◄ Carnets Geol. 22 (21) ►
![]()
Outline:
[1. Introduction]
[2. Methods]
[3. Systematic paleontology]
[4. Associated LBF, ages, and paleoenvironments] and ... [Bibliographic references]
Department of Geography & Geology, The University of the West Indies, Mona, Kingston 7 (Jamaica, W.I.)
U.S. Geological Survey, 3321 College Avenue, Fort Lauderdale, FL 33314 (USA)
Published online in final form (pdf) on December 25, 2022
DOI 10.2110/carnets.2022.2221
![]()
[Editor:
Simon F. Mitchell; technical editor: Bruno R.C. Granier]
![]()
We describe two larger benthic foraminiferal taxa collected from wells drilled in the subsurface Eocene rocks of southeastern Florida that are new to peninsular Florida and the Caribbean region. Saudia floridana n.sp. is characteristic of a foraminiferal assemblage, along with Helicostegina gyralis, wide forms of the Cushmania americana group, and Gunteria floridana, in an upper part of the Oldsmar Formation. Globogypsinoides browardensis n.gen. n.sp. occurs in a second foraminiferal assemblage, along with Borelis cf. floridanus, Coskinolina cf. yucatanensis, and as-yet undescribed large rotaliids, in a middle part of the Oldsmar Formation. The foraminiferal assemblage of the middle Oldsmar unit is ascribed an Ypresian age and the assemblage of the upper Oldsmar unit a Lutetian age. These two assemblages indicate inner shelf water depths of 40 m or less on the Florida Platform during the Early to Middle Eocene deposition of the middle to upper part of the Oldsmar Formation.
larger benthic foraminifera;
Eocene;
Florida;
Oldsmar Formation
Robinson E. & Cunningham K.J. (2022).- New larger benthic foraminifera from the subsurface Lower to Middle Eocene Oldsmar Formation of southeastern Florida (USA).- Carnets Geol., Madrid, vol. 22, no. 21, p. 857-865.
Nouveaux grands foraminifères benthiques de la Formation d'Oldsmar (Éocène inférieur à moyen) du sud-est de la Floride (États-Unis d'Amérique).- Nous décrivons ici deux taxons de grands foraminifères benthiques provenant de puits forés dans les couches éocènes du sud-est de la Floride. Il s'agit d'espèces nouvelles pour la Floride péninsulaire et la région caribéenne. Saudia floridana n.sp. est caractéristique d'une association de foraminifères de la partie supérieure de la Formation d'Oldsmar, comprenant également Helicostegina gyralis, de grandes formes du groupe Cushmania americana, et Gunteria floridana. Globogypsinoides browardensis n.gen. n.sp. apparaît dans une seconde association de foraminifères de la partie médiane de la Formation d'Oldsmar, accompagnée de Borelis cf. floridanus, Coskinolina cf. yucatanensis et de grands rotaliidés non encore décrits à ce jour. Un âge yprésien est attribué à l'association de foraminifères de l'unité médiane d'Oldsmar et un âge lutétien à celle de l'unité supérieure d'Oldsmar. Ces deux associations indiquent des bathymétries de -40 m ou moins pour la partie interne de la plate-forme de Floride pendant le dépôt des couches de l'Éocène inférieur à moyen des parties moyenne à supérieure de la Formation Oldsmar.
grands foraminifères benthiques ;
Éocène ;
Floride ;
Formation Oldsmar
The Paleogene carbonate rocks of peninsular Florida contain varied assemblages of larger benthic foraminifera (LBF) which are detailed in several local and regional studies, particularly since the mid-20th Century (e.g., Cole,
1938, 1941, 1942;
E.R. Applin & Jordan,
1945; Levin, 1957; Bowen-Powell,
2010; Cotton et al.,
2018). These rocks form the northernmost component of a much larger Caribbean and Central American faunal province extending into the offshore areas of northern South America (Barker & Grimsdale,
1936; Vaughan, 1945; Frost & Langenheim,
1974; Wong, 1976; Butterlin,
1981, 1990; Robinson & Wright,
1993; Caudri,1996; Serra-Kiel et al.,
2007; BouDagher-Fadel & Price,
2010; Vicedo et al.,
2014; Torres-Silva et al.,
2019; Mitchell et al.,
2022). The Eocene LBF assemblages, although diverse, show marked differences from the LBF zonation established by the Shallow Benthic Zone (SBZ) scheme of Serra-Kiel et al.
(1998 and references therein) for the Tethys region. The large nummulitids, which historically have formed the backbone of Tethys biostratigraphy, are mostly absent from the Western Hemisphere, apart from a few isolated monospecific occurrences in the northern Caribbean (Rutten,
1935; Robinson & Wright,
1993; Mitchell et al.,
2022). The principal taxa of much of the Floridan, Caribbean and northern South American Eocene assemblages are dominated by the Lepidocyclinidae that are amenable to forming the basis of a LBF zonation parallel to SBZ zonation of the Tethys region (Mitchell et al.,
2022). As part of an ongoing subsurface stratigraphic study in southeast Florida (Cunningham,
2015; Cunningham & Robinson,
2017; Cunningham et al.,
2018a, 2018b), we describe two Eocene LBF that appear to be related to Tethys groups but have not been previously reported from the Western Hemisphere. They occur in several wells drilled in the southeastern part of peninsular Florida
(Fig. 1 ![]() ), in the Lower to lower Middle Eocene Oldsmar Formation (P.L.
Applin & E.R. Applin, 1944; Levin, 1957; Cunningham et al.,
2018b).
), in the Lower to lower Middle Eocene Oldsmar Formation (P.L.
Applin & E.R. Applin, 1944; Levin, 1957; Cunningham et al.,
2018b).
|
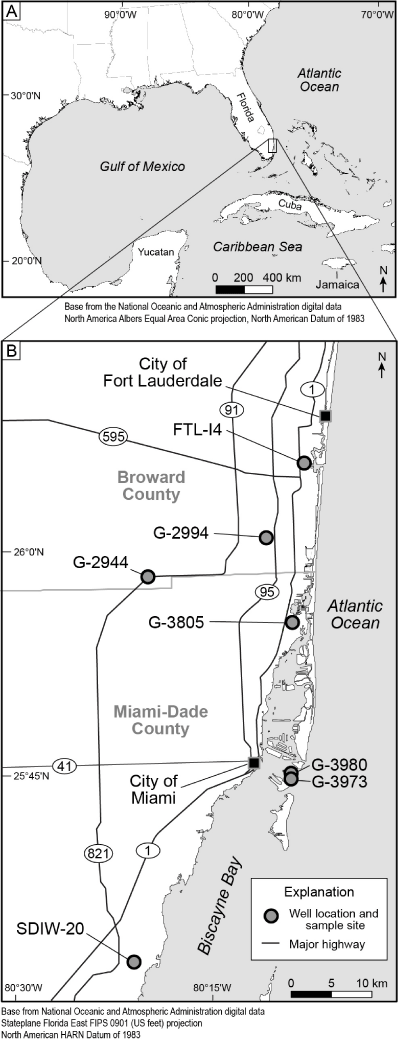
Figure 1:
Geographical location of study area. 1, Regional map showing location of study area. 2, General map showing location of 7 wells sampled for foraminiferal specimens. |
The material for the study consisted of thin sections of samples taken, either as cuttings or cores from several wells in Southeast Florida, as listed for each taxon in the Systematic section. The thin sections were prepared primarily for sedimentological studies (Cunningham, 2015; Cunningham et al., 2018a, 2018b) and the LBF specimens examined for the present paper occurred in these as thin sections that in many cases were not acquired specifically for LBF study. Standard transmitted-light petrography was used to examine thin sections containing specimens. Measurements of the dimensions of specimens were completed and images acquired using a trinocular Amscope T120C microscope with an eyepiece micrometer and MU1000 digital camera.
Suprageneric classification follows Kaminski, 2014.
Order Loftusiida Kaminski & Mikhalevich, 2004
Superfamily Loftusoidea Brady, 1884
Family Spirocyclinidae Munier-Chalmas, 1887
Genus Saudia Henson, 1948
Type species: Saudia discoidea Henson, 1948
Saudia discoidea Henson, 1948
(Fig. 2 ![]() )
)
1948 Henson, p. 97-98; Pl. XII, figs. 1-4 (paratypes), 5.
Remarks: Saudia discoidea was described and named by Henson (1948) for a species of spirocyclinid collected on the Saudi Arabian border, from localities reported (in his text) to be "in Lower Eocene limestones with Dictyoconus sp." (Henson, 1948, p. 98) It was designated the type species for the monotypic genus Saudia, described as having cyclical chambers with a complex exoskeleton consisting of several orders of beams and rafters forming a cellular structure, diagrammatically illustrated by Henson (1948) in his Figure 14. The published illustrations consisted of two subequatorial sections of flabelliform (reniform) microspheric (B-form) paratypes; (Henson, 1948, Pl. XII, figs. 1-2) and three axial sections of fragments of the annular chambers, two of which were designated as paratypes, captioned as being "Lutetian" (Henson, 1948, Pl. XII, figs. 3-5). The holotype was not illustrated. We cannot confirm which, if either, of the two ages quoted by Henson above for his types is correct.
As Drobne and Hottinger (1971) aptly remarked in their redescription of Saudia (translated from the German): "The photographic documentation for the description is so modest that you have to rely on the drawing interpretation of the structure as the basis for the generic definition".
In the present paper, we illustrate the holotype of Saudia discoidea for the first time
(Fig. 2 ![]() ). As is evident from the illustrations, the holotype has been extensively affected by diagenesis but is an example of a reniform, possibly discoidal, megalospheric (A-form) individual. In referring our species to the genus Saudia, we follow Drobne and Hottinger
(1971) in placing emphasis on Henson's written generic description
(Henson, 1948, p. 97) and his
Figure 14 (ibid., p. 98).
). As is evident from the illustrations, the holotype has been extensively affected by diagenesis but is an example of a reniform, possibly discoidal, megalospheric (A-form) individual. In referring our species to the genus Saudia, we follow Drobne and Hottinger
(1971) in placing emphasis on Henson's written generic description
(Henson, 1948, p. 97) and his
Figure 14 (ibid., p. 98).
|
Figure 2:
Saudia discoidea Henson, 1948, Holotype. NHMUK PM P36023. 1, Subequatorial (top) view; 2, side view. Accompanying scale bar is 0.5 mm for both 1 and 2. The holotype was not illustrated along with
Henson's (1948) original published description. Photographs, including scale bar, by Dr. Stephen Stukins, senior curator, Natural History Museum, London and courtesy of the Trustees of the Natural History Museum, London. |
Saudia floridana n.sp.
2018b Thomasella cf. T. labyrinthica (Grimsdale); Cunningham et al., text in Figs. 3, 10; Thomasella n.sp. Table 3.
Material: Numerous incomplete specimens in thin sections cut from cores and cuttings from wells FTL-I4, G-2994, G-2944, G-3805, G-3973, and SDIW-20
(Fig. 1 ![]() ).
).
Description: The large reniform (discoidal?) agglutinated test is bilaterally symmetrical with specimens varying from slightly to distinctly biconcave with a central swelling. The test margin is rounded. The species is strongly dimorphic. Two centred axial specimens of the megalospheric generation have diameters of 9.6 and 8.8 mm
(Fig. 3.4-3.5 ![]() ), while a single equatorial section has a diameter of 7 mm
(Fig. 3.1
), while a single equatorial section has a diameter of 7 mm
(Fig. 3.1 ![]() ). On megalospheric specimens the polar region is inflated with examples reaching a thickness of about 1.3 mm over the embryonic region at the pole, 0.8 to 0.9 mm over the first annular chambers and 0.8 to 1.2 mm at the test margin. The complete subaxial section of a microspheric (B form) individual has a diameter of 27 mm
(Fig. 4.4
). On megalospheric specimens the polar region is inflated with examples reaching a thickness of about 1.3 mm over the embryonic region at the pole, 0.8 to 0.9 mm over the first annular chambers and 0.8 to 1.2 mm at the test margin. The complete subaxial section of a microspheric (B form) individual has a diameter of 27 mm
(Fig. 4.4 ![]() ), a thickness over the polar region of 0.6 mm, and of 2.0 to 2.2 mm at the periphery. Other microspheric specimens exceed 20 mm in equatorial diameter, based on radius and partial diameter measurements.
), a thickness over the polar region of 0.6 mm, and of 2.0 to 2.2 mm at the periphery. Other microspheric specimens exceed 20 mm in equatorial diameter, based on radius and partial diameter measurements.
The nuclear zone in the A form generation is poorly preserved in the specimens available
(Fig. 3.1-3.5 ![]() ). It is irregularly subspherical with a polar diameter of 0.9 to 1.1 mm and an equatorial diameter of 1.0 to 1.5 mm. It consists of an inner, oblate spheroidal chamber
(sphaeroconch of Hottinger, 1967, and Drobne and Hottinger,
1971) with an approximate ratio equatorial diameter/height of 1.1 to 1.2, pierced by multiple foramina
(Fig. 3.2-3.3
). It is irregularly subspherical with a polar diameter of 0.9 to 1.1 mm and an equatorial diameter of 1.0 to 1.5 mm. It consists of an inner, oblate spheroidal chamber
(sphaeroconch of Hottinger, 1967, and Drobne and Hottinger,
1971) with an approximate ratio equatorial diameter/height of 1.1 to 1.2, pierced by multiple foramina
(Fig. 3.2-3.3 ![]() ) averaging 0.04 to 0.05 mm in diameter and uniformly distributed over its surface, embraced by an outer chamber or chambers divided by radial pillars or partitions that appear to support the sphaeroconch
(Fig. 3.2
) averaging 0.04 to 0.05 mm in diameter and uniformly distributed over its surface, embraced by an outer chamber or chambers divided by radial pillars or partitions that appear to support the sphaeroconch
(Fig. 3.2 ![]() ). The embryon is succeeded by 1-2 reniform or spiral chambers, then by annular chambers. The character of the embryonic region of the B generation was not determined.
). The embryon is succeeded by 1-2 reniform or spiral chambers, then by annular chambers. The character of the embryonic region of the B generation was not determined.
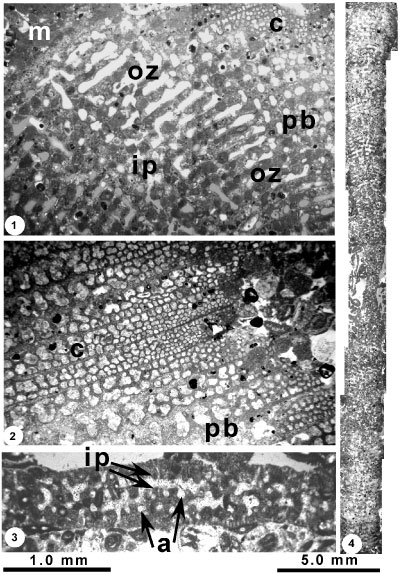
The nuclear zone is surrounded by up to 40 annular chambers in the megalospheric test with an average length of 0.13-0.14 mm. About 66 chambers were counted for the microspheric example, with an average length of 0.19 mm. Each chamber is divided into a central or endoskeletal zone containing interseptal pillars, separated above and below by narrow spaces (the "open zones" of Grimsdale,
1952, p. 231) from a marginal exoskeleton of beams and rafters. The radially directed interseptal pillars of the endoskeleton are close to continuous from chamber to chamber
(Figs. 3.1 ![]() ,
4.1
,
4.1 ![]() ), increasing distally from one to two, rarely three tiers, resembling the axial fragment illustrated by Henson
(1948, Pl. XII, fig. 4), alternating in position with one or more rows of stolons
(Fig. 4.3
), increasing distally from one to two, rarely three tiers, resembling the axial fragment illustrated by Henson
(1948, Pl. XII, fig. 4), alternating in position with one or more rows of stolons
(Fig. 4.3 ![]() ). The open zones, about 0.05 mm high, are free of skeletal elements except for the chamber septa, which are pierced by multiple, radially directed foramina about 0.18 mm apart and with diameters of about 0.05 to 0.07 mm
(Fig. 4.1
). The open zones, about 0.05 mm high, are free of skeletal elements except for the chamber septa, which are pierced by multiple, radially directed foramina about 0.18 mm apart and with diameters of about 0.05 to 0.07 mm
(Fig. 4.1 ![]() ). Each marginal exoskeletal zone consists of a subepidermal network of radial beams and concentric rafters. These increase in number towards the marginal wall. On tangential cuts, the test surface resembles a honeycomb-like meshwork of cellules, like that described by Grimsdale
(1952, p. 231, Pl. 21, fig. 3) for his Saudia labyrinthica, and structurally like those seen on the conical genus Cushmania (e.g., Robinson,
1974,
Fig. 3). There are at least two orders of beams
(Fig. 4.1
). Each marginal exoskeletal zone consists of a subepidermal network of radial beams and concentric rafters. These increase in number towards the marginal wall. On tangential cuts, the test surface resembles a honeycomb-like meshwork of cellules, like that described by Grimsdale
(1952, p. 231, Pl. 21, fig. 3) for his Saudia labyrinthica, and structurally like those seen on the conical genus Cushmania (e.g., Robinson,
1974,
Fig. 3). There are at least two orders of beams
(Fig. 4.1 ![]() ). The primary beams are the most prominent element of the exoskeleton and alternate from chamber to chamber as in S. labyrinthica (Grimsdale,
1952) initially about 0.08 to 0.09 mm thick and spaced 0.17 to 0.18 mm apart
(Fig. 4.2
). The primary beams are the most prominent element of the exoskeleton and alternate from chamber to chamber as in S. labyrinthica (Grimsdale,
1952) initially about 0.08 to 0.09 mm thick and spaced 0.17 to 0.18 mm apart
(Fig. 4.2 ![]() ). Near the thinner margin, second- and third-order beams are inserted, together with much thinner rafters, so that at the margin the number of cellules may reach as high as 6 to 7 across the chamber length
(Fig. 4.2
). Near the thinner margin, second- and third-order beams are inserted, together with much thinner rafters, so that at the margin the number of cellules may reach as high as 6 to 7 across the chamber length
(Fig. 4.2 ![]() ).
).
|
Figure 3: Saudia floridana
n.sp. 1, Equatorial section, designated as holotype, showing the central nuclear region surrounded by annular chambers and radially directed interseptal pillars, exoskeletal zone of beams and rafters visible around the periphery of the test, and rectangular area at upper left is enlarged on
Fig. 4 |
|
Figure
4: Saudia floridana n.sp. 1, part of the holotype at Fig. 3.1 |
Remarks: Saudia floridana
n.sp. is distinguished from other complex discoidal LBF in the Florida Paleogene (E.R.
Applin & Jordan, 1945, p. 141) by three features in the
ultrastructure: (1) the agglutinated, particulate nature of the wall (Figs.
3 ![]() - 4
- 4 ![]() ); (2) the complex irregular cellular structure of the wall, seen in Paleogene and Mesozoic spirocyclinids (Hottinger,
1967) and many of the conical dictyoconids, such as Cushmania; and (3) the more complex prolocular region
(sphaeroconch). These features are not present in the porcelaneous peneroplid group (ER observations; Loeblich & Tappan, 1987). It is likely that fragmental S. floridana has been confused with fragments of wide forms of the Cushmania americana group in the past as these are common at the same stratigraphic level as S. floridana (E.R.
Applin and Jordan, 1945, p. 136). Compare the sketch of Figure 2 in
E.R. Applin and Jordan (1945) with the exoskeletal structure visible in images 1 and 2 of our
Figure 4
); (2) the complex irregular cellular structure of the wall, seen in Paleogene and Mesozoic spirocyclinids (Hottinger,
1967) and many of the conical dictyoconids, such as Cushmania; and (3) the more complex prolocular region
(sphaeroconch). These features are not present in the porcelaneous peneroplid group (ER observations; Loeblich & Tappan, 1987). It is likely that fragmental S. floridana has been confused with fragments of wide forms of the Cushmania americana group in the past as these are common at the same stratigraphic level as S. floridana (E.R.
Applin and Jordan, 1945, p. 136). Compare the sketch of Figure 2 in
E.R. Applin and Jordan (1945) with the exoskeletal structure visible in images 1 and 2 of our
Figure 4 ![]() ). Grimsdale (1952, p. 231) emphasized, in his well-illustrated description, that his Saudia labyrinthica was differentiated from Saudia discoidea solely by the extreme complexity of the endoskeleton. Sirel
(1998) also emphasized the greater complexity of the exoskeleton of S. labyrinthica to support the type for his new genus Thomasella. Because Henson
(1948) had described the exoskeleton of S. discoidea as being complex, with multiple beams and rafters, the types of Saudia Henson and Thomasella Sirel
(1998) were examined by the first author
(E.R.) at the Natural History Museum, United Kingdom. The details of the published type specimens of S. discoidea are difficult to evaluate due to diagenetic influences. None appear to display more than two orders of beams in the exoskeleton, but possible further multiplication is obscured by rock matrix.
). Grimsdale (1952, p. 231) emphasized, in his well-illustrated description, that his Saudia labyrinthica was differentiated from Saudia discoidea solely by the extreme complexity of the endoskeleton. Sirel
(1998) also emphasized the greater complexity of the exoskeleton of S. labyrinthica to support the type for his new genus Thomasella. Because Henson
(1948) had described the exoskeleton of S. discoidea as being complex, with multiple beams and rafters, the types of Saudia Henson and Thomasella Sirel
(1998) were examined by the first author
(E.R.) at the Natural History Museum, United Kingdom. The details of the published type specimens of S. discoidea are difficult to evaluate due to diagenetic influences. None appear to display more than two orders of beams in the exoskeleton, but possible further multiplication is obscured by rock matrix.
Differences from other species: Saudia floridana
n.sp. has similar exoskeletal features as seen in Saudia labyrinthica but differs in the absence of the labyrinthic structural complexity and the less prominent open zone. It resembles Saudia discoidea in the relative simplicity of the endoskeleton with scattered interseptal pillars, the relatively narrow open zone and a test with less pronounced biconcavity. The size of the sphaeroconch is about the same. The B-form of S. floridana
(Fig. 4.4 ![]() ) is much larger than those illustrated for S. discoidea.
) is much larger than those illustrated for S. discoidea.
The specimen in Figure 3.1 ![]() , designated as the holotype, is from a core from well FTL-I4 at 2,297.25 ft (700.2 m) depth. Paratypes
(Fig. 3.4-3.5
, designated as the holotype, is from a core from well FTL-I4 at 2,297.25 ft (700.2 m) depth. Paratypes
(Fig. 3.4-3.5 ![]() ) are from the FTL-I4 core at 2,296.70 ft (700.0 m) and well G-3973 core at 2,564.80 ft (781.8 m), respectively. The microspheric specimen
(Fig. 4.4
) are from the FTL-I4 core at 2,296.70 ft (700.0 m) and well G-3973 core at 2,564.80 ft (781.8 m), respectively. The microspheric specimen
(Fig. 4.4 ![]() ) is from a core from well FTL-I4 at 2,358.40 ft (718.8 m). See
Figure 1
) is from a core from well FTL-I4 at 2,358.40 ft (718.8 m). See
Figure 1 ![]() for well locations.
for well locations.
Depository: The type specimens, along with other reference specimens will be deposited in the collections of the U.S. National Museum, Washington - DC.
Family Acervulinidae Schultze, 1854
Genus Globogypsinoides n.gen.
Type species: Globogypsinoides browardensis n.gen. n.sp.
Diagnosis: A small globular acervulinid with a central proloculus followed by a short nepionic coil of arcuate chambers, succeeded in the adult stage by one or more series of streptospirally arranged orbitoidiform chambers.
Although the overall architecture bears a close resemblance to the genus Orduella Sirel, 1999, we do not include Globogypsinoides n.gen. in that genus, nor in the family Orduellidae Sirel, because the nepionic chambers are distinctly arcuate, not subrectangular, at least in the megalospheric generation. For this reason and until more material becomes available for examination, we retain Globogypsinoides in the Acervulinidae. The genus differs from the globular genus Sphaerogypsina Galloway, 1933, in possessing a multiple spiral development of orbitoid-like chambers whereas the chambers in Sphaerogypsina develop in a radial pattern. The new genus differs from Protogypsina Matsumaru & Sarma, 2010, in the test shape, which is consistently globular, and in the size and arrangement of the nepionic coiling.
Globogypsinoides browardensis n.sp.
2018b Orduella cf. O. sphaerica Sirel; Cunningham et al., Figs. 3, 10,Table 3
Material: About 60 specimens in thin sections from wells G2994, G3805 and G3973
(Fig. 1 ![]() ) are available.
) are available.
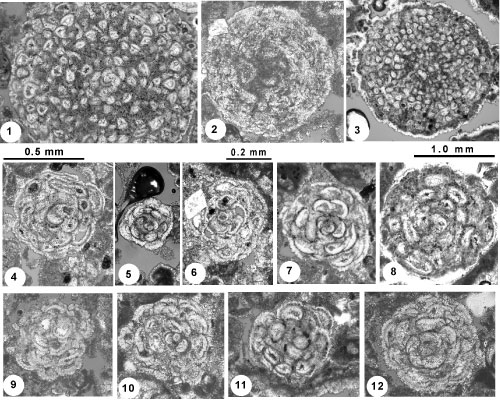
Description: The sub-globular tests are small, with mostly off-centred sections of megalospheric specimens ranging in diameter from about 0.2-0.5 mm. The largest centred megalospheric specimens are 0.51 and 0.52 mm in diameter. Centered megalospheric tests have proloculi ranging in diameter from about 0.05 up to 0.08 mm. The initial chambers are followed by two to three whorls of spirally arranged arcuate chambers then by arcuate (orbitoidiform?) chambers arranged in multiple streptospires to form the globular test. The species is dimorphic, and the microspheric form is rare. No centred microspheric tests were encountered.
Figure 5.2 ![]() shows a near-centered, probable microspheric form but the preservation is poor. The largest
uncentred, presumed microspheric specimen has a diameter of 1.7 mm (Fig.
5.3
shows a near-centered, probable microspheric form but the preservation is poor. The largest
uncentred, presumed microspheric specimen has a diameter of 1.7 mm (Fig.
5.3 ![]() ).
).
The individual in Figure 5.6 ![]() , designated as holotype, is from well G-2994 at 2,936.7 ft (895.1 m).
Individuals in Figure 5.3-5.4, 5.7, 5.12
, designated as holotype, is from well G-2994 at 2,936.7 ft (895.1 m).
Individuals in Figure 5.3-5.4, 5.7, 5.12 ![]() are designated as paratypes from well G-3973 at 2,622.82 ft (799.4 m), G-3973 at 2,621.25 ft (799.0 m), well G3805 at 2,734.3ft (833.4 m), and well G-2994 at 2,938.9 ft (895.8 m), respectively. See
Figure 1
are designated as paratypes from well G-3973 at 2,622.82 ft (799.4 m), G-3973 at 2,621.25 ft (799.0 m), well G3805 at 2,734.3ft (833.4 m), and well G-2994 at 2,938.9 ft (895.8 m), respectively. See
Figure 1 ![]() for well locations.
for well locations.
Depository: The type specimens, along with other reference specimens will be deposited in the collections of the U.S. National Museum, Washington - DC.
|
Figure
5: 1-12, Globogypsinoides browardensis n.gen. n.sp. Random sections through twelve
specimens. 1-3 are probable microspheric specimens and 4-12 are megalospheric specimens. Location and depths for 6, the holotype, and
3-4, 7 and 12, paratypes, are given in the text. 1-2, 5, 8 are from well G-3805 at a depth of 2,732.7 ft (832.9 m), well G-2994 at 2,930.3 ft (893.2 m), well G3805 at 2,737.8 ft (834.5 m) and from well G-3973 at a depth of 2,780.3ft (847.4 m) respectively. 9-12 are from well G2994 at depths of 2,930.3 ft (893.2 m), 2,938.9 ft (895.8 m), 2,934.2 ft (894.3 m) and 2,938.9 ft (895.8 m) respectively. The 0.5 mm scale is for 1; 1 mm scale for 3; and 0.2 mm scale for 2, 4-12. See
Figure 1 |
This is the first report of Saudia sp. from peninsular Florida and the Caribbean region. In southeastern Florida S. floridana is accompanied by Helicostegina gyralis Barker & Grimsdale,
1936, Gunteria floridana Cushman & Ponton,
1933, common wide forms of Cushmania americana (Cushman,
1919), Fallotella cookei (Moberg,
1928), and Fallotella floridana (Cole, 1941), which constitutes a foraminiferal assemblage in an upper part of the Oldsmar Formation in wells FTL-I4, G-3805, G3980, and G-3973
(Fig. 1 ![]() ). Apart from Helicostegina gyralis, a widespread although not exclusive marker for the Oldsmar Formation (Winston,
1993), the accompanying species have also been recorded from the Middle Eocene Avon Park Formation, including the Lake City limestone of authors
P.L. Applin and E.R. Applin (1944) and
E.R. Applin and Jordan (1945). We therefore assign an early Middle Eocene (Lutetian) age to the assemblage in the upper part of the Oldsmar Formation that includes S. floridana. This age assignment is similar to the age suggested by Henson
(1948, Pl. XII, 3-5) for the type species of the genus from the Middle Eastern part of the Tethys bioprovince.
). Apart from Helicostegina gyralis, a widespread although not exclusive marker for the Oldsmar Formation (Winston,
1993), the accompanying species have also been recorded from the Middle Eocene Avon Park Formation, including the Lake City limestone of authors
P.L. Applin and E.R. Applin (1944) and
E.R. Applin and Jordan (1945). We therefore assign an early Middle Eocene (Lutetian) age to the assemblage in the upper part of the Oldsmar Formation that includes S. floridana. This age assignment is similar to the age suggested by Henson
(1948, Pl. XII, 3-5) for the type species of the genus from the Middle Eastern part of the Tethys bioprovince.
Globogypsinoides browardensis n.gen. n.sp. is a member of a foraminiferal assemblage within a middle part of the Oldsmar Formation in wells G2994, G3805, and G3973
(Fig. 1 ![]() ), accompanied by Coskinolina cf. yucatanensis (Vicedo et al.,
2014), and by large species of undescribed
rotaliids, including the form called Rotalia trochidiformis of Levin (1957), and in wells G-3805 and G-3973
(Fig. 1
), accompanied by Coskinolina cf. yucatanensis (Vicedo et al.,
2014), and by large species of undescribed
rotaliids, including the form called Rotalia trochidiformis of Levin (1957), and in wells G-3805 and G-3973
(Fig. 1 ![]() ) also by Borelis cf. floridanus Cole, 1941. The latter is a species usually considered to be a Cedar Keys Formation marker of Paleocene age but also reported by
Levin (1957) in the Oldsmar Formation in his Zone IV. We assign an Early Eocene (Ypresian) age to the foraminiferal assemblage of the middle part of the Oldsmar Formation.
) also by Borelis cf. floridanus Cole, 1941. The latter is a species usually considered to be a Cedar Keys Formation marker of Paleocene age but also reported by
Levin (1957) in the Oldsmar Formation in his Zone IV. We assign an Early Eocene (Ypresian) age to the foraminiferal assemblage of the middle part of the Oldsmar Formation.
In all seven southeastern Florida wells (Fig. 1 ![]() ), conical agglutinated taxa dominate the two foraminiferal assemblages in the middle and upper parts of the Oldsmar Formation. In contrast, there is a lack of orbitoidal genera and nummulitids. These observations indicate that inner shelf water depths of 40 m or less (Hottinger,
1997) were the rule during the Early and Middle Eocene deposition of the middle to upper part of the Oldsmar Formation in our study area of the southeastern Florida peninsula.
), conical agglutinated taxa dominate the two foraminiferal assemblages in the middle and upper parts of the Oldsmar Formation. In contrast, there is a lack of orbitoidal genera and nummulitids. These observations indicate that inner shelf water depths of 40 m or less (Hottinger,
1997) were the rule during the Early and Middle Eocene deposition of the middle to upper part of the Oldsmar Formation in our study area of the southeastern Florida peninsula.
Saudia floridana n.sp. and Globogypsinoides browardensis n.gen. n.sp. are important members of foraminiferal assemblages in the Oldsmar Formation with occurrences in the upper part and middle part, respectively. The recongition of Saudia in Florida, could be useful in extending biostratigraphic correlations beyond southeastern Florida and other areas of the Caribbean where LBF assemblages characteristic of very shallow water paleoenvironments lack nummulitid and orbitoidal components.
The Miami-Dade Water and Sewer Department, U.S. Geological Survey, and Broward County Environmental Planning and Community Resilience Division provided the funding for this study. The type collections of Saudia and Thomasella were made available for study in April 2018 through the courtesy of Dr. Stephen Stukins, Natural History Museum, UK, who also provided images of the holotype of Saudia discoidea Henson. Prior to journal submission, Pamela Hallock and Viviana Diaz provided reviews that improved the manuscript. Richard Westcott assisted with preparation of
Figure 1 ![]() . We thank Ercan Özcan and one other anonymous reviewer for their comments on the initial manuscript submitted.
. We thank Ercan Özcan and one other anonymous reviewer for their comments on the initial manuscript submitted.
Applin E.R. & Jordan L. (1945).- Diagnostic foraminifera from subsurface formations in Florida.- Journal of Paleontology, Cambridge (UK), vol. 19, no. 2, p. 129-148.
Applin P.L. & Applin E.R. (1944).- Regional subsurface stratigraphy and structure of Florida and southern Georgia.- Bulletin of the American Association of Petroleum Geologists, Tulsa - OK, vol. 28, no. 12, p. 1673-1753 (5 Pls., 38 Figs.).
Barker R.W. & Grimsdale T.F. (1936).- A contribution to the phylogeny of the orbitoidal foraminifera, with descriptions of new forms from the Eocene of Mexico.- Journal of Paleontology, Cambridge (UK), vol. 10, p. 231-247 (Pls. 30-38).
BouDagher-Fadel M.K. & Price G.D. (2010).- Evolution and paleogeographic evolution of the lepidocyclinids.- Journal of Foraminiferal Research, Lawrence - KS, vol. 40, no. 1, p. 79-108.
Bowen-Powell J. (2010).- Larger foraminiferal biostratigraphy, systematics and paleoenvironments of the Avon Park Formation and Ocala Limestone, Highlands County, Florida.- PhD dissertation, Florida International University, Miami - FL, 239 p.
Butterlin J. (1981).- Claves para la determinacion de Macroforaminiferos de Mexico y del Caribe, del Cretacico superior al Mioceno medio.- Instituto Mexicano del Petroleo, Mexico - DC, 219 p. (51 Pls.).
Butterlin J. (1990).- Problèmes posés par la systématique de la famille Lepidocyclinidae (Foraminiferida).- Revista Española de Micropaleontología, Madrid, vol. 22, p. 101-126.
Caudri C.M.B. (1996).- The larger foraminifera of Trinidad (West Indies).- Eclogae Geologicae Helvetiae, Basel, Band 89, Heft 3, p. 1137-1309 (30 Pls.). DOI: https://dx.doi.org/10.5169/seals-167940
Cole W.S. (1938).- Stratigraphy and micropaleontology of two deep wells in Florida.- Florida Geological Survey, Geological Bulletin, Tallahassee - FL, no. 16, p. 1-48 (12 Pls.).
Cole W.S. (1941).- Stratigraphic and paleontologic studies of wells in Florida.- Florida Geological Survey, Geological Bulletin, Tallahassee - FL, no. 16, p. 1-53 (18 Pls.).
Cole W.S. (1942).- Stratigraphic and paleontologic studies of wells in Florida - no. 2.- Florida Geological Survey, Geological Bulletin, Tallahassee - FL, no. 20, p. 1-55.
Cotton L.J., Eder W. & Floyd J. (2018).- Larger foraminifera of the Devil's Den and Blue Hole sinkholes, Florida.- Journal of Micropalaeontology, London, vol. 37, p. 347-356.
Cunningham K.J. (2015).- Seismic-sequence stratigraphy and geologic structure of the Floridan aquifer system near "Boulder Zone" deep wells in Miami-Dade County, Florida.- U.S. Geological Survey Scientific Investigations Report 2015-5013, 28 p. (2 Pls.). DOI: http://dx.doi.org/10.3133/sir20155013
Cunningham K.J. & Robinson E. (2017).- Lithofacies and sequence stratigraphic description of the upper part of the Avon Park Formation and the Arcadia Formation in U.S. Geological Survey G-2984 test corehole, Broward County, Florida.- U.S. Geological Survey Open File-Report, Fort Lauderdale - FL, 2017-1074, 139 p. DOI: https://doi.org/10.3133/ofr20171074
Cunningham K.J., Dixon J.F., Westcott R.L., Norgard S. & Walker C. (2018a).- Three-dimensional seismic characterization of karst in the Floridan aquifer system, southeastern Miami-Dade County, Florida.- U.S. Geological Survey Scientific Investigations Report, Fort Lauderdale - FL, 2018-5117, 39 p. DOI: https://doi.org/10.3133/sir20185117
Cunningham K.J., Kluesner J.W., Westcott R.L., Robinson E., Walker C. & Khan S.A. (2018b).- Sequence stratigraphy, seismic stratigraphy, and seismic structures of the lower intermediate confining unit and most of the Floridan aquifer system, Broward County, Florida (ver. 1.1, January 2018).- U.S. Geological Survey Scientific Investigations Report, Fort Lauderdale - FL, 2017-5109, 71 p. (21 Pls.). DOI: https://doi.org/10.3133/sir20175109
Cushman J.A. (1919).- Fossil foraminifera from the West Indies.- Carnegie Institution, Washington - DC, Publication 291m, p. 21-71.
Cushman J.A. & Ponton G.M. (1933).- A new genus of the foraminifera, Gunteria, from the middle Eocene of Florida.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon - MA, vol. 9, no. 2, p. 25-30. URL: https://cushmanfoundation.org/PersonifyEbusiness/Portals/0/pdf/pubarchive/cclfr/9cclfr2.pdf
Drobne K. & Hottinger L. (1971).- Broeckinella und Saudia (Foraminifera) aus dem nordwestlichen teil Jugoslawiens, ihre Morphologie und ihre Stratigraphische Verbreitung.- Razprave IV. razreda SAZU, Ljubljana, vol. 14, p. 213-238.
Frost S.H. & Langenheim R.L. (1974).- Cenozoic reef biofacies. Tertiary larger foraminifera and scleractinian corals from Mexico.- Northern Illinois University Press, DeKalb - IL, 388 p. (123 Pls.).
Galloway J.J. (1933).- A Manual of the Foraminifera.- Principia Press, Bloomington - IN, 483 p.
Grimsdale T.F. (1952).- Cretaceous and Tertiary Foraminifera from the Middle East.- Bulletin of the British Museum (Natural History), Geology, London, vol. 1, no. 8, p. 223-247 (Pls. 20-25).
Henson F.R.S. (1948).- Larger imperforate Foraminifera of South-western Asia.- British Museum (Natural History), London, 127 p.
Hottinger L. (1967).- Foraminifères imperforés du Mésozoïque marocain.- Notes et Mémoires du Service Géologique, Rabat, no. 209, p. 7-168.
Hottinger L. (1997).- Shallow benthic foraminiferal assemblages as signals for depth of their deposition and their limitations.- Bulletin de la Société géologique de France, Paris, vol. 168, no. 4, p. 491-505.
Kaminski M.A. (2014).- The year 2010 classification of the agglutinated foraminifera. In: Advances in agglutinated foraminiferal research: The Ninth International Workshop on Agglutinated Foraminifera, IWAF-9.- Micropaleontology, New York - NY, vol. 60, no. 1, p. 89-108.
Levin H.L. (1957).- Micropaleontology of the Oldsmar limestone (Eocene) of Florida.- Micropaleontology, New York - NY, vol. 3, no. 2, p. 137-154 (chart 1).
Loeblich A.R. & Tappan H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York - NY, 970 p. (847 Pls.).
Matsumaru K. & Sarma A. (2010).- Larger foraminiferal biostratigraphy of the lower Tertiary of Jaintia Hills, Meghalaya, NE India.- Micropaleontology, New York - NY, vol. 56, no. 6, p. 539-565.
Mitchell S.F., Robinson E., Özcan E., Jiang M.M. & Robinson N. (2022).- A larger benthic foraminiferal zonation for the Eocene of the Caribbean and central American region.- Carnets Geol., Madrid, vol. 22, no. 11, p. 409-565. DOI: https://doi.org/10.2110/carnets.2022.2211
Moberg M.W. (1928).- New species of Coskinolina and Dictyoconus (?) from Florida.- Florida State Geological Survey, 19th Annual Report, Tallahassee - FL, p. 166-175.
Robinson E. (1974).- Some larger foraminifera from the Eocene limestone at Red Gal Ring, Jamaica. In: Jung P. (ed.), Contributions to the geology and paleobiology of the Caribbean and adjacent areas.- Verhandlungen der Naturforschenden Gesellschaft, Basel, vol. 84, no. 1, p. 281-292.
Robinson E. & Wright R.M. (1993).- Jamaican Paleogene larger foraminifera. In: Wright R.M. & Robinson E. (eds.), Biostratigraphy of Jamaica.- Geological Society of America, GSA Memoirs, Boulder - CO, vol. 182, p. 283-345.
Rutten M.G. (1935).- Larger foraminifera of Northern Santa Clara Province Cuba.- Journal of Paleontology, Cambridge (UK), vol. 9, p. 527-545.
Serra-Kiel J., Ferràndez-Cañadell C., García-Senz J. & Hernaiz Huerta P.P. (2007).- Cainozoic larger foraminifers from Dominican Republic.- Boletín Geológico y Minero, Madrid, vol. 118, no. 2, p. 359-384. URL: https://www.igme.es/boletin/2007/118_2_2007/ART.%2010.pdf
Serra-Kiel J., Hottinger L., Caus E., Drobne K., Ferrandez C., Jauhri A. K., Less G., Pavlovec R., Pignatti J., Samso J.M., Schaub H., Sirel E., Stougo A., Tambareau Y., Tosequella J. & Zakrevskya E. (1998).- Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene.- Bulletin de la Société géologique de France, Paris, vol. 169, no. 2, p. 281-299.
Sirel E. (1998).- Foraminiferal and biostratigraphy of the Paleocene - lower Eocene shallow water limestones and discussion on the Cretaceous - Tertiary boundary in Turkey.- General Directorate of the Mineral Research and Exploration, Monography Series (ser. 2), Ankara, 117 p. (68 Pls.).
Sirel E. (1999).- Four new genera (Haymanella, Kayseriella, Elazigella and Orduella) and one new species of Hottingerina from the Paleocene of Turkey.- Micropaleontology, New York - NY, vol. 45, no. 2, p. 113-137.
Torres-Silva A.I., Eder W., Hohenegger J. & Briguglio A. (2019).- Morphometric analysis of Eocene nummulitids in western and central Cuba: Taxonomy, biostratigraphy and evolutionary trends.- Journal of Systematic Palaeontology, vol. 17, no. 7, p. 557-595.
Vaughan T.W. (1945).- American old and middle Tertiary larger Foraminifera and corals: Part I - American Paleocene and Eocene larger Foraminifera.- Geological Society of America, GSA Memoirs, Boulder - CO, vol. 9, 175 p.
Vicedo V., Berlanga J.A. & Serra-Kiel J. (2014).- Paleocene larger foraminifera from the Yucatán Peninsula (SE Mexico).- Carnets Geol., Madrid, vol. 14, no. 4, p. 41-68. DOI: https://doi.org/10.4267/2042/53527
Winston G.O. (1993).- The Paleogene of Florida, Volume 2: A regional analysis of the Oligocene-Eocene section of the peninsula using vertical lithologic stacks.- Miami Geological Society, Coral Gables - FL, 33 p.
Wong T.E. (1976).- Tertiary stratigraphy and micropaleontology of the Guiana Basin.- Doctoral thesis, University of Utrecht; Geologie Mijnbouwkundige Dienst, Suriname, Paramaribo, 105 p.
| Nomenclatural note: Life Sciences Identifier (LSID) https://zoobank.org/References/0deaa536-4324-492f-aa9b-43654966f6ab Genus Group Species Group |