◄ Carnets Geol. 22 (20) ►
![]()
Outline:
[1. Introduction]
[2. Material and methods]
[3. Geological and palaeontological background]
[4. Systematics]
[5. Discussion and conclusions]
and ... [Bibliographic references]
Dipartimento di Scienze della Terra,
Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Museo di Storia Naturale, Universitŕ
di Pisa, via Roma 79, 56011 Calci (Italy)
Dipartimento di Scienze della Terra,
Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Dottorato Regionale in Scienze della
Terra "Pegaso", via Santa Maria 53, 56126 Pisa (Italy)
Department of Paleontology, Calvert
Marine Museum, PO Box 97, Solomons, Maryland 20688 (U.S.A.);
Research Associate, National Museum
of Natural History, Smithsonian Institution, Washington, DC 20560 (U.S.A.)
Dipartimento di Scienze della Terra,
Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy);
Museo di Storia Naturale, Universitŕ
di Pisa, via Roma 79, 56011 Calci (Italy)
Published online in final form (pdf) on December 25, 2022
DOI 10.2110/carnets.2022.2220
![]()
[Editor:
Brian Pratt; technical editor: Bruno R.C. Granier]
![]()
Faeces produced by marine vertebrates and macro-invertebrates contain sufficient organic matter to represent a usable food source for a wide array of macroscopic animals. In some extant marine environments, coprophagy even represents a crucial trophic interaction in food webs. In ancient ecosystems, coprophagy by macroscopic animals is occasionally exemplified by coprolites that exhibit biting traces or burrows. Here, we report Gnathichnus pentax on an exquisitely preserved vertebrate-bitten vertebrate coprolite from the marine calcareous deposits of the Pietra leccese (Miocene, southern Italy). This unusual occurrence is interpreted as evidence of the feeding activity of a regular echinoid; in particular, it may represent either exploratory coprophagy or the browsing of an algal (microbial) film that locally developed on the exterior of the faeces. Strengthening the former interpretation, the development of microbial communities on submerged faeces often leads to their destruction; furthermore, some extant Antarctic echinoderms are well known to ordinarily feed on vertebrate faeces, and coprophagy is believed to be fairly widespread among sea urchins. Supporting the algal browsing hypothesis, in turn, only a limited area of the external surface of the faeces was subject to grazing, and the resulting trace is neatly defined, which suggest that the feeding sea urchin targeted a precise location on the dung's exterior when the latter was already rather firm. To our knowledge, the G. pentax specimen studied here represents the first published record of this ichnotaxon on a coprolite.
• bromalite;
• coprophagy;
• Echinodermata;
• palaeoichnology;
• Pascichnia;
• Pietra leccese;
• sea urchin;
• Vertebrata
Collareta A., Peri E., Godfrey S.J. & Bianucci G. (2022).- Just a different place to graze? An unusual occurrence of the echinoid feeding trace Gnathichnus pentax on a marine vertebrate coprolite (Miocene, Italy) and its palaeoethological implications.- Carnets Geol., Madrid, vol. 22, no. 20, p. 847-855.
Juste un endroit différent pour brouter ? Une occurrence inhabituelle de la trace d'alimentation d'un échinoďde Gnathichnus pentax sur un coprolithe de vertébrés marins (Miocčne, Italie) et ses implications paléoéthologiques.- Les matičres fécales produites par les vertébrés et les macro-invertébrés marins contiennent une quantité suffisante de matičre organique pour représenter une source de nourriture utilisable pour un large éventail d'animaux macroscopiques. Dans certains environnements marins modernes, la coprophagie représente une interaction trophique cruciale dans les réseaux trophiques. Dans les écosystčmes anciens, la coprophagie par des animaux macroscopiques est parfois illustrée par des coprolithes qui présentent des traces de morsure ou des terriers. Ici, nous rapportons un spécimen de Gnathichnus pentax présent sur un coprolithe de vertébré, parfaitement préservé et lui-męme mordu par des vertébrés, provenant des dépôts calcaires marins de la Pietra leccese (Miocčne, sud de l'Italie). Cette occurrence inhabituelle est interprété comme une preuve de l'activité alimentaire d'un échinoďde régulier; en particulier, il peut résulter soit d'une coprophagie exploratoire, soit du broutage d'un film algal (microbien) qui s'était développé localement ŕ l'extérieur des fčces. Renforçant la premičre interprétation, le développement de communautés microbiennes sur les matičres fécales submergées conduit souvent ŕ leur destruction ; en outre, certains échinodermes vivants antarctiques sont bien connus pour se nourrir habituellement d'excréments de vertébrés et la coprophagie semble ętre assez répandue chez les oursins. D'autre part, ŕ l'appui de l'hypothčse du broutage algal, seule une zone limitée de la surface externe des fčces était sujette au pâturage, et la trace résultante est bien définie, ce qui suggčre que l'oursin en train de se nourrir visait un endroit précis ŕ l'extérieur de l'excrément alors que le dernier était déjŕ assez raide. Ŕ notre connaissance, le spécimen de G. pentax étudié ici représente le premier enregistrement publié de cet ichnotaxon sur un coprolithe.
• bromalithe ;
• coprophagie ;
• Echinodermata ;
• paléoichnologie ;
• Pascichnia ;
• Pietra leccese ;
• oursin ;
• Vertébrés
It is well known that the faeces produced by marine vertebrates and macro-invertebrates contain sufficient organic matter to represent a usable food source for a wide array of macroscopic animals (Newell, 1965; Frankenberg & Smith, 1967; Frankenberg et al., 1967; Robertson, 1982; Parrish, 1989; Sazima et al., 2003). In some extant marine environments, including coral reefs, the consumption of faeces (i.e., coprophagy) is so important that it is sometimes a crucial kind of trophic interaction in food webs (Bailey & Robertson, 1982; Fuller & Parsons, 2019). In ancient ecosystems, coprophagy by macroscopic animals is occasionally envisaged on the basis of fossilised faeces (i.e., coprolites) that display feeding-related modifications like tooth marks (e.g., Godfrey & Smith, 2010; Godfrey & Palmer, 2015; Godfrey & Frandsen, 2016; Dentzien-Dias et al., 2018, 2021; Frandsen & Godfrey, 2019; Cueille et al., 2020; Godfrey et al., 2020; Rozada et al., 2021; Rummy et al., 2021) and burrows (Bradley, 1946; Chin & Gill, 1996; Chin, 2007; Milŕn et al., 2012; Godfrey et al., 2022; Godfrey & Collareta, 2022). Overall, the fossil record demonstrates that faecal matter has been an important source of nutrition in the marine realm since the Cambrian (Kimmig & Pratt, 2018; Hunt et al., 2021).
With the aim of contributing to the short but growing list of coprolites with associated feeding traces, Collareta et al. (2019) described fossilised vertebrate (shark?) dung from the Miocene of southern Italy featuring tooth marks referred to both cartilaginous and bony fishes. The recent reappraisal of this coprolite led to the identification of an additional trace originating from the grazing activity of a macro-invertebrate, namely, a sea urchin. Here, we characterize and describe this echinoid feeding trace from this remarkable coprolite.
The studied coprolite is currently housed in the Museo di Storia Naturale dell'Universitŕ di Pisa (=MSNUP, Calci, Pisa Province, Italy) with accession number MSNUP I-17604. Photographs and microphotographs of MSNUP I-17604 were taken using a Nikon D5200 digital camera equipped with a Sigma 50 mm F2.8 macro lens and a Sony a6000 digital camera equipped with a Sony 50 mm F1.8 lens. A 3D model of the specimen was created by aligning 133 photos through the photogrammetric software Agisoft Metashape v1.70. The 3D model is freely available via figshare at the following internet address: https://doi.org/10.6084/m9.figshare.17161235.
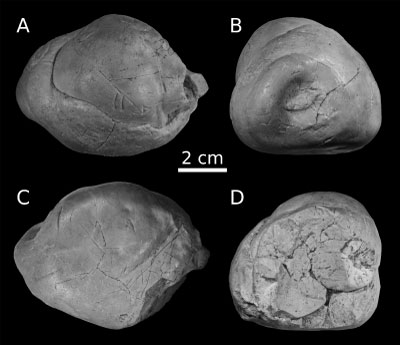
MSNUP
I-17604 (Fig. 1 ![]() ) is an oval, spiral, heteropolar coprolite (sensu Häntzschel
et al., 1968, and Hunt
& Lucas, 2012). It is
almost complete, only lacking some fragments (likely because of post-burial
breakage) at one end. It measures 79 mm in maximum length (estimated total
length if missing parts are included ~85 mm), 58 mm in width, and 54 mm in
thickness. Hand-held energy dispersive X-ray fluorescence analyses demonstrated
that MSNUP I-17604 is very rich in calcium (ca. 36 wt%) and phosphorous (ca. 14
wt%) (Collareta et al., 2019). A number of
different traces occur on the surface of MSNUP I-17604, mostly consisting of
tooth incisions from indeterminate bony fishes and sharks (Collareta et
al., 2019).
) is an oval, spiral, heteropolar coprolite (sensu Häntzschel
et al., 1968, and Hunt
& Lucas, 2012). It is
almost complete, only lacking some fragments (likely because of post-burial
breakage) at one end. It measures 79 mm in maximum length (estimated total
length if missing parts are included ~85 mm), 58 mm in width, and 54 mm in
thickness. Hand-held energy dispersive X-ray fluorescence analyses demonstrated
that MSNUP I-17604 is very rich in calcium (ca. 36 wt%) and phosphorous (ca. 14
wt%) (Collareta et al., 2019). A number of
different traces occur on the surface of MSNUP I-17604, mostly consisting of
tooth incisions from indeterminate bony fishes and sharks (Collareta et
al., 2019).
|
Figure 1:
MSNUP I-17604, trace-bearing vertebrate coprolite from the Miocene Pietra
leccese of southern Italy, in (A-D) four different views, including (B,D) two
terminal views. |
All that is
known about the geographic and stratigraphic origin of MSNUP I-17604 is its
provenance from the Miocene Pietra leccese formation (Collareta et al.,
2019),
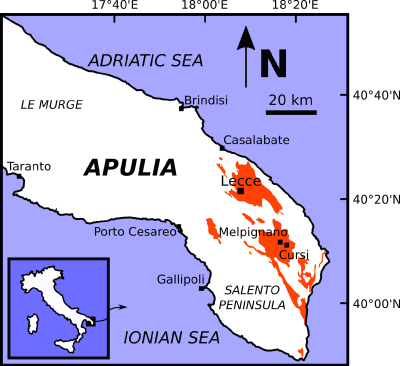
which crops out in the Salento peninsula (Apulia Region, southern Italy; Fig.
2 ![]() ). This informal lithostratigraphic unit consists mostly of foraminiferal
biomicrites and biosparites that were deposited between the late Burdigalian and
the early Messinian (Foresi et al., 2002; Bossio
et al., 2005, 2006; Mazzei
et al., 2009; Margiotta,
2015). A
few major depositional hiatuses occur within the Pietra leccese, being often
marked by glauconite-rich horizons (Balenzano et al.,
2002; Foresi
et al., 2002; Mazzei
et al., 2009). The
depositional environment is considered to be the deepest part of the continental
shelf (Bossio et al., 2005,
2006).
). This informal lithostratigraphic unit consists mostly of foraminiferal
biomicrites and biosparites that were deposited between the late Burdigalian and
the early Messinian (Foresi et al., 2002; Bossio
et al., 2005, 2006; Mazzei
et al., 2009; Margiotta,
2015). A
few major depositional hiatuses occur within the Pietra leccese, being often
marked by glauconite-rich horizons (Balenzano et al.,
2002; Foresi
et al., 2002; Mazzei
et al., 2009). The
depositional environment is considered to be the deepest part of the continental
shelf (Bossio et al., 2005,
2006).
The Pietra leccese is home to an impressive marine vertebrate fossil assemblage. Most of the collected fossils belong to cetaceans, with both toothed and baleen whales well represented (Moncharmont Zei, 1950, 1956; Bianucci et al., 1992, 1994, 2016; Bianucci & Varola, 1994; Bianucci, 2001; Bianucci & Landini, 2006; Bisconti & Varola, 2006; Peri et al., 2019, 2020, 2021, 2022). Sirenians and marine reptiles (turtles and crocodilians) are also present (Costa, 1853, 1856; Aldinio, 1896; Bianucci et al., 2003; Chesi et al., 2007). Fishes are represented by elasmobranchs (sharks and rays) and teleosts (bony fishes; Menesini, 1969; Vigliarolo, 1891; Carnevale et al., 2002; Collareta et al., 2021). Finally, but not the least of which, Digestichnia of marine vertebrates have also been reported, including putative gastroliths as well as the coprolite studied herein (Tavani, 1973; Collareta et al., 2019).
|
Figure 2:
Schematic geological map of the Salento peninsula (Apulia,
southern Italy). Orange-shaded areas indicate the exposures of the Pietra leccese, the Miocene limestone from which the fossil specimen MSNUP
I-17604
originates. Redrawn and modified after the original illustration by Calia
et al. (2014). |
Gnathichnus Bromley, 1975
Remarks. The monotypic ichnogenus Gnathichnus is a grazing structure (Pascichnia) that usually occurs on calcareous invertebrate shells and hardgrounds (e.g., Bromley, 1975; Fürsich & Wendt, 1977; Martinell, 1982; Bromley & Asgaard, 1993a, 1993b; Gibert et al., 2007; Wilson, 2007; Belaústegui et al., 2017; Angseesing, 2021) but has also been described from consolidated sediments such as burrow walls (Mayoral & Muńiz, 1996) and, rarely, vertebrate bones (Twitchett, 1994; Meyer, 2011; Reolid et al., 2015; Jagt et al., 2020).
Höpner and Bertling (2017) argued that since the nutrition of grazers is nonselective regarding the substrate, the nature of the latter is not a valuable ichnotaxobase for naming grazing traces. Such a statement would imply that all grazing traces do originate from feeding upon organisms that encrust or bore into the substrate, and not upon the substrate itself and/or remains of the associated tissues. As discussed below, this premise is controversial, but identifying cases of deliberate targeting of specific substrates by ancient grazers often leads to questionable conclusions, hence our assignment of the gnathichnian trace that occurs on MSNUP I-17604 to the ichnogenus Gnathichnus.
Gnathichnus almost invariably occurs in multiples of several overlapping grooves intersecting at angles of ~72° as a result of the star-shaped arrangement of the five calcium-carbonate teeth that comprise the "Aristotle's lantern", i.e., the chewing organ of most sea urchins (Bromley, 1975).
In spite of its low preservation potential, Gnathichnus even gives its name to an archetypal marine ichnofacies (e.g., Bromley & Asgaard, 1993a, 1993b; Gibert et al., 2007; MacEachern et al., 2007).
Gnathichnus pentax Bromley, 1975
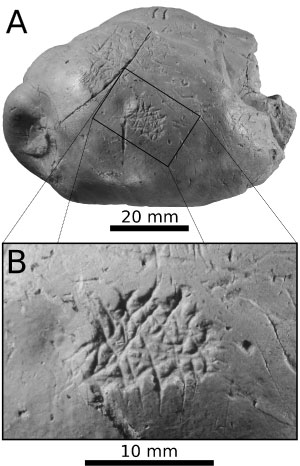
(Fig. 3 ![]() )
)
Referred material. A single cluster of grooves preserved on the surface of MSNUP I-17604, a marine vertebrate coprolite.
Occurrence. MSNUP I-17604 originates from the Pietra leccese, a Miocene (upper Burdigalian to lower Messinian) calcareous formation of southern Italy (Apulia Region, Salento peninsula) (see above for more details).
Description. This Gnathichnus pentax trace consists of a cluster of sub-linear, partly overlapping grooves that are a few millimetres long and meet each other at angles of ~72° (after Bromley, 1975; Fig. 3). The individual grooves have sub-millimetric transverse widths, and their distal terminations are often acuminate. The whole trace fossil appears as a densely sculptured area measuring ~10 mm × 8 mm. The G. pentax trace seemingly obliterates a pre-existing shark tooth mark (ichnospecies Linichnus bromleyi; see Muńiz et al., 2020, and Godfrey & Lowry, 2021).
Remarks. The Pietra leccese preserves remains of regular echinoids (cidaroids and temnopleuroids; Ragaini, 1994) that may account for the production of the gnathichnian trace observed on MSNUP I-17604. Because the morphological quality of the latter is as high as in similar representatives of G. pentax from hard substrates such as oyster valves (e.g., Wilson, 2007: Fig. 20.10), the dung was likely firm when grazing took place. Additionally, the faeces would have had a fairly rigid consistency to facilitate the tube feet attachment (Hennebert et al., 2012) to anchor the sea urchin to the curved surface of the dung so that its teeth could excavate the surface.
|
Figure 3: MSNUP I-17604, trace-bearing vertebrate coprolite from the
Miocene Pietra leccese of southern Italy. (A) Lateral view of the coprolite,
featuring the occurrence of the echinoid feeding trace Gnathichnus pentax
(black rectangle); (B) close-up of the G. pentax specimen. |
To our knowledge, the Gnathichnus pentax specimen studied here represents the first published record of this ichnotaxon on a coprolite, and this is quite remarkable in its own right. A cursory screening of the many marine vertebrate coprolites stored in the palaeontological collections of the Calvert Marine Museum (Solomons, Maryland, USA) revealed no specimens preserving gnathichnian traces (SJG, personal observation), though other kinds of traces (including bite marks) are present (e.g., Godfrey & Smith, 2010; Godfrey et al., 2020, 2022). Thus, fossilised vertebrate faeces are a very rare substrate for the preservation of G. pentax.
What kind of behaviour is responsible for this unusual occurrence? Coprophagy certainly represents an intriguing explanation. Some extant Antarctic echinoderms, including the echinid sea urchin Sterechinus neumayeri, are known to feed on vertebrate faeces such as seal droppings (Pearse & Giese, 1966). Furthermore, according to McClintock (1994), "[t]he ingestion of faecal material is not unique to Antarctic echinoderms, but to asteroids and echinoids in general." In light of these neontological observations, the G. pentax specimen observed on MSNUP I-17604 may be suggestive of coprophagy by a regular echinoid. As such, it would represent the first fossil evidence for faeces consumption by a sea urchin; in addition, it would demonstrate that coprophagy has been practiced by regular echinoids at least since the Miocene.
On the other hand, browsing of an algal (microbial) film that locally developed on the exterior of the faeces is also a possibility. Indeed, regardless of the nature of the substrate, gnathichnian traces have invariably been interpreted as evidence of the feeding activity of regular echinoids, and especially the grazing of epilithic (encrusting) and/or endolithic (boring) organisms (mostly algae) (Bromley, 1975; Fürsich & Wendt, 1977; Bromley & Asgaard, 1993b; Gibert et al., 2007; Wilson, 2007; Jagt et al., 2020; Angseesing, 2021), though feeding on residual organic matter associated with vertebrate and invertebrate hardparts or scavenging of bone tissues for phosphate has also been proposed (e.g., Breton et al., 1992; Reolid et al., 2015). Considering the trace that occurs on MSNUP I-17604, support for the algal browsing hypothesis may come from observing that only a limited area was subject to grazing, which suggests that the feeding echinoid targeted a precise location on the dung's exterior; furthermore, as mentioned, the faeces were likely firm when feeding took place. That said, even this scenario is not without concerns: indeed, the development of microbial communities on submerged faeces often leads to their destruction, whereas the exquisite state of preservation of MSNUP I-17604 evokes the rapid burial and early lithification of the dung (Bradley, 1946; Thulborn, 1991; Dentzien-Dias et al., 2012, 2018).
As detailed elsewhere (Collareta et al., 2019), the studied coprolite also displays shark and bony fish tooth marks, which in turn suggests that for some reason it represented a target for exploratory consumption for a variety of marine organisms, including both vertebrate and invertebrate taxa. Thus, this new find further strengthens the notion that the analysis of coprolites is a powerful tool for investigating the feeding behaviours of ancient organisms, though sometimes in largely unexpected ways.
We are grateful to Chiara Sorbini (Museo di Storia Naturale, Universitŕ di Pisa) for providing access to the palaeontological material studied in the present work. Constructive comments by Mark A. Wilson and an anonymous reviewer contributed to shape this work into its present form – thank you very much! Not least, thanks are due to Brian Pratt and to Bruno Granier for their valuable and thorough editorial support.
As many other vertebrate fossils from the Pietra leccese, the coprolite MSNUP I-17604 was collected by the late and missed Angelo Varola in the framework of the scientific activities of the Gruppo Naturalisti Salentini.
Aldinio P. (1896).- Sul Tomistoma (Gavialosuchus) lyceensis del Calcare miocenico di Lecce.- Atti dell'Accademia Gioenia di Scienze Naturali, Catania, vol. IX, memoria XV, p. 1-11 (2 Pls.). URL: https://archive.org/details/mobot31753002141809/page/n257/mode/2up
Angseesing J. (2021).- Gnathichnus trace fossils on three Middle Jurassic terebratulid assemblages of the Cotswold Hills.- Proceedings of the Cotteswold Naturalists' Field Club, London & Gloucester, vol. 48, p. 348-356.
Bailey T.G. & Robertson D.R. (1982).- Organic and caloric levels of fish feces relative to its consumption by coprophagous reef fishes.- Marine Biology, Berlin, vol. 69, p. 45-50.
Balenzano F., Margiotta S. & Moresi M. (2002).- Significato di un deposito glauconitico-fosfatico appartenente ad una Unitŕ Miocenica del Salento (Puglia).- Atti della Societŕ Toscana Di Scienze Naturali, Memorie (Serie A), Pisa, vol. 108, p. 7-21.
Belaústegui Z., Muńiz F., Nebelsick J.H., Domčnech R. & Martinell J. (2017).- Echinoderm ichnology: Bioturbation, bioerosion and related processes.- Journal of Paleontology, Cambridge (UK), vol. 91, no. 4, p. 643-661.
Bianucci G. (2001).- A new genus of kentriodontid (Cetacea: Odontoceti) from the Miocene of South Italy.- Journal of Vertebrate Paleontology, vol. 21, p. 573-577.
Bianucci G. & Angelo V. (2014).- I cetacei fossili della Pietra leccese nei musei del Salento.- Museologia Scientifica Memorie, Firenze, vol. 13, p. 130-134.
Bianucci G., Collareta, A., Post K., Varola A. & Lambert O. (2016).- A new record of Messapicetus from the Pietra leccese (late Miocene, southern Italy): Antitropical distribution in a fossil beaked whale (Cetacea, Ziphiidae).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 122, no. 1, p. 63-74. URL: https://riviste.unimi.it/index.php/RIPS/article/view/6930
Bianucci G. & Landini W. (2006).- Killer sperm whale: A new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy.- Zoological Journal of the Linnean Society, London, vol. 148, p. 103-131.
Bianucci G., Landini W. & Varola A. (1992).- Messapicetus longirostris, a new genus and species of Ziphiidae (Cetacea) from the Late Miocene of "Pietra leccese" (Apulia, Italy).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 31, no. 2, p. 261-264.
Bianucci G., Landini W. & Varola A. (1994).- New remains of Cetacea Odontoceti from the "Pietra leccese" (Apulia, Italy).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 33, no. 2, p. 215-230.
Bianucci G., Landini W. & Varola A. (2003).- New records of Metaxytherium (Mammalia: Sirenia) from the late Miocene of Cisterna quarry (Apulia, southern Italy).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 42, p. 59-64.
Bianucci G., Landini W. & Varola A. (2004).- First discovery of the Miocene northern Atlantic sperm whale Orycterocetus in the Mediterranean.- Géobios, Villeurbanne, vol. 37, p. 569-573.
Bisconti M. & Varola A. (2006).- The oldest eschrichtiid mysticete and a new morphological diagnosis of Eschrichtiidae (gray whales).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 112, p. 447-457.
Bossio A., Foresi L.M., Margiotta S., Mazzei R., Salvatorini G. & Donia F. (2006).- Stratigrafia neogenico-quaternaria del settore nord-orientale della Provincia di Lecce (con rilevamento geologico alla scala 1:25.000).- Geologica Romana, vol. XXXIX, p. 63-88. URL: https://www.dst.uniroma1.it/geologicaromana/Volumi/VOL%2039/3%20Bossio.pdf
Bossio A., Mazzei R., Monteforti B. & Salvatorini G. (2005).- Stratigrafia del Neogene e Quaternario del Salento Sud-orientale (con rilevamento geologico alla scala 1:25.000).- Geologica Romana, vol. XXXVIII, p. 31-60. URL: https://www.dst.uniroma1.it/geologicaromana/Volumi/VOL%2038/6%20bossio.pdf
Bradley W.H. (1946).- Coprolites from the Bridger Formation of Wyoming: Their composition and microorganisms.- American Journal of Science, New Haven - CT, vol. 244, no. 3, p. 215-239.
Breton G., Neraudeau D. & Cuenca-Boulat C. (1992).- Gnathichnus stellarum ichnosp. nov., trace de broutage d'un echinide du Campanien des Charentes (France).- Revue de Paléobiologie, Genčve, vol. 11, no. 1, p. 219-229.
Bromley R.G. (1975).- Comparative analysis of fossil and recent echinoid bioerosion.- Palaeontology, London, vol. 18, part 4, p. 725-739.
Bromley R.G. & Asgaard U. (1993a).- Two bioerosion ichnofacies produced by early and late burial associated with sea-level change.- Geologische Rundschau, Berlin, vol. 82, no. 2, p. 276-280.
Bromley R.G. & Asgaard U. (1993b).- Endolithic community replacement on a Pliocene rocky coast.- Ichnos, Boca Raton - FL, vol. 2, no. 2, p. 93-116.
Calia A., Laurenzi Tabasso M., Mecchi A.M. & Quarta G. (2014).- The study of stone for conservation purposes: Lecce stone (southern Italy).- Geological Society, London, vol. 391, Special Publications, p. 139-156.
Carnevale G., Sorbini C., Landini W. & Varola A. (2002).- Makaira cf. M. nigricans Lacépčde, 1802 (Teleostei: Perciformes: Istiophoridae) from the Pietra leccese, Late Miocene, Apulia, Southern Italy.- Palaeontographia Italica, Pisa, vol. 88, p. 63-75.
Chesi F., Delfino M., Varola A. & Rook L. (2007).- Fossil sea turtles (Chelonii, Dermochelyidae and Cheloniidae) from the Miocene of Pietra Leccese (late Burdigalian–early Messinian), Southern Italy.- Geodiversitas, Paris, vol. 29, no. 2, p. 231-333.
Chin K. (2007).- The paleobiological implications of herbivorous dinosaur coprolites from the Upper Cretaceous Two Medicine Formation of Montana: why eat wood?- Palaios, Lawrence - KS, vol. 22, p. 554-566.
Chin K. & Gill B.D. (1996).- Dinosaurs, dung beetles, and conifers: participants in a Cretaceous food web.- Palaios, Lawrence - KS, vol. 11, p. 280-285.
Collareta A., Gemelli M., Varola A. & Bianucci G. (2019).- Trace fossils on a trace fossil: A vertebrate-bitten vertebrate coprolite from the Miocene of Italy.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 293, no. 2, p. 117-126.
Collareta A., Peri E., Carnevale G., Bosselaers M. & Bianucci G. (2021).- On the presence of an ocean sunfish (Tetraodontiformes, Molidae) in the Miocene Pietra Leccese formation of Southern Italy.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 301, no. 2, p. 147-155.
Costa O.G. (1853).- Paleontologia del Regno di Napoli. Parte I.- Atti dell'Accademia Pontaniana, Naples, vol. 5, p. 233-433.
Costa O.G. (1856).- Paleontologia del Regno di Napoli. Parte II.- Atti dell'Accademia Pontaniana, Naples, vol. 7, p. 1-378.
Cueille M., Green E., Duffin C.J. Hildebrand C. & Benton M.J. (2020).- Fish and crab coprolites from the latest Triassic of the UK: From Buckland to the Mesozoic Marine Revolution.- Proceedings of the Geologists' Association, London, vol. 131, no. 6, p. 699-721.
Dentzien-Dias P.C., de Figueiredo A.E.Q., Horn B., Cisneros J.C. & Schultz C.L. (2012).- Paleobiology of a unique vertebrate coprolites concentration from Rio do Rasto formation (Middle/Upper Permian), Paraná Basin, Brazil.- Journal of South American Earth Sciences, vol. 40, p. 53-62.
Dentzien-Dias P., Carrillo-Briceńo J.D., Francischini H. & Sánchez R. (2018).- Paleoecological and taphonomical aspects of the Late Miocene vertebrate coprolites (Urumaco Formation) of Venezuela.- Palćogeography, Palćoclimatology, Palćoecology, vol. 490, p. 590-603.
Dentzien-Dias P., Hunt A.P., Lucas S.G., Francischini H. & Gulotta M. (2021).- Coprolites from shallow marine deposits of the Nanjemoy Formation, Lower Eocene of Virginia, USA.- Lethaia, Oslo, vol. 54, no. 1, p. 26-39.
Foresi L.M., Margiotta S. & Salvatorini G. (2002).- Bio-cronostratigrafia a foraminiferi planctonici della Pietra leccese (Miocene) nell'area tipo di Cursi-Melpignano (Lecce, Puglie).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 41, no. 2-3, p. 175–185.
Frandsen G. & Godfrey S.J. (2019).- A gar-bitten coprolite from the Eocene Green River Formation near Kemmerer, Wyoming, U.S.A.- The Ecphora, Solomons - MD, vol. 34, p. 2-4.
Frankenberg D. & Smith K.L. Jr (1967).- Coprophagy in marine animals.- Limnology and Oceanography, vol. 12, no. 3, p. 443-450.
Frankenberg D., Coles S.L. & Johannes R.E. (1967).- The potential trophic significance of Callianassa major fecal pellets.- Limnology and Oceanography, vol. 12, no. 1, p. 113-120.
Fuller L.N. & Parsons G.R. (2019).- A note on associations observed between sharks and teleosts.- Southeastern Naturalist, Steuben - ME, vol. 18, no. 3, p. 489-498.
Fürsich F.T. & Wendt J. (1977).- Biostratinomy and palaeoecology of the Cassian formation (Triassic) of the Southern Alps.- Palćogeography, Palćoclimatology, Palćoecology, vol. 22, no. 4, p. 257-323.
Gibert J.M. de, Domčnech R. & Martinell J. (2007).- Bioerosion in shell beds from the Pliocene Roussillon Basin, France: Implications for the (macro) bioerosion ichnofacies model.- Acta Palaeontologica Polonica, Warsaw, vol. 52, no. 4, p. 783-798. URL: https://www.app.pan.pl/article/item/app52-783.html
Godfrey S.J. & Collareta A. (2022).- A new ichnotaxonomic name for burrows in vertebrate coprolites from the Miocene Chesapeake Group of Maryland, U.S.A.- Swiss Journal of Palaeontology, vol. 141, article 9, 9 p. DOI: https://doi.org/10.1186/s13358-022-00250-6
Godfrey S.J., Alford A., Collareta A. & Weems R.E. (2020).- A Paleocene vertebrate-bitten crocodilian coprolite from Liverpool Point, Maryland, USA.- Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, Stuttgart, vol. 296, no. 3, p. 237-244.
Godfrey S.J., Collareta A. & Nance J.R. (2022).- Coprolites from Calvert Cliffs: Miocene fecal pellets and burrowed crocodilian droppings from the Chesapeake Group of Maryland, U.S.A.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 128, no. 1, p. 69-79. DOI: https://doi.org/10.54103/2039-4942/17064
Godfrey S.J. & Lowry A.J. (2021).- The ichnospecies Linichnus bromleyi on a Miocene baleen whale radius preserving multiple shark bite-shake traces suggests scavenging.- Carnets Geol., Madrid, vol. 21, no. 17, p. 391-398. DOI: https://doi.org/10.2110/carnets.2021.2117
Godfrey S.J. & Frandsen G. (2016).- Vertebrate-bitten coprolite from South Carolina. - The Ecphora, Solomons - MD, vol. 31, p. 12-14.
Godfrey S.J. & Palmer B.T. (2015).- Gar-bitten coprolite from South Carolina, USA.- Ichnos, Boca Raton - FL, vol. 22, no. 2, p. 103-108.
Godfrey S.J. & Smith J.B. (2010).- Shark-bitten vertebrate coprolites from the Miocene of Maryland.- Naturwissenschaften, Berlin, vol. 97, no. 5, p. 461-467.
Häntzschel W., El-Baz F. & Amstutz G.C. (1968).- Coprolites: An annotated bibliography.- Memoirs of the Geological Society of America, Boulder - CO, vol. 108, 132 p.
Hennebert E., Santos R. & Flammang P. (2012).- Echinoderms don't suck: Evidence against the involvement of suction in tube foot attachment.- Zoosymposia, Auckland, vol. 7, p. 25-32.
Höpner S. & Bertling M. (2017).- Holes in bones: Ichnotaxonomy of bone borings.- Ichnos, Boca Raton - FL, vol. 24, no. 4, p. 259-282.
Hunt A.P. & Lucas S.G. (2012).- Descriptive terminology of coprolites and recent feces.- New Mexico Museum of Natural History and Science Bulletin, Albuquerque - NM, vol. 57, p. 153-160.
Hunt A.P. & Lucas S.G. (2021).- The ichnology of vertebrate consumption: Dentalites, gastroliths and bromalites.- New Mexico Museum of Natural History and Science Bulletin, Albuquerque - NM, vol. 87, p. 1-215.
Jagt J.W., Deckers M.J., De Leebeeck M., Donovan S.K. & Nieuwenhuis E. (2020).- Episkeletozoans and bioerosional ichnotaxa on isolated bones of Late Cretaceous mosasaurs and cheloniid turtles from the Maastricht area, the Netherlands.- Geologos, Poznań, vol. 26, no. 1, p. 39-49. DOI: https://doi.org/10.2478/logos-2020-0003
Kimmig J. & Pratt B.R. (2018).- Coprolites in the Ravens Throat River Lagerstätte of northwestern Canada: Implications for the middle Cambrian food web.- Palaios, Lawrence - KS, vol. 33, no. 4, p. 125-140.
MacEachern J.A., Bann K.L., Pemberton S.G. & Gingras M.K. (2007).- The Ichnofacies paradigm. High-resolution paleoenvironmental interpretation of the rock record. In: MacEachern J.A., Bann K.L., Gingras M.K. & Pemberton S.G. (eds.), Applied ichnology.- SEPM Short Course Notes, Lawrence - KS, vol. 52, p. 103-138.
Margiotta S. (2015).- Inquadramento geologico e territoriale della pietra leccese.- Thalassia Salentina, Lecce, vol. 37, p. 17-28.
Martinell J. (1982).- Echinoid bioerosion from the Pliocene of NE Spain.- Géobios, Villeurbanne, vol. 15, no. 2, p. 249-253.
Mayoral E. & Muńiz F. (1996).- The Gnathichnus ichnofacies in the southwestern sector of the Guadalquivir Basin (Lepe, Huelva, Spain).- Coloquios de Paleontología, Madrid, vol. 48, p. 87-102. URL: https://dialnet.unirioja.es/servlet/articulo?codigo=169686
Mazzei R., Margiotta S., Foresi L.M., Riforgiato F. & Salvatorini G. (2009).- Biostratigraphy and chronostratigraphy of the Miocene Pietra Leccese in the type area of Lecce (Apulia, southern Italy).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 48, no. 2, p. 129-145.
McClintock J.B. (1994).- Trophic biology of Antarctic shallow-water echinoderms.- Marine Ecology Progress Series, Oldendorf, vol. 111, no. 1, p. 191-202.
Menesini E. (1969).- Ittiodontoliti Miocenici di Terra d'Otranto.- Palaeontographia Italica, Pisa, vol. 65, p. 1-61.
Meyer C.A. (2011).- Amazing graze–Grazing traces of sea urchins on turtles – An example from the Late Jurassic of Switzerland.- Annalen des Naturhistorischen Museums in Wien (Serie A), vol. 113, p. 555-565.
Milŕn J. (2012).- Crocodylian scatology–a look into morphology, internal architecture, inter-and intraspecific variation and prey remains in extant crocodylian feces.- New Mexico Museum of Natural History and Science Bulletin, Albuquerque - NM, vol. 57, p. 65-71.
Moncharmont Zei M. (1950).- Sopra una nuova specie di Eurhinodelphis della Pietra leccese.- Rendiconti dell'Accademia di Scienze Fisiche e Matematiche della Societŕ Nazionale di Scienze, Lettere ed Arti di Napoli, vol. 4, p. 1-11.
Moncharmont Zei M. (1956).- Hesperoina dalpiazi n.gen. et n.sp., Platanistidae, Cetacea, della Pietra leccese.- Memorie dell'Istituto di Geologia e Mineralogia dell'Universitŕ di Padova, vol. 19, p. 1-10.
Muńiz F., Belaústegui Z., Toscano A., Ramirez-Cruzado S. & Gámez Vintaned J.A. (2020).- New ichnospecies of Linichnus Jacobsen & Bromley, 2009.- Ichnos, Boca Raton - FL, vol. 27, no. 3, p. 344-351.
Newell R. (1965).- The role of detritus in the nutrition of two marine deposit feeders, the prosobranch Hydrobia ulvae and the bivalve Macoma balthica.- Proceedings of the Zoological Society of London, London, vol. 144, no. 1, p. 25-45.
Parrish J.D. (1989).- Fish communities of interacting shallow-water habitats in tropical oceanic regions.- Marine Ecology Progress Series, Oldendorf, vol. 58, no. 1-2, p. 143-160.
Pearse J.S. & Giese A.C. (1966).- Food, reproduction and organic constitution of the common Antarctic echinoid Sterechinus neumayeri (Meissner).- The Biological Bulletin, Chicago, vol. 130, no. 3, p. 387-401.
Peri E., Collareta A., Aringhieri G., Caramella D., Foresi L.M. & Bianucci G. (2022).- A new physeteroid cetacean from the Lower Miocene of southern Italy: CT imaging, retrodeformation, systematics and palaeobiology of a sperm whale from the Pietra leccese.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 61, no. 2, p. 187-206.
Peri E., Collareta A. & Bianucci G. (2020).- A new record of Physeteroidea from the upper Miocene of the Pietra leccese (southern Italy): Systematics, paleoecology and taphonomy of a fossil macroraptorial sperm whale.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 126, no. 3, p. 751-769. URL: https://riviste.unimi.it/index.php/RIPS/article/view/14284
Peri E., Collareta A., Insacco G. & Bianucci G. (2019).- An Inticetus-like (Cetacea: Odontoceti) postcanine tooth from the Pietra leccese (Miocene, southeastern Italy) and its palaeobiogeographical implications.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 291, no. 2, p. 221-228.
Peri E., Falkingham P.L., Collareta A. & Bianucci G. (2021).- Biting in the Miocene seas: Estimation of the bite force of the macroraptorial sperm whale Zygophyseter varolai using finite element analysis.- Historical Biology, vol. 34, no. 10, p. 1916-1927. DOI: https://doi.org/10.1080/08912963.2021.1986814
Ragaini L. (1994).- Neogene echinoids from Salento (Apulia, Southern Italy).- Bollettino della Societa Paleontologica Italiana, Modena, spec. vol. 2, p. 287-290.
Reolid M., Santos A. & Mayoral E. (2015).- Grazing activity as taphonomic record of necrobiotic interaction: A case study of a sea turtle carapace from the Upper Jurassic of the Prebetic (south Spain).- Revista Mexicana de Ciencias Geológicas, Mexico City - DC, vol. 32, no. 1, p. 21-28. URL: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1026-87742015000100003
Robertson D.R. (1982).- Fish feces as fish food on a Pacific coral reef.- Marine Ecology Progress Series, Oldendorf, vol. 7, no. 3, p. 253-265.
Rozada L., Allain R., Vullo R., Goedert J., Augier D., Jean A., Marchal J., Peyre de Fabrčgues C., Qvarnström M. & Royo-Torres R. (2021).- A Lower Cretaceous Lagerstätte from France: A taphonomic overview of the Angeac-Charente vertebrate assemblage.- Lethaia, Oslo, vol. 54, no. 2, p. 141-165.
Rummy P., Halaclar K. & Chen H. (2021).- The first record of exceptionally-preserved spiral coprolites from the Tsagan-Tsab formation (Lower Cretaceous), Tatal, western Mongolia.- Scientific Reports, London, vol. 11, no. 1, p. 1-17. DOI: https://doi.org/10.1038/s41598-021-87090-5
Sazima I., Sazima C. & Silva J.M. (2003).- The cetacean offal connection: feces and vomits of spinner dolphins as a food source for reef fishes.- Bulletin of Marine Science, Miami - FL, vol. 72, no. 1, p. 151-160.
Tavani G. (1973).- Ipotesi sulla presenza di grossi frammenti di gneiss e di calcare nella "Pietra leccese" della Puglia.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 80, p. 121-125.
Thulborn R.A. (1991).- Morphology, preservation and palaeobiological significance of dinosaur coprolites.- Palćogeography, Palćoclimatology, Palćoecology, vol. 83, no. 4, p. 341-366.
Twitchett RJ. (1994).- Preparation of a disarticulated Ophthalmosaurus skeleton to retain important taphonomic details.- Geological Curator, London, vol. 6, no. 1, p. 7-10. URL: https://www.geocurator.org/images/resources/geocurator/vol6/geocurator_6_1.pdf
Vigliarolo G. (1891).- Monografia dei Pristis fossili con la descrizione di una nuova specie nel calcare di Lecce.- Atti della Reale Accademia delle Scienze Fisiche e Matematiche di Napoli, vol. 4 (app. no. 32), p. 1-28.
Wilson M.A. (2007).- Macroborings and the evolution of marine bioerosion. In: Miller W. III (ed.), Trace fossils: Concepts, problems, prospects.- Elsevier, Amsterdam, p. 356-367.