◄ Carnets Geol. 22 (19) ►
![]()
Outline:
[1. Introduction]
[2. Study area]
[3. Material and methods]
[4. Results]
[5. Discussion]
[6. Concluding remarks]
and ... [Bibliographic references]
Departamento de Biología y Geología, Física
y Química Inorgánica, ESCET, Universidad Rey Juan Carlos, c/ Tulipán s/n,
28933 Móstoles (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
corresponding author;
Research Center in Historical, Cultural and
Natural Heritage;
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain
Departamento de Biología y Geología, Física
y Química Inorgánica, ESCET, Universidad Rey Juan Carlos, c/ Tulipán s/n,
28933 Móstoles (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Departamento de Ciencias de la Tierra,
Universidad de Huelva, Avda. 3 de marzo, 21071 Huelva (Spain)
Published online in final form (pdf) on December 25, 2022
DOI 10.2110/carnets.2022.2219
![]()
[Editor:
Brian Pratt; technical editor: Bruno R.C. Granier]
![]()
This paper investigates the paleoenvironmental evolution of a long core extracted in a small cove located in the Tinto-Odiel estuary (SW Spain). The inferred reconstruction is supported by sedimentological, geochemical, paleontological data and dating. Seven phases have been identified, with the transit from Late Neogene marine environments to a subrecent freshwater pond and a final anthropic filling. On the basis of these data, this area was flooded during the maximum of the MIS-1 transgression (6.5-5.2 cal. kyr BP), with the temporary presence of a subtidal channel with phanerogam meadows. During this evolution, three geochemical peaks have been detected, which correspond to 1) the first evidence of mining activities (~4.5 cal. kyr BP), 2) an interval of intensive mining (1850-1960) and 3) an industrial period (1966-1985), affected by the dumping of highly polluting waste into this estuary.
• texture;
• historical pollution;
• paleontological record;
• shelf-marsh evolution;
• MIS-1 transgression;
• SW Iberian Peninsula
Abad M., Arroyo M., Ruiz F., González-Regalado M.L., Rodríguez Vidal J., Cáceres L.M., Izquierdo T., Toscano A., Gómez P., Gómez G. & Romero V. (2022).- Miocene-Holocene paleoenvironmental changes in the Tinto River estuary (SW Spain) evidenced by sedimentology, geochemistry and fauna.- Carnets Geol., Madrid, vol. 22, no. 19, p. 825-845.
Mise en évidence de changements paléoenvironnementaux au Miocčne-Holocčne dans l'estuaire du Rio Tinto (Sud-Ouest de l'Espagne) par des données sédimentologiques, géochimiques et faunistiques.- Cet article étudie l'évolution paléoenvironnementale des couches d'un carottage d'une vingtaine de mčtres effectué dans une petite anse de l'estuaire de Tinto-Odiel (Sud-Ouest de l'Espagne). La reconstitution des environnement est étayée par des données sédimentologiques, géochimiques et paléontologiques, ainsi que par des datations absolues. On y a distingué sept phases avec le passage des milieux marins du Néogčne supérieur ŕ un bassin d'eau douce sub-récent et un remplissage anthropique final. D'aprčs ces données, cette zone a été ennoyée lors du maximum de la transgression MIS-1 (6500-5200 ans calibrés AP), avec la présence temporaire d'un chenal infralittoral avec des herbiers ŕ phanérogames. Au cours de cette évolution, trois pics géochimiques ont été détectés ; ils correspondent successivement 1) aux premiers indices d'activités miničres (~4500 ans calibrés AP), 2) ŕ un épisode d'exploitation miničre intensive (1850-1960) et 3) ŕ une période industrielle (1966-1985) caractérisée par le déversement dans cet estuaire de déchets trčs polluants.
• texture ;
• pollution
historique ;
• registre
paléontologique ;
• évolution
de plate-forme ŕ marais ;
• trangression MIS-1 ;
• Sud-Ouest de la Péninsule
ibérique
In this century, numerous investigations were focused the paleoenvironment evolution of coastal environments during the Pleistocene (Sciuto, 2014; Sciuto et al., 2015) and the Holocene (Zaibi et al., 2011; Stephan et al., 2019; Hardage et al., 2021). For this purpose, different proxies have been used, such as sedimentological analyses (Ruiz et al., 2021; Jiwarungrueankul et al., 2022), geomorphological mapping (Maya et al., 2017; McSweeny et al., 2017), the paleontological record (Baichtal et al., 2021; Pereira et al., 2021) or geophysical datasets (Ronchi et al., 2018; Trobec et al., 2018), among others. This growing research is very useful to infer the paleogeographic reconstructions of estuaries (Desruelles et al., 2021), lagoons (Marks et al., 2020), deltas (Ta et al., 2021) or barrier island systems (Schaumann et al., 2021). In addition, they are essential to test sea-level changes during the Pleistocene and Holocene (Ghandour et al., 2021). Molluscs, foraminifera and ostracods are some of the most used groups in these studies. The analysis of their key species or assemblages can identify ocean water cooling/warming (Pereira et al., 2022), salinity variations (Gildeeva et al., 2021), sea-level changes (Williams et al., 2021) or even high-energy events (Matamales-Andreu, 2019).
The southwestern Atlantic coast of Spain is made up by large sandy beaches (>250 km long), only interrupted by the presence of estuarine mouths (Guadiana, Piedras, Tinto-Odiel, Guadalquivir, Guadalete). During the maximum of the MIS-1 transgression, approximately 6.5 cal. kyr BP (Zazo et al., 1994), these estuaries were flooded and a number of large bays and lagoons were formed on a Neogene substrate (Dabrio et al., 2000; Boski et al., 2002; Pozo et al., 2010). The late Holocene evolution of these coastal systems was characterized by the progressive clogging of these estuaries, due to the growth of sandy spits near their mouths and the appearance of large marshes, mudflats and islands in their innermost areas (Ruiz et al., 2010; Cáceres et al., 2018; Caporizzo et al., 2021).
This paper carries out a multidisciplinary analysis of a continuous sediment core extracted in the Tinto-Odiel estuary. The main objectives are: i) the definition and paleoenvironmental interpretation of its sedimentary facies; ii) the reconstruction of its paleoenvironmental evolution; and iii) the identification of macrofaunal and microfaunal assemblages as proxies of the MIS-1 transgression in this area.
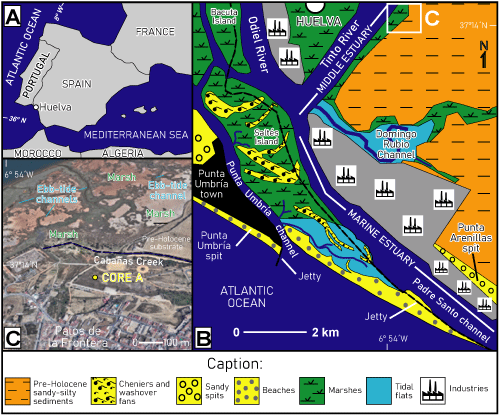
The
Tinto-Odiel River estuary is a 25-km-long incised bar-built
system (Fairbridge, 1980) located on the southwestern Spanish coast (Fig.
1.A-B ![]() ). This coastal environment is characterized by wide tidal flats, salt
marshes and barrier islands, some of which are composed of a complex system of
cheniers and washover fans (Fig. 1.B
). This coastal environment is characterized by wide tidal flats, salt
marshes and barrier islands, some of which are composed of a complex system of
cheniers and washover fans (Fig. 1.B ![]() : Saltés Island; González-Regalado
et al., 2019a). These inner areas are connected with the Atlantic Ocean through
two main channels (Punta Umbría channel and Padre Santo channel) and they are
partially protected by two sandy spits: i) the Punta Umbría spit, to the west;
and ii) the Punta Arenillas spit, developed on the eastern margin and directly
linked with Neogene cliffs, which include some inner small coves (e.g., Fig.
1.B-C
: Saltés Island; González-Regalado
et al., 2019a). These inner areas are connected with the Atlantic Ocean through
two main channels (Punta Umbría channel and Padre Santo channel) and they are
partially protected by two sandy spits: i) the Punta Umbría spit, to the west;
and ii) the Punta Arenillas spit, developed on the eastern margin and directly
linked with Neogene cliffs, which include some inner small coves (e.g., Fig.
1.B-C ![]() : La Fontanilla cove) excavated by the fluvial dynamics during the
Pleistocene. The Holocene estuarine sediments of this estuary were deposited on
Miocene-Pliocene siliciclastic sediments deposited in marine environments (Civis
et al., 1987). This Tertiary
succession is composed of Miocene clays and silts (Gibraleon Clay Formation) and
Pliocene fine sandy silts and sands (Huelva Sand Formation).
: La Fontanilla cove) excavated by the fluvial dynamics during the
Pleistocene. The Holocene estuarine sediments of this estuary were deposited on
Miocene-Pliocene siliciclastic sediments deposited in marine environments (Civis
et al., 1987). This Tertiary
succession is composed of Miocene clays and silts (Gibraleon Clay Formation) and
Pliocene fine sandy silts and sands (Huelva Sand Formation).
|
Figure 1: A-B.
Location and geomorphological map of the Tinto River estuary, with location of
La Fontanilla cove; C. La Fontanilla cove: main geomorphological features and location of
core A. |
Fluvial discharge and tidal regime are the fundamental hydrodynamic processes in the Tinto-Odiel Estuary. The freshwater discharge of the Tinto River is subject to seasonal changes. The highest runoff occurs from December to February, while this discharge extremely scarce during the summer months (~1 hm3/month). The tidal regime is mesotidal (mean range: 2.15 m) and semidiurnal with a low diurnal amplitude (Borrego et al., 1995). Interaction between both fresh and marine waters delimits three sectors (Borrego, 1992): i) Fluvial estuary, dominated by the riverine action; b) Middle Estuary, well mixed during spring tides and partially stratified during neap tides; and iii) Marine Estuary, a tide-dominated zone.
The
drainage basin of the Tinto River includes the Iberian Pyrite Belt, one of the
most important mining-metallurgical provinces in the world. These deposits have
been mined for al least 4,500 years, with two main periods: i) Roman period
(2.1-1.7 kyr BP), with more than 20 Mt extracted (Tornos,
2008); and ii)
Recent period (1850-1960), with almost 300 Mt extracted in more than twenty
mines. This historical contamination has been increased since 1966 due to
the effluents coming from two industrial concentrations located in its margins (Fig.
1.B ![]() ). The final results of these two pollution sources is the presence of
heavily polluted bottom sediments by trace elements such as Cu (up to 3,000 mg
kg-1), Pb (up to 4,860 mg kg-1) or Zn (up to 3,300 mg kg-1)
(Ruiz, 2001; Carro et al.,
2011).
). The final results of these two pollution sources is the presence of
heavily polluted bottom sediments by trace elements such as Cu (up to 3,000 mg
kg-1), Pb (up to 4,860 mg kg-1) or Zn (up to 3,300 mg kg-1)
(Ruiz, 2001; Carro et al.,
2011).
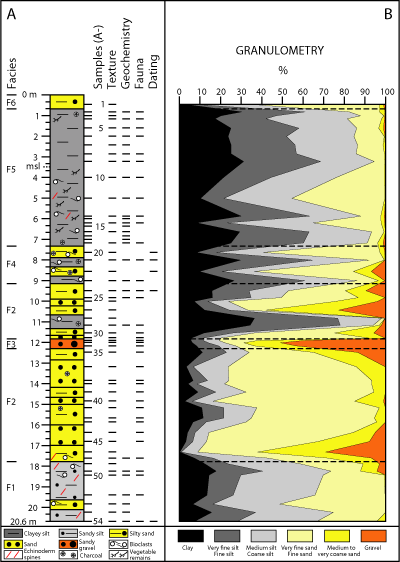
Core
A (Fig. 2.A ![]() : 20.6 m length;
37°13'58"N-6°53'49"W; + 4 m m.s.l.)
was extracted in 2013 near the transition between the alluvial deposits and the
marshes located in the middle estuary of the Tinto River, near the town of Palos
de la Frontera (Fig. 1.C
: 20.6 m length;
37°13'58"N-6°53'49"W; + 4 m m.s.l.)
was extracted in 2013 near the transition between the alluvial deposits and the
marshes located in the middle estuary of the Tinto River, near the town of Palos
de la Frontera (Fig. 1.C ![]() ). It was obtained by usual rotary drilling techniques
with a continuous recovery of sediment and a barrel diameter of 11.6 mm.
Fifty-four samples (2 cm thickness) have been collected for a multidisciplinary
analysis, according to the visual study of the different sedimentary facies and
their boundaries. These selected samples cover all sedimentary facies observed
in core A and the main bioclastic levels. The visual lithological description
has been complemented with the grain-size analysis of fifty samples (Fig.
2.A
). It was obtained by usual rotary drilling techniques
with a continuous recovery of sediment and a barrel diameter of 11.6 mm.
Fifty-four samples (2 cm thickness) have been collected for a multidisciplinary
analysis, according to the visual study of the different sedimentary facies and
their boundaries. These selected samples cover all sedimentary facies observed
in core A and the main bioclastic levels. The visual lithological description
has been complemented with the grain-size analysis of fifty samples (Fig.
2.A ![]() ),
selected according to the vertical distribution of sedimentary facies.
Grain-size distribution was determined by wet sieving for the coarser fractions,
which were sieved with
pressured water and floated using sieves of
63 μm, 125 μm, 250 μm, 500 μm, 1000 μm, and 2000 μm
diameter. Fractions lesser than 63 μm were analyzed with a particle counter
(model Mastersizer-2000) at the CIDERTA center (University of Huelva, Spain).
Na-hexametaphosphate has been used as a dispersing agent.
),
selected according to the vertical distribution of sedimentary facies.
Grain-size distribution was determined by wet sieving for the coarser fractions,
which were sieved with
pressured water and floated using sieves of
63 μm, 125 μm, 250 μm, 500 μm, 1000 μm, and 2000 μm
diameter. Fractions lesser than 63 μm were analyzed with a particle counter
(model Mastersizer-2000) at the CIDERTA center (University of Huelva, Spain).
Na-hexametaphosphate has been used as a dispersing agent.
The
geochemical contents of twenty-nine samples (Fig. 2.A ![]() ) were performed and
certified by MS Analytical, Langley (Canada). In a first step, an acid digestion
(hydrochloric, nitric, perchloric and hydrofluoric acids) was applied to each
sample. Concentrations of five major elements (in %; Al, Ca, Fe, Na, S), four
trace metals (in mg kg-1; As, Cu, Pb, Zn) and phosphorus (in mg kg-1)
were obtained by ICP Atomic Emission Spectrometry, with less than 5% variation
between different replicate samples and a quality control based on more than 30
sample references (e.g., OREAS 904). In these samples, detection limits are: Al
(0.01%), Ca (0.01%), Fe (0.01%), Na (0.01%), S (0.01%), As (0.02 mg kg-1),
Cu (0.2 mg kg-1), Pb (0.5 mg kg-1), Zn (2
mg kg-1) and P (10 mg kg-1).
) were performed and
certified by MS Analytical, Langley (Canada). In a first step, an acid digestion
(hydrochloric, nitric, perchloric and hydrofluoric acids) was applied to each
sample. Concentrations of five major elements (in %; Al, Ca, Fe, Na, S), four
trace metals (in mg kg-1; As, Cu, Pb, Zn) and phosphorus (in mg kg-1)
were obtained by ICP Atomic Emission Spectrometry, with less than 5% variation
between different replicate samples and a quality control based on more than 30
sample references (e.g., OREAS 904). In these samples, detection limits are: Al
(0.01%), Ca (0.01%), Fe (0.01%), Na (0.01%), S (0.01%), As (0.02 mg kg-1),
Cu (0.2 mg kg-1), Pb (0.5 mg kg-1), Zn (2
mg kg-1) and P (10 mg kg-1).
|
Figure 2:
A.
Core A: Log and multidisciplinary sampling; B. Core A: Grain size distribution. |
Thirty-seven
samples were selected for microfaunistic analysis (Fig. 2.A ![]() ). In each sample,
twenty grams of sediment were separated and levigated through a 125 μm mesh
sieve. This sieve was selected for three reasons: a) this sieve retains
practically all of the malacofauna, for whose study sieves with a larger mesh
diameter are usually used (> 250 μm mesh diameter in most cases; Alexandrowicz,
2017; Cristini & De Francesco,
2017); b) the benthic
foraminiferal assemblages picked from this fraction provide useful information
on prevailing environmental conditions and it is useful for an overview of
paleoenvironmental changes in coastal settings (Lo Giudice Capelli & Austin,
2019), two of the main objectives of this paper; and c) this sieve is adequate
for screening of the ostracod fauna and prevent the presence of juveniles
(stages 1-4) of these microcrustaceans, with important difficulties in
identifying them (Danielopol et al., 2002).
). In each sample,
twenty grams of sediment were separated and levigated through a 125 μm mesh
sieve. This sieve was selected for three reasons: a) this sieve retains
practically all of the malacofauna, for whose study sieves with a larger mesh
diameter are usually used (> 250 μm mesh diameter in most cases; Alexandrowicz,
2017; Cristini & De Francesco,
2017); b) the benthic
foraminiferal assemblages picked from this fraction provide useful information
on prevailing environmental conditions and it is useful for an overview of
paleoenvironmental changes in coastal settings (Lo Giudice Capelli & Austin,
2019), two of the main objectives of this paper; and c) this sieve is adequate
for screening of the ostracod fauna and prevent the presence of juveniles
(stages 1-4) of these microcrustaceans, with important difficulties in
identifying them (Danielopol et al., 2002).
The residue was dried in an oven at a constant temperature of 40°C for a period of not less than one day. The microfaunal content (foraminifera and ostracods) was also extracted and determined. Foraminifera were classified according to Loeblich & Tappan (1987), Milker & Schmiedl (2012), and the World Record of Marine Species (WoRMS). Ostracoda were classified according to Ruiz et al. (1997, 2000). This micropaleontological record has been compared with: (i) the assemblages of both groups present in the Neogene formations found in the core A vicinity (González-Regalado & Ruiz, 1996, Ruiz & González-Regalado, 1996), in order to identify reworked specimens; and (ii) the distribution of both groups in the estuaries of southwestern Spain (Ruiz et al., 2000; González-Regalado et al., 2001), a database to recognize paleoenvironmental changes or anthropogenic impacts in this core. These studies have analyzed the foraminiferal and ostracod assemblages of the main sedimentary facies of these estuaries (high salt marsh, low salt marsh, ebb-tide channel, main channel, tidal plain, beach).
Five dates were produced at the National Center of Accelerators (CNA, Seville, Spain), with a subsequent calibration using CALIB version 8.2 and the reservoir effect correction (-108 ± 31 14C yr) calculated by Martins & Soares (2013) for this area. The final results correspond to calibrated ages using 2σ intervals. These calibrated ages have been completed with the comparison between the vertical geochemical variations and the main periods of mining extraction and industrial discharges (see Study Area), in order to obtain additional age references.
Six
main sedimentary facies have been distinguished within core A (Fig. 2.A ![]() ). The
lowermost 2.8 m of this core consist of grayish to greenish massive sandy silts
(F1; Munsell
scale: 10Y 7/2 to 7/6) with intercalations of silty sands (Munsell
scale: 10Y 9/8). Macrofauna is very scarce, with rare fragments of the bivalve Parvamussium
cristatellum (Dautzenberg & Bavay, 1912) and isolated
remains of scaphopods. The upper boundary of F1 (Fig. 2.A
). The
lowermost 2.8 m of this core consist of grayish to greenish massive sandy silts
(F1; Munsell
scale: 10Y 7/2 to 7/6) with intercalations of silty sands (Munsell
scale: 10Y 9/8). Macrofauna is very scarce, with rare fragments of the bivalve Parvamussium
cristatellum (Dautzenberg & Bavay, 1912) and isolated
remains of scaphopods. The upper boundary of F1 (Fig. 2.A ![]() : 17.8 m depth) is
characterized by an erosional discontinuity, which separates it from the
overlying brownish silty sands (F2; 17.8-12.3 m depth and 11.7-9.1 m depth; Munsell
scale: 10Y 3/0) with some muddy intercalations (11.3-10.7 m depth). F2
presents frequent quartzite clasts (up to 2 mm diameter) and some charcoal-rich
layers, while no macrofaunal remains have been observed. F3 (12.3-11.8 m depth)
has an erosive base and it is made up of yellow to whitish quartzite sandy
gravels (Munsell scale: 5Y 8/8) within a silty-clayey matrix (gravel: 55%; sand:
25%; silt+clay: 20%). Macrofauna is absent in this facies.
: 17.8 m depth) is
characterized by an erosional discontinuity, which separates it from the
overlying brownish silty sands (F2; 17.8-12.3 m depth and 11.7-9.1 m depth; Munsell
scale: 10Y 3/0) with some muddy intercalations (11.3-10.7 m depth). F2
presents frequent quartzite clasts (up to 2 mm diameter) and some charcoal-rich
layers, while no macrofaunal remains have been observed. F3 (12.3-11.8 m depth)
has an erosive base and it is made up of yellow to whitish quartzite sandy
gravels (Munsell scale: 5Y 8/8) within a silty-clayey matrix (gravel: 55%; sand:
25%; silt+clay: 20%). Macrofauna is absent in this facies.
F4
(9.1-7.3 m depth) consists of bioclastic greenish silty sands and sandy silts (Munsell
scale: 10Y 3/2) with important percentages of clays (up to 30%). This facies
includes frequent quartzite clasts and fragments of charcoal. Macrofauna is
abundant, with numerous specimens of the gastropods Peringia
ulvae (Pennant, 1877) and Bittium
reticulatum (Mendes da Costa, 1778). The overlying clayey
silts of F5 (7.3-0.7 m depth; Munsell scale: 5Y 8/4) can be divided into two
subfacies: i) subfacies F5a (7.3-4 m depth), characterized by numerous roots and
frequent specimens of Peringia ulvae;
and ii) subfacies F5b (4-0.7 m depth), with very massive sediments, higher silty
percentages and absence of macrofaunal remains (Fig. 2.B ![]() ). The uppermost 0.7 m of
core A are mainly composed of azoic silty sands (F6; sand: 56%; Munsell
scale:
10YR 7/10).
). The uppermost 0.7 m of
core A are mainly composed of azoic silty sands (F6; sand: 56%; Munsell
scale:
10YR 7/10).
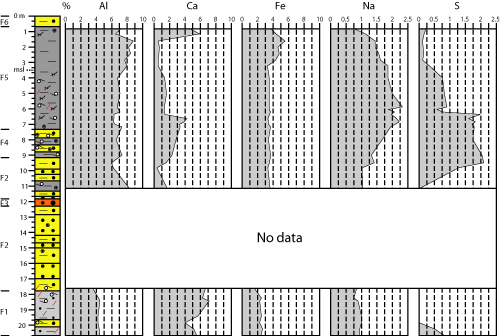
4.2.1. Major elements
F1
is characterized by high percentages of Ca (Fig. 3 ![]() : 4-7%), as well as low to
very low contents of Al (3.5-4.5%), Fe (1.5-2.6%), Na (0.75-1%) and S
(0.04-0.82%). These last four elements increase in F2 (Al: 6-8.2%; Fe: 3.2-3.7%;
Na: 1-1.4%; S: up to 2.1%), while Ca decreases noticeably (0.4-1.7%). Transition
from F2 to F4 is marked by a progressive increase in Ca (up to 3.3%) and Na (up
to 1.9%).
: 4-7%), as well as low to
very low contents of Al (3.5-4.5%), Fe (1.5-2.6%), Na (0.75-1%) and S
(0.04-0.82%). These last four elements increase in F2 (Al: 6-8.2%; Fe: 3.2-3.7%;
Na: 1-1.4%; S: up to 2.1%), while Ca decreases noticeably (0.4-1.7%). Transition
from F2 to F4 is marked by a progressive increase in Ca (up to 3.3%) and Na (up
to 1.9%).
Calcium decreases upward within F5a from more than 4% near its base to less than 1.4% near the limit with F5b (4 m depth). This decrease is very abrupt for S (2% to 0.5%) and milder for Na (2.2% to 1.8%). The F5a-F5b boundary is clearly delimited by a significant increase in Fe (3.5% to 4.7-5.5%) and Al (up to 8.8%), while S decreases in F5b (<0.2%). Except for sulphur, all other major elements decrease near the boundary between F5b and F6.
|
Figure 3: Down-core
variations of major elements in core A. |
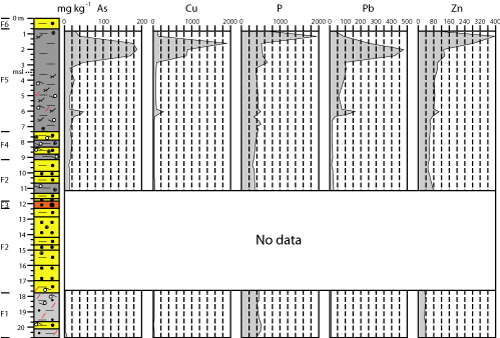
4.2.2. Trace elements
The
vertical variations of the five elements show similar trends (Fig.
4 ![]() ). The
lowest values of As (< 20 mg kg-1), Cu (< 20 mg kg-1),
P (< 500 mg kg-1), Pb (< 20 mg kg-1) and Zn (< 70
mg kg-1 in most samples) were measured in F1, F2 (except in some
muddy intercalations) and F4, although these contents increase upward. A first
peak of four trace elements is observed within F5a at 6.2-6 m depth (As: 42-49
mg kg-1; Cu: 155-252 mg kg-1; Pb: 117-164 mg kg-1;
Zn: 89-110 mg kg-1). Concentrations of these elements decrease
remarkably in the remaining samples of F5a, although the last two elements
slowly increase to the limit between F5a and F5b.
). The
lowest values of As (< 20 mg kg-1), Cu (< 20 mg kg-1),
P (< 500 mg kg-1), Pb (< 20 mg kg-1) and Zn (< 70
mg kg-1 in most samples) were measured in F1, F2 (except in some
muddy intercalations) and F4, although these contents increase upward. A first
peak of four trace elements is observed within F5a at 6.2-6 m depth (As: 42-49
mg kg-1; Cu: 155-252 mg kg-1; Pb: 117-164 mg kg-1;
Zn: 89-110 mg kg-1). Concentrations of these elements decrease
remarkably in the remaining samples of F5a, although the last two elements
slowly increase to the limit between F5a and F5b.
The
highest trace element contents were measured within F5b, with two peaks. The
second geochemical peak of core A (Fig.
4 ![]() : 2.4-1.6 m depth) is characterized by
the highest concentrations of As (170-187 mg kg-1), Cu (853-1893 mg
kg-1) and Pb (253-475 mg kg-1), as well as by high values
of Zn (131-285 mk kg-1). A third peak (1.2 m depth) is located near
the F5a-F5b boundary, with the highest concentrations of P (up to 1940 mg kg-1)
and Zn (up to 394 mg kg-1). In this boundary, all these contents
decrease (As: 27 mg kg-1; Cu: 97 mg kg-1; P: 843 mg kg-1;
Pb: 35 mg kg-1; 85 mg kg-1).
: 2.4-1.6 m depth) is characterized by
the highest concentrations of As (170-187 mg kg-1), Cu (853-1893 mg
kg-1) and Pb (253-475 mg kg-1), as well as by high values
of Zn (131-285 mk kg-1). A third peak (1.2 m depth) is located near
the F5a-F5b boundary, with the highest concentrations of P (up to 1940 mg kg-1)
and Zn (up to 394 mg kg-1). In this boundary, all these contents
decrease (As: 27 mg kg-1; Cu: 97 mg kg-1; P: 843 mg kg-1;
Pb: 35 mg kg-1; 85 mg kg-1).
|
Figure
4: Down-core
variations of trace elements in core A. |
4.3.1. General remarks
The microfossil distribution can be consulted in Table 1. Microfauna is abundant in F1, with numerous individuals of benthic foraminifera and frequent planktonic species of this group (mainly Globigerina bulloides Orbigny, 1826, Globigerinoides obliquus Bolli, 1957, and Orbulina universa Orbigny, 1839). These planktonic species are not found in the remaining facies. In addition, this facies presents numerous spines of echinoderms and some undifferentiated fragments of ostracods. Conversely, F2 is almost devoid of microfauna, which has only been observed near its lower (sample A-47; 17.6 m depth) and upper (sample A-24: 9.5 m depth) limits. No microfossils have been found in F3 and most samples of F5b, while benthic foraminifera are well represented in F4 and, to a lesser extent, in F5a.
4.3.2. Benthic foraminifera
A total of 661 shells of benthic foraminifera belonging to 33 species were extracted from fifteen samples, with more than 76% of the individuals concentrated in F1 and F4 (Table 1). The foraminiferal record of F1 (48-121 individuals/20 g; 7-19 species/sample) is dominated by Ammonia inflata (Seguenza, 1862), Bulimina elongata Orbigny, 1846, Marginulinopsis costata (Batsch, 1791), Nonion faba (Fichtel & Moll, 1798) and some species of Lenticulina. Some of them are also present in the basal sample of F2 (A-47; 17.6 m depth), together with scarce shells of other species also observed in F1, such as Globobulimina auriculata (Bailey, 1894) or Heterolepa bellincionii (Giannini & Tavani, 1960). In the remaining samples of this facies, benthic foraminifera are only observed near the upper boundary with F4 (sample A-24; 9.5 m depth), with very frequent shells of Hansenisca soldanii (Orbigny, 1826) and rare specimens of Ammonia tepida (Cushman, 1926), Haynesina spp., Nonion faba (Fichtel & Moll, 1798) and Trochammina inflata (Montagu, 1808). These microorganisms are absent in F3.
The foraminiferal record of F4 (42-98 individuals/20 g; 5-6 species/sample) is mainly composed of Ammonia tepida (Cushman, 1926) (22-35 individuals/20 g), Haynesina depressula (Walker & Jacob, 1798) and Haynesina germanica (Ehrenberg, 1840), with an increasing number of individuals upward. These species are accompanied by Ammonia beccarii (Linnaeus, 1758) and scarce elphidids. The three first species are the almost only benthic foraminifera observed in F5a (12-38 individuals/20 g; 3-4 species/sample), with a decreasing density towards the top of this subfacies. On the contrary, samples of F5b are characterized by the absence of these microorganisms and the presence of rare individuals of Nonion faba (Fichtel & Moll, 1798), Ortomorphina tenuicostata (Costa, 1853) and Quinqueloculina seminulum (Linnaeus, 1758).
Table 1:
Distribution
of benthic foraminifera and ostracoda in core A.
Tableau 1 :
Distribution des foraminifčres benthiques et des ostracodes dans la carotte A.
| FORAMINIFERA | |||||||||||||||||
| SPECIES/SAMPLES | A-2 | A-3 | A-5 | A-10 | A-11 | A-12 | A-14 | A-19 | A-20 | A-21 | A-23 | A-24 | A-47 | A-49 | A-50 | A-51 | A-54 |
| Ammonia beccarii (LINNAEUS, 1758) | 4 | 9 | 2 | ||||||||||||||
| Ammonia inflata (SEGUENZA, 1862) | 4 | 11 | 12 | 4 | |||||||||||||
| Ammonia tepida (CUSHMAN, 1926) | 8 | 7 | 10 | 20 | 17 | 35 | 25 | 22 | 4 | ||||||||
| Bannerella gibbosa (ORBIGNY, 1826) | 3 | 3 | |||||||||||||||
| Bolivina catanensis (SEGUENZA, 1862) | 3 | 5 | |||||||||||||||
| Bolivina spathulata (WILLIANSON, 1858) | 5 | 10 | |||||||||||||||
| Bulimina elongata ORBIGNY, 1846 | 1 | 11 | 24 | 2 | |||||||||||||
| Cassidulina carinata SILVESTRI, 1896 | 11 | 3 | |||||||||||||||
| Cibicidoides pseudoungerianus (CUSHMAN, 1922) | 1 | ||||||||||||||||
| Cribroelphidium williamsonii (HAYNES, 1973) |
1 | ||||||||||||||||
| Elphidium advenum (CUSHMAN, 1922) | 1 | 1 | 1 | ||||||||||||||
| Elphidium macellum (FICHTEL & MOLL, 1798) |
2 | ||||||||||||||||
| Fursenkoina acuta (CZJZEK, 1848) | 14 | 3 | |||||||||||||||
| Globobulimina auriculata (BAILEY, 1894) | 1 | 5 | 3 | ||||||||||||||
| Hansenisca soldanii (ORBIGNY, 1826) | 14 | 2 | |||||||||||||||
| Heterolepa bellincionii (GIANNINI & TAVANI, 1960) |
1 | 7 | 6 | 1 | |||||||||||||
| Haynesina depressula (WALKER & JACOB, 1798) |
2 | 3 | 8 | 11 | 10 | 29 | 16 | 5 | 3 | ||||||||
| Haynesina germanica (EHRENBERG, 1840) |
2 | 3 | 4 | 7 | 3 | 30 | 24 | 3 | 1 | ||||||||
| Hopkinsina bononiensis (FORNASINI, 1888) |
2 | ||||||||||||||||
| Lenticulina calcar (LINNAEUS, 1758) | 1 | 12 | 4 | 5 | 1 | ||||||||||||
| Lenticulina cultrata (MONTFORT, 1808) | 17 | 4 | 3 | 4 | |||||||||||||
| Lenticulina curvisepta (SEGUENZA, 1880) |
1 | ||||||||||||||||
| Lenticulina inornata (ORBIGNY, 1846) | 4 | 4 | 1 | ||||||||||||||
| Marginulinopsis costata (BATSCH, 1791) | 1 | 9 | 10 | 3 | |||||||||||||
| Melonis pompilioides (FICHTEL & MOLL, 1798) |
2 | ||||||||||||||||
| Nonion faba FICHTEL & MOLL, 1798) | 1 | 3 | 8 | 21 | 2 | ||||||||||||
| Orthomorphina tenuicostata (COSTA, 1853) |
1 | 1 | |||||||||||||||
| Pullenia bulloides (ORBIGNY, 1846) | 7 | 2 | |||||||||||||||
| Quinqueloculina seminulum (LINNAEUS, 1758) |
1 | ||||||||||||||||
| Sahulia neorugosa (THALMANN, 1950) | 7 | 3 | |||||||||||||||
| Trochammina inflata (MONTAGU, 1808) | 3 | 3 | |||||||||||||||
| Vaginulina striatissima SCHRODT, 1890 | 3 | 1 | |||||||||||||||
| Valvulineria bradyana (FORNASINI, 1900) | 3 | ||||||||||||||||
| NUMBER OF INDIVIDUALS/20 g | 3 | 12 | 13 | 22 | 38 | 31 | 98 | 71 | 42 | 28 | 9 | 48 | 121 | 110 | 15 | ||
| NUMBER OF SPECIES | 3 | 3 | 3 | 3 | 3 | 4 | 6 | 6 | 5 | 6 | 6 | 7 | 19 | 18 | 7 | ||
| OSTRACODA | |||||||||||||||||
| SPECIES/SAMPLES | A-2 | A-3 | A-5 | A-10 | A-11 | A-12 | A-14 | A-19 | A-20 | A-21 | A-23 | A-24 | A-47 | A-49 | A-50 | A-51 | A-54 |
| Cyprideis torosa (JONES, 1850) | 1 | ||||||||||||||||
| Cypridopsis vidua (O.F. MULLER, 1776) | 17 | ||||||||||||||||
| Cytherois fischeri (SARS, 1866) | 1 | 3 | |||||||||||||||
| Ilyocypris gibba (RAMDOHR, 1808) | 1 | ||||||||||||||||
| Leptocythere castanea (SARS, 1866) | 3 | ||||||||||||||||
| Loxoconcha elliptica BRADY, 1867 | 1 | 3 | 61 | ||||||||||||||
| NUMBER OF INDIVIDUALS/20 g | 18 | 1 | 1 | 4 | 67 | ||||||||||||
| NUMBER OF SPECIES | 2 | 1 | 1 | 2 | 3 | ||||||||||||
| FACIES | F5b | F5a | F4 | F2 | F1 | ||||||||||||
4.3.3. Ostracoda
This group is represented by six species and 91 valves and carapaces, although no specimens were found in F2 and F3. This group is abundant in the lower-middle samples of F4 (9-8 m depth), with numerous adults and moults of Loxoconcha elliptica Brady, 1867, together with rare valves of Cytherois fischeri (Sars, 1866) and Leptocythere castanea (Sars, 1866). An isolated valve of Loxoconcha elliptica Brady, 1867, was extracted in the uppermost sample of F5a, while Cypridopsis vidua (O.F. Müller, 1776) is the main species of the uppermost sample of F5b (sample A-2; 17 individuals/20 g), together with isolated valves of Ilyocypris gibba (Ramdohr, 1808) and Cyprideis torosa (Jones, 1850).
Five dates have been obtained from F1, F2 and F4 (Table 2). According to these data, a first approximation to the ages of the different facies can be made: i) F1 was deposited more than 50,000 years ago; ii) the ages of F2 and F3 are older than ~6.31 cal. kyr BP; iii) the bioclastic deposits of F4 were deposited between ~6.2 cal. kyr BP and ~5.4 cal. kyr BP; and iv) the ages of the uppermost facies (F5a, F5b and F6) are younger than 5.4 cal. kyr BP.
Table
2:
Radiocarbon
database of core A.
Tableau 2 :
Datations de la carotte A.
| Laboratory code | Sample material | ∂ 13C% |
14C age (yr BP) |
Calibrated yr BP/ IntCal20/Marine20 (2σ) |
Mean calibrated age (kyr BP) |
| CNA-4277 | Organic matter | -26.7 | 6407 ± 32 | 5470–5325 | 5.4 |
| CNA-4278 | Organic matter | -26.2 | 6993 ± 33 | 5980–5785 | 5.88 |
| CNA-4279 | Shell | -0.4 | 5690 ± 30 | 6190−5990 | 6.09 |
| CNA-4280 | Organic matter | -28.0 | 7437 ± 33 | 6390–6240 | 6.31 |
| CNA-4283 | Shell | -1.2 | Out of range | Out of range | Out of range |
5.1.1. Molluscs
F1 is characterized by fragmented valves of the bivalve Parvamussium cristatellum (Dautzenberg & Bavay, 1912), a highly mobile, epifaunal species (Danise et al., 2010). It was very common [as Amussium cristatum (Bronn, 1827)] in the Miocene silty-clayey sediments of the Gibraleon Clay Formation (Civis et al., 1987), deposited from slope to shelf paleoenvironments (González-Regalado et al., 2019c). It has been also found in Pliocene monospecific-paucispecific beds of southeastern Spain formed in mesotrophic mid-shelf settings (Aguirre, 2017), although it has been cited in Miocene and Pleistocene shallower paleoenvironments of Italy (Bernasconi & Robba, 1993).
The
gastropods Peringia ulvae (Pennant,
1877) (Fig. 5.A-B ![]() ) and Bittium reticulatum (Mendes da Costa, 1778)
(Fig. 5.C
) and Bittium reticulatum (Mendes da Costa, 1778)
(Fig. 5.C ![]() ) are
very abundant in F4. The first species is frequent in the inner, silty-sandy
mesolittoral areas of estuaries and marshes with low hydrodynamics and important
fluvial inputs (Gómez, 2017). It is
specially adapted to these changing conditions and it crawl about on the surface
of the mud then the burrow for several hours during low tide (Newell,
2009). This species has been mainly
cited in Pleistocene to Holocene sediments around Europe (Gofas et
al., 2001; Danukalova et al., 2015).
) are
very abundant in F4. The first species is frequent in the inner, silty-sandy
mesolittoral areas of estuaries and marshes with low hydrodynamics and important
fluvial inputs (Gómez, 2017). It is
specially adapted to these changing conditions and it crawl about on the surface
of the mud then the burrow for several hours during low tide (Newell,
2009). This species has been mainly
cited in Pleistocene to Holocene sediments around Europe (Gofas et
al., 2001; Danukalova et al., 2015).
Both species coexist in mesolittoral phanerogam meadows or in its vicinity, since Bittium reticulatum (Mendes da Costa, 1778) feeds on the organic matter that covers the beam of the leaves and the soil of these meadows (Borja, 1986; Luque & Templado, 2004). In Spain, this species has been found present in Miocene to Holocene sediments (González Delgado, 1986; Vázquez et al., 1990; González-Regalado et al., 2019b).
|
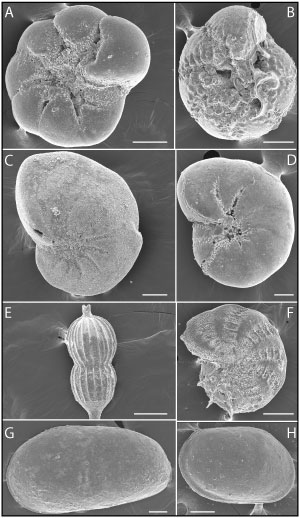
Figure
5: Gastropods. A-B: Bittium reticulatum (Mendes da Costa,
1778); C: Peringia ulvae (Pennant, 1877). Scale: 1 cm. |
5.1.2. Benthic foraminifera
Ammonia
inflata (Seguenza, 1862), Bulimina elongata Orbigny, 1846,
Marginulinopsis costata
(Batsch, 1791) and
Nonion faba (Fichtel & Moll, 1798)
(Fig. 6.C ![]() ) are the
most abundant species of F1. Ammonia inflata (Seguenza, 1862) is a temporary motile
epifaunal species (morphotype B sensu Langer, 1993) that prefers shallow marine (paleo-)environments (Colom,
1974; Mateu-Vicens
et al., 2014). In southern Spain, it
is common in Miocene silty-clayey sediments and Pliocene sandy silts (González-Regalado,
1989; Corbí, 2010). Bulimina
elongata Orbigny, 1846, and Nonion
faba (Fichtel & Moll, 1798) are currently found together
in the outer inner shelf (12-40 m depth) of the southwestern Spanish-Portuguese
littoral (Mendes et al., 2004),
although these species have been cited from deeper Miocene paleoenvironments (up
to 80-100 m depth) of southeastern Germany (Pipper & Reichenbaker,
2009) and Morocco (Tulbure et al., 2017) and recent sediments from the Alboran Sea (García Sanz & Usera,
2015). Bulimina elongata Orbigny,
1846, is common in low-oxygen (paleo-)environments (cf. Rögl & Spezzaferri,
2003), while Nonion faba (Fichtel
& Moll, 1798) is considered intermediate infaunal and associated with eutrophic
environments and high organic matter of low quality from terrestrial runoff (see
García Gallardo, 2017, and references therein). Marginulinopsis
costata (Batsch, 1791) is an infaunal species frequently cited in
circalittoral and slope (paleo-)environments under dysoxic conditions (Corbí,
2010; Amakrane et al., 2016; González-Regalado
et al., 2019b), mainly in soft bottoms
enriched in organic matter (Sciuto & Baldanza,
2020). It has
been cited in Late Oligocene to Recent sediments (e.g., Carboni & Palagi,
1998).
) are the
most abundant species of F1. Ammonia inflata (Seguenza, 1862) is a temporary motile
epifaunal species (morphotype B sensu Langer, 1993) that prefers shallow marine (paleo-)environments (Colom,
1974; Mateu-Vicens
et al., 2014). In southern Spain, it
is common in Miocene silty-clayey sediments and Pliocene sandy silts (González-Regalado,
1989; Corbí, 2010). Bulimina
elongata Orbigny, 1846, and Nonion
faba (Fichtel & Moll, 1798) are currently found together
in the outer inner shelf (12-40 m depth) of the southwestern Spanish-Portuguese
littoral (Mendes et al., 2004),
although these species have been cited from deeper Miocene paleoenvironments (up
to 80-100 m depth) of southeastern Germany (Pipper & Reichenbaker,
2009) and Morocco (Tulbure et al., 2017) and recent sediments from the Alboran Sea (García Sanz & Usera,
2015). Bulimina elongata Orbigny,
1846, is common in low-oxygen (paleo-)environments (cf. Rögl & Spezzaferri,
2003), while Nonion faba (Fichtel
& Moll, 1798) is considered intermediate infaunal and associated with eutrophic
environments and high organic matter of low quality from terrestrial runoff (see
García Gallardo, 2017, and references therein). Marginulinopsis
costata (Batsch, 1791) is an infaunal species frequently cited in
circalittoral and slope (paleo-)environments under dysoxic conditions (Corbí,
2010; Amakrane et al., 2016; González-Regalado
et al., 2019b), mainly in soft bottoms
enriched in organic matter (Sciuto & Baldanza,
2020). It has
been cited in Late Oligocene to Recent sediments (e.g., Carboni & Palagi,
1998).
|
Figure
6: Microfauna.
Foraminifera. A: Ammonia tepida (Cushman, 1926) (ventral view);
B:
Ammonia
beccarii (Linnaeus, 1758) (ventral view); C: Nonion
faba (Fichtel & Moll, 1798);
D: Haynesina
germanica (Ehrenberg, 1840); E: Amphycorina
scalaris (Batsch, 1791); F: Elphidium
advenum (Cushman, 1922); Ostracoda. G: Left valve of Cyprideis
torosa (Jones, 1850); H:
Right valve of Loxoconcha elliptica Brady, 1867. Scale: 100 μm. |
Ammonia
tepida (Cushman, 1926) (Fig. 6.A ![]() ), Haynesina depressula (Walker & Jacob, 1798) and
Haynesina
germanica (Ehrenberg, 1840) (Fig.
6.D
), Haynesina depressula (Walker & Jacob, 1798) and
Haynesina
germanica (Ehrenberg, 1840) (Fig.
6.D ![]() ) are well represented in both
F4 and F5a. Ammonia tepida (Cushman, 1926) is an endopelic species,
living in intertidal mudflats of brackish environments, such as estuaries or
lagoons (Debenay et al., 1998; Ruiz
et al., 2012; Wukovits et al., 2018; Pascual et
al., 2019). In southern Europe, this species was described in Miocene to
Recent sediments (González Delgado et al., 1986; González-Regalado,
1989; Bossio
et al., 2003-2004).
) are well represented in both
F4 and F5a. Ammonia tepida (Cushman, 1926) is an endopelic species,
living in intertidal mudflats of brackish environments, such as estuaries or
lagoons (Debenay et al., 1998; Ruiz
et al., 2012; Wukovits et al., 2018; Pascual et
al., 2019). In southern Europe, this species was described in Miocene to
Recent sediments (González Delgado et al., 1986; González-Regalado,
1989; Bossio
et al., 2003-2004).
Haynesina depressula (Walker & Jacob, 1798) is one of the most common paralic-shallow water marine species, which tolerates polyhaline waters and stressed environments (see Aiello et al., 2018, and references therein). It is well represented in marshes, intertidal mudflats or bays with freshwater inputs (Cearreta, 1989; Pascual et al., 2002; Calvo & Langer, 2010). It was found in Middle Miocene sediments from eastern Spain (García-Sanz et al., 2019), as well as in Pliocene-Pleistocene sections from Italy (Di Bella et al., 2000) and Pliocene clays from Crete (Drinia et al., 2005).
Haynesina germanica (Ehrenberg, 1840) is a common, shallow-water, benthic foraminifer that has been reported from recent estuaries, bays, salt marshes, and mudflats from around the globe (see Goldstein & Richardson, 2018, and references therein). These environments have large diurnal salinity variations (0-35‰), a temperature range of 0-32şC and depth from intertidal to 20 m (Murray, 2006). It has been collected in Tortonian to Holocene sediments from America, Europe and Asia (Gibson, 1994; Usera et al., 2000; Melis et al., 2015).
5.1.3. Ostracoda
Cypridopsis vidua (O.F. Müller, 1776) was only found in the uppermost sample of F5b (0.8 m depth). It is a freshwater, phytophilous species (Roca & Danielopol, 1991), very frequent in ponds displaying high vegetation cover (Martínez et al., 2016b), springs (Martínez et al., 2016a), lakes and reservoirs (Kulkoyluoglu, 2003), among other freshwater (paleo-)environments. In southern Europe, this species has been collected in fossil sediments from the Upper Miocene to the Holocene (Bekkali et al., 2003; Ruiz et al., 2004a; Sciuto et al., 2015).
Cytherois
fischeri (Sars, 1866), Leptocythere
castanea (Sars, 1866) and Loxoconcha
elliptica Brady, 1867 (Fig. 6.H ![]() ) are the only ostracod species
extracted from F4. Cytherois fischeri (Sars,
1866) is an euryhaline species (Athersuch et al.,
1990), frequent in silty sand flats, low marshes or lagoons
(Zaibi et al.,
2011; Martínez et al., 2013; Altinsacli
et al., 2015). In recent sediment of southwestern Spain, it was found from the
middle estuaries to the innermost shelf (Ruiz et al.,
1997; Mattos et al.,
2016). This species has been observed in Pleistocene and
Holocene cores and sections from Spain, France and Greece (Ruiz et
al., 2007; Devillers et al., 2019; Papadopoulou et al.,
2019).
) are the only ostracod species
extracted from F4. Cytherois fischeri (Sars,
1866) is an euryhaline species (Athersuch et al.,
1990), frequent in silty sand flats, low marshes or lagoons
(Zaibi et al.,
2011; Martínez et al., 2013; Altinsacli
et al., 2015). In recent sediment of southwestern Spain, it was found from the
middle estuaries to the innermost shelf (Ruiz et al.,
1997; Mattos et al.,
2016). This species has been observed in Pleistocene and
Holocene cores and sections from Spain, France and Greece (Ruiz et
al., 2007; Devillers et al., 2019; Papadopoulou et al.,
2019).
Leptocythere castanea (Sars, 1866) is very common in inner estuarine and salt marsh environments (Athersuch et al., 1990), with periodic supply of seawater (Martín-Rubio et al., 2006) and a highly variable salinity (1‰-32‰; Pascual et al., 1999). This species has been found in Pleistocene and Holocene cores and sections from southern Spain (Ruiz et al., 2002; Torres et al., 2003). Loxoconcha elliptica Brady, 1867, is a euryhaline species, very abundant in lagoons, estuaries, marshes or ebb-tide channels from the Baltic sea to the Turkey (Martín-Rubio et al., 2004; Bassler-Veit et al., 2013; Pint & Frenzel, 2017). This species has been cited in Pleistocene and Holocene sediments from Germany, Spain, Italy, Tunisia and Turkey (Ruiz et al., 2004b; Viehberg et al., 2008; Salihoglu et al., 2018; Catanzariti & Da Prato, 2020).
A first, high sedimentation rate (SR: 2.03 cm/yr) can be estimated for F4 between samples A-24 (9 m depth; ~6.09 cal. kyr BP) and A-20 (7.6 m depth; ~5.4 cal. kyr BP). If this SR is extrapolated to the total thickness of F4, this facies would have been deposited between ~6.09 cal. kyr BP and ~5.2 cal. kyr BP.
In the uppermost meters of core A, three geochemical peaks has been observed at 6.2-6 m depth, 2.4-1.6 m depth and 1.2 m depth, respectively (see section B.2. Trace elements). The first peak would correspond to the first evidence of mining activity in this region (~4.5 cal. kyr BP; Rothenberg & Blanco Freijero, 1980), according to: i) the age of sample A-20 (7.6 m depth; ~5.4 cal. kyr BP), close to the F4-F5a boundary (7.3 m depth); and ii) the depth of this geochemical peak (~4.2 m depth) detected in other cores of the Tinto estuary (Arroyo et al., 2021b). According to these data and the mean calibrated age of sample A-20 (7.6 m depth; ~5.4 cal. kyr BP), the estimated SR between ~5.4 cal. kyr BP and ~4.5 cal. kyr BP (6.2 m depth) was around 1.55 cm/year. This decreasing trend has been observed in other cores of this estuary after 6.5 cal. kyr BP until the present (Lario et al., 2002).
The
second peak starts at 2.4 m depth and leads to a notable rise (even over an
order of magnitude) in the concentrations of As, Cu and Pb, which remain very
high in the following 0.8 m above (Fig.
4 ![]() ). These very high contents are
attributed to the increase in intensive mining in the Iberian Pyrite Belt,
especially from 1850 to 1980. In the middle estuary of the Tinto river, this
peak has been observed in other nearby cores at 2 m depth (Arroyo et
al., 2021b). Consequently, a sedimentation rate (~0.84 cm/yr) can be
estimated between ~4.5 cal. kyr BP (6.2 m depth) and 1850 (~2.4 m depth), which
confirms the decreasing trend of SR mentioned above. According to this estimated
SR, the calibrated age of the F5a-F5b boundary (4 m depth) is around 1.9 cal.
kyr BP.
). These very high contents are
attributed to the increase in intensive mining in the Iberian Pyrite Belt,
especially from 1850 to 1980. In the middle estuary of the Tinto river, this
peak has been observed in other nearby cores at 2 m depth (Arroyo et
al., 2021b). Consequently, a sedimentation rate (~0.84 cm/yr) can be
estimated between ~4.5 cal. kyr BP (6.2 m depth) and 1850 (~2.4 m depth), which
confirms the decreasing trend of SR mentioned above. According to this estimated
SR, the calibrated age of the F5a-F5b boundary (4 m depth) is around 1.9 cal.
kyr BP.
The
third peak (1.2 m depth) is characterized by the highest concentrations of P and
Zn. This peak would be due to the wastes dumped by the industrial concentrations
located on the banks of the estuary (Fig. 1.B ![]() ), especially from 1966 to 1985 (Ruiz
et al., 2004b). These concentrations
include the largest complex in the European Union producing phosphoric acid, as
well as Zn-rich pyrite processing plants, which explain the high contents of
these elements. Since 1985, this zone is subject of a Corrective Plan for
Control of Industrial Waste Disposal, with an improvement in environmental
conditions that is reflected in the lower levels of contamination near the limit
between F5b and F6 (Fig.
4
), especially from 1966 to 1985 (Ruiz
et al., 2004b). These concentrations
include the largest complex in the European Union producing phosphoric acid, as
well as Zn-rich pyrite processing plants, which explain the high contents of
these elements. Since 1985, this zone is subject of a Corrective Plan for
Control of Industrial Waste Disposal, with an improvement in environmental
conditions that is reflected in the lower levels of contamination near the limit
between F5b and F6 (Fig.
4 ![]() ).
).
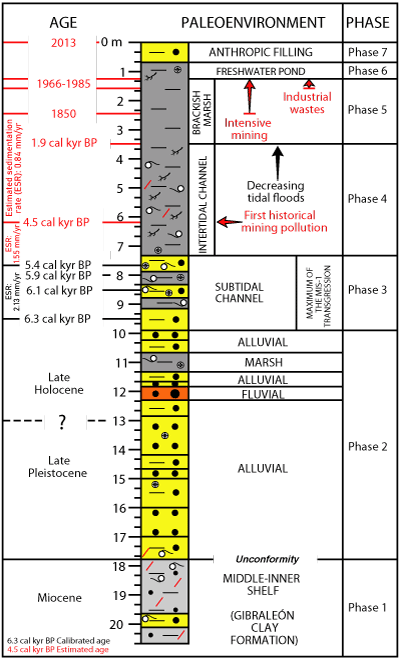
The
integrated analysis of core A (sedimentology, geochemistry, paleontology, dating)
allows to separate seven phases in the paleoenvironmental evolution of core A (Fig.
7 ![]() ).
).
Phase 1 (F1; Miocene). The sedimentological and paleontological features of F1 are very similar to those observed in cores and surface sections that include the upper part of the Gibraleón Clay Formation. The main features of these sediments are: i) dominance of predominance of massive silts and sandy silts; ii) frequent presence of the bivalve Parvamussium cristatellum (Dautzenberg & Bavay, 1912); iii) a foraminiferal assemblage dominated by Ammonia spp., Bulimina elongata Orbigny, 1846 and Nonion faba (Fichtel & Moll, 1798), very frequent in middle-inner shelf (paleo-)environments; and iv) a Late Messinian age, according to their magnetostratigraphic chronology (Civis et al., 1987; García, 2002; Pérez-Asensio et al., 2012; Larrasoańa et al., 2014).
Phase 2 (F2-F3; Late Pleistocene-6.1 cal. kyr BP). The F1-F2 boundary (17.8 m depth) is characterized by a clear unconformity, indicating a partial erosion of F1 as attested by the presence of F1 benthic foraminifera within the basal sample of F2 (Table 1). In this area, the boundary between the Gibraleon Clay Formation and the overlying Huelva Sand Formation was located around +7 m m.s.l. in surface sections (Arroyo et al., 2021a) and consequently about 25 m of F1 were eroded during this new phase. It is characterized by alluvial sands (F2) and fluvial gravels (F3) with very scarce fauna or absence of macrofaunal remains, also detected in other cores of this estuary and arranged on a Neogene substrate (Borrego et al., 1999; Arroyo et al., 2021a, 2021b). These coarser sediments would begin to be deposited during the Late Pleistocene-Early Holocene, according to the dating of other cores and due to the domain of fluvial dynamics in this estuary during this period (Dabrio et al., 2000). The geochemical analysis of these two facies has been studied by Arroyo et al. (2021a, 2021b) in other cores from this estuary and they show very low values of all the metals studied here, close to the regional background (Ruiz et al., 1997).
Phase 3 (F4; 6.1-5.2 cal. kyr BP). The abundant macrofaunal and microfaunal record of F4 points to a subtidal brackish paleoenvironment during this phase (see section 5.A), with the presence of phanerogam meadows with abundant individuals of gastropods (Bittium, Peringia) and frequent foraminifera (Ammonia, Haynesina) and ostracods (mainly Loxoconcha) in the bottom sediments (Borja, 1986; Luque & Templado, 2004; Wukovits et al., 2018; Pascual et al., 2019). This phase coincided with the maximum of the MIS-1 transgression in this area (6.5-5 cal. kyr BP; Zazo et al., 1994), which was invaded by the sea with the creation of cliffs formed by Neogene formations (Dabrio et al., 2000).
Phase 4 (F5a; 5.2 cal. kyr BP-1.9 cal. kyr BP). This phase is characterized by clayey-silty sediments, frequent specimens of Peringia ulvae (Pennant, 1877) and a foraminiferal assemblage dominated by Ammonia tepida (Cushman, 1926), Haynesina depressula (Walker & Jacob, 1798) and Haynesina germanica (Ehrenberg, 1840). These features are similar to those observed in estuarine intertidal channels of southwestern Spain with low hydrodynamic gradient (González-Regalado et al., 2001; Gómez, 2017).
During
this phase, the start of mining activities at ~4.5 cal. kyr BP (sample A-14; 6.2
m depth) caused a small pollution of this estuary, mainly by As, Cu, Pb and Zn (Fig.
4 ![]() ). A subsequent decrease in foraminiferal abundance was observed towards
the top of this subfacies, as in other polluted estuaries (Alve,
1995; Nagendra
& Reddy, 2019). This decrease may also be related to an increasing
clogging and lower tidal inputs (Murray, 2006; Cearreta et
al., 2006).
). A subsequent decrease in foraminiferal abundance was observed towards
the top of this subfacies, as in other polluted estuaries (Alve,
1995; Nagendra
& Reddy, 2019). This decrease may also be related to an increasing
clogging and lower tidal inputs (Murray, 2006; Cearreta et
al., 2006).
Phase 5 (F5b; 1.9 cal. kyr BP-1985). Transit to F5b (4 m depth; ~1.9
cal kyr BP) is marked by: i) a further decrease in grain size (Fig.
2 ![]() ); ii)
lower concentrations of Ca, Na and S (Fig. 3
); ii)
lower concentrations of Ca, Na and S (Fig. 3 ![]() ); iii) a small increase in Pb
content (Fig.
4
); iii) a small increase in Pb
content (Fig.
4 ![]() ); iv) the disappearance of macrofauna, foraminifera and spines
of echinoderms (Figs. 2.A
); iv) the disappearance of macrofauna, foraminifera and spines
of echinoderms (Figs. 2.A ![]() - 3
- 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() ). All these features point to an increasing
confinement of this area and a transition to marsh paleoenvironments, probably
about mean sea level (González-Regalado et al.,
2001; Ruiz et al.,
2007). The presence of fossil foraminifera (Table 1: sample A-3) are attributed
to the erosion of the Neogene formations that make up the slopes near core A.
). All these features point to an increasing
confinement of this area and a transition to marsh paleoenvironments, probably
about mean sea level (González-Regalado et al.,
2001; Ruiz et al.,
2007). The presence of fossil foraminifera (Table 1: sample A-3) are attributed
to the erosion of the Neogene formations that make up the slopes near core A.
The
higher Pb content could be due to pollution produce by Roman mining activities,
very intense around 1900 cal. kyr BP (Arroyo et al.,
2021b). In addition, this phase
includes the highest concentrations of heavy metals, derived from industrial
effluents discharged into this estuary between 1966 and 1985 (Fig.
4 ![]() ; see
previous chapter).
; see
previous chapter).
|
Figure 7: Paleoenvironmental reconstruction of core A. |
Phase 6 (uppermost part of F5b: subrecent). This phase is characterized by lower concentrations of major and trace elements and the presence of freshwater ostracods [mainly Cypridopsis vidua (O.F. Müller, 1776)]. This phase would end the regression that started at the end of the MIS-1 transgression, with a transition from a subtidal channel (F4) to this inferred freshwater pond (Fernandes et al., 2010; Martínez et al., 2015), without connection with the tidal dynamics.
Phase 7 (F6: last decades). The uppermost 0.7 m constitute the anthropic filling carried out in recent decades in this area, with the construction of a model airplane circuit and a soccer field.
The integrated analysis (sedimentology, geochemistry, paleontology, dating) of a long core collected in the middle estuary of the Tinto-Odiel estuary (SW Spain) reveals a complex evolution from marine to freshwater paleoenvironments. The lowermost meters of this core are composed of Miocene sandy silts, which were deposited on a middle-inner shelf. A significant thickness of these Neogene sediments were eroded during the Pleistocene-Early Holocene due the dominant fluvial dynamics during this period, with the deposit of fluvial and alluvial sands and gravels. This middle estuary was flooded during the maximum of the MIS-1 transgression (~6.5-5.2 cal kyr BP), with the implantation of seagrass meadows in a subtidal channel with typical associations of gastropods (Peringia, Bittium), foraminifera (Ammonia, Haynesina) and ostracods (mainly Loxoconcha). The subsequent regressive evolution included the transit from an intertidal channel to mudflats and finally to a freshwater pond, with a recent anthropic filling in the uppermost meter of the core. This paper also includes one of the oldest historical evidence of mining pollution (~4.5 cal. kyr BP), as well as two strong polluted levels derived from an intensive mining interval during the 19th and 20th centuries and a recent industrial period.
This work has been mainly financed by the Palos de la Frontera Council. It is also carried out through the following projects: a) DGYCIT project CTM2006-06722/MAR; b) DGYCIT project CGL2006-01412; c) "Roman cities of the Baetica. CORPVS VRBIVM BAETICARVM (I) (CUB)" (Andalusian Gouvernement); d) "From the Atlantic to the Tyrrhenian. The Hispanic ports and their commercial relations with Ostia Antica. DEATLANTIR II - HAR2017-89154-P - (Plan Nacional de I+D+i)"; and e) FEDER 2014-2020 project UHU-1260298. Other funds have come from Andalusian Government (groups HUM-132, RNM-238, and RNM-293). It is a contribution to the Research Center in Historical, Cultural and Natural Heritage (CIPHCN) of the University of Huelva.
The authors would like to acknowledge the constructive comments of the anonymous reviewers and the advice of the editors, Dr. Brian Pratt and Dr. Bruno Granier, which have contributed to a significant improvement of this paper.
Aguirre J. (2017).- Amusium cristatum shell beds in the lower Pliocene deposits of Almería-Níjar Basin (SE Spain).- Spanish Journal of Palaeontology, Madrid, vol. 32, no. 1, p. 3-16. DOI: https://doi.org/10.7203/sjp.32.1.16985
Aiello G., Barra D., Parisi R., Isaia R. & Marturano A. (2018).- Holocene benthic foraminiferal and ostracod assemblages in a paleo- hydrothermal vent system of Campi Flegrei (Campania, South Italy).- Palaeontologia Electronica, Amherst - MA, article no. 21.3.41A, 71 p. DOI: https://doi.org/10.26879/835
Alexandrowicz W.P. (2017).- Malacofauna of the Holocene tufa in the valleys of the Ociemny Stream (Pieniny Mts, southern Poland).- Geology, Geophysics & Environment, Kraków, vol. 43, p. 5-18. URL: https://journals.agh.edu.pl/geol/article/view/2373
Altinsacli S., Percin-Pacal F. & Altinsacli S. (2015).- Diversity, spatiotemporal distribution, abundance, species composition and habitat preferences of Ostracoda in Akbük and Akdeniz coastal mesosaline lagoons (Muğla, the South Aegean Region, Turkey).- Oceanological and Hydrobiological Studies, Warsaw, vol. 44, p. 206-222. DOI: https://doi.org/10.1515/ohs-2015-0020
Alve E. (1995).- Benthic foraminiferal response to estuarine pollution: A review.- Journal of Foraminiferal Research, Lawrence - KS, vol. 25, p. 190-203.
Amakrane J., Azdimousa A., Rezqi H., El Hammouti K., El Ouhabi M. & Fagel N. (2016).- Paleoenvironment and sequence stratigraphy of the late Miocene from the Guercif basin (Northeastern of Morocco).- Bulletin de l'Institut Scientifique, Rabat, Section Sciences de la Terre, vol. 38, p. 95-110.
Arroyo, M., Ruiz F., Campos J.M., Bermejo J., González-Regalado M.L., Rodríguez Vidal J., Cáceres L.M., Olías M., Abad M., Izquierdo T., Gómez, P., Toscano A., Romero V. & Gómez G. (2021a).- Where did Christopher Columbus start? The estuarine scenario of a historical date.- Estuarine, Coastal & Shelf Science, vol. 250, article no. 107162, 12 p.
Arroyo M., Ruiz, F., González-Regalado M.L., Rodríguez Vidal J., Cáceres L.M., Olías M., Campos J.M., Fernández L., Abad M., Izquierdo T., Gómez P., Toscano A., Romero V. & Gómez G. (2021b).- Natural and anthropic pollution episodes during the Late Holocene evolution of the Tinto River estuary (SW Spain).- Scientia Marina, vol. 85, no. 2, p. 113-123. DOI: https://doi.org/10.3989/scimar.05131.011
Athersuch J., Horne D.J. & Whittaker J.E. (1990).- Marine and brackish water ostracods.- Synopses of the British fauna, Leiden, vol. 43, 343 p.
Baichtal J.F., Lesnek A.J., Carlson R.J., Smuck N.S., Smith J.L., Landwehr D.J. & Briner J.P. (2021).- Late Pleistocene and early Holocene sea-level history and glacial retreat interpreted from shell-bearing marine deposits of southeastern Alaska, USA.- Geosphere, Tulsa - OK, vol. 17, no. 6, p. 1590-1615. DOI: https://doi.org/10.1130/GES02359.1
Bassler-Veit B., Barut I.F., Meric E., Avsar N., Nazik A., Kapan-Yesilyurt S. & Yildiz A. (2013).- Distribution of microflora, meiofauna, and macrofauna assemblages in the hypersaline environment of northeastern Aegean Sea coasts.- Journal of Coastal Research, Fort Lauderdale - FL, vol. 29, no. 4, p. 883-898.
Bekkali R, Nachite D., Rodríguez-Lázaro J. & Martín-Rubio M. (2003).- Los ostrácodos lacustres del Mioceno terminal de Salar (Margen NO de la cuenca de Granada, Espańa).- Geogaceta, Madrid, vol. 34, p. 91-94. URL: https://sge.usal.es/archivos/geogacetas/Geo34/Art23.pdf
Bernasconi M.P. & Robba E. (1993).- Molluscan paleoecology and sedimentogical features: An integrated approach from the Miocene Meduna section, northern Italy.- Palćogeography, Palćoclimatology, Palćoecology, vol. 100, p. 267-290.
Borja A. (1986).- Biologia y ecologia de tres especies de moluscos gasterópodos intermareales: Rissoa parva, Barleeia unifasciata y Bittium reticulatum. I: Estructura y dinamica de poblaciones.- Cahiers de Biologie marine, Roscoff, vol. 27, p. 491-507.
Borrego J. (1992, unpublished).- Sedimentología del estuario del río Odiel (Huelva, SO Espańa).- PhD Thesis, Universidad de Sevilla, 311 p.
Borrego J., Morales J.A. & Pendón J.G. (1995).- Holocene estuarine facies along the mesotidal coast of Huelva, south-western Spain. In: Fleming B.W. & Bartholoma A. (eds.), Tidal signatures in modern and ancient sediments.- International Association of Sedimentologists, Special Publication, vol. 24, p. 151-170.
Borrego J., Ruiz, F., González-Regalado M.L., Pendón J.G. & Morales J.A. (1999).- The Holocene transgression into the estuarine central basin of the Odiel River mouth (Cadiz gulf, SW, Spain): Lithology and faunal assemblages.- Quaternary Science Reviews, vol. 18, p. 769-788.
Boski T., Moura D., Veiga-Pires C., Camacho S., Duarte D., Scott D.B. & Fernandes S.G. (2002).- Postglacial sea-level rise and sedimentary response in the Guadiana Estuary, Portugal/Spain border.- Sedimentary Geology, vol. 150, p. 103-122.
Bossio A., Foresi L.M., Mazzei R., Salvatorini G., Sandrelli F., Bilotti M., Colli A. & Rosetto R. (2003-2004).- Geology and stratigraphy of the southern sector of the Neogene Albegna river basin (Grosseto, Tuscany, Italy).- Geologica Romana, vol. XXXVII, p. 165-173. URL: https://www.dst.uniroma1.it/geologicaromana/Volumi/VOL%2037/12-Bossio.pdf
Cáceres L.M., Gómez, P., González-Regalado M.L., Clemente M.J., Rodríguez Vidal J., Toscano A., Monge G., Abad M., Izquierdo T., Monge A.M., Ruiz F., Campos J.M., Bermejo J., Martínez-Aguirre A. & López G. (2018).- Modelling the mid-late Holocene evolution of the Huelva Estuary and its human colonization, south-western Spain.- Marine Geology, vol. 406, p. 12-26.
Calvo L. & Langer M.R. (2010).- Beaching biogeographic barriers: The invasión of Haynesina germanica (Foraminifera, Protista) in the Bahía Blanca estuary, Argentina.- Biological Invasions, vol. 12, p. 3299-3306.
Caporizzo C., Gracia F.J., Aucelli P.P.C., Barbero L., Martín-Puertas C., Lagóstena L., Ruiz, J.A., Alonso C., Mattei G., Galán-Ruffoni I., López-Ramírez J.A. & Higueras-Milena A. (2021).- Late-Holocene evolution of the Northern Bay of Cádiz from geomorphological, stratigraphic and archaeological data.- Quaternary International, vol. 602, p. 92-109.
Carboni M.G. & Palagi I. (1998).- The Neogene - Quaternary deposits of the coastal belt between the Tafone and Marta Rivers (Northern Latium).- Bolletino della Societá Paleontologica Italiana, Milano, vol. 37, no. 1, p. 41-60.
Carro B., Borrego J., López-González N., Grande J.A., Gómez T., De la Torre M.L. & Valente T. (2011).- Impact of acid mine drainage on the hydrogeochemical characteristics of the Tinto-Odiel Estuary (SW Spain).- Journal of Iberian Geology, vol. 37, p. 87-96.
Catanzariti R. & Da Prato S. (2020).- New biostratigraphic and lithological data on the Neogene and Quaternary of the Livorno area (Tuscany, central Italy).- Rivista Italiana di Paleontologia e Stratigrafia, vol. 126, p. 725-749. DOI: https://doi.org/10.13130/2039-4942/14277
Cearreta A. (1989).- Foraminiferal assemblages in the ria of San Vicente de La Barquera (Cantabria, Spain).- Revista Espańola de Micropaleontología, Madrid, vol. 21, p. 67-80.
Cearreta A., Monge-Ganuzas M. & Iriarte E. (2006).- Análisis micropaleontológico y evolución ambiental holocena del estuario superior del río Oka (área de Portuzarra, Gernika-Lumo).- Illunzar, Gernika-Lumo, no. 6, p. 57-68.
Civis J., Sierro F.J., González-Delgado J.A., Flores J.A., Andrés I., Porta J. & Valle M.F. (1987).- El Neógeno marino de la Provincia de Huelva: Antecedentes y definición de sus unidades litoestratigráficas. In: Civis J. (ed.), Paleontología del Neógeno de Huelva (W Cuenca del Guadalquivir).- Universidad de Salamanca, p. 5-16.
Colom G. (1974).- Foraminíferos ibéricos. Introducción al estudio de las especies bentónicas recientes.- Investigaciones Pesqueras, Barcelona, t. 38, no. 1, 245 p. URL: http://scimar.icm.csic.es/scimar/index.php/secId/8/IdArt/1834/
Corbí H. (2010).- Los foraminíferos de la cuenca neógena del Bajo Segura (sureste de Espańa). Bioestratigrafía y cambios paleoambientales en relación con la crisis de salinidad messiniense.- PhD Thesis, Universidad de Alicante, 289 p.
Cristini P.A. & De Francesco C.G. (2017).- Molluscan taphonomy patterns below the sediment-water interface in freshwater shallow lakes from the southeastern Pampa plain, Argentina.- Palaios, Lawrence - KS, vol. 32, p. 528-542.
Dabrio C.J., Zazo C., Goy J.L., Sierro F.J., Borja F., Lario J., González J.A. & Flores J.A. (2000).- Depositional history of estuarine infill during the last postglacial transgression Gulf of Cadiz, Southern Spain.- Marine Geology, vol. 162, p. 381- 404.
Danielopol D.L., Ito E., Wansarf G., Kamiya T., Cronin T.M. & Baltanás A. (2002).- Techniques for collection and study of Ostracoda. In: Holmes J.A. & Chivas A.R. (eds.), The Ostracoda: Applications in Quaternary research.- Geophysical Monograph Series, Washington - DC, vol. 131, p. 65-97.
Danise S., Dominici S. & Betocco U. (2010).- Mollusk species at a Pliocene shelf whale fall (Orciano Pisano, Tuscany).- Palaios, Lawrence - KS, vol. 25, p. 449-456.
Danukalova G., Hallégouet B., Lefort J.P., Monnier J.-L. & Osipova E. (2015).- Reconstruction of the Middle-Late Pleistocene and Holocene palaeoenvironments of the Piégu Palaeolithic site (Brittany, France) based on biostratigraphical data.- Archaeosciences, Rennes, vol. 39, p. 7-30.
Debenay J.-P., Bénéteau E., Zhang J., Stouff V., Geslin E., Redois F. & Fernández-González M. (1998).- Ammonia beccarii and Ammonia tepida (Foraminifera): Morphofunctional arguments for their distinction.- Marine Micropaleontology, vol. 34, p. 235-244.
Desruelles S., Ghilardi M., Cloquier C., Delanghe D., Buchot Y., Hermitte D., Parisot J.-C. & Hoeblich J.M. (2021).- Mid-to Late Holocene landscape reconstruction of the Maye Estuary (Picardy, Northern France) and its implications for human occupation.- Quaternary International, vol. 601, p. 49-65.
Devillers B., Bony G., Degeai J.-P., Gascó J., Lachenal T., Bruneton H., Yung F., Oueslati H. & Thierry A. (2019).- Holocene coastal environmental changes and human occupation of the lower Hérault River, southern France.- Quaternary Science Reviews, vol. 222, article 105912, 16 p.
Di Bella L., Carboni M.G. & Bergamin L. (2000).- Pliocene-Pleistocene Foraminiferal assemblages of the middle and lower Tiber Valley: Stratigraphy and paleoecology.- Geologica Romana, vol. XXXVI, p. 129-145. URL: https://www.dst.uniroma1.it/geologicaromana/Volumi/VOL%2036/GR_36_129_145_Di%20Bella%20et%20al.pdf
Drinia H., Koskeridou E. & Antonarakou A. (2005).- Late Pliocene benthic foraminifera and mollusks from the Atsipades Section, Central Crete; Palaeoecological distribution and use in palaeoenvironmental assessment.- Géobios, vol. 38, p. 315-324.
Fairbridge R.W. (1980).- The estuary: Its definition and geodynamic cycle. In: Olausson E. & Cato I. (eds.), Chemistry and biochemistry of estuaries.- Wiley, New York - NY, p. 1-36.
Fernandes M.J., Namiotko T., Cabral M.C., Fatela F. & Boavida M.J. (2010).- Contribution to the knowledge of the freshwater Ostracoda fauna in continental Portugal, with an updated checklist of Recent and Quaternary species.- Journal of Limnology, Pavia, vol. 69, p. 160-173.
García E.X.M. (2002).- Estudio paleontológico de condrichtios y osteichtios del Neógeno (Plioceno Inferior) de la Formación "Arenas de Huelva" (suroeste de Espańa). MSc Thesis, Universidad de Guadalajara, 139 p.
García Gallardo A., Grunert P., Voelker A.H.L., Mendes I. & Piller W.E. (2017).- Re- evaluation of the "elevated epifauna" as indicator of Mediterranean outlflow water in the Gulf of Cadiz using stable isotopes (δ13C, δ18O).- Global and Planetary Change, vol. 155, p. 78-97.
García Sanz I. & Usera J. (2015).- Introducción al estudio de los foraminíferos del Cuaternario reciente del mar de Alborán.- Isurus, Alcoy, vol. 8, p. 56-63. URL: https://rua.ua.es/dspace/handle/10045/118524
García Sanz I., Usera J., Guillem J. & Alberola C. (2019).- Middle Miocene foraminifera from Canals (Valencia, western Mediterranean). Biostratigraphic and palaeoenvironmental aspects.- Spanish Journal of Palaeontology, Madrid, vol. 35, p. 89-106. DOI: https://doi.org/10.7203/sjp.35.1.17237
Ghandour I.M., Al-Zubier A.G., Basaham A.S., Mannaa A.A., Al-Dubai T.A. & Jones B.G. (2021).- Mid-Late Holocene paleoenvironmental and sea level reconstruction on the Al Lith Red Sea Coast, Saudi Arabia.- Frontiers in Marine Science, Lausanne, vol. 8, article 677010, 20 p. DOI: https://doi.org/10.3389/fmars.2021.677010
Gibson T.G. (1994).- Neogene and Quaternary Foraminifera and paleoenvironments of a corehole from Horn Island, Mississippi.- U.S. Geological Survey Open-file Report, Washington - DC, vol. 94-702, p. 1-24.
Gildeeva O., Akita L.G., Biehler J., Frenzel P. & Alivernini M. (2021).- Recent brackish water Foraminifera and Ostracoda from two estuaries in Ghana, and their potential as (palaeo)environmental indicators.- Estuarine, Coastal and Shelf Science, vol. 256, article 107270, 13 p.
Gofas S., Le Renard J. & Bouchet P. (2001).- In: Costello M.J., Emblow C. & White R. (eds.), European register of marine species. A check-list of the marine species in Europe and a bibliography of guides to their identification.- Mollusca.- Patrimoines Naturels, Paris, vol. 50, p. 180-213.
Goldstein S.T. & Richardson E.A. (2017).- Fine structure of the foraminifer Haynesina germanica (Ehrenberg) and its sequestered chloroplasts.- Marine Micropaleontology, vol. 138, p. 63-71.
Gómez G. (2017).- Guía de los moluscos marinos de Huelva y el Golfo de Cádiz.- Diputación de Huelva, Huelva, 300 p.
González Delgado J.A. (1986).- Estudio sistemático de los gasterópodos del Plioceno de Huelva (SW de Espańa). II: Mesogastropoda (Rissoacea, Cerithiacea).- Stvdia Geologica Salmanticensia, vol. 23, p. 61-120.
González Delgado J.A., Civis J., Valle M.F., Sierro F.J. & Flores J.A. (1986).- Distribución de los foraminíferos, moluscos y ostrácodos en el Neógeno de la Cuenca del Duero. Aspectos más significativos.- Stvdia Geologica Salmanticensia, vol. 22, p. 277-291.
González-Regalado M.L. (1989).- Estudio sistemático de los foraminíferos bentónicos de las arenas fosilíferas del Plioceno de Huelva: Su significado paleoecológico.- Estudios Geológicos, Madrid, vol. 45, nos. 1-2, p. 101-119. DOI: https://doi.org/10.3989/egeol.89451-2485
González-Regalado M.L., Gómez P., Ruiz F., Cáceres L.M., Clemente M.J., Rodríguez Vidal J., Toscano A., Monge G., Abad M., Izquierdo T., Campos J.M., Bermejo J., Martínez-Aguirre A., Prudencio M.I., Dias M.I., Marques R. & Muńoz J.M. (2019a).- Facies analysis, foraminiferal record and chronostratigraphy of Holocene sequences from Saltés Island (Tinto-Odiel estuary, SW Spain): The origin of high-energy deposits.- Estuarine, Coastal and Shelf Science, vol. 218, p. 95-105.
González-Regalado M.L., Gómez P., Ruiz F., Cáceres L.M., Clemente M.J., Rodríguez Vidal J., Toscano A., Monge G., Abad M., Izquierdo T., Monge Soares A.M., Campos J.M., Bermejo J., Martínez A. & López G.I. (2019b).- Holocene palaeoenvironmental evolution of Saltés Island (Tinto and Odiel estuary, SW Spain) during the Roman period (1st century BC-5th century AD).- Journal of Iberian Geology, vol. 45, p. 129-145.
González-Regalado M.L., Romero V., Abad M., Tosquella J., Izquierdo T., Gómez, P., Clemente M.J., Toscano A., Rodríguez Vidal J., Cáceres L.M., Muńoz J.M., Prudencio M.I., Dias M.I., Marques R., García E.X.M., Carretero M.I., Ruiz F. & Monge G. (2019c).- Late Tortonian-Middle Messinian palaeoenvironmental changes in the western Betic Strait (SW Spain).- Ameghiniana, Buenos Aires, vol. 56, p. 336- 360.
González-Regalado M.L. & Ruiz F. (1996).- Les foraminiféres benthiques de la baie dusud-ouest de l'Espagne pendant le Néogčne supérieur : Le Mio-Pliocčne de Huelva.- Revue de Paléobiologie, Genčven vol. 15, p. 109-120.
González-Regalado M.L., Ruiz F., Baceta J.I., González-Regalado E. & Muńoz J.M. (2001).- Total benthic foraminifera assemblages in the southwestern Spanish estuaries.- Géobios, vol. 34, p. 39-51.
Hardage K., Street J., Herrera-Silveira J.A., Oberle F.K.J & Paytan A. (2021).- Late Holocene environmental change in Celestun Lagoon, Yucatan, Mexico.- Journal of Paleolimnology, vol. 67, p. 131-162.
Jiwarungrueangkul T., Kirapinyakul A., Sompongchiyakul P., Zhao S. & Rattanakom R. (2022).- Response of sediment grain size to sea-level rise during the middle Holocene on the west coast of the Gulf of Thailand.- Arabian Journal of Geosciences, vol. 15, article no. 132, 10 p.
Kulkoyluoglu O. (2003).- Ecology of Freshwater Ostracoda (Crustacea) from Lakes and Reservoirs in Bolu, Turkey.- Journal of Freshwater Ecology, vol. 3, p. 343- 347.
Langer M.R. (1993).- Epiphytic foraminifera.- Marine Micropaleontology, vol. 20, p. 235-265.
Lario J., Zazo C., Goy J.L., Dabrio C.J., Borja F., Silva P.G., Sierro F.J., González A., Soler V. & Yll E. (2002).- Changes in sedimentation trends in SW Iberia Holocene estuaries.- Quaternary International, vol. 93-94, p. 171-176.
Larrasoańa J.C., Liu Q., Hu P., Roberts A.P., Mata P., Civis J., Sierro F.J. & Pérez-Asensio J.N. (2014).- Paleomagnetic and paleoenvironmental implications of magnetofossil occurrences in late Miocene marine sediments from the Guadalquivir Basin, SW Spain.- Frontiers in Microbiology, Lausanne, vol. 5, article 71, 15 p. DOI: https://doi.org/10.3389/fmicb.2014.00071
Lo Giudice Capelli E. & Austin W.E.N. (2019).- Size matters: Analyses of benthic Foraminiferal assemblages across Differing Size Fractions.- Frontiers in Marine Science, Lausanne, vol. 6, article 752, 8 p. DOI: https://doi.org/10.3389/fmars.2019.00752
Loeblich A.R. & Tappan H. (1987).- Foraminiferal Genera and Their Classification.- Van Nostrand Rienhold Co., New York - NY, 2013 p.
Luque A.A. & Templado J.T. (2004).- Praderas y bosques marinos de Andalucía.- Consejería de Medio Ambiente, Sevilla, 336 p.
Marks L., Welc F., Woronko B., Krzyminska J., Rogoz-Matyszczak A., Szymakek M., Holusa J., Nitychoruk J., Chen Z. & Salem A. (2022).- High-resolution insight into the Holocene environmental history of the Burullus Lagoon in northern Nile delta, Egypt.- Quaternary Research, Cambridge (UK), vol. 107, p. 87-103.
Martín-Rubio M., Elorza-Ramón M., Rodríguez-Lázaro J. & Pascual A. (2006).- Distribución areal y ecología de las asociaciones de ostrácodos recientes en la marisma Joyel (Cantabria).- Geogaceta, vol. 40, p. 87-90. URL: https://sge.usal.es/archivos/geogacetas/Geo40/Geo40-47.pdf
Martín-Rubio M., Rodríguez-Lázaro J. & Pascual A. (2004).- Distribución areal y ecología de las asociaciones de ostrácodos recientes en la marisma Victoria (Noja, Cantabria).- Geogaceta, Madrid, vol. 36, p. 155-158. URL: https://sge.usal.es/archivos/geogacetas/Geo36/Art39.pdf
Martínez B., Pascual A., Rodríguez-Lázaro J., Martín-Rubio M. & Rofes J. (2013).- The Ostracoda (Crustacea) of the Tina Menor estuary (Cantabria, southern Bay of Biscay): Distribution and ecology.- Journal of Sea Research, vol. 83, p. 111-122.
Martínez B., Ordiales A., Pérez A., Muńoz A., Luzón A. & Murelaga X. (2016a).- Caracterización ecológica de los ostrácodos holocenos de los Ojos de Ańavieja (Soria, Espańa).- Geogaceta, Madrid, vol. 59, p. 55-58. URL: https://sge.usal.es/archivos/geogacetas/geo59/geo59_14p55_58.pdf
Martínez B., Suárez O. & Murelaga X. (2015).- Estudio de las asociaciones de ostrácodos en sedimentos actuales de las Bardenas Reales de Navarra.- Geogaceta, Madrid, vol. 58, p. 59-62. URL: https://sge.usal.es/archivos/geogacetas/geo58/geo58pag55-58.pdf
Martínez B., Suárez O. & Murelaga X. (2016b).- Distribución y ecología de los ostrácodos recientes de las balsas de Bardenas Reales (Navarra, N de Espańa). In: Meléndez-Hevia G., Núńez A. & Tomás M. (eds.), Actas de las XXXII Jornadas de la Sociedad Espańola de Paleontología.- Instituto Geológico y Minero de Espańa, Madrid, p. 451-455.
Martins J.M.M. & Soares A.M.M. (2013).- Marine radiocarbon reservoir effect in Southern Atlantic Iberian coast.- Radiocarbon, Cambridge (UK), vol. 55, p. 1123-1134.
Matamales-Andreu R. (2019).- Historical review together with stratigraphical and taphonomical considerations on the upper Pleistocene deposit of Arenal de son Servera (Mallorca, Balearic Islands).- Carnets Geol., Madrid, vol. 19, no. 14, p. 277-285. DOI: https://doi.org/10.4267/2042/70497
Mateu-Vicens G., Khokhova A. & Sebastián-Pastor T. (2014).- Epiphytic foraminiferal indices as bioindicators in Mediterranean seagrass meadows.- Journal of Foraminiferal Research, Lawrence - KS, vol. 44, p. 325-339.
Mattos L.L., Martins I., Belart P., Alves M.V., Frontalini F., Mattos V., Gomes A., Toski T., Lorini M.L., Fortes R.R. & Rodrigues M.C. (2016).- Multiproxies (foraminifera, ostracods and biopolymers) approach to identify the environmental partitioning of the Guadiana River estuary (Iberian Peninsula).- Journal of Sedimentary Environments, vol. 1, p. 178-195.
Maya K., Mohan V., Limaye R.B., Padmalal D. & Navnith K.P. (2017).- Geomorphic response to sea level and climate changes during Late Quaternary in a humid tropical coastline: Terrain evolution model from Southwest India.- PLos One, vol. 12, no. 5, article e0176775, 32 p. DOI: https://doi.org/10.1371/journal.pone.0176775
McSweeney S.L., Kennedy D.M. & Rutherford I.D. (2017).- A geomorphic classification of intermittently open/closed estuaries (IOCE) derived from estuaries in Victoria, Australia.- Progress in Physical Geography: Earth Environment, London, vol. 41, p. 421-449.
Melis R., Bernasconi M.P., Colizza E., Di Rita F., Equini E., Forte E., Montenegro M.E., Pugliese N. & Ricci M. (2015).- Late Holocene palaeoenvironmental evolution of the northern harbour at the Elaiussa Sebaste archaeological site (south-eastern Turkey): Evidence from core ELA6.- Turkish Journal of Earth Sciences, Ankara, vol. 24, no. 6, article 2, p. 566-584. DOI: https://doi.org/10.3906/yer-1504-10
Mendes I., González R., Dias J.M.A, Lobo F. & Martins V. (2004).- Factors influencing recent benthic foraminifera distribution on the Guadiana shelf (Southwestern Iberia).- Marine Micropaleontology, vol. 51, p. 171-192.
Milker Y. & Schmiedl G. (2012).- A taxonomic guide to modern benthic shelf foraminifera of the western Mediterranean Sea.- Palaeontologia Electronica, Amherst - MA, article no. 15.2.16A, 134 p. URL: https://palaeo-electronica.org/content/2012-issue-2-articles/223-taxonomy-foraminifera
Murray J.W. (2006).- Ecology and palaeoecology of benthic Foraminifera.- Routledge, London, 396 p.
Nagendra R. & Reddy A.N. (2019).- Benthic Foraminifera Response to Ecosystem Pollution in the Uppanar Estuary, Tamil Nadu Coast, India.- Journal of the Geological Society of India, vol. 93, p. 555-566.
Newell R. (2009).- Behavioural aspects of the ecology of Peringia (=Hydrobia) ulvae (Pennant) (Gastropoda, Prosobranchia).- Journal of Zoology, vol. 138, p. 49-75.
Papadopoulou P., Iliopoulos I., Protopapas D., Spyropoulos S., Karanika K., Tsoni M. & Koukouvelas I. (2019).- Formation, evolution and demise of a tectonically controlled volcanic lake: A case study from the lower Pleistocene Sousaki succession.- Géobios, vol. 55, p. 41-55.
Pascual A., Martínez-García B. & Mendicoa J. (2019).- Cambios medioambientales de origen antrópico en el estuario de Joyel (Cantabria) detectados mediante asociaciones vivas de foraminíferos y ostrácodos.- Geogaceta, Madrid, vol. 66, p. 71-74. URL: https://sge.usal.es/archivos/geogacetas/geo66/Geo66_19.pdf
Pascual A., Rodríguez-Lázaro J., Weber O. & Jouanneau J.M. (2002).- Late Holocene pollution in the Gernika estuary (southern Bay of Biscay) evidenced by the study of Foraminifera and Ostracoda.- Hydrobiologia, vol. 475/476, p. 477-491.
Pascual A., Weber O., Caballero F., Rodríguez-Lázaro J. & Jouanneay J.M. (1999).- Análisis integral (micropaleontología y sedimentología) de un sondeo supramareal Holoceno en la ría de Gernika (Golfo de Vizcaya).- Geogaceta, Madrid, vol. 26, p. 75-78. URL: https://sge.usal.es/archivos/geogacetas/Geo26/Art19.pdf
Pereira R., Dillenburg S.R., Savian J.F. & Pereira J.C. (2021).- The Santa Vitória Alloformation: An update on a Pleistocene fossil-rich unit in Southern Brazil.- Brazilian Journal of Geology, Săo Paulo, vol. 51, no. 1, article e2020065, 18 p. DOI: https://doi.org/10.1590/2317-4889202120200065
Pereira R., Ritter M.N., Guimaraes E., Correa M.L., Rebello S. & Caron F. (2022).- The influence of coastal evolution on the paleobiogeography of the bivalve Anomalocardia flexuosa (Linné, 1767) along the southwestern Atlantic Ocean.- Journal of South America Earth Sciences, vol. 113, article 103662, 16 p.
Pérez-Asensio J.N., Aguirre J., Schmiedl G. & Civis J. (2012).- Messinian paleoenvironmental evolution in the lower Guadalquivir Basin (SW Spain) based on benthic foraminifera.- Palćogeography, Palćoclimatology, Palćoecology, vol. 326-328, p. 135-151.
Pint A. & Frenzel P. (2017).- Ostracod fauna associated with Cyprideis torosa - an overview.- Journal of Micropalaeontology, vol. 36, p. 113-119.
Pippér M. & Reichenbacher B. (2009).- Biostratigraphy and paleoecology of benthic foraminifera from the Eggenburgian "Ortenburger Meeressande" of southeastern Germany (Early Miocene, Paratethys).- Neues Jahrburch für Geologie und Paläontologie, Stuttgart, vol. 254, p. 41-61.
Pozo M., Ruiz F., Carretero M.I., Rodríguez Vidal J., Cáceres L.M., Abad M. & González-Regalado M.L. (2010).- Mineralogical assemblages, geochemistry and fossil associations of Pleistocene-Holocene complex siliciclastic deposits from the Southwestern Dońana National Park (SW Spain): A palaeoenvironmental approach.- Sedimentary Geology, vol. 225, p. 1-18.
Roca J.R. & Danielopol D.L. (1991).- Exploration of interstitial habitats by the phytophilous Ostracod Cypridopsis vidua (O.F. Müller): Experimental evidence.- Annales de Limnologie, Les Ulis, vol. 27, no. 3, p. 243-252. DOI: https://doi.org/10.1051/limn/1991018
Rögl F. & Spezzaferri S. (2003).- Foraminiferal paleoecology and biostratigraphy of the Muhlbach section (Gaindorf Formation, Lower Badenian), Lower Austria.- Annalen des Naturhistorischen Museums in Wien (serie A), Bd. 104, p. 23-75. URL: https://www.zobodat.at/pdf/ANNA_104A_0023-0075.pdf
Ronchi L., Fontana A. & Correggiari A. (2018).- Characteristics and potential application of Holocene tidal inlets in the northern Adriatic shelf (Italy). In: Quaternary: Past, Present, Future - AIQUA Conference 2018.- Alpine and Mediterranean Quaternary, Firenze, vol. 31, p. 31-34. URL: https://amq.aiqua.it/index.php/amq/article/view/182
Rothenberg B. & Blanco Freijero A. (1980).- Ancient copper mining and smelting at Chinflon. In: Craddock P.T. (ed.), Scientific studies in early mining and extractive metallurgy.- British Museum, London, p. 41-62.
Ruiz F. (2001).- Trace metals in estuarine sediments from the southwestern Spanish coast.- Marine Pollution Bulletin, vol. 42, p. 482-490.
Ruiz F., Borrego J., López-González N., Abad, M., González-Regalado M.L., Carro B., Pendón J.G., Rodríguez Vidal J., Cáceres L.M., Prudencio M.I. & Dias M.I. (2007).- The geological record of a mid-Holocene marine storm in southwestern Spain.- Géobios, vol. 40, p. 689-699.
Ruiz F., Carretero M.I., Pozo M., González-Regalado M.L., Monge G., Rodríguez Vidal J., Cáceres L.M., Abad M., Tosquella J., Izquierdo T., Muńoz J.M., Prudencio M.I., Dias M.I., Marques R., Gómez P., Toscano A., Romero V. & Arroyo M. (2021).- Texture and mineralogy as proxies of late Holocene tsunami deposits from the Dońana National Park (SW Spain).- Italian Journal of Geosciences, Roma, vol. 140, no. 2, p. 312-324.
Ruiz F. & González-Regalado M.L. (1996).- Les ostracodes du Golfe Mio-Pliocene du sud-ouest de l'Espagne.- Révue de Micropaléontologie, vol. 39, p. 137-151.
Ruiz F., González-Regalado M.L., Baceta J.I. & Muńoz J.M. (2000).- Comparative ecological analysis of the ostracod faunas from low- and high-polluted southwestern Spanish estuaries: A multivariate approach.- Marine Micropaleontology, vol. 40, p. 345-376.
Ruiz F., González-Regalado M.L., Borrego J., Abad M. & Pendón J.G. (2004b).- Ostracoda and foraminifera as short-term tracers of environmental changes in very polluted areas: The Odiel Estuary (SW Spain).- Environmental Pollution, vol. 129, p. 49-61.
Ruiz F., González-Regalado M.L., Galán E., González I., Prudencio M.I., Dias M.I., Abad M., Toscano A., Prenda J. & García E.X.M. (2012).- Benthic foraminifera as bioindicators of anthropogenic impacts in two north African lagoons: A comparison with ostracod assemblages.- Revista Mexicana de Ciencias Geológicas, Mexico City, vol. 29, no. 3, p. 527-533. URL: http://www.rmcg.unam.mx/index.php/rmcg/article/view/49/40
Ruiz F., González-Regalado M.L. & Muńoz J.M. (1997).- Multivariate analysis applied to total and living fauna: Seasonal ecology of recent benthic Ostracoda off the North Cádiz Gulf coast (southwestern Spain).- Marine Micropaleontology, vol. 31, p. 183-203.
Ruiz F., Rodríguez Ramírez A., Cáceres L.M., Rodríguez Vidal J., Carretero M.I., Clemente L., Muńoz J.M., Yáńez C. & Abad M. (2004a).- Late Holocene evolution of the southwestern Dońana Nacional Park (Guadalquivir Estuary, SW Spain): A multivariate approach.- Palćogeography, Palćoclimatology, Palćoecology, vol. 204, p. 47-64.
Ruiz F., Rodríguez Ramírez A., Cáceres L.M., Rodríguez Vidal J., Yáńez C., Clemente L., González-Regalado M.L., Abad M. & De Andrés J.A. (2002).- Cambios paleoambientales en la desembocadura del río Guadalquivir durante el Holoceno reciente.- Geogaceta, Madrid, vol. 31, p. 167-170. URL: https://sge.usal.es/archivos/geogacetas/Geo31/Art42.pdf
Ruiz F., Pozo M., Carretero M.I., Abad M., Gonzalez-Regalado M.L., Muńoz J.M., Rodríguez Vidal J., Cáceres L.M., Pendón J.G., Prudencio M.I. & Dias M.I. (2010).- Birth, Evolution and Death of a Lagoon: Late Pleistocene to Holocene Palaeoenvironmental Reconstruction of the Dońana National Park (SW Spain). In: Friedman A.G. (ed.), Lagoons: Biology, management and environmental impact.- Nova Science Publishers, New York - NY, p. 371-396.
Salihoglu R., Okcer-on S., Langer M. & Viehberg F. (2018).- Holocene environmental reconstruction of Lake Bafa (SW, Anatolia) by statistical analyses of Ostracod and benthic foraminifer assemblages.- Geophysical Research Abstracts, Munich, vol. 20, EGU2018-1108-1, p. 1108.
Schaumann R.M., Capperucci R.M., Bungenstock F., McCan T., Enters D., Wehrmann A. & Bartholoma A. (2021).- The Middle Pleistocene to early Holocene subsurface geology of the Norderney tidal basin: New insights from core data and high-resolution sub-bottom profiling (Central Wadden Sea, southern North Sea).- Netherlands Journal of Geosciences, Utrecht, vol. 100, article e15, 20 p. DOI: https://doi.org/10.1017/njg.2021.3
Sciuto F. (2014).- Ostracods of the Upper Pliocene-Pleistocene Punta Mazza succession (NE Sicily) with special focus on the Family Trachyleberididae Sylvester-Bradley, 1948, and description of a new species.- Carnets Geol., Madrid, vol. 14, no. 1, p. 1-13. DOI: https://doi.org/10.4267/2042/53172
Sciuto F., Rosso A., Sanfilippo R. & Maniscalco R. (2015).- New faunistic data on the Pleistocene environmental evolution of the south-western edge of the Hyblean Plateau (SE Sicily).- Carnets Geol., Madrid, vol. 15, p. 41-57. DOI: https://doi.org/10.4267/2042/56401
Sciuto F. & Baldanza A. (2020).- Full restoration of marine conditions after the late Messinian Mediterranean Lago-Mare phase in Licodia Eubea and Villafranca Tirrena areas (east Sicily).- Carnets Geol., Madrid, vol. 20, no. 6, p. 107-123. DOI: https://doi.org/10.4267/2042/70760
Stéphan P., Verdin F., Arnaud-Fassetta G., Bertrand F., Eynaud F., García-Artola A., Bosq M., Culioli C., Suanez S., Coutelier C., Bertran P. & Costa S. (2019).- Holocene coastal changes along the Gironde estuary (SW France): New insights from the North Médoc península beach/dune system.- Quaternaire, Paris, vol. 30, no. 1, p. 47-75. DOI: https://doi.org/10.4000/quaternaire.11172
Ta T.K.O., Nguyen V.L., Saito Y., Gugliotta M., Tamura T., Nguyen T.M.L., Truong M.H. & Bui T.L. (2021).- Latest Pleistocene to Holocene stratigraphic record and evolution of the Paleo-Mekong incised valley, Vietnam.- Marine Geology, vol. 433, article 106406, 16 p.
Tornos F. (2008).- La geología y metalogenia de la Faja Pirítica Ibérica.- Macla, Madrid, vol. 10, p. 13-23. URL: http://www.semineral.es/websem/PdfServlet?mod=archivos&subMod=publicaciones&archivo=Macla10_13.pdf
Torres T., Ortiz J.E., Soler V., Reyes E., Delgado A., Valle M., Llamas J.F., Cobos R., Juliá R., Badal E., García de la Morena M.A., García-Martínez M.J., Fernández-Gianotti J., Calvo J.P. & Cortés A. (2003).- Pleistocene lacustrine basin of the east domain of Guadix-Baza basin (Granada, Spain): Sedimentology, chronostratigraphy, and palaeoenvironment. In: Valero-Garcés B.L. (ed.), Limnogeology in Spain: A tribute to Kerry R. Kelts.- Consejo Superior de Investigaciones Científicas, Madrid, p. 151-185.
Trobec A., Busetti M., Zgur F., Baradello L., Babich A., Cova A., Gordini E., Romeo R., Tomini I., Poglajen S., Diviacco P. & Vrabec M. (2018).- Thickness of marine Holocene sediment in the Gulf of Trieste (northern Adriatic Sea).- Earth System Science Data, vol. 10, no. 2, p. 1077-1092. DOI: https://doi.org/10.5194/essd-10-1077-2018
Tulbure M.A., Capella W., Barhoun N., Flores J.A., Hilgen F.J., Krijgsman W. & Kouwenhoven T. (2017).- Age refinement and basin evolution of the North Rifian Corridor (Morocco): No evidence for a marine connection during the Messinian Salinity Crisis.- Palćogeography, Palćoclimatology, Palćoecology, vol. 485, p. 416-432.
Usera J., Molina E., Montoya P., Robles R. & Santisteban C. (2000).- Límites entre sistemas y pisos en la provincia de Alicante. In: Cańaveras J.C., García del Cura M.A. & Meléndez A. (eds.), Itinerarios geológicos por la provincia de Alicante y limítrofes.- Universidad de Alicante, p. 43-58.
Vázquez A., Plana F., De Porta J., Zamarreńo I. & Macpherson I. (1990).- Caracterizacion paleoclimatica de la serie margosa Langhiense de Can Rosell (Barcelona).- Acta Geologica Hispanica, Barcelona, vol. 25, p. 209-219. URL: https://revistes.ub.edu/index.php/ActaGeologica/article/view/4490
Viehberg F.A., Frenzel P. & Hoffmann G. (2008).- Succession of late Pleistocene and Holocene ostracode assemblages in a transgressive environment: A study at a coastal locality of the southern Baltic Sea (Germany).- Palćogeography, Palćoclimatology, Palćoecology, vol. 264, p. 318-329.
Williams S., Garrett E., Moss P., Bartlett R. & Gehrels R. (2021).- Development of a training set of contemporary salt-marsh Foraminifera for Late Holocene sea-level reconstructions in southeastern Australia.- Open Quaternary, London, vol. 7, article 4, 29 p. DOI: http://doi.org/10.5334/oq.93
Wukovits J., Bukenberger P., Enge A.J., Gerg M., Wanek W., Watzka M. & Heinz P. (2018).- Food supply and size class depending variations in phytodetritus intake in the benthic foraminifer Ammonia tepida.- Biology Open, Cambridge (UK), vol. 7, no. 4, article bio030056, 10 p. DOI: https://doi.org/10.1242/bio.030056
Zaibi C., Kamoun F., Carbonel P. & Montacer M. (2011).- Ostracods distribution in the subsurface sediments of El-Guettiate Sebkha (Skhira, gulf of Gabes). Interest for Holocene palaeoenvironmental reconstruction.- Carnets Geol., Madrid, vol. 11, no. A03 (CG2011_A03), p. 63-81. DOI: https://doi.org/10.4267/2042/42327
Zazo C., Goy J.L., Somoza L., Dabrio C.J., Belloumini G., Improta S., Lario J., Bardají T. & Silva P.G. (1994).- Holocene sequence of sea-level fluctuations in relation to climatic trends in the Atlantic-Mediterranean linkage coast.- Journal of Coastal Research, Fort Lauderdale - FL, vol. 10, no. 4, p. 933-945.