◄ Carnets Geol. 23 (1) ►
![]()
Outline:
[1. Introduction] [2.
Geological context] [3. The conceptualization of the Albian]
[4. The Aptian-Albian boundary in CSA, ESA, and BNE sedimentary basins] [5.
Discussion]
[6. Conclusion] [Bibliographic references] and ... [Supplementary data]
Visiting researcher at the Universidade Federal do Rio de Janeiro
(UFRJ), Rua Sara Braune, 40/101 a; ZIP Code 28611-020, Nova Friburgo, Rio de Janeiro (Brazil)
Visiting researcher at the Universidade Federal do Rio de Janeiro
(UFRJ), Rua Lopes da Cruz, 167/601; ZIP Code 20720-170, Rio de Janeiro (Brazil)
Instituto Tecnol�gico de Paleoceanografia e Mudan�as Clim�ticas - itt
Oceaneon, Universidade do Vale do Rio dos Sinos (UNISINOS University), Av.
Unisinos, 950 / Sector C11, ZIP Code 93. 022-750, S�o Leopoldo, Rio Grande do Sul (Brazil)
Published online in final form (pdf) on January 1, 2023
DOI 10.2110/carnets.2023.2301
![]()
[Editor: Bruno
R.C. Granier]
![]()
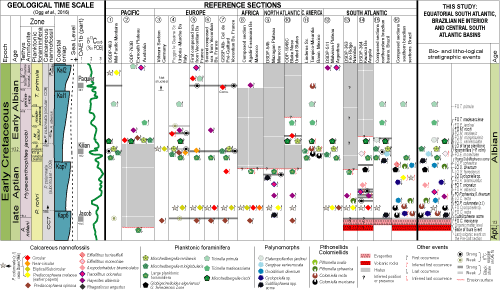
The use of taxonomic-phylogenetic criteria established for planktonic foraminifera in the 2000's and the definition of the Albian Global Stratotype Section Point (GSSP-Alb) have resulted in a major change in the interpretation of the carbonate sections overlying the giant layer of salt present in basins of the South Central Atlantic (CSA) and their equivalent strata in the Equatorial South Atlantic (ESA), and interior of northeastern Brazil (BNE). These post-salt carbonates have long been considered Albian in age, but they contain a planktonic foraminifera association characteristically Aptian. Great conflicts arise, however, when this faunal association is compared with biostratigraphic successions based on other fossil groups or with lithostratigraphic and geochronological data. Controversies similar to those observed at sites 363 and 364 of the Deep Sea Drilling Project (DSDP) leg 40, drilled almost 45 years ago, have resurfaced. Thus, it is paradoxical that the remarkable disappearance of large species of planktonic foraminifera, associated with the top of the Paraticinella rohri Zone (of the upper Aptian), occurred stratigraphically above a typical Albian calcareous nannofossil succession (as the First Stratigraphic Occurrences of Hayesites albiensis, Tranolithus orionatus, Axopodorhabdus biramiculatus, and Eiffellithus turriseiffelli) or the FOs of three known species of pelagic calcispheres, all assumed to be of Albian age. Another notorious conflict lies in the fact that these carbonates rest directly on the salt layer onlapping the South Atlantic Middle Barrier (SAMB), where trachyandesite has been dated at 113.2 0.1 Ma, identical to the value established for the GSSP-Alb. Detailed examination of 16 stratigraphic sections from around the world shows that the difficulties of fully applying the GSSP-Alb criteria are not limited to the CSA, ESA, and BNE basins. The explanation of these controversies may lie in the specific conditions of the water mass of the primitive South Atlantic that may have influenced morphological alterations or affected the temporal amplitude of taxa. But until geochronological, biostratigraphic, and lithostratigraphic incompatibilities can be clarified, it is here recommended to use the base of the evaporitic layer as the reference for the Aptian/Albian transition in CSA, ESA, and BNE basins.
� South Atlantic basins;
� biochronostratigraphic and geochronological conflict;
� planktonic foraminifera;
�
calcareous nannofossils;
� Albian Global Boundary Stratotype Section and Point
Azevedo R.L.M., Antunes R.L. & Bruno D.R. (2023).- Issues in the identification of the Aptian/Albian boundary in South Atlantic basins and beyond.- Carnets Geol., Madrid, vol. 23, no. 1, p. 1-42.
Probl�mes li�s � l'identification de la limite Aptien/Albien dans les bassins sud-atlantiques et au-del�.- L'utilisation de crit�res taxinomiques et phylog�n�tiques �tablis pour les foraminif�res planctoniques dans les ann�es 2000 et la d�finition du Point Stratotypique Mondial de l'Albien (PSM Albien) ont entra�n� un changement majeur dans l'interpr�tation des d�p�ts � dominante calcaire post�rieurs � l'�norme accumulation salif�re pr�sente dans l'Atlantique Centre-Sud (CSA), ainsi que dans celle des couches �quivalentes de l'Atlantique Sud �quatorial (ESA) et de l'int�rieur du nord-est du Br�sil (BNE). Ces "carbonates post-salif�res" ont longtemps �t� consid�r�s comme d'�ge albien alors qu'ils rec�lent une association de foraminif�res planctoniques caract�ristique de l'Aptien. Toutefois, des d�saccords apparaissent quand cette association faunistique est compar�e � des successions biostratigraphiques dat�es � partir d'autres groupes fossiles ou fond�es sur des donn�es lithostratigraphiques et g�ochronologiques. Des controverses similaires � celles observ�es dans l'interpr�tation des sites 363 et 364, Leg 40 du Deep Sea Drilling Project (DSDP), for�s il y a pr�s de 45 ans, refont surface. Paradoxalement, l'�v�nement remarquable que constitue la disparition de grandes esp�ces de foraminif�res planctoniques au sommet de la Zone � Paraticinella rohri (de l'Aptien sup�rieur) survient stratigraphiquement apr�s une s�rie d'apparitions de nannofossiles calcaires typiquement albiens (tels que les esp�ces Hayesites albiensis, Tranolithus orionatus, Axopodorhabdus biramiculatus et Eiffellithus turriseiffelli) ou apr�s l'apparition de trois esp�ces de calcisph�res p�lagiques, toutes consid�r�es comme d'�ge albien. Un autre conflit notoire r�side dans le fait que ces carbonates reposent directement sur la couche de sel, elle-m�me superpos�e � la Barri�re M�diane de l'Atlantique Sud (SAMB), o� une trachyandesite a �t� dat�e � 113,2 + 0,1 Ma, soit une valeur identique � celle qui est retenue dans l'�chelle des Temps G�ologiques 2020 pour le PSM Albien. L'examen d�taill� de 16 coupes stratigraphiques provenant de diff�rentes localit�s dans le monde montre que les difficult�s de l'application stricte des crit�res du PSM Albien ne se limitent pas aux bassins CSA, ESA et BNE. Il se peut que les r�ponses � ces controverses r�sident dans les conditions particuli�res de la masse d'eau de l'Atlantique Sud primitif, induisant des alt�rations morphologiques ou encore dans l'amplitude temporelle des taxons. Mais jusqu'� ce que les incompatibilit�s g�ochronologiques et bio-lithostratigraphiques soient clarifi�es, il est recommand� d'utiliser la base de la couche �vaporitique comme r�f�rence pour la limite Aptien/Albien dans les bassins CSA, ESA et BNE.
� bassins de l'Atlantique Sud ;
� conflits biochronostratigraphiques et g�ochronologiques ;
� foraminif�res planctoniques ;
�
nannofossiles calcaires ;
� Point Stratotypique Mondial de l'Albien
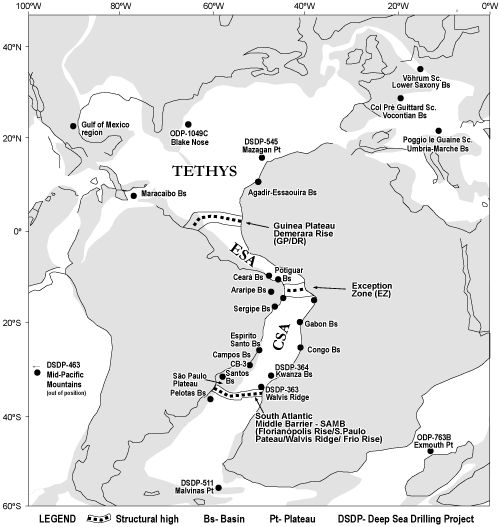
Sedimentary basins in the South Atlantic Ocean situated along the continental margins of Africa and South America, as well as some interior basins in the Brazilian Northeast (BNE), contain key elements for understanding Early Cretaceous evolution of the Central South Atlantic (CSA) and Equatorial South Atlantic (ESA). During this time, basins in the CSA lay between two barriers transverse to the main axis of Gondwana break-up: to the south, the Florian polis
High/S�o Paulo Plateau/Walvis and Frio Ridge, here referred to as the South Atlantic Middle Barrier (SAMB); and to the north, the Exception Zone (EZ), as defined by Barbosa et al. (2008). Water masses of numerous sedimentary basins throughout the ESA in the area extending from the EZ to the Guinea Plateau/Demerara Rise (Heine & Brune,
2014) were also connected to interior basins
(Fig. 1
![]() ).
).
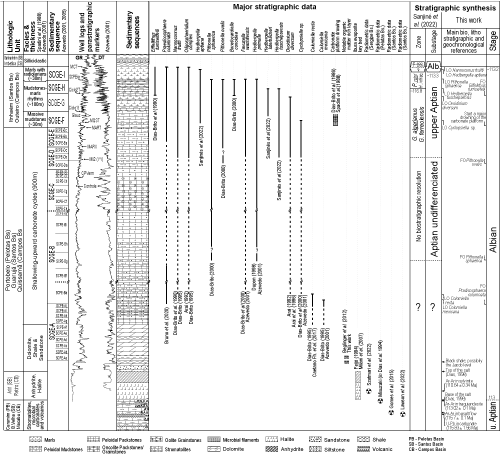
This paper discusses the chronostratigraphy of the first marine deposits in the CSA, ESA, and BNE based on a broad survey of the geological literature. Concepts, consistencies, and inconsistencies in 16 geological sections containing the Aptian-Albian boundary, including several from elsewhere, were examined, with special attention given to the biochronology of ammonites, planktonic foraminifera, calcareous nannofossils, pithonellids, colomiellids, and palynomorphs. Lithostratigraphic units and geochronological information for these basins have also been analyzed in the search for additional elements for calibration with Albian Global Stratotype Section Point (GSSP-Alb).
Many oil fields have been discovered in the CSA, ESA, and BNE basins over the last half-century. However, interest in the area increased in the last decade because of the discovery of the Pre-Salt hydrocarbon province along the southeastern Brazilian margin, one of the largest such discoveries of the 21st century (Jones & Chaves, 2011; Petersohn, 2019). Thus, given its economic importance, it is imperative that the age of the evaporites and overlying carbonates be precisely known. It is these carbonates that comprise the main focus of this study.
|
|
Figure 1: Location of the basins and sections analyzed in the Central South Atlantic (CSA) and Equatorial South Atlantic
(ESA), as well as in other regions around the world. Paleogeography is for 113 Ma ago (modified from
http://www.odsn.de). |
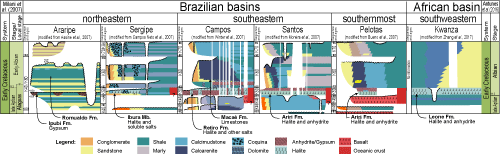
The break-up of Gondwana during the Early Cretaceous led to the development of a relatively common sedimentary succession in the basins from both sides of the CSA and ESA. During this time, a large sedimentary pile formed in these basins associated with the rift, sag, and drift phases of break-up (Milani et al.,
2007). In the rift phase, non-marine siliciclastic sediments were deposited, including black shales, together with grainstones, rudstones, and interlayered volcanic rocks
(Fig. 2
![]() ). Lacustrine deposits formed during this period allowed creation of a local chronostratigraphic scheme, whose stages were defined mostly by ostracod bioevents (Schaller,
1969; Viana et al., 1971). Yet five decades later, the equivalence between these units and international ones remains uncertain (Antunes et al.,
2018).
). Lacustrine deposits formed during this period allowed creation of a local chronostratigraphic scheme, whose stages were defined mostly by ostracod bioevents (Schaller,
1969; Viana et al., 1971). Yet five decades later, the equivalence between these units and international ones remains uncertain (Antunes et al.,
2018).
The youngest of these local stages, the Alagoas Stage, is a siliciclastic package covered by microbial carbonates-travertines and a very thick and widespread salt layer, which comprises the sag sequence and marks the beginning of the establishment of marine conditions in the CSA. The evaporitic package is dominated by halite and anhydrite, deposited during a time span of ~600 ka to 1 Ma (e.g., Dias,
1998,
2005; Fran a et al.,
2007; Moreira et al., 2007; Winter et al.,
2007), or as short as 530 ka according to Rodrigues et al.
(2018). This layer is around 2,000 m thick and is spread over an area of approximately 1,500 km by 500 km area, limited to the south by the SAMB and to the north by the EZ
(Fig. 1
![]() ). Scattered coeval evaporites, though not as thick, can also be seen in offshore and onshore deposits of the ESA (Asmus & Campos,
1983; Regali, 1989). Dias
(1998,
2005) created the useful term Ibura Event to unify analysis of all marine incursions responsible for the almost
"instantaneous" deposition of these salts, which have distinct lithostratigraphic designations in each basin.
). Scattered coeval evaporites, though not as thick, can also be seen in offshore and onshore deposits of the ESA (Asmus & Campos,
1983; Regali, 1989). Dias
(1998,
2005) created the useful term Ibura Event to unify analysis of all marine incursions responsible for the almost
"instantaneous" deposition of these salts, which have distinct lithostratigraphic designations in each basin.
Marine carbonates deposited in shallow waters and open seas are the main components at the beginning of the drift phase in the CSA; they can also be found in basins of the ESA and BNE (Milani et al.,
2007). Until the 2010's, these carbonates were dated as Albian in the synthetic stratigraphic charts of Brazilian basins published by Petrobras (Feij� ,
1994; Milani et al.,
2007). Since then, this entire carbonate section has been reclassified as Aptian because of biochronostratigraphic revisions mainly affecting planktonic foraminifera in the Santos, Campos, and
Esp�rito Santo basins (Lima et al.,
2018; Viviers et al., 2018; Sanjin s et al.,
2022). This was the result of both the application of taxonomic-phylogenetic criteria established for this protist group (Huber & Leckie,
2011) and the definition of the GSSP-Alb (Kennedy et al.,
2017). However, this interpretation conflicts with other renowned regional or global chronostratigraphic data, particularly that of calcareous nannofossils at Site 364, Kwanza Basin (Bruno et al.,
2020). Moreover, in the extreme south of the CSA, the salt layer onlaps volcanic rocks in the Florian polis High, where a trachyandesite was dated by the Ar/Ar method at
113.2 0.1 Ma (Mizusaki, 1993, fide Dias et al.,
1994), almost identical to the value established in the Geological Time Scale
2020 - GTS2020 (Gale et al.,
2021) for the base of the Albian (GSSP-Alb). Because of this, Antunes et al. (2018) proposed an alternative solution by adopting the Ibura evaporite as marking the Aptian-Albian boundary for basins in the CSA, ESA, and BNR, which justifies the chronostratigraphic discrepancies between the two extreme columns illustrated in
Figure 2
![]() .
.
|
|
Figure 2:
Stratigraphic chart for the Araripe (located in the BNE), Sergipe, Campos, Santos, Pelotas and Kwanza basins during the late
Aptian-Early Albian. For the Brazilian basins, stratigraphic adjustment was made considering geochronological interpretations and the position of the sequence boundaries described by Milani et al. (2007), who used the geological time scale of Gradstein et al. (2004). The right column illustrates the alternative proposal by Antunes et al. (2018), who suggested the base of the Ibura evaporite as the marker for the Aptian-Albian boundary. |
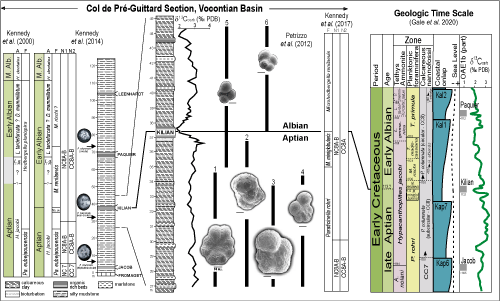
The Concise Geologic Time Scale 2016 (CGTS2016), by Ogg et al. (2016), integrates the detailed studies carried out by Kennedy et al. (2000), Petrizzo et al. (2012), and Kennedy et al. (2014) in outcrops of the Col de Pr�-Guittard, France, which were key for the proposal of that area as the Albian Global Stratotype Section Point (Petrizzo et al., 2016; Kennedy et al., 2017; GPS coordinates [according to Granier, 2017]: 44 30'28.3"N 5 17'50.1"E, not 44 29' 47.78"N 5 18'42.36"E). Differently from previous suggestions (e.g., Br�h�ret et al., 1986; Hart et al., 1996; Owen, 2002) and the geological time scales from 2004 and 2012 (GTS2004 and GTS2012; Ogg et al., 2004 and 2012; respectively), the CGTS2016 discarded the option of using the First Stratigraphic Occurrence (FO) of the calcareous nannofossil Prediscosphaera columnata as the datum for the base of the Albian. A perceived diachronism regarding the FO of P. columnata contributed to this decision after Bown (in Kennedy et al., 2000) recognized three morphological categories for the species based on their outlines: elliptical-subcircular, subcircular, and circular.
The new datum used to indicate the base of the Albian was the FO of the planktonic foraminifera Microhedbergella renilaevis
(Fig. 3
![]() ). This bioevent occurs during the notable biotic turnover around the Aptian-Albian boundary that strongly affected the evolution of these protists, led to a drastic decrease in both foraminifera abundance as well as the planktonic/benthic ratio and occurred during the last 1 Ma of the Aptian (Huber & Leckie,
2011). Another important factor is that the FO of M. renilaevis in Col de Pr�-Guittard occurs in the Killian anoxic level and is accompanied by other, stratigraphically close black shale layers: the Jacob Level, below, and the Paquier and Leenhardt levels, above, as described by Br�h�ret (1983). The age inferred for the GSSP-Alb was
113.1 0.3 Ma by the 238U/206Pb method, as determined in zircon from volcanic ash at V�hrum, Germany
(Selby et al., 2009). Distant about 1000km from Col Pr�-Guittard, the V�hrum section does not contain planktonic foraminifera, so that support for the chronocorrelation between the two outcrops came from ammonite associations and the FO of Prediscosphaera columnata (subcircular category) a few centimeters above the ash layer
(Mutterlose et al., 2003).
). This bioevent occurs during the notable biotic turnover around the Aptian-Albian boundary that strongly affected the evolution of these protists, led to a drastic decrease in both foraminifera abundance as well as the planktonic/benthic ratio and occurred during the last 1 Ma of the Aptian (Huber & Leckie,
2011). Another important factor is that the FO of M. renilaevis in Col de Pr�-Guittard occurs in the Killian anoxic level and is accompanied by other, stratigraphically close black shale layers: the Jacob Level, below, and the Paquier and Leenhardt levels, above, as described by Br�h�ret (1983). The age inferred for the GSSP-Alb was
113.1 0.3 Ma by the 238U/206Pb method, as determined in zircon from volcanic ash at V�hrum, Germany
(Selby et al., 2009). Distant about 1000km from Col Pr�-Guittard, the V�hrum section does not contain planktonic foraminifera, so that support for the chronocorrelation between the two outcrops came from ammonite associations and the FO of Prediscosphaera columnata (subcircular category) a few centimeters above the ash layer
(Mutterlose et al., 2003).
In the updated version of the Geological Time Scale (GTS2020) of Gale et al. (2021), a few alterations were introduced to the Albian, of which the three most notable were: changing the age of the GSSP-Alb from 113.1 0.3 to 113.2 0.3; not indicating limits for Albian substages; and promoting changes in the succession of ammonite zones More noteworthy still was the change in age from 126.3 to 121.4 Ma for the base of the Aptian, maintain the magnetic polarity-Chron M0 as datum. This clearly impacted the ages of the data that subdivide this stage. We mainly follow the GTS2020 in this paper but adopt the CGTS2016 criteria for identification of Albian substages where necessary
|
|
Figure 3:
The GSSP-Alb major stratigraphic data (modified from Petrizzo et al.,
2012; Kennedy et al.,
2017), FOs of the Prediscosphaera columnata varieties: elliptical-subcircular, subcircular, and circular (modified from Bown, in Kennedy et al., 2000,
2014). δ13Ccarb curve and stratigraphic amplitude of some planktonic foraminifera: 1) Paraticinella eubejaouaensis (=P. rohri), 2) Pseudoguembelitria blakenosensis, 3) Hedbergella infracretacea, 4) H. aptiana, 5) Microhedbergella miniglobularis, and 6) M. renilaevis (modified from Petrizzo et al.,
2012); scale bar is = 30 m, except for P. eubejaouaensis). And the CGTS2020, showing the biostratigraphic units defined with planktonic foraminifera and calcareous nannofossils; main black shale layers (Jacob, Kilian, Paquier, and Leenhardt); coastal onlap and sea level curves; and the δ13Ccarb reference curve. |
Here we summarize data from numerous studies that attributed Aptian and/or Albian ages to the marine sections of sedimentary basins of the CSA, ESA, and BNE. We also discuss lithological units having potential chronostratigraphic value and absolute ages of volcanic and sedimentary rocks that provide geochronological references for geological interpretations of these basins.
The pioneering studies with ammonites illustrated the difficulty in attributing ages for the oldest, post-salt marine sediments of the CSA, ESA, and BNE basins, sometimes dating them as late Aptian, sometimes as Early or younger Albian (e.g., Beurlen, 1961, fide Schaller,
1969; Kennedy & Cooper,
1975; Wiedmann & Neugebauer,
1978; Bolli,
1978; Bolli et al., 1978a,
1978b; Reyment & Dingle,
1987). The most recent paper by Bengtson et al. (2018) for the Sergipe Basin
(Fig. 1
![]() ) attributed an Aptian age to the association zones marked by Epicheloniceras and Eodouvilleiceras. They chronocorrelate these units with the Parahoplites melchioris Zone, Epicheloniceras martini Zone, and the basal portion of the Hypacanthoplites jacobi Zone, which are part of the standard international zonation scheme of the
GTS2020. They are equivalent to the planktonic foraminifera from the Leopoldina cabri (upper part) to Microhedbergella miniglobularis zones; to almost the entire range of Chiastozygus litterarius (CC7) and to the lower part of Prediscosphaera columnata (CC8), among the calcareous nannofossils. Bengtson et al. (2018) also consider the Douvilleiceras mammillatum Interval Zone as being Early Albian, corresponding to the upper part of the H. jacobi and D. mammillatum zones. They are equivalent to the Microhedbegella renilaevis and the lower part of the Ticinella primula zones in the planktonic foraminiferal biostratigraphic succession, and to a large part of Prediscosphaera columnata Zone. This set of zones comprises almost the entirety of the upper Aptian and the Lower Albian. However, because of doubts regarding species provincialism, lack of good outcrops, and uncertain stratigraphic positioning of many recovered specimens, Bengtson et al. (2018) regarded as
"tentative" their calibration between the Sergipe Basin ammonites and the standard international scheme.
) attributed an Aptian age to the association zones marked by Epicheloniceras and Eodouvilleiceras. They chronocorrelate these units with the Parahoplites melchioris Zone, Epicheloniceras martini Zone, and the basal portion of the Hypacanthoplites jacobi Zone, which are part of the standard international zonation scheme of the
GTS2020. They are equivalent to the planktonic foraminifera from the Leopoldina cabri (upper part) to Microhedbergella miniglobularis zones; to almost the entire range of Chiastozygus litterarius (CC7) and to the lower part of Prediscosphaera columnata (CC8), among the calcareous nannofossils. Bengtson et al. (2018) also consider the Douvilleiceras mammillatum Interval Zone as being Early Albian, corresponding to the upper part of the H. jacobi and D. mammillatum zones. They are equivalent to the Microhedbegella renilaevis and the lower part of the Ticinella primula zones in the planktonic foraminiferal biostratigraphic succession, and to a large part of Prediscosphaera columnata Zone. This set of zones comprises almost the entirety of the upper Aptian and the Lower Albian. However, because of doubts regarding species provincialism, lack of good outcrops, and uncertain stratigraphic positioning of many recovered specimens, Bengtson et al. (2018) regarded as
"tentative" their calibration between the Sergipe Basin ammonites and the standard international scheme.
Although late Aptian planktonic foraminifera have been reported in the South Atlantic in scattered occurrences (Cear Basin, Della F vera et al., 1984; Deep Sea Drilling Project - DSDP Leg 40, Sites 363-364, Kwanza Basin, Caron, 1978) between 1980 and 2000, many studies in the CSA basins and also some in the ESA basins involving this protist group attributed an Albian age to the post-salt carbonate section (e.g., Azevedo et al., 1987; Koutsoukos, 1987; Viviers, 1987; Viviers & Beurlen, 1987; Dias-Brito et al., 1990; Azevedo, 2001). This concept ended up enshrined in the stratigraphic charts of these basins.
However, in important work involving cores and outcrops of the Sergipe Basin, Koutsoukos (1989, 1992) identified zones of planktonic foraminifera attributed to the Aptian in the basal portion of carbonates covering evaporites of the Ibura Event: the composite zones Globigerinelloides barri-Hedbergella (H.) gorbachikae and Globigerinelloides ex. gr. maridalensis-Hedbergella (H.) similis, both associated with the uppermost Aptian. Above these, the composite zone Globigerinelloides cushmani-Ticinella bejaouaensis and the acme zone Ticinella bejaouensis were considered to be Early Albian. One year later, Koutsoukos and Bengtson (1993) made the first attempt to integrate foraminifera and ammonite zonations for the Sergipe Basin, recognizing it as a provisional proposal for the Aptian-Albian interval as well.
Analyzing post-salt carbonates on the African side of the CSA, Caron
(1978) dated the lower section of Site 364 as
Aptian, based on recognition of the planktonic foraminiferal Globigerinelloides algerianus Zone and the Hedbergella gorbachikae-Hedbergella trocoidea composite zone
(Fig. 4
![]() ). This author placed the base of the Lower-Middle Albian in a distinct stratigraphic horizon defined by the FOs of Ticinella bejaouaensis, T. roberti, and T. primula, occurring alongside Globigerinelloides bentonensis, G. texomaensis,
Ticinella raynaudi, and Clavehedbergella simplicissima, species that are currently associated with younger ages
(Young et al., 2022b).
). This author placed the base of the Lower-Middle Albian in a distinct stratigraphic horizon defined by the FOs of Ticinella bejaouaensis, T. roberti, and T. primula, occurring alongside Globigerinelloides bentonensis, G. texomaensis,
Ticinella raynaudi, and Clavehedbergella simplicissima, species that are currently associated with younger ages
(Young et al., 2022b).
|
|
Figure 4:
Biochronostratigraphic interpretations for DSDP-364, based on different fossil groups, alongside results for TOC and δ13Ccarb
(Supplementary data). |
For calcareous nannofossils in the same section, Proto-Decima et al. (1978) noted the FOs of Parhabdolithus angustus (= Rhagodiscus angustus), Hayesites albiensis, Tranolithus orionatus, and Prediscosphaera cretacea (= P. columnata) in the G. algerianus Zone. They identified the P. angustus and P. cretacea zones as being Early Albian and Early-Middle Albian in age in stratigraphic levels that Caron (1978) had considered early Aptian. Further adding to the controversy, Proto-Decima et al. (1978) also observed that the FO of Podorhabdus albianus (= Axopodorhabdus biramiculatus), another typical Albian species, was observed in the composite zone Hedbergella gorbachikae-Hedbergella trocoidea, which Caron (1978) had associated with the late Aptian.
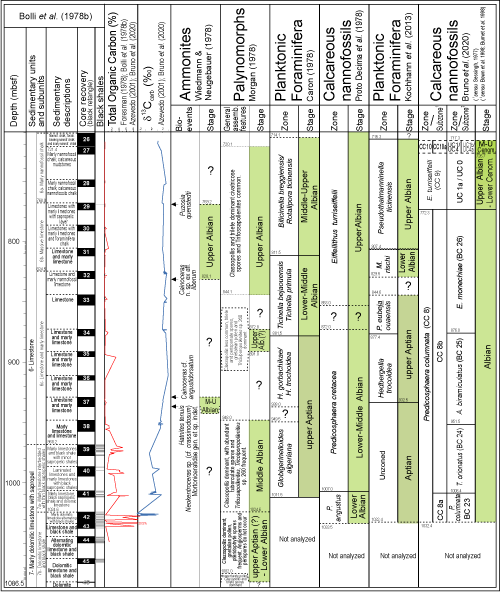
Conflicting biochronostratigraphies were also observed at Site 363, Leg 40 - DSDP between planktonic foraminifera and calcareous nannofossil events, the former suggesting Aptian ages and the Early-Middle Albian ones (Caron, 1978; Proto-Decima et al., 1978). During the integration of biochronostratigraphic data from sites 363 and 364, an Early Albian age was indicated for the carbonates at the base of both sections via better adjustment of the calcareous nannofossils with the classical zonation observed in Southern France (Bolli, 1978; Bolli et al., 1978a, 1978b).
With the discovery by Dias-Brito (1985) of the equivalence of the semiquantitative distribution of pithonellids from Site 363 and those observed in the Campos Basin, the Albian interpretation adopted by Bolli
(1978) became the reference for the southeastern and eastern Brazilian margin
(Fig. 5
![]() ). A few years later, Dias-Brito et al. (1990) consolidated a biostratigraphic zonation using data from the different planktonic fossils found throughout the thick post-salt interval continuously sampled in the core of the 3-BO-3-RJS well from the Campos Basin. The carbonate section was subdivided into a lower unit of Early to Middle Albian age and a younger one dated as
Late Albian. Finally, Dias-Brito
(2000) published a comprehensive study on the worldwide spatial and stratigraphic distribution of pithonellids in the Cretaceous, where he demonstrated that several species found in the CSA did indeed first appear in the
Albian.
). A few years later, Dias-Brito et al. (1990) consolidated a biostratigraphic zonation using data from the different planktonic fossils found throughout the thick post-salt interval continuously sampled in the core of the 3-BO-3-RJS well from the Campos Basin. The carbonate section was subdivided into a lower unit of Early to Middle Albian age and a younger one dated as
Late Albian. Finally, Dias-Brito
(2000) published a comprehensive study on the worldwide spatial and stratigraphic distribution of pithonellids in the Cretaceous, where he demonstrated that several species found in the CSA did indeed first appear in the
Albian.
|
|
Figure 5:
Cored carbonate section of the 3-BO-3-RJS well, with the distribution record for different fossil groups. In Tedeschi et al. (2017)' and Sanjin s et al. (2022)' papers this well was designated BO-3 and CB-01, respectively. The geophysical profile and parastratigraphic data are from Azevedo
(2001). The depths were recorded with reference to the
drilling floor ship level (+12m), to standardize the set of information. |
Dias-Brito (1994) also highlighted the affinity that many of the post-salt carbonate facies had with those described in Albian sections from the Gulf of Mexico with emphasis on the presence of Colomiella recta and C. mexicana in well 1-SPS-6 in the Santos Basin
(Fig. 1
![]() ). These taxa do not yet have well-defined ranges but are frequently considered to be markers for the end of the Aptian and beginning of the Albian, an interpretation adopted in the biochronostratigraphy for the basins of the southeastern margin of Brazil using colomiellids and pithonellids (Dias-Brito,
1999). More recently, with the support of isotopic data of 87Sr/86Sr, Caetano-Filho et al. (2017) reached the same conclusion for the chronostratigraphic positioning of C. recta in a distal section of the Campos Basin, where this species was also observed in strata immediately above the salt layer.
). These taxa do not yet have well-defined ranges but are frequently considered to be markers for the end of the Aptian and beginning of the Albian, an interpretation adopted in the biochronostratigraphy for the basins of the southeastern margin of Brazil using colomiellids and pithonellids (Dias-Brito,
1999). More recently, with the support of isotopic data of 87Sr/86Sr, Caetano-Filho et al. (2017) reached the same conclusion for the chronostratigraphic positioning of C. recta in a distal section of the Campos Basin, where this species was also observed in strata immediately above the salt layer.
Dupont (1996) studied planktonic foraminifera and other microfossils from dozens of wells drilled in the Gabon Basin
(Fig. 1
![]() ) and tentatively associated the Hedbergella cf. sigali - Globigerinelloides aff. maridalensis composite subzone and the lower portion of the Hedbergella occulta subzone with the Aptian. The presence of Colomiella recta also contributed to this chronostratigraphic interpretation, with the author also highlighting the affinity among fossil assemblages from Gabon, the Gulf of Mexico and the Sergipe Basin.
) and tentatively associated the Hedbergella cf. sigali - Globigerinelloides aff. maridalensis composite subzone and the lower portion of the Hedbergella occulta subzone with the Aptian. The presence of Colomiella recta also contributed to this chronostratigraphic interpretation, with the author also highlighting the affinity among fossil assemblages from Gabon, the Gulf of Mexico and the Sergipe Basin.
Corroborating the observations of Caron (1978), Dupont (1996) noted the co-occurrence of Ticinella bejaouaensis, T. roberti, and T. primula. Currently, the taxa designated as T. bejaouensis and T. eubejaouaensis (or, later, Paraticinella eubejaouaensis), are considered synonymous with T. roberti and Paraticinella rohri, respectively, with both having distinct, incompatible amplitudes: P. rohri is associated with the latest Aptian, while the FO of T. roberti lies in the Early-Middle Albian and its Last Stratigraphic Occurrence (LO) in the Late Albian (Moullade et al., 2002; Premoli-Silva et al., 2009; Huber & Leckie, 2011; Ando et al., 2013; Kennedy et al., 2017). This nomenclatural problem resulted in the choice of the T. primula FO as the datum for the base of Middle Albian by some (e.g., Leckie, 1984; Caron, 1985; Br�h�ret et al., 1986; Sliter, 1989), and for other authors, to indicate the uppermost part of the Lower Albian (e.g., Luciani et al., 2004; Huber & Leckie, 2011; CGTS2016 - Ogg et al., 2016).
Recently, new planktonic foraminifera and calcareous nannofossil studies in the basal section at Site 364 by Kochhann et al. (2013) and Bruno et al. (2020), respectively, confirmed the incompatibility between these two fossil groups in light of the biochronostratigraphic patterns in the CGTS2020 and the GSSP-Alb.
Kochhann et al. (2013) identified the Hedbergella trocoidea Zone and Paraticinella eubejaouaensis (= P. rohri) Zone, both associated with the upper Aptian
(Fig. 4
![]() ). The top of the latter also coincides with the extinction of other long-lived species having large tests that marks the beginning of the biotic turnover recognized just before the end of the Aptian in the GSSP-Alb. According to Kochhann et al. (2013), the Microhedbergella rischi Zone and Pseudothalmanninella ticinensis Zone represent strata from the Lower and Upper Albian, respectively, with an expressive gap indicated by the absence of the Middle Albian. Approximately 13 m of the drill core were not recovered, thereby impeding possible identification of the Microhedbergella miniglobularis and M. renilaevis zones or of other biotic events associated with the turnover, which could have better defined the Aptian-Albian boundary, according to the GSSP-Alb.
). The top of the latter also coincides with the extinction of other long-lived species having large tests that marks the beginning of the biotic turnover recognized just before the end of the Aptian in the GSSP-Alb. According to Kochhann et al. (2013), the Microhedbergella rischi Zone and Pseudothalmanninella ticinensis Zone represent strata from the Lower and Upper Albian, respectively, with an expressive gap indicated by the absence of the Middle Albian. Approximately 13 m of the drill core were not recovered, thereby impeding possible identification of the Microhedbergella miniglobularis and M. renilaevis zones or of other biotic events associated with the turnover, which could have better defined the Aptian-Albian boundary, according to the GSSP-Alb.
Kochhann et al. (2013) indicated the FO of Paraticinella eubejaouaensis four meters above the level at which Caron
(1978) observed the FO of T. bejaouensis, T. roberti, T. primula, and other taxa attributed to the
Middle-Late Albian
(Fig. 4
![]() ). Kochhann et al. (2013) revised the taxonomic description of T. primula, considering it to be closer to that of T. madecassiana (Microhedbergella rischi trans. madecassiana). In addition, they stated that
"Caron's figures of T. cf. T. roberti and T. praeticinensis do not yield a typical
'ticinellid wall structure', being [t]herein considered Muricohedbergella astrepta and M. praelibyca, respectively". They did not, however, discuss the taxa G. bentonensis, G. texomaensis, T. raynaudi, and C. simplicissima described by Caron
(1978), also associated with a Middle-Upper Albian, which they would be present in the interval in which Paraticinella eubejaouaensis occurs.
). Kochhann et al. (2013) revised the taxonomic description of T. primula, considering it to be closer to that of T. madecassiana (Microhedbergella rischi trans. madecassiana). In addition, they stated that
"Caron's figures of T. cf. T. roberti and T. praeticinensis do not yield a typical
'ticinellid wall structure', being [t]herein considered Muricohedbergella astrepta and M. praelibyca, respectively". They did not, however, discuss the taxa G. bentonensis, G. texomaensis, T. raynaudi, and C. simplicissima described by Caron
(1978), also associated with a Middle-Upper Albian, which they would be present in the interval in which Paraticinella eubejaouaensis occurs.
Studying calcareous nannofossils, Bruno et al. (2020) identified in core 42, lower portion of Site 364, the circular morphotype of Prediscosphaera columnata
(Fig. 4
![]() ; Supplementary data, no. 2). The presence of all the biozones described by Bown et al. (1998) characterizing the Albian (BC24 to BC27 zones) suggests that continuous sedimentation took place during this time interval, which contradicts the hypothesis of the hiatus described by Kochhann et al. (2013). The extinctions of large planktonic foraminifera at the top of the P. eubejaouaensis Zone are coeval to the Eiffellithus monechiae (BC26) Zone, whose base coincides with the beginning of the
Late Albian, according to the CGTS2016 (GTS2020 did not indicate the limits of the Albian substages). Thus, once more, while studies with foraminifera classify the interval as Aptian, calcareous nannofossils point to an Albian age.
; Supplementary data, no. 2). The presence of all the biozones described by Bown et al. (1998) characterizing the Albian (BC24 to BC27 zones) suggests that continuous sedimentation took place during this time interval, which contradicts the hypothesis of the hiatus described by Kochhann et al. (2013). The extinctions of large planktonic foraminifera at the top of the P. eubejaouaensis Zone are coeval to the Eiffellithus monechiae (BC26) Zone, whose base coincides with the beginning of the
Late Albian, according to the CGTS2016 (GTS2020 did not indicate the limits of the Albian substages). Thus, once more, while studies with foraminifera classify the interval as Aptian, calcareous nannofossils point to an Albian age.
As commented before, since 1990, the cored section of the 3-BO-3-RJS well has become the biochronostratigraphic reference for the basins of the southeastern margin of the CSA for studies on associations of planktonic foraminifers, calcareous nannofossils, pithonellids. As it could not be otherwise, there was object of studies by
Tedeschi et al. (2017) and Sanjin s et al. (2022). Tedeschi et al. (2017) were the first to propose a zonation scheme following the taxonomic-phylogenetic criteria of Huber
and Leckie
(2011) for a well drilled on the Brazilian margin. Based on the presence of the Globigerinelloides ferreolensis-G. algerianus zones, these authors dated a major part of a thick interval recovered from core CB-3 (the nickname adopted by
Tedeschi et al.,
2017, and Azevedo, 2001, to 3-BO-3-RJS) as late Aptian
(Fig. 5
![]() ). Below this, the presence of Favusella ex. gr. washitensis and Lenticulina ex. gr. nodosa was considered as indicative of an Aptian age or older. Lacking marker species for the uppermost part of the well, Tedeschi et al. (2017) used isotopic correlations to attribute an Aptian age to almost the entirety of these post-salt carbonates.
). Below this, the presence of Favusella ex. gr. washitensis and Lenticulina ex. gr. nodosa was considered as indicative of an Aptian age or older. Lacking marker species for the uppermost part of the well, Tedeschi et al. (2017) used isotopic correlations to attribute an Aptian age to almost the entirety of these post-salt carbonates.
Sanjin s et al. (2022) analyzed this section, which they designated as
CB-01 [*], in great detail (Fig. 5
![]() ), where they described an association of planktonic foraminifera, reproducing what they observed in a wide set of wells in the Santos, Campos, and
Esp�rito Santo basins. Above the salt, following an unspecified interval, they identified the Globigerinelloides ferreolensis-G. algerianus and Hedbergella infracretacea-Paraticinella rohri composite zones, formally defined by the LOs of these species or of coeval species
(Fig. 5
), where they described an association of planktonic foraminifera, reproducing what they observed in a wide set of wells in the Santos, Campos, and
Esp�rito Santo basins. Above the salt, following an unspecified interval, they identified the Globigerinelloides ferreolensis-G. algerianus and Hedbergella infracretacea-Paraticinella rohri composite zones, formally defined by the LOs of these species or of coeval species
(Fig. 5
![]() ). The top of both zones are located in the uppermost section of the post-salt carbonates, characterized by the dominance of pelagic and hemipelagic sediments that until then were exclusively associated with the Upper Albian. In the stratigraphic charts of the Santos and Campos basins, these rocks comprise the Itanha m and Outeiro formations, respectively (Moreira et al.,
2007; Winter et al., 2007).
). The top of both zones are located in the uppermost section of the post-salt carbonates, characterized by the dominance of pelagic and hemipelagic sediments that until then were exclusively associated with the Upper Albian. In the stratigraphic charts of the Santos and Campos basins, these rocks comprise the Itanha m and Outeiro formations, respectively (Moreira et al.,
2007; Winter et al., 2007).
According to Sanjin s et al. (2022), a mixed Aptian-Albian fauna was registered "immediately above the Aptian section that could be related to rework[ing] or, eventually, misidentification. Depending on new research or integrations with [other] (bio)stratigraphic methods, it could represent an unconformity or even a condensed section. Due to this undetermined interval, the P. rohri Zone is probably not complete, and the Lower-Middle Albian biozones were not characterized". Therefore, to date, the species that define the Microhedebergella miniglobularis and Microhedbergella renilaevis zones have not been recorded in the southeastern basins of the CSA. This regional unconformity, which would compartmentalize the K70 Sequence encompassing these fine carbonates, has not been recognized in the stratigraphic charts of the Santos and Campos basins (Moreira et al., 2007, and Winter et al., 2007, respectively).
As in Caron (1978), Dupont (1996), and Kochhann et al. (2013), the paper by Sanjin s et al. (2022) records a high number of species classified as conferatum (cf.), affinis (aff.) or doubtful (?). This supports the idea that perhaps conditions in the primitive CSA, a restricted sea with lagoonal circulation (Azevedo, 2001, 2004) favors morphological variation and greater longevity for the species that came to occupy that environmental niche. At any rate, Sanjin s et al. 's (op cit.) observation of coexisting species having seemingly incompatible ranges in core samples from CB-01 (=3-BO-3-RJS), such as Hedbergella tuschepsensis or H. mitra versus H. gorbacikae or H. trochoidea, attests to the potential that CSA sections hold for contributing to the improvement of the geological range of many planktonic foraminifera.
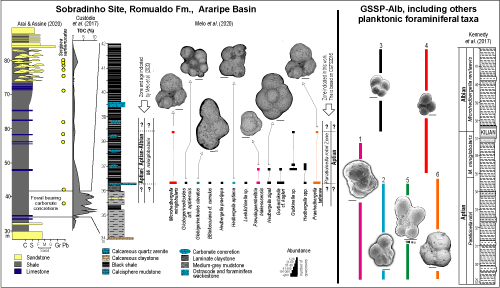
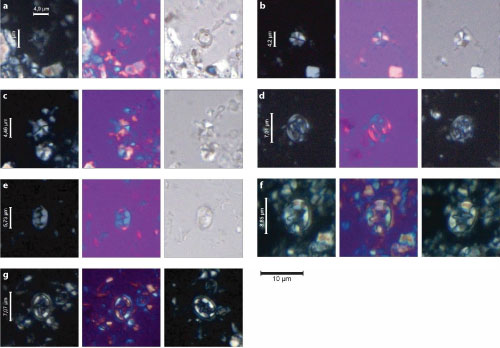
It is important to mention the remarkable discovery of a planktonic foraminifera assemblage in the Romualdo Formation at the Sobradinho Site, Araripe Basin (Melo et al.,
2020), a Cretaceous Konservat-Lagerst tten
[**]
(Figs. 1
![]() - 2
- 2
![]() ,
6
,
6
![]() ). The Romualdo Formation overlies thin evaporites of the Ipubi Formation related to the thick salt packets of the Ibura Event (Asmus & Campos,
1983; Assine,
2007).
). The Romualdo Formation overlies thin evaporites of the Ipubi Formation related to the thick salt packets of the Ibura Event (Asmus & Campos,
1983; Assine,
2007).
At the Sobradinho Site, almost all species have their LOs associated with the late Aptian Paraticinella rohri Zone. However, Melo et al. (2020) identified Microhedbergella miniglobularis in the assemblages, a species ranging from latest Aptian to Early Albian. The creates a paradox, considering that in the GSSP-Alb, M. miniglobularis does not coexist with P. rohri and other large species whose LOs define the P. rohri Zone
(Fig. 3
![]() ). Reworking and redeposition of these large foraminifera into a younger stratigraphic level may explain this observation. This, however, is unlikely, given the anoxic/dysoxic environment, not the least propitious to sedimentary remobilization, in which these planktonic fossils were deposited. Alternatively, the FO of M. miniglobularis may well appear earlier at Sobradinho than indicated in the GSSP-Alb. The fact that this same stratigraphic inversion is also recorded at Site 511 (Huber & Leckie,
2011) and Poggio le Guaine, Italy (Coccioni et al.,
2014) is coherent with this interpretation.
). Reworking and redeposition of these large foraminifera into a younger stratigraphic level may explain this observation. This, however, is unlikely, given the anoxic/dysoxic environment, not the least propitious to sedimentary remobilization, in which these planktonic fossils were deposited. Alternatively, the FO of M. miniglobularis may well appear earlier at Sobradinho than indicated in the GSSP-Alb. The fact that this same stratigraphic inversion is also recorded at Site 511 (Huber & Leckie,
2011) and Poggio le Guaine, Italy (Coccioni et al.,
2014) is coherent with this interpretation.
Melo et al. (2020) found M. miniglobularis in great abundance at the Sobradinho Site, even though their illustrations do not allow for indisputable differentiation from other, older taxa, for example, the Praehedbergellidae. In this respect, it is important to note that Praehedbergella and Microhedbergella are considered as parts of a distantly related homeomorphic lineage. Furthermore, Sobradinho specimens are larger than expected for M. miniglobularis and exhibit great morphological affinity to Praehedbergella tatianae, whose last occurrence was near the end of the Aptian (Banner & Desai, 1988). Therefore, in the present study, we consider the entire planktonic foraminifera assemblage described by Melo et al. (2020) at the Sobradinho Site as belonging to the Paraticinella rohri Zone.
Another important study has been carried out in core section SER-03 in the onshore portion of the Sergipe Basin (Fauth et al., 2022). For the first time in the CSA, FO of Microhedbergella renilaevisis is indicated accompanied by an ideal succession of Lower Albian planktonic foraminifera- the FOs of M. rischi, Ticinella madecassiana, and T. primula. Below the M. renilaevis datum, there is an interval with low δ13Ccarb values preceded by a strong positive excursion, where occurs the FO of Prediscosphaera columnata ? (subcircular). Fauth et al. (2022) also described the FOs of Braarudosphaera batiliformis, B. africana, B. regularis, and B. pseudobatiliformis, and Nannoconus quadriangulus apertus, N. quadriangulus quadriangulus and N. troelsenii below to the M. renilaevis datum.
Fauth et al. (2022) indicated an age of latest Aptian to Early Albian for the SER-03 section, based on astronomical cyclicity analysis, using these planktonic foraminifera and calcareous nannofossils as references for time calibration. Furthermore, we were not identified any exclusive Aptian planktonic foraminiferal taxon below FO of M. renilaevisis.
The list of species of other fossil groups that have their LOs close to the Aptian-Albian boundary presented by Lima et al. (2018) created new controversies. The calcareous nannofossils Braarudosphaera batilliformis, B. pseudobatilliformis, and Nannoconus troelsenii, considered to be restricted to the Aptian by Alves et al. (2017), were observed in Albian strata at Site 364 (Bruno et al., 2020) and in the SER-03 section Fauth et al. (2022). Without detailing their stratigraphic positions, Alves et al. (op cit.) also reported the presence of Nannoconus bucheri, N. circularis, N. globulus, Micrantholithus hoschulzii, and Rhagodiscus achylostaurion, all of which, except for the last, have amplitudes not extending beyond the Aptian (Young et al., 2022a). Likewise, other species considered Aptian by Lima et al. (2018) also reach the Albian, such as Colomiella recta and C. mexicana and the crinoid Microcalamoides diversus (Dias-Brito, 1995, 1999; Dias-Brito & Ferr , 2001; Le n-Francisco et al., 2013; Madhavaraju et al., 2013; Fekete et al., 2017). Additional taxa with poorly defined ranges were also added to this list (e.g., Cadosina ex gr. semiradiata, and Colomisphaera heliosphaera (Young et al., 2022a; https://paleobotany.ru/ accessed: 2022/12/06).
Pollen, spores, dinoflagellate cysts, and other palynomorphs are all found in strata of local Alagoas age, which are tentatively correlated with the late Aptian-Early Albian interval (e.g., Schaller, 1969; Antunes et al., 2018). These deposits are transition from a continental to a marine environment in the CSA, ESA, and BNE basins. The palynozones subdividing the sedimentary record of this period in the inner Brazilian basins and in the marginal basins of northeastern and equatorial Brazil are distinct from those of the marginal basins of southeastern and eastern Brazil. Regali & Viana (1989), Arai et al. (1989), Dino et al. (1999), and Azevedo (2001) provide a summary of this discussion, including information on basins from the African margin.
The most relevant taxa for correlations and for attempts to delineate the Aptian-Albian boundary are elaterate pollen (particularly Elateropollenites jardinei), the gymnosperm pollen Sergipea variverrucata, and the dinoflagellates that make up the Subtilisphaera spp. ecozone.
Elaterate pollen first appeared at the end of the Aptian and became widely dispersed during the Albian (Dino et al., 1999). E. jardinei is the taxon the range of which is best calibrated with international chronostratigraphy; its FO has been observed in South American and African basins postdating the beginning of the Albian (Atta-Peters, 2013). As discussed by Dias-Brito et al. (1987), E. jardinei is not mentioned among the palynomorphs in the type-section of the local Alagoas stage, which was defined in a subsurface section of the Sergipe Basin (Schaller, 1969).
The LO of Sergipea variverrucata, an endemic species described by Regali et al. (1974), marks the top of the zone of the same name that makes up part of the Alagoas stage. For Regali and Viana (1989), this bioevent occurred in the late Aptian and was followed, upwards, by the FO of E. jardinei and Cretaceiporites polygonalis, which mark the beginning of the Albian. Arai et al. (1989), in a different interpretation, place the LO of S. variverrucata at the Aptian-Albian boundary. Above this horizon, the FO of E. jardinei would already lie within Lower Albian strata.
The correlation of S. variverrucata with planktonic foraminifera in marine deposits of the Sergipe Basin (Koutsoukos,
1989) allowed it to be associated with the later portion of the Aptian
(Regali & Silva Santos, 1999). Nevertheless, the recent identification of the Paraticinella rohri Zone at the Sobradinho Site, Araripe Basin (Melo et al.,
2020), impacts the interpretations of Regali
and Viana (1989) and reinforces the chronostratigraphic solution proposed by Arai et al. (1989). As demonstrated by Arai
and Assine (2020), at Sobradinho S. variverrucata extends above the limit of the P. rohri Zone
(Fig. 6
![]() ), noting that this biostratigraphic unit occurs shortly before the FO of Microhedbergella renilaevis, the GSSP-Alb
datum.
), noting that this biostratigraphic unit occurs shortly before the FO of Microhedbergella renilaevis, the GSSP-Alb
datum.
|
|
Figure 6:
Stratigraphic data for the Romualdo Formation, Sobradinho Site, Araripe Basin, and for Col de Pr
Guittard, GSSP-Alb, with modifications. For the Araripe Basin are displayed the range of Sergipea variverrucata (Arai & Assine,
2020), the TOC profile (Cust dio et al.,
2017), and the planktonic foraminifera succession (Melo et al.,
2020). For GSSP-Alb are shown some of the planktonic foraminifera registered by
Kennedy et al., 2014): Pseudoguembelitria blakenosensis (1), Hedbergella aptiana (2), Microhedbergella renilaevis (3) and M. miniglobularis (4). The ranges of the taxa Globigerinelloides clavatus (5) and Praehedbergella tatianae (6) are from the Mikrotax Catalogue
(URL: https://mikrotax.org/pforams/index.php?id=110128, accessed: 2021/01/31) and Banner
and Desai (1988), respectively. The scale bar = 30 m, except for G. clavatus, which represents 100 m. To the right, in the central column, the specimen classified as Microhedbergella miniglobularis by Melo et al. (2020) is represented under the taxonomic designation Praehedbergella tatianae. |
The Subtilisphaera spp. ecozone was first described in the Cear Basin by Regali (1989) and is notable for the high number of cysts it contains members of this dinoflagellate genus. The rocks containing these cysts were considered late Aptian in age. Since then, this ecozone has been observed in many other basins, particularly in the BNE, including as far south as the Aptian-Albian of the Esp�rito Santos Basin (Michels et al., 2018). This evidence of dinoflagellate blooms lies either directly upon or slightly above the Ibura evaporites, although in some basins, Subtilisphaera spp. blooms are also noted in younger, Albian strata (Arai, 2009, 2014; Carvalho et al., 2016; Michels et al., 2018). This ecozone has also been found in Senegal, Morocco, and Venezuela (Arai, 2014, and papers cited therein).
These anomalous, episodic Subtilisphaera blooms have been associated with restricted shallow marine and mixohaline environments (e.g., Regali, 1989; Arai et al., 1994; Carvalho et al., 2016; Helmond et al., 2014). According to Carvalho et al. (2016), the phenomenon would have been related to a "humid period, increased freshwater input, a high rate of nutrient supply, and the formation of a pycnocline separating slightly less saline surface waters from higher saline deeper waters. These conditions would have impeded vertical circulation". In the drill cores from the Sergipe Basin studied by these authors, the two intervals in which Subtilisphaera dominate exhibits moderate to high ratios of amorphous organic matter versus phytoclasts, particularly the older one, which led these authors to associate these conditions with restricted marine environments and reduced terrigenous influence.
This accords well with the occurrence of the Subtilisphaera ecozone immediately or slightly above the evaporitic layers. Hence, it can be considered a good marker for the end of the evaporite event apparently caused by the arrival of lower salinity waters carrying the nutrients that allowed these dinoflagellate blooms and led to stratification of the water body. Therefore, more than just representing an environmental anomaly, this ecozone also has chronostratigraphic value for not only basins in the CSA, ESA, and BNE, but, potentially, for basins on the margins of southern Tethys as well.
In the basins of the eastern and southeastern Brazilian margins, other palynomorphs subdivide the post-salt carbonate section and the overlying marls and shales. These are the classic biozones Cyclopsiella sp. (Early-Middle Albian), Hystrichosphaeridum maranhensis and Din D/Din H
(Late Albian), and Classopollis major (Cenomanian), all defined by the LO of their respective homonyms. Their stratigraphic positions were indicated by considering the well-defined ranges of other fossil groups observed in the cored section of 3-BO-3-RJS (Uesugui, 1976, fide Arai,
1992; Dias-Brito et al.,
1990; Dias-Brito, 1995; Arai et al.,
1996; Azevedo,
2001). Palynological analyses made by Morgan
(1978) at Site 364 also contributed to establishing the age of these post-salt limestones as Albian
(Fig. 4
![]() ).
).
Therefore, with current palynological knowledge it is not possible to assert with certainty that marine Aptian strata are present above the evaporites on both margins of the meridian portion of the CSA, ESA, and BNE. Inferences of an Aptian age for the post-salt limestones are mainly a corollary of the correlation between palynozones and the planktonic foraminiferal biounits identified by Koutsoukos (1989). Thus, palynomorphs are inserted as coadjuvants within the controversies regarding the chronostratigraphic attributions of these protists and other fossil groups that have internationally recognized ranges.
The option of indicating lithological horizons to define chronostratigraphic boundaries is incorporated in the North American Commission on Stratigraphic Nomenclature - NASC
(2005). A relevant example of here is the Kilian Level, an organic-rich shale comprising the main lithological
datum associated with the GSSP-Alb
(Fig. 3
![]() ).
).
According to Tissot et al. (1980), Herbin et al. (1987), and Beglinger et al. (2012), there is but a single notable organic-rich layer in the basins of the CSA, ESA, and BNE. Exceptions to this occur in the Sergipe-Alagoas, Gabon, and Kwanza basins, where other similar layers have been identified in the Albian (Beglinger, op cit.). At Site 364, Kwanza Basin, the most organic-rich layer, with more than 20% Total Organic Carbon (TOC), occurs in the lower portion of the drill core, where black shales intercalated with dolomites overlie the Ibura evaporites
(Fig. 4
![]() ). TOC values greater than 2% are found in recurring layers in the overlying first 50 meters, above which TOC values become extremely low throughout the Albian (Kumar et al.,
1977; Azevedo,
2001; Bruno et al.,
2020). In the Campos and Santos basins, rocks with TOC values greater than 2% in the post-salt limestones are rare, other than those that immediately overlying the Ibura salts (Azevedo,
2001).
). TOC values greater than 2% are found in recurring layers in the overlying first 50 meters, above which TOC values become extremely low throughout the Albian (Kumar et al.,
1977; Azevedo,
2001; Bruno et al.,
2020). In the Campos and Santos basins, rocks with TOC values greater than 2% in the post-salt limestones are rare, other than those that immediately overlying the Ibura salts (Azevedo,
2001).
For chronocorrelation purposes, the most efficient lithological datum for basins in the CSA, ESA, and BNE is the base of the Ibura Event (Fig. 2). The age indicated for this stratigraphic horizon is related to the definition of the Curumim Formation, Pelotas Basin (Dias et al.,
1994). This lithostratigraphic unit is formed by trachyandesites, covered discordantly by both Portobelo carbonates and Ariri evaporites
(Figs. 2
![]() ,
7
,
7
![]() ). Its type-section is in the 1-SCS-2 well. These volcanic rocks are attributed to the Alagoas Stage on the basis of
Ar/Ar dating of a sample from the 1-SCS-1 well at the northern end of the Pelotas Basin (Mizusaki, 1993,
fide Dias et al., 1994). In both 1-SCS-1 and
1-SCS-2, there is no salt layer, and the Portobelo carbonates cover the volcanics discordantly. The complete stratigraphic succession defining the stratigraphic relationships between the three lithological units occurs in well 1-SCS-3B, chosen by Dias et al. (1994) as type-sections for the Ariri (evaporites) and Portobelo (carbonates) formations. This succession is covered by fine-grained sediments exhibiting a greater terrigenous influence that defines the Atl ntida Formation, chronoequivalent to pelagic and hemipelagic carbonates in which Sanjin s et al. (2022) recorded the diverse fauna of planktonic foraminifera assigned to the Aptian in the Santos, Campos, and
Esp�rito Santo basins.
). Its type-section is in the 1-SCS-2 well. These volcanic rocks are attributed to the Alagoas Stage on the basis of
Ar/Ar dating of a sample from the 1-SCS-1 well at the northern end of the Pelotas Basin (Mizusaki, 1993,
fide Dias et al., 1994). In both 1-SCS-1 and
1-SCS-2, there is no salt layer, and the Portobelo carbonates cover the volcanics discordantly. The complete stratigraphic succession defining the stratigraphic relationships between the three lithological units occurs in well 1-SCS-3B, chosen by Dias et al. (1994) as type-sections for the Ariri (evaporites) and Portobelo (carbonates) formations. This succession is covered by fine-grained sediments exhibiting a greater terrigenous influence that defines the Atl ntida Formation, chronoequivalent to pelagic and hemipelagic carbonates in which Sanjin s et al. (2022) recorded the diverse fauna of planktonic foraminifera assigned to the Aptian in the Santos, Campos, and
Esp�rito Santo basins.
Demercian (1996) showed that both the evaporitic and carbonate layers thin towards the volcanic high on the southern edge of the S�o Paulo Plateau and its extension to the continent, the Florian polis High. Based on this, Dias (1998) pointed out the onlap character of the base of the evaporitic package onto the volcanic high making up the SAMB, even noting the difficulty of seismic resolution in layers of anhydrite, more frequent in the limits of the occurrence of the salt over the Florian polis High. Although lacking direct evidence, this information contains the basic elements that led Dias (1998) to estimate an age for evaporite deposition between 113 and 112 Ma, a little younger than that of the trachyandesite in 1-SCS-1-SC. In 2007, Petrobras updated the stratigraphic maps of all Brazilian basins, establishing the age obtained by Mizusaki (1993, fide Dias et al., 1994) and published by Dias et al. (1994) as a geochronological reference for the salt base in the CSA basins (e.g., Bueno et al., 2007; Fran a et al., 2007; Moreira et al., 2007; Winter et al., 2007).
Szatmari and Milani
(2016) describe the particularities and succession of Pre-Salt volcanic events in the offshore portion of the basins of the southeast margin of the CSA
(Fig. 7
![]() ). They cite an age of
115.7 1 Ma obtained by the Ar-Ar method for a thick basalt flood sampled in the Santos Basin
(Gomes et al., 2016 -
Petrobras Internal Report). Keeping in mind the limited number of radiometric age determinations,
Szatmari and Milani (2016) proposed that
"volcanic activity in the CSA, as in the North Atlantic (Keen et al.,
2012), started almost simultaneously, about 10 Ma after the start of the rifting process; it peaked at about 135 to 130 Ma, and continued alternating with sedimentation until about 113 Ma, shortly before the beginning of the salt deposition". Recently, Szatmari et al. (2021) reaffirmed the 113 Ma Ar-Ar age for the volcanic rocks below the evaporites of the Santos Basin. However, they also report a
110.64 0.34 Ma age for an intra-salt unconformity at the carnallite/sylvinite contact in the Sergipe Basin, suggesting that salt deposition took longer than previously thought.
Figure 8
). They cite an age of
115.7 1 Ma obtained by the Ar-Ar method for a thick basalt flood sampled in the Santos Basin
(Gomes et al., 2016 -
Petrobras Internal Report). Keeping in mind the limited number of radiometric age determinations,
Szatmari and Milani (2016) proposed that
"volcanic activity in the CSA, as in the North Atlantic (Keen et al.,
2012), started almost simultaneously, about 10 Ma after the start of the rifting process; it peaked at about 135 to 130 Ma, and continued alternating with sedimentation until about 113 Ma, shortly before the beginning of the salt deposition". Recently, Szatmari et al. (2021) reaffirmed the 113 Ma Ar-Ar age for the volcanic rocks below the evaporites of the Santos Basin. However, they also report a
110.64 0.34 Ma age for an intra-salt unconformity at the carnallite/sylvinite contact in the Sergipe Basin, suggesting that salt deposition took longer than previously thought.
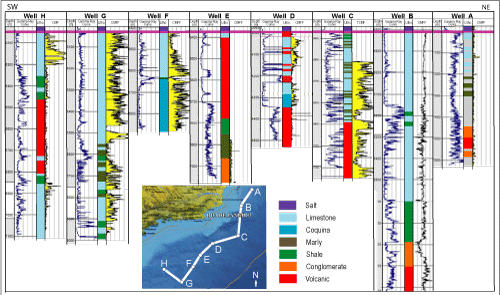
Figure 8
![]() shows the stacking of some of these volcanic events
identified in deep water wells drilled in the Santos and Campos basins.
shows the stacking of some of these volcanic events
identified in deep water wells drilled in the Santos and Campos basins.
|
|
Figure 7:
Schematic stacking of sedimentary succession and other geological parameters with respect to the
Pelotas, Santos, Campos and Kwanza basins. The most relevant stratigraphic data justify the discrepancies between the solution proposed by Sanjin s et al.
(2022) and the one offered in this work. Lithological and geophysical profiles are from the type-section of the
post-salt carbonate in well 1-RJS-135, Campos Basin (Rangel et al.,
1994). The parastratigraphic data are from Azevedo
(2001) and the ages from CGTS2020. |
|
|
Figure 8:
Well section illustrating the Pre-Salt reservoir from the Campos and Santos basins. Well D is notable for the presence of a volcanic layer overlain by the
Ibura Event (modified from Matias et al.,
2015). |
Lawson et al. (2022) recently offered further geochronological elements favoring an age assignment of 113 Ma to the base of the salt. Although focusing on the Pre-Salt carbonates (Barra Velha Fm, Santos Basin), these authors provide a good review of the complexity involved in dating the evaporites that cover them. They register that one "of the main challenges in developing a robust understanding of these basins is the total absence of reliable time markers to place any environmental constraints into temporal context. This has led to significant uncertainty on the age of key sequences within the stratigraphy. The Ariri Formation that represents the Santos Basin equivalent of the regional evaporite sequence, for example, has been proposed to be as young as 111 Ma based on an Ar-Ar age of sylvinite of 110.64 0.3 Ma from the Sergipe Basin (Szatmari et al., 2021). Further, evaporites unconformably overly volcanic rocks dated to 113.2 0.1 Ma in the northern Pelotas Basin (Dias et al., 1994; Davison, 2007). However, biostratigraphic data from the DSDP Site 364 suggest that the first marine deposition above these evaporites occurred during the late Aptian prior to 113 Ma (Kochhann et al., 2013). This has been further supported by the study of Sanjin s et al. (2022), who also report upper Aptian aged fauna in the stratigraphy immediately overlying the evaporites in numerous wells within the Santos, Campos, and Esp�rito Santo basins offshore Brazil. Alternatively, these evaporites have been proposed to be as old as 120-125 Ma and associated with the Aptian Ocean Anoxic Event (OAE) 1a (Tedeschi et al., 2017)".
Lawson et al. (2022) conclude that we
"have constrained the depositional age of the upper Barra Velha Fm to be
115.83
1.56 Ma through isotope dilution U-Pb dating. This age is consistent with overlapping laser ablation U-Pb ages of
114.46 4.72 Ma and 109.73 9.26 Ma from two separate carbonate platforms in the Santos Basin". These values would rule out in advance the age indication proposed by Tedeschi et al. (2017) for the evaporitic layer, of the order of 120 to 125 Ma, fundamentally supported by isotopic correlation. Considering the margin of error, the age of
115.83
1.56 Ma for the sediments of the upper portion of the Pre-Salt carbonates, but not their top, is more compatible with the
113.2 0.1 Ma measured in the trachyandesite of the 1 -SCS-1 than the value of
116.8 Ma (according to GTS2020) of the top of the Globigerinelloides ferreolensis-Globigerinelloides algerianus composite biozone (Sanjin s et al.,
2022). The top of this biozone is almost 1,000 m above the evaporite layer and it is estimating a time interval of the order of 6 Ma among these horizons. This value considers the minimum time suggested for deposition of the salt bed
(~0.5 Ma, according to Rodrigues et al., 2018) as well as the 5.5 Ma attributed to 55 short eccentricity cycles (~100 ka each one) recognized by Azevedo
(2001,
2005) in the Quissam Formation (41 cycles) and Outeiro Formation (14 cycles) in well 1-RJS-135, type-section of the Maca carbonates in the Campos Basin (Rangel et al.,
1984;
Fig. 7
![]() ).
).
Despite controversies over the age of the salt, the unique character of the Ibura evaporite led Antunes et al. (2018) suggest using the base of the salt as the datum to the Aptian-Albian transition for the CSA basins. It is arguably one of the most notorious lithological data in these basins, deposited over a short period of time with its base onlapping upon trachyandesite sampled in the SAMB, dated at 113.2 0.1 Ma. As already shown, support also comes from biostratigraphic successions involving other groups of fossils, such as pelagic calcispheres and calcareous nannofossils, which attest to the Albian age for post-salt carbonates. The recognition at Site 364 of the FO of Prediscosphaera columnata (circular) just above the salt and black shale layer permits associating these organic-rich accumulations with the Jacob Level, which by this interpretation would also be Early Albian age. It is always worth recalling that Prediscosphaera columnata was for a long time considered as a potential marker to define the base of the Albian.
An important additional argument derives from the fact that sea level remained low with only small-magnitude oscillations for almost 20 Myr, from the late Barremian to the late Aptian (Haq, 2014). At the end of the Albian this changed as the sea level rose rapidly to reach its apex, an episode of global eustatic elevation represented in CSA by the fine sediments in the upper portion of the post-salt carbonates, traditionally related to Late Albian in age (e.g., Dias-Brito & Azevedo, 1986; Spadini et al., 1988; Spadini & Pra a, 1989; Spadini, 1992).
Figure 7 shows a schematic sedimentary succession with the major data that justify the two quite different chronostratigraphic interpretations. Information on the evaporitic package and the carbonate section comes from the 1-RJS-135 well and is complemented by data from sections in the Pelotas, Santos, Campos, and Kwanza basins.
The papers by Tedeschi et al. (2017) and Sanjin s et al. (2022) show that, unlike previous thinking, it is not just the lower part of the post-salt carbonate section, but practically the entire section that would be Aptian in age according to more recent international biochronostratigraphic charts and GSSP-Alb criteria. In this scenario, only a thin Upper Albian section would have been preserved in the southeastern part of the CSA. Yet, even if doubts about the presence of putative Aptian planktonic foraminifera and even ammonites (according to the GSSP-Alb criteria) in the CSA, ESA, and BNE basins, are overcome, other new ones present themselves. For example, how does one explain the subversion in the successions of different fossil groups, or even among LO's of species of the same group, as in the case of the planktonic foraminifera? Or, the "incompatible" radiometric ages obtained by Mizusaki (1993, fide Dias et al., 1994), Szatmari et al. (2021) and Lawson et al. (2022)? Some speculative answers might include bioprovincialism in the South Atlantic; or an inappropriately old age indication for the GSSP-Alb, where a younger value should be considered? It is more important for the moment, however, to know if these "stratigraphic anomalies" also occur elsewhere on the globe.
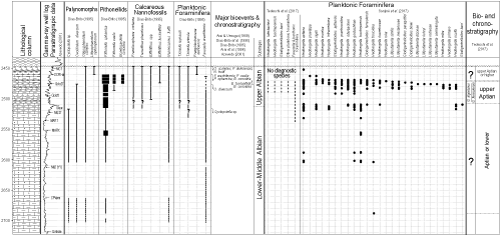
Analysis of 16 sedimentary sections from different regions of the world
[Only papers showing sections with the planktonic foraminifera reviewed according to the taxonomic and phylogenetic revisions of Huber and Leckie
(2011) were considered] allowed us to observe and evaluate relationships between biostratigraphic, chemostratigraphic and lithologic data regarding the latest Aptian-Early Albian
(Fig. 9
![]() ). The stratigraphic reference used for the dataset was the FO of Microhedbergella renilaevis, as well as, whenever possible, the position of the other events indicated in the
GTS2020. Among these, the LOs of large planktonic foraminifera (~Paraticinella rohri
Zone), indicative of the last portion of the Aptian, deserves to be highlighted. When conflicting geological information presented itself, biostratigraphic references were chosen to define stratigraphic positioning rather than other data, such as black shale beds or carbon isotope events. Supplementary data
inform the stratigraphic position and the studies supporting the succession in each section in
Figure 9
). The stratigraphic reference used for the dataset was the FO of Microhedbergella renilaevis, as well as, whenever possible, the position of the other events indicated in the
GTS2020. Among these, the LOs of large planktonic foraminifera (~Paraticinella rohri
Zone), indicative of the last portion of the Aptian, deserves to be highlighted. When conflicting geological information presented itself, biostratigraphic references were chosen to define stratigraphic positioning rather than other data, such as black shale beds or carbon isotope events. Supplementary data
inform the stratigraphic position and the studies supporting the succession in each section in
Figure 9
![]() .
.
|
|
Figure 9:
Biostratigraphic, chemostratigraphic and lithological events for the latest
Aptian-Early Albian interval. The columns to the left are from CGTS2020; the ones at the center are the 16 analyzed sections; to the right is an idealized section for the CSA, ESA and BNE encompassing the chronostratigraphic and geochronological limits suggested in this study. The highlighted black shales and δ13C excursions are limited to three OAE1b levels; the carbon isotopic data where tentatively differentiated among strong (2 ) and weak (between 1 and 2 ) oscillations. Bibliographic references and reference sections can be found in Supplementary data. |
The initial aim of indexing the stratigraphic relationships between different datasets was to evaluate the chronostratigraphic consistency of the FO of Prediscophaera columnata. Throughout the development of the numerous Geologic Time Scales (from the 90s to GTS2012), the FO of this species was always considered relevant, either as a secondary
datum, or as a reference to informally define the Aptian-Albian boundary. After Bown (in Kennedy et al., 2000) showed that P. columnata consisted of elliptical-subcircular, subcircular, and circular morphological categories, the idea that its FO was diachronous became the norm. The erratic distribution of these morphotypes points to the high degree of subjectivity in this taxonomic option, with a strong impact on biochronostratigraphic interpretations. Not even in neighboring outcrops, such as those in European sections
(Fig. 9
![]() , columns 3-7), can the synchronicity that was once attributed to the FO of P. columnata be observed (e.g., Br�h�ret et al., 1986; Hart et al.,
1996; Owen,
2002; Ogg et al., 2004, 2012).
, columns 3-7), can the synchronicity that was once attributed to the FO of P. columnata be observed (e.g., Br�h�ret et al., 1986; Hart et al.,
1996; Owen,
2002; Ogg et al., 2004, 2012).
On the other hand, if one groups these three morphotypes into one taxon, Prediscosphaera columnata
(sensu lato), it is possible to confirm the value of the FO of this species as a chronostratigraphic
datum, present in 11 of the 16 analyzed sections (Fig. 9
![]() , columns 4-10, 12-15). This morphotype group is part of the P. spinosa-P. columnata s.l. plexus, which has a wide, global distribution, with the FOs of these taxa being separated by the Jacob Level, as recognized in the GSSP-Alb (e.g., Herrle & Mutterlose,
2003; Kennedy et al.,
2017). P. columnata s.l. is found outside of this stratigraphic level only at Site 463
(Fig. 9
, columns 4-10, 12-15). This morphotype group is part of the P. spinosa-P. columnata s.l. plexus, which has a wide, global distribution, with the FOs of these taxa being separated by the Jacob Level, as recognized in the GSSP-Alb (e.g., Herrle & Mutterlose,
2003; Kennedy et al.,
2017). P. columnata s.l. is found outside of this stratigraphic level only at Site 463
(Fig. 9
![]() , column 1) and V�hrum
(Fig. 9
, column 1) and V�hrum
(Fig. 9
![]() , column 3). In the composite section for the southeastern Brazilian margin
(Fig. 9
, column 3). In the composite section for the southeastern Brazilian margin
(Fig. 9
![]() , column 16), it was recorded in well-core samples that recovered just part of the Albian, which did not allow precise identification of its full stratigraphic range (Dias-Brito et al.,
1990; Antunes, 1996;
Fig. 5
, column 16), it was recorded in well-core samples that recovered just part of the Albian, which did not allow precise identification of its full stratigraphic range (Dias-Brito et al.,
1990; Antunes, 1996;
Fig. 5
![]() ). A hiatus prevents the analysis of the FO of P. columnata s.l. at Site 763B, from the Ocean Drilling Program, ODP
(Fig. 9
). A hiatus prevents the analysis of the FO of P. columnata s.l. at Site 763B, from the Ocean Drilling Program, ODP
(Fig. 9
![]() , column 2), and there is no information about calcareous nannofossils for the Linderos section
(Fig. 9
, column 2), and there is no information about calcareous nannofossils for the Linderos section
(Fig. 9
![]() , column 11).
, column 11).
Worthy of note are the FOs of P. columnata s.l. in the V�hrum outcrop (Germany), in the composite section of the Vocontian Basin (France), in the Agadir-Essaouira Basin (Morocco), and at sites 463 (Pacific) and 364 (CSA). At Site 463
(Fig. 9
![]() , column 1) and in V�hrum
(Fig. 9
, column 1) and in V�hrum
(Fig. 9
![]() , column 3), P. columnata s.l. occurs at later stratigraphic levels than expected. The V�hrum outcrop was a candidate reference-section for the base of the Albian Stage (Owen,
2002). It has an ash layer dated as
113.1 0.3 Ma (Selby et al., 2009) that is today used to mark the beginning of the Albian in the GSSP-Alb. Mutterlose et al. (2003) identified the FO of P. columnata s.l. a few centimeters above this tuff, which defines its position in
Figure 9. It occurs in an assemblage containing few specimens, amid the low specific diversity of calcareous nannofossils in the Boreal region.
, column 3), P. columnata s.l. occurs at later stratigraphic levels than expected. The V�hrum outcrop was a candidate reference-section for the base of the Albian Stage (Owen,
2002). It has an ash layer dated as
113.1 0.3 Ma (Selby et al., 2009) that is today used to mark the beginning of the Albian in the GSSP-Alb. Mutterlose et al. (2003) identified the FO of P. columnata s.l. a few centimeters above this tuff, which defines its position in
Figure 9. It occurs in an assemblage containing few specimens, amid the low specific diversity of calcareous nannofossils in the Boreal region.
In the composite section for the Vocontian Basin described by Herrle & Mutterlose
(2003), the only P. columnata morphotype identified is circular and occurs slightly above the Jacob Level
(Fig. 9
![]() , column 6). Its stratigraphic position is distinct from those described by Kennedy et al. (2000) and Kennedy et al. (2014) for the Col de Pr� Guittard
(Fig. 9
, column 6). Its stratigraphic position is distinct from those described by Kennedy et al. (2000) and Kennedy et al. (2014) for the Col de Pr� Guittard
(Fig. 9
![]() , columns 5-7). In the Moroccan sections
(Fig. 9
, columns 5-7). In the Moroccan sections
(Fig. 9
![]() , column 8), Hassanein
(2016) did not observe any stratigraphic ordering between the FOs of the P. columnata s.l. varieties as described by Bown (in Kennedy et al., 2000). Finally, as already mentioned, Bruno et al. (2020) found the circular morphotype of this species in Core 42, Site 364, not much more than dozens meters above the inferred salt layer for the area
(Fig. 9
, column 8), Hassanein
(2016) did not observe any stratigraphic ordering between the FOs of the P. columnata s.l. varieties as described by Bown (in Kennedy et al., 2000). Finally, as already mentioned, Bruno et al. (2020) found the circular morphotype of this species in Core 42, Site 364, not much more than dozens meters above the inferred salt layer for the area
(Fig. 9
![]() , column 14). Towards the top of the section are the FOs of H. albiensis, T. orionatus, A. biramiculatus, E. monechiae, and E. turriseiffelli, which perfectly follows the characteristic stratigraphic succession of calcareous nannofossil data for the Albian (Bown et al.,
1998). Except the FO E. turriseiffelli, the other ones bioevents occur beneath the LO of the large planktonic foraminifera considered as being a reference for the upper portion of the Aptian (CGTS2020), thereby conflicting with the standard established for the GSSP-Alb
(Figs. 3
, column 14). Towards the top of the section are the FOs of H. albiensis, T. orionatus, A. biramiculatus, E. monechiae, and E. turriseiffelli, which perfectly follows the characteristic stratigraphic succession of calcareous nannofossil data for the Albian (Bown et al.,
1998). Except the FO E. turriseiffelli, the other ones bioevents occur beneath the LO of the large planktonic foraminifera considered as being a reference for the upper portion of the Aptian (CGTS2020), thereby conflicting with the standard established for the GSSP-Alb
(Figs. 3
![]() -
4
-
4
![]() ,
9
,
9
![]() ).
).
This striking biochronostratigraphic discrepancy appears to extend far beyond Site 364, with Hayesites albiensis, Tranolithus orionatus, Axopodorhabdus biramiculatus, and even Eiffellithus turriseiffelli occurring below the turnover of large planktonic foraminifera in North Atlantic and South Atlantic sections
(Fig. 9
![]() , columns 9-10, 13-14). However, in the Pacific, European, Agadir-Essaouira Basin and the Site 511 sections
(Fig. 9
, columns 9-10, 13-14). However, in the Pacific, European, Agadir-Essaouira Basin and the Site 511 sections
(Fig. 9
![]() , columns 2, 4-6, 8,
12) these taxa follow the standard succession and occur above this turnover. As the only exception in the South Atlantic, the succession observed in the 511 confirming the thesis of the isolation of the waters of the CSA in relation to the southernmost waters of this ocean.
, columns 2, 4-6, 8,
12) these taxa follow the standard succession and occur above this turnover. As the only exception in the South Atlantic, the succession observed in the 511 confirming the thesis of the isolation of the waters of the CSA in relation to the southernmost waters of this ocean.
This phenomenon needs to be understood. Maybe it points to precocious phylogenetic evolution of the nannoplankton in the CSA, or maybe it can be explained taphonomically, or even by a sampling constrains. It must be considered that the set of North Atlantic and South Atlantic sections used samples that came from deep wells and the European, Agadir-Essaouira Basin sections were studied outcrop samples. However, the presence of Hayesites albiensis and Tranolithus orionatus at the 763 and 511' drilled sections in their predicted stratigraphic positions in the GTS2020 rules out a sampling constrains influence and reinforcing the idea that evolutionary or taphonomic particularities should have been the cause of theses biochronostratigraphic regional discrepancies.
The succession of stratigraphic data for the ESA and BNE basins
(Fig. 9
![]() , column 15) incorporates a lot of information from outcrops, particularly regarding the planktonic foraminifera, palynomorphs, and geochemical information from the Sobradinho Site, Araripe Basin
(Fig. 6
, column 15) incorporates a lot of information from outcrops, particularly regarding the planktonic foraminifera, palynomorphs, and geochemical information from the Sobradinho Site, Araripe Basin
(Fig. 6
![]() ). The planktonic foraminifera are here interpreted as belonging to the Paraticinella rohri Zone and occur in a black shale layer with high TOC values (Cust dio et al.,
2017; Melo et al., 2020), making it chronocorrelative to the Jacob Level. Furthermore, Cunha & Koutsoukos
(1998) associated the FO of P. columnata s.l. with the G. ex gr. maridalensis - H. (H.) similis Zone (Koutsoukos,
1989; Koutsoukos & Bengtson,
1993), suggesting that this calcareous nannofossil species may have occurred below the LO of the large planktonic foraminifera in Sergipe Basin. In turn, the completed biostratigraphy succession of planktonic foraminifera described by Fauth et al.
(2022) in a well also drilled in the Sergipe Basin assures perfect adjustment with the Albian succession shown in the GTS2020
). The planktonic foraminifera are here interpreted as belonging to the Paraticinella rohri Zone and occur in a black shale layer with high TOC values (Cust dio et al.,
2017; Melo et al., 2020), making it chronocorrelative to the Jacob Level. Furthermore, Cunha & Koutsoukos
(1998) associated the FO of P. columnata s.l. with the G. ex gr. maridalensis - H. (H.) similis Zone (Koutsoukos,
1989; Koutsoukos & Bengtson,
1993), suggesting that this calcareous nannofossil species may have occurred below the LO of the large planktonic foraminifera in Sergipe Basin. In turn, the completed biostratigraphy succession of planktonic foraminifera described by Fauth et al.
(2022) in a well also drilled in the Sergipe Basin assures perfect adjustment with the Albian succession shown in the GTS2020
The composite section for the southeastern Brazilian basins (Fig. 9, column 16, and
Fig. 7
![]() ) is based, mainly, on information from drill cores 3-BO-3-RJS from the Campos Basin and 1-SPS-6 from the Santos Basin, the latter of which did not reach the Ibura evaporites. The former provided most information related to the calcareous nannofossils, palynomorphs, and pithonellids, and 1-SPS-6 furnished the data related to the
colomiellids (Dias-Brito et al., 1990; Dias-Brito,
1994, 1995, 1999,
2000). Tedeschi et al. (2017) and Sanjin s et al. (2022) complete the dataset with their reports of large planktonic foraminifera (~Paraticinella rohri) and species related to Globigerinelloides algerianus-G. ferreolensis Zone in the
CSA's upper portion of the post-salt carbonates.
) is based, mainly, on information from drill cores 3-BO-3-RJS from the Campos Basin and 1-SPS-6 from the Santos Basin, the latter of which did not reach the Ibura evaporites. The former provided most information related to the calcareous nannofossils, palynomorphs, and pithonellids, and 1-SPS-6 furnished the data related to the
colomiellids (Dias-Brito et al., 1990; Dias-Brito,
1994, 1995, 1999,
2000). Tedeschi et al. (2017) and Sanjin s et al. (2022) complete the dataset with their reports of large planktonic foraminifera (~Paraticinella rohri) and species related to Globigerinelloides algerianus-G. ferreolensis Zone in the
CSA's upper portion of the post-salt carbonates.
Microbiofacies analyses carried out on core samples from well 1-SPS-6 by Dias-Brito
(1994, 1995) demonstrated the similarity between the faunas and floras of the lower portion of the post-salt carbonates of the Guaruj Formation and the Tamaulipas Formation, Mexico. Based on this affinity, Dias-Brito
(1995) placed the FO of Colomiella mexicana in the uppermost portion of the upper Aptian and the FO of C. recta at the base of the Lower Albian. These occur at a much lower stratigraphic level than the Itanha m Formation, where Tedeschi et al. (2017) and Sanjin s et al. (2022) recognized the LO of the large planktonic foraminifera in the southeastern Brazilian basins
(Fig. 7
![]() ). Guti�rrez-Puente et al. (2021), on the other hand, observed the FO of both Colomiella species slightly before the LO of P. rohri and in the middle of the Microhedbergella Zone, though it bears mentioning that his taxonomic description of the foraminifera was based on thin-sections
(Fig. 9
). Guti�rrez-Puente et al. (2021), on the other hand, observed the FO of both Colomiella species slightly before the LO of P. rohri and in the middle of the Microhedbergella Zone, though it bears mentioning that his taxonomic description of the foraminifera was based on thin-sections
(Fig. 9
![]() , column 11).
, column 11).
The bioevents shown in the right column of figure 9 are based on the calcareous nannofossils, planktonic foraminifera, pithonellids, and colomiellids observed at sites 363 and 364 and in the composite sections of the northeastern and southeastern Brazilian basins
(Fig. 9
![]() , columns 13-16). Also shown in that column are volcanic events, evaporites, and intercalations between organic shales and dolomites, with the two latter being widespread in the meridional portion of the CSA. Volcanic rocks, the salt and black shales bed are indicated at sites 363 and 364 based on geophysical and geochemical evidence (Bolli et al., 1978a,
1978b). The last column on the right side of Figure 9
, columns 13-16). Also shown in that column are volcanic events, evaporites, and intercalations between organic shales and dolomites, with the two latter being widespread in the meridional portion of the CSA. Volcanic rocks, the salt and black shales bed are indicated at sites 363 and 364 based on geophysical and geochemical evidence (Bolli et al., 1978a,
1978b). The last column on the right side of Figure 9
![]() registers the chronostratigraphic proposal put forward in this study, which uses the 113 Ma age for the base of the Albian, coinciding with initiation of the Ibura
Event. There were not indicated formal subdivision to Albian Age according to the GTS2020 decision, and because of the difficulty in reconciling established planktonic foraminifera biochronostratigraphic positions with those of calcareous nannofossils.
registers the chronostratigraphic proposal put forward in this study, which uses the 113 Ma age for the base of the Albian, coinciding with initiation of the Ibura
Event. There were not indicated formal subdivision to Albian Age according to the GTS2020 decision, and because of the difficulty in reconciling established planktonic foraminifera biochronostratigraphic positions with those of calcareous nannofossils.
The results obtained from the 16 sections do not support the idea that the Kilian and Paquier layers are chronostratigraphically consistent on a global scale. Besides occurring in Col de
Pr�-Guittard, organ-rich shales are also present in isolated levels in Teronne outcrops (Kennedy et al., 2000) and in Tardendol and
l'Arboudeysse (Herrle & Mutterlose,
2003), all within the Vocontian Basin
(Fig. 9
![]() , columns 5-7). The Kilian Level was not observed in either Site 463, V�hrum, and Linderos sections
(Fig. 9
, columns 5-7). The Kilian Level was not observed in either Site 463, V�hrum, and Linderos sections
(Fig. 9
![]() , columns 1, 3, 11, respectively), or in any of the South Atlantic sections
(Fig. 9
, columns 1, 3, 11, respectively), or in any of the South Atlantic sections
(Fig. 9
![]() , columns 12-16). An erosional hiatus does not permit speculation regarding its presence at Site 763B, Agadir-Essaouira, Site 545, Site 1049C, or Site 363
(Fig. 9
, columns 12-16). An erosional hiatus does not permit speculation regarding its presence at Site 763B, Agadir-Essaouira, Site 545, Site 1049C, or Site 363
(Fig. 9
![]() , columns 2, 8-10, 13).
, columns 2, 8-10, 13).
In Poggio le Guaine, Site 545, Site 1049C, Site 364, and the composite section of Northeastern Brazilian basins
(Fig. 9
![]() , columns 4, 9-10,
14-15), the occurrence of horizons rich in organic matter are not compatible with the succession of bioevents associated with the
Kilian Level in the GSSP-Alb. In Poggio le Guaine, this organic layer is situated above the FO of M. renilaevis (Coccioni et al.,
2014). At Site 1049C and in the composite section of Northeastern Brazilian basins the black shales are located above the FO of M. rischi, while at Site 545 these deposits are located below the LO of the large planktonic foraminifer, associated with the P. rohri Zone, whose top coincides with an erosional surface (Huber & Leckie,
2011; Trabucho Alexandre et al.,
2010). At the upper section of Site 364, the only horizon with TOC greater than 2% also occurs below the disappearance of these protists
(Fig. 4
, columns 4, 9-10,
14-15), the occurrence of horizons rich in organic matter are not compatible with the succession of bioevents associated with the
Kilian Level in the GSSP-Alb. In Poggio le Guaine, this organic layer is situated above the FO of M. renilaevis (Coccioni et al.,
2014). At Site 1049C and in the composite section of Northeastern Brazilian basins the black shales are located above the FO of M. rischi, while at Site 545 these deposits are located below the LO of the large planktonic foraminifer, associated with the P. rohri Zone, whose top coincides with an erosional surface (Huber & Leckie,
2011; Trabucho Alexandre et al.,
2010). At the upper section of Site 364, the only horizon with TOC greater than 2% also occurs below the disappearance of these protists
(Fig. 4
![]() ).
).
The Paquier Level is observed in sections of the Vocontian Basin, where it was first described
(Fig. 9
![]() , columns 5-7), but its presence in Poggio le Guaine, Umbria-March Basin, is controversial. Differently from what is seen in the GSSP-Alb, the FO of Hayesites albiensis occurs within the upper part of this black shale, named the Urbino Level
(Fig. 9
, columns 5-7), but its presence in Poggio le Guaine, Umbria-March Basin, is controversial. Differently from what is seen in the GSSP-Alb, the FO of Hayesites albiensis occurs within the upper part of this black shale, named the Urbino Level
(Fig. 9
![]() , column 4). The fourth organic-rich horizon in Poggio le Guaine was attributed by Coccioni et al. (2014) to the Leenhardt Level, but is within the Microhedbergella rischi Zone, hence below the FO of Ticinella primula, opposite the order of events defined in the CGTS2020. This illustrates the limited spatial distribution of the Paquier Level, even in Europe, which is only accompanied by an occurrence of a layer rich in organic matter at Site 463
(Fig. 9
, column 4). The fourth organic-rich horizon in Poggio le Guaine was attributed by Coccioni et al. (2014) to the Leenhardt Level, but is within the Microhedbergella rischi Zone, hence below the FO of Ticinella primula, opposite the order of events defined in the CGTS2020. This illustrates the limited spatial distribution of the Paquier Level, even in Europe, which is only accompanied by an occurrence of a layer rich in organic matter at Site 463
(Fig. 9
![]() , column 1). However, this layer does not extend to Site 763B
(Fig. 9
, column 1). However, this layer does not extend to Site 763B
(Fig. 9
![]() , column 2), also drilled in the Pacific, nor to the Moroccan sections
(Fig. 9
, column 2), also drilled in the Pacific, nor to the Moroccan sections
(Fig. 9
![]() , column 8) or to South Atlantic sections where hiatuses are absent
(Fig. 9
, column 8) or to South Atlantic sections where hiatuses are absent
(Fig. 9
![]() , columns 12,
15-16).
, columns 12,
15-16).
The Jacob Level is present in almost all European sections
(Fig. 9
![]() , columns 4-7) and apparently has chronostratigraphic equivalents in sections of the CSA
(Fig. 9
, columns 4-7) and apparently has chronostratigraphic equivalents in sections of the CSA
(Fig. 9
![]() , columns 13-14, 16) and in the inner Brazilian basins
(Fig. 9
, columns 13-14, 16) and in the inner Brazilian basins
(Fig. 9
![]() , column 15), having organic-rich layers overlying the Ibura evaporites. Its presence is inferred at Sites 363 and 364
(Fig. 9
, column 15), having organic-rich layers overlying the Ibura evaporites. Its presence is inferred at Sites 363 and 364
(Fig. 9
![]() , columns 13-14) based on
undirected geological evidence (Bolli et al., 1978a,
1978b). It does not occur at sites 463, 545, 1049C, and 511
(Fig. 9
, columns 13-14) based on
undirected geological evidence (Bolli et al., 1978a,
1978b). It does not occur at sites 463, 545, 1049C, and 511
(Fig. 9
![]() , columns 1, 9-10,
12), nor in the Agadir-Essaouira and Linderos outcrops
(Fig. 9
, columns 1, 9-10,
12), nor in the Agadir-Essaouira and Linderos outcrops
(Fig. 9
![]() , columns 8, 11). An eroded interval inhibits inferring its presence at Site 763B
(Fig. 9
, columns 8, 11). An eroded interval inhibits inferring its presence at Site 763B
(Fig. 9
![]() , column 2).
, column 2).
The results from Site 463 in the Pacific (Fig. 9
![]() , column 1) demonstrate the lack of correlation between organic-rich layers and expressive δ13Ccarb excursions (Matsumoto et al.,
2020). Despite the excellent correlation of carbon isotope values with the Poggio le Guaine outcrop, the TOC values for the section at Site 463 are too low in all but one organic-rich horizon, tentatively associated with the Paquier Level. δ13Ccarb excursions are also present at the Site 545 section
(Fig. 9
, column 1) demonstrate the lack of correlation between organic-rich layers and expressive δ13Ccarb excursions (Matsumoto et al.,
2020). Despite the excellent correlation of carbon isotope values with the Poggio le Guaine outcrop, the TOC values for the section at Site 463 are too low in all but one organic-rich horizon, tentatively associated with the Paquier Level. δ13Ccarb excursions are also present at the Site 545 section
(Fig. 9
![]() , column 9) and in the many horizons with high TOC values at Site 364 (Foresman,
1978; Azevedo,
2001; Bruno et al.,
2020;
Fig. 9
, column 9) and in the many horizons with high TOC values at Site 364 (Foresman,
1978; Azevedo,
2001; Bruno et al.,
2020;
Fig. 9
![]() , column 14). However, the anomalous condition observed in this well is not seen in its neighboring section, Site 363
(Fig. 9
, column 14). However, the anomalous condition observed in this well is not seen in its neighboring section, Site 363
(Fig. 9
![]() , column 13), which suggests that local paleogeographic factors, instead of global ones, influenced deposition of these black shales. In turn, the absence of important δ13Ccarb negative excursion in the Jacob Level, in contrast to what occurs in the Kilian and Paquier levels in European sections
(Fig. 9
, column 13), which suggests that local paleogeographic factors, instead of global ones, influenced deposition of these black shales. In turn, the absence of important δ13Ccarb negative excursion in the Jacob Level, in contrast to what occurs in the Kilian and Paquier levels in European sections
(Fig. 9
![]() , columns 5, 7) and is consolidated in the
CGTS2020's δ13Ccarb curve, demonstrates the difficulty in understanding the effective modeling agents of carbon isotope fractionation during the formation of these black shales.
, columns 5, 7) and is consolidated in the
CGTS2020's δ13Ccarb curve, demonstrates the difficulty in understanding the effective modeling agents of carbon isotope fractionation during the formation of these black shales.
The information presented in Figure 9 demonstrates the difficulty in applying the model established by the GSSP-Alb to other regions of the world outside of the Vocontian Basin. Although the planktonic foraminifera succession indicated in the GSSP-Alb has proved to be widely applicable, adjustments are required with other fossil groups, particularly with calcareous nannofossils. The peculiarities of this situation justify the proposal of Antunes et al. (2018) to adopt the base of the salt as the Aptian-Albian boundary, with an age of around 113 Ma, for the basins of the CSA, ESA, and BNE. Until the significant biochronological incompatibilities of key taxa for these stratigraphic stages are resolved, it is their proposal that best aligns with the geochronology indicated in the GTS2020 (113.2 0.3 Ma).
A "Scientific Revolution" presented itself to researchers of the fauna and flora preserved in the earliest marine sediments in the South Central Atlantic (CSA), Equatorial South Atlantic (ESA), and interior of northeastern Brazil (BNE), with the identification of a diversified assemblage of planktonic foraminifera of Aptian age, according the Albian Global Stratotype Section Point (GSSP-Alb). This assembly extends up to the uppermost portion of the post-salt carbonates, hitherto considered to be of Albian age. As happens when a challenge to "normal science" presents itself, it was necessary to test the new information against already established information involving the same research object (Kuhn, 1962). In this case, information on planktonic foraminifera was integrated with biochronostratigraphic data from different fossil groups, lithostratigraphic data, and geochronological data. This revealed a paradoxical conflict among this information, a problem particularly notable in the CSA, ESA, and BNE basins but not exclusive to this region of the world.
The analysis of 16 sections from sedimentary basins at different localities showed the difficulty in recognizing numerous markers data in the GSSP-Alb. For example, the diagnostic datum for the Aptian-Albian boundary, the first occurrence (FO) Microhedbergella renilaevis, did not show up as frequently as did the disappearance of the large planktonic foraminifera, the most globally consistent event observed among this protist group in the analysed sections. The few sure occurrences of the Kilian, Paquier and Leenhardt levels observed in this study exhibited limitations as global stratigraphic markers. Only the Jacob level appears to occur below the consistent FO of Prediscosphaera columnata s.l. on a global scale. The use of an expanded taxonomic concept for this calcareous nannofossil confirms its stratigraphic value as a candidate for marking the base of the Albian, as suggested a few years ago.
In relation to the CSA, ESA, and BNE basins, the controversy between the biochronostratigraphy offered by planktonic foraminifera in relation to those indicated by calcareous nannofossils and pithonellids has been presented in detail. This problem was first recorded about 45 years ago in the Initial Report on Leg 40 of the Deep Sea Drilling Project (DSDP). At stratigraphic levels below the disappearance of the large planktonic foraminifera (~Paraticinella rohri Zone), forms recognized as having much younger and exclusively Albian amplitudes occur, such as Hayesites albiensis, Tranolithus orionatus, and Axopodorhabdus biramiculatus, among the calcareous nannofossils, and Pithonella sphaerica, P. ovalis, and Bonetocardiella conoidea, among the pithonellids. Moreover, the description of successions of Aptian planktonic foraminifera in wells showing no indication of reworking documents the coexistence of species traditionally considered as from different time intervals, once again reinforcing the need to review many of the LOs recorded in chronostratigraphic tables.
These divergences would be less relevant were it not for the 113.2 0.1 Ma age determined on trachyandesite samples overlain by the Ibura evaporites in the Florian polis High in the CSA, which is almost same age indicated for the GSSP-Alb by the GTS2020.More recent drilling in the Campos and Santos basins has recognized Pre-Salt strata intercalated with basic volcanic flows, also covered by the salt layer. The youngest of these volcanic rocks confirm the 113 Ma age for the base of this salt. Thus, in order to build a globally consistent chronostratigraphy, the geochronology dictates for the GGSP-Alb cannot fail to consider the many possibilities of volcanic flows close to the base of the salt limit CSA sections.
Changing the focus, for now, from the international GSSP-Alb definition, we recommend adopting the base of the salt of the Ibura Event as the reference for the Aptian-Albian boundary in the CSA, ESA, and BNE basins. Of unequivocal stratigraphic value, it is easily recognizable and coeval in these basins, contrary to the suggested criteria for the GSSP-Alb, whose many limitations have been discussed above. Furthermore, the magnitude of the processes leading to salt formation, as well as those succeeding it during the opening and expansion of the South Atlantic Ocean, likely left records in other parts of the globe, that merit proper exploration.
The authors sincerely acknowledge Malcolm Hart, Bruno Granier, Pierre Masse, and Mike Simmons for reviewing and providing key insights that improved the quality of the paper. They are also grateful to colleagues Dimas Dias-Brito, Peter Szatmari, Ismar de Souza Carvalho, Thomas Rich Fairchild, Marcos A.B.S. Filho, Mitsuru Arai, L cio Riogi Tokutake, Robbyson Mendes Melo, David K. Watkins, and Karlos G.D. Kochhann for reviews, suggestions, and/or criticism of specific parts or all of the text. The authors also acknowledge Petr leo Brasileiro S.A., Universidade Federal do Rio de Janeiro, and Universidade do Vale do Rio dos Sinos for logistical, financial support and/or scientific training for the authors.
Alves C.F., Lima F.H. de O. & Shimabukuro S. (2017).- New Aptian calcareous nannofossil species from Brazil.- Journal of Nannoplankton Research, vol. 37, p. 15-23.
Ando A., Huber B.T. & Premoli Silva I. (2013).- Paraticinella rohri (Bolli, 1959) as the valid name for the latest Aptian zonal marker species of planktonic foraminifera traditionally called bejaouaensis or eubejaouaensis.- Cretaceous Research, vol. 45, p. 275-287.
Antunes R.L. (1996).- Biozonas de Nanof�sseis do Cret�ceo da Margem Continental Brasileira: Problemas e poss�vel solu��es.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 10, no. 1/4, p. 19-42.
Antunes R.L. & Azevedo R.L.M. & Lobo J.T. (2018).- Reflex�es Sobre a S�rie Rec�ncavo, Brasil.- Anu�rio do Instituto de Geoci�ncias - UFRJ, Rio de Janeiro, vol. 41, p. 276-296. DOI: https://doi.org/10.11137/2018_2_276_296
Arai M. (1992).- Dinoflagellates from the middle Cretaceous in the offshore Campos Basin, southeastern Brazil. In: 2o Simp�sio sobre as Bacias Cret�cicas Brasileiras.- Resumos expandidos, UNESP, Rio Claro, p. 27-29.
Arai M. (2009).- Paleogeografia do Atl�ntico Sul no Aptiano: um novo modelo a partir de dados micropaleontol�gicos recentes.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 17, no. 2, p. 331-351.
Arai M. (2014).- Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: A paleontological perspective.- Brazilian Journal of Geology, S�o Paulo, vol. 44, no. 2, p. 339-350. DOI: https://doi.org/10.5327/Z2317-4889201400020012
Arai M. & Assine M.L. (2020).- Chronostratigraphic constraints and paleoenvironmental interpretation of the Romualdo Formation (Santana Group, Araripe Basin, Northeastern Brazil) based on palynology.- Cretaceous Research, vol. 116, article 104610, 13 p.
Arai M., Hashimoto A.T. & Uesugui N. (1989).- Significado cronoestratigr�fico da associa��o microflor�stica do Cret�ceo Inferior no Brasil.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 3, no. 1/2, p. 87-103.
Arai M., Lana C.C. & Pedr o E. (1994).- Ecozona Subtilisphaera: Registro EoCret�ceo de um importante epis�dio ecol�gico do oceano Atl�ntico primitivo.- Acta Geologica Leopoldensia, São Leopoldo, vol. 32, no. 2, p. 521-538.
Arai M., Shimabukuro S. & Viviers M.C. (1996).- Caracteriza��o do Vraconiano (Albiano superior, Cret�ceo Inferior) no Brasil: Uma contribui��o paleomicroplanct�nica. In: Boletim do 4o Simp�sio Sobre o Cret�ceo do Brasil.- UNESP, Rio Claro, p. 39-45.
Asmus H.E. & Campos D.A. (1983).- Stratigraphic division of the Brazilian continental margin and its paleogeographic significance.- Zitteliana, M�nchen, vol. 10, p. 265-276.
Assine M.L. (2007).- Bacia do Araripe.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 371-389.
Atta-Peters D. (2013).- Occurrences of elaterate pollen from the Lower Cretaceous of Ghana: Implications for biostratigraphy and palaeoclimatology.- International Letters of Natural Sciences, Bach, vol. 4, p. 54-66. DOI: https://doi.org/10.56431/p-c1m8f9
Azevedo R.L.M. (2001, unpublished).- O Albiano no Atl�ntico Sul: Estratigrafia, paleoceanografia e rela��es globais.- PhD Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, 401 p.
Azevedo R.L.M. (2004).- Paleoceanografia e a evolu��o do Atl�ntico Sul no Albiano.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 12, no. 2, p. 231-249
Azevedo R.L.M. (2005).- Os ciclos sedimentares e seus significados na forma��o das sequ�ncias deposicionais carbon�ticas da Bacia de Campos.- Revista Brasileira de Geoci�ncias, S�o Paulo, vol. 35, p. 127-138. URL: https://www.ppegeo.igc.usp.br/index.php/rbg/article/view/9393
Azevedo R.L.M., Gomide J., Viviers M.C. & Hashimoto A.H. (1987).- Bioestratigrafia do Cret�ceo marinho da Bacia de Campos, Brasil.- Revista Brasileira de Geoci�ncias, S�o Paulo, vol. 17, no. 2, p. 147-153. https://www.ppegeo.igc.usp.br/index.php/rbg/article/view/11902
Banner F.T. & Desai D. (1988).- A review and revision of the Jurassic-Early Cretaceous Globigerinina, with especial reference to the Aptian assemblages of Speeton (North Yorkshire, England).- Journal of Micropalaeontology, London, vol. 7, p. 143-185. DOI: https://doi.org/10.1144/jm.7.2.143
Barbosa J.A., Lima Filho M., Neumann V.H., Neto J.C.J. & Ara jo J.A.A. (2008).- Potencial explorat�rio das bacias da Para�ba e da plataforma de Natal, NE do Brasil. In: Rio Oil & Gas Expo and Conference 2008.- Instituto Brasileiro de Petr�leo, G�s e Biocombust�veis.- IBP, Rio de Janeiro, 8 p.
Beglinger S.E., Doust H. & Cloetingh S. (2012).- Relating petroleum system and play development to basin evolution: Brazilian South Atlantic Margin.- Petroleum Geoscience, vol. 18, p. 315-336.
Below R. (1984).- Aptian to Cenomanian Dinoflagellate Cysts from the Mazagan Plateau, Northwest Africa (Sites 545 and 547, Deep Sea Drilling Project Leg 79). In: Hinz K., Winterer E.L., Baumgartner P.O., Bradshaw M.J., Channell J.E.T., Jaffrezo M., Jansa L.F., Leckie R.M., Moore J.N., Rullk tter J., Schaftenaar C., Steiger T.H., Vuchev V. & Wiegand G.E. (eds.), Leg 79 of the cruises of the Drilling Vessel Glomar Challenger. Las Palmas, Grand Canary Island, to Brest, France. April-May 1981.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXIX, p. 621-649. DOI: https://doi.org/10.2973/dsdp.proc.79.123.1984
Bengtson P., Zucon M.H. & Sobral A. da C.S. (2018).- Cretaceous ammonite zonation of the Sergipe Basin, northeastern Brazil.- Cretaceous Research, vol. 88, p. 111-122.
Bolli H.M. (1978).- Synthesis of the Leg 40 biostratigraphy and paleontology. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 1063-1067. DOI: https://doi.org/10.2973/dsdp.proc.40.131.1978
Bolli H.M., Ryan W.B.F., Foresman J.B., Hottman W.E., Kagami H., Longoria J.F., McKnight B.K., Melguen M., Natland J., Proto-Decima F. & Siesser W.G. (1978a).- Walvis Ridge-Sites 362 and 363. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 183-356. DOI: https://doi.org/10.2973/dsdp.proc.40.103.1978
Bolli H.M., Ryan W.B.F., Foresman J.B., Hottman W.E., Kagami H., Longoria J.F., McKnight B.K., Melguen M., Natland J., Proto-Decima F. & Siesser W.G. (1978b).- Angola continental margin-Sites 364 and 365. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 357-455. DOI: https://doi.org/10.2973/dsdp.proc.40.104.1978
Bown P.R., Rutledge D.C., Crux J.A. & Gallagher L.T. (1998).- Lower Cretaceous. In: Bown P.R. (ed.), Calcareous nannofossil biostratigraphy.- Kluwer Academic Publishers, Dordrecht, p. 86-131.
Bralower T.J. & Siesser W.G. (1992).- Cretaceous calcareous nannofossil biostratigraphy of sites 761, 762, and 763, Exmouth and Wombat plateaus, northwest Australia. In: Rad U. von, Haq B.U., Kidd R.B. & O'Connell S. (eds.), Exmouth Plateau. Leg 122 of the cruises of the Drilling Vessel JOIDES Resolution, Singapore, Republic of Sing., to Singapore, Republic of Sing., Sites 759-764, 28 June 1988-28 August 1988.- Proceedings of the Ocean Drilling Program, Scientific Results, Washington - DC, vol. 122, p. 529-556. DOI: https://doi.org/10.2973/odp.proc.sr.122.169.1992
Br�h�ret J.G. (1983).- Sur des niveaux de black shales dans l'Albien inf�rieur et moyen du domaine vocontien (Sud-Est de la France) : �tude de nannofaci�s et signification des pal�oenvironments.- Bulletin du Museum National d'Histoire Naturelle, Paris, vol. 1, p. 113-159.
Br�h�ret J.G., Caron M. & Delamette M. (1986).- Niveaux riches en mati�re organique dans l'Albien Vocontien ; quelques caract�res du pal�oenvironnement ; essai d'interpr�tation g�n�tique.- Documents du Bureau de Recherches G�ologiques et Mini�res, Orl�ans, vol. 110, p. 141-191.
Browning E.L. & Watkins D.K. (2008).- Elevated primary productivity of calcareous nannoplankton associated with ocean anoxic event 1b during the Aptian/Albian transition (Early Cretaceous).- Paleoceanography, vol. 23, no. 2, article PA2213, 11 p. DOI: https://doi.org/10.1029/2007PA001413
Bruno M.D.R., Fauth G., Watkins D.K. & Savian J.F. (2020).- Albian-Cenomanian calcareous nannofossils from DSDP Site 364 (Kwanza Basin, Angola): Biostratigraphic and paleoceanographic implications for the South Atlantic.- Cretaceous Research, vol. 109, article 104377, 12 p.
Bueno G.V., Zacharias A.A., Oreiro S.G., Cupertino J.A., Falkenhein F.U.H. & Martins Neto M.M. (2007).- Bacia de Pelotas.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 551-559.
Caetano-Filho S., Dias-Brito D., Rodrigues R. & Azevedo R.L.M. (2017).- Carbonate microfacies and chemostratigraphy of a late Aptian-early Albian marine distal section from the primitive South Atlantic (SE Brazilian continental margin): Record of global ocean-climate changes? Cretaceous Research, vol. 74, p. 23-44.
Caron M. (1978).- Cretaceous planktonic foraminifers from DSDP Leg 40, Southeastern Atlantic Ocean. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 651-678. DOI: https://doi.org/10.2973/dsdp.proc.40.114.1978
Caron M. (1985).- Cretaceous planktic foraminifera. In: Bolli H.M., Saunders J.B. & Perch-Nielsen K. (eds.), Plankton stratigraphy.- Cambridge University Press, Cambridge (UK), p. 17-86.
Carvalho M. de A., Bengtson P. & Lana C.C. (2016).- Late Aptian (Cretaceous) paleoceanography of the South Atlantic Ocean inferred from dinocyst communities of the Sergipe Basin, Brazil.- Paleoceanography, vol. 31, no. 1, p. 2-26. DOI: https://doi.org/10.1002/2014PA002772
Cep�k P. (1981).- Mesozoic calcareous nannoplankton stratigraphy of the Central North pacific (Mid-Pacific mountains and Hess Rise) Deep Sea Drilling Project 62. In: Thiede J., Vallier T.L., Adelseck C.G., Boersma A., Cep�k P., Dean W.E., Fujii N., Koporulin V.I., Rea D.K., Sancetta C.A., Sayre W.O., Seifert K.E., Schaaf A., Schmidt R.R., Windom K.E. & Vincent E. (eds.), Leg 62 of the cruises of the Drilling Vessel Glomar Challenger. Marjuro Atoll, Marshall Islands to Honolulu, Hawaii. July-September 1978.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXII, p. 397-418. DOI: https://doi.org/10.2973/dsdp.proc.62.111.1981
Coccioni R., Sabatino N., Frontalini F., Gardin S., Sideri M. & Sprovieri M. (2014).- The neglected history of Oceanic Anoxic Event 1b: Insights and new data from the Poggio le Guaine section (Umbria-Marche Basin).- Stratigraphy, Flushing - NY, vol. 11, p. 245-282.
Cunha A.A.S. & Koutsoukos E.A.M. (1998).- Calcareous nannofossils and planktic foraminifers in the upper Aptian of the Sergipe Basin, northeastern Brazil: palaeoecological inferences.- Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 142, nos. 3-4, p. 175-184.
Cust dio M.A., Quaglio F., Warren L.V., Sim es M.G., F rsich F.T., Perinotto J.A.J. & Assine M.L. (2017).- The transgressive-regressive cycle of the Romualdo Formation (Araripe Basin): Sedimentary archive of the Early Cretaceous marine ingression in the interior of Northeast Brazil.- Sedimentary Geology, vol. 359, p. 1-15.
Davison I. (2007).- Geology and tectonics of the South Atlantic Brazilian salt basins.- Geological Society, London, Special Publications, vol. 272, p. 345-359.
Dean W.E., Claypool G.E. & Thiede J. (1981).- Origin of organic Carbin rich mid-Cretaceous limestone, Mid-Pacific Mountains and Southern Hess Hise. In: Thiede J., Vallier T.L., Adelseck C.G., Boersma A., Cep�k P., Dean W.E., Fujii N., Koporulin V.I., Rea D.K., Sancetta C., Sayre W.O., Seifert K., Schaaf A., Schmidt R.R., Windom K. & Vincent E. (eds.), Leg 62 of the cruises of the Drilling Vessel Glomar Challenger. Marjuro Atoll, Marshall Islands to Honolulu, Hawaii. July-September 1978.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXII, p. 877-890. DOI: https://doi.org/10.2973/dsdp.proc.62.144.1981
Della F vera J.C., Medeiros R.A. Appi C.J., Beurlen G., Viviers M.C. & Hashimoto A.T. (1984).- An�lise estratigr�fica do Andar Alagoas na Bacia do Cear�.- Petrobras, CENPES, Rio de Janeiro, Internal Report.
Demercian L.S. (1996, unpublished).- A halocinese na evolu��o da Bacia de Santos do Aptiano ao Cret�ceo Superior.- MSc Master of Science, Universidade Federal do Rio Grande do Sul, Porto Alegre, 201 p.
Dias J.L. (1998, unpublished).- An�lise sedimentol�gica e estratigr�fica do Andar Aptiano em parte da margem leste do Brasil e no Plat� das Malvinas - Considera��es sobre as primeiras incurs�es marinhas e ingress�es marinhas do oceano Atl�ntico Sul Meridional.- PhD Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, 145 p.
Dias J.L. (2005).- Tect�nica, estratigrafia e sedimenta��o no Andar Aptiano da margem leste brasileira.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 13, no. 1, p. 7-25.
Dias J.L., Sad, A.R.R., Fontana R.L. & Feij� F.J. (1994).- Bacia de Pelotas.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 8, no. 1, 235-245.
Dias-Brito D. (1985).- Calcisphaerulidae do Albiano da Bacia de Campos, Rio de Janeiro, Brasil. Investiga��es taxon�micas: biocronoestratigr�ficas e paleoambientais.- Colet�nea de Trabalhos Paleontol�gicos, Geologia (Geologia 27, Paleontologia, Estratigrafia 2), Bras�lia, p. 295-306.
Dias-Brito D. (1994).- Compara��o dos carbonatos pel�gicos do Cret�ceo m�dio da Margem Atl�ntica Brasileira com os do Golfo do M�xico: Novas evid�ncias do T�tis Sul-Atlantiano. In: Boletim do 3o Simp�sio Sobre o Cret�ceo do Brasil.- UNESP, Rio Claro, p. 11-18.
Dias-Brito D. (1995, unpublished).- Calcisferas and microf�cies em rochas carbon�ticas pel�gicas mesocret�ceas.- PhD Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil, 688 p.
Dias-Brito D. (1999).- Usando pitonelidos e colomielidos para dividir o Albiano: um estudo a partir da margem sudeste do Brasil. In: Boletim do 5o Simp�sio Sobre o Cret�ceo do Brasil/1oo Simp�sio Sobre El Cret�cecico de America del Sur.- UNESP, Rio Claro, p. 213-220.
Dias-Brito D. (2000).- Global stratigraphy, palaeobiogeography and palaeoecology of Albian-Maastrichtian pithonellid calcispheres: Impact on Tethys configuration.- Cretaceous Research, vol. 21, p. 315-349.
Dias-Brito D., Arai M., Shimabukuro S. & Azevedo R.L.M. (1990).- Estratigrafia do pl�ncton Albiano da Bacia de Campos: Um reconhecimento a partir do po�o 3-BO-3-RJS. In: Boletim de Resumos do 1o Simp�sio sobre as Bacias Cret�ceas Brasileiras.- UNESP, Rio Claro, p. 41-42 (abstract).
Dias-Brito D. & Azevedo R.L.M. (1986).- As seq��ncias deposicionais marinhas da Bacia de Campos sob a �tica paleoecol�gica. In: Anais do 34o Congresso Brasileiro de Geologia, Goiania.- Sociedade Brasileira de Geologia (SBG), Rio de Janeiro, p. 38-49.
Dias-Brito D. & Ferr B. (2001).- Rovearinids (stemless crinoids) in the Albian carbonates of the offshore Santos Basin, southeastern Brazil: Stratigraphic, palaeobiogeographic and palaeoceanographic significance.- Journal of South American Earth Sciences, vol. 14, p. 203-218.
Dias-Brito D., Uesugui N. & Hashimoto A.T. (1987).- Uma reflex�o hist�rica em torno do Andar Alagoas - �mportante e problem�tica unidade cronoestratigr�fica do Cret�ceo Inferior do Brasil.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 1, p. 111-115.
Dino R., Pocknall D.T. & Dettmann M.E. (1999).- Morphology and ultrastructure of elater-bearing pollen from the Albian to Cenomanian of Brazil and Ecuador: Implications for botanical affinity.- Review of Palaeobotany and Palynology, vol. 105, p. 201-235.
Dupont G. (1996).- Principaux foraminif�res planctoniques du Cr�tac� gabonais (Aptien � Campanien inf�rieur).- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, M�moire 16, p. 83-121.
Fauth G., Krahl G., Kochhann K.G.D., Bom M.H.H., Fauth S.B., Bruno M.D.R., Guerra R.M., Ceolin D., Santos A.S., Martin, J.V., Strohschoen O. Jr, Savian J.F., Leandro C.G., Mello R.G. & Lima F.H.O. (2022).- Astronomical calibration of the latest Aptian to middle Albian in the South Atlantic Ocean.- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 602, article 111175, 13 p.
Feij� F.J. (1994).- Introdu��o.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 8, no. 1, p. 5-8.
Fekete K., Sot�k J., Boorov� D., Lintnerov� O., Michal�k J. & Grabowski J. (2017).- An Albian demise of the carbonate platform in the Man n Unit (Western Carpathians, Slovakia).- Geologica Carpathica, vol. 68, p. 385-402. DOI: https://doi.org/10.1515/geoca-2017-0026
Foresman J.B. (1978).- Organic geochemistry DSDP Leg 40, Continental Rise of Southwest Africa. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 557-567. DOI: https://doi.org/10.2973/dsdp.proc.40.111.1978
Fran�a R.L., Rey A.C. del, Tagliari C.V., Brand o J.R. & Fontanelli P. de R. (2007).- Bacia do Esp�rito Santo.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 501-509.
Gale A.S., Mutterlose J. & Baternburg S. (2021).- The Cretaceous period. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale.- Elsevier, Amsterdam, p. 1023-1086.
Gomes L.C., Lima F.M., Vieira I.S., Lobo J.T. et al. (2016).- Magmatismo Jiqui� e Alagoas na Bacia de Santos.- Petrobras, CENPES, Rio de Janeiro, Internal Report.
Gradstein F.M., Ogg J.G., Smith A.G., Bleeker W. & Lourens L.J. (2004).- A new Geologic Time Scale with special reference to Precambrian and Neogene.- Episodes, Bangalore, vol. 27, p. 83-100.
Granier B. (2017).- Introductory remarks. In: Granier B. (ed.), Some key Lower Cretaceous sites in Dr�me (SE France).- Carnets de G�ologie, Madrid, Book 2017/01 (CG2017_B01), p. 2-45. DOI: https://doi.org/10.4267/2042/62543
Guti�rrez-Puente N.A., Barrag�n R. & N��ez-Useche F. (2021).- Paleoenvironmental changes and biotic response to Aptian-Albian episodes of accelerated global change: Evidence from the western margin of the proto-North Atlantic (central-eastern Mexico).- Cretaceous Research, vol. 126, article 104883, 27 p.
Haq B.U. (2014).- Cretaceous eustasy revisited.- Global and Planetary Change, vol. 113, p. 44-58.
Hart M.B., Am dro F. & Owen H. (1996).- The Albian stage and substage boundaries.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, vol. 66-SUPP, p. 45-56.
Hassanein W. (2016).- The Aptian-Albian transgression in the Agadir-Essaouira Basin, western Morocco.- PhD Thesis, Universit Grenoble-Alpes Grenoble, 268 p.
Heine C. & Brune S. (2014).- Oblique rifting of the Equatorial Atlantic: Why there is no Saharan Atlantic ocean.- Geology, Boulder - CO, vol. 42, no. 3, p. 211-214. DOI: https://doi.org/10.1130/G35082.1
Helmond N.A.G.M. van, Sluijs A., Reichart G.J., Damst J.S.S., Slomp C.P. & Brinkhuis H. (2014).- A perturbed hydrological cycle during Oceanic Anoxic Event 2.- Geology, Boulder - CO, vol. 42, no. 2, p. 123-126. DOI: https://doi.org/10.1130/G34929.1
Herbin J.-P., M ller C., Graciansky P.C. de, Jacquin T. & Magniez-Jannin F., Unterneher P. (1987).- Cretaceous Anoxic Events in the South Atlantic.- Revista Brasileira de Geoci�ncias, S�o Paulo, vol. 17, no. 2, p. 92-99. URL: https://ppegeo.igc.usp.br/index.php/rbg/article/view/11882/11427
Herrle J.O. & Mutterlose J. (2003).- Calcareous nannofossils from the Aptian-Lower Albian of southeast France: Palaeoecological and biostratigraphic implications.- Cretaceous Research, vol. 24, p. 1-22.
Herrle J.O., K ler P., Friedrich O., Erlenkeuser H. & Hemleben C. (2004).- High-resolution carbon isotope records of the Aptian to Lower Albian from SE France and the Mazagan Plateau (DSDP Site 545): A stratigraphic tool for paleoceanographic and paleobiologic reconstruction.- Earth and Planetary Science Letters, vol. 218, p. 149-161.
Huber B.T. & Leckie R.M. (2011).- Planktic foraminiferal species turnover across deep-sea Aptian/Albian boundary sections.- Journal of Foraminiferal Research, Lawrence - KS, vol. 41, p. 53-95.
Jones C.M. & Chaves H.A.F. (2011).- Assessment of yet-to-find oil in the Brazilian Pre-Salt region. In: Brazil Offshore Conference 2011. Maca , Rio de Janeiro, Brazil. 14-17 June 2011.- Society of Petroleum Engineers, Richardson - TX, SPE 143911, p. 978-985.
Keen C.E., Dickie K. & Dehler S.A. (2012).- The volcanic margins of the northern Labrador Sea: Insights to the rifting process.- Tectonics, Washington - DC, vol. 31, no. 1, article TC1011, 13 p. DOI: https://doi.org/10.1029/2011TC002985
Kennedy W.J. & Cooper M. (1975).- Cretaceous ammonite distributions and the opening of the South Atlantic.- Journal of the Geological Society, London, vol. 131, no. 3, p. 283-288.
Kennedy J.W., Gale A.S., Bown P.R., Caron M., Davey R.J., Gr cke D.R. & Wray D.S. (2000).- Integrated stratigraphy across the Aptian-Albian boundary in the Marnes Bleues, at the Col de Pr�-Guittard, Arnayon (Dr�me), and at Tartonne (Alpes-de-Haute-Provence), France: A candidate global boundary stratotype section and boundary point for the base.- Cretaceous Research, vol. 21, p. 591-720.
Kennedy J.W., Gale A.S., Huber B.T., Petrizzo M.R., Bown P.R., Barchetta A. & Jenkyns H.C. (2014).- Integrated stratigraphy across the Aptian/Albian boundary at Col de Pr�-Guittard (southeast France): A candidate global boundary stratotype section.- Cretaceous Research, vol. 51, p. 248-259.
Kennedy J.W., Gale A.S., Huber B.T., Petrizzo M.R., Bown P.R. & Jenkyns H.C. (2017).- The Global Boundary Stratotype Section and Point (GSSP) for the base of the Albian Stage, of the Cretaceous, the Col de Pr�-Guittard section, Arnayon, Dr�me, France.- Episodes, Beijing, vol. 40, p. 177-188. DOI: https://doi.org/10.18814/epiiugs/2017/v40i3/017021
Kochhann K.G.D., Koutsoukos E.A.M., Fauth G. & Sial A.N. (2013).- Aptian-Albian planktic Foraminifera from DSDP Site 364 (Offshore Angola): Biostratigraphy, paleoecology, and paleoceanographic significance.- Journal of Foraminiferal Research, Lawrence - KS, vol. 43, no. 4, p. 443-463.
Koutsoukos E.A.M. (1987).- A �rea Nordeste da Bacia de Campos, Brasil, do mesocret�ceo ao Neocret�ceo: evolu��o paleoambiental e paleogeogr�fica pelo estudo de foramin�feros.- Revista Brasileira de Geoci�ncias, S�o Paulo, vol. 17, no. 2, p. 168-172. URL: https://ppegeo.igc.usp.br/index.php/rbg/article/view/11880/11425
Koutsoukos E.A.M. (1989, unpublished).- Mid- to late Cretaceous microbiostratigraphy, palaeoecology and palaeogeography of the Sergipe basin, northeastern Brazil.- PhD Thesis, Council for Nacional Academic Awards, Polytechnic South West, Plymouth, 886 p. URL: https://pearl.plymouth.ac.uk/bitstream/handle/10026.1/2016/Eduardo%20Apostolos%20Machado%20Koutsoukos_V2.pdf?sequence=2
Koutsoukos E.A.M. (1992).- Late Aptian to Maastrichtian foraminiferal biogeography and palaeoceanography of the Sergipe Basin, Brazil.- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 92, p. 295-324.
Koutsoukos E.A.M. & Bengtson P. (1993).- Towards an integrated biostratigraphy of the upper Aptian-Maastrichtian of the Sergipe Basin, Brazil.- Documents des Laboratoires de G�ologie de Lyon, vol. 125, p. 241-262. URL: https://www.persee.fr/doc/geoly_0750-6635_1993_act_125_1_1560
Kumar N., Gamb a L.A.P., Schreiber B.C. & Mascle J. (1977).- Geologic history and origin of Sao Paulo Plateau (Southeastern Brazilian Margin), comparison with the Angolan margin, and the early evolution of the northern South Atlantic. In: Supko P.R., Perch-Nielsen K., Neprochnov Y.P., Zimmerman H.B., McCoy F., Kumar N., Thiede J., Bonatti E., Fodor R., Boersma A., Dinkelman M.G. & Carlson R.L. (eds.), Leg 39 of the cruises of the Drilling Vessel Glomar Challenger. Amsterdam, Netherlands to Cape Town, South Africa. October-December 1974.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XXXIX, p. 927-945. DOI: https://doi.org/10.2973/dsdp.proc.39.140.1977
Kuhn T.S. (1962).- The structure of scientific revolutions.- 4th edition (2012), The University of Chicago, Chicago - IL, 264 p.
Lawson M., Sitgreaves J., Rasbury T., Wooton K., Esch W., Marcon V., Henares S., Konstantinou A., Kneller E., Gombosi D., Torres V., Silva A., Alevato R., Wren M., Becker S. & Eiler J. (2022).- New age and lake chemistry constraints on the Aptian pre-salt carbonates of the central South Atlantic.- GSA Bulletin, Boulder - CO, 13 p. DOI: https://doi.org/10.1130/B36378.1
Leckie R.M. (1984).- Mid-Cretaceous planktonic foraminiferal biostratigraphy off central Morocco. In: Hinz K., Winterer E.L., Baumgartner P.O., Bradshaw M.J., Channell J.E.T., Jaffrezo M., Jansa L.F., Leckie R.M., Moore J.N., Rullk tter J., Schaftenaar C., Steiger T.H., Vuchev V. & Wiegand G.E. (eds.), Leg 79 of the cruises of the Drilling Vessel Glomar Challenger. Las Palmas, Grand Canary Island, to Brest, France. April-May 1981.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXIX, p. 579-620. DOI: https://doi.org/10.2973/dsdp.proc.79.122.1984
Le n-Francisco J.M., Murilo-Mu eton G. & Franco-Navarrete S.P. (2013).- Stratigraphic/sedimentological analysis of a Lower Cretaceous carbonate succession in the Actopan Platform, Hidalgo State, Mexico: Documentation on an early Albian deepening event.- American Geophysical Union, Washington - DC, Spring Meeting 2013, id. PP41C-02, 1 p. (abstract). URL: https://ui.adsabs.harvard.edu/abs/2013AGUSMPP41C..02L/abstract
Lima F.H. de O., Sanjin s A.E.S., Maizatto J.R., Ferreira, E.P., Ng C., Costa D.S., Anjos-Zerfass G. de S., Alves C.F., Strohschoen O. Jr, Viviers M.C., Lana C.C., Arai M., Beurlen G., Shimabukuro S. & Galm P.C. (2018).- Aptian marine post-salt rocks in Santos, Campos and Esp�rito Santo basins, Brazil: a biochronostratigraphical approach. In: RIO 2018, 49o Congresso Brasileiro de Geologia.- Sociedade Brasileira de Geologia (SBG), Rio de Janeiro, p. 2023 (abstract). URL: http://cbg2018anais.siteoficial.ws/resumos/4924.pdf
Luciani V., Cobianchi M. & Jenkyns H.C. (2004).- Albian high-resolution biostratigraphy and isotope stratigraphy: The Coppa della Nuvola pelagic succession of the Gargano Promontory (Southern Italy).- Eclogae Geologicae Helvetiae, Basel, vol. 97, p. 77-92. DOI: https://doi.org/10.1007/s00015-004-1106-9
Madhavaraju J., Sial A.N., Gonz lez-Le n C.M. & Nagarajan R. (2013).- Carbon and oxygen isotopic variations in early Albian limestone facies of the Mural Formation, Pitaycachi section, northeastern Sonora, Mexico.- Revista Mexicana de Ciencia Geol gicas, Quer taro, vol. 30, no. 3, p. 526-539. URL: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1026-87742013000300005
Matias H.C., Ninci B., Guangsheng X., Pessoa M.C., Mattos F., Margem R., Carballo J., Esteban M., Tritlla J., Loma R., & Pimentel G. (2015).- Unlocking Pandora - Insights from pre-salt reservoirs in Campos and Santos Basins (offshore Brazil). In: 77th EAGE Conference and Exhibition 2015: Earth Science for Energy and Environment.- European Association of Geoscientists and Engineers, Bunnik, p. 1691-1695.
Matsumoto H., Kuroda J., Coccioni R., Frontalini F., Sakai S., Ogawa N.O. & Ohkouchi N. (2020).- Marine Os isotopic evidence for multiple volcanic episodes during Cretaceous Oceanic Anoxic Event 1b.- Scientific Reports, vol. 10, no. 12601, 10 p. DOI: https://doi.org/10.1038/s41598-020-69505-x
McAnena A., Fl gel S., Hofmann P., Herrle J.O., Griesand A., Pross J., Talbot H.M., Rethemeyer J., Wallmann K. & Wagner T. (2013).- Atlantic cooling associated with a marine biotic crisis during the mid-Cretaceous period.- Nature Geoscience, vol. 6, p. 558-561.
Melo R.M., Guzm n J., Almeida-Lima D., Piovesan E.K., Neumann V.H. de M.L. & Sousa A. de J. (2020).- New marine data and age accuracy of the Romualdo Formation, Araripe Basin, Brazil.- Scientific Reports, vol. 10, no. 15779, 15 p. DOI: https://doi.org/10.1038/s41598-020-72789-8
Michels F.H., Souza P.A. de & Premaor E. (2018).- Aptian-Albian palynologic assemblages interbedded within salt deposits in the Esp�rito Santo Basin, eastern Brazil: Biostratigraphical and paleoenvironmental analysis.- Marine and Petroleum Geology, vol. 91, p. 785-799.
Milani E.J., Rangel H.D., Bueno G.V., Stica J.M., Winter W.R., Caixeta J.M. & Pessoa Neto O. da C. (2007).- Bacias sedimentares brasileiras - Cartas estratigr�ficas.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 183-205.
Moreira J.L.P., Madeira C.V., Gil J.A. & Machado M.A.P. (2007).- Bacia de Santos.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 531-549.
Morgan R. (1978).- Albian to Senonian Palynology of Site 364, Angola Basin. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 915-951. DOI: http://deepseadrilling.org/40/volume/dsdp40_26.pdf
Moullade M., Bellier J.-P. & Tronchetti G. (2002).- Hierarchy of criteria, evolutionary processes and taxonomic simplification in the classification of Lower Cretaceous planktonic foraminifera.- Cretaceous Research, vol. 23, p. 111-148.
Mutterlose J., Bornemann A., Luppold F.W., Owen H.G., Ruffell A., Weiss W. & Wray D. (2003).- The V�hrum section (northwest Germany) and the Aptian/Albian boundary.- Cretaceous Research, vol. 24, p. 203-252.
NASC (1983).- North American Stratigraphic Code.- AAPG Bulletin, Tulsa - OK, vol. 67, no. 5, p. 841-875. URL: https://ngmdb.usgs.gov/Info/NACSN/05_1547.pdf
Norris R.D., Kroon D., Klaus A. et al. (1998).- Section 4: CORES. Site 1049. In: LEG 171B. Blake Nose Paleoceanographic Transect. Sites 1049-1053.- Proceedings of the Ocean Drilling Program, Initial Reports, Washington - DC, vol. 171B, p. 363-400. DOI: https://doi.org/10.2973/odp.proc.ir.171B.103.1998
Ogg J., Agterberg F.P. & Gradstein F. (2004).- The Cretaceous Period. In: Gradstein F., Ogg J. & Smith A. (eds.), A geologic time scale 2004.- Cambridge University Press, Cambridge (UK), p. 344-383.
Ogg J.G. & Hinnov L.A. (2012).- Cretaceous. In: Gradstein F., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), The geologic time scale 2012.- Elsevier, Amsterdam, p. 793-853.
Ogg J.G., Ogg G.M. & Gradstein F.M. (2016).- 13 - Cretaceous. In: Ogg J.G., Ogg G.M. & Gradstein F.M. (eds.), A concise geologic time scale 2016.- Elsevier, Amsterdam, p. 167-186.
Owen H.G. (2002).- The base of the Albian Stage; comments on recent proposals.- Cretaceous Research, vol. 23, p. 1-13.
Petersohn E. (2019).- Pre-Salt super play: Leading Brazil into the World's top 5 oil suppliers.- AAPG Latin America and Caribbean Region Geoscience Technology Workshop, Rio de Janeiro, 1 p. (abstract).
Petrizzo M.R., Huber B.T., Gale A.S., Barchetta A. & Jenkyns H.C. (2012).- Abrupt planktic foraminiferal turnover across the niveau kilian at col de pr�-Guittard (Vocontian Basin, southeast France): New criteria for defining the Aptian/Albian boundary.- Newsletters on Stratigraphy, vol. 45, no. 1, p. 55-74.
Petrizzo M.R., Wagreich M. & Haggart J. (2016).- Annual Report 2016 of the International Subcommission on Cretaceous Stratigraphy.- ISCS, 13 p. URL: https://cretaceous.stratigraphy.org/files/ICS2016_Report_Creta.pdf
Premoli-Silva I., Caron M., Leckie M.R., Petrizzo M.R., Soldan D. & Verga D. (2009).- Paraticinella n.gen. and taxonomic revision of Ticinella bejaouaensis Sigal.- Journal of Foraminiferal Research, Lawrence - KS, vol. 39, no. 2, p. 126-137.
Proto-Decima F., Medizza F. & Todesco L. (1978).- Southeastern Atlantic Leg 40 calcareous nannofossils. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 571-634. DOI: https://doi.org/10.2973/dsdp.proc.40.112.1978
Rangel H.D., Martins F.A.L., Esteves F.R. & Feij� F.J. (1994).- Bacia de Campos.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 8, no. 1, 203-217.
Regali M. da S.P. (1989).- A idade dos evaporitos da plataforma continental do Cear�, Brasil, e sua rela��o com os outros evaporitos das bacias nordestinas.- Boletim do IG-USP, S�o Paulo, Publica��o Especial, vol. 7, p. 139-143. DOI: https://doi.org/10.11606/issn.2317-8078.v0i7p139-143
Regali M. da S.P. & Viana C.F. (1989).- Sedimentos do NeoJur�ssico-EoCret�ceo do Brasil: Idade e correla��o com a escala internacional.- Ed. G�vea, Petrobras, Rio de Janeiro, 95 p.
Regali M. da S.P. & Silva Santos P.R. (1999).- Palinoestratigrafia e geocronologia dos sedimentos Albo-Aptianos das bacias de Sergipe e de Alagoas - Brasil. In: Boletim do 5o Simp�sio Sobre o Cret�ceo do Brasil.- UNESP, Rio Claro, p. 411-419.
Regali M. da S.P., Uesugui N. & Silva Santos A. (1974).- Palinologia dos sedimentos Meso-cenoz�icos do Brasil.- Boletim T�cnico da Petrobras, Rio de Janeiro, vol. 17, no. 3, p 177-191.
Reyment R.A. & Dingle R.V. (1987).- Palaeogeography of Africa during the Cretaceous Period.- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 59, p. 93-116.
Rodriguez C.R., Jackson C.A-L., Rotevatn A., Bell R.E & Francis M. (2018).- Dual tectonic-climatic controls on salt giant deposition in the Santos Basin, offshore Brazil.- Geosphere, Boulder - CO, vol. 14, no. 1, p. 215-242. DOI: https://doi.org/10.1130/GES01434.1
Roth P.H. (1981).- Mid-Cretaceous calcareous nannoplankton from the central Pacific: implication for paleoceanography. In: Thiede J., Vallier T.L., Adelseck C.G., Boersma A., Cep�k P., Dean W.E., Fujii N., Koporulin V.I., Rea D.K., Sancetta C., Sayre W.O., Seifert K., Schaaf A., Schmidt R.R., Windom K. & Vincent E. (eds.), Leg 62 of the cruises of the Drilling Vessel Glomar Challenger. Marjuro Atoll, Marshall Islands to Honolulu, Hawaii. July-September 1978.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXII, p. 471-489. DOI: https://doi.org/10.2973/dsdp.proc.62.113.1981
Sabatino N., Coccioni R., Manta D.S., Baudin, F., Vallefuoco M., Traina A. & Sprovieri M. (2015).- High-resolution chemostratigraphy of the late Aptian-early Albian oceanic anoxic event (OAE 1b) from the Poggio le Guaine section (Umbria-Marche Basin, central Italy).- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 426, p. 319-333.
Sanjin s A.E.S, Viviers M.C, Costa D.S., Anjos-Zerfass G.S., Burlen G. & Strohschoen O. Jr (2022).- Planktonic foraminifera from the Aptian section of the southeastern Brazilian Atlantic margin.- Cretaceous Research, vol. 134, article 105141, 16 p.
Schaller H. (1969).- Revis�o estratigr�fica da Bacia de Sergipe/Alagoas.- Boletim T�cnico da Petrobras, Rio de Janeiro, vol. 12, no. 1, p. 21-86.
Selby D., Mutterlose J. & Condon D.J. (2009).- U-Pb and Re-Os geochronology of the Aptian/Albian and Cenomanian/Turonian stage boundaries: Implications for timescale calibration, osmium isotope seawater composition and Re-Os systematics in organic-rich sediments.- Chemical Geology, vol. 265, nos. 3-4, p. 394-409.
Sliter W.V. (1989).- Biostratigraphic zonation for Cretaceous planktonic foraminifera examined in thin section.- Journal of Foraminiferal Research, Lawrence - KS, vol. 19, no. 1, p. 1-19.
Spadini A.R. (1992, unpublished).- Processos deposicionais e ciclicidade em carbonatos albianos de plataforma rasa da Bacia de Campos.- MSc, Disserta��o de mestrado, Universidade Federal do Rio de Janeiro, 139 p.
Spadini A.R., Esteves F.R., Dias-Brito, D., Azevedo R.L.M. & Rodrigues R. (1988).- The Maca� Formation, Campos Basin, Brazil: Its evolution in the context of the initial history of the South Atlantic.- Revista Brasileira de Geoci�ncias, S�o Paulo, vol. 18, no. 3, p. 261-272.
Spadini A.R. & Pra a U.M. (1989).- Glauconita em uma anomalia radioativa em calc�rios do Albiano da bacia de Campos: Um marco estratigr�fico com significado paleoambiental. In: Anais do XI Congresso Brasileiro de Paleontologia, Curitiba.- Sociedade Brasileira de Paleontologia, Rio de Janeiro, p. 1017-1028.
Szatmari P. & Milani E.J. (2016).- Tectonic control of the oil-rich large igneous-carbonate-salt province of the South Atlantic rift.- Marine and Petroleum Geology, vol. 77, p. 567-596.
Szatmari P., Mor de Lima C., Fontaneta G., Melo Lima N. de, Zambonato E., Menezes M.R., Bahniuk J., Coelho S.L., Figueiredo M., Florencio C.P. & Gontijo R. (2021).- Petrography, geochemistry and origin of South Atlantic evaporites: The Brazilian side.- Marine and Petroleum Geology, vol. 127, article 104805, 31 p.
Tedeschi L.R., Jenkyns H.C., Robinson S.A., Sanjin s A.E.S., Viviers M.C., Quintaes C.M.S.P. & Vazquez J.C. (2017).- New age constraints on Aptian evaporites and carbonates from the South Atlantic: Implications for Oceanic Anoxic Event 1a.- Geology, Boulder - CO, vol. 45, no. 6, p. 543-546. DOI: https://doi.org/10.1130/G38886.1
Tissot B., Demaison G.J., Masson P., Delteil J.R. & Combaz A. (1980).- Paleoenvironment and petroleum potential of middle Cretaceous black shales in Atlantic basins.- AAPG Bulletin, Tulsa - OK, vol. 24, no. 12, p. 2051-2063.
Trabucho Alexandre J., Tuenter E., Henstra G.A., Zwan K.J. van der, Wal R.S.W. van de, Dijkstra H.A. & Boer P.L. de (2010).- The mid-Cretaceous North Atlantic nutrient trap: Black shales and OAEs.- Paleoceanography, vol. 25, no. 4, PA4201, 14 p. DOI: https://doi.org/10.1029/2010PA001925
Viana C.F., Gama E.G. Jr, Sim es I.A., Moura J.A., Fonseca J.R. & Alves R.J. (1971).- Revis�o estratigr�fica da Bacia do Rec�ncavo/Tucano.- Boletim T�cnico da Petrobras, Rio de Janeiro, vol. 14, no. 3/4 p. 57-92.
Viviers M.C. (1987).- Foramin�feros planct�nicos no Cret�ceo m�dio da Bacia de Santos, Brasil. Revista Brasileira de Geologia, S�o Paulo, vol. 17, no. 2, p. 154-161. URL: https://www.ppegeo.igc.usp.br/index.php/rbg/article/view/11903
Viviers M.C. & Beurlen G. (1987).- Sobre alguns n veis de correla o bioestratigr fica regional (foramin feros planct nicos) no Cret ceo brasileiro, Revista Brasileira de Geologia, S�o Paulo, vol. 17, no. 2, p. 106-110. URL: https://www.ppegeo.igc.usp.br/index.php/rbg/article/view/11891
Viviers M.C., Sanjin s A.E.S., Costa D.S., Anjos-Zerfass G. de S., Beurlen G. & Strohschoen O. Jr (2018).- Aptian planktonic foraminifera occurrence in the southeastern brazilian marginal basins. In: RIO 2018, 49o Congresso Brasileiro de Geologia.- Sociedade Brasileira de Geologia (SBG), Rio de Janeiro, p. 2027 (abstract). URL: http://cbg2018anais.siteoficial.ws/resumos/7442.pdf
Wagner T., Herrle J.O., Damst J.S.S., Schouten S., St sser I. & Hofmann P. (2008).- Rapid warming and salinity changes of Cretaceous surface waters in the subtropical north Atlantic.- Geology, Boulder - CO, vol. 36, no. 3, p. 203-206.
Wiedmann J. & Neugebauer J. (1978).- Lower Cretaceous ammonites from the South Atlantic Leg 40 (DSDP), their stratigraphic value and sedimentological properties. In: Bolli H.M., Ryan W.B.F., McKnight B.K., Kagami H., Melguen M., Siesser W.G., Natland J.H., Longoria J.F., Proto Decima F., Foresman J.B. & Hottman W.E. (eds.), Leg 40 of the cruises of the Drilling Vessel Glomar Challenger. Cape Town, South Africa to Abidjan, Ivory Coast. December 1974-February 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XL, p. 709-734. DOI: https://doi.org/10.2973/dsdp.proc.38394041s.312.1978
Wiegand G.E. (1984).- Cretaceous Nannofossils from the Northwest African Margin, Deep Sea Drilling Project Leg 79. In: Hinz K., Winterer E.L., Baumgartner P.O., Bradshaw M.J., Channell J.E.T., Jaffrezo M., Jansa L.F., Leckie R.M., Moore J.N., Rullk tter J., Schaftenaar C., Steiger T.H., Vuchev V. & Wiegand G.E. (eds.), Leg 79 of the cruises of the Drilling Vessel Glomar Challenger. Las Palmas, Grand Canary Island, to Brest, France. April-May 1981.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXIX, p. 563-578. DOI: https://doi.org/10.2973/dsdp.proc.79.121.1984
Winter W.R., Jahnert R.J. & Fran a A.B. (2007).- Bacia de Campos.- Boletim de Geoci�ncias da Petrobras, Rio de Janeiro, vol. 15, no. 2, p. 511-529.
Wise S.W. (1983).- Mesozoic and Cenozoic calcareous nannofossils recovered by DSDP Leg 71 in the Falkland Plateau region, Southwest Atlantic Ocean. In: Wise S.W. Jr, Blakeslee J.H. & Lee M. (eds.), Leg 71 of the cruises of the Drilling Vessel Glomar Challenger. Valparaiso, Chile, to Santos, Brazil. January-February, 1980.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXI, p. 481-550. DOI: https://doi.org/10.2973/dsdp.proc.71.121.1983
Yamina B. [sic], Nabiha B.H.A. [sic], Saloua R. [sic] & Kamal T. [sic] = Bourgeoini Y., Ben Haj Ali N., Razgallah S. & Tajeddine K. (2002).- �tude biostratigraphique du Cr�tac� inf�rieur (Barr�mien sup�rieur-Albien) du Haut Atlas occidental (Maroc).- Estudios Geol�gicos, vol. 58, 109-116. URL: https://estudiosgeol.revistas.csic.es/index.php/estudiosgeol/article/view/116/112
Young J.R., Bown P.R. & Lees J.A. (eds., 2022a).- Nannotax3 website.- International Nannoplankton Association.- Accessed: 2022/12/06. URL: http://www.mikrotax.org/Nannotax3
Young J.R., Wade B.S. & Huber B.T. (eds., 2022b).- pforams@mikrotax website.- Accessed: 2022/12/06. URL: http://www.mikrotax.org/pforams
mbsf = meters below surface; FO = first occurrence datum; LO = last occurrence datum; Bs = basin; Sc = section.
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO P. cretacea (= P. columnata) | #55-1 | 480.70 | Cep k (1981); Roth (1981) |
| Paquier black shale (top) | #56-1 | 490.95 | Dean et al. (1981) |
| FO T. primula | #58-3 | 513.54 | Matsumoto et al. (2020) |
| FO M. rischi | #59-3 | 522.83 | Matsumoto et al. (2020) |
| δ13Ccarb excursion - Kilian Level | #60-1 | 529.33 | Matsumoto et al. (2020) |
| FO M. renilaevis | #60-2 | 529.56 | Matsumoto et al. (2020) |
| LO M. miniglobularis | #60-2 | 530.19 | Matsumoto et al. (2020) |
| LO Large planktonic foraminifers (~LO P. rohri) | #60-2 | 530.19 | Matsumoto et al. (2020) |
| δ13Ccarb excursion - Jacob Level | #62-1 | 538.64 | Matsumoto et al. (2020) |
Comments: The δ13Ccarb excursions related to the Jacob and Kilian levels are dissociated from any organic-rich layer; the black shale occurring in this section has been associated with the Paquier Level but is not accompanied by any relevant oscillation in δ13Ccarb.
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO T. primula | #34R-1 | 494.22 | Huber & Leckie (2011) |
| FO T. orionatus | #34X-1 | 494.28 | Bralower & Siesser (1992) |
| FO T. madecassiana | #35R-5 | 509.69 | Huber & Leckie (2011) |
| FO H. albiensis | #36R | 517.80 | Bralower & Siesser (1992) |
| FO P. spinosa | #37X-2 | 524.29 | Bralower & Siesser (1992) |
| FO M. miniglobularis | #37R-3 | 525.74 | Huber & Leckie (2011) |
| FO M. renilaevis | #37R-3 | 525.74 | Huber & Leckie (2011) |
| FO P. columnata | #37X-5 | 528.75 | Bralower & Siesser (1992) |
| FO M. rischi | #37R-5 | 529.45 | Huber & Leckie (2011) |
| Unconformity | #37R-5 | between 529.45 and 530.23 | Huber & Leckie (2011) |
| LO large planktonic foraminifers (~LO P. rohri) | #37R-5 | 530.23 | Huber & Leckie (2011) |
Comments: Supported by Huber and Leckie (2011), the last occurrence (LO) of P. transitoria is adopted as the datum for the large planktonic foraminifera (P. rohri Zone), covered by the M. rischi Zone, which characterizes a hiatus; the absence of P. spinosa and the Jacob Level determined the position of the base of the hiatus; occurrences of M. renilaevis, M. miniglobularis, P. spinosa and P. columnata were considered as reworked or in a stratigraphic position above their first occurrences (FOs); the section has low TOC values (< 2%), and no δ13Ccarb information was found.
1.3. V�hrum Sc., Lower Saxony Basin, Germany
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO P. columnata (subcircular) | Vo 15 | 2.15 | Mutterlose et al. (2003) |
| Ash layer | Vo 15 | 2.12 | Mutterlose et al. (2003) |
| FO P. spinosa | Vo 1 | 0.1 | Mutterlose et al. (2003) |
Comments: No planktonic foraminifera were recovered in the section and the palynomorphs present have no chronostratigraphic value; the section has low TOC values (< 1.3%) and no δ13Ccarb information was obtained.
1.4. Poggio le Guaine Sc., Umbria-Marche Basin, Italy
| Event | Depth (mbsf) |
Reference |
| T. primula | 21 | Coccioni et al. (2014) |
| Black shale (top) | 18 | Coccioni et al. (2014) |
| δ13Ccarb excursion | 17.71-18 | Coccioni et al. (2014) |
| Black shale (base) | 17.71 | Coccioni et al. (2014) |
| Urbino black shale (top) | 16.37 | Coccioni et al. (2014) |
| FO H. albiensis | 16.3 | Coccioni et al. (2014) |
| δ13Ccarb excursion (Urbino) | 16.12-16.37 | Coccioni et al. (2014) |
| Urbino black shale (base) | 16.12 | Coccioni et al. (2014) |
| FO P. columnata (circular) | 15.5 | Coccioni et al. (2014) |
| FO M. rischi | 10.8 | Coccioni et al. (2014) |
| Black shale (top) | 8.37 | Coccioni et al. (2014) |
| δ13Ccarb excursion | 7.97-8.37 | Coccioni et al. (2014) |
| Black shale (base) | 7.97 | Coccioni et al. (2014) |
| FO M. renilaevis | 7.6 | Coccioni et al. (2014) |
| FO P. spinosa | 6.5 | Coccioni et al. (2014) |
| FO P. columnata (near-circular) | 6.5 | Coccioni et al. (2014) |
| LO large planktonic foraminifers (~LO P. rohri) | 6.5 | Coccioni et al. (2014) |
| FO M. miniglobularis | 6.42 | Coccioni et al. (2014) |
| FO P. columnata (subcircular) | 4 | Coccioni et al. (2014) |
| 113/Jacob black shale (top) | 3.55 | Coccioni et al. (2014); Sabatino et al. (2015) |
| δ13Ccarb excursion (Jacob) | 3.47-3.55 | Coccioni et al. (2014); Sabatino et al. (2015) |
| 113/Jacob black shale (base) | 3.47 | Coccioni et al. (2014); Sabatino et al. (2015) |
Comments: A biostratigraphic inversion was observed in this section, with the FO of M. miniglobularis preceding the LO of P. rohri; the black shale associated to the Kilian level identified by Coccioni et al. (2014) occurring a few meters above the FO of M. renilaevis, outside of the position defined by the GSSP-Alb; the same problem occurs with the Urbino Layer, with high TOC values, considered by these authors as being chronocorrelated to the Paquier Level even though it is located below the FO of H. albiensis; the occurrence of another horizon rich in organic matter above, but below the FO of T. primula (indicated as being 2 m above the studied section), makes it impossible to chronocorrelate it with the Leenhardt Level.
1.5. First composed sections from Vocontian Basin, France (Col de Pr�-Guittard and Tartonne sections)
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO T. orionatus | PG29 | 125.5 | P. Guittard Sc, Kennedy et al. (2000). Biostratigraphic chart. Depth from Fig. 21 |
| FO T. primula | PG17 | 72.5 | P. Guittard Sc, Kennedy et al. (2000). Depth from Fig. 21 |
| FO P. columnata (circular) | PG17 | 72.5 | P. Guittard Sc, Kennedy et al. (2000). Biostratigraphic chart. |
| Paquier black shale (top) | below PG16 | 70 | P. Guittard Sc, Kennedy et al. (2000). Depth from Fig. 21. TOC >2 (Fig. 23) |
| δ13Ccarb excursion Paquier | PG15-PG16 | 68-70 | P. Guittard Sc, Kennedy et al. (2000). Smooth negative δ13C excursion (Fig. 25) |
| Paquier black shale (base) | above PG15 | 68 | P. Guittard Sc, Kennedy et al. (2000). Depth from Fig. 21. TOC >2 (Fig. 23) |
| Kilian black shale (top) | below PG9 | 38.5 | P. Guittard Sc, Kennedy et al. (2000). TOC >2 (Fig. 23) |
| δ13Ccarb on the Kilian Level | PG8-PG9 | 38.5-37.5 | P. Guittard Sc, Kennedy et al. (2000). No δ13C excursion detected (Fig. 25) |
| Kilian black shale (base) | above PG8 | 37.5 | P. Guittard Sc, Kennedy et al. (2000). TOC >2 (Fig. 23) |
| FO P. columnata (near-circular) | PG8 | 36 | P. Guittard Sc, Kennedy et al. (2000). Biostratigraphic chart |
| LO large planktonic foraminifers (~LO P. rohri) | PG6 | 26 | P. Guittard Sc, Kennedy et al. (2000) |
| FO P. columnata (subcircular) | PG2 | 6 | P. Guittard Sc, Kennedy et al. (2000) |
| Jacob level (top) | below PG2 | 4 | P. Guittard Sc, Kennedy et al. (2000). Low values of TOC (Fig. 23) |
| δ13 Ccarb on the Jacob Level | PG1-PG2 | 4-2.5 | P. Guittard Sc, Kennedy et al. (2000). No δ13C excursion detected (Fig. 25) |
| Jacob level (base) | above PG1 | 2.5 | P. Guittard Sc, Kennedy et al. (2000). Low values of TOC (Fig. 23) |
| FO P. spinosa | PG1 | 1 | P. Guittard Sc, Kennedy et al. (2000). Biostratigraphic chart. Depth from Fig. 21 |
| Paquier black shale (top) | above TO20 | 33 | Tartone Sc, Kennedy et al. (2000). High values of TOC (Fig. 24) |
| δ13Ccarb on the Paquier Level | TO10 | 33-21.8 | Tartonne Sc, Kennedy et al. (2000). Depth from Fig. 24. Smooth negative δ13C excursion (Fig. 26) |
| Paquier black shale (base) | TO10 | 21.8 | Tartonne Sc, Kennedy et al. (2000). High values of TOC (Fig. 24) |
Comments: Kennedy et al. (2000)' paper was published before the planktonic foraminifera taxonomic review of Huber and Leckie (2011), which justifies the limited number of key species; Col de Pr�-Guittard is the section where the Jacob, Kilian, Paquier and Leenhardt levels were described (Br�h�ret et al., 1986), which required the adjustment of the biostratigraphic positions to these lithological data, which justifies the FO T. primula outside the expected position in the GTS2020; TOC and δ13Ccarb information come from samples separated by metric spacing, which may have masked the detection of excursions of this last variable; only the Paquier Level recorded relevant variations of δ13Ccarb.
1.6. Second composed sections from Vocontian Basin, France (Serre Chaitieu, Gaubert, Tarendol, Pr�-Guittard, Les Oustaus, l'Arboudeysse, and Col de Palluel sections)
| Event | Depth (mbsf) |
Reference |
| Paquier black shale (top) | 281.5 | l'Arboudeysse Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| Paquier black shale (base) | 280.5 | l'Arboudeysse Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| FO H. albiensis | 244.5 | Herrle & Mutterlose (2003). Depth from Fig. 5 |
| Kilian bed (top) | 241.6 | C. P. Guittard Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| Kilian bed (base) | 241.6 | C. P. Guittard Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| LO large planktonic foraminifers (~LO P. rohri) | 234.5 | Composite lithological column. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| FO P. columnata (circular) | 214.5 | Tarendol Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| Jacob black shale (top) | 201 | Tarendol Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| Jacob black shale (base) | 200 | Tarendol Sc. Depth from Fig. 5 (Herrle & Mutterlose, 2003) |
| FO P. spinosa | 194.5 | Herrle & Mutterlose (2003). Depth obtained in Fig. 5 |
Comments: The sections were studied seeking to establish the succession of calcareous nannofossils in the Vocontian Basin; the circular shape of P. columnata was the variety found in the oldest horizon; planktonic foraminifera biostratigraphy was based on Moullade et al. (1998) and Kennedy et al. (2000); no information on δ13Ccarb values was available.
1.7. Col de Pr�-Guittard Sc., GSSP-Alb, Vocontian Basin, France
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO M. rischi | 73 | Kennedy et al. (2017). Depth from Fig. 4 | |
| Paquier black shale (top) | 70 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| δ13C Paquier excursion | 25 | 68-70 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) |
| Paquier black shale (base) | 68 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| FO P. columnata (circular - consistent) | 17 | 66.6 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) |
| Kilian bed (top) | 37.8 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017). | |
| δ13C excursion Kilian | 37-37.8 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| FO M. renilaevis | 13 | 37.4 | Kennedy et al. (2017). Depth from Fig. 4 |
| Kilian bed (base) | 37 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| FO M. miniglobularis | 8 | 35 | Kennedy et al. (2017). Depth from Fig. 4 |
| LO large planktonic foraminifers (~LO P. rohri) | 7 | 34.75 | Kennedy et al. (2017). Depth from Fig. 4 |
| FO P. columnata (circular) | 5 | 29.5 | Kennedy et al. (2017). Depth from Fig. 4 |
| FO P. columnata (subcircular) | 3 | 6 | Kennedy et al. (2017). Depth from Fig. 4 |
| Jacob black shale (top) | 201 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| No δ13C on the Jacob level | 200-201 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| Jacob black shale (base) | 200 | Herrle et al. (2004) and Ogg et al. (2016). Depth from Fig. 4 (Kennedy et al., 2017) | |
| FO P. spinosa | PG1 | 1 | Depth from Kennedy et al. (2000) |
Comments: This is the GSSP-Alb's reference section; the position of the FO of P. spinosa was based on Kennedy et al. (2000); the FO M. rischi was not found; the names of the black shales with δ13Ccarb excursions were taken from previous studies.
1.8. Composed section from Agadir-Essaouira Basin, Marocco (Ida w Shayq, Tissakatine Center, Anzate, and Tinfoul)
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| LO large planktonic foraminifers (~LO P. rohri) ? | EAB | These sections belong to Oeud Tidzi/Tamgo Fms, where the planktonic foraminifera turnover occurs (Hassanein, 2016; Yamina et al. [sic Bourgeoini et al.], 2002) | |
| FO H. albiensis | ISA25 | 29.05 | Ida w Shayq sc. Above the D4 unconformity (Hassanein, 2016) |
| FO P. columnata (subcircular) | ISA24 | 28.7 | Ida w Shayq sc (Hassanein, 2016) |
| FO P. columnata (near circular) | ISA23 | 27.9 | Ida w Shayq sc (Hassanein, 2016) |
| FO P. columnata (circular) | ISA22 | 26.6 | Ida w Shayq sc (Hassanein, 2016) |
| FO H. albiensis | TKC66a | 41 | Tissakatine sc. Above the D4 unconformity. Depth from Fig. 3.11 (Hassanein, 2016) |
| FO P. columnata (subcircular) | TKC63a | 37 | Tissakatine sc. Depth from Fig. 3.11 (Hassanein, 2016) |
| FO H. albiensis | TF16 | 49.5 | Tinfoul sc. Above the D4 unconformity Depth from Fig. 3.13 (Hassanein 2016) |
| FO P. columnata (circular and near-circular) | TF7 | 28 | Tinfoul sc. Depth from Fig. 3.13 (Hassanein, 2016) |
| FO P. spinosa | TF2 | 16.5 | Tinfoul sc. Depth from Fig. 3.13 (Hassanein, 2016) |
Comments: COT values are low throughout the section; the long-spaced intervals between samples and diagenetic processes around the D4 discontinuity does not permit indicate reliable δ13Ccarb excursions; the LOs large planktonic foraminifers have dubious indications based on lithostratigraphic and ammonites biozones correlations.
1.9. Site 545-DSDP, Mazagan Plateau, Morocco
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| Unconformity | 40-6 | between 377.06 and 377.11 |
Wiegand (1984); Huber & Leckie (2011) |
| LO large planktonic foraminifers (~LO P. rohri) | 40-6 | 377.11 | Huber & Leckie (2011) |
| Black shale (top) | 42 | 388.8 | Wagner et al. (2008); Trabuco-Alexandre et al (2011); McAnena et al. (2013) |
| δ13C excursion | 42 | 388.8-390.45 | Herrle et al. (2004); Wagner et al. (2008) |
| Black shale (base) | 42 | 390.45 | Wagner et al. (2008); Trabuco-Alexandre et al (2011); McAnena et al. (2013) |
| Acme Subtilisphaera spp. | 43-1 | 401.2 | Below (1984) |
| FO T. orionatus | 44-1 | 406.63 | Wiegand (1984) |
| FO H. albiensis | 45-4 | 422.2 | Wiegand (1984) |
| FO P. cretacea (= P. columnata) | 47-3 | 439.3 | Wiegand (1984) |
| Acme Subtilisphaera spp. | 48-6 | 453.2 | Below (1984) |
Comments: An unconformity is indicated in this section based on the absence of several zones in the interval between the disappearance of the large foraminifera (~P. rohri) and the FOs of many species related to Late Albian in age (Huber & Leckie, 2011); there is also an inversion in the biochronostratigraphic succession, with the FOs of H. albiensis and T. orionatus preceding the large foraminifera turnover; also the anoxic event has its stratigraphic position marked by biostratigraphic data and cannot be related to any of the OAE1b levels.
1.10. Site 1049C-ODP, Blake Nose, USA
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| Unconformity | 11X-2 | 132.31-132.44 | Huber & Leckie (2011) |
| FO T. madecassiana | 11cc | 134.53 | Huber & Leckie (2011) |
| Black shale (top) | 12X-2 | 142,75 | Measure on core description (Norris et al., 1998) |
| No δ13C excursion | 12X-2 | 142.75-143.1 | Huber & Leckie (2011) |
| Black shale (base) | 12X-2 | 143.1 | Measure on core description (Norris et al., 1998) |
| FO P. columnata s.s. | 12 | 143.2 | Browning & Watkins (2008). Depth from Fig. 2 |
| FO M. rischi | 12X-4 | 145.26 | Huber & Leckie (2011) |
| FO M. renilaevis | 12X-4 | 145.26 | Huber & Leckie (2011) |
| Unconformity | 11X-2 | 145.26-145.29 | Huber & Leckie (2011) |
| LO large planktonic foraminifers (~LO P. rohri) | 12X-4 | 145.29 | Huber & Leckie (2011) |
| FO H. albiensis | 12X-5 | 145.4 | Browning & Watkins (2008). Depth from Fig. 2 |
| FO P. columnata s.l. | 12X-5 | 146.4 | Browning & Watkins (2008). Depth from Fig. 2 |
| FO P. spinosa | 12X-6 | 146.8 | Browning & Watkins (2008). Depth from Fig. 2 |
Comments: Two unconformities are inferred based on bioevents successions; the first is indicated by the absence of the M. miniglobularis Zone and the fact that M. renilaevis and M. rischi have their local FO happening at the same time; the second unconformity is defined by the presence of Campanian fossils succeeding the T. madecassiana Zone (Huber & Leckie, 2011); there is also an inversion in the biochronostratigraphic succession, with the FO of H. albiensis preceding the disappearance of the large foraminifera (~P. rohri); the biostratigraphic information's shows that the anoxic event is not related to any of the OAE1b levels.
1.11. Linderos Section, Tampico-Misantla Basin, Mexico
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| LO Colomiella recta | LIN 66.1 | 64.5 | Guti�rrez-Puente et al. (2021) |
| FO T. primula | LIN 51 | 49 | Guti�rrez-Puente et al. (2021) |
| FO T. madecassiana | LIN 48.2 | 46.5 | Guti�rrez-Puente et al. (2021) |
| FO Colomiella recta | LIN 47.2 | 45.5 | Guti�rrez-Puente et al. (2021) |
| LO Colomiella mexicana | LIN 47.2 | 45.5 | Guti�rrez-Puente et al. (2021) |
| FO Microhedbergella Zone | LIN 47 | 45 | Guti�rrez-Puente et al. (2021) |
| LO large planktonic foraminifers (~LO P. rohri) | LIN 46.2 | 44.5 | Guti�rrez-Puente et al. (2021) |
| FO Colomiella mexicana | LIN 45.1 | 43.2 | Guti�rrez-Puente et al. (2021) |
Comments: The taxonomic classification was based exclusively on thin sections; the identification of Microhedbergella was limited to the generic level; there is no information about calcareous nannofossils, COT or δ13Ccarb.
1.12. Site 511-DSDP, Malvinas Plateau, Argentina
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO T. orionatus | 51-cc | 451 | Wise (1983) |
| FO H. albiensis | 51-cc | 451 | Wise (1983) |
| FO M. rischi | 55-4 | 484.75 | Huber & Leckie (2011), Table 2 |
| FO M. renilaevis | 55-4 | 485.11 | Huber & Leckie (2011), Table 2 |
| LO large planktonic foraminifers (~LO P. rohri) | 55-5 | 486.14 | Huber & Leckie (2011), Table 2 |
| LO M. miniglobularis | 55-1 | 486.7 | Huber & Leckie (2011), Table 2 |
| FO P. cretacea (=P. columnata) | 56-1 | 491 | Wise (1983) |
| FO P. spinosa | 56-1 | 491 | Wise (1983) |
Comments: A biostratigraphic inversion is noted here, in which the FO of M. miniglobularis precedes the LO of P. rohri.
1.13. Site 363-DSDP, Walvis Ridge
| Event | Stratigraphic position |
Depth (mbsf) |
Reference |
| FO E. turriseiffelli | #34-3, 41-42 | 596 | Proto Decima et al. (1978) |
| Unconformity | #34-3, 41-42 | 596 | This study |
| LO large planktonic foraminifers (~LO P. rohri) ? | #34-3, 41-42 | 596 | This study |
| FO H. albiensis | #38-1, 90-91 | 668.41 | Proto Decima et al. (1978) |
| FO. P. cf sphaerica | #39-2, 92-94 | 688.94 | Bolli (1978) |
| FO P. cretacea | #38-1, 90-91 | 669.25 | Proto Decima et al. (1978) |
| Black shale | Not drilled | Inferred by Bolli (1978) | |
| Evaporite layer | Not drilled | Inferred by Bolli (1978) | |
| Volcanic layer | Not drilled | Inferred by Bolli (1978) |
Comments: An unconformity marks the top of the H. algeriana/T. bejaouensis zone, defined in Caron (1978); it is indicated by the presence of the FOs of Effeillitus turriseiffelli (Proto Decima et al., 1978) and Clavihedbergella simplissima (Caron, 1978), Late Albian age.
1.14. Site 364-DSDP, Kwanza Basin, Angola
| Event | Stratigraphic level |
Depth (mbsf) |
Reference |
| FO E. turriseiffelli | #29-4 | 772.3 | Bruno et al. (2020) |
| FO P. ticinensis | #31 | 807.4 | Kochhann et al. (2013) |
| Unconformity | #31 | 807.4 | Kochhann et al. (2013) |
| FO M. rischi | #32 | 829.5 | Kochhann et al. (2013) |
| LO large planktonic foraminifers (~LO P. rohri) | #33-4 | 844.6 | Kochhann et al. (2013) |
| FO E. monechiae | #34-3 | 876.0 | Bruno et al. (2020) |
| LOs H. trochoidea and G. barri | #34 | 877.4 | Kochhann et al. (2013) |
| Black shale | #35-3 | 895.0 | Bruno et al. (2020) |
| FO A. biramiculatus | #38.3 | 951.5 | Bruno et al. (2020) |
| FO T. orionatus | #41.3 | 1008.4 | Bruno et al. (2020) |
| δ13Ccarb excursion | #41-3 | 1007.9-1009.2 | Azevedo (2001); Bruno et al. (2020) |
| FO H. albiensis | #42-5 | 1031.2 | Bruno et al. (2020) |
| δ13Ccarb strong negative excursion | #42-1, #42-1 | 1025.4-1026.9 | Azevedo (2001); Bruno et al. (2020) |
| FO P. columnata (circular) | #42-6 | 1032.4 | Bruno et al. (2020) |
| Several black shales | #39-1, #43-1 | 968.45-1034.6 | Foresman (1978); Bolli et al. (1978b); Azevedo (2001); Bruno et al. (2020) |
| Black shale | #44 - #46 | 1043.0-1086.5 | Bolli et al. (1978b) |
| Evaporite layer | Not drilled | Inferred by Bolli et al. (1978b) | |
| Volcanic layer | Not drilled | Inferred by Bolli et al. (1978b) |
Comments: The incompatibility between the biostratigraphic successions of planktonic foraminifera and calcareous nannofossils, previously recognized in 1978 and confirmed by Kochhann et al. (2013) and Bruno et al. (2020), makes the chronostratigraphic interpretation for the section difficult; It is elected to follow the stratigraphic positioning of the planktonic foraminifera, to the detriment of the international calcareous nannofossil markers; LOs H. trochoidea and G. barri are indicating by the G. ferreolensis/G. algerianus Zone symbol; sampling gap between P. eubejaouensis (= P. rohri) and M. rischi zones (844.6 and 829.5 mbsf) may have prevented the identification of the M. miniglobularis and M. renilaevis zones; the P. ticinensis Zone, Late Albian in age, rests in an unconformity over the M. rischi Zone; the presence of successive black shales levels in the last 120 mbsf of the perforated section is accompanied by at least two δ13Ccarb excursions.
1.15. Composite section of northeastern Brazilian basins
| Event | Stratigraphic position | Position or depth (mbsf) |
Reference |
| LO T. primula | Well SER-03. Sergipe Bs. | 117.51 | Fauth et al. (2022) |
| LO T. madecassiana | Well SER-03. Sergipe Bs. | 122.57 | Fauth et al. (2022) |
| Black shale | Well SER-03. Sergipe Bs. | 153.07 | Fauth et al. (2022) |
| LO E. jardinei | Equatorial South Atlantic, Brazilian northern interior and African basins | Relative position | Arai et al. (1989); Atta-Peters (2013) |
| FO Microhedbergella rischi | Well SER-03. Sergipe Bs. | 168.14 | Fauth et al. (2022) |
| FO Microhedbergella renilaevis | Well SER-03. Sergipe Bs. | 171.55m | Fauth et al. (2022) |
| FO Microhedbergella microglobularis | Well SER-03. Sergipe Bs. | 171.55m | Fauth et al. (2022) |
| δ13Ccarb positive excursion | Well SER-03. Sergipe Bs. | 178.4-176.62 | Fauth et al. (2022) |
| LO S. variverrucata | Equatorial South Atlantic, Brazilian northern interior and African basins | Relative position | Arai et al. (1989) |
| LO large planktonic foraminifers (~LO P. rohri) | Sobradinho Sc., Araripe Bs. | 38.4 | Melo et al. (2020) and this study |
| FO Upper acme zone of Subtilisphaera spp. | Well 1-CES-42A, Cear Bs. | ~2330-2600 | Regali (1989) |
| FO. P. columnata | Sergipe Bs. | Relative position | Cunha & Koutsoukos (1998) |
| FO Lower acme zone of Subtilisphaera spp. | Well 1-CES-46, Cear Bs. | ~1200-1700 | Regali (1989); Arai et al. (1994); Arai (2009, 2014) |
| Black shale | Sobradinho Sc., Araripe Bs. | ~35-42 | Cust dio et al. (2017) |
| Evaporite layer | Well 1-CES-46, Cear Bs. | ~1700-2400 | Regali (1989) |
Comments: The position of the FO of P. columnata was defined through stratigraphic correspondence with the G. ex gr. maridalensis - H. (H.) similis Zone (Koutsoukos, 1989; Koutsoukos & Bengtson, 1993); a younger acme of Subtilisphaera spp. were recognized by Arai et al. (1994) and Arai (2009; 2014); alternative interpretation is adopted in this study to indicate the LO large planktonic foraminifers (~LO P. rohri), based on Melo et al. (2020) informations; the top of the G. ex gr. maridalensis - H. (H.) similis Zone (Koutsoukos, 1989; Koutsoukos & Bengtson, 1993) also support the presence of this bioevent; the stratigraphic position to the youngest level of black shale is dubious.
1.16. Composite section of southeastern Brazilian basins
| Event | Stratigraphic position | Depth (mbsf) | Reference |
| FO T. primula | Well 3-BO-3, Campos Bs | Relative position | Dias-Brito (1999); Sanjin et al. (2022), Fig. 10 |
| LO large planktonic foraminifers (~LO P. rohri) |
Well CB-3 (=CB-3), Campos Bs. | 2450 | Tedeschi et al. (2017). Depth from Fig. 2 |
| Top of G. algerianus/G. ferreolensis Zone | Well CB-3 (=CB-3), Campos Bs. | 2484 | Tedeschi et al. (2017) |
| FO E. turriseiffelli | Well 3-BO-3, Campos Bs. | 2473 | Dias-Brito (1995). Depth from Fig. 92 |
| LO Ovoidinium diversum | Well 3-BO-3, Campos Bs. | 2476.6 | Dias-Brito et al. (1990) |
| FO P. cretacea (P. columnata) | Well 3-BO-3, Campos Bs. | 2515 | Dias-Brito (1995). Depth from Fig. 92 |
| LO Cyclopsiella spp. | Well 3-BO-3, Campos Bs. | 2516,9 | Dias-Brito et al. (1990) |
| FO P. sphaerica | Well 3-BO-3, Campos Bs. | 2602 | Dias-Brito (1995). Depth from Fig. 92 |
| FO Ovoidinium diversum | Well 3-BO-3, Campos Bs. | 2602 | Dias-Brito (1995). Depth from Fig. 92 |
| LO Colomiella recta | Well 1-SPS-6, Santos Bs. | 5220 | Dias-Brito (1999) |
| LO Colomiella mexicana | Well 1-SPS-6, Santos Bs. | 5487.7 | Dias-Brito (1999) |
| FO Cyclopsiella spp. | Integrate data | Relative position | Azevedo (2001) |
| FO Colomiella recta | Well 1-SPS-6, Santos Bs. | 5595 | Dias-Brito (1999) |
| FO Colomiella mexicana | Well 1-SPS-6, Santos Bs. | > 5595 | Dias-Brito (1999) |
| Black shale | Well X, Santos Bs. | 2600-2756 | Tedeschi et al. (2017) |
| Evaporite layer | Well 1-RJS-117A, Campos Bs | Relative position | Rangel et al. (1994) |
| Volcanic layer | Well 1-SCS-1, Santos Bs. | Not informed | Mizusaki (1993) fide Dias et al. (1994) |
Comments: There is no information about δ13Ccarb strong excursions nor high TOC values in the section.
|
|
a) Prediscosphaera columnata (DSDP Site 364 - depth 1,032.37
mbsf); b) Hayesites albiensis (DSDP Site 364 - depth 790.41 mbsf);
c) Hayesites albiensis (DSDP Site 364 - depth 1,031.21 mbsf); d)
Axopodorhabdus biramiculatus (DSDP Site 364 - depth 951.47 mbsf); e)
Tranolithus orionatus (DSDP Site 364 - depth 1008.41 mbsf); f) Eiffellithus turriseiffelii (DSDP Site 364
- depth 772.34 mbsf); g) Eiffellithus monechiae (DSDP Site 364 - depth 875.96
mbsf). |
| Core | Section | Interval (mbsf) |
Depth (mbsf) |
TOC (%) |
Reference | δ13Ccarb | δ18Ocarb |
| 26 | 5 | 73-77 | 717.23 | 0.13 | Bruno et al. (2020) | 1.14 | -4.17 |
| 26 | 6 | 115-117 | 719.15 | 0.11 | Bruno et al. (2020) | 1.06 | -3.29 |
| 27 | 2 | 125-128 | 722.75 | 0.13 | Bruno et al. (2020) | ||
| 27 | 3 | 67-71 | 723.67 | 0.08 | Bruno et al. (2020) | 1.53 | -3.28 |
| 27 | 4 | 74-77 | 725.24 | 0.09 | Bruno et al. (2020) | 1.71 | -2.89 |
| 27 | 5 | 66-70 | 726.66 | 0.12 | Bruno et al. (2020) | ||
| 28 | 1 | 69-73 | 749.19 | 0.11 | Bruno et al. (2020) | ||
| 28 | 2 | 70-74 | 750.7 | 0.12 | Bruno et al. (2020) | ||
| 28 | 2 | 130 | 751.3 | 0.1 | Bolli et al. (1978a) | ||
| 28 | 2 | 150 | 751.5 | 0.1 | Bolli et al. (1978a) | ||
| 28 | 3 | 87-91 | 752.37 | 0.05 | Bruno et al. (2020) | 2.57 | -2.90 |
| 28 | 4 | 18 | 753.18 | 0 | Bolli et al. 1978a | ||
| 28 | 4 | 64-68 | 753.64 | 0.10 | Bruno et al. (2020) | 2.17 | -3.70 |
| 28 | cc | Bottom of the box | 755 | 0.06 | Foresman (1978) | ||
| 29 | 1 | 69-71 | 768.19 | 0.09 | Bruno et al. (2020) | 1.84 | -4.49 |
| 29 | 2 | 33-37 | 769.33 | 0.13 | Bruno et al. (2020) | 2.14 | -3.37 |
| 29 | 3 | 67-71 | 771.17 | 0.12 | Bruno et al. (2020) | ||
| 29 | 4 | 15 | 772.15 | 0.1 | Bolli et al. (1978a) | ||
| 29 | 4 | 34-37 | 772.34 | 0.10 | Bruno et al. (2020) | ||
| 29 | cc | 774.5 | 0.09 | Foresman (1978) | |||
| 30 | 1 | 66-70 | 787.16 | 0.07 | Bruno et al. (2020) | ||
| 30 | 2 | 94-98 | 788.94 | 0.49 | Bruno et al. (2020) | ||
| 30 | 2 | 130 | 789.3 | 0.1 | Bolli et al. (1978a) | ||
| 30 | 2 | 150 | 789.5 | 0.1 | Bolli et al. (1978a) | ||
| 30 | 3 | 91-94 | 790.41 | 0.14 | Bruno et al. (2020) | ||
| 30 | cc | 792 | 0.1 | Foresman (1978) | |||
| 31 | 1 | 27-29 | 805.28 | 1.03 | Foresman (1978) | ||
| 31 | 1 | 79-82 | 806.29 | 0.19 | Bruno et al. (2020) | ||
| 31 | 2 | 40-42 | 807.4 | 0.19 | Bruno et al. (2020) | 1.96 | -3.25 |
| 31 | 3 | 41-44 | 808.91 | 0.11 | Bruno et al. (2020) | ||
| 31 | 4 | 52 | 810.52 | 0.1 | Bolli et al. (1978a) | ||
| 31 | 4 | 94-98 | 810.94 | 0.18 | Bruno et al. (2020) | 2.07 | -3.58 |
| 32 | 1 | 20-21 | 824.7 | Azevedo (2001) | 2.28 | -3.71 | |
| 32 | 1 | 61-61 | 825.11 | Azevedo (2001) | 2.38 | -3.68 | |
| 32 | 1 | 101-102 | 825.5 | Azevedo (2001) | 2.07 | -4.38 | |
| 32 | 1 | 123-127 | 825.73 | 0.08 | Bruno et al. (2020) | 2.11 | -4.08 |
| 32 | 1 | 143-143 | 825.93 | Azevedo (2001) | 2.6 | -2.17 | |
| 32 | 2 | 40-44 | 826.4 | 0.17 | Azevedo (2001) | 2.24 | -4.11 |
| 32 | 2 | 40-44 | 826.4 | Bruno et al. (2020) | 2.33 | -3.60 | |
| 32 | 2 | 79-80 | 826.79 | Azevedo (2001) | 2.77 | -2.27 | |
| 32 | 2 | 100-101 | 827 | Azevedo (2001) | 2.16 | -3.52 | |
| 32 | 2 | 120-121 | 827.2 | Azevedo (2001) | 2.68 | -2.11 | |
| 32 | 2 | 126-128 | 827.27 | 0.61 | Foresman (1978) | ||
| 32 | 3 | 53-56 | 828.05 | 0.09 | Foresman (1978) | ||
| 32 | 3 | 90-92 | 828.4 | 0.10 | Bruno et al. (2020) | ||
| 32 | 3 | 130 | 828.8 | 0.1 | Bolli et al. (1978a) | ||
| 32 | 3 | 150 | 829 | 0.2 | Bolli et al. (1978a) | ||
| 32 | 4 | 46-50 | 829.46 | 0.07 | Bruno et al. (2020) | 2.57 | -3.62 |
| 32 | 4 | 92 | 829.92 | 0.1 | Bolli et al. (1978a) | ||
| 33 | 1 | 107-111 | 844.57 | 0.06 | Bruno et al. (2020) | 2.21 | -4.13 |
| 33 | 2 | 92-95 | 845.92 | 0.34 | Bruno et al. (2020) | 2.42 | -3.69 |
| 33 | 3 | 75 | 847.25 | 0.7 | Bolli et al. (1978a) | ||
| 33 | 3 | 93-95 | 847.43 | 0.15 | Bruno et al. (2020) | 2.72 | -2.78 |
| 33 | 4 | 63-67 | 848.63 | 0.15 | Bruno et al. (2020) | ||
| 33 | 5 | 65-69 | 850.15 | 0.07 | Bruno et al. (2020) | 2.25 | -3.92 |
| 33 | 5 | 100-101 | 850.5 | Azevedo (2001) | 2.68 | -3.18 | |
| 33 | 5 | 121-121 | 850.71 | Azevedo (2001) | 2.4 | -3.51 | |
| 33 | 5 | 140-141 | 850.9 | Azevedo (2001) | 2.17 | -4.71 | |
| 33 | cc | 0-1 | 851 | Azevedo (2001) | 2.19 | -4.62 | |
| 34 | 1 | 100-101 | 873 | Azevedo (2001) | 1.92 | -4.3 | |
| 34 | 1 | 121-121 | 873.21 | Azevedo (2001) | 2.68 | -2.55 | |
| 34 | 1 | 133-136 | 873.33 | 0.22 | Bruno et al. (2020) | ||
| 34 | 2 | 19-20 | 873.69 | Azevedo (2001) | 2.02 | -4.11 | |
| 34 | 2 | 60-61 | 874.1 | Azevedo (2001) | 2.13 | -3.6 | |
| 34 | 2 | 93-96 | 874.43 | 0.90 | Bruno et al. (2020) | ||
| 34 | 2 | 100-101 | 874.5 | Azevedo (2001) | 2.23 | -3.52 | |
| 34 | 2 | 140-141 | 874.9 | Azevedo (2001) | 2.22 | -3.52 | |
| 34 | 3 | 96-99 | 875.96 | Bruno et al. (2020) | 2.50 | -3.84 | |
| 34 | 3 | 130 | 876.3 | 0.6 | Bolli et al. (1978a) | ||
| 34 | 4 | 53 | 877.03 | 0.3 | Bolli et al. (1978a) | ||
| 34 | 4 | 92-96 | 877.42 | 0.21 | Bruno et al. (2020) | ||
| 35 | 1 | 114-117 | 892.14 | 0.15 | Bruno et al. (2020) | ||
| 35 | 2 | 46-49 | 892.96 | 0.19 | Bruno et al. (2020) | ||
| 35 | 3 | 99-101 | 894.99 | 2.89 | Bruno et al. (2020) | ||
| 36 | 1 | 125-128 | 911.25 | 0.46 | Bruno et al. (2020) | 2.94 | -2.69 |
| 36 | 3 | 75-77 | 913.75 | 0.13 | Bruno et al. (2020) | ||
| 37 | 1 | 66-68 | 929.66 | 0.10 | Bruno et al. (2020) | ||
| 37 | 1 | 130 | 930.3 | 0.1 | Bolli et al. (1978a) | ||
| 37 | 2 | 120-122 | 931.7 | 0.12 | Bruno et al. (2020) | 1.99 | -3.76 |
| 37 | 3 | 45-47 | 932.45 | 0.46 | Bruno et al. (2020) | 2.05 | -3.36 |
| 37 | 4 | 94-97 | 934.44 | 0.15 | Bruno et al. (2020) | 1.94 | -3.77 |
| 37 | 5 | 98-101 | 935.98 | 0.09 | Bruno et al. (2020) | ||
| 38 | 1 | 20-21 | 948.2 | Azevedo (2001) | 1.55 | -5.05 | |
| 38 | 1 | 90-93 | 948.9 | 0.19 | Bruno et al. (2020) | ||
| 38 | 1 | 99-100 | 948.99 | Azevedo (2001) | 1.73 | -4.8 | |
| 38 | 2 | 39-40 | 949.89 | Azevedo (2001) | 1.54 | -4.14 | |
| 38 | 2 | 80-81 | 950.3 | Azevedo (2001) | 1.32 | -2.76 | |
| 38 | 2 | 100-101 | 950.5 | Azevedo (2001) | 1.23 | -4.44 | |
| 38 | 2 | 103-105 | 950.53 | 0.20 | Bruno et al. (2020) | 1.37 | -2.68 |
| 38 | 2 | 120-12 | 950.7 | Azevedo (2001) | 1.23 | -4.41 | |
| 38 | 2 | 140-141 | 950.9 | Azevedo (2001) | 1.62 | -4.8 | |
| 38 | 3 | 20-21 | 951.2 | Azevedo (2001) | 2 | -4.53 | |
| 38 | 3 | 40-41 | 951.4 | Azevedo (2001) | 1.56 | -4.71 | |
| 38 | 3 | 47-49 | 951.47 | 1.13 | Bruno et al. (2020) | ||
| 38 | 3 | 60-61 | 951.6 | Azevedo (2001) | 1.6 | -4.16 | |
| 38 | 3 | 80-81 | 951.8 | Azevedo (2001) | 1.17 | -3.41 | |
| 38 | 3 | 100-101 | 952 | Azevedo (2001) | 1.59 | -4.64 | |
| 38 | 3 | 120-12 | 952.2 | Azevedo (2001) | 1.54 | -4.14 | |
| 38 | 3 | 139-140 | 952.39 | Azevedo (2001) | 1.65 | -4.55 | |
| 38 | 4 | 27-30 | 952.77 | Bruno et al. (2020) | 1.57 | -3.86 | |
| 38 | 4 | 143 | 953.93 | 1.2 | Bolli et al. (1978a) | ||
| 38 | 5 | 53-55 | 954.53 | 0.28 | Bruno et al. (2020) | 1.56 | -3.98 |
| 38 | 6 | 25-28 | 955.75 | 1.27 | Bruno et al. (2020) | ||
| 39 | 1 | 145-148 | 968.45 | 3.02 | Bruno et al. (2020) | ||
| 39 | 2 | 114-119 | 969.64 | 4.60 | Bruno et al. (2020) | ||
| 39 | 3 | 118-122 | 971.18 | 1.58 | Bruno et al. (2020) | ||
| 39 | 4 | 66-69 | 972.16 | 1.81 | Bruno et al. (2020) | ||
| 39 | 4 | 91 | 972.41 | 4.7 | Bolli et al. (1978a) | ||
| 39 | 5 | 89-92 | 973.89 | 1.44 | Bruno et al. (2020) | ||
| 39 | 5 | 127 | 974.27 | 1.5 | Bolli et al. (1978a) | ||
| 39 | 5 | 150 | 974.5 | 2.4 | Bolli et al. (1978a) | ||
| 39 | 6 | 85 | 975.35 | 1.4 | Bolli et al. (1978a) | ||
| 39 | 6 | 99-101 | 975.49 | 1.22 | Bruno et al. (2020) | -0.42 | -4.58 |
| 40 | 1 | 142-147 | 987.42 | 0.63 | Bruno et al. (2020) | 0.15 | -3.05 |
| 40 | 2 | 92-96 | 988.42 | 3.85 | Bruno et al. (2020) | 1.36 | -4.11 |
| 40 | 3 | 71-75 | 989.71 | 3.71 | Bruno et al. (2020) | ||
| 40 | 4 | 44 | 990.94 | 4.9 | Bolli et al. (1978a) | ||
| 40 | 4 | 66-68 | 991.16 | 1.02 | Bruno et al. (2020) | ||
| 40 | 5 | 63-66 | 992.63 | 2.39 | Bruno et al. (2020) | 0.12 | -5.07 |
| 41 | 1 | 141-145 | 1006.41 | 3.03 | Bruno et al. (2020) | ||
| 41 | 2 | 19-20 | 1006.7 | Azevedo (2001) | 0.72 | -4.06 | |
| 41 | 2 | 46-50 | 1006.96 | 2.26 | Bruno et al. (2020) | ||
| 41 | 2 | 60-61 | 1007.1 | Azevedo (2001) | 0.52 | -5.15 | |
| 41 | 2 | 79-80 | 1007.3 | Azevedo (2001) | 0.74 | -5.46 | |
| 41 | 2 | 100-01 | 1007.5 | Azevedo (2001) | -0.02 | -4.98 | |
| 41 | 2 | 141-142 | 1007.91 | Azevedo (2001) | 0.77 | -4.59 | |
| 41 | 3 | 40-41 | 1008.4 | Azevedo (2001) | 0.03 | -5.33 | |
| 41 | 3 | 41-44 | 1008.41 | 1.50 | Bruno et al. (2020) | -0.21 | -5.47 |
| 41 | 3 | 80-81 | 1008.8 | Azevedo (2001) | 0.22 | -3.89 | |
| 41 | 3 | 119-120 | 1009.2 | Azevedo (2001) | -0.52 | -5.14 | |
| 41 | 3 | 130 | 1009.3 | 0.7 | Bolli et al. (1978a) | ||
| 41 | 3 | 150 | 1009.5 | 6 | Bolli et al. (1978a) | ||
| 41 | 4 | 19-20 | 1009.7 | Azevedo (2001) | 1.73 | -3.42 | |
| 41 | 4 | 29-32 | 1009.79 | 3.47 | Bruno et al. (2020) | ||
| 41 | 4 | 32 | 1009.82 | 3.8 | Bolli et al. (1978a) | ||
| 42 | 1 | 137-139 | 1025.37 | 0.28 | Bruno et al. (2020) | -2.59 | -4.85 |
| 42 | 2 | 20-21 | 1025.7 | Azevedo (2001) | 1.15 | -5.75 | |
| 42 | 2 | 60-61 | 1026.1 | Azevedo (2001) | -1.99 | -5.28 | |
| 42 | 2 | 81-84 | 1026.31 | 8.91 | Bruno et al. (2020) | ||
| 42 | 2 | 100-101 | 1026.5 | Azevedo (2001) | 0.78 | -4.06 | |
| 42 | 2 | 140-141 | 1026.9 | Azevedo (2001) | -2.37 | -5.17 | |
| 42 | 3 | 24-28 | 1027.24 | 4.58 | Bruno et al. (2020) | ||
| 42 | 4 | 75 | 1029.25 | 8.4 | Bolli et al. (1978a) | ||
| 42 | 4 | 100-125 | 1029.5 | 0.19 | Bruno et al. (2020) | ||
| 42 | 5 | 121-125 | 1031.21 | 0.54 | Bruno et al. (2020) | ||
| 42 | 6 | 32 | 1031.82 | 22.5 | Bolli et al. (1978a) | ||
| 42 | 6 | 87-90 | 1032.37 | 0.40 | Bruno et al. (2020) | ||
| 43 | 1 | 130 | 1034.6 | 7.4 | Bolli et al. (1978a) | ||
| 43 | 1 | 150 | 1034.8 | 0.3 | Bolli et al. (1978a) | ||
| 44 | 4 | 10 | 1047.6 | 1.2 | Bolli et al. (1978a) |
[**] The term Konservat-Lagerst tten was coined by A. Seilacher, in 1970, to deposits known for the exceptional preservation of fossilized organisms or traces (https://en.wikipedia.org/wiki/Lagerst tte, accessed: 2022/11/15).