◄ Carnets Geol. 23 (3) ►
![]()
Outline:
[1. Introduction]
[2. Geographic and geological setting]
[3. Materials and methods]
[4. Results]
[5. Discussion]
[6. Conclusion]
[Bibliographic references]
and ...
[Plate]
Laboratory of Geosciences and Applications, Hassan II University, Faculty of
Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of
Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Scientific Institute, University Mohammed V in Rabat, Avenue Ibn Batouta, B.P. 703, 10106 Rabat-Agdal (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Laboratory of Geosciences and Applications, Hassan II University, Faculty of Sciences Ben M'sick Casablanca, B.P. 7955, Sidi Othmane Casablanca (Morocco)
Published online in final form (pdf) on April 1, 2023
DOI 10.2110/carnets.2023.2303
![]()
[Editor: Bruno R.C. Granier; language editor: Stephen Eagar]
![]()
During the Late Miocene, the Rifian Corridor was one of the two main corridors allowing exchanges between the Atlantic and the Mediterranean Seas. Sedimentary records accumulated along this corridor and specifically, in the Sais Basin, constitute case-study material for understanding the impact of the South Rifian Corridor on the evolution of the fauna, phytoplankton and environment during the Late Miocene. In order to reconstruct the evolution of marine paleoenvironments in the Moulay Yakoub region and to compare this evolution with other areas of the Sais basin, we conducted an integrated biostratigraphic and palynological study of the Upper Miocene of borehole KA 102. High-resolution biostratigraphy based on planktonic foraminifera revised and defined the chronostratigraphic framework of the Upper Miocene sedimentary series of the Moulay Yakoub region providing an age of 7.8 Ma to 6.52 Ma (Late Tortonian to Early Messinian). In the Late Tortonian, the planktonic foraminiferal assemblages suggest an open and relatively deep marine environment. At the Tortonian/Messinian boundary, planktonic foraminiferal and dinoflagellate associations, as well as distal indices, suggest a relatively deep and neritic distal marine environment with a decrease in diversity and an increase in reworked taxa. Surface waters are warm with a minor cooling trend observed just below this boundary. In comparison to previously studied sections in the Sais Basin, the Lower Messinian is characterized by a deeper and more distal marine environment. However, proximal trends and warm surface water conditions persisted in all studied areas from the basin during this period.
• Late Miocene;
• Foraminifera;
• palynology;
• paleoenvironment;
• Sais Basin;
• southern Rifian;
• Morocco
Targhi S., Barhoun N., Bachiri Taoufiq N., Achab M., Essamoud R., Bahaj H., Rahmouna J. & Berry N. (2023).- Late Miocene biostratigraphy and paleoenvironment of the Sais Basin (southern Rifian Corridor, Morocco): New insights from the Moulay Yakoub area.- Carnets Geol., Madrid, vol. 23, no. 3, p. 53-75.
Biostratigraphie et paléoenvironnement du Miocčne supérieur du bassin de Sais (Corridor sud-rifain, Maroc) : Nouvelles données sur la région de Moulay Yakoub.- Le Corridor rifain constituait, au Miocčne supérieur, l'un des deux principaux corridors d'échanges entre l'Atlantique et la Méditerranée. Les dépôts sédimentaires accumulés le long de ce corridor et plus particuličrement dans le bassin du Saďs représentent un terrain d'étude permettant de comprendre le rôle du Corridor sud-rifain dans l'évolution des environnements, des faunes et du phytoplancton au Miocčne supérieur. Une étude biostratigraphique et palynologique intégrée du Miocčne supérieur du forage KA 102 a été réalisée afin de reconstituer l'évolution des paléoenvironnements marins dans la région de Moulay Yakoub et de comparer au mieux cette évolution avec d'autres zones du bassin du Saďs. La biostratigraphique ŕ haute résolution ŕ base des foraminifčres planctoniques a permis de réviser et de préciser le cadre chronostratigraphique des séries sédimentaires de Miocčne supérieur de la région de Moulay Yakoub et de leur attribuer un âge allant de 7,8 Ma ŕ 6,52 Ma (du Tortonien supérieur au Messinien inférieur). Au Tortonien supérieur, les associations de foraminifčres planctoniques documentent un environnement marin ouvert et relativement profond. Ŕ la limite Tortonien-Messinien, les associations des foraminifčres planctoniques et de dinoflagellés ainsi que les indices de distalité suggčrent un environnement marin relativement profond et néritique distal avec une baisse de la diversité et une augmentation des remaniements de taxons. Les eaux de surface étaient chaudes avec une légčre tendance au refroidissement observée juste en dessous de cette limite. Par comparaison avec les coupes étudiées précédemment dans le bassin de Saďs, le Messinien inférieur est caractérisé par un environnement marin plus profond et plus distal. Cependant, les tendances proximales et les conditions d'eaux de surface chaudes ont persisté dans toutes les secteurs étudiés dans ce bassin au cours de cette période.
• Miocčne supérieur ;
• foraminifčres ;
• palynologie ;
• paléoenvironnement ;
• bassin de Saďs ;
• Corridor sud-rifain;
• Maroc
Marine gateways have had a critical impact on climate and oceans throughout geological history. However, water circulation across oceans, marine environment, faunal, floral and phycological evolution, and climatic systems have been strongly influenced by such gate closures and openings (Flecker et al., 2015; Berg, 2016). During the Late Miocene, the Mediterranean Sea was linked to the Atlantic Ocean by several marine gateways lying respectively in the north of Morocco, along the Rifian range, and in the south of Spain, along the Betic chain (Benson et al., 1991; Krijgsman et al., 1999a; Martín et al., 2001, 2009, 2014; Do couto et al., 2014; Flecker et al., 2015; Achalhi et al., 2016; Tulbure et al., 2017; Capella et al., 2018b; Berg et al., 2018). Combination of tectonic phenomena, mostly caused by the convergence between African and European plates, together with sea level and climatic variations, has gradually led to the restriction and closure of these corridors (Krijgsman et al., 1999a; Gutscher et al., 2002; Duggen et al., 2003; Hilgen et al., 2007; Garcia-Castellanos & Villaseńor, 2011; Laan et al., 2012; Manzi et al., 2013; Suc & Frizon de Lamotte, 2019; Kontakiotis et al., 2019). After the closure of the Betic Corridor (Martín et al., 2001, 2014), the Rifian Corridor has been hypothesized to be the last existing marine link between the Atlantic Ocean and the Mediterranean Sea before the Messinian salinity crisis (Martín et al., 2001; Flecker et al., 2015). This event generated the deposition of a considerable mass of primary lower gypsum (5.97-5.6 Ma) in the Mediterranean marginal basins, followed by thick halite series (5.6-5.55 Ma) in deep basins (Roveri et al., 2014). The south Rifian Corridor basins have been covered with detrital inputs from the backland, i.e., the Rifian domain in the North and the Meseto-Atlasic domain in the South (Michard, 1976; Cirac, 1987). Among them, one might consider the Sais basin, mainly characterized by its geographical location in the west of the Taza sill, the closure point of the southern Rifian Corridor at the end of the Messinian. This basin, more particularly the Moulay Yakoub area in this work, stands at the edge of several structural domains, namely the pre-Rifian ridges and the Mesoto-Atlasic domain. Despite its late and relatively weak structuration compared to the whole Rifian chain (Bargach, 2011), the Miocene sedimentation in the Moulay Yakoub area was influenced by the reactivation of the bordering faults of the southern Rifian ridge (e.g., Jbel Zerhoune, Jbel Trhat and Jbel Zalarh). The Sais basin comprises Upper Miocene sediments recording numerous biostratigraphic, environmental and climatic events that occurred just before the Messinian salinity crisis. Due to its strategic location, this area is hence considered as a key site to better understand the evolution of the Southern Rifian Corridor during the Late Miocene.
The majority of the Upper Miocene surface sections within the Sais basin are discontinuous; the successions are poorly exposed and they do not extend over the entire Tortonian-Messinian interval. To trace marine palaeoenvironments in Moulay Yakoub and also to compare trends with other sites from the Sais basin, concerning the narrowing of the southern Rifian Corridor, an integrated biostratigraphic and palynological study of the Upper Miocene sediments from borehole KA 102 was carried out.
To further refine the age of the Moulay Yakoub deposits, the planktonic foraminiferal biostratigraphy requires further evaluation based on biozonal schemes developed for the Mediterranean and adjacent Atlantic areas (Sierro et al., 1993, 2001; Krijgsman et al., 1995, 2000; Sprovieri et al., 1999; Hilgen et al., 2000a, 2000b; Lirer et al., 2019).
The present paper provides, through a high-resolution biostratigraphic study (based on planktonic foraminifers) and a palynological survey (based on dinocysts), a new contribution allowing to reconstruct the evolution of the Tortono-Messinian environments in the Moulay Yakoub region. This area is located between two pre-Rifian ridges that had been potentially affected by tectonic deformation. Therefore, it is considered a crucial area for environmental reconstruction. This will be useful to better understand the narrowing mechanisms of the southern Rifian Corridor at the Tortonian-Messinian transition as well as to explain the depositional conditions of these sediments.
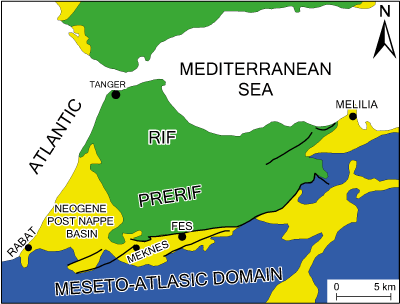
The Moulay Yakoub borehole KA 102 is located in the NW of Fes city on the northern margin of Sais Basin (Fig. 1 ![]() ). It lies in a depression of about 30 km wide on average, stretching from East to West over about 110 km, and extending from the El Kansera sill and the Oued Beht valley in the West to the Touahar pass towards Taza in the East. The Sais plain (which corresponds to the central part of the Southern Rifian Corridor) is located at the southern piedmont of the Prerif. It is limited by the pre-Rifian domain and the South-Rifian ridges (overlapping the Sais plain) to the north, by the Tazekka reliefs and the Taza sill to the East, by the Oued Beht valley and the El Kansra sill to the West, and by the eastern edge of the central
Meseta and the Middle Atlas plateau to the South. Both of these structural domains, clearly distinguished by their deformation pattern, had played a major role in the ante- and post-Neogene tectono-sedimentary evolution of the SRC (Abdelkhalikl,
1997; Sendide, 2000; Zizi,
2000; Winckel, 2002; Charroud et al.,
2007).
). It lies in a depression of about 30 km wide on average, stretching from East to West over about 110 km, and extending from the El Kansera sill and the Oued Beht valley in the West to the Touahar pass towards Taza in the East. The Sais plain (which corresponds to the central part of the Southern Rifian Corridor) is located at the southern piedmont of the Prerif. It is limited by the pre-Rifian domain and the South-Rifian ridges (overlapping the Sais plain) to the north, by the Tazekka reliefs and the Taza sill to the East, by the Oued Beht valley and the El Kansra sill to the West, and by the eastern edge of the central
Meseta and the Middle Atlas plateau to the South. Both of these structural domains, clearly distinguished by their deformation pattern, had played a major role in the ante- and post-Neogene tectono-sedimentary evolution of the SRC (Abdelkhalikl,
1997; Sendide, 2000; Zizi,
2000; Winckel, 2002; Charroud et al.,
2007).
During the Middle Tortonian, the palaeogeography of the Rifian Corridor was strongly controlled by low-intensity tectonic processes (Capella et al., 2018b). The Miocene of the Sais basin lies on a Jurassic basement structured into horsts and grabens limited by a network of NE and NW faults (Charroud et al., 2007). Its northern edge is essentially characterized by tangential tectonics with S and SW tilting, allowing the placement of the Prerif structure (Charroud et al., 2007).
The Sais subsiding basin is interpreted as a result of a distension phase that caused the basin collapse and drove the Tortonian transgression.
During the Late Tortonian, between 8.37 and 7.92 Ma, the pre-Rifian nappes had taken place in the current areas of the Sais and Gharb basins, causing the flexure of the marginal foreland and generating open marine conditions (Krijgsman et al., 1999a; Hilgen et al., 2000a; Dayja et al., 2005; Capella et al., 2017a).
As a result of this distension phase, detrital inputs of Rifian, Mesetian and Atlasian origins were triggered; thus, since 8 Ma, the sedimentation in the Sais basin is characterized by a widespread deposit of "blue marls" with diverse terrigenous interlayers (Wernli, 1988; Krijgsman et al., 1999a; Gelati et al., 2000; Hilgen et al., 2000a; Dayja et al., 2002, 2005; Barhoun & Bachiri Taoufiq 2008; Achalhi et al., 2016; Capella et al., 2017a). Bathymetric estimations in the Sais basin suggest that the maximal depth of these marly sequences was 400-600 m at the end of the Tortonian (Dayja et al., 2002).
A high-intensity tectonic phase was initiated at the Late Tortonian and Early Messinian, during which the inherited faults of the Hercynian basement had been reactivated, causing an uplift in-depth, and therefore restricting the Rifian Corridor to limited depocenters by shallow sills (Morley, 1987; Gomez et al., 2000; Sani et al., 2000, 2007; Capella et al., 2017b). Furthermore, the change in the tectonic process is mainly associated with the relative progression of convergence between Africa and Europe (Morel, 1989; Frizon de Lamotte et al., 1991; Jolivet et al., 2006; Capella et al., 2017b, 2018a).
During the Early Messinian, the northern Rifian Corridor was an emergent land, while water exchanges between the Atlantic Ocean and the Mediterranean Sea occured through the southern Rifian Corridor. The marine deposition had only developed in the deepest depocenters of the southern Rifian Corridor such as in Guercif, Sais and Gharb. Progressive closure of the Taza strait terminated exchanges between the Atlantic Ocean and the Mediterranean Sea. Since then, the Sais basin has mostly been connected with the Atlantic area.
The age of the Rifian Corridor closure is estimated ranging around 7.1-6.9 Ma, and the whole Mediterranean-Atlantic connection is closed and uplifted.
|
|
Figure 1:
A: Geographical and B: geological setting from Sais basin (simplified after the Rifian geological map
1/500,000; Suter, 1980). |
3.1 Materials
The current study relies on cutting samples from the borehole KA 102, collected by the National Office of Hydrocarbons and Mines (ONHYM) during a petroleum exploration mission. This borehole is located in the north of Sais basin (Fig. 1 ![]() ),
some 5 km WSW of the Moulay Yakoub area, its geographical coordinates are
34°04'35.5"N
5°13'55.8"W.
),
some 5 km WSW of the Moulay Yakoub area, its geographical coordinates are
34°04'35.5"N
5°13'55.8"W.
3.2 Lithological description of the studied borehole
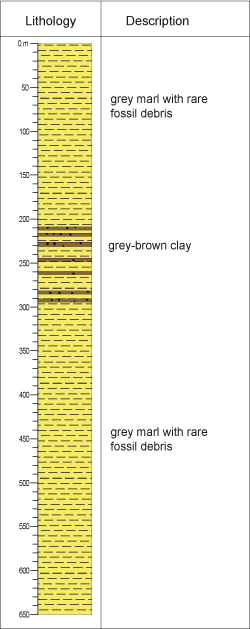
Borehole KA 102 was drilled by ONHYM in 1976. It is mainly designed to recognize the Sais depression formations and the ridges between Jbel Zerhoune and Jbel Traht, and to evaluate the petroleum interest in this structure.
The sediments in the borehole are essentially represented by grey marls with rare fossil debris, intercalated by grey-brown clay levels, slightly carbonated, observed between 199 m and 291 m depth (unpublished report from ONHYM) (Fig. 2 ![]() ).
).
|
|
Figure 2: Lithological description of borehole KA-102. |
3.3 Micropaleontology
90 samples collected for the micropaleontological study, wherein only 64 of the 90 samples processed had a sufficient number of planktonic foraminifera for biostratigraphic analysis. The samples were washed and sieved according to the method used in our laboratory (Geosciences and Applications Laboratory). The qualitative and quantitative analysis are carried out on a portion containing 300 individuals in the 150 μm fraction.
For paleoenvironmental reconstruction purposes, two ecological indices were calculated: diversity index and pelagic index or planktonic-benthic ratio (P/B+P).
P/P+B ratio
This ratio expressed by the calculation of P/((B + P) x 100) is commonly used for paleobathymetric reconstructions (Gibson, 1989) and also provides information on possible upwelling phenomenon that enhances high productivity (Mathieu, 1986, 1988).
Diversity index
Species diversity in planktonic foraminiferal assemblages is established from the Shannon-Weaver function (Shannon & Weaver, 1949). The Shannon diversity index is commonly used to characterize the diversity of species within a population.
Isc= -Σ (qi / Q x Ln qi / Q) with qi = number of individuals counted for each species and Q = Total number of individuals for each level.
Shannon diversity index of a sample provide numerous information about the structure of the taphocoenosis held by the sample and how the individuals are distributed among various species (Daget, 1979; Murray, 1991).
3.4 Palynology
57 samples were treated using standard palynological preparation. 20 g of sediment was decalcified with dilute HCl (20%) and then treated with HF (40%) to remove silicates. After neutralization and rinsing, the samples were filtered through a 10µm screen using an ultrasonic bath to remove fine-silt and clay particles. The residues were concentrated by centrifugation for 10 min at 1,000 rpm and finally diluted in glycerin. A volume of 50 µl was mounted between slide and coverslip.
Among the 57 treated samples, 12 delivered palynomorphs. Most samples are richer in terrestrial palynomorphs (Pinaceae are very dominant while other pollen are poorly represented) compared to marine palynomorphs. In this study, we counted all palynomorphs, so our attention was focused on both pollen and dinoflagellates. Palynomorphs were identified using a Nikon microscope (x1,000 magnification).
At least 160 dinocysts and 80 pollen grains per sample (excluding over-represented taxa such as Pinaceae) were counted. The frequencies of each dinocyst taxon are represented in a detailed diagram. Regarding dinoflagellate cysts, we are mainly interested in taxa with an ecological significance, undetermined cysts are grouped with other dinocysts.
Dinoflagellate cysts (dinocysts) have proven to be excellent tools for estimating temperature variations, salinity and primary productivity changes in surface waters (Wall et al., 1977; Morzadec-Kerfourn, 1979, 1983, 1988; Turon, 1984; Turon & Londeix, 1988; Benzakour, 1992; Harland, 1994; Marret & Turon, 1994; Versteegh, 1994, 1997; Vernal et al., 1994, 1997, 2001; Londeix, 1995, 2007; Zonneveld, 1995, 1999; Warny, 1999; Marret et al., 2001a).
In this work some indices were calculated: the synthetic diagram with Pinaceae which (advantaged by transport) are opposed to halophytes (littoral plants), the D/S ratio, the G/P ratio, the distality index (IN/ON), and the temperature index.
Synthetic diagram with Pinus and indeterminable Pinaceae
Pinaceae (bisaccate pollen grains) are ecologically insignificant because their infra-generic determination is not possible, whereas the genus Pinus is species representative of the entire thermal range. In general, their abundance increases in distal areas and decreases in proximal areas (Heusser & Balsam, 1977; Suc & Drivaliari, 1991). In contrast to the Pinaceae, halophytes are good indicators of littoral environments.
D/S ratio
The curve D/S is used to appreciate the marine influence compared to the continental influence (Warny, 1999). According to following formula: D/S = nD/(nD+nS) where n is the number of taxa counted, S corresponds to pollen and spores, and D to Dinocysts.
G/P ratio
The curve resulting from the ratio between these two families (Gonyaulacoids and Peridinoids) is used to detect the inshore - offshore direction. The ratio is calculated as follows: G/P=nG/(nG+nP) with G = Gonyaulacoids, P = Peridinoids, and n = number of counted taxa.
Distality index (IN/ON)
The IN/ON distality index considers internal neritic platform assemblages (IN) (such as Lingulodinium machaerophorum, Tectatodinium pellitum, Operculodinium israelianum, and Tuberculodinium vancampoae) and external neritic platform assemblages (ON) defined by the sum of Spiniferites spp., using following formula: IN/ON= n IN/(nIN+nON).
Temperature index (W/C ratio)
The ratio of thermophilic dinocyst assemblages (W) to less thermophilic dinocyst assemblages (C) is used to appreciate temperature variations in surface waters. In this work, Spiniferites spp., Operculodinium israelianum, Selenopemphix nephroides, Melitaspheridium coanophorum, Lingulodinium machaerophorum, Polyspheridium zoharyi, Tuberculodinium vancompoae, Tectatodinium pellitum, Impagidinium patulum, Imp. aculeatum, and the Peridiniaceae are considered as markers of warm surface waters. In contrast (Operculodinium centrocarpum, Impagidinium sphearicum, Nematosphaeropsis lemniscata, Nem. labyrinthea, Bitectatodinium tepikiense) are cold surface water indicators.
The W/C ratio represents the number of warm indicator taxa divided by the number of cold indicator taxa, using following formula: W/C = (nW/(nW + nC)). with n = numbers of counted taxa, W = species indicating warm sea surface water temperature, and C = species indicating a cold sea surface water temperature.
4.1. Foraminiferal biostratigraphic and paleoecological results from borehole KA 102
Biostratigraphic analysis considered herein is based on the qualitative and quantitative study of planktonic foraminiferal assemblages. Specific identification is conducted by using specialized references (Sierro et al., 1987, 1993, 1997; Wernli, 1988; Iaccarino, 1985; Barhoun, 1999, 2000; Iaccarino et al., 2007).
In order to establish a precise biostratigraphic framework for the studied sediments, we adopted high-resolution, astronomically calibrated biostratigraphy, recently used within the Mediterranean domain and enabling precise datations and correlations (Krijgsman et al.,
1994, 1995, 1999a;
Hilgen et al., 1995, 2012; Sierro et al.,
2001; Lourens et al., 2004; Lirer et al.,
2019). This biochronology is very similar to the one developed for the adjacent Atlantic domain by Sierro
(1985) and Sierro et al.
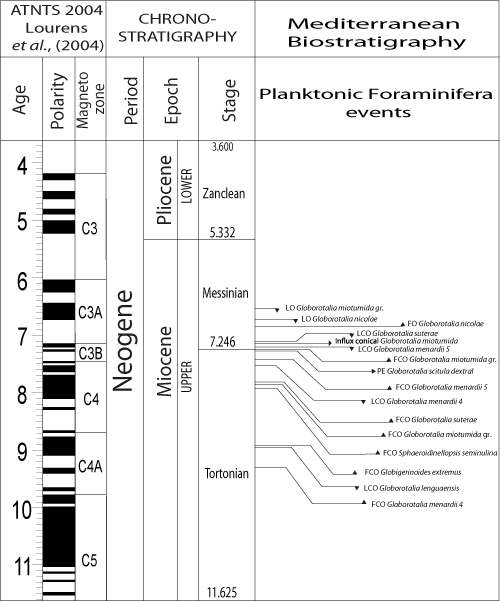
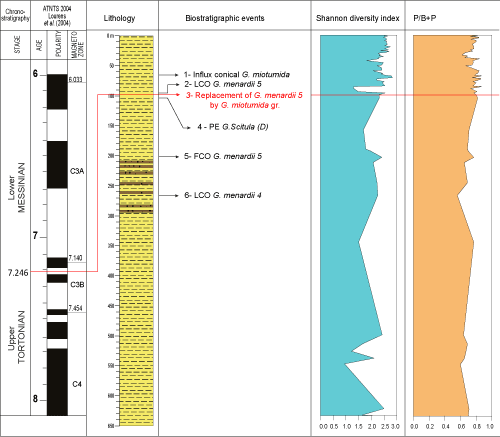
(1993). The succession of the main biostratigraphic events characterizing the Late Miocene is illustrated in Figure 3 ![]() .
.
|
|
Figure 3: Planktonic Foraminiferal events of the Late Miocene. FO= First Occurrence; FCO= First Common Occurrence; LO = Last Occurrence; LCO= Last Common Occurrence, PE : Paracme end. |
The biostratigraphic analysis is based on the qualitative study by specific determination of all specimens and the quantitative one by the whole counting of individuals of each species in a portion containing about 300 specimens from the 150 μm fraction of each washing residues. The taxonomic approach adopted in this work considers, in addition, the vertical distribution of the marker species, the change in coiling of Globorotalia and Neogloboquadrina, the presence and absence of keeled and non-keeled Globorotalia.
4.1.1. Paleoecological results of the studied borehole
The micropaleontological study of the sediments cut-drilled by borehole KA 102 revealed a remarkable richness in both species and individuals, especially in the upper part of the borehole. 26 species from 12 genera were identified.
Between depths 96 m and 604 m, 27 samples were studied, only 17 provided a relatively rich and diversified microfauna. The
Shannon diversity index fluctuates between low diversity levels below 2 (in 7 samples) and normal diversity levels between 2 and 2.4 (in 8 samples) (Fig. 4 ![]() ). The P/B+P ratio varies between 60 and 77%, except for the 101 m depth, where the ratio is around 82%.
). The P/B+P ratio varies between 60 and 77%, except for the 101 m depth, where the ratio is around 82%.
The planktonic foraminiferal assemblages are characterized by a high diversity between 1.5 m and 96 m depth and Shannon diversity index varying between 2 and 2.78. This diversity becomes low between 83 m and 91 m and at level 42 m. The planktonic/benthic ratio varies generally between 73% and 87% in the upper part of the borehole, from 1.5 m to 96 m, except for the samples located between 29 m and 37 m and in the 50 m level where ratio values oscillate around 62% to 69%.
|
|
Figure 4: Ecological parameters recorded in borehole KA-102. |
4.1.2. Biostratigraphic results from borehole KA 102
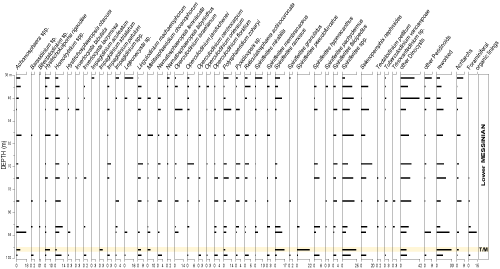
Microfaunal analysis reveals that, from 96 m to 605 m, the planktonic foraminiferal assemblages are dominated by the following species: Globoturborotalita decoraperta, Globot. nepenthes, Globigerinoides obliquus, Globorotalia scitula, Globor. menardii, and Neogloboquadrina acostaensis. Whereas, in the 1.5 m to 96 m depth, the most represented species of planktonic foraminifera are: Globigerinoides bulloides, Globigerina falconensis, Globoturborotalita decoraperta, Globot. nepenthes, Globigerinoides obliquus, Globi. extremus, Globigerinella obesa, Globigerinita uvula, Gl. glutinata, Globorotalia scitula, Globor. miotumida, and Neogloboquadrina acostaensis. We also noticed the sporadic presence of Globorotalia menardii until 62 m. Some reworked forms were observed in some samples.
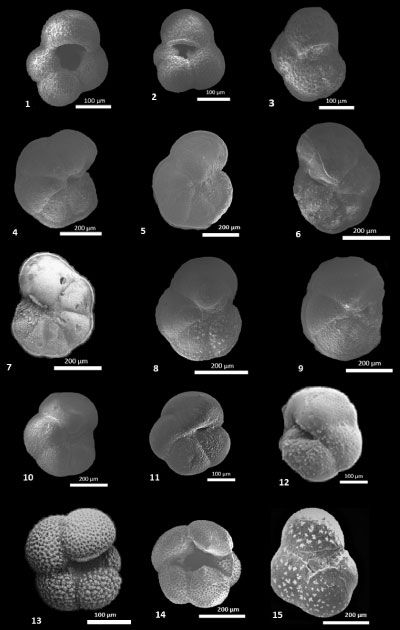
The vertical distribution of planktonic foraminiferal species is indicated in Figure 5 ![]() . The planktonic foraminiferal associations are mainly composed of the following species: Globigerina bulloides Orbigny,
1826 (Pl. 1, fig. 1
. The planktonic foraminiferal associations are mainly composed of the following species: Globigerina bulloides Orbigny,
1826 (Pl. 1, fig. 1 ![]() ), Globi. praebulloides Blow, 1959, Globi. falconensis Blow,
1959 (Pl. 1, fig. 2
), Globi. praebulloides Blow, 1959, Globi. falconensis Blow,
1959 (Pl. 1, fig. 2 ![]() ), Globi. foliata Bolli, 1957, Globoturborotalita apertura (Cushman, 1918), Globot. decoraperta (Takayanagi & Saito, 1962), Globot. nepenthes (Todd,
1957) (Pl. 1, fig. 3
), Globi. foliata Bolli, 1957, Globoturborotalita apertura (Cushman, 1918), Globot. decoraperta (Takayanagi & Saito, 1962), Globot. nepenthes (Todd,
1957) (Pl. 1, fig. 3 ![]() ), Globigerinoides bollii (Blow, 1959), Globi. bulloides Crescenti, 1966, Globi. extremus Bolli & Bermúdez, 1957, Globi. obliquus Bolli, 1957, Globi. quadrilobatus (Orbigny, 1946), Globigerinoides sacculifer (Brady, 1877), Globi. trilobus (Reuss, 1850), Globigerinita glutinata (Egger, 1893), Gl. uvula (Ehrenberg, 1861), Globigerinella obesa (Bolli, 1957), Globorotalia miotumida Jenkins,
1960 (Pl. 1, fig. 9
), Globigerinoides bollii (Blow, 1959), Globi. bulloides Crescenti, 1966, Globi. extremus Bolli & Bermúdez, 1957, Globi. obliquus Bolli, 1957, Globi. quadrilobatus (Orbigny, 1946), Globigerinoides sacculifer (Brady, 1877), Globi. trilobus (Reuss, 1850), Globigerinita glutinata (Egger, 1893), Gl. uvula (Ehrenberg, 1861), Globigerinella obesa (Bolli, 1957), Globorotalia miotumida Jenkins,
1960 (Pl. 1, fig. 9 ![]() ), Globor. menardii (Parker et al.,
1865) (Pl. 1, figs. 4-7
), Globor. menardii (Parker et al.,
1865) (Pl. 1, figs. 4-7 ![]() ), Globor. scitula (Brady,
1882)(Pl. 1, figs. 11-12
), Globor. scitula (Brady,
1882)(Pl. 1, figs. 11-12 ![]() ), Neogloboquadrina acostaensis (Blow,
1959) (Pl. 1, fig. 13
), Neogloboquadrina acostaensis (Blow,
1959) (Pl. 1, fig. 13 ![]() ), Neogloboquadrina humerosa (Takayanagi & Saito, 1962), Dentoglobigerina altispira (Cushman & Jarvis,
1936) (Pl. 1, fig. 14
), Neogloboquadrina humerosa (Takayanagi & Saito, 1962), Dentoglobigerina altispira (Cushman & Jarvis,
1936) (Pl. 1, fig. 14 ![]() ), Orbulina universa Orbigny, 1851, Sphaeroidinellopsis seminulina (Schwager,
1866) (Pl. 1, fig. 15
), Orbulina universa Orbigny, 1851, Sphaeroidinellopsis seminulina (Schwager,
1866) (Pl. 1, fig. 15 ![]() ), and Turborotalita quinqueloba (Natland, 1938).
), and Turborotalita quinqueloba (Natland, 1938).
|
|
Figure 5: Planktonic foraminiferal distribution recorded in borehole KA-102. |
The biostratigraphic division of the studied samples is carried out downhole from top to bottom, mainly based on the last regular presence of marker species. Such a method is applied by 'economic' micropalaeontologists engaged in oil and gas exploration (Granier, 2013). According to the stratigraphic distribution and quantitative analysis of planktonic foraminiferal species, especially the markers Globorotalia menardii, Globor. miotumida, Globor. scitula, and Neogloboquadrina acostaensis, six biostratigraphic events were identified in this borehole (Table 1).
Table 1: Succession of planktonic foraminiferal events in KA-102 borehole.
| Planktonic foraminifer bioevents | Depth (m) | Age (Ma) | References | |
| Early Messinian | 1- influx of conical Globorotalia miotumida group | 66 m | 7.18 - 7.22 Ma | Sierro et al., 2001; Lourens et al., 2004 |
| 2- Last Common Occurrence of Globorotalia menardii 5 | 93 m | 7.23 Ma | Lourens et al., 2004; Hilgen et al., 2000a, 2000b; Lirer et al., 2019 | |
| T/M boundary | 3- replacement of Globorotalia menardii 5 by Globorotalia miotumida group | 96 m | 7.24 Ma | Sierro, 1985; Sierro et al., 1993, 2001; Lourens et al., 2004; Hilgen et al., 2000a, 2000b; Lirer et al., 2019 |
| Upper Tortonian | 4- S/D coiling change of Globorotalia scitula group | 101 m | 7.28 Ma | Sierro, 1985; Sierro et al., 1993; Krijgsman et al., 1995; Hilgen et al., 1995, 2000a, 2000b; Lourens et al., 2004; Lirer et al., 2019 |
| 5- First Common Occurrence (FCO) of Globorotalia menardii 5 | 195 m | 7.36 Ma | Sierro, 1985; Sierro et al., 1993; Krijgsman et al., 1995; Hilgen et al., 1995, 2000a, 2000b; Lourens et al., 2004; Lirer et al., 2019 | |
| 6- Last Common Occurrence (LCO) of Globorotalia menardii 4 | 235 m | 7.51 Ma | Sierro, 1985; Sierro et al., 1993; Hilgen et al., 1995, 2000a, 2000b; Lourens et al., 2004; Lirer et al., 2019 |
The upper part of the borehole is characterized by the presence and occasionally the abundance of Neogloboquadrina acostaensis group with sinistral coiling. We have carefully looked at non-keeled Globorotalia to find Globor. nicolae species; however, no typical form of this species has been found. Furthermore, the presence of the Globor. miotumida group was noted from the top of the borehole
(1.5 m) and persisted until 93 m. The quantitative analysis of the Globor. miotumida group, including Globor. conomiozea (Pl. 1, fig.
8 ![]() ), reveals an influx in this group (influx of conical Globor. miotumida) at 66 m depth. This bioevent (event 1) has been identified by Sierro et al.
(2001) and Lourens et al.
(2004) with an average age comprised between 7.18 Ma and 7.20 Ma.
), reveals an influx in this group (influx of conical Globor. miotumida) at 66 m depth. This bioevent (event 1) has been identified by Sierro et al.
(2001) and Lourens et al.
(2004) with an average age comprised between 7.18 Ma and 7.20 Ma.
A further relevant criterion applied in our biochronological framework concerns the evolution of the percentage of the coiling direction in Globor. menardii. Thus, the latest common occurrence of Globor. menardii form 5 was recorded at 93 m depth in the studied borehole. LCO Globor. menardii form 5 corresponds to "event 2", dated to 7.23 Ma, and was identified by Lourens et al. (2004), Hilgen et al. (2000a, 2000b), and Lirer et al. (2019).
In our samples, the replacement of Globorotalia menardii 5 (dextral) by Globor. miotumida group was reported at 96 m. This event (event 3), which was identified by Sierro (1985) and Sierro et al. (1993), has been proposed to locate the Tortonian/Messinian boundary.
Concerning non-keeled Globorotalia, we controlled the percentage evolution of sinistral and dextral forms of the Globor. scitula group in all studied samples. Hence the coiling changes from sinister to dexter of Globorotalia group (S/D coiling change of Globor. scitula group) are recorded at 101 m. This bioevent (event 4), also known as the top paracme of dextral Globor. scitula, was previously reported by Hilgen et al. (2000a, 2000b), Lourens et al. (2004), and Lirer et al. (2019). By chrono-correlation the age is estimated at around 7.28 Ma.
We diligently looked for the first occurrence of dextral Globor. menardii (form 5), its positioning in the borehole was done with much precaution. In our studied samples, we noted that the first common occurrence of Globor. menardii form 5 (" dextral coiling ") occurred at about 195 m. The FCO Globor. menardii form 5 (event 5) was been recorded at 7.35 Ma in the Atlantic and Mediterranean domains (Sierro, 1985; Sierro et al., 1993; Hilgen et al., 1995, 2000a, 2000b; Krijgsman et al., 1995, 1999a, 1999b; Lourens et al., 2004; Lirer et al., 2019).
In the sediments at the bottom of the borehole, the percentage trends reveal the presence and the abundance of sinistral forms of Globor. menardii (form 4) until their decrease and eventual disappearance. The LCO of Globor. menardii 4 (event 6), with a postulated age of 7.51 Ma, was observed in the adjacent Atlantic (Sierro, 1985; Sierro et al., 1993; Hilgen et al., 2000a, 2000b; Lourens et al., 2004) and in the Mediterranean basin (Hilgen et al., 1995; Krijgsman et al., 1995, 1999a, 1999b; Lourens et al., 2004; Lirer et al., 2019).
4.2. Datation of borehole KA 102
The main objective of this research is to refine the chronostratigraphic framework of the studied sedimentary series and, additionally, to contribute to a more rigorous palaeoenvironmental and palaeogeographic interpretation of the Moulay Yakoub area (northern part of the Sais Basin) during the Late Miocene.
The high-resolution biostratigraphy adopted in this work allowed us to highlight the succession of 6 biostratigraphic events (Table 1); furthermore, we could pinpoint the Tortonian/Messinian boundary within the analyzed borehole, recognize the Upper Tortonian and Messinian, and precisely date the investigated strata.
The succession of two biostratigraphical events characterizes the top of the borehole from 1.5 m to 96 m: the influx conical Globor. miotumida (7.18 Ma to 7.20 Ma) and LCO Globor. menardii form 5 (7.23 Ma). We also noticed that the Globorotalia miotumida group occurs from the top of the borehole (at 1.5 m depth). These deposits are attributed to the Lower Messinian, correlated age estimated between 7.24 Ma and 6.52 Ma.
The Tortonian/Messinian boundary in the studied borehole from loose cuttings is estimated at 96 m. It corresponds to the replacement of the Globor. menardii group (form 5) by the Globorotalia miotumida group. Recently this bioevent has been used as a datum to localize this boundary in the Rifian domain, Morocco (Tulbure et al., 2017; Capella et al., 2018a, 2018b). Indeed, the Tortonian/Messinian boundary GSSP is positioned at the bottom of reddish layer in cycle no.15 and has an astronomical age of 7.24 Ma. It coincides with the FCO of the Globor. miotumida group and lies in the middle of Chron C3br.1r on the geomagnetic polarity time scale of Hilgen et al. (2000a, 2000b), this latter event is widely used to identify the Tortonian/Messinian boundary in the Mediterranean and adjacent North Atlantic (Hilgen et al., 2000a, 2000b; Sierro et al., 2001; Lourens et al., 2004; Lirer et al., 2019).
Moreover, the sediments between 96 m and 602 m present the succession of three bioevents: S/D coiling change of Globor. scitula group (7.28 Ma), FCO of Globor. menardii 5 (7.36 Ma), and LCO of Globor. menardii 4 (7.51 Ma). This material is attributed to the Upper Tortonian, with an estimated age between 7.24 Ma and 7.8 Ma.
This datation improved the chronostratigraphic framework of borehole KA 102, which was attributed to the Miocene in the unpublished ONHYM report. It also allowed providing biostratigraphic details on the Upper Miocene of the Moulay Yakoub region, where previous works mentioned the presence of Tortonian, Lower and Upper Messinian (Lakhdar et al., 2006; Wernli, 1988). Furthermore, these biostratigraphic results support recent work conducted in the Sais basin (Capella et al., 2018a, 2018b).
Although the sampling is not fine enough to follow the details of variation in the sedimentation rate, we tried to calculate this rate by averaging time intervals between two successive events. During the Late Tortonian, the sedimentation rate is in the range of 266 m to 1,175 m/MA, whereas in the Messinian it is about 300m/Ma.
4.3. Palynological results
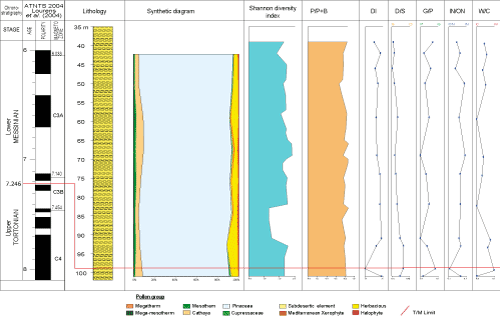
Samples from borehole KA 102 contained a phytoplankton rich in dinocysts and especially Pollen of Pinaceae other pollen grains are present to rare. Dinocyst assemblages are moderately diverse, 40 taxa are found in the sediments of this borehole, the dinocyst diversity is in the range of 48 to 68% (Figs. 6 ![]() - 7
- 7 ![]() ) except for the 99m level that shows low diversity (24%).
) except for the 99m level that shows low diversity (24%).
The dinocyst assemblage (Fig. 6 ![]() ) is dominated by species of the genus Spiniferites (22.83 to 61.33%), Selenopemphix nephroides (0 to 20%) Polysphaeridium zoharyi (1.44 to 13.69%), Operculodinium israelianum (0 to 13.69%), Homotryblium spp. (2.67 to 13.64%), Lingulodinium machaerophorum (1.to 9.45%), and Hystrichokolpoma rigaudiae (1 to 9.46%). Other taxa such as Impagidinium, Nematosphaeropsis spp., Pyxidinopsis, Tectatodinium pellitum, Tuberculodinium vancampoae, do not exceed 4%. The frequency of reworked taxa is important and varies between 8 and 33%, it is dominant at the 93 m and 99 m levels (varying from 28.61% to 32.61 %).
) is dominated by species of the genus Spiniferites (22.83 to 61.33%), Selenopemphix nephroides (0 to 20%) Polysphaeridium zoharyi (1.44 to 13.69%), Operculodinium israelianum (0 to 13.69%), Homotryblium spp. (2.67 to 13.64%), Lingulodinium machaerophorum (1.to 9.45%), and Hystrichokolpoma rigaudiae (1 to 9.46%). Other taxa such as Impagidinium, Nematosphaeropsis spp., Pyxidinopsis, Tectatodinium pellitum, Tuberculodinium vancampoae, do not exceed 4%. The frequency of reworked taxa is important and varies between 8 and 33%, it is dominant at the 93 m and 99 m levels (varying from 28.61% to 32.61 %).
Acritarchs are generally poorly represented (0-8%). foraminiferal linings are present to rare in most of the borehole samples, their percentage increases at the 93 m level.
The D/S ratio values are low and fluctuated between 0.12 and 0.36. The dominant pollens are Pinaceae. Other pollen grains and spores are less frequent. We note that in samples 39 m, 46 m, 93 m, and 99 m, pollen grains (except Pinaceae) are rare and dinocysts are present.
The IN/ON distality index varies between 0.1 and 0.6. It is at the 74 m, 69 m, and 50 m levels that proximal neritic dinocysts are noted in equal or more quantity than distal neritic dinocysts.
The G/P curve shows high ratios in the order of 0.9 and reflects the predominance of Gonyaulacoids in comparison to Peridinoids.
The temperature index of the borehole KA 102 varies between 0.92 and 1 and indicates the predominance of warm sea surface waters dinocyst taxa. A minor cooling trend is observed at the 101 m level due to the increase of cold marker taxa percentage (Operculodinium centrocarpum, Impagidinium sp., Nematosphaeropsis spp.)
The synthetic diagram with Pinus + indeterminable Pinaceae shows a high abundance of Pinaceae reaching a frequency of 90%. The percentage of other pollen grains varies between 10 and 18 % and at some levels, it is low (Fig. 7 ![]() ).
).
Interpretation: The dinocyst assemblages from borehole KA 102 (Fig. 6 ![]() ) from the
Tortonian-Messinian transition to the Early Messinian are characterized by the diversity of neritic taxa such as Lingulodinium machaerophorum, Tectatodinium pellitum, Polysphaeridium zoharyi, Operculodinium israelianum, Tuberculodinium vancampoae, Homotryblium spp., Spiniferites spp. (Percentage of this genus varies between 22.83% and 61.33%). The abundance of neritic taxa and the rarity of oceanic taxa document a neritic environment.
) from the
Tortonian-Messinian transition to the Early Messinian are characterized by the diversity of neritic taxa such as Lingulodinium machaerophorum, Tectatodinium pellitum, Polysphaeridium zoharyi, Operculodinium israelianum, Tuberculodinium vancampoae, Homotryblium spp., Spiniferites spp. (Percentage of this genus varies between 22.83% and 61.33%). The abundance of neritic taxa and the rarity of oceanic taxa document a neritic environment.
The D/S curve shows low ratios ranging from 0.12 to 0.36, reflecting the terrestrial palynomorph concentrations (especially Pinaceae) are higher than that of dinocyst ones and that the sedimentation environment was epicontinental from the Tortonian-Messinian transition to the Early Messinian.
The high G/P ratios reveal that Peridiniacae are in a low proportion and support the existence of an open marine environment.
The dominance of distal neritic taxa in the borehole KA 102 and the rarity of oceanic taxa implies the existence of a distal neritic environment. This is justified by a low IN/ON distality index in most samples, except at levels 50 m, 69 m, and 74 m where proximal taxa are equal to or slightly dominant over distal ones. This implies that in the Early Messinian all sediments were deposited in an outer platform environment with trends towards an inner platform environment.
In the synthetic diagram with Pinus + indeterminate Pinaceae of borehole KA 102, Pinaceae (which generally increase in distal areas) are very abundant while halophytes are poorly represented. The Pinaceae pollen richness in these sediments indicates a distal depositional environment.
The temperature index is high; this shows the predominance of warm sea surface taxa and consequently warm surface water conditions.
To conclude, all of the KA 102 indices cited above show a similarity pointing to the existence of an outer platform neritic environment with trends to inner platform and warm surface waters in the Early Messinian.
|
|
Figure 6: Detailed dinocyst diagram of borehole KA-102. |
The main purpose of this work is to reconstruct paleoenvironmental evolution tendencies in the Moulay Yakoub region from the Late Tortonian to the Early Messinian. This reconstruction is based mainly on the analysis of planktonic foraminiferal associations, dinocyst assemblages, and the comparison of these data to previous works conducted in Sais basin. The paleoenvironment evolution will be presented from the Late Tortonian to Early Messinian (Fig. 7 ![]() ).
).
|
|
Figure 7: Quantitative results for paleoenvironmental and paleoclimatic reconstructions across
Tortonian/Messinian boundary in borehole KA-102. |
Sediments between 96 m and 602 m are assigned to the upper Tortonian with estimated ages between 7.8 Ma and 7.24 Ma. These deposits are characterized by the succession of three bioevents: S/D coiling change of Globorotalia scitula group (7.28 Ma), FCO Globor. menardii 5 (7.35 Ma) and LCO Globor. menardii 4 (7.51 Ma). The calculated sedimentation rate is 267 to 1,175 m /Ma (the highest rate is between 7.36 Ma and 7.28 Ma). From 96 m to 602 m P/B+P ratio varies between 60 and 77% (except for the 101 m level where the ratio reaches 82%), showing the dominance of planktonic foraminifers in comparison to benthic foraminifers. The diversity index is normal to low at some levels. In the sample 101 m, Globigerina bulloides and Neogloboquadrina are dominant, with rare Globigerinoides. This group of species is characteristic of cold waters (Be, 1977; Hemleben et al., 1989; Pujol & Grazzini, 1995).
These two parameters indicate a relatively deep open marine to oceanic environment. Lower diversity levels, presence of reworked forms, and variation in sedimentation rate along the borehole are probably due to tectonic activities. The study of the Moulay Yakoub outcrop by Wernli (SCP/ERICO report, 1991) and revised by Capella et al. (2018a, 2018b) suggests a depth of 300-500 m indicating a suprabathyal environment (Capella et al., 2018a, 2018b).
The same environmental conditions (~150-400 m depth) are observed at Douyet, central Sais basin, and Jenanat, southern edge of Sais basin (Dayja, 2002).
It can be concluded that in the Late Tortonian, marine conditions were open with estimates of 300-500 m in depth.
Samples processed between 114 m and 682 m attributed to the Upper Tortonian did not yield palynomorphs, the organic matter is poorly preserved and therefore was not considered in this study.
Previous work on Moulay Yakoub outcrop, Douyet (northern margin of the Sais Basin in the vicinity of borehole KA 102), and Ain Lorma boreholes (western part of the Sais basin), show normal foraminiferal species diversity and high P/B+P ratios similar to those of KA 102. However, in this latter there are some levels with low diversity. Sedimentation rate between 7.28 Ma and 7.35 of the same sections (Douyet 4,000 m/Ma, Barhoun & Bachiri Taoufiq, 2008) or ~3,700 m/Ma (Dayja et al., 2005) Ain Lorma 2,014 m/Ma (Targhi et al., 2021) is variable. These high sedimentation rates in these different sections could reflect high sediment inputs due to tectonic uplift in the Late Tortonian (Capella et al., 2018a, 2018b). However, on the southeastern margin of the Sais Basin, sedimentation conditions are very different in the Jenanat section, with quite low rates (300 to 700 m/Ma) compared to other sections. This area was protected from sedimentary inputs (Dayja et al., 2005). Between 7.28 and 7.35 Ma and throughout the Sais basin, the sedimentation rate is variable in relation to the position of the studied area which is exposed or quite protected from sediment inputs according to the basin topography.
The Tortonian/Messinian boundary is positioned at level 96 m. It corresponds to the replacement of Globorotalia menardii 5 by Globor. miotumida group and a correlated age of 7.24 Ma. This limit is characterized by the regression of the diversity index and the P/B+P ratio reaches 70% reflecting an open and relatively deep marine environment. The D/S ratio is low, indicating an epicontinental setting. At level 99 m, dinocyst diversity decreases. The high G/P ratio indicates the opening of the deposit environment.
From 93 m to 101 m IN/ON distality index oscillates between 0.4 and 0.1 and shows that distal dinocysts are superior or approximately equal to proximal ones. The abundance of Pinaceae and the rarity of halophytes confirm that the depositional environment of borehole KA 102 is distal. Concerning the temperature index, the W/C ratio curve shows values higher than 0.9, which means that surface water was warm (Fig. 7 ![]() ). We noticed at level 101 m a slight cooling detected by increase of cold indicator.
). We noticed at level 101 m a slight cooling detected by increase of cold indicator.
Comparison with data from Douyet (Barhoun & Bachiri Taoufiq, 2008) and Ain Lorma (Targhi et al., 2021) shows that in KA 102 site the environment is maintained more or less distal and relatively deep, while the tendencies at Douyet and Ain Lorma are proximal at the Tortono-Messinian transition. Species diversity decreased at all three sites, while the reworked forms increase at the levels 93 m and 99 m of borehole KA 102 implying increased tectonic activity. This tectonic stress mentioned in previous work (Capella et al., 2017b, 2018a, 2018b, 2019a, 2019b), that began at the end of the Late Tortonian and continued at the T/M boundary.
A succession of 2 biostratigraphic events: the influx conical Globorotalia miotumida (7.18 Ma to 7.20 Ma), and LCO Globorotalia menardii form 5 (7.23 Ma), characterizes the upper part of the borehole from 1.5 m to 96 m. The estimated age for these deposit ranges between 6.52 Ma and 7.24 Ma and they are attributed to the Early Messinian. The sedimentation rate is around 300 m/Ma and significantly lower than the Late Tortonian. The higher planktono-benthic ratio and high to medium diversity indicate that the Early Messinian environment was relatively deep and open. This is in agreement with the palynological data, which also points towards a distal neritic environment with proximal tendencies. So, the D/S ratio is always low where terrestrial inputs (which are dominated by bisaccate pollen grains that are presumably transported by the wind) dominate dinocyst. Gonyaulacoid dinocysts are the most represented and underlines the open nature of the environment. All the levels show the abundance of neritic dinocyst whereas the oceanic ones are not very frequent, announcing consequently that this is a platform. Distal neritic dinocysts dominates the proximal ones in most samples and suggests an external platform with some inclinations towards an internal platform. Comparison with data from the Moulay Yakoub section shows that this was at a ranging of 50 to 100 m in the Early Messinian and thus reflecting a shallowing upward into a middle shelf environment or an intermediate environment straddling the outer and inner platform (Capella et al., 2018a, 2018b). Whereas in borehole KA 102 the environment corresponded on a whole to an outer platform. Douyet borehole indicates a relatively deep epibathyal depositional environment (200 to 400 m) (Bouab 1992; Barhoun, 2000; Dayja, 2002; Barhoun & Bachiri Taoufiq, 2008) and less distal than that of KA 102 where continental inputs transported by rivers are much less important (in KA 102 pollen grains represented mainly by Pinaceae moved usually by the wind) than in Douyet (Barhoun & Bachiri Taoufiq, 2008), Jbel Kaitone (Bachiri Taoufiq et al., 2000), and Ain Lorma (Targhi et al., 2021). This may suggest that the environment of KA 102 is deeper and more distal than other sections in the vicinity (Douyet, Moulay Yakoub section) and south of the Sais basin (Ain Lorma borehole, Jbel Kaitone section) which are characterized by abundant fluvial inputs.
The impact of the restriction of the South Rifian Corridor in the Moulay Yakoub region was recorded mainly around the Tortonian/Messinian boundary, whereas during the Lower Messinian the microfauna suggests a deep and relatively protected environment from environmental disturbances. This study site could represent the deepest area of the Sais basin. Marine sedimentation prevailed at the top of the borehole and the continental marine transition, represented by the transition from carbonate-rich lagoonal to lacustrine sediments, observed in other areas, is not present in borehole KA 102.
This study emphasizes the importance to integrate previous work (sedimentological, biostratigraphical, structural, …) to the current study (based on planktonic foraminifera and dinocysts) in order to understand the evolution of foreland basins and their gateway.
A qualitative and quantitative study of the planktonic foraminiferal associations in the sediments of the borehole KA 102 evidenced a noteworthy richness in species and individuals, particularly in the upper part of the borehole (26 species belonging to 12 genera were identified).
The high-resolution biostratigraphy processed in this work allowed us to distinguish the succession of six biostratigraphic events. Therefore, we could locate precisely the Tortonian/Messinian boundary in the studied borehole, recognize the Late Tortonian and the Messinian and also accurately date the studied sediments. These data improved the chronostratigraphic framework of borehole KA 102, formerly assigned to the Miocene in the ONHYM internal report. It also provided biostratigraphic details on the Upper Miocene of the Moulay Yakoub area, where previous works mentioned the presence of Tortonian, Lower, and Upper Messinian (Wernli, 1988; Lakhdar et al., 2006). On the other hand, these biostratigraphic results validate recent work in the Sais Basin (Capella et al., 2018a, 2018b).
The palynological study of the samples from borehole KA 102 highlights the presence of a phytoplankton rich in dinocysts and a microflora especially rich in Pinaceae, the other pollen grains being poorly represented.
The Late Tortonian environment of the Moulay Yakoub area is characterized by a normal diversity of planktonic foraminifera reflecting a relatively deep marine depositional environment, with a variable sedimentation rate as high as in other sections located in the Sais basin (e.g., Douyet). Depositional conditions were much more perturbed because of the location near the pre-Rifian ridges during the structuring phase.
Overall distality and temperature indexes from borehole KA 102 demonstrated the existence of an outer shelf neritic environment and warm surface waters at the TMT (Tortonian-Messinian Transition). Just below this boundary, a slight cooling is detected in the surface waters. In the Early Messinian, the environment is consistently distal with warm sea surface conditions, trending toward an inner platform.
One of the most important characteristics of the studied sediments in the upper part of borehole KA 102 (TMT and Early Messinian), is the remarkable richness of planktonic foraminifera, dinocysts, and the presence of other pollen grains except Pinaceae (highly represented).
The sedimentation rate and reworking remain high, indicating important tectonic activity. The depositional environment was probably more distal and deeper in comparison with the sections of the Moulay Yakoub, Douyet, Ain Lorma, and Jbel Kaitone areas.
We thank ONHYM for providing borehole samples and necessary technical support. We are deeply grateful to Bruno Granier for his valuable contributions and precious recommendations all along the editorial process. The relevant and constructive remarks of anonymous reviewers have considerably improved the manuscript; they are warmly acknowledged.
Abdelkhaliki L. (1997).- Évolution tectono-sédimentaire des dépôts gravitaires dans le Prérif interne et l'unité du Habt (Rif externe occidental, Maroc) : Mise en place dans les bassins néogčnes d'avant-fosse.- Thčse de Doctorat, Université Mohammed V, Rabat, 308 p.
Achalhi M., Münch P., Cornée J.-J., Azdimousa A., Melinte-Dobrinescu M., Quillévéré F., Drinia H., Fauquette S., Jiménez-Moreno G., Merzeraud G., Moussa A., Ben El Kharim Y. & Feddi N. (2016).- The late Miocene Mediterranean-Atlantic connections through the North Rifian Corridor: New insights from the Boudinar and Arbaa Taourirt basins (northeastern Rif, Morocco).- Palćogeography, Palćoclimatology, Palćoecology, vol. 459, p. 131-152.
Bachiri Taoufiq N. (2000, unpublished).- Les environnements marins et continentaux du corridor rifain au Miocčne supérieur d'aprčs la palynologie.- Thčse d'État es Sciences, Université Hassan II-Mohammedia, Casablanca, 206 p.
Bachiri Taoufiq N., Barhoun N. & Suc J.-P. (2008).- Les environnements continentaux du corridor rifain (Maroc) au Miocčne supérieur d'aprčs la palynologie.- Geodiversitas, Paris, vol. 30, no. 1, p. 41-58.
Bargach K. (2011).- Les déformations plio-quaternaires dans la partie frontale de la chaîne du Rif (Rides prérifaines, et bassin du Saďs) un exemple de la tectonique en coins expulsés.- Thčse de Doctorat, Université Mohammed V, Rabat, no. 2523, 215 p.
Barhoun N. (2000, unpublished).- Biostratigraphie et paléoenvironnement du Miocčne supérieur et du Pliocčne inférieur du Maroc septentrional : Apport des foraminifčres planctoniques.- Thčse d'État es Sciences, Université Hassan II-Mohammedia, Casablanca, 272 p.
Barhoun N. & Bachiri Taoufiq N. (2008).- Événements biostratigraphiques et environnementaux enregistrés dans le corridor sud rifain (Maroc septentrional) au Miocčne supérieur avant la crise de salinité messinienne.- Geodiversitas, Paris, vol. 30, no. 1, p. 21-40.
Barhoun N. & Wernli R. (1999).- Biostratigraphie du Mio-Pliocčne du bassin de Boudinar par les foraminifčres planctoniques (Rif nord-oriental, Maroc).- Revue de Paléobiologie, Genčve, vol. 18, no. 2, p. 491-508.
Bé A.W.H. & Hutson W.H. (1977).- Ecology of planktonic foraminifera and biogeographic patterns of life and fossil assemblages in the Indian Ocean.- Micropaleontology, New York - NY, vol. 23, no. 4, p. 369-414.
Benson R.H., Rakic-El Bied K. & Bonaduce G. (1991).- An important current reversal (influx) in the Rifian Corridor (Morocco) at the Tortonian-Messinian boundary: The end of Tethys Ocean.- Paleoceanography, vol. 6, no. 1, p. 165-192.
Benzakour M. (1992).- Palynologie du Néogčne dans la région du détroit de Sicile, Méditerranée centrale.- Thčse de Doctorat en Sciences de l'Environnement, Université de Québec, Montréal, p. 176
Berg B.C.J. van den, Sierro F.J., Hilgen F.J., Flecker R., Larrasoańa J.C., Krijgsman W., Flores J.A. & Mata M.P. (2018).- Imprint of Messinian Salinity Crisis events on the Spanish Atlantic margin.- Newsletters on Stratigraphy, vol. 51, no. 1, p. 93-115.
Bouab B. (1992).- Les ostracodes du Néogčne supérieur du Maroc nord-oriental : Évolution des paléoenvironnements, paléogéographie.- Thčse de Doctorat, Université Bordeaux I, 278 p.
Capella W., Barhoun N., Flecker R., Hilgen F.J., Kouwenhoven T., Matenco L.C., Sierro F.J., Tulbure M.A., Yousfi M.Z. & Krijgsman W. (2018a).- Data on lithofacies, sedimentology and palaeontology of South Rifian Corridor sections (Morocco).- Data in Brief, vol. 19, p. 712-736.
Capella W., Barhoun N., Flecker R., Hilgen F.J., Kouwenhoven T., Matenco L.C., Sierro F.J., Tulbure M.A., Yousfi M.Z. & Krijgsman W. (2018b).- Palaeogeographic evolution of the late Miocene Rifian Corridor (Morocco): Reconstructions from surface and subsurface data.- Earth-Science Reviews, vol. 180, p. 37-59.
Capella W., Flecker R., Hernández-Molina F.J., Simon D., Meijer P.T., Rogerson M., Sierro F.J. & Krijgsman W. (2019a).- Mediterranean isolation preconditioning the Earth System for late Miocene climate cooling.- Scientific Reports, vol. 9, no. 1, p. 1-8.
Capella W., Hernández-Molina F.J., Flecker R., Hilgen F.J., Hssain M., Kouwenhoven T.J., Oorschot M. van, Sierro F.J., Stow D.A.V., Trabucho-Alexandre J., Tulbure M.A., Weger W. de, Yousfi M.Z. & Krijgsman W. (2017b).- Sandy contourite drift in the late Miocene Rifian Corridor (Morocco): Reconstruction of depositional environments in a foreland-basin seaway.- Sedimentary Geology, vol. 355, p. 31-57.
Capella W., Matenco L., Dmitrieva E., Roest W.M.J., Hessels S., Hssain M., Chakor-Alami A., Sierro F.J. & Krijgsman W. (2017a).- Thick-skinned tectonics closing the Rifian Corridor.- Tectonophysics, vol. 710-711, p. 249-265.
Capella W., Spakman W., Hinsbergen D.J.J. van, Chertova M.V. & Krijgsman W. (2019b).- Mantle resistance against Gibraltar slab dragging as a key cause of the Messinian Salinity Crisis.- Terra Nova, vol. 32, no. 2, p. 141-150.
Charroud M., Cherai B., Benabdelhadi M. & Falgučres C. (2007).- Impact de la néotectonique quaternaire sur la dynamique sédimentaire du Saďs (Maroc) : Du bassin d'avant fosse pliocčne au plateau continental quaternaire.- Quaternaire, Paris, vol. 18, no. 4, p. 327-334.
Cirac P. (1987).- Le bassin sud-rifain occidental au Néogčne supérieur. Évolution de la dynamique sédimentaire et de la paléogéographie au cours d'une phase de comblement.- Thčse de Doctorat d'État, Université Bordeaux 1, no. 837; Mémoires de l'Institut de Géologie du Bassin d'Aquitaine, Bordeaux, no. 21, 271 p.
Daget J. (1979).- Les modéles mathématiques en écologie.- Masson, Paris, 172 p.
Dayja D. (2002).- Les foraminifčres néogčnes, témoins de la chronologie, de la bathymétrie et de l'hydrologie du Corridor Rifain (Maroc septentrional).- Thčse de Doctorat, Université Pierre et Marie Curie (Paris VI), 340 p.
Dayja D., Janin M.-C. & Boutakiout M. (2005).- Biochronolgy and correlation between the Neogene basins of the South Rifian Corridor (Morocco) based on planktonic foraminifer and calcareous nannofossil events.- Revue de Micropaléontologie, Paris, vol. 48, no. 3, p. 141-157.
Do Couto D., Popescu S.-M., Suc J.-P. Melinte-Dobrinescu M.C., Barhoun N., Gorini C., Jolivet L., Poort J., Jouannic G. & Auxietre J.-L. (2014).- Lago Mare and the Messinian Salinity Crisis: Evidence from the Alboran Sea (S. Spain).- Marine and Petroleum Geology, vol. 52, p. 57-76.
Duggen S., Hoernie K., Bogaard P. van den, Rüpke L. & Morgan J.-P. (2003).- Deep roots of the Messinian salinity crisis.- Nature, vol. 422, p. 602-606.
Flecker R., Krijgsman W., Capella W., Castro Martíns C. de, Dmitrieva E., Mayser J.P., Marzocchi A., Modestu S., Ochoa D., Simon D., Tulbure M., Berg B. van den, Schee M. van der, Lange G. de, Ellam R., Govers R., Gutjahr M., Hilgen F., Kouwenhoven T. & Yousfi M.Z. (2015).- Evolution of the Late Miocene Mediterranean-Atlantic gateways and their impact on regional and global environmental change.- Earth-Science Reviews, vol. 150, p. 365-392.
Frizon de Lamotte D., Andrieux J. & Guézou J.-C. (1991).- Cinématique des chevauchements néogčnes dans l'Arc bético-rifain ; discussion sur les modčles géodynamiques.- Bulletin de la Société géologique de France, Paris, t. 162, no. 4, p. 611–626.
Garcia-Castellanos D. & Villaseńor A. (2011).- Messinian salinity crisis regulated by competing tectonics and erosion at the Gibraltar arc.- Nature, vol. 480, article 7377, p. 359-363.
Gelati R., Moratti G. & Papani G. (2000).- The Late Cenozoic sedimentary succession of the Taza-Guercif Basin, South Rifian Corridor, Morocco.- Marine and Petroleum Geology, vol. 17, no. 3, p. 373-390.
Gibson T.G. (1989).- Planktonic benthonic foraminiferal ratios: Modern patterns and Tertiary applicability.- Marine micropaleontology, vol. 15, no. 1-2, p. 29-52.
Gomez F., Barazangi M. & Demnati A. (2000).- Structure and evolution of the Neogene Guercif Basin at the junction of the Middle Atlas Mountains and the Rif Thrust Belt, Morocco.- AAPG Bulletin, Boulder - CO, vol. 84, no. 9, p. 1340-1364.
Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds., 2012).- The Geologic Time Scale 2012.- Cambridge University Press, 1144 p.
Granier B. (2013).- Micropaleontology from the STRATI 2013 perspective.- Cięncias da Terra, Lisboa, vol. 18, p. 89-94. URL: https://cienciasdaterra.novaidfct.pt/index.php/ct-esj/article/view/288/321
Gutscher M.A., Malod J., Rehault J.-P., Contrucci I., Klingelhoefer F., Mendes-Victor L. & Spakman W. (2002).- Evidence for active subduction beneath Gibraltar.- Geology, Boulder - CO, vol. 30, no. 12, p. 1071-1074
Harland R. (1979).- 22. Dinoflagellate biostratigraphy of Neogene and Quaternary sediments at holes 400/400A in the Bay of Biscay (Deep Sea Drilling Project Leg 48). In: Montadert L. et al. (eds.), Leg 48 of the cruises of the Drilling Vessel Glomar Challenger. Brest, France to Aberdeen, Scotland. May-July 1976- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XLVIII, p. 531-546.
Harland R. (1994).- Dinoflagellate cysts from the Glacial/Postglacial transition in the northeast Atlantic Ocean.- Palaeontology, London, vol. 37, no. 2, p. 263-283.
Hemleben C., Spindler M. & Anderson O.R. (1989).- Modern planktonic foraminifera.- Springer-Verlag New York Inc., 363 p.
Heusser L.E. & Balsam W.L. (1977).- Pollen distribution in the Northeast Pacific Ocean.- Quaternary Research, vol. 7, no. 1, p. 45–62.
Hilgen F.J., Bissoli L., Iaccarino S., Krijgsman W., Meijer R., Negri A. & Villa G. (2000a).- Integrated stratigraphy and astrochronology of the Messinian GSSP at Oued Akrech (Atlantic Morocco).- Earth and Planetary Science Letters, vol. 182, no. 3-4, p. 237-251.
Hilgen F.J., Iaccarino S., Krijgsman W., Villa G., Langereis C.G. & Zachariasse W.J. (2000b).- The global boundary stratotype section and point (GSSP) of the Messinian Stage (uppermost Miocene).- Episodes, Beijing, vol. 23, no. 3, p. 172-178.
Hilgen F.J., Krijgsman W., Langereis C.G., Lourens L.J., Santarelli A. & Zachariasse W.J. (1995).- Extending the astronomical (polarity) time scale into the Miocene.- Earth and Planetary Science Letters, vol. 136, no. 3-4, p. 495-510.
Hilgen F.J., Krijgsman W., Raffi I., Turco E. & Zachariasse W.J. (2000c).- Integrated stratigraphy and astronomical calibration of the Serravallian/Tortonian boundary section at Monte Gibliscemi (Sicily, Italy).- Marine Micropaleontology, vol. 38, no. 3-4, p. 181-211.
Hilgen F., Kuiper K., Krijgsman W., Snel E. & Laan E. van der (2007).- Astronomical tuning as the basis for high-resolution chronostratigraphy: The intricate history of the Messinian Salinity Crisis.- Stratigraphy, Flushing - NY, vol. 4, no. 2-3, p. 231-238.
Hilgen F.J., Lourens L.J., Dam J.A. van, Beu A.G., Boyes A.F., Cooper R.A., Krijgsman W., Ogg J.G., Piller W.E. & Wilson D.S. (2012).- Chapter 29. The Neogene Period. In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (eds.), The Geologic Time Scale 2012.- Cambridge University Press, p. 923-978.
Iaccarino S.M. (1989).- Mediterranean Miocene and Pliocene planktic foraminifera. In: Bolli H.M., Saunders J.B. & Perch-Nielsen K. (eds.), Plankton Stratigraphy. 1.- Cambridge University Press, p. 283-314.
Iaccarino S.M., Premoli Silva I., Biolzi M., Foresi L.M., Lirer F., Turco E. & Petrizzo M.R. (2007).- Practical manual of Neogene planktonic foraminifera. In: International School on Planktonic Foraminifera (Neogene Planktonic Foraminifera).- Universitŕ di Perugia Press, 180 p.
Jolivet L., Augier R., Robin C., Suc J.-P. & Rouchy J.-M. (2006).- Lithospheric-scale geodynamic context of the Messinian salinity crisis.- Sedimentary Geology, vol. 188-189, p. 9-33.
Kontakiotis G., Besiou E., Antonarakou A., Zarkogiannis S.D., Kostis A., Mortyn P.G., Moissette P., Cornée J.-J., Schulbert C., Drinia H., Anastasakis G. & Karakitsios V. (2019).- Decoding sea surface and paleoclimate conditions in the eastern Mediterranean over the Tortonian-Messinian transition.- Palćogeography, Palćoclimatology, Palćoecology, vol. 534, article 109312, 16 p.
Krijgsman W., Garcés M., Agustí J., Raffi I., Taberner C. & Zachariasse W.J. (2000).- The "Tortonian salinity crisis" of the eastern Betics (Spain).- Earth and Planetary Science Letters, 181, no. 4, p. 497-511.
Krijgsman W., Hilgen F.J., Langereis C.G. & Zachariasse W.J. (1994).- The age of the Tortonian/Messinian boundary.- Earth and Planetary Science Letters, vol. 121, p. 533-547.
Krijgsman W., Hilgen F.J., Langereis C.G., Santarelli A. & Zachariasse W.J. (1995).- Late Miocene magnetostratigraphy, biostratigraphy and cyclostratigraphy in the Mediterranean.- Earth and Planetary Science Letters, vol. 136, no. 3-4, p. 475-494.
Krijgsman W., Hilgen F.J., Marabini S. & Vai G.B. (1999a).- New paleomagnetic and cyclostratigraphic age constraints on the Messinian of the Northern Apennines (Vena del Gesso Basin, Italy).- Memorie della Societŕ Geologica Italiana, Roma, vol. 54, p. 25-33.
Krijgsman W., Hilgen F.J., Raffi I., Sierro F.J. & Wilson D.S. (1999b).- Chronology, causes and progression of the Messinian salinity crisis.- Nature, vol. 400, article 6745, p. 652-655.
Laan E. van der, Hilgen F.J., Lourens L.J., Kaenel E. de, Gaboardi S. & Iaccarino S. (2012).- Astronomical forcing of Northwest African climate and glacial history during the late Messinian (6.5-5.5 Ma).- Palćogeography, Palćoclimatology, Palćoecology, vol. 313-314, p. 107-126.
Lakhdar A., Ntarmouchant A., Ribeiro M.L., Beqqali M., El Ouadeihe K., Benaabidate L., Dahire M. & Driouch Y. (2006).- Nouvelle approche geologique et geodynamique du complexe hydrothermal de Moulay Yacoub (bordure septentrionale du Sillon Sud Rifain).- Comunicaçőes Geologicas, Lisboa, t. 93, p. 185-204.
Lirer F., Foresi L.M., Iaccarino S.M., Salvatorini G., Turco E., Cosentino C., Sierro F.J. & Caruso A. (2019).- Mediterranean Neogene planktonic foraminifer biozonation and biochronology.- Earth-Science Reviews, vol. 196, article 102869, 36 p.
Londeix L. (1995).- Les kystes de dinoflagellés du Golfe de Hammamet (Tunisie) ; contribution ŕ la reconstitution paléoenvironnementale du Pliocčne méditerranéen.- Comptes Rendus de l'Académie des Sciences (Série 2), Paris, vol. 320, no. 12, p. 1233-1240.
Londeix L., Benzakour M., Suc J.-P. & Turon J.-L. (2007).- Messinian palaeoenvironments and hydrology in Sicily (Italy): The dinoflagellate cyst record.- Geobios, Villeurbanne, vol. 40, no. 3, p. 233-250.
Lourens L., Hilgen F.J. Shackleton N.J., Laskar J. & Wilson D. (2004).- The Neogene period. In: Gradstein F.M., Ogg J.G. & Schmitz M. (eds.), The Geologic Time Scale 2004.- Cambridge University Press, p. 409-440.
Manzi V., Gennari R., Hilgen F., Krijgsman W., Lugli S., Roveri M. & Sierro F.J. (2013).- Age refinement of the Messinian salinity crisis onset in the Mediterranean.- Terra Nova, vol. 25, no. 4, p. 315-322.
Marret F. & Turon J.-L. (1994).- Paleohydrology and paleoclimatology off Northwest Africa during the last glacial-interglacial transition and the Holocene: Palynological evidences.- Marine Geology, vol. 118, no. 1-2, p. 107-117.
Marret F., Vernal A. de, Bendera F. & Harland R. (2001a).- Late Quaternary Sea surface conditions at DSDP Hole 594 in the southwest Pacific Ocean based on dinoflagellate cyst assemblages.- Journal of Quaternary Science, vol. 16, no. 7, p. 739-751.
Marret F., Vernal A. de, Pedersen T.F. de & McDonald D. (2001b).- Middle Pleistocene to Holocene palynostratigraphy of ocean drilling program site 887 in the Gulf of Alaska, northeastern North Pacific.- Canadian Journal of Earth Sciences, Ottawa, vol. 38, no. 3, p. 373-386.
Martín J.M., Braga J.C., Aguirre J. & Puga-Bernabéu A. (2009).- History and evolution of the North-Betic Strait (Prebetic Zone, Betic Cordillera): A narrow, early Tortonian, tidal-dominated, Atlantic-Mediterranean marine passage.- Sedimentary Geology, vol. 216, no. 3-4, p. 80-90.
Martín J.M., Braga J.C. & Betzler C. (2001).- The Messinian Guadalhorce corridor: The last northern, Atlantic-Mediterranean gateway.- Terra Nova, vol. 13, no. 6, p. 418-424.
Martín Martín J.M. Puga Bernabéu Á., Aguirre Rodríguez J. & Braga Alarcón J. (2014).- Miocene Atlantic-Mediterranean seaways in the Betic Cordillera (southern Spain).- Revista de la Sociedad Geológica de Espańa, Madrid, vol. 27, no. 1, p. 175-186.
Mathieu R. (1986).- Sédiments et foraminifčres actuels de la marge continentale atlantique du Maroc.- Thčse de Doctorat d'État čs Sciences naturelle, Université Pierre et Marie Curie (Paris VI); Mémoires des Sciences de la Terre, Paris, no. 86-14, 419 p.
Mathieu R. (1988).- Foraminifčres actuels et résurgences côtičres sur la marge continentale atlantique du Maroc. In: Benthos'86.- Revue de Paléobiologie, Genčve, vol. spécial, no. 2, Partie II, p. 845-850.
Michard A. (1976).- Éléments de géologie marocaine.- Notes et Mémoires du Service géologique du Maroc, Rabat, no. 252, 408 p.
Morel J.-L. (1989).- États de contrainte et cinématique de la chaîne rifaine (Maroc) du Tortonien ŕ l'actuel.- Geodinamica acta, vol. 3, no. 4, p. 283-294.
Morley C.K. (1987).- Origin of a major cross-element zone: Moroccan Rif.- Geology, Boulder - CO, vol. 15, no. 8, p. 761-764.
Morzadec-Kerfourn M.T. (1979).- Les kystes de Dinoflagellés. In: La mer pélagienne. Étude sédimentologique et écologique du plateau tunisien et de ses abords.- Géologie méditerranéenne, Mareille, t. VI, no. 1, p. 221-246. URL: https://www.persee.fr/doc/geolm_0397-2844_1979_num_6_1_1083
Morzadec-Kerfourn M.T. (1983).- Intéręt des kystes de dinoflagellés pour l'établissement de reconstitution paléogéographique : Exemple du Golfe de Gabčs (Tunisie).- Cahiers de Micropaléontologie, année 1983, no. 4, p. 15-22.
Morzadec-Kerfourn M.T. (1988).- Paléoclimats et paléoenvironnements du Tardiglaciaire au Récent, en Méditerranée orientale, ŕ l'est du Delta du Nil : L'apport des microfossiles ŕ membrane organique.- Bulletin du Centre de Recherches Exploration-Production elf-Aquitaine, Pau, Numéro spécial, vol. 12, p. 267-275.
Murray J.W. (1991).- Ecology and palaeoecology of benthic foraminifera.- Longman Scientific and Technical, Harlow, 408 p.
Pujol C. & Grazzini C.V. (1995).- Distribution patterns of live planktic foraminifers as related to regional hydrography and productive systems of the Mediterranean Sea.- Marine Micropaleontology, vol. 15, no. 2-3, p. 187-217.
Roveri M., Flecker R., Krijgsman W., Lofi J., Lugli S., Manzi V., Sierro F.J., Bertini A., Camerlenghi A., Lange G. de, Govers R., Hilgen F.J., Hübscher C., Meijer P.T. & Stoica M. (2014).- The Messinian Salinity Crisis: Past and future of a great challenge for marine sciences.- Marine Geology, vol. 352, p. 25-58.
Sani F., Ventisette C. del, Montanari D., Bendkik A. & Chenakeb M. (2007).- Structural evolution of the Rides Prerifaines (Morocco): Structural and seismic interpretation and analogue modelling experiments.- International Journal of Earth Sciences, vol. 96, p. 685-706.
Sani F., Zizi M. & Bally A.W. (2000).- The Neogene-Quaternary evolution of the Guercif Basin (Morocco) reconstructed from seismic line interpretation.- Marine and Petroleum Geology, vol. 17, no. 3, p. 343-357.
SCP/ERICO report (1991).- Étude de synthčse géologique et géophysique du bassin du Gharb.- ONHYM internal report, Rabat, 533 p.
Sendide O. (2002).- Étude qualitative des eaux de la nappe phréatique du bassin de Fčs-Meknés, caractérisation, évaluation, modélisation mathématique et moyen de protection.- Thčse Doctorat, Université Tunis El Manar, 356 p.
Shannon C.E. & Weaver W. (1949).- The mathematical theory of communication.- University of Illinois Press, Urbana - IL, 144 p.
Sierro F.J. (1985).- The replacement of the "Globorotalia menardii" group by the Globorotalia miotumida group: An aid to recognizing the Tortonian-Messinian boundary in the Mediterranean and adjacent Atlantic.- Marine Micropaleontology, vol. 9, no. 6, p. 525-535.
Sierro F.J., Flores J.A., Civis J. & Gonzalez Delgado J.A. (1987).- New criteria for the correlation of the Andalusian and Messinian stages.- Annales Instituti Geologici Publici Hungarici, Budapest, vol. LXX, p. 355-361.
Sierro F.J., Flores J.A., Civis J., González Delgado J.A. & Francés G. (1993).- Late Miocene globorotaliid event-stratigraphy and biogeography in the NE-Atlantic and Mediterranean.- Marine Micropaleontology, vol. 21, no. 1-3, p. 143-167.
Sierro F.J., Flores J.A., Zamarreno I., Vázquez A., Utrilla R., Francés G., Hilgen F. & Krijgsman W. (1997).- Astronomical cyclicity and sapropels in the pre-evaporitic Messinian of the Sorbas basin (Western Mediterranean).- Geogaceta, Madrid, no. 21, p. 199-202.
Sierro F.J., Hilgen F.J., Krijgsman W. & Flores J.A. (2001).- The Abad composite (SE Spain): A Messinian reference section for the Mediterranean and the APTS.- Palćogeography, Palćoclimatology, Palćoecology, vol. 168, no. 1-2, p. 141-169.
Sprovieri M., Bellanca A., Neri R., Mazzola S., Bonanno A., Patti B. & Sorgente R. (1999).- Astronomical calibration of late Miocene stratigraphic events and analysis of precessionally driven palaeoceanographic changes in the Mediterranean basin.- Memorie della Societŕ Geologica Italiana, Roma, vol. 54, p. 7-24.
Suc J.-P. & Drivaliari A. (1991).- Transport of bisaccate coniferous fossil pollen grains to coastal sediments: An example from the earliest Pliocene Orb Ria (Languedoc, Southern France).- Review of Palaeobotany and Palynology, vol. 70, no. 3, p. 247-253.
Suc J.-P. & Frizon de Lamotte D. (2019).- Paléoenvironnements méditerranéens : Changements et bouleversements environnementaux depuis 23 Ma.- Géochronique, Paris, no. 151, p. 16-17.
Suter G. (1980).- Carte géologique de la chaîne rifaine (1/500 000).- Notes et Mémoires du Service géologique du Maroc, Rabat, no. 245.
Targhi S., Barhoun N., Bachiri Taoufiq N., Achab M., Ait Salem A. & Yousfi M.Z. (2021).- Vegetation climate and marine environmental reconstruction in the western Mediterranean (southern Rifian corridor, Morocco) over the Tortonian-Messinian transition.- Heliyon, vol. 7, no. 12, article e08569, 19 p.
Tulbure M.A., Capella W., Barhoun N., Flores J.A. Hilgen F.J., Krijgsman W., Kouwenhoven T., Sierro F.J. & Yousfi M.Z. (2017).- Age refinement and basin evolution of the North Rifian Corridor (Morocco): No evidence for a marine connection during the Messinian Salinity Crisis.- Palćogeography, Palćoclimatology, Palćoecology, vol. 485, p. 416-432.
Turon J.-L. (1984).- Le palynoplancton dans l'environnement actuel de l'Atlantique nord-oriental. Évolution climatique et hydrologique depuis le dernier maximum glaciaire.- Mémoires de l'Institut géologique du Bassin d'Aquitaine, Talence, no. 17, 313 p.
Turon J.-L. & Londeix L. (1988).- Les assemblages de kystes de Dinoflagelles en Méditerranée occidentale (Mer d'Alboran) : Mise en évidence de l'évolution des paléoenvironnements depuis le dernier maximum glaciaire.- Bulletin des Centres de Recherche Exploration-Production elf-Aquitaine, Pau, vol. 12, no. 1, p. 313-344.
Vernal A. de, Henry M., Matthiessen J., Mudie P.J., Rochon A., Boessenkool K.P., Eynaud F., Grřsfjeld K., Guiot J., Hamel D., Harland R, Head M.J., Kunz-Pirrung M., Levac E., Loucheur V., Peyron O., Pospelova V., Radi T., Turon J.-L. & Voronina E. (2001).- Dinoflagellate cyst assemblages as tracers of sea-surface conditions in the Northern North Atlantic, Arctic and sub-Arctic seas: The new "n = 677" data base and its application for quantitative palaeoceanographic reconstruction.- Journal of Quaternary Science, vol. 16, no. 7, p. 681-698. https://doi.org/10.1002/jqs.659
Vernal A. de, Rochon A., Turon J.-L. & Matthiessen J. (1997).- Organic-walled dinoflagellate cysts: Palynological tracers of sea-surface conditions in middle to high latitude marine environments.- Geobios, Villeurbanne, vol. 30, no. 7, p. 905-920.
Vernal A. de, Turon J.-L. & Guiot J. (1994).- Dinoflagellate cyst distribution in high-latitude marine environments and quantitative reconstruction of sea-surface salinity, temperature, and seasonality.- Canadian Journal of Earth Sciences, Ottawa, vol. 31, no. 1, p. 48-62.
Versteegh G.J.M. (1994).- Recognition of cyclic and non-cyclic environmental changes in the Mediterranean Pliocene: A palynological approach.- Marine Micropaleontology, vol. 23, no. 2, p. 147-183.
Versteegh G.J.M. (1997).- The onset of major Northern Hemisphere glaciations and their impact on dinoflagellate cysts and acritarchs from the Singa section, Calabria (southern Italy) and DSDP Holes 607/607A (North Atlantic).- Marine Micropaleontology, vol. 30, no. 4, p. 319-343.
Wall D., Dale B., Lohmann G.P. & Smith W.K. (1977).- The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the North and South Atlantic oceans and adjacent seas.- Marine Micropaleontology, vol. 2, p. 121-200.
Warny S. (1999).- Mio-Pliocene palynology of the Gibraltar Arc: A new perspective on the Messinian Salinity Crisis.- Thčse de Doctorat, Université Catholique de Louvain, 307 p.
Wernli R. (1988).- Micropaléontologie du Néogčne post-nappes du Maroc septentrional et description systématique des foraminifčres planctoniques.- Notes et Mémoires du Service géologique du Maroc, Rabat, no. 331, 270 p.
Winckel A. (2002).- Essai d'établissement d'une typologie de sources thermales par une approche hydrochimique, isotopique et tectonique : Exemple du Maroc.- Thčse de Doctorat, Université Paris XI, Orsay, 326 + 88 p.
Zizi M. (2002).- Triassic-Jurassic extensional systems and their neogene reactivation in northern Morocco: The Rides Prérifaines and Guercif Basin.- Notes et Mémoires du Service géologique du Maroc, Rabat, no. 146, 138 p.
Zonneveld K.A.F. (1995).- Palaeoclimatical and palaeo-ecological changes during the last deglaciation in the Eastern Mediterranean - implications for dinoflagellate ecology.- Review of Palaeobotany and Palynology, vol. 84, no. 3-4, p. 221-253.
Zonneveld K.A.F., Höll C., Janofske D., Karwath B., Kerntopf B., Rühlemann C. & Willems H. (1999).- Calcareous dinoflagellate cysts as paleo-environmental tools. In: Fischer G. & Wefer G. (eds.), Use of proxies in paleoceanography.- Springer, p. 145-164.
|
Plate 1: 1: Globigerina bulloides (KA-102/49m); 2: Globigerina falconensis (KA-102/20m); 3: Globoturborotalita nepenthes (KA-102/91m); 4-5: Globorotalia menardii with sinistral coiling (KA-102/476m); 6-7: Globorotalia menardii with dextral coiling (KA-102/195m); 8: Globorotalia conomiozea (KA-102/66m); 9: Globorotalia miotumida (KA-102/66m); 10: Globorotalia suterae (KA-102/329m); 11: Globorotalia scitula with sinistral coiling (KA-102/329m); 12: Globorotalia scitula with dextral coiling (KA-102/185m); 13: Neogloboquadrina acostaensis (KA-102/70m); 14: Globoquadrina altispira (KA-102/35m); 15: Sphaeroidinellopsis seminulina (KA-102/203m).
|
|