various modes of
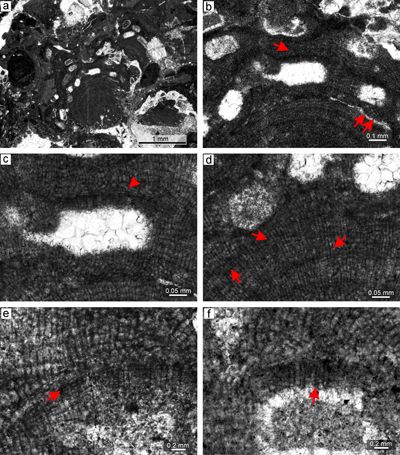
thallial division by applanate branching. Applanate protuberances loosely adhered to the substrate resulting in numerous primary voids (Fig. 2a ![]() ). Superimposed thalli form crusts which are up to 2mm thick. It also develops small warty protuberances.
). Superimposed thalli form crusts which are up to 2mm thick. It also develops small warty protuberances.
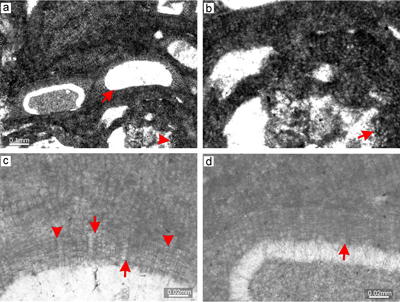
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 2b), consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upward to the peripheral region where it grows roughly perpendicular to the substrate. Branching pattern of the filaments is monopodial from rectangular cells or pseudodichotomous from bladed cells. Ventral core is non-coaxial, 29-110 μm thick, and consists of six to 15 filaments. Cells measure 17-30 μm (mean 23 μm � 3.6) in length and 6-12 μm in diameter (mean 9 μm � 1.5) (n = 23). Peripheral region is of variable thickness and consists of cells which are 7-21 μm (mean 14 μm � 2.8) long and 7-12 μm (mean 10 μm � 1.3) in diameter (n = 61). Cells of adjacent filaments are laterally connected by fusions (Fig. 2c). We have observed a single layer of flattened and flared cells terminating the filaments (Fig. 2c). Cells are 4-9 μm long (mean 6 μm � 1.3) and 10-13 μm in diameter (mean 11 μm � 1.2) in diameter (n = 12). Cells below them are as long, slightly elongate, or shorter than the cells immediately subtending them. We consider these cells tentatively as epithallial and the subtending layer as meristematic cells.
Calcified sporangial compartments are arranged in sori built by tens of compartments buried in the thallus (Fig. 2a). No solitary sporangial compartment was found. Calcified sporangial compartments are borne above the layer of more-or-less
elongate cells (Fig. 2d). These cells are 17-24 μm long (mean, 20 μm � 2.1) while the cells immediately subtending them are 14-20 μm long (mean 16 μm � 1.6)
(n = 12 for each). Compartments are 24-44 μm in diameter (mean 32 μm � 5.8) and 56-81 μm high (mean 67 μm � 6.1)
(n = 23). Compartments are separated by paraphyses consisting of 3 elongate cells (Fig. 2d). Some empty chambers bear remains of single stalk cell. No sporangial complexes were observed. Pore canals are
conical with diameter 7-12 μm at the base and are 17-25 μm long. The proposed gametophyte encrusts other coralline algae (Fig. 2e.2 ![]() ) and contains uniporate chambers with central pedestal (Fig. 2f). Chambers are 23-28 μm high and 74-98 μm in diameter. Pore canal is 21 μm in diameter at its base and is tapered towards its opening. The roof development involves only filaments peripheral to the fertile area. Since central pedestal (or columella) is absent in spermatangial conceptacles and could be present in carpogonial one, we tentatively consider this plant as female gametophyte of Sporolithon lugeonii.
) and contains uniporate chambers with central pedestal (Fig. 2f). Chambers are 23-28 μm high and 74-98 μm in diameter. Pore canal is 21 μm in diameter at its base and is tapered towards its opening. The roof development involves only filaments peripheral to the fertile area. Since central pedestal (or columella) is absent in spermatangial conceptacles and could be present in carpogonial one, we tentatively consider this plant as female gametophyte of Sporolithon lugeonii.
Remarks. Identified sporolithoid algae from the Paleocene limestones of the Western Carpathians are Archaeolithothamnium nummuliticum (G�mbel) Rothpletz, 1891, Archaeolithothamnium cf. oulianovi Pfender, 1926, Archaeolithothamnium lugeoni Pfender, 1926, Archaeolithothamnium gunteri Johnson & Farris, 1948, and Archaeolithothamnium sp. (Schalekov�, 1962). All of them are fertile and bear calcified sporangial compartments arranged in sori and lacking sporangial complexes. Following Townsend et al. (1995), Ghosh and Maithy (1996), and Moussavian and Kuss (1990), all these occurrences should be reassigned to the genus Sporolithon. The diagnosis of the studied specimen matches the characteristics of S. lugeonii which was recently neotypified by Aguirre et al. (2011). Unfortunately, the specimen described and figured in Schalekov� (1962) is lost. Nevertheless, morphological characteristics and provided figure suggests this specimen may represents S. lugeonii.
|
|
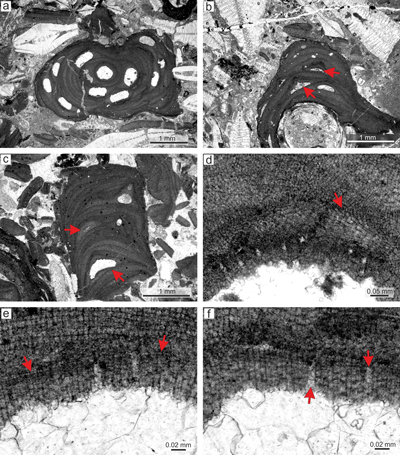
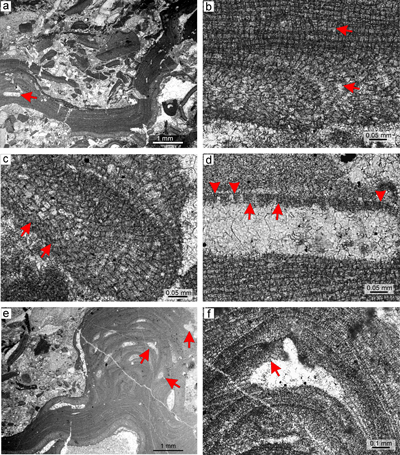
Figure 2: Sporolithon lugeonii (Pfender) Gosh & Maithy, 1996. a) Encrusting-layered
growth form with numerous applanate branches which are loosely adhered to the substrate, giving rise to numerous primary voids (arrows). Note tens of the compartments arranged in sori on the right bottom corner. b) Monomerous and non-coaxial thallus with embedded sporangial compartments arranged in sori. c) Proposed epithallial cells (arrows) developed above the meristematic cells which are as long, shorter or longer than cells immediately subtending them. Note the flared morphology of epithallial cells. d)
Elongate cells below the calcified sporangial compartments (arrow). Compartments are separated by 3-celled paraphyses. e) Supposed gametophyte (arrow) with encrusting
growth form. Gametophyte overgrows another coralline alga. f) Enlarged thallus with uniporate conceptacles bearing central columella (arrows). |
Sporolithon nummuliticum (G�mbel) Ghosh & Maithy, 1996
(Fig. 3 ![]() )
)
Selected specimens examined. Čaplovka: fertile specimens from the thin sections 31003, 31005, 31016, 31017, 31024. Ježov vrch: thin sections 28082, 28090.
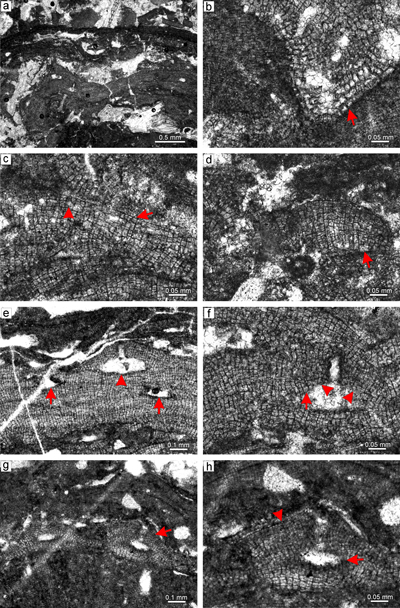
Material. Sporolithon nummuliticum is a common species in many thin sections from both studied localities. It typically occurs as a crust above or within the multispecific rhodoliths. It also develops small monospecific warty rhodoliths (Fig. 3a). Species is described based on the specimen from thin section 31003. Proposed gametophyte is described based on the specimen from thin section 31016.
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 3b), consisting of single system of branched filaments which run parallel to the substrate in ventral core and bend upward forming the peripheral region of the thallus. Branching pattern of the filaments is monopodial and pseudodichotomous. Ventral core is non-coaxial. In section it consists of six to 13 filaments and is 47-87 μm thick. Cells are rectangular and measure 14-26 μm (mean, 19 μm � 3.3) in length and 5-11 μm (mean 8 μm � 1.8) in diameter (n = 13). The peripheral region is 271-465 μm thick and consists of cells which are 10-26 μm (mean 17 μm � 4.2) long and 7-14 μm (mean 11 μm � 1.6) in diameter (n = 28). Cells of the adjacent filaments are laterally connected by fusions in both ventral core (Fig. 3b) and peripheral region (Fig. 3c). Epithallial cells are flattened, rectangular to flared (Fig. 3c). One to two layers of epithallial cells are present above the meristematic cells, which are of variable size.
Calcified sporangial compartments are arranged in the sori which are built by tens of compartments (Fig. 3a). No solitary sporangial compartment was found. Numerous calcified sporangial compartments are borne above the layer of elongate cells (Fig. 3d). These cells are 15-37 μm long (mean 26 μm � 6.1) (n = 11) while cells immediately subtending them are 15-26 μm long (mean, 20 μm � 5) (n = 6). Compartments are 39-67 μm in diameter (mean 48 μm � 7.8) and 81-124 μm high (mean 99 μm � 13) (n = 17). Some empty chambers bear remains of single stalk cell. Compartments are separated by three to five celled paraphyses. Pore canals are cylindrical and 9 μm in diameter. Sori are buried in the thallus. No sporangial complexes were observed. Proposed gametophyte bears two types of conceptacles, the first small with flat floor (Fig. 3e), the second larger with flat floor (Fig. 3f). Roof and pore canal anatomy suggest development from filaments located peripherally to fertile area. We tentatively consider these conceptacles as gametophytes. Small conceptacles are 16-27 μm high and 60-88 μm in diameter. Large conceptacles are 38-55 μm high and 128-208 μm in diameter.
Remarks. This species has a cosmopolitan distribution in Eocene strata (Ghosh & Maithy, 1996; Hrabovsk� et al., 2022; Aguirre et al., 2020). It is known from rhodolith beds (Aguirre et al., 2020) as well as from seagrass meadows (Hrabovsk� et al., 2022). We have observed S. nummuliticum in sequences of large multispecific rhodoliths from both studied sites as well as isolated fragments of protuberant and encrusting growth forms. The specimen from Oravice which was described and figured by Schalekov� (1962) matches our specimens, although it does not show the elongate cells below the sporangial compartments that we document here.
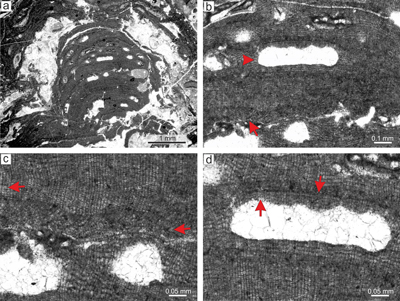
|
|
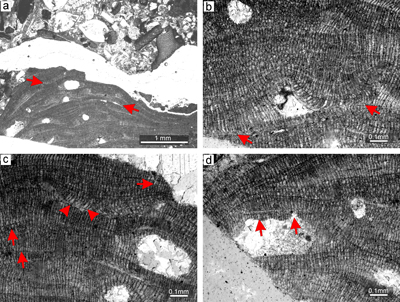
Figure 3: Sporolithon nummuliticum (G�mbel) Gosh & Maithy, 1996. a) Encrusting warty-protuberant
growth form. Arrow points to the development of encrusting growth form above the damaged protuberance. Note the tens of the calcified sporangial compartments building each sorus. b) Monomerous non-coaxial thallus with common lateral fusions of adjacent cells in ventral core filaments (arrow). c) Lateral fusion of cells of the adjacent filaments in the peripheral region (right arrow). Arrow on the left points to the epithallial cells preserved below the ventral core filaments. Note that the epithallial cells are developed above the layer of meristematic cells which are of variable lengths. d) Calcified sporangial compartments are developed above the
elongate cells (arrow). e) Proposed gametophyte showing small conceptacles. Note that the conceptacle floor is flat (arrow). f) Proposed gametophyte with larger conceptacles. Arrow points to the central part of the conceptacle flat floor. Note central part of the chamber with remains of the matter of uncertain origin. |
Sporolithon sp. 1
(Fig. 4 ![]() )
)
Selected specimens examined. Čaplovka: one fertile specimen in thin section 31018 and numerous sterile specimens from thin sections 31002, 31003, 31016.
Material. Sporolithon sp. 1 fertile specimens are rare in the studied material and completely absent in the Ježov vrch material. Sporolithon sp. 1 was found on the surface of multispecific rhodoliths (Fig. 4a). These crusts are generally thin. Thalli show scar-like structures which are determined by rows of cells with irregular shape and size. These cells appear to represent renewed meristematic activity which produced new perithallial filaments. Growth form is encrusting-layered with numerous applanate branches developed from damaged thallial surfaces (Fig. 4a-b). Species is described based on the specimen from thin section 31018.
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 4b), consisting of single system of branched filaments running parallel to the substrate in the ventral core and bending upward to form the peripheral region where it grows perpendicular to the substrate. Branching pattern of the filaments is monopodial and pseudodichotomous. Ventral core is non-coaxial. In section it consists of five to eight filaments and is 42-74 μm thick. Cells are rectangular or bladed and measure 13-18 μm (mean 16 μm � 1.9) in length and 5-8 μm (mean 7 μm � 0.7) in diameter (n = 11). Peripheral region is 129-151 μm thick and consists of rectangular to square cells which are 7-19 μm (mean 12 μm � 2.6) long and 7-11 μm (mean 9 μm � 0.9) in diameter (n = 49). Cells of the adjacent filaments are laterally joined with fusions in both ventral core and peripheral region (Fig. 4b). We have observed a single layer of flattened and rectangular cells terminating perithallial filaments which are commonly preserved below the overlying secondary ventral core (Fig. 4b). Cells are 6-7 μm (mean 6 μm � 0.3) long and 9-12 μm (mean 10 μm � 1.4) in diameter (n = 3). Cells below them are as long, slightly elongate, or shorter than the cells immediately subtending them. We tentatively identify the flattened cells as the epithallial cells and the subtending layer as the meristem.
All studied specimens which were observed overlapping each other are sterile, with the exception of the single plant described here, which bears calcified sporangial compartment observed on the two spots of the thallus. Proposed sori are raised above the thallial surface and consist of several badly preserved compartments filled by vegetative cells (Fig. 4c). Solitary compartments are also found (Fig. 4d). None of the observed compartments is borne above a layer of elongate cells. Sporangial compartments are 51-52 μm high and 21-28 μm in diameter. Gametophytes were not observed.
Remarks. Species is known from the locality Čaplovka and is absent in Ježov vrch. Size of sporangial compartments, absence of elongate cells below them, growth form and characteristics associated with the vegetative filaments collectively separate this species from other known sporolithoids documented in the Tich� Dolina valley.
|
|
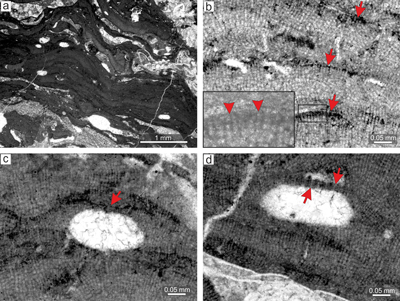
Figure 4: Sporolithon
sp. 1. a) Studied specimen is growing at the surface of the multispecific rhodolith. Arrow points to the specimen with encrusting-layered
growth form. Note multiple overgrowths and applanate branches at the base and the margin of the thallus. b) Detail of multiple overgrowths. Thalli continue to grow from damaged surfaces and overlap old one below, which is delineated by layers of epithallial cells (arrows). Arrowheads point to the cell fusions in ventral core as well as in peripheral filaments. Note non-coaxial arrangement of ventral core filaments. c) Protruding structure that is considered to be sorus (arrow). Note the badly preserved outline of the proposed sporangial compartments which are borne above the
non-elongate cells (arrowhead). Empty chambers are filled by vegetative filaments. Note the lightened scar-like structures at the lower part and top right corner of the figure. This structures could be indicative of flaked-off empty sori. d) Solitary calcified sporangial compartment partly filled by vegetative cells (arrow). |
Sporolithon sp. 2
(Fig. 5 ![]() )
)
Selected specimens examined. The species is present only in thin section 28080. Similar sterile thalli are found in other thin sections but cannot be identified adequately. Species description is based on the specimen from thin section 28080.
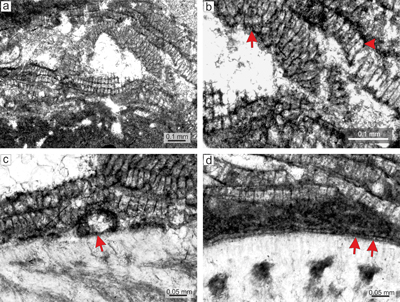
Material. Species is non-geniculate with encrusting-layered growth form (Fig. 5a) and develops thin crusts with numerous applanate branches (Fig. 5b). The greatest thickness (about 350 μm) was measured on fertile parts of the thallus where calcified sporangial compartments markedly protrude above the thallial surface. Species is present in sequences of larger multispecific rhodoliths.
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upward to peripheral region where they grow perpendicular to the substrate. The branching pattern of filaments is monopodial and pseudodichotomous. The ventral core is non-coaxial, up to 144 μm thick, and in section it consists of three to 14 filaments. Cells are rectangular or bladed and measure 16-33 μm (mean 2 5 μm � 5.3) in length and 7-15 μm (mean 11 μm � 2.4) in diameter (n = 19). The peripheral region is thin, usually as thin as or thinner than the ventral core and consists of rectangular, polygonal to square cells which are 12-26 μm (mean 17 μm � 4.1) long and 7-16 μm (mean 12 μm � 2.2) in diameter (n = 18). Pseudodichotomous branching pattern occurs in polygonal or bladed cells. Cells of the adjacent filaments are laterally connected by fusions. We have observed flattened and rectangular cells terminating the filaments (Fig. 5b). Cells are 5-7 μm (mean 6 μm � 0.8) long and 10-13 μm (mean 12 μm � 1.4) in diameter (n = 4). Cells below them are as long, slightly elongate, or shorter than the cells immediately subtending them. We tentatively identify the flattened cells as epithallial and the subtending layer as the meristem.
Calcified sporangial compartments are arranged in the sori built by a few (less than ten) compartments (Fig. 5c). A solitary sporangial compartment was also found (Fig. 5c). Calcified sporangial compartments are not borne above the layer of elongate cells. Sori protrude 141-187 μm above the thallial surface and their external diameter reaches 528-578 μm. Compartments are 46-53 μm in diameter (mean 50 μm � 2.8) and 77-102 μm high (mean 92 μm � 11.21) (n = 4). No sporangial complexes were observed. Pore canals were not detected. However, well-rounded chambers can be present below four-celled and 17 μm-long filaments (Fig. 5d). No gametophytes were observed.
Remarks. This species is similar to S. nummuliticum as it possesses cells and sporangial compartments with overlapping dimensions. However, there are three diagnostic features which enable us to separate S. nummuliticum and Sporolithon sp. 2. The first is the absence of elongate cells below the compartments. Second, the calcified sporangial compartments forming the sorus are markedly protruding above the thallial surface. Third is the presence of thin thalli with numerous applanate branches and encrusting-layered growth form. Since the first two characters have high diagnostic value and are commonly used by biologists as well as palaeontologists in the diagnosis of Sporolithon species we elected to separate the two species. Specimens, including gametophytes, from the Tich� Dolina locality, an historical collection, are described in Hrabovsk� et al. (2022).
|
|
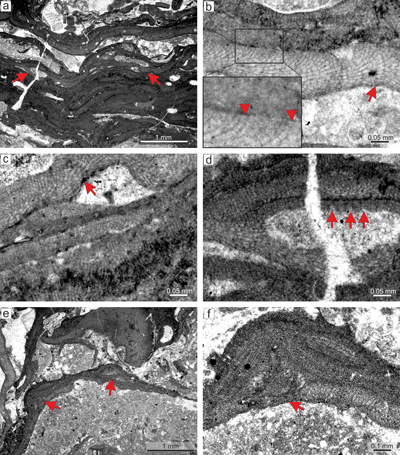
Figure 5: Sporolithon
sp. 2. a) Encrusting-layered growth form with sori protruding above the thallial surface. Specimen overgrows another coralline alga (arrow). b) Monomerous thallus construction with non-coaxial arrangement of ventral core filaments. Note the cell fusions (arrows). c) Fertile
thallus. Arrows point to the cells below the calcified sporangial compartments that are not
elongate. Note the solitary compartment at the top of the figure. d) Protruding sorus built by few - less than ten compartments (arrow). |
Order Hapalidiales Nelson et al., 2015
Family Lithothamniaceae Haas, 1886
Genus Lithothamnion Heydrich, 1897
Type species: Lithothamnion muelleri Lenormand ex Rosanoff, 1866, Westernport Bay, Victoria, Australia, Recent.
Lithothamnion camarasae Pfender, 1926
(Fig. 6 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31001, 31003, 31006; Ježov vrch: thin sections 28092, 28021, 28086, 28084.
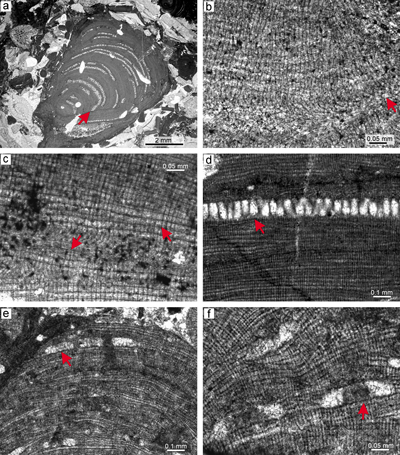
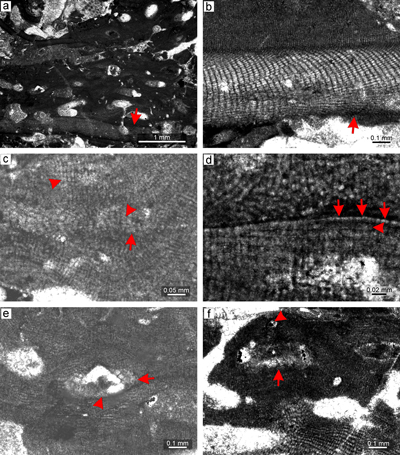
Material. Species description is based on the bi/tetrasporangial specimens from thin section 28081 (Fig. 6a). The proposed male gametophyte is described based on the specimen from thin section 28086 (Fig. 6b). The proposed female-carposporangial gametophyte is described based on the specimen from thin section 31001 (Fig. 6c). The species is non-geniculate, displaying an encrusting fruticose or warty-protuberant growth form. Applanate branches were not observed.
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upward into the peripheral region where they grow perpendicular to the substrate. Branching pattern of the filaments is monopodial or pseudodichotomous. Secondary ventral core is present in this specimen only (Fig. 6d). The secondary ventral core consists of six to 10 filaments and is up to 40-86 μm thick in its thickest portion. Cells are poorly preserved. They are rectangular and measure 12-26 μm (mean, 19 μm � 4) in length and 5-10 μm (mean 7 μm � 1.4) in diameter (n = 19). Cells in the peripheral region are rectangular to square, 6-17 μm (mean 10 μm � 2.5) long and 5-10 μm (mean 8 μm � 0.9) in diameter (n = 45). Cells of the adjacent filaments are laterally connected by fusions (Fig. 6e). Epithallial cells are rectangular or, in some cases, flared cells were observed (Fig. 6e). Epithallial cells are 2-4 μm long (mean 3 μm � 0.5) and 6-8 μm in diameter (mean 7 μm � 0.9) (n = 8). Epithallial cells are preserved on the sporangial conceptacle roof. Meristematic cells were not observed.
Reproduction. Multiporate sporangial conceptacles are borne on the sides and the tips of protuberances (Fig. 6a). Conceptacles very slightly protrude above the thallial surface. They are embedded within the thallus by continual growth of the perithallial filaments from the sides of the conceptacles or from their roofs and can leave small openings above the roof. Chambers are rectangular with rounded corners. Chambers are empty, though remains of adjacent large cells are present. Their internal diameter is 400-529 μm (mean 450 μm � 69.5) and they are 167-200 μm (mean 179 μm � 18.5) high. Pore canals are cylindrical (Fig. 6f). Roof filaments are four- to five-celled. Roof cells are 5-11 μm (mean 7 μm � 1.6) long and 4-7 μm (mean 6 μm � 0.8) in diameter (n = 44). Pore canals are lined by cells which are similar in size to other roof cells (Fig. 6f). Pore canal lining cells are 5-10 μm (mean 8 μm � 1.5) long and 4-9 μm (mean 6 μm � 1.3) in diameter (n = 26). Proposed male gametophyte bears uniporate conceptacles with a flat floor (Fig. 6b).The largest chamber of the conceptacle with visible pore canal is 300 μm in diameter and 60 μm high. The proposed female-carposporangial gametophyte bears few conceptacles of different morphology (Fig. 6c) borne at the tip of the protuberance, and only one of them has a visible pore canal. Conceptacles have a flat or convex floor. Unfortunately, none of the thin sections is oriented to show this feature satisfactorily. Conceptacles are 92-477 μm in diameter and 102-135 μm high. We tentatively identify the small, more-or-less flattened to triangular chamber as a carpogonial conceptacle while the single large one as a carposporangial conceptacle.
Remarks. Described specimen matches the neotypified L. camarasae described in Aguirre et al. (2011). It also matches the lower Priabonian specimen from the remote �trba locality (Hrabovsk� et al., 2022). Note that almost all studied specimen bears well-preserved pore-canal-lining cells, at least, in some conceptacles. Epithallial cells also commonly occur. We cannot confirm the presence of the species in the historical collection for the following reasons: the species from thin section 11Ad/304/ (p. 131, Fig. 62, Schalekov�, 1962) described in Schalekov� (1962) has fruticose protuberant growth form, small and flat conceptacles, sporangial conceptacles with thin roofs and a zonation pattern different from the neotype diagnosis (see Aguirre et al., 2011). This species, formerly described as L. cf. camarasae, does not match the specimens from Tich� Dolina and �trba section (Hrabovsk� et al., 2022, this work).
|
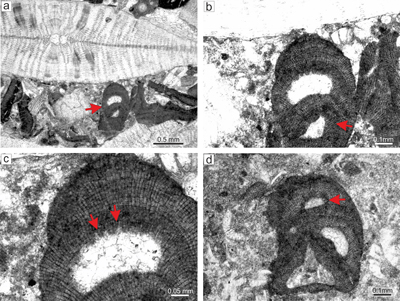
|
Figure 6: Lithothamnion camarasae Pfender, 1926. a) Sporophyte with
fruticose-protuberant growth form. Thallus exhibits numerous sporangial conceptacles borne on the sides as well as tips of protuberance. b) Proposed male
gametophyte. Uniporate male conceptacles are marked by arrows. c) Proposed
female-carposporangial gametophyte. Note the two types of conceptacles, a small one on the upper part of the thallus (arrow) and the large one below (arrow). d) Secondary ventral core (arrow). e) Relicts of epithallial cells which are rectangular to flared (arrows). f) Detail of cylindrical pore canals (arrows) which are lined by cells similar in morphology with adjacent roof cells. |
Lithothamnion cf. corallioides (P.L. Crouan & H.M. Crouan) P.L. Crouan & H.M. Crouan, 1867
(Fig. 7 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31001, 31002, 31004, 31022, 31016, 31021, 31018, 31025, 31005, 31006, 31012, 31015; Ježov vrch: thin sections 28088, 28089, 28079, 28086. Among tens of sporophytes four gametophytes are known.
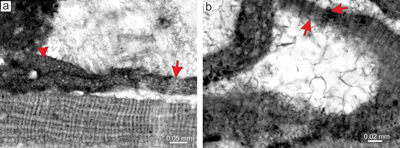
Material. Lithothamnion cf. corallioides is a common species in the samples from both sites. The description of the species is based on the bi/tetrasporangial specimen from thin section 31016 and the gametophyte from thin section 31004. Additional description of epithallial cells is based on the specimen from thin section 31016 (Supplement 1). Species has fruticose protuberant growth form with simple protuberances up to 1 mm long and 0.5 mm wide (Fig. 7a). It is not predominant in any sample.
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments which form the ventral core and bend upward forming the peripheral region where they grow perpendicular to the substrate (Fig. 7b). Most of the observed specimens, including the described one, have secondary ventral cores above the conceptacles or damaged thalli while others are fragments lacking the ventral core. Secondary ventral cores consist of a maximum of five filaments in the section and are 30-47 μm thick. Cells are rectangular and measure 11-24 μm (mean 17 μm � 3.8) in length and 6-8 μm (mean 6 μm � 0.5) in diameter (n = 10). Cells in the peripheral region are rectangular, square to bladed, 6-22 μm (mean 13 μm � 3.8) long and 5-12 μm (mean 8 μm � 1.5) in diameter (n = 35). Cells of the adjacent filaments are laterally joined by numerous fusions (Fig. 7c). Branching pattern of the filaments is monopodial and pseudodichotomous. The latter is common and was observed in elongate large cells at the sides of the conceptacles as well as in the peripheral filaments. These cells have bladed morphology and produce two filaments with smaller and thinner cells. Epithallial and meristematic cells were not observed in this specimen but flared epithallial cells were detected in gametophytes from the same thin section (Supplement 1).
Reproduction. Multiporate sporangial conceptacles are developed on the tips of protuberances and protrude slightly above the thallial surface. Conceptacles are embedded within the thallus by continual growth of the perithallial filaments from the sides of the conceptacles, leaving openings above the chambers. Some conceptacles do not have such openings and the roof appears to have vanished. Chambers are rectangular with rounded corners. Their diameter is 214-257 μm (mean 233 μm � 22) and are 91-111 μm high (mean 101 μm � 10). Pore canals are cone-shaped (Fig. 7c) with diameter about 10 μm. Roof filaments are three- to four-celled. Pore-lining filaments are of the same length and consist of the same number of similar cells as observed in the roof filaments (Fig. 7c). Roof cells are 5-10 μm long and 5-8 μm in diameter (n = 18). Pore canal cells are 6-9 μm long and 5-8 μm in diameter (n = 10). Proposed gametophyte bears few conceptacles borne at the tip of the protuberance, of which only one has a visible single pore canal (Fig. 7d). A conceptacle with visible pore canal has flat to slightly convex floor. This conceptacle protrudes 250 μm above the thallial surface and its external diameter is 462 μm. Chamber is 274 μm in diameter and 114 μm high. Pore canal is 143 μm long and 82 μm wide at the base and only 18 μm at the top. The smaller conceptacle at the top of the protuberance shows enlarged cells at the sides.
Remarks. Lithothamnion cf. corallioides is one of the most frequent species in the Čaplovka and Ježov vrch localities, but it is scarce in the other lower Priabonian site, �trba, where it is represented by just two specimens (Hrabovsk� et al., 2022). Gametophytes are known only from recent study. Morphological characteristics suggest this species can be well-defined. Based on the available data, new collections and historical collections (Schalekov�, 1962) we can conclude that 1) the species is absent from the historical collections and 2) there are no other non-geniculate coralline algae in the studied and other known material from CCPB with protuberances of such morphology and dimensions. Unfortunately, there is only one gametophyte specimen with preserved flared epithallial cells.
|
|
Figure 7: Lithothamnion cf. corallioides
(P.L. Crouan & H.M. Crouan) P.L. Crouan & H.M. Crouan, 1867. a) Fragment of fruticose-protuberant
growth form (arrow). b) Secondary monomerous non-coaxial ventral core filaments (arrow). c) Multiporate sporangial conceptacle embedded within the
thallus. Arrows point to the cells lining the cone-shaped pore canals. These cells are same as other roof cells (arrows). d) Supposed gametophyte with uniporate conceptacles. Arrow points to the large cells at the sides of the chamber. Two conceptacles may be considered as carpogonial (small one) and carposporangial (large one). |
Lithothamnion prascoi Mastrorilli, 1968
(Fig. 8 ![]() )
)
Selected specimens examined. Čaplovka: one fertile specimen in each of the thin sections 31007 and 31024.
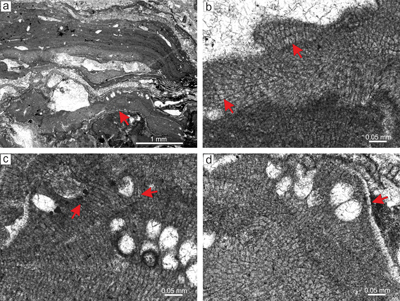
Material. The species is described based on the specimen from thin section 31024. This species is rare in the studied material. L. prascoi forms crusts in large multispecific rhodoliths. It is non-geniculate with an encrusting to lumpy or warty-protuberant growth form with applanate branches (Fig. 8a).
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 8b), consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upward forming the peripheral region where they grow perpendicular to the substrate. Ventral core is non-coaxial. It consists of five to 10 filaments and is 31-62 μm thick. Branching pattern of ventral core filaments is monopodial. Cells are rectangular, 7-15 μm (mean 11 μm � 2.3) in length and 4-6 μm (mean 5 μm � 0.7) in diameter (n = 20). Cells in the peripheral region are rectangular to square, 5-15 μm (mean 10 μm � 2.6) long and 4-9 μm (mean 7 μm � 1) in diameter (n = 47). Peripheral filaments are weakly stratified (Fig. 8a) or stratification may be absent from some places (Fig. 8c). Cells of the adjacent filaments are laterally joined by fusions. Epithallial cells are flattened, rectangular and occasionally flared, 3-4 μm long and 6-7 μm in diameter (Fig. 8d).
Multiporate sporangial conceptacles with visible pore canals are 140-210 μm in diameter and 79-102 μm high. Roof filaments are mostly three-celled but some are four-celled. All of the observed roofs are badly preserved and discrimination of cells was problematic. Only a few well-preserved filaments were found and measured. Roof cells are 5-9 μm long and 5-8 μm in diameter (n = 11). The roof filaments are 27-28 μm long. Proposed pore canal cells are of comparable dimensions to other roof cells (Fig. 8c-d). Pore canals are cylindrical with diameter 6-7 μm. Cells at the sides of conceptacles are not markedly elongate. Gametophytes were not observed.
Remarks. This species is clearly distinguished from other Lithothamnion species by 1) the encrusting-lumpy to warty-protuberant growth form, 2) the thin ventral core, 3) the development of conceptacles in encrusting as well as protuberant growth forms and 4) the small conceptacles, each with 5) a thin roof. It is worth noting that the morphology of Lithothamnion prascoi matches most of the morphological characteristics of Mesophyllum sp. Features that separate them are flared epithallial cells observed in some parts of L. prascoi specimens, the non-coaxial ventral core and the absence of specialized pore canal cells, which are often observed in Mesophyllum sp. However, many conceptacles do not show pore canals with visible pore-canal-lining filaments and do not bear preserved epithallial cells. Hence, only the ventral core arrangement is able to distinguish these species in many occurrences. We were not able to detect any L. prascoi in the historical collection.
|
|
Figure 8: Lithothamnion prascoi Mastrorilli, 1968. a) Proposed encrusting layered
growth form. Two sporangial conceptacles are protruding above the thallial surface. Note weak stratification of the peripheral filaments. b) Monomerous thallus construction with badly preserved non-coaxially arranged ventral core filaments (arrow). c) Part of the thallus where stratification of peripheral filaments lacks. Note multiporate sporangial conceptacle with badly visible pore canals. d) Detail of the multiporate sporangial conceptacles with preserved roofs and visible cylindrical pore canals. Proposed pore canals lined with filaments (arrows) consisting of cells similar in morphology with adjacent roof cells. Note the single layer of flattened to flared epithallial cells at the conceptacle roof (middle arrow). |
Lithothamnion roveretoi Airoldi, 1932
(Fig. 9 ![]() )
)
Selected specimens examined. Caplovka, thin section 31018.
Material. The species is described based on the specimen from thin section 31018. This is the only thin section from our collections that bears L. roveretoi. The species is non-geniculate with an encrusting-layered to lumpy-protuberant growth form (Fig. 9a).
Description. Small protuberances are developed at the fertile places on the thallus. Multiple overgrowths of thalli may develop lumpy protuberances. Applanate branches were not observed. The thallus is pseudoparenchymatous with monomerous construction (Fig. 9b), consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upward to the peripheral region where they grow perpendicular to the substrate. Branching pattern of the filaments is monopodial. The ventral core is non-coaxial and consists of five to 10 filaments and is 40-55 μm thick in its thickest portions. Cells are rectangular and measure 16-20 μm (mean 18 μm � 1.7) in length and 5-9 μm (mean 7 μm � 1.2) in diameter (n = 18). Cells in the peripheral region are rectangular to square, 5-13 μm (mean 10 μm � 1.9) long and 5-8 μm (mean 7 μm � 0.8) in diameter (n = 36). Cells of adjacent filaments are laterally joined by fusions (Fig. 9c). Epithallial cells are rectangular 3-4 μm long and 4-7 μm in diameter (Fig. 9d). Epithallial cells are preserved above the elongate cells at the conceptacle roof.
Reproduction. Multiporate sporangial conceptacles are embedded within the thallus by continual growth of the perithallial filaments from the roofs and sides of the conceptacles. Chambers with visible pore canals are rectangular with rounded corners.
Chambers are 94-117 μm high (mean 103 μm � 9.4) and their diameter is 242-451 μm (mean 308 μm � 97.3) (n = 4). Pore canals are cylindrical with 7-11 μm diameter. Roof filaments are three- to four-celled and 32-43 μm long. Pore-lining filaments are three- to four-celled also and consist of cells similar to the other roof cells. Roof cells are 6-11 μm long and 5-8 μm in diameter (n = 21), i.e., similar to the cells in the peripheral region (Fig. 9d). Pore-canal-lining cells are 5-9 μm long and 6-7 μm in diameter (n = 8). Gametophytes not observed.
Remarks. Species match with the diagnosis of the type provided by Vannucci et al. (2000). The exceptions are some conceptacles with a smaller internal diameter (about 80 μm) than those reported for the type. These conceptacles are most likely cut at their periphery. However, according to Vannucci et al. (2000), roof filaments of the Lithothamnion roveretoi multiporate sporangial conceptacles have similar dimensions to perithallial cells. This character is diagnostic in separating L. roveretoi from other known Lithothamnion species found in the lower Priabonian of CCPB. We were not able to detect any L. roveretoi in the historical collection.
|
|
Figure 9: Lithothamnion roveretoi Airoldi, 1932. a) Lumpy-protuberant
growth form with numerous conceptacles. b) Monomerous thallus construction with non-coaxially arranged ventral core filaments (arrow). Note the slightly
elongate cells at the conceptacle side (arrowhead). c) Cell fusions in ventral core and peripheral region of the thallus (arrows). d) Epithallial cells (arrow at the top) developed above the layer of
elongate cells. Note the cylindrical pore canal lined by cells which are similar with other roof cells (lower arrow). |
Lithothamnion cf. ramosissimum (Reuss) Piller, 1994
(Fig. 10 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31001, 31002, 31022, 31003, 31016, 31021, 31017, 31005, 31006, 31007, 31008, 31011; Ježov vrch: thin sections 28093, 28094, 28095, 28098, 28090, 28091, 28084, 28085.
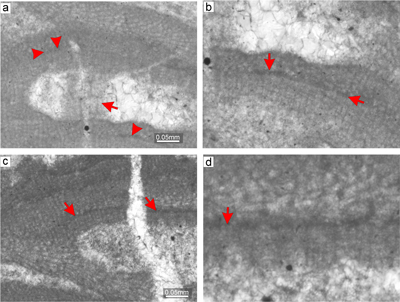
Material. Lithothamnion cf. ramosissimum is a frequent species at both sites. Bi/tetrasporangial as well as gametophytic plants are described based on the specimens from thin section 28091 where both plants are apparently fused. However, the protuberant growth form is better exposed in the thallus from thin section 28094 (Supplement 2). The species forms a rigid framework with numerous primary voids, in association with some other melobesioids. The species is non-geniculate with encrusting (Fig. 10a) to warty to fruticose-protuberant growth form. Applanate branches were not observed.
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments running parallel to the substrate in the ventral core and bending upwards to form the peripheral region, where they grow perpendicular to the substrate. Branching pattern of the filaments is monopodial. The ventral core is coaxial (Fig. 10b) to non-coaxial (Fig. 10c), consists of 11-25 filaments, and is 158-185 μm thick in its thickest portion (Fig. 10b). Cells are rectangular and measure 22-47 μm (mean 32 μm � 8.7) in length and 9-15 μm (mean 12 μm � 1.8) in diameter (n = 12). Cells in the peripheral region are rectangular to square, 13-27 μm (mean 17 μm � 3.5) long and 10-16 μm (mean 13 μm � 1.5) in diameter (n = 27). Cells of adjacent filaments are laterally joined with fusions (Fig. 10c). Epithallial cells are rectangular and occasionally exhibit a flared morphology (Fig. 10d, Supplement 2). Meristematic cells were not observed.
Reproduction. A single multiporate sporangial conceptacle was observed. The conceptacle protrudes 108 μm above the thallial surface and its external diameter is 952 μm. The conceptacle chamber is rectangular with rounded corners and is not embedded within the thallus. It is 129 μm high and 585 μm in diameter. Pore canals are cylindrical with 9-13 μm diameter. Roof filaments are made of three or four cells 31-33 μm long (Fig. 10d). Pore-lining filaments are made of three or four cells similar to those of the roof (Fig. 10d). Roof cells are 6-11 μm long and 6-9 μm in diameter (n = 10). Pore-canal-lining cells are barely visible (Fig. 10d). They are 5-10 μm long and 6-10 μm in diameter (n = 17). The proposed gametophyte bears numerous conceptacles which are developed in the fruticose protuberant thallus (Fig. 10e). Conceptacles are 120-300 μm in diameter and are 100-120 μm high. Conceptacles are borne mostly at the tips of protuberance and are embedded in the thallus. Pore canals are up to 100 μm long and 40 μm in diameter. The chambers have flat floors (Fig. 10f). The roofs of these conceptacles are developed by filaments peripheral to the fertile area (Fig. 10f).
Remarks. The studied specimens broadly match the type described by Aguirre et al. (1996) from the Badenian of the Vienna Basin. However, they differ from the type in 1) length of the ventral core cells in Priabonian species that exceeds the length of the cells of the type material; 2) fruticose-protuberant growth form of the type while encrusting to warty to fruticose-protuberant form occurs in the studied specimens, and 3) coaxial to noncoaxial arrangement of the ventral core filaments. In fact, there are numerous protuberant growth forms in the studied Priabonian limestone that suggest an unattached growth habit in this species. Unattached rhodoliths of Langhian-Serravallian L. ramosissimum (Aguirre et al., 1996; Hrabovsk� & Fordin�l, 2013) are characteristic. Therefore, its presence in the rigid coralline algal framework calls into question the presence of L. ramosissimum in studied samples. This conclusion is supported by the presence of long cells of the ventral core which are absent from the type species. The species was found in the historical collection - Tich� dolina material of Schalekov� (1962) and published in Hrabovsk� et al. (2022). The species was not found in the remainder of the historical collection.
|
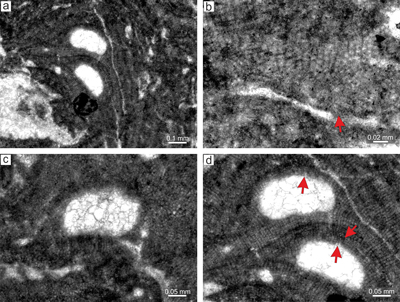
|
Figure 10: Lithothamnion cf. ramosissimum (Reuss) Piller, 1994. a) Portion of the thallus with encrusting
growth form. Arrow indicate multiporate sporangial conceptacle embedded in the
thallus. b) Common lateral fusion of cells in adjacent filaments (arrows). Note coaxial arrangement of the ventral core filaments. c) Portion of the thallus with non-coaxial arrangement of the ventral core filaments. Arrows point to the rows where division of cells was asynchronous. d) Multiporate sporangial conceptacle with pore canals lined by cells which are same as other roof cells (arrows). Note relicts of flared epithallial cells at the top of the conceptacle (arrowheads). e) Proposed gametophyte with encrusting
fruticose-protuberant thallus bearing numerous uniporate conceptacles (arrows). f) Detail of the gametangial conceptacle with visible pore canal and flat to slightly convex floor. The roof and the pore canal were formed by filaments peripheral to fertile area (arrow). |
Lithothamnion sp.
(Fig. 11 ![]() )
)
Selected specimens examined. Ježov vrch: thin sections 28098 and 28088.
Material. This species displays a fertile specimen in thin section 28098. Sterile thallus from 28088 bears the same anatomical characteristics of the vegetative filaments and growth form. The species is non-geniculate with an encrusting to lumpy-protuberant growth form without applanate branches (Fig. 11a).
Description. The thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments which run parallel to the substrate in the ventral core and bend upwards to form the peripheral region where they grow perpendicular to the substrate. Only the secondary ventral core is present. The secondary ventral core is very thin and consists of two or three filaments (Fig. 11a-c). Cells are rectangular, 11-53 μm (mean 33 μm � 10.6) in length and 7-21 μm (mean 14 μm � 3.5) in diameter (n = 25). Cells in the peripheral portion of the thallus are rectangular to square, 10-44 μm (mean 23 μm � 8) long and 9-22 μm (mean 14 μm � 2.7) in diameter (n = 38) (Fig. 11b). Cells change markedly in length in the vertical and horizontal planes and hence exhibit irregular stratification of peripheral filaments. Cells of adjacent filaments are laterally joined by fusions (Fig. 11c). Branching pattern of the perithallial filaments is monopodial, occasionally pseudodichotomous (Fig. 11c). Epithallial cells are rectangular or flared, 5-10 μm long and 12-16 μm in diameter (n = 11). However, their outline is poorly visible. Meristematic cells are elongate.
Reproduction. Two multiporate sporangial conceptacles with probable visible pore canals were observed (Fig. 11c-d). Conceptacle chambers are 323-449 μm in diameter and 186-189 μm high. Roof filaments are two- to four-celled (Fig. 11d). Roof cells are 5-15 μm long and 5-12 μm in diameter (n = 14). The roof filaments are 41-58 μm long. Pore canal filaments are also two- to four-celled, with cells which are 7-12 μm long and 8-11 μm in diameter (n = 8). Thus, the roof and the pore-canal cells are of comparable size. Pore canals are cylindrical with a diameter of 16-17 μm.
Remarks. This species is readily distinguished from other Lithothamnion species based on 1) the length of the cells of vegetative filaments and 2) the thin two- or three-celled ventral core. Sterile portion of the thallus may be erroneously interpreted as Titanoderma because of the rarity of lateral fusion between the cells of adjacent filaments and the apparent patchily dimerous internal organization. Presence of multiporate sporangial conceptacles places this species within the Order Hapalidiales. Flared epithallial cells, non-specialized pore-canal cells and non-coaxial ventral core place this specimen in the genus Lithothamnion within the family Lithothamniaceae. On the other hand, the reduced ventral core suggests that the species belongs to the subfamily Melobesioideae within the family Hapalidiaceae. Here, Exilicrusta bears flared epithallial cells and non-specialized pore-canal-lining cells such that it is very similar to the observed Lithothamnion sp. Lack of data about the primary ventral core prevents confident classification of the studied specimens. However, we tentatively place it within the genus Lithothamnion. This species is absent from the historical collection.
|
|
Figure 11: Lithothamnion sp. a) Encrusting to lumpy-protuberant
growth form with embedded multiporate conceptacles. Arrows point to the preserved secondary ventral core that is significantly reduced. b) Section through the thallus with
elongate cells. Arrows point to the thin secondary ventral core. c) Few cell fusions (arrows) between the cells of adjacent filaments. Arrowheads point to the secondary ventral core where thallus construction appears almost as
dimerous. d) Multiporate sporangial conceptacle. Arrows point to the pore canals lined by badly preserved cells appearing of the same size as other roof cells. |
Genus Phymatolithon Foslie, 1898
Type species: Phymatolithon calcareum (Pallas) Adey & McKibbin ex Woelkerling & Irvine, 1986, Falmout Harbour, Cornwall, England, Recent.
Phymatolithon sp.
(Fig. 12 ![]() )
)
Selected specimens examined. Ježov vrch: thin section 28097.
Material. The species occurs in only one sample from the Ježov vrch locality, thin section 28097, and is described based on this specimen. It is non-geniculate with encrusting to lumpy-protuberant growth form (Fig. 12a) with applanate branches (Fig. 12b).
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 12b), consisting of a single system of branched filaments which run parallel to the substrate to form the ventral core and bending upwards forming the peripheral zone where they grow perpendicular to the substrate. Branching pattern is monopodial. The ventral core is non-coaxial and consists of three to nine filaments and is 28-58 μm thick. Ventral core cells are rectangular, 10-23 μm (mean 16 μm � 3.2) in length and 5-11 μm (mean 7 μm � 1.3) in diameter (n = 26). Cells in the peripheral filaments are rectangular to square, 6-16 μm (mean 11 μm � 2.1) long and 5-10 μm (mean 8 μm � 1.2) in diameter (n = 52). Cells change markedly in length in the vertical and horizontal planes. Cells of adjacent filaments are laterally joined by fusions. Epithallial cells are rounded 3-4 μm long and 6-7 μm in diameter (n = 4) (Fig. 12b). However, their outline is barely distinguishable. Meristematic cells are shorter than cells immediately subtending them. They are 5 μm long and 6 μm in diameter (n = 4).
Reproduction. Multiporate sporangial conceptacles are 216-225 μm in diameter and 109-115 μm high. The roof consists of a peripheral rim and a sunken pore plate (Fig. 12c). In some cases the roof displays flat morphology (Fig. 12d). Pore plate-diameter is up to 93 μm and is depressed 9 μm below the rim. The external diameter of the conceptacles is 418 μm and conceptacles rise up to 71 μm above the thallus. Roof filaments are three-celled but, where the pore canals are not visible, they could be up to five-celled (Fig. 12d). Roof cells are 4-9 μm long and 3-8 μm in diameter (n = 31). Pore-canal filaments are also made of two to three cells that are 4-7 μm long and 4-6 μm in diameter (n = 20). Thus, the roof and the pore-canal cells are of comparable size (Fig. 12d). Pore canals are cylindrical with a diameter of up to 7 μm.
Remarks. This species matches the description of Phymatolithon sp. from the lower Priabonian locality �trba (Hrabovsk� et al., 2022). This species was not observed in the historical collection.
|
|
Figure 12: Phymatolithon sp. a) Encrusting to lumpy-protuberant
growth form. b) Detail of the thallus shows that the protuberances and thickened thallus is formed by numerous superimposed thalli (arrows). Note that the non-flared epithallial cells are developed in 2 layers above the meristematic cells which are as long as, or shorter than cells immediately subtending them (arrowheads). c) Multiporate sporangial conceptacle with peripheral rim and distinct pore plate (arrow). d) Multiporate sporangial conceptacle with flat roof. Arrows point to the proposed pore canals which are lined by cells similar to other roof cells. Note secondary ventral core growing from the sides of the conceptacles and leaving a small space above the conceptacles roof. |
Family Mesophyllaceae Athanasiadis, 2016
Genus Mesophyllum Lemoine, 1928
Type species: Mesophyllum lichenoides (Ellis) Lemoine, 1928, Cornwall, England, Recent.
Mesophyllum fructiferum Airoldi, 1932
(Fig. 13 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31011, 31012, 31014; Ježov vrch: thin section 28095.
Material. This species occurs at both localities. It is represented mostly by fragments of encrusting thalli. The species is described based on the largest crust observed in thin section 31012. This species is non-geniculate with an encrusting growth form, which does not produce applanate branches (Fig. 13a).
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments running parallel to the substrate, forming the ventral core and bending upwards to form the peripheral portion of the thallus where they grow perpendicular to the substrate. Branching pattern of vegetative filaments is monopodial and pseudodichotomous. Ventral core is poorly preserved and its large basal part was chemically dissolved as indicated by the presence of microstylolites. However, the coaxial ventral core is present in these places (Fig. 13b). The preserved ventral core is up to, 197 μm thick and consists of cells 12-23 μm long (mean, 19 μm � 2.9) and 6-11 μm in diameter (mean 8 μm � 1.2) (n = 27). The peripheral region is up to 1.4 mm thick with a distinct growth pattern. Cells are 8-20 μm long (mean 13 μm � 3.3) and 7-11 μm in diameter (mean 8 μm � 1) (n = 22). Cells are laterally joined by fusions (Fig. 13c). Proposed epithallial cells were observed on the roof of a multiporate sporangial conceptacle (Fig. 14d). Epithallial cells are rounded to flattened but not flared. Cells are 3-4 μm long (mean 3 μm � 0.4) and 6-8 μm in diameter (mean 7 μm � 0.8) (n = 5).
Multiporate bi/tetrasporangial conceptacles are present in rows within the thallus (Fig. 13a). Chambers are oval with a diameter of 396-481 μm and they are 138-175 μm high. The roof is flat and without a peripheral rim. The roof filaments are four-celled and 25-43 μm long, and consist of cells which are identical to cells lining the pore canals (Fig. 13d). Pore canals are cylindrical. Gametophytes are not present in the studied material.
Remarks. The described species matches the type revised by Basso et al. (1998) as well as the description provided by Hrabovsk� et al. (2022) for specimens from the �trba section. The species does not occur in the historical collection studied.
|
|
Figure 13: Mesophyllum fructiferum Airoldi, 1932. a) Encrusting
growth form. Note that the thallus is massive and thick with conceptacles developed in rows. b) Part of the fertile thallus with ventral core and peripheral filaments. Arrows point to the section where more-or-less coaxial arrangement may be visible. c) Fusion of the cells (arrows) in ventral core and peripheral filaments. d) Detail of the multiporate sporangial conceptacle roof. Arrow points to the supposed pore canal that is lined by cells which appear similar as other roof cells. Note the proposed single layer of rounded to flattened epithallial cells (arrowheads). |
Mesophyllum cf. engelhartii (Foslie) Adey, 1970
(Fig. 14 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31001, 31022, 31018, 31017, 31024; Ježov vrch: 28093, 28097, 28098, 28090, 28091, 28092, 28084, 28085.
Material. The bi/tetrasporangial plant is described based on the specimen from thin section 28097; gametophytes are present in thin section 28091. The species is non-geniculate with encrusting-layered growth form producing numerous applanate branches (Fig. 14a). Occasionally, it displays a lumpy-protuberant growth form. The species occurs in the coralline algal framework facies or within large multispecific rhodoliths.
Description. The thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments running parallel to the substrate and forming the ventral core, bending upwards to form the peripheral portion of the thallus. Branching pattern of vegetative filaments is monopodial and pseudodichotomous. The ventral core is predominantly coaxial but some sections may exhibit a non-coaxial arrangement (Fig. 14b-c). This may be an artefact of the orientation of the section through the thallus. Alternatively, the division of the cells in the ventral core filaments may have been asynchronous during at least short periods of the core's growth. The ventral core is 114-143 μm thick and consists of cells which are 17-30 μm long (mean 22 μm � 4) and 7-11 μm in diameter (mean 9 μm � 1.1) (n = 15). The peripheral portion of the thallus is thin and without distinct growth patterns. Cells are 11-17 μm long (mean 13 μm � 2.0) and 6-13 μm in diameter (mean 10 μm � 1.7) (n = 12). Proposed epithallial cells were observed in the roof of the multiporate sporangial conceptacle and above elongate cells (Fig. 14b, Supplement 3). Epithallial cells are flattened and rounded to hemispherical, but not flared. These cells are 3-5 μm long (mean 4 μm � 0.5) and 6-9 μm in diameter (mean 7 μm � 0.9) (n = 8).
The multiporate bi/tetrasporangial conceptacle markedly protrudes above the thallial surface (110 μm) and reaches an external diameter of 773 μm (Fig. 14d). The chamber is oval with a diameter of 652 μm and height of 128 μm. The roof is flat and without a peripheral rim. The roof filaments are four-celled and 38 μm long and consist of cells which are 9-13 μm long (mean 11 μm � 1.3) and 7-11 μm in diameter (mean 9 μm � 1.5). Pore canals are cylindrical with a 9 μm diameter and are lined by cells which are similar to the other roof cells. Openings to the pore canals are not sunken. A specimen with two distinct uniporate conceptacles, one smaller triangular and another larger with weak central pedestal is present in the material from the Ježov vrch (Fig. 14e). Our observations suggest the larger conceptacle developed from the smaller one by enlarging and destroying the cells at the sides of the chamber (Fig. 14f). Both of the conceptacles bear a pore canal and display a roof which suggests a development from initials (filaments) interspersed around the fertile area. We tentatively consider this specimen to be a female gametophyte. The absence of male conceptacles indicates that the gametophyte may be dioecious.
Remarks. The described specimen matches Mesophyllum cf. engelhartii from the Priabonian of �trba. Both bi-tetrasporangial and gametophytic plants are present. However, the coaxial arrangement of the ventral core is more distinct in the specimen from Oravice than in the �trba specimens. Inclusion of this specimen within the genus Mesophyllum is based on the morphological similarities to the Recent Mesophyllum engelhartii, as described by Woelkerling and Harvey (1993) (Hrabovsk� et al., 2022). However, the studied species bears a predominantly coaxial ventral core, a single layer of non-flared epithallial cells and elongate meristematic cells, carposporangial conceptacle with weak but preserved central pedestal and most likely dioecious gametophytes. All these characteristics are consistent with the genus Mesophyllum (Athanasiadis & Ballantine, 2014). This species does not occur in the historical collection.
|
|
Figure 14: Mesophyllum cf. engelhartii. a) Encrusting-layered
growth form (arrows) with applanate branches. b) Coaxial ventral core filaments (arrow) and rounded to flattened epithallial cells (arrowheads). c) Part of the thallus where coaxial arrangement of vegetative filaments is well-visible (arrow). d) Multiporate sporangial conceptacle with visible pore canals (arrows) lined by pore canal cells which are similar to the other roof cells. e) Fertile
female-carposporangial gametophyte with carposporangial conceptacle on the left (arrow) and carpogonial conceptacle on the right (arrow). f) Detail of the
female-carposporangial conceptacles borne above the ventral core (arrow). Note the weak central pedestal above the arrow. |
Mesophyllum sp.
(Fig. 15 ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31002, 31016, 31025, 31024, 31018, 31011; Ježov vrch: thin sections 28093, 28096, 28095, 28098, 28087, 28088, 28089, 28091, 28086, 28084, 28085.
Material. The species description is based on the specimen from thin section 31016. The species is non-geniculate with an encrusting to encrusting warty-protuberant growth form, without applanate branches (Fig. 15a). Species is frequent but never dominant in the studied material.
Description. The thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments that runs parallel with the substrate to form the ventral core and bending upwards to form the peripheral portion of the thallus. Branching pattern of vegetative filaments is monopodial and pseudodichotomous. The ventral core is thin, and discrimination between the coaxial and non-coaxial arrangement is problematic (Fig. 15b). However, some patches exhibit filaments with cells that point rather to synchronous division of these filaments although the full arches of cells are not present (Fig. 15b, Supplement 4). Secondary ventral core filaments above the conceptacles are often present (Fig. 15c). Ventral core is 29-63 μm thick and consists of bladed to rectangular cells which are 13-27 μm long (mean, 19 μm � 3.3) and 4-11 μm in diameter (mean 7 μm � 1.7) (n = 24). Peripheral region of the thallus is thick and zoned (Fig. 14a). Cells are 8-19 μm long (mean 14 μm � 2.3) and 6-10 μm in diameter (mean 8 μm � 1.3) (n = 18). Cells of the adjacent filaments are joined by fusions in the ventral core (Fig. 15c) and peripheral filaments (Fig. 15d). Epithallial cells are rounded and hemispherical to flattened, but not flared (Fig. 15d). Meristematic cells are elongate (Fig. 15c, e).
Multiporate bi/tetrasporangial conceptacles slightly protrude up to 54 μm above the thallial surface and were later embedded within the thallus by lateral overgrowths of the peripheral filaments (Fig. 15a-b). Their external diameter is up to 365 μm (Fig. 15a, e). Chambers are oval with a diameter of 174-260 μm (mean 217 μm � 60.2) and height of 118 μm. The roof is flat and lacks a peripheral rim. The roof filaments are four- to five-celled and 38 μm long, consisting of cells which are 6-8 μm long (mean 8 μm � 0.9) and 5-8 μm in diameter (mean 7 μm � 1.4). Pore canals are cylindrical 11 to 14 μm in diameter, and lined by specialized thinner-wider cells differing from other roof cells (Fig. 15f). Openings of the pore canals are not sunken. Gametophytes not observed.
Remarks. Mesophyllum sp. bears characteristics that are consistent with the diagnosis of Recent Mesophyllum, i.e., specialized pore-canal-lining cells in pore canals of multiporate sporangial conceptacles, elongate meristematic cells and non-flared epithallial cells. However, a permanently coaxial ventral core was not distinguished (see Athanasiadis & Ballantine, 2014). The species matches the �trba specimen described as Mesophyllum sp. 1 in Hrabovsk� et al. (2022). We have observed Mesophyllum sp. as fragments of protuberances in the biodetrital matrix as well as a part of multispecific rhodoliths. It is a frequent but not dominant species in the studied material. This species was not observed in the historical collection.
|
|
Figure 15: Mesophyllum sp. a)
Fruticose-protuberant growth form. b) Spots in the secondary ventral core filaments where division of cells was synchronous (arrows). c) Secondary ventral core above the conceptacle roof. d) Zoned peripheral region with numerous fusion of cells of the adjacent filaments (arrows). e) Preserved epithallial cells which are rounded and flattened (arrow). Note
elongate cells subtending them on some spots. f) Pore canals lined by specialized pore canal lining cells (arrow). |
Order Corallinales Silva & Johansen, 1986
Family Hydrolithaceae Townsend & Huisman, 2018
Subfamily Chamberlainoideae Caragnano et al., 2018
Genus Chamberlainium Foetisch et al., 2018
Type species: Chamberlainium tumidum (Foslie) Caragnano et al., 2018. Monterey, California, Recent
Chamberlainium lemoinei (Miranda) comb. nov.
(Fig. 16 ![]() )
)
Basionym. Melobesia lemoinei Miranda, 1935 (Algas coralin�ceas f�siles del Terciario de San Vicente de la Barquera (Santander); p. 284-285; Fig. 3A-3B, Pl. 38, Fig. 1).
2011 Hydrolithon lemoinei (Miranda) Aguirre et al., p. 282, fig. 6A-C.
Selected specimens examined. Fertile thalli are known only from the locality Čaplovka: thin sections 31018, 31020, 31005.
Material. Chamberlainium lemoinei is common in numerous samples where multispecific rhodoliths are present. Description of the bi/tetrasporangial plant and vegetative characteristics is based on the specimens in thin section 31020. The proposed gametophyte is described based on the specimen in thin section 31018. Sterile thalli from both studied sites are easily recognized because of the presence of large and often polygonal cells in thalli with dimerous internal organization and pseudodichotomous to monopodial branching pattern of the vegetative filaments. These thalli often bear hemispherical or flattened but not flared epithallial cells. The species is non-geniculate and has an encrusting growth form (Fig. 16a). Applanate branches were not observed.
Description. The thallus is pseudoparenchymatous with dimerous construction (Fig. 16b), consisting of the two systems of filaments, one basal single-layered primigenous and, perpendicularly to the primigenous filaments, the second a growth of postigenous filaments. Cells in the primigenous filaments are rectangular, squarish, or polygonal. Palisade cells were not observed. The cells measure 11-24 μm long (mean 17 μm � 3.8) and 11-24 μm in diameter (mean 16 μm � 3.6) (n = 18). The postigenous filaments consist of cells which are 8-27 μm long (mean 17 μm � 4.2) and 9-20 μm in diameter (mean 14 μm � 2.5) (n = 37). Postigenous filaments show monopodial and pseudodichotomous branching patterns (Fig. 16c). Epithallial cells are flattened or hemispherical but not flared (Fig. 16c). Cells are 3-7 μm long (mean 5 μm � 1) and 6-13 μm in diameter (mean 9 μm � 2.4) (n = 14). Cells of adjacent filaments are laterally joined by fusion (Fig. 16d).
The probable bi/tetrasporangial conceptacles are uniporate. The roof and the pore canal of the bi/tetrasporangial conceptacles are developed from filaments located peripherally to the fertile area (Fig. 16e). Small chambers are considered as initial stages of conceptacle primordia and are embedded within the thallus (Fig. 16e). Similar primordia were figured in extant Chamberlainium by Puckree-Padua et al. (2021). The large, empty, probably mature bi/tetrasporic conceptacles are markedly raised from the surrounding thallial surface (Fig. 16e). Their external diameter is 492-540 μm and they are raised 119-141 μm above the thallial surface (n = 2). Chambers are 238-259 μm in diameter and up to 91 μm in height with a pore canal 53-82 μm long and 48-49 μm wide. There is another embedded conceptacle within the thallus in which the pattern of the roof and the pore canal can be misidentified because in some places the roof filaments are oriented near perpendicular to the chamber while in others the roof filaments are oriented parallel to the chambers (Fig. 16f). Chamber is 181 μm in diameter and 72 μm high with pore canal 77 μm long and 24 μm in diameter. The roof filaments of all studied conceptacles are less than eight-celled and protrude into the pore canal (Fig. 16f).
We have observed one possible gametophytic plant in thin section 31018 (Fig. 16g). This specimen bears small flask-shaped conceptacles only. The morphology of these conceptacles as well as their pore canals differs from those described above. Chambers are 97-106 μm in diameter and 37-38 μm high (n = 2). The extended cylindrical pore canals are 116-119 μm long and 31-34 μm in diameter (Fig. 16h). The roof and the pore canals suggest the same pattern of development as observed in the bi/tetrasporic conceptacles described above. Tentatively we consider this plant to be a female gametophyte. Lack of the male conceptacles may suggest that the gametophyte is dioecious.
Remarks. Studied bi/tetrasporangial specimens match the neotype of H. lemoinei described in Aguirre et al. (2011) as well as the specimens from the lower Priabonian of �trba (Hrabovsk� et al., 2022) and Middle Eocene of Spain (Aguirre et al., 2020). This species is common in the Middle Miocene limestones of the central Paratethys (Chelaru and Bucur, 2016; Hrabovsk� et al., 2016; Hrabovsk�, 2019). Specimens from the historical collection were described and figured in Hrabovsk� et al. (2022). With respect to the description of the subfamily Hydrolithoideae by R�sler et al. (2016), specimens known as Hydrolithon lemoinei do not fit within the diagnosis because all known specimens bear pore canals surrounded by filaments that protrude into the pore canal (Aguirre et al., 2011; Hrabovsk� et al., 2016, 2022; Chelaru & Bucur, 2016; Hrabovsk�, 2019). Also, roof and pore-canal formations are not well-known and are often described as involving cells oriented roughly perpendicular to the chamber. However, figured specimens have roofs formed rather from filaments peripheral to the fertile area (Aguirre et al., 2011; Aguirre et al., 2020; Hrabovsk�, 2019). The same arrangement exists in the embedded conceptacle of the studied specimen. Re-examination of known Paratethys specimens led us to the conclusion that roof formation has previously been incorrectly interpreted. According to available data, thin dimerous thalli and filaments protruding into the pore canals are diagnostic of the subfamily Neogoniolithoideae (R�sler et al., 2016) or Chamberlainoideae (Caragnano et al., 2018). Therefore, the systematic position of this species is questioned. Following the Caragnano et al. (2018) definition of Chamberlainium and further consideration of fossil species in Coletti et al. (2020) for C. pentagonum and Spongites fruticulosus we re-assign H. lemoinei to C. lemoinei within the Corallinaceae based on the dimerous thallus, pore-canal filaments of sporangial conceptacles protruding into the pore canal, dimensions of conceptacles, roof filaments consisting of fewer than eight cells, manner of formation of the roof and the pore canal of sporophyte conceptacles, and dioecious gametophytes. Chamberlainium lemoinei, therefore, represents the second exclusively fossil species of Chamberlainium, together with C. pentagonum (Coletti et al., 2020).
|
|
Figure 16: Chamberlainium lemoinei
(Miranda) comb.
nov. a) Encrusting-layered growth form. b) Dimerous thallus construction with non-palisade cells in primigenous filaments (arrow). c) Layer of epithallial cells which are not flared (arrow). Arrowhead points to the pseudodichotomous branching pattern of the postigenous filaments. d) Fusion of cells (arrow). e) Proposed uniporate
bi/tetrasporangial conceptacle (arrowhead) and conceptacle primordia (arrows) suggesting the roof and the pore canal formation by filaments peripheral to fertile area. f) Detail of the proposed
bi/tetrasporangial conceptacle with filaments running more or less parallel to the chamber (arrowheads) as well as perpendicularly to the
thallial surface (arrow). g) Proposed female gametophyte with carpogonial conceptacles raised above the
thallial surface. h) Detail of the carpogonial conceptacle. Arrow points to the filaments which run parallel to the chamber. Note flattened hemispherical epithallial cells (arrowheads). |
Subfamily Mastophoroideae Setchell, 1943
Genus Lithoporella (Foslie) Foslie, 1909
Type species: Lithoporella melobesioides (Foslie) Foslie, 1909; South Nilandu Island, Maldives, Recent.
Lithoporella melobesioides (Foslie) Foslie, 1909
(Fig. 17a-b ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31025, 31017, 31020; Ježov vrch: thin sections 28091, 28084, 28080.
Material. Lithoporella melobesioides occurs in many thin sections from both localities, generally within large multispecific rhodoliths or around bioclasts. We have observed fertile specimens growing around brachiopods as well as upon nummulitids and other coralline algae. There are four fertile specimens among the studied samples. The description of the species is based on the specimen from thin section 31017 which displays the best preserved conceptacles and epithallial cells. The species is non-geniculate with an encrusting-layered growth form. The thallus shows numerous applanate branches and is predominantly monostromatic (Fig. 17a).
Description. Thallus consisting of two systems of filaments, primigenous and postigenous. Postigenous filaments are developed sporadically, only locally or around the conceptacles (Fig. 17a). These filaments show a monopodial branching pattern. The thallus mostly consists of primigenous cells and single layer of epithallial cells (Fig. 17b). Cells of primigenous filaments are rectangular to palisade, 14-76 μm in height (mean 49 μm � 23.5) and 13-29 μm in length (mean, 19 μm � 3.7). The length to the height ratio is 1:5. Epithallial cells are mostly hemispherical (Fig. 17b), rarely rounded or flattened. Cells are 10-17 μm long and 14-18 μm in diameter.
Proposed bi/tetrasporangial conceptacles protrude above the thallial surface for their whole height (Fig. 17a). Their external diameter is 612-683 μm and they are raised 269-306 μm. Chambers are 342-415 μm in diameter and 159-162 μm high. The roofs consist of two or three cells oriented perpendicular to the chamber. Pore canals are conical, 95-151 μm long and about 95 μm in diameter at the top. Gametophytes were not observed.
Remarks. Species is known from the Eocene and Oligocene limestones of the peri-Mediterranean area (Coletti et al., 2018; Rasser & Piller, 1999; Aguirre et al., 2020 as Lithoporella sp.). It is one of the most frequent species in the studied limestones commonly co-occurring with sporolithoids. Sterile thalli are common in the historical collection.
|
|
Figure 17: a-b) Lithoporella
melobesioides Foslie. a) Fertile thallus with proposed uniporate conceptacle. Species forms monostromatic thalli with numerous applanate branches overlapping each other. b) Rounded and hemispherical epithallial cells (arrowhead) and roof filaments perpendicularly oriented to the chamber (arrow). c-d) Lithoporella cf. minus Johnson,
1964. c) Fertile thallus with small chamber proposed to be
bi/tetrasporangial conceptacle. However, conceptacle is barely
visible (arrow). d) Monostromatic thallus with palisade cells (arrows). |
Lithoporella cf. minus Johnson, 1964
(Fig. 17c-d ![]() )
)
Selected specimens examined. Čaplovka: thin sections 31025, 31019; Ježov vrch: thin section 28097.
Material. This species is not as common as Lithoporella melobesioides but, in some samples, develops multiple stacked thalli forming thick crusts. The species commonly occurs encrusting pebbles. It is common in the studied material but fertile specimens are rare and restricted to the thin sections examined (see above). The description of the species is based on the specimen from thin section 28097.
Description. Non-geniculate species with an encrusting-layered growth form. The thallus shows numerous applanate branches and is exclusively monostromatic (Fig. 17c). The thallus has dimerous construction, consisting of two systems of filaments, primigenous and postigenous. Postigenous filaments are rarely developed so that primigenous filaments are usually terminated by epithallial cells. We were not able to observe the branching pattern of vegetative filaments. Cells of the primigenous filaments are rectangular to palisade, 5-10 μm in height (mean 7 μm � 1.3) and 3-9 μm in length (mean 4 μm � 1.6) (Fig. 17d). The length to the height ratio is 1:2.4. Epithallial cells are mostly hemispherical, rarely rounded or flattened.
A propable bi/tetrasporangial conceptacle protrudes above the thallial surface with its whole height. Its external diameter is 146 μm and it is raised 42 μm. The chamber is 86 μm in diameter and 40 μm in height. The roof filaments are single-celled and are oriented perpendicular to the chamber. Pore canals are not observed. Roof cells are 7-10 μm long and 6-8 μm in diameter.
Remarks. Lithoporella cf. minus matches specimens known in the �trba section as well as the Middle Miocene specimens (Hrabovsk�, 2019; Hrabovsk� et al., 2022). Specimens from the historical collection were published in Hrabovsk� et al. (2022). The size of the conceptacle is the smallest amongst known species of Lithoporella and is rather similar to that of Lithoporella sauvageaui (Foslie) Adey, 1970. Further study of new specimens is necessary for a proper identification of this species.
Corallinophycidae sp. 1
(Fig. 18 ![]() )
)
Selected specimens examined. Ježov vrch: thin section 28094.
Material. This species is described from thin section 28094, its only known occurrence. The species is non-geniculate with an encrusting-layered growth form (Fig. 18a) and numerous applanate branches.
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branching filaments which runs roughly parallel to the substrate where they form the ventral core of the thallus and then bend upward to develop the peripheral region. The ventral core is coaxial to non-coaxial (Fig. 18b). Secondary ventral core observed above the damaged portion of the thallus can be arranged coaxially or non-coaxially (Fig. 18c). Ventral core is 72-317 μm thick and consists of seven to 35 filaments. A lower number of filaments was observed in the secondary ventral core. Branching pattern of the filaments is monopodial. Cells in the ventral core are rectangular, 23-34 μm long (mean 28 μm � 2.5) and 9-17 μm in diameter (mean 13 μm � 2.4) (n = 36). Cells in the peripheral region are square to rectangular, 5-19 μm long (mean 12 μm � 2.5) and 7-11 μm in diameter (mean 8 μm � 1.4) (n = 73). Cells of adjacent filaments are laterally connected by fusions. Epithallial and meristematic cells are poorly preserved (Fig. 18d). However, visible epithallial cells appear as rounded to flattened and the meristematic cells are as long as the cells immediately subtending them.
Two types of conceptacles occur on the thallus. The small conceptacles are triangular and have chambers at their sides filled by large cells (Fig. 18e). The roof and the pore canal of the conceptacles are formed by filaments that run parallel to the conceptacle chamber suggesting their development from the initials (filaments) peripheral to the fertile area. Chambers have a weak central pedestal and are up to 112 μm high and 227 μm in diameter. The large conceptacles probably developed from the smaller ones through the destruction of the large cells. Chambers of the large conceptacles have a central pedestal (Fig. 18f). Height from the chamber floor to the roof is 140-193 μm while the height from the central pedestal to the roof is up to 127 μm. Pore canal is conical with a diameter at its base of 114 μm and at its top of 54 μm. Pore canals are 64-216 μm long. No other plants were observed.
Remarks. This species shows vegetative and reproductive characteristics consistent with either an unrecognized gametophyte or bi/tetrasporophytes, with the roofs of bi/tetrasporangial conceptacles developed from filaments peripheral to the fertile area. These are 1) a coaxial ventral core, 2) lateral fusion of the cells in vegetative filaments, 3) the shape of the epithallial cells and 4) conceptacle-style formation. Among known coralline algal genera, the gametophytes of Mesophyllum are characterized by a permanently coaxial ventral core and distinct central pedestal in the carposporangial conceptacles. Therefore, we conclude that the observed specimen may represent the carpogonial-carposporangial plant of the genus Mesophyllum. This suggestion is supported by the occurrence of non-flared epithallial cells. However, meristematic cells are not elongate. On the other hand, a coaxial ventral core, non-flared epithallial cells and short meristematic cells are characteristics of Crustaphytum (see Appendix) but little is known about its gametophyte, especially the morphology of the carposporangial conceptacles. Alternatively, Tectolithon bears meristematic cells of variable length and carposporangial conceptacles with a central pedestal but has a non-coaxial ventral core (see Appendix). Our identification is therefore hampered by the absence of bi/tetrasporangial plant and/or reproductive structures that rarely fossilize (Woelkerling et al., 2012; Teichert et al., 2019).
|
|
Figure 18: Corallinophycidae
sp. 1. a) Encrusting growth form. Note the thick thallus with numerous conceptacles and distinct ventral core. Arrow points to the coaxially arranged ventral core filaments. b) Coaxial to non-coaxial arrangement of ventral core filaments. Arrow points to the base of the ventral core where cells exhibit asynchronous division. c) Secondary ventral core and proposed applanate branch (arrow). Arrowheads point to the cell fusions. d) Hardly visible rounded epithallial cells (arrows) and meristematic cells (arrowhead) which are of the same length as the cells immediately subtending them. e) Triangular conceptacle with enlarged cells at the sides of the chamber (arrow). Arrowhead points to the weak central pedestal. f) Large conceptacle with preserved central pedestal (arrow). Arrowhead points to the single large pore canal. |
Corallinophycidae sp. 2
(Fig. 19 ![]() )
)
Selected specimens examined. Ježov vrch: thin section 28097.
Material. This species is described based on a single specimen from thin section 28097. It is non-geniculate with an encrusting growth form (Fig. 19a) and numerous applanate branches.
Description. Thalli are 44-251 μm thick and are thickest at fertile parts. The thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branching filaments which runs roughly parallel with the substrate where they form the ventral core of the thallus and bend upward to develop the peripheral zone. The branching pattern of the vegetative filaments is monopodial. The ventral core is non-coaxial. It is 25-34 μm thick and consists of five to 10 filaments. Cells in the ventral core are rectangular, 7-14 μm long (mean 11 μm � 1.9) and 3-7 μm in diameter (mean 5 μm � 0.9) (n = 18). Cells in the peripheral region are square to rectangular, 6-13 μm long (mean 9 μm � 2) and 6-8 μm in diameter (mean 7 μm � 0.8) (n = 13). Cells of adjacent filaments are laterally connected by fusions. Epithallial and meristematic cells are poorly preserved, hindering a proper description of their morphology. Nevertheless, they appear as rectangular and flattened and not rounded or hemispherical. Cells below the epithallial cells are elongate.
Observed conceptacles bear a single pore canal. The roof and the pore canal of the conceptacles are formed by filaments that run parallel to the conceptacle chamber and the pore canal, suggesting their development from the initials (filaments) peripheral to the fertile area (Fig. 19b). Conceptacles are raised 70-120 μm above the thallial surface and their external diameter is 192-292 μm. Chambers have a central pedestal and are up to 112 μm high and 227 μm in diameter (Fig. 19a).
Remarks. In the identification of this specimen, we encountered similar problems as in Coralliniophycidae sp. 1. Differences are in the arrangement of the ventral core and presence of the single type of conceptacle which provides a larger number of possible bi/tetrasporangial plants.
|
|
Figure 19: Corallinophycidae
sp. 2. a) Two uniporated conceptacles. Conceptacle on the left bears central pedestal (arrow) while conceptacle on the right has flat floor. b) Detail of the conceptacle shows that the roof is formed by filaments running parallel to the chamber (arrow). |
Corallinophycidae sp. 3
(Fig. 20 ![]() )
)
Selected specimens examined. Ježov vrch: thin section 28098.
Material. This species is described based on the specimen of thin section 28098. There are only two occurrences of this species and both are from Ježov vrch. The species is non-geniculate and displays a warty-protuberant growth form (Fig. 20a).
Description. Thallus is pseudoparenchymatous with monomerous construction (Fig. 20b), consisting of a single system of branching filaments which run roughly parallel to the substrate where they form a ventral core of the thallus and then bend upward to develop a peripheral zone. Branching pattern of the vegetative filaments is monopodial. The ventral core is non-coaxial. Cells are rectangular, 11-20 μm long (mean 15 μm � 2.4) and 4-10 μm in diameter (mean 7 μm � 1.1) (n = 18). Cells in the peripheral region are square to rectangular, 6-14 μm long (mean 8 μm � 2.2) and 4-8 μm in diameter (mean 6 μm � 0.9) (n = 39). Cells of adjacent filaments are laterally connected by numerous fusions (Fig. 20c). Epithallial and meristematic cells were not observed.
Conceptacles are uniporate. The orientation of the section precludes proper understanding of the formative mechanism of the conceptacle (Fig. 20d). Only one conceptacle with visible pore canal is present in this specimen. The chamber has a central pedestal and is up to 112 μm high from the floor and 227 μm in diameter.
Remarks. The poorly preserved reproductive anatomy does not allow assignment of this specimen to a known genus.
|
|
Figure 20: Corallinophycidae
sp. 3. a) Warty protuberant
growth form. b) Non-coaxial arrangement of ventral core filaments (arrow). c) Numerous cell fusions (arrows). d) Uniporate conceptacle. Arrow points to its roof. |
Hapalidiales sp.
(Fig. 21 ![]() )
)
Studied specimens examined. Ježov vrch thin section 28097.
Material. The species is described based on the fertile part of the specimen in thin section 28097. Other occurrences are sterile thalli which are compared with this species based on the morphology of the thallus, size of vegetative cells and thin ventral core, all features found to be characteristic of this species. The species is non-geniculate with encrusting-layered growth form and applanate branches (Fig. 21a).
Description. Thallus is pseudoparenchymatous with monomerous construction, consisting of a single system of branched filaments which run parallel to the substrate to form the ventral core and bending upward to form the peripheral region, where they grow perpendicular to the substrate. Branching pattern of the filaments is monopodial and pseudodichotomous. Thalli are 49-85 μm thick. The ventral core consists of two to five filaments and is 7-50 μm thick in its thickest portion. The thallus is often built exclusively by the ventral core. Cells are rectangular or bladed and measure 8-22 μm (mean 16 μm � 3.4) in length and 5-9 μm (mean 7 μm � 1.1) in diameter (n = 26). Cells in the peripheral filaments are rectangular to square, 7-12 μm (mean 9 μm � 1.5) long and 7-9 μm (mean 8 μm � 0.6) in diameter (n = 11). Cells of adjacent filaments are laterally joined by fusions. Epithallial and meristematic cells were not observed.
Reproduction. A single multiporate sporangial conceptacle was observed (Fig. 21b). The margin of the conceptacle is missing, so that it was not possible to measure its diameter correctly. Its whole height protrudes above the surrounding thallial surface. The roof consists of a sunken plate and a peripheral rim. The roof is 22 μm thick and the filaments in the pore plate are up to five-celled. The roof is crossed by cylindrical pore canals which may be lined by five-celled filaments (Fig. 21b). However, the outlines of the cells in the roof and pore-canal-lining filaments is not clear in most cases. In some places the straight pore canals show cells which appear to be specialized, but these spots are bleached and prevent measuring of cell dimensions. The basal cells of both filament types appear slightly elongate. Measurements of the available cell morphology, however, suggest that the cells in the roof and pore-canal filaments are of comparable size. Roof cells are 5-6 μm long and 4-5 μm in diameter (n = 6). Pore-canal-lining cells are 4-6 μm long and 4-5 μm in diameter (n = 7). Gametophytes were not observed.
Remarks. This species can be easily separated from other algae based on 1) its growth form, 2) thallus thickness, 3) features of the ventral core and 4) its sporangial conceptacle roof which bears a distinct pore plate and peripheral rim. Lack of diagnostic features in this species prevents more specific classification within the Order Hapalidiales. This is because 1) epithallial-cell morphology is unknown, 2) length of the meristematic cells is unknown, 3) arrangement of ventral core filaments is visible only rarely, 4) morphology of the cells lining the pore canal lacks adequate measurements and 5) gametophytes are unknown. What we know, however, about this species is that its thallus has monomerous construction that excludes Melobesioideae with dimerous thallus construction (Athanasiadis, 2016) and straight pore canals in multiporate sporangial conceptacles which may exclude some genera, e.g., Melyvonnea and Synarthophyton with conical pore canals and Carlskottspergia with pyriform pore canals. Also, the encrusting non-protuberant growth form that is applanately branched confirms that the species does not belong to the genus Melyvonnea (Athanasiadis & Ballantine, 2014). Unfortunately, only one fertile specimen is known from Ježov vrch and the species is absent from the historical collection. The study of specimens from other collections could shed light on its systematic position within the Order Hapalidiales.
|
|
Figure 21: Hapalidiales
sp. 1. a) Encrusting-layered growth form. Note that the peripheral region is thin or not developed (arrow). Applanate branching is marked with arrowhead. b) Fragment of multiporate sporangial conceptacle with peripheral rim and distinct pore plate. Note the pore canal lining cells (arrows) which are similar to the other roof cells. |
Our research enabled the identification of seventeen coralline algal species belonging to six genera and three orders. These are: Sporolithon lugeonii, Sporolithon nummuliticum, Sporolithon sp. 1, Sporolithon sp. 2 (Order Sporolithales), Lithothamnion camarasae, Lithothamnion cf. corallioides, Lithothamnion prascoi, Lithothamnion cf. ramosissimum, Lithothamnion roveretoi, Lithothamnion sp., Phymatolithon sp., Mesophyllum fructiferum, Mesophyllum cf. engelhartii, Mesophyllum sp. (Order Hapalidiales), Chamberlainium lemoinei, Lithoporella melobesioides and Lithoporella cf. minus (Order Corallinales). Specimens are described based on morphological characteristics which provide suitable criteria for fossil species delimitation (De Queiroz, 2007). Classification at the higher taxonomic ranks was facilitated by the proposed key for Cenozoic coralline algae from the Central Carpathian Basin (Supplement - Appendix: Identification key). Criteria for species delimitation, however, may be different in the biological sciences. Modern biological studies, for examples, are chiefly based on molecular genetics, sometimes supported by morpho-anatomical characters, ecology or biogeography (e.g., Kato et al., 2011; Kato et al., 2013; Athanasiadis & Ballantine, 2014; Adey et al., 2015, 2018; Caragnano et al., 2018; Athanasiadis, 2020). Very often, new coralline algal taxa are exclusively defined based on molecular genetics as they are considered almost indistinguishable on purely morphological grounds (Maneweldt et al., 2017; Caragnano et al., 2018; Nelson et al., 2021). However, some new coralline algal genera have been erected based on morphological studies lacking genetic analyses (see Athanasiadis, 2017, 2019). Thus, the present state of knowledge either avoids delimitation of coralline algal taxa solely on the basis of morphology or provides improved morphological description of many others whose identification is possible. Thanks to phycological research on modern coralline algae, the taxonomy of fossil taxa has advanced significantly, integrating new characters and providing clear and updated definitions for several genera and a large number of species (Braga & Aguirre, 1995; Basso et al., 1996, 1998; Rasser & Piller, 1999; Quaranta et al., 2007; Coletti et al., 2016, 2020; Hrabovsk� et al., 2019). Encrusting coralline algal genera which can, at least for now, be discriminated in the CCPB based on morphological characteristics are:
1) Lithothamnion: monomerous thallus construction, multiporate sporangial conceptacles without weakly calcified walls of filaments consisting of elongate sterile cells, and flared epithallial cells terminating vegetative filaments.
2) Mesophyllum: predominantly coaxial arrangement of the ventral core, filaments in the thalli with simple and unbranched protuberances growing from an encrusting base, filaments with monopodial branching pattern, non-flared epithallial cells above the elongate cells of the meristem, multiporate sporangial conceptacles with specialized pore-canal-lining cells, and pore-canal filaments with usually more than three to five cells where basal cell which is not markedly elongate. Gametophytes are dioecious and carposporangial conceptacles bear distinct central pedestal.
3) Phymatolithon: bears encrusting as well as protuberant growth forms, short meristematic cells and non-flared epithallial cells together with multiporate sporangial conceptacles with pore canals lined by non-specialized cells.
4) Lithoporella: monostromatic thallus with primigenous filaments consisting of palisade cells, numerous cell fusions, uniporate bi/tetrasporangial conceptacles developed from filaments peripheral to and interspersed among sporangial initials and hemispherical epithallial cells.
5) Chamberlainium: mainly dimerous thallus construction without palisade cells in primigenous filaments, bi/tetrasporangial conceptacles less than 300 μm in diameter, roof filaments less than eight-celled, bi/tetrasporangial conceptacles developed from initials peripheral to fertile area, pore-canal filaments protruding into the pore canal and cells of adjacent filaments connected by lateral fusions. Gametophytes are dioecious.
6) Hydrolithon: uniporate conceptacles developed from filaments peripheral to and interspersed among sporangial initials, pore canal lined by filaments which do not protrude into the pore canal, trichocytes not developed in large tightly packed horizontal fields and thallus predominantly dimerous.
7) Sporolithon: sporangial compartments separated by paraphyses and lacking multiple stalk cell and involucre.
The genera discussed above - Lithothamnion, Phymatolithon, Mesophyllum, Lithoporella, Chamberlainium, Hydrolithon and Sporolithon - are common in the bioclastic units of the lower Priabonian of the CCPB and all of them, to some degree, can be discriminated from other known coralline algal genera when diagnostic characteristics are preserved. Since these key morpho-anatomical characters, useful to separate these genera, are in most cases supported by genetic studies on living algae, it may be possible to discriminate other taxa of coralline algae in the fossil record. It may be hoped that further studies will be able to identify, e.g., Corallinapetra (Order Corallinapetrales) by the unique morphology of its multiporate sporangial conceptacles that bear filaments of sterile cells separating sporangia (Jeong et al., 2021), Dawsoniolithon in its possession of large type-2 sporangial conceptacles with more than eight-celled roof filaments in a monomerous thallus with a coaxial ventral core (Caragnano et al., 2018), Floiophycus by its fusions between the meristem cells and trichocytes in the form of megacells (Townsend & Huisman, 2018a), and Exilicrusta by its multiporate sporangial conceptacles, dimerous thallus construction, absence of palisade cells and flared epithallial cells (Chamberlain, 1992), while Melyvonnea and Leptophytum can be identified by the presence of specialized pore-canal-lining cells in multiporate sporangial conceptacles (Athanasiadis & Ballantine, 2014) (Table 1). In light of the above points, the application of accepted and confirmed morphological characteristics in palaeontology is important and enriching in the understanding of these organisms, which played a major role in the Earth's natural history.
Table 1: List of genera of encrusting coralline algae with characters of high fossilization potential.
| Genus | Diagnostic character or combination of characters |
| Corallinapetra | Sporangial conceptacles with weakly calcified filaments of sterile cells, flared epithallial cells |
| Capensia | Sympodial branching in the venral core filaments |
| Melyvonnea | Bifurcating protuberances, up to 5 specialized pore canal cells, basal cell elongate, corposporangial conceptacle without central pedestal, monoecy |
| Phragmope | Secondary aporous roof of sporangial conceptacles |
| Carlskottsbergia | Monostromatic ventral core |
| Crustaphytum | Short meristematic cells and strictly coaxial arrangement of ventral core filaments |
| Rhizolamellia | Coaxial ventral core, elongate meristematic cells, non-specialized pore canal cells, long pore canal of carposporangial conceptacle |
| Synarthrophyton | Mostly non-coaxial, elongate meristematic cells, 1-2 epithallial cell, non-specialized / thinner pore canal cells, carposporangial conceptacle with pedestal |
| Neopolyporolithon | Non-coaxial, elongate meristematic cells, 2-3 epithallial cells, non-specialized pore canal cells, carposporangial conceptacle with flat floor |
| Leptophytum | Monomerous non-coaxial, short meristematic cells, specialized pore canal lining cells, carposporangial conceptacle with flat floor |
| Harveylithon | Monomerous non-coaxial, solitary trichocytes, type 2 conceptacle, not protruding filaments into the pore canal |
| Dawsoniolithon | Coaxial ventral core, large type 2 conceptacle, protruding filaments into the pore canal, meristematic cells, non-stratified peripheral region |
| Floiophycus | Fusion of meristematic cells and thickness of the non-coaxial ventral core, trichocytes in the form of megacells |
| Crusticorallina | Flask-shaped bi/tetrasporangial uniporate conceptacles |
| Parvicellularium | Coaxial ventral core, short meristematic cells, large columellated bi/tetrasporic conceptacles, roof filaments more than 10-celled |
Our observations show, however, that fossil specimens commonly do not bear sufficient morpho-anatomically diagnostic characteristics, making their classification at the generic rank and even at the subfamilial level almost impossible. Diagnostic features are only partially preserved, e.g., in specimens described here as Lithothamnion cf. corallioides which mostly do not have preserved epithallial and meristematic cells. Mesophyllum species may either have no preserved epithallial cells, specialized pore-canal-lining cells, meristematic cells, or gametophytes with carposporangial conceptacles and even the predominantly coaxial ventral core which is considered as diagnostic in emended diagnosis of Mesophyllum s.s. (Athanasiadis & Ballantine, 2014). Accordingly, none of the Mesophyllum specimens described in this work bears all diagnostic features. Also, gametophytes are unknown in numerous other identified fossil species, preventing their proper classification. These arguments justify the questioning of the validity of our classification that places tens of the studied specimens within the group incertae sedis. In light of the above, we agree that the simple classification of fossil specimens based on morpho-anatomical features may lead to errors and that the systematic position of some fossil species (specimens) could be, at least for now, inaccurate (Hind et al., 2016; Nelson et al., 2021). Nevertheless, we tentatively assign fossil specimens to well-described species where they at least preserve the morphological characteristics of fertile specimens.
It is worth noting that palaeoecological studies of coralline-algal-rich limestones is based on quantification of certain genera or subfamilies. As demonstrated palaeoecologically, the previously mentioned approach works well at higher (supraspecific) taxonomic ranks, especially when associated with sedimentological and other palaeontological data (e.g., Braga & Aguirre, 2001; Checconi et al., 2010; Quaranta et al., 2012; Coletti et al., 2018). Recently, fossil coralline algae were investigated in the lower Priabonian limestones at the �trba section (Hrabovsk� et al., 2022). A comparison of �trba and the preliminary results from the Tich� Dolina assemblages (this work) indicates, e.g., that 1) S. nummuliticum, Lithoporella melobesioides and Lp. cf. minus are more frequent and abundant at Tich� Dolina than at �trba, 2) diversified sporolithoids with gametophytes are known from Tich� Dolina only, while only few sporophytes of S. nummuliticum are known at the �trba section, 3) fertile thalli of Chamberlainium are known from Tich� Dolina and are absent from the �trba section, 4) Mesophyllum is rare in Tich� Dolina while dominated at �trba, 5) L. camarasae is more frequent at Tich� Dolina and 6) geniculate species are almost absent from Tich� Dolina (Table 2). The observed differences based on quantification of coralline algae may be related to the palaeoecological settings of these areas, i.e., the presence of upwelling of cooler water as a major factor controlling the benthic assemblage of the �trba section, resulting in the dominance of Mesophyllum over the Lithoporella-Sporolithon association which is dominant at Tich� Dolina (Hrabovsk� et al., 2022).
Table 2: Preliminary comparison of the early Priabonian coralline algal limestones from CCPB.
| �trba | Oravice |
| Sporolithon - Lithoporella association absent | Common Sporolithon - Lithoporella association |
| Few Sporolithon nummuliticum only | Diversified Sporolithon assemblage |
| Mesophyllum dominant assemblages | Mesophyllum rare assemblage |
| Mesophyllum gametophytes | Sporolithon and Mesophyllum gametophytes |
| Few sterile Chamberlainium | Chamberlainium with gametophytes |
| Facies rich with geniculate algae | Geniculate algae almost absent |
| Rare L. camarasae | Frequent L. camarasae |
| Monospecific rhodoliths, hooked structures | Coralline algal framework, multispecific rhodoliths |
| Suspension feeders | Large benthic foraminifers |
Finally, DNA-based studies have enabled the recognition of more fossil genera than listed, e.g., in Braga et al. (1993). At the same time, our knowledge of coralline algal ecology and biogeography has improved also (e.g., Athanasiadis & Ballantine, 2014; Caragnano et al., 2018). Therefore, morphologically defined fossil species and their quantification have great potential in improving and strengthening of palaeoecological reconstructions. However, questions still arise as to whether and how use of new techniques (e.g., computed tomography) or statistics could incorporate new information into palaeontological studies. This point is crucial also in understanding of the evolution of coralline algae and their recent biogeography.
Preliminary results from Tich� Dolina show a rich coralline-algal assemblage with diversified Sporolithales and Hapalidiales accompanied with minor Corallinales. The assemblage consists of 17 non-geniculate species: Sporolithon lugeonii, Sporolithon nummuliticum, Sporolithon sp. 1, Sporolithon sp. 2, Lithothamnion camarasae, Lithothamnion cf. corallioides, Lithothamnion prascoi, Lithothamnion cf. ramosissimum, Lithothamnion roveretoi, Lithothamnion sp., Phymatolithon sp., Mesophyllum fructiferum, Mesophyllum cf. engelhartii, Mesophyllum sp., Chamberlainium lemoinei, Lithoporella melobesioides and Lithoporella cf. minus and four species classified within the group incertae sedis.
Morpho-anatomical characteristics of the coralline algae enabled: 1) recognition of the new combination - Chamberlainium lemoinei (Miranda) comb. nov., formerly classified within the genus Hydrolithon but having roof cells projecting into the pore canal - a feature which is not consistent with the modified definition of the genus Hydrolithon. This species represents the second exclusively fossil record of Chamberlainium which most likely has existed at least since the Bartonian (as Hydrolithon cf. lemoinei Aguirre et al., 2020); 2) identification of a unique coralline alga of the genus Lithothamnion bearing elongate cells and a reduced ventral core which is two- to three-filaments thick.
Research was supported by project VEGA 2/0070/22 and APVV 20-0079. We are very grateful to Juraj �urka (Slovak Academy of Sciences, Bansk� Bystrica) for providing samples collected in the Tich� Dolina valley and to Nat�li� Hud�čkov� (Faculty of Sciences of Comenius University in Bratislava) for providing historical collections of Paleogene coralline algae from Central Carpathian Paleogene Basin. We thank two anonymous reviewers for providing helpful comments as well as significant and well-founded suggestions on the earlier draft of the manuscript. Last but not least, we thank Stephen Carey for his conscientious proofreading.
Adey W.H., Hernandez-Kantun J.J., Johnson G. & Gabrielson P.W. (2015).- DNA sequencing, anatomy, and calcification patterns support a monophyletic, subarctic, carbonate reef-forming Clathromorphum (Hapalidiaceae, Corallinales, Rhodophyta).- Journal of Phycology, Moss Landing - CA, vol. 51, p. 189-203.
Adey W.H., Hernandez-Kantun J.J., Gabrielson P.W., Nash M.C. & Hayerk L.-A.C. (2018).- Phymatolithon (Melobesioideae, Hapalidiales) in the boreal-subarctic transition zone of the North Atlantic. A correlation of plastid DNA markers with morpho-anatomy, ecology, and biogeography.- Smithsonian Institution Scholarly Press, Washington - DC, 90 p.
Adey W.H. & Johansen H.W. (1972).- Morphology and anatomy of Corallinaceae with special reference to Clathromorphum, Mesophyllum, and Neopolyporolithon gen. nov. (Rhodophyceae, Cryptonemiales).- Phycologia, Lawrence - KS, vol. 11, p. 159-180.
Adey W.H., Masaki T. & Akioka H. (1974).- Ezo epiyassoense, a new parasitic genus and species of Corallinaceae (Rhodophyta, Cryptonemiales).- Phycologia, Lawrence - KS, vol. 13, no. 4, p. 329-344.
Adey W.H. & Sperapani C.P. (1971).- The biology of Kvaleya epilaeve, a new parasitic genus and species of Corallinaceae.- Phycologia, Lawrence - KS, vol. 10, p. 29-42.
Aguirre J., Braga J.C. & Piller W.E. (1996).- Reassessment of Palaeothamnium Conti, 1946 (Corallinales, Rhodophyta).- Review of Palaeobotany and Palynology, vol. 94, p. 1-9.
Aguirre J. & Braga J.C. (1998).- Redescription of Lemoine's (1939) types of coralline algal species from Algeria.- Palaeontology, London, vol. 41, p. 489-507.
Aguirre J., Braga J.C. & Bassi D. (2011).- Taxonomic assessment of coralline algal species (Rhodophyta; Corallinales and Sporolithales) described by Pfender, Lemoine, and Miranda from northern Spain type localities.- Annalen des Naturhistorischen Museums in Wien, Series A, Vienna, vol. 113, p. 267-289.
Aguirre J., Braga J.C., Pujalt V., Orue-Etxebarria X., Salazar-Ortiz E., Rinc�n-Mart�nez D., Abad M. & P�rez-Valera F. (2020).- Middle Eocene rhodoliths from tropical and mid-latitude regions.- Diversity, Basel, vol. 12, no. 13, article 117, 20 p.
Aguirre J., Riding R. & Braga J.C. (2000).- Diversity of coralline red algae: Origination and extinction patterns from the Early Cretaceous to the Pleistocene.- Paleobiology, Cambridge - MA, vol. 26, p. 651-667.
Andrusov D. (1937).- R�le des Thallophytes dans la constitution des roches s�dimentaires des Carpathes tch�coslovaques.- Věstn�k Kr�lovsk� Česk� Spolučnosti Nauk, Tř�da Matematicko-Př�rodovědeck�, Praha, vol. 1938, p. 1-32.
Athanasiadis A. (2001).- Lectotypification of Lithophyllum arcticum (Corallinales, Rhodophyta) and a study of its relationships within the Melobesioideae.- Nordic Journal of Botany, vol. 21, p. 93-112.
Athanasiadis A. (2016).- Phycologia Europaea. Rhodophyta. Vol. I.- Published and distributed by the author, ISBN I 978-91-639-1448-5, xxxxviii + 762 p.
Athanasiadis A. (2017).- Capensia fucorum (Esper) gen. et comb. nov. (Mesophyllaceae, Corallinales, Rhodophyta), a hemiparasite on Gelidium from South Africa.- Botanica Marina, Berlin, vol. 60, no. 5, p. 555-565.
Athanasiadis A. (2019).- Carlskottsbergia antarctica (Hooker fil. & Harv.) gen. & comb. nov., with a re-assessment of Synarthrophyton (Mesophyllaceae, Corallinales, Rhodophyta).- Nova Hedwigia, vol. 108, nos. 3-4, p. 291-320.
Athanasiadis A. (2020).- Phragmope discrepans, gen. & comb. nov. (Mesophyllaceae, Corallinales, Rhodophyta), the species known as "Mesophyllum engelhartii" from South Africa.- Marine Biology Research, vol. 16, nos. 6-7, p. 532-549.
Athanasiadis A. & Adey W.H. (2006).- The genus Leptophytum (Melobesioideae, Corallinales, Rhodophyta) of the Pacific coast of North America.- Phycologia, Lawrence - KS, vol. 45, p. 71-115.
Athanasiadis A. & Ballantine D.L. (2014).- The genera Melyvonnea gen. nov. and Mesophyllum s.s. (Melobesioideae, Corallinales, Rhodophyta) particularly from the central Atlantic Ocean.- Nordic Journal of Botany, vol. 32, p. 385-436.
Athanasiadis A., Lebednik P.A. & Adey W.H. (2004).- The genus Mesophyllum (Melobesioideae, Corallinales, Rhodophyta) on the northern Pacific coast of North America.- Phycologia, Lawrence - KS, vol. 43, p. 126-165.
Bahia R.G., Maneveldt G.W., Amado-Filho G.M. & Yoneshigue-Valentin Y. (2015).- New diagnostic characters for the Order Sporolithales (Corallinophycidae, Rhodophyta).- Journal of Phycology, Moss Landing - CA, vol. 51, p. 1137-1146.
Bailey J.C. (1999).- Phylogenetic position of Lithophyllum incrustans and itanoderma pustulatum (Corallinaceae, Rhodophyta) based on 18S rRNA gene sequence analyses, with a revised classification of the Lithophylloideae.- Phycologia, Lawrence - KS, vol. 38, p. 208-216.
Bailey J.C., Gabel J.E. & Freshwater D.W. (2004).- Nuclear 18S rRNA gene sequence analyses indicate that the Mastophoroideae (Corallinaceae, Rhodophyta) is a polyphyletic taxon.- Phycologia, Lawrence - KS, vol. 43, no. 1, p. 3-12.
Bar�th I. & Kov�č M. (1995).- Systematics of gravity-flow deposits in the marginal Paleogene formations between Marku�ovce and Kluknava villages (Horn�d Depression).- Mineralia Slovaca, Geovestn�k, Bratislava, vol. 27, p. 6 [in Slovak].
Basso D., Fravega P. & Vannucci G. (1996).- Fossil and living Corallinaceans related to the Mediterranean endemic species Lithophyllum racemus (Lamarck) Foslie.- Facies, Erlangen, v. 35, p. 275-292.
Basso D., Fravega P. & Vannuci G. (1997).- The taxonomy of Lithothamnium ramosissimum (G�mbel non Reuss) Conti and Lithothamnium operculatum (Conti) Conti (Rhodophyta, Corallinaceae).- Facies, Erlangen, vol. 37, p. 167-182.
Basso D., Fravega P., Piazza M. & Vannucci G. (1998).- Revision and re-documentation of M. Airoldi's species of Mesophyllum from the Tertiary Piedmon Basin (NW Italy).- Rivista Italiana di Paleontoogia e Stratigrafia, Milano, vol. 106, p. 85-94.
Basso D., Caragnano A. & Graziella R. (2014).- Trichocytes in Lithophyllum kotschyanum and Lithophyllum spp. (Corallinales, Rhodophyta) from the NW Indian Ocean.- Journal of Phycology, Moss Landing - CA, vol. 50, p. 711-717.
Basso D., Coletti G., Bracchi V.A. & Yazdi-Moghadam M. (2019).- Lower Oligocene coralline algae of the Uromieh section (Qom Formation, NW Iran) and the oldest record of Titanoderma pustulatum (Corallinophycidae, Rhodophyta).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 125, no. 1, p. 197-218.
Bittner L., Payri C.E., Maneweldt G.W., Couloux A., Cruaud C., Reviers B. de& Le Gall L. (2011).- Evolutionary history of the Corallinales (Corallinophycidae, Rhodophyta) inferred from nuclear, plastidial and mitochondrial genomes.- Molecular Phylogenetics and Evolution, Cambridge - MA, vol. 61, p. 697-713.
Braga J.C, Bosence D.W.J. & Steneck R.S. (1993).- New anatomical characters in fossil coralline algae and their taxonomic implications.- Palaeontology, London, vol. 36, p. 535-547.
Braga J.C. & Aguirre J. (1995).- Taxonomy of fossil coralline algal species: Neogene Lithophylloideae (Rhodophyta, Corallinaceae) from southern Spain.- Review of Palaeobotany and Palynology, vol. 86, p. 265-285.
Braga J.C. & Aguirre J. (2001).- Coralline algal assemblages in upper Neogene reef and temperate carbonates in Southern Spain.- Palæogeography, Palæoclimatology, Palæoecology, vol. 175, p. 27-41.
Buček S., Filo I. & Laurinc D. (2013).- Litofaci�lne členenie borovsk�ho s�vrstvia Liptovskej kotliny a z�padnej časti Popradskej kotliny. In: Hra�ko Ľ. (ed.), Aktualiz�cia geologickej stavby probl�mov�ch �zem� Slovenskej republiky v mierke 1:50 000.- Ministerstvo Životn�ho Prostredia Slovenskej Republiky, �t�tny Geologick� �stav Dion�za �t�ra, Bratislava. Geofond Archive, archive number: 91733-37, p. 50.
Caragnano A., Foetisch A., Maneweldt G.W., Millet L., Liu L-C., Lin S-M., Rodondi G. & Payri C.E. (2018).- Revision of Corallinaceae (Corallinales, Rhodophyta): Recognizing Dawsoniolithon gen. nov., Parvicellularium gen. nov. and Chamberlainoideae subfam. nov. containing Chamberlainium gen. nov. and Pneophyllum.- Journal of Phycology, Moss Landing - CA, vol. 54, p. 391-409.
Caragnano A., Rodondi G., Basso D., Pe�a V., Le Gall L. & Rindi F. (2020).- Circumscription of Lithophyllum racemus (Corallinales, Rhodophyta) from the western Mediterranean Sea reveals the species Lithophyllum pseudoracemus sp. nov.- Phycologia, Lawrence - KS, vol. 59, no. 6, p. 584-597.
Chamberlain Y.M. (1983).- Studies in the Corallinaceae with special reference to Fosliella and Pneophyllum in the British Isles.- Bulletin of the British Museum (Natural History), Botany, vol. 11, no. 4, p. 291-463.
Chamberlain Y.M. (1984).- Spore size and germination in Fosliella, Pneophyllum and Melobesia (Rhodophyta, Corallinaceae).- Phycologia, Lawrence - KS, vol. 23, no. 4, p. 433-442.
Chamberlain Y.M. (1991).- Historical and taxonomic studies in the genus Titanoderma (Rhodophyta, Corallinales) in the British Isles.- Bulletin of the British Museum (Natural History) (series Botany), London, vol. 21, no. 1, p. 1-80.
Chamberlain Y.M. (1992).- Observations on two Melobesioid crustose coralline red algal species from the British Isles: Exilicrusta parva, a new genus and species, and Lithothamnion sonderi Hauck.- British Phycological Journal, vol. 27, p. 185-201.
Checconi A., Bassi D., Carannante G. & Monaco P. (2010).- Re-deposited rhodoliths in the Middle Miocene hemipelagic deposits of Vitulano (Southern Apennines, Italy): Coralline assemblages characterization and related trace fossils.- Sedimentary Geology, vol. 225, p. 50-66.
Chelaru R. & Bucur I.I. (2016).- The taxonomy of Middle Miocene red algae from the G�brova de Sus Formation (Transylvania Basin, Romania).- Carnets Geol., Madrid, vol. 16, no. 11, p. 307-336. DOI : 10.4267/2042/60121
Coletti G., Hrabovsk� J. & Basso D. (2016).- Lithothamnion crispatum: Long-lasting species of non-geniculate coralline algae (Rhodophyta, Hapalidiales).- Carnets Geol., Madrid, vol. 16, no. 3, p. 27-41. DOI: 10.4267/2042/58720
Coletti G., Basso D. & Corselli C. (2018).- Coralline algae as depth indicator in the Sommi�res Basin (early Miocene, southern France).- G�obios, Villeurbanne, vol. 51, p. 15-30.
Coletti G., Hrabovsk� J. & Basso D. (2020).- Chamberlainium pentagonum (Conti) comb. nov. and Spongites fruticulosus (Corallinales, Rhodophyta) in the Miocene carbonates of the western Mediterranean.- Carnets Geol., Madrid, vol. 20, no. 11, p. 223-240. DOI: 10.4267/2042/70837
De Queiroz K. (2007).- Species concepts and species delimitation.- Systematic Biology, vol. 56, no. 6, p. 879-886.
Filipek A., Wysocka A. & Barski M. (2017).- Depositional setting of the Oligocene sequence of the western Carpathians in the Polish Spisz region - a reinterpretation based on integrated palynofacies and sedimentological analyses.- Geological Quarterly, Warsaw, vol. 61, p. 859-876.
Filo I. & Sir�ňov� Z. (1996).- The Tom�ovce Member - a new lithostratigraphic unit of the Subtatric Group.- Geologicke Pr�ce, Spr�vy, Ko�ice, vol. 102, p. 41-49 [in Slovak with English summary].
Filo I. & Sir�ňov� Z. (1998).- Horn�d and Chrasť Member - new regional lithostratigraphic units of the Sub-Tatric Group.- Geologicke Pr�ce, Spr�vy, Ko�ice, vol. 103, p. 35-51 [in Slovak with English summary].
Filo I., Buček S. & Sir�ňov� Z. (2009).- Litofaci�lne členenie borovsk�ho s�vrstvia podtatranskej skupiny (oravsk� paleog�n).- Manuskript - arch�v �G�D�, Bratislava.
Fravega P., Piazza M. & Vannucci G. (1993).- Three new species of coralline algae (genera Lithothamnion and Lithophyllum) from the Tertiary Piedmont Basin.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 98, p. 453-466.
Garecka M. (2005).- Calcareous nannoplankton from the Podhale Flysh (Oligocene-Miocene, Inner Carpathians, Poland).- Studia Geologica Polonica, Warsaw, vol. 124, p. 353-369.
Gedl P. (2000).- Biostratigraphy of the Podhale Paleogene (Inner Carpathians, Poland) in the light of palynological studies. Part II. Summary and systematic descriptions.- Studia Geologica Polonica, Warszaw, vol. 117, p. 155-303.
Ghosh A.K. & Maithy P.K. (1996).- On the present status of coralline red alga Archaeolithothamnium Roth. from India.- The Palaeobotanist, Lucknow, vol. 45, p. 64-70.
Gross P. (2008).- Lithostratigraphy of western Carpathians: Paleogene - Podtatransk� Group.- �t�tny Geologick� �stav Dion�za �t�ra, Bratislava [in Slovak with English summary].
Gross P., K�hler E. et al. (1980).- Geology of Liptovsk� kotlina (depression).- �t�tny Geologick� �stav Dion�za �t�ra, Bratislava, 242 p. [in Slovak with English summary].
Gross P., K�hler E.& Samuel O. (1984).- A new lithostratigraphic division of the Inner-Carpathian Paleogene.- Geologick� Pr�ce, Spr�vy, Ko�ice, vol. 81, p. 113-117 [in Slovak with English summary].
Gross P., K�hler E., Mello J., Ha�ko J., Halouzka R. & Nagy A. (1993).- Geol�gia Južnej a V�chodnej Oravy (Geology of Southern and Eastern Orava), G�D�, Bratislava, 319 p. [in Slovak with English summary].
Harvey A.S., Broadwater S.T., Woelkerling W.J. & Mitrovski P.J. (2003).- Choreonema (Corallinales, Rhodophyta): 18S rDNA phylogeny and resurrection of the Hapalidiaceae for the subfamilies Choreonematoideae, Austrolithoideae, and Melobesioideae.- Journal of Phycology, Moss Landing - CA, vol. 39, p. 988-998.
Harvey A.S. & Woelkerling W.J. (1995).- An account of Austrolithon intumescens gen. et. sp. nov. and Boreolithon van-heurckii (Heydrich) gen. et comb. nov. (Austrolithoideae subfam. nov., Corallinaceae, Rhodophyta).- Phycologia, Lawrence - KS, vol. 34, no. 5, p. 362-382.
Harvey A.S., Woelkerling W.J. & Millar A.J.K. (2009).- The genus Lithophyllum (Lithophylloideae, Corallinaceae, Rhodophyta) in south-eastern Australia, with the description of L. riosmenae, sp. nov.- Australian Systematic Botany, Clayton South - VIC, vol. 22, p. 296-317.
Hind K.H., Gabrielson P.W., Jensen C.P. & Martone P.T. (2016).- Crusticorallina gen. nov., a nongeniculate genus in the subfamily Corallinoideae (Corallinales, Rhodophyta).- Journal of Phycology, Moss Landing - CA, vol. 52, p. 929-941.
Hrabovsk� J. (2019).- Reproductive phases of Miocene algae from central Paratethys and their bearing on systematics.- Acta Palaeontologica Polonica, Warsaw, vol. 40, p. 417-439.
Hrabovsk� J., Basso D.& Coletti G. (2019).- The first identification of fossil Mesophyllum in accordance to the modern taxonomic concepts in coralline algae.- Acta Paleontologica Polonica, Warsaw, vol. 64, p. 897-909.
Hrabovsk� J. & Fordin�l K. (2013).- Paleoecological evaluation of the algal limestones in Stupava-Vrchn� hora locality (Vienna Basin, Slovakia).- Mineralia Slovaca, Ko�ice, vol. 45, p. 11-22.
Hrabovsk� J., Basso D. & Dol�kov� N. (2016).- Diagnostic characters in fossil coralline algae (Corallinophycidae: Rhodophyta) from the Miocene of southern Moravia (Carpathian foredeep, Czech Republic).- Journal of Systematic Palaeontology, London, vol. 14, p. 499-525.
Hrabovsk� J., Starek D., Holcov� K. & Z�gor�ek K. (2022).- Early Priabonian Mesophyllum dominated coralline algal assemblage from the coastal upwelling settings (Central Carpathian Paleogene Basin, Slovakia).- Palaeobiodiversity and Palaeoenvironments, Berlin, vol. 102, p. 1-38.
Janočko J. & Jacko S. (1999).- Marginal and deep-sea deposits of Central Carpathian Paleogene Basin, Spi� Magura region, Slovakia: Implication for basin history.- Slovak Geological Magazine, Ko�ice, vol. 4, p. 281-292.
Jeong S.Y., Nelson W.A., Sutherland J.E., Pe�a V., Le Gall L., Diaz-Pulido G., Won B.Y. & Cho T.O. (2021).- Corallinapetrales and Corallinapetraceae: A new order and family of coralline red algae including Corallinapetra gabrielii comb. nov.- Journal of Phycology, Moss Landing - CA, vol. 57, p. 849-862.
Jesionek M.B., Bahia R.G., Lyra M.B., Le�o L.A.B., Oliviera M.C. & Amado-Filho G.M. (2020).- Newly discovered algae in southeast Brazil: Tectolithon flumiense gen. et sp. nov. and Crustaphytum atlanticum sp. nov. (Hapalidiales, Rhodophyta).- Phycologia, Lawrence - KS, vol. 59, no. 2, 101-115.
Johansen H.W. (1981).- Coralline algae, a first synthesis.- CRC Press, Boca Raton - FL, 249 p.
Kato A. & Baba M. (2019).- Distribution of Lithophyllum kuroshioense sp. nov., Lithophyllum subtile and L. kaiseri (Corallinales, Rhodophyta), but not L. kotschyanum, in the northwestern Pacific Ocean.- Phycologia, Lawrence - KS, vol. 58, no. 6, p. 648-660.
Kato A., Baba M. & Suda S. (2011).- Revision of the Mastophoroideae (Corallinales, Rhodophyta) and polyphyly in nongeniculate species widely distributed on Pacific coral reefs.- Journal of Phycology, Moss Landing - CA, vol. 47, p. 662-672.
Kato A., Baba M. & Suda S. (2013).- Taxonomic circumscription of heterogeneous species Neogoniolithon brassica-florida (Corallinales, Rhodophyta) in Japan.- Phycological Research, vol. 61, p. 15-26.
K�zm�r M., Dunkl I., Frisch W., Kuhlemann J. & Ozsv�rt P. (2003).- The Palaeogene forearc basin of the eastern Alps and western Carpathians: Subduction erosion and basin evolution.- Journal of the Geological Society of London, London, vol. 160, p. 413-428.
Kov�č M., Pla�ienka D., Sot�k J., Vojtko R., Oszczypko N., Less Gy., Ćosović V., F�genschuh B. & Kr�likov� S. (2016).- Paleogene palaeogeography and basin evolution of the western Carpathians, northern Pannonian domain and adjoining areas.- Global and Planetary Change, vol. 140, p. 9-27.
Kulka A. (1985).- Arni sedimentological model in the Tatra Eocene.- Geological Quarterly, Warsaw, vol. 29, no. 1, p. 31-64. URL: https://gq.pgi.gov.pl/article/view/8946/pdf_972
Le Gall L., Payri C.E., Bittner L. & Saunders G.W. (2010).- Multigene phylogenetic analyses support recognition of the Sporolithales ord. nov.- Molecular Phylogenetics and Evolution, vol. 54, p. 302-305.
Lemoine M. (1928).- Un nouveau genre de M�lob�si�es : Mesophyllum.- Bulletin de la Soci�t� Botanique de France, Paris, vol. 75, no. 2, p. 251-254.
Lemoine P. (1934).- Algues calcaires de la famille des Corallinac�es recueillies dans les Carpathes occidentales par M.D. Andrusov.- Věstn�k St�tn�ho Geologick�ho �stavu ČSR, Praha, vol. 26, p. 269-289.
Liu L-C., Lin S-M., Caragnano A. & Payri C. (2018).- Species diversity and molecular phylogeny of non-geniculate coralline algae (Corallinophycidae, Rhodophyta) from Taoyuan algal reefs in northern Taiwan, including Crustaphytum gen. nov. and three new species.- Journal of Applied Phycology, vol. 30, no. 6, p. 3455-3469.
Maneweldt G.W., Gabrielson P.W. & Kangwe J. (2017).- Sporolithon indopacificum sp. nov. (Sporolithales, Rhodophyta) from tropical western Indian and western Pacific oceans: First report, confirmed by DNA sequence data, of a widely distributed species of Sporolithon.- Phytotaxa, Auckland, p. 115-128.
Maneveldt G.W. & Merwe E. van der (2012).- Heydrichia cerasina sp. nov. (Sporolithales, Corallinophycidae, Rhodophyta) from the southernmost tip of Africa.- Phycologia, Lawrence - KS, vol. 51, no. 1, p. 11-21.
Marshalko R. (1970).- The research of sedimentary textures, structures, and palaeocurrent analysis of basal formations (central Western Carpathian Paleogene, N of Spi�sko-gemersk� rudohorie Mts.).- Acta Geologica et Geographica Universitatis Comenianae, Bratislava, vol. 19, p. 129-163.
Mastrorilli V.I. (1968).- Nuovo contributo allo studio delle Corallinacee dell'Oligocene Ligure-Piemontese: I reperti della tavoletta Ponzone.- Atti dell'Istituto di Geologia della Universit� di Genova, vol. 5, no. 2, 153-406.
Mateo-Cid L.E., Mendoza-Gonz�lez A.C. & Gabrielson P.W. (2014).- Neogoniolithon (Corallinales, Rhodophyta) on the Atlantic coast of Mexico, including N. siankanensis sp. nov.- Phytotaxa, Auckland, vol. 190, no. 1, p. 64-93.
Melbourne L.A., Hern�ndez-Kant�n J.J., Russell S. & Brodie J. (2017).- There is more maerl than meets the eye: DNA barcoding reveals a new species in Britain, Lithothamnion erinaceum sp. nov. (Hapalidiales, Rhodophyta).- European Journal of Phycology, London, vol. 52, p. 166-178.
Moussavian E. & Kuss J. (1990).- Typification and status of Lithothamnion aschersoni Schwager, 1883 (Corallinaceae, Rhodophyta) from Paleocene limestones of Egypt. A contribution to the synonymy and priority of the genera Archaeolithothamnium Rothpletz and Sporolithon Heydrich.- Berliner Geowissenschaftliche Abhandlungen, Berlin, v. 120, no. 2, p. 929-942.
Nelson W.A., Neill K.F., Twist B.A. & Southerland J.E. (2021).- A new genus and species in the Sporolithales (Corallinophycidae, Rhodophyta) from northern New Zealand: Roseapetra farriae sp. nov.- Phytotaxa, Auckland, vol. 490, no. 1, p. 35-46.
Nelson W.A., Sutherland J.E., Farr T.J., Hart D.R., Neil K.F., Kim H.J. & Yoon H.S. (2015).- Multi-gene phylogenetic analyses of New Zealand coralline algae: Corallinapetra novaezelandiae gen. et sp. nov. and recognition of the Hapalidiales ord. nov.- Journal of Phycology, Moss Landing - CA, vol. 51, p. 454-468.
Olszewska B.W. & Wieczorek J. (1998).- The Paleogene of the Podhale Basin (Polish Inner Carpathians) - micropaleontological perspective.- Przegled Geologičny, Warsaw, vol. 46, p. 721-728.
Pe�a V., Le Gall L., R�sler A., Payri C.E. & Braga J.C. (2019).- Adeylithon bosencei gen. et sp. nov. (Corallinales, Rhodophyta): A new reef-building genus with anatomical affinities with the fossil Aethesolithon.- Journal of Phycology, Moss Landing - CA, vol. 55, p. 134-145.
Pezzolesi L., Pe�a V., Le Gall L., Gabrielson P.W., Kaleb S., Hughey J.R., Rodondi G., Hernandez-Kantun J.J., Falace A., Basso D., Cerrano C. & Rindi F. (2019).- Mediterranean Lithophyllum stictiforme (Corallinales, Rhodophyta) is a genetically diverse species complex: Implications for species circumscription, biogeography and conservation of coralligenous habitat.- Journal of Phycology, Moss Landing - CA, vol. 55, p. 473-492.
Puckree-Padua C.A., Gabrielson P.W. & Maneveldt G.W. (2021).- DNA sequencing reveals three new species of Chamberlainium (Corallinales, Rhodophyta) from South Africa, all formerly passing under Spongites yendoi.- Botanica Marina, Berlin, vol. 64, no. 1, p. 19-40. DOI: 10.1515/bot-2020-0074
Quaranta F., Tomasetti L., Vannucci G. & Brandano M. (2012).- Coralline algae as environmental indicators: A case study from the Attard member (Chattian, Malta).- Geodiversitas, Paris, vol. 34, no. 1, p. 151-166. URL: https://sciencepress.mnhn.fr/sites/default/files/articles/pdf/g2012n1a9.pdf
Quaranta F., Vannucci G. & Basso D. (2007).- Neogoniolithon contii comb. nov. based on the taxonomic re-assessment of Mastrorilli's original collections from the Oligocene of NW Italy (Tertiary Piedmont Basin).- Rivista Italiana di Paleontolog�a e Stratigrafia, Milano, vol. 113, no. 1, p. 43-55. URL: https://riviste.unimi.it/index.php/RIPS/article/view/6357/6312
Rasser M.W. & Piller W.E. (1999).- Application of neontological taxonomic concepts to Late Eocene coralline algae (Rhodophyta) of the Austrian Molasse Zone.- Journal of Micropalaeontology, London, vol. 18, p. 67-80.
Richards J.L., Bahia R.G., Jesionek M.B. & Fredericq S. (2019).- Sporolithon amadoi sp. nov. (Sporolithales, Rhodophyta), a new rhodolith-forming non-geniculate coralline alga from offshore the northwestern Gulf of Mexico and Brazil.- Phytotaxa, Auckland, vol. 423, no. 2, p. 049-067.
Richards J.L., Gabrielson P.W. & Fredericq S. (2014).- New insights into the Genus Lithophyllum (Lithophylloideae, Corallinaceae, Corallinales) from deepwater rhodolith beds offshore the NW Gulf of Mexico.- Phytotaxa, Auckland, vol. 190, no. 1, p. 162-175.
Richards J.L., Vieira-Pinto T., Schmidt W.E., Sauvage T., Gabrielson P.W., Oliveira M.C. & Fredericq S. (2016).- Molecular and morphological diversity of Lithothamnion spp. (Hapalidiales, Rhodophyta) from deepwater rhodolith beds in the northwestern Gulf of Mexico.- Phytotaxa, Auckland, vol. 278, no. 2, p. 81-114.
R�sler A., Perfectti F., Pe�a V. & Braga J.C. (2016).- Phylogenetic relationships of Corallinaceae (Corallinales, Rhodophyta): Taxonomic implications for reef-building corallines.- Journal of Phycology, Moss Landing - CA, vol. 52, p. 412-431.
R�sler A., Pretković V., Novak V., Renema W. & Braga J.C. (2015).- Coralline algae from the Miocene Mahakam delta (east Kalimantan, southeast Asia).- Palaios, Lawrence - KS, vol. 30, p. 83-93.
Samuel O. & Fus�n O. (1992).- Reconstruction of subsidence and sedimentation of Central Carpathian Paleogene.- Z�padn� Karpaty, S�ria Geol�gia, Bratislava, vol. 16, p. 7-46 [in Slovak with English summary].
Schalekov� A. (1962).- Fytog�nne v�pence mezozoika a terci�ru Slovenska.- Dissertation, Comenius University, Bratislava.
Schalekov� A. (1964).- New informations on the calcareous algae in the bioherm limestones of the Paleocene-Eocene in the western and central Slovakia.- Geologica Carpathica, Bratislava, vol. 15, p. 57-73.
Schneider C.W. & Wynne M.J. (2019).- Fourth addendum to the synoptic review of red algal genera.- Botanica Marina, Berlin, vol. 62, no. 4, p. 355-367.
Sciuto K., Moschin E., Alongi G., Cecchetto M., Schiaparelli S., Caragnano A., Rindi F. & Moro I. (2021).- Tethysphytum antarcticum gen. et sp. nov. (Hapalidiales, Rhodophyta), a new non-geniculate coralline alga from Terra Nova Bay (Ross Sea, Antarctica): Morpho-anatomical characterization and molecular phylogeny.- European Journal of Phycology, vol. 56, no. 4, p. 416-427.
Sliva Ľ. (2005).- Sedimentary facies of the Central Carpathian Paleogene Basin from Spi�sk� Magura.- PhD Thesis, Manuscript, Department of Geology and Paleontology - Faculty of Natural Sciences CU, Bratislava, 137 p. [in Slovak]
Sot�k J. (1998).- Sequence stratigraphy approach to the Central Carpathian Paleogene (Eastern Slovakia): Eustasy and tectonics as controls of deep-sea fan deposition. Slovak Geological Magazine, Ko�ice, vol. 4, p. 185-190
Sot�k J., Pereszlenyi M., Marschalko R., Milička J. & Starek D. (2001).- Sedimentology and hydrocarbon habitat of the submarine-fan deposits of the Central Carpathian Paleogene Basin (NE Slovakia).- Marine and Petroleum Geology, vol. 18, p. 87-114.
Starek D. & Fuksi T. (2017a).- Distal turbidite fan/lobe succession of the Late Oligocene Zuberec Fm. - architecture and hierarchy (central Western Carpathians, Orava-Podhale basin).- Open Geosciences, Berlin, vol. 9, p. 385-406.
Starek D. & Fuksi T. (2017b).- Statistical analysis as a tool for identification of depositional palaeoenvironments in deep-sea fans (Palaeogene formations, central Western Carpathians, north Slovakia).- Acta Geologica Slovaca, Bratislava, vol. 9, p. 149-162.
Starek D., Andreyeva-Grigorovich A.S. & Sot�k J. (2000).- Suprafan deposits of the Biely Potok Formation in the Orava region: Sedimentary facies and nannoplankton distribution.- Slovak Geological Magazine, Ko�ice, vol. 6, p. 188-190.
Starek D., Sliva L. & Vojtko R. (2004).- The channel-levee sedimentary facies and their synsedimentary deformation: A case study from Huty Formation of the Podtatransk�skupina Group (Western Carpathians).- Slovak Geological Magazine, Ko�ice, vol. 10, p. 177-182.
Starek D., Sot�k J., Jablonsk� J.& Marschalko R. (2013).- Large-volume gravity flow deposits in the Central Carpathian Paleogene Basin (Orava region, Slovakia): Evidence for hyperpycnal river discharge in deep-sea fans.- Geologica Carpathica, Bratislava, vol. 64, p. 305-326.
Starek D., �imo V., Antol�kov� S. & Fuksi T. (2019).- Turbidite sedimentology, biostratigraphy and paleoecology: A case study from the Oligocene Zuberec Fm. (Liptov Basin, central Western Carpathians).- Geologica Carpathica, Bratislava, vol. 70, no. 4, p. 279-297.
�urka J., Sliva Ľ. & Sot�k J. (2012).- Facial development of the Borov� Formation in the area of Biely Potok at the town of Ru�omberok and at Komjatn� village (Western Carpathians, Slovakia).- Mineralia Slovaca, Ko�ice, vol. 44, p. 267-278.
Teichert S., Woelkerling W.J. & Munnecke A. (2019).- Coralline red algae from the Silurian of Gotland indicate that the Order Corallinales (Corallinophycidae, Rhodophyta) is much older than previously thought.- Palaeontology, London, vol. 4, p. 599-613.
Townsend R.A. (1979).- Synarthrophyton, a new genus of Corallinaceae (Cryptonemiales, Rhodophyta) from the southern hemisphere.- Journal of Phycology, Moss Landing - CA, vol. 15, p. 251-259.
Townsend R.A., Chamberlain Y.M. & Keats D.W. (1994).- Heydrichia woelkerlingii gen. et sp. nov., a newly discovered non-geniculate red alga (Corallinales, Rhodophyta) from Cape Province, South Africa.- Phycologia, Lawrence - KS, vol. 33, no. 3, p. 177-186.
Townsend R.A. & Huisman J.M. (2004).- Epulo multipedes gen. et sp. nov. (Corallinaceae, Rhodophyta), a coralline parasite from Australia.- Phycologia, Lawrence - KS, vol. 43, no. 3, p. 288-295.
Townsend R.A. & Huisman J.M. (2018a).- Coralline algae. In: Huisman J.M. (ed.).- Algae of Australia. Marine Benthic Algae of north-western Australia, 2. Red Algae. Canberra, ABRS & Melbourne, CSIRO Publishing, ISBN 978-1-486-30954-2.
Townsend R.A. & Huisman J.M. (2018b).- Transfer of Oztralia gabrielsonii and Oztralia orbiculata to Dawsoniolithon (Corallinales, Rhodophyta).- Notulae Algarum, vol. 78, p. 1-2.
Townsend R.A., Woelkerling W.J., Harvey A.S. & Borowitzka M. (1995).- An account of the red algal genus Sporolithon (Sporolithaceae, Corallinales) in southern Australia.- Australian Systematic Botany, Clayton South - VIC, vol. 8, p. 85-121.
Vannucci G., Piazza M., Fravega P. & Basso D. (2000).- Revision and re-documentation of M. Airoldi's species of Archaeolithothamnium from the Tertiary Piedmont Basin (NW Italy).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 106, p. 191-202.
Verheij E. (1993).- The genus Sporolithon (Sporolithaceae fam. nov., Coralliales, Rhodophyta) from the Spermonde Archipelao, Indonesia.- Phycologia, Lawrence - KS, vol. 32, p. 184-196.
Westwalewicz-Mogilska E. (1986).- A new look at the genesis of the Podhale Flysh.- Przegląd Geologiczny, Warsaw, vol. 34, p. 690-698 [in Polish with English summary].
Wieczorek J. (1989).- The Hecho model for Podhale flysh?.- Przegląd Geologiczny, Warsaw, vol. 37, p. 419-422 [in Polish].
Woelkerling W.J. (1988).- The coralline red algae: An analysis of the genera and subfamilies of nongeniculate Corallinaceae.- British Museum (Natural History) and Oxford University Press, London and Oxford, ISBN 0-19-854249-6, xi + 268 p.
Woelkerling W.J., Bassi D. & Iryu Y. (2012).- Hydrolithon braganum sp. nov. (Corallinaceae, Rhodophyta), the first known exclusively fossil semi-endophytic coralline red alga.- Phycologia, Lawrence - KS, vol. 51, no. 6, p. 604-611.
Woelkerling W.J. & Ducker S.C. (1987).- Lesueuria minderiana gen. et sp. nov. (Corallinaceae, Rhodophyta) from southern and western Australia.- Phycologia, Lawrence - KS, vol. 26, no. 2, p. 192-204.
Woelkerling W.J., Granier B. & Dias-Brito D. (2014).- Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), a 100 million year old fossil coralline red alga from north-eastern Brazil, a new Hauterivian record of Sporolithon from Switzerland.- Carnets Geol., Madrid, vol. 14, no. 7, p. 139-158. DOI: 10.4267/2042/53736
Woelkerling W.J. & Harvey A. (1992).- Mesophyllum incisum (Corallinaceae, Rhodophyta) in southern Australia: Implications for generic and specific delimitation in the Melobesioideae.- British Phycological Journal, vol. 27, p. 381-399.
Woelkerling W.J. & Harvey A. (1993).- An account of southern Australian species of Mesophyllum (Corallinaceae, Rhodophyta).- Australian Systematic Botany, Clayton South - VIC, vol. 6, p. 571-637.
Woelkerling W.J., Irvine L.M. & Harvey A.S. (1993).- Growth-forms in non-geniculate coralline red algae.- Australian Systematic Botany, Clayton South - VIC, vol. 6, p. 277-293.
Supplement
1: Lithothamnion cf. corallioides |
|
Supplement
2: Lithothamnion cf. ramosissimum |
|
Supplement
3: Mesophyllum cf. engelhartii |
|
Supplement
4: Mesophyllum sp. |
|
| Tetrasporangia borne in sporangial compartments, sporangia cruciately divided | ► Order Sporolithales [2] |
| Bi/tetrasporangia borne in multiporate conceptacles, sporangia zonately divided and separated by weakly calcified filaments of sterile cells | ► Order Corallinapetrales |
| Bi/tetrasporangia borne in uniporate conceptacles, sporangia zonately divided | ► Order Corallinales |
| Bi/tetrasporangia borne in multiporate conceptacles, sporangia zonately divided but not separated by weakly calcified filaments of sterile cells | ► Order Hapalidiales |
Sporolithales with tetrasporangia which are either enclosed in modified filaments (involucre) to form sporangial complexes or sporangia are not enclosed by modified filaments and sporangial complexes are absent ► Family SPOROLITHACEAE
- Non-geniculate thallus with monomerous non-coaxial ventral core, branching pattern of vegetative filaments is monopodial to pseudodichotomous, epithallial cells flared, meristematic cells elongate or short, cells of adjacent filaments are laterally joined with secondary cell fusions and secondary pit connections, sporangial complexes consisting of one or up to several stalk cells and involucre embedding the spore is present ► Heydrichia and ► Roseapetra [3]
- Non-geniculate thallus with monomerous construction which show both, coaxial and non-coaxial organization of ventral core filaments or with dimerous construction, palisade cells unknown, branching pattern of vegetative filaments is monopodial to pseudodichotomous, epithallial cells flared, meristematic cells elongate or short, cells of adjacent filaments are laterally joined with secondary cell fusions and secondary pit connections, sporangial complexes absent, chambers are separated by paraphyses, each chamber produces single spore borne on a single stalk cell ► Sporolithon [4]
Sporangia are borne in multiporate conceptacles and undergo zonate division. Weakly calcified groups of sterile filaments composed of elongate cells are interspersed among developing sporangia. Pore canals blocked by apical plugs. Cells of adjacent filaments joined with fusion and lacking secondary pit connections ► Family CORALLINAPETRACEAE
- Non-geniculate, monomerous thallus with non-coaxial ventral core, meristematic cells elongate, epithallial cells are flared ► Corallinapetra
1) Corallinales with predominantly geniculate thalli which lack stratification ► Family CORALLINACEAE [6]
Specimens mostly geniculate with encrusting holdfast. Cells of adjacent filaments connected by cell fusions, secondary pit connections absent. Development of bi/tetrasporangial conceptacles from filaments surrounding fertile area. In geniculate forms, branches are multiaxial, genicula with single tier of non-calcified cells, intergenicula with 1-40 tiers of cells ► Subfamily CORALLINOIDEAE
- Non-geniculate coralline algae, sunken uniporate bi/tetrasporangial conceptacles flask-shaped, cells of adjacent filaments joined with fusions, lacking secondary pit connections, epithallial cells in one or two layers, flared or unflared, ventral core of basal filaments occupies 50% or more of thallial thickness, elongate meristematic cells, trichocytes absent ► Crusticorallina
2) Corallinales with predominantly non-geniculate thalli, cells of adjacent filaments joined with fusion ► Family SPONGITIDACEAE
2.1) Thallial construction is dimerous, primigenous filaments are composed of palisade cells throughout thallus. Secondary pit connections absent or rare, cell fusions between cells of adjacent filaments are common. Development of bi/tetrasporangial conceptacles from filaments surrounding fertile area or developed from both, peripheral to and interspersed among sporangial initials ► Subfamily MASTOPHOROIDEAE [7]
- Non-geniculate coralline algae, bi/tetrasporangial conceptacles are developed from filaments peripheral to sporangial initials. Conceptacles project strongly above thallial surface ► Mastophora [8]
- Non-geniculate coralline algae, bi/tetrasporangial conceptacles developed from filaments peripheral to and interspersed among sporangial initials ► Lithoporella
2.2) Thallial construction is monomerous. Development of bi/tetrasporangial conceptacles from filaments surrounding fertile area. Filaments lining pore canal are oriented parallel to oblique to thallial surface and may protrude laterally into pore canal ► Subfamily NEOGONIOLITHOIDEAE
- Non-geniculate coralline algae, ventral core filaments arranged coaxially. Trichocytes solitary or in horizontal fields or vertical stacks ► Neogoniolithon [9]
- Non-geniculate coralline algae, thallus with monomerous construction and non-coaxial organization or dimerous construction with non-palisade cells in primigenous filaments. Trichocytes when present, occur singly or in horizontal field. Conceptacle roof filaments are less than eight-celled and chambers are more than 300 μm in diameter. Bi/tetrasporangia borne peripherally around central columella ► Spongites [10]
2.3) Thallial construction dimerous or monomerous but not both. Dimerous thallus without palisade cells. In tangential section may appear in some species palisade like. Some but not all cells of adjacent filaments are joined with fusions, secondary pit connections absent. Trichocytes absent or present. When present, trichocytes are single or paired, always terminal and never intercalary in peripheral filaments. Gametangial plants dioecious. Bi/tetrasporangial conceptacles formed either by filaments surrounding the fertile area or from filaments surrounding and interspersed among the sporangial initials but not both on the same specimen ► Subfamily CHAMBERLAINOIDEAE
- Non-geniculate coralline algae, epithallial cells mostly in single layer, sometimes multi-layered, cells vary in shape from domed to flattened but not flared. Sub-epithallial initials vary from rectangular to ovoid. Bi/tetrasporangial conceptacles uniporate and raised above the surrounding thallial surface. Central columella present or absent. Conceptacle roof filaments are less than 8-celled and formed from filaments peripheral to developing sporangial initials. Gametangial plant dioecious ► Chamberlainium [11]
- Non-geniculate coralline algae, thallial construction primarily dimerous with basal layer comprised of non-palisade cells. Epithallial cells form in one layer, cells are lenticular (short and wide). Subepithallial initials generally rectangular. Cells of adjacent basal and erect filaments joined with fusions, secondary pit connection absent or rare. Trichocytes present or absent, when present single or in pairs. Bi/tetrasporangial conceptacles raised above surrounding thallial surface. Chambers less than 350 μm diameter. Columella present or absent. Conceptacle roof consists of less than 8 celled filaments and formed from filaments both, peripheral to or interspersed among sporangial initials. Conceptacle pore canal lined by cells oriented more or less parallel to thallial surface and protruding into canal as papillae that sometimes also form distinct corona that projects above pore opening. Gametangial plant monoecious ► Pneophyllum [11]
2.4) Thallial internal organization is primarily dimerous, but monomerous can occur as well. Outline of cell filaments entirely lost in large portions due to pervasive and extensive cell fusions. Trichocytes present or absent but not in large tightly packed horizontal fields. Bi/tetrasporangial conceptacles roofs are formed by filaments surrounding and interspersed among sporangial initials. Pore canals of sporangial conceptacles are lined by a ring of elongate cells that do not protrude into pore canal and are oriented more or less perpendicular to roof surface ► Subfamily HYDROLITHOIDEAE [12]
- Non-geniculate coralline algae with predominantly monomerous thallial construction. Trichocytes disposed in large postulate horizontal fields, although they are not tightly packed and do not become buried in the thallus. Presence of layers of large polygonal cells, which resulted from extensive lateral fusions of cells in peripheral filaments interspersed among layers of vegetative cells ► Adeylithon [13]
- Non-geniculate coralline algae with thallus construction predominantly dimerous. Cells of adjacent filaments sometimes joined by fusion but secondary pit connections absent. If trichocytes occur they are either terminal or intercalary. Peripheral filaments lacking. No layers of large polygonal cells interspersed among layers of vegetative cells ► Fosliella [14]
- Thallial construction predominantly dimerous. Trichocytes present but not terminal. Layers of large polygonal cells interspersed among layers of vegetative cells absent ► Hydrolithon
2.5) Thallus either geniculate or non-geniculate. Geniculate specimens with conspicuous mucilaginous caps and genicula of many-celled untiered filaments (Metagoniolithon). Encrusting specimens are monomerous, lateral cell fusions present, secondary pit connections absent, trichocytes occur singly or in horizontal fields. Bi/tetrasporangial conceptacles uniporate with roofs and pore canal formed by filaments peripheral to fertile area and interspersed among sporangial initials ► Subfamily METAGONIOLITHOIDEAE [11]
- Thallial construction monomerous with predominantly coaxial ventral core filaments. Epithallial cells in one layer, cells ovoid to elliptical. Subepithallial initials vary from isodiametric to drop-like. Cells of adjacent filaments joined by fusions, secondary pit connections absent. Trichocytes abundant and occur singly or paired. Bi/tetrasporic conceptacles uniporate and raised above thallial surface. Bi/tetrasporangial chambers elliptical, mostly larger than 300 μm in diameter with central columella. Conceptacle roof consisting of less than eight-celled filaments and formed from filaments peripheral to and interspersed amongst developing sporangial initials. Conceptacle pore canal lined by cells oriented more or less parallel to the thallial surface, protruding into canal without forming papillae ► Dawsoniolithon [15]
- Thallus monomerous with plumose ventral core. Trichocytes usually occur singly and may became buried in thallus. Cells lining pore canal of bi/tetrasporangial conceptacles are oriented perpendicular to thallial surface and do not protrude into pore canal ► Harveylithon [12]
- Thallus dimerous. Trichocytes usually occur in tightly packed horizontal fields. Cells lining pore canal of bi/tetrasporangial conceptacles perpendicular to thallial surface and not protrude into the pore canal ► Porolithon [12]
2.6) Uniporate bi/tetrasporangial conceptacles, type of formation unknown, thallus with megacytes, cells of meristem occasionally to frequently fusing prior to production of peripheral filaments of cell ► Subfamily FLOIOPHYCOIDEAE [16]
- Non-geniculate coralline algae, ventral core filaments non-coaxial and less than 20% of thallial thickness ► Floiophycus
2.7) Family SPONGITIDACEAE, incertae sedis [17]
- Non-geniculate, monomerous with coaxial ventral core, single layer of flattened epithelial cells, subepithallial cells shorter than cells immediately subtending them, cells of adjacent filaments joined with fusion, secondary pit connections absent. If trichocytes occur they are either solitary or arranged in vertical rows. Tetrasporangial conceptacles are large and raised above thallial surface and have poorly developed columella. Pore canal lined by cells oriented more or less perpendicular to thallial surface and protruding into the pore canal without formation of papillae. Gametophytes seemingly dioecious ► Parvicellularium
3) Corallinales with non-geniculate thalli, cells of adjacent filaments joined by secondary pit-connections ► Family LITHOPHYLLACEAE [18]
Roofs of bi/tetrasporangial conceptacles developed from filaments peripheral to and interspersed among sporangial initials. Conceptacles uniporate, tetrasporangia zonately divided and apical plug absent. Cell fusion absent and secondary pit connections present in cells of adjacent filaments ► Subfamily LITHOPHYLLOIDEAE
- Non-geniculate coralline algae, primigenous filaments mostly non-palisade, solitary trichocytes if any ► Lithophyllum
- Non-geniculate coralline algae, primigenous filaments elongate and palisade, solitary trichocytes if any ► Titanoderma
- Non-geniculate coralline algae, parasitic, haustoria present, thin epithallium, thallus dimerous ► Ezo
1) Ventral core reduced, usually monostromatic or composed of few filaments ► Family HAPALIDIACEAE [19]
1.1) Specimens with bi/tetrasporangial conceptacles bearing multiporate acellular plates ► Subfamily CHOREONEMATOIDEAE
- Non-geniculate, parasitic, secondary cell connections absent, multiporate plate acellular at maturity consisting of calcium carbonate matrix, parasitic ► Choreonema [20]
1.2) Endophytic, diffuse, partly pseudoparenchymatous and monomerous thallus. Secondary cell-connections absent. Bi/tetrasporangial conceptacles with multiporate cellular plates ► Subfamily AUSTROLITHOIDEAE [21]
- Non-geniculate, haustoria unknown, roof of multiporate conceptacles consists of two-celled filaments, carpogonial disc consist of seven cells ► Austrolithon
- Non-geniculate, haustoria present, roof of multiporate conceptacles consists of three-celled filaments, carpogonial disc consists of four to six cells ► Epulo
1.3) Thallial construction dimerous. Bi/tetrasporangial conceptacles with multiporate cellular plates, sporangia lacking separation by filaments consisting of sterile cells ► Subfamily MELOBESIOIDEAE [22]
- Non-geniculate, endophytic, secondary cell connections absent ► Boreolithon
- Non-geniculate, cell fusion between adjacent cells present, epithallial cells are non-flared ► Melobesia
- Non-geniculate, cell fusion between adjacent cells present, subepithallial initials variable in length, epithallial cells with flared outer cell walls ► Exilicrusta [23]
2) Ventral core of basal filaments is polystromatic. Carposporangia borne from periphery of central fertile zone, spermatangia predominantly simple ► Family MESOPHYLLUMACEAE [24]
3) Ventral core of basal filaments is polystromatic. Carposporangia borne across fertile zone, spermatangia predominantly dendroid ► Family LITHOTHAMNIACEAE [25]
4) Order HAPALIDIALES incertae sedis [26]
[1] Most recent study of Jeong et al. (2021) recognizes five orders within Corallinophycidae - Rhodogorgonales, Sporolithales, Corallinopetrales, Hapalidiales and Corallinales - based on the bi/tetrasporic conceptacle types and division patterns of tetraspores, including four orders (excluding Rhodogorgonales) of calcified encrusting corallines. On the other hand, Athanasiadis (2016) divided encrusting calcified coralline algae into just two orders based on the division patterns of tetraspores - Sporolithales and Corallinales.
[2] Bahia et al. (2015) refined the diagnosis of the Order Sporolithales with references to characteristics of the carposporophyte. According to Bahia et al. (2015) Sporolithales includes specimens bearing mature carposporangial conceptacles which lack a central fusion cell and having one to two-celled filaments that are distributed across the conceptacle chamber floor and walls. Central pedestal (or columella) in carposporangial conceptacles is as yet unknown.
[3] According to Nelson et al. (2021) two genera - Roseapetra and Heydrichia share common characteristics and cannot be separated based on their morphology. Both, monoecy and dioecy is known in Heydrichia gametophytes (Townsend et al., 1994; Maneveldt & Merwe, 2012) but this information lack for Roseapetra (Nelson et al., 2021). Fossil species - Heydrichia poignantii bi/tetrasporophyte is described in Woelkerling et al. (2014).
[4] Sporolithon is single genus from family Sporolithaceae which can be well-discriminated based on sporangia developed on single stalk cell and not embedded in involucre (Vannucci et al., 2000; Nelson et al., 2021). Pseudodichotomous branching pattern of vegetative filaments was figured in the thallus of Sporolithon amadoi (Richards et al., 2019). This information lack in the description for most of the members of the genera Sporolithon, Heydrichia and Roseapetra. However, its presence can be confirmed based on the figured species of Heydrichia (e.g., Maneveldt & Merwe, 2012, Figs. 8-9, p. 14).
[5] Recently monotypic order with single genus Corallinapetra within the family Corallinapetraceae. Jeong et al. (2021) described the order as non-geniculate. However, this character is most likely diagnostic at the genus rank (e.g., Hind et al., 2016; R�sler et al., 2016). Therefore we use it in the genus description. Epithallial cell morphology was described in the type species but was not mentioned in the genus description (Jeong et al., 2021). This character is known in all members of the Order Sporolithales but in the certain genera of the Order Hapalidiales. Thus, flared morphology of epithallial cells was placed in the genus description in this study. For further information see Jeong et al. (2021).
[6] According e.g. Townsend and Huisman (2018a) the Order Corallinales includes six families: Corallinaceae, Hydrolithaceae, Lithophyllaceae, Mastophoraceae, Porolithaceae and Spongitaceae. Thereof, Corallinaceae with single subfamily Corallinoideae includes only geniculate species. Townsend and Huisman (2018a) classify the given families based on the type of the roof and pore canal formation of the sporangial conceptacles as well as presence/absence of genicula. Such classification however appears useless (see Caragnano et al., 2018; Hind et al., 2016). Recently established genus Crusticorallina represents single encrusting non-geniculate coralline algal genus within the subfamily Corallinoideae (Hind et al., 2016) and subfamily Chamberlainoideae includes genera with both types of sporangial conceptacles (Type 1 and Type 2 according to Johansen, 1981) (Caragnano et al., 2018). By contrast, Athanasiadis (2016) included within the family Spongitidaceae all coralline algal subfamilies with uniporate sporangial conceptacles that are predominantly non-geniculate while predominantly geniculate taxa which bear non-stratified peripheral filaments he placed within the family Corallinaceae. Flask-shaped bi/tetrasporic conceptacles seems to be unique for Crusticorallina.
[7] Mastophoroid coralline algae were recently revised in many publications including e.g. Kato et al. (2011) or R�sler et al. (2016). Subfamily was considered polyphyletic and study revealed new subfamilies (R�sler et al., 2016). Therefore the distributional model of coralline algae (Aguirre et al., 2000) including four subfamilies - Sporolithoideae, Melobesioideae, Mastophoroideae and Lithophylloideae should be considered in paleoclimatological interpretations with caution. Subfamily recently include three genera Lithoporella, Mastophora and Lesueuria. Thereof, Lesueuria is parasite with haustoria penetrating the host cells and pseudoparenchymatous to filamentous thallus. Genus had been assessed to Mastophoroideae by Woelkerling and Ducker (1987) but was not considered in most recent works on coralline algal systematic (e.g., R�sler et al., 2016).
[8] Mastophora and Lithoporella share common characteristics as are monostromatic or thin thallus with dimerous internal construction and non-flared epithallial cells. However, they are separated by the pattern of the bi/tetrasporic conceptacle roof formation. Gametophytes of Lithoporella are more difficult to recognize to palaeontologists because their roof formation is same as in bi/tetrasporangial conceptacles of Mastophora. Lithoporella DNA sequence study lack yet (R�sler et al., 2016; Caragnano et al., 2018).
[9] For detail discussion on the status of Neogoniolithon see R�sler et al. (2016). Fossil Neogoniolithon contii documented in complete life cycle is described in Quaranta et al. (2007). Only species which classified within the genus Neogoniolithon can bear combination of following characteristics: trichocytes arranged in vertical stacks, coaxial arrangement of the ventral core filaments and the type 1 of the bi/tetrasporic conceptacles (Kato et al., 2013; Mateo-Cid et al., 2014; Caragnano et al., 2018). However, lacking of trichocytes or their arrangement in vertical stacks would hamper classification based on the morphology. Nevertheless, trichocytes in vertical stacks can be preserved in some fossil Neogoniolithon (R�sler et al., 2015).
[10] Further details in Caragnano et al. (2018) and R�sler et al. (2016). According to most recent knowledge, Spongites may bears monomerous as well as dimerous thallus. Thereof, monomerous is dominant. Therefore, Spongites is similar with Chamberlainium. However, two genera are separated by smaller conceptacles and less than 8-celled roof filaments of bi/tetrasporangial conceptacles in Chamberlainium (Caragnano et al., 2018). Moreover, Spongites bears trichocytes arranged singly or in horizontal fields while Chamberlainium has trichocytes developed singly or in pairs (Caragnano et al., 2018). First fossil species C. pentagonum was described in Coletti et al. (2020).
[11] Chamberlainium and Pneophyllum are recently placed in the subfamily Chamberlainoideae. They differ in the pattern of the roof and the pore canal formation of the bi/tetrasporangial conceptacles (Caragnano et al., 2018). Similar occurs in Mastophoroideae, i.e., genera Mastophora and Lithoporella.
[12] Subfamilies Hydrolithoideae and Metagoniolithoideae were designated in Kato et al. (2011). Thereof the Metagoniolithoideae was emended in R�sler et al. (2016). Based on their research the Metagoniolithoideae has priority over the Porolithoideae and includes two genera Porolithon and Metagoniolithon (R�sler et al. 2016). Caragnano et al. (2018) established the new genus Dawsoniolithon based on the examination of the type species of Hydrolithon conicum, and placed it in to the subfamily Metagoniolithoideae. There are yet three similar genera in the Hydrolithoideae and Metagoniolithoideae - Hydrolithon, Harveylithon and Porolithon. These genera are separated by presence of trichocytes in large tightly packed horizontal fields found yet in Porolithon genus only. Primarily dimerous thallus construction in Hydrolithon and monomerous in Harveylithon R�sler et al. (2016). None of these genera bear pore canal of bi/tetrasporangial conceptacles lined by filaments protruding into it.
[13] Adeylithon was recently designated in Pe�a et al. (2019). Although authors did not find any morphological character which would separate extant Adeylithon from extinct Aethesolithon, absence of DNA have provided argument which support validity of both (Pe�a et al., 2019). Decision is strongly subjective because absence of DNA may serve as argument which support conspecifity of the two genera as well with nomenclature priority of extant taxa.
[14] Description of the genus can be found in Chamberlain (1983, 1984) or Woelkerling (1988). More recent informations on the status of the genus is provided in Athanasiadis (2016). Recently, the genera Fosliella and Hydrolithon cannot be well-separated in fossil coralline algae (Braga et al., 1993). However, the roof and the pore canal formation of bi/tetrasporangial conceptacles is unknown in the genus Fosliella (Woelkerling, 1988). Athanasiadis (2016) suggested that the peripheral filaments in the genus Fosliela lack (with the exception of single species) and the trichocytes are terminal.
[15] Dawsoniolithon was recently established in Caragnano et al. (2018). There is no other genus morphologically identical with Dawsoniolithon. However, Caragnano et al. (2018) compare it with certain Neogoniolithon species and consider that the topography of spermatangia is crucial in discrimination between the two genera. Nevertheless, type of the roof formation in Dawsoniolithon and Neogoniolithon does not matches.
[16] See Townsend and Huisman (2018a, 2018b). Townsend and Huisman (2018a) erected new genus and subfamily Ozralia and Oztralioideae including O. gabrielsonii, O. orbiculata and O. conica. Thereof, the last mentioned was based on Hydrolithon conicum. Based on this species, however, Caragnano et al. (2018) erected new genus Dawsoniolithon, few months earlier. Therefore the genus name Oztralia as well as subfamily Oztralioideae became illegitimate and Townsend and Huisman (2018b) transferred remaining two Oztralia species to the genus Dawsoniolithon as well. Oztralioideae formerly included two genera Oztralia and Floiophycus. Second mentioned became the type genus of the new subfamily Floiophycoideae (Townsend & Huisman, 2018b).
[17] See Caragnano et al. (2018). Althought the systematic position within the Corallinales is yet uncertain, this genus can be distinguished from some similar genera based on the bi/tetrasporic conceptacles morphology. Caragnano et al. (2018) did not created the new subfamily because of the small number of included specimens.
[18] Lithophylloid coralline algae are well-separated from other known non-parasitic coralline algae by absence of secondary cells fusions between the cells of adjacent filaments (e.g., Woelkerling, 1988; Bittner et al., 2011; Harvey et al., 2009). However, Richards et al. (2014) point out that the subfamily should be genetically reassessed. Excluding the parasitic genus Ezo (Adey et al., 1974), subfamily Lithophylloideae include genera Lithophyllum and Titanoderma (Athanasiadis, 2016). Genera are usually separated based on presence of palisade cells in primigenous filaments found in the Titanoderma (Woelkerling, 1988; Chamberlain, 1991; Bailey et al., 1999). DNA sequence study suggests that two genera may be separated, however, boundaries which separate them are yet unresolved and validity of the name Titanoderma could not be made with confidence (Richards et al., 2014). Oldest fossil record of the Titanoderma is in Oligocene (Basso et al., 2019). Diagnosis of the genus Lithophyllum was emended by Basso et al. (2014) based on the trichocytes occurrences. Fossil Lithophyllum in complete life cycle are well recognized (Basso et al., 1996).
[19] See Nelson et al. (2015).
[20] Further information in Harvey et al. (2003).
[21] Austrolithon and Boreolithon were described in Harvey and Woelkerling (1995). The genus Epulo was designated by Townsend and Huisman (2004). All three genera were placed within the subfamily Austrolithoideae which encompass specimens with multiporate sporangial conceptacles and lacking secondary cell connections (Harvey & Woelkerling, 1995). However, Athanasiadis (2016) placed Boreolithon within the subfamily Melobesioideae which encompass specimens with dimerous thallus construction including e.g., Exilicrusta. Genera Austrolithon and Epulo are separated by number of cells in the roof filaments of sporangial conceptacles, number of cells in carpogonial disc (diameter) and presence/absence of haustoria, all of which could be to some degree preserved in fossil record, as well as number of nuclei per cell in vegetative filaments which are not fossilized (Townsend & Huisman, 2004).
[22] Subfamily Melobesioideae formerly encompassed other well-known fossil representatives of the genera Lithothamnion, Phymatolithon or Mesophyllum (e.g., Woelkerling, 1988; Aguirre et al., 2000). These genera are recently placed in families Lithothamniaceae (subfamily Lithothamnioideae) and Mesophyllumaceae (Athanasiadis, 2016; Schneider & Wynne, 2019). See comments to 6 - subfamily Mastophoroideae.
[23] Exilicrusta and Melobesia are genera with dimerous thallus internal construction. Thereof, Exilicrusta bears epithallial cells which are flared (Chamberlain, 1992).
[24] Schneider and Wynne (2019). Family Mesophyllumaceae encompass genera Leptophytum (Athanasiadis & Adey, 2006), Capensia (Athanasiadis, 2017), Mesophyllum (Athanasiadis & Ballantine, 2014), Melyvonnea (Athanasiadis & Ballantine, 2014), Synarthrophyton (Townsend, 1979; Athanasiadis, 2019), Clathromorphum (Adey et al., 2015), Callilithophytum (Adey et al., 2015), Neopolyporolithon (Adey & Johansen, 1972; Adey et al., 2015), Carlskottsbergia (Athanasiadis, 2019), Phragmope (Athanasiadis, 2020) and parasitic Kvaleya (Adey & Sperapani, 1971). Thereof, Capensia and Melyvonnea are designated based on the study of coralline algae formerly assigned to the genus Mesophyllum while Carlskottsbergia was designated based on the study of coralline algae formerly placed in Synarthrophyton. Carlskottsbergia and Capensia, however, bears monostromatic ventral core and monomerous thallus construction, hence the ventral core appear as reduced. Recently designated genus Tectonolithon bears two features which enable classify it within the Mesophyllumaceae based on classification of Athanasiadis (2016). These are unbranched spermatangia and carposporangia borne from the periphery of the central fertile zone (see Jesionek et al., 2020). Discrimination of the genera within the family Mesophyllumaceae is based on the growth form, morphology of pore canal lining cells, number of cells in pore canal filaments, morphology of the floor of carposporangial conceptacles, pattern in branching of vegetative filaments, morphology of meristematic cells, number of epithallial cells layers and fate of the conceptacles roof all of which are morphological characteristics hence observable on fossils. However, there are characteristics included as are those associated with the type and topography of spermatangia which cannot be recognized in fossil recerd, or their occurrence is extremely limited. Thereof, Carlskottsbergia and Phragmope bears unique characteristics which may facilitate their recognition. These are secondary aporous plate of sporangial conceptacles of Phragmope and monostromatic hypothallium of Carlskottsbergia.
[25] Family Lithothamniaceae include Phymatolithon with meristematic cells morphology similar to Leptophytum bud specialized pore canal lining cells are absent and Lithothamnion with flared epithallial cells and elongate meristematic cells (e.g., Woelkerling, 1988; Braga et al., 1993; Athanasiadis, 2001; Adey et al., 2015). Characteristics of the family Lithothamniaceae as are carposporangia borne across the fertile zone and predominantly dendroid spermatangia lack in the known fossil specimens contrary to the characteristics of the genera included (Basso et al., 1997; Coletti et al., 2018). Crustaphytum includes specimens with gametophytes bearing dendroid spermatangia. Detail description in Liu et al. (2018) and Jesionek et al. (2021).
[26] This group includes Tethysphytum with yet unknown gametophytes (see Sciuto et al., 2021).