◄ Carnets Geol. 22 (18) ►
![]()
Outline:
[1. Introduction]
[2. Geographic and geologic framework of the study area]
[3. Material and methods]
[4. Brief description of the study deposits]
[5. Systematics]
[6. Concluding remarks]
[7. Episkeletobionts (encrustings)]
[8. Taphonomic remarks and palaeoenvironmental interpretation)]
[9. Conclusion)]
and ... [Bibliographic references]
École Normale Supérieure, Laboratory of Animal Ecobiology (LEBA), Kouba, DZ-16000 Algiers (Algeria)
Faculty of Natural and Life Sciences, University Mustapha Stambouli of Mascara, DZ-29000 Mascara (Algeria)
2 rue Guy de Maupassant, F-76800 Saint Étienne du Rouvray (France)
University of Science and Technology Houari Boumediene (USTHB), Geodynamics of the Lithosphere, DZ-16000 Algiers (Algeria)
École Normale Supérieure, Kouba, DZ-16000 Algiers (Algeria)
Published online in final form (pdf) on December 25, 2022
DOI 10.2110/carnets.2022.2218
![]()
[Editor: Beatriz Bádenas; language editor: Stephen Eagar;
technical editor: Bruno R.C. Granier]
![]()
A hundred specimens of pteriomorph bivalves were collected from the marine deposits of the marly-sandy formations of Algiers Sahel, which correspond to transitional facies between the Piacenzian deep marly deposits and the Astian molassic deposits. They are herein analysed from a systematic, taphonomic, palaeoecological and paleoenvironmental point of view. The preliminary inventory list consists of 27 species belonging to eight families: Arcidae, Nuculidae, Glycymeridae, Spondylidae, Pectinidae, Plicatulidae, Gryphaeidae, and Ostreidae. Two sedimentary units with different bivalve biodiversity are recognized: the first: a shallow infralittoral unit characterised by large Flabellipecten alessii, Aequipecten angelonii, Ostrea lamellosa and O. edulis; the second: A deeper unit of circalittoral environment mainly includes Amusium cristatum. These bivalve shells display sclerobiont traces. Bioerosion traces are mostly assignable to clionid sponges (Entobia isp.), polychaete worms (Maeandropolydora isp. and Caulostrepsis isp.), bivalves (Gastrochaenolites isp.) and predatory gastropods (Oichnus isp.). Identified encrusting organisms are juvenile oyster shells, cirripedes, polychaete worms (serpulids), and indeterminate bryozoans. Analysis of boring and encrusting traces indicate a progressive environmental change from a relative high energy setting to a lower energy and deeper water setting, and correspond to a shallow sea with well-oxygenated waters.
• pteriomorph bivalves;
• systematics;
• taphonomy and paleoenvironment;
• marine Pliocene;
• Algeria
Talmat S., Benyoucef M., Ferré B., Bouzeguella S. & Ouchene F.-Z. (2022).- Pliocene bivalves (Pteriomorphia) of Algiers Sahel (Algeria): Systematics and palaeoecology.- Carnets Geol., Madrid, vol. 22, no. 18, p. 795-824.
Bivalves ptériomorphes pliocčnes du Sahel d'Alger (Algérie) : Systématique et paléo-écologie.- Une centaine de spécimens de bivalves ptériomorphes ont été récoltés dans les dépots marins des formations marno-sableuses pliocčnes du Sahel d'Alger qui correspondent ŕ des facičs de transition entre les dépots marneux profonds du Plaisancien et les dépots molassiques de l'Astien. Ils sont ici analysés sur le plan taxonomique, taphonomique, paléoécologique et paléoenvironemental. L'inventaire taxonomique préliminaire comprend 27 espčces appartenant ŕ huit familles : Arcidae, Nuculidae, Glycymeridae, Spondylidae, Pectinidae, Plicatulidae, Gryphaeidae et Ostreideae. Deux unités sédimentaires avec une biodiversité différente des bivalves ont été reconnues : La premičre peu profonde de milieu infralittoral est caractérisée par Flabellipecten alessii, Aequipecten angelonii, Ostrea lamellosa et O. edulis ; la seconde, plus profonde, circalittorale comprend principalement Amusium cristatum. Les coquilles de ces bivalves comportent des traces de sclérobiontes. Les traces de bioérosion sont principalement attribuables ŕ des éponges clionides (Entobia isp.), des polychčtes (Maeandropolydora isp. et Caulostrepsis isp.), des bivalves (Gastrochaenolites isp.) et ŕ des gastéropodes prédateurs (Oichnus isp.). Les organismes encroűtants identifiés sont des valves d'huîtres juvéniles, des balanes, des polychčtes (serpulidés) et des bryozoaires indéterminés. L'analyse de ces traces majoritairement observées sur les bivalves de grande taille indiquerait plutôt un changement environnemental progressif, d'un milieu ŕ haute énergie vers une ŕ basse énergie et des eaux plus profondes, et devrait correspondre ŕ une mer plutôt peu profonde avec des eaux bien oxygénées.
• bivalves ptériomorphes ;
• systématique ;
• taphonomie et paléo-environnement ;
• Pliocčne marin ;
• Algérie
The Pliocene deposits of Algiers Sahel are composed of marls and sandy marls with intercalated sandstones and limestones, which are rich in benthonic fauna, especially bivalves. Quite a number of studies in this area reported the abundance of these bivalves but without any precise information on their systematics and palaeoecology (e.g., Delage, 1888; Glangeaud, 1932, 1936, 1968; Dalloni, 1934, 1954; Aymé, 1948, 1955, 1963; Muraour, 1949, 1951; Glangeaud et al., 1952; Aymé et al., 1953; Cabot, 1968; Yassini, 1973, 1975; Saadallah, 1981; Saoudi, 1989; Djediat, 2013).
During the construction of residential buildings in the southeastern suburb of Algiers in 2012-2013, extensive earthwork in the Pliocene marly-sandy deposits exposed two lithological sections that allowed us to collect a rich and diversified bivalve assemblage associated with some gastropods, brachiopods and bryozoans. The present study aims to describe the collected bivalve specimens, to define their systematics and to interpret their palaeoecological and palaeobiogeographical framework.
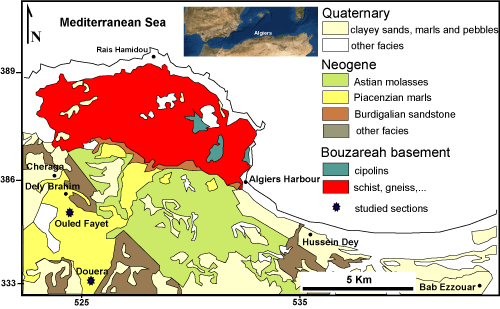
The Algiers Sahel belongs to the inner zones of the Maghrebides range that represents the northern part of the Peri-Mediterranean Alpine orogen (Durand Delga,
1971). Of NE-SW direction, along the Mediterranean shore, the Algiers Sahel runs over an anticline structure and builds a high plateau, ranging between 200 m and 250 m in height, between the Chenoua and the Algiers massifs in the north and the Mitidja plain in the south. The main tectono-sedimentary units of the Algiers massif are as follows
(Fig. 1 ![]() ):
):
1) The Palaeozoic metamorphic massif of Bouzareah, consisting of gneiss, schists, marble, and micaschists. It displays traces of Hercynian and Alpine intense tectonics dividing the basement rocks into seven tectono-metamorphic units (Saadallah, 1981);
2) The Cenozoic strata represented by transgressive formations that overlie unconformably the metamorphic terrains. Their succession can be subdivided in three parts:
the Miocene, more extensively outcropping in the south of the Mitidja basin. In the Oued Mazafran, Aymé et al. (1953) characterised a sedimentary series, including from base to top, Burdigalian sandy marls (350 m), a sandstone marker bed underlining the transition to the Upper Miocene, and a thick Upper Miocene marly succession, gypsiferous at its base (250 m), with thin sandstone intercalations at its top (300 m);
the Pliocene deposits including from base to top: Piacenzian gypsiferous marls (1,000 m), a ca. 100 m thick Astian succession of glauconitic sandstones, sandy marls rich in benthonic fauna, and massive calcareous sandstones with large bivalves. They are followed by a reefal or subreefal complex, and marine and continental detrital beds (ca. 30 m), that initiated the Villafranchian infilling of the Mitidja basin (Glangeaud, 1932; Glangeaud et al., 1952);
the Quaternary displaying several regressive phases led to the deposition of diversified facies structured in the northern flank of the Algiers Sahel: the Pleistocene, which is composed of Calabrian, Sicilian and Tyrrhenian marine terraces (Saoudi, 1989), and the Holocene which is characterised from base to top by a shelly facies, coastal silty mudstones rich in microfauna and red clayey sandy soils, resulting from the weathering of sandstone-carbonate facies, marls and gravels (Djediat, 2013).
|
Figure 1:
Geological map of Algiers and its surroundings (after Aymé,
1963), showing the location of the study area. |
The studied material was collected in two selected sections of the Astian sandy marls of Algiers Sahel
(Fig. 2 ![]() ), the first located at Ouled Fayet (UMTS coordinates:
36°42'37"N
2°56'08"E), and the second one at Douera (UMTS coordinates: 36°38'18"N
2°56'18"E). Both localities correspond to outcrops exposed thanks to intensive earthwork made for residential buildings
purposes during the years 2012-2013. A hundred bivalve shells were collected and housed at the Laboratory of Animal Ecobiology (LEBA) from the École Normale Supérieure of Kouba (Algiers, Algeria). The collected shells were cleaned, numbered (ST: Sciences Terre), and photographed with a digital picture camera Nikon d3100.
), the first located at Ouled Fayet (UMTS coordinates:
36°42'37"N
2°56'08"E), and the second one at Douera (UMTS coordinates: 36°38'18"N
2°56'18"E). Both localities correspond to outcrops exposed thanks to intensive earthwork made for residential buildings
purposes during the years 2012-2013. A hundred bivalve shells were collected and housed at the Laboratory of Animal Ecobiology (LEBA) from the École Normale Supérieure of Kouba (Algiers, Algeria). The collected shells were cleaned, numbered (ST: Sciences Terre), and photographed with a digital picture camera Nikon d3100.
Respective measurements of the shell valves are as follows: h= antero-posterior length; l= umbo-ventral length; c= hinge length when available.
The study area belongs to the Astian of Algiers Sahel, which is mainly formed of sandy marls. The synthetic lithological succession at the Ouled Fayet section, displays from base to top three units
(Fig. 2.B ![]() ): 10 m of compact grey marls (unit a); ca. 30 m of brownish, fossiliferous, sandy marls with intercalated lumachellic levels rich in Amusium cristatum (unit b) and ca 25 m of yellowish, fossiliferous sandy marls (unit c). At the Douera section, the retrieved bivalve specimens only came from the yellow, fossiliferous, sandy marls of unit c (ca. 10 m); this succession is
inaccessible today since the building of a high school soon after our fossil collecting.
Figure 2.B
): 10 m of compact grey marls (unit a); ca. 30 m of brownish, fossiliferous, sandy marls with intercalated lumachellic levels rich in Amusium cristatum (unit b) and ca 25 m of yellowish, fossiliferous sandy marls (unit c). At the Douera section, the retrieved bivalve specimens only came from the yellow, fossiliferous, sandy marls of unit c (ca. 10 m); this succession is
inaccessible today since the building of a high school soon after our fossil collecting.
Figure 2.B ![]() illustrates the stratigraphic distribution of the identified species collected in both localities.
illustrates the stratigraphic distribution of the identified species collected in both localities.
|
Figure 2:
A (A1 and A2). Aerial and panoramic view of the Ouled Fayet section.
B. Synthetic lithological log showing the stratigraphic distribution of the respective species collected in this study. |
The 27 bivalve species listed in this work are recorded in Figure 2.B ![]() and illustrated in Figures 3
and illustrated in Figures 3 ![]() to 7
to 7 ![]() . The shells are in good state of preservation. We used the classification of Pteriomorphia established by Waller
(1978).
. The shells are in good state of preservation. We used the classification of Pteriomorphia established by Waller
(1978).
Class Bivalvia Linnaeus, 1758
Subclass Pteriomorpha Beurlen, 1944
Order Arcoida Stoliczka, 1871
Superfamily Arcacea Lamarck, 1809
Family Arcidae Lamarck, 1809
Subfamily Anadarinae Reinhart, 1935
Genus Anadara Gray, 1847
Type species: Arca antiquata Linnaeus, 1758
Anadara diluvii (Lamarck, 1805)
(Fig. 3a ![]() )
)
1805 Arca diluvii Lamarck, VI, p. 219.
1898 Anadara diluvii (Lamarck).- Sacco, XXVI, p. 20, Pl. IV, figs. 7-12.
1914 Arca (Anadara) diluvii Lamarck.- Cossmann & Peyrot, p. 149, Pl. VIII, figs. 3-6; Pl. X, figs. 53, 56.
1963 Arca (Arca) diluvii Lamarck.- Venzo & Pelosio, p. 139, Pl. XLII, figs. 6-8.
1975 Anadara (Anadara) diluvii (Lamarck).- Feki, p. 28, Pl. III, fig. 1.a-d.
1987 Anadara (Anadara) diluvii (Lmk).- Freneix et al., p. 12, Pl. I, fig. 9.
1994 Anadara (Anadara) diluvii (Lamarck, 1805).- Ben Moussa, p. 63, Pl. 1, figs. 19-22.
2012 Anadara (Anadara) diluvii (Lamarck, 1805).- Satour, p. 5, Fig. 4K.
2021 Anadara diluvii sensu Sacco, 1898, non Lamarck, 1805.- Benyoucef et al., Fig. 4A.
Material: Two right valves (ST136, l=34 mm, h=25 mm, c=25 mm; ST139, l=8 mm, h=10 mm, c=10 mm) and one left valve (ST138, l=33 mm, h=28 mm, c=23 mm), all coming from the yellow sandy marls of Ouled Fayet (unit c).
Description: The three collected valves are relatively small, oval, with rounded anterior side and clearly shorter than the posterior side. The rounded umbones are slightly prosogyrous. The ligament area is narrow and rather wide. The outer side of the valves is ornamented by some thirty unequal radial costulae wider than the interspaces.
Stratigraphic range: A. diluvii was first recorded in the Paratethys from the Oligocene of Hungary (Malatesta, 1974). It is also known in the Aquitanian of the Atlantic-Mediterranean domain. It had a wide geographic distribution during the Miocene (Nordic, Atlantic, Mediterranean, and Paratethys domains). In the Pliocene, this taxon was restricted to the Atlantic-Mediterranean domain. Nowadays it is still scarce: Mediterranean Sea, Atlantic coast of Iberia, West coast of Africa to Angola, and Cape Verde Islands (Freneix et al., 1987).
Ecology: This taxon is suspensivorous, slow burrowing, surficial as generally is the genus Anadara (Stanley, 1970). According to Pérčs & Picard (1964), A. diluvii characterises rather the circalittoral zone than the infralittoral one, mobile seabed and the biocoenosis of offshore detrital sea bottoms. It is abundant between -80 m and -100 m, less common in organogenic muds of the Algerian precontinent (Caulet, 1972). It is also reported between -20 m and -50 m in the continental shelf of the eastern Rif in Morocco (Saubade & Tesson, 1981).
|
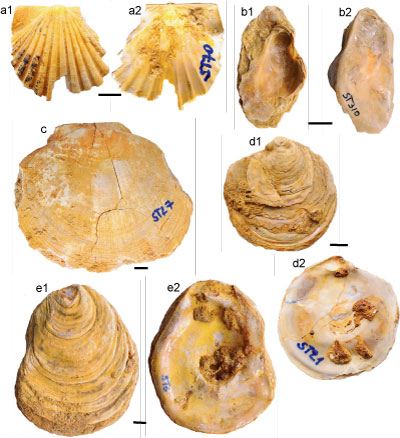
Figure 3:
Bivalve taxa from the Pliocene of the Algiers Sahel. a, Anadara diluvii (Lamarck, 1805), specimen ST136; a1, external view, right valve; a2, internal view, right valve; b, Nucula (Nucula) sp., specimen ST91; b1, external view, right valve; b2, dorsal view; c, Glycymeris bimaculata deshayesi (Mayer,
1868), specimen ST60; c1, external view, left valve; c2, external view, right valve; d, Spondylus crassicosta Lamarck,
1819, specimen ST37; complete shell; e, Talochlamys multistriata (Poli, 1795), specimen ST48; e1, external view, right valve; e2, internal view, right valve; f, Mimachlamys varia (Linnaeus, 1758), specimen ST51; f1, external view, left valve; f2, internal view, left valve; g, Mimachlamys varia (Linnaeus, 1758), specimen ST102; g1, external view, right valve; g2, internal view, right valve. Scale bar= 10 mm for all. |
Order Nuculoida Dall, 1889
Family Nuculidae Gray, 1824
Genus Nucula Lamarck, 1799
Subgenus Nucula Lamarck, 1799
Type species: Arca nucleus Linnaeus, 1758
Nucula (Nucula) sp.
(Fig. 3b ![]() )
)
Material: A single specimen (ST91) from the yellowish sandy marls of Ouled Fayet (unit c).
Description: The sampled specimen is an inner mould of small size (l=23 mm, h=25 mm, c=12 mm), of oval-triangular shape, slightly higher than wide and oblique, with opisthogyrous umbones displaced with respect to one another.
Stratigraphic range: Nucula (Nucula) sp. ranges from the Middle Miocene to the Recent.
Ecology: Species of the subgenus Nucula (Nucula) are ubiquitous, euryhaline and depositivorous, and live superficially burrowing in sandy bottoms, in depths not exceeding 100 m (Parenzan, 1974). They tolerate low salinity and can live in lagoons (Saubade & Tesson, 1981), but mostly they are currently found offshore, on fine-sandy, coarse sand and silty bottoms around -180 m.
Superfamily Limopsacea Dall, 1895
Family Glycymerididae Newton, 1922
Subfamily Glycymeridinae Newton, 1922
Genus Glycymeris Costa, 1778
Subgenus Glycymeris Costa, 1778
Type species: Glycymeris glycymeris Linnaeus, 1758
Glycymeris (Glycymeris) bimaculata deshayesi (Mayer, 1868)
(Fig. 3c ![]() )
)
1795 Arca bimaculata.- Poli, p. 143, Pl. XXV, figs. 17-18;
1868 Pectunculus Deshayesi.- Mayer, p. 53; 1 46
1963 Glycymeris bimaculata.- Tavani & Tongiorgi, p. 10, Pl. 1, fig. 1; Pl. 2, fig. 3; Pl. 4, figs. 1, 7-8.
1987 Glycymeris (Glycymeris) bimaculata (Poli, 1793) deshayesi (Mayer, 1868).- Freneix et al., p. 14, Pl. II, fig. 1.
1994 Glycymeris (Glycymeris) bimaculata deshayesi (Mayer, 1868).- Ben Moussa, p. 66, Pl. 1, fig. 26.
2012 Glycymeris (Glycymeris) bimaculata deshayesi (Poli, 1795).- Satour, p. 23, Pl. 1, fig. 4.
2021 Glycymeris cf. bimaculata ((Poli, 1795).- Benyoucef et al., p. 8-9, Fig. 4D.
Material: A complete shell (ST60, l=50 mm, h=50 mm, c=30 mm) and three inner moulds (ST76, l=40 mm, h=39 mm, c=30 mm; ST77, l=44 mm, h=40 mm, c=30 mm; ST106, l=55 mm, h=50 mm, c=40 mm) sampled from Ouled Fayet, as well as valve fragments from Douera (unit c).
Description: The complete specimen (ST60) displays a very thick and relatively large shell with two valves slightly offset on the dorsal side with respect to one another. The triangular ligament area shows a clearly visible taxodont hinge. These three inner moulds are of similar size. The shell outline is orbicular and displays a straight and slightly bulging umbo. The cardinal plate is wide and slightly arcuate. The outer surface is covered by fine concentric growth lines.
Stratigraphic range: G. bimaculata bimaculata appeared in the Mediterranean Pliocene, replacing G. bimaculata deshayesi, and ranges in the Atlantic, Mediterranean Sea and the Black Sea (Freneix et al., 1987).
Ecology: This subspecies lives in infralittoral, soft sea bottoms and can be one of the characteristic components of the biocoenosis of fine and coarse sands under the influence of bottom water currents (Pérčs & Picard, 1964).
Family Spondylidae Gray, 1825
Genus Spondylus Linnaeus, 1758
Type species: Spondylus gaederopus Linnaeus, 1758
Spondylus (Spondylus) crassicosta Lamarck, 1819
(Fig. 3d ![]() )
)
1819 Spondylus crassicosta.- Lamarck, VI, p. 193.
1898 Spondylus crassicosta Lamarck.- Sacco, XXV, p. 5, Pl. I, fig. 17; Pl. II, figs. 1-2.
1920 Spondylus crassicosta Lamarck.- Dollfus & Dautzenberg, p. 452, Pl. XLVI, figs. 1-4.
1963 Spondylus crassicosta Lamarck.- Venzo & Pelosio, p. 161, Pl. II, fig. 1.
1974 Spondylus (Spondylus) crassicosta Lamarck.- Malatesta, p. 60, Pl. V, fig. 3.a-b.
1987 Spondylus (Spondylus) crassicosta Lamarck, 1819.- Freneix et al., p. 35, Pl. VIII, figs. 2-3.
1994 Spondylus (Spondylus) crassicosta crassicosta Lamarck, 1819.- Ben Moussa, p. 105, Pl. 6, fig. 4.
Material: One single shell (ST37, l=80 mm, h=85 mm, C=50 mm) collected in the sandy marls unit of Ouled Fayet (unit c).
Description: This single specimen is thick, ovoid and of large size. It is clearly characterised by its strong ribs (4-7), evenly spaced and ornamented by wide scaly spines. The interspaces, much wider than the ribs, have two, underdeveloped but clearly visible, secondary ribs bearing quite numerous spines.
Stratigraphic range: S. crassicosta is a species of the Atlantic-Mediterranean Neogene, which appeared under its ancestral form, S. crassicosta aquitanica Cossmann & Peyrot, in the Aquitanian from Aquitaine, France. From the Middle Miocene, this taxon acquired its derived form, S. crassicosta crassicosta, and became abundant and widespread in the Atlantic, Mediterranean Sea and the Paratethys. It reached its acme during the Pliocene, and at the upper boundary of its stratigraphic extension, this taxon became circum-Mediterranean and disappeared at the end of the Pliocene (Zavarei, 1973).
Ecology: As well as recent spondyles, this species was a suspensivorous and cemented its valves to hard substrates, living in shallow settings (down to -50 m) on coralligenous bottoms. S. crassicosta dwelled coralligenous bottoms between -25 and -30 m, likewise its live homologous sister taxon, S. princeps Reeve from the Gulf of California and Panama (Freneix et al., 1987).
Order Pterioida Newell, 1965
Superfamily Pectinacea Rafinesque, 1815 (emend.)
Family Pectinidae Rafinesque, 1815 (emend.)
Genus Talochlamys Iredale, 1929
Type species: Chlamys islandica Müller, 1776
Talochlamys multistriata (Poli, 1795)
(Fig. 3e ![]() )
)
1758 Ostrea pusio Linnaeus, X, p. 698.
1939 Chlamys multistriata Poli.- Roger, p. 165, Pl. XXII, figs. 5-7, 11-15; Pl. XXIII, fig. 5; Pl. XXIV, figs. 8-9.
1961 Chlamys multistriata Poli.- Veiga Ferreira, p. 452, Pl. XI, figs. 65-66.
1970 Chlamys (Chlamys) multistriata Poli.- Raffi, p. 108, Pl. XXX, figs. 5-17.
1987 Chlamys (Mimachlamys) pusio (Linnaeus, 1758).- Freneix et al., p. 22, Pl. III, 2.
1994 Chlamys (Chlamys) multistriata (Poli, 1795).- Ben Moussa, p. 76, Pl. 3, fig. 3.
2003 Crassodoma multistriata (Poli, 1795).- Courville & Bongrain, p. 138, Pl. 3, figs. 3-4.
2012 Crassodoma multistriata (Poli, 1795).- Satour, p. 40, Pl. 1, fig. 13.
2017 Talochlamys multistriata (Poli, 1895).- Spadini, p. 10.
Material: Seven right valves (ST48, l=25 mm, h=30 mm, c=10 mm; ST54, l=20 mm, h=22 mm, c=10 mm; ST156, l=25 mm, h=25 mm, c=12 mm; ST183, l=19 mm, h=20 mm, c=12 mm; ST219, l=29 mm, h=26 mm, c=15 mm; ST220, l=22 mm, h=24 mm, c=10 mm; ST222, l=16 mm, h=21 mm, c=10 mm) collected in the sandy marls of Ouled Fayet and Douera (unit c).
Description: This taxon is easily identifiable thanks to its umbono-palleal oval, convex, slightly squashed shape, with some 40 fine, unequal radial ribs, of yellowish to brownish colour. The radial ridges are more or less rounded and the margin is slightly undulated. The dimensions of the different valves are small when compared to those of modern representatives, suggesting that these are juvenile specimens. Both auricles are unequal; the anterior one is three times larger than the posterior one. This latter is obliquely truncated and bears small spines, as well as fine ribs of the Camptonectes type.
Stratigraphic range: Talochlamys multistriata is a very common species, reported from the Aquitanian to the Recent, in the Boreal, Nordic, Lusitanian, Mauritanian, and eastern South-Atlantic bioprovinces, as well as in the Mediterranean, Indo-Mediterranean and St. Helena, southwards to South Africa (Dijkstra & Goud, 2002).
Ecology: This taxon is suspensivorous, epibyssal or cemented, eurytopic, living from the infralittoral to the bathyal zones; however, it is more frequent in depths ranging from -10 m to -100 m. It bears numerous fine costulae developing by division on its right valve and by intercalation on its left valve. In the Mediterranean, this shell keeps its regular shape, but in the Atlantic at the adult stage, it cements to the substratum by its right valve that gets extremely irregular and strongly squamose, while its left valve flattens. By this moment, it exhibits the outline of the recent uncommon genus Hinnites (Roger, 1939).
Genus Mimachlamys Iredale, 1929
Type species: Pecten asperrimus Lamarck, 1819
Mimachlamys varia (Linnaeus, 1758)
(Fig. 3f-g ![]() )
)
1758 Ostrea varia.- Linnaeus, X, p. 698.
1939 Chlamys varia Linnaeus.- Roger, p. 197, Pl. XXII, figs. 21-23.
1957 Chlamys varia Linnaeus.- Perrodon, p. 207.
1975 Chlamys varia Linnaeus.- Feki, p. 40, Pl. X, fig. 5.
1989 Chlamys (Chlamys) varia (Linnaeus).- Lauriat-Rage et al., p. 125.
1994 Chlamys (Aequipecten) radians (Nyst & Westendrop, 1839).- Ben Moussa, p. 73, Pl. 3, fig. 6.
2012 Mimachlamys varia (Linnaeus).- Satour, p. 33.
2017 Mimachlamys varia (Linnaeus).- Spadini, p. 21-22.
2021 Mimachlamys varia (Linnaeus, 1758).- Benyoucef et al., p. 12, Fig. 5E.
Material: Two groups of specimens were collected from the brown sandy marls of Ouled Fayet (unit b). The first group is represented by two right valves (ST51, l=27 mm, h=29 mm, c=16 mm; ST156, l=21 mm, h=25 mm, c=12 mm). The second group includes a single complete shell (ST56, l=40 mm, h=41 mm, c=12 mm), a single left valve (ST102, l=44 mm, h=47 mm, c=20 mm) and a single right valve (ST143, l=31 mm, h=35 mm, c=15 mm).
Description: The shell resembles that of Talochlamys multistriata as it is inequivalve, higher than wider, of medium size, and carries 23 thin, rounded, radial ribs. The auricles are very unequal, finely crenulated with a distinct and rounded byssal sinus, with a ctenolium in the anterior auricle twice bigger than the posterior one, topped by three fine teeth on the posterior edge. The second group of specimens is characterised by a medium-size, equivalve, slightly higher than wide, and thicker shell. The valves are convex to rather flat and display rounded radial ribs and deep interspaces. The auricles are unequal and of triangular shape with fine ribs crossed by growth lines. The smaller posterior auricle is usually broken.
Stratigraphic range: Mimachlamys varia, as well as T. multistriata, are known from the western Mediterranean Sea since the Miocene (Roger, 1939). M. varia is reported from the Aquitanian of Portugal to the Recent (Veiga Ferreira, 1961). Widespread during the whole Neogene, it reached the Nordic domain in the Pleistocene.
Ecology: This taxon, optionally epibyssal, dwells the different biotopes of the infralittoral and circalittoral zones. It ranges from southern Norway to the northern Gulf of Guinea. Nowadays this species dwells the Atlantic (frequent), Mediterranean Sea and the Adriatic.
According to Caulet (1972), it occurs frequently on the detrital sea bottoms of the Algerian shelf.
Genus Manupecten Monterosato, 1872
Type species: Manupecten pesfelis (Linnaeus, 1758)
Manupecten pesfelis (Linnaeus, 1758)
(Fig. 4a ![]() )
)
1758 Ostrea pesfelis.- Linnaeus, p. 697.
1939 Chlamys pesfelis Linnaeus.- Roger, p. 178, Pl. XXVI, figs. 1-2.
1970 Chlamys (Manupecten) pesfelis (Linnaeus).- Raffi, p. 122, Pl. 27, fig. 8.
1994 Chlamys (Manupecten) pesfelis (L.).- Ben Moussa, p. 86, Pl. 3, fig. 14.
2014 Manupecten pesfelis (Linnaeus, 1758).- Satour, p. 38.
2017 Manupecten pesfelis (Linnaeus, 1758).- Spadini, p. 10.
Material: Two left valves (ST49, l=26 mm, h=28 mm, c=10 mm; ST146, l=24 mm, h=24 mm, c=4 mm) collected from the brown sandy marls of Ouled Fayet (unit b).
Description: Manupecten pesfelis resembles Pseudamussium clavatum. The sampled valves are of small size and of yellowish to brownish colour. The ribs (5) are flatter and the auricles are slightly less developed than those of the former species, with the anterior auricle more developed.
Stratigraphic range: M. pesfelis is reported from the Pliocene of Cyprus (Reed, 1935) and the Plio-Pleistocene of Syria (Roman, 1940).
Ecology: It dwells the coastal detrital sea bottoms of the circalittoral zone, mostly in the Mediterranean Sea, as well as the adjacent area of the eastern Atlantic from Portugal southwards to Cape Verde (Pérčs & Picard, 1964).
|
Figure
4: Bivalve taxa from the Pliocene of the Algiers Sahel. a, Manupecten pesfelis (Linnaeus, 1758), specimen ST49; a1, external view, left valve; a2, internal view, left valve; b, Pseudamussium clavatum (Poli, 1795), specimen ST85; b1, external view, right valve; b2, internal view, right valve; c, Aequipecten opercularis (Linnaeus, 1758), specimen ST136; c1, external view, right valve; c2, internal view, right valve; d, Aequipecten angelonii Stefani & Pantanelli,
1878, specimen ST207; d1, external view, left valve; d2, internal view, left valve; e, Aequipecten scabrella (Lamarck, 1819), specimen ST82; e1, external view, right valve; e2, internal view, right valve; f, Aequipecten bollenensis (Mayer,
1876), specimen ST217; f1, external view, right valve; f2, internal view, right valve. Scale bar= 10 mm for all.
|
Genus Pseudamussium Mörch, 1853
Type species: Pecten septemradiatus Müller, 1776
Pseudamussium clavatum (Poli, 1795)
(Fig. 4b ![]() )
)
1795 Chlamys clavata (Poli).- Peplum clavatum (Poli, 1795).
1795 Ostrea clavata Poli.- p. 160, Pl. 28, fig. 17.
1795 Ostrea inflexa Poli.- p. 160, Pl. 28, figs. 4-5.
1993 Pseudamussium clavatum (Poli, 1795).- Poppe & Goto, p. 70, Pl. 10, fig. 6.a-b.
1999 Pseudamussium clavatum (Poli, 1795).- Ardovini & Cossignani, p. 91, 2 Figs.
2002 Pseudamussium clavatum (Poli, 1795).- Dijkstra & Gould, p. 56, Fig. 53.
2017 Pseudamussium clavatum (Poli, 1795).- Spadini, p. 10.
Material: A single left valve (ST47, l=18 mm, h=19 mm, c=9 mm) and two right valves (ST74, l=18 mm, h=19 mm, c=9 mm; ST85, l=22 mm, h=23 mm, c=8 mm) collected from the brown sandy marls of Ouled Fayet (unit b), and also from the yellow sandy marls of both localities (unit c).
Description: Pseudamussium clavatum displays different macro- and microconic shells. The shell is slightly unequilateral, wider than high, with an anterior margin slightly wider than the posterior one, the right valve being more concave than the left one. The umbo is triangular and slightly protruding on the cardinal edge. While the valves are less curvaceous, their ventral margins are really curved, showing a spoon-like shape and five marked ribs wider than the interspace among which, the three in the middle being much wider. The ribs are rounded close to the umbo and flatten towards the outer margin. Both auricles are smaller but the anterior one being slightly larger than the posterior one. They display radial ribs, as well as growth lines oriented perpendicularly to the cardinal edge.
Stratigraphic range: Pseudamussium clavatum appeared in the Miocene.
Ecology: It occurs on the sandy and gravelly detrital sea bottoms of the circalittoral zone in the Mediterranean Sea and the eastern Atlantic.
Genus Aequipecten Fischer, 1886
Type species: Ostrea opercularis Linnaeus, 1758
Aequipecten opercularis (Linnaeus, 1758)
(Fig. 4c ![]() )
)
1758 Ostrea opercularis.- Linnaeus, X, p. 698.
1939 Chlamys opercularis (L.).- Roger, p. 131, Pl. 16, figs. 6-7; Pl. 17, figs. 3-5.
1972 Chlamys (Aequipecten) opercularis (L.).- Caprotti, p. 58, Pl. 1, fig. 5.
1978 Chlamys (Aequipecten) opercularis (L.).- Steininger & Schultz, p. 341.
1987 Aequipecten opercularis (Linnaeus, 1758).- Freneix et al., p. 24, Pl. III, fig. 6.
1994 Chlamys (Aequipecten) opercularis (L.).- Ben Moussa, p. 78.
2017 Aequipecten opercularis (Linnaeus, 1758).- Spadini, p. 10.
Material: Five right valves (ST52, l=37 mm, h=38 mm, c=21 mm; ST55, l=46 mm, h=45 mm, c=27 mm; ST83, l=32 mm, h=32 mm, c=14 mm; ST134, l=25 mm, h=25 mm, c=11 mm; ST167, l=40 mm, h=39 mm, c=23 mm) and three left valves (ST50, l=38 mm, h=38 mm, c=20 mm; ST168, l=34 mm, h=35 mm, c=17 mm; ST182, l=30 mm, h=30 mm, c=12 mm) collected from the grey sandy marls of Ouled Fayet (unit b).
Description: The shell is rounded, almost equivalve and equilateral. The valve is easily recognisable thanks to the imprint of the adductor muscle and the byssal sinus of the anterior auricle, as well as to the set of small teeth building the ctenolium. The valve is weakly convex with a flat umbonal area. The umbo is straight and not contorted. Ornamentation consists of 21 radial ribs wider than the interspaces, terminated by spines and with less accentuated relief. The auricles are unequal and costate. Some forms bear triangular-outlined ribs.
Stratigraphic range: Aequipecten opercularis is a common and widespread taxon, ranging from the Miocene to the Recent, in the Nordic and Boreal bioprovinces, as well as in the Lusitanian-Mauritanian, Mediterranean, and central Paratethys ones. Nowadays it is found from Norway and the Faroe Islands to the Iberian Peninsula, the Açores, and the Canaries (Tebble, 1976).
Ecology: This species is suspensivorous and lives attached by a byssus or free-lying. It is an active swimmer, ranging from several metres deep down to -2,000 m (Roger, 1939). In the Algerian continental margin, it is reported between -10 m and -150 m (Caulet, 1972).
Aequipecten angelonii Stefani & Pantanelli, 1878
(Fig. 4d ![]() )
)
1858 Pecten angelonii Meneghini (in litt.).
1878 Mimachlamys angelonii.- Stefani & Pantanelli, p. 31.
1939 Chlamys angelonii Meneghini.- Roger, p. 138, Pl. XVIII, figs. 7-10; Pl. XX, figs. 5-9.
1970 Chlamys (Aequipecten) angelonii (Stefani & Pantanelli).- Raffi, p. 112, Pl. 27, fig. 9.
1987 Aequipecten angelonii (Stefani & Pantanelli, 1878).- Freneix et al., p. 25, Pl. IE, fig. X.
1994 Chlamys (Aequipecten) angelonii (Meneghini, 1859).- Ben Moussa, p. 81.
2017 Aequipecten angelonii (Stefani & Pantanelli, 1758).- Spadini, p. 10.
Material: Five left valves (ST118, 1=70 mm, h=70 mm, c=30 mm; ST202, l=74 mm, h=71 mm, c=15 mm; ST207, l=60 mm, h=60 mm, c=28 mm; ST210, l=52 mm, h=50 mm, c=28 mm; ST218, l=46 mm, h=48 mm, c=25 mm) collected from Ouled Fayet (unit b).
Description: The shell is of orange-yellow to brown, sometimes grayish in colour, of circular shape and variable size. It is inequivalve and displays 22 radial ribs slightly wider than the interspaces, with a spinose ornamentation consisting of two spine rows slanted towards the outer margin on both sides of the median row, especially developed close to the margin. The auricles are unequal and costate.
Stratigraphic range: It ranges from the Helvetian to the Pliocene, especially in the Auversian of Belgium, but its distribution is generally Mediterranean, with its acme in the Pliocene. This species also appears in the Burdigalian of SE France and spreads into the Atlantic and the Paratethys during the Middle Miocene. In the Pliocene, it was widespread in the Atlantic and the Mediterranean Sea, and disappears at the end of this period. In North Africa, it is reported from the "Sahelian" of Morocco (Ben Moussa, 1994).
Ecology: A. angelonii belongs to a species group adapted to shallow and agitated environments. According to Demarcq (1979), it occurs in the Burdigalian of the Rhône valley, often "in the platforms and well exposed shores, as well as in rocky shores". It dwells, like C. radians, both coarse and fine detrital facies.
Aequipecten scabrella (Lamarck, 1819)
(Fig. 4e ![]() )
)
1819 Pecten seniensis.- Lamarck, p. 182.
1819 Pecten scabrellus.- Lamarck, p. 183.
1939 Chlamys scabrella (Lamarck).- Roger, p. 332, 342.
1987 Chlamys seniensis (Lamarck).- Freneix et al., p. 24, Pl. III, figs. 7-10.
1994 Chlamys (Chlamys) varia (Linnaeus, 1758).- Ben Moussa, p. 77, Pl. 3, figs. 4-5, 10-11.
2009 Aequipecten scabrella (Lamarck).- Jimenez et al., p. 7, figs. 3.f-i, 4.a-b.
2012 Pecten seniensis (Lamarck, 1819).- Satour, p. 35, Pl. 1, figs. 10-11.
2017 Aequipecten scabrellus (Lamarck, 1819).- Spadini, p. 10.
2021 Aequipecten scabrella (Lamarck, 1819).- Benyoucef et al., p. 11, Fig. 5B.
Material: Five right valves (ST79, l=32 mm, h=29 mm, c=15 mm); ST80, l=30 mm, h=26 mm, c=10 mm; ST181, l=45 mm, h=39 mm, c=12 mm; ST82, l=36 mm, h=34 mm, c=20 mm; ST185, l=38 mm, h=36 mm, c=15 mm) and a single left valve (ST135, l=30 mm, h=27 mm, c=17 mm) collected from the brown sandy marls of Ouled Fayet (unit b) and the yellow sandy marls of both localities (unit c).
Description: The valves are wider than high with a medium to rather high convexity, almost even for both valves. The number of ribs varies from 14 to 17; they are thinner than the interspaces and have rounded edges. Concentric lamellae are visible on the ribs and in the interspaces; their marked undulation gives birth to visible scales first in the interspaces and then in the rib flanks. A. scabrella displays wide morphological range. The size, number of ribs, convexity, and secondary ornamentation vary from an individual to another. The auricles are unequal, the left auricle being wider and thinner with a small triangular, byssal sinus. The auricles bear radial ribs cut by growth lines.
Stratigraphic range: Aequipecten scabrella is very frequent in the Pliocene deposits of the Mediterranean Sea, ranging from the Miocene (Tortonian). It was replaced at the Pleistocene by Aequipecten commutatus (Monterosato, 1875).
Ecology: It dwells the sandy/muddy sea bottoms of western Mediterranean Sea in depths ca -30 to -40 m (Benyoucef et al., 2021).
Aequipecten bollenensis (Mayer, 1876)
(Fig. 4f ![]() )
)
1876 Pecten (Neitha) Bollenensis.- Mayer-Eymar, p. 169, Pl. VI, fig. 2.
1939 Chlamys bollenensis Mayer-Eymar.- Roger, p. 107, Pl. XIII, figs. 13-15; Pl. XV, figs. 9-11.
1961 Chlamys bollenensis Mayer-Eymar.- Veiga Ferreira, p. 447, Pl. n, figs. 5-9.
1975 Chlamys scabrella sarmenticia Goldfus.- Feki, p. 38, Pl. IX, fig. 4.
1994 Chlamys (Argopecten) bollenensis (Mayer-Eymar, 1876).- Ben Moussa, p. 85, Pl. 3, fig. 9.
2017 Aequipecten bollenensis Mayer, 1876.- Spadini, p. 10.
Material: Six right valves (ST137, l=31 mm, h=29 mm, c=11 mm; ST142, l=32 mm, h=30 mm, c=ind; ST157, l=25 mm, h=25 mm, c=10 mm; ST169, l=24 mm, h=23 mm, c=12 mm; ST217, l=27 mm, h=25 mm, c=13 mm; ST184, l=25 mm, h=25 mm, c=14 mm) and three left valves (ST47, l=18 mm, h=19 mm, c=9 mm; ST81, l=35 mm, h=34 mm, c=18 mm; ST84, l=34 mm, h=31 mm, c=14 mm) all collected in the grey sandy marls of Ouled Fayet (unit b) and the yellow sandy marls of both localities (unit c).
Description: The shell is unequilateral with a triangular shape, smaller than A. scabrellus. The right valve is more convex than the left one. Ribs (18) are slightly protruding, quadrate in section and wider than the interspaces.
Stratigraphic range: A. bollenensis has the same biogeographic range than A. scabrella but is shifted stratigraphically with respect to this latter. It appeared in the Burdigalian and got extinct in the Pleistocene. In the Miocene, it reached the Paratethys and even Minor Asia. In the Pliocene, this species became quite abundant, especially in the Mediterranean Sea (Ben Moussa, 1994).
Ecology: This taxon, more ubiquitous, usually occurs in the same facies and appears to dwell the same living environment than A. scabrella.
Aequipecten radians (Nyst & Westendrop, 1839)
(Fig. 5a ![]() )
)
1839 Pecten radians.- Nyst & Westendrop, p. 15, Pls. H-I, fig. 19.
1939 Chlamys radians Nyst.- Roger, p. 134, Pl. XVI, figs. 8-12; Pl. XVII, fig. 9.
1945 Pecten (Aequipecten) radians Nyst, 1839.- Glibert, p. 65, Pl. III, fig. 5.
1970 Chlamys radians (Nyst).- Bongrain, p. 35, Pls. A-B.
1989 Chlamys (Aequipecten) radians (Nyst).- Lauriat-Rage et al., p. 125, Pl. II, fig. 8.
1994 Chlamys (Aequipecten) radians (Nyst & Westendrop, 1839).- Ben Moussa, p. 73, Pl. 3, fig. 6.
2003 Aequipecten radians (Nyst).- Courville & Bongrain, p. 139, Pl. 3, figs. 6-8.
Material: Two complete shells (ST46, l=40 mm, h=42 mm, c=1.7 mm; ST78, l=43 mm, h=40 mm, c=22 mm) collected from the sandy marls of Ouled Fayet (unit b).
Description: Species of medium size. Both valves, slightly convex, build an equilateral shell in fan-shape, with 18 rounded-side ribs widening towards the outer margin. The shell surface is ornamented by fine and deep ribs. The unequal auricles display circular and oblique ribs and oblique ribs on the side areas. This species, wrongly called seniensis (Roger, 1939), can be confused with A. scabrella.
Stratigraphic range: A. radians, mentioned in several Miocene basins of the Atlantic-Mediterranean domain, was first described in the Aquitanian of Bazadais (Glibert & Poel, 1965). During the Miocene, it was quite widespread in the Atlantic from Belgium (Anversian) to Morocco, and in the Mediterranean Sea. In the Pliocene, it occurred in the Mediterranean Sea and is more widespread in the Atlantic and disappeared before the Pleistocene.
Ecology: This taxon, quite common and widespread, frequently occurs in detrital facies, most likely in shallow settings (Ben Moussa, 1994).
|
Figure
5 : Bivalve taxa from the Pliocene of the Algiers Sahel. a, Aequipecten radians (Nyst & Westendrop,
1839), specimen ST78; a1, external view, left valve; a2, external view, right valve; b, Korobkovia oblonga (Philippi, 1844), specimen ST177; b1, external view, right valve; b2, internal view, right valve; c, Amusium cristatum (Bronn,
1827), specimen ST162; c1, external view, left valve; c2, internal view, left valve; d, Pecten bipartitus (Foresti,
1876), specimen ST116; d1, external view, left valve; d2, internal view, left valve; e, Pecten maximus (Linnaeus, 1758), specimen ST86; e1, external view, left valve; e2, internal view, left valve. Scale bar= 10 mm for all. |
Genus Korobkovia Glibert & Poel, 1965
Type species: Korobkovia oblonga Philippi, 1844
Korobkovia oblonga (Philippi, 1844)
(Fig. 5b ![]() )
)
1844 Pecten oblongum.- Philippi, p. 300, Pl. XVI, fig. 7.
1876 Pecten comitatus.- Fontannes, p. 94
1897 Pseudoamussium oblongum Philippi.- Sacco, XXV, p. 52, Pl. XIV, figs. 40-43.
1928 Amusium oblungum Philippi.- Depéret & Roman, p. 186, Pl. XXVIII, figs. 2-3, 3.a-b.
1977 Korobkovia oblonga (Philippi).- González Donoso & Porta, p. 53, Pls. H-I, fig. 7.
1987 Amusium oblongum (Philippi, 1841).- Kharrim, p. 138, Pl. 12, fig. 2.
2017 Korobkovia oblonga (Philippi, 1844).- Spadini, p. 10.
Material: Three left valves (ST44, l=48 mm, h=54 mm, c=17 mm; ST164, l=76 mm, h=56 mm, c=31 mm; ST184, l=30 mm, h=36 mm, c=14 mm) and a single right valve (ST177, l= ind, h=50 mm, c=11 mm) all collected from the brownish sandy marls of Ouled Fayet (unit b).
Description: This is a thin, oval, small to medium size, higher than wide, rather flat, shell with a faintly protruding umbo and thin growth lines. It is characterised by the lack of costate ornamentation on the outer side of both valves and the inner side of the right valve. The left valve displays some 30 fine ribs on its inner side. Both tiny auricles are identical and draw a rectilinear cardinal margin unlike Amusium cristatum.
Stratigraphic range: K. oblonga is characteristic of the Pliocene of the western Mediterranean Sea: for example, in Spain, this species is restricted to the silts and fine-grained calcarenites of the base of the Pliocene (Jiménez et al., 2009).
Ecology: It should have lived in the same biotopes than A. cristatum (see below).
Genus Amusium Röding, 1798
Type species: Ostrea pleuronectes Linnaeus, 1758
Amusium cristatum (Bronn, 1827)
(Fig. 5c ![]() )
)
1827 Pecten cristatus.- Bronn, p. 542, Pl. 6, fig. 3.
1928 Amusium cristatum Bronn.- Depéret & Roman, p. 171, Pl. XXV, fig. 1.
1951 Amusium cristatum Bronn.- Veiga Ferreira, p. 160, Pl. V, figs. 15, 18-20.
1974 Amusium cristatum (Br.).- Malatesta, p. 39, Pl. 4, fig. 2.
1987 Amusium (Amussium) cristatum (Bronn, 1827).- Freneix et al., p. 21, Pls. H-I, fig. 1.
1994 Amusium (Amussium) cristatum (Bronn, 1827).- Ben Moussa, p. 103, Pl. 6, fig. 3.
2009 Amusium cristatum (Bronn, 1827).- Jiménez et al., p. 15, Fig. 6.a-b.
2017 Amusium cristatus (Brocchi, 1814).- Spadini, p. 10.
Material: Four right valves (ST162, l=85 mm, h=90 mm, c=25 mm; ST163, l=95 mm, h/2=45 mm, c/2=30 mm; ST178, l= ind, h= ind, c=30 mm; ST175, l=35 mm, h=35 mm, c=ca 10 mm) and also several valve fragments all collected from the brown and yellow sandy marls of Ouled Fayet (units b and c).
Description: Amusium cristatum is represented by a fine and translucent, equivalve, equilateral, slightly wider than high, shell, often of large size. The outer smooth shell displays on its inner side 26 to 30 scaly ribs with pointed edges, irregularly spaced and sometimes geminated, fading towards the umbo and ending shortly before the outer margin. Both valves, identical and slightly convex, display fine growth lines and a smooth outline. The auricles are almost equal without any byssal sinus, but with spines along the cardinal line, that form an obtuse angle with the umbo. According to Philippi (1844), the juvenile forms resemble Korobkovia oblonga.
Stratigraphic range: A. cristatum is essentially Mio-Pliocene, disappearing in the Pleistocene. It is mostly Mediterranean known from the Burdigalian. During the Miocene, it spread through the whole Mediterranean Sea, reached the Paratethys and extended into the Atlantic. In the Pliocene, it was widespread in all fine-grained facies of the Atlantic-Mediterranean domain.
Ecology: A. cristatum lived in calm waters, sandy-muddy soft sea bottoms, in depth ranging from -50 to -100 m. It is an excellent swimmer, with loose-sided valves and free adult stage, devoid of byssal attachment (Freneix et al., 1987).
Genus Pecten Müller, 1776
Subgenus Pecten Müller, 1776
Type species: Ostrea maxima Linnaeus, 1758
Pecten bipartitus Foresti, 1876
(Fig. 5d ![]() )
)
1873 Vola jacobaea var.- Cocconi, p. 338.
1876 Pecten (Pecten) bipartitus.- Foresti, p. 51, Tav. I, figs. 21-23.
1897 Pecten reghiensis.- Sacco, XXIV, p. 59, tav. XVIII, figs. 11-14.
1963 Pecten bipartitus (For.).- Tavani & Tongiorgi, p. 30, Pl. 1; Pl. 2; Pl. 3; Pl. 4, figs. 1-2.
1987 Pecten bipartitus (Foresti, 1876).- Freneix et al., p. 27, Pl. IV, figs. 2-3.
2012 Pecten bipartitus (Foresti, 1876).- Satour, p. 47.
2017 Pecten bipartitus Foresti, 1876.- Spadini, p. 10.
Material: One complete shell (ST88-212, l=43 mm, h=40 mm, c=20 mm), four right valves (ST75, l=58 mm, h=51 mm, c=3 mm; ST116, l=56 mm, h=50 mm, c=33 mm; ST203, l=53 mm, h=48 mm, c=27 mm; ST208, l=52 mm, h=48 mm, c=28 mm) and a single left valve (ST211, l=45 mm, h=43 mm, c=27 mm) all collected from the brown and yellow sandy marls of Ouled Fayet (units b and c).
Description: The shells are especially thin, inequivalve, slightly unequilateral and of medium to large size. The umbo-outer margin is shorter than the anterior-posterior one. The right valve is convex with a prominent umbo strongly protruding the cardinal line. The costulation consists of 13 ribs, sub-rectangular with rounded edge, and two thin ribbings on both sides of the valve. Interspaces are twice narrower than the ribs. Very fine, tightly spaced, growth lines run through the ribs and particularly the interspaces. They protrude on the ventral side. From a distance of the umbo, a longitudinal ridge divides the rib into two, sometimes three costulae, but such a costulation is not visible in juvenile forms. The left valve is flat to concave and hollow in the umbo area. It bears 13 ribs narrower than the interspaces, cut by thin, serrated and protruding, growth lines like in the right valve. The auricles are of medium size and subequal; the anterior auricle is costulated and shows a small byssal sinus and five to six radial ribs cut by thin, dense growth lines, all concentric and parallel on the ventral margin.
Stratigraphic range: P. bipartitus was first recorded in the Middle Miocene in southern Italy, being uncommon in the Tortonian (Sardinia, Morocco) and the Messinian (Algeria). It is mostly Pliocene in age in Italy, France, Spain, Algeria, Creta, and Syria (Freneix et al., 1987).
Ecology: This species is found in calcarenite facies and medium- to coarse-grained sands.
Pecten maximus (Linnaeus, 1758)
(Fig. 5e ![]() )
)
1758 Ostrea maxima.- Linnaeus, X, p. 696.
1778 Pecten vulgaris.- Costa, p. 140, Pl. IX, fig. 3.
1888 Pecten medius Daniel.- Arnould Locart, p. 150.
1905 Pecten maximus (L).- Choffat & Dollfus, p. 179-181.
1952 Pecten maximus (L).- Lecointre, p. 5-173.
1989 Pecten maximus (L).- Lauriat-Rage, p. 217.
2009 Pecten maximus Linnaeus, 1758.- Jiménez, p. 19, Figs. 8.c-d.
Material: Three left valves (ST12, l=85 mm, h=77 mm, c=40 mm; ST86, l=110 mm, h=92 mm, c=50 mm; ST88, l=80 mm, h=67 mm, c=34 mm) and a single right valve (ST120, l= ind, h=80 mm, c= ind), all collected from the yellow sandy marls of Ouled Fayet and Douera (unit c).
Description: This species bears rather thick, asymmetrical valves, with an upper (left) valve, flat or slightly concave in the umbo area, and a lower (right) curved valve slightly exceeding the left one on the edge. The dorsal-ventral length is always smaller than the antero-posterior one. Both valves bear wide radiating ribs (18 ribs on the left valve) characterised by their rounded section on the right valve and their quadrate section on the left valve, and by the occurrence of ridges in rib interspaces, numerous concentric undulations and fine striae. Growth lines are marked on both valves at interspaces, especially on the left valve. The auricles are equal and slightly costate. There is a slight byssal sinus and the ctenolium is missing.
Stratigraphic range: The species is reported in the northern Atlantic, from Norway through the Channel, and the Iberian Peninsula to Sudan. It is rather common in the Pliocene deposits of Almerěa (Jiménez et al., 2009).
Ecology: Pecten maximus lives freely on the surface of sandy and fine-gravelly sea bottoms to a depth of approximately -150 to -200 m. This species is preferentially found in calcarenites, median- and coarse-grained sandstones and conglomerates.
Pecten (Pecten) jacobaeus (Linnaeus, 1758)
(Fig. 6a ![]() )
)
1758 Ostrea Jacobea.- Linnaeus, X, p. 696. Lig., XXIV, pag. 58, Tav. XIII, figs. 1-10.
1902 Pecten jacobaeus Linnaeus.- Depéret & Roman, p. 58, Pl. V, fig. 1, 1a.
1907 Pecten jacobaeus.- Cerruli Irelli, I, p. 98, Tav. VI, fig. 14.a-b.
1957 Pecten jacobaeus (Linnaeus).- Perrodon, p. 207.
1975 Pecten jacobaeus (Linnaeus).- Feki, p. 32, Pl. V, fig. 5.
1987 Pecten jacobaeus (Linnaeus, 1758).- Kharrim, p. 90, Pl. 5, fig. 1.
2017 Pecten jacobaeus (Linnaeus, 1758).- Spadini, p. 10.
Material: Two, incomplete and thick, right valves (ST28, l=85 mm, h=80 mm, c= ind; ST70, l= ind, h= ind, c=50 mm) and a single inner mould (ST38, l=100 mm, h=98 mm, c=50 mm), all collected in the yellow sandy marls of Ouled Fayet and Douera (unit c).
Description: The collected specimens display an impressive size and are convex with a prominent umbo, but shorter than in P. bipartitus. The right valve bears 15 ribs with rectangular section, wider than the interspaces. Ribbings (4) run through the ribs and are lacking in the interspaces. Auricles are almost equal.
Stratigraphic range: This species appeared in the Pliocene, replacing P. grayi Michelotti, 1847. From the early Pliocene, it is found abundantly in the Atlantic and the Mediterranean Sea where it still lives nowadays (Ben Moussa, 1994).
Ecology: P. jacobaeus lives freely set on the sea bottom with its right valve and can swim actively. This is an exclusive species of the coastal detrital biocoenosis (Caulet, 1972).
|
Figure
6: Bivalve taxa from the Pliocene of the Algiers Sahel. a, Pecten jacobaeus (Linnaeus, 1758), specimen ST70; a1, external view; a2, internal view; b, Plicatula mytilina (Philippi,
1836), specimen ST310; b1, external view, left valve; b2, internal view, left valve; c, Flabellipecten alessii (Philippi,
1836), specimen ST27; d, Ostrea lamellosa (Brocchi,
1814), specimen ST21; d1, external view, right valve; d2, internal view, right valve; e, Ostrea edulis (Linnaeus, 1758), specimen ST6; e1, external view, right valve; e2, internal view, right valve. Scale bar= 10 mm for all. |
Genus Flabellipecten Sacco, 1897
Type species: Pecten alessii Philippi, 1836
Flabellipecten alessii (Philippi, 1836)
(Fig. 6c ![]() )
)
1910-1912 Flabellipecten alessii (Philippi, 1836).- Depéret et al., p. 141, Pl. XVIII, fig. 3, 3a.
2009 Flabellipecten alessii (Philippi, 1836).- Jiménez et al., p. 17, 15.
2017 Flabellipecten alessii (Philippi, 1836).- Spadini, p. 10.
Material: A single complete shell (ST27, l=125 mm, h=110 mm, c=99 mm) collected from the yellow sandy marls of Douera (unit c).
Description: The studied specimen is a complete, equivalve and equilateral shell, slightly wider than high. Both valves are weakly bulging, especially around the umbo, with a somewhat slightly flat left valve. The surface of this smooth shell displays fine ridges enhancing the position of ribs (about fifty) sometimes arranged in couplets. Their surface is ornamented by fine growth lines. Both auricles, devoid of jagged ridges, are almost identical and show a bundle of growth lines cut by radial ribs. The anterior auricle displays a shallow byssal sinus. Depéret & Roman (1910) considered F. alessii as a variant of F. flabelliformis (Brocchi, 1814) and described various intermediate morphologies between both species.
Stratigraphic range: Flabellipecten alessii was described by Philippi (1836) in the Pliocene of Castrogiovanni, Sicily. It also occurs in Liguria, in the Piacenzian of Bordighera, in the surroundings of Nice, in the Piacenzian of Villeneuve-Loubet and in the Astian of Asti, Modena, Zenzano and Calabria. In summary, the species is exclusively limited to Sicily, Italy and Nice, and can be found in the Piacenzian and Astian (Depéret & Roman, 1910-1912).
Family Plicatulidae Gray, 1854
Genus Plicatula Lamarck, 1801
Type species: Spondylus plicatus Linnaeus, 1758
Plicatula mytilina Philippi, 1836
(Fig. 6b ![]() )
)
1836 Plicatula mytilina.- Philippi, p. 86, Pl. 6, fig. 1.
1914 Plicatula mytilina Philippi.- Cossmann & Peyrot, Pl. 20, figs. 39-42.
1965 Plicatula mytilina Philippi.- Glibert & Poel, p. 45.
1988 Plicatula mytilina Philippi 1836.- Studencka & Studencki, p. 296.
2003 Plicatula mytilina Philippi.- Lozouet et al., p. 6, Pl. 14, figs. 1-6.
Material: Two right valves (ST301, l=30 mm, h=45 mm; ST310, l=31 mm, h=62 mm) collected from the yellow sandy marls of Ouled Fayet (unit c).
Description: Both valves are thick, flat, oval and dorso-ventrally elongated with a small, slightly opisthogyrous escutcheon. Irregular and rather spaced, growth lines are visible on the dorsal side. The inner surface displays a tiny, triangular, ligament area, base of which varies from 12 to 14 mm and shows an open, anterior-posteriorly elongated slit of some 5 mm. Vermiculate chomata are arranged outside the ligament area, along the anterior and posterior parts of the valve, much more visible in specimen ST301. The postero-anterior muscle scar in dorso-posterior position is subcircular, 7 mm in diameter and a bit more elongated dorso-ventrally in specimen ST310. Several other left valves, sometimes complete, are cemented on the outer surface of the right valve.
Stratigraphic range: Plicatula mytilina is reported from the Early Miocene to the Middle Pliocene (Studencka & Studencki, 1988).
Ecology: This is an epifaunal suspensivorous taxon preferring hard substrates.
Order Ostreoida Férussac, 1822 (emend.)
Suborder Ostreina Férussac, 1822 (emend.)
Superfamily Ostreacea Rafinesque, 1815
Family Ostreidae Rafinesque, 1815
Genus Ostrea Linnaeus, 1758
Subgenus Ostrea Linnaeus, 1758
Type species: Ostrea edulis Linnaeus, 1758
Ostrea (Ostrea) lamellosa offreti Kilian, 1889
(Fig. 6d ![]() )
)
1814 Ostrea lamellosa.- Brocchi, p. 564.
1897 Ostrea edulis Linnaeus var. lamellosa Brocchi.- Sacco, p. 7, Pl. H, figs. 3-4.
1889 Ostrea lamellosa Brocchi.- Kilian, p. 730, Pl. XVI, figs. 1-2.
1920 Ostrea edulis Linnaeus. var. boblayei Deshayes.- Dollfus & Dautzenberg, p. 462, Pl. 98, figs. 1-13; Pl. 101, fig. 1.
1965 Ostrea (s.s.) edulis Linnaeus lamellosa (Brocchi).- Glibert & Poel, p. 58.
1975 Ostrea lamellosa Brocchi.- Feki, p. 43, Pl. XII, fig. 5.
1988 Ostrea (Ostrea) lamellosa Brocchi, 1814.- Freneix et al., p. 5, Pl. I, fig. 5.
1994 Ostrea (Ostrea) edulis lamellosa Brocchi, 1814.- Ben Moussa, p. 116, Pl. 7, fig. 6.
2012 Ostrea lamellosa (Brocchi, 1814).- Satour, p. 72, Pl. 3, figs. 4-5.
Material: A complete shell (ST6, l=90 mm, h=110 mm) and two right valves (ST42, l=80 mm, h=90 mm; ST69, l=80 mm, h=90 mm), all collected in the yellow sandy marls of Ouled Fayet and Douera (unit c).
Description: The large-sized complete shell (ST6) consists of a left valve with dense concentric and undulated lamellae of loose radial folds, while the right (opercular) valve displays more or less spaced, concentric imbricated lamellae. The two right valves are of relatively large size, thicker with a flat shape, more elongated from umbo to outer margin with a slight posterior elongation without any ornamentation, with a gyrostreiform escutcheon, a tiny triangular, ligament area, ca. 35 mm in diameter, and with fine striae. The muscle scar, sub-triangular and slightly stretched posteriorly, is subcentral and of ca. 25 mm in diameter. The edge of the right valve is thick and folded. The outer surface bears strongly marked and non-homogenous, lamellar growth lines.
Stratigraphic range: The subspecies disappeared by the end of the Pliocene and was replaced by O. edulis edulis Linnaeus, 1758, still living today (Ben Moussa, 1994).
Ecology: This subspecies should have had the same environment as O. edulis edulis, which is found cemented to hard substrates, in oxygenated waters of the infralittoral zone, with quite variable salinity. It is suspensivorous, avoids very fine sediments and excessively turbid waters, and is adapted to various muddy, gravelly and rocky sea substrates, always immersed, ranging from -20 to -85 m in water depth. The water temperature must remain below 15°C in summer, the salinity being at least 25‰ (Laurain, 1984). However, it was less eurythermal than O. edulis, latitudinal distribution of which ranges from Norway to the Mediterranean Sea.
Ostrea edulis Linnaeus, 1758
(Fig. 6e ![]() )
)
1758 Ostrea edulis.- Linnaeus, p. 482, fig. 70.
1814 Ostrea edulis L.- Brocchi, p. 562.
1819 Ostrea edulina.- Lamarck, p. 218, no. 22.
1836 Ostrea edulis Linnaeus.- Deshayes, p. 244-245, no. 22 note 2.
2000 Ostrea edulis Linnaeus.- Goulven, p. 186.
2021 Ostrea edulis Linnaeus, 1758.- Benyoucef et al., p. 9, Fig. 6.C.
Material: A single complete shell (ST67, l=75 mm, h=95 mm) and two right valves (ST5, l=80 mm, h=85 mm; ST21, l=65 mm, h=60 mm), all collected from the yellow sandy marls of Ouled Fayet and Douera (unit c).
Description: The complete, rather large-size, shell displays flat and subcircular valves bearing imbricated, irregularly spaced concentric lamellae towards the outer edge, and having a folded and chalky structure. The robust escutcheon, of ostreiform type, displays on its inner side a triangular ligament area covered by fine growth lines; this latter consists of a deep median pit in between two ridges. The muscle scar, kidney-shaped and slightly stretched backwards, is subcentral.
Stratigraphic range: Ostrea edulis is known from the Pliocene to the Recent (Benyoucef et al., 2021). It is found from Norway to near Moroccan waters, along the Mediterranean shores and in the Black Sea. It is also reported in the Lower Chelif basin (NW Algeria).
Ecology: It dwells on sea bottoms not exceeding -40 m or ranges from the intertidal to the infralittoral zones. The adult forms can live attached to hard substrates (rocks, shell debris), but also on soft ones of sandy/muddy type (Global Biodiversity Information Facility, 2022).
Ostrea fimbriata Grateloup in Raulin & Delbos, 1855
(Fig. 7a ![]() )
)
1855 Ostrea fimbriata Grateloup.- Raulin & Delbos, p. 1158, Pl. 16, figs. 1-7; Pl. 17, figs. 1-4.
1914 Ostrea fimbriata Grateloup.- Cossmann & Peyrot, Pl. 19, figs. 19-21; Pl. 20, figs. 14-15.
1914 Ostrea (Cubitostrea) producta Raulin & Delbos.- Cossmann & Peyrot, Pl. 19, figs. 1-4.
1914 Ostrea (Ostreola) duvergieri.- Cossmann & Peyrot, Pl. 20, figs. 23-28.
2001 Ostrea fimbriata Raulin & Delbos.- Schultz, p. 353, Pl. 52, figs. 4.a-b, 5.a-b.
2002 Ostrea producta.- Rocher, p. 41, Pl. 10, fig. 13.
2003 Ostrea fimbriata (Grateloup, 1855).- Lozouet et al., p. 7, Pl. 16, figs. 1-7; Pl. 17, figs. 1-4.
2014 Ostrea fimbriata (Raulin & Delbos, 1855).- Križnar & Mikuž, p. 55.
Material: A single complete shell (ST36, l=84 mm, h=95 mm) and two left valves (ST1, l=115 mm, h=110 mm; ST4, l=70 mm, h=70 mm), hollow and of brownish colour, all collected from the yellow sandy marls of Douera (unit c).
Description: These shells are of rather large size, quite thick, with a slightly concave and subcircular shape. They display an opisthogyrous umbo of ostreiform type. The palleal edge of the costulated valve displays both crenulated ornamentation and edge, as well as lamellar growth lines. The ligament area is small and has a triangular shape. The umbonal cavity is missing. There is a large central, kidney-shaped muscle scar.
Stratigraphic range: O. fimbriata is only known at the Aquitanian.
Ecology: These individuals of O. fimbriata seem to be developed only on small-size hard substrates, separating them from soft substrates (fine sands, marls) or linked to the agitation of the environment (small pebbles, wood debris, etc.). The associated fauna is always littoral, with oligohaline affinity. On Mediterranean shores, we often use to collect individuals with aspect similar to that of O. fimbriata and assigned to the species O. edulis. Such a morphology is related to a series of environmental conditions: rather high temperature, normal salinity, water turbulence, and attachment on mobile substrate (Laurain, 1984).
|
Figure 7: Bivalve taxa from the Pliocene of the Algiers Sahel. a, Ostrea fimbriata
Grateloup in Raulin & Delbos,
1855, specimen ST1; a1, external view, left valve; a2, internal view, left valve; b, Ostrea stentina (Payraudeau,
1826), specimen ST225; b1, external view, left valve; b2, internal view, left valve; b3, detail of dorsal part; c, Saccostrea cucullata (Born,
1778), specimen ST215; c1, external view, left valve; c2, internal view, left valve; d, Neopycnodonte navicularis (Brocchi,
1814), specimen ST204; d1, external view, left valve; d2, internal view, left valve; e, Neopycnodonte cochlear (Poli,
1791), specimen ST68; e1, external view, left valve; e2, internal view, left valve. Scale bar= 10 mm for all. |
Ostrea stentina Payraudeau, 1826
(Fig. 7b ![]() )
)
1826 Ostreola stentina.- Payraudeau, Pl. I, figs. 6, 7.a-b.
1962 Ostrea stentina (Payr.).- Pasteur-Humbert, p. 37, Pl. 15, fig. 54.
1971 Ostrea (Ostrea) stentina (Payr.).- Stenzel, p. 1139, Fig. J 112, 1.a-e, 2.a-e.
1979 Lopha (Ostreola) stentina (Payr.).- Rosso, p. 472.
1985 Ostreola stentina (Payr.).- Harry, p. 142, Fig. 20.
Material: Four left valves (ST214, l=30 mm, h=42 mm; ST222, l=35 mm, h=50 mm; ST302, l=30 mm, h=50 mm; ST307, l=30 mm, h=40 mm) collected from the yellow sandy marls of Ouled Fayet (unit c).
Description: The collected valves are oval and rather small. They display undulated growth lines as more or less developed folds, lamellae of which are raised in places as spines. The folds build a characteristic serrated marginal edge. The inner side of the shell displays an umbonal cavity. The ligament area is rather narrow with a resilifer quite as wide as each bulge, except on specimen ST222. The escutcheon is slightly to strongly opisthogyrous. Vermiculate chomata are visible on both sides of the inner side of the shell and outside the ligament area. The rather large, adductor muscle insertion area is subcircular to kidney-shaped, and close to the dorsal margin.
Stratigraphic range: Ostrea stentina is known from the Neogene and mostly mentioned from the Quaternary in the Mediterranean basin. Nowadays it dwells the Mediterranean, the eastern Atlantic coast from Portugal down south to Angola, the western coast from North Carolina to Argentina, and the eastern Pacific coast from Alaska to Panama; but O. stentina is still unreported in the Indopacific domain, according to Harry (1985).
Ecology: O. stentina is a mostly infralittoral (from several metres down to -35 m) oyster, but can reach the upper limit of the continental shelf and cannot stand any salinity decrease; it dwells in the wide-open bays in tropical, subtropical and temperate regions (Freneix, 1988).
Saccostrea cucullata Born, 1778
(Fig. 7c ![]() )
)
1778 Ostrea cucullata.- Born, p. 114, Pl. 6, figs. 11-12.
1835 Ostrea echinata.- Quoy & Gaimard, p. 455, Pl. 76, figs. 13-14.
1931 Ostrea echinata.- Awati & Rai, p. 1-107.
1971 Saccostrea cucullata.- Stenzel, N1134-N1135, Fig. J106.
1985 Saccostrea cucullata.- Morris, p. 125-128, Pl. 3, figs. E-G.
2009 Saccostrea cucullata.- Lam & Morton, p. 110-112, Pls. 11-12.
Material: Three left valves (ST215, l=30 mm, h=30 mm; ST225, l=45 mm, h= ind; ST303, l=45 mm, h= ind) collected in the yellow sandy marls of Ouled Fayet (unit c).
Description: The collected left valves are of small size, slightly convex and of typical subtriangular shape. Only specimen ST215 is complete; specimens ST225 and ST303 are broken on their ventral side. The outer shape of the shells bears growth lines and undulations sometimes building spines at rib intersections. The shell edge is irregularly folded likewise the edge of the genus Lopha. The ligament area is small and of triangular shape, following the opisthogyrous escutcheon bend, and shows a crura at its lower base (specimen ST225). There is an umbonal cavity; the subcircular muscle scar, slightly dorsal-ventrally elongated (ca. 5 mm) is 15 mm in diametre. Vermiculate chomata are arranged outside the ligament area and are extending on the inner, anterior and posterior, parts of the valve.
Stratigraphic range: S. cucullata is a typical species of the Neogene of the Mediterranean Sea and reported in the Miocene and Pliocene sediments (Sacco, 1898) and in the Pleistocene of East Africa, the southern Arabic Peninsula, Java, Middle East, Syria, Egypt, Cyprus and Turkey. This species is also reported in the subtropical and tropical Indo-Pacific domain, southern Japan and Australia (Lam & Morton, 2009).
Ecology: S. cucullata dominates the eulittoral zone of the rocky shores like Hoi Ha Wan, Starfish Bay, Tso Woo Hang, Deep Bay, and Tai Tam Bay, and the mangroves like Ting Kok (Morton, 1990; Chiu, 1998). It is scarce on exposed shores like Cheung Sha and Cape Aguilar. This taxon also occurs on immersed and protection concrete structures of the East Ping Chau pier, and individually scattered in the barnacle environment of Big Wave Bay, Hong Kong islands.
Family Gryphaeidae Vyalov, 1936
Subfamily Pycnodonteinae Stenzel, 1971
Genus Neopycnodonte Stenzel, 1971
Type species: Neopycnodonte cochlear Poli, 1795
Neopycnodonte navicularis Brocchi, 1814
(Fig. 7d ![]() )
)
1814 Ostrea navicularis.- Brocchi, Pl. 565.
1897 Pycnodonte cochlear.- Sacco, p. 22, Pl. 8, figs. 1-14.
1965 Pycnodonte (Pycnodonte) cochlear navicularis (Brocchi).- Gilbert & Poel, p. 67.
1988 Neopycnodonte navicularis (Brocchi, 1814).- Freneix et al., p. 2, Pl. I, figs. 1, 2.a-b.
1994 Neopycnodonte navicularis (Brocchi, 1814).- Ben Moussa, p. 115, Pl. 7, figs. 4-5.
2012 Neopycnodonte navicularis (Brocchi, 1814).- Satour, p. 64, Pl. 2, figs. 13-14.
Material: Two complete shells (ST68, l=57 mm, h=70 mm, ST204 right valve, l=60 mm, h=70 mm; ST213 left valve, l=42 mm, h=60 mm), two left valves (ST172, l=67 mm, h=90 mm; ST208, l=40mm, h=60 mm) and one right valve (ST124, l=55 mm, h=65 mm), all collected from the yellow sandy marls of Ouled Fayet (unit c).
Description: These are thin or thick, large-sized shells of ovoid shape. The left valve is very concave (hollow in boat-shape), whereas the right valve is flat or slightly concave. The outer surface is smooth, sometimes slightly folded (specimen ST68) and bears concentric, lamellar growth lines. The escutcheon is slightly opisthogyrous. The ligament area, sometimes twinned (specimen ST208), of triangular shape, displays two lateral bulges defining a median resilium. Vermiculate chomata, when present (lacking in specimens ST172 and ST208), are situated outside the ligament area, along the lateral, anterior and posterior, ridges. The oval muscular scar of the left valve is located towards the posterior end. The shape of the adductor muscle scar of the right valve is much more circular. This latter bears an additional trace of adductor muscle as a gutter (specimen ST213) in the middle and below the ligament area.
Stratigraphic range: N. navicularis is known from the Early Miocene in the Mediterranean basin and the Paratethys to enter the Boreal domain (Belgium, Netherlands) during the Middle Miocene (Freneix et al., 1988). In the Pliocene, it is widespread in the Mediterranean Sea; it disappeared in the late Pliocene and was replaced by N. cochlear, this latter still living today.
Ecology: The taxon dwelled attached or free-lying on various sea bottoms in depths ranging from -50 and -300 m and in temperate waters of low turbidity (Malatesta, 1974).
Neopycnodonte cochlear (Poli, 1795)
(Fig. 7e ![]() )
)
1795 Ostrea cochlear.- Poli, p. 179, 26, p. iii (index).
1961 Pycnodonte cochlear (Poli).- Lecointre & Ranson, p. 29, Pl. 7, fig. 11.
1981 Pycnodonte cochlear (Poli).- Torigoe, p. 320, Pl. 9, figs. 1-7; Pl. 31, fig. 1.
2003 Neopycnodonte cochlear (Poli, 1791).- Videt, p. 22, Pl. 4, figs. 1-5.
2012 Neopycnodonte cochlear (Poli, 1791).- Satour, p. 67, Pl. 2, fig. 15.
Material: A single complete shell (ST61 right valve, l=42 mm, h=70 mm; ST62 left valve, l=46 mm, h=80 mm) and four left valves (ST158, l=34 mm, h=50 mm; ST194, l=40 mm, h=56 mm; ST196, l=45 mm, h=72mm; ST197, l=35 mm, h=45 mm; ST209, l=34 mm, h=45 mm) all collected from the yellow sandy marls of Ouled Fayet and Douera (unit c).
Description: This taxon displays the same morphological characteristics than its ancestor, Neopycnodonte navicularis. The oval shell, thick or thin, or even sometimes chalky (specimen ST97), is inequivalve with concave left valve and plano-concave right one. Growth lines are lamellar, irregular and rather tight on the anterior side. The escutcheon is opisthogyrous. The triangular, ligament area displays two lateral ridges defining a shallow, median, resilifer. Vermiculate chomata are arranged outside the ligament area, along the lateral, anterior and posterior, ridges of the valve. The adductor muscle scar is subcircular in posterior position, closer to the ligament area than the ventral side.
Stratigraphic range: N. cochlear is ubiquitous species, widespread in the Mediterranean Sea, eastern and western Atlantic shores, Pacific Ocean, Indian Ocean, and the Red Sea. It lives in deep waters along the continental margins and slopes. It is especially abundant in the lower part of the circalittoral zone and, more particularly, between -25 and -150 m, well cemented to hard substrates (Global Biodiversity Information Facility, 2022).
The Pliocene bivalve specimens of Algiers Sahel are weakly affected by boring and encrusting traces of organisms.
The bioerosive traces, observable on some bivalve specimens, are attributable to both endobionts and predators, and are mostly related to six ichnogenera:
6.1. Ichnogenus Caulostrepsis Clarke, 1908
Caulostrepsis taeniola Clarke, 1908
(Fig. 8a ![]() )
)
The figured specimens correspond to irregular, large-size, boring traces, with shape varying from simple, slightly curved galleries to U-shaped tubes with clavate (specimen ST403), rounded (specimen ST402) and sometimes single-pointed (specimen ST173) or double-pointed (specimen ST204) ends. This structure ranges from 2 to 17 mm in length by 1 to 4 mm in width. This ichnospecies, associated to Entobia isp., is preserved on both the outer and inner valve surfaces of Neopychnodonte cochlear, N. navicularis and Aequipecten angelonii. Caulostrepsis taeniola is a domichnion produced by several polychete families like siphonids (Parras & Casadio, 2005).
|
Figure 8: Bioerosion traces in Pliocene bivalves from the Algiers
Sahel. a, outer surface of the left valve of Neopycnodonte navicularis (Brocchi,
1814) showing Caulostrepsis taeniola (Clarke,
1908), specimen ST195; b, Entobia
isp. on the right valve of Ostrea lamellosa (Brocchi,
1814), specimen ST42; c-e, Oichnus simplex (Bromley,
1981) on left valve of Neopycnodonte navicularis (Brocchi,
1814), specimen ST41; left valve of Pecten bipartitus (Foresti,
1876), specimen ST117, right valve of Neopycnodonte cochlear (Poli, 1795), specimen ST173; f, left valve of Neopycnodonte cochlear (Poli, 1795) showing Gastrochaenolites torpedo (Kelly & Bromley,
1984), specimen ST197. Scale bar= 10 mm for all. |
6.2. Ichnogenus Entobia Bronn, 1838
Entobia isp.
(Fig. 8b ![]() )
)
This is the most abundant trace fossil. It is widely developed on the outer and some inner valve surfaces of Ostrea lamellosa (specimen ST42) and in articulated Neopycnodonte navicularis (specimen ST208). The genus Entobia is represented by a network of holes irregularly distributed, circular to elliptical and of variable size. The hole diameter ranges from 0.5 to 1 mm, devoid of any ornamentation. The drilling chambers do not penetrate the whole shell wall. In the present study we assign the trace material to Entobia isp., because it lacks diagnostic features. Borings assigned to the genus Entobia are produced by drilling sponges (Clionidae; Bromley & Tendal, 1973; Taylor & Wilson, 2003).
6.3. Ichnogenus Oichnus Bromley, 1981
Oichnus simplex Bromley, 1981
(Fig. 8c-e ![]() )
)
These are circular, subcircular to slightly oval borings, varying from 0.7 to 1 mm in diametre, perpendicular to sub-perpendicular to the outer surface of the right valves of the oyster O. edulis. Oichnus specimens are usually interpreted as Praedichnia (sensu Ekdale, 1985) of the gastropod families Naticidae and Muricidae respectively (Bromley, 1981; Pickerill & Donovan, 1998). O. paraboloides likely originates from naticid gastropods, whereas O. simplex is produced by muricid ones (Pickerill & Donovan, 1998).
6.4. Ichnogenus Gastrochaenolites Leymerie, 1842
Gastrochaenolites torpedo Kelly & Bromley, 1984
(Fig. 8f ![]() )
)
This ichnotaxon is less represented. It shows an elongate, claviform structure on the outer part of the left valve of Neopycnodonte cochlear (specimen ST197). It is 10 mm-long and 4 mm-wide.
Specimens of Gastrochaenolites are interpreted by Seilacher (1953) as Domichnia. Its origin refers usually to endolithic bivalves that bore mechanically or chemically shelly substrates such as Pholadidae, Gastrochaenidae and Mytilidae (Warme, 1975; Kelly & Bromley, 1984; Fischer, 1990a, 1990b). Gastropods of the genus Coralliophila and some sipunculids can also produce structures belonging to the ichnogenus Gastrochaenolites (Donovan, 2013). The oldest record of this ichnogenus is of Early Ordovician age (Ekdale & Bromley, 2001).
6.5. Ichnogenus Maeandropolydora Voigt, 1965
The genus Maeandropolydora differs from the genus Caulostrepsis (which appears as simple, U-shaped tube), by its complex and sinuous structure, as well as by the well-developed cylindrical shape of its galleries (e.g., Bromley & Alessandro, 1983).
Maeandropolydora sulcans Voigt, 1965
(Fig. 9a ![]() )
)
This ichnotaxon is particularly visible on the outer valve surfaces of Neopycnodonta cochlear and N. navicularis. It consists of one or several irregular, sinuous and sometimes coiled, cylindrical galleries, some 10 to 20 mm-long and 0.5 to 1 mm-wide. The ichnogenus Maeandropolydora is interpreted as resulting from the activity of annelid worms (Bromley & Alessandro, 1983).
6.6. Ichnogenus Pinaceocladichnus Mayoral,
1988 (Fig. 9b ![]() )
)
Visible on the left valve of Neopycnodonte navicularis (specimen ST402) as thin, regular, 1 mm-long, funnels with numerous bifurcations, giving a rhombic appearance. These traces are reported from the Oligocene to the Recent, and are assigned to ctenostome bryozoans (Mayoral, 1988).
|
Figure
9: Epibionts on Pliocene bivalves from the Algiers Sahel. a, outer surface of right valve of Neopycnodonte cochlear (Poli,
1795) showing traces of Maeandropolydora sulcans (Voigt,
1965), specimen ST402; b, traces of Pinaceocladichnus (Mayoral,
1988) on the same specimen ST402; c, external surface of the left valve of Ostrea fimbriata (Grateloup in Raulin & Delbos,
1855) encrusted with barnacles, specimen ST4; d, external surface of the right valve of Ostrea lamellosa encrusted with ferruginised
serpulids, specimen ST3; e, outer surface of right valve of Ostrea edulis (Linnaeus, 1758) encrusted with juvenile oyster shells, specimen ST15. Scale bar= 10 mm for all. |
Encrusting traces are seldom on Pliocene bivalve shells of Algiers Sahel. When available, they are usually concentrated on oyster specimens. Four types of encrusting organisms are identified among the study material:
7.1. Barnacles (Fig. 9c ![]() )
)
These are the most abundant encrusting traces on pteriomorph bivalve shells of Algiers Sahel. They belong to the genus Balanus and are well developed as aggregates on both the outer and inner valve surfaces of O. edulis (specimens ST4). Barnacles generally dwell any shallow marine settings (El-Sorogy et al., 2003).
7.2. Polychetes (Fig. 9d ![]() )
)
The first encrusting type is serpulid worms. Their preservation is lame and they scarcely occur on the outer surface of the left valves of O. lamellosa (specimen ST3) and O. edulis as thin limonitised tubes. Their dimensions range from 10 to 20 mm in length with 1 mm in diametre.
7.3. Juvenile oyster valves (Fig.
9e ![]() )
)
Numerous juvenile oyster valves are visible arranged in a circular pattern on the external side of the left valve. These juvenile valves represent several ontogenic stages, umbo-marginal dimensions range from 10 to 29 mm for specimen ST15, and from 5 to 10 mm for specimen ST4. The outer surface of N. cochlear (specimen ST15) and of O. edulis (specimen ST4) is encrusted by various ontogenetic stages of other disarticulated oyster specimens. As a rule, oysters are cementing on the substrate by their left valve in a downward-convex position (Benyoucef et al., 2021).
7.4. Bryozoans
Bryozoans represent the less abundant group among the studied encrusting organisms. They are mostly represented by whole colonies spreading from the outer part of the umbo of Neopycnodonte navicularis (specimen ST402). Bryozoans are sclerobionts attached to various hard substrates (Taylor, 1979, 2016; Breton, 2017).
The study of the Pliocene deposits of the Algiers Sahel in the surroundings of Douera and those of Ouled Fayet shows a relative abundance of bivalves (Fig. 2B) unevenly distributed among the brown (unit b) and yellow sandy marls (unit c).
8.1. Unit b
At the base of this succession, the brown sandy marls yielded small to medium-size bivalve shells, except for some valves of Amusium cristatum that can reach up to 10 cm in height and 10 cm in width (specimens ST162 and ST163) and build monospecific shell beds in some places. The bivalve fauna consists mostly of a pectinid assemblage dominated by: Amusium cristatum (35%), Pecten bipartitus (30%) and Aequipecten opercularis (20%), but also contains in decreasing order: Aequipecten bollenensis, A. radians, Mimachlamys varia, Aequipecten scabrella, Korobkovia oblonga, Pseudamussium clavatum, and Manupecten pesfelis. The gryphaeids are represented by Neopycnodonte navicularis. The highest fragmentation rate of bivalve shells within the brown marly sands of Ouled Fayet is observed on those of Amusium cristatum and Korobkovia oblonga due to the brittleness of their valves, with fragmentation occurring by breakage firstly on its periphery and secondly on the valve surface. The shell bed, represent biological concentrations because the low fragmentation, absence of abrasion and many shells preserve the patterns of the original colouration, disarticulated shells occur concave-up and down, and they are unaffected by encrusting organisms or boring (Kidwell, 1991). As for the rest of the bivalve assemblage, the specimens at hand display shells complete or slightly disarticulated with a weak to moderate fragmentation, supporting a preferential preservation within weak hydrodynamic settings. The wear of the pectinid bivalves is low with fragments showing non-blunt edges, non-altered colorations and slightly to non-deteriorated ornamentation, all that supporting a scarce post-mortem transport or even none. Furthermore, the occurrence of complete, non-worn and still connected, shells with populations showing various ontogenetic stages, and the presence of bioerosive traces attributable to both endobionts and predators (barnacles, gastropods) on the outer surface of valves, all document a very short transport from the original life habitat to the fossilisation/burial site; subsequently, these assemblages are direct evidence of original environmental conditions. Likewise drilling and biogenic encrusting are abundant in depositional settings with low sedimentation rate and indurated sea bottom (Lescinsky et al., 2002).
The pectinids (Pecten bipartitus, Aequipecten opercularis, A. bollenensis, A. radians, Mimachlamys varia, and Aequipecten scabrella) occur indifferently in both calm and agitated environments, on soft or hard substrates, and dwell all kind of marine settings from the infralittoral down to the circalittoral zones (Freneix et al., 1987, 1988; Saint Martin et al., 2000; Lacour et al., 2002). In the environments of the Mediterranean Sea, Pseudamussium clavatum and Manupecten pesfelis mostly characterise the detrital, sandy and gravelly, sea bottoms of the circalittoral zone (Pérčs & Picard, 1964).
Large-size, vagile and epibenthonic, specimens of Amusium cristatum and of Korobkovia oblonga suggest calm, muddy sea bottoms of the circalittoral zone; likewise, the oyster Neopycnodonte navicularis is a typical form of circalittoral sea bottoms, of depth ranging from -50 to -100 m (Freneix et al., 1988; Lacour, 1999; Saint Martin et al., 2000; Lacour et al., 2002; Videt, 2003).
8.2. Unit c
The yellow marly sands of both localities of Douera and Ouled Fayet, yield a bivalve assemblage dominated by large-size specimens such as Flabellipecten alessii, Ostrea fimbriata and Spondylus crassicosta. Other associated pteriomorph bivalves are: Pecten maximus, P. jacobaeus, Glycymeris bimaculata, Ostrea lamellosa, O. edulis, and Neopycnodonte cochlear.
Small-size pectinids are represented by Talochlamys multistriata, Mimachlamys varia, Aequipecten angelonii, A. scabrella, A. bollenensis, and Amusium cristatum. They form assemblages with Ostrea stentina, O. cuculatta, Anadara diluvii, Nucula sp., and Plicatula mytilina.
The state of preservation of shells within the yellow sandy marls of Douera and Ouled Fayet is relatively fair, with complete shells showing articulated, or slightly shifted, valves. The abrasion of pectinid valves is faint to nil with non-altered colours and ornamentation in excellent state of preservation (spines, ribs). All sampled fragments show non-blunt angles. Among the family Ostreidae, fragmentation is mainly observed in the marginal area and occurs as layered structures of the valves, regardless environmental hydrodynamics but rather related to intrinsic shell conditions. The assemblage of small-size pectinids such as Talochlamys multistriata, A. angelonii, A. scabrella, A. bollenensis, and Mimachlamys varia, indicate an infralittoral to bathyal zone. Whereas T. multistriata is frequent within depths ranging from -10 and -100 m, A. angelonii belongs to the group of species craving agitated waters (Ben Moussa, 1994). G. bimaculata dwells the fine to coarse sandy sea bottoms of the infralittoral zone under the influence of bottom currents. The occurrence of Spondylus crassicosta in this level indicates a median infralittoral type environment. Plicatula mytilina is an epifaunal, suspensivorous species dwelling hard substrates, likewise Ostrea cucullata dwelling the eulittoral zone of rocky shores. Ostrea stentina is essentially an infralittoral and intertidal oyster, ranging from several metres down to -35 m.
Large-size pectinids are represented by Pecten maximus, P. jacobaeus and Flabellipecten alessii. Like in present-day, these scallops live free-lying on the surface of sandy to fine-to-coarse gravel substrates. Ostrea lamellosa dwelled the intertidal to infralittoral zones down to -40 m water depth. This species records the last marine to brackish influences in the studied sections, considering that it could withstand temporary salinity decrease of at least 25‰ (Freneix et al., 1988). O. fimbriata is associated to a littoral fauna with oligohaline affinity. N. navicularis, replaced by N. cochlear, usually dwells water depths ranging from -25 to -150 m, plainly attached to hard substrates.
The assemblage of large-size bivalves characterises the shallow inner shelf environments under the influence of high hydrodynamics and deposition of coarse detrital sediments; it corresponds to the thriving of large-size Pectinidae, Gryphaeidae and Ostreidae in environmental settings situated in the shallowest part of the infralittoral zone.
The bivalve assemblage within the yellow sandy marls suggests a shallow, agitated, marine setting, with some 'washover' influence evidenced by the occurrence of Spondylus crassicosta (Lacour et al., 2002). These observations are supported by the frequency of boring and encrusting traces mostly affecting large-size bivalves: the ichnogenus Entobia is strongly developed on the outer valve surfaces of Ostrea lamellosa and Neopycnodonte navicularis; Oichnus simplex is developed on the outer surfaces of O. edulis and Pecten bipartitus; Caulostrepsis taeniola and Maeandropolydora sulcans are mostly observed on the outer valve surface of Neopychnodonte cochlear and N. navicularis; Gastrochaenolites torpedo and Pinaceocladichnus occur on the outer valve surface of Neopychnodonte navicularis. As a whole, these observed fossil traces would indicate a shallow marine setting with well-oxygenated waters.
Bivalves form the main constituent of the fossil assemblages of the Pliocene formations of the Algiers Sahel. In this present study, 27 pteriomorph bivalve taxa, among which 13 are exclusively fossil species without modern representatives, six bioerosive ichnogenera and four encrusting ichnogenera, are identified. Some of these bivalve species (Aequipecten angelonii, A. radians, A. bollenensis, Pecten bipartitus, Plicatula mytilina, Korobkovia oblonga, Ostrea fimbriata, O. stentina, O. cucullata, and O. navicularis) are described for the first time, supplementing the early (preliminary) inventory list of Pliocene bivalves of Algeria. The biodiversity of bivalve fauna in platform environments is expressed through two units: the first shallow infralittoral unit, is characterised by large Flabellipecten alessii, Aequipecten angelonii, Ostrea lamellosa and O. edulis; whereas the second deeper unit, of circalittoral environment, comprises Amusium cristatum, Pseudamussium clavatum, Mimachlamys varia, and Manupecten pesfelis. The Amusium cristatum shell beds represent biological concentrations.
Taphonomical data evidence short transport with faint valve wear and slight abrasion of ornamentation. Analysis of boring and encrusting traces, mostly observed on large-size bivalves, would indicate a progressive environmental change from a high energy setting to deeper water conditions with lower energy, and correspond to a rather shallow sea with well-oxygenated waters.
The authors wish to express their most sincere gratitude to Pr Beatriz Bádenas (the Associate Editor) for her kind assistance in the editorial process, to Dr Stephen Eagar (the Language Editor) for his valuable assistance in ironing up the final version of this manuscript and to the Reviewers, Pr Pedro Callapez and Dr Pierre Lozouet, for their constructive advice and highlighting comments.
Ardovini R. & Cossignani T. (1999).- Atlante delle conchiglie di profonditŕ del Mediterraneo.- L'Informatore Piceno, Ancona, 111 p.
Arnould Locart E.A. (1888).- Monographie des espčces appartenant au genre Pecten.- Publications de la Société Linnéenne de Lyon, no. 34, p. 133- 287.
Awati P.R. & Rai H.S. (1931).- The Indian Zoological Memoirs on Indian Animal Types: III Ostrea cucullata (The Bombay Oyster).- Methodist Publishing House, Lucknow, 107 p.
Aymé A. (1948).- Contribution ŕ l'étude des terrasses marines entre le Cap-Matifou et l'Oued Isser.- Afrique du Nord, Alger, vol. 39, p. 97-100.
Aymé A., Aymé J.- M. & Magné J. (1953).- Étude des terrains néogčnes de la cluse du Mazafran (Sahel d'Alger), Algérie.- Publication du Service de la carte géologique de l'Algérie (nouvelle série), Alger, Bulletin des travaux des collaborateurs, no. 1, fasc. II, p. 129-150.
Aymé A. (1955).- Contribution ŕ l'étude de la plaine de la Mitidja occidentale et de sa bordure atlantique : Origine des eaux alcalines-accidents tectoniques-Séismicité de la région de Blida, Algérie.- Publication du Service de la carte géologique de l'Algérie (nouvelle série), Alger, Bulletin des travaux des collaborateurs, no. 8, p. 347-362.
Aymé A. (1963).- Carte géologique d'Algérie au 1/50 000, feuille no 21 d'Alger.- Service de la carte géologique de l'Algérie, Alger.
Ben Moussa A. (1994).- Les bivalves néogčnes du Maroc septentrional (façades atlantique et méditerranéenne) : Biostratigraphie, paléobiogéographie et paléoécologie.- Documents des Laboratoires de Géologie de Lyon, Villeurbanne, no. 132, 281 p.
Benyoucef M., Bendella M., Brunetti M., Ferré B., Koči T., Bouchemla F., Slami R. & Ghenim A.F. (2021).- Upper Pliocene bivalve shell concentrations from the Lower Chelif basin (NW Algeria): Systematics, sedimentologic and taphonomic framework.- Annales de Paléontologie, Paris, vol. 107, no. 4, p. 1-23.
Beurlen K. (1944).- Beiträge zur Stammensgeschichte der Muscheln.- Bayerische Akademie Wissenschaft, Sitzungsberichten, München, vol. 1-2, p. 133-145.
Bongrain M. (1970).- Pectinidés des Faluns de la Loire : Le groupe de Chlamys opercularis dans le bassin de Noyant-sous-le-Lude.- Travaux du Laboratoire de Paléontologie, Faculté des Sciences d'Orsay, Université Paris XI, 73 p.
Born I. (1778).- Index Rerum Naturalium Musei Caesarei Vindobonensis, pars prima, Testacea.- Officina Krausiana, Vienna, 442 p. URL: https://www.biodiversitylibrary.org/page/12009661
Breton G. (2017).- The sclerobionts of the Upper Cenomanian oysters from Le Mans (Sarthe, France).- Annales de Paléontologie, Paris, vol. 103, p. 173-183.
Brocchi G. (1814).- Conchiologia fossile subappennina con osservazioni geologiche sugli Appennini e sul suolo adiacente. 2.- Stamperia Reale, Milano, p. 241-712. URL: https://www.biodiversitylibrary.org/page/12009099
Bromley R.G. (1981).- Concepts in ichnotaxonomy illustrated by small round holes in shells.- Acta Geologica Hispanica, Barcelona, vol. 16, p. 55-64. URL: https://revistes.ub.edu/index.php/ActaGeologica/article/view/4222/5155
Bromley R.G. & Alessandro A. d' (1983).- Bioerosion in the Pleistocene of Southern Italy. Ichnogenera Caulostrepsis and Maeandropolydora.- Rivista Italiana di Paleontologia e Stratigrafia, Roma, vol. 89, p. 283-309.
Bromley R.G. & Tendal O.S. (1973).- Example of substrate competition and phototropism between two clionid sponges.- Journal of Zoology, vol. 169, no. 2, p. 151-155.
Bronn H.G. (1827).- Ergebnisse meiner naturhistorisch-öconomischen Reisen. Erster Theil. Briefe aus der Schweiz, Italien und Südfrankreich, im Sommer 1824.- Karl Groos, Heidelberg, xxii + 654 p. (VII Pls.).
Bronn H.G. (1838).- Lethaea geognostica: oder, Abbildungen und Beschreibung der für die Gebirgs-Formationenbezeichnendsten.- E. Schweizerbart, Stuttgart, 545 p.
Cabot J. & Prenant A. (1968).- Observations sur le Quaternaire dans le Sahel d'Alger.- Annales algériennes de géographie, Alger, vol. 3, no. 5, p. 77-92.
Caprotti E. (1972).- I Bivalvi dello stratotipo Piacenziano (Castell Arquato, Piacenza).- Natura, Milano, vol. 63, no. 1, p. 47-86.
Caulet J. (1972).- Les sédiments organogčnes du Précontinent algérien.- Mémoires du Muséum national d'Histoire naturelle, série C (Sciences de la Terre), Paris, 289 p.
Cerulli-Irelli S. (1907-1909).- Fauna malacologia mariana.- Palaeontographia Italica, Pisa, vol. 14 (1908), p. 1-63; vol. 15 (1909), p. 125-213.
Choffat P. & Dollfus G. (1905).- Quelques cordons littoraux marins du Pleistocčne du Portugal.- Communicaçőes du Service Géologique du Portugal, Lisboa, vol. 6, p. 158-173.
Chiu H.M.C. (1998).- Population dynamics of Saccostrea cucullata (Bivalvia: Ostreidae) from five shores in Hong Kong.- Asian Marine Biology, Hong Kong, vol. 15, p. 73-103.
Cocconi G. (1873).- Enumerazione sistematica dei molluschi miocenici e pliocenici delle provincie di Parma e di Piacenza.- Memorie della Accademia delle Scienze dell'Instuto di Bologna, vol. 3, no. 3, p. 409-776.
Clarke J.M. (1908).- The beginnings of dependent life.- Bulletin New York State Museum, vol. 121, p. 146-196.
Cossmann M. & Peyrot A. (1909-1914).- Conchologie néogénique de l'Aquitaine. Pélécypodes.- Actes de la Société Linnéenne de Bordeaux, t. I, vol. 1, p. 1-220, 1909 (LXIII); t. I, vol. 2, p. 221-428, 1911 (LXIV); t. I, vol. 3, p. 429-714, 1912 (LXV); t. II, vol. 1, p. 1-204, 1913 (LXVI); t. II, vol. 2, p. 205-410, 1914 (LXVII); t. III et suppl., p. 411-496, 1914 (LXVIII). URL: https://www.biodiversitylibrary.org/bibliography/165529
Costa E.M. da (1778).- Historia naturalis testaceorum Britanniae, or, the British conchology; containing the descriptions and other particulars of natural history of the shells of Great Britain and Ireland. I-XII.- privately published, London, 254 p. URL: https://www.biodiversitylibrary.org/bibliography/13179
Courville P. & Bongrain M. (2003).- Les Pectinida miocčnes des faluns (Ouest de la France). Intéręts biostratigraphiques des associations.- Annales de Paléontologie, Paris, vol. 89, no. 3, p. 125-151.
Dall W.H. (1889).- A preliminary catalogue of the shells bearing marine mollusks and Brachiopoda of the Southeastern coast of the United States, with illustrations of many species.- United States National Museum Bulletin, Washington - DC, vol. 37, 221 p.
Dall W.H. (1895).- Synopsis of a review of the genera of Recent and Tertiary Mactridae and Mesodesmatidae.- Malacological Society of London, Proceedings, vol. 1, no. 5, p. 203-213.
Dalloni M. (1934).- Le Pliocčne du Sahel oriental et de la Kabylie.- Bulletin de la Société d'Histoire Naturelle de l'Agrique du Nord, Alger, t. XXIV, p. 9-18.
Dalloni M. (1954).- Sur quelques problčmes du Quaternaire méditerranéen.- Bulletin de la Société d'Histoire Naturelle d'Afrique du Nord, Alger, t. XLV, nos. 3-4, p. 134-169.
Delage M. (1888).- La géologie du Sahel.- L'association générale des étudiants des chroniques de Languedoc, Montpellier, 216 p.
Demarcq G. (1979a).- Quelques observations paléoécologiques comparées sur des Pectinidés du Néogčne méditerranéen. In: VIIth intern. R.C.MJV.S. Congr., Athčnes.- Annales Géologiques des Pays Helléniques (h.s.), Athčnes, no. 1, p. 295-304.
Depéret C. & Roman F. (1902-1928).- Monographie des Pectinidés néogčnes de l'Europe et des régions voisines.- Mémoire de la Société Géologique de France, Paris, Mémoire 26, Genre Pecten, t. X, fasc. 1, 1902, p. 1-74, Figs. 1-33, Pls. I-VIII ; Genre Pecten, t. XIII, fasc. 2, 1905, p. 75-104, Figs. 34-43, Pls. IX-XI ; Genre Flabellipecten, t. XVIII, fasc. 2, 1910, p. 105-138, Figs. 44-60, Pls. XII-XVII ; Genre Flabellipecten, t. XIX, fasc. 1, 1912, p. 139-168, Figs. 61-71, Pls. XVIII-XXIII ; Genre Amussium, t. IV (n. sér.), 1928, p. 169-194, Pls. XXIV-XXVIII.
Deshayes G.P. (1836).- Les Mollusques. In: G. Cuvier (ed.), Le rčgne animal distribué d'aprčs son organisation, pour servire de base ŕ l'histoire naturelle des animaux, et d'introduction ŕ l'anatomie comparée.- Masson, Paris, 266 p.
Dijkstra H.H. & Goud J. (2002).- Pectinoidea (Bivalvia, Propeamussiidae and Pectinidae) collected during the Dutch Cancap and Mauritania expeditions in the south-eastern region ofthe North Atlantic Ocean.- Basteria, Amsterdam, vol. 66, p. 31-82.
Djediat Y. (2013).- Le Domaine margino-littoral algérois. Organisation et évolution néogčne et quaternaire, implications géotechniques et prospects pour une occupation rationnelle des espaces.- Thčse Doctorat Université des Sciences et de la Technologie Houari Boumedične, Alger, 106 p.
Dollfus G. & Dautzenberg P. (1902-1920).- Conchyliologie du Miocčne moyen du Bassin de la Loire. I: Pélécypodes.- Mémoires de la Société Géologique de France, Paléontologie, Paris, vol. 27, no. 1, p. 1-106, Pls. 1-5 ; 1902, no. 2, p. 107-162, Pls. 6-10 ; 1904, no. 3, p. 163-240, Pls. 11-15 ; 1906, no. 4, p. 241-296, Pls. 16-22 ; 1909, no. 5, p. 297-378, Pls. 23-33 ; 1913, no. 6, p. 379-500, Pls. 34-51.
Donovan S.K. (2013).- Neoichnology of the parish church of All Saints, Freshwater, Isle of Wight. Wight studies.- Proceedings of the Isle of Wight Natural History and Archaeological Society, Newport, vol. 27, p. 70-75.
Durand Delga M. (1971).- Les unités mésozoďques métamorphiques d'El Milia ŕ Texena (Algérie) et leur cadre structural.- Bulletin de la Société Géologique de France (7čme série), Paris, vol. 13, p. 328-377.
Ekdale A.A. (1985).- Paleoecology of the marine endobenthos.- Palćogeography Palćoclimatology, Palćoecology, vol. 50, p. 63-81.
Ekdale A.A. & Bromley R.G. (2001).- Bioerosional innovation for living in carbonate hardgrounds in the Early Ordovician of Sweden.- Lethaia, Oslo, vol. 34, p. 1-12.
El-Sorogy A.S., Ziko A., Saber N. & Nour H.E. (2003).- The most common rocky shore invertebrate dwellers of the Red Sea Coast.- Egyptian Journal of Paleontology, Cairo, vol. 3, p. 271-284.
Feki M. (1975).- Paléoécologie du Pliocčne marin au Nord de la Tunisie.- Annales des Mines et de la Géologie, no. 27, Thčse Sciences, Paris, 195 p.
Férussac A.É. d'A. de (1822).- Tableaux systématiques des animaux mollusques classés en familles naturelles, dans lesquels on a établi la concordance de tous les systčmes ; suivis d’un prodome général pour tous les mollusques terrestres ou fluviatiles, vivants ou fossiles.- Arthus Bertrand, Paris, 110 p. DOI: https://doi.org/10.5962/bhl.title.10558
Fischer P.H. (1886).- Manuel de Conchyliologie et de Paléontologie Conchyliologique, ou Histoire Naturelle des Mollusques Vivants et Fossiles suivi d'un Appendice sur les Brachiopodes par D.P. Oehlert. Avec 23 planches contenant 600 figures dessinées par S.P. Woodward.- F. Savy, Paris, fasc. 10, p. 897-1008. Published in 11 parts (fascicules), xxiv + 1369 p. (1138 Figs., 23 Pls.).
Fischer R. (1990a).- Significado paleoecologico y geologico de perforaciones fosiles de bivalvos.- Revista de la Sociedad Mexicana de Paleontologia, Mexico City, vol. 3, p. 79-95.
Fischer R. (1990b).- Paläoökologische und geologische Bedentungfossiler Muschelbohrungen.- Arbeitskreis Palaontologie, Hannover, vol. 18, p. 1-28.
Fontannes F. (1876).- Études stratigraphiques et paléontologiques pour servir a l'histoire de la Période Tertiaire dans le Bassin du Rhone. II. Les terrains Tertiaires Supérieurs du Haut Comtat-Venaissin.- F. Savy, Paris, 97 p. URL: https://gallica.bnf.fr/ark:/12148/bpt6k5439003t.texteImage
Fontannes F. (1879-1882).- Les Mollusques pliocčnes de la vallée du Rhône et du Roussillon. (Les Invertébrés du bassin tertiaire du sud-est de la France). T. II. Acéphales des formations marines et saumâtres.- F. Savy, Paris, 321 p.
Foresti L. (1876).- Cenni geologici e paleontologici sul Pliocene antico di Castrocaro.- Memorie della Accademia delle Scienze dell'Istituto di Bologna (Serie Terza), vol. 6, p. 521-574.
Freneix S., Saint Martin J.-P. & Moissette P. (1987).- Bivalves Ptériomorphes du Messinien d'Oranie (Algérie occidentale).- Bulletin du Muséum National d'Histoire Naturelle (4čme série), Paris, Section C1, vol. 9, p. 3-61.
Freneix S., Saint Martin J.-P. & Moisette P. (1988).- Huîtres du Messinien d'Oranie (Algérie occidentale) et Paléobiologie de l'ensemble de la faune de Bivalves.- Bulletin du Muséum National d'Histoire Naturelle, Paris, (4čme série), section C, vol. 10, no. 1, p. 1-21.
Glibert M. (1945).- Faune malacologique du Miocčne de la Belgique.- Mémoire du Musée Royal d'Histoire Naturelle de Belgique, Bruxelles, no. 103, 266 p.
Glibert M. & Poel L. van de (1965-1970).- Les Bivalves fossiles du Cénozoďque étranger des collections de l'Institut Royal des Sciences Naturelles de Belgique.- Mémoire du Musée Royal d'Histoire Naturelle de Belgique, Bruxelles, vol. 77 (1965), 112 p. ; vol. 78, 105 p. ; vol. 81 (1966), vol. 82 p. ; vol. 82, 108 p. ; vol. 83 (1967), 152 p. ; vol. 84 (1970), 185 p.
Glangeaud L. (1932).- Étude géologique de la région littorale de la province d'Alger.- Publication du Service de la Carte Géologique de l'Algérie (2čme série), Alger, no. 8, 627 p.
Glangeaud L. (1936).- Coupes structurales du Nord-Ouest de la Province d'Alger au 1/200 000.- Publication du Service de la Carte Géologique de l'Algérie, Alger, no. 8 bis.
Glangeaud L., Aymé A., Caire A., Mattauer M. & Muraour P. (1952).- Histoire géologique de la province d'Alger.- Publications du XIX Congrčs Géologique International, Alger, Monographies Régionales (1čre Série), vol. 25, 142 p.
Glangeaud L. (1968).- Les méthodes de la géodynamique et leurs applications aux structures de la Méditerranée Occidentale.- Revue de Géographie Physique et de Géologie Dynamique, Paris, vol. 10, p. 83-135.
Global Biodiversity Information Facility (2022).- URL: https://www.gbif.org/
González Donoso J. & Porta J.D. (1977).- Datos preliminares sobre afloramiento de materiales pliocenicos en Estepona (Provincia de Malaga).- Studia geologica Salmanticensia, Salamanca, vol. 13, p. 35-57.
Goulven L. (2000).- Paléontologie(s) et évolution au début du XIXe sičcle. Cuvier et Lamarck.- Asclepio, Madrid, vol. 52, no. 2, p. 133-212. DOI: https://dx.doi.org/10.3989/asclepio.2000.v52.i2.211
Gray J.E. (1824).- Supplement to the appendix of Captain Parry's voyage for the discovery of a North-West Passage, in the years 1819-20, containing an account of the subjects of Natural History, Shells.- John Murray, London, p. 240-246. URL: https://www.biodiversitylibrary.org/page/53407756
Gray J.E. (1825).- A list and description of some species of shells not taken notice of by Lamarck.- Annals of Philosophy (New Series), London, vol. 9, p. 134-140.
Gray J.E. (1847).- A list of the genera of Recent Mollusca, their synonyma and types.- Proceedings of the Zoological Society of London, vol. 15, p. 129-219.
Gray J.E. (1854).- On the division of ctenobranchous gasteropodous Mollusca into larger groups and families.-Proceedings of the Zoological Society of London, vol. 21, p. 32-44.
Harry H.W. (1985).- Synopsis of the supraspecific classification of living Oysters (Bivalvia: Gryphaeidae and Ostreidae).- The Veliger, Berkeley, vol. 28, no. 2, p. 21-158.
Jiménez A.P., Aguirre J. & Rivas P. (2009).- Taxonomic study of scallops (Pectinidae: Mollusca, Bivalvia) from Pliocene deposits (Almería, SE Spain).- Revista Espańola de Paleontología, Madrid, vol. 24, no. 1, p. 1-30. DOI: https://dx.doi.org/10.7203/sjp.24.1.20338
Kelly S.R.A. & Bromley R.G. (1984).- Ichnological nomenclature of clavate borings.- Palaeontology, Cambridge (UK), vol. 27, p. 793-807.
Kharrim M.R. (1987).- Pectinidés du Néogčne du bassin de Boudinar (Rif oriental, Maroc).- Thčse 3čme Cycle, Lyon 1, 251 p.
Kidwell S.M. (1991).- The stratigraphy of shell concentrations. In: Allison P.A. & Briggs D.E.G. (eds.), Taphonomy: Releasing the data locked in the fossil record.- Plenum Press, New York – NY, vol. 9, p. 211- 290.
Kilian M. (1889).- Mission d'Andalousie. II - Études paléontologiques sur les terrains secondaires et tertiaires de l'Andalousie.- Mémoires de l'Académie des Sciences de Paris, t. XXX, p. 601-750. URL: https://gallica.bnf.fr/ark:/12148/bpt6k9322916
Križnar M. & Mikuž V. (2014).- Lipovica quarry and its interesting paleontological characteristics.- Scopolia, Journal of the Slovenian Museum of Natural History, Ljubljana, no. 82, 120 p.
Iredale T. (1929).- Strange molluscs in Sydney Harbour.- Australian Zoologist, Sidney, vol. 5, no. 4, p. 337-352.
Lacour D. (1999, unpublished).- Évolution de la biodiversité des échinides et des bivalves méditerranéens du Miocčne terminal ŕ l'Actuel : Application au Messinien du bassin de Sorbas (Sud-Est de l'Espagne).- D.E.A., Université Aix-Marseille 1, Marseille, France, 50 p.
Lacour D., Lauriat-Rage A., Saint Martin J.-P., Videt B., Néraudeau D., Goubert E. & Bongrain M. (2002).- Les associations de bivalves (Mollusca, Bivalvia) du Messinien du bassin de Sorbas (SE Espagne).- Geodiversitas, Paris, vol. 24, no. 3, p. 416-657. URL: https://sciencepress.mnhn.fr/sites/default/files/articles/pdf/g2002n3a6.pdf
Lam K. & Morton B. (2009).- Oysters (Bivalvia: Ostreidae and Gryphaeidae) recorded from Malaysia and Singapore. University of Singapore.- The Raffles Bulletin of Zoology, Singapore, vol. 2, p. 481-494.
Lamarck J.-B.P. A. de M. de (1799).- Prodrome d'une nouvelle classification des coquilles, comprenant une rédaction appropriée des caractčres génériques, et l'établissement d'un grand nombre de genres nouveaux.- Mémoires de la Société d'Histoire Naturelle de Paris, Paris, 1, p. 63-91.
Lamarck J.-B.P. A. de M. de (1805).- Suite des mémoires sur les fossiles des environs de Paris.- Annales du Muséum d'Histoire Naturelle, Paris, vol. 6, p. 117-126, p. 214-221, p. 337-345, p. 407-415.
Lamarck J.-B.P. A. de M. de (1809).- Philosophie zoologique, ou exposition des considérations relatives ŕ l'histoire naturelle des Animaux ; ŕ la diversité de leur organisation et des facultés qu'ils en obtiennent ; aux causes physiques qui maintiennent en eux la vie et donnent lieu aux mouvemens qu'ils exécutent ; enfin, ŕ celles qui produisent, les unes le sentiment, et les autres l'intelligence de ceux qui en sont doués.- Muséum d'Histoire Naturelle, Paris, t. II, 473 p.
Lamarck J.-B.P. A. de M. de (1819).- Histoire naturelle des animaux sans vertčbres.- Paris, t. VI, 1čre partie, 345 p. URL: https://www.biodiversitylibrary.org/page/41659588
Laurain M. (1984).- Bivalves ostréidés.- Nouvelles archives du Muséum d'Histoire naturelle de Lyon, vol. 22, p. 74-78. URL: https://dx.doi.org/10.3406/mhnly.1984.1658
Lauriat-Rage A. (1989).- Les Bivalves du Pliocčne de Normandie.- Bulletin du Muséum national d'Histoire Naturelle (4čme série), Paris, sect. C, vol. 1, p. 3-51.
Lecointre G. (1952).- Recherches sur le Néogčne et le Quaternaire marin de la côte atlantique du Maroc. Paléontologie.- Notes et Mémoires du Service Géologique du Maroc, Rabat, vol. 99, 173 p.
Lecointre G. & Ranson G. (1961).- Ostrea offreti Kilian, 1889 du Miocčne circumméditerranéen.- Bulletin de la Société Géologique de France (7čme série), Paris, vol. 3, p. 288-289.
Lescinsky H.L., Edinger E. & Risk M.J. (2002).- Mollusc shell encrustation and bioerosion rates in a modern epeiric sea: Taphonomy experiments in the Java Sea, Indonesia.- Palaios, Lawrence - KS, vol. 17, p. 171-191.
Linnaeus C. (1758-1767).- Systema naturae, per regna tria naturae, secundum classes, ordines, genera, species cum characteribus differentilis synonumis, locis,…- Typis Ioannis Thomae, Vienna, t. I, part. I, (1758), 824 p.; t. I, part II, (1767), p. 533-1328. URL: https://www.biodiversitylibrary.org/bibliography/559
Lozouet P., Lesport J.- F., Favia R., Renard P. & Rocher P. (2003).- Les bivalvia de l'Aquitainien de Saucats "Lariey" (Gironde). I.- Pteriomorpha.- Cossmanniana, Paris, vol. 9, no. 1-4, p. 1-45.
Malatesta A. (1974).- Malacofauna pliocenica Umbra.- Cossmanniana, Paris, vol. 13, 498 p.
Mayer-Eymar C. (1868).- Catalogue systématique et descriptif des fossiles de terrains tertiaires qui se trouvent au Musée fédéral de Zürich, vol. 3. Mollusques Famille des Arcides.- Librairie Schabelitz, Zurich, 124 p.
Mayoral E. (1988).- Pennatichnus nov. icnogen., Pinaceocladichnus nov. icnogen. e Iramena. Huellas de bioerosión debidasa Bryozoa perforantes (Ctenostomata, Plioceno inferior) en la cuenca del Bajo Guadalquivir.- Revista Espańola de Paleontología, Madrid, vol. 3, p. 13-23.
Meneghini G. (1858).- Paléontologie de l'île de Sardaigne. In: La Marmora A. (ed.), Voyage en Sardaigne.- Imprimerie Royale, Turin, p. 53-144.
Michelotti G. (1847).- Description des fossiles des terrains miocčnes de l'Italie septentrionale.- Société Hollandaise des Sciences, Leide, 408 p.
Monterosato T.A. (1872).- Notizie intorno alle conchiglie fossili di Monte Pellegrino e Ficarazzi.- Ufficio Tipografico M. Amenta, Palermo, 44 p.
Monterosato T.A. (1875).- Nuova rivista delle Conchiglie Mediterranee.- Atti dell'Accademia di Scienze, Lettere ed Arti di Palermo (New Series), vol. 5, p. 1-50.
Mörch O.A.L. (1853).- Catalogus conchyliorum quae reliquit D. Alphonso d'Aguirra & Gadea Comes de Yoldi, Regis Daniae Cubiculariorum Princeps, Ordinis Dannebrogici in Prima Classe & Ordinis Caroli Tertii Eques. Fasciculus secundus. Acephala, Annulata, Cirripedia, Echinodermata.- L. Kleinii, Copenhag, 76 p. URL: https://www.biodiversitylibrary.org/page/11720383
Morris S. (1985).- Preliminary guide to the oysters of Hong Kong.- Asian Marine Biology, Hong Kong, vol. 2, p. 119-138.
Morton B. (1990).- Life cycle and sexual strategy of Saccostrea cucullata (Bivalvia: Ostreidae) from a Hong Kong mangrove.- American Malacological Bulletin, Clamont - CA, vol. 8, p. 1-8. URL: https://www.biodiversitylibrary.org/page/45930983
Müller O.F. (1776).- Zoologiae Danicae Prodromus, seu Animalium Daniae et Norvigiae Indigenarum, characteres, nomina et synonyma imprimis popularum Havniae.- Typis Hallageriis, Copenhag, 274 p. URL: https://www.biodiversitylibrary.org/page/41499690
Muraour P. (1949).- Contribution ŕ l'étude micropaléontologique des marnes plaisanciennes d'Alger.- Éditions du Service Géologique National, Alger, p. 93-150.
Muraour P. (1951).- Les phénomčnes de fausses discordances dans l'Astien d'Alger.- Éditions du Service Géologique National, Alger, 26 p.
Newell N.D. (1965).- Classification of the Bivalvia.- American Museum Novitates, New York – NY, no. 2206, 25 p. URL: http://hdl.handle.net/2246/4472
Newton R.B. (1922).- Eocene Mollusca from Nigeria.- Geological survey of Nigeria, Abuja, vol. 3, p. 7-112.
Nyst P.H. & Westendrop G.D. (1839).- Nouvelles recherches sur les fossiles de la province d'Anvers.- Bulletin de l'Académie des Sciences, Lettres et Beaux-Arts de Belgique, Brussells, vol. 6, no. 10,393 p.
Parenzan P. (1974-1976).- Carta d'identitŕ delle conchiglie del Mediterraneo, II: Bivalvia.- Ed. Bios Taras, Taranto, part I (1974), 227 p.; part II (1976), p. 283-546.
Parras A. & Casadio S. (2005).- Taphonomy and sequence stratigraphic significance of oyster-dominated concentrations from the San Julián formation, Oligocene of Patagonia, Argentina.- Palćogeography, Palćoclimatology, Palćoecology, vol. 217, p. 47-66.
Pasteur-Humbert C. (1962).- Les Mollusques marins testacés du Maroc. II. Les Lamellibranches et les Scaphopodes.- Travaux de l'Institut scientifique chérifien (série Zoologie), Rabat, vol. 28, 184 p.
Payraudeau R.C. (1826).- Catalogue descriptif et méthodique des Annélides et des Mollusques de l'île de Corse.- Béchet, Paris, 218 p. URL: https://www.biodiversitylibrary.org/page/12960795
Pérčs J.M. & Picard J. (1964).- Nouveau manuel de bionomie benthique de la Méditerranée.- Recueil des Travaux de la Station Marine d'Endoume, vol. 31, 137 p.
Perrodon A. (1957).- Étude géologique des bassins néogčnes sub-littoraux de l'Algérie occidentale.- Bulletin du Service de la Carte géologique de l'Algérie, Alger, vol. 12, 323 p.
Philippi R.A. (1836-1844).- Enumeratio Molluscorum Siciliae, cum viventium tuum tellure tertiaro fossilium quae un itinere suo observavit auctor.- Berolini, S. Schroppii, (1), 268 p.; 1836; (2), 303 p. URL: https://www.biodiversitylibrary.org/page/46935803
Pickerill R.K. & Donovan S.K. (1998).- Ichnology of the Pliocene Bowden shell bed, southeast Jamaica.- Contributions to Tertiary and Quaternary Geology, Leyden, no. 35, p. 3-8.
Poli G.S. (1791-1827).- Testacea utriusque Siciliae eorumque historia et anatome tabulis aensis illustrata.- Parma, 3 vols. + 1 atlas (57 Pls.). URL: https://www.biodiversitylibrary.org/bibliography/79042
Poppe G.T. & Goto Y. (1993).- European seashells (Scaphopoda, Bivalvia, Cephalopoda), Vol. II.- Verlang Christa Hemmen, Wiesbaden, 221 p.
Quoy J.R.C. & Gaimard J.-P. (1834).- Voyage de découvertes de l'Astrolabe exécuté par ordre du Roi pendant les années 1826, 1827, 1828, 1829, sous le commandement de M. J. Dumont d'Urville. Zoologie. Tome 3 (part I).- Tastu, Paris, 366 p. URL: https://www.biodiversitylibrary.org/page/2076481
Raffi S. (1970).- I Pettinidé del Pliocene Calabriano delle'Emilia occidentale (Piacentino e Parmense).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 9, p. 97-135.
Rafinesque C.S. (1815).- Analyse de la Nature ou Tableau de l'univers et des corps organisés.- Aux dépens de l'auteur, Palerme, 226 p. URL: https://gallica.bnf.fr/ark:/12148/bpt6k98061z
Raulin V. & Delbos J. (1855).- Extrait d'une monographie des Ostrea des terrains tertiaires de l'Aquitaine.- Bulletin de la Société Géologique de France (2čme série), Paris, t. XII, p. 1144-1164.
Reed F.R.C. (1935).- Notes on the Neogene faunas of Cyprus. II.- Annals and Magazine of Natural History (tenth series), London, vol. XV, no. 85, p. 489-524.
Reinhart P.W. (1935).- Classification of the pelecypod family Arcidae.- Bulletin du Musée Royal d'Histoire Naturelle de Belgique, Brussels, vol. 11, no. 13, p. 1-68.
Rocher P. (2002).- Fossiles typiques du Bordelais.- Muséum d’Histoire naturelle de Bordeaux, 60 p.
Röding P.F. (1798).- Museum Boltenianum sive catalogues cimeliorum e tribus regnis naturae quae olim collegerat Joa. Fried. Bolten, M.D.p.d., Pars Secunda.- Hamburgi Typis Johan. Christi. Trappii, Hamburg, 199 p. DOI: https://doi.org/10.5962/bhl.title.16250
Roger J. (1939).- Le genre Chlamys dans les formations néogčnes de l'Europe.- Mémoire de la Société Géologique de France (nouvelle série), Paris, no. 40, 294 p.
Roman F. (1940).- Listes raisonnées des faunes du Pliocčne et du Miocčne de Syrie et faune du Liban.- Revue de Géographie physique et Géologie dynamique, Notes et Mémoires, Paris, vol. 3, p. 353-400.
Rosso J.C. (1979).- Mollusques testacés (Macrofaune). In: Burollet P.F., Clairefond P. & Winnock E. (eds.), La mer pélagienne. Étude sédimentologique et écologique du Plateau tunisien et du Golfe de Gabčs.- Géologie méditerranéenne, Marseille, t. VI, no. 1, p. 143-170. URL: https://www.persee.fr/doc/geolm_0397-2844_1979_num_6_1_1080
Saadallah A. (1981).- Le massif cristallophyllien d'El-Djazaďr (Algérie) : Évolution d'un charriage ŕ vergence nord dans les internides des Maghrébides.- Thčse Doctorat, Université des Sciences et de la Technologie Houari Boumedične, Alger, 160 p.
Sacco F. (1890-1904).- I Molluschi dei terreni terziari del Piemonte e délia Liguria.- Carlo Clausen, Torino, vol. 6, 30 p.
Saint Martin J.-P., Néraudeau D., Lauriat-Rage A., Goubert E., Secretan S., Babinot J.- F., Boukli-Hacene S., Pouyet S., Lacour D., Pestrea S. & Conesa G. (2000).- La faune interstratifiée dans les gypses messiniens de Los Yesos (Bassin de Sorbas, SE Espagne) : Implications.- Géobios, Lyon, vol. 33, no. 5, p. 637-649.
Saoudi N. (1989).- Pliocčne et Pléistocčne inférieur et moyen du Sahel occidental d'Alger.- Entreprise Nationale du Livre, Alger, no. 2020/85, 197 p.
Satour L. (2012).- Les bivalves néogčnes de l'Algérie nord-occidental : Systématique et paléoécologie.- Thčse Doctorat, Université d'Oran, Algérie, 312 p.
Saubade A.M. & Tesson M. (1981).- Étude préliminaire de la malacofaune du plateau continental du Rif oriental (Maroc), entre la côte et 50 mčtres.- Bulletin Géologique du Bassin d'Aquitaine, Bordeaux, vol. 29, p. 113-131.
Schultz O. (2001).- Bivalvia neogenica (Nuculacea-Unionacea).- Catalogus Fossilium Austriae, Vienna, Band 1, 379 p.
Seilacher A. (1953).- Studien zur Palichnologie I. Die fossilen Ruhespuren (Cubichnia).- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 98, p. 87-124.
Spadini V. (2017).- I pettinidi del senese.- 4o convegno Geo-Paleontologico and Malacologico Toscano Naturaliter Etruria, 58 p.
Stanley S.M. (1970).- Relation of shell form to life habits in the Bivalvia (Mollusca).- Memoirs of the Geological Society of America, Boulder - CO, vol. 125, 296 p.
Stefani C. de & Pantanelli D. (1878).- Molluschi pliocenici dei dintorni de Siena.- Bollettino della Societa Malacologica Italiana, Pisa, vol. 4, p. 5-215 [p. 31].
Stenzel H.B. (1971).- Oysters. In: Moore R.T. (ed.), Part N, vol. 3. Mollusca 6.- Treatise on Invertebrate Paleontology, Geological Society of America, Boulder - CO, University of Kansas, Lawrence - KS, N953-N1224, p. l-20.
Steininger F. & Schultz O. (1978).- Palaeobiologische charakteristik des Badenien. In: Papp A., Cicha I., Senes J. & Steininger F. (eds.), Chronostratigraphie und Neostratotypen. Miozän der zentralen Paratethys. Bd. IV, Mollusca, Pectinidae (Steininger).- Verlag der Slowakischen Akademie der Wissenschaften, Bratislava, p. 340-347.
Stoliczka F. (1871).- Cretaceous fauna of southern India, Vol. 3. The pelecypoda, with a review of all known genera of this class, fossils and Recent.- Geological Survey of India, Palaeontologica Indica, Calcutta, vol. 6, 537 p.
Studencka A. & Studencki W. (1988).- Middle Miocene (Badenian) bivalves from the carbonate deposits of the Wojcza-Pinczow Range (southern slopes of the Cross Mountains, Central Poland).- Acta Geologica Polonica, Warsaw, vol. 38, no. 1-4, p. 1-44.
Tavani G. & Tongiorgi M. (1963).- La fauna miocenica delle "Arenarie di Ponsano" (Volterra, Provincia di Pisa). I parte. Lamellibranchiata.- Palaeontogia Italica, Pisa, vol. 58, p. 1-43.
Taylor P.D. (1979).- Palaeoecology of the encrusting epifauna of some British Jurassic bivalves.- Palćogeography, Palćoclimatology, Palćoecology, vol. 28, p. 241-262.
Taylor P.D. (2016).- Competition between Encrusters on marine hard substrates and its fossil record.- Palaeontology, London, vol. 59, p. 481-497.
Taylor P.D. & Wilson M.A. (2003).- Palaeoecology and evolution of marine hard substrate communities.- Earth Science Reviews, vol. 62, p. 1-103.
Tebble N. (1976).- British Bivalve Shells.- Royal Scottish Museum Publication, Edimburg, 212 p.
Torigoe K. (1981).- Oyster in Japan.- Journal of Science of The Hiroshima University, vol. 29, no. 2, p. 291-419.
Veiga Ferreira O. da (1961).- Pectinideos do Miocénico da Bacia do Tejo.- Comunicaçőes dos Serviços Géológícos de Portugal, Lisboa, vol. 45, p. 419-465.
Venzo S. & Pelosio G. (1963).- La malacofauna tortoniana del Colle di Vigoleno (Preappennino Piaceneto).- Paleontolographia Italica, Pisa, vol. 58, p. 43-213.
Videt B. (2003).- Dynamique des paléoenvironnements ŕ huîtres du Crétacé Supérieur nord aquitain (SO France) et du Mio-Pliocčne andalou (SE Espagne) : Biodiversité, analyse séquentielle, biogéochimie.- Doctorat d'Université, Université de Rennes 1, 305 p.
Voigt E. (1965).- Über parasitische Polychaeten in Kreide-Austern sowie einige andere in Muschelschalen bohrende Würmer.- Paläontologische Zeitschrift, Stuttgart, vol. 39, p. 193-211.
Vyalov P.O.S. (1936).- Sur la classification des huîtres.- Comptes Rendus (Doklady) de l'Académie des Sciences de l'URSS, Moskwa, vol. 4, p. 17-20.
Waller T.R. (1978).- Morphology, morphoclines and a new classification of the Pteriomorphia (Mollusca; Bivalvia).- Philosophical Transactions of the Royal Society, London, B 284, p. 345-365.
Warme J.E. (1975).- Borings as trace fossils, and the processes of marine bioeorosion. In: Frey R.W. (ed.), The study of trace fossils.- Springer-Verlag, Berlin, p. 181-227.
Yassini I. (1973).- Nouvelles données stratigraphiques et microfaunistiques sur la limite Pliocčne inférieur-Pliocčne moyen (Plaisancien-Astien) dans la région d'Alger.- Revue de Micropaléontologie, Paris, vol. 16, no. 4, p. 229-248.
Yassini I. (1975).- Répartition des ostracodes dans une série marine régressive d'âge pliocčne dans la région d'Alger, Algérie.- Revue de Micropaléontologie, Paris, vol. 22, no. 2, p. 89-124.
Zavarei A. (1973).- Monographie des Spondylidae (Lamellibranches) actuels et fossiles.- Centre d'études et de recherches de paléontologie biostratigraphique (CERPAB), Notes et contributions, Orsay, no. 4, 233 p.