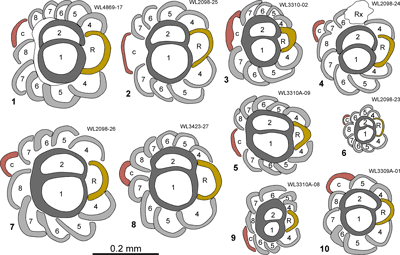
◄ Carnets Geol. 22 (11) ►
![]()
Outline:
[1. Introduction]
[2. Methodology]
[3. Investigated sections in Jamaica]
[4. Zonation scheme]
[5. Application to selected sections elsewhere in the American biogeographic province]
[6. Classification of LBFs and taxonomic notes]
[7. Conclusions]
[Bibliographic references]
and ...[Appendix]
Department of Geography and Geology, The University of the West Indies, Mona, Kingston
7 (Jamaica)
Department of Geography and Geology, The University of the West Indies, Mona, Kingston 7 (Jamaica)
İstanbul Technical University, Faculty of Mines, Department of Geological Engineering, Maslak 34469, İstanbul (Türkiye)
Ellington Geological Services, 1414 Lumpkin Road, Houston, TX 77043 (U.S.A.)
Department of Geography and Geology, The University of the West Indies, Mona, Kingston 7 (Jamaica)
Published online in final form (pdf) on July 1, 2022
DOI 10.2110/carnets.2022.2211
![]()
[Editor:
Daniela Basso; technical editor: Bruno R.C. Granier]
![]()
We have undertaken a study of the larger benthic foraminifera (LBF) of the Eocene of Jamaica that involved collecting free specimens and calibrating their occurrence against chronostratigraphy using calcareous nannofossils identified from sample splits or adjacent samples. The results have been plotted stratigraphically and allow a detailed zonation of the Eocene (14 zones) to be developed based largely on lepidocyclinids and similar forms. The zonation can be shown to be applicable to the Americas extending from northern South America, to Central America, the Caribbean and southern North America. The LBF biostratigraphy is based on the evolution of various groups. Three zones are recognized in the Ypresian, six zones in the Lutetian, three zones in the Bartonian, and two zones in the Priabonian. In order to classify the LBF we have revised the important groups used for the zonation scheme that occur in the Eocene and figure numerous examples. Our revision rearranges described genera into evolutionary, monophyletic clades to which we give the hierarchy of family and subfamily. We provide remarks on published taxa explaining how they are recognized and describe all the new taxa (two families, one subfamily, four genera and five species). The following families, genera and species are new: Butterliniana gen. nov., Helicosteginidae fam. nov., Helicostegina minor sp. nov., Helicostegina jamaicensis sp. nov., Helicostegina jeannemairae sp. nov., Pseudolepidinidae fam. nov., Orbitoininae subfam. nov., Planorbitoinella gen. nov., Polyorbitoinella gen. nov., Polyorbitoinella lilyfieldensis sp. nov., Orbitoina wrighti sp. nov., and Nephronummulites gen. nov.
• Lepidocyclinidae;
• Nummulitidae;
• Helicosteginidae;
• Eocene;
• biostratigraphy;
• American Larger Benthic Foraminifer (LBF) zones
Mitchell S.F., Robinson E., Özcan E., Jiang M.M. & Robinson N. (2022).- A larger benthic foraminiferal zonation for the Eocene of the Caribbean and central American region.- Carnets Geol., Madrid, vol. 22, no. 11, p. 409-565.
Zonation de l'Éocčne de la région des Caraďbes et de l'Amérique centrale ŕ l'aide des grands foraminifčres benthiques.- Le matériel de notre étude des grands foraminifčres benthiques (GFB) de l'Éocčne de la Jamaďque provient de la récolte de spécimens dégagés. Leur distribution a été calée sur une échelle chronostratigraphique fondée sur les nannofossiles calcaires identifiés ŕ partir de broyats des męmes échantillons ou d'échantillons adjacents. Les résultats de ce calibrage biostratigraphique permettent d'élaborer une zonation détaillée de l'Éocčne, soit 14 zones fondées en grande partie sur les lépidocyclinides et formes affines. On démontre que cette zonation peut s'appliquer aux Caraďbes et aux Amériques, depuis la partie septentrionale de l'Amérique du Sud et, via l'Amérique centrale, jusqu'ŕ la partie méridionale de l'Amérique du Nord. La biostratigraphie par GFB s'appuie sur l'évolution des différents groupes. Trois zones sont ainsi identifiées pour l'Yprésien, six pour le Lutétien, trois pour le Bartonien et deux pour le Priabonien. Afin d'organiser taxinomiquement ces GFB, nous avons révisé les groupes importants présents ŕ l'Éocčne et utilisés dans notre zonation. Ils sont pour la plupart abondamment figurés. Notre révision réorganise les genres décrits en clades évolutifs et monophylétiques auxquels nous attribuons le niveau hiérarchique de famille et sous-famille. Nous fournissons des précisions sur les taxons publiés en donnant les critčres qui ont servi ŕ leur identification. Quelques taxons nouveaux (deux familles, une sous-famille, quatre genres et cinq espčces) sont introduits ici ; il s'agit de Butterliniana n. gen., Helicosteginidae n. fam., Helicostegina minor n. sp., Helicostegina jamaicensis n. sp., Helicostegina jeannemairae n. sp., Pseudolepidinidae n. fam., Orbitoininae n. subfam., Planorbitoinella n. gen., Polyorbitoinella n. gen., Polyorbitoinella lilyfieldensis n. sp., Orbitoina wrighti n. sp. et Nephronummulites n. gen.
• Lepidocyclinidae ;
• Nummulitidae ;
• Helicosteginidae ;
• Éocčne ;
• biostratigraphie ;
• zones de Grands Foraminifčres Benthiques (GFB) américains
With a duration of 22.1 Myr the Eocene is the longest epoch in the 66 Myr-long Cenozoic and is traditionally zoned using planktic foraminifers (e.g., Bolli, 1957a, 1957b; Bolli & Saunders, 1985; Wade et al., 2011; Vandenderghe et al., 2012; Ogg et al., 2016; Speijer et al., 2020) and calcareous nannofossils (e.g., Martini, 1971; Bukry, 1973, 1975; Okada & Bukry, 1980; Vandenderghe et al., 2012; Agnini et al., 2014; Ogg et al., 2016). Yet these groups are difficult to use in shallow-water platform limestones because they are either rare or absent, or because of the generally lithified nature of these deposits. In the Old World (i.e., Europe and the Middle East), a high-resolution, larger benthic foraminiferal (LBF) zonation has been developed for the shallow-water Tethys Paleogene and early Neogene (Cahuzac & Poignant, 1997; Serra-Kiel et al., 1998; Less & Özcan, 2012; Papazzoni et al., 2017), yet no comparable scheme has been developed for the New World (Americas), although LBF have been used to biostratigraphically calibrate successions.
Work in Florida (P.L. Applin & E.R. Applin, 1944; Cole, 1941, 1942, 1944, 1945; Cole & E.R. Applin, 1964) suggested a series of guide fossils for the Eocene. Later work showed that the ranges of many of these 'guide fossils' were stratigraphically more extensive in the Caribbean (e.g., Cole & Gravell, 1952), probably due to diachroneity of facies.
In Jamaica, Matley (1925) had attempted to zone the White Limestone using LBF, and Hose and Versey (1956) had recognized a series of zones and fossil bands, many of which were based on foraminifers that had been evaluated by Cole (1956). Robinson (1977, 1988, 1993) suggested a series of zones for the Eocene of central Jamaica, and subsequently Robinson and Jiang (1995) and Robinson (1996, 1997) demonstrated the value of evolution as a high-resolution biostratigraphic tool within the Eulinderina-Polylepidina-Lepidocyclina lineage in Florida and Alabama, as well as in parts of the Caribbean. In 1999, Robinson and Mitchell suggested a series of LBF assemblages for the Eocene and early Oligocene of Jamaica, and in 2004, Robinson presented a range chart showing the stratigraphic distribution of Cenozoic LBF species for Jamaica.
Elsewhere in the American region, Butterlin (1961, 1971, 1981) studied the LBF and suggested a zonation for the Eocene of Mexico (Butterlin, 1981). He recognized one subzone (B2: Pseudophragmina zaragosensis subzone) in the lower Eocene, four subzones (Ca: Helicostegina dimorpha subzone; C2: Lepidocyclina (Polylepidina) antillea subzone; C3: Lepidocyclina (Neolepidina) ariana subzone; C4: Lituonella grandicamerata subzone in the C: Discocyclina marginata zone) in the middle Eocene, and two subzones (D1: Dictyoconus americana subzone, D2: Heterodictyoconus cookei subzone in the D: Asterocyclina minima zone) in the upper Eocene. The middle Eocene zonation followed that recognized in Florida by Cole and E.R. Applin (1964), whereas the upper Eocene zonation followed Robinson (1977).
A zonation has also been proposed for north-eastern Brazil (Mello e Sousa et al., 2003), however, many of the specimens illustrated are random sections and some species (e.g., Helicostegina dimorpha and Amphistegina lopeztrigoi) do not seem to be correctly identified.
While all these schemes allow some biostratigraphic resolution for parts of the Eocene, it is far below that of the comparable scheme in the Old World. In this paper we develop a high-resolution biostratigraphic zonation for the Eocene based on LBF and chronostratigraphically calibrate it using calcareous nannofossils. We have studied sections in Jamaica that expose a more-or-less complete Eocene succession in a variety of deposition environments: shallow-water platform deposits, platform edge deposits and shelf and basin deposits. Although small unconformities are developed in the shallow-water platform successions, the deeper-water successions appear to be more complete, although some short hiatuses (or faults?) have been recognized in this study. Jamaica also lies in the centre of the evolutionary area for many of the LBF used in the zonation, and therefore Jamaica seems ideally suited to develop a zonation scheme based on LBF for the Eocene. Our zonation has a comparable resolution to the zonation developed in the Old World at least for the late Ypresian to Priabonian (Serra-Kiel et al., 1998).
In this paper, we describe our methodology, describe sections used in Jamaica to define the Eocene scheme, and lay out the zonation scheme. We then show how the scheme can be used for the Eocene in selected areas of the American biogeographical province using our own observations and previous published data. Finally, we provide an updated taxonomy of the LBF used in the zonation.
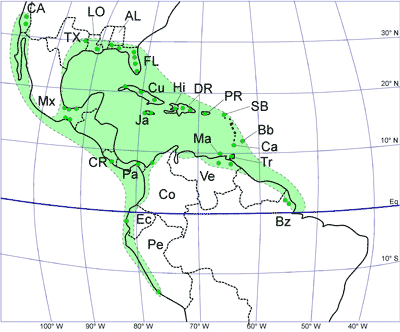
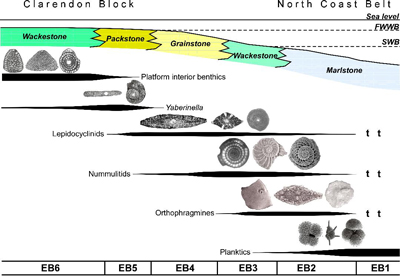
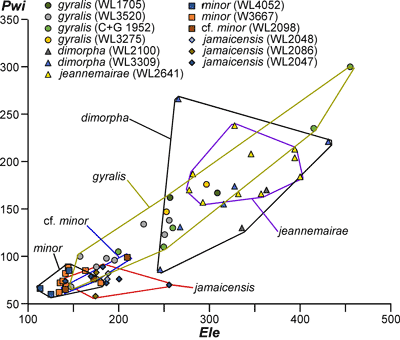
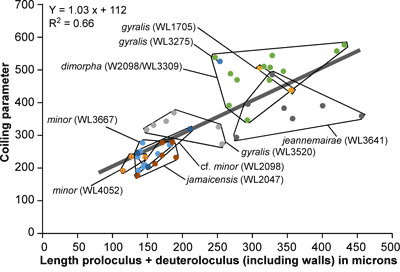
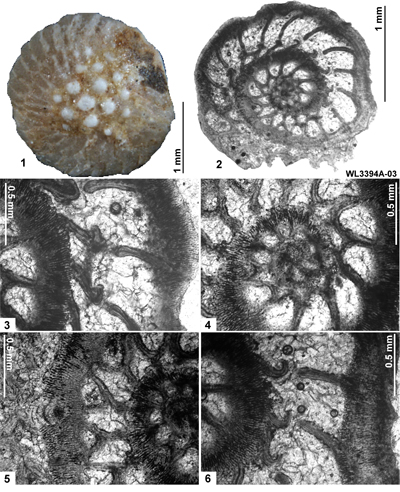
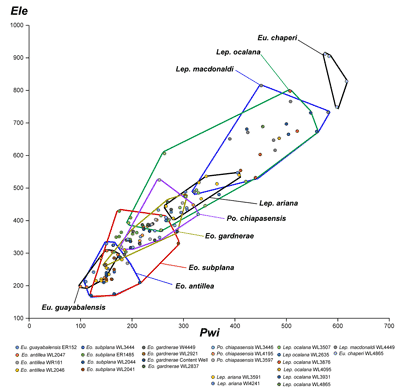
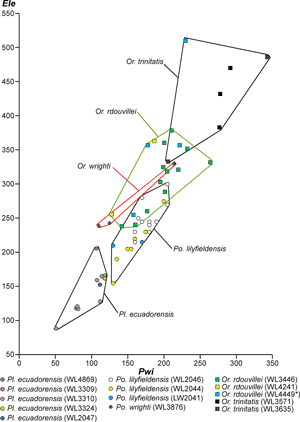
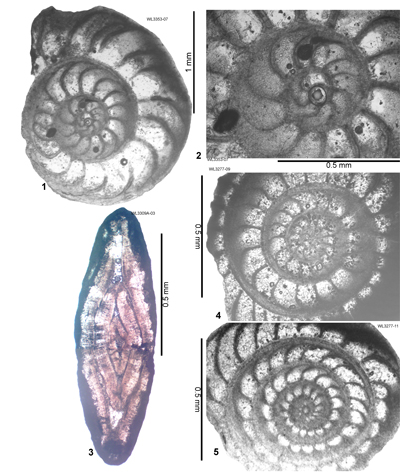
LBF are abundant in shallow-water Eocene rocks of the Caribbean and middle American region (Fig. 1 ![]() ) and range from shallow-water restricted environments to the deeper parts of the photic zone (Eva, 1976; Wright, 1974; Robinson & Mitchell, 1999; Table 1). Those inhabiting the platform margin and shallow-water photic zone (e.g., lepidocyclinids and nummulitids) have the widest palaeoecological distribution (Fig. 2
) and range from shallow-water restricted environments to the deeper parts of the photic zone (Eva, 1976; Wright, 1974; Robinson & Mitchell, 1999; Table 1). Those inhabiting the platform margin and shallow-water photic zone (e.g., lepidocyclinids and nummulitids) have the widest palaeoecological distribution (Fig. 2 ![]() ) and are selected as the principal taxa for the development of a zonation scheme in this paper.
) and are selected as the principal taxa for the development of a zonation scheme in this paper.
|
|
Figure 1: Map of the distribution of Larger Benthic Foraminifers in the American (Caribbean) lepidocyclinid bio-geographical province using the current plate configuration. Dots indicate selected occurrences of
LBF. AL, Alabama; Bb, Barbados; Bz, Brazil; CA, California; Co, Colombia; CR, Costa Rica; Ca,
Carriacou; Cu, Cuba; DR, Dominican Republic; Ec, Ecuador; Fl, Florida; Hi, Haiti;
Ja, Jamaica; LO, Louisiana; Ma, Margarita; Mx, Mexico; Pa, Panama; Pe, Peru; PR, Puerto Rico; SB, St Bartholomew;
Tr, Trinidad; TX, Texas; Ve, Venezuela. |
Table 1: Foraminiferal biofacies from the Eocene of Jamaica. In situ assemblages (EB) and transported assemblages (EB-t) can be recognized. Based on Eva (1976), Wright (1974), and Robinson and Mitchell (1999) with amendments.
| Biofacies | Description and interpretation |
| EB1 | Foraminiferal assemblage consists of planktic and smaller benthic foraminifers. LBF absent except as transported assemblages in turbidites and storm beds (EB2-t, EB3-t and EB4-t). Deposition below the photic zone (lower ramp). |
| EB2 (EB2-t as transported assemblage down slope) | LBF dominated by diverse assemblages of orthophragmines together with generally small numbers of Asterigerinoidea and other taxa (e.g., dictyoconids, nummulitids). Deposition in lower part of the photic zone (middle ramp). |
| EB3 (EB3-t as transported assemblage down slope) | LBF dominated by lepidocyclinids and nummulitids with only a low diversity (but often high abundance) assemblage of orthophragmines (notably Stenocyclina). Middle photic zone (middle ramp) |
| EB4 (EB4-t as transported assemblage down slope) | LBF dominated by lepidocyclinids (orthophragmines absent) and some nummulitids. Middle to upper photic zone, middle to upper ramp and open shelf edge. |
| EB5 | LBF assemblage comprises lepidocyclinids and platform interior forms (typically Yaberinella) in broadly equal abundance. Upper photic zone, inner ramp, and protected platform edge settings. |
| EB6 | LBF assemblage dominated by platform interior forms (Yaberinella, Pseudofabularia, Fabularia, dictyoconids, miliolids, etc.). Upper photic zone, inner platform settings. |
|
|
Figure 2: Palaeoecological distribution of selected Larger Benthic Foraminifers in the Eocene based on transects across the Clarendon Block and North Coast Belt of Jamaica. Revised from Eva (1976), Wright (1974), and Robinson
and Mitchell (1999). T
- specimens transported in
turbidites/debris flows. |
Extensive new material (about 100 samples) has been collected from Jamaica over the last 15 years, which we integrate with previously collected material from Jamaica and elsewhere within the Caribbean-American region (Fig. 1 ![]() ). In general we have tried to collect free specimens of LBF so that external ornamentation can be recorded, and orientated sections (equatorial and axial) can be cut. This allows us to evaluate whether species can be determined in random orientations in thin sections cut from rock samples.
). In general we have tried to collect free specimens of LBF so that external ornamentation can be recorded, and orientated sections (equatorial and axial) can be cut. This allows us to evaluate whether species can be determined in random orientations in thin sections cut from rock samples.
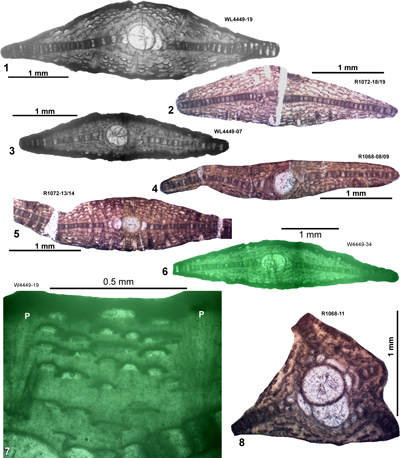
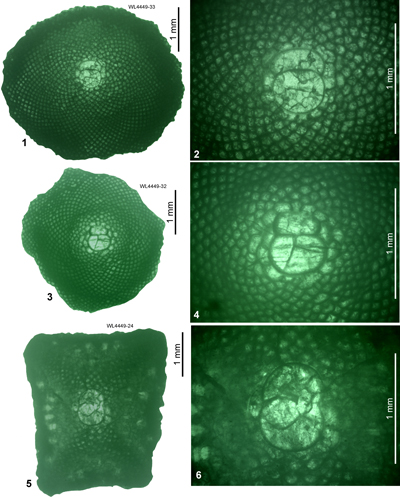
Typically large samples (a one US gallon or 3.78 litre sized bag) were collected by one of us (SFM, labelled WL) or by two of us (SFM and EÖ, also labelled WL) and broken down by drying and soaking in water, or by repeated freeze-thaw cycles in water. Following breakdown, samples were washed through a 0.5 mm sieve and the LBF were picked. For samples with low yields (or low yields of critical species) further samples were collected and disaggregated. Specimens were sorted into morphospecies based on exterior characteristics and selected specimens were then cut on a glass plate with 600 grade corundum grit to provide orientated equatorial and axial sections. During the cutting process observations were also made of the presence of features (e.g., lateral chamberlets and piles/pillars) that would not be present in the final orientated equatorial thin-section. UWIGM.WL samples are preserved in the collections of the University of the West Indies Geological Museum, Jamaica. EO.WL samples are preserved in the paleontological collections of the Geological Engineering Department of İstanbul Technical University, Turkey. USNM refers to specimens housed in the Smithsonian Institute, Washington - DC. Additional material collected by two of us (ER and NR) is also used in this study, this is prefixed by the letters 'ER' and 'R' respectively. The ER material was collected at various times between the late 1950s and 2005. The NR material was collected in 2017-2018. Some of the ER and NR material includes orientated sections, but most is of unorientated specimens in thin section. A detailed list of samples is provided in the sections below.
Selected samples were also studied for nannofossils. These either represented samples from continuous sections or spot samples associated with particular LBF assemblages. Previous work on nannofossils (e.g., Jiang & Robinson, 1987; Robinson & Jiang, 1997) has been updated for inclusion in this work.
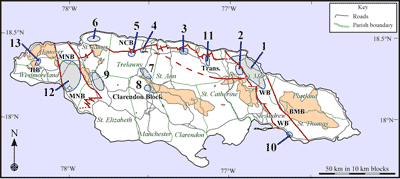
We present brief details of the investigated sections here together with the lithostratigraphic schemes currently in use. Sections are included either because they contain intervals with both LBF and nannofossils, because they yielded free specimens of LBF, or because they are important historically for the descriptions of LBF. We correlate these sections using our LBF zones (American Benthic Zones or
ABZ) that are calibrated to the chronostratigraphy using calcareous nannofossils. This paper is concerned with the Eocene which covers zones AE3 to AE16 (Fig. 4 ![]() ) and zones for the Paleocene and Oligocene to Miocene will be presented elsewhere. The zones are formally described after the description of the sections. A revision of the lithostratigraphic units will be presented elsewhere, as it is beyond the scope of this paper. Numbers for localities refer to Fig. 3
) and zones for the Paleocene and Oligocene to Miocene will be presented elsewhere. The zones are formally described after the description of the sections. A revision of the lithostratigraphic units will be presented elsewhere, as it is beyond the scope of this paper. Numbers for localities refer to Fig. 3 ![]() .
.
|
|
Figure 3: Eocene areas in Jamaica studied for the development of the
zonation. 'HB' - Hanover Block; BMB - Blue Mountains Block (Cretaceous rocks, same colour elsewhere); MNB
- Montpelier-New Market Belt; NCB - North Coast Belt; Trans. - Transitional zone between North Coast Belt and Clarendon Block; WB
- Wag Water Belt. Areas studied: 1, northern Wag Water Belt; 2, Rio
Sambre; 3, Lilyfield-Bamboo area; 4, Swanswick-Clarks Town area; 5, Sherwood Content area;
6, Salt Spring
- Flower Hill area; 7, Albert Town-Litchfield area; 8, Coleyville area;
9, Western Clarendon Block; 10, Grants Pen area (St Thomas); 11, N-S Highway (St Ann);
12, Montpelier-New Market Belt; 13, Jerusalem Mountain area. |
|
|
Figure 4: Correlation of formations across Jamaica using Larger Benthic Foraminifers (see zonation below) and calcareous
nannofossils. Numbers as in Fig. 3 |
Area 1. Northern Wag Water Belt (parish of St Mary): Ypresian to Priabonian
This area of the northern part of the Wag Water Belt (Fig. 3 ![]() ) consists of small-scale tectonic blocks separated by faults. As such, a continuous stratigraphy is not present. The lithostratigraphy follows Burke et al. (1969) and Mann
and Burke (1990). LBF have been recovered from the Wag Water Formation, Richmond Formation and the Yellow Limestone (Palmetto Grove and Preston Hill formations) and White Limestone ("Bonny Gate" Formation) groups. The LBF typically occur as transported specimens in turbidites in deep-water shales and marlstone-limestone deposits, which we consider are broadly contemporaneous with the sedimentary rocks they are found in. Chronostratigraphic correlation is based on updated previous nannofossil records (Jiang & Robinson, 1987; Robinson, 1996, 2004) together with additional samples investigated in this work. The samples are listed in Table 2 (with details of the nannofossils in Appendix 1) and a correlation of the formations in this area is shown in Fig. 4
) consists of small-scale tectonic blocks separated by faults. As such, a continuous stratigraphy is not present. The lithostratigraphy follows Burke et al. (1969) and Mann
and Burke (1990). LBF have been recovered from the Wag Water Formation, Richmond Formation and the Yellow Limestone (Palmetto Grove and Preston Hill formations) and White Limestone ("Bonny Gate" Formation) groups. The LBF typically occur as transported specimens in turbidites in deep-water shales and marlstone-limestone deposits, which we consider are broadly contemporaneous with the sedimentary rocks they are found in. Chronostratigraphic correlation is based on updated previous nannofossil records (Jiang & Robinson, 1987; Robinson, 1996, 2004) together with additional samples investigated in this work. The samples are listed in Table 2 (with details of the nannofossils in Appendix 1) and a correlation of the formations in this area is shown in Fig. 4 ![]() .
.
A calcareous conglomerate in the Wag Water Formation (at Devon Pen) yielded a LBF assemblage characterised by Tremastegina with orthophragmines (Jiang & Robinson, 1987). This would indicated ABZ3. The Richmond Formation yields LBF in turbidites or shell beds (Robinson & Mitchell, 1999) and indicate zones ABZ3 and ABZ4. Samples from the Richmond Formation around Albany yield ABZ3 foraminifers with nannofossils indicating Zone NP12 (Jiang & Robinson, 1987). Samples from mudstones around Langley yield LBF indicating ABZ4 and nannofossils indicating Zone NP14a (Jiang & Robinson, 1987; Appendix 1). The Wag Water Formation and Richmond Formation, at least in this area, must interfinger; with the Wag Water Formation representing slope deposits that pass into the basinal sandstones and shales of the Richmond Formation (Mitchell, 2021).
The Yellow Limestone Group is represented by the Palmetto Grove and Preston Hill formations (Burke et al., 1969; Robinson & Mitchell, 1999). One sample was collected from the Palmetto Grove Formation (WL3315) and four samples were collected from the Preston Hill Formation (ER152 (= original ER sample R78), ER161, WL4869 and WL4870). No nannofossil data are available for WL3315 (which contains ABZ5 LBF) or ER152/R78 (which contains ABZ7 LBF). Planktic foraminiferal and nannofossil data were published for ER161 (P11, upper NP15, and CP13-lower CP14a; ABZ7-ABZ8 transition) by Robinson (1996). Nannofossil data for samples WL4869 and WL4870, which yield Eulinderina guayabalensis and Planorbitoinella ecuadorensis n. gen. et sp., indicate lower Zone NP15b (Table 2, Appendix 1).
Table 2: Details of LBF and calcareous nannofossil zone for samples from area 1 (northern Wag Water Belt). Column 1 indicates LBF zone assignment based on proximity of samples or fauna. WL GPS positions were measured in the field (ER GPS positions picked from georeferenced original field maps).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ3 | Devon Pen | c. 18°12.3'N; 76°48.9'W | Devon Pen | Wag Water | - | ABZ3 * |
| " | WL3157 | 18°17.789'N; 76°52.263'W | Albany | Richmond | - | ABZ3 |
| " | WL3156 | 18°17.444'N; 76°53.085'W | Albany | Richmond | - | ABZ3 |
| " | WL3304 | 18°17.720'N; 76°51.768'W | Albany | Richmond | - | ABZ3 |
| " | ER1547 | c. 18°17.62.N; 76°50.95'W | Albany | Richmond | NP12 * | - |
| " | ER1548 | c. 18°17.72'N; 76°51.10'W | Albany | Richmond | NP12 * | - |
| " | ER1549 | c. 18°17.17'N; 76°51.88'W | Albany | Richmond | NP12 * | - |
| " | ER1550 | c. 18°17.59'N; 76°51.81'W | Albany | Richmond | NP12 * | - |
| " | ER1551 | c. 18°18.32'N; 76°51.83'W | Albany | Richmond | NP12 * | - |
| ABZ4 | WL3158 | 18°15.176'N; 76°52.748'W | Richmond | Richmond | - | ABZ4 |
| " | WL3159 | 18°14.507'N; 76°52.666'W | Richmond | Richmond | - | ABZ4 |
| " | WL3160 | 18°14.507'N; 76°52.666'W | Richmond | Richmond | - | ABZ4 |
| " | WL3148B | 18°13.353'N; 76°50.987'W | Rock River | Wag Water - Richmond transition | NP14a | - |
| " | WL3394A | 18°19.056'N; 076°55.387'W | Langley | Richmond | NP14a | ABZ4 |
| " | WL3154 | 18°18.649'N; 76°54.904'W | Langley | Richmond | - | ABZ4 |
| ABZ5 | WL3155 | 18°18.252'N; 76°55.237'W | Hampstead | Palmetto Grove | - | ABZ5 |
| " | WL3316 | " | " | " | - | ABZ5 |
| " | WL3381 | " | " | " | - | ABZ5 |
| " | WL3155 | " | " | " | - | ABZ5 |
| ABZ7 | ER152 (R78) | c. 18°23.24'N; 76°54.25'W | Grants Town | Preston Hill | - | ABZ7 |
| " | WL4869 | 18°23.242'N; 76°54.247'W | Grants Town | Preston Hill | Lower NP15b | ABZ7 |
| " | WL4870 | 18°23.257'N; 76°54.314'W | Grants Town | Preston Hill | Lower NP15b | ABZ7 |
| " | ER161 | c. 18°19.57'N; 76°55.81'W | Preston Hill | Preston Hill | Upper NP15+ | ABZ7/8 |
| ABZ11 | WL3416 | 18°20.116'N; 076°59.907'W | Rio Nuevo, Gayle | 'Bonny Gate' | - | ABZ11 |
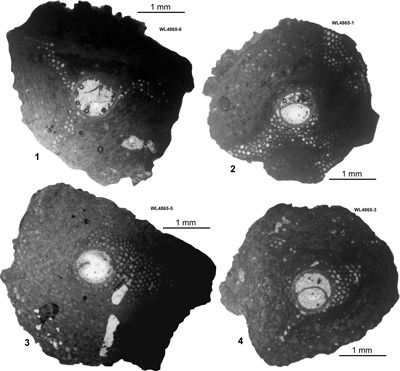
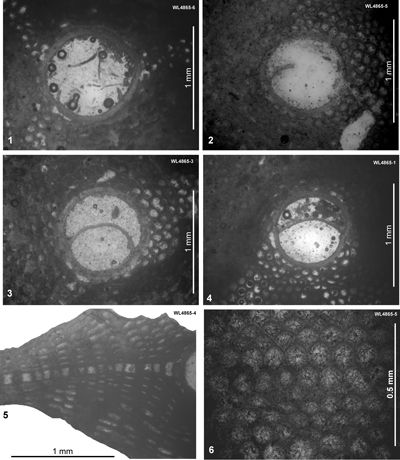
| ABZ16 | ER176 | c. 18°20.08'N; 76°59.05'W | Dressike | 'Bonny Gate' | NP21 * | ABZ16 |
| " | WL4865 | 18°20.079'N; 76°59.045'W | Dressike | 'Bonny Gate' | - | ABZ16 |
* Data from Jiang & Robinson (1987); + Data from Robinson (1996).
The "Bonny Gate" Formation in this area has also yielded some useful samples. These are referred to the early Bartonian (WL3416: ABZ11) and late Priabonian (ER176, WL4865, ABZ16). Nannofossil data for ER176 (zone NP21) were published by Robinson (2004).
Area 2. North Coast Belt, Rio Sambre Gorge, and area around (parish of St Mary): Ypresian to Lutetian
This is among the most important areas for establishing the succession of faunas from the late Ypresian to the late Lutetian because of the continuous nature of the successions and quality of the exposures present. The lower part of the succession (Guys Hill, Richmond, and lower Palmetto Grove formations) is exposed in the bed and banks of the Rio
Sambre, about 0.5 to 2 km downstream from the bridge between Palmetto Grove and
Kilancholly, and the upper part (upper Palmetto Grove and Preston Hill formations) is exposed in road cuts between Farm Turn, Kilancholly and Palmetto Grove in St Mary (Fig. 3 ![]() ). The section in the Rio Sambre has been the subject of several reports (e.g., Burke et al., 1969; Wise & Constans, 1976; Robinson & Mitchell, 1999). Detailed geological mapping indicates that the section begins with the Pembroke Hall Formation (of Burke et al., 1969), passes through the Guys Hill, Richmond and Palmetto Grove formations and terminates in the Preston Hill Formation. A summary section of the Richmond and Palmetto Grove formations was published in Robinson and Mitchell (1999,p. 38-39).
). The section in the Rio Sambre has been the subject of several reports (e.g., Burke et al., 1969; Wise & Constans, 1976; Robinson & Mitchell, 1999). Detailed geological mapping indicates that the section begins with the Pembroke Hall Formation (of Burke et al., 1969), passes through the Guys Hill, Richmond and Palmetto Grove formations and terminates in the Preston Hill Formation. A summary section of the Richmond and Palmetto Grove formations was published in Robinson and Mitchell (1999,p. 38-39).
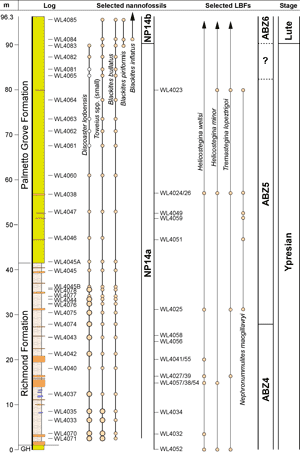
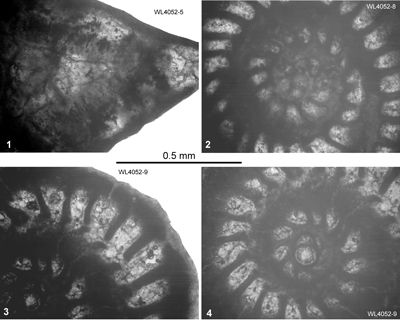
We have re-measured and recollected this entire section. Three separate sections were logged in the Rio
Sambre. Section 1 (18° 17.411" N; 76° 56.039" W) extends from the Richmond Formation (samples WL4054-WL4056: ABZ4) into the lower part of the Palmetto Grove Formation. Section 2 (18° 16.903" N; 76° 55.547" W to 18° 16.991" N; 76° 55.618" W) extends from the Richmond Formation (WL4071-4070, WL4033, WL4035, WL4037, WL4041: ABZ4) into the lower Palmetto Grove Formation (samples WL4049, WL4026: ABZ5). Section 3 (18° 16.930" N; 76° 55.621" W to 18° 16.903" N; 76° 55.547" W) extends from the Richmond Formation (samples WL4057, WL4027, WL4058: ABZ4; sample WL4025: ABZ5) into the lower Palmetto Grove Formation (samples WL4059, WL4024, WL4023: ABZ5). The different sections can be correlated using turbidites (Fig. 5 ![]() ) which can be traced between sections. Samples were collected for calcareous nannofossils to chronostratigraphically calibrate the succession. This indicates the presence of zone NP14a and basal NP14b. Additionally upsection reductions in the abundances of small Toweius
spp. and Discoaster lodoensis suggest that nearly the entire thickness of Zone NP14a is present (Fig. 5
) which can be traced between sections. Samples were collected for calcareous nannofossils to chronostratigraphically calibrate the succession. This indicates the presence of zone NP14a and basal NP14b. Additionally upsection reductions in the abundances of small Toweius
spp. and Discoaster lodoensis suggest that nearly the entire thickness of Zone NP14a is present (Fig. 5 ![]() ). The appearance of Blackites inflatus near the top of the section (Fig. 5
). The appearance of Blackites inflatus near the top of the section (Fig. 5 ![]() ) indicates the base of the
Lutetian. Details of all samples are provided in Fig. 5
) indicates the base of the
Lutetian. Details of all samples are provided in Fig. 5 ![]() and Table 3.
and Table 3.
|
Figure 5: Composite of logged sections for the Rio Sambre Gorge showing distribution of selected nannofossils and Larger Benthic Foraminifers. Dot sizes for nannofossils indicates relative abundance; white dots indicate sporadic occurrences. Dots for foraminifers indicate presence. |
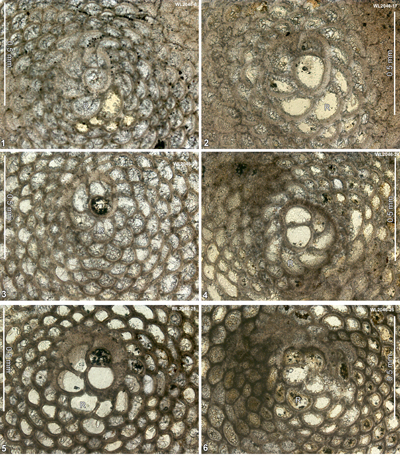
The upper part of the succession is limited to small exposures along roads around Farm Turn, and Kilancholly, where the upper part of the Palmetto Grove Formation and the lower part of the Preston Hill Formation are exposed. Spot samples along these roads have been analysed for their LBFs and zones ABZ6 to ABZ8 are represented (Table 3). These samples have been tied into the chronostratigraphy using calcareous nannofossil identifications from sample splits. Very rich assemblages of LBF are present including dictyoconids, orthophragmines, helicosteginids, nummulitids and Fabiania, amongst others. Table 3 shows the breakdown of sample numbers and zone based on LBF and calcareous nannofossils for samples above the Rio Sambre section. Details of the nannofossils are given in Appendix 1.
Table 3: Details of LBF and calcareous nannofossil zone for samples (above the Rio Sambre section) from area 2 (western St Mary).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ6 | WL4066 | 18°17.351'N; 76°55.645'W | Farm Turn | Palmetto Grove | NP14b | - |
| " | WL3307 | 18°17.267'N; 76°55.667'W | Farm Turn | Preston Hill | - | - |
| " | WL3317 | 18°17.603'N; 76°55.499'W | Farm Turn | Preston Hill | NP14b | - |
| " | WL3308A | 18°17.324'N; 76°55.566'W | Farm Turn | Preston Hill | NP14b | ABZ6 |
| ABZ7 | WL3305 | 18°17.212'N; 76°55.611'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL3306 | 18°17.221'N; 76°55.619'W | Farm Turn | Preston Hill | - | ABZ7 |
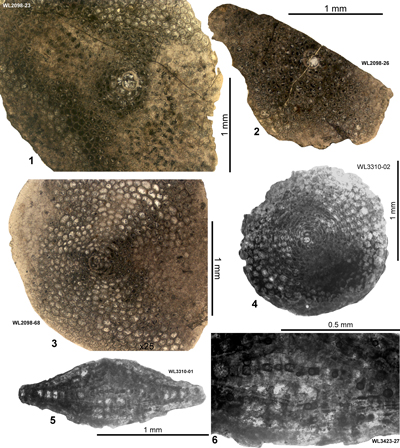
| " | WL2098 | 18°17.542'N; 76°55.452'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL3309 | 18°17.542'N; 76°55.452'W | Farm Turn | Preston Hill | NP15b | ABZ7 |
| " | WL3309A | 18°17.542'N; 76°55.452'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL3310 | 18°17.632'N; 76°55.487'W | Farm Turn | Preston Hill | NP15b | ABZ7 |
| " | WL3310A | 18°17.632'N; 76°55.487'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL2099 | 18°17.632'N; 76°55.487'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL3311 | 18°17.648'N; 76°55.438'W | Farm Turn | Preston Hill | NP15b | ABZ7 |
| " | WL4068 | 18°17.872'N; 76°55.127'W | Farm Turn | Preston Hill | - | ABZ7 |
| " | WL3405 | 18°16.642'N; 76°55.304'W | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL4028 | 18°16.638'N; 76°55.153'W | Kilocholly | Preston Hill | - | ABZ7 |
| " | WL4029 | 18°16.861'N; 76°55.105'W | Kilocholly | Preston Hill | - | ABZ7 |
| " | WL3030 | 18°16.890'N; 76°55.078'W | Kilocholly | Preston Hill | - | ABZ7 |
| " | WL4031 | 18°16.941'N; 76°55.100'W | Kilocholly | Preston Hill | - | ABZ7 |
| " | WL3695 | 18°16.631'N; 76°55.289'W (Series of samples taken along road way) |
Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3696 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3697 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3698 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3699 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3700 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3701 | " | Kilocholly | Preston Hill | NP15b | ABZ7 |
| " | WL3423 | 18°16.631N; 76°55.289'W | Kilocholly | Preston Hill | - | ABZ7 |
| ABZ8 | WL3312 | 18°17.664'N; 76°55.416'W | Farm Turn | Preston Hill | NP15b | ABZ8 |
| " | WL2100 | 18°17.664'N; 76°55.416'W | Farm Turn | Preston Hill | - | ABZ8 |
| " | WL3314 | 18°17.605'N; 76°55.295'W | Farm Turn | Preston Hill | NP15b | ABZ8 |
| " | WL4069 | 18°17.606N; 76°55.291'W | Farm Turn | Preston Hill | - | ABZ8 |
| " | WL3313 | 18°17.661N; 76°55.353'W | Farm Turn | Preston Hill | - | ABZ8 |
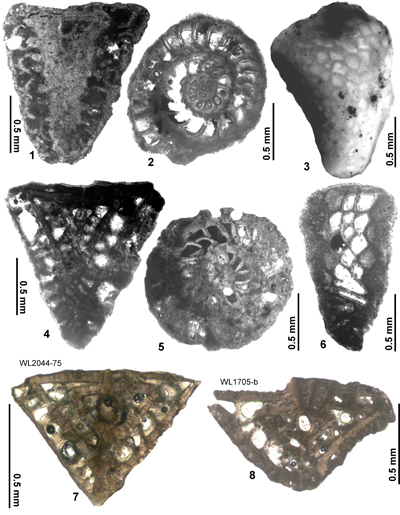
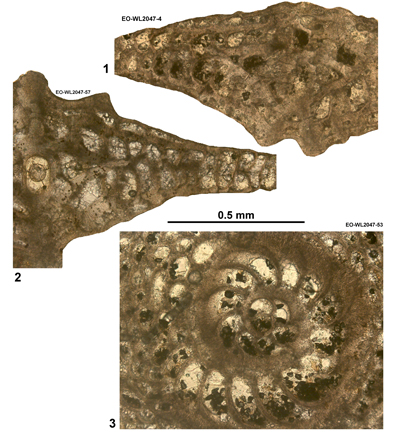
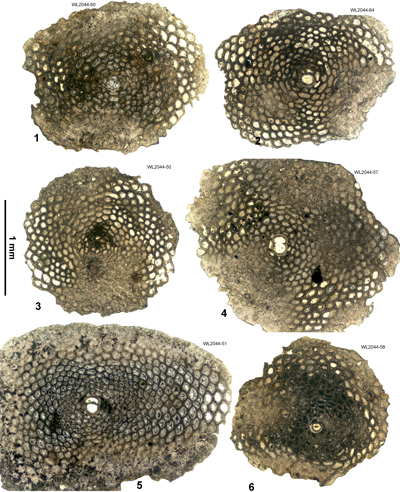
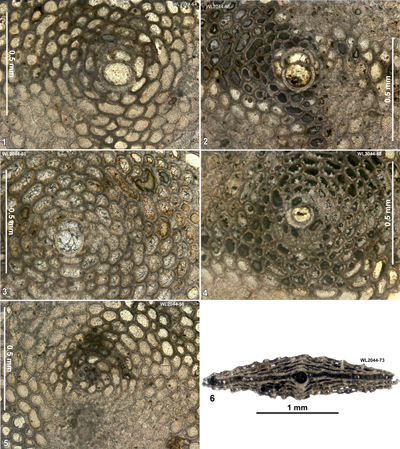
Area 3. Lilyfield-Bamboo Area (North Coast Belt, parish of St Ann): Lutetian
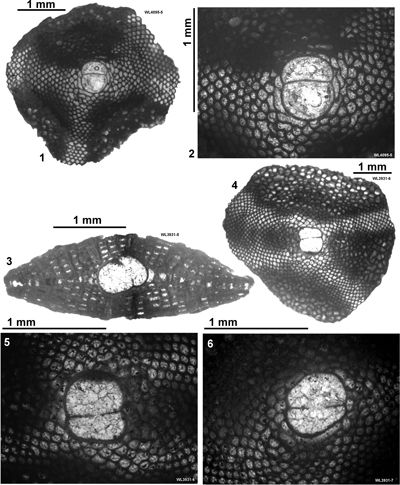
This area is historically significant because Cole (1956, 1963) presented information on the LBF from here. Özcan et al. (2019,
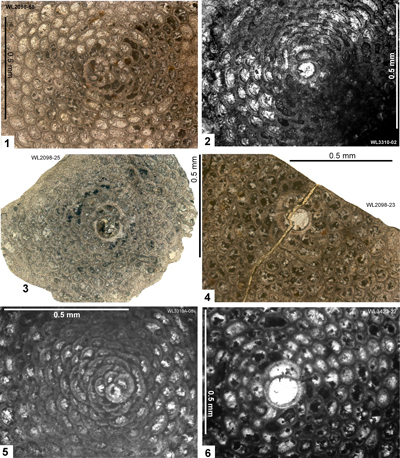
2022a) also presented data on the orthophragmines from here. This area (Fig. 3 ![]()
![]() ) yields good faunas from the Preston Hill and
"Chapelton" formations. Above this is a typical shallow-water Clarendon Block stratigraphy which consists of the Troy,
"Swanswick" s.l. (see area 4 below), Claremont and Somerset formations; these are hard limestones and were not sampled for LBF because they lack free specimens (they are included on Fig. 4
) yields good faunas from the Preston Hill and
"Chapelton" formations. Above this is a typical shallow-water Clarendon Block stratigraphy which consists of the Troy,
"Swanswick" s.l. (see area 4 below), Claremont and Somerset formations; these are hard limestones and were not sampled for LBF because they lack free specimens (they are included on Fig. 4 ![]() for completeness). We did not carry out any nannofossil analyses for this area, because this part of the stratigraphy was covered in the area around Farm Turn (Area 2) and the LBF allow a straight forward correlation (Fig. 4
for completeness). We did not carry out any nannofossil analyses for this area, because this part of the stratigraphy was covered in the area around Farm Turn (Area 2) and the LBF allow a straight forward correlation (Fig. 4 ![]() ). Zones ABZ8 and ABZ9 are represented in this area (Fig. 4
). Zones ABZ8 and ABZ9 are represented in this area (Fig. 4 ![]() ). Details of the samples, LBF zones and nannofossil zone are given in Table 4 and Appendix 1.
). Details of the samples, LBF zones and nannofossil zone are given in Table 4 and Appendix 1.
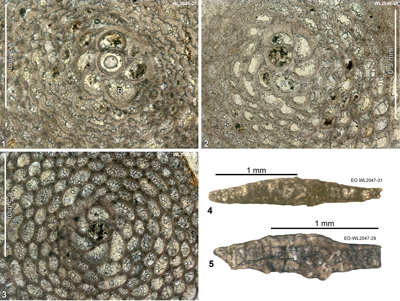
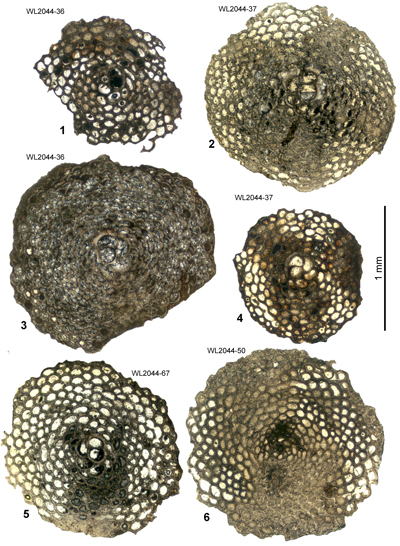
Table 4: Details of LBF zone for samples from area 3 (Lilyfield area). (Nannofossils were not studied from this area.)
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ8 | WL3427 | 18°25.288'N; 077°16.427'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| " | WL2086 | 18°23.581'N; 077°14.094'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| " | WL3426 | 18°25.150'N; 077°16.452'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| " | WL2048 | 18°25.074'N; 077°16.641'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| " | WL2047 | 18°25.049'N; 077°16.674'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| " | WL2047A | 18°25.035'N; 077°16.677'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ8 |
| ABZ9 | WL2049 | 18°25.043'N; 077°16.549'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ9 |
| " | WL2049B | 18°25.035'N; 077°16.543'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ9 |
| " | WL2049C | 18°25.042'N; 077°16.549'W | Elgin Hall/Lilyfield | Preston Hill Fm | - | ABZ9 |
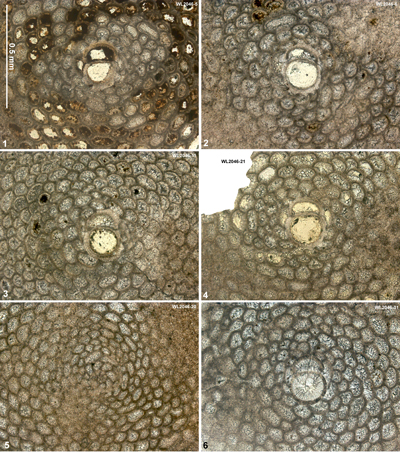
| " | WL2046A | 18°24.993'N; 077°16.692'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2046 | 18°24.992'N; 077°16.693'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2045A | 18°24.975'N; 077°16.700'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2045A | 18°24.975'N; 077°16.700'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2045C | 18°24.968'N; 077°16.701'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2044 | 18°24.944'N; 077°16.699'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL2041 | 18°24.800'N; 077°16.743'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
| " | WL4379 | 18°24.464'N; 077°15.792'W | Elgin Hall/Lilyfield | Chapelton Fm | - | ABZ9 |
Area 4. Swanswick and Clarks Town (North Coast Belt/Clarendon Block): Lutetian, Bartonian and Priabonian
This area is significant because it contains the type section of the Swanswick Formation of Hose and
Versey (1956
- see also Versey in Zans et al., 1963, where the type section is defined). Unfortunately, the faunas from the type Swanswick Formation do not correlate with the Swanswick Formation as mapped across much of Jamaica (e.g., Versey in Zans et al., 1963; Robinson & Mitchell, 1999; Mitchell, 2013a, 2016). The type Swanswick section contains two different stratigraphic intervals: the lower level consists of yellow-brown impure wackestones (a typical Yellow Limestone
lithology) and yields Helicostegina gyralis in association with Linderina floridensis and nummulitids (Wright & Switzer, 1971; Robinson & Mitchell, 1999; herein) and can be assigned to the early Lutetian (ABZ7); in contrast, the upper part of the succession consists of pale cream packstones and yields Heterostegina ocalana and very rare Fabularia verseyi allowing a correlation with ABZ16 (Fig. 4 ![]() ); an interval between consists of lime mudstones and is best assigned to the top of the Claremont Formation. Across much of the Clarendon Block, the white grainstones that have been mapped as Swanswick Formation occur between the Troy Formation
(dolostones and recrystallized limestones) and the Claremont Formation (predominately lime mudstones: Mitchell, 2004, 2013a), which yield an ABZ15 LBF assemblage. Thus the type Swanswick (here designated as
"Swanswick" s.s.) includes only early Lutetian (ABZ7) and late Priabonian (ABZ16) LBF with a probable fault (maybe
an unconformity) between, whereas the widely mapped Swanswick Formation (designated here as
"Swanswick" s.l.) yields a lower ABZ15 LBF assemblage. It is notable that to the south of Clarkes Town (SW of Swanswick itself) the Preston Hill Formation is exposed and yields late Lutetian LBF (ABZ10, herein) indicating that the Yellow Limestone in the lower part of the type Swanswick Formation is a limestone level developed within the Preston Hill Formation. A revision to the lithostratigraphy is required, and this will be published elsewhere. No nannofossil work was undertaken by us on these successions because of the numerous faults and the generally indurated nature of the
limestones. Samples are listed in Table 5.
); an interval between consists of lime mudstones and is best assigned to the top of the Claremont Formation. Across much of the Clarendon Block, the white grainstones that have been mapped as Swanswick Formation occur between the Troy Formation
(dolostones and recrystallized limestones) and the Claremont Formation (predominately lime mudstones: Mitchell, 2004, 2013a), which yield an ABZ15 LBF assemblage. Thus the type Swanswick (here designated as
"Swanswick" s.s.) includes only early Lutetian (ABZ7) and late Priabonian (ABZ16) LBF with a probable fault (maybe
an unconformity) between, whereas the widely mapped Swanswick Formation (designated here as
"Swanswick" s.l.) yields a lower ABZ15 LBF assemblage. It is notable that to the south of Clarkes Town (SW of Swanswick itself) the Preston Hill Formation is exposed and yields late Lutetian LBF (ABZ10, herein) indicating that the Yellow Limestone in the lower part of the type Swanswick Formation is a limestone level developed within the Preston Hill Formation. A revision to the lithostratigraphy is required, and this will be published elsewhere. No nannofossil work was undertaken by us on these successions because of the numerous faults and the generally indurated nature of the
limestones. Samples are listed in Table 5.
Table 5: Details of LBF and calcareous nannofossil zone for samples from area 4 (type Swanswick Formation).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
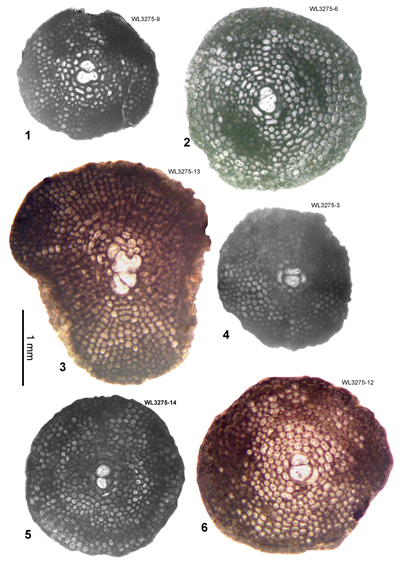
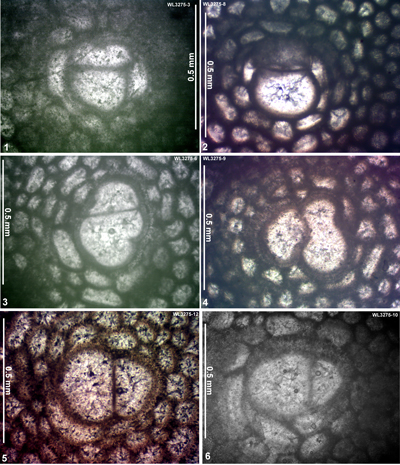
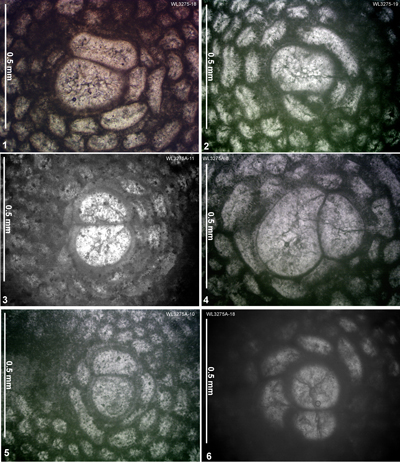
| ABZ7 | WL3275 | 18°24.982'N; 077°31.915'W | Swanswick House | Yellow Limestone | - | ABZ7 |
| " | WL3275A | 18°24.982'N; 077°31.915'W | Swanswick House | Yellow Limestone | - | ABZ7 |
| ABZ9 | WL3277 | 18°23.845'N; 077°32.206'W | Clarkes Town | Preston Hill | - | ABZ9 |
| ABZ16 | WL3265 | 18°24.917'N; 077°31.983'W | Swanswick House | Swanswick s.s. | - | ABZ16 |
Area 5. Sherwood Content (North Coast Belt, parish of Trelawny): Lutetian and Bartonian
Sherwood Content lies in the transition zone between the Clarendon Block and the North Coast Belt (Fig. 3 ![]() ) and faunas were described from here by Vaughan (1928) and Cole (1956). The succession consists of three units: the Preston Hill Formation, Chapelton Formation and Healthy Hill Formation. LBF assemblages indicate a succession through the Bartonian (Fig. 4
) and faunas were described from here by Vaughan (1928) and Cole (1956). The succession consists of three units: the Preston Hill Formation, Chapelton Formation and Healthy Hill Formation. LBF assemblages indicate a succession through the Bartonian (Fig. 4 ![]() ) with ABZ11, ABZ13 and ABZ14 represented. No nannofossil work was undertaken in this area because much of the succession here consists of indurated
limestones. Samples are indicated in Table 6.
) with ABZ11, ABZ13 and ABZ14 represented. No nannofossil work was undertaken in this area because much of the succession here consists of indurated
limestones. Samples are indicated in Table 6.
Table 6: Details of LBF and calcareous nannofossil zone for samples from area 5 (North Coast Belt, parish of Trelawny).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ11 | WL3958 | 18°24.464'N; 77°36.151'W | Sherwood Content | Chapelton | - | ABZ11 |
| ABZ13 | WL3772 | 18°23.912'N; 77°37.500'W | Sherwood Content | Chapelton | - | ABZ13 |
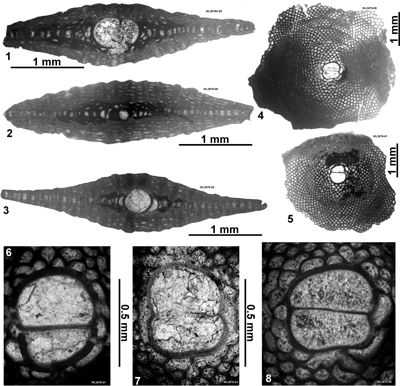
| ABZ15 | R.1068 | 18°24.043'N; 77°37.517W | Sherwood Content | Chapelton | - | ABZ14 |
| " | R.1072 | 18°24.070'N; 77°37.515'W | Sherwood Content | Chapelton | - | ABZ14 |
| " | WL3774 | 18°24.071'N; 77°37.514'W | Sherwood Content | Chapelton | - | ABZ15 |
Area 6. Spot Valley, Salt Spring and Flower Hill (North Coast Belt): Bartonian
Eocene rocks belonging to the Yellow Limestone and White Limestone groups occur in a structurally complex area to the SE of Montego Bay (Fig. 3 ![]() ) in what has been called the Montego Bay Anticline (Meyerhoff & Krieg, 1977). There is a transition here from marlstones of the Preston Hill Formation to the overlying chalks of the "Bonny Gate" Formation (Fig. 4
) in what has been called the Montego Bay Anticline (Meyerhoff & Krieg, 1977). There is a transition here from marlstones of the Preston Hill Formation to the overlying chalks of the "Bonny Gate" Formation (Fig. 4 ![]() ), which occurs in ABZ11. The overlying chalks of the "Bonny Gate" Formation have yielded LBF indicating ABZ12 (at Spots Valley) and ABZ14 (south of Flower Hill). Nannofossil samples were taken from the marlstone-chalk sequence in ABZ11 (indicating zone NP16 and CP14a) and from the chalks in ABZ12 at Spots Valley (which were barren). Correlations are shown in Fig. 4
), which occurs in ABZ11. The overlying chalks of the "Bonny Gate" Formation have yielded LBF indicating ABZ12 (at Spots Valley) and ABZ14 (south of Flower Hill). Nannofossil samples were taken from the marlstone-chalk sequence in ABZ11 (indicating zone NP16 and CP14a) and from the chalks in ABZ12 at Spots Valley (which were barren). Correlations are shown in Fig. 4 ![]() . Samples and LBF and nannofossil zones are listed in Table 7 and Appendix
2.
. Samples and LBF and nannofossil zones are listed in Table 7 and Appendix
2.
Table 7: Details of LBF and calcareous nannofossil zone for samples from area 6 (Spot Valley and Flower Hill, North Coast Belt, parish of St James).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
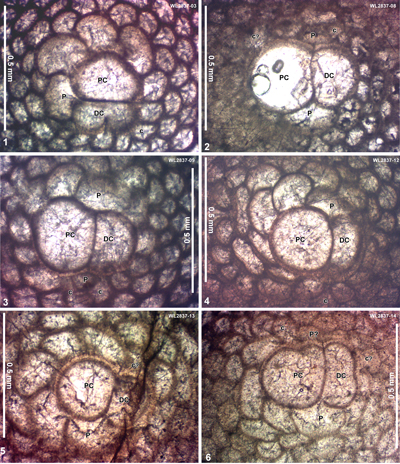
| ABZ11 | WL2837 | 18°29.525'N; 77°51.791'W | Salt Spring | Preston Hill | - | ABZ11 |
| " | WL4234 | 18°29.526'N; 77°51.793'W | Salt Spring | Preston Hill | Barren | ABZ11 |
| " | WL2836 | 18°29.627'N; 77°51.622'W | Salt Spring | 'Bonny Gate' | - | ABZ11 |
| " | WL4236 | 18°29.679'N; 77°51.373'W | Salt Spring | 'Bonny Gate' | NP16 | ABZ11 |
| " | WL4237 | 18°29.621'N; 77°51.203'W | Salt Spring | 'Bonny Gate' | NP16 | ABZ11 |
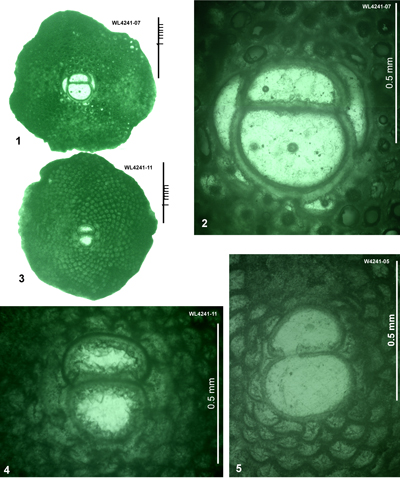
| ABZ12 | WL4241 | 18°30.401'N; 77°47.827'W | Spot Valley | 'Bonny Gate' | Barren | ABZ12 |
| " | WL4243 | 18°30.422'N; 77°47.710'W | Spot Valley | 'Bonny Gate' | Barren | ABZ12 |
| ABZ14 | WL2819 | 18°29.135'N; 77°50.385'W | Flower Hill | 'Bonny Gate' | - | ABZ14 |
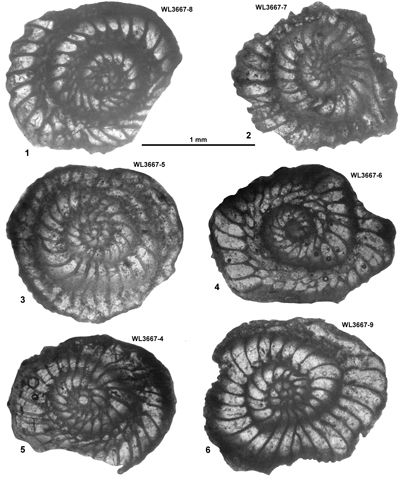
Area 7. Yellow Limestone around Albert Town and Litchfield (Clarendon Block): late Lutetian and early Bartonian
The Yellow Limestone around the north-western margin of the Central Inlier (Mitchell, 2013b; Fig. 3 ![]() ) consists of four formations: Freemans Hall,
Stettin, Litchfield and Chapelton formations (Fig. 4
) consists of four formations: Freemans Hall,
Stettin, Litchfield and Chapelton formations (Fig. 4 ![]() ) (Robinson, 1996; Robinson & Mitchell, 1999; Mitchell, 2016). Samples from the Stettin Formation yield H. minor in the absence of nummulitids and can be assigned to the earliest Lutetian lower ABZ6 (Fig. 4
) (Robinson, 1996; Robinson & Mitchell, 1999; Mitchell, 2016). Samples from the Stettin Formation yield H. minor in the absence of nummulitids and can be assigned to the earliest Lutetian lower ABZ6 (Fig. 4 ![]() ). The overlying Litchfield Formation lacks zonally significant
LBF, but the Chapelton Formation contains LBF indicating zones ABZ9 and ABZ11, indicating a hiatus representing ABZ10 (Fig. 4
). The overlying Litchfield Formation lacks zonally significant
LBF, but the Chapelton Formation contains LBF indicating zones ABZ9 and ABZ11, indicating a hiatus representing ABZ10 (Fig. 4 ![]() ). Some nannofossil work on the Chapelton Formation has previously been presented by Robinson (1996) for this area and indicates zone NP16. Samples are detailed in Table 8.
). Some nannofossil work on the Chapelton Formation has previously been presented by Robinson (1996) for this area and indicates zone NP16. Samples are detailed in Table 8.
Table 8: Details of LBF and calcareous nannofossil zone for samples from area 7 (Albert Town and Litchfield, Clarendon Block, parish of Trelawny).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
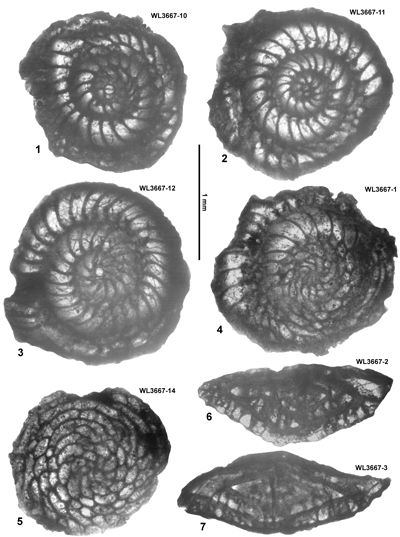
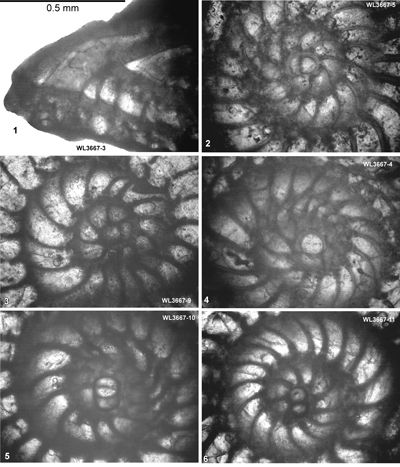
| ABZ6 | WL3666 | 18°17.225'N; 77°31.959'W | Stettin | Stettin | - | ABZ6 |
| " | WL3667 | 18°17.318'N; 77°31.886'W | Stettin | Stettin | - | ABZ6 |
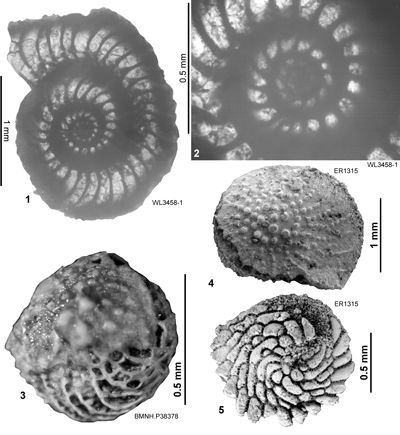
| " | WL3458 | 18°12.508'N; 77°30.051'W | Coleyville | Stettin | - | ABZ6 |
| ABZ9 | WL3444 | 18°17.519'N; 77°32.634'W | Albert Town | Chapelton | - | ABZ9 |
| ABZ11 | WL3445 | 18°17.566'N; 77°32.780'W | Albert Town | Chapelton | - | ABZ11 |
| " | ER1356 | c. 18°17.66'N; 77°32.85'W | Albert Town | Chapelton | NP16 * | ABZ11 |
| " | WL3447 | 18°17.659'N; 77°32.853'W | Albert Town | Chapelton | - | ABZ11 |
| " | WL4195 | 18°15.271'N; 77°29.995'W | Litchfield | Chapelton | - | ABZ11 |
* Data from Robinson (1996).
Area 8. Yellow Limestone at Coleyville (Clarendon Block): Bartonian
The area around Coleyville (Fig. 3 ![]() ) has a similar lithostratigraphy to that of the area around Albert Town and Litchfield, although the LBF assemblages are different (further from the platform edge). Zonal indicators are largely absent (except for very rare H. minor in the Stettin Formation) until the upper part of the Chapelton Formation. The upper part of the Chapelton Formation at the Forest Reserve yields abundant lepidocyclinids indicating ABZ13. No nannofossil work was undertaken here and correlations are shown in Fig. 4
) has a similar lithostratigraphy to that of the area around Albert Town and Litchfield, although the LBF assemblages are different (further from the platform edge). Zonal indicators are largely absent (except for very rare H. minor in the Stettin Formation) until the upper part of the Chapelton Formation. The upper part of the Chapelton Formation at the Forest Reserve yields abundant lepidocyclinids indicating ABZ13. No nannofossil work was undertaken here and correlations are shown in Fig. 4 ![]() . Samples are described in Table 9.
. Samples are described in Table 9.
Table 9: Details of LBF and calcareous nannofossil zone for samples from area 8 (Coleyville, Clarendon Block, parish of Manchester).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ6 | WL3458 | 18°12.508'N; 77°30.051'W | Coleyville | Stettin | - | ABZ6 |
| ABZ13 | WL4109 | 18°11.570'N; 77°30.798'W | Coleyville | Chapelton | - | ABZ13 |
Area 9. Western side of the Clarendon Block, St Elizabeth and St James: Lutetian and Bartonian
Rocks of the Yellow Limestone Group occur in a series of inliers in northern St Elizabeth and southern St James where the rocks of the White Limestone Group have been eroded (Fig. 3 ![]() ). The succession has similarities with the succession around the Central
Inlier, but the Bartonian has thickened substantially and can be mapped in more detail. The formations present here need revision, but are currently assigned to the Stettin Formation, Helicostegina-beds, Litchfield Formation, Chapelton Formation, Healthy Hill Formation, and
'Ipswich' Formation (Versey, 1957; Atkinson, 1969). The
'Ipswich' Formation can be divided into three distinct mappable units: a lower unit of pale, thickly-bedded wackestone with a platform-type LBF assemblage; a series of brown and locally red packstones and grainstones carrying a lepidocyclinid assemblage (the type Ipswich Formation of Versey, 1957), and an upper unit passing back into wackestones with some lepidocyclinids and platform interior
LBF.
). The succession has similarities with the succession around the Central
Inlier, but the Bartonian has thickened substantially and can be mapped in more detail. The formations present here need revision, but are currently assigned to the Stettin Formation, Helicostegina-beds, Litchfield Formation, Chapelton Formation, Healthy Hill Formation, and
'Ipswich' Formation (Versey, 1957; Atkinson, 1969). The
'Ipswich' Formation can be divided into three distinct mappable units: a lower unit of pale, thickly-bedded wackestone with a platform-type LBF assemblage; a series of brown and locally red packstones and grainstones carrying a lepidocyclinid assemblage (the type Ipswich Formation of Versey, 1957), and an upper unit passing back into wackestones with some lepidocyclinids and platform interior
LBF.
ABZ6 is found in the Stettin and basal part of the Helicostegina-beds, zones ABZ7
- ABZ8 in the middle and upper parts of the Helicostegina-beds, ABZ10 in the Chapelton Formation, and zones ABZ13 to ABZ14 are represented in the middle and upper
'Ipswich' Formation (Fig. 4 ![]() ). The Yellow Limestone is overlain by a typical lower White Limestone succession which includes the Troy,
'Swanswick' sensu lato, Claremont and Somerset formations (Fig. 4
). The Yellow Limestone is overlain by a typical lower White Limestone succession which includes the Troy,
'Swanswick' sensu lato, Claremont and Somerset formations (Fig. 4 ![]() ). This demonstrates that the Ipswich Formation is demonstrably older than the Swanswick Formation, although the two were mapped as a single unit by the Jamaican Geological Survey (Bateson, 1972). Limited nannofossil data has been provided by Robinson et al. (2018). Samples are listed in Table 10.
). This demonstrates that the Ipswich Formation is demonstrably older than the Swanswick Formation, although the two were mapped as a single unit by the Jamaican Geological Survey (Bateson, 1972). Limited nannofossil data has been provided by Robinson et al. (2018). Samples are listed in Table 10.
Table 10: Details of LBF and calcareous nannofossil zone for samples from area 9 (western Clarendon Block, parishes of St James and St Elizabeth).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
| ABZ6 | WL3520 | 18°22.470'N; 077°51.220'W | Springvale | Yellow Lmst | - | ABZ6 |
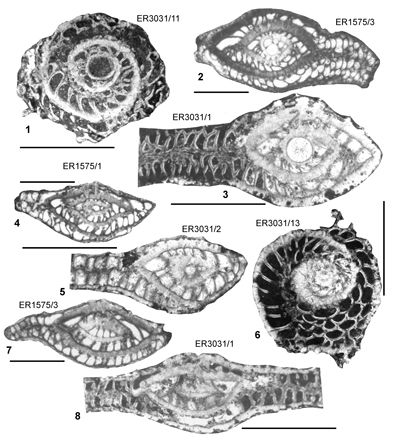
| ABZ9 | ER1575 | c. 18°21.54'N; 77°49.11'W | Point | Yellow Lmst | - | ABZ9 |
| " | ER3031 | c. 18°21.54'N; 77°49.11'W | Point | Yellow Lmst | - | ABZ9 |
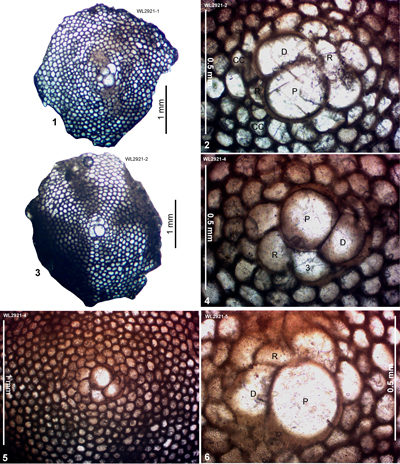
| ABZ10 | WL2921 | 18°09.880'N; 77°44.270'W | Elderslie | Chapelton | - | ABZ10 |
| ABZ13 | WL2564 | 18°12.019'N; 77°50.771'W | Ipswich | Ipswich | - | ABZ13 |
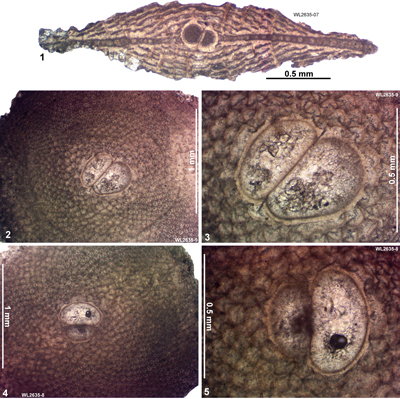
| " | WL2635 | 18°11.603'N; 77°49.246'W | Ipswich | Ipswich | - | ABZ13 |
| " | WL3501 | 18°11.631'N; 77°48.959'W | Railway cutting | Ipswich | - | ABZ13 |
| " | WL3503 | 18°11.700'N; 77°49.354'W | Railway cutting | Ipswich | - | ABZ13 |
| " | WL3507 | 18°09.880'N; 77°44.270'W | Appleton | Ipswich | - | ABZ13 |
Area 10. Grants Pen, St Andrew (SW Wag Water Belt): Bartonian
The Grants Pen area of St Thomas (Fig. 3 ![]() ) is a complex area which has overturned beds and probably significant faulting. The area contains a suite of Yellow Limestone marlstones that are overlain by chalks with at least one chert band. The area is important because it yields
LBF. Pseudolepidina trimera occurs in a hard limestone that cannot be dated using calcareous
nannofossils; whereas Lepidocyclina ariana (ABZ12) occurs in a Yellow Limestone marlstone which has been chronostratigraphically calibrated using nannofossils to zone NP16 in this work (Appendix
2). Correlations are shown in Fig. 4
) is a complex area which has overturned beds and probably significant faulting. The area contains a suite of Yellow Limestone marlstones that are overlain by chalks with at least one chert band. The area is important because it yields
LBF. Pseudolepidina trimera occurs in a hard limestone that cannot be dated using calcareous
nannofossils; whereas Lepidocyclina ariana (ABZ12) occurs in a Yellow Limestone marlstone which has been chronostratigraphically calibrated using nannofossils to zone NP16 in this work (Appendix
2). Correlations are shown in Fig. 4 ![]() , and samples are listed in Table 11.
, and samples are listed in Table 11.
Table 11: Details of LBF and calcareous nannofossil zone for samples from area 10 (Grants Pen, parish of St Thomas).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
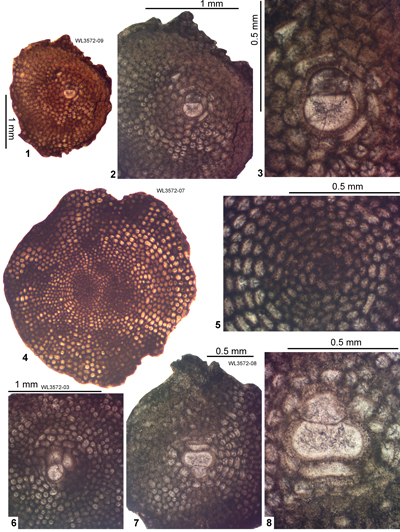
| ABZ7 | WL3572 | 17°54.730'N; 76°38.107'W | Grants Pen | Yellow Lmst | - | ABZ7 |
| " | WL3714 | 17°54.732'N; 76°38.107'W | Grants Pen | Yellow Lmst | - | ABZ7 |
| " | WL3715 | 17°54.784'N; 76°38.213'W | Grants Pen | Yellow Lmst | - | ABZ7 |
| " | WL4631 | 17°54.749'N; 77°38.113'W | Grants Pen | Yellow Lmst | - | ABZ7 |
| " | WL4631 | 17°54.753'N; 76°38.114'W | Grants Pen | Yellow Lmst | - | ABZ7 |
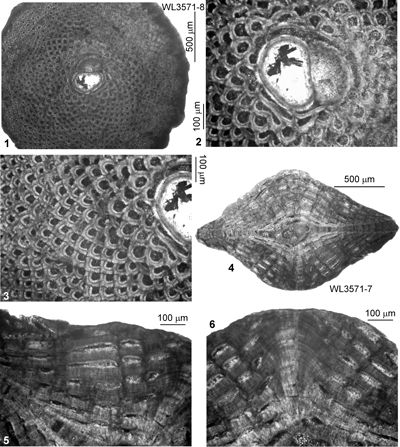
| ABZ12 | WL3571 | 17°54.590'N; 76°37.906'W | Grants Pen | Yellow Lmst | - | ABZ12 |
| " | WL3711 | 17°54.589'N; 76°37.907'W | Grants Pen | Yellow Lmst | - | ABZ12 |
| " | WL4628 | 17°54.540'N; 77°36.888'W | Grants Pen | Yellow Lmst | NP17 | Barren |
| " | WL4629 | 17°54.589'N; 77°37.904'W | Grants Pen | Yellow Lmst | Barren | ABZ12 |
| " | WL4630 | 17°54.589'N; 77°37.904'W | Grants Pen | Yellow Lmst | NP16 | ABZ12 |
Area 11. Highway 2000 around the Golden Vale Junction, parish of St. Ann (Clarendon Block): Priabonian
The road cuts created during the construction of the North-South Leg of Highway 2000 in the parish of St Ann (Fig. 3 ![]() ) showed good sections through the Swanswick
s.l., Claremont, Somerset, and Walderston Formations. The lower three of these were composed of relatively weakly cemented limestones from which free specimens of LBF could be extracted. These sections were close to the shelf edge in Late Eocene time and contain rich lepidocyclinid assemblages. These were used to understand the succession of lepidocyclinids through the
Priabonian, with zones ABZ15 and ABZ16 represented (Fig. 4
) showed good sections through the Swanswick
s.l., Claremont, Somerset, and Walderston Formations. The lower three of these were composed of relatively weakly cemented limestones from which free specimens of LBF could be extracted. These sections were close to the shelf edge in Late Eocene time and contain rich lepidocyclinid assemblages. These were used to understand the succession of lepidocyclinids through the
Priabonian, with zones ABZ15 and ABZ16 represented (Fig. 4 ![]() ). No nannofossil work was undertaken on these samples. Samples are described in Table 12.
). No nannofossil work was undertaken on these samples. Samples are described in Table 12.
Table 12: Details of LBF and calcareous nannofossil zone for samples from area 11 (road cuts along northern N-S leg of Highway 2000).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
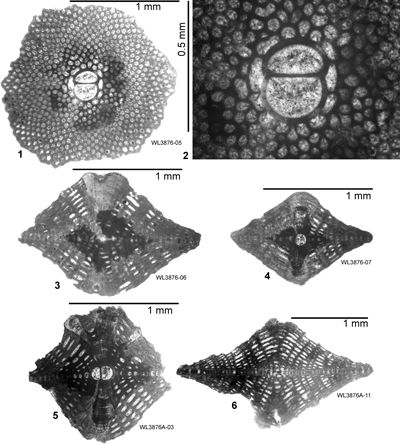
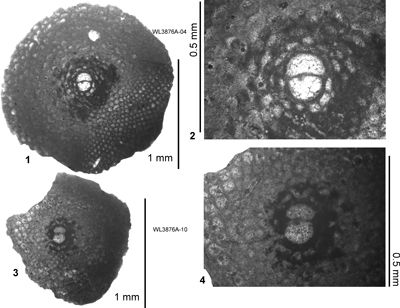
| ABZ15 | WL3876 | 18°21.538'N; 77°09.500'W | Highway2000 | 'Swanswick s.l.' | - | ABZ15 |
| " | WL3876A | 18°21.538'N; 77°09.500'W | Highway2000 | 'Swanswick s.l.' | - | ABZ15 |
| " | WL4182 | 18°22.800'N; 77°09.517'W | Highway2000 | 'Swanswick s.l.' | - | ABZ15 |
| " | WL4095 | 18°20.790'N; 77°08.917'W | Highway2000 | Claremont | - | ABZ16 |
| ABZ16 | WL3931 | 18°19.905'N; 77°08.525'W | Highway2000 | Somerset | - | ABZ16 |
Area 12. The central and southern Montpelier-New Market Belt: late Bartonian to Priabonian
The succession in the Montpelier-New Market Belt (Fig. 3 ![]() ) covers the interval from the late Lutetian to the
Priabonian. Samples were used from the Content-1 well which penetrated the lower part of the succession
(Chapelton Formation and overlying marlstones) and from surface outcrops. Surface outcrops can be divided into five Eocene units: Chapelton Formation, marlstones, lower unit of chalks, unit of chalks with
cherts, and upper unit of chalks. New road cuts associated with the improvement work on the Mackfield to Ferris Cross main road facilitated the collection of samples from weakly
indurated, freshly exposed rocks. A series of nannofossil samples was analysed from here from the late Bartonian into the
mid-Priabonian. ABZ10 (samples from the Content Well: Eva, 1980), ABZ14, ABZ15 and ABZ16 are represented in this area (Fig. 4
) covers the interval from the late Lutetian to the
Priabonian. Samples were used from the Content-1 well which penetrated the lower part of the succession
(Chapelton Formation and overlying marlstones) and from surface outcrops. Surface outcrops can be divided into five Eocene units: Chapelton Formation, marlstones, lower unit of chalks, unit of chalks with
cherts, and upper unit of chalks. New road cuts associated with the improvement work on the Mackfield to Ferris Cross main road facilitated the collection of samples from weakly
indurated, freshly exposed rocks. A series of nannofossil samples was analysed from here from the late Bartonian into the
mid-Priabonian. ABZ10 (samples from the Content Well: Eva, 1980), ABZ14, ABZ15 and ABZ16 are represented in this area (Fig. 4 ![]() ). Samples are listed in Table 13 and nannofossil records are included in Appendix
2.
). Samples are listed in Table 13 and nannofossil records are included in Appendix
2.
Table 13: Details of LBF and calcareous nannofossil zone for samples from area 12 (Montpelier-New Market Belt).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
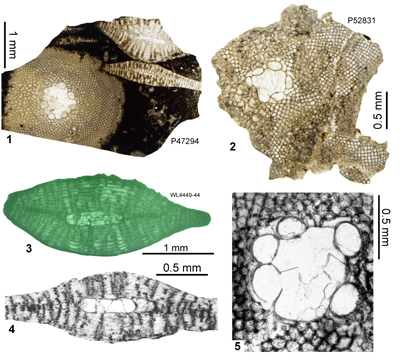
| ABZ14 | WL4444 | 18°16.742'N; 77°59.423'W | Cold Spring | Marlstones | - | ABZ14 |
| " | WL4445 | 18°16.725'N; 77°59.440'W | Cold Spring | 'Bonny Gate' | NP17 | |
| " | WL4449 | 18°16.697'N; 77°59.636'W | Cold Spring | 'Bonny Gate' | - | ABZ14 |
| ABZ15 | WL4618 | 18°18.267'N; 78°01.061'W | Haddo | 'Bonny Gate' | NP18 | ABZ15 |
| " | WL4620 | 18°18.172'N; 78°01.122'W | Haddo | 'Bonny Gate' | NP18 | ABZ15 |
| " | WL4621 | 18°17.887'N; 78°01.275'W | Haddo | 'Bonny Gate' | NP18 | ABZ15 |
| " | WL4622 | 18°17.864'N; 78°01.285'W | Haddo | 'Bonny Gate' | NP18 | ABZ15 |
| ABZ16 | WL4602 | 18°11.493'N; 78°00.770'W | Bath | 'Bonny Gate' | NP19 | ABZ16 |
| " | WL4624A | 18°16.938'N; 78°01.605'W | Line Gate | 'Bonny Gate' | NP19 | ABZ16 |
| " | WL4617 | 18°18.383'N; 78°00.957'W | Mackfield | 'Bonny Gate' | NP19 | ABZ16 |
| " | WL4616 | 18°18.735'N; 78°00.795'W | Mackfield | 'Bonny Gate' | NP19 | ABZ16 |
Area 13. Jerusalem Mountain - Upper Rock Spring Area (parish of Westmoreland)
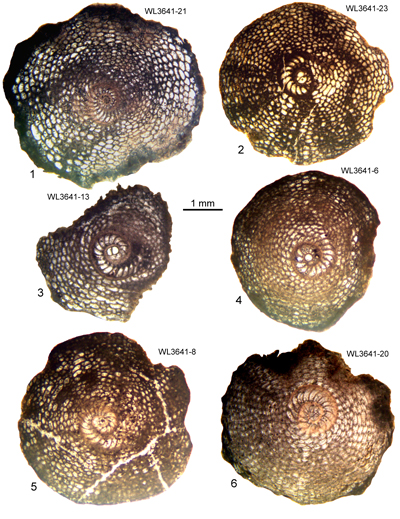
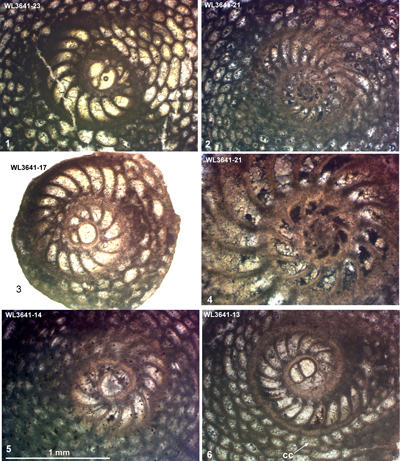
The Chapelton Formation is exposed around Rock Spring to the north of the Jerusalem Mountain Inlier (Fig. 3 ![]() ; Mitchell & Edwards, 2016). The Chapelton Formation here yields abundant free specimens of Helicostegina jeannemairae associated with Operculinoides sp. and can be assigned to ABZ9 (Fig. 4
; Mitchell & Edwards, 2016). The Chapelton Formation here yields abundant free specimens of Helicostegina jeannemairae associated with Operculinoides sp. and can be assigned to ABZ9 (Fig. 4 ![]() ). No nannofossil work was carried out here, and the sample is listed in Table 14.
). No nannofossil work was carried out here, and the sample is listed in Table 14.
Table 14: Details of LBF and calcareous nannofossil zone for the sample from area 13 (Jerusalem Mountain - Upper Rock Spring, parish of Westmoreland).
| Zone | Sample | Lat/Long | Location | Formation | Nanno zone | LBF zone |
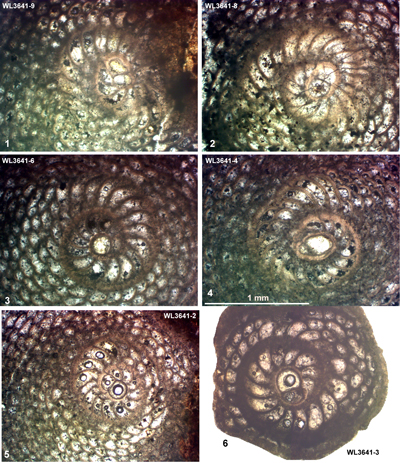
| ABZ9 | WL3641 | 18°20.757'N; 77°13.870'W | Rock Spring | Chapelton | - | ABZ9 |
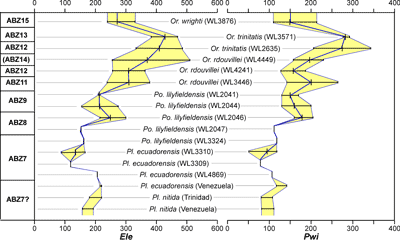
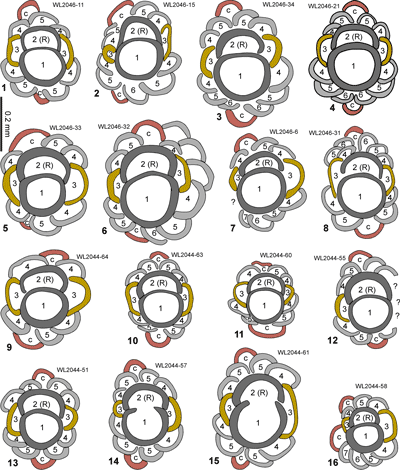
We develop a zonation scheme for the Eocene of the American biogeographic province based on the stratigraphic distribution of selected
taxa. The zonation is based on the first appearance datum (FAD) and last appearance datum (LAD) of selected
taxa. Planktic foraminiferal zones are indicated by the prefixes
'P' (Berggren et al., 1995) and 'E' (Pearson et al., 2006) and calcareous nannofossil zones by the prefixes
'NP' (Martini, 1971), 'CN' (Okada & Bukry, 1980) and
'CNE' (Agnini et al., 2014). The American Benthic Zones
(ABZs) are defined below, and their ages are constrained largely using calcareous nannofossils related to the
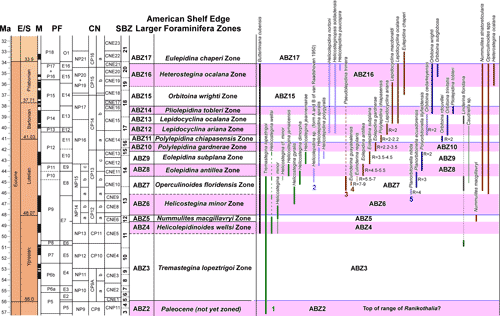
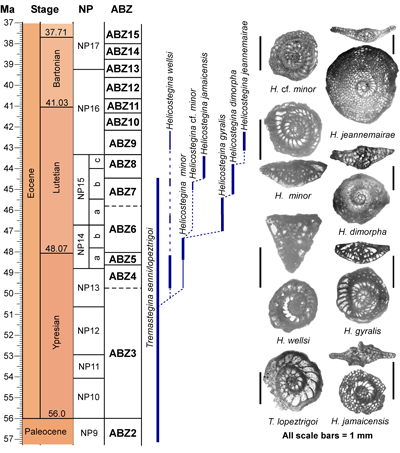
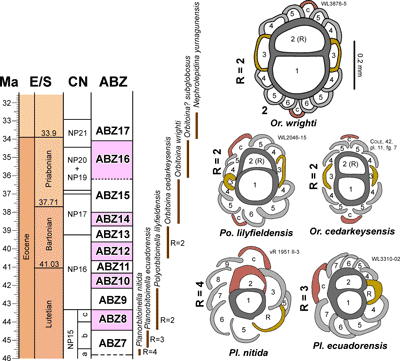
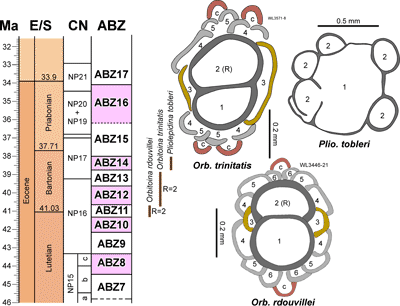
'NP' zonation. The zonation scheme for the Eocene is shown in Fig. 6 ![]() and consists of 14 zones (ABZ3 to ABZ16). The fourteen Eocene zone are defined below, and the zonation for the Paleocene, Oligocene and early to middle Miocene will be presented in a separate paper.
and consists of 14 zones (ABZ3 to ABZ16). The fourteen Eocene zone are defined below, and the zonation for the Paleocene, Oligocene and early to middle Miocene will be presented in a separate paper.
| Figure 6:
Distribution of selected Larger Benthic Foraminifers in relation to the Eocene
zonation scheme calibrated against planktic foraminifers and calcareous
nannofossil zonations. The Standard Benthic zonation (SBZ) scheme for the
Tethys is shown for comparison (Serra-Kiel et al.,
1998; Speijer et al., 2020). |
ABZ3. Tremastegina lopeztrigoi Zone. From the LAD of Ranikothalia catenula (which defines ABZ2) to the FAD of H. wellsi. Small specimens tentatively assigned to T. lopeztrigoi are found in the Nonsuch Formation of Jamaica where they co-occur with Ranikothalia (Robinson & Jiang, 1990; Robinson & Wright, 1993). ABZ3 is
recognized by the presence of T. lopeztrigoi, in the absence of either Ranikothalia (ABZ2) or Helicostegina wellsi (ABZ4). The associated LBF assemblage is
characterised by long-ranging forms, particularly
orthophragmines, such as Orbitoclypeus, Stenocyclina, Pseudophragmina and Asterocyclina (see Özcan et al., 2019,
2022a, for a revision of the
orthophragmines). The reference section for the zone is in the Richmond Formation near Albany, northern Wag Water Belt, Jamaica (Fig. 3 ![]() ), and is assigned to NP12 (Fig. 6
), and is assigned to NP12 (Fig. 6 ![]() ; Jiang & Robinson, 1987). Blanco-Bustamentel et al. (1999) suggested that Ranikothalia and Eoconuloides (= Helicostegina herein) wellsi occurred throughout the lower Eocene, but these results have not been proved elsewhere and a reassessment of these successions in Cuba is needed.
; Jiang & Robinson, 1987). Blanco-Bustamentel et al. (1999) suggested that Ranikothalia and Eoconuloides (= Helicostegina herein) wellsi occurred throughout the lower Eocene, but these results have not been proved elsewhere and a reassessment of these successions in Cuba is needed.
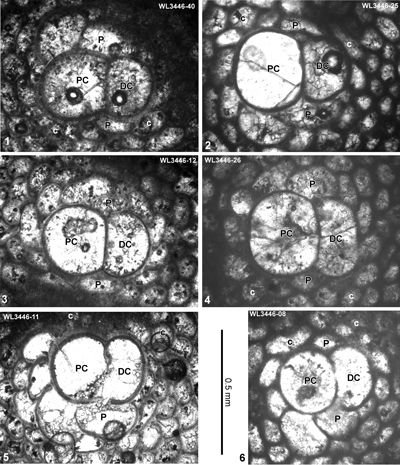
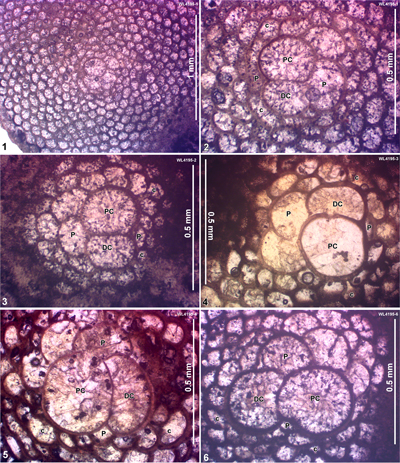
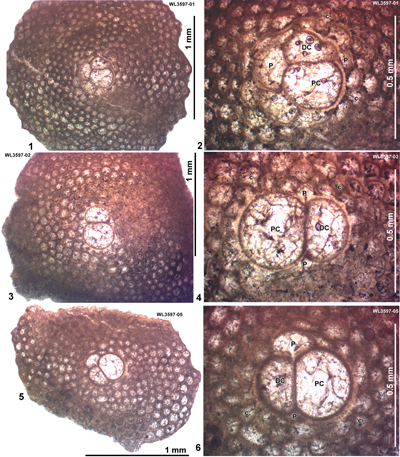
ABZ4. Helicostegina wellsi Zone. From the FAD of H. wellsi to the FAD of Nephronummulites macgillavryi. In shelf edge localities, the LBF of this zone are dominated by H. wellsi and T. lopeztrigoi, locally with H. minor, Fallotella and Cushmania. In the deeper shelf succession of the Richmond Formation the zone contains common H. wellsi and T. lopeztrigoi along with a high-diversity LBF assemblage including orthophragmines (Asterocyclina, Stenocyclina, Pseudophragmina and Orbitoclypeus), Fabiania, Cushmania, Fallotella and Eorupertia, and locally H. minor. The reference locality for the zone is in the Rio Sambre section, parish of St Mary, Jamaica (Robinson & Mitchell, 1999; Fig. 5). Our nannofossil determinations indicate that ABZ4 belongs to the lower part of Zone NP14a (Fig. 6 ![]() ; Appendix 1). This is consistent with previous age assignments (NP14 and P9: Wise & Constans, 1976; Robinson & Mitchell, 1999). We tentatively place the boundary between ABZ3 and ABZ4 within Zone NP13 as we do not have LBF assemblages indicating Zone NP13. It is unclear if NP13 is absent due to a hiatus, or, more likely, cut out by faulting.
; Appendix 1). This is consistent with previous age assignments (NP14 and P9: Wise & Constans, 1976; Robinson & Mitchell, 1999). We tentatively place the boundary between ABZ3 and ABZ4 within Zone NP13 as we do not have LBF assemblages indicating Zone NP13. It is unclear if NP13 is absent due to a hiatus, or, more likely, cut out by faulting.
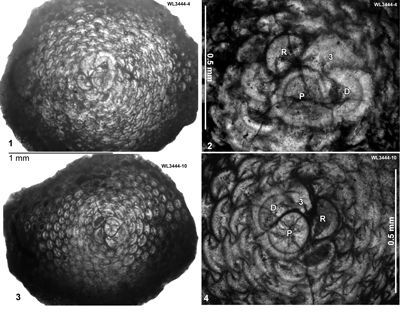
The occurrence of H. wellsi (as Amphistegina elliotti) in Ecuador as recorded by Cushman and Stainforth (1946) is a typical shallow-water occurrence, where H. wellsi is the only species present. H. wellsi is a distinctive species and can easily be recognized in random thin sections. The presence of Tremostegina senni and H. minor in the absence of Operculinoides (Senn, 1940; Caudri, 1972; Jones, 2009) in the Murphy's Beds of Barbados (Upper Scotland Group) suggests assignment to ABZ4 (or possibly lower ABZ6).
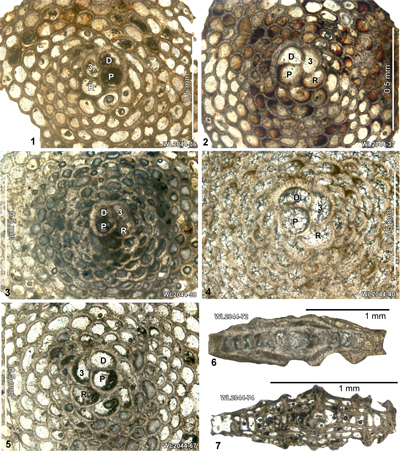
ABZ5. Nephronummulites macgillavryi Zone. From the FAD to LAD of N. macgillavryi. The reference section lies in the upper part of the Richmond Formation and lower part of the Palmetto Grove Formation in the Rio Sambre Gorge, parish of St Mary, Jamaica (Figs. 5 - 6). In the lower part of the zone, N. macgillavryi co-occurs with H. wellsi, whereas H. minor occurs throughout the zone. The zone is here correlated with the upper part of the Ypresian (upper Zone NP14a below the entry of the nannofossil Blackites inflatus in Zone NP14b (Figs. 5 - 6). The zone is recognized in Cuba (Rutten, 1935), Jamaica (Robinson & Wright, 1993; herein), and Nicaragua (Andji et al., 2018). Many assemblages are characterised only by N. macgillavryi (as reported by Rutten, 1935; Robinson & Wright, 1993; Andji et al., 2018), and these probably represent shelf edge assemblages lacking the deeper-water orthophragmine component of the fauna. N. macgillavryi is a distinctive species and can easily be recognized in hand specimen and in random thin sections.
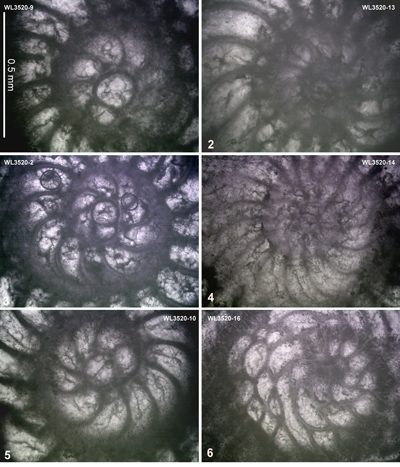
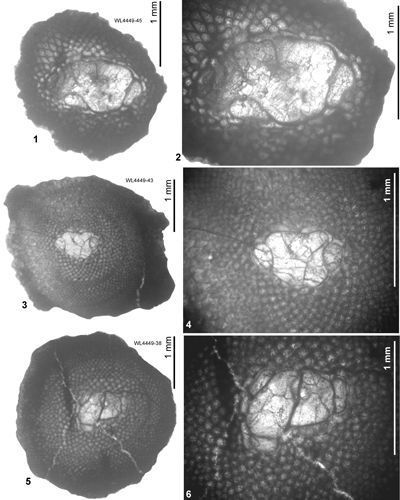
ABZ6. Helicostegina minor Zone. From the LAD of N. macgillavryi to the FAD of Operculinoides
spp. This is a low diversity interval above the Lower-Middle Eocene boundary, and amongst the Asterogerinoidea only Helicostegina and T. lopeztrigoi are present; longer ranging taxa include orthophragmines (including Stenocyclina advena and Asterocyclina
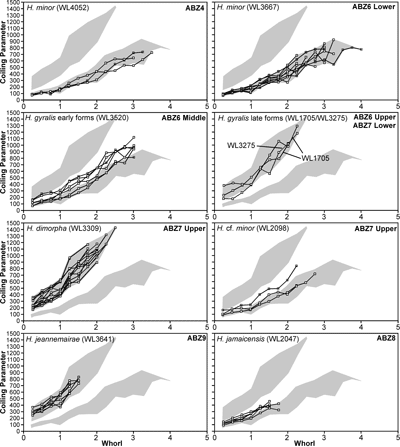
spp.), Fallotella and Cushmania. There is a succession of species of Helicostegina, H. minor occurs in the lower part of the zone and H. gyralis in the upper part of the zone (Fig. 6 ![]() ) and thus it can be divided into lower (ABZ6A) and upper (ABZ6B) parts. The reference locality for the zone lies in the upper part of the Palmetto Grove Formation above the Rio Sambre in the parish of St Mary, Jamaica. The zone also occurs in shelf edge assemblages where H. minor occurs in association with inner platform LBF (Verseyella jamaicensis (Cole, 1956), Coleiconus zansi Robinson, 1993, and Fabularia colei Robinson, 1969) in the Stettin Formation of the Central Inlier (Robinson & Wright, 1993; Robinson & Mitchell, 1999). In the Rio Sambre section, the entire zone yields the nannofossil Blackites inflatus indicating Zone NP14b (Fig. 6
) and thus it can be divided into lower (ABZ6A) and upper (ABZ6B) parts. The reference locality for the zone lies in the upper part of the Palmetto Grove Formation above the Rio Sambre in the parish of St Mary, Jamaica. The zone also occurs in shelf edge assemblages where H. minor occurs in association with inner platform LBF (Verseyella jamaicensis (Cole, 1956), Coleiconus zansi Robinson, 1993, and Fabularia colei Robinson, 1969) in the Stettin Formation of the Central Inlier (Robinson & Wright, 1993; Robinson & Mitchell, 1999). In the Rio Sambre section, the entire zone yields the nannofossil Blackites inflatus indicating Zone NP14b (Fig. 6 ![]() ; Appendix 1). Zone NP15a has not been recorded in samples from transects in this area, nor elsewhere in Jamaica, and there appears to be a minor hiatus (representing zone NP15a) between ABZ6 and ABZ7 in Jamaica.
; Appendix 1). Zone NP15a has not been recorded in samples from transects in this area, nor elsewhere in Jamaica, and there appears to be a minor hiatus (representing zone NP15a) between ABZ6 and ABZ7 in Jamaica.
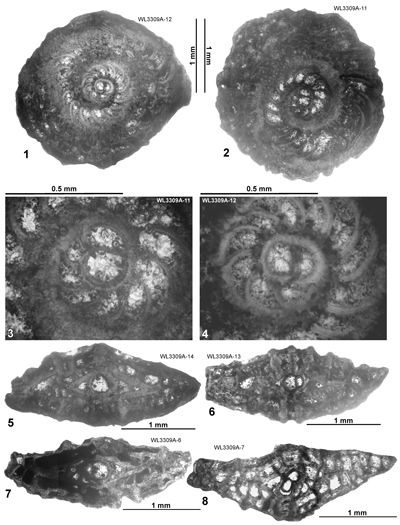
In Cuba, Cole and Bermúdez (1944) recorded an identical assemblage of LBFs from the Peńon Formation, which we can assign to the upper part of ABZ6 based on the presence of H. gyralis in the absence of Operculinoides spp. Torres-Silva et al. (2018) recorded a second assemblage from the Peńon Formation in Cuba in which Operculinoides spp. occurred with H. wellsi (reported as E. wellsi), which we would place in ABZ7.
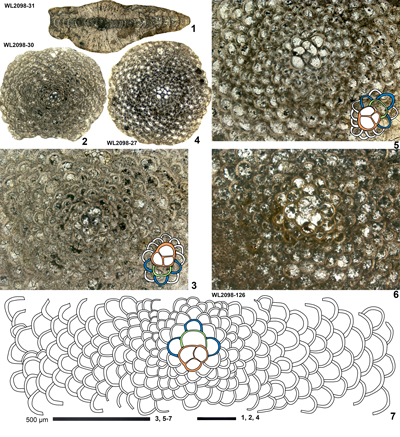
ABZ7. Operculinoides spp. Zone. From the FAD of Operculinoides ex gr. floridensis and Nummulites ex gr. striatoreticulatus to the FAD of Eolepidina antillea (with
R < 5.5). The reference section lies on the road from Farm Turn to Hampstead, parish of St Mary, Jamaica. The zone sees the appearance of numerous taxa including Eulinderina guayabalensis, Planorbitoinella ecuadorensis and Pseudolepidina trimera suggesting an important migration event at this time. The area from where these taxa migrated from is unknown, but could be in the Pacific, and it represents the most significant faunal change in the Eocene of the Americas. Helicostegina gyralis occurs in the lower part of the zone, whereas H. dimorpha occurs in the upper part of the zone. The appearance of Operculinoides ex gr. floridensis (e.g., Op. jennyi
Barker) and Nummulites striatoreticulatus (e.g., N. guayabalensis
Barker) in large numbers is particularly conspicuous and the genera can easily be recognized in random thin sections. In the Rio Sambre area Operculinoides ex gr. floridensis appears at the base of Zone NP15b, but Zone NP15a is missing. We tentatively put the ABZ6/ABZ7 boundary within Zone NP15a (Fig. 6 ![]() ), pending further research. In Mexico, Eu. guayabalensis occurs in association with Operculinoides prenummulitiformis and Nummulites guayabalensis (Barker, 1939, p. 325) indicating ABZ7. The association of H. wellsi and T. senni (cited as Eoconuloides senni) with Operculinoides sp. in Calyx wells 50 and 50A in Trinidad (Caudri,
1996, Fig. 7
), pending further research. In Mexico, Eu. guayabalensis occurs in association with Operculinoides prenummulitiformis and Nummulites guayabalensis (Barker, 1939, p. 325) indicating ABZ7. The association of H. wellsi and T. senni (cited as Eoconuloides senni) with Operculinoides sp. in Calyx wells 50 and 50A in Trinidad (Caudri,
1996, Fig. 7 ![]() ) also suggests assignment to ABZ7.
) also suggests assignment to ABZ7.
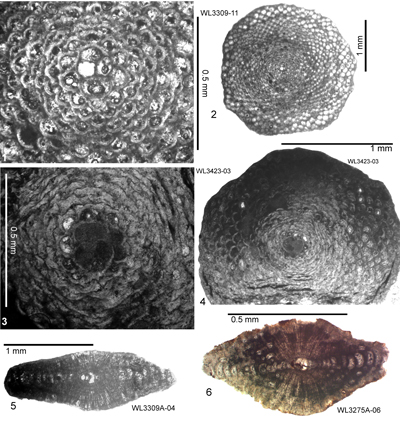
ABZ8. Eolepidina antillea Zone. From the FAD of Eo. antillea (with
R < 5.5) to the FAD of Eo. subplana (with R <
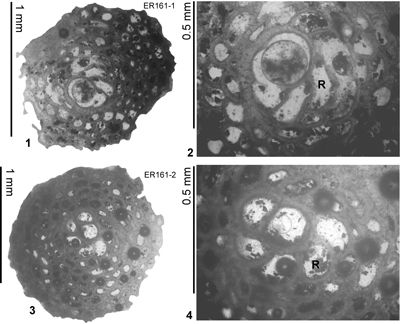
4.5). The reference section lies on the road from Farm Turn to Hampstead, parish of St Mary, Jamaica. At Farm Turn (Rio Sambre area) the transition from ABZ7 to ABZ8 occurs in the upper part of Zone NP15b (Fig. 6 ![]() ; Appendix 1). Robinson (1996) reported that a sample from the type locality of the Preston Hill Marl (his locality 2c, ER161) contained Eolepidina with
R = 5.5 (n = 14), which is at the transition between Eu. guayabalensis and Eo. antillea (i.e., the ABZ7/ABZ8 boundary) as defined here. This locality has yielded the following
plankton correlations: P11, upper NP15, and CP13-lower CP14a (Robinson, 1996) which is consistent with the upper part of Zone NP15c. We therefore place the boundary between ABZ7 and ABZ8 in the upper part of Zone NP15b (Fig. 6
; Appendix 1). Robinson (1996) reported that a sample from the type locality of the Preston Hill Marl (his locality 2c, ER161) contained Eolepidina with
R = 5.5 (n = 14), which is at the transition between Eu. guayabalensis and Eo. antillea (i.e., the ABZ7/ABZ8 boundary) as defined here. This locality has yielded the following
plankton correlations: P11, upper NP15, and CP13-lower CP14a (Robinson, 1996) which is consistent with the upper part of Zone NP15c. We therefore place the boundary between ABZ7 and ABZ8 in the upper part of Zone NP15b (Fig. 6 ![]() ). Sample USGS 6897 (collected by
S.B. Senn) from St. Bartholomew was reported (Robinson, 1996) to have Eo. antillea with
R = 5.08 (n = 12) and was very tentatively attributed to Zone P11/P12 (Westercamp & Andreieff, 1983; Robinson, 1996); it is consistent with the age of ABZ8 as determined here.
). Sample USGS 6897 (collected by
S.B. Senn) from St. Bartholomew was reported (Robinson, 1996) to have Eo. antillea with
R = 5.08 (n = 12) and was very tentatively attributed to Zone P11/P12 (Westercamp & Andreieff, 1983; Robinson, 1996); it is consistent with the age of ABZ8 as determined here.
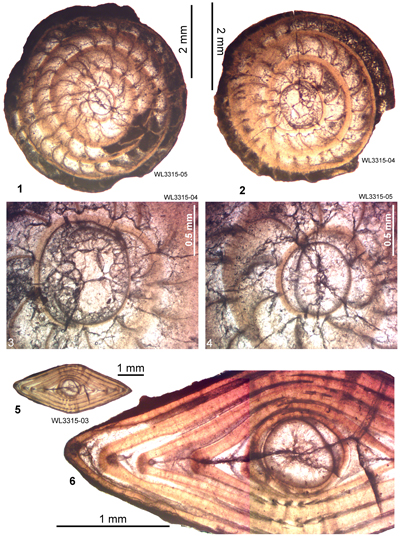
ABZ9. Eolepidina subplana Zone. From the FAD of Eo. subplana (with
R < 4.5) to the FAD of Eolepidina gardnerae (with
R = 3.5 or less). The type section lies at Lilyfield in the parish of St Ann, Jamaica. The zone can be
recognized in Alabama (Robinson & Jiang, 1995), St. Bartholomew (Robinson, 1996), Jamaica (Robinson, 1996), Mexico (Barker & Grimsdale, 1936), and Margarita Island, Venezuela (Caudri, 1974). A sample from Albert Town (ER1366 from Robinson, 1996, locality 2B) containing Eo. subplana (indicating ABZ9) yielded a few calcareous
nannofossils, including Sphenolithus furcatolithoides, Pemma papillatum and Discoaster barbadiensis and was assigned to Zone NP16 (Robinson, 1996). In the Bay Minette Corehole (Baldwin County, Alabama) Eo. subplana with
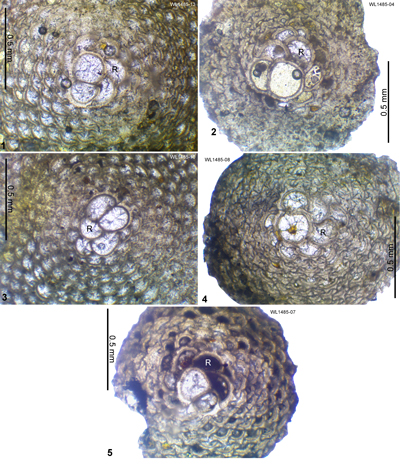
R = 4 occurs in samples at depths of 529.7 m, 543.2 m and 644.4-646.6 m and the nannofossils indicate assignment to Zone NP16 (Robinson & Jiang, 1995). We therefore place the boundary between ABZ8 and ABZ9 at about the NP15/NP16 boundary (Fig. 6 ![]() ). Sample ER1485 from St. Bartholomew (Sample 3A: Robinson, 1996) from the "Upper Group" (near
Lurin) was reported (Robinson, 1996) to have a R
= 3.82 (n = 11) and was tentatively attributed to Zone P12 (Westercamp & Andreieff, 1983; Robinson, 1996), which is consistent with the age of ABZ9 as determined here. On Margarita Island, Venezuela (Caudri, 1974), the following species are present (identifications updated from Caudri, 1974): Eo. subplana (=
Lepidocyclina
(Polylepidina) antillea of Caudri, 1974), Orbitoina sp., Helicolepidina polygyralis and Operculinoides
spp. and can be assigned to ABZ9.
). Sample ER1485 from St. Bartholomew (Sample 3A: Robinson, 1996) from the "Upper Group" (near
Lurin) was reported (Robinson, 1996) to have a R
= 3.82 (n = 11) and was tentatively attributed to Zone P12 (Westercamp & Andreieff, 1983; Robinson, 1996), which is consistent with the age of ABZ9 as determined here. On Margarita Island, Venezuela (Caudri, 1974), the following species are present (identifications updated from Caudri, 1974): Eo. subplana (=
Lepidocyclina
(Polylepidina) antillea of Caudri, 1974), Orbitoina sp., Helicolepidina polygyralis and Operculinoides
spp. and can be assigned to ABZ9.
ABZ10. Eolepidina gardnerae Zone. From the FAD of Eolepidina gardnerae (with R <
3.5) to the FAD of Polylepidina chiapasensis (with R
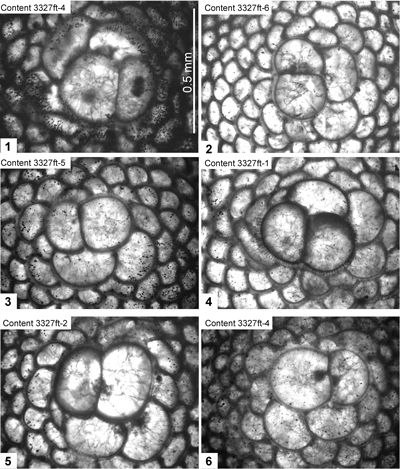
> 2.2). The reference section lies at a depth of 3327 feet
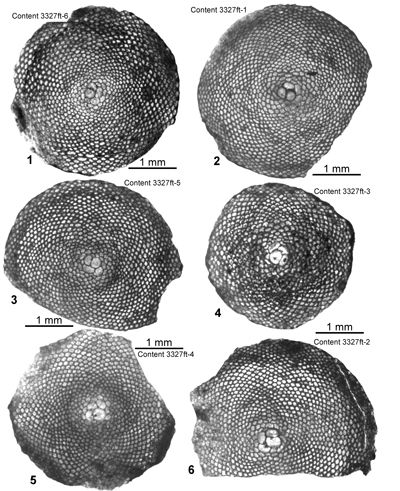
(= 1037.6 m) in the Content #1 Well, at Content, parish of Westmoreland, Jamaica. Eolepidina from here (as Polylepidina) have been described by Eva (1980) and Robinson (1996) and are also figured here. Robinson (1996) recorded biostratigraphic data from the level that yielded Eo. gardnerae in the Content-1 Well (his locality 2A): core samples from depths of 1010.4 m, 1030.8 m, 1040.6 m and 1070.8 m had rich, but overgrown, nannofossils (including Sphenolithus furcatolithoides, Cribrocentrum reticulatum, Reticulofenestra umbilica, Helicosphaera compacta, Dictyococcites bisectus (small form; = D. scrippsae) and Pemma papillatum: Robinson & Jiang, 1995; Robinson, 1996) that indicate Zone NP16; whereas a sample from 1040.6 m had calcareous nannofossils (including Sphenolithus furcatolithoides, Pemma papillatum and Reticulofenestra umbilica) and was assigned to Zone CP14a (determinations by
W.E. Steinkraus in Robinson, 1996). In the Bay Minette Corehole (Baldwin County, Alabama) Eo. gardnerae (ABZ10) with R
close to 3 occurs in three samples collected at depths of 515.1 m, 518.8 m and 522.1
m. These samples are assigned to Zone NP16 (Robinson & Jiang, 1995; Robinson, 1996), but the appearance of Cribrocentrum reticulata at 518.8 m (Robinson & Jiang, 1995) means that the uppermost sample that yielded P. gardnerae can be assigned to a level in Zone CP14a. This means that the boundary between zones ABZ9 and ABZ10 must occur within Zone NP16, probably within the upper part of Zone CP13c (Fig. 6 ![]() ).
).
ABZ11. Polylepidina chiapasensis Zone. From the FAD of Polylepidina chiapasensis (with R
= 2 - 2.2) to the FAD of Lepidocyclina ariana. The reference section lies on the road from Salt Spring to Flower Hill in the parish of St James, Jamaica. Samples WL4234 and WL4236 from the Preston Hill Formation (Salt Spring to Flower Hill transect) yield Po. chiapasensis and have yielded nannofossils indicating Zone NP16 and CP14a (Appendix 1). We therefore assign the ABZ10 - ABZ11 boundary to an interval within Zone NP16 (within Zone CP14a) (Fig. 6 ![]() ).
).
ABZ12. Lepidocyclina ariana Zone. From the FAD of Lepidocyclina ariana to the FAD of Lepidocyclina ocalana. At Grants Pen in St Thomas (Jamaica), L. ariana (Sample WL4630) occurs with nannofossils indicating upper Zone NP16 (CP14a) whereas a slightly higher sample without LBF (WL4628) yielded nannofossils possibly indicating Zone NP17 (Appendix 1). In the Bay Minette Corehole (Baldwin County, Alabama) L. ariana occurs with nannofossils indicating Zone NP17 (Robinson & Jiang, 1995). We therefore place the ABZ11- ABZ12 boundary in the upper part of Zone NP16 (CP14a). The reference section for the zone lies at Grants Pen in St Thomas, Jamaica.
This zone is widely developed being found in Jamaica, Alabama (Robinson & Jiang, 1995; Robinson, 1997), Florida (Cole, 1944) and Margarita Island, Venezuela (Caudri, 1974). In Margarita Island the LBF assemblage consists of advanced L. ariana, Orbitoina
spp., Helicolepidina spiralis trinitatis [= nortoni Vaughan], Helicosteginopsis soldadensis. Caudriella ospinae and Operculinoides
spp. (Caudri, 1974, identifications updated); this assemblage occurs with the planktic foraminifer Orbulinoides beckmanni indicating Zone P13 (Caudri, 1974)
- this is in complete agreement with the age calibration of ABZ12 herein based on calcareous nannofossils (Fig. 6 ![]() ).
).
ABZ13. Lepidocyclina ocalana Zone. From the FAD of L. ocalana to the FAD of Pliolepidina tobleri. The assemblage is
characterised by the appearance of species of Lepidocyclina with adauxiliary chambers, in contrast to the assemblage present in ABZ12. The reference locality for the zone is in the "Ipswich" Limestone in the Ipswich area, parish of St Elizabeth, Jamaica. A sample from the Ipswich Limestone (ER3103) yielded a sparse
nannoflora, including Chiasmolithus grandis and small Cribrocentrum reticulatum, indicating a range from upper Zone NP16 to Zone NP17 (Robinson et al., 2018). Strontium isotope ratios from this level (Robinson et al., 2018) suggest an age in the range 31.25-38.8 Ma consistent with Zone NP17. We therefore correlate ABZ13 primarily to lower-mid Zone NP17 (Fig. 6 ![]() ).
).
ABZ14. Pliolepidina tobleri Zone. From the FAD of Pl. tobleri to the LAD of Pl. tobleri. The reference locality of this zone lies around the marlstones/chalk boundary at Cold Spring in the Montpelier-New Market Belt, parish of Westmoreland, Jamaica. A sample (WL4444) from the marlstones at Cold Spring, just below a sample (WL4449) with common Pl. tobleri yielded a nannoflora indicating zone NP17 (Appendix 1). We therefore assign Zone ABZ14 to Zone NP17 (Fig. 6 ![]() ). Strontium isotope ratios from a sample with Pl. tobleri from Appleton, St Elizabeth, Jamaica (Robinson et al., 2018) gave ages which are too young in stratigraphic terms suggesting diagenetic alteration.
). Strontium isotope ratios from a sample with Pl. tobleri from Appleton, St Elizabeth, Jamaica (Robinson et al., 2018) gave ages which are too young in stratigraphic terms suggesting diagenetic alteration.
ABZ15. Orbitoina wrighti Zone. From the LAD of Pl. tobleri to the FAD of Heterostegina ocalana. The reference section for the zone lies in the Swanswick Formation
s.l. on the N-S Highway 2000 in the parish of St Ann, Jamaica. Nannofossil data from the Montpelier-New Market Belt indicate that ABZ15 ranges from Zone NP18 to the lower part of Zone NP19/20 (sensu Martini, 1971, non Speijer et al., 2020; Fig. 6 ![]() ).
).
ABZ16. Heterostegina ocalana Zone. From the FAD of H. ocalana to the LAD of H. ocalana. The reference locality lies in the White Limestone at
Dressikie, parish of St Mary, Jamaica. ABZ16 occurs in the type Swanswick (Robinson & Wright, 1993) and Somerset formations (Robinson, 1974) of the marginal facies of the Clarendon Block, as well as in deep-water turbidites in the Wag Water Belt (Robinson, 2004). Localities in the NW Wagwater Belt (ER176 from deep-water chalks of the White Limestone Group at
Dressikie) that have yielded H. ocalana contain a nannoflora assignable to Zone NP21 (Robinson, 2004; Robinson et al., 2018). In Cuba, Torres-Silva et al. (2017) suggested a range of H. ocalana from Zone NP17 to Zone NP19-20/CP15 (but the individual zones were not separated suggesting limited precision) and possibly Zone NP21 (CP16) and P18 (early
Rupelian). These data do not seem to relate well to Jamaica and provisionally we suggest that ABZ16 ranges from Zone NP19/20 to lower Zone NP21 (Fig. 6 ![]() ). Strontium isotopes from the Claremont Formation (which we would attribute to ABZ16) gave ages of 35.05-35.95 Ma (Robinson et al., 2018), which would indicate a level in lower Zone NP19-20.
). Strontium isotopes from the Claremont Formation (which we would attribute to ABZ16) gave ages of 35.05-35.95 Ma (Robinson et al., 2018), which would indicate a level in lower Zone NP19-20.
Occurrences of what have been regarded as typical late Eocene LBF indicators (e.g., orthophragmines, H. ocalana and Fabularia verseyi) in the Oligocene are likely due either to alteration of Sr-isotope values (giving an age that is too young) or due to reworking of the LBFs into the Oligocene (Robinson, 2004; Molina et al., 2015; Robinson et al., 2018; Torres-Silva et al., 2017). The top of the zone is notable for the disappearance of many typical Eocene taxa including H. ocalana, Fabularia verseyi, Coskinolina cf. douvillei, Pellatispirella matleyi and Butterliniana cubensis.
Having established a zonation for the Eocene strata of Jamaica based on
LBF, in this section we show how it can be used in selected areas of the American Bioprovince based on previous studies (that is, our interpretations of previously figured specimens) of
LBF. Some areas are difficult to comment on, because of tectonic deformation and associated reworking of faunas. The Cuban arc underwent a collision with the North American Plate (Iturralde-Vinent et al., 2016) in the mid-late Eocene and there is extensive reworking of material; it is therefore difficult to work out successions in Cuba other than for the comments we have provided above. With further work, we beleave that the scheme can be used in Cuba, but will require extensive consideration of reworked material. A considerable amount of work has been undertaken in NE Brazil (e.g., Mello e Sousa et al., 2003), but is largely based on random sections of LBF in thin sections; as such it is difficult to apply the scheme there without a reinvestigation of the material available. Our regional correlation is shown in Fig. 6 ![]() .
.
5.1. Florida. Extensive studies concern the Eocene LBF succession in Florida (e.g., P.L. Applin & E.R. Applin, 1941; Cole, 1941, 1942, 1944, 1945; E.R. Applin & Jordan, 1945; Levin, 1957; Cole & E.R. Applin, 1964; Powell, 2010; Cotton et al., 2018). We have reviewed the occurrences and illustrations in these works and present our reinterpretation of the biostratigraphy in Fig. 7 ![]() .
.
The Oldsmar Formation consists of a lower unit of grey-brown dolostone and an upper unit of white to grey, interbedded layers of lime mudstone, wackestones, packstones and tan dolostones (Miller, 1986). The basal boundary is sharp. In Cedar Keys No. 2 well, Linderina floridensis occurs at 983-1068 ft. (= 300-326 m) and Hel. gyralis and L. floridensis occur at 1301-1308 ft (= 397-399 m) (Cole, 1942, p. 15), but no nummulitids were recorded from this well (although it is possible that nummulitids may occur in more open water environments that Helicostegina). Levin (1957) illustrated typical Hel. gyralis from the Oldsmar Formation. This suggests a correlation with upper ABZ6, or possibly lower ABZ7 (if nummulitids are absent due to facies control). However, the similar embryonic sizes of H. gyralis as given by Levin (1957) with specimens from upper ABZ6 suggests an assignment to upper ABZ6.
The Avon Park Formation of P.L. Applin & E.R. Applin (1944, p. 1686) includes the Lake City Formation of P.L. Applin & E.R. Applin (1944, p. 1693) (Miller, 1986). The formation consists of light-brown limestones and dolostones, and has an unconformable relationship with the underlying Oldsmar Formation (Miller, 1986). The best foraminiferal record from the Avon Park Formation comes from Hilliard Turpentine Company well W-366 (Cole, 1944,p. 33-34) where the following species and levels were indicated (species identifications updated): Eolepidina antillea (R = 5/6, Cole, 1944, Pl. 10, figs. 1-3, 5, 7) at a depth of 1340-1350 ft (= 408-411 m) (ABZ8); Eo. subplana (R = 4, Cole, 1944, Pl. 10, figs. 4, 6; Pl. 12, fig. 3) at a depth of 1300-1305 ft (= 396-398 m) (ABZ9); Lep. ariana (R = 2, Cole, 1944, Pl. 16, figs. 11-13) at a depth of 1065-1070 ft (= 324-326 m) (ABZ12) and Orbitoina cedarkeyensis Cole, 1944, p. 34) at a depth of 1005-1025 ft (= 306-312 m) (ABZ12 or ABZ13?). Above 1005 ft. (= 306 m), there is a change in fauna, with the upper part of the Avon Park Formation being characterised by conical agglutinated foraminifers including Coskinolina, Fallotella and Cushmania (Cole, 1944). Specimens of Cushmania from the Avon Park Formation (e.g., Cole, 1944, Pl. 4, fig. 6) have a relatively large (400 μm) first chamber, and this morphology characterises ABZ13-ABZ15 in Jamaica. Thus the Avon Park Formation probably ranges from ABZ8 to at least ABZ13.
The Ocala Formation consists of white- to cream-coloured, highly fossiliferous wackestones, packstones and grainstones with a basal unconformity with the underlying Avon Park Formation (Miller, 1986). It yields a fauna characterised by Lep. ocalana, Het. ocalana and Eu. chaperi (Cole, 1941, 1942, 1944, 1945; Powell, 2010; Cotton, 2018). This assemblage indicates ABZ16.
5.2. Alabama. Robinson and Jiang (1996) provided biostratigraphic data on calcareous nannofossils and LBFs from the Bay Minette corehole and the section at Little Stave Creek in Alabama (Fig. 7 ![]() ). We recognize four depositional units, the upper three of which can be constrained by LBF. The appearance of Operculinoides gravelli indicates either ABZ7 or ABZ8, whereas the presence of Eo. subplana and Eo. gardnerae indicate ABZ9 and ABZ10, respectively. There is a hiatus representing ABZ11 and Lep. ariana indicates ABZ12. There is a further hiatus representing ABZ13-ABZ15 whereas the presence of Lep. montgomeriensis possibly suggests ABZ16. There is a general resemblance to the stratigraphy of Florida with similar hiatuses/unconformities.
). We recognize four depositional units, the upper three of which can be constrained by LBF. The appearance of Operculinoides gravelli indicates either ABZ7 or ABZ8, whereas the presence of Eo. subplana and Eo. gardnerae indicate ABZ9 and ABZ10, respectively. There is a hiatus representing ABZ11 and Lep. ariana indicates ABZ12. There is a further hiatus representing ABZ13-ABZ15 whereas the presence of Lep. montgomeriensis possibly suggests ABZ16. There is a general resemblance to the stratigraphy of Florida with similar hiatuses/unconformities.
5.3. Panama. Cole (1952) described the foraminifers of the Gatuncillo Formation from the Panama Canal Zone, and Woodring (1957) provided additional information on the samples. The samples of Cole (1952) indicate three zones: ABZ14 with Plio. tobleri; ABZ15 without either Plio. tobleri or Heter. ocalana (but some samples with Eu. chaperi); and ABZ16 with Heter. ocalana and Eu. chaperi (Fig. 7 ![]() ).
).
5.4. Mexico (state of Veracruz). The species recorded by Barker and
Grimsdale (1936, 1937) and Barker (1939) allow the identification of some zones in the state of Veracruz, Mexico (Fig. 7 ![]() ). The association of Hel. gyralis with Pseudolepidina trimera and Operc. jennyi at Jenny Station No. 1573 (11 km southeast of Sabaneta, state of Veracruz, Mexico) indicates lower ABZ7, whereas the presence of Hel. dimorpha indicates upper ABZ7 or ABZ8. The presence of Eu. guayabalensis in association with Operculinoides prenummulitiformis and Nummulites guayabalensis (Barker, 1939, p. 325) also indicates ABZ7. The presence of Eulinderina semiradiata (=
Eolepidina antillea) and Eulinderina guayabalensis regularis (= Eu.
regularis) in association with Tremastegina would indicate a level around the ABZ7-ABZ8 boundary. Polylepidina chiapasensis var. subplana (= Eo. subplana) and Polylepidina
'chiapasensis' (= Eo. gardnerae) indicate zones ABZ9 and ABZ10, respectively. Frost and
Langenheim (1974) figured specimens of Eo. gardnerae as Lepidocyclina (Polylepidina) antillea from central Chiapas, Mexico, which would also indicate ABZ10.
). The association of Hel. gyralis with Pseudolepidina trimera and Operc. jennyi at Jenny Station No. 1573 (11 km southeast of Sabaneta, state of Veracruz, Mexico) indicates lower ABZ7, whereas the presence of Hel. dimorpha indicates upper ABZ7 or ABZ8. The presence of Eu. guayabalensis in association with Operculinoides prenummulitiformis and Nummulites guayabalensis (Barker, 1939, p. 325) also indicates ABZ7. The presence of Eulinderina semiradiata (=
Eolepidina antillea) and Eulinderina guayabalensis regularis (= Eu.
regularis) in association with Tremastegina would indicate a level around the ABZ7-ABZ8 boundary. Polylepidina chiapasensis var. subplana (= Eo. subplana) and Polylepidina
'chiapasensis' (= Eo. gardnerae) indicate zones ABZ9 and ABZ10, respectively. Frost and
Langenheim (1974) figured specimens of Eo. gardnerae as Lepidocyclina (Polylepidina) antillea from central Chiapas, Mexico, which would also indicate ABZ10.
5.5. Margarita Island, Venezuela. Brönnimann (1944), Butterlin (1970) and Caudri (1974) described Eocene LBF assemblages from the Punta Mosquito Formation of Margarita Island. Caudri (1974) figured the fauna in detail from three samples and these forms can be placed in the zonation scheme (Fig. 7 ![]() ). Sample B yielded Eo. subplana (=
Lepidocyclina (Polylepidina) antillea of Caudri, 1974), Orbitoina sp., Helicolepidina polygyralis and Operculinoides spp. (Caudri, 1974, identifications updated) and can be placed in ABZ9. Sample A yielded L. ariana, Orbitoina spp. and Operculinoides spp. (Caudri, 1974) and can be assigned to ABZ12. Sample P.J.B.145 yielded L. ariana, Orbitoina spp., Helicolepidina spiralis trinitatis [=
nortoni Vaughan], Helicosteginopsis soldadensis, Caudriella ospinae and Operculinoides spp. (Caudri, 1974, identifications updated) and can also be assigned to ABZ12.
). Sample B yielded Eo. subplana (=
Lepidocyclina (Polylepidina) antillea of Caudri, 1974), Orbitoina sp., Helicolepidina polygyralis and Operculinoides spp. (Caudri, 1974, identifications updated) and can be placed in ABZ9. Sample A yielded L. ariana, Orbitoina spp. and Operculinoides spp. (Caudri, 1974) and can be assigned to ABZ12. Sample P.J.B.145 yielded L. ariana, Orbitoina spp., Helicolepidina spiralis trinitatis [=
nortoni Vaughan], Helicosteginopsis soldadensis, Caudriella ospinae and Operculinoides spp. (Caudri, 1974, identifications updated) and can also be assigned to ABZ12.
5.6. Venezuela. Raadshooven (1951) described a series of LBF assemblages from Venezuela. The earliest Eocene assemblage from the Quebrada Grande Limestone (Baralt District, State of Zulia, western Venezuela) consisted of Planorbitoinella nitida
(= Lepidocyclina sp. A of Raadshooven) which occurred with Helic. gyralis and Operculinoides cf. oliveri which suggests lower ABZ7 (Fig. 7 ![]() ). The San Juan Limestone (Baralt District, State of Zulia, western Venezuela) yields Poly. ecuadorensis in association with Operculinoides spp. (cited as O. jennyi by Raadshooven, 1951); this suggests upper ABZ7 (Fig. 7
). The San Juan Limestone (Baralt District, State of Zulia, western Venezuela) yields Poly. ecuadorensis in association with Operculinoides spp. (cited as O. jennyi by Raadshooven, 1951); this suggests upper ABZ7 (Fig. 7 ![]() ).
).
5.7. Saint Bartholomew (St. Barths). The Eocene succession in Saint Bartholomew has been described in many works (e.g., Lewis & Robinson, 1976; Westercamp & Andreieff, 1983; Caron et al., 2019; Cornée et al., 2020) and details of the LBFs have been given in works such as Cushman (1919), Cole (1960), Robinson (1996) and Cornée et al. (2020). The succession consists of alternations of shallow-water limestones and volcaniclastic sedimentary rocks which have been intruded by various igneous plutons (Westercamp & Andreieff, 1983; Legendre et al., 2018). The stratigraphy for the Eocene rocks has varied from paper to paper. Lewis and
Robinson (1976) placed all the Eocene limestones in the Middle Eocene. Westercamp and
Andreieff (1983) recognized six Eocene limestone horizons which they divided into a lower group (lower two limestones) and an upper group (upper four limestones), all of which were assigned to the Upper Lutetian. Caron et al. (2019) named the six limestone horizons from base to top: Flamands (1), Mangin (2), Cayes (3), Gouverneur (4) and Lurin (5-6); they assigned these to the upper Lutetian to Bartonian. In contrast, Cornée et al. (2020) divided the succession into three limestones: a Lower Limestone of Lutetian age, an Intermediate Limestone of late Bartonian/early-late Priabonian age, and an Upper Limestone of Priabonian age. The ages of the limestones as given by Cornée et al. (2020) were largely based on LBF identified in random thin sections to genus level, and judging by their illustrations, there are clearly some misidentifications of species and genera (e.g., Eoconuloides wellsi, Helicostegina, Heterostegina ocalana). Detailed investigations of orientated sections of LBFs from two of the limestones have been given by Cole (1960) and Robinson (1996) and these can be placed within the zonal scheme presented here. The Flamands Limestone (1) of Caron et al. (2019) at Pointe Lézarde yields Eolepidina antillea (it is the type locality: Cole, 1960; Robinson, 1996), which allows assignment to ABZ8 (Fig. 7 ![]() ); whereas the Lurin Limestone (5-6) of Caron et al. (2019) yields E. subplana (Robinson, 1996) and can be assigned to ABZ9 (Fig. 7
); whereas the Lurin Limestone (5-6) of Caron et al. (2019) yields E. subplana (Robinson, 1996) and can be assigned to ABZ9 (Fig. 7 ![]() ). Assuming that the LBFs are not reworked, this would suggest that the limestone sequence on Saint Bartholomew is solely of mid to mid-late Lutetian age (although higher levels could be represented in the upper part of the Lurin Limestone) and a study of orientated sections of LBF from other limestone horizons is needed. Miocene limestones occur above this interval but are not relevant to this work.
). Assuming that the LBFs are not reworked, this would suggest that the limestone sequence on Saint Bartholomew is solely of mid to mid-late Lutetian age (although higher levels could be represented in the upper part of the Lurin Limestone) and a study of orientated sections of LBF from other limestone horizons is needed. Miocene limestones occur above this interval but are not relevant to this work.
5.8. Trinidad. It is difficult to apply the zonation scheme to Trinidad because of the thin successions of Eocene rocks and the extensive reworking of specimens of LBFs. This is clearly seen in Soldado Rock (e.g., Caudri, 1975, 1996) where there are extensive slump deposits and the reworking of older foraminifers into
younger layers (Kugler & Caudri, 1975). Ignoring the slump masses, the first appearances of specific foraminifers can be used to suggest some correlations for Soldado Rock. The presence of Operculinoides spp. in lower bed 3 suggests ABZ7 or younger, whereas the absence of typical Lutetian markers suggests that Bed 3 may be as young as Bartonian. The appearance of Pliolepidina tobleri in bed 7 would suggest a correlation with ABZ14. Because of the extensive reworking, we do not show Trinidad on Fig. 7 ![]() .
.
5.9. Barbados. The Scotland District of Barbados also contains Eocene LBFs, but as with Trinidad there is extensive reworking. The Murphys Beds however yield T. senni in association with H. minor without Operculinoides spp. or Nummulites spp. (Brönnimann, 1950). This would suggest an assignment to either ABZ4 or ABZ6. Paleocene Ranikothalia occur throughout (Cizancourt, 1948; Caudri, 1972) indicating extensive reworking from the Paleocene. As a consequence, we do not show Barbados on Fig. 7 ![]() .
.
| Figure 7:
Our interpretation of representative sections in selected areas across the
American biogeographical Province based on our zonal scheme presented here. |
Previous classifications of the orbitoidiform foraminifers considered here can be found in Adams (1987), Loeblich and Tappan (1988), Butterlin (1988), and BouDagher-Fadel (2018), with the phylogeny suggested by Barker and Grimsdale (1936) having been generally accepted in all subsequent works. With regard to the Lepidocyclininae, previous workers (e.g., Grimsdale, 1959; Vlerk, 1959; Sirotti, 1983; Butterlin, 1987) have suggested different clades. Our studies of the Asterigerinoidea Orbigny (forms surrounding Amphistegina, Lepidocyclina and their allies) clearly demonstrate that the current classification is polyphyletic, and also that the simple phylogenetic scheme presented by Barker and Grimsdale (1936) is too simplistic, ignores the different forms of test construction in the various groups, and thus should not be the basis for the classification of these foraminifers into families and subfamilies. Here, we introduce a revised classification (based on foraminiferal test morphology and our detailed stratigraphic occurrences) that we think provides a suitable monophyletic scheme and gives an appropriate classification rank to the various clades. We feel that the current classification of taxa into the Helicolepidininae and Lepidocyclininae is not appropriate and prefer to base families on clearly defined phylogenetic series regardless of whether orbitoidiform growth has not or has been achieved. We do not formally describe each genus or species (unless new), but provide synonyms, diagnostic criteria, and discussions that hopefully allow genera and species to be properly identified. We illustrate numerous specimens (some well preserved and some more poorly preserved) of each species so that there is no ambiguity in our specific interpretations. For ease of reference, we also list the bewildering array of synonyms at the generic level that have been previously introduced. The classification scheme adopted for the taxa we use in our zonation is laid out below.
6.1. Terminology
We follow standard guides on terminology for the classification of the foraminifers described here (e.g., Adams, 1987; Hottinger, 2006). We discuss some of these terms in more detail below.
The terms used for the orientation of the tests of foraminifers are not uniform between different morphologies, and separate schemes are used for trochospiral and orbitoidiform taxa (Hottinger, 2006). In the Helicosteginidae fam. nov. the test is trochospiral, and in the early forms (e.g., Helicostegina wellsi), one side of the test is elevated, and the other side of the test is flat or even concave (Fig. 8 ![]() .1). In Helicostegina wellsi, the proloculus is situated close to the flat (or concave) side of the test (Fig. 8
.1). In Helicostegina wellsi, the proloculus is situated close to the flat (or concave) side of the test (Fig. 8 ![]() .1), which by definition makes this the
'spiral' or dorsal side of the test (Hottinger, 2006). In H. wellsi the subsidiary chamberlets are formed within the alar prolongations (or replace the septa forming the alar prolongations) on the
'umbilical' or ventral side of the test (Fig. 8
.1), which by definition makes this the
'spiral' or dorsal side of the test (Hottinger, 2006). In H. wellsi the subsidiary chamberlets are formed within the alar prolongations (or replace the septa forming the alar prolongations) on the
'umbilical' or ventral side of the test (Fig. 8 ![]() .1). In later species of Helicostegina (e.g., H. gyralis, H. dimorpha, H. jeannemairae sp. nov.) the ventral side of the test can be recognized by the layer of subsidiary chamberlets (Fig. 8
.1). In later species of Helicostegina (e.g., H. gyralis, H. dimorpha, H. jeannemairae sp. nov.) the ventral side of the test can be recognized by the layer of subsidiary chamberlets (Fig. 8 ![]() .2-3). In forms with an orbitoidiform test, such as the Lepidocyclinidae, there is an equatorial layer with lateral chamberlets on both sides (Fig. 8
.2-3). In forms with an orbitoidiform test, such as the Lepidocyclinidae, there is an equatorial layer with lateral chamberlets on both sides (Fig. 8 ![]() .4). It is only in the earliest forms (Eulinderina guayabalensis) that the proloculus lies closer to one (dorsal) side of the test and there is a hint of low trochospiral coiling (Fig. 8
.4). It is only in the earliest forms (Eulinderina guayabalensis) that the proloculus lies closer to one (dorsal) side of the test and there is a hint of low trochospiral coiling (Fig. 8 ![]() .5); in later forms the dorsal and ventral sides of the test cannot be distinguished and an equatorial layer is overlain by two sets of lateral chamberlets (Fig. 8
.5); in later forms the dorsal and ventral sides of the test cannot be distinguished and an equatorial layer is overlain by two sets of lateral chamberlets (Fig. 8 ![]() .4). Thus the subsidiary chamberlets in the Helicosteginidae are equivalent to one set of lateral chamberlets as developed in the Lepidocyclinidae. Thus the
'complete flange of equatorial chamberlets' described in H. dimorpha by Adams (1987, p. 294) represents a cut passing from the primary spire into the subsidiary layer of ventral chamberlets. In later species (e.g., H. jeannemairae sp. nov., H. jamaicensis sp. nov.) a true flange of equatorial chamberlets is developed from the primary spire and a subsidiary layer of ventral chamberlets (which have the same morphology as a true equatorial layer) is developed on the ventral side giving the flange a two-layered structure (Fig. 8
.4). Thus the subsidiary chamberlets in the Helicosteginidae are equivalent to one set of lateral chamberlets as developed in the Lepidocyclinidae. Thus the
'complete flange of equatorial chamberlets' described in H. dimorpha by Adams (1987, p. 294) represents a cut passing from the primary spire into the subsidiary layer of ventral chamberlets. In later species (e.g., H. jeannemairae sp. nov., H. jamaicensis sp. nov.) a true flange of equatorial chamberlets is developed from the primary spire and a subsidiary layer of ventral chamberlets (which have the same morphology as a true equatorial layer) is developed on the ventral side giving the flange a two-layered structure (Fig. 8 ![]() .3).
.3).
Following Adams (1987, p. 289) we use the terms proloculus and deuteroloculus for the first two chambers in spiral forms (Asterigerinoidea, Nummulitidae) where the second chamber (deuteroloculus) has only one aperture or stolon (R = 3 or more; Fig. 8 ![]() .6). We use the term protoconch and deuteroconch in forms where the deuteroconch has two (R = 2) apertures or stolons (Fig. 8
.6). We use the term protoconch and deuteroconch in forms where the deuteroconch has two (R = 2) apertures or stolons (Fig. 8 ![]() .7). In many of these forms (whether they consist of a proloculus and deuteroloculus or a protoconch and a deuteroconch) the first two chambers frequently have a thicker and darker wall than the succeeding chambers and this represents the embryo (e.g., Adams, 1987; Krüger et al., 1997; Langer & Hottinger, 2000). We use the form of the embryo and subsequent growth as generic and specific characters. In axial sections either the first two chambers (Figs. 8.1-2, 8.5), or one of the first two chambers may be visible (Fig. 8
.7). In many of these forms (whether they consist of a proloculus and deuteroloculus or a protoconch and a deuteroconch) the first two chambers frequently have a thicker and darker wall than the succeeding chambers and this represents the embryo (e.g., Adams, 1987; Krüger et al., 1997; Langer & Hottinger, 2000). We use the form of the embryo and subsequent growth as generic and specific characters. In axial sections either the first two chambers (Figs. 8.1-2, 8.5), or one of the first two chambers may be visible (Fig. 8 ![]() .3-4).
.3-4).
We use the term chamber for successive growth stages of spiral foraminifers. In some forms, the alar prolongations of the chambers are divided up into smaller chamberlets on the ventral side of the test in trochospiral forms. In some trochospiral forms, the alar prolongations of the chambers on the ventral side of the test interfinger with short ventral chamberlets (Amphistegina, Tremastegina: Fig. 8 ![]() .8); the term stellar chamberlets has been used for these short ventral chamberlets (Hottinger, 2006), but may not be appropriate and they may have had a separate origin. We tentatively use the term stellar chamberlet here pending further research on these forms. In other trochospiral forms (e.g., Helicostegina gyralis), the alar prolongations on the ventral side of the test are divided up into a series of subsidiary chamberlets (Fig. 8
.8); the term stellar chamberlets has been used for these short ventral chamberlets (Hottinger, 2006), but may not be appropriate and they may have had a separate origin. We tentatively use the term stellar chamberlet here pending further research on these forms. In other trochospiral forms (e.g., Helicostegina gyralis), the alar prolongations on the ventral side of the test are divided up into a series of subsidiary chamberlets (Fig. 8 ![]() .9) connected by a series of ventral stolons. In this case the lumina of the chambers with undivided dorsal alar prolongations can be seen interfingering with the subsidiary chamberlets on the ventral side of the test (H. minor, Fig. 8
.9) connected by a series of ventral stolons. In this case the lumina of the chambers with undivided dorsal alar prolongations can be seen interfingering with the subsidiary chamberlets on the ventral side of the test (H. minor, Fig. 8 ![]() .9). In later forms (H. jeannemairae) the ventral layer of subsidiary chamberlets passes into a ventral sheet of chamberlets which mimics the growth in the equatorial layer of orbitoidiform foraminifers (Fig. 8
.9). In later forms (H. jeannemairae) the ventral layer of subsidiary chamberlets passes into a ventral sheet of chamberlets which mimics the growth in the equatorial layer of orbitoidiform foraminifers (Fig. 8 ![]() .3). In contrast, the equatorial layer of orbitoidiform foraminifers has lateral chamberlets on both sides (Fig. 8
.3). In contrast, the equatorial layer of orbitoidiform foraminifers has lateral chamberlets on both sides (Fig. 8 ![]() .4) of the equatorial layer which are separated by irregular or regular walls and connected by diagonal stolons (Hottinger, 2006).
.4) of the equatorial layer which are separated by irregular or regular walls and connected by diagonal stolons (Hottinger, 2006).
| Figure 8:
Terminology used in this paper. 1 Helicostegina wellsi (EO.WL2044-75)
showing position of proloculus and ventral subsidiary chamberlets (axial
section). 2 Helicostegina gyralis (BMNH.P51952) and 3 Helicostegina jeannemairae
sp. nov. (UWIGM.WL3641-12) showing ventral layers of subsidiary chamberlets
(axial sections). 4 Lepidocyclina yurnagunensis morganopsis (BMNH
PS47315) showing equatorial layer with lateral chamberlets (axial section). 5 Eulinderina guayabalensis
(BMNH.P51959) showing proloculus closer to one side (?dorsal) than the other
(axial section) and a lack of lateral chamberlets. 6 Eulinderina subplana
(UWIGM.WL1485-04) showing embryo consisting of proloculus and deuteroloculus
with the deuteroloculus having one budding chamber (3), equatorial section. 7 Lepidocyclina ariana
(UWIGM.WL3571-01) showing embryo with protoconch and deuteroconch with the
deuteroconch having two budding chambers (R) and symmetrical spires of peri-embryonic
chambers, equatorial section. 8 Tremastegina lopeztrigoi (USNM 544392)
showing stellar chamberlets and piles/pillars, ventral exterior view. 9 Helicostegina minor
(ER1315), decortized specimen showing lumina of dorsal chambers extending onto
ventral side, septa between chambers, ventral subsidiary chamberlets, and the
spiral wall between whorls, ventral view. Key to symbols: 1 - protoconch or
proloculus; 2 - deuteroloculus or deuteroconch; 3, 4, 5 - subsequent chambers/chamberlets.
D - dorsal; DAP - dorsal alar projections; E - embryo; ECls - equatorial
chamberlets; LCls - lateral chamberlets; LDCs - lumina of dorsal chambers; Ls
- lumina; P - protoconch/proloculus or deuteroloculus/deuteroconch; R - first
chamber with a retrovert aperture; S - septa; V - ventral; VSCls - ventral
subsidiary chamberlets. |
The equatorial layer in more primitive members of many megalospheric forms of various lineages starts with a proloculus and a deuteroloculus followed by a primary spire of equatorial chambers (Fig. 9 ![]() .1). In many forms, the primary spire persists to the edge of the test, but in others the spire terminates in a chamber (R) with two apertures (Fig. 9
.1). In many forms, the primary spire persists to the edge of the test, but in others the spire terminates in a chamber (R) with two apertures (Fig. 9 ![]() .1) from which a spiral and a counterspiral of equatorial chambers encircle either the embryo (of two chambers) or the earlier chambers of the initial spire (following the embryo) and terminate in a closing chamber (Fig. 9
.1) from which a spiral and a counterspiral of equatorial chambers encircle either the embryo (of two chambers) or the earlier chambers of the initial spire (following the embryo) and terminate in a closing chamber (Fig. 9 ![]() .1). Each chamber in these two spires has two apertures which gives rise to equatorial chamberlets and initiates orbitoidiform growth. In many lineages, the length of the primary spire shows a progressive reduction over time (nepionic acceleration); this may give rise to one PAC (in which case a proloculus and deuteroloculus are still present and R = 3) or two PACs (in which a protoconch and deuteroconch are present and R = 2; Fig. 9
.1). Each chamber in these two spires has two apertures which gives rise to equatorial chamberlets and initiates orbitoidiform growth. In many lineages, the length of the primary spire shows a progressive reduction over time (nepionic acceleration); this may give rise to one PAC (in which case a proloculus and deuteroloculus are still present and R = 3) or two PACs (in which a protoconch and deuteroconch are present and R = 2; Fig. 9 ![]() .2). Once an embryo and two PACs are developed, either three of four unequal spires of peri-embryonic chambers encircle the embryo, with two closing chambers; Fig. 9
.2). Once an embryo and two PACs are developed, either three of four unequal spires of peri-embryonic chambers encircle the embryo, with two closing chambers; Fig. 9 ![]() .2). In more primitive forms the three or four spirals are asymmetrical (Fig. 9
.2). In more primitive forms the three or four spirals are asymmetrical (Fig. 9 ![]() .2), but over time four symmetrical spirals of peri-embryonic chambers encircle the embryo (Fig. 9
.2), but over time four symmetrical spirals of peri-embryonic chambers encircle the embryo (Fig. 9 ![]() .3). Subsequently, some forms develop adauxiliary chambers (Fig. 9
.3). Subsequently, some forms develop adauxiliary chambers (Fig. 9 ![]() .4) which are connected to the protoconch and/or the deuteroconch by stolons; in these forms short spirals and multiple closing chambers are developed. This progressive evolution of the equatorial layer is repeated in many lineages and this gives rise to homeomorphs which are not phylogenetically related. We use other criteria to separate these different clades.
.4) which are connected to the protoconch and/or the deuteroconch by stolons; in these forms short spirals and multiple closing chambers are developed. This progressive evolution of the equatorial layer is repeated in many lineages and this gives rise to homeomorphs which are not phylogenetically related. We use other criteria to separate these different clades.
The size of the embryo has commonly been used in separating species in the Americas (e.g., Cole, 1952; Barker & Grimsdale, 1936; Colletti et al., 2019). However, the size of the embryo has both phylogenetic significance and environmental significance, with variations due to salinity, temperature, and water depth (e.g., Nigam & Rao, 1987; Bryan, 1995; Yu et al., 2016). Given the variation in the size of the embryo in populations of the same species from different environments, the size of the embryo, in itself, is not diagnostic for species identification (we show examples of this in our work here). Notwithstanding this, the size of the embryo helps in distinguishing between different lineages (e.g., Lepidocyclina and Orbitoina) and needs to be taken into consideration (with other characters).
Equatorial chamberlets in an orbitoidiform growth plan are developed from the primary spiral(s) of chambers or from the peri-embryonic equatorial chambers (Fig. 9 ![]() .5). Their morphology is dependent on shape (i.e., how the chambers interlock), the radial distance versus the concentric distance of the connecting walls, and the position of the connecting stolons between radially adjacent equatorial chamberlets. The form of
equatorial chamberlets in American lepidocyclinids has been investigated by Hohenegger and
Torres Silva (2020), but we do not consider this in detail in this work. We illustrate the equatorial chamberlets in the figures of the species that follow.
.5). Their morphology is dependent on shape (i.e., how the chambers interlock), the radial distance versus the concentric distance of the connecting walls, and the position of the connecting stolons between radially adjacent equatorial chamberlets. The form of
equatorial chamberlets in American lepidocyclinids has been investigated by Hohenegger and
Torres Silva (2020), but we do not consider this in detail in this work. We illustrate the equatorial chamberlets in the figures of the species that follow.
Lateral chamberlets (seen in axial sections) may form regular stacks (tiers) or alternate. They may be separated by thin walls, thick walls or piles (pillars). The surface of the test may be smooth, have irregular anastomosing patterns representing the walls between the lateral chamberlets, or have piles (pillars or pustules). We illustrate the form of the lateral chamberlets in the figures for each taxon below. In some forms (e.g., Tremastegina, Helicostegina), small pustules may follow the septa on the dorsal and/or ventral side/s of the test. In some forms, the ventral side of the test may be divided into chamberlets or stellar chamberlets (Fig. 8 ![]() .8).
.8).
| Figure 9:
Forms of equatorial chambers and chamberlets. 1-4, line drawings showing
equatorial chambers. 1, Proloculus (1) and deuteroloculus (2) followed by a
short spire of equatorial chambers ending in a chamber with a retrovert
aperture (R) from which two spires (spire and counterspire) of equatorial
chambers develop and meet in a closing chamber (c). 2, Form with a protoconch
and deuteroconch and two Principal Auxiliary Chambers (PACs) with four
asymmetrical spires meeting in two closing chambers (c). 3, Form with a
protoconch and deuteroconch and two Principal Auxiliary Chambers (PACs) with
four symmetrical spires meeting in two closing chambers (c). 4, Form with four
adauxiliary chamberlets and multiple closing chambers. 5, Form with equatorial
chamber arrangement as in 3, showing equatorial chamberlets developing from
the equatorial chambers. |
Various measurements have been made on the embryos of LBF. This has varied both by geographical region (e.g., Tethys versus the Americas) and by group. For instance, in Cretaceous Orbitoides (Fig. 10 ![]() .1) the length and width of the nucleoconch excluding the thickness of the walls has been measured (e.g., Hinte, 1965; Gorsel, 1978). For
orthophragmines (Fig. 10
.1) the length and width of the nucleoconch excluding the thickness of the walls has been measured (e.g., Hinte, 1965; Gorsel, 1978). For
orthophragmines (Fig. 10 ![]() .2) the thickness of the wall has been included in the measurements of the width of the protoconch and the deuteroconch (e.g., Less, 1987). In contrast, for
Tethys lepidocyclinids (Fig. 10
.2) the thickness of the wall has been included in the measurements of the width of the protoconch and the deuteroconch (e.g., Less, 1987). In contrast, for
Tethys lepidocyclinids (Fig. 10 ![]() .3) the width of the protoconch and deuteroconch (including half the thickness of the walls) has been used (e.g., Vlerk, 1959; Drooger, 1993). Subsequent work has tended to follow a particular
scheme for a particular group so as to allow comparisons (e.g., Mitchell, 2006; Less et al., 2018; Özcan et al., 2018,
2021a). For American lepidocyclinids and their allies, different measurements have been made. These are typically the internal widths (Pwi and Dwi) and internal lengths (Pli and Dli) of the protoconch/proloculus and deuteroconch/deuterolocus (i.e., excluding the wall) and the length and width of the embryo (including the wall: Ele, Ewe). Some workers have only given parameters for the embryo, Ele and sometimes Ewe (e.g., Gorter & Vlerk, 1932; Gravel, 1933; Vaughan, 1933; Cole & Ponton, 1934; Ritten & Vermunt, 1932;Rutten, 1935; Eames et al., 1962; Caudri, 1996; Serra-Kiel et al., 2007); some workers have only given lengths (Pli, Dli) or widths (Pwi, Dwi) of the proloculus/protoconch and sometimes deuteroloculus/deuteroconch (e.g., Frost & Langenheim, 1974; Robinson & Jiang, 1995); and some workers both (e.g., Berry, 1930, 1932; Barker, 1932, 1934; Barker & Grimsdale, 1936; Cole & Bermudez, 1944; Cole & Vaughan, 1945; Cole, 1952; Butterlin, 1961; Cole, 1963; Eames et al., 1968; Colletti et al., 2019). In the Tethys the terms height and width have been used, whereas in the Americas the terms length and width have been used. In order to compare our results with those that have been made before in the Americas, we provide details of the width of the protoconch/proloculus (Pwi) and the length of the embryo (Ele) herein. These measurements are shown in Fig. 10
.3) the width of the protoconch and deuteroconch (including half the thickness of the walls) has been used (e.g., Vlerk, 1959; Drooger, 1993). Subsequent work has tended to follow a particular
scheme for a particular group so as to allow comparisons (e.g., Mitchell, 2006; Less et al., 2018; Özcan et al., 2018,
2021a). For American lepidocyclinids and their allies, different measurements have been made. These are typically the internal widths (Pwi and Dwi) and internal lengths (Pli and Dli) of the protoconch/proloculus and deuteroconch/deuterolocus (i.e., excluding the wall) and the length and width of the embryo (including the wall: Ele, Ewe). Some workers have only given parameters for the embryo, Ele and sometimes Ewe (e.g., Gorter & Vlerk, 1932; Gravel, 1933; Vaughan, 1933; Cole & Ponton, 1934; Ritten & Vermunt, 1932;Rutten, 1935; Eames et al., 1962; Caudri, 1996; Serra-Kiel et al., 2007); some workers have only given lengths (Pli, Dli) or widths (Pwi, Dwi) of the proloculus/protoconch and sometimes deuteroloculus/deuteroconch (e.g., Frost & Langenheim, 1974; Robinson & Jiang, 1995); and some workers both (e.g., Berry, 1930, 1932; Barker, 1932, 1934; Barker & Grimsdale, 1936; Cole & Bermudez, 1944; Cole & Vaughan, 1945; Cole, 1952; Butterlin, 1961; Cole, 1963; Eames et al., 1968; Colletti et al., 2019). In the Tethys the terms height and width have been used, whereas in the Americas the terms length and width have been used. In order to compare our results with those that have been made before in the Americas, we provide details of the width of the protoconch/proloculus (Pwi) and the length of the embryo (Ele) herein. These measurements are shown in Fig. 10 ![]() .4. In the Eocene taxa we are dealing with, the proloculus/protoconch and deuteroloculus/deuteroconch are generally of similar sizes; we therefore do not provide
parameters for Pli, Dli nor Dwi at the present time. To compare the tightness of coiling in the Helicosteginidae, we also construct coiling diagrams (e.g., Robinson, 1993) using measurements every 90° (Fig. 10
.4. In the Eocene taxa we are dealing with, the proloculus/protoconch and deuteroloculus/deuteroconch are generally of similar sizes; we therefore do not provide
parameters for Pli, Dli nor Dwi at the present time. To compare the tightness of coiling in the Helicosteginidae, we also construct coiling diagrams (e.g., Robinson, 1993) using measurements every 90° (Fig. 10 ![]() .5).
.5).
| Figure 10:
Measurements and coiling diagrams for selected LBFs. Note that either the
internal dimensions (i), external dimensions (e) or middle of the wall (m) are
used in different schemes. 1, measurement for Cretaceous Orbitoides
(after Hinte, 1965; Gorsel,
1978). 2, measurements for Tethys orthophragmines (after Less,
1987). 3, Measurements for Tethys lepidocyclinids (after Vlerk,
1959; Drooger, 1993). 4, measurements for American
lepidocyclinids (after various authors). Note that different measurements are
used for different groups in different biogeographic provinces. For the
Americas, we use the external length of the embryo (Ele) and the internal
width of the proloculus/protoconch (Pwi). 5, measurements for construction of
a coiling diagram: primary axis is the embryonic axis (EA), with measurements
of length (from centre of proloculus) to edge of spiral wall being made at
90° intervals (as shown); coiling diagrams are produced by plotting length
versus whorl and the gradient of the linear regression (with intercept not set
to the origin) gives the coiling parameter. |
6.2. Species concepts
Species concepts in American LBF have varied greatly among various specialists using orientated equatorial sections, with some accepting very broad species definitions (e.g., Cole, 1963; Frost & Langenheim, 1974; Robinson & Wright, 1993), while others have narrow species definitions (Cole, 1956, 1958a, 1958b; Robinson & Jiang, 1995, Robinson, 1996, 1997). This is partly due to the material available for study, with random sections in rock samples being much more difficult to identify than isolated specimens from which orientated thin sections can be cut. It may also be due to the reworking of specimens where populations from different stratigraphic levels have been mixed together (e.g., Cole, 1963; Caudri, 1975, 1996). In order to achieve a high-resolution biostratigraphy, multiple isolated specimens are clearly preferable, because this allows detailed measurements of features of the test to be made. We have found that there is generally a uniformity in equatorial sections and 4 or 5 specimens generally allow species to be identified, but (many) more specimens are desirable.
There are also the differences between a typological approach and a statistical approach to be considered. This can be illustrated by considering the evolutionary development within Eolepidina in specimens from populations used in this study. Consider, for instance a typological approach to a set of specimens using the criterion R. A 'population' might include specimens with R = 5, R = 4, and R = 3, which, under a typological approach, might be assigned to Eo. antillea and Eo. subplana, and Eo. gardnerae, respectively. Under a statistical approach, the mean value of R can be calculated, which, with values for R defined for each species) would allow a population to be placed in a single species (so in the previous population if the mean R was 4.2, it would allow the population to be assigned to Eo. subplana). Our samples, albeit often with only a limited number of specimens, calibrated using nannofossils, demonstrate that mean values of R are very useful biostratigraphy, and we define limits for R for different species in this paper.
In the following sections, we describe the various families, genera, and species of LBF that we use in the zonation scheme. We illustrate each species with numerous specimens (some well preserved, others more poorly preserved) so that our species concepts can be used by other workers.
Superfamily ASTERIGERINOIDEA Orbigny, 1839
Family AMPHISTEGINIDAE Cushman, 1927
Remarks. Loeblich and Tappan (1988) included only Amphistegina in the family Amphisteginidae. We place the genus Butterliniana gen. nov. also in this family, but further work may indicate that it belongs elsewhere.
Genus Amphistegina Orbigny, 1826
Type species. Amphistegina quoyii Orbigny, 1826 (by monotypy: Loeblich & Tappan, 1988, p. 610) from the Recent of the island of Rawack, Papua province, Indonesia.
Remarks. Species such as Amphistegina lopeztrigoi Palmer and Nummulites parvula Cushman have previously been placed in Amphistegina; in this work we place these taxa in other/new genera. Caudri's (1975) species grimsdalei (and associated forms), from the Eocene of Trinidad might be an Amphistegina, but this needs more work to confirm and is beyond the scope of this paper.
Genus Butterliniana Mitchell, E. Robinson & Özcan gen. nov.
Type species. Amphistegina cubensis Palmer, 1934, from the Eocene of Cuba.
Diagnosis. Low trochospiral coil, embryo consists of a proloculus and deuteroloculus followed by a spiral arrangement of chambers; stellar chamberlets absent; stolons between chambers without countersepta.
Origin of name. Named after Jacques Butterlin for his work on the foraminifers of the Caribbean region.
Discussion. Carefully prepared thin sections cut perpendicular to the axis on the ventral side (although invariable close to the thick wall of the test and difficult to illustrate) show that the septa divide a short distance from the umbo, but typical stellar chamberlets, as seen in Amphistegina are not present. Butterliniana differs from Amphistegina in lacking stellar chamberlets, and also in having simple stolons between chambers without the development of countersepta. We retain Butterliniana in the Amphisteginidae for the present but recognize that the lack of stellar chamberlets might require placement in a new family in the future. This can only be determined once studies of well-preserved, early forms of the Amphisteginidae have been completed.
Butterliniana cubensis (Palmer, 1934)
Previous descriptions. Palmer, 1934, p. 256-257, Figs. 15-16, Pl. 15, fig. 2; Butterlin, 1970, p. 293, Pl. 3, figs. 5-6; Robinson & Wright, 1993, p. 309, Figs. 18.3-5.
Remarks. Cushman (1919) figured four specimens as Nummulites parvula (his Pl. 4, figs. 3-6) from St. Bartholomew which are preserved in the Smithsonian. The Smithsonian catalogue records his Pl. 4, fig. 3 (USNM.MO.328183) as holotype and the other specimens (his Pl. 4, figs. 4-6: USNM.PAL27143, PAL.27143 [number duplicated], and PAL.321801) as paratypes. Cushman (1919, p. 51) clearly stated that his Pl. 4, fig. 3 was the type specimen, and that the other three specimens may not represent the same species; as such these should not be considered part of the type series or as paratypes. The species 'parvula' should therefore only be interpreted based on the holotype. The holotype is a natural, approximately equatorial section in which no indication of the original ornament of the test is visible. It has a diameter of 1.75 mm (as measured from the plate) and shows 5 whorls, with maybe 20 chambers in the final whorl.
Cole (1958a, p. 201, Pl. 25, figs. 17-19; 1969, p. 80, Pl. 16, figs. 1, 3, 5-7, 10-12; Pl. 17, fig. 5) interpreted 'parvula' based on Cushman's (1919) Pl. 4, figs. 3, 6, probably fig. 4, but not fig. 5. He interpreted 'parvula' as a senior synonym of Amphistegina lopeztrigoi Palmer, 1934. Exterior features of A. lopeztrigoi were given by Palmer (1934) and interior features of topotype material were provided by Cole and Gravell (1952). Cole's (1958a) Pl. 25, figs. 17-18, show thick-tested forms in axial section with small chambers which are similar to Cushman's (1919) Pl. 4, fig. 5; whereas his Pl. 25, fig. 19 (and Cole, 1969, Pl. 12) shows an equatorial section with 4Ľ whorls, which appears to show forwardly orientated (away from the protoconch) septal necks (countersepta) and 17 chambers in the final whorl. Cole (1958a, 1969) does not provide information on the external ornament of specimens, but he included Cushman's (1919) Pl. 4, fig. 6, in synonymy, and this form clearly lacks pustules and is smooth.
A second interpretation of Cushman's (1919) Nummulites parvula has followed Butterlin (1970), p. 294, Pl. 3, figs. 5-6 (e.g., Robinson & Wright, 1993, p. 309, Figs. 18.3-5; Serra-Kiel et al., 2007, p. 369, Pl. 2, figs. 11-12, 15). This interpretation effectively includes Cushman's (1919), Pl. 4, fig. 5, and Cole (1958a), Pl. 25, figs. 17-18. This form was named Amphistegina cubensis by Palmer (1934), p. 256-257, Figs. 15-16, Pl. 15, fig. 2.
Loeblich and Tappan (1988, p. 611) placed Cushman's Pl. 4, figs. 3, 6 (but not 5), in synonymy with 'Amphistegina' lopeztrigoi Palmer, 1934. Clearly Cushman's, 1919, Pl. 4, fig. 3, is not related to what we name here as Butterliniana, but we are unable to directly assign this specimen to lopeztrigoi because of its very weathered nature.
It is clear that an interpretation of Cushman's (1919) species Nummulites parvula is made difficult because the name-bearing type is so poorly preserved. The holotype might be close to Amphistegina lopeztrigoi as maintained by Cole (1958a, 1969), but would appear to lack ornament (as judged from other illustrated sections that might be attributed to that species) and it is difficult to interpret as that species. Similarly, one of Cushman's (1919) figured specimens (his Pl. 4, fig. 5) is equivalent to the two axial sections illustrated by Cole (1958a, Pl. 25, figs. 17-18) and was named Amphistegina cubensis by Palmer (1934). The name parvula is therefore surrounded by nomenclature issues that make it an ambiguous species of debatable affinity. It seems the best course of action would be to prepare a submission to the ICZN for the suppression of the species parvula in the combination Nummulites parvula Cushman and that Amphistegina lopeztrigoi Palmer and Amphistegina cubensis Palmer should be placed on the list of accepted names. A case will be prepared for this. We use the specific name 'cubensis' for this species here.
Butterliniana cubensis is a distinctive long-ranging species appearing in the late Ypresian and apparently becoming extinct near, or at, the top of the Eocene. Specimens vary from forms which are relatively symmetrical in axial section (e.g., Cushman, 1919, Pl. 4, fig, 5; Butterlin, 1970, Pl. 3, figs. 5-6; Cole, 1958a, Pl. 25, figs. 17-18; Robinson & Wright, 1993, Figs. 18.3-5; Serra-Kiel et al., 2007, Pl. 2, figs. 11-12, 15) to forms that are strongly asymmetrical (e.g., Palmer, 1934, Pl. 15, fig. 2). Asymmetrical forms are present at some levels in the mid and late Eocene in Jamaica, but without an analysis of populations it is not possible to determine if more than one species might be recognizable at this time. The number of chambers in the final whorl also varies significantly; without the study of populations it is not possible to determine if this is due to natural variation or might be stratigraphically significant.
| Figure 11: Butterliniana cubensis (Palmer,
1934). 1, axial section (UWIGM.WL3446-33); 2, orientated equatorial section
(UWIGM.WL3446-39); 3, equatorial section (UWIGM.WL3444-32): Chapelton
Formation (ABZ11), Albert Town. 4 (UWIGM.WL3265A), random axial section, type
Swanswick Formation (ABZ16), Swanswick House, Trelawny. 5, off-centre, axial
section (UWIGM.WL4368-2); 6, equatorial section (UWIGM.WL4368-1), turbidite
bed in chalk (ABZ11), Windsor Castle, Portland, Jamaica. |
| Figure 12: Butterliniana cubensis
(Palmer, 1934). 1, section cut parallel to the
equatorial plane on the spiral (ventral side) of the test (UWIGM.WL3446-37)
showing diverging septa but no stellar chamberlets, Chapelton Formation
(ABZ11), Albert Town, Trelawny, Jamaica. 2, axial section (UWIGM.WL2046A-8),
Chapelton Formation (ABZ9), Lilyfield, St Ann, Jamaica. 3, axial section
(UWIGM.WL3876-10), Swanswick (s.l.) Formation (ABZ15), N-S Highway 2000, St
Ann, Jamaica. 4, axial section (UWIGM.WL4052-2), Guys Hill Formation (ABZ4),
Rio Sambre Gorge, St Mary, Jamaica. 5-6, SEM photomicrographs, close to type
locality of Nummulites parvula Cushman, St Bartholomew (ABZ8); 5, view
of chambers on ventral side of test, 6, view of ventral aspect of test (in
opposite view to other specimens because of the light direction in the SEM)
showing indications of diverging septa between chambers. |
Family HELICOSTEGINIDAE Mitchell, E. Robinson & Özcan fam. nov.
Diagnosis. The family is characterised by involute to secondarily orbitoidiform taxa which have an early stage of trochospiral coiling. There is a progressive development from multiple stolons into a spiral sheet of subsidiary chamberlets on the ventral side in early forms. The development of chamberlets in the peripheral primary spire may lead to the development of an equatorial layer and in some forms a retrovert chamber may develop leading to orbitoidiform growth. In some forms the spiral sheet of equatorial chambers may form a second 'orbitoidiform' layer composed of orbitoidiform chambers on the ventral side of the equatorial layer.
Genera included. Tremastegina Brönnimann, 1950; Helicostegina Barker & Grimsdale, 1936.
Remarks. This family shows the development of advanced forms which either show low trochospiral growth with an equatorial layer or orbitoidiform growth with the ancestral form being Tremastegina. Below, we synonymise Eoconuloides with Helicostegina, and this leaves the genus Boreloides (type species Boreloides cubensis Cole & Bermúdez, 1947) as the only genus in the family Boreloididae Reiss, 1963, a family which we regard as of uncertain affinity at the present time. Further research on Boreloides is needed to determine its affinity.
Table 15 shows the length (including the walls) of the first two chambers (embryo consisting of a proloculus and a deuteroloculus; Ele) for different species belonging to the family. Table 16 shows the width (excluding the walls) of the first chamber (proloculus; Pwi) for different species belonging to the family.
Figure 13 ![]() tracks the evolutionary development of the family. Figure 14
tracks the evolutionary development of the family. Figure 14 ![]() shows the size of the embryo and protoconch in selected species of Helicostegina.
Figure 15
shows the size of the embryo and protoconch in selected species of Helicostegina.
Figure 15 ![]() shows a scatter plot of protoconch width versus embryo length, and Fig. 16 shows coiling diagrams for selected species of Helicostegina.
shows a scatter plot of protoconch width versus embryo length, and Fig. 16 shows coiling diagrams for selected species of Helicostegina.
| Figure 13:
Evolutionary development of the family Helicosteginidae through the Ypresian
and Lutetian based on the occurrences of species in Jamaica. |
Table 15: Measurements (in μm) of the lengths of the first two chambers (embryo) including the chamber walls (Ele) for species of Tremastegina and Helicostegina from Jamaica (WL numbers) and reported values (1) and our measurements from figures (2) from other locations.
| Taxon | Zone | Min | Mean | Max | SD | N | Source |
| Tremastegina lopeztrigoi | ABZ7 | 215 | - | 230 | - | - | 1 Barker & Grimsdale, 1936 |
| Helicostegina wellsi | ABZ4 | - | 140 | - | - | 1 | Unnamed lmst (WL3324) |
| H. wellsi | ABZ6 | - | 100 | - | - | - | 1 Cole & Bermudez, 1944 |
| H. wellsi | ABZ6 | 78 | 101 | 135 | 25.54 | 4 | 2 Cole & Gravell, 1952 |
| H. minor | ABZ4 | 114 | 128.7 | 146 | 16.17 | 3 | Richmond Fm (WL4052) |
| H. minor | ABZ6 | 135 | 150 | 181 | 14.97 | 11 | Stettin Fm (WL3667 & others) |
| H. gyralis | ABZ6 | 148 | 200 | 256 | 40.97 | 8 | Helicostegina beds (WL3520) |
| H. gyralis | ABZ6 | 312 | 334 | 355 | 30.4 | 2 | Palmetto Grove Fm (WL1705) |
| H. gyralis | ABZ6 | - | 235 | - | - | 1 | 2 Florida, Levin, 1957 |
| H. gyralis | ABZ6 | 200 | 316 | 455 | 111.9 | 5 | 2 Cole & Gravell, 1952 |
| H. gyralis | ABZ7 | 185 | - | 240 | - | - | 1 Barker & Grimsdale, 1936 |
| H. gyralis | ABZ7 | 253 | 275 | 297 | 31.11 | 2 | Swanswick House (WL3275) |
| H. dimorpha | ABZ7 | 246 | 308.6 | 431 | 61.65 | 7 | Preston Hill Fm (WL3309) |
| H. dimorpha | ABZ7 | - | 260 | - | - | - | 1 Barker & Grimsdale, 1936 |
| h. dimorpha | ABZ8 | 336 | 350 | 363 | 19.1 | 2 | Preston Hill Fm (WL2100) |
| H. cf. minor | ABZ7 | 135 | 167.5 | 210 | 32.79 | 4 | Preston Hill Fm (WL2098) |
| H. jamaicensis | ABZ8 | 135 | 165.6 | 190 | 20.38 | 9 | Preston Hill Fm (WL2047) |
| H. jamaicensis | ABZ8 | 175 | 176 | 176 | 0.7 | 2 | Preston Hill Fm (WL2086) |
| H. jamaicensis | ABZ8 | 188 | 189 | 189 | 0.7 | 2 | Preston Hill Fm (WL2048) |
| N. jeannemairae | ABZ9 | 278 | 340.4 | 400 | 46.4 | 10 | Rock Spring (WL3641) |
Table 16: Measurements (in μm) of the width of the first chamber (proloculus) excluding the chamber walls (Pwi) for species of Tremastegina and Helicostegina from Jamaica (WL numbers) and reported values (1) and our measurements from figures (2) from other locations.
| Taxon | Zone | Min | Mean | Max | SD | N | Source |
| Tremastegina lopeztrigoi | ABZ7 | 115 | - | 160 | - | - | 1 Barker & Grimsdale, 1936 |
| Helicostegina wellsi | ABZ4 | - | 27 | - | - | 1 | Unnamed lmst (WL3324) |
| H. wellsi | ABZ6 | 40 | - | 60 | - | - | 1 Cole & Bermudez, 1944 |
| H. wellsi | ABZ6 | 30 | 39 | 45 | 6.68 | 4 | 2 Cole & Gravell, 1952 |
| H. minor | ABZ4 | 60 | 70 | 85 | 13.05 | 3 | Richmond Fm (WL4052) |
| H. minor | ABZ6 | 62 | 76 | 89 | 8.99 | 8 | Stettin Fm (WL3667 & others) |
| H. gyralis | ABZ6 | 68 | 106 | 138 | 23.99 | 8 | Helicostegina beds (WL3520) |
| H. gyralis | ABZ6 | 162 | 165 | 167 | 3.5 | 2 | Palmetto Grove Fm (WL1705) |
| H. gyralis | ABZ6 | - | 100 | - | - | - | 2 Florida, Levin, 1957 |
| H. gyralis | ABZ6 | 105 | 176 | 300 | 87.13 | 5 | 2 Cole & Gravell, 1952 |
| H. gyralis | ABZ7 | 110 | - | 160 | - | - | 1 Barker & Grimsdale, 1936 |
| H. gyralis | ABZ7 | 147 | 162 | 176 | 20.51 | 2 | Swanswick House (WL3275) |
| H. dimorpha | ABZ7 | 86 | 172 | 266 | 64.20 | 6 | Preston Hill Fm (WL3309) |
| H. dimorpha | ABZ7 | - | 140 | - | - | - | 1 Barker & Grimsdale, 1936 |
| H. dimorpha | ABZ8 | 130 | 150 | 170 | 28.3 | 2 | Preston Hill Fm (WL2100) |
| H. jamaicensis | ABZ8 | 58 | 71 | 83 | 17.7 | 2 | Preston Hill Fm (WL2086) |
| H. jamaicensis | ABZ8 | 76 | 79 | 82 | 4.2 | 2 | Preston Hill Fm (WL2048) |
| H. cf. minor | ABZ7 | 65 | 80 | 99 | 17.3 | 3 | Preston Hill Fm (WL2098) |
| H. jamaicensis | ABZ8 | 70 | 77 | 89 | 6.8 | 6 | Preston Hill Fm (WL2047) |
| N. jeannemairae | ABZ9 | 157 | 189 | 238 | 25.89 | 10 | Rock Spring (WL3641) |
| Figure 14:
Length of the embryo (Ele) in Helicosteginidae including published material (B & G, 1936 = Barker & Grimsdale,
1936; C & G = Cole & Gravell,
1952) and populations from Jamaica (WL
numbers). |
| Figure 15:
Scatter plot of width of protoconch (Pwi in μm) versus length of embryo (Ele
in μm) for selected populations of Helicostegina from Jamaica. |
| Figure 16:
Coiling diagrams (coiling parameter versus whorls) for selected species and
populations of the Helicosteginidae from Jamaica. Grey areas are the fields
for Helicostegina minor (sample WL3667) and H. dimorpha
(samples WL3309/WL2098). |
| Figure 17:
Coiling parameter plotted against length of embryo (proloculus +
deuteroloculus including walls) for selected species of Helicostegina.
Coiling becomes looser (larger coiling parameter values) as the length of the
first two chambers increases. The regression line relates to all data points. |
Genus Tremastegina Brönnimann, 1950
Type species. Amphistegina senni Cushman (in Vaughan, 1945) from the Eocene of the Murphys Beds, Upper Scotland Group, Barbados.
Diagnosis. The genus is characterised by 'stellar' chamberlets on the ventral side in combination with forwardly (away from the protoconch) directed septal necks (counter septa) around the stolons between dorsal chambers. The
'stellar' chamberlets are connected to each other by a series of basal stolons, with the most peripheral
stolon(s) connecting to the dorsal chamber (Fig. 18 ![]() ).
).
| Figure 18:
Tremastegina senni
(Cushman in Vaughan, 1919).
1-2 (USNM.MO.625411), holotype (1, ventral; 2, dorsal), Senn locality S.34C,
Murphys Beds, Upper Scotland Group, Barbados (image from Smithsonian websire,
used by permission). 3-5 (USNM.0001), Senn locality 34C, Murphys Beds, Upper
Scotland Group, Barbados. Note that there are one or two stolons connecting
the 'dorsal' chambers (which are separated from the ventral, stellar
chamberlets by a stolon with counter septum) and a string of stolons
connecting the ventral stellar chamberlets. Also note the finely perforate
test. |
Remarks. Amphistegina differs from Tremastegina in having backwardly (towards the protoconch directed septal necks: O'Herne, 1974), whereas in Tremastegina the septal necks (counter septa) point away from the protoconch (Barker & Grimsdale, 1936; Fig. 13). These septal necks reflect the condition seen in other members of the Helicosteginidae fam. nov., but Tremastegina lacks the spiral sheet of subsidiary chamberlets developed on the ventral side in Helicostegina. Tremastegina is further distinguished from Amphistegina by the presence of the basal stolons, which we regard as the first step in the development of the spiral sheet of chamberlets seen in Helicostegina. We therefore regard Tremastegina as a primitive member of the Helicosteginidae. We provisionally recognize Tremastegina senni (Cushman in Vaughan, 1945) and T. lopeztrigoi (Palmer, 1934) as separate species and discuss this below.
Tremastegina senni (Cushman in Vaughan, 1945)
(Fig. 18 ![]() )
)
Previous descriptions. Cushman in Vaughan, 1945, p. 49, Pl. 19, figs. 1-4; Brönnimann, 1950, p. 257-265, Figs. 1-7.
Recognition. Tremastegina senni is a relatively small species of Tremastegina (typically less than 1 mm in diameter) with a small embryo.
Remarks. Cushman (in Vaughan, 1945, p. 49) stated that the holotype of A. senni came from Senn station S. 34B, but in the plate descriptions (Vaughan, 1945, Pl. 19, figs. 1-4) all the specimens were stated to come from Senn station S. 34C. Brönnimann (1950, p. 255) stated that the holotype did come from Senn station S. 34C, and that the statement by Cushman (in Vaughan, 1945, p. 49) was an error (this was probably communicated to Brönnimann by Senn who sent him material of the species to study from station S. 34C in 1946: Brönnimann, 1950, p. 255). Therefore, Brönnimann's (1950) descriptions refer to topotype material of T. senni.
T. senni was originally compared to T. lopeztrigoi by Senn (1940) who referred to the species as Amphistegina cf. lopeztrigoi following a personal communication from Vaughan (Senn, 1940, p. 1550). T. senni is, however, significantly smaller (about half the size) than T. lopeztrigoi, although this may be partly due to the loss of the final whorl in many specimens (Brönnimann, 1950, p. 259). Cole and Gravell (1952) regarded size of relatively little importance and synonymised 'A'. senni with 'A'. lopeztrigoi. Brönnimann (1950, p. 263) gives the diameter of the first chamber as 34 to 58 μm in 'A.' senni. This compares to the internal size of the first chamber in T. lopeztrigoi of 140-160 μm with walls from 10-30 μm thick (Barker & Grimsdale, 1936, p. 236). The two species may therefore be distinct.
Vaughan (1945) used the presence of 'A'. senni to argue for a middle Eocene age for that part of the Scotland Group in Barbados based on the range of 'A'. lopeztrigoi in Mexico (Barker & Grimsdale, 1936) and Florida (Cole, 1942, 1944). Both Caudri (1972) and Jones (2009) also considered that T. senni was part of the Middle Eocene fauna. However, Tremastegina has a relatively long range first appearing in the late Paleocene (Robinson & Wright, 1993) and extending up to ABZ7 in Jamaica. As noted by Cizancourt (1948) and Brönnimann (1950), the LBFs from the Scotland Group of Barbados have been strongly abraded; Brönnimann (1950, p. 259) stated that most of the specimens of T. senni had either lost their outer whorl or had it significantly worn away showing the interior structure. Furthermore, Ranikothalia occurs in the same deposits as T. senni in Barbados, and the former species is considered reworked from the late Paleocene (Caudri, 1972). Small specimens that could be attributed to T. senni occur in the late Paleocene Nonsuch Limestone of Jamaica (Robinson & Wright, 1993) adding potential support that T. senni may be an older species. There could be a progressive size increase (as well as increase in the size of the proloculus) from the late Paleocene to the mid Lutetian, which may make sequential populations of Tremastegina useful for biostratigraphy. However, until a section is found where the evolution of Tremastegina can be determined, and the variation in the size of the early chambers can be measured, it is impossible to determine if T. senni is a Paleocene, Ypresian or early Lutetian species.
Tremastegina lopeztrigoi (Palmer, 1934)
(Fig. 19 ![]() )
)
Previous descriptions. Palmer (1934), p. 255, Pl. 15, figs. 6, 8; Barker & Grimsdale, 1936, p. 233, Pl. 30, figs. 1-2; Pl. 32, figs. 1-3; Pl. 34, fig. 1; Pl. 38, fig. 3; Robinson & Wright, 1993, p. 311, Figs. 19.6-9; Hottinger, 2006, Figs. 33E-G.
Recognition. Tremastegina lopeztrigoi is a relatively large species of Tremastegina (typically more than 1 mm in diameter) with a large embryo.
Remarks. Cushman (1919) erected Nummulites parvula which Cole (1969) regarded as a senior synonym of A. lopeztrigoi. As discussed above, we regard the holotype of N. parvula as indeterminate and suggest suppression of the specific name. T. lopeztrigoi ranges at least from the mid early Eocene to the mid Lutetian (ABZ7) in Jamaica. It is the nominate taxon for ABZ3, which represents a low-diversity interval for LBFs other than for dictyoconids and orthophragmines.
| Figure 19: Tremastegina lopeztrigoi
(Palmer). 1, exterior of cotype (USNM.MO.544392),
showing stellar chamberlets, Cuba (image from Smithsonian website, used by
permission). 2-6, slightly off-centred (on the ventral side) section parallel
to the equatorial plane (WL3394A-03), Richmond Formation, Langley, St Mary,
Jamaica. 2, showing complete test. 3, detail showing counter septa directed
away from the proloculus. 4, detail showing stellar chamberlets developed on
the ventral side. 5, detail showing stellar chamberlets developed on the
ventral side and part of a pile. 6, detail showing counter septa directed away
from the proloculus. Note the finely perforate test in 3-6. |
Genus Helicostegina Barker & Grimsdale, 1936
Synonyms. Helicolepidinoides Tan, 1936 (type species Helicostegina gyralis Barker & Grimsdale, 1936) [junior synonym]; Eoconuloides Cole & Bermúdez, 1944 (type species Eoconuloides wellsi Cole & Bermúdez, 1944) [junior synonym].
Diagnosis. A lenticular to conical helicosteginid with an involute test throughout growth; the primary spire extends to the edge of the test; the alar prolongations on the dorsal side are separated by radial septa; the alar prolongations on the ventral side are represented by strongly spiral septa that merge into the spiral sheet of subsidiary chamberlets in early species, but are not recognizable in later species. The subsidiary chamberlets on the ventral side are separated by arcuate (in the direction of spiral growth) chamberlet walls. The youngest forms show the arcuate septa of the subsidiary chamberlets directed in the direction of growth, whereas in later species they show a radial (orbitoidiform) arrangement.
Remarks. Barker and Grimsdale (1936, p. 234) clearly described the morphology of this genus; their description states "Test multichambered, the earliest chambers coiled in an involute trochoid spire, the chambers of the later coils subdividing ventrally into two or more subsidiary chambers or chamberlets" and "In … H. gyralis the chamberlets are restricted to the ventral region of the test, upon which they form a continuous layer but in … [H. dimorpha] this layer of chamberlets extends peripherally as a more or less undulating compressed flange one cell thick, almost surrounding the early spiral portion which forms eccentric umbones on the dorsal and ventral surfaces of the test." The latter statement is a little misleading (as shown by Barker & Grimsdale, 1936, Pl. 32, figs. 6-7) where a narrow double-layered (in part) flange is developed, with one layer corresponding to the ventral sheet of subsidiary chamberlets and the other corresponding to the alar prolongations on the dorsal side of the test or to the primary spire or equatorial chambers. Certainly in more derived forms (e.g., H. jeannemairae sp. nov.) this would appear to be developed from the primary spire of equatorial chambers.
The relationships between Helicostegina dimorpha and H. gyralis have always been difficult to determine because of the figures of H. dimorpha given by Barker and Grimsdale (1936); yet all specimens of H. dimorpha came from Jenny station No. 1592, and all specimens of H. gyralis came from Jenny station No. 1573. Axial sections of H. dimorpha (Barker & Grimsdale, 1936, Pl. 32, figs. 6-7) show forms in which a thickened wall extends to the margin of the shell, and which contain a few peripheral subsidiary chamberlets in the outer whorl. Yet equatorial sections (Barker & Grimsdale, 1936, Pl. 34, figs. 7, 9) seem to show a relatively short spire and a 'wide flange' of orbitoidiform chambers. However, if straight lines are drawn on the axial sections corresponding to the equatorial plane, the trochospiral coiling of the test means that the equatorial sections will not pass through the spire across the whole of the section, but will also include the peripheral ventral portions of the test with subsidiary chamberlets. This is confirmed from numerous sections of Jamaican specimens, which have identical axial sections, but in relatively symmetrical tests (i.e., very low trochospiral coiling) the complete spire can be seen in equatorial section.
Adams (1987) maintained H. gyralis and H. dimorpha as separate species, but stated (p. 294) that "Cole (1960) considered that H. gyralis should be placed in synonymy with H. dimorpha …", but that "These were not, however, from the same sample or locality, and while it is true that temporal gradation between these species may occur, two individuals seen in median section do not constitute proof, neither does their synonymy warrant the possible loss of important biostratigraphical information." Yet, Adams himself figured specimens from the type locality of H. dimorpha (Jenny station No. 1592) as both H. dimorpha (Adams, 1987, Pl. 1, figs. 1-3) and H. gyralis (Adams, 1987, Pl. 1, figs. 4-8), and a specimen from the type locality of H. gyralis (Jenny station No. 1573) as H. gyralis (Adams, 1987, Pl. 1, fig. 9), thus confusing the issue.
Furthermore, Adams (1987) description of the genus Helicostegina states that the equatorial layer passes into an orbitoidiform layer with a chamber having a retrovert aperture. This is quite a different interpretation to the description given by Barker and Grimsdale (1936) who clearly stated that the lateral chambers on the ventral side of the test are divided into subsidiary chamberlets. We agree with the interpretation of Barker and Grimsdale (1936) and illustrate material herein that demonstrates this without ambiguity. Adams (1987) erroneous interpretation of the structure of H. dimorpha led him to place forms that had been separated as Helicolepidina (e.g., H. spiralis) within Helicostegina. However, the test construction in Helicolepidina is completely different to that of Helicostegina, and the two genera cannot be closely related.
Tan (1936) established the genus Helicolepidinoides for H. gyralis. Tan (1936), however, failed to realise that the difference between H. dimorpha and H. gyralis is due to the equatorial sections cut through a trochospiral test, and this is clearly indicated by the axial sections of H. dimorpha as illustrated by Barker and Grimsdale (1936) as well as by the topotype material illustrated by Adams (1987, Pl. 1, figs. 1-8). Since there is no major constructional difference between H. dimorpha and H. gyralis (other that the extension of the chamberlets into chamber lumina), and particularly since random sections are difficult to identify as either H. gyralis or H. dimorpha, there is no need for the genus Helicolepidinoides as pointed out by Adams (1987). We therefore synonymise Helicolepidinoides with Helicostegina herein.
Eoconuloides wellsi shows a similar test construction to H. gyralis and only differs in a stronger trochospiral coiling giving rise to a conical test; we therefore synonymise Eoconuloides with Helicostegina here. Cole and Bermudez (1944, p. 340) stated that Eoconuloides wellsi lacked the ventral subsidiary chamberlets seen in Helicostegina, yet they are clearly present in the holotype as figured here (Fig. 20.3). We do not regard the conical shape as of generic significance, particularly as microspheric specimens of Helicostegina minor sp. nov. show a weakly conical test. We therefore transfer E. wellsi to the genus Helicostegina, and synonymise Eoconuloides with Helicostegina herein.
The two more derived species of Helicostegina, H. jamaicensis sp. nov. and H. jeannemairae sp. nov., develop orbitoidiform growth in the equatorial layer. While this could be a justification for the establishment of new genera, they are clearly the terminations of two separate lineages. For the present, we feel it is best to leave these two terminal species within Helicostegina, particularly as the trochospiral coiling makes it difficult to distinguish orbitoidiform growth from spiral growth when a spiral layer of subsidiary chamberlets is present. We distinguish species here based on three criteria: the tightness of the coiling, the size of the embryo, and the distribution of subsidiary chamberlets.
Helicostegina wellsi (Cole & Bermúdez, 1944)
(Fig. 20 ![]() )
)
Previous descriptions and figures. Cole & Bermúdez, 1944, p. 341-342, Pl. 1, figs. 4-10; Cole & Gravell, 1952, p. 713, Pl. 92, figs. 1-10; Robinson & Wright, 1993, p. 311, Pl. 18.1-2, Pl. 19.1-5.
Recognition. The species is easily recognized because of its strongly conical form with the ventral side being highly expanded to form a tall pointed conical umbo and the dorsal side being gently convex, flat, or even gently concave. The ventral surface of the test is covered by small papillae, which are difficult to see in poorly preserved specimens. There are about 3˝ whorls in megalospheric specimens, and the final whorl has about 20 or 21 chambers. The embryo (the first two chambers - proloculus and deuteroloculus) is relatively small with a length (including the walls) of 100 to 255 μm.
Synonyms. Amphistegina elliotti Cushman & Stainforth, 1946 [junior synonym].
Remarks. Helicostegina wellsi has a test constructed in exactly the same way as H. gyralis and H. minor sp. nov. and they often occur in the same samples; the only difference is that H. wellsi is conical with a flat, weakly concave or very weakly convex dorsal side, whereas H. gyralis is lenticular. H. wellsi is a highly distinctive form which ranges from forms in which the axial height is slightly greater than the diameter to forms in which the axial height is twice that of the diameter. The relative height of the test may have some biostratigraphic significance, but this has yet to be worked out. H. wellsi is easily recognizable, although specimens may be rather small and missed if too coarse a sieve is used in sample preparation. The species can be recognized in the field with a hand lens. Amphistegina elliotti Cushman & Stainforth (1946) has the same morphology (their Pl. 20, fig. 5 shows the alar prolongations on the dorsal side, whereas their Pl. 20, fig. 6 shows the ventral layer of subsidiary chamberlets), and falls within the range of variation of H. wellsi. The ornamentation of H. wellsi consists of small papillae scattered across the whole surface of the test; this contrasts with the central crown of large pustules in H. gyralis (although early examples of H. minor from ABZ4 have an ornament closer to H. wellsi).
We use H. wellsi as the nominate taxon for ABZ4. H. wellsi has its peak abundance in ABZ4 and lower ABZ5 in Jamaica, but also occurs in very small numbers in the upper part of ABZ6 and in zones ABZ8 and ABZ9. The association of H. wellsi with H. gyralis (cited as Eoconuloides senni) and Operculinoides sp. in Calyx wells 50 and 50A in Trinidad (Caudri, 1996, Fig. 7) suggests assignment to ABZ7. Reworked(?) specimens also occur in the Upper Scotland Group in Barbados according to Brönnimann (1950, p. 264).
Kalia and Banerjee (1995) recorded 'Eoconuloides' wellsi from India. They recorded associated planktic foraminifers as indicating zone P12 and P14 (equivalent to ABZ9 to ABZ11), which is a partly younger age range than the range of H. wellsi in the Caribbean as indicated here (ABZ4 to ABZ9). We identify their Pl. 1, figs. 5 and 10 as Asterigerina sp., whereas other specimens are typical Nummulites. We doubt the occurrence of H. wellsi in India.
| Figure 20: Helicostegina wellsi
(Cole & Bermudez). 1,
off-centre axial section (WL3324-16) showing typical conical form and
chamberlets of the spiral sheet. 2, equatorial section through proloculus and
primary spire of a megalospheric specimen (WL3324-8), note that chamberlets of
the spiral sheet are not seen at this level. 3, holotype (USNM PAL 338960),
exterior view, photographed under water to show chamberlets of the spiral
sheet. 4, axial section (WL3324-7). 5, equatorial section below (i.e.,
on ventral side of) the nucleoconch (WL3324-18) showing well-developed
chamberlets. 6, tangential section parallel to side of test (WL3324-17)
showing chamberlets developed in the spiral sheet. 7 (EO.WL2044-75), axial
section, relatively low conical form with a weakly convex dorsal surface,
Chapelton Formation, Lilyfield, St Ann. 8 (EO.WL1705-b), axial section, very
low, conical specimen. 1-2, 4-6, unnamed limestone above the Pembroke Hall
Formation and below the Richmond Formation (ABZ4), Darlands, parish of St
Mary, Jamaica (ABZ4). 3, road cut on road from Managuaco to Nazareno, Habana
Province, Cuba (monospecific assemblage tentatively attributed to ABZ4 or ABZ6
here). 7, Chapelton Formation (ABZ9), Lilyfield, St Ann, Jamaica. 8, Palmetto
Grove Formation (ABZ6), near Gayle, St Mary, Jamaica. |
Helicostegina minor Mitchell, E. Robinson & Özcan, sp. nov.
Figs. 21 ![]() , 22
, 22 ![]() ,
23
,
23 ![]() ,
24
,
24 ![]() ,
25
,
25 ![]() ,
26
,
26 ![]() )
)
1945 Amphistegina senni Cushman in Vaughan, in part?
1993 Helicostegina gyralis Barker & Grimsdale; Robinson, Pl. 5, figs. 11, 13.
1993 Helicostegina gyralis Barker & Grimsdale; Robinson & Wright, p. 313-314, Fig. 18.6-8.
1993 Eoconuloides lopeztrigoi (Palmer); Robinson & Wright, in part, p. 311, Fig. 19.7 only.
Diagnosis. A species of Helicostegina with a very small proloculus followed by a smaller deuteroloculus and then a primary spiral that extends to the margin of the test; tightly coiled. Alar prolongations radial on the dorsal side and undivided; alar prolongations on the ventral side are spiral with more-or-less continuous walls and subdivided into arcuate subsidiary chamberlets.
Origin name. 'Minor' after its small proloculus.
Type specimens. Holotype: UWIGM.WL3667-5; Paratypes: UWIGM.WL3667-8, UWIGM.WL3667-7, UWIGM.WL3667-6, UWIGM.WL3667-4, UWIGM.WL3667-9; all from sample WL3667, Stettin Formation (ABZ6), Stettin, Trelawny, Jamaica.
Description. Microspheric forms have a diameter of up to 2.0 mm, megalospheric forms have a diameter of up to 1.6 mm. The ornament consists of a central crown of pustules with small pustules along the septa. Microspheric forms have a very small proloculus, with about five whorls; there are 33 chambers in the final whorl. Megalospheric specimens have the first two chambers (proloculus and deuteroloculus) with a length of 114-181 um (including the walls), with means of 129 um (sample WL4052) and 150 um (sample WL3667). The internal width of the proloculus (Pwi) ranges from 60-89 μm (Table 16). The first two chambers are followed by 3˝ to 4˝ whorls, with 27 to 31 chambers in the final whorl. Chambers on the dorsal side undivided with simple arcuate septa; dorsal chambers visible from ventral side (where they form the marginal series of chambers). Septa on the ventral side of the test can be traced through the subsidiary chamberlets and are strongly spirally arranged. Alar prolongations of chambers on the ventral side divided into subsidiary chamberlets, with arcuate walls, and extending up to the spire of the previous whorl. Axial sections range from relatively symmetrical to asymmetrical forms. Septa on the dorsal side are only occasionally visible in orientated axial sections, so that most of the alar prolongations on the dorsal side of the test are 'open.' In axial section, the walls of the ventral chamberlets are closely spaced and fill the alar prolongations on the ventral side of the test.
Remarks. H. minor can be distinguished from T. lopeztrigoi by the presence of subsidiary chamberlets in or replacing the alar prolongations on the ventral side, which extend up to the keel of the primary spire of the previous whorl. The well-marked septa forming the alar prolongations are retained on the ventral side such that a spiral pattern is quite obvious. This is lost in later species where a true spiral sheet of subsidiary chamberlets is developed which have a more typical orbitoidiform arrangement of the chamberlets. H. minor differs from H. wellsi by the presence of large tubercles on the surface of the test that are concentrated at the centre (crown) and the fine series of papillae that mark the courses of the septa across the flanks of the test on the dorsal side.
| Figure 21: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov., megalospheric specimens. 1 (UWIGM.WL4052-6), 2 (UWIGM.WL4052-7),
3
(UWIGM.WL4052-1), 4 (UWIGM.WL4052-8), 6 (UWIGM.WL4052-4), 7
(UWIGM.WL4052-9):
equatorial sections. 5 (UWIGM.WL4052-5): axial section. Sample WL4052 (ABZ4),
Richmond Formation, Rio Sambre Gorge, St Mary, Jamaica. |
| Figure 22: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov., megalospheric specimens. 1 (UWIGM.WL4052-5): detail of axial
section. 2 (UWIGM.WL4052-8), 3-4 (UWIGM.WL4052-9): details of equatorial
sections. Sample WL4052 (ABZ4), Richmond Formation, Rio Sambre Gorge, St Mary,
Jamaica. |
| Figure 23: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov., megalospheric specimens. 1 (UWIGM.WL3667-8), paratype; 2
(UWIGM.WL3667-7), paratype; 3 (UWIGM.WL3667-5), holotype; 4 (UWIGM.WL3667-6),
paratype; 5 (UWIGM.WL3667-4), paratype; 6 (UWIGM.WL3667-9) paratype:
equatorial sections. Sample WL3667, Stettin Formation (ABZ6), Stettin,
Trelawny, Jamaica. |
| Figure 24: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov., megalospheric specimens. 1 (UWIGM.WL3667-10), 2 (UWIGM.WL3667-11),
3
(UWIGM.WL3667-12): equatorial sections. 4 (UWIGM.WL3667-1), 5
(UWIGM.WL3667-14): sections cut through the spiral sheet of subsidiary
chamberlets. 6 (UWIGM.WL3667-2), 7 (UWIGM.WL3667-3): axial sections. Sample
WL3667, Stettin Formation (ABZ6), Stettin, Trelawny, Jamaica. |
| Figure 25: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov., megalospheric specimens. 1 (UWIGM.WL3667-3): detail of axial
section. 2 (UWIGM.WL3667-5), 3 (UWIGM.WL3667-9), 4 (UWIGM.WL3667-4),
5
(UWIGM.WL3667-10), 6 (UWIGM.WL3667-11): details of equatorial sections. Sample
WL3667, Stettin Formation (ABZ6), Stettin, Trelawny, Jamaica. |
| Figure 26: Helicostegina minor Mitchell,
E. Robinson & Özcan
sp. nov. 1-2 (UWIGM.WL3458-1), microspheric form, equatorial section, Stettin
Formation (ABZ6), Coleyville, Jamaica. 3 (BMNH.P38378), exterior view (ventral
side), partially eroded to show chamberlets, Murphys' Beds, Scotland
Formation, Barbados. 4 (ER1315), SEM, dorsal side showing crown of pustules
and smaller pustules extending along septa, Stettin Formation (ABZ6), Stettin,
Jamaica. 5 (ER1315), exterior view (ventral side) of decortitized specimen
showing ventral sheet of subsidiary chamberlets, Stettin Formation (ABZ6),
Stettin, Jamaica. |
Helicostegina minor ranges from ABZ4 to ABZ6. It is replaced in the upper part of ABZ6 by H. gyralis. The species is recorded from Jamaica (Robinson & Wright, 1993, herein) and Barbados (Fig. 26.3).
Helicostegina cf. minor Mitchell, E. Robinson & Özcan, sp. nov.
(Fig. 27 ![]() )
)
Remarks. Four specimens from middle ABZ7 show an equatorial section with a small proloculus and small deuteroloculus combined with a spiral wall that extends to the margin of the test. We do not have an axial section so cannot determine if they have an incipient double layer, and we attribute these specimens to Helicostegina cf. minor sp. nov. Further material will need to be collected to determine whether this species should be retained in H. minor sp. nov. or placed in a new species transitional between H. minor sp. nov. and H. jamaicaensis sp. nov.
| Figure 27: Helicostegina
cf. minor Mitchell, E. Robinson
& Özcan sp. nov. megalospheric specimens. 1
(EO.WL2098-117), 2 (EO.WL2098-118), 3 (EO.WL2098-119): equatorial sections.
Preston Hill Formation (ABZ7), Farm Turn, St Mary, Jamaica. |
Helicostegina gyralis Barker & Grimsdale, 1936
Figs. 28 ![]() ,
29
,
29 ![]() ,
30
,
30 ![]() ,
31
,
31 ![]() ,
32
,
32 ![]() , 33
, 33 ![]() )
)
Previous descriptions and figures. Barker & Grimsdale, 1936, p. 236-237, Pl. 30, figs. 3-5; Pl. 32, figs. 4-5; Pl. 34, figs. 2-6; Pl. 37, fig. 6; Cole & Gravell, 1952, p. 713, Pl. 92, figs. 11-21.
Recognition. Helicostegina gyralis is a lenticular species, but shows a low trochospiral coil such that in many specimens it is difficult to cut true equatorial sections, and the sections often pass from the primary spire in the centre of the test into part of the spiral sheet of subsidiary chamberlets. Barker and Grimsdale (1936, p. 236) stated that the first two chambers have a length of 185-240 μm and there are about 25-30 chambers in the 'final whorl'. Measurements of populations from Jamaica are given in Tables 15 and 16. The dorsal surface of the test is ornamented with coarse piles (pillars) on the central area of the test with finer papillae aligned along the septa separating the alar prolongations/chambers. Axial sections of most specimens lack chamberlets at the periphery of the test, although a few specimens (e.g., Barker & Grimsdale, 1936, Pl. 32, fig. 4: reproduced in Fig. 28.4 here) show a few chamberlets at the periphery, but not the clear double row of chamberlets seen in H. dimorpha.
Remarks. H. gyralis can be distinguished from T. lopeztrigoi by the presence of a spiral sheet of subsidiary chamberlets on the ventral side. These can be seen during section preparation even if the resulting equatorial section apparently shows none (which is the case in relatively symmetrical specimens). H. gyralis differs from H. minor in having a larger embryo and in being less tightly coiled. H. gyralis differs from H. wellsi by its shape, and by the presence of large tubercles on the surface of the test that are concentrated at the test's centre and the fine series of papillae that mark the courses of the septa across the flanks of the test on the dorsal side.
The two populations of H. gyralis (upper ABZ6 and lower ABZ7) from Jamaica plot in different areas on Coiling Parameter vs. Embryo Length diagrams (Fig. 17). Since we have not measured the Coiling Parameter on specimens of the type population of H. gyralis from Mexico, and since the coiling parameter is highly dependent on reproduction of figures at the correct scale in publications (unless a scale bar is present), we do not separate the Jamaican populations at the present time. Pwi of specimens from upper ABZ6 is smaller than for lower ABZ7 (Table 16, Fig. 14), and the specimen of H. gyralis from Florida figured by Levin (1957) falls within the range of the upper ABZ6 population from Jamaica; this may be of stratigraphic significance and populations from Florida need to be measured to determine this.
H. gyralis ranges through the middle of ABZ6 to the lower part of ABZ7 in Jamaica. The type locality of H. gyralis is Jenny Station No. 1573 (11 km southeast of Sabaneta, state of Veracruz, Mexico: Barker & Grimsdale, 1936); this locality has also yielded Pseudolepidina trimera (Barker & Grimsdale, 1937) and Operculinoides jennyi (Barker, 1939) and can therefore be assigned to lower ABZ7. H. gyralis is widely distributed across the American region being reported in Mexico (Barker & Grimsdale, 1936), Florida (Cole, 1942; Levin, 1957), probably Venezuela (Raadshooven, 1951), Cuba (Cole & Bermudez, 1944; Cole & Gravel, 1952) and Jamaica (herein). Previous records from the Stettin Formation of Jamaica (e.g., Robinson, 1993; Robinson & Wright, 1993) are here placed in Helicostegina minor sp. nov.
| Figure 28:
Reproduction of the original illustrations of Helicostegina gyralis
from Barker & Grimsdale
(1936). 1 (Pl. 34, fig. 2), equatorial section. 2 (Pl. 34, fig. 3) section cut
on ventral side of the equatorial plane showing ventral sheet of chamberlets.
3 (Pl. 34, fig. 6), equatorial section. 4 (Pl. 32, fig. 4) axial section.
5
(Pl. 32, fig. 5) axial section. Note that in 4 the ventral chamberlets extend
into the periphery of the test, whereas in 5 they do not. Guayabal Formation, Jenny
Section No. 1573, 11 km southeast of Sabaneta, state of Veracruz, Mexico.
Scale bar = 1 mm. |
| Figure 29: Helicostegina gyralis Barker
& Grimsdale 1936. Topotypes. 1 (BMNH.P.39151-2),
section cut on the dorsal side of the equatorial layer showing the pillars. 2
(BMNH.P.39151-1), equatorial section. 3 (BMNH.P.39150-3), equatorial section.
4 (BMNH.P.39152-1), axial section. 5 (BMNH.P.39152), axial section.
6
(BMNH.P.39152-1), axial section. All from Eleven km SE of Sabaneta, State of
Veracruz (Collection of Dr. H. Jenny, No. 1573),
Mexico. Note that the lumina of the chambers do not contain subsidiary
chamberlets in 5 and 6, but do in 4. |
| Figure 30: Helicostegina gyralis Barker & Grimsdale, 1936.
1 (UWIGM.WL3520-9), equatorial section. 2 (UWIGM.WL3520-2), equatorial section.
3 (UWIGM.WL3520-10), equatorial section. 4 (UWIGM.WL3520-14), equatorial section.
5 (UWIGM.WL3520-13), equatorial section. 6 (UWIGM.WL3520-16), section parallel to the equatorial plane passing through the ventral sheet of subsidiary
chamberlets. 'Helicostegina-beds' (upper ABZ6), road from Springfield to Mount
Horeb, parish of St James, Jamaica. |
| Figure 31: Helicostegina gyralis Barker & Grimsdale, 1936. Details of embryos.
1 (UWIGM.WL3520-9), equatorial section. 2 (UWIGM.WL3520-13), equatorial section.
3 (UWIGM.WL3520-2), equatorial section. 4 (UWIGM.WL3520-14), equatorial section.
5 (UWIGM.WL3520-10), equatorial section. 6 (UWIGM.WL3520-16), section parallel to the equatorial plane passing through the ventral sheet of subsidiary
chamberlets. 'Helicostegina-beds' (ABZ6), road from Springfield to Mount Horeb, parish of St James, Jamaica. |
| Figure 32: Helicostegina gyralis Barker & Grimsdale, 1936.
1-2
(UWIGM.WL3520-3), axial section. 3 (UWIGM.WL3520-7), axial section. 4
(UWIGM.WL3520-5), axial section. Note that the subsidiary chamberlets on the ventral side only extend to the periphery of the previous whorl.
'Helicostegina-beds' (ABZ6), road from Springfield to Mount Horeb, parish of St James,
Jamaica. |
| Figure 33: Helicostegina gyralis Barker
& Grimsdale, 1936. 1-2 (EO.WL1705-17),
microspheric specimen, equatorial section. 3-4 (EO.WL1705-18), megalospheric
specimen, equatorial section. Upper Palmetto Grove Formation (sample WL1705),
upper ABZ6, near Gayle, St Mary, Jamaica. |
Helicostegina dimorpha Barker & Grimsdale, 1936
Previous descriptions and figures. Barker & Grimsdale. 1936, p. 235, Pl. 30, fig. 6; Pl. 32, figs. 6-7; Pl. 33, fig. 9; Pl. 34, figs. 7, 9; Pl. 37, fig. 3.
Recognition. According to Barker and Grimsdale (1936), the spire of H. dimorpha has a length of 18 to 21 chambers, inclusive of the proloculus. However, because of the trochospiral coiling it is probable that this is only the part of the spire that is cut in the section and in reality the spire is longer extending to the margin of the test. This is confirmed by Adams' (1987) Pl. 1, fig. 2, a topotype, which shows the spiral wall extending to the periphery of the test. Barker and Grimsdale (1936, p. 236) reported that the first two chambers (proloculus and deuteroloculus) had a length of 240 μm. There is no orbitoidiform layer and the apparent presence of one is due to equatorial sections cutting the spiral sheet of subsidiary chamberlets because of the trochospiral coiling of the test. Axial sections show the presence of subsidiary chamberlets in the peripheral area of the dorsal chambers, although the primary septa of the spire continue to the margin of the test.
Remarks. The type material of Barker and Grimsdale (1936) is confusing because the two equatorial sections pass from the primary spire into the spiral sheet of subsidiary chamberlets and this makes it look like the primary spire passes into an orbitoidiform layer. This is clearly not the case in the two axial sections that are presented (Barker & Grimsdale, 1936, Pl. 32, figs. 6-7, reproduced here in Fig. 35.3-4). Adams (1987) illustrated topotype material that shows that the primary spire extends to the margin of the test, yet even he talked about an orbitoidal layer. H. dimorpha occurs in upper ABZ7 (samples WL2098 and WL3309/WL3309A) and lower ABZ8 (sample WL2100) in Jamaica.
| Figure 34: Reproduction of illustrations of Helicostegina dimorpha Barker & Grimsdale, 1936 (Pls. 32 and 34).
1-2, equatorial sections. 3-4, axial sections. Nine km SE of
Sabaneta, Veracruz, Mexico. Scale bar 1 mm. |
| Figure 35: Helicostegina dimorpha Barker & Grimsdale, 1936.
1 (EO.WL2098-14), 2 (EO.WL2098-112), 3 (EO.WL2098-86),
4 (EO.WL2098-20), 5 (EO.WL2098-123), 6 (EO.WL2098-111): equatorial sections. Sample WL2098 (ABZ7), Farm Turn, St Mary,
Jamaica. |
| Figure 36: Helicostegina dimorpha Barker & Grimsdale, 1936.
1 (EO.WL2098-19), 2 (EO.WL2098-16): equatorial sections. 3 (EO.WL2098-122),
4 (EO.WL2098-110): axial sections. 4 shows a slightly longer flange, but occurs in a population which is assigned to H. dimorpha. Sample WL2098 (ABZ7), Farm Turn, St Mary,
Jamaica. |
| Figure 37:
Helicostegina dimorpha Barker & Grimsdale, 1936.
1, 4 (UWIGM.WL3309A-12), equatorial section. 2-3 (UWIGM.WL3309A-11) equatorial section.
5 (UWIGM.WL3309A-14), axial section. 6 (UWIGM.WL3309A-13), axial section. 7 (UWIGM.WL3309A-6), axial section.
8 (UWIGM.WL3309A-7), axial section. Preston Hill Formation (ABZ7), Farm Turn (sample WL3309A), parish of St Mary,
Jamaica. |
Helicostegina dimorpha has only been found in the middle and upper parts of ABZ7 and lower ABZ8 in Jamaica, where it is locally common. It is distinguished from H. gyralis by the acquisition of subsidiary chamberlets in the periphery of the chamber which, with the equatorial chamberlets, produces an incipient double layer.
Helicostegina jamaicensis Mitchell, E. Robinson & Özcan, sp. nov.
Diagnosis. A species of Helicostegina with a small embryo (proloculus and deuteroloculus), a thickened spiral wall which terminates before the margin of the test, and with two layers (equatorial, and ventral sheet of subsidiary chamberlets) in the flange. Low lateral chambers present during adult growth.
Type specimens. Holotype: EO.WL2047-51; Paratypes: EO.WL2047-21, EO.WL2047-22, EO.WL2047-23, EO.WL2047-26, EO.WL2047-4, EO.WL2047-41, EO.WL2047-53, EO.WL2047-57, EO.WL2047-57, UWIGM.WL3532C-07, UWIGM.WL3532C-08. Samples WL2047 and WL3532C (both from the same level), Preston Hill Formation, Lilyfield, St Ann, Jamaica.
Description. Only megalospheric specimens seen. Megalospheric specimens are up to 1.5 mm in diameter and 0.8 mm thick. The surface is covered with regularly arranged piles (pillars). The equatorial layer consists of the embryo and a primary spire of 17-20 chambers (including the proloculus and deuteroloculus) occupying 1˝ to 1ľ whorls. The length of the embryo (Ele) ranges from 135 to 190 μm (including the walls). The primary spire has a thickened wall which does not extend to the margin of the test. The last chamber in the primary spire has two apertures and gives rise to orbitoidiform growth of the equatorial layer. In axial sections the alar prolongations of the spiral chambers on the dorsal side are not divided into chamberlets. The ventral side of the test is divided into subsidiary chamberlets, and this extends as a layer parallel to the equatorial layer giving the test a double flanged appearance. 'Lateral' chamberlets (i.e., additional layers of dorsal and ventral chamberlets) are added in the adult stage, but do not extend onto the flange which just has a thickened wall.
Remarks. The species is distinguished from H. gyralis and H. dimorpha by its small embryo, and in the spiral wall not reaching the margin of the test. It is distinguished from H. minor sp. nov. and H. cf. minor sp. nov. by the fact that the spiral wall does not reach the edge of the test whereas in the latter forms the spiral wall extends to the margin of the test; thus orbitoidiform growth is neither achieved in H. minor sp. nov. nor in H. cf. minor sp. nov., but is in H. jamaicensis sp. nov.
Distribution. Only recorded from ABZ8, upper Lutetian, along the deep-water facies of the Preston Hill Formation, Lilyfield, St Ann, northern Jamaica.
| Figure 38: Helicostegina jamaicensis Mitchell, E. Robinson & Özcan sp.
nov. 1 (EO.WL2047-21, paratype), 2 (EO.WL2047-22, paratype), 3 (EO.WL2047-23,
paratype), 4 (EO.WL2047-53, paratype), 5 (EO.WL2047-51, holotype),
6 (EO.WL2047-26,
paratype): equatorial sections. Note that the thickened spiral wall terminates before the margin of the test. Sample WL2047 (ABZ8), Preston Hill Formation,
Lilyfield, St Ann, Jamaica. |
| Figure 39: Helicostegina jamaicensis Mitchell, E. Robinson & Özcan sp.
nov. Paratypes. 1 (UWIGM.WL3532C-08, paratype), equatorial section. 2 (UWIGM.WL3532C-07),
3 (EO.WL2047-4), 4 (EO.WL2047-57), 5 (EO.WL2047-41), 6 (EO.WL2047-57), axial sections. Samples WL2047 and WL3532C (ABZ8), Preston Hill Formation,
Lilyfield, St Ann, Jamaica. |
| Figure 40: Helicostegina jamaicensis Mitchell, E. Robinson & Özcan sp.
nov. Paratypes. 1 (EO.WL2047-4), 2 (EO.WL2047-57), details of axial sections; note the addition of lateral chambers.
3 (EO.WL2047-53), details of axial section showing the embryo
(proloculus and deuteroloculus). Sample WL2047 (ABZ8), Preston Hill Formation,
Lilyfield, St Ann, Jamaica. |
Helicostegina jeannemairae E. Robinson, Mitchell & Özcan, sp. nov.
(Figs. 41 ![]() ,
42
,
42 ![]() ,
43
,
43 ![]() ,
44
,
44 ![]() ,
45
,
45 ![]() ,
46
,
46 ![]() ,
47
,
47 ![]() )
)
Diagnosis. Early growth stage as in Heterostegina dimorpha with alar prolongations open on the dorsal side and merging into subsidiary chamberlets on the ventral side. Adult test shows purely annular growth (alar prolongations absent in adult test) with an orbitoidiform (equatorial) layer and a ventral layer of subsidiary chamberlets giving a two-layered structure in axial section.
Type specimens. Holotype: UWIGM.WL3641-13 (Figs. 41 ![]() .3,
43
.3,
43 ![]() .6). Paratypes: UWIGM.WL3641-2 to 6; UWIGM.UWLGM3641-8 to 9; UWIGM.WL3641-12; UWIGM.WL3641-14, WL3641-16 to 21; UWIGM.WL3641-23 to 25. All from Chapelton Formation, Yellow Limestone (sample WL3641), Upper Rock
Spring, parish of Westmoreland, Jamaica. There are several hundred additional paratypes in sample WL3641 that have not been numbered or cut.
.6). Paratypes: UWIGM.WL3641-2 to 6; UWIGM.UWLGM3641-8 to 9; UWIGM.WL3641-12; UWIGM.WL3641-14, WL3641-16 to 21; UWIGM.WL3641-23 to 25. All from Chapelton Formation, Yellow Limestone (sample WL3641), Upper Rock
Spring, parish of Westmoreland, Jamaica. There are several hundred additional paratypes in sample WL3641 that have not been numbered or cut.
Description. Tests are flat lenticular with swollen umbos and a diameter of up to 6.4 mm. The ornament consists of small scattered piles (pillars) across the whole test. The population is dominated by megalospheric specimens with very rare microspheric specimens. Microspheric specimens are only slightly larger (6.4 mm) than the megalospheric specimens (largest specimen is 5.7 mm).
Microspheric specimens have a very small proloculus that is followed by a primary spire of about 40 chambers. The spire is terminated by a retrovert chamber with two solons and this gives rise to the orbitoidiform layer.
The megalospheric specimens have an embryo consisting of two chambers with a thickened chamber wall. The length of the embryo ranges from 278 to 400 μm (mean 340, SD 46.4, n = 10), with the proloculus being slightly larger than the deuteroloculus. The primary spire consists of one and a half whorls, is involute, and contains 15 to 23 chambers including the embryo (mean 19.1, st. dev. 2.88, n = 10). The primary spire contains subsidiary chamberlets on the ventral side, whereas the dorsal chambers extend onto the acute keel of the spire (this is developed as in Helicostegina dimorpha). The primary spire has a thickened wall which terminates at the point where orbitoidiform growth commences. The primary spire gives rise to two spirals (primary and secondary) through a chamber with two stolons (one forward directed and one retrovertly directed), and these spires coil around the periphery of the test and end in a closing chamber. Each chamber in the secondary spires gives rise to additional chamberlets which initiates orbitoidiform growth of the equatorial layer. The orbitoidiform chamberlets are low arcuate. The orbitoidiform layer occupies about four-fifths of the diameter of the test in large specimens. The ventral side of the test is composed of a single sheet of subsidiary chamberlets that develop from the sheet of secondary chamberlets on the ventral side of the primary spire. The chamberlets in the ventral subsidiary sheet have a typical orbitoidal form with two stolons as in the equatorial layer. Once orbitoidiform growth is initiated, there are no more alar prolongations and the test wall is compact without lateral chambers which results in the formation of a wide double flange. This indicates that growth of the adult test was facilitated by annular addition of orbitoidiform chamberlets to both the equatorial layer and to the ventral sheet of subsidiary chamberlets.
Remarks. The species is named after the late Jeanne Mair Robinson who, with her children, discovered the first specimens of this species at Point, St James (Jamaica), although the type locality is situated in Westmoreland Jamaica where free specimens are abundant. The population from Point
(Fig. 41 ![]() ) has more whorls in the spire than specimens from the type population and many have a
one-biop morphology (Fig. 41
) has more whorls in the spire than specimens from the type population and many have a
one-biop morphology (Fig. 41 ![]() .2-5, 7) in axial section. We regard these specimens as transitional to H. dimorpha, but because of the wide flange we place them in H. jeannemairae sp. nov. The species is common in some areas of western Jamaica (St. James and Westmoreland), but has not been found in the marlstones (Preston Hill Formation) or limestones (Chapelton Formation) along the North Coast Belt of Jamaica where H. jamaicensis sp. nov. is common. It may be that H. jeannemairae sp. nov. developed as a shallow-water species and H. jamaicensis sp. nov. as a deeper-water species.
.2-5, 7) in axial section. We regard these specimens as transitional to H. dimorpha, but because of the wide flange we place them in H. jeannemairae sp. nov. The species is common in some areas of western Jamaica (St. James and Westmoreland), but has not been found in the marlstones (Preston Hill Formation) or limestones (Chapelton Formation) along the North Coast Belt of Jamaica where H. jamaicensis sp. nov. is common. It may be that H. jeannemairae sp. nov. developed as a shallow-water species and H. jamaicensis sp. nov. as a deeper-water species.
Distribution. Helicostegina jeannemairae sp. nov. has only been found in sections in western Jamaica that lack zonally diagnostic forms. In St James it occurs in the Yellow Limestone above levels yielding H. cf. dimorpha and Eolepidina (probably lower ABZ8) and below levels yielding Eo. gardnerae (ABZ10), so an attribution to ABZ9 seems likely. At Rock Spring (Westmoreland) it occurs with only rare specimens of Operculinoides sp.
| Figure 41: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Specimens in random orientations in thin section of rock. 1, 6, off-centred equatorial sections.
2-5, 7-8,
off-centre axial sections. Note that some off-centred axial sections (3-7) show a single
'wing' and all axial sections show more whorls that in the type population of H. jeannemairae; we regard the Point population as transitional to H. dimorpha. Note that the flange is more strongly developed in specimens from sample ER3031 that in ER1575, with specimens from ER1575 be closer to H. dimorpha. Yellow Limestone, samples ER1575 and ER3031 (?lower ABZ9), Point, St James,
Jamaica. |
| Figure 42: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population. Equatorial sections of microspheric (1: UWIGM.WL3641-21) and megalospheric
(2, UWIGM.WL3641-23; 3, UWIGM.WL3641-13; 4, UWIGM.WL3641-6;
5, UWIGM.WL3641-8; 6, UWIGM.WL3641-20). Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
| Figure 43: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population. Equatorial sections of megalospheric individuals (1, UWIGM.WL3641-2;
2, UWIGM.WL3641-14; 3, UWIGM.WL3641-24; 4, UWIGM.WL3641-25). Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
| Figure 44: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population, details of spire. Equatorial sections of microspheric (2,
4, UWIGM.WL3641-21) and megalospheric individuals (1, UWIGM.WL3641-23;
3, UWIGM.WL3641-17; 5, UWIGM.WL3641-14; 6, UWIGM.WL3641-13). Note closing chamber developed from two spires coiling around the shell wall of the primary spire. Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
| Figure 45: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population, details of spire. Equatorial sections of megalospheric individuals
(1, UWIGM.WL3641-9; 2, UWIGM.WL3641-8; 3, UWIGM.WL3641-6;
4, UWIGM.WL3641-4; 5. UWIGM.WL3641-2; 6, UWIGM.WL3641-3). Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
| Figure 46: Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population. Axial sections of megalospheric individuals (1-3, UWIGM.WL3641-18;
4, UWIGM.WL3641-5; 5, UWIGM.WL3641-12).
sw, spiral wall; oc, orbitoidiform chamber; sc, subsidiary chamberlet. Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
| Figure 47:
Helicostegina jeannemairae E. Robinson, Mitchell & Özcan sp.
nov. Type population. 1 (UWIGM.WL3641-23), detail showing stolons in the equatorial layer of
chamberlets. 2 (UWIGM.WL3641-19), detail showing stolons in the subsidiary ventral layer of
chamberlets. 3 (UWIGM.WL3641-19), section cut parallel to the equatorial plane through a rather symmetrical specimen showing the development of the subsidiary chamberlets in the ventral sheet of subsidiary chamberlets (a typical equatorial layer was ground away during the making of the section).
4 (UWIGM.WL3641-16), tangential section through the ventral sheet of subsidiary
chamberlets. Chapelton Formation (?upper ABZ9), Yellow Limestone (sample WL3641), Upper Rock Spring, parish of Westmoreland,
Jamaica. |
Family HELICOLEPIDINIDAE Tan, 1936
Diagnosis. The family is characterised by forms showing lenticular growth with the spiral and umbilical sides generally not determinable in axial sections. The equatorial layer consists of a primary spiral of chambers with equatorial chamberlets developing on the outside of the primary spiral. The alar prolongations on the ventral (spiral) side of the test are not subdivided into subsidiary chamberlets as in the Helicosteginidae, and lateral chamberlets are present on each side of the equatorial layer.
Genera included. Helicosteginopsis Caudri, 1975; and Helicolepidina Tobler, 1922.
Remarks. The family is separated from the Helicosteginidae by the lack of a subsidiary sheet of chamberlets on the ventral (spiral) side of the test and the presence of lateral chamberlets on either side of the equatorial layer. We do not describe these forms as they are rare in our samples, but do provide our best estimates of ages.
Genus Helicosteginopsis Caudri, 1975
Type species. Helicostegina soldadensis Grimsdale in Vaughan & Cole, 1941.
Helicosteginopsis soldadensis (Grimsdale in Vaughan & Cole, 1941)
(Fig. 48 ![]() .1, 3-4)
.1, 3-4)
Previous descriptions. Grimsdale in Vaughan & Cole (1941); Caudri (1975, 1996); Adams (1987).
Age. The material from Trinidad comes from levels that have been assigned to the upper Eocene (Vaughan & Cole, 1941; Caudri, 1975, 1996), but contains reworked material from Paleocene and middle Eocene levels such that the age of the species cannot be determined from Trinidad. In Margarita Island (Caudri, 1974), Venezuela, H. soldadensis occurs in sample P.J.B.145 with L. ariana, Orbitoina spp. and Helicolepidina spiralis trinitatis [= nortoni Vaughan] and can be assigned to ABZ12. In Panama, Cole (1941) recorded H. soldadensis with Eu. chaperi and H. ocalana indicating ABZ16. We provisionally give a range estimate of zones ABZ12 to ABZ16.
Genus Helicolepidina Tobler, 1922
Type species. Lepidocyclina (Helicolepidina) spiralis Tobler, 1922.
Synonyms. Helicocyclina Tan, 1936 (type species: Helicolepidina paucispira Barker & Grimsdale, 1936).
Helicolepidina sp. A
(Fig. 48 ![]() .2)
.2)
Remarks. Barker and Grimsdale (1936, Pl. 35, fig. 4) figured a specimen that they ascribed to Eulinderina guayabalensis var. regularis Barker & Grimsdale, but differs in having some equatorial chamberlets developing on the outside of the primary spire (Fig. 49.4). This form does not have a thick spiral wall. We ascribe this specimen (from the ABZ7- ABZ8 boundary zone) to Helicolepidina sp. A.
Helicolepidina spiralis Tobler, 1922, and Helicolepidina nortoni Vaughan, 1936
(Fig. 49 ![]() .1)
.1)
This group of species has been discussed by Raadshooven (1951) who recognized six forms (morphotypes) with some stratigraphic significance; his forms C and D correspond to H. spiralis Tobler and his forms E and F, to H. nortoni (= H. spiralis trinitatis Brönnimann). Caudri (1975) recorded H. nortoni (her H. spiralis trinitatis) from ABZ12 on Margarita Island, Venezuela. Cole (1952) recorded H. nortoni (as H. spiralis) from Panama in samples lacking both Pl. tobleri and H. ocalana (ABZ15) and in samples with H. ocalana (ABZ16). Thus we infer a range of zones ABZ12 to ABZ16 to H. nortoni. We assign tentative ages of ABZ7 to ABZ8 for H. sp. 1 (forms A and B of Raadshooven, 1951) and ABZ8 and ABZ10 for H. spiralis, based on Raadshooven (1951); further work is needed to confirm the ranges.
Helicolepidina polygyralis Barker, 1932
(Fig. 49 ![]() .3-4)
.3-4)
Remarks. Barker (1932, 1934) placed megalospheric forms with diameters of 1.6 - 2.4 mm and 2.5 - 3 whorls in this species; it therefore has more whorls than other species of Helicolepidina. Caudri (1974) recorded H. polygyralis from ABZ9 on Margarita Island, Venezuela.
Helicolepidina paucispira Barker & Grimsdale, 1936
(Fig. 49 ![]() .2)
.2)
Remarks. Barker and Grimsdale referred H. paucispira to the upper Eocene. Cole (1945, p. 17) reported this species from water wells at Tallahassee in the Upper Eocene of Florida which can be assigned to either upper ABZ15 or ABZ16 due to the presence of Eu. chaperi.
| Figure 48:
1, 3-4, Helicosteginopsis soldadensis (Grimsdale):
1 (BMNH.P49515-16), megalospheric specimen, equatorial section; 3
(BMNH.P49515-16), megalospheric specimen, axial section: note symmetrical
development of lateral chambers and single spiral (equatorial) layer. 4
(BMNH.P51953), megalospheric specimen, equatorial section; Soldado Rock,
Trinidad (T. F. Grimsdale collection). 2, Helicolepidina
sp. (reproduced from figure of Barker & Grimsdale,
1936, Pl. 35, fig. 4, Eulinderina guayabalensis var. regularis),
megalospheric form, equatorial section, close to village of Yecautla, Veracruz,
Mexico. |
| Figure 49:
1, Helicolepidina spiralis Tobler, 1922 (BMNH.P30247), megalospheric specimen, equatorial section; El Alto, north-western Peru.
2, Helicolepidina paucispira Barker & Grimsdale, 1936 (BMNH.P47311), megalospheric specimen, equatorial section; Ecuador.
3-4 (BMNH.P51955), Helicolepidina polygyralis Barker, 1932:
3, megalospheric specimen, axial section (note the single spiral-equatorial layer);
4, megalospheric specimen, equatorial section: Ancon Peninsula, Santa Elena Peninsula,
Ecuador. |
Family PSEUDOLEPIDINIDAE Mitchell, E. Robinson & Özcan fam. nov.
Diagnosis. The family is characterised by the development of a double equatorial layer together with lateral chambers on both the dorsal (umbilical) and ventral (spiral) sides. The double equatorial layer develops from an initial single equatorial layer.
Remarks. The biloculine embryo is joined by a further chamber on the ventral side that we consider to be part of the embryo. There is/are one or two PACs developed in addition to this third embryonic chamber. Pseudolepidina developed a lepidocyclinid orbitoidiform morphology before the Lepidocyclinidae and therefore cannot be derived from that stock. We also see it difficult to generate Pseudolepidina from the Helicosteginidae or the Helicolepidinae, again because of its advanced equatorial plan. The subfamily appears to be a separate radiation, presumably from an unknown rotalid ancestor.
Genera included. Pseudolepidina Barker & Grimsdale, 1937 (type species: Pseudolepidina trimera Barker & Grimsdale, 1937); Triplalepidina Vaughan & Cole, 1938 (type species: Triplalepidina veracruziana Vaughan & Cole, 1938). We do not consider Triplalepidina in this work as we do not have specimens of this genus.
Genus Pseudolepidina Barker & Grimsdale, 1937
Type species. Pseudolepidina trimera Barker & Grimsdale, 1937 from the Lutetian of Jenny station 1573, 11 km SE of Sabaneta, state of Veracruz, Mexico.
Diagnosis. A genus of Pseudolepidinidae developing a double equatorial layer, but lacking a compact shell layer between the two parts of the equatorial layer; with a 'third PAC' developed; and well-developed lateral chamberlets.
Remarks. Pseudolepidina is distinguished from Triplalepidina in lacking a compact shell layer between the two parts of the double equatorial layer.
Pseudolepidina trimera Barker & Grimsdale, 1937
(Figs. 50 ![]() ,
51
,
51 ![]() ,
52
,
52 ![]() ,
53
,
53 ![]() ,
54
,
54 ![]() ,
55
,
55 ![]() )
)
Previous descriptions and figures. Barker & Grimsdale, 1937, p. 172, Pl. V. figs. 1-3; Pl. VIII, figs. 1-5; Adams, 1987, p. 299, Pl. 3, figs. 11-16.
Recognition. The double equatorial layer as seen in axial sections is diagnostic, particularly when the third PAC is visible in such sections. Oblique sections of Lepidocyclina or Orbitoina may give the impression of showing two equatorial layers and can be confused with Pseudolepidina, so the presence of the additional PAC is critical to identifications of specimens in random orientations in limestone thin sections. There is considerable variation in the embryo and PACs with specimens having one or two PACs when cut in equatorial section; the position of the 'third PAC' (which we regard as part of the embryo) seems variable and it may also be seen in some equatorial sections.
Remarks. The type series of Barker and
Grimsdale (1937) came from Jenny station 1573, 11 km SE of
Sabaneta, state of Veracruz, Mexico, where it was associated with H. gyralis and Operculinoides jennyi (Barker & Grimsdale, 1936, 1937; Barker, 1939) and can be assigned to ABZ7. Adams (1987, Pl. 3, figs. 15-16, rephotographed in Fig. 54.8 here) figured a specimen from Goldschmidt station 837, close to the village of Yecuatla, state of Veracruz, Mexico, where it was associated with Eulinderina regularis and Eolepidina antillea (reported as Eulinderina semiradiata: Barker & Grimsdale, 1936); this can be assigned to the ABZ7- ABZ8 transition interval. Robinson (2000) recorded Ps. trimera from the type Swanswick Formation and from a level in the Chapelton Formation of Jamaica. We have specimens from the type Swanswick Formation at Swanswick House and this can be assigned to ABZ7. The occurrence in the Chapelton Formation needs confirmation, and we would assign this to the early Bartonian (maybe ABZ12 or ABZ13)
- we do not show this occurrence on the stratigraphic chart (Fig. 6 ![]() ) since it needs confirmation.
) since it needs confirmation.
| Figure 50: Pseudolepidina trimera Barker & Grimsdale, 1937.
1-3 (UWIGM.WL3572-09), megalospheric forms, equatorial section showing two PACs and four unequal spirals of epiembryonic chambers.
4-5 (UWIGM.WL3572-07), microspheric form, equatorial section. 6 (UWIGM.WL3572-03), megalospheric form showing one obvious PAC.
7-8 (UWIGM.WL3572-08), megalospheric form with deformed embryo showing three chambers with four PACs. Yellow Limestone (ABZ7), Grants Pen, St Thomas,
Jamaica. |
| Figure 51: Pseudolepidina trimera Barker & Grimsdale, 1937. Axial sections of megalospheric
(1, 3-8) and microspheric (2) forms. 1 (UWIGM.WL3572-06), 2 (UWIGM.WL3572-05),
3 (UWIGM.WL3572-01), Yellow Limestone (sample WL3572), Grants Pen (attributed to ABZ7), St Thomas, Jamaica.
4 (UWIGM.3275-4), 5 (UWIGM.3275-11), 6 (UWIGM.3275-19),
7 (UWIGM.3275A-14), Yellow Limestone, type section of the
'Swanswick' Formation (samples WL3275 and WL3275A), Swanswick House (ABZ7),
Trelwany, Jamaica. 8 (BMNH.PS1960), locality with Eulinderina regularis and Eolepidina guayabalensis (ABZ7), close to the village of
Yecuatla, state of Veracruz, Mexico. |
| Figure 52: Pseudolepidina trimera Barker & Grimsdale, 1937. Megalospheric forms.
1 (UWIGM.WL3275-9), 2 (UWIGM.WL3275-6), 3 (UWIGM.WL3275-13),
4 (UWIGM.WL3275-3), 5 (UWIGM.WL3275-14), 6 (UWIGM.WL3275-12): Yellow Limestone, type section of the
'Swanswick' Formation (sample WL3275), Swanswick House (ABZ7), Trelwany,
Jamaica. |
| Figure 53: Pseudolepidina trimera Barker & Grimsdale, 1937. Embryo in equatorial section (some specimens rotated so that
peri-embryonic chambers can be shown). 1 (UWIGM.WL3275-3), specimen with
'two' PACs; 2 (UWIGM.WL3275-8), specimen with 'two' PACS; 3 (UWIGM.WL3275-6), specimen with irregular
peri-embryonic chambers); 4 (UWIGM.WL3275-9) specimen with irregular embryo;
5 (UWIGM.WL3275-12), specimen with one large chamber (third PAC?); 6 (UWIGM.WL3275-10), specimen with one? PAC: Yellow Limestone, type section of the
'Swanswick' Formation (sample WL3275), Swanswick House (ABZ7), Trelwany,
Jamaica. |
| Figure 54: Pseudolepidina trimera Barker & Grimsdale, 1937. Embryo in equatorial section (some specimens rotated so that
peri-embryonic chambers can be shown). 1 (UWIGM.WL3275-18), specimen with
'two' PACs and irregular spires; 2 (UWIGM.WL3275-19), specimen with irregular embryo;
3 (UWIGM.WL3275A-11), specimen with one? PAC); 4 (UWIGM.WL3275A-8) specimen with irregular embryo and two PACs;
5 (UWIGM.WL3275A-10), specimen with two PACs; 6 (UWIGM.WL3275A-18), Pseudolepidina cf. trimera, specimen with one PAC and a clear asymmetrical spire developed from it: Yellow Limestone, type section of the
'Swanswick' Formation (samples WL3275 and WL3275A), Swanswick House (ABZ7),
Trelwany, Jamaica. |
| Figure 55: Pseudolepidina trimera Barker & Grimsdale, 1937. Equatorial chamberlets shown in equatorial section.
1 (UWIGM.WL3275-12), 2 (UWIGM.WL3275-13), 3 (UWIGM.WL3275-6),
4 (UWIGM.WL3275-5): Yellow Limestone, type section of the
'Swanswick' Formation (sample WL3275), Swanswick House (ABZ7), Trelwany, Jamaica. Chamberlets are broadly arranged in radial rows (at least for short sections) and consist of both wide and narrow forms.
Radially, chamberlets get progressively wider until they are replaced by two rows of
chamberlets. |
Family LEPIDOCYCLINIDAE Scheffen, 1932
Diagnosis. A family developed from a spiral (eulinderine) ancestor without alar prolongations. We recognize two subfamilies (Lepidocyclininae and Orbitoininae subfam. nov.) which each show a progressive reduction in the primary spire from eulinderine to polylepidine to lepidocycline to nephrolepidine or eulepidine. All but the most primitive forms have 'lateral' chamberlets developed.
Remarks. The Lepidocyclinidae show clear and separate radiations of genera that are obvious from the detailed work we have undertaken in Jamaica. This has been suggested previously for the Eocene lepidocyclines (e.g., Grimsdale, 1959; Vlerk, 1959; Sirotti, 1983; Butterlin, 1987), but age relationships between different forms have previously been poorly constrained. From the samples collected from Jamaica we can see two clear radiations, which we place in separate subfamilies: the Lepidocyclininae and Orbitoininae. The largest problem is with homeomorphy, as each radiation gives rise to advanced lepidocycline morphologies which are difficult to distinguish from each other on morphological features (particularly in random thin sections). This classification has the advantage of retaining all lepidocycline-like forms in a single family, but separating them into their separate radiations. Although the orbitoidiform plan has been used to define families or subfamilies previously (e.g., Loeblich & Tappan, 1964, 1988; Adams, 1987; Boudagher-Fadel, 2018), we believe that individual phylogenetic radiations should not be arbitrarily split into subfamilies based on the acquisition of an orbitoidiform growth plan. If such a course was taken, at least four separate subfamilies would be required in the Lepidocyclinidae. We therefore give subfamily status to the two individual evolving clades that show a progressive change from cyclic to orbitoidiform growth.
Each radiation shows a progressive change up section. Early forms have an embryo consisting of a proloculus and a deuteroloculus followed by a short spire of large, peri-embryonic chambers (eulinderine forms) that gives rise to a chamber (R) with a retrovert aperture from which two spires (primary and secondary) of relatively large peri-embryonic chambers emanate. Up section, the position or R progressively decreases until the development of first one PAC (unilepidine) and then two PACs (polylepidine). With one PAC two spirals of peri-embryonic chambers extend around the embryo and meet at a closing chamber. With two PACs three or four, asymmetrical spirals of peri-embryonic chambers extend around the embryo with two closing chambers (Eva, 1980). A true orbitoidiform growth plan (lepidocycline) is achieved when the four spires emanating from the PACs become bilaterally symmetrical about the embryonic (protoconch-deuteroconch) axis. Subsequent development sees additional stolons connecting adauxilary chambers to the protoconch and/or deuteroconch, and a reduction in the size (and consequent increase in number) of the peri-embryonic chambers. Further development sees an increase in the width of the deuteroconch which then partially envelops the protoconch (nephrolepidine to eulepidine forms).
Equatorial chambers and chamberlets can be divided into several types. The first two chambers represent the embryo. In the earliest forms there is only a single stolon connecting the proloculus to the deuteroloculus. Subsequent chambers in the primary spire of early forms have two stolons: a single stolon connecting back to the embryo, or earlier chambers and a single stolon connecting to subsequent chambers. Once an R chamber, one PAC, or two PACs are developed (or for that matter adauxilary chambers), these chambers and any subsequently peri-embryonic chambers have three stolons, one connecting to the chamber that the chamber arises from and two connecting to subsequent chambers/chamberlets (note, that the presence of adauxiliary chambers adds to the numbers of stolons connecting to the protoconch and/or deuteroconch). The closing chamber(s) mark(s) a change to four or six stolons and all subsequent chamberlets after this have either four stolons or six stolons. We define equatorial chambers as those representing the embryo and the peri-embryonic chambers (including any closing chambers) and equatorial chamberlets as those that arise from the peri-embryonic and closing chambers.
There are three broad arrangements of equatorial chamberlets when viewed in equatorial sections. The first has a predominantly radial form and occurs in Eolepidina and Orb. rdouvillei (similar patterns are seen in Pseudolepidina). The second only shows arcs of equatorial chamberlets with neither a prominent radial nor a prominent concentric arrangement of chamberlets and occurs in Lep. ariana. The third is characterised by a series of concentric or annular rings of equatorial chamberlets and occurs in Lep. ocalana, Eulepidina chaperi and Eulepidina undosa. These arrangements result from the way that chamberlets are arranged, from the chamberlet shapes and from the distribution of stolons (four versus six) connecting chamberlets. The form of these arrangements is not of generic significance, since they repeatedly occur at similar stages in the development of different lineages.
Subfamily LEPIDOCYCLININAE Scheffen, 1932
The breakdown of the Lepidocyclininae into genera has been demonstrated by Robinson & Jiang (1995) and Robinson (1996, 1997) and we follow their scheme here, with modifications, recognizing eulinderine (R > 7, no lateral chambers), eolepidine (R > 3, with lateral chambers), unilepidine (R = 3), polylepidine (R = 2), lepidocycline (R = 2) and eulepidine morphologies (Fig. 56 ![]() ). Different stages of development are given generic status and have useful biostratigraphic value. The described genera are somewhat arbitrary and are retained here for historical reasons rather than because they are fully merited. We discuss the use of generic names under each genus below. The proloculus/protoconch and embryo sizes in the Lepidocyclininae gets larger through the Eocene, although there are periods of stasis (Figs. 57
). Different stages of development are given generic status and have useful biostratigraphic value. The described genera are somewhat arbitrary and are retained here for historical reasons rather than because they are fully merited. We discuss the use of generic names under each genus below. The proloculus/protoconch and embryo sizes in the Lepidocyclininae gets larger through the Eocene, although there are periods of stasis (Figs. 57 ![]() ,
58
,
58 ![]() ,
59
,
59 ![]() ), and the embryo is generally larger than the embryo size of co-occurring populations of the Orbitoininae (other than for Pliolepidina).
), and the embryo is generally larger than the embryo size of co-occurring populations of the Orbitoininae (other than for Pliolepidina).
Frost and Langenheim (1974) synonymised many species in this group (although their figured populations can often be assigned to individual species as recognized here) and their scheme clearly obscures the biostratigraphic value demonstrated by Robinson and Jiang (1995) and Robinson (1996, 1997). We do not follow Frost and Langenheim's (1974) synonyms here, but develop our own classification of these forms. Many studies are based on random sections in thin sections of rock samples (e.g., Robinson & Wright, 1993; Serra-Kiel et al., 2007); yet many random sections are difficult to place in the evolutionary scheme as understood here and, if possible, workers should try to obtain free specimens to produce orientated sections for better biostratigraphic resolution. An alternative is to cut large numbers of random sections hoping to get some more-or-less equatorial sections by chance. Yet some genera (e.g., Eulinderina) and species (e.g., Lepidocyclina macdonaldi) are very distinctive in random sections, and provide some useful biostratigraphic resolution.
| Figure 56: Evolution and stratigraphic distribution of the Lepidocyclininae showing typical equatorial arrangements of chambers for each
species. |
| Figure 57:
Range in R values and length of embryos (Ele) in populations of Lepidocyclininae from Jamaica (and St Bartholomew). Note periods of
stasis in embryo size punctuated by significant changes at the top of ABZ9, ABZ11 and ABZ14 (all of which correspond to sequence boundaries) and within ABZ16 (which is due to the appearance of Eu. chaperi). |
| Figure 58:
Range in R values and width of proloculus/protoconch (Pwi) in populations of Lepidocyclininae from Jamaica (and St Bartholomew). Note periods of
stasis in proloculus/protoconch size punctuated by significant changes at the top of ABZ9, ABZ11 and ABZ14 (all of which correspond to sequence boundaries) and within ABZ16 (which is due to the appearance of Eu. chaperi). |
| Figure 59: Scatter plot of length of embryo
(Ele) versus width of proloculus/protoconch (Pwi) for populations of Lepidocyclininae from Jamaica (and St Bartholomew) with fields for species indicated. Note that there is extensive overlap of the different species
fields. |
Robinson (1996, 1997) introduced mean R values (R), being the mean number of chambers (including the protoloculus and deuteroloculus) to the first retrovert chamber in populations of eulinderine and polylepidine forms. He showed that R values fell within narrow limits within populations, and many additional populations analysed from Jamaica in this work show a similar narrow limit for R in any one sample. We therefore define limits for mean R size for identifying species here (Table 17). We also provide measurements for the embryo (combined length of the proloculus and deuterolocus or protoconch and deuteroconch, including the thickness of the walls; Ele, Table 18) and width of the proloculus/protoconch excluding walls (Pwi, Table 19) for different species as given in the literature and in measured specimens/populations for Jamaica. The transition from typical four-spired lepidocyclines (L. ariana) to lepidocyclines with adauxilary chambers (L. macdonaldi, L. ocalana), to nephrolepidine-like forms (Eu. chaperi) is not straight forward and needs to be investigated in more detail. Other recent papers (e.g., Molina et al., 2015, Pl. 1.5; Hohenegger & Torres Silva, 2020, Fig. 2) confuse isolepidine forms (L. ocalana) with adauxilary chambers for nephrolepidine forms (Eu. chaperi).
Numerous generic names have been proposed for the Lepidocyclininae, many of which have been synonymised. The names retained here break the lineage(s) up into a series of genera, and the generic names have a long historical usage. Our placement of species into different genera is based on our revised definitions of the genera as indicated below.
Genera included. Eulinderina Barker & Grimsdale, 1936; Eolepidina Tan, 1939; Polylepidina Vaughan, 1924; Lepidocyclina Gümbel, 1868; Eulepidina Douvillé, 1911.
Table 17: Criteria for distinguishing species of Eulinderina, Eolepidina and Polylepidina based on mean R values.
| Species | Range of mean R values | 'Lateral' chamberlets? |
| Eulinderina guayabalensis (Nuttall) | 7 - 9 | No |
| Eulinderina regularis (Barker & Grimsdale) | 5.5 - 7 | No |
| Eolepidina antillea (Cushman) | 4.5 - 5.5 | Yes |
| Eolepidina subplana (Barker & Grimsdale) | 3.5 - 4.5 | Yes |
| Eolepidina gardnerae (Cole) | 2.2 - 3.5 | Yes |
| Polylepidina chiapasensis (Vaughan) | 2 - 2.2 | Yes |
| Lepidocyclina spp. | 2 | Yes |
Table 18: Measurements of mean R and the length of the nucleoconch (or first two chambers) including the walls for specimens/species of Lepidocyclininae from Jamaica and elsewhere. Measurements include nucleoconch (chamber) walls.
| Species | Sample | R | Zone | Length of nucleus (μm) | Source | |||
| range | mean | SD | n | |||||
| Eu. guayabalensis | Mexico | 7 - 9 | ABZ7 | 160 - 230 | - | - | - | B & G, 1936 |
| Eu. guayabalensis | ER152/WL4869 | 8.25 | ABZ7 | 242 - 332 | 269 | 51.0 | 4 | Our measurements |
| Eu. regularis | Mexico | 6 - 7 | ABZ7/6 | 220 - 260 | - | - | - | B & G, 1936 |
| Eo. 'semiradiata' | Mexico | 3 - 5 | ABZ7/6 | 330 - 430 | - | - | - | B & G, 1936 |
| Eo. antillea | WL2047A | 5 | ABZ8 | 196 - 307 | 262 | 44.6 | 9 | Our measurements |
| Eo. antillea | ER161 | 5 | ABZ8 | 308 - 323 | 316 | 21.3 | 2 | Our measurements |
| Eo. antillea | WL2046 | 4.7 | ABZ8 | 215 - 310 | 243 | 33.5 | 9 | Our measurements |
| Eo. subplana | WL3444 | 4 | ABZ9 | 220 - 300 | 279 | 56.6 | 4 | Our measurements |
| Eo. subplana | ER1485 | 3.82 | ABZ9 | 304 - 429 | 371 | 38.8 | 9 | Our measurements |
| Eo. subplana | WL2044 | 4 | ABZ9 | 170 - 289 | 220 | 38.2 | 8 | Our measurements |
| Eo. gardnerae | WL4449 † | 3 | ABZ14 | 263 - 367 | 319 | 40.3 | 5 | Our measurements |
| Eo. gardnerae | WL2921 | 3 | ABZ10 | 381 - 477 | 414 | 54.9 | 3 | Our measurements |
| Eo. gardnerae | Content Well | 2.7 | ABZ10 | 317 - 386 | 358 | 25.9 | 6 | Our measurements |
| Eo. gardnerae | WL2837 | 2.5 | ABZ10 | 340 - 457 | 399 | 41.0 | 6 | Our measurements |
| Po. chiapasensis | WL3446 | 2 | ABZ11 | 318 - 439 | 381 | 42.1 | 10 | Our measurements |
| Po. chiapasensis | WL4195 | 2 | ABZ11 | 338 - 525 | 412 | 67.8 | 6 | Our measurements |
| Po. chiapasensis | WL3597 | 2 | ABZ11 | 319 - 454 | 381 | 52.8 | 5 | Our measurements |
| Lep. ariana | WL3571 | 2 | ABZ12 | 371 - 463 | 422 | 41.9 | 5 | Our measurements |
| Lep. ariana | WL4241 | 2 | ABZ12 | 425 - 547 | 464 | 53.4 | 5 | Our measurements |
| Lep. ariana | Margarita | 2 | ABZ12 | 385 - 605 | 449 | 77.4 | 8 | *Butterlin, 1970 |
| Lep ocalana | WL2635 | 2 | ABZ13 | 479 | - | - | 1 | Our measurements |
| Lep. ocalana | WL3507 | 2 | ABZ13 | 371 - 606 | 459 | 67.5 | 11 | Our measurements |
| Lep. ocalana | WL3876/3876A | 2 | ABZ15 | 532 - 798 | 632 | 107.6 | 5 | Our measurements |
| L. montgomeriensis | Moodys Branch | 2 | ABZ15 | 433 - 833 | - | - | - | G & H, 1935 |
| Lep. macdonaldi | WL4449 | 2 | ABZ15 | 488 - 766 | 660 | 124.1 | 4 | Our measurements |
| Lep. macdonaldi | Jamaica | 2 | 520 - 720 | 620 | - | 3 | *Cole, 1956 | |
| Lep. ocalana | WL4095 | 2 | ABZ15 | 673 | - | - | 1 | Our measurements |
| Lep. ocalana | WL3931 | 2 | ABZ16 | 665 - 731 | 690 | 25.9 | 5 | Our measurements |
| Eulepidina chaperi | WL4865 | 2 | ABZ16 | 749 - 912 | 849 | 76.5 | 4 | Our measurements |
* specimens measured from illustrations in publications (which is dependent on accuracy of plate reproduction when only magnifications were given). † specimens regarded as reworked. B & G, 1936 = Barker & Grimsdale, 1936; G & H, 1935 = Gravell & Hanna, 1935.
Table 19: Measurements of mean R and the width of the proloculus/protoconch (Pwi) excluding the walls for specimens/species of Lepidocyclininae from Jamaica and elsewhere. Measurements include nucleoconch (chamber) walls. † specimens regarded as reworked.
| Species | Sample | R | Zone | Width of Proloculus/Protoconch (μm) | Source | |||
| range | mean | SD | n | |||||
| Eu. guayabalensis | Mexico | 7 - 9 | ABZ7 | 85 - 115 | - | - | - | B & G, 1936 |
| Eu. guayabalensis | ER152/WL4869 | 8.25 | ABZ7 | 212 - 329 | 259 | 49.7 | 4 | Our measurements |
| Eu. regularis | Mexico | 6 - 7 | ABZ7/6 | 120 | - | - | - | B & G, 1936 |
| Eo. 'semiradiata' | Mexico | 3 - 5 | ABZ7/6 | 180 - 220 | - | - | - | B & G, 1936 |
| Eo. antillea | WL2047 | 5 | ABZ8 | 196 - 307 | 262 | 44.6 | 9 | Our measurements |
| Eo. antillea | ER161 | 5 | ABZ8 | 269 - 324 | 297 | 38.9 | 2 | Our measurements |
| Eo. antillea | WL2046 | 4.7 | ABZ8 | 215 - 310 | 246 | 34.6 | 8 | Our measurements |
| Eo. subplana | WL3444 | 4 | ABZ9 | 220 - 330 | 279 | 56.6 | 4 | Our measurements |
| Eo. subplana | ER1485 | 3.82 | ABZ9 | 288 - 429 | 356 | 40.9 | 14 | Our measurements |
| Eo. subplana | WL2044 | 4 | ABZ9 | 170 - 289 | 220 | 38.2 | 8 | Our measurements |
| Eo. gardnerae | WL4449 † | 3 | ABZ14 | 263 - 367 | 319 | 40.3 | 5 | Our measurements |
| Eo. gardnerae | WL2921 | 3 | ABZ10 | 381 - 477 | 414 | 54.9 | 3 | Our measurements |
| Eo. gardnerae | Content Well | 2.7 | ABZ10 | 317 - 386 | 358 | 25.9 | 6 | Our measurements |
| Eo. gardnerae | WL2837 | 2.5 | ABZ10 | 340 - 457 | 399 | 41.0 | 6 | Our measurements |
| Po. chiapasensis | WL3446 | 2 | ABZ11 | 318 - 439 | 381 | 42.1 | 10 | Our measurements |
| Po. chiapasensis | WL4195 | 2 | ABZ11 | 338 - 525 | 412 | 67.8 | 6 | Our measurements |
| Po. chiapasensis | WL3597 | 2 | ABZ11 | 319 - 454 | 381 | 52.8 | 5 | Our measurements |
| Lep. ariana | WL3571 | 2 | ABZ12 | 407 - 540 | 489 | 50.7 | 7 | Our measurements |
| Lep. ariana | WL4241 | 2 | ABZ12 | 425 - 547 | 464 | 53.4 | 5 | Our measurements |
| Lep ocalana | WL2635 | 2 | ABZ13 | 400 - 480 | 440 | 56.6 | 2 | Our measurements |
| Lep. ocalana | WL3507 | 2 | ABZ13 | 371 - 606 | 459 | 67.5 | 11 | Our measurements |
| Lep. ocalana | WL3876/3876A | 2 | ABZ15 | 532 - 798 | 632 | 107.6 | 5 | Our measurements |
| Lep. macdonaldi | WL4449 | 2 | ABZ15 | 488 - 815 | 673 | 100.7 | 8 | Our measurements |
| Lep. ocalana | WL4095 | 2 | ABZ15 | - | 653 | - | 1 | Our measurements |
| Lep. ocalana | WL3931 | 2 | ABZ16 | 665 - 731 | 690 | 25.9 | 5 | Our measurements |
| Lep. ocalana | WL4865 | 2 | ABZ16 | - | 689 | - | 1 | Our measurements |
| Eulepidina chaperi | WL4865 | 2 | ABZ16 | 749 - 912 | 849 | 76.5 | 4 | Our measurements |
Genus Eulinderina Barker & Grimsdale, 1936
Diagnosis. Lepidocyclininid with a primary spire comprising six or more chambers before the first retrovert chamber (i.e., the chamber that gives rise to two separate spires); lateral chamberlets absent.
Remarks. We formally define limits of R for the two species of Eulinderina here for use for biostratigraphic purposes (Table 17). The two taxa assigned to Eulinderina, Eu. guayabalensis (Nuttall, 1930) (R = 7 - 9) and Eu. regularis Barker & Grimsdale, 1936 (R = 5.5 - 7), appear to be rare outside of Mexico and are not currently incorporated into the zonation. It is likely that the Lepidocyclininae initially evolved in Mexico (close to where Jamaica was situated at the time: Pindell, 2009; Mitchell, 2020) during the Lutetian and subsequently spread across the American LBF bioprovince.
Eulinderina guayabalensis (Nuttall, 1930)
Previous descriptions. Barker & Grimsdale, 1936, p. 238, Pl. 32, figs. 8-9; Pl. 34, figs. 8, 10-11.
Recognition. Eu. guayabalensis is recognized by its long primary spire before the development of a retrovert aperture, and also by its lack of lateral chamberlets and a laminate shell seen in axial section.
Remarks. Eu. guayabalensis appears to be rare outside of Mexico. We have several specimens from the Preston Hill Formation (samples ER152 and WL4869) from Grants Town (St Mary, Jamaica) with R values of 7 and 9 (mean = 8.5, n = 4). Barker and Grimsdale (1936) give the length of the embryo of Eu. guayabalensis as 160-230 μm, which is notably smaller than specimens of this species from Jamaica that have an embryo with a length of 212-329 μm. The age of the material described by Barker and Grimsdale (1936) is difficult to assign to our biostratigraphic zonation because they did not list associated species (particularly nummulitids); it could be that their material was from ABZ6 rather than ABZ7, but this will have to await studies of topotype material. Samples ER152 and WL4869, both from the Preston Hill Formation at Grants Town, yield Eu. guayabalensis in association with Operculinoides and orthophragmines (Robinson & Mitchell, 1999, p. 35; herein) and are assigned to ABZ7 and NP15b. For the present, we assume that Eu. guayabalensis is likely restricted to ABZ7.
| Figure 60:
A-B, Eulinderina guayabalensis (Nuttall,
1930), megalospheric forms. 1-2 (ER152a), equatorial section (R = 7). 3
(ER152b), equatorial section (R = 9). 4-5 (WL4869-1), equatorial
section (R = 9). Preston Hill Formation (samples ER152 and WL4869), Grants
Town, parish of St Mary, Jamaica. |
| Figure 61:
1-5
Eulinderina guayabalensis (Nuttall, 1930) and 58.6-8 Eulinderina regularis Barker & Grimsdale, 1936.
1 (topotype: BMNH.P51958), megalospheric specimen, equatorial section (R = 8).
2 (reproduction of original specimen of Barker & Grimsdale, 1936, Pl. 34, fig. 8) equatorial section (R = 9),
3 (BMNH.P51959), megalospheric specimen, axial section (note absence of lateral chambers).
4 (reproduction of original specimen of Barker & Grimsdale, 1936, Pl. 32, fig. 9), axial section of megalospheric form.
5 (reproduction of original specimen of Barker & Grimsdale, 1936, Pl. 32, fig. 8), axial section of microspheric form.
6 -7 (reproduction of original specimen of Barker & Grimsdale, 1936, Pl. 35,
figs. 10-11), axial sections of megalospheric specimens. 8 (topotype: BMNH.PS1960), equatorial section (R = 8).
1-5, close to the village of Yecuatla, state of Veracruz, Mexico. 6-8,
Mexican Eagle Oil Company Well No. 5 (4,590 ft depth), Mexico. 5, Union Oil Company Miahuapan No. 3-A well (3,100 to 4,100 ft depth),
Mexico. |
Eulinderina regularis Barker & Grimsdale, 1936
(Fig. 61 ![]() .6-9)
.6-9)
Previous descriptions. Barker & Grimsdale, 1936, p. 238, Pl. 30, figs. 7, 9; Pl. 32, figs. 10-11; Pl. 35, figs. 4-5; Pl. 37, fig. 5.
Recognition. Eu. regularis is recognized by its shorter primary spire before the development of a retrovert aperture, and also by its lack of lateral chamberlets and a compact shell seen in axial section.
Remarks. Barker and Grimsdale (1936) recorded Eu. regularis (their Eu. guayabalensis var. regularis) from the same locality as Eolepidina semiradiata (= Eolepidina antillea); this suggests that this locality (assuming the material was from the same sample, rather than locality) contains an assemblage of specimens at the transition between Eulinderina and Eolepidina. We have not recorded Eu. regularis from Jamaica. In Mexico, its occurrence with Eolepidina antillea (reported as Eu. semiradiata) and Ps. trimera (Barker & Grimsdale, 1936; Adams, 1987) indicates it occurs around the ABZ7 to ABZ8 transition.
Genus Eolepidina Tan, 1939
Type species. Eulinderina semiradiata Barker & Grimsdale, 1936 (junior synonym of Lepidocyclina antillea Cushman, 1919) from the Lutetian, close to the village of Yecuatla, state of Veracruz, Mexico (Barker & Grimsdale, 1936).
Diagnosis. A Lepidocyclininae with a short primary spire of equatorial chambers (from 3 to 6), none or one PAC, and vacuole-like lateral chambers.
Discussion. Eolepidina is distinguished from Eulinderina by the presence of lateral chamberlets. The type locality of Eo. semiradiata (= Eo. antillea) appears to show specimens in transition between Eulinderina and Eolepidina. Because the presence or absence of lateral chamberlets is easily determined in random sections (as seen in thin sections of rock samples), the separation of the two genera is certainly warranted for biostratigraphic purposes.
We recognize three species of Eolepidina: Eo. antillea, Eo. subplana and Eo. gardnerae. The last species has one PAC and has been placed in Polylepidina by some workers (e.g., Cole, 1929). We define Polylepidina as containing two PACs with three or four asymmetric spires of peri-embryonic chambers, which leads us to place the species gardnerae in the genus Eolepidina.
Eolepidina antillea (Cushman, 1919)
(Figs. 62 ![]() ,
63
,
63 ![]() ,
64
,
64 ![]() ,
65
,
65 ![]() ,
66
,
66 ![]() )
)
Previous descriptions and figures. Cole, 1960, Pl. 11, figs. 6, 13; Pl. 12, figs. 1-8.
Remarks. Eo. antillea has an ornament consisting of scattered pustules, and we define that R = 5.5 - 4.5. The secondary spire of chambers developed from the first retrovert chamber is often short with small chambers and contrasts with the primary spire developed from the first retrovert chamber which is long with larger chambers. We follow the revision of Eo. antillea as proposed by Cole (1960) and Robinson (1996). We synonymise Eulinderina semiradiata Barker & Grimsdale, 1936 with Eo. antillea here. Eo. antillea occurs in the Preston Hill Formation in the North Coast Belt and northern Wag Water Belt in Jamaica, where it is associated with Operculinoides and small Nummulites. It is the nominate taxon for ABZ8. At the type locality in St Bartholomew (recollected as samples ER1524-ER1526), Eo. antillea occurs with Fallotella cf. cookie.
Some specimens show a thick spiral wall which extends up to the chamber before the chamber with the first retrovert aperture (e.g., Fig. 66.2). Other specimens (e.g., Fig. 66.3-4) lack a thickened spiral wall, but occur with specimens which have a thickened spiral wall. Further work is needed to determine if these are separate morphospecies, or simply variants of a single morphospecies.
| Figure 62: Eolepidina antillea (Cushman, 1919).
1
(unfigured specimen from Cole, 1960), equatorial section.
2 (original of Cole, 1960, Pl. 13, fig. 5, figured as Lepidocyclina (Polylepidina) antillea Cushman), equatorial section.
3 (UWIGM.WL3532C-09), equatorial section. 4 (UWIGM.WL3532C-09), detail of equatorial section showing primary spire with R = 5.
5 (UWIGM.WL3532C-13), axial section (note presence of irregular lateral
chamberlets. A-B, Saint Bartholomew. C-E, Preston Hill Formation, Bamboo, parish of St Ann,
Jamaica. |
| Figure 63: Eolepidina antillea (Cushman, 1919).
1-2 (UWIGM.ER161-1), equatorial sections. 3-4 (UWIGM.ER161-2), equatorial sections. Sample ER161, stratotype of the Preston Hill Formation (which disappeared due to a landslide), Preston Hill, parish of St Mary,
Jamaica. |
| Figure 64:
Eolepidina antillea (Cushman, 1919), equatorial sections, megalospheric forms.
1 (EO.WL2047-2), R = 5. 2 (EO.WL2047-19), R = 5. 3 (EO.WL2047-18), R = 5.
4 (EO.WL2046-24), R = 5. 5 (EO.WL2046-17) R = 5. 6 (EO.WL2046-25), R = 5. Note differences in preservation between the two samples. Samples WL2046 and WL2047, Preston Hill Formation (ABZ8),
Lilyfield, St Ann, Jamaica. |
| Figure 65: Eolepidina antillea (Cushman, 1919), details of equatorial sections, megalospheric forms.
1 (EO.WL2046-8), R = 5. 2 (EO.WL2046-17), R = 5. 3 (EO.WL2046-23), R = 5.
4 (EO.WL2046-24), R = 6). 5 (EO.WL2046-25), R = 5. 6 (EO.WL2046-26), R = 5. Sample WL2046, Preston Hill Formation (ABZ8),
Lilyfield, St Ann, Jamaica. |
| Figure 66: Eolepidina antillea (Cushman, 1919), megalospheric forms.
1 (EO.WL2046-27), equatorial section, R = 5. 2 (EO.WL2046-28), equatorial section, R = 5.
3 (EO.WL2046-30), equatorial section, R = 5. 4 (EO.WL2047-31), axial section.
5 (EO.WL2047-29), axial section. Preston Hill Formation (ABZ8),
Lilyfield, St Ann, Jamaica. |
Eolepidina subplana (Barker & Grimsdale, 1936)
Previous descriptions. Barker & Grimsdale, 1936, p. 241, Pl. 31, figs. 1-2; Pl. 32, figs. 12-13; Pl. 33, fig. 8; Pl. 35, fig. 6; Pl. 37, fig. 2.
Recognition. Eo. subplana is recognized by its low, lenticular test with lateral chambers and a primary spire consisting of typically 4 chambers. We place populations with R = 4.5 - 3.5 in Eo. subplana.
Remarks. The ornament of Eo. subplana is variable. Some forms have an ornament consisting of regular pustules that extend across the entire surface, whereas other forms (Chapelton Formation at Lilyfield) have an ornament that consists of a central crown of pustules with radiating furrows in the marginal zone. These ornament types are also seen in Eo. antillea. Eo. subplana is the nominate species for ABZ9. The species has been recorded from Mexico (Veracruz), Jamaica, Florida, and St Bartholomew.
| Figure 67:
Eolepidina subplana (Barker & Grimsdale, 1936).
1 (EO.WL2044-36), equatorial section. 2 (EO.WL2044-37), equatorial section.
3 (EO.WL2044-36), equatorial section. 4 (EO.WL2044-37), equatorial section.
5 (EO.WL2044-67), equatorial section. 6 (EO.WL2044-50), equatorial section. Chapelton Formation (ABZ9),
Lilyfield, St Ann, Jamaica. |
| Figure 68: Eolepidina subplana (Barker & Grimsdale, 1936).
1 (EO.WL2044-36),
equatorial section, R = 4. 2 (EO.WL2044-37), equatorial section, R = 4.
3 (EO.WL2044-38), equatorial section, R = 4. 4 (EO.WL2044-40), equatorial section, R =
4. 5 (EO.WL2044-67), equatorial section, R = 4. 6 (EO.WL2044-72), axial section. 7 (EO.WL2044-74), axial section. Chapelton Formation (ABZ9),
Lilyfield, St Ann, Jamaica. |
| Figure 69:
Eolepidina subplana (Barker & Grimsdale, 1936). Equatorial sections, megalospheric forms.
1-2 (UWIGM.WL3444-4), 3-4 (UWIGM.WL3444-10): Chapelton Formation (ABZ9), Albert Town,
Trelawny, Jamaica. |
| Figure 70:
Eolepidina subplana (Barker & Grimsdale, 1936).
1 (ER1485-13), 2 (ER1485-04), 3 (ER1485-10), 4 (ER1485-08),
5 (ER1485-07): megalospheric specimens, equatorial sections. Locality ER1485 of Robinson (1996), ABZ9, St
Bartholomew. |
Eolepidina gardnerae (Cole, 1929)
Previous descriptions and figures. Cole, 1929, p. 60-62, Pl. 1, figs. 1-6; Pl. 2, figs. 1-2.
Recognition. Eo. gardnerae has one PAC and two subequal spires. This is clearly visible in material from the type series (Cole, 1929, Pl. 1, fig. 5; Pl. 2, fig. 2). We define R as ranging from 2.2-3.5. If a second PAC is present (e.g., Figs. 26.3, 26.6) it either produces no spires or very short ones.
Remarks. We place the species gardnerae in the genus Eolepidina as we regard the development of two PACs as being of generic value. Eo. gardnerae is distinctive and easily distinguished from P. chiapasensis, based on the number of PACs, when several specimens are available for study. Although there is undoubtedly a graduation from Eo. gardnerae to P. chiapasensis to Lepidocyclina ariana, the separation of Eo. gardnerae from P. chiapasensis is useful biostratigraphically. Eo. gardnerae is the nominate species for ABZ10. The species has been recorded from Florida/Alabama (as Polylepidina: Robinson & Jiang, 1995), Jamaica and Veracruz (as Polylepidina chiapasensis by Barker & Grimsdale, 1936, Pl. 35, figs. 7-8) and Chiapas (as Polylepidina antillea by Frost & Langenheim, 1974, Pls. 30-31), Mexico.
| Figure 71: Eolepidina gardnerae
(Cole, 1929). Megalospheric forms, equatorial
sections. 1 (UWIGM.Content 3327ft-6), 2 (UWIGM.Content
3327ft-1), 3 UWIGM.Content 3327ft-5), 4 (UWIGM.Content
3327ft-3), 5 (UWIGM.Content 3327ft-4), 6 UWIGM.Content
3327ft-2): Content Well (depth 3327ft = 1014 m), ABZ10, Content, Westmoreland,
Jamaica. |
| Figure 72: Eolepidina gardnerae (Cole, 1929). Megalospheric forms, equatorial sections (details).
1 (UWIGM.Content 3327ft-4), 2 (UWIGM.Content 3327ft-6), 3 UWIGM.Content 3327ft-5),
4
(UWIGM.Content 3327ft-1), 5 (UWIGM.Content 3327ft-2), 6 UWIGM.Content 3327ft-4): Content Well (depth 3327ft = 1014 m), ABZ10, Content, Westmoreland,
Jamaica. |
| Figure 73: Eolepidina gardnerae (Cole, 1929).
Megalospheric forms, equatorial sections. 1 (UWIGM.WL2921-1), specimen with R =
3. 2-3 (UWIGM.WL2921-2), specimen with
R = 2. 4-5 (UWIGM.WL2921-4), specimen with R = 4). 6 (UWIGM.WL2921-5), specimen with
R = 3. This population yields specimens that can be attributed on a typological approach to Eo. subplana, Eo. gardnerae and Po.
chiapasensis, but to Eo. gardnerae using a statistical approach. Chapelton Formation (ABZ10), north of
Elderslie, St James, Jamaica. |
| Figure 74: Eolepidina gardnerae (Cole, 1929). Megalospheric forms, equatorial sections.
1-2 (UWIGM.WL4449-03),
R = 3. 3-4 (UWIGM.WL4449-30). Specimens of typical Eo. gardnerae that we regard as reworked in ABZ14 sediments (with Plio. tobleri), chalk without
cherts, Cold Spring, Westmoreland, Jamaica. |
Genus Polylepidina Vaughan, 1924
Type species. Polylepidina chiapasensis Vaughan from the Middle Eocene of Chiapas, Mexico.
Diagnosis. A lepidocyclininid with two PACs and three or four asymmetrical primary spires of periequatorial chambers. Lateral chamberlets developed on both sides, with low numbers (usually 3 to 5, but up to 6 or 7) of tiers and with vacuole-like chamberlets.
Discussion. We follow Robinson (1997) in regarding the presence of two PACs and asymmetrical primary spires as the diagnostic criterion for discrimination at the generic level.
Polylepidina chiapasensis Vaughan, 1924
(Figs. 75 ![]() ,
76
,
76 ![]() ,
77
,
77 ![]() ,
78
,
78 ![]() ,
79
,
79 ![]() ,
80
,
80 ![]() )
)
Previous descriptions and figures. Vaughan, 1924, p. 807-808, Fig. 4; 1929a, p. 288-289, Fig. 3.
Recognition. Po. chiapasensis is flat discoidal and has numerous small to large pustules extending across most, or all, of the surface, yet sometimes it has a marginal band with radial striations formed from very-fine granules (as in Eo. semiradiata (Barker & Grimsdale, 1936) = Eo. antillea (Cushman, 1919)). Here, we define R as ranging from 2.0 - 2.2, and typically three or four asymmetrical peri-embryonic spirals develop from the two PACs.
Remarks. P. chiapasensis is externally similar to Lepidocyclina ariana and can only be distinguished in equatorial sections (P. chiapasensis is characterised by R = 2.0 - 2.2 with three or four asymmetrical peri-embryonic spirals, whereas L. ariana has R = 2 with two sets of two symmetrical peri-embryonic spirals). In most specimens, the three or four peri-embryonic spirals are markedly differently developed, with one spiral usually being dominant and having larger chambers. In a few specimens (e.g., Fig. 80.3-4) the spirals have chambers of equal sizes and on first glance the spirals appear symmetrical; yet a closer inspection reveals the spirals are asymmetric as the closing chambers (c) lie closer to one PAC (P) that the other.
Polylepidina chiapasensis is the nominal species for ABZ11, and its presence indicates a level in the lower Bartonian. The species is widespread in Jamaica in the Chapelton Formation at Albert Town and Litchfield, and in the Preston Hill Formation at Salt Spring.
| Figure 75: Specimens from the type material of Polylepidina chiapasensis Vaughan, 1923.
1,
off-centre equatorial section (USNM 269273), cut from same block as type specimens.
2,
off-centre equatorial section (USNM 324726), figured by Vaughan,
1929a, Fig. 3. Note that both have two PACs (although difficult to see in 1) and that the PACs are of similar size in 2; note two spirals with asymmetrical closing chamber (cc) in 2. Since these are
off-centre sections we do not see all the peri-embryonic spires that are developed; 2 clearly shows two spires developing from two large PACs and a closing chamber (cc) that is asymmetrical to the spire
development. |
| Figure 76: Polylepidina chiapasensis Vaughan, 1924 from Jamaica.
1-2 (UWIGM.WL3446-08), 3-4 (UWIGM.WL3446-12), 5 (UWIGM.WL3446-11),
6 (UWIGM.WL3446-03): megalospheric specimens, equatorial sections. Sample WL3446 (ABZ11), Chapelton Formation (ABZ11), Albert Town,
Trelawny, Jamaica. |
| Figure 77: Polylepidina chiapasensis Vaughan, 1924 from Jamaica.
1 (UWIGM.WL3446-40), 2 (UWIGM.WL3446-25), 3 (UWIGM.WL3446-12),
4 (UWIGM.WL3446-26), 5 (UWIGM.WL3446-11), 6 (UWIGM.WL3446-08): megalospheric specimens, equatorial sections. Note all specimens have two PACs (P) (although difficult to see in 5) and two closing chambers (c). Sample WL3446 (ABZ11), Chapelton Formation (ABZ11), Albert Town,
Trelawny, Jamaica. |
| Figure 78: Polylepidina chiapasensis Vaughan, 1924.
1-2 (UWIGM.WL4195-01), 3 (UWIGM.WL4195-2), 4 (UWIGM.WL4195-03),
5 (UWIGM.WL4195-4), 6 (UWIGM.WL4195-6), megalospheric specimens, equatorial sections. Sample WL4195, Chapelton Formation (ABZ11), Litchfield,
Trelawny, Jamaica. |
| Figure 79: Polylepidina chiapasensis Vaughan, 1924.
1 (UWIGM.WL2837-03), 2 (UWIGM.WL2837-08), 3 (UWIGM.WL2837-09),
4 (UWIGM.WL2837-12), 5 (UWIGM.WL2837-13), 6 (UWIGM.WL2837-14): megalospheric specimens, equatorial sections. Sample WL2837 (ABZ11), Preston Hill Formation, Salt Spring, St James,
Jamaica. |
| Figure 80: Polylepidina chiapasensis Vaughan, 1924.
1-2 (UWIGM.WL3597-01), 3-4 (UWIGM.WL3597-02), 5-6 (UWIGM.WL3597-05): megalospheric specimens, equatorial sections. Sample WL3597 (ABZ11), Preston Hill Formation, Salt Spring, St James,
Jamaica. |
Genus Lepidocyclina Gümbel, 1868
Type species. Nummulites mantelli Morton, 1833 from the Marianna limestone (lower Oligocene), near Claiborne, Alabama, USA (Morton, 1833; Vaughan, 1927).
Synonyms. Isolepidina Douvillé, 1915 (type species Nummulites mantelli Morton, 1833) [objective synonym]; Amphilepidina Douvillé, 1922 (type species Orbitoides sumatrensis Brady, 1875) [junior synonym]; Polyorbitoina Geyn & Vlerk, 1935 (type species Lepidocyclina (Polylepidina) proteiformis Vaughan, 1924) [junior synonym]; Multilepidina Hanzawa, 1932 (non Silvestri, 1937) (type species Lepidocyclina (Multilepidina) irregularis Hanzawa, 1932) [junior synonym].
Remarks. We restrict Lepidocyclina here to forms which have two PACs and an embryo with the protoconch and deuteroconch divided by a more-or-less straight to gently curved wall. The more 'primitive' forms lack adauxiliary chambers and include Lep. ariana Cole & Ponton. The more 'advanced' forms have adauxiliary chambers and include Lep. ocalana Cushman and Lep. macdonaldi Cushman. We do not think that separating these forms into separate subgenera (or genera) is useful as we are probably dealing with several distinct lineages that show a progression from forms lacking adauxiliary chambers to forms bearing adauxiliary chambers. The relationships with other 'simple' lepidocyclinids, such as Lep. canellei Lemoine & R. Douvillé from the Oligocene, are also unclear at the present time and we provisionally include all these forms in Lepidocyclina pending further work.
Lepidocyclina ariana Cole & Ponton, 1934
Previous description and figures. Cole & Ponton, 1934, p. 142-143, Pl. 2, figs. 1-5; Cole, 1944, p. 61-62, Pl. 1, fig. 14; Pl. 13, fig. 3; Pl. 14, figs. 1-7; Pl. 16, figs. 11-13; Pl. 17, fig. 13.
Remarks. In equatorial view L. ariana is characterised by a typical four-spiral peri-embryonic chamber plan developed from two PACs; it lacks adauxilary chamberlets and has semi-circular equatorial chamberlets. In axial section, it possesses relatively few lateral chamberlets that have thick walls with small irregular vacuoles.
Occurrence. In Jamaica, L. ariana occurs during a strongly regressive interval, linked either to a major sea-level fall, or to local tectonics, and has only been found in deposits within basins; equivalent deposits on the platforms are either lacking, or represented by platform interior facies without lepidocyclinids. In Florida and Alabama the species is common at some localities (e.g., Cole, 1944, 1945; Robinson & Jiang, 1995; Robinson, 1997). The species, with its distinctive lateral chamberlets is restricted to ABZ12.
| Figure 81: Lepidocyclina ariana Cole & Ponton, 1934, equatorial sections from the Yellow Limestone (ABZ12), Grants Pen, parish of St Thomas, Jamaica.
1 (UWIGM.WL3571-6); 2, 6 (UWIGM.WL3571-10); 3 (UWIGM.WL3571-11);
4 (UWIGM.WL3571-1); 5 (UWIGM.WL3571-2). Note the regular 4-spire plan in equatorial
section. |
| Figure 82: Lepidocyclina ariana Cole & Ponton, 1934, orientated axial sections from the Yellow Limestone (ABZ12), Grants Pen, parish of St Thomas, Jamaica. 1, 4 (UWIGM.WL3571-3),
2, 5 (UWIGM.WL3571-5), 3 (UWIGM.WL3571-13). Note the vacuolar appearance of the lateral
chamberlets. |
| Figure 83: Lepidocyclina ariana Cole & Ponton, 1934, details of equatorial sections from the Yellow Limestone (ABZ12), Grants Pen, parish of St Thomas, Jamaica.
1 (UWIGM.WL3571-1), megalospheric form; 2 (UWIGM.WL3571-6), megalospheric form;
3 (UWIGM.WL3571-2), megalospheric form; 4 (UWIGM.WL3571-14), microspheric
form. |
| Figure 84: Lepidocyclina ariana Cole & Ponton, 1934. 1-2 (UWIGM.WL4241-07),
3-4 (UWIGM.WL4241-11), 5 (UWIGM.WL4241-05), megalospheric specimens, equatorial sections. Sample WL4241 (ABZ12), chalks with scattered
cherts, Spot Valley, parish of St James, Jamaica. |
Lepidocyclina macdonaldi Cushman, 1919
Previous description and figures. Cushman, 1919, p. 94, Pl. 40, figs. 1-6; Cushman, 1920, p. 77, Pl. 34, figs. 1-3; Cole, 1952, p. 16, Pl. 7, figs. 1-19; Pl. 8, figs. 1-4; Pl. 14, fig. 11.
Remarks. Cole's (1952) material from Panama includes several species which show differences in the form of the lateral chambers. In 1956, Cole (p. 221) stated that three species of Lepidocyclina, namely Lep. ariana, Lep. proteiformis and Lep. macdonaldi could be separated on the basis of their lateral chamberlets: the first (Lep. ariana) had relatively thick but straight floors and roofs of the lateral chamberlets, and the chamberlet cavities were rather low; the second (Lep. proteiformis) had relatively thin and straight lateral floors and the cavities of the lateral chamberlets were open; the third (Lep. macdonaldi) had much thicker and curved floors and roofs of the lateral chamberlets and the cavities of the chamberlets were more appressed. Lep. ariana as figured here is relatively distinct, because of its basic four-spiral arrangement of peri-embryonic chambers and its thick walls to the lateral chamberlets that have thin spaced vacuole-like chambers. Lep. macdonaldi is also relatively distinct, in the form of its lateral chamberlets (thin) and thick curved roofs and floors; it is very distinctive in thin section. Cole (1956, Pl. 27, figs. 4-5) shows equatorial sections of Lep. macdonaldi from Jamaica, which appear to show the dark lines (cavities) above the equatorial layer, with the thick curved floors showing a bright area inside the cavities. Yet, the embryo is relatively similar to Lep. proteiformis and forms such as Lep. ocalana. The equatorial section of Lep. proteiformis illustrated by Cole (1956, Pl. 27, fig. 6) has an embryonic length of about 675 μm (as measured from his illustration), and therefore falls well outside the range of forms such as Lep. ariana and within the range of forms such as Lep. montgomeriensis and Lep. ocalana, although it has a simple four-spiral plan of peri-embryonic chambers. In fact, the lateral chamberlets of forms attributed to Lep. proteiformis from Jamaica and Mexico by Cole (1956) have lateral chambers similar to Lep. ocalana. We therefore tentatively attribute Lep. proteiformis of Cole, 1956 to the Lep. ocalana group.
Cole (1956, Pl. 27, figs. 3-5) illustrated three equatorial sections he referred to Lep. macdonaldi; these had embryo lengths of 520, 720 and 620 μm, respectively (as determined by measurement of the figures with the scale given). Four equatorial sections from sample WL4449 (ABZ14) from Cold Spring have an embryo length ranging from 488 μm to 766 μm (mean 660 μm, standard deviation 124.1 μm) and show the development of one or two adauxilary chambers. Both Cole's (1956) illustrations and our sectioned specimens have larger embryos than typical of Lep. ariana, and fall within the range of Lep. ocalana. It seems that Lep. macdonaldi and Lep. ocalana are parallel evolving lineages in the mid to late Eocene.
Our material shows the very distinctive features of the lateral chamberlets when seen in axial sections. The lateral chamberlets are strongly arcuate, offset in an en-echelon fashion, and do not show regular vertical tiers. Scattered piles (pillars) are also seen in the axial sections. This is consistent with material from the type locality in Mexico. Note that our specimens do not show an annular arrangement of equatorial chamberlets. This species resembles the arrangement seen in Lep. ariana in this respect rather than Lep. ocalana.
Range. Specimens of Lep. macdonaldi are common in ABZ14 (sample UWIGM.WL4449) at Cold Spring and Sherwood Content (samples UWIGM.R1068 and UWIGM.R1072) and sparingly so in the type Swanswick section at Swanswick House (ABZ16). In Panama, judging by a reinterpretation of the specimens illustrated by Cole (1952), the species ranges from zones ABZ14 to ABZ16.
| Figure 85: Lepidocyclina macdonaldi Cushman,
1920. 1 (WL4449-19), axial section, megalospheric form. 2 (R1072-18/19),
off-centred, axial section, microspheric form. 3 (WL4449-07), axial section, megalospheric specimen. 4 (R1068-08/09), axial section, megalospheric form.
5 (R1072-13/14), axial section, megalospheric form. 6 (WL4449-34) axial section, megalospheric form.
7 (WL4449-19), detail of axial section (same as 82.1), showing form of lateral chamberlets at two pillars (P).
8 (R1068-11), equatorial section, slightly off
centre, megalospheric form with selliform morphology. 1, 3, 6-7: sample WL4449, Cold Spring, Hanover (ABZ14).
2, 4-5, 8: samples R1068 and R1072, Sherwood Content, Trelawny
(ABZ14). |
| Figure 86: Lepidocyclina macdonaldi Cushman,
1920. 1-2 (WL4449-33), equatorial sections, megalospheric specimen. 3-4
(WL4449-32), equatorial sections, megalospheric specimen. 5-6 (WL4449-24), equatorial sections, weakly
selliform megalospheric specimen. Sample WL4449, Cold Spring, Hanover (ABZ14). |
Lepidocyclina ocalana Cushman, 1920
Previous description and figures. Cushman, 1920, p. 70-71, Pl. 27, figs. 1-4; Pl. 28, figs. 1-3; Cole, 1941, p. 41-43, Pl. 13, figs. 1-7; Pl. 16, figs. 1-4, 6-10, 15. Molina et al. (2015, Pl. 1.5) and Hohenegger and Torres Silva (2020, Fig. 2) illustrated specimens of Lep. ocalana erroneously under the name Lepidocyclina chaperi.
Recognition. Lep. ocalana in equatorial section combines a relatively large embryo, with a straight boundary between the protoconch and deuteroconch, with adauxilary chambers on the protoconch and deuteroconch, a relatively large number of peri-embryonic chambers, and a generally annular arrangement of equatorial chamberlets. At any level, there is a gradation from forms with four peri-embryonic spires to forms with adauxilary chambers; we place all these forms in Lep. ocalana and consider them natural variants of a single morphospecies. In axial section Lep. ocalana has a small number of layers of lateral chamberlets (up to about 7), which have open vacuoles, which are about as wide as the floors/roofs. Piles (pillars) are well developed.
Remarks. The species differs markedly from Lep. macdonaldi in axial section, with the later having arc-shaped, thin lateral chamberlets. Axial sections of the two species cannot be confused, but equatorial sections can. Equatorial sections sometimes show one larger and one smaller embryonic chamber (e.g., Fig. 87.4-5), but this may be due to the embryo being tilted at an angle to the equatorial plane (e.g., Fig. 87.1).
Range. In Florida Lep. ocalana occurs in the Ocala Limestone with Eulepidina chaperi and Heterostegina ocalana (Cole, 1941, 1943). In Panama, this species occurs with Pliolepidina tobleri, Eulepidina chaperi and Heterostegina ocalana (Cole, 1952). We have specimens in Jamaica from the middle unit of the Ipswich Formation (although these have a smaller embryo; Fig. 87), the Swanswick Formation sensu lato, the Claremont Formation and the Somerset Formation. The species therefore ranges through ABZ13 to ABZ16, and there is an increase in the size of the embryo passing from ABZ13 to ABZ14/ABZ15.
| Figure 87: Lepidocyclina ocalana Cushman, 1920, megalospheric specimens.
1 (UWIGM.WL2635-07), axial section, showing the embryo at an inclined angle to the equatorial plane.
2-3 (UWIGM.WL2635-9), equatorial section. 4-5 (UWIGM.WL2635-8), equatorial section; the difference in size between the first two chambers is probably due to the embryo being inclined as in
87.1. Ipswich Formation (ABZ13), Ipswich Railway Cutting, Ipswich, St Elizabeth,
Jamaica. |
| Figure 88: Lepidocyclina ocalana Cushman, 1920, megalospheric specimens.
1 (UWIGM.WL3876A-5), axial section. 2 (UWIGM.WL3876-09), axial section. 3 (UWIGM.WL3876-03), axial section. 4, 8 (UWIGM.WL3876-08), equatorial section.
5-6 (UWIGM.WL3876-01), equatorial section, note development of one adauxilary chamber on
protoconch. 7 (UWIGM.WL3876-02), equatorial section. Swanswick Formation s.l. (ABZ15), Highway 2000,
St Ann,
Jamaica. |
| Figure 89: Lepidocyclina ocalana Cushman, 1920, megalospheric specimens.
1-2 (UWIGM.WL4095-05), equatorial section, weakly selliform with equatorial chamberlets in annular rings.
3 (UWIGM.3931-5), axial section. 4-5 (UWIGM.3931-6), equatorial section, weakly
selliform variety. 6 (UWIGM.3931-7), equatorial section. 1-2, Claremont Formation (ABZ15);
3-6, Somerset Formation (ABZ16); Highway 2000, St Ann,
Jamaica. |
Genus Eulepidina Douvillé, 1911, p. 59
Type species. Orbitoides dilatata Michelotti, 1861 from the early Miocene of northern Italy (Michelotti, 1861).
Synonyms. Trybliolepidina ven der Vlerk, 1928 (type species Orbitoides (Lepidocyclina) ephippioides Jones & Chapman, 1900) [junior synonym].
Remarks. We follow Robinson (2004) in placing Lepidocyclina chaperi as the earliest species of Eulepidina.
Eulepidina chaperi (Lemoine & R. Douvillé, 1904)
Previous descriptions and figures. Cole, 1952, p. 23-27, Pl. 8, figs. 5-8; Pl. 9, figs. 3-19; Pl. 10, figs. 1-10; Pl. 11, figs. 1-8; Pl. 12, figs. 1-15; Pl. 20, figs. 8-10; Pl. 23, figs. 8, 11-12.
Recognition. The species is easily distinguished in equatorial section by the deuteroconch partially enveloping the protoconch, by the small and numerous peri-embryonic chambers and by the annular arrangement of the equatorial chamberlets. Specimens range from lenticular to weakly selliform (saddle-shaped). The species is distinguished from Oligocene species (such as, Eu. undosa and Eu. favosa) by its relatively similar diameters of the protoconch and deuteroconch, whereas in the Oligocene species the deuteroconch has a much greater diameter than the protoconch. The length of the embryo of Eu. chaperi is notably larger than that of co-occurring specimens of Lep. ocalana. Eu. chaperi occurs in upper ABZ15, ABZ16, and in the earliest Oligocene (ABZ17).
| Figure 90: Eulepidina chaperi (Lemoine & R. Douvillé, 1904), rather poorly preserved.
1 (UWIGM.WL4865-6); 2 (UWIGM.WL4865-1); 3 (UWIGM.WL4865-5);
4 (UWIGM.WL4865-3) : megalospheric specimens, equatorial sections. Sample WL4865, chalk (ABZ16),
Dressikie, parish of St Mary, Jamaica. |
| Figure 91: Eulepidina chaperi (Lemoine & R. Douvillé, 1904), rather poorly preserved specimens.
1 (UWIGM.WL4865-6); 2 (UWIGM.WL4865-5); 3 (UWIGM.WL4865-3);
4 (UWIGM.WL4865-1): details of embryo, with the walls of the protoconch partially broken away in 1 and 2
- note the thickened embryo wall. 5 (UWIGM.WL4865-4), axial section, showing lateral
chamberlets. 6 (UWIGM.WL4865-5), details of equatorial chamberlets showing annular arrangement. Sample WL4865 (ABZ16), chalk,
Dressikie, parish of St Mary, Jamaica. |
Subfamily ORBITOININAE Mitchell, E. Robinson & Özcan subfam. nov.
Diagnosis. The subfamily shows a parallel development to the Lepidocyclininae through eolepidine, unilepidine, polylepidine and lepidocycline forms to nephrolepidine forms. Early members of the subfamily have a relatively large number of well-developed lateral chamberlets in contrast to the early members of the Lepidocyclininae, which have none. The earliest member of the subfamily is Planorbitoinella gen. nov., which has well-developed lateral chamberlets. It is likely that the Orbitoinidae initially evolved in north-eastern South America during the Lutetian and subsequently spread across the American LBF bioprovince.
Genera included. Planorbitoinella gen. nov., Polyorbitoinella gen. nov., Orbitoina Schenck & Frizzell, 1936, Pliolepidina Douvillé, 1915, Nephrolepidina Douvillé, 1911.
Remarks. The evolutionary development of this subfamily is parallel to that of the Lepidocyclininae and gives rise to
homeomorphs. Details of the Orbitoininae are give in
Tables 20-21 and Figs. 92 ![]() ,
93
,
93 ![]() ,
94
,
94 ![]() ,
95
,
95 ![]() . Planorbitoinella gen. nov. gives rise to Polyorbitoinella gen. nov., which in turn gives rise to Orbitoina, and Orbitoina gives rise to Pliolepidina. Nephrolepidina (not shown) with its small embryo, probably evolved from Orbitoina, but this requires further work. Here we describe the forms that have so far been found in Jamaica and comment on selected forms from the American LBF bioprovince. Furthermore, as we get higher in the Eocene, the distinction between the Lepidocyclininae and the Orbitoininae becomes more difficult because of convergent morphologies, and assignment of specimens to a genus becomes more problematic.
Table 20 provides measurements of the length of the embryo, including the walls (Ele) for different species belonging to the
Orbitoininae. Table 21 provides measurements of the width of the proloculus/protoconch, excluding the walls (Pwi) for different species belonging to the Orbitoininae. Variations of the length of the embryo (Ele) and width of the proloculus/protoconch are shown in
Fig. 92
. Planorbitoinella gen. nov. gives rise to Polyorbitoinella gen. nov., which in turn gives rise to Orbitoina, and Orbitoina gives rise to Pliolepidina. Nephrolepidina (not shown) with its small embryo, probably evolved from Orbitoina, but this requires further work. Here we describe the forms that have so far been found in Jamaica and comment on selected forms from the American LBF bioprovince. Furthermore, as we get higher in the Eocene, the distinction between the Lepidocyclininae and the Orbitoininae becomes more difficult because of convergent morphologies, and assignment of specimens to a genus becomes more problematic.
Table 20 provides measurements of the length of the embryo, including the walls (Ele) for different species belonging to the
Orbitoininae. Table 21 provides measurements of the width of the proloculus/protoconch, excluding the walls (Pwi) for different species belonging to the Orbitoininae. Variations of the length of the embryo (Ele) and width of the proloculus/protoconch are shown in
Fig. 92 ![]() and a plot of Ele versus Pwi is shown in Fig. 93
and a plot of Ele versus Pwi is shown in Fig. 93 ![]() .
.
Table 20: Measurements of the length of the embryo (Ele) for species of Orbitoininae
| Species | Sample | Zone | Length of nucleus (μm) | Source | |||
| range | mean | SD | n | ||||
| Pl. nitida | Venezuela | ABZ7 | 150 - 195 | - | - | 2 | *Raadshooven, 1951 |
| Pl. ecuadorensis? | Venezuela | ABZ7 | 210 - 220 | - | - | 2 | *Raadshooven, 1951 |
| Pl. ecuadorensis | WL4869 Grants Town | ABZ7 | - | 206 | - | 1 | Our measurements |
| Pl. ecuadorensis | WL3309 Farm Turn | ABZ7 | - | 118 | - | 1 | Our measurements |
| Pl. ecuadorensis | WL3310 Farm Turn | ABZ7 | 89 - 166 | 135 | 29 | 7 | Our measurements |
| Pl. ecuadorensis | WL3324 Kilancholly | ABZ7 | - | 162 | - | 1 | Our measurements |
| Pl. ecuadorensis | Clay Pebble Bed, Ecuador | - | 120 - 150 | - | - | - | Hofker, 1956 |
| Po. lilyfieldensis | WL2047 Jamaica | ABZ8 | - | 153 | - | 1 | Our measurements |
| Po. lilyfieldensis | WL2046 Jamaica | ABZ8 | 215 - 310 | 149 | 23.5 | 12 | Our measurements |
| Po. lilyfieldensis | WL2044 Jamaica | ABZ9 | 155 - 275 | 214 | 32.6 | 9 | Our measurements |
| Po. lilyfieldensis | WL2041 Jamaica | ABZ9 | 210 - 215 | 213 | 3.5 | 2 | Our measurements |
| O. cedarkeysensis | Florida | ABZ13? | 160 - 200 | - | - | - | Cole, 1944 |
| O. wrighti | WL3876/3876A | ABZ15 | 239 - 330 | 2714 | 42.0 | 4 | Our measurements |
| O. rdouvillei | Types (Peru) | - | 265 - 295 | - | - | - | Todd & Barker, 1932 |
| O. rdouvillei | Borthworth's (Peru) | - | 222 - 240 | - | - | - | Todd & Barker, 1932 |
| O. rdouvillei | Cardo Grits (Peru) | - | 204 - 307 | - | - | - | Todd & Barker, 1932 |
| O. rdouvillei | Organos Grits (Peru) | - | 207 - 307 | - | - | - | Todd & Barker, 1932 |
| O. rdouvillei | Yapato Grits (Peru) | - | 248 - 315 | - | - | - | Todd & Barker, 1932 |
| O. rdouvillei | All specimens (Peru) | - | 204 - 340 | 276 | - | - | Todd & Barker, 1932 |
| O. rdouvillei | Peru | - | 145 - 260 | 195 | - | 9 | Colletti et al., 2019 |
| O. rdouvillei | WL3446 Jamaica | ABZ11 | 238 - 378 | 310 | 45.5 | 12 | Our measurements |
| O. rdouvillei | Florida | ABZ11 | 240 - 460 | 315 | 77.1 | 8 | Cole, 1944 |
| O. rdouvillei | WL4241 Jamaica | ABZ12 | 256 - 363 | 310 | 75.7 | 2 | Our measurements |
| O. sherwoodensis | Jamaica | - | 240 - 400 | - | - | - | Vaughan, 1928 |
| O. rdouvillei | WL4449 (reworked) | ABZ14 | 255 - 510 | 370 | 105.1 | 4 | Our measurements |
| O. trinitatis | Trinidad | - | 300 - 350 | - | - | - | Douvillé, 1924 |
| O. trinitatis | Trinidad | - | 400 - 650 | - | - | - | Eames et al., 1962 |
| O. trinitatis | WL3571 Jamaica | ABZ12 | 383 - 470 | 428 | 43.6 | 3 | Our measurements |
| O. kugleri | Venezuela | - | 350 - 400 | - | - | - | Vlerk, 1932 |
| O. trinitatis | WL2635 Jamaica | ABZ13 | 333 - 486 | 410 | 108.2 | 2 | Our measurements |
*Specimens measured from illustrations in publications (which is dependent on accuracy of plate reproduction when only magnifications were given). + Specimens considered reworked.
Table 21: Width of Proloculus/Protoconch (Pwi) for species of Orbitininae from Jamaica
| Species | Sample | Zone | Width of Proloculus/Protoconch (μm) | Source | |||
| range | mean | SD | n | ||||
| Pl. ecuadorensis | WL4869 Grants Town | ABZ7 | - | 107 | - | 1 | Our measurements |
| Pl. ecuadorensis | WL3309 Farm Turn | ABZ7 | - | 79 | - | 1 | Our measurements |
| Pl. ecuadorensis | WL3310 Farm Turn | ABZ7 | 51 - 118 | 95 | 24.7 | 7 | Our measurements |
| Pl. ecuadorensis? | WL3324 Kilancholly | ABZ7 | - | 118 | - | 1 | Our measurements |
| Pl. ecuadorensis? | WL2047 Jamaica | ABZ8 | - | 112 | - | 1 | Our measurements |
| Po. lilyfieldensis | WL2046 Jamaica | ABZ8 | 160 - 205 | 179 | 15.2 | 12 | Our measurements |
| Po. lilyfieldensis | WL2044 Jamaica | ABZ9 | 130 - 200 | 160 | 23.0 | 8 | Our measurements |
| Po. lilyfieldensis | WL2041 Jamaica | ABZ9 | 130 - 170 | 150 | 28.3 | 2 | Our measurements |
| Pa. wrighti | WL3876/3876A | ABZ15 | 110 - 214 | 150 | 56.2 | 3 | Our measurements |
| O. rdouvillei | WL3446 Jamaica | ABZ11 | 142 - 264 | 201 | 31.9 | 12 | Our measurements |
| O. rdouvillei | WL4241 Jamaica | ABZ12 | 128 - 187 | 158 | 41.7 | 2 | Our measurements |
| O. rdouvillei | WL4449 Jamaica | ABZ14 + | 158 - 230 | 197 | 34.6 | 4 | Our measurements |
| O. trinitatis | WL3571 Jamaica | ABZ12 | 277 - 292 | 282 | 8.4 | 3 | Our measurements |
| O. trinitatis | WL2635 Jamaica | ABZ13 | 206 - 343 | 275 | 96.9 | 2 | Our measurements |
| Figure 92: Length of embryo
(Ele) and width of proloculus/protoconch (Pwi) for the evolution of the Orbitoininae plotted against
zone. |
| Figure 93: Cross plot of length of embryo
(Ele in μm) versus width of proloculus/protoconch (Pwi in μm) for species of Orbitoininae from Jamaica. *includes indigenous and reworked
specimens. |
| Figure 94: Evolution of the Orbitoininae with small embryos. vR = van Raadshooven. |
| Figure 95: Evolution of the Orbitoina with large
embryos. |
Genus Planorbitoinella Mitchell, E. Robinson & Özcan gen. nov.
Type species. Lepidocyclina ecuadorensis Hofker, 1956 from the Clay Pebble Beds of the sea cliffs at Ancon, Ecuador (Hofker, 1956).
Diagnosis. An Orbitoininae with an eolepidine to unilepidine morphology, a small proloculus and many, well-developed tiers of lateral chambers.
Remarks. Planorbitoinella appears before Eolepidina or Polylepidina, and has lateral chamberlets, which are absent in early species of the Lepidocyclininae such as Eulinderina that occur at the same time. Planorbitoinella and Eolepidina are homeomorphs in the evolutionary development of different lineages (subfamilies) of the Lepidocyclinidae. These are the roots of Grimsdale's (1959) Lepidocyclina branches X and Y, respectively. We establish a parallel series of genera to those seen in the Lepidocyclininae and erect Planorbitoinella for populations with mean R = 3 and 4, and Polyorbitoinella for forms with mean R = 2 and asymmetrical spires of peri-embryonic chambers.
Planorbitoinella nitida (Caudri, 1996)
(Fig. 96 ![]() .6-10)
.6-10)
Previous descriptions. Lepidocyclina (Polylepidina) nitida: Caudri, 1996, p. 1215-1216, Pl. 2.12-13, Pl. 14.3-8, Pl. 30.3; Lepidocyclina spec. A: Raadshooven, 1951, p. 482, Figs. k-l, Pl. I, fig. 6; Pl. II, figs. 1, 3-4.
Recognition. Planorbitoinella nitida can be recognized by having a mean R of 3.5 or more, in conjunction with the presence of multiple tiers of lateral chamberlets.
Discussion. Planorbitoinella nitida (Caudri, 1996) is the earliest species of Planorbitoinella that we are aware of. The species shows an eolepidine morphology combined with well-developed lateral chamberlets. In Venezuela, Raadshooven (1951) reported Lepidocyclina spec. A (= Pl. nitida) as occurring in the Quebrada Grande Limestone (Baralt District, State of Zulia, western Venezuela) with Helic. gyralis and Operculinoides cf. oliveri, but at a lower level than Lep. spec. B (= Pl. ecuadorensis). The associated foraminifers would suggest an assignment to a level low in ABZ7. This assemblage has not been seen by us in Jamaica. In Trinidad, Pl. nitida occurs with Operculinoides spp. (Caudri, 1996) and can therefore be assigned to ABZ7.
The material illustrated by Caudri (1996; Fig. 96 ![]() .10-12) shows an embryo succeeded by one chamber and then the chamber with the first retrovert aperture; thus mean R = 4. Three specimens of Lep. spec. A were illustrated by Raadshooven (1951); two have R = 4 (e.g., Fig. 96
.10-12) shows an embryo succeeded by one chamber and then the chamber with the first retrovert aperture; thus mean R = 4. Three specimens of Lep. spec. A were illustrated by Raadshooven (1951); two have R = 4 (e.g., Fig. 96 ![]() .8) and one has R = 3 and the secondary spire from R has only one chamber
(Fig. 96
.8) and one has R = 3 and the secondary spire from R has only one chamber
(Fig. 96 ![]() .9). This would suggest a provisional assignment to Pl. nitida pending the study of more specimens.
.9). This would suggest a provisional assignment to Pl. nitida pending the study of more specimens.
| Figure 96: Drawings from published photomicrographs of the equatorial chamber arrangements of previously illustrated specimens of Planorbitoinella nitida and Pl. ecuadorensis from Venezuela, Ecuador, and Trinidad.
1-2, original interpretations of Lep. spec. B (= Pl. ecuadorensis) as given by Raadshooven, 1951,
Fig. 1 m-n. 3-4, our interpretation of the illustrations of the same specimens of Pl. ecuadorensis draw from figures in Raadshooven (1951), Pl. II, figs. 5-6 (same as 95.1-2, respectively).
5, specimen of Pl. ecuadorensis figured by Hofker, 1956, from Venezuela.
6-7, original interpretations of Lep. spec. A (= Pl. nitida) as given by Raadshooven, 1951,
Fig. 1 k-l. 8-9, our interpretation of the illustrations of the same specimens of Pl. nitida drawn from figures in Raadshooven (1951), Pl. II, figs. 3-4 (same as 95.8-9, respectively).
10-12, drawings from specimens of Pl. nitida figured by Caudri (1996): holotype - Pl. 2, fig. 12; other specimens: Pl. 14, fig. 7; Pl. 14, fig. 8, respectively. See text for locations of specimens and discussion. vR = Raadshooven; Hof = Hofker; C = Caudri. |
Planorbitoinella ecuadorensis (Hofker, 1956)
(Figs. 96 ![]() .1-5,
97
.1-5,
97 ![]() ,
98
,
98 ![]() ,
99
,
99 ![]() )
)
Synonymy. Lepidocyclina spec. B: Raadshooven, 1951, p. 481, Figs. m-n, Pl. 1, fig. 7; Pl. 2, figs. 2, 5-6; Lepidocyclina ecuadorensis Hofker, 1956, p. 938-939, Figs. 71-73.
Previous descriptions and figures. Hofker, 1956, p. 938-939, Figs. 71-73.
Type specimens. Hofker (1956, p. 940) stated there were four syntypes placed in the U.S. National Museum (Smithsonian). Online, the Smithsonian indicates that the holotype is USNM PR 4811, which is an uncut free specimen. Only one additional entry is present (USNM PR 4811A). The species must be interpreted based on Hofker's (1956) illustrations of an axial section (Hofker,
1956, Fig. 73b) and its arrangement of peri-embryonic chamberlets as seen in equatorial section (Hofker,
1956, Fig. 72); this shows a unilepidine arrangement (Fig. 96 ![]() .6).
.6).
Recognition. The species is recognized by its unilepidine arrangement (mean R = 2.2 to 3.5) of equatorial chambers in combination with a large number of lateral chamberlets with well-developed, open vacuoles. The equatorial chamberlets tend to be arranged in short radial rows. Unfortunately, Jamaican specimens are not easy to photograph, so we present a series of drawings showing the arrangements of equatorial chambers (Fig. 97 ![]() ) to complement the photomicrographs.
) to complement the photomicrographs.
Remarks. Hofker (1956) illustrated additional specimens in his
Fig. 73, but these illustrations are difficult to interpret, and our interpretation has been made based on Hofker's
(1956) Fig. 72. Raadshooven (1951) illustrated three specimens of Lep. spec. B; two of these show a unilepidine morphology (e.g.,
Fig. 96 ![]() .3) and one shows a polylepidine morphology
(Fig. 96
.3) and one shows a polylepidine morphology
(Fig. 96 ![]() .4); given the dominance of unilepidine forms (even with only three specimens), this material is assigned to Pl. ecuadorensis; this population may be a little younger than the Jamaican material assigned to Pl. ecuadorensis, since the latter only includes unilepidine forms
(Figs. 96
.4); given the dominance of unilepidine forms (even with only three specimens), this material is assigned to Pl. ecuadorensis; this population may be a little younger than the Jamaican material assigned to Pl. ecuadorensis, since the latter only includes unilepidine forms
(Figs. 96 ![]() ,
97
,
97 ![]() ,
98
,
98 ![]() ,
99
,
99 ![]() ).
).
Hofker (1956) and Cushman and Stainforth (1951) attributed the occurrence of Pl. ecuadorensis in the Clay Pebble Beds in Ecuador to the upper middle Eocene, but placed it before the appearance of the planktic foraminifer Hantkenina (which they took to mark the base of the upper Eocene). Hantkenina appears in planktic foraminifer zone E10 (P12) and suggests that the occurrence of Pl. ecuadorensis is older than this. This is consistent with the occurrence of Pl. ecuadorensis in Jamaica which is correlated with planktic foraminifer zone E8 (upper P9).
In Jamaica, Pl. ecuadorensis occurs with Operculinoides spp. in the Preston Hill Formation (ABZ7) at Grants Town (St Mary), Farm Turn (St Ann) and Kilancholly (St Ann). In Venezuela, Pl. ecuadorensis occurs in the early middle Eocene San Juan Limestone in the Baralt District, State of Zulia, western Venezuela, where it is also associated with Operculinoides spp. (cited as O. jennyi by Raadshooven, 1951); it can therefore also be assigned to ABZ7. We therefore suspect that Pl. ecuadorensis is a characteristic species of upper ABZ7 (whereas Pl. nitida is probably a lower ABZ7 species). The lack of Pl. nitida in Jamaica may be due to the hiatus between ABZ6 and ABZ7 (with NP15a not being recognized).
Cole (1960) regarded Lep. ecuadorensis as a synonym of Helicolepidina spirals Tobler; whereas Adams (1987, p. 293) stated that Lep. ecuadorensis was a problematic species which is neither a true Lepidocyclina, nor a typical Polylepidina, while the absence of an extended primary spire prevented assignment to Helicolepidina. As shown here, Pl. ecuadorensis is close to the origin of the Orbitoininae, and is unrelated to either Helicolepidina or Polylepidina.
| Figure 97: Planorbitoinella ecuadorensis (Hofker, 1956).
1 (EO.WL2098-23), equatorial section, megalospheric specimen. 2 (EO.WL2098-26), equatorial section, megalospheric specimen.
3 (EO.WL2098-48), equatorial section, microspheric specimen. 4 (UWIGM.WL3310-02), equatorial section, megalospheric specimen.
5-6 (UWIGM.WL3310-01), axial section (slightly off-center), megalospheric specimen. Preston Hill Formation, Farm Turn, St Mary,
Jamaica. |
| Figure 98: Planorbitoinella ecuadorensis (Hofker, 1956) from the Preton Hill Formation of Jamaica.
1 (EO.WL2098-68), equatorial section, microspheric form. 2 (UWIGM.WL3310-02), equatorial section, megalospheric form.
3 (EO.WL2098-25), equatorial section, megalospheric form. 4 (EO.WL2098-23), equatorial section, megalospheric form. 5
(UWIGM.WL3310A-08), equatorial section, megalospheric form with a very small embryo.
6 (UWIGM.WL3423-27), equatorial section, megalospheric form. WL2098, WL3010 and WL3010A from the Preston Hill Formation (ABZ7), Farm Turn, St Mary, Jamaica. WL3423 from the Preston Hill Formation (ABZ7),
Kilancholly, St Mary, Jamaica. |
| Figure 99: Line drawings of the equatorial chamber arrangement as seen in specimens of Planorbitoinella ecuadorensis (Hofker, 1956) from Jamaica. All specimens show a unilepidine morphology.
1
(UWIGM.WL4869-17), 2 (EO.WL2098-25), 3 (UWIGM.WL3310-02), 4 (EO.WL2098-24),
5 (UWIGM.WL3310A-09), 6 (EO.WL2098-23), 7 (EO.WL2098-26),
8 (UWIGM.WL3423-27), 9 (UWIGM.WL3310A-08), 10 (UWIGM.WL3309A-01). Preston Hill Formation (ABZ7): WL4869 from Grants Town; WL2098, WL3309A, WL3310 and WL3310A from Farm Turn; WL3423 from
Kilancholly, St Mary, Jamaica. Rx, recrystallized area. |
Genus Polyorbitoinella Mitchell, E. Robinson & Özcan gen. nov.
Type species. Polyorbitoinella lilyfieldensis sp. nov. from the Chapelton Formation (zones ABZ7 and ABZ8) at Lilyfield, St Mary, Jamaica.
Diagnosis. An Orbitoininae with a polylepidine morphology (mean R = 2.0 - 2.2 with asymmetrical spires of epiembryonic chamberlets), a small proloculus and many, well-developed tiers of lateral chambers.
Remarks. Polyorbitoinella shares the same relationship with Planorbitoinella as Polylepidina does to Eolepidina. We regard the presence of two PACS in combination with asymmetrical spires of peri-embryonic chambers as being of generic significance by analogy with the Lepidocyclininae.
Polyorbitoinella lilyfieldensis Mitchell, E. Robinson & Özcan sp. nov.
(Figs. 100 ![]() ,
101
,
101 ![]() ,
102
,
102 ![]() ,
103
,
103 ![]() ,
104
,
104 ![]() ,
105
,
105 ![]() )
)
Diagnosis. As for genus.
Type specimens. Holotype: OE.WL2046-11 (Figs. 100 ![]() .6,
101
.6,
101 ![]() .3,
105
.3,
105 ![]() .1); Paratypes: EO.WL2046.5, WL2046.6, WL2046.10, WL2046.15, WL2046.19, WL2046.21, WL2046.29, WL2046.31-34
(Figs. 100
.1); Paratypes: EO.WL2046.5, WL2046.6, WL2046.10, WL2046.15, WL2046.19, WL2046.21, WL2046.29, WL2046.31-34
(Figs. 100 ![]() ,
101
,
101 ![]() ,
102
,
102 ![]() ,
105
,
105 ![]() ); sample WL2046, Preston Hill Formation, Lilyfield area, St Ann (ABZ8).
); sample WL2046, Preston Hill Formation, Lilyfield area, St Ann (ABZ8).
Origin of name. From the district of Lilyfield in the parish of St Ann, Jamaica, where the species is found.
Description. Megalospheric specimens predominate, whereas microspheric specimens are relatively rare. Specimens are typically lenticular with a rounded
umbo and a gradual thinning towards the relatively acute periphery. The ornament consists of a series of fine pustules extending across the surface of the test. Megalospheric specimens have diameters of up to 2.8 mm and thicknesses of up to 0.6 mm. Orientated equatorial sections show the nepiont in detail
(Figs. 100 ![]() ,
101
,
101 ![]() ,
103
,
103 ![]() ,
104
,
104 ![]() ). The embryo consists of two chambers and has a mean length of 213
- 249 μm with the embryo of individual specimens ranging from 155 - 310 μm. The deuteroconch is slightly smaller than the protoconch; the protoconch is circular and the deuteroconch weakly arcuate (i.e., the wall is convex towards the deuteroconch). There are two PACs, one is often larger, or they may be of similar sizes. Two peri-embryonic spires develop from each PAC. At least one pair of asymmetrical spires is developed in most specimens, usually with one more chamber than the opposite spire
(Fig. 105
). The embryo consists of two chambers and has a mean length of 213
- 249 μm with the embryo of individual specimens ranging from 155 - 310 μm. The deuteroconch is slightly smaller than the protoconch; the protoconch is circular and the deuteroconch weakly arcuate (i.e., the wall is convex towards the deuteroconch). There are two PACs, one is often larger, or they may be of similar sizes. Two peri-embryonic spires develop from each PAC. At least one pair of asymmetrical spires is developed in most specimens, usually with one more chamber than the opposite spire
(Fig. 105 ![]() .2,
.5-9, .11, .16). Spires consist of from 1 to 4 chambers excluding the closing chambers, but usually consist of 1, 2 or 3 chambers. In a few specimens, the spires have symmetrical numbers of chambers (typically 2 to 3 chambers), but have the closing chambers arranged asymmetrically with respect to the protoconch-deuteroconch axis (e.g.,
Fig. 105
.2,
.5-9, .11, .16). Spires consist of from 1 to 4 chambers excluding the closing chambers, but usually consist of 1, 2 or 3 chambers. In a few specimens, the spires have symmetrical numbers of chambers (typically 2 to 3 chambers), but have the closing chambers arranged asymmetrically with respect to the protoconch-deuteroconch axis (e.g.,
Fig. 105 ![]() .1,
.3, .10, .14-15). In a few cases (Fig. 105
.1,
.3, .10, .14-15). In a few cases (Fig. 105 ![]() .4,
.13) the peri-embryonic spires are symmetrical around the protoconch-deuteroconch axis. The three samples (WL2041, WL.2044, WL2046) are interpreted to represent natural populations dominated by forms with two PACs and predominantly asymmetrically developed spires of peri-embryonic chambers or closing chambers. Orbitoidiform growth follows from the peri-embryonic spires and the equatorial chamberlets are arcuate. Axial sections are regularly elliptical
(Figs. 102
.4,
.13) the peri-embryonic spires are symmetrical around the protoconch-deuteroconch axis. The three samples (WL2041, WL.2044, WL2046) are interpreted to represent natural populations dominated by forms with two PACs and predominantly asymmetrically developed spires of peri-embryonic chambers or closing chambers. Orbitoidiform growth follows from the peri-embryonic spires and the equatorial chamberlets are arcuate. Axial sections are regularly elliptical
(Figs. 102 ![]() .6,
104
.6,
104 ![]() .6) and have 4 to 6 layers of lateral chamberlets. The lateral chamberlets range from relatively narrow to moderately wide and overlap chamberlets of the previous layer. Piles (pillars) can be seen developed in the axial sections. Microspheric forms are rare
(Figs. 101
.6) and have 4 to 6 layers of lateral chamberlets. The lateral chamberlets range from relatively narrow to moderately wide and overlap chamberlets of the previous layer. Piles (pillars) can be seen developed in the axial sections. Microspheric forms are rare
(Figs. 101 ![]() .5,
103
.5,
103 ![]() .3,
104
.3,
104 ![]() .5). The nepionic stage consists of 2-3 whorls of uniserial equatorial chambers, with a thickened wall which terminates in a chamber with a retrovert aperture. From this chamber a primary spiral of equatorial chambers encircles the thickened wall and terminates in a closing chamber of the short counter spiral. Orbitoidiform growth consisting of equatorial chamberlets in microspheric forms is similar to megalospheric specimens.
.5). The nepionic stage consists of 2-3 whorls of uniserial equatorial chambers, with a thickened wall which terminates in a chamber with a retrovert aperture. From this chamber a primary spiral of equatorial chambers encircles the thickened wall and terminates in a closing chamber of the short counter spiral. Orbitoidiform growth consisting of equatorial chamberlets in microspheric forms is similar to megalospheric specimens.
Distribution. Zones ABZ8 - ABZ9, Lilyfield, parish of St Ann, Jamaica, where it occurs with Eolepidina antillea (ABZ8) and Eo. subplana (ABZ9).
Remarks. Polyorbitoinella lilyfieldensis gen. nov., sp. nov. occurs in Jamaica through zones ABZ8-ABZ9 in the Preston Hill and Chapelton formations at Lilyfield, parish of St Ann; all levels are characterised by specimens that have predominantly asymmetrical spires or asymmetrical closing chambers (specimens with symmetrical spires and closing chambers are rare and considered part of the normal populations). The species shows a similar nepionic development to Polylepidina chiapasensis (ABZ11), but occurs with more primitive species of the Lepidocyclininae (Eo. antillea and Eo. subplana). Po. lilyfieldensis can be distinguished from Polylepidina by its smaller embryo and its greater number of layers of lateral chamberlets.
| Figure 100: Polyorbitoinella lilyfieldensis sp.
nov. 1 (EO.WL2046-6), 2 (EO.WL2046-5), 3 (EO.WL2046-15),
4 (EO.WL2046-29), 5 (EO.WL2046-21), 6 (EO.WL2046-11): equatorial sections. Preston Hill Formation (ABZ8),
Lilyfield, parish of St Ann, Jamaica. |
| Figure 101: Polyorbitoinella lilyfieldensis sp.
nov. Details of equatorial sections. 1 (EO.WL2046-5), 2 (EO.WL2046-4),
3 (EO.WL2046-11), 4 (EO.WL2046-21), 5 (EO.WL2046-20),
6 (EO.WL2046-31). 1-4, 6, megalospheric specimens, 5, microspheric specimen. Preston Hill Formation (ABZ8),
Lilyfield, parish of St Ann, Jamaica. |
| Figure 102: Polyorbitoinella lilyfieldensis sp.
nov. 1 (EO.WL2046-19), 2 (EO.WL2046-15), 3 (EO.WL2046-34), 4 (EO.WL2046-31),
5 (EO.WL2046-32), 6 (EO.WL2046-16). 1-5, equatorial sections;
6, axial section. Preston Hill Formation (ABZ8),
Lilyfield, parish of St Ann, Jamaica. |
| Figure 103: Polyorbitoinella lilyfieldensis sp.
nov. Equatorial sections. 1 (EO.WL2044-60), 2 (EO.WL2044-64), 3
(EO.WL2044-50), 4 (EO.WL2044-57), 5 (EO.WL2044-51), 6 (EO.WL2044-58).
1-2, 4-6, megalospheric specimens; 3, microspheric specimen. Preston Hill Formation (ABZ9),
Lilyfield, parish of St Ann, Jamaica. |
| Figure 104: Polyorbitoinella lilyfieldensis sp.
nov. 1 (EO.WL2044-64), 2 (EO.WL2044-63), 3 (EO.WL2044-60),
4 (EO.WL2044-68), 5 (EO.WL2044-50), 6 (EO.WL2044-73). 1-4, megalospheric specimens, equatorial section; 5, microspheric specimen, equatorial section; 6, axial section of megalospheric specimen. Preston Hill Formation (ABZ9),
Lilyfield, parish of St Ann, Jamaica. |
| Figure 105: Polyorbitoinella lilyfieldensis sp.
nov. Line drawings showing coiling of peri-embryonic equatorial chambers in equatorial section; note presence of two PACs and generally asymmetrical coiling.
1 (EO.WL2046-11), 2 (EO.WL2046-15), 3 (EO.WL2046-34), 4 (EO.WL2046-21), 5 (EO.WL2046-33), 6 (EO.WL2046-32),
7 (EO.WL2046-6), 8 (EO.WL2046-31): ABZ8. 9 (EO.WL2044-64),
10 (EO.WL2044-63), 11 (EO.WL2044-60), 12 (EO.WL2044-55),
13 (EO.WL2044-51), 14 (EO.WL2044-57), 15 (EO.WL2044-61),
16 (EO.WL2044-58): ABZ9. Preston Hill Formation,
Lilyfield, parish of St Ann, Jamaica. |
Genus Orbitoina Schenck & Frizzell, 1936
Type species. Isolepidina trinitatis Douvillé, 1924 from the middle or late Eocene of Trinidad (by original designation).
Synonymy. Orbitoina Geyn & Vlerk, 1935 [nomen nudum]; Isorbitoina Geyn & Vlerk, 1935 [nomen nudum]; Isorbitoina Schenck & Frizzell, 1936 (type species Isolepidina trinitatis Douvillé, 1924) [objective synonym]; Isorbitoina Thalmann, 1938 (type species Isolepidina trinitatis Douvillé, 1924) [junior objective synonym]; Neolepidina Brönnimann, 1947 (type species Isolepidina trinitatis Douvillé, 1924) [subjective junior synonym].
Remarks. Numerous species have been described that can be assigned to Orbitoina, but many of these need to be synonymised; this is beyond the scope of this paper. Here we describe some of the species that occur in the Middle and Upper Eocene that have biostratigraphic value. The main characters of importance in the recognition of species are: the size of the embryo, the form of the peri-embryonic chambers, the distribution of pillars and the form of the axial section.
Orbitoina cedarkeysensis (Cole, 1942)
(Fig. 94 ![]() )
)
Previous descriptions and figures. Cole, 1942, p. 43-45, Pl. 3, figs. 1-2; Pl. 8, fig. 11; Pl. 11, figs. 6-7; Pl. 12, figs. 1-7; Cole, 1944, p. 62, Pl. 7, figs. 16-17, 22.
Recognition. This species was described by Cole (1942, 1944) from wells in Florida, and is distinguished from species of Orbitoina from similar stratigraphic levels by its small embryo (160 to 200 μm).
Remarks. Cole (1942, 1944) described this species from three wells in Florida, the Sholtz No.1, the Cedar Key No. 2, and the St Mary's Oil Company, Hillard Turpentine Company No. 1. It seems to occur in a distinct stratigraphic interval in Florida, which is best interpreted in the St Mary's Oil Company well. Here it occurs above Lepidocyclina ariana, but below the more restricted foraminiferal facies with dictyoconids and Fabularia vaughani (i.e., Avon Park Formation). This would suggest a likely assignment to zones ABZ12 or ABZ13. The species has not been recorded in Jamaica.
Orbitoina rdouvillei (Lisson, 1921)
(Fig. 106 ![]() )
)
Synonyms. Lepidocyclina rdouvillei Lisson, 1921, p. 53-55, Pls. 4-5; Lepidocyclina peruviana Cushman (in Bosworth, 1922), p. 138, Pl. XXIV, fig. 1; Lepidocyclina antillea Cushman (in Bosworth, 1922) (non Lepidocyclina antillea Cushman, 1919), p. 137-138, Pl. XXIV, fig. 2; Lepidocyclina rdouvillei var. armata L. Rutten, 1928, p. 944-945, Figs. 24l-m, 29a-d, Pl. II, figs. 27-28; Lepidocyclina pervuviana Todd & Wright, 1932, p. 535-537, Pl. XLI, figs. 1-4; Pl. XLII, figs. 2-3, 5-7; Lepidocyclina sherwoodensis Vaughan, 1928, p. 287-288, Pl. 48, figs. 4-8.
Previous descriptions and figures. Lisson, 1921, p. 53-55, Pls. 4-5; Todd & Wright, 1932, p. 535-537, Pl. XLI, figs. 1-4; Pl. XLII, figs. 2-3, 5-7; Coletti et al., 2019, Fig. 10.
Recognition. An Orbitoina with an embryo with a length of 204 to 340 μm, with a relatively thick wall, two PACs and four symmetrical spirals of peri-embryonic chambers. Equatorial sections often show a radial arrangement of chamberlets. Axial sections show well-developed lateral chamberlets with up to 10 layers presents; the pillars are well developed.
Remarks. Lepidocyclina peruviana was described by Cushman (in Bosworth, 1922), but only external details were given. Todd and Barker (1932) provided the details of the interior based on some of the type series together with additional material. They stated (Todd & Barker, 1932, p. 536) that the embryo had a mean length of 276 μm and a range from 204 μm to 340 μm. Their table (Todd & Barker, 1932, p. 539), however, lists details of the length of the embryo in five populations that they place in Lepidocyclina peruviana: the type material (265 - 295 μm), Bosworth's material of 'antillea' (222 - 240 μm), material from the Cardo Grits (204 - 307 μm), specimens from the Organos Grits (207 - 307 μm) and material from the Yapato Grits (248 - 315 μm). We suspect a typographic error in the table and that 240 μm for Bosworth's 'antillea' should read 340 μm (although we cannot confirm this). Todd and Barker (1932) argued that L. antillea Cushman (in Bosworth, 1922) and Lepidocyclina rdouvillei var. armata L. Rutten, 1928 had similar embryo lengths and were synonyms of L. peruviana Cushman (in Bosworth, 1922). Lepidocyclina armata L. Rutten, 1928 was re-described by Eames et al. (1968) based on the type series. They demonstrated that the embryo ranged from 270 to 320 μm, and therefore matched the dimensions seen in L. peruviana. Coletti et al. (2019) clearly recognized that L. rdouvillei Lisson (Lepidocyclina rdouvillei Lisson) is a senior synonym of L. peruviana, recognizing that Todd and Barker (1932) relied on measurements given by Gorter and Vlerk (1931) rather than the description and figures of L. rdouvillei as given by Lisson (1921). We therefore place L. peruviana Cushman in Bosworth, 1922, L. antillea Cushman in Bosworth, 1922 and L. rdouvillei var. armata L. Rutten, 1928 in synonymy with L. rdouvillei Lisson, and place L. rdouvillei in the genus Orbitoina. We also consider Lepidocyclina sherwoodensis a synonym of O. rdouvillei. Vaughan (1928) recorded the length of the nucleus in three specimens as being 0.4 mm, 0.25 and 0.3 mm.
Coletti et al. (2019) recorded O. rdouvillei in southern Peru as occurring in zone CNE14 which is equivalent to parts of NP16 and CN14a. This occurrence of O. rdouvillei, with Polylepidina with two PACs (Coletti et al., 2019), would suggest an assignment to ABZ11. Or. rdouvillei occurs in Jamaica in zone ABZ11.
| Figure 106: Orbitoina rdouvillei (Lisson, 1921).
1-2, equatorial section
(UWIGM.WL3446-21); 3, axial section (UWIGM.WL3446-23); 4, equatorial section (UWIGM.WL3446-2); 5, equatorial section (UWIGM.WL3446-18); 6, equatorial section (UWIGM.WL3446-19). Scale bar for 2, 4-6 is the 0.5 mm scale above 5. Chapelton Formation, Albert Town, Jamaica,
ABZ11. |
Orbitoina trinitatis (Douvillé, 1924)
(Fig. 107 ![]() )
)
Previous descriptions and illustrations. Vaughan & Cole, 1941 (described as Isolepidina pustulosa Douvillé), p. 65-66, Pls. 25-32; Caudri, 1996 (described as Lepidocyclina pustulosa Douvillé and L. pustulosa trinitatis Douvillé), p. 1218-1219, Pl. 15.3, 15.9.
Recognition. This species is recognized on the basis of its highly inflated axial section with well-developed pillars, and its equatorial section with broad, narrow, often double-arched PACs and equatorial chambers. Previous descriptions of the length of the nucleoconch are given as 300 to 480 μm by Douvillé (1917, 1924; Todd & Barker, 1932) and 400 to 650 μm (Eames et al., 1962).
Remarks. Douvillé (1924) clearly distinguished L. pustulosa from L. trinitatis on the basis of the lateral chamberlets seen on the surface. L. trinitatis (both megalospheric and microspheric generations) is characterised by numerous polygonal pillars surrounded only by a small number of lateral chambers; in contrast L. pustulosa (stating only microspheric specimens being available) is characterised a large number of large pustules (300 µm) at the umbo, each surrounded by about fifteen small rounded chambers 70 to 120 µm in diameter. Douvillé (1924) gives measurements of the length of the embryo as ranging from 300 μm to 350 μm in L. trinitatis, and from 90 μm to 100 μm in L. pustulosa (despite stating that he only had microspheric forms). Vaughan and Cole (1941) made no mention of the lateral chamberlets in their discussion of the two species.
Eames et al. (1962) recognized that Vaughan and Cole (1941) had only figured specimens which would be attributable to L. trinitatis in their work under the name L. pustulosa, and this has been the source of much subsequent confusion. Eames et al. (1962, p. 302) gave the length of the nucleoconch for L. pustulosa as 0.25 - 0.35 mm with thick-walls and that for L. trinitatis as 0.40 - 0.65 mm (rarely over 0.55 mm) with thin walls. While we agree with these two groups, their L. pustulosa is equivalent to L. peruviana of Caudri (1975, 1996) which we have placed in Or. rdouvillei, and not with Douvillé's (1917, 1924) L. pustulosa which has a much smaller nucleoconch. L. pustulosa will remain enigmatic until the type material is studied (if it is still available), but should probably be attributed to Orbitoina.
In general form Or. trinitatis is similar to Or. wrighti sp. nov. However, Or. trinitatis has a larger embryo and the equatorial chamberlets are narrow and more strongly arched than those in Or. wrighti sp. nov.
| Figure 107: Orbitoina trinitatis Douvillé, 1924.
1-3, equatorial section (WL3571-8), note the narrow PACs and the elongate two-humped chamberlets that for the spirals.
4-6, axial section (WL3571-7), note the pillars and regular tiers of
chamberlets. Limestones above the Grants Pen Clay (Yellow Limestone), Grants Pen, St Thomas, Jamaica
(ABZ12). |
Orbitoina wrighti Mitchell, E. Robinson & Özcan sp. nov.
1993 Lepidocyclina pustulosa Douvillé; Robinson & Wright (in part), p. 317, Figs. 23.1, 23.3 only.
Type Material. Holotype (UWIGM.WL3876.5). Paratypes (UWIGM.WL3876-06, WL3876-07, WL3876A-3, WL3876A-4, WL3876A-10, WL3876A-11). All from the Swanswick Formation sensu lato, ABZ15, North-South Leg of Highway 2000, north of the Golden Vale Toll Plaza, parish of St Ann, Jamaica.
Diagnosis. A small species of Orbitoina with a thick diamond-shaped test in axial section; a relatively small embryo in equatorial section, with four simple spirals emanating from the two PACs and closing in symmetrical closing chambers. A few thick piles (pillars) producing pustules at the crown; and numerous rows of open lateral chamberlets.
Origin of name. From the late Raymond Wright, for his work on the foraminifers and stratigraphy of the White Limestone of Jamaica.
Description. A small species of Orbitoina with a strongly inflated test. Megalospheric specimens reach a diameter of 1.7 mm and a thickness of 1.2 mm. In equatorial sections it displays a typical Orbitoina arrangement of chambers. The nucleoconch is relatively small and comprises a protoconch that is slightly larger than the deuteroconch, with the protoconch and deuteroconch separated by an almost straight, or gently curved wall. The deuteroconch is marginally wider that the protoconch. The size of the embryo is relatively small (length 254 μm in the holotype). Two PACs are present which are slightly larger than the other peri-embryonic chambers and are regularly globular. There are four primary spires, two developed from each PAC, with the spires around the protoconch consisting each of three peri-embryonic chambers and a closing chamber, and those around the deuteroconch consisting of two peri-embryonic chambers and a closing chamber. There are no adauxilary chambers. The equatorial chamberlets are diamond shaped. In axial section, the test is very thick with the thickness nearly equaling or exceeding the diameter. One, two or rarely three pillars are seen in axial sections and extend from near the embryo towards the crown of the test where they form pustules with a diameter of up to 290 μm. The equatorial layer is relatively thin and shows only minor thickening towards the margin of the test. There are up to 13 rows of lateral chambers developed over the embryo, with the chambers arranged in tiers; the thickness of the chamber floors and chambers are about the same and the floors are straight and not curved.
Remarks. Specimens belonging to this species have typically been placed in 'Lepidocyclina' pustulosa or 'L.' trinitatis in previous studies (e.g., Robinson & Wright, 1993, Fig. 23.1-3). In fact the species is clearly well-defined amongst Orbitoina in the late Eocene. Or. cedarkeyensis is less inflated that Or. wrighti sp. nov. This form is easily recognizable in random cuts in thin section due to its distinctive axial section and the presence of relatively few pillars projecting up towards the crowns.
Range. In Jamaica, the species occurs commonly in the Swanswick Formation sensu lato on N-S Highway 2000 (ABZ15), in the Swanswick Formation at Dallas Mountain (ABZ15), and in the type Swanswick Formation at Swanswick House (ABZ16). It also occurs in the middle part of the Ipswich Formation in St Elizabeth, Jamaica (ABZ14). The species serves as an index for ABZ15, although it ranges from ABZ14 to ABZ16.
| Figure 108: Orbitoina wrighti sp.
nov. 1-2, equatorial section, megalospheric specimen (holotype: UWIGM.WL3876-05).
3, slightly
off-centre axial section, megalospheric specimen (Paratype: UWIGM.WL3876-06). 4, axial section, megalospheric specimen
(Paratype: UWIGM.WL3876-07). 5, axial section, megalospheric specimen
(Paratype UWIGM.WL3876A-03). 6, axial section, microspheric specimen
(Paratype: UWIGM.WL3876A-11). Swanswick Formation sensu lato (ABZ15), North-South Leg of Highway 2000, just north of the Goldon Vale Toll Plaza, parish of St Ann,
Jamaica. |
| Figure 109: Orbitoina wrighti sp.
nov. 1-2, equatorial section, megalospheric specimen (holotype: UWIGM.WL3876A-04).
3-4, equatorial section, megalospheric specimen
(Paratype: UWIGM.WL3876A-10). Swanswick Formation sensu lato (ABZ15), North-South Leg of Highway 2000, just north of the Goldon Vale Toll Plaza, parish of St Ann,
Jamaica. |
Genus Pliolepidina Douvillé, 1915
Type species. Lepidocyclina (Pliolepidina) tobleri Douvillé, 1917 (by subsequent monotypy: Douvillé, 1917) from the middle or late Eocene of Trinidad (Caudri, 1975, 1996).
Synonyms. Multicyclina Cushman, 1918 (type species: Lepidocyclina (Multicyclina) duplicata Cushman, 1918 = Lepidocyclina (Pliolepidina) tobleri Douvillé, 1917) [junior synonym]; Multilepidina Silvestri, 1937 (non Hanzawa, 1932) (type species: Pliolepidina tobleri Douvillé, 1917) [objective junior synonym].
Pliolepidina tobleri Douvillé, 1917
Previous descriptions and figures. Douvillé, 1917, p. 844, Fgs. 5-6 ; Vaughan & Cole, 1941, p. 66-67, Pl. 24; Caudri, 1975, p. 575, Pl. 27.4-7; Caudri, 1996, p. 1219, Pl. 15.7, 15.12.
Synonyms. Lepidocyclina panamaensis Cushman, 1918; Lepidocyclina (Multicyclina) duplicata Cushman, 1918.
Remarks. This is a very distinctive species with a short stratigraphic range and has been reported widely in the literature: Trinidad (Douvillé, 1915, 1917, 1924; Vaughan & Cole, 1941; Caudri, 1975, 1996), Panama (Cushman, 1918; Cole, 1952), Venezuela (Eames et al., 1968) and Jamaica (Robinson & Mitchell, 1999; Robinson et al., 2018). Due to extensive reworking in sections in Trinidad, it is difficult to determine the succession of benthic larger foraminiferal assemblages, and it appears that O. rdouvillei (= Lep. peruviana sensu Caudri), O. trinitatis (= Lep. pustulosa sensu Caudri) and Pl. tobleri, which characterise different levels in the successions in Jamaica (herein) and Panama (Cole, 1952) occur as a mixed assemblage (indicating reworking) in Trinidad (Vaughan & Cole, 1941; Caudri, 1975, 1996). Pl. tobleri is the nominate species for ABZ14.
| Figure 110: Pliolepidina tobleri Douvillé, 1917.
1, random sections (equatorial and axial) in limestone (NHM.PM.P.47294), Peńas Blancas Limestone, Rio
Chacual, Venezuela. 2, equatorial section (NHM.PM.P.52831), Nicaragua. 3, axial section (sample UWIGM.WL4449-44), limestone above marlstones, Cold Spring, Westmoreland, Jamaica.
4, axial section, and 5, equatorial section, upper Ipswich Formation (ER2756), former Western Cement Company, Appleton, St Elizabeth, Jamaica. ABZ14 (reproduced from Robinson & Mitchell,
1999). |
| Figure 111: Pliolepidina tobleri Douvillé, 1917, equatorial sections from sample WL4449 (ABZ14), the deep-water limestones and the marlstones at Cold Spring, Westmoreland, Jamaica.
1-2 (UWIGM.WL4449-45), 3-4 (UWIGM.WL4449-43), 5-6 (UWIGM.WL4449-38). |
Superfamily ORBITOIDOIDEA Schwager, 1876
Family LINDERINIDAE Loeblich & Tappan, 1984
Remarks. See Ferrŕndez-Cańadell & Serra-Kiel (1999) and Özcan et al. (2021b, 2022b) for a reclassification of the Linderinidae and a description of the embryo of Linderina.
Genus Linderina Schlumberger, 1893
Type species. Linderina brugesi Schlumberger, 1893 (by monotypy), from the Eocene of Bruges, Gironde, southwestern France.
Linderina floridensis Cole, 1942
Previous descriptions and illustrations. Cole, 1942, p. 36-37, Pl. 11, fig. 8; Pl. 15, figs. 7-11.
Remarks. Cole (1942) nominated two specimens as cotypes, his Pl. 15, figs. 9 (axial section) and 10 (equatorial section) which therefore form the type series. We do not nominate a lectotype at present because a restudy of the type material is needed.
The embryo of L. floridensis (Figs. 112 ![]() ,
113
,
113 ![]() .1) is comparable to the embryo of the type species of the genus, Linderina brugesi Schlumberger, as figured by Ferrŕndez-Cańadell and
Serra-Kiel (1999) justifying assignment to Linderina. Hose and
Versey (1956) defined a Linderina band in the limestone successions of Jamaica; however our studies indicate that Linderina floridensis is a rather long-ranging species occurring at at least three different stratigraphic levels (mid
Ypresian, mid Lutetian, early Bartonian) and is not very useful in high-resolution biostratigraphy (it also occurs in the
mid-Ypresian in Turkey: Özcan et al., 2021b).
.1) is comparable to the embryo of the type species of the genus, Linderina brugesi Schlumberger, as figured by Ferrŕndez-Cańadell and
Serra-Kiel (1999) justifying assignment to Linderina. Hose and
Versey (1956) defined a Linderina band in the limestone successions of Jamaica; however our studies indicate that Linderina floridensis is a rather long-ranging species occurring at at least three different stratigraphic levels (mid
Ypresian, mid Lutetian, early Bartonian) and is not very useful in high-resolution biostratigraphy (it also occurs in the
mid-Ypresian in Turkey: Özcan et al., 2021b).
| Figure 112: Linderina floridensis Cole, 1942.
1 (EO.WL2098-31), axial section. 2-3 (EO.WL2098-30), equatorial section.
4-5 (EO.WL2098-27), equatorial section. 6-7 (EO.WL2098-126), equatorial section. Sample WL2098, Preston Hill Formation (ABZ7), Farm Turn, St Mary, Jamaica. Notice that three chamberlets are formed consistently at the second growth stage following the embryonic apparatus
(chamberlets with blue walls). |
| Figure 113: Linderina and Caudriella.
1-5, Linderina floridensis Cole, 1942: 1-2, equatorial section (WL3309-11);
3-4, equatorial section (WL3423-03); 5, axial section (WL3309A-04).
1-2, 5: Preston Hill Marl, Farm Turn, St Mary, ABZ7. 3-4: Preston Hill Marl,
Kilancolly, St Mary, ABZ7. Note the embryonic apparatus in 1 and 3 (with drawings), which is typical for the genus Linderina (Ferrŕndez-Cańadell & Serra-Kiel, 1999).
6, Caudriella sp., axial section (WL3275A-06), Swanswick House,
Trelawny, Jamaica, ABZ7. |
Genus Caudriella Haman & Huddleston, 1984
Synonyms. Margaritella Caudri, 1974 (type species Margaritella ospinae Caudri, 1974) [preoccupied, non Margaritella Meek & Hayden, 1860 (a mollusc)].
Type species. Margaritella ospinae Caudri, 1974, from the early mid Bartonian of Margarita Island, Venezuela.
Caudriella sp.
(Fig. 113 ![]() .6)
.6)
Previous descriptions and illustrations. Özcan et al., 2022b, p. 26-27, Fig. 9J.
Remarks. Ferrŕndez-Cańadell & Serra-Kiel (1999) placed Caudriella in the Linderinidae based on the similarity of the embryos of C. ospinae and L. brugesi. Caudriella has lateral chamberlets whereas Linderina lacks lateral chamberlets. We have found Caudriella sp. in only one sample in Jamaica. Interestingly, samples from the type section of the Swanswick Formation at Swanswick House contain both Linderina and Caudriella (with a few lateral chamberlets:
Fig. 113 ![]() .6) demonstrating that there is unlikely to be a simple evolution from Linderina to Caudriella.
.6) demonstrating that there is unlikely to be a simple evolution from Linderina to Caudriella.
Superfamily NUMMULITOIDEA Blainville, 1827
Family NUMMULITIDAE Blainville, 1827
Genus Nephronummulites Mitchell, E. Robinson & Özcan gen. nov.
Type species. Nummulites macgillavryi M. G. Rutten, 1935, from the ?Ypresian of northern Santa Clara Province, Cuba.
Diagnosis. A genus of Nummulitidae with strongly dimorphic generations, and with the megalospheric forms having a large rounded proloculus and a thin crescent-shaped deuteroloculus.
Remarks. The genus differs from other American and Tethys Nummulites by the shape of the deuteroloculus. In Nummulites the deuteroloculus is only slightly smaller than the proloculus whereas in Nephronummulites the deuteroloculus is a narrow crescent-shaped chamber (Fig. 115).
Nephronummulites macgillavryi (M.G. Rutten, 1935)
(Fig. 114 ![]() )
)
Previous descriptions. Rutten, 1935, p. 530, Pl. 59, figs. 6-10; Robinson & Wright, 1993, p. 331, Fig. 29.1-3.
Remarks. Nephronummulites macgillavryi is a very distinctive species characterised by its large megalospheric form with a large proloculus and its very large microspheric form (Rutten, 1935); it cannot be confused with any other described American nummulite. Orientated equatorial and axial sections of free megalospheric specimens are illustrated here for the first time and show that the deuteroloculus is small, crescent-shaped and embraces the protoconch. The species has a short stratigraphic range in the latest part of the Ypresian (ABZ5), is easily recognizable (even before sectioning) and is a valuable biostratigraphic marker.
| Figure 114: Nephronummulites macgillavryi (M. G. Rutten, 1935).
1-4, megalospheric equatorial sections, 1, 4, WL3315-05; 2-3, WL3315-04.
5-6, megalospheric axial section (WL3315-03). Palmetto Grove Formation (ABZ5), SW of Hampstead, St Ann,
Jamaica. |
Genus Nummulites Lamarck, 1801
Type species. Camerina laevigata Bruguičre, 1792 (ICZN, 1945, Op. 192), from the middle Eocene (Lutetian) of Cayenne, France.
Nummulites striatoreticulatus L. Rutten, 1928
(Fig. 115 ![]() )
)
Previous descriptions. Rutten, 1928, p. 1068; Robinson & Wright, 1993, p. 331, Figs. 29.5, 30.6.
Remarks. Small species of Nummulites are abundant from the mid Lutetian to the late Priabonian. They have been split into numerous taxa (e.g., Barker, 1939) many of which have subsequently been synonymised (e.g., Cole, 1958b). Some studies (e.g., Wright & Swizer, 1971; Torres-Silva et al., 2018) have attempted statistical studies on American Nummulites, but mostly using isolated populations. What is needed is a statistical treatment of populations collected sequentially from the mid Lutetian to the late Priabonian, but this is beyond the scope of this paper. The results of Torres-Silva et al. (2018) offer hope that some stratigraphic resolution can be achieved. We place all these small Nummulites from the Lutetian, Bartonian and Priabonian in the group of N. striatoreticulatus here.
| Figure 115: Nummulites striatoreticularis Rutten,
1928, from the mid Lutetian to Bartonian of Jamaica. 1 (UWIGM.WL3444-17),
2 (UWIGM.WL3275-22), 6 (UWIGM.WL4449-26): equatorial sections;
3 (UWIGM.WL3444-18), 4 (UWIGM.WL3275-21), 5 (UWIGM.WL3275-2),
7 (UWIGM.WL4449-25): axial sections. Sample WL3275: ABZ7, Swanswick type section,
Trelawny, Jamaica; WL3444: ABZ9, Chapelton Formation, Albert Town, Trelawny, Jamaica; WL4449: ABZ14, limestones above marlstones, Cold Spring, Westmoreland,
Jamaica. |
Genus Operculinoides Hanzawa, 1935
Type species. Nummulites (Nummulina) willcoxi Heilprin, 1882, from the Priabonian of Florida.
Remarks. Although various studies have looked at Operculinoides (e.g., Wright & Switzer, 1971; Torres-Silva et al., 2018), there is too little biostratigraphic resolution at the present time (Torres-Silva et al., 2018). The appearance of Operculinoides (with Nummulites striatoreticularis) at the base of ABZ7 is an important biostratigraphic datum in the mid Lutetian of the American region.
Operculinoides spp.
(Fig. 116 ![]() )
)
Remarks. Details of various species of Operculinoides are present in Barker (1939) and Cole (1958b), with recent statistical treatments given in Torres-Silva et al. (2018). Individual species names are not assigned here. Small Operculinoides spp. appear in ABZ7 and range up to ABZ16.
| Figure 116: Operculinoides
spp. 1-2 (UWIGM.WL3353-07), equatorial sections, ABZ9, Preston Hill Formation, Jeffrey Town, St Mary, Jamaica.
3 (UWIGM.WL3309A-03), axial section, ABZ7, Preston Hill Formation, Farm Turn, St Mary, Jamaica.
4 (UWIGM.WL3277-09), 5 (UWIGM.WL3277-11), equatorial sections, ABZ9, Preston Hill Formation, Clarkes Town,
Trelawny, Jamaica. |
Genus Heterostegina Orbigny, 1826
Type species. Heterostegina depressa Orbigny, 1826 (by subsequent designated by Parker et al., 1865, p. 36) from the Recent.
Remarks. The genus Heterostegina appears within the Eocene with two groups which are probably unrelated. H. cubana Cizancourt, 1948, is restricted to the upper Bartonian in Cuba (Torres-Silva et al., 2017). H. ocalana is a widespread species in the upper part of the Priabonian (Robinson & Wright, 1993; Robinson, 2004; Torres-Silva et al., 2017).
Heterostegina ocalana Cushman, 1920
(Fig. 117 ![]() )
)
Previous descriptions and figures. Cushman, 1920, p. 130-131, Pl. XXI, figs. 15-18; Robinson, 2004, Fig. 16A-B; Torres-Silva et al., 2017, p. 586-587, Fig. 18A, B, D-H.
Remarks. The species is highly distinctive and easily identifiable even in random orientations in thin section (Fig. 117.4).
| Figure 117: Heterostegina ocalana Cushman, 1920.
1-3 (ER176), split specimens, equatorial sections: 1, 3,
megalospheric, 2, microspheric; deep-water White Limestone (Chalk), ABZ16,
Dressikie, St Ann, Jamaica. 4 (UWIGM.WL3265B), ABZ16, random section in rock, Swanswick House,
Trelawny, Jamaica. |
Our detailed work on selected LBF that have been calibrated against the nannofossil stratigraphy has allowed us to develop a high-resolution zonation for the Eocene of the Americas. This has also allowed us to revise the classification and phylogeny of the rapidly evolving clades that are useful in zoning these deposits.
The zonation reflects a series of migration events and the in situ evolution of different clades. Migration events are particularly associated with the nummulitids, with important migrations involving short-lived appearances of Nephronummulites and Heterostegina and a major influx of Nummulites and Operculinoides. In contrast, the Helicosteginidae and Lepidocyclinidae show a progressive evolution that is the main basis of the zonation scheme.
Future work will involve collecting larger populations to adequately characterise each species and more work on other groups (e.g., orthophragmines and nummulitids) to improve the zonal resolution. We hope that our zonal scheme will be of value to the work of others and will help towards correlating the Eocene deposits across the Americas.
SFM developed the project, logged and collected material from Jamaica, cut the LBFs, worked on the LBF taxonomy and developed the zonal scheme. ER worked on the taxonomy of the LBFs and provided extensive images of material. EÖ reviewed the taxonomy and provided extensive photomicrographs and measurements of specimens. MMJ provided nannofossil zonations on spot samples and samples collected from logged/collected sections. NR helped collect some of the samples, processed some of the material, cut the thin sections from Sherwood Content, and photographed material in the BMNH. The main text was prepared by SFM, but all authors contributed to the text and agree with the conclusions.
Thanks to Julian Williams and Roshaun Brown for providing field support to SFM while mapping, collecting material and processing material for LBFs. We would particularly like to thank NROCC for allowing us access to collect along N-S Highway 2000 and Long Pond for allowing us access to collect on the type section of the Swanswick Formation at Swanswick House. We thank Shavel Watson and Kenrick McFralane for helping to collect specimens from N-S leg of Highway 2000. EÖ thanks İstanbul Technical University BAP Unit for financial support to visit The University of the West Indies, Mona for a 3-month duration in 2012. We thank Brian Huber of the Smithsonian Institute, Washington - DC, for giving permission to use the images of the types of Tremastegina senni and T. lopeztrigoi. We thank Mike Simmons and Antonino Briguglio for their detailed, constructive and useful reviews of the manuscript which have allowed us to make significant improvements throughout. We thank the editors (Daniela Basso and Bruno Granier) for their help with bringing this paper to publication.
Adams C.G. (1987).- On the classification of the Lepidocyclinidae (Foraminiferida) with redescriptions of the unrelated Paleocene genera Actinosiphon and Orbitosiphon.- Micropaleontology, New York - NY, vol. 33, no. 4, p. 289-317.
Agnini C., Fornaciari E., Raffi I., Catanzariti R., Pälike H., Backman J. & Rio D. (2014).- Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes.- Newsletters on Stratigraphy, Stuttgart, vol. 47, p. 131-181.
Andji G., Baumgartner-Mora C., Baumgartner P.O. & Petrizzo M.R. (2018).- Tectono-stratigraphic response of the Sandino Forearc Basin (N-Costa Rica and W-Nicaragua) to episodes of rough crust and oblique subduction.- The Depositional Record, Hoboken - NJ, vol. 4, no. 1, p. 90-132. DOI: 10.1002/dep2.40
Applin P.L. & Applin E.R. (1944).- Regional subsurface stratigraphy and structure of Florida and southern Georgia.- Bulletin of the American Association of Petroleum Geologists, Tulsa - OK, vol. 28, no. 12, p. 1673-1753.
Applin E.R. & Jordan L. (1945).- Diagnostic Foraminifera from subsurface formations in Florida.- Journal of Paleontology, Tulsa - OK, vol. 19, no. 2, p. 129-148.
Atkinson T.C. (1969).- The geology of the country around Maldon and Maroon Town, St. James, Jamaica.- Journal of the British Speleological Association, Settle, vol. 6, p. 90-95.
Barker R.W. (1932).- Larger Foraminifera from the Eocene of Santa Elena Peninsula, Ecuador.- Geological Magazine, Cambridge - MA, vol. 69, p. 302-310 (Pls. 21-22).
Barker R.W. (1934).- Some notes on the genus Helicolepidina Tobler.- Journal of Paleontology, Cambridge - MA, vol. 8, no. 3, p. 344-351.
Barker R.W. (1939).- Species of the foraminiferal family Camerinidae in the Tertiary and Cretaceous of Mexico.- Proceedings of the United States National Museum, Washington - DC, vol. 86, no. 3052, p. 305-329. URL: https://www.biodiversitylibrary.org/page/7569269
Barker R.W. & Grimsdale T.F. (1936).- A contribution to the phylogeny of the orbitoidal Foraminifera, with descriptions of new forms from the Eocene of Mexico.- Journal of Paleontology, Tulsa - OK, vol. 10, no. 4, p. 231-247 (Pls. 30-38).
Barker R.W. & Grimsdale T.F. (1937).- Studies of Mexican fossil Foraminifera.- The Annals and Magazine of Natural History (ser. 10), London, vol. 19, 161-174 (Pls. 5-9).
Bateson J.H. (Compiler, 1972).- Geological Sheet 6. Maggoty.- 1:50,000 Jamaica, Jamaican Geological Survey, Kingston, Jamaica.
Berggren W.A., Kent D.V., Swisher C.C. III & Aubry M.-P. (1995).- A revised Cenozoic geochronology and chronostratigraphy.- In: Berggren W.A., Kent D.V., Aubry M.-P. & Hardenbol J. (Eds.), Geochronology, time scales and global stratigraphic correlation.- SEPM Special Publication, Tulsa - OK, vol. 54, p. 129-212.
Berry W. (1930).- The larger Foraminifera of the Atascadero Limestone of North-West Peru, S. America.- Eclogae Geologicae Helvetiae, Basel, vol. 23, no. 2, p. 489-496. DOI: 10.5169/seals-158931
Berry W. (1932).- The larger Foraminifera of the Talara shale of north-western Peru.- Journal of the Washington Academy of Sciences, Washington - DC, vol. 22, no. 1, p. 1-9. URL: https://www.biodiversitylibrary.org/page/39917719
Blainville H.M. de (1827).- Manuel de malacology et de conchyliologie (1825).- E.G. Levrault, Paris, Strasbourg, VII+664 p. (5 Pls.).
Blanco-Bustamente S., Fernandez-Rodríguez G. & Fluegeman R.H. (1999).- A note on the biostratigraphy of Paleocene-Eocene larger Foraminifera from western Cuba.- Micropaleontology, New York - NY, vol. 45 (supplement 2), p. 19-26.
Bolli H.M. (1957a).- The genera Globigerina and Globorotalia in the Paleocene-Lower Eocene Lizard Springs Formation of Trinidad, B.W.I.- Bulletin of the United States National Museum, Washington - DC, vol. 215, p. 61-82. URL: https://www.biodiversitylibrary.org/part/70796
Bolli H.M. (1957b).- Planktonic Foraminifera from the Eocene Navet and San Fernando Formations of Trinidad, B.W.I.- Bulletin of the United States National Museum, Washington - DC, vol. 215, p. 155-172. URL: https://www.biodiversitylibrary.org/page/32376667
Bolli H.M. & Saunders J.B. (1985).- Oligocene to Holocene low latitude planktic Foraminifera. In: Bolli H.M., Saunders J.B., Perch-Nielsen K. (Eds.), Plankton stratigraphy.- Cambridge University Press, Cambridge - MA, p. 155-262.
Bosworth T.O. (1922).- Geology of the Tertiary and Quaternary periods in the north-west part of Peru.- MacMillan and Co., London, 434 p.
BouDagher-Fadel M.K. (2018).- Evolution and geological significance of Larger Benthic Foraminifera, 2nd edition.- UCL Press, London, 702 p. DOI: 10.2307/j.ctvqhsq3
Brady H.B. (1875).- On some fossil Foraminifera from the west-coast district of Sumatra.- Geological Magazine, Cambridge - MA, vol. 2, p. 532-539 (Pls. 13-14).
Brönnimann P. (1944).- Ein neues Subgenus von Orbitocyclina aus Iran nebst Bemerkungen uber Helicolepidina Tobler und verwandte Formen.- Schweizerische Paläontologische Abhandlungen, Basel, vol. 64, p. 2-42.
Brönnimann P. (1947).- Zur Neu-Definition von Pliolepidina H. Douvillé, 1915.- Eclogae Geologicae Helvetiae, Basel, vol. 39, p. 373-379. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001:1946:39#400
Brönnimann P. (1950).- Tremastegina, ein neues genus der familie Asterigerinidae d'Orbigny.- Verhandlungen der Schweizerische Naturforschende Gesellschaft, Basel, vol. 166, p. 255-265.
Bruguičre J.G. (1792).- Camerine. In: Encyclopédie méthodique : Histoire naturelle des Vers, vol. 1, p. 395-400. URL: https://www.biodiversitylibrary.org/page/8892383
Bryan J.R. (1995).- Life history and development of Oligocene Larger Benthic Foraminifera: A test of the environmental control on heterochrony.- Tulane Studies in Geology and Paleontology, New Orleans - LA, vol. 27, p. 101-118. URL: https://journals.tulanE.edu/tsgp/article/view/667/585
Bukry D. (1973).- Low latitude coccolith biostratigraphic zonation.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. 15, p. 658-677. URL: http://deepseadrilling.org/15/volume/dsdp15_16.pdf
Bukry D. (1975).- Coccolith and silicoflagellate stratigraphy, northwestern Pacific Ocean.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. 32, p. 67-701. URL: http://deepseadrilling.org/32/volume/dsdp32_24.pdf
Burke K., Coates A. G. & Robinson E. (1969).- Geology of the Benbow Inlier and surrounding areas, Jamaica. In: Saunders J.B. (Ed.), Transactions of the Fourth Caribbean Geological Conference 28th March - 12th April 1965 Port-of-Spain, Trinidad and Tobago.- Caribbean Printers, Arima, Trinidad (dated 1968, published 1969), p. 299-307.
Butterlin J. (1961).- Grandes Foraminiferos del Pozo Palizada No. 2, Municipio de Palizada, Estado de Campeche.- Paleontologia Mexicana, Mexico C.D., no. 10, p. 1-59. URL: http://www.ojs-igl.unam.mx/index.php/Paleontologia/article/view/18/16
Butterlin J. (1970).- Macroforaminiferos y edad de la Formacion Punta Mosquito (Grupo Punta Carnero), de la Isla de Margarita, Venezuela.- Boletin Informativo, Caracas, vol. 13, no. 10, p. 273-308.
Butterlin J. (1971).- Contribution ŕ la connaissance du Paléogčne marin du Nord-Ouest de la Colombie, basée sur les Macroforaminifčres.- Eclogae Geologicae Helvetiae, Basel, vol. 64, p. 13-27. URL: https://www.e-periodica.ch/cntmng?pid=egh-001%3A1971%3A64%3A%3A80
Butterlin J. (1981).- Claves para la determinacion de Macroforaminiferos de Mexico y del Caribe, del Cretacico superior al Mioceno medio.- Instituto Mexicano del Petroleo, Mexico C.D., 219 p. (51 Pls.).
Butterlin J. (1987).- Origine et évolution des Lépidocyclines de la région des Caraibes. Comparaisons et relations avec les Lépidocyclines des autres régions du monde.- Revue de Micropaléontologie, Paris, vol. 29, p. 203-219.
Butterlin J. (1988).- A reexamination of the stratigraphic distribution of the larger Foraminifera in the Caribbean region during the Paleogene and Miocene periods and the implications on the geodynamic history of this region.- Transactions 11th Caribbean Geological Conference, Barbados, July 1986, p. 4:1-4:9.
Cahuzac B. & Poignant A. (1997).- Essai de biozonation de l'Oligo-Miocčne dans les bassins européens ŕ l'aide des grands foraminifčres néritiques.- Bulletin de la Société Géologique de France, Paris, vol. 168, p. 155-169.
Caron V., Bailleul J., Chanier J. & Mahieux G. (2019).- Demise and recovery of Antillean marine carbonate factoires adjacent to active submarine volcanoes (Lutetian-Bartonian).- Sedimentary Geology, vol. 387, p. 104-125.
Caudri C.M.B. (1972).- The larger Foraminifera of the Scotland district of Barbados.- Eclogae Geologicae Helvetiae, Basel, vol. 65, p. 221-234. URL: https://www.e-periodica.ch/cntmng?pid=egh-001:1972:65::898
Caudri C.M.B. (1974).- The larger Foraminifera of Punta Mosquito, Margarita Island, Venezuela.- Verhandlungen der Naturforschenden Gesellschaft, Basel, vol. 84, no. 1, p. 293-318 (15 Pls.).
Caudri C.M.B. (1975).- Geology and paleontology of Soldado Rock, Trinidad (West Indies). Part 2: The larger Foraminifera.- Eclogae geologicae Helvetiae, Basel, vol. 68, p. 533-589. URL: https://www.e-periodica.ch/cntmng?pid=egh-001%3A1975%3A68%3A%3A985
Caudri C.M.B. (1996).- The larger Foraminifera of Trinidad (West Indies).- Eclogae geologicae Helvetiae, Basel, vol. 89, no. 3, p. 1137-1309 (30 Pls.). URL: https://www.e-periodica.ch/cntmng?pid=egh-001%3A1996%3A89%3A%3A1545
Cizancourt M. de (1948).- Nummulites de l'île de Barbade (Petites Antilles).- Mémoire de la Société Géologique de France (nouvelle série), Paris, vol. 27, no. 57, p. 1-40.
Cole W.S. (1929).- Three new Claiborne fossils.- Bulletins of American Paleontology, Ithaca - NY, vol. 15, no. 56, p. 1-10 (Pls. 1-2).
Cole W.S. (1941).- Stratigraphic and Paleontologic Studies of Wells in Florida.- United Brotherhood of Carpenters and Joiners of America, Power House Well 2. Peninsular Oil and Refining Company's J. W. Cory 1. With description of species of Foraminifera from another well.- Florida Geological Survey Bulletin, Tallahassee - FL, no. 19, p. 1-91. URL: https://ufdc.ufl.edu/UF00000451/00001
Cole W.S. (1942).- Stratigraphic and Paleontologic Studies of Wells in Florida - No. 2. Suwannee Petroleum Corporations's Sholtz No. 1, Florida Oil Discovery Company's Cedar Keys No. 2.- Florida Geological Survey Bulletin, Tallahassee - FL, no. 20, p. 1-89. URL: https://ufdc.ufl.edu/UF00000445/00001
Cole W.S. (1944).- Stratigraphic and paleontologic studies of wells in Florida - no. 3. City of Quincy water well, St. Mary's oil corporation, Hilliard turpentine company no. 1 well.- Florida Geological Survey Bulletin, Tallahassee - FL, no. 29, p. 1-168. URL: https://ufdc.ufl.edu/UF00000240/00001
Cole W.S. (1945).- Stratigraphic and Paleontologic Studies of Wells in Florida - No. 4. City of Tallahassee water well No. 6, Dale Mabry Field water well "B", Ravlin-Brown, V. G. Philips No. 1 Well.- Florida Geological Survey Bulletin, Tallahassee - FL, no. 28, p. 1-160. URL: https://original-ufdc.uflib.ufl.edu/UF00000456/00001
Cole W.S. (1952).- Eocene and Oligocene larger Foraminifera from the Panama Canal Zone and vicinity.- U.S. Geological Survey Professional Paper, Washington - DC, no. 244, p. 1-41. URL: https://pubs.usgs.gov/pp/0244/report.pdf
Cole W.S. (1956).- Jamaican larger Foraminifera.- Bulletins of American Paleontology, Ithaca - NY, vol. 36, p. 203-233. URL: https://www.biodiversitylibrary.org/item/40434
Cole W.S. (1958a).- Names of and variation in certain American larger Foraminifera, no. 1.- Bulletins of American Paleontology, Ithaca - NY, vol. 38, no. 170, p. 175-213 (Pls. 18-25). URL: https://www.biodiversitylibrary.org/item/91637
Cole W.S. (1958b).- Names of and variation in certain American larger Foraminifera, particularly the Camerinids - No. 2.- Bulletins of American Paleontology, Ithaca - NY, vol. 38, no. 173, 261-284 (Pls. 32-34). URL: https://www.biodiversitylibrary.org/item/89065
Cole W.S. (1960).- Revision of Helicostegina, Helicolepidina and Lepidocyclina (Polylepidina).- Contributions from the Cushman Foundation for Foraminiferal Research, Lawrence - KS, vol. 11, p. 57-63 (Pls. 10-13). URL: https://cushmanfoundation.org/PersonifyEbusiness/Portals/0/pdf/pubarchive/ccffr/11ccffr2.pdf
Cole W.S. (1963).- Illustraions of the conflicting interpretations of the biology and classification of certain larger Foraminifera.- Bulletins of American Paleontology, vol. 46, p. 6-63. URL: https://www.biodiversitylibrary.org/item/40515#page/9/mode/1up
Cole W.S. (1969).- Internal structure, stratigraphical range, and phylogenetic relationships of certain American Eocene Foraminifera.- Contributions from the Cushman Foundation for Foraminiferal Research, Lawrence - KS, vol. 20, p. 77-86. URL: https://cushmanfoundation.org/PersonifyEbusiness/Portals/0/pdf/pubarchive/ccffr/20ccffr3.pdf
Cole W.S. & Applin E.R. (1964).- Problems of the geographic and stratigraphic distribution of American middle Eocene large Foraminifera.- Bulletins of American Paleontology, Ithaca - NY, vol. 47, (no. 212), p. 1-48. URL: https://www.biodiversitylibrary.org/item/91774
Cole W.S. & Bermudez P.J. (1944).- New foraminiferal genera from the Cuban Middle Eocene.- Bulletins of American Paleontology, Ithaca - NY, vol. 28, (113), p. 333-334. URL: https://www.biodiversitylibrary.org/item/40554
Cole W.S. & Bermudez P.J. (1947).- Eocene Discocyclinidae and other Foraminifera from Cuba.- Bulletins of American Paleontology, Ithaca - NY, vol. 31, (120), p. 1-36. URL: https://www.biodiversitylibrary.org/item/40477
Cole W.S. & Gravell D.W. (1952).- Middle Eocene Foraminifera from Peńon Seep, Matanzas Province, Cuba.- Journal of Paleontology, Tulsa - OK, vol. 26, p. 708-727.
Cole W.S. & Ponton G.M. (1934).- New species of Fabularia, Asterocyclina, and Lepidocyclina from the Florida Eocene.- American Midland Naturalist, Notre Dame - IN, vol. 15, no. 2, p. 138-147.
Coletti G., Bosio G., Collareta A., Malinverno E., Bracchi V.A., Di Celma C., Basso D., Stainbank S., Spezzaferri S., Cannings T. & Bianucci G. (2019).- Biostratigraphic, evolutionary, and paleoenvironmental significance of the southernmost lepidocyclinids of the Pacific coast of South America (East Pisco Basin, southern Peru).- Journal of South American Earth Sciences, vol. 96, 102372.
Cornée J.-J., BouDagher-Fadel M., Philippon M., Léticée J.L., Legendre L., Maincent G., Lebrun J.-F. & Münch P. (2020).- Paleogene carbonate systems of Saint Barthélemy, Lesser Antilles: Stratigraphy and general organization.- Newsletters on Stratigraphy, Stuttgart, vol. 53, no. 4, p. 461-478.
Cotton L.J., Eder W. & Floyd J. (2018).- Larger Foraminifera of the Devil's Den and Blue Hole sinkholes, Florida.- Journal of Micropalaeontology, vol. 37, no. 1, p. 347-356. URL: https://jm.copernicus.org/articles/37/347/2018/
Cushman J.A. (1918).- The larger fossil Foraminifera of the Panama Canal Zone.- Bulletin of the U.S. National Museum, Washington - DC, vol. 103, p. 89-102 (Pls. 34-45).
Cushman J.A. (1919).- Fossil Foraminifera from the West Indies. In: Vaughan T.W. (Ed.), Contributions to the geology and paleontology of the West Indies.- Carnegie Institution, Publications, Washington - DC, no. 291, p. 23-71 (15 Pls.).
Cushman J. (1920).- The American species of Orthophragmina and Lepidocyclina.- USGS Professional Paper, Reston - VA, no. 125-D, 39-105. DOI: 10.3133/pp125d
Cushman J.A. (1927).- An outline of a re-classification of the Foraminifera.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon - MA, vol. 3, p. 1-105. URL: https://cushmanfoundation.org/PersonifyEbusiness/Portals/0/pdf/pubarchive/cclfr/1cclfr1.pdf
Cushman J.A. & Stainforth R.M. (1946).- A new species of Amphistegina from the Eocene of Ecuador.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon - MA, vol. 22, no. 4, p. 117-119. URL: https://cushmanfoundation.org/PersonifyEbusiness/Portals/0/pdf/pubarchive/cclfr/22cclfr4.pdf
Cushman J.A. & Stainforth R.M. (1951).- Tertiary Foraminifera of coastal Ecuador: Part I, Eocene.- Journal of Paleontology, Tulsa - OK, vol. 25, no. 2, p. 129-164.
Douvillé H. (1911).- Les foraminifčres dans le Tertiaire des Philippines.- The Philippine Journal of Science, Manila, vol. VI, no. 2, p. 53-80. URL: https://www.biodiversitylibrary.org/page/34354759
Douvillé H. (1915).- Les orbitoďdés du Danien et du Tertiaire : Orthophragmina et Lepidocyclina.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 161, p. 721-728. URL: https://gallica.bnf.fr/ark:/12148/bpt6k3114b/f721.item
Douvillé H. (1917).- Les Orbitoďdes de l'Ile de la Trinité.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 164, p. 841-849. URL: http://gallica.bnf.fr/ark:/12148/bpt6k31178/f841.image
Douvillé H. (1922).- Les Lépidocyclines et leur évolution : Un genre nouveau "Amphilepidina".- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 175, p. 550-555. https://gallica.bnf.fr/ark:/12148/bpt6k3128v/f550.item
Douvillé H. (1924).- Les Orbitoďdes et leur évolution en Amérique.- Bulletin de la Société Géologique de France (4e Série), Paris, vol. 23, no. 7/8, p. 369-376. URL: https://www.biodiversitylibrary.org/page/31475656#page/451/mode/1up
Drooger C.W. (1993).- Radial Foraminifera: Morphometrics and evolution.- Verhandelingen der Koninklijke Nederlandse Akademie van Westenschappen, Afdeling Natuurkunde (Eerste Reeks), Amsterdam, Deel 41, 242 p.
Eames F.E, Banner F.T., Blow W.H., Clarke W.J. & Smout A.H. (1962).- Morphology, taxonomy, and stratigraphic occurrence of the Lepidocyclininae.- Micropaleontology, New York - NY, vol. 8, no. 3, p. 289-322 (Pls. 1-8).
Eames F.E., Clarke W.J., Banner F.T., Smout A.H. & Blow W.H. (1968).- Some larger Foraminifera from the Tertiary of Central America.- Palaeontology, Tulsa - OK, vol. 11, p. 283-305. URL: https://www.palass.org/publications/palaeontology-journal/archive/11/2/article_pp283-305
Eva A.N. (1976).- The palaeoecology and sedimentology of middle Eocene larger Foraminifera in Jamaica.- Maritime Sediments Special Publication, Fredericton - NB, no. 1, 467-475.
Eva A.N. (1980).- Pre-cyclical chamber arrangement in the foraminiferal genus Polylepidina Vaughan 1924.- Micropaleontology, New York - NY, vol. 26, p. 90-94.
Ferrŕndez-Cańadell C. & Serra-Kiel J. (1999).- Morphostructure and systematics of Linderina brugesi Schlumberger, 1893 (Foraminifera, Eocene).- Géobios, Villeurbanne, vol. 32, p. 525-537.
Frost S.H. & Langenheim R.L. (1974).- Cenozoic reef biofacies. Tertiary larger Foraminifera and scleractinian corals from Mexico.- Northern Illinois University Press, DeKalb - IL, 388 p. (123 Pls.).
Geyn W.A.E. & Vlerk I.M. van der (1935).- A monograph on the Orbitoididae, occurring in the Tertiary of America, compiled in connection with an examination of a collection of larger Foraminifera from Trinidad.- Leidsche geologische Mededeelingen, vol. 7, no. 2, p. 221-272. URL: http://www.repository.naturalis.nl/record/505742
Gorsel J.T. van (1978).- Late Cretaceous orbitoidal Foraminifera. In: Hedley R.G. & Adams C.G. (Eds.), Foraminifera 3.- Academic Press, Cambridge - MA, p. 1-120.
Gorter N.E. & Vlerk I.M. van der (1932).- Larger Foraminifera from Central Falcon (Venezuela).- Leidsche Geologische Mededeelingen, vol. 4, no. 2, p. 94-122
Gravell D.W. (1933).- Tertiary larger Foraminifera of Venezuela.- Smithsonian Miscellaneous Collections, Washington - DC, vol. 89, no. 11, p. 1-44.
Gravell D.W. & Hanna M.A. (1938).- Subsurface Tertiary zone of correlation through Mississippi, Alabama, and Florida.- Bulletin of the American Association of Petroleum Geologists, Tulsa - OK, vol. 22, p. 984-1013.
Grimsdale T.F. (1959).- Evolution in the American Lepidocyclinidae (Cainozoic Foraminifera): An interim view.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, ser. B, vol. 62, p. 8-33.
Gümbel C.W. (1870).- Beiträge zur Foraminiferenfauna der Nordalpinen Eocängebilde.- Abhandlungen der Mathematisch-Physicalischen Classe der Koniglich Bayerischen Akademie der Wissenschaften, vol. 10 (for 1868), p. 581-730 (Pls. 1-4).
Haman D. & Huddleston R.W. (1984).- Caudriella, a new name for Margaritella Caudri, 1974 (Foraminiferida), non Meek and Hayden, 1860 (Mollusca).- Proceedings of the Biological Society of Washington, Washington - DC, vol. 97, p. 126.
Hanzawa S. (1932).- A new type of Lepidocyclina with a multilocular nucleoconch from the Taito Mountains, Taiwan (Formosa).- Proceedings of the Imperial Academy of Japan, Tokyo, vol. 8, p. 446-449.
Hanzawa S. (1935).- Some fossil Operculina and Miogypsina from Japan and their stratigraphical significance.- Science Reports of the Tohoku University (ser. 2, Geology), Sendai, vol. 18, no. 1, p. 1-29. URL: http://hdl.handle.net/10097/30260
Heilprin A. (1882).- On the occurrence of nummulitic deposits in Florida, and the association of Nummulites with a fresh-water fauna.- Proceedings of the Academy of Natural Sciences of Philadelphia, Philadelphia - PA, vol. 34, no. 2, p. 189-193.
Hinte J.E. van (1965).- An approach to Orbitoides.- Proceedings of the Koninklijke Nederlandse Akademie Van Wetenschappen (Series B), Amsterdam, vol. 68, p. 57-70.
Hofker J. (1956).- Tertiary Foraminifera of coastal Ecuador, Pt. II, additional notes on the Eocene species.- Journal of Paleontology, Tulsa - OK, vol. 30, p. 891-958.
Hohenegger J. & Torres Silva A.I. (2020).- Methods for testing ontogenetic changes of neanic chamberlets in lepidocyclinids.- Journal of Foraminiferal Research, Lawrence - KS, vol. 50, no. 2, p. 182-194.
Hose H.R. & Versey H.R. (1956).- Palaeontological and lithological divisions of the Lower Tertiary limestones of Jamaica.- Colonial Geology and Mineral Resources, London, vol. 6, p. 19-39.
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, no. M02 (CG2006_M02), p. 1-126. DOI: 10.4267/2042/5832
ICZN (International Commission on Zoological Nomenclature) (1945).- Opinion 192. Suspension of Rules for Nummulites Lamarck, 1801 (Class Rhizopoda, Order Foraminifera).- Opinions and Declarations Rendered by the International Commission on Zoological Nomenclature, vol. 3, no. 11, p. 137-160. URL: https://www.biodiversitylibrary.org/part/149718
Iturralde-Vinent M.A. García-Casco A., Rojas-Agramonte Y., Proenza J.A., Murphy J.B. & Stern R.J. (2016).- The geology of Cuba: A brief overview and synthesis.- GSA Today, Boulder - CO, vol. 26, no. 10, p. 4-10. URL: https://www.geosociety.org/gsatoday/archive/26/10/pdf/i1052-5173-26-10-4.pdf
Jiang M.-J. & Robinson E. (1987).- Calcareous nannofossils and larger Foraminifera in Jamaican rocks of Cretaceous to early Eocene age. In: Ahmad R. (Ed.), Proceedings of a workshop on the status of Jamaican geology.- Geological Society of Jamaica, Signart Printing House, Kingston, Jamaica, p. 24-51.
Jones R.W. (2009).- Stratigraphy, palaeoenvironmental interpretation and uplift history of Barbados based on foraminiferal and other palaeontological evidence.- Journal of Micropalaeontology, London, vol. 28, p. 37-44.
Jones T.R. & Chapman F. (1900).- On the Foraminifera of the orbitoidal limestones and reef rocks of Christmas Island. In: Andrews C.W. (ed.), A monograph of Christmas Island (Indian Ocean).- British Museum (Natural History), London, p. 226-264.
Kalia P. & Banerjee A. (1995).- Occurrence of the genus Eoconuloides Cole and Bermudez 1944 in Rajasthan, India and Its paleobiogeographic significance.- Micropaleontology, New York - NY, vol. 41, no. 2, p. 187-194.
Krüger R., Röttger R., Lietz R. & Hohenegger J. (1997).- Biology and reproductive processes of the larger foraminiferan Cycloclypeus carpenteri (Protozoa, Nummulitidae).- Archiv für Protistenkunde, vol. 147, nos. 3-4, p. 307-321.
Kugler H.G. & Caudri C.M.B. (1975).- Geology and paleontology of Soldado Rock Trinidad (West indies). Part 1 Geology and biostratigraphy.- Eclogae Geologicae Helvetiae, Basal, vol. 68, no. 2, p. 365-430. URL: https://www.e-periodica.ch/cntmng?pid=egh-001%3A1975%3A68%3A%3A934
Lamarck J.B. de (1801).- Systęme des Animaux sans vertčbres ou Tableau général des classes, des ordres et des genres de ces animaux.- Paris, p. 432 p. URL: https://archive.org/download/systmedesanima00lama/systmedesanima00lama.pdf
Langer M.R. & Hottinger L. (2000).- Biogeography of selected "larger" Foraminifera.- Micropaleontology, New York - NY, vol. 46, Suppl. 1, p. 105-127.
Legendre L., Philippon M., Münch P., Léticée J.L., Nouy M., Maincent G., Cornée J.-J., Caravati A., Lebrun J.-F. & Mazabraud Y. (2018).- Trench bending initiation: Upper plate strain pattern and volcanism. Insights from the Lesser Antillea Arc, St. Barthélemy Island, French West Indies.- Tectonics, vol. 37, p. 2777-2797. DOI: 10.1029/2017TC004921
Lemoine P. & Douvillé R. (1904).- Sur le genre Lepidocyclina Gümbel.- Mémoires de la Société Géologique de France (Paléontologie), no. 32, p. 1-41 (Pls. 1-3). URL: https://patrimoine.sorbonne-universite.fr/idurl/1/2830
Less G. (1987).- Paleontology and stratigraphy of the European Orthophragminae.- Geologica Hungarica (series Palaeontologica), Budapest, vol. 51, 373 p.
Less G., Frijia G., Özcan E., Saraswati P.K., Parente M. & Kumar P. (2018).- Nummulitids, lepidocyclinids and Sr-isotope data from the Oligocene of Kutch (western India) with chronostratigraphic and paleobiogeographic evaluations.- Geodinamica Acta, vol. 30 no. 1, p. 183-211. DOI: 10.1080/09853111.2018.1465214
Less G. & Özcan E. (2012).- Bartonian-Priabonian larger benthic foraminiferal events in the Western Tethys.- Austrian Journal of Earth Sciences, Vienna, vol. 105, no. 1, p. 129-140. URL: http://www.univie.ac.at/ajes/archive/volume_105_1/less_oezcan_ajes_v105_1.pdf
Levin H.L. (1957).- Micropaleontology of the Oldsmar Limestone (Eocene) of Florida.- Micropaleontology, New York - NY, vol. 3, p. 137-154.
Lewis J. & Robinson E. (1976).- A revised stratigraphy and geological History of the Lesser Antilles.- Transactions of the VII Conference Géologique des Caraďbes, St. Francois, Guadeloupe, p. 339-344.
Lisson C.I. (1921).- Contribución al estudio de algunos foraminiferos terciarios provenientes de la región del Norte del Perú.- Archivos de la Asociación peruana para el Progreso de la Ciencia, Lima, Tomo I, fasc. 1, p. 52-55 (Pls. III-V). URL: https://marinespecies.org/foraminifera./aphia.php?p=sourceget&id=146612
Loeblich A.R. Jr & Tappan H. (1964).- Protista 2: Sarcodina chiefly "Thecamoebians" and Foraminiferida. In: Moore R.C. (Ed.), Treatise on Invertebrate Paleontology. Part C.- Geological Society of America, Boulder - CO; University of Kansas, Lawrence - KS, Part C, vols. 1-2, 550 p.
Loeblich A.R. Jr & Tappan H. (1984).- Suprageneric Classification of the Foraminiferida (Protozoa).- Micropaleontology, New York - NY, vol. 30, no. 1, p. 1-70.
Loeblich A.R. Jr & Tappan H. (1988).- Foraminiferal genera and their classification.* Van Nostrand Reinhold Co., New York - NY, 730 p.
Mann P. & Burke K. (1990).- Transverse intra-arc rifting: Palaeogene Wagwater Belt, Jamaica.- Marine and Petroleum Geology, vol. 7, p. 410-427.
Martini E. (1971).- Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: Farinacci A. (Ed.), Proceedings of the II Planktonic Conference, Roma, 1969.- Tecnoscienza, Rome, p. 739-785.
Matley C.A. (1925).- Recent geological work in Jamaica.- Report to the British Association (Toronto 1924), p. 391-392.
Meek F.B. & Hayden F.V. (1860).- Systematic catalogue with synonyma, etc., of Jurassic, Cretaceous and Tertiary fossils collected in Nebraska, by the exploring expeditions under the command of Lieut. G.K. Warren of U.S. Topographical Engineers.- Proceedings of the National Academy of Sciences, Philadelphia - PA, 1860, p. 417-432. URL: https://archive.org/details/jstor-4059386
Mello e Sousa S.H. de, Fairchild T.R. & Tibana P. (2003).- Cenozoic biostratigraphy of Larger Foraminifera from the Foz Do Amazonas Basin, Brazil.- Micropaleontology, New York - NY, vol. 49, no. 3, p. 253-266.
Meyerhoff A.A. & Krieg E.A. (1977).- Petroleum potential of Jamaica.- Ministry of Mining and Natural Resources, Kingston, Jamaica, 131 p.
Michelotti G. (1861).- Études sur le Miocčne inférieur de l'Italie septentrionale.- Les Héritiers Loosjes, Harlem, 184 p. (16 Pls.).
Miller J.A. (1986).- Hydrogeologic framework of the Floridan aquifer system in Florida and in parts of Georgia, South Carolina, and Alabama.- USGS Professional Paper 1403-B, p. B1-B91. URL: https://pubs.usgs.gov/pp/1807/pdf/pp1807.pdf
Mitchell S.F. (2004).- Lithostratigraphy and palaeogeography of the White Limestone Group. In: Donovan S.K. (Ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Rotterdam, vol. 3, p. 5-29.
Mitchell S.F. (2013a).- Stratigraphy of the White Limestone of Jamaica.- Bulletin de la Société Géologique de France, vol. 184, no. 1-2, p. 111-118.
Mitchell S.F. (2013b).- Lithostratigraphy of the Central Inlier, Jamaica.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 46, p. 31-42. URL: http://caribjes.com/CJESpdf/CJES46-03-MitchellCentral%20InlieR.pdf
Mitchell S.F. (2016).- Geology of the western margin of the Benbow Inlier - implications for the relationship between the Yellow Limestone and White Limestone groups (with the description of the Litchfield Formation, new name).- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 48, p. 19-25. URL: http://caribjes.com/CJESpdf/CJES48-2-Mitchell2016WhiteLimestoneUnconformity.pdf
Mitchell S.F. (2020).- Cretaceous geology and tectonic assembly of Jamaica. In: Davison I, Hull J.N.F. & Pindell J. (Eds.), The basins, orogens and evolution of the southern Gulf of Mexico and northern Caribbean.- Geological Society of London, Special publication, SP504, p. 507-547.
Mitchell S.F. (2021).- The first Paleogene transgression onto the Clarendon Block (Jamaica).- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 53, p. 1-10. URL: http://caribjes.com/CJESpdf/CJES53-1-Mitchell-Bottom-Leinster-Inlier.pdf
Mitchell S.F. & Edwards T.C.P. (2016).- Geology of the Maastrichtian (Upper Cretaceous) succession of the Jerusalem Mountain Inlier in western Jamaica.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 48, p. 29-36. URL: http://caribjes.com/CJESpdf/CJES48-4-Mitchell-Edwards-JerusalemMountain.pdf
Molina E., Torres-Silva A.I., Ćorić S. & Briguglio A. (2015).- Integrated biostratigraphy across the Eocene/Oligocene boundary at Norońa, Cuba, and the question of the extinction of orthophragminids.- Newsletters on Stratigraphy, vol. 49 (2016), p. 27-40.
Morton S.G. (1833).- Supplement to the "Synopsis of the organic remains of the ferruginous sand formation of the United States," contained in vols. XVII and XVIII of this journal.- American Journal of Science and Arts, vol. 23, p. 288-294. URL: https://biodiversitylibrary.org/page/27909388
Nigam R. & Rao A.S. (1987).- Proloculus size variation in recent benthic Foraminifera: Implications for paleoclimatic studies.- Estuarine, Coastal and Shelf Science, vol. 24, no. 5, p. 649-655.
Nuttall W.L.F. (1930).- Eocene Foraminifera from Mexico.- Journal of Paleontology, Tulsa - OK, vol. 4, p. 271-293 (Pls. 23-25).
Ogg J.G., Ogg G.M. & Gradstein F.M. (2016).- A concise geologic time scale.- Elsevier, Amsterdam, 234 p.
O'Herne L. (1974).- A reconsideration of Amphistegina lessonii d'Orbigny, 1826, sensu Brady, 1884 (Foraminifera).- Scripta Geologica, Leiden, vol. 26, p. 1-53. URL: https://repository.naturalis.nl/pub/317469
Okada H. & Bukry D. (1980).- Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry 1973; 1975).- Marine Micropaleontology, vol. 51, p. 321-325. URL: https://doi.org/10.1016/0377-8398(80)90016-X
Orbigny A.D. d' (1826).- Tableau méthodique de la classe des Céphalopodes.- Annales des Sciences Naturelles, Paris, vol. 7, p. 96-169, 245-314.
Orbigny A.D. d' (1839).- Foraminifčres. In: Sagra R. de la (Ed.), Histoire physique, politique et naturelle de l'ile de Cuba.- Arthus Bertrand, Paris, 224 p. URL: https://gallica.bnf.fr/ark:/12148/bpt6k975884/f1.item
Özcan E., Ercan K., Gül M.A., Özkan R., Yücel A.O. & Acar Ş. (2021b).- The first record of genus Linderina Schlumberger (Foraminifera) from the Ypresian of Neo-Tethys: Data from the Kozaklı Basin, Nevşehir, Central Turkey.- Mediterranean Geoscience Reviews, vol. 3, p. 253-268.
Özcan E., Kumar Saraswati P., Yücel A.O., Ali N. & Hanif M. (2018).- Bartonian orthophragminids from the Fulra Limestone (Kutch, W India) and coeval units in Sulaiman Range, Pakistan: A synthesis of shallow benthic zone (SBZ) 17 for the Indian Subcontinent.- Geodinamica Acta, vol. 30, no. 1, p. 137-162.
Özcan E., Mitchell S.F., Less G., Robinson E., Bryan J.R., Pignatti J. & Yücel A.O. (2019).- A revised suprageneric classification of American orthophragminids with emphasis on late Paleocene representatives from Jamaica and Alabama.- Journal of Systematic Palaeontology, vol. 17, p. 1551-1579.
Özcan E., Yücel A.O., Catanzariti R., Kayğılı S., Okay A.I., Simmons M.D., Pignatti J., Abbasi İ.A. & Erbil Ü. (2021a).- Multiple Orbitoides d'Orbigny lineages in the Maastrichtian? Data from the Central Sakarya Basin (Turkey) and Arabian Platform successions (Southeastern Turkey and Oman).- Swiss Journal of Palaeontology, volume 140, article no. 8, 30 p. DOI: 10.1186/s13358-021-00219-x
Özcan E., Yücel A.O., Erkizan L.S., Gültekin M.N., kayğılı S.& Yurtsever S. (2022a).- Atlas of the Tethyan orthophragmines.- Mediterranean Geoscience Reviews, vol. 4, p. 3-213. DOI: https://doi.org/10.1007/s42990-022-00072-1
Özcan E., Yücel A.O., Mitchell S.F., Pignatti J., Simmons M.D., Okay A.I., Erkızan L.S. & Gültekin M.N. (2022b).- New records of Caudriella Haman and Huddleston from the middle and late Eocene of Neo-Tethys: Taxonomic and paleobiogeographic implications.- Journal of Foraminiferal Research, Lawrence - KS, vol. 52, p. 21-39.
Palmer D.K. (1934).- Some large fossil Foraminifera from Cuba.- Memorias de Ia Sociedad Cubana de Historia Natural "Felipe Poey", vol. 8, p. 235-264 (Pls. 12-16).
Papazzoni C.A., Ćosović V., Briguglio A. & Drobne K. (2017).- Towards a calibrated larger Foraminifera biostratigraphic zonation: Celebrating 18 years of the application of shallow benthic zone.- Palaios, Lawrence - KS, vol. 32, p. 1-5.
Parker W.K., Jones T.R. & Brady H.B. (1865).- On the nomenclature of the Foraminifera. Pt. XII. The species enumerated by d'Orbigny in the "Annales des Sciences Naturelles," vol. vii. 1826.- Annals and Magazine of Natural History (ser. 3), London, vol. 16, p. 15-41.
Pearson P.N., Olsson R.K., Huber B.T., Hemleben C. & Berggren W.A. (2006).- Atlas of Eocene Planktonic Foraminifera.- Cushman Foundation Special Publication, Lawrence - KS, no. 41, p. 514 p.
Pindell J. & Kennan L. (2009).- Tectonic evolution of the Gulf of Mexico, Caribbean and northern South America in the mantle reference frame: An update.- Geological Society London Special Publications, no. 328, p. 1-55.
Powell J.B. (2010).- Larger foraminiferal biostratigraphy, systematics and paleoenvironments of the Avon Park Formation and Ocala Limestone, Highlands County, Florida.- PhD thesis, Florida International University, Miami - FL, 239 p. URL: https://digitalcommons.fiu.edu/etd/288/
Raadshooven B. van (1951).- On some Paleocene and Eocene larger Foraminifera of western Venezuela.- Third World Petroleum Congress, The Hague, Sect. 1, p. 476-489 (Pls. 1-3).
Reiss Z. (1963).- Reclassification of perforate Foraminifera.- Bulletin of the Geological Survey of Israel, Jerusalem, vol. 35, p. 1-111.
Robinson E. (1974).- Some larger Foraminifera from the Eocene limestones at Red Gal Ring, Jamaica.- Verhandlungen der Naturforschenden Gesellschaft in Basel, vol. 84, p. 281-292.
Robinson E. (1977).- Larger imperforate foraminiferal zone of the Eocene of central Jamaica. In: Segundo Congreso Latinoamericano de Geología, Caracas, Venezuela, 11 al 16 de Noviembre de 1973, Tomo III.- Ministerio de Minas e Hidrocarburos, Caracas, p. 1413-1421.
Robinson E. (1988).- Early Tertiary larger Foraminifera and platform carbonates of the northern Caribbean. In: Barker L. (ed.), Transactions of the 11th Caribbean Geological Conference, Barbados, July 20-26, 1986.- Energy and Natural Resources Division, National Petroleum Corporation, Barbados, p. 5:1-5:12.
Robinson E. (1993).- Some imperforate larger Foraminifera from the Paleogene of Jamaica and the Nicaragua Rise.- Journal of Foraminiferal Research, Lawrence - KS, vol. 23, no. 1, p. 47-65.
Robinson E. (1996).- Using larger foraminifers in high resolution biostratigraphy: An example from the Eocene of the Gulf of Mexico and northern Caribbean.- Palaios, Lawrence - KS, vol. 11, p. 220-229.
Robinson E. (1997).- The Eocene larger foraminifer Lepidocyclina ariana Cole and Ponton from the so-called Polylepidina gardnerae horizon, Little Stave Creek, Alabama.- Journal of Paleontology, Tulsa - OK, vol. 71, p. 1-5.
Robinson E. (2004).- Zoning the White Limestone Group of Jamaica using larger foraminiferal genera: A review and proposal. In: Donovan S.K. (Ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Rotterdam, vol. 3, no. 1-2, p. 39-75. URL: https://natuurtijdschriften.nl/pub/541702
Robinson E. & Jiang M.-J. (1990).- Paleogene calcareous nannofossils from western Portland, and the ages and significance of the Richmond and Mooretown formations of Jamaica.- Journal of the Geological Society of Jamaica, Kingston, Jamaica, vol. 27, p. 17-25.
Robinson E. & Jiang M.-J. (1995).- Evolution of the foraminiferal genus Lepidocyclina in the Middle Eocene and its implication for Gulf Coast stratigraphy.- Gulf Coast Association of Geological Societies Transactions, vol. 45, p. 509-517.
Robinson E. & Mitchell S.F. (1999).- Middle Eocene to Oligocene stratigraphy and palaeogeography in Jamaica: A window on the Nicaragua Rise.- Contributions to Geology, UWI, Mona, Kingston, Jamaica, no. 4, 47 p. URL: http://www.sfmgeology.com/pdfs/MiddleEocenetoOligoceneStratigraphy.pdf
Robinson E., Paytan A., Chien C.-T. & Broach K. (2018).- Dating the White Limestone of Jamaica using Sr isotope stratigraphy: a progress report.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 49, p. 11-21. URL: http://caribjes.com/CJESpdf/CJES49-2-RobinsonSrIsotopes.pdf
Robinson E. & Wright R.M. (1993).- Jamaican Paleogene larger Foraminifera. In: Wright R.M. & Robinson E. (Eds.), Jamaican biostratigraphy.- Geological Society of America Memoir, Boulder - CO, vol. 182, p. 283-345.
Rutten L.M.R. (1928).- On Tertiary rocks and Foraminifera from north-west Peru.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (ser. B), Amsterdam, vol. 31, p. 931-946 (Pls. 1-2).
Rutten M.G. (1935).- Larger Foraminifera of Northern Santa Clara Province Cuba.- Journal of Paleontology, Tulsa - OK, vol. 9, p. 527-545.
Rutten M.G. & Vermunt L.W.J. (1932).- The Serce di Cueba limestone from Curaçao.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, vol. 35, p. 227-240.
Scheffen W. (1932).- Zur Morphologie und Morphogenese der Lepidocyclinen.- Paläontologische Zeitschrift, vol. 14, p. 233-256, (Pls. 9-10).
Schenck H.G. & Frizzell D. L. (1936).- Subgeneric nomenclature in Foraminifera.- American Journal of Science (ser. 5), New Haven - CT, vol. 31, p. 363-366.
Schlumberger C. (1893).- Note sur les genres Trillina et Linderina.- Bulletin de la Société Géologique de France (3e Série), Paris, vol. 21, no. 2, p. 118-123. URL: http://www.marinespecies.org/foraminifera/aphia.php?p=sourceget&id=146841
Schwager C. (1876).- Saggio di una classificazione dei foraminiferi avuto riguardo alle loro famiglie naturali.- Bolletino R. Comitato Geologico d'Italia, Firenze, vol. 7, p. 475-485. URL: https://biodiversitylibrary.org/page/53477189
Senn A. (1940).- Paleogene of Barbados and its bearing on history and structure of the Antillean Caribbean region.- Bulletin of the American Association of Petroleum Geologists, Tulsa - OK, vol. 24, no. 9, p. 1548-1610.
Serra-Kiel J., Hottinger L., Caus E., Drobne K., Ferrŕndez C., Kumar Jauhri A., Less G., Pavlovec R., Pignatti J., Samsó J.M., Schaub H., Sirel E., Strougo A., Tambareau Y., Tosquella J. & Zakrevskaya E. (1998).- Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene.- Bulletin de la Société Géologique de France, Paris, vol. 169, p. 281-299.
Serra-Kiel J., Ferrŕndez-Cańadell C., García-Senz J. & Hernaiz Huerta P.P. (2007).- Cainozoic larger foraminifers from Dominican Republic.- Boletín Geológico y Minero, Madrid, vol. 118, no. 2, p. 359-384. URL:https://www.igme.es/boletin/2007/118_2_2007/ART.%2010.pdf
Silvestri A. (1937).- Foraminiferi dell'Oligocene e del Miocene della Somalia.- Palaeontographia Italica, Pisa, vol. 32, sup. 2, p. 45-264 (Pls. 4-22).
Sirotti A. (1983).- Phylogenetic classification of Lepidocyclinidae; a proposal.- Bollettino della Societa Paleontologica Italiana, Milano, vol. 21 (for 1982), no. 1, p. 99-112.
Speijer R.P., Pälike H., Hollis C.J., Hooker J.J. & Ogg J.GG. (2020).- Chapter 28 - The Paleogene Period. In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (Eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 1087-1140.
Tan S.H. (1936).- Zur Kenntnis der Lepidocycliniden.- Natuurkundig Tijdschrift voor Nederlandsch-Indie, Batavia (= Jakarta), vol. 96, p. 235-280.
Tan S.H. (1939).- On Polylepidina, Orbitocyclina, and Lepidorbitoides.- De Ingenieur in Nederlandsch-Indië (IV - Mijnbouw en Geologie), Batavia (= Jakarta), vol. 6, p. 53-84.
Thalmann H.E. (1938).- Bibliography and index to new genera, species and varieties of Foraminifera for 1935.- Journal of Paleontology, Tulsa - OK, vol. 12, p. 177-208.
Tobler A. (1922).- Helicolepidina, ein neues Subgenus von Lepidocyclina.- Eclogae Geologicae Helvetiae, Basel, vol. 17, p. 380-384. URL: https://www.e-periodica.ch/cntmng?pid=egh-001%3A1922%3A17%3A%3A750
Todd J.U. & Barker R.W. (1932).- Tertiary orbitoids from northwestern Peru.- Geological Magazine, Cambridge - MA, vol. 69, p. 529-543 (Pls. 39-42).
Torres-Silva A.I., Hohenegger J., Ćorić S., Briguglio A. & Eder W. (2017).- Biostratigraphy and evolutionary tendencies of Eocene heterostegines in western and central Cuba based on morphometric analyses.- Palaios, Lawrence - KS, vol. 32, p. 44-60.
Torres-Silva A.I., Eder W., Hohenegger J. & Briguglio A. (2018).- Morphometric analysis of Eocene nummulitids in western and central Cuba: Taxonomy, biostratigraphy and evolutionary trends.- Journal of Systematic Palaeontology, vol. 17, no. 7, p. 557-595. DOI: 10.1080/14772019.2018.1446462
Vandenberghe N., Hilgen F.J. & Speijer R.P. (2012).- The Paleogene Period. In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (Eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 855-921.
Vaughan T.W. (1924).- American and European Tertiary larger Foraminifera.- Bulletin of the Geological Society of America, Boulder - CO, vol. 35, p. 785-822 (Pls. 30-38).
Vaughan T.W. (1927).- Notes on the types of Lepidocyclina mantelli (Morton) Gümbel and on topotypes of Nummulites floridanus Conrad.- Proceedings of the Academy of Natural Sciences of Philadelphia, Philadelphia - PA, vol. 79, p. 299-303.
Vaughan T.W. (1928).- Species of large arenaceous and orbitoidal Foraminifera from the Tertiary deposits of Jamaica.- Journal of Paleontology, Tulsa - OK, vol. 1, no. 4, p. 277-298.
Vaughan T.W. (1929).- Studies of orbitoidal Foraminifera: The subgenus Polylepidina of Lepidocyclina and Orbitocyclina, a new genus.- Proceedings of the National Academy of Sciences, Washington - DC, vol. 15, no. 3, p. 288-295. DOI: 10.1073/pnas.15.3.288
Vaughan T.W. (1933).- Studies of American species of Foraminifera of the genus Lepidocyclina.- Smithsonian Miscellaneous Collections, Washington - DC, vol. 89, no. 10, p. 1-53 (32 Pls.). URL: https://repository.si.edu/handle/10088/26127
Vaughan T.W. (1936).- Helicolepidina nortoni, a new species of Foraminifera from a deep well in St. Landry Parish, Louisiana.- Journal of Paleontology, Tulsa - OK, vol. 10, no. 4, p. 248-252 (Pls. 39-40).
Vaughan T.W. (1945).- Part 1 - American Paleocene and Eocene larger Foraminifera.- Geological Society of America, Memoir, Boulder - CO, vol. 9, 175 p.
Vaughan T.W. & Cole W.S. (1938).- Triplalepidina veracruziana, a new genus and species of orbitoidal Foraminifera from the Eocene of Mexico.- Journal of Paleontology, Tulsa - OK, vol. 12, no. 2, p. 167-169.
Vaughan T.W. & Cole W.S. (1941).- Preliminary Report on the Cretaceous and Tertiary Larger Foraminifera of Trinidad British West Indies.- Geological Society of America Special Papers, Boulder - CO, 137 p.
Versey H.R. (1957).- The Ipswich Limestone of Jamaica and its structural significance.- Congreso Geologico Internacional XX Sesión - Ciudad de México, 1956, Seccion V - Relaciones entre la Tectónica y la Sedimentación (Segundo Tomo), Mexico - D.F., p. 519-524.
Vlerk I.M. van der (1928).- Het genus Lepidocyclina in het Indo-Pacifische gebied.- Wetenschappelijke mededeelingen van de Dienst van den Mijnbouw in Nederlandsch-Oost-Indië, vol. 8, p. 7-88.
Vlerk I.M. van der (1959).- Modification de l'ontogénčse pendant l'évolution des Lépidocyclines.- Bulletin de la Societé Géologique de France (7e Série), Paris, vol. I, no. 7, p. 669-673.
Wade B.S., Pearson P.N., Berggren W.A. & Pälike H. (2011).- Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale.- Earth-Science Reviews, vol. 104, p. 111-142.
Westercamp D. & Andreieff P. (1983).- Saint-Barthélemy et ses îlets, Antilles françaises : Stratigraphie et évolution magmato-structurale.- Bulletin de la Société Géologique de France (7e Série), Paris, vol. XXV, no. 6, p. 873-883.
Wise S.W. Jr & Constans R.E. (1976).- Mid-Eocene planktonic correlations: Northern Italy-Jamaica, W.I.- Gulf Coast Association of Geological Societies, Transactions, Shreveport - LA, vol. 26, p. 144-155.
Woodring W.P. (1957).- Geology and paleontology of Canal Zone and adjoining parts of Panama: Geology and description of Tertiary mollusks (gastropods: Trochidae to Turritellidae).- Geological Survey Professional Paper, Washington - DC, vol. 306-C, 145 p. URL: https://ufdc.ufl.edu/AA00022346/00001
Wright R.M. (Ed., 1974).- Field guide to selected Jamaican geological localities.- Ministry of Mining and Natural Resources, Mines and Geology Division, Kingston, Jamaica, Special Publication no. 1, 57 p.
Wright R.M. & Switzer P. (1971).- Numerical classification applied to certain Jamaican Eocene nummulitids.- Journal of the International Association for Mathematical Geology, Washington - DC, vol. 3, p. 297-311.
Yu Zhoufei, Li Yanli & Li Tiegang (2016).- Mean proloculus size as a salinity index in benthic Foraminifera Ammonia aomoriensis: based on culture and seasonal studies.- Journal of the Palaeontological Society of India, Lucknow, vol. 61, no. 2, p. 215-223. URL: http://palaeontologicalsociety.in/2016/2016_61_2/4.%20China_Ms._To_Review-R1-submit.pdf
Zans V.A., Chubb L.J., Versey H.R., Williams J.B., Robinson E. & Cooke D.L. (1963).- Synopsis of the geology of Jamaica an explanation of the 1958 provisional geological map of Jamaica.- Geological Survey Department, Bulletin, Kingston, Jamaica, no. 4, p. 1-72 [Dated on front cover and title page 1962, Printers imprint at bottom of front page is 1963].
Two subsidiary files are available showing the distribution of calcareous nannofossils in selected samples
analysed for this project. The first file (Appendix
A1 ![]() ) shows analyses of samples from central and eastern Jamaica. The second file
(Appendix A2
) shows analyses of samples from central and eastern Jamaica. The second file
(Appendix A2 ![]() ) shows analyses of samples from western Jamaica.
) shows analyses of samples from western Jamaica.