◄ Carnets Geol. 23 (2) ►
![]()
Outline:
[1. Introduction]
[2. Geological background and material]
[3. Description of fossils]
[4. Discussion] [5. Conclusions]
and ...
[Bibliographic references]
Institute of Ecology and Earth Sciences,
University of Tartu, Ravila 14A, 50411, Tartu (Estonia)
Institute
of Geological Sciences, Jagiellonian University, ul. Gronostajowa
3a, 30-387 Kraków (Poland)
Faculty of Science, Charles University in Prague,
Albertov 6, 128 43 Praha 2 (Czech Republic)
Published online in final form (pdf) on February 15, 2023
DOI 10.2110/carnets.2023.2302
![]()
[Editor: Bruno
R.C. Granier; language editor: Stephen Carey]
![]()
The fossils described here are from the Tithonian-lower Berriasian Štramberk Limestone of the Czech Republic. The specimens consist of densely packed, straight to slightly curved short tubes. Each tube is composed of a single valve rather than two valves. Some tubes show constrictions in the wall, generally on just one side. The lumen is almost completely filled with numerous densely packed slightly to strongly concave tabulae. In some tubes, the tabulae form complete internal floors while in others they connect to the wall with another tabula or seal the concavity in the underlying tabula. The tubes somewhat resemble tubeworms, rudists, vermetids and calcareous algae. However, based on morphological similarities such as the tube-like morphology in longitudinal section and the shapes of the lamellae and tabulae, these tubes resemble oysters described from the Albian of Brazil. Accordingly, we infer that the Štramberk tubes represent oysters or oyster-related bivalves.
• Bivalves;
• tubular organisms;
• problematic fossils;
• biological affinities;
• Jurassic
Vinn O., Kołodziej B. & Kozlová Z. (2023).- Peculiar tube-like bivalve with densely packed concave tabulae (Štramberk Limestone, Tithonian-Berriasian).- Carnets Geol., Madrid, vol. 23, no. 2, p. 43-51.
Sur un bivalve tubulaire particulier ŕ tabulae concaves densément empilées (Calcaires de Štramberk, Tithonien-Berriasien).- Les fossiles décrits proviennent des Calcaires de Štramberk d'âge Tithonien-Berriasien inférieur de République tchčque. Ces spécimens sont constitués de tubes courts, droits ŕ faiblement incurvés et densément empilés. Chaque tube est composé d'une seule valve, plutôt que de deux valves, vraisemblablement cimentée au substrat ; l'autre valve est manquante. Certains comportent des constrictions au niveau de la paroi, habituellement sur seulement un côté du tube. Le lumen de ces tubes est presque entičrement rempli de nombreuses tabulae, faiblement ŕ fortement concaves, densément empilées. Elles peuvent former des planchers complets ŕ l'intérieur de certains tubes ; dans d'autres, elles joignent simplement la paroi du tube ŕ une autre tabula ou scellent la concavité dans la tabula inférieure. Les tubes décrits ressemblent quelque peu ŕ des annélides, des rudistes, des vermetidés, voire ŕ des algues calcaires. Toutefois, sur la base de similitudes morphologiques telles que la morphologie tubulaire en section longitudinale et la forme des lamelles et des tabulae, ces tubes ressemblent également aux huîtres décrites dans l'Albien du Brésil. En conclusion, nous en déduisons que les tubes de Štramberk représentent des huîtres ou des bivalves apparentés aux huîtres.
• bivalves ;
• organismes tubulaires ;
• fossiles problématiques ;
• affinités biologiques ;
• Jurassique
The Paleozoic is well known for its problematic tubular fossils and, similarly, the biological affinities of Mesozoic tubular fossils can be sometimes difficult to ascertain. Tubular skeletons occur in many Mesozoic marine invertebrate groups such as serpulids, sabellids, vermetids and rudists, to name a few. In addition, fossils of other groups such as calcareous algae could resemble tubes. The external morphology of tubular fossils usually does not provide enough morphological details to identify the organisms with confidence. The internal structure of tubular organisms is more often of value in revealing clues to their systematic affinities. The identified biological affinities of fossils help us to better understand the paleoecology of these organisms and reconstruct paleo-ecosystems.
The Late Jurassic was a time of widespread development of carbonate platforms and reefs. Most European studies are concerned with the upper Oxfordian-lower Kimmeridgian of the northern Tethys shelf. Less is known about the carbonate platforms of the Tethys domain and of Tithonian age, exemplified by the Štramberk Carbonate Platform. The sample of previously unknown tubular fossils (henceforth called the Štramberk tubes) studied here was collected from the Tithonian-lower Berriasian Štramberk Limestone (Carpathians, Czech Republic). These limestones were deposited on the intra-Tethys carbonate platform and contain numerous fossils (Vašíček & Skupien, 2004, 2005). Many fossils were described in the nineteenth century, and were summarized by Blaschke (1911). The Štramberk Limestone is famous for its reef faunas that dwelled on the northern margin of the Tethys. We interpret these tubular fossils to be bivalves. Boehm (1883) described 90 bivalve species (including species in open nomenclature) from the Štramberk Limestone and Štramberk-type limestones from Cieszyn Silesia (Poland), but none of them resembles the tubes addressed here.
The aim of the present paper is to: 1) describe tubular fossils from the Štramberk Limestone; and 2) discuss their biological affinities.
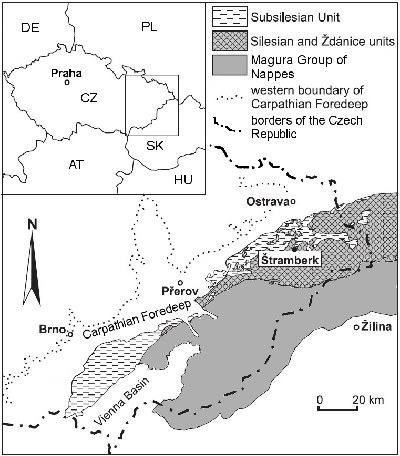
Megablocks of the Štramberk Limestone in Moravia (Czech Republic) are fragments of the carbonate platform embedded in the deep-water siliciclastic flysch of the Silesian Nappe of the Western Carpathians (Picha et al., 2006). Until the 1980s the Štramberk Limestone was considered to be of Tithonian age, but both calpionellid (Houša, 1990) and ammonite data (e.g., Vašíček & Skupien, 2013, 2014, 2016) indicate a Tithonian-early Berriasian age (latest Jurassic-earliest Cretaceous). The Štramberk Limestone is exposed in several quarries in the immediate vicinity of the city of Štramberk near Ostrava. The largest one is the active Kotouč Quarry where the sample studied was collected. Limestones, creamy beige in colour, occur in the form of carbonate megablocks, breccias and conglomerates. The geology of the Štramberk area and the nature of the megablocks are controversial. The carbonate megablocks have been interpreted as a tectonic klippe (Houša, 1990) or as carbonate bodies embedded in base-of-slope conglomerates and slump bodies formed between the Tithonian and the Turonian (e.g., Eliáš & Eliášová, 1986; Svobodová et al., 2011; Vašíček & Skupien, 2014). A combination of mass movement (gravitational slides and turbidity currents) and tectonic activity was inferred by Picha et al. (2006).
Traditionally, the Štramberk Limestone is considered as a reefal limestone, but it consists of diverse facies of the carbonate platform and its slope. Most common are detrital limestones (Eliáš & Eliášová, 1984, 1986; Hoffmann et al., 2017; Vaňková et al., 2019). The spatial and temporal relations of particular limestone olistoliths and smaller blocks are usually unknown. Similar to the Štramberk Limestone, in terms of lithology and age, are the Štramberk-type limestones occurring within the uppermost Jurassic-Oligocene flysch as pebbles and small blocks (so-called exotics; sporadically small klippen) in the Polish Outer Carpathians (Hoffmann et al., 2021). The Štramberk Limestone and the Štramberk-type limestones were developed on narrow platforms attached to intra-basinal highs (e.g., Baška Ridge, Silesian Ridge) in the Carpathian Basin of the Neotethys realm. These platforms are collectively termed the Štramberk Carbonate Platform (Eliáš & Eliášová, 1984, 1986; Hoffmann et al., 2021). The Štramberk Limestone contains numerous fossils (about 1,000 species), studied since the middle of the 19th century (see Blaschke, 1911; Vašíček & Skupien, 2004, 2005). Especially common and diversified in the Štramberk Limestone and in the Štramberk-type limestones are corals (ca. 120 species belonging to 50 genera). They are the world's most diverse reef assemblages from the Jurassic-Cretaceous transition (see Vašíček & Skupien, 2004, 2005; Eliáš & Eliášová, 1984, 1986; Eliášová, 2008; Kołodziej, 2015; Kozlová, 2022).
The sample examined was collected in the active Kotouč Quarry from
level 7 (see Vašíček & Skupien,
2013, Fig. 2) (Fig. 1 ![]() ).
Five thin sections of standard size (40 x 27 mm) and four larger thin sections
(60 x 50 mm), as well as six polished slabs, were prepared. The specimen and
thin sections are housed at the Institute of Geological
Sciences, Jagiellonian University in Kraków.
).
Five thin sections of standard size (40 x 27 mm) and four larger thin sections
(60 x 50 mm), as well as six polished slabs, were prepared. The specimen and
thin sections are housed at the Institute of Geological
Sciences, Jagiellonian University in Kraków.
|
Figure 1:
Location
of the study area on the tectonic map of the Outer Carpathians (based on Vašíček
and Skupien, 2013, simplified). |
The specimen with densely packed
tubes measures 140 mm x 80 mm in transverse section and ca. 60 mm in length.
Straight to slightly curved tubular shells are of diameter ranging from 3 to 8
mm (Fig. 2.A-D ![]() ) and are 15 to 25 mm long. The larger tubes (probably representing
the left valve) are more typically bivalve-like in appearance than the smaller
tubes (it is not clear whether the latter represent the left or right valve).
The preservation of larger and smaller tubes also differs somewhat as the
calcite of large tubes is dark and that of small tubes are lighter (Fig.
2.A
) and are 15 to 25 mm long. The larger tubes (probably representing
the left valve) are more typically bivalve-like in appearance than the smaller
tubes (it is not clear whether the latter represent the left or right valve).
The preservation of larger and smaller tubes also differs somewhat as the
calcite of large tubes is dark and that of small tubes are lighter (Fig.
2.A ![]() ).
).
|
Figure 2:
Densely packed tubes from the Štramberk
Limestone; Kotouč Quarry. A. Polished surface
of tubes in transverse section. B. Polished surfaces of tubes in longitudinal
section. Scale bar 1 cm. |
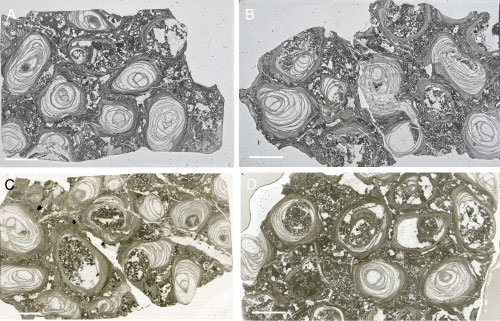
Most of the tube lumen is filled
with numerous densely packed, slightly to strongly concave tabulae (Figs. 3 ![]() - 4
- 4 ![]() ).
There are eight to 13 tabulae per 5 mm (Fig. 4.A-D
).
There are eight to 13 tabulae per 5 mm (Fig. 4.A-D ![]() ). The tabulae can form
complete floors inside the tube, or connect another tabula to the tube wall, or
seal the concavity in the tabula below (Fig. 4.A-D
). The tabulae can form
complete floors inside the tube, or connect another tabula to the tube wall, or
seal the concavity in the tabula below (Fig. 4.A-D ![]() ). The tabulae do not have any
perforations (Fig. 4.A-D
). The tabulae do not have any
perforations (Fig. 4.A-D ![]() ). The space between tabulae is filled with light
sparitic cement (Figs. 3
). The space between tabulae is filled with light
sparitic cement (Figs. 3 ![]() - 4
- 4 ![]() ).
).
|
Figure 3:
A-D. Transverse sections of the tubes showing lamellar tube wall and numerous
tabulae in the interior of tubes. Štramberk Limestone; Kotouč Quarry. Scale bar 5 mm. |
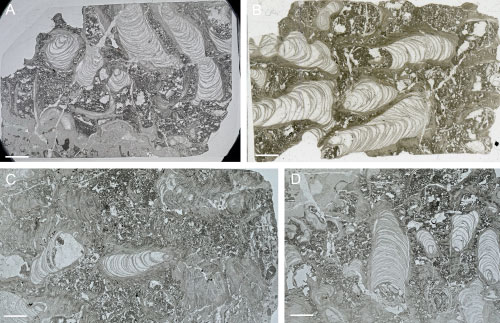
The large tubes are filled with
sediment (Fig.
2.A ![]() ). Some specimens show constrictions in the tube wall which
usually affect just one side of the tube (Fig. 4.A-B
). Some specimens show constrictions in the tube wall which
usually affect just one side of the tube (Fig. 4.A-B ![]() ). The tube wall is composed
of loosely packed lamellae that decline from the tube axis at 40-70°(Fig. 4.A-D
). The tube wall is composed
of loosely packed lamellae that decline from the tube axis at 40-70°(Fig. 4.A-D ![]() ). Some lamellae are straight but most are slightly curved in longitudinal
section with their convex side oriented towards the tube aperture. The ends of
single growth lamellae or groups of lamellae are separated from each other and
project away from the tube wall. All tubes are cylindrical or slightly wider
proximally. Tube bases are flat to slightly undulating in longitudinal section (Fig. 4.A-D
). Some lamellae are straight but most are slightly curved in longitudinal
section with their convex side oriented towards the tube aperture. The ends of
single growth lamellae or groups of lamellae are separated from each other and
project away from the tube wall. All tubes are cylindrical or slightly wider
proximally. Tube bases are flat to slightly undulating in longitudinal section (Fig. 4.A-D ![]() ).
).
Between tubes a shallow-water bioclastic grainstone occurs. However, because of the small size of the limestone sample it is not possible to determine whether the sample is from a reefal or other shallow-water facies.
|
Figure 4:
A-D. Longitudinal sections of the tubes showing numerous tabulae in the interior
of tubes and tube wall composed of loosely packed lamellae that decline from the
tube axis at 40-70°. Štramberk Limestone; Kotouč
Quarry. Scale bar 5 mm. |
Bivalves
Among the shells of molluscs, only rudists, vermetids and some oysters show any similarity to the tubes from Štramberk. The straight to slightly curved lamellae in the tube wall are consistent with the shape of lamellae in most molluscs. The earliest rudists (diceratids) are known from the Upper Jurassic (Kauffman & Johnson, 1988), including the Štramberk Limestone. Somewhat similar shells occur in small rudists such as Mathesia darderi (diameter: 0.6 to 2.4 cm) from the Barremian (Early Cretaceous) of Bulgaria (Fenerci-Masse et al., 2011). Rudist shells are often tubular (Kauffman & Johnson 1988) and could resemble tubeworms, but they usually are of much smaller diameter than that observed in the tubes. Rudist shells are attached to the substrate with one of their two valves (Kauffman & Johnson, 1988). It is likely that the Štramberk tubes were also cemented to a hard substrate as suggested by the flattened or undulating relief of the base of the tubes. Tabulae (i.e., more or less horizontal floors) may exist in the interior of the attached valve as also occurs in the Štramberk tubes (Fenerci-Masse et al., 2011). The shells of rudists are usually composed of two layers, one aragonitic and the other calcitic (Kauffman & Johnson, 1988). However, the tubes from Štramberk have only one shell layer, of calcite, making rudistan affinity for the Štramberk tubes unlikely. The major diagnostic characters of rudists are sockets and teeth that served to connect the two valves (Kauffman & Johnson, 1988; Skelton & Smith, 2000). In the Štramberk tubes, there is no evidence of sockets or teeth and thus rudist affinities can be ruled out with confidence.
The
inner layer of the oyster shell often contains chambers and vesicles and
sometimes chalky deposits; they can also have extraordinarily thick shells of
large size (Vermeij, 2014). Typically oysters do not have tubular shells
but their shell morphology is very variable. Oysters with a chambered inner
shell layer are known from the Lower Jurassic of northern Chile (Malchus
& Aberhan, 1998). The shape of the lamellae and tabulae in the Štramberk
tubes are similar particularly to those of oysters described from the lower-middle
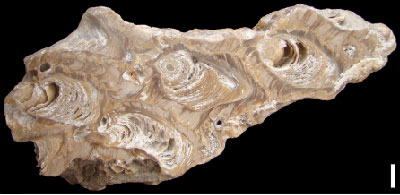
Albian of Brazil (Fig. 5 ![]() ). The Brazilian oysters likewise show a tube-like
morphology in longitudinal section (Granier & Dias-Brito,
2015, p. 126, Fig.
3A
). The Brazilian oysters likewise show a tube-like
morphology in longitudinal section (Granier & Dias-Brito,
2015, p. 126, Fig.
3A ![]() ; here Fig. 5
; here Fig. 5 ![]() ). Based on these similarities it is likely that
the Štramberk tubes belong to oysters or oyster-related bivalves (Fig. 5
). Based on these similarities it is likely that
the Štramberk tubes belong to oysters or oyster-related bivalves (Fig. 5 ![]() ). The
problem for the assignment of the Štramberk tubes to ostreids is the absence of
a former, inner aragonitic shell layer. A possibility is that the tabulae may
represent the remains of formerly aragonitic layer in question, which enclosed
the soft body. This hypothesis is here illustrated by a close-up photograph
showing the connection between the calcitic outer shell layer and several
adjacent tabulae, verifying that the soft body was not in direct contact with
the calcitic shell layer (Fig. 6
). The
problem for the assignment of the Štramberk tubes to ostreids is the absence of
a former, inner aragonitic shell layer. A possibility is that the tabulae may
represent the remains of formerly aragonitic layer in question, which enclosed
the soft body. This hypothesis is here illustrated by a close-up photograph
showing the connection between the calcitic outer shell layer and several
adjacent tabulae, verifying that the soft body was not in direct contact with
the calcitic shell layer (Fig. 6 ![]() ).
).
|
Figure 5:
Polished slab with several large calcite-dominated shells from Fazenda Cafuz,
Sergipe Basin, Brazil; Riachuelo Formation, ?lower-middle Albian (from Granier
and Dias-Brito, 2015, p. 126, Fig.
3A |
|
Figure 6:
Detail showing the connection between the calcitic outer shell layer and several
adjacent tabulae. Scale bar 2 mm. |
Vermetids
Vermetid gastropods have tubular shells that lack coiling in their adult part. Vermetids appeared in the Late Cretaceous (Bandel & Kowalke, 1997) and their oldest representatives are much younger than the Štramberk tubes. Vermetid shells have a spirally coiled protoconch, imperforate tabulae and a wall structure with one or more superimposed layers, which are characteristic of all mollusks (Sanfilippo et al., 2021). The wall structure of vermetids and the presence of imperforate tabulae resemble features of the Štramberk tubes. However, there is no sign of a spirally coiled protoconch in our material, though this could have been missed in the thin sections studied. Moreover, the tabulae of vermetids are always complete anteriorly concave floors inside the tube lumen (Sanfilippo et al., 2021), while in our material tabulae can be incomplete and closing just the concavity in another tabula or a section between the tabula and the tube wall. The vermetid tubes do not exhibit any constriction and are not wider in their proximal parts. All these features suggest that the Štramberk tubes are not related to vermetid gastropods.
Polychaetes
The straight to slightly curved lamellae in the longitudinal section of the tube wall in the Štramberk tubes are not consistent with the chevron-shaped lamellae of serpulids and therefore serpulid affinities can be excluded with confidence. The occurrence of numerous densely packed tabulae in the tube lumen is also unknown in serpulids. The shape of rare perforate tabulae in serpulids differs from that of the imperforate tabulae in the Štramberk tubes. However, straight lamellae that are slightly tilted from the direction of the tube longitudinal axis occur in calcareous sabellids of the genus Glomerula (Vinn et al., 2008). On the other hand, calcareous sabellids do not have tabulae in their tubes. Moreover, sabellid tubes lack constrictions; they are narrower in their proximal part and wider in their distal part. The shape of sabellid tubes is different from that of the Štramberk tubes, such that they are usually irregularly curved and attached to the substrate along their entire length; alternatively they may form knots of various sizes (Jäger, 1983, 2004). Thus, sabellid affinity of the Štramberk tubes can be ruled out. The tabulae of cirratulid tubes somewhat resemble those of the Štramberk tubes but the former always form complete floors inside the tube lumen (Fisher et al., 2000; Vinn, 2009). Calcareous cirratulid tubes have lamellar tube walls but their tube lamellae do not have loose ends projecting away from the tube external surface. The microstructure of cirratulid tubes is characterized by peloidal lamellae forming a stromatolitic fabric and intercalated lenses of fibrous aragonite that differs from that of the Štramberk tubes (Fisher et al., 2000; Vinn et al., 2009). Almost certainly the Štramberk tubes do not belong to cirratulids.
Barbafera is a probable polychaete fossil genus known from Triassic reefs (Senowbari-Daryan, 1997). The tubes of Barbafera resemble those from Štramberk in containing multiple chambers. However, in Barbafera, the chambers do not fill the tube lumen but surround it and are located inside the tube wall (Senowbari-Daryan, 1997). The loose ends of lamellae on the exterior surface of the tube somewhat resemble those of the Štramberk tubes. However, given the regular shape of the chambers and the fact that these chambers are not inside the tube lumen, it is unlikely that the Štramberk tubes are closely related to the genus Barbafera.
Is it possible that Štramberk tubes were originally organic tubes, like those of chaetopterids (Kiel & Dando, 2009) or siboglinids (Georgieva et al., 2019)? Organic tubes could mineralize during the fossilization process. However, chaetopterids and siboglinids do not have numerous tabulae in their tubes (Georgieva et al., 2019). The other major difference is in the morphology of the external tube wall. The external tube wall of chaetopterids or siboglinids is not covered by the free ends of the growth lamellae. Thus, even if the tubes were originally not biomineralized they are not comparable to the organic tubes of the polychaetes.
Calcareous algae
One should explore the possibility that the Štramberk tubes may not be invertebrate fossils but calcareous algae. The calcareous algae in thin sections may be superficially similar to the Štramberk tubes (Scholle & Ulmer-Scholle, 2005). However, sections of algae do not display free margins of the growth lamellae on their external surfaces. Thus, most likely the Štramberk tubes do not belong to calcareous algae but are invertebrate skeletons.
The tube lumen is almost completely filled with numerous densely packed, slightly to strongly concave tabulae. The Štramberk tubes exhibit some similarity to calcareous algae, tubular mollusk shells and calcareous tubeworms. The sections of algae do not show loose ends of growth lamellae on their external surfaces, making algal affinities for the Štramberk tubes unlikely. The tubes of the possible polychaete Barbafera resemble those of Štramberk in containing multiple chambers but, in Barbafera, the chambers surround the tube lumen rather than filling it and are located inside the tube wall. The Štramberk tubes also differ from those of any other tubeworm; therefore they are unlikely to be closely related to any of them. In the Štramberk tubes, there is no evidence of sockets or teeth; therefore rudist affinities can be ruled out with certainty. It is most likely that the Štramberk tubes are not closely related to vermetid gastropods since their tabulae differ. Based on morphological similarities such as the tube-like morphology of the longitudinal section of the shell, the shape of the lamellae, and the shape and number of tabulae, the Štramberk tubes resemble oysters discussed in Granier and Dias-Brito (2015) and therefore the Štramberk tubes may belong to oysters or other oyster-related bivalves.
We are grateful to Bruno Granier for suggestions about the affinity of the Štramberk tubes. Financial support to O.V. was provided by an Institute of Ecology and Earth Sciences (University of Tartu) Research Grant and a Paleontological Society Sepkoski Grant. We would like to acknowledge Petr Skupien (Ostrava), Justyna Kowal-Kasprzyk (Kraków) and Jan Geist (Praha) for assistance in field studies. We are grateful to journal reviewers Bruno Ferré and Jean-Pierre Masse for constructive comments on the manuscript.
Bandel K. & Kowalke T. (1997).- Cretaceous Laxispira and a discussion on the monophyly of vermetids and turritellids (Caenogastropoda, Mollusca).- Geologica et Palaeontologica, Marburg, vol. 31, p. 257-274.
Blaschke F. (1911).- Zur Tithonfauna von Stramberg in Mähren.- Annalen des Naturhistorischen Museums in Wien, Vienna, vol. 25, p. 143-222.
Boehm G. (1883).- Die Bivalven der Stramberger Schichten.- Palaeontographica, Supplement Abtheilung, Stuttgart, vol. 2, no. 4, p. 493-680.
Eliáš M. & Eliášová H. (1984).- Facies and palaeogeography of the Jurassic in the western part of the Outer Flysch Carpathians in Czechoslovakia.- Sborník gelogických Vĕd, Prague, vol. 39, p. 105-170.
Eliáš M. & Eliášová H. (1986).- Elevation facies of the Malm in Moravia.- Geologicky Zbornik, Prague, vol. 37, p. 533-550.
Eliášová H. (2008).- Corals from the Štramberk Limestone (Silesian Unit, Western Outer Carpathians, Czech Republic). In: Krobicki M. (ed.), Utwory przełomu jury i kredy w zachodnich Karpatach fliszowych polsko-czeskiego pogranicza. Jurassica VII, Żywiec/Štramberk, 27-29/09/2008.- Geologia, Prague, vol. 34, p. 151-160.
Fenerci-Masse M., Masse J.-P., Kołodziej B., Ivanov M. & Idakieva V. (2011).- Mathesia darderi (Astre) (Bivalvia, Hippuritoidea, Monopleuridae): Morphological, biogeographical and ecological changes in the Mediterranean domain during the late Barremian-Albian.- Cretaceous Research, vol. 32, no. 4, p. 407-421.
Fischer R., Pernet B. & Reitner J. (2000).- Organomineralization of cirratulid annelid tubes - Fossil and recent examples.- Facies, Erlangen, vol. 42, p. 35-50.
Georgieva M.N., Little C.T.S., Watson J.S., Sephton M.A., Ball A.D. & Glover A.G. (2019).- Identification of fossil worm tubes from Phanerozoic hydrothermal vents and cold seeps.- Journal of Systematic Palaeontology, vol. 17, p. 287-329.
Granier B. & Dias-Brito D. (2015).- End of a modern geological myth: There are no rudists in Brazil! Paleobiogeographic implications.- Carnets Geol., Madrid, vol. 15, no. 11, p. 123-136. DOI: 10.4267/2042/56880
Hoffmann M., Kołodziej B. & Kowal-Kasprzyk J. (2021).- A lost carbonate platform deciphered from clasts embedded in flysch: Štramberk-type limestones, Polish Outer Carpathians.- Annales Societatis Geologorum Poloniae, Kraków, vol. 91, p. 203-251.
Hoffmann M., Kołodziej B. & Skupien P. (2017).- Microencruster-microbial framework and synsedimentary cements in the Štramberk Limestone (Carpathians, Czech Republic): Insights into reef zonation.- Annales Societatis Geologorum Poloniae, Kraków, vol. 87, p. 325-347.
Houša V. (1990).- Stratigraphy and calpionellid zonation of the Štramberk Limestone and associated Lower Cretaceous beds.- Atti del secondo convegno internazionale Fossili, Evoluzione, Ambient, Pergola 25-30 ottobre 1987, p. 365-370.
Jäger M. (1983).- Serpulidae (Polychaeta sedentaria) aus der norddeutschen höheren Oberkreide-Systematik, Stratigraphie, Ökologie.- Geologisches Jahrbuch, Reihe A, Hannover, vol. 68, p. 3-219.
Jäger M. (2004).- Serpulidae und Spirorbidae (Polychaeta sedentaria) aus Campan und Maastricht von Norddeutschland, den Niederlanden, Belgien und angrenzenden Gebieten.- Geologisches Jahrbuch, Reihe A, Hannover, vol. 157, p. 121–249.
Kauffman E.G. & Johnson C.C. (1988).- The morphological and ecological evolution of Middle and Upper Cretaceous reef-building rudistids.- Palaios, Lawrence - KS, vol. 3, no. 2, p. 194-216.
Kiel S. & Dando P.R. (2009).- Chaetopterid tubes from vent and seep sites: Implications for fossil record and evolutionary history of vent and seep annelids.- Acta Palaeontologica Polonica, Warsaw, vol. 54, p. 443-448.
Kołodziej B. (2015).- Corals of the Štramberk-type limestones from Poland: taxonomic and palaeoecological aspects.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 276, p. 181-199.
Kozlová Z. (2022).- Corals from the Štramberk carbonate platform (Tithonian-Berriasian, Poland, Carpathians): An overview. In: Pálfy J. and Főzy I. (eds.), 11th International Congress on the Jurassic System, 29/08-02/09/2022, Budapest.- Abstracts, Program and Field Trip Guide, p. 64.
Malchus N. & Aberhan M. (1998).- Transitional gryphaete/exogyrate oysters (Bivalvia: Gryphaeidae) from the Lower Jurassic of northern Chile.- Journal of Paleontology, Tulsa - OK, vol. 72, p. 619-631.
Picha F.J., Stráník Z. & Krejčí O. (2006).- Geology and hydrocarbon resources of the Outer Western Carpathians and their foreland, Czech Republic. In: Golonka J. & Picha F.J. (eds), The Carpathians and their foreland: Geology and hydrocarbon resources.- AAPG Memoir, Tulsa - OK, vol. 84, p. 49-175.
Sanfilippo R., Kočí T., Bosio G., Collareta A., Ekrt B., Malinverno E., Di Celma C., Urbina M. & Bianucci G. (2021).- An investigation of vermetid reefs from the Miocene of Peru, with the description of a new species.- Journal of South American Earth Sciences, Article 103233, 13 p.
Scholle P.A. & Ulmer-Scholle D. (2005).- A color guide to the petrography of carbonate rocks: Grains, textures, porosity, diagenesis.- AAPG Memoir, Tulsa - OK, vol. 77, 486 p.
Senowbari-Daryan B. (1997).- Barbafera carnica Senowbari-Daryan, 1980: A Triassic worm-tube.- Facies, Erlangen, vol. 36, p. 57-68.
Skelton P.W. & Smith A.B. (2000).- A preliminary phylogeny for rudist bivalves: Sifting clades from grades. In: Harper E.M., Taylor J.D. & Crame J.A. (eds.), The evolutionary biology of the Bivalvia.- Geological Society Special Publication, vol. 177, p. 97-127.
Svobodová M., Švábenická L., Skupien P. & Hradecká L. (2011).- Biostratigraphy and paleoecology of the Lower Cretaceous sediments in the Outer Western Carpathians (Silesian Unit, Czech Republic).- Geologica Carpathica, Bratislava, vol. 62, p. 309-332.
Vaňková L., Elbra T., Pruner P., Vašíček Z., Skupien P., Reháková D., Schnabl P., Košťák M., Švábenická L., Svobodová A., Bubík M., Mazuch M., Čížková K. & Kdýr Š. (2019).- Integrated stratigraphy and palaeoenvironment of the Berriasian peri-reefal limestones at Štramberk (Outer Western Carpathians, Czech Republic).- Palćogeography, Palćoclimatology, Palćoecology, vol. 532, article 109256, 21 p.
Vašíček Z. & Skupien P. (2004).- The Štramberk fossil site (uppermost Jurassic/Lower Cretaceous, Outer Western Carpathians) - two centuries of the geological and paleontological research. Sborník Vědeckých Prací Vysoké školy Báňské - Technické Univerzity Ostrava.- Řada Hornicko-Geologická, Ostrava, vol. 50, p. 83-102 [in Czech, with English summary].
Vašíček Z. & Skupien P. (2005).- Supplements to history of geological and paleontological research of Štramberk territory. Sborník Vědeckých Prací Vysoké školy Báňské - Technické Univerzity Ostrava.- Řada Hornicko-Geologická, Ostrava, vol. 51, p. 1-6 [in Czech, with English summary].
Vašíček Z. & Skupien P. (2013).- Early Berriasian ammonites from the Štramberk Limestone in the Kotouč Quarry (Outer Western Carpathians, Czech Republic).- Annales Societatis Geologorum Poloniae, Kraków, vol. 83, p. 329-342.
Vašíček Z. & Skupien P. (2014).- Recent discoveries of Tithonian ammonites in the Štramberk Limestone (Kotouč Quarry, Outer Western Carpathians).- Annales Societatis Geologorum Poloniae, Kraków, vol. 84, p. 131-141.
Vašíček Z. & Skupien P. (2016).- Tithonian-early Berriasian perisphinctoid ammonites from the Štramberk Limestone at Kotouč Quarry near Štramberk, Outer Western Carpathians (Czech Republic).- Cretaceous Research, vol. 64, p. 12-29.
Vermeij G.J. (2014).- The oyster enigma variations: A hypothesis of microbial calcification.- Paleobiology, vol. 40, p. 1-13.
Vinn O. (2009).- The ultrastructure of calcareous cirratulid (Polychaeta, Annelida) tubes.- Estonian Journal of Earth Sciences, Tallinn, vol. 58, no. 2, p. 153-156.
Vinn O., Hove H.A. ten & Mutvei H. (2008).- On the tube ultrastructure and origin of calcification in sabellids (Annelida, Polychaeta).- Palaeontology, vol. 51, no. 2, p. 295-301.