◄ Carnets Geol. 22 (17) ►
![]()
Outline:
[1. Introduction]
[2. The studied area]
[3. Materials and methods ...]
[4. Biostratigraphy]
[5. Discussion]
[6. Systematic paleontology]
[Bibliographic references] and ... [Plates]
Société Axonaise de Paléontologie, 43, rue du château 02110 Bohain-en-Vermandois (France)
Published online in final form (pdf) on November 11, 2022
DOI 10.2110/carnets.2022.2217
![]()
[Editor: Michel
Moullade; language editor: Simon F. Mitchell; technical editor: Bruno Granier]
![]()
An 148 m composite section located in the Gargas area
stratotype in southeastern France has been studied for its calcareous
nannofossil content. The four sections span the upper Bedoulian to lower
Gargasian with the stadial boundary located within the basal section south of
the village of Clavaillan. A total of 75 samples was processed to inventory
taxonomy and stratigraphic distributions.
Eprolithus floralis is present at the base of the stratigraphic succession. The lowest
occurrence of this species, which marks the base of Subzone NC7A, has been
documented in the uppermost lower Aptian (Bedoulian) in other sections in
southeastern France. The highest occurrence of the genus Micrantholithus
has been utilized to delineate the base of Subzone NC7B. This subzonal boundary
was placed at 48 m in the Les Gays I section (113.5 m in the composite) below
the lowest occurrence of the foraminifer Globigerinelloides
ferreolensis in the suprajacent sample (50 m). The lowest
occurrence of Braarudosphaera africana
is identified as a regional biohorizon in lower Subzone NC7A and a local proxy
for the Bedoulian/Gargasian boundary. This event was placed at 15.5 m in the
Clavaillan section at the base of the Dufrenoyia furcata Zone and within the
"Niveau Blanc inférieur" marker bed (NB1).
The genus Nannoconus is abundant to very
abundant in all samples examined. Taxonomic rigor has resulted in the
recognition of five main morphologic groups (A-E), including all but one of the
15 species discriminated over this relatively short stratigraphic interval. Four
main Nannoconus assemblage biozones - with one subdivision - have been
distinguished through semi-quantitative analyses and organized relative to these
taxonomic groupings. Assemblage Biozone B is restricted to the Bedoulian and has
been correlated to the lower portion of Subzone NC7A
(i.e., NC7A1). Assemblage biozones GI
and GII (A-B) have been correlated to the upper portion of Subzone NC7A
(i.e., NC7A2) and Biozone GIII to
Subzone NC7B within the Gargasian.
Assipetra is another solution-resistant genus included in semi-quantitative
analyses, where both its species were separated into small and large forms based
on a size of 10 μm. The highest percentages of large morphotypes are within
the Bedoulian in the lower 10.5 m of the Clavaillan section, roughly coeval to
an acme of large Assipetra observed in the basal portion of the Serre
Chaitieu section in the nearby Vocontian Basin.
• Aptian stratotype;
• Gargasian;
•
Bedoulian/Gargasian boundary;
• calcareous nannofossils;
• Nannoconus;
•
taxonomy;
• biostratigraphy
Lambert B.C. (2022).- Calcareous nannofossils of the uppermost Bedoulian and lower Gargasian of La Tuiličre - St-Saturnin-lčs-Apt (area of the Aptian stratotype, Vaucluse, SE France).- Carnets Geol., Madrid, vol. 22, no. 17, p. 745-793.
Nannofossiles calcaires du Bédoulien sommital et
du Gargasien inférieur de La Tuiličre - St-Saturnin-lčs-Apt (région du
stratotype de l'Aptien, Vaucluse, Sud-Est de la France).- Une coupe composite de
148 m localisée dans la région stratotypique de Gargas (Sud-Est de la France)
a été étudiée pour son contenu en nannofossiles calcaires. Les quatre coupes
couvrent le Bédoulien supérieur et le Gargasien inférieur. La limite entre
Aptien inférieur (Bédoulien) et Aptien supérieur (Gargasien) a été reconnue
ŕ la partie inférieure de la coupe basale au sud du village de Clavaillan. Un
total de 75 échantillons ont été préparés pour l'analyse taxinomique et
stratigraphique.
Eprolithus floralis est présent dans les premiers échantillons de la succession
stratigraphique. La premičre apparition de cette espčce qui marque la base de
la sous-zone NC7A a été documentée dans la partie sommitale de l'Aptien inférieur
(Bédoulien) dans d'autres coupes du Sud-Est
de la France. La derničre apparition du genre Micrantholithus
a été utilisée pour identifier la base de la sous-zone NC7B. Cette limite de
sous-zone a été placée ŕ 48 m dans la coupe Les Gays I (113.5m dans la coupe
composite) en dessous de la premičre apparition du foraminifčre planctonique Globigerinelloides
ferreolensis dans l'échantillon du dessus (50 m). La premičre
apparition de Braarudosphaera africana
est identifiée comme un horizon régional ŕ l'intérieur de la partie inférieure
de la sous-zone NC7A et un repčre local pour la limite Bédoulien/Gargasien.
Cet événement a été placé ŕ 15,5 m dans la coupe de Clavaillan ŕ la base
de la Zone ŕ Dufrenoyia
furcata (ammonite) et dans le "Niveau Blanc inférieur" (marqueur
NB1).
Une attention particuličre a été portée aux
Nannoconidés toujours abondants ŕ trčs abondants dans tous les échantillons. Sur cet intervalle stratigraphique relativement court, cinq groupes
morphologiques principaux (A-E) avec une quinzaine d'espčces ont été
identifiés.
Quatre biozones ŕ Nannoconus, dont une subdivisée en deux sous-zones, ont été
distinguées. La biozone B est limitée au Bédoulien et a
été corrélée ŕ la partie inférieure de la sous-zone NC7A (i.e.,
NC7A1). Les biozones GI et GII (A-B) ont été corrélées ŕ la partie supérieure
de la sous-zone NC7A (i.e., NC7A2) et la biozone GIII ŕ la sous-zone
NC7B dans le Gargasien.
Assipetra, un autre genre résistant ŕ
la dissolution, a également été pris en considération dans les analyses
semi-quantitatives, dont les représentants ont été séparés en petites et
grandes formes sur la base d'un seuil de taille de 10 μm. Les taux les plus élevés de grands morphotypes sont reconnus dans les
10,5 m inférieurs de la coupe de Clavaillan (Bédoulien). Cet épisode est ŕ
peu prčs équivalent de l'acmé ŕ grande Assipetra observée dans la
partie basale de la coupe de Serre Chaitieu dans le bassin Vocontien voisin.
• Aptien stratotype ;
• Gargasien ;
•
limite Gargasien/Bédoulien ;
• nannofossiles calcaires ;
•
Nannoconus ;
• taxinomie ;
• biostratigraphie
The Aptian calcareous nannofossils of southeastern France have been previously studied by several researchers. Deflandre and Deflandre-Rigaud (1962, 1967) published their pioneering taxonomic work on Nannoconus from Carniol ("Haute-Provence"). The first attempts at Lower Cretaceous biostratigraphy by means of calcareous nannofossils, with reference to the stratotype areas, were by Manivit (1971) and Thierstein (1971, 1973). The latter established the first Lower Cretaceous nannofossil biozonation. The detailed taxonomical study of the calcareous nannofossils of two Aptian substage type sections (Cassis-La Bédoule, Lower Aptian and Gargas, Upper Aptian) was carried out by Barrier (1977a, 1977b). More recently, in a collective volume dedicated to the Lower Aptian stratotype, Bergen (1998) published a thorough inventory of the Bedoulian calcareous nannofossils, as well as data on stratigraphic ranges and semi-quantitative distributions. The Aptian-Albian calcareous nannofossil biostratigraphy of the neighbouring Vocontian area was reviewed by Herrle and Mutterlose (2003) and most recently for the lower to upper Aptian by Giraud et al. (2018).
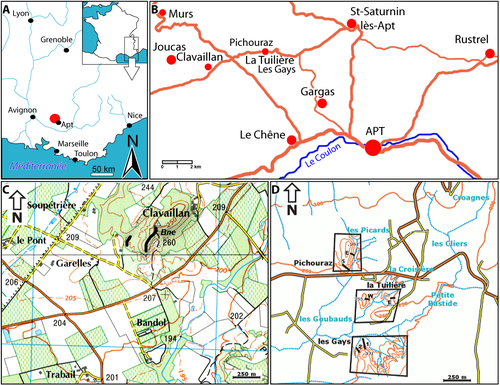
The
four studied sections are located near the hamlet of La Tuiličre, included in
the village of Saint-Saturnin-lčs-Apt (Fig. 1 ![]() ). These sections are the
Clavaillan Hill, south Pichouraz Hill and the Les Gays Farm area (two
sections). For further information about their geographic location,
relationships and geological setting, see Dutour (2005), and Moullade
et al. (2006, 2008, 2017).
). These sections are the
Clavaillan Hill, south Pichouraz Hill and the Les Gays Farm area (two
sections). For further information about their geographic location,
relationships and geological setting, see Dutour (2005), and Moullade
et al. (2006, 2008, 2017).
|
|
Figure 1: Geographic
location of the area studied. A) Apt area in SE France; B) studied sections;
C) Clavaillan
area; D) Pichouraz East and South; La Tuiličre W; Les Gays 1 and 2. |
By compositing these four sections, we estimate the total thickness of Aptian sediments measured in this area at around 148 m (with a gap between the top of Clavaillan and the base of Pichouraz East sections, see Moullade et al., 2017):
The average spacing between samples is about 2 m.
Moullade et al. (2006, 2017) divided the relatively homogeneous lithostratigraphy in this area into five main units from the base to top:
Details of the composite section and sample levels, in addition to the ammonite and foraminiferal stratigraphies, are summarized in Moullade et al. (2017).
Nannofossil preservation is heterogenous in the material studied. For Nannoconus, there is no special problem due to the relatively great size and robustness of the specimens, especially in the lower part of the studied sections (Clavaillan). As for the other nannofossils, preservation appears related to the calcium carbonate content. In the Clavaillan section, the sediments are composed of shaly limestones, where the high content in CaCO3 seems to be directly related to the abundance of Nannoconus tests and the poor preservation of the other nannofossils components (especially small specimens). In the other sections (i.e., South Pichouraz and Les Gays), marls and calcareous shales are the dominant lithologies and the nannofossils preservation is very good. This basic contrast in nannofossils preservation introduces bias in calcareous nannofossil distributions.
All analyses undertaken on our material are based on optical observations only (no SEM analyses were performed). Two smear slides were prepared for each sample: One slide with a higher concentration of sediment to investigate all nannofossil components and one slide with a relatively dilute amount of sediment for semi-quantitative analyses. No special cleaning methods were used in sample preparations.
The "semiquantitative" approach employed is unconventional (see Lambert & Laporte-Galaa, 2005, for further details). This approach did not account for the actual nannofossil abundance, but instead are counts of an individual taxon or a group of taxa to the total nannofossil population. For this exercise, two types of estimates were made:
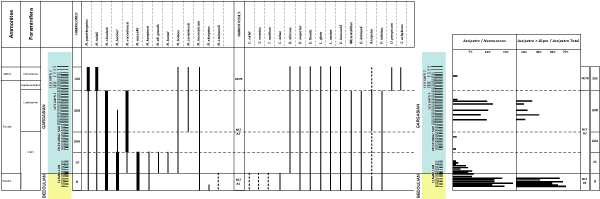
Tremolada and Erba (2002) have illustrated the importance of the size variation of both A. infracretacea and A. terebrodentatorius during Aptian time. These authors noted that large specimens are more frequent in the lower Aptian. We have carried out a semi-quantitative distribution of these two species. Threshold given by these authors is 7.5 µm. However, for operational reasons (technical limitations related to the microscope) we have applied a 10 µm threshold.
We have introduced a new index, the ratio between A.i + A.t and Nannoconus. The estimated frequency have been obtained after counting 500 specimens of Nannoconus. This index allows us to provide a relative frequency of these two nannofossils. The A.i + A.t (big / small with a threshold at 10 µm) ratio have been obtained if the number of specimens is equal to or greater than 50 specimens of Assipetra (in one smear slide). Four intervals can be distinguished:
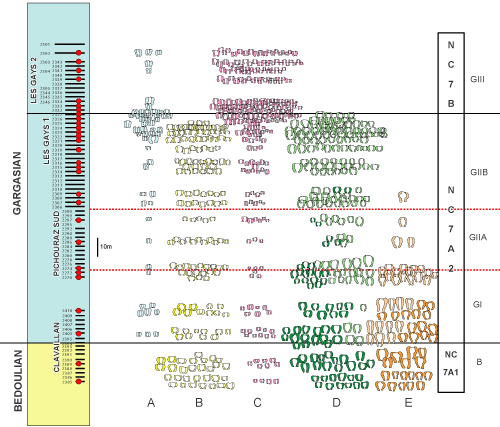
Nannoconid
assemblage biozones. Four local Nannoconus
assemblage zones - including one divided into two subzones - have been
recognized in the materials studied (Figs. 2 ![]() , 4
, 4 ![]() ) and integrated with the event
stratigraphy (see § "Nannofossil biozonation" below):
) and integrated with the event
stratigraphy (see § "Nannofossil biozonation" below):
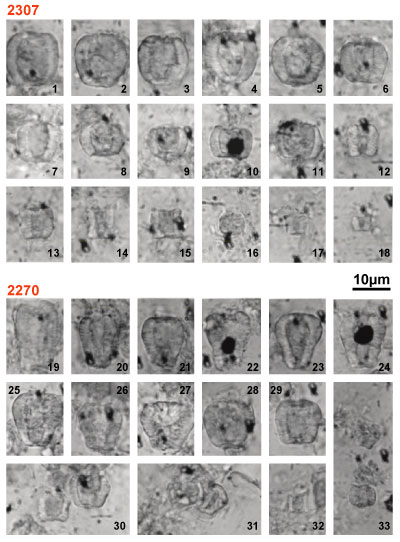
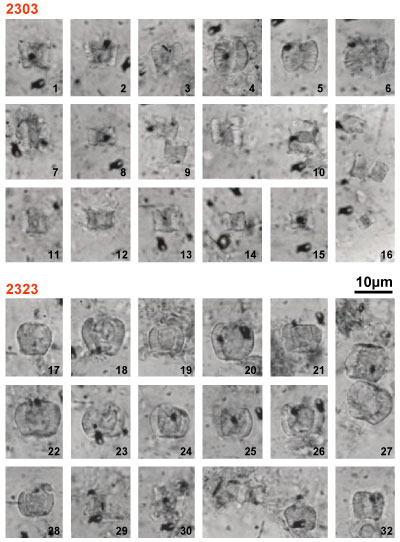
1. Biozone B (Bedoulian): Clavaillan 2385 to 2393 samples
2. Biozone GI (Gargasian): Clavaillan samples 2394 to 2410 samples, south Pichouraz samples 2270 and 2272. The complete Nannoconus association is as follows:
3a. Biozone GII (Subzone GIIA): south Pichouraz samples 2274 to 2292
3b. Biozone GII (Subzone GIIB): south Pichouraz samples 2295 to 2300, Les Gays 1 Samples 2306 to 2327
4. Biozone GIII, samples Les Gays 1 samples 2332 to 2343, Les Gays 2 samples 2346 to 2301
|
|
Figure 2: semi-quantitative Nannoconus distributions.
|
Nannofossil biozonation: The standard NC zonation of Roth
(1978), as
emended by Bralower et al.
(1993), is employed herein for consistency with previous regional studies
in southeastern France. The following stratigraphic subdivisions are proposed
for this study (Fig. 3 ![]() ):
):
The interval from the first occurrence of Eprolithus floralis (present at the base of the composite section) to the first occurrence of Braarudosphaera africana. The lowest occurrence of Br. africana is in Sample 2394 (15.5 m) in the Clavaillan section. This interval contains Nannoconus assemblage Biozone B. The highest abundances of Assipetra and highest percentages of large morphotypes (> 10 μm) relative to small are restricted to the lower 10.5 m (sample 2390 and below) of the Clavaillan section. Sporadic occurrences of Nannoconus steinmannii and Conusphaera rothii were observed throughout. Nannoconus elongatus is restricted to the lower part of this interval.
The interval from the first occurrence of Braarudosphaera africana to the last occurrence of Micrantholithus spp. This interval includes Nannoconus assemblage biozones GI and GII. The lowest occurrences of N. boletus and N. vocontiensis were observed at the base of this interval. The highest occurrences of Diazomatolithus lehmanii and "wide" Nannoconus (N. circularis, N. vocontiensis, N. bucheri) were observed at the top of this interval. The ratio of large to small Assipetra is higher in the upper portion of this interval (Nannoconus assemblage Biozone GIIB).
The interval between the last occurrence of Micrantholithus spp. and the first occurrence of Prediscosphaera columnata (not present at the top of the composite section). This interval contains Nannoconus assemblage Biozone GIII. Short Nannoconus with wide axial canals (N. truittii and N. quadriangulus) are very abundant throughout this subzone, but rare below. A significant change in the nannoconid populations at the base of this subzone between samples 2327 and 2332 in the Les Gays I section is further reinforced by highest common occurrences and tops of both N. circularis and N. vocontiensis, in addition to the top of N. bucheri. The lowest occurrences of Stoverius achylosus and Orastrum perspicuum were also observed at the base of this subzone.
|
|
Figure 3: Nannoplankton
stratigraphic distributions. |
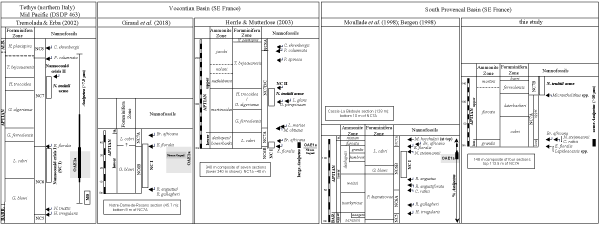
Regional nannofossil studies: The first comprehensive Lower Cretaceous nannofossil
biostratigraphic scheme by Thierstein (1971) was founded on a composite
of fourteen sections located in southeastern France. Thierstein (1973)
later published range charts of this research and the five outcrops encompassing
the Aptian included the Bedoulian and Gargasian historical stratotypes. These
data and interpretations provided the foundation for the Aptian zonal
stratigraphy both regionally and globally for the bases of NC6 to NC8. More
recent Aptian nannofossil biostratigraphic and paleoecologic studies in
southeastern France are summarized in Figure 4 ![]() , along with the integrated
ammonite and foraminiferal biozonations.
, along with the integrated
ammonite and foraminiferal biozonations.
Multidisciplinary stratigraphic research on the lower Aptian historical stratotype in the Cassis-La Bédoule area (South Provencal Basin) is summarized by Moullade et al. (1998). Bergen (1998) published the calcareous nannofossil biostratigraphy and abundance trends of some key genera in the same volume and bracketed the section from the upper portion of Subzone NC5C to the lower 6 m of Subzone NC7A (M. hoschulzii ranges to the top of the section, Bed 175B). The lowest occurrence of Eprolithus floralis (Bed 169A) marked the base of Subzone NC7A, followed upwards by the bases of Braarudosphaera africana (Bed 172B), Radiolithus planus (Bed 173), and Chiastozygus platyrhethum (Bed 174B).
Two biostratigraphic-paleoecologic studies (Herrle
& Mutterlose, 2003; Giraud et al.,
2018) in the nearby Vocontian Basin overlap
stratigraphically in lower Subzone NC7A (Fig. 4 ![]() ). The multidisciplinary study on
the Notre-Dame-de-Rosans section by Giraud et
al. (2018) assessed early Aptian paleoecologic trends across the
OAE1a event within their detailed chronologic framework. The majority of the
45.5 m measured section was placed in Subzone NC6B based on the presence of Rhagodiscus
angustus at its base and the absence of Conusphaera
rothii throughout. The base of Subzone NC7A was placed 4 m above
the black shales of the "Niveau Goguel" (~9.5 m below the top
of the section) on the lowest occurrence of Eprolithus
floralis and roughly equivalent to the lowest occurrence of the
foraminifer Leupoldina cabri.
They also reported the following sequence of lowest occurrences within Subzone
NC7A relative to the top of the "Niveau Goguel": Radiolithus
planus (8 m), Eprolithus
apertior (12.5 m), and Braarudosphaera
africana (13 m). Micrantholithus
hoschulzii, the extinction of which marks the top of Subzone NC7A,
ranges to the top of this section.
). The multidisciplinary study on
the Notre-Dame-de-Rosans section by Giraud et
al. (2018) assessed early Aptian paleoecologic trends across the
OAE1a event within their detailed chronologic framework. The majority of the
45.5 m measured section was placed in Subzone NC6B based on the presence of Rhagodiscus
angustus at its base and the absence of Conusphaera
rothii throughout. The base of Subzone NC7A was placed 4 m above
the black shales of the "Niveau Goguel" (~9.5 m below the top
of the section) on the lowest occurrence of Eprolithus
floralis and roughly equivalent to the lowest occurrence of the
foraminifer Leupoldina cabri.
They also reported the following sequence of lowest occurrences within Subzone
NC7A relative to the top of the "Niveau Goguel": Radiolithus
planus (8 m), Eprolithus
apertior (12.5 m), and Braarudosphaera
africana (13 m). Micrantholithus
hoschulzii, the extinction of which marks the top of Subzone NC7A,
ranges to the top of this section.
Herrle and Mutterlose (2003) investigated the nannofossil biostratigraphy and assemblages from a 340 m composite of seven sections spanning the upper lower Aptian (Zone NC6) to lowermost Albian (Subzone NC8B). They documented a sequence of 23 nannofossil appearance/extinction events, in addition to abundance trends of the genera Nannoconus, Assipetra, and Repagulum. These results were integrated to the lithostratigraphy (esp. black shales and other key beds), ammonite and foraminiferal biozonations, and interpreted eustatic cycles. The lowest occurrence of Eprolithus floralis (base Subzone NC7A) was placed 1 m above the "Niveau Goguel" black shales near the base of the composite section. The highest occurrence of Micrantholithus obtusus was used to mark the top Subzone NC7A, as opposed to the original marker species (M. hoschulzii) proposed by Bralower et al. (1993). This event was placed 7.7 m below the Niveau Noire Calcaire 2 in the same section. Within the lower half of the approximately 40 m of Subzone NC7A in this Serre Chaitieu section, the following nannofossil radiation was documented from bottom to top: Radiolithus planus, Eprolithus apertior, Braarudosphaera africana, Br. hockwoldensis, and Eprolithus varolii/Corollithion acutum. In addition, a 13 m interval from the Niveau Goguel (OAE1a) to the Niveau Blanc straddling the NC6/NC7A boundary was observed to be nearly devoid of Nannoconus, but containing elevated percentages of large Assipetra infracretacea (> 6 μm) and A. terebrodentarius (> 10 μm). Herrle and Mutterlose (2003) equated this interval to the nannoconid crisis of Erba (1994) and formerly designated it as nannoconid crisis I (NCI).
Herrle and Mutterlose (2003) could not differentiate Subzone NC7C because the lowest occurrence of Rhagodiscus achlyostaurion was not observed in the Serre Chaitieu section. Their combined Subzone NC7B/C, approximately 170 m thick and spanning three sections, was delineated at its top by the lowest occurrence of Prediscosphaera columnata. Within this combined subzone, a sequence of five lowest occurrences are distributed into three discrete intervals from bottom to top: (1) Lapideacassis mariae followed above by Prediscosphaera sp. near the base; (2) Orastrum perspicuum followed immediately above by Lapideacassis glans in the middle; and (3) Prediscosphaera spinosa near the top of Subzone NC7B/C.
Mutterlose (1989, 1991) described a late Aptian acme in Nannoconus truittii, noting its occurrence in both Boreal and Tethys sections. Herrle and Mutterlose (2003) documented a nannoconid acme in the lower upper Aptian of their composite in southeastern France, ranging from the middle Globigerinelloides algerianus to the base of the Ticinella bejaouaensis foraminiferal zones. Herrle and Mutterlose (2003) defined the rapid decrease in Nannoconus spp. at the top of this acme as nannoconid crisis II (NCII).
Tremolada and Erba
(2002) studied the Aptian biostratigraphy, paleoecology,
and morphometrics of Assipetra infracretacea
and Assipetra terebrodentarius
from individual cores in northern Italy (Cismon) and the mid Pacific (DSDP Site
453), providing insight and calibration for the current work (Fig. 4 ![]() ). They
found that the highest percentages of large specimens (> 7.5 μm) relative
to normal-sized specimens for both species occurred within the upper lower
Aptian "Selli" event (OAE1a). This acme of large Assipetra
occurred in the upper portion of Zone NC6 is also associated with the first
nannoconid crisis (NCI). Tremolada and Erba
(2002) summarized the
abundance and stratigraphy of Assipetra from the uppermost Barremian
(NC5) to lower Albian (lower NC8) relative to the magnetostratigraphy and
foraminiferal zones. Their definitions of zones NC6 to NC8 (Fig. 4
). They
found that the highest percentages of large specimens (> 7.5 μm) relative
to normal-sized specimens for both species occurred within the upper lower
Aptian "Selli" event (OAE1a). This acme of large Assipetra
occurred in the upper portion of Zone NC6 is also associated with the first
nannoconid crisis (NCI). Tremolada and Erba
(2002) summarized the
abundance and stratigraphy of Assipetra from the uppermost Barremian
(NC5) to lower Albian (lower NC8) relative to the magnetostratigraphy and
foraminiferal zones. Their definitions of zones NC6 to NC8 (Fig. 4 ![]() ) are
congruent with the aforementioned research in southeastern France (Bergen,
1998; Moullade et al., 1998; Herrle & Mutterlose,
2003; Giraud et
al., 2018), as well as the calibration of the late Aptian Nannoconus truittii acme
(NCII) with Herrle and Mutterlose
(2003).
) are
congruent with the aforementioned research in southeastern France (Bergen,
1998; Moullade et al., 1998; Herrle & Mutterlose,
2003; Giraud et
al., 2018), as well as the calibration of the late Aptian Nannoconus truittii acme
(NCII) with Herrle and Mutterlose
(2003).
|
|
Figure 4: Comparisons to
relevant nannofossil research. |
The composite of four sections in the South Provencal Basin spans the upper portion of Subzone NC7A to the lower
portion of Subzone NC7B (see § "Nannofossil biozonation" above). The
lowest occurrence of Braarudosphaera
africana can be used as an event to further subdivide Subzone
NC7A in southeastern France, whereas Subzone NC7C (base on the first
occurrence of Rhagodiscus achlyostaurion)
was not delineated in these same regional studies (Fig. 4 ![]() ).
).
Lapideacassis and Eprolithus floralis
were observed throughout the entire composite of four sections studied (Fig. 3 ![]() ).
The presence of both L. mariae
and L. glans at
the base of the composite in Subzone NC7A are both lower (in NC7B/C) than
reported by Herrle and Mutterlose
(2003) in the nearby Vocontian
Basin. Conversely, the lowest occurrence of Stoverius
achylosus was determined to be higher in this study (base Subzone
NC7B) relative to original zonal framework (NC6A, Bralower et al.,
1993) and the historical Bedoulian stratotype (NC6B,
Bergen, 1998).
).
The presence of both L. mariae
and L. glans at
the base of the composite in Subzone NC7A are both lower (in NC7B/C) than
reported by Herrle and Mutterlose
(2003) in the nearby Vocontian
Basin. Conversely, the lowest occurrence of Stoverius
achylosus was determined to be higher in this study (base Subzone
NC7B) relative to original zonal framework (NC6A, Bralower et al.,
1993) and the historical Bedoulian stratotype (NC6B,
Bergen, 1998).
Two stratigraphic anomalies in the lower half of the
Clavaillan section are: (1) the occurrences of Conusphaera rothii and Nannoconus
steinmannii with Eprolithus
floralis; and (2)
increased percentages of large Assipetra.
Bralower et al.
(1993) defined the top of Subzone NC6A on the highest occurrence of Conusphaera
rothii and placed the top of Nannoconus steinmannii
immediately above in the basal portion of Subzone NC6B; they defined the base of
Subzone NC7A on the lowest occurrence of Eprolithus
floralis. Bergen (1998) confirmed this subzonal
stratigraphy in the historical Bedoulian stratotype, but placed the top of Nannoconus
steinmannii much higher - immediately below the base of Subzone NC7A.
Both N. steinmannii and Conusphaera
rothii are extremely rare in Clavaillan samples. When contrasted to
the continuous presence of Eprolithus floralis,
redeposition is a likely explanation for these extremely rare occurrences. Tremolada
and Erba (2002) documented the highest percentage of large versus
normal-sized Assipetra in the upper
part of Zone NC6 below the lowest occurrence of Eprolithus
floralis. They associated the "acmes"
of these large forms with maximum TOC values during OAE1a in cores in northern
Italy and the mid Pacific. Disparate taxonomy (i.e., size) and counting
methodologies could explain the apparent age discrepancies in the "acmes"
of these large morphotypes. However, the possibility of diachroneity - or a
second, late early Aptian acme in large Assipetra - must be considered. Herrle
and Mutterlose (2003) also observed an acme in large Assipetra in
lowermost Subzone NC7A in the Vocontian Basin (Fig. 4 ![]() ).
).
The lowest occurrence of Braarudosphaera
africana at 15.5 m above the base of the Clavaillan section
correlates to the base of Dufrenoyia furcata
Zone and falls within the "Niveau Blanc inférieur" (NB1) marker
(see Moullade et al., 2017). Elsewhere in the South Provencal Basin at the Cassis-La Bédoule section,
this event was placed 2.5 m above the base of the Dufrenoyia furcata
Zone and 3 m above the base of NC7A (Moullade et
al., 1998; Bergen, 1998). Ten other nannofossil events
associated with the Braarudosphaera
africana horizon at Clavaillan (Fig. 3 ![]() ) indicate a disconformity
and would explain this slight discrepancy in correlation to the ammonite zonal
stratigraphy in the basin. In the Vocontian Basin, the lowest occurrence of Br. africana was placed 9 m above the base of NC7A in the
Notre-Dame-de-Rosans section by Giraud et al. (2018) and approximately 17 m above the base of NC7A
in the Serre Chaitieu section by Herrle and Mutterlose
(2003).
) indicate a disconformity
and would explain this slight discrepancy in correlation to the ammonite zonal
stratigraphy in the basin. In the Vocontian Basin, the lowest occurrence of Br. africana was placed 9 m above the base of NC7A in the
Notre-Dame-de-Rosans section by Giraud et al. (2018) and approximately 17 m above the base of NC7A
in the Serre Chaitieu section by Herrle and Mutterlose
(2003).
The highest occurrence of the genus Micrantholithus
in sample 2327 (48 m) in the Les Gays I section is used herein to mark the
base of Subzone NC7B; the lowest occurrence of the foraminifer Globigerinelloides
ferreolensis is immediately above in sample 2332 (50 m; Moullade
et al., 2017). Seven other nannofossil bioevents (lowest and
highest occurrences) have been associated with the boundary between subzones
NC7A and NC7B in this study; abundance changes of five Nannoconus species
were also observed at this stratigraphic level (Fig. 3 ![]() ). All these data indicate
a significant change between samples 2327 and 2332 in the Les Gays I section. In the Vocontian
Basin, Herrle and Mutterlose
(2003) used the highest occurrence of
Micrantholithus obtusus
to mark the base of Subzone NC7B, as opposed to the highest occurrence of the
genus. The strong abundance increase in short nannoconids with wide axial canals
at the base of Subzone NC7B could be related to the base of the Nannoconus
truittii acme correlated to the middle of the Globigerinelloides algerianus foraminiferal Zone (Tremolada & Erba,
2002; Herrle & Mutterlose,
2003). However, it is associated
with the base of the underlying Globigerinelloides
ferreolensis foraminiferal Zone herein.
). All these data indicate
a significant change between samples 2327 and 2332 in the Les Gays I section. In the Vocontian
Basin, Herrle and Mutterlose
(2003) used the highest occurrence of
Micrantholithus obtusus
to mark the base of Subzone NC7B, as opposed to the highest occurrence of the
genus. The strong abundance increase in short nannoconids with wide axial canals
at the base of Subzone NC7B could be related to the base of the Nannoconus
truittii acme correlated to the middle of the Globigerinelloides algerianus foraminiferal Zone (Tremolada & Erba,
2002; Herrle & Mutterlose,
2003). However, it is associated
with the base of the underlying Globigerinelloides
ferreolensis foraminiferal Zone herein.
The Mesozoic classification of Bown and Young (1997) is mainly followed, with some modification to the ordinal classification proposed by Watkins and Raffi (2020). Perch-Nielsen (1985) and Young et al. (1997, 2022) were also used for classification.
Order EIFFELLITHALES Rood et al., 1971
Imbricate muroliths.
Family CHIASTOZYGACEAE (Rood et al., 1973) Varol & Girgis, 1994
Genus Staurolithites Caratini, 1963
Imbricate muroliths with a central axial cross. Vagalapilla Bukry, 1969, Vekshinella Loeblich & Tappan, 1963, and Staurorhabdus Noël, 1972, are considered junior synonyms.
Staurolithites handleyi Lees, 2007
Pl. 3 ![]() , fig. 13
, fig. 13
Small coccolith (< 4 μm) with a delicate cross.
Distribution: Sporadic in the Gargasian marls.
Staurolithites mutterlosei Crux, 1989
Pl. 3 ![]() , fig. 14
, fig. 14
This relatively large coccolith has a spiraled extinction pattern. The bright cross is slightly off-axis, but the arms are at right angles. It occurs sporadically throughout the Gargasian.
Staurolithites rectus Black, 1971a
Pl. 3 ![]() , figs. 15-16
, figs. 15-16
This relatively large coccolith (< 6 μm) has a wide central area spanned by a prominent axial cross. Optically, the relatively broad, parallel-sided arms are divided by longitudinal sutures.
Distribution: Sporadic in all the samples.
Genus Bukrylithus Black, 1971a
Bukrylithus ambiguus Black, 1971a
Pl. 3 ![]() , fig. 17
, fig. 17
The well-developed axial cross broadens towards the center; its base arched on the proximal surface.
Distribution: Sporadic in the Gargasian marls.
Genus Rhabdophidites (Manivit, 1971) Lambert, 1987
The murolith base is minute relative to the elongate, bladed stem. For further discussion about this genus, see Covington and Wise (1987, p. 632).
Rhabdophidites moeslensis Manivit, 1971
Pl. 3 ![]() , figs. 20-21
, figs. 20-21
Stem is greater than 10 times the coccolith length (< 1 μm). The stem tapers in both directions as in Lithraphidites, but that genus has no basal coccolith.
Distribution: Sporadic in the samples studied. Manivit (1971) has reported this nannofossil from the Albian.
Rhabdophidites parallelus (Wind & Čepek, 1979) Lambert, 1987
Pl. 3 ![]() , figs. 18-19, 22
, figs. 18-19, 22
The basal coccolith is 1-2 μm and the distal shield composed of around 20 elements (Lambert, 1987). Stem length is about 3-4 times the coccolith lengh.
Distribution: Sporadic in all the samples.
Genus Chiastozygus Gartner, 1968
The central area is occupied by a diagonal cross; the angle between the arms is variable.
Chiastozygus platyrhetum Hill, 1976
Pl. 4 ![]() , figs. 4-5
, figs. 4-5
Description: "Central area spanned by a central cross, the arms of which are oriented symmetrically about the principal axes of the ellipses. The crossbars and the rim are approximately equal in width" (Hill, 1976, p. 129). The crossbar is located slightly above the rim and each arm appears bisected longitudinally in cross-polarized light.
Distribution: Sporadic in the Gargasian marls. Bergen (1998) placed the lowest occurrence of this species in the upper Bedoulian within the lower portion of Zone NC7.
Chiastozygus aff. tenuis Black, 1971a
Pl. 4 ![]() , figs. 1-3, ? 9
, figs. 1-3, ? 9
This taxon mostly closely resembles Chiastozygus tenuis, from which it differs by having a stem and a cross that slightly deviates from the main ellipse axes. The cross arms are orthogonal. Specimens observed appear conspecific to the specimen of "Zygolithus" litterarius (Górka) in Thierstein (1971, Pl. 1, figs. 3-4), but Chiastozygus litterarius (Górka, 1957) Manivit, 1971, is a Maastrichtian species with a bright, spiraled rim extinction pattern (see Reinhardt & Górka, 1967; Bergen, 1998).
Distribution: Sporadic in all the samples.
Genus Tegumentum Thierstein in Roth & Thierstein, 1972
Tegumentum stradneri Thierstein in Roth & Thierstein, 1972
Pl. 4 ![]() , figs. 6-7
, figs. 6-7
The arms form an acute angle to the minor ellipse axis. Specimens were
observed with crosses that were asymmetric (Pl. 4 ![]() , fig. 6) to symmetric
(Pl. 4
, fig. 6) to symmetric
(Pl. 4 ![]() ,
fig. 7) to the ellipse axes.
,
fig. 7) to the ellipse axes.
Distribution: Sporadic throughout the samples.
Genus Percivalia Bukry, 1969
Percivalia is characterized by an inner rim area composed of several narrow tiers of elements (Bukry, 1969). The lath-shaped elements of these tiered cycles are upright and exhibit a first order white birefringence. The central area may be occupied by a variety of central structures.
Percivalia fenestratus (Worsley, 1971) Wise, 1983
Pl. 7 ![]() , fig. 3
, fig. 3
This species has an imperforate central area; in cross-polarized light, two nodes may be noticed in the minor axis of the central area.
Distribution: Sporadic in all the samples.
Percivalia hauxtonensis Black, 1973
Pl. 7 ![]() , figs. 1-2
, figs. 1-2
Percivalia hauxtonensis is a medium to large species with two longitudinal central perforations and a distal boss. Black (1973) illustrated five electron photomicrographs (proximal and distal views) and provided measurements of nine specimens (7.2-9.0 μm in length). The longitudinal central openings of the lower Cenomanian holotype are rimmed by a cycle of elements, akin to the two specimens illustrated herein. The light photomicrographs of an upper Albian specimen illustrated by Bralower and Bergen (1998) is smaller (5.7 μm) and lacks a distal boss. The upper Albian specimen illustrated by Burnett in Gale et al. (1996) and again in Burnett (1998) does not exhibit the rim birefringence pattern of Percivalia.
Distribution: This species was recorded within Percivalia fenestratus during sample analyses.
Genus Zeugrhabdotus Reinhardt, 1965
Murolith with a bridge along the short axis (distal view). Without any electron photographs it has been impossible to take into consideration the proximal ultrastructure (grid or other element) to differentiate the genera Zygodiscus and Zeugrhabdotus (sensu Lambert, 1987).
Zeugrhabdotus bicrescenticus (Stover, 1966) Burnett in Gale et al., 1996
Pl. 4 ![]() , fig. 19
, fig. 19
Massive bridge composed of two distinct calcitic series. Some specimens belonging to this species have been previously put in the species Z. diplogrammus (for instance, see Manivit, 1971, Pl. 13, figs. 5-6). But this author places into the same species name (Z. diplogrammus) some coccoliths clearly different (e.g., Tranolithus manifestus Stover, 1966, see Manivit, 1971, Pl. 13, figs. 2, 4, 12-13). See discussion about Zeugrhabdotus diplogrammus here below.
Distribution: Sporadic in all the samples.
Zeugrhabdotus choffatii Rood et al., 1973
Pl. 4 ![]() , fig. 15
, fig. 15
A very small to small coccolith characterized by a simple bridge with a central opening corresponding to a hollow stem base. The Bathonian holotype is very small (2.4 μm). Mutterlose and Wise (1990) provided an excellent electron photomicrograph of a Valanginian specimen recovered from the east Antarctic margin (3.5 μm); the Aptian specimen illustrated herein is 3.2 μ in length.
Distribution: Sporadic in the Gargasian marls.
Zeugrhabdotus cf. diplogrammus (Deflandre in Deflandre & Fert, 1954) Burnett in Gale et al., 1996
Pl. 4 ![]() , figs. 10-11, 18
, figs. 10-11, 18
Much confusion exists about the species Zygodiscus diplogrammus described by Deflandre in 1954 based on poorly illustrated North Africa Mio-Pliocene specimens. The initial description indicated "elliptique ŕ marge étroite, ŕ bord lisse; aire centrale occupée selon son petit axe par deux barres parallčles". We put in cf. diplogrammus sensu Hill, 1976, coccoliths close to those illustrated by Hill.
Distribution: Sporadic in the Gargasian shales.
Zeugrhabdotus howei Bown in Kennedy et al., 2000
Pl. 4 ![]() , fig. 17
, fig. 17
Bown (in Kennedy et al., 2000) described this species to account for specimens identified as "Zygodiscus" elegans (Gartner, 1968) Bukry, 1969. Bergen (1998) illustrated a Bedoulian specimen as Lordia sp.
Distribution: The species is sporadic in the Aptian samples examined.
Zeugrhabdotus moulladei Bergen, 1998
Pl. 4 ![]() , fig. 14
, fig. 14
Small murolith with a transverse bar composed by two opposing bright plates separated by a dark central stem.
Distribution: Sporadic in the Gargasian marls.
Zeugrhadotus noeliae Rood et al., 1971
Pl. 4 ![]() , figs. 16, 23-25
, figs. 16, 23-25
The Oxfordian holotype of Zeugrhabdotus noeliae is an electron photomicrograph (distal view) of a very small specimen (2.9 μm). Light microscope identification of this species by various Mesozoic researchers has been highly variable (see Young et al., 2022). Very small muroliths (2-3 μm) with a transverse central bar have been assigned to this species herein. This group constitutes one of the main components of the recovered nannoflora (up to 25-35% of the assemblage).
Zeugrhabdotus xenotus (Stover, 1966) Burnett in Gale et al., 1996
Pl. 3 ![]() , fig. 26
, fig. 26
The specimen illustrated is oriented parallel to the polarizing direction.
Distribution: Sporadic in the Gargasian marls.
Zeugrhabdotus sp. 1
Pl. 4 ![]() , figs. 21-22
, figs. 21-22
Murolith with a narrow, bicyclic rim and a large central area. The outer rim cycle is broader and less birefringent than the bright inner rim cycle. The inner rim cycle broadens around the contacts with a narrow central transverse bar. The bar appears constructed of two elements and supports a solid distal stem which has an obloing base in plan view. Zeugrhabdotus clarus Bown, 2005, is an Albian-Cenomanian species described from the northwest Pacific Ocean. This species has a broader rim and corresponding smaller central area; its more prominent bright inner rim cycle has strongly spiraled extinction. The bar elements of Z. trivectis Bergen, 1994, are optically grouped into a bundle of three elements slightly offset from each other, as opposed to the two-element bundle of Zeugrhabdotus sp. 1.
Distribution: Sporadic in all the samples.
Genus Parhabdolithus Deflandre, 1952
Murolith with a massive wall, which differentiates it from Zeugrhabdotus, and a bridge supporting a generally well developed central process, which differentiates it from Rhagodiscus.
Parhabdolithus embergeri (Noël, 1958) Stradner, 1963
Pl. 4 ![]() , figs. 12-13
, figs. 12-13
A size variation was observed between smaller
(Pl. 4 ![]() , fig. 13) and larger
specimens (Pl. 4
, fig. 13) and larger
specimens (Pl. 4 ![]() , fig. 12).
, fig. 12).
Distribution: Sporadic to frequent in all the samples.
Parhabdolithus imperfossus (Black, 1972) Lambert, 1987
Pl. 5 ![]() , figs. 13-15
, figs. 13-15
Very
tiny nannofossils (2-4 µm) with very long and thin stem are observed in our
samples (Pl. 5 ![]() , fig. 14, distal view, and
Pl. 5
, fig. 14, distal view, and
Pl. 5 ![]() , fig. 15, profile view). Lambert
(1987) discussed the possible link between Parabdolithus imperfossus
initially described in SEM and Rhagodiscus achlyostaurion described in
photonic microscopy. From a structural point of view with the absence of hole in
the proximal rim, the species could be considered as a Rhagodiscus (see Bergen,
1998, p. 246). However, Lambert
(1987) described several morphotypes
based on the size of the coccolith and the form of the coccosphere.
, fig. 15, profile view). Lambert
(1987) discussed the possible link between Parabdolithus imperfossus
initially described in SEM and Rhagodiscus achlyostaurion described in
photonic microscopy. From a structural point of view with the absence of hole in
the proximal rim, the species could be considered as a Rhagodiscus (see Bergen,
1998, p. 246). However, Lambert
(1987) described several morphotypes
based on the size of the coccolith and the form of the coccosphere.
Distribution: Sporadic in all the Gargasian samples.
Parabdolithus infinitus (Worsley, 1971) Thierstein in Roth & Thierstein, 1972
Pl. 9 ![]() , figs. 2-3, 7-8
, figs. 2-3, 7-8
This species exhibits a low birefringence (first order grey) and has two
elliptical central openings aligned with the major ellipse axis. This
nannofossil is frequently overgrown and can be difficult to discern from Calcicalathina
erbae when the two openings are obliterated (see also Bergen,
1998). The
size criterion could be used to discriminate both species: Parabdolithus infinitus
(Pl. 9 ![]() , figs. 2-3, 7-8) is smaller than
Calcicalathina erbae (Pl. 9
, figs. 2-3, 7-8) is smaller than
Calcicalathina erbae (Pl. 9 ![]() , figs. 4-5, 10).
, figs. 4-5, 10).
Distribution: Frequent in the Bedoulian, sporadic in the Gargasian.
Genus Rhagodiscus Reinhardt, 1967
This diverse genus is reserved for loxoliths with low distal rim and a central area filled by a granular plate. The central plate may be perforate and a distel stem may or may not be present. Specimens referred to this genus typically represent between 5-10% of assemblages recovered during this study.
Rhagodiscus angustus (Stradner, 1963) Reinhardt, 1971
Pl. 5 ![]() , figs. 6, 9;
Pl. 9
, figs. 6, 9;
Pl. 9 ![]() , figs. 24-25
, figs. 24-25
This species has a subrectangular outline and a wide hollow stem. The central area is reduced to where the stem periphery may touch the inner distal rim margin.
Distribution: Bergen (1998) placed the lowest occurrence of R. angustus in the middle Bedoulian within Subzone NC6B of Bralower et al. (1993, 1995) based on its evolution from R. gallagheri.
Rhagodiscus asper (Stradner, 1963) Reinhardt, 1967
Pl. 5 ![]() , figs. 1-2, 4-5, 8
, figs. 1-2, 4-5, 8
Coccolith size is variable from small (3-5 μm) to large specimens up to 10 μm. The distal rim is broad; a hollow, circular stem is present.
Distribution: Frequent in all studied samples.
Rhagodiscus gallagheri Rutledge & Bown, 1996
Pl. 5 ![]() , figs. 10-11
, figs. 10-11
This small species (3.5-5 μm) has a narrowly elliptical outline.
Distribution: Present in all samples.
Rhagodiscus splendens (Deflandre, 1953) Verbeek, 1977
Pl. 5 ![]() , figs. 3, 7
, figs. 3, 7
This species has a long hollow, tapered distal stem. The birefringent stem base has a serrate periphery.
Distribution: Sporadic in the samples examined.
Genus Tranolithus (Stover, 1966) Lambert, 1987
Tranolithus gabalus Stover, 1966
Pl. 4 ![]() , fig. 20
, fig. 20
Small to medium-sized coccolith characterized by the presence of two central "block" oriented along the minor ellipse axis.
Distribution: Sporadic in the rich Gargasian marls.
Genus Eiffellithus Reinhardt, 1965
Eiffellithus hancockii Burnett, 1997
Pl. 9 ![]() , fig. 15
, fig. 15
This small nannofossil has a broad, bicyclic rim; the bright inner rim cycle nearly closes the central area. It has a simple axial cross atypical for the genus Eiffellithus.
Distribution: This species occurs sporadically thoughout all the samples, including the Uppermost Bedoulian. Bergen (1998) placed its lowest occurrence in the middle Bedoulian (lower NC6B Subzone).
Genus Calcicalathina Thierstein, 1971
Large loxoliths with a relatively narrow, imbricated distal shield and a large central area filled by a mass of granular elements that rise above the distal rim.
Calcicalathina erbae Bergen, 1998
Pl. 9 ![]() , figs. 4-5, 9-10
, figs. 4-5, 9-10
The diagnosis by Bergen (1998) is "a species of Calcicalathina with a low central area showing sharp distinction between rim (1st order yellow birefringence) and coarsely-granular central area".
Distribution: C. erbae occurs continuously in the Bedoulian (Clavaillan section, Deshayesites grandis Zone) and is rare in the Gargasian marls.
Order STEPHANOLITHIALES Bown & Young, 1997
Family STEPHANOLITHICEAE Black, 1968
Genus Rotelapillus Noël, 1972
Rotelapillus crenulatus (Stover, 1966) Perch-Nielsen, 1984
Pl. 8 ![]() , fig. 2
, fig. 2
This distinct species has a circular to subcircular outline, eight radial central bars, and lateral nodes extending from the rim periphery.
Distribution: This species occurs sporadically in samples examined.
Genus Stoverius Perch-Nielsen, 1986
The diagnosis of the genus is "round to broadly elliptical coccoliths with a wall of more or less vertical elements, a cycle of proximal elements and a central cross." The cross and inner rim cycle exhibit a first order white birefringence, whereas the outer rim cycle exhibits a very faint birefringence. Perch-Nielsen (1986) differentiated Corollithion Stradner, 1961, by its hexagonal outline and Rotelapillus Noël, 1972, by its eight central bars. There is a clear phylogenetic relationship between the genera Stoverius (with St. achylosus) and Corollithion (with C. protosignum and C. signum), see Lambert, 1993.
Stoverius achylosus (Stover, 1966) Perch-Nielsen, 1986
Pl. 7 ![]() , figs. 16-19
, figs. 16-19
The species was described for circular to broadly elliptical coccoliths; the holotype has an orthogonal cross aligned with the ellipse axes.
Distribution: The lowest occurrence of this species was observed in sample 2327 (Les Gays 1 section) at the base of the nannofossil Subzone NC7B. Bergen (1998) placed the lowest occurrence within the middle Bedoulian in Subzone NC6B, immediately below the lowest occurrence of Rhagodiscus angustus. Bralower et al. (1993) placed its base further down in the lower Aptian, immediately below the top of Subzone NC6A.
Genus Corollithion Stradner, 1961
Corollithion ? madagaskarensis Perch-Nielsen, 1973
Pl. 4 ![]() , fig. 8
, fig. 8
Distribution: Rare in the Gargasian marls.
Genus Stradnerlithus Black, 1971a
Stradnerlithus ellipticus (Bukry, 1969) Perch-Nielsen, 1984
Pl. 7 ![]() , fig. 15;
Pl. 8
, fig. 15;
Pl. 8 ![]() ,
fig. 9
,
fig. 9
Distribution: This species occurs sporadically in the Gargasian marls.
Order PODORHABDALES (Rood et al., 1971) Bown, 1987
Family CRETARHABDACEAE Thierstein, 1973
Genus Cretarhabdus Bramlette & Martini, 1964
The central area is occupied by an axial cross and net.
Cretarhabdus conicus Bramlette & Martini, 1964
Pl. 5 ![]() , figs. 12, 18-20
, figs. 12, 18-20
There is variability in coccolith size and central area construction.
Large specimens with distinct axial crosses and vaulted central areas (Pl. 5 ![]() ,
fig. 12) are more typical of the Maastrichtian holotype.
,
fig. 12) are more typical of the Maastrichtian holotype.
Distribution: This species occurs sporadically in samples examined.
Genus Microstaurus Black, 1971a
The thick proximal shield is noticeably smaller than the distal shield; the central opening contains an axial cross-structure.
Microstaurus chiastius (Worsley, 1971) Bralower et al., 1989
Pl. 5 ![]() , fig. 16
, fig. 16
Bralower et al. (1989) emended the species to include specimens with small central areas less than one-half the coccolith length.
Distribution: The species occurs sporadically in the Gargasian.
Microstaurus quadratus Black, 1971a
Pl. 5 ![]() , fig. 21
, fig. 21
Specimens with central areas greater than one-half the coccolith length are referred to this species, following Bralower et al. (1989).
Distribution: M. quadratus occurs sporadically in the samples examined.
Genus Retecapsa Black, 1971a
The genus is distinguished by having an axial cross with lateral bars.
Retecapsa schizobrachiata (Gartner, 1968) Grün in Grün & Allemann, 1975
Pl. 5 ![]() , fig. 17
, fig. 17
The bar is the major axis trifurcates at the rim margin and the one in the minor axis bifurcates. Coccolith size is variable.
Distribution: The species is sporadic in Gargasian samples.
Genus Grantarhabdus Black, 1971a
The genus is distinguished by its diagonal central cross.
Grantarhabdus meddii Black, 1971a
Pl. 5 ![]() , fig. 22
, fig. 22
The diagonal cross is symmetric to the ellipse axes and the bars form an acute angle to the minor ellipse axis. The holotype of Grantarhabdus unicornis (Stover, 1966) Black, 1972, has a diagonal cross asymmetric to the ellipse axes.
Distribution: This species occurs sporadically in the samples examined.
Order PODORHABDALES (Rood et al., 1971) Bown, 1987
Family AXOPODORHABDACEAE Bown & Young, 1997
Genus Axopodorhabdus Wind & Wise in Wise & Wind, 1977
Axopodorhabdus dietzmannii (Reinhardt, 1965) Wind & Wise, 1983
Pl. 6 ![]() , figs. 4, ? 8
, figs. 4, ? 8
This medium to large species of Axopodorhabdus is distinguished by its narrowly elliptical outline and broad transverse bars.
Distribution: It occurs sporadically in the Gargasian.
Genus Tetrapodorhabdus Black, 1971a
Tetrapodorhabdus coptensis Black, 1971a
Pl. 6 ![]() , figs. 2-3, ? 5, 6
, figs. 2-3, ? 5, 6
Small to medium-sized species with bars that form an acute angle with the minor ellipse axis.
Distribution: It occurs sporadically in the samples examined.
Tetrapodorhabdus decorus (Deflandre in Deflandre & Fert,
1954)
Wind & Wise in Wise
& Wind, 1977
Pl. 6 ![]() , figs. ? 1, 7
, figs. ? 1, 7
The holotype is a light photomicrograph in profile. In plan view, it has been associated with a symmetric diagonal cross where the orthogonal bars flare near the rim margin and form circular openings.
Distribution: Sporadic in the samples.
Genus Acaenolithus Black, 1973
This genus (first described by Black for Albian specimens) is related to the Family Arkhangelskiellacea well represented in Upper Cretaceous series.
Acaenolithus ? sp.
Pl. 3 ![]() , fig. 23
, fig. 23
We ascribed to this genus very small coccoliths with alternate extinction of the cross arms. In the absence of observations in SEM, it seems very difficult to precisely determine the ultra structure.
Distribution: Sporadic in the richest Gargasian samples.
aff. Misceomarginatus Wind & Wise in Wise & Wind, 1977
Pl. 3 ![]() , figs. 24-25
, figs. 24-25
The general aspect including rims and central cross is close to Misceomarginatus (and also to the close genus Diloma Wind & Cepek, 1979). The quadrants seem free or spanned by a diagonal bar.
Distribution: Sporadic in the samples.
Family COCCOLITHACEAE Noël, 1965
Circular or elliptical coccoliths composed by two superimposed rim composed by radial calcitic elements with a strong angle between them in the external part (junction of the two rims). This particularity allows a very good cohesion between the coccoliths on the coccosphere. The two rims are linked in their internal sides by a calcitic tube. The central area is occupied (or not) by variable filling taking place indistinctly on the proximal or distal rims.
Genus Flabellites Thierstein, 1973
Original diagnosis: "Presence of an inner cycle of the distal shield composed of elongated calcitic elements, one or two clusters of these elements often extend beyond the elliptical periphery of the coccolith" (Thierstein, 1973, p. 41).
Flabellites biforaminis Thierstein, 1973
Pl. 6 ![]() , figs. 9-11
, figs. 9-11
Morphologic variation observed includes coccolith size and outline, as well as the size and shape of the central openings.
Distribution: This species is present in all our samples.
Family BISCUTACEAE Black, 1971a
Genus Biscutum Black in Black & Barnes, 1959
Biscutum is retained for non-imbricating, elliptical placoliths constructed of two broad shields and an inner distal tube cycle. In cross-polarized light, the shields are faint and the narrow tube cycle is bright. Palaeopontosphaera Noël, 1965, is considered a junior synonym herein; the two species included within Biscutum are taxonomically problematic due to uncertainties regarding their holotypes. Discussion of these genera and their included species is a topic beyond the current study (see Lambert, 1993, p. 206).
Biscutum constans (Górka, 1957) Black, 1968
Pl. 6 ![]() , fig. 17
, fig. 17
Górka (1957) described this species from the upper Maastrichtian and indicated a length of 5-9 μm. It is founded on a rudimentary drawing of a normally elliptical specimen (1.38 aspect ratio) shown to have a single cycle of fifteen radial elements (19-20 described) and an open center (described as smooth) approximately 0.44 of the coccolith width. Current usage has evolved into a small to medium (3-8 µm), normally elliptical species with a narrow central area, bright tube cycle and dark central plate (see Young et al., 2022). That concept is followed herein.
Distribution: Biscutum constans and B. dubium have been counted as a single taxon and are frequent in samples (5-15% of assemblages).
Biscutum dubium (Noël, 1965) Grün in Grün et al., 1974
Pl. 6 ![]() , fig. 22
, fig. 22
Small elliptical coccoliths with acentral area nearly filled by a distal boss are identified as this species (see Grün & Zweili, 1980; Kaenel & Bergen, 1993). Biscutum ellipticum (Górka, 1957) is not appropriate because the original description indicates a very large size (length 18 μm) and the accompanying sketch of an Upper Maastrichtian specimen shows ten rim elements with curved sutures and an open center. The presence of a central boss distinguishes B. dubium from B. constans.
Genus Discorhabdus Noël, 1965
Discorhabdus ignotus (Górka, 1957) Perch-Nielsen, 1968
Pl. 6 ![]() , fig. 16
, fig. 16
Very small circular placolithswith faint birefringence and a closed center were identified as this constant background species (2.5-5% of assemblages).
Discorhabdus serratus Worsley, 1971
Pl. 9 ![]() , figs. 1, 6
, figs. 1, 6
Shields composed by 8 to 12 elements with blunt terminations. The central area is occupied by a stem (distal view) and not by a central open area as mentionned by Worsley (1971).
Distribution: This form was found only in the lower part of the Clavaillan section (Deshayesites grandis Zone).
Genus Haqius Roth, 1978
Large circular coccolith wih a reduced central area.
Haqius circumradiatus (Stover, 1966) Roth, 1978
Pl. 6 ![]() , figs. 18, 23
, figs. 18, 23
This large circular coccolith is easily recognizable. The rim is composed of 30-35 elements and exhibits a faint first order white birefringence.
Distribution: This species occurs sporadically in samples.
Genus Watznaueria Reinhardt, 1964
Three species were assigned to this elliptical genus, which comprises 20-35% of the nannoplankton population in samples examined.
Watznaueria barnesae (Black, 1959) Perch-Nielsen, 1968
Distribution: This species is common to abundant in samples.
Watznaueria britannicus (Stradner, 1963) Reinhardt, 1964
Pl. 6 ![]() , figs. 20-21
, figs. 20-21
Specimens with disjunct bridges were assigned to this species, which is much less frequent than Watznaueria barnesae.
Distribution: Present in all samples.
Watznaueria ovata Bukry, 1969
Pl. 7 ![]() , fig. 11
, fig. 11
Specimens observed with large central openings were typically smaller than the medium-sized specimens originally illustrated by Bukry (1969).
Distribution: The species occurs sporadically in the Gargasian.
Genus Pickelhaube Applegate et al. in Covington & Wise, 1987
Pickelhaube furtiva (Roth, 1983) Applegate et al. in Covington & Wise, 1987
Pl. 7 ![]() , fig. 4
, fig. 4
This large, delicate nannofossil was often observed broken in Gargasian sediments, where it is very rare.
Genus Crucibiscutum Jakubowski, 1986
This genus is reserved for elliptical Biscutaceae with a central tube and axial central structure.
Crucibiscutum salebrosum (Black, 1971a) Jakubowski, 1986
Pl. 7 ![]() , figs. 5-9
, figs. 5-9
The broad species concept maintained herein includes variations in coccolith size, the ratio of the central area size to rim width, and the construction of the axial cross. Crucibiscutum salebrosum has priority over other Cretaceous species that may be distinguished on these characteristics.
Distribution: Present throughout the Gargasian.
Genus Manivitella Thierstein, 1971
Manivitella pemmatoidea (Deflandre in Manivit, 1965) Thierstein, 1971
Pl. 7 ![]() , fig. 14
, fig. 14
The similar large species Tubodiscus jurapelagicus (Worsley, 1971) Roth, 1973, was not differentied during sample analyses.
Distribution: This taxon occurs sporadically in the samples examined.
Genus Tubodiscus Thierstein, 1973
Tubodiscus burnettiae Bown in Kennedy et al., 2000
Pl. 7 ![]() , figs. 12-13
, figs. 12-13
This medium-sized species of Tubodiscus has a relatively broad inner rim cycle.
Genus Diazomatolithus Noël, 1965
Description: Circular to subcircular coccolith with a central area always empty.
Diazomatolithus lehmanii Noël, 1965
Pl. 6 ![]() , figs. 12-15
, figs. 12-15
Outside of size, variants with narrow (Pl. 6 ![]() , figs. 12-13) and broad
(Pl. 6
, figs. 12-13) and broad
(Pl. 6 ![]() ,
figs. 14-15) rims were observed. Those with narrow rims and large centers may
represent Diazomatolithus galicianus
Kaenel & Bergen, 1996, but were not differentiated
during sample analyses.
,
figs. 14-15) rims were observed. Those with narrow rims and large centers may
represent Diazomatolithus galicianus
Kaenel & Bergen, 1996, but were not differentiated
during sample analyses.
Distribution: Although Diazomatolithus does not reach the NC7B subzone in our samples, this Late Jurassic genus is recorded above the Aptian.
Order BRAARUDOSPHAERALES (Aubry, 2013) Lees & Bown, 2016
Lees and Bown (2016) included Braarudosphaeraceae Deflandre, 1947, and Nannoconaceae Deflandre, 1959, within their emended definition of this order. Young et al. (2022) tentatively placed the Polycyclolithaceae Forchheimer, 1972, within this order.
Family BRAARUDOSPHAERACEAE Deflandre, 1947
Pentagonal coccoliths constructed of superimposed plates. Genera are distinguished by the intersection of the sutures and periphery. The present-day species Braarudosphaera bigelowii is composed of a single layer of pentagonal elements (12 per coccosphere), whereas fossil species can be more complex with individual coccoliths composed of numerous layers (see Lambert, 1986).
Genus Braarudosphaera Deflandre, 1947
Braarudosphaera africana Stradner, 1961
Pl. 7 ![]() , fig. 22
, fig. 22
Distribution: This species was not observed in the lower part of the Clavaillan section (Bedoulian) and is sporadic in the Gargasian. Bergen (1998) placed the base of Br. africana in lowermost Subzone NC7A, immediately above the base of Eprolithus floralis.
Braarudosphaera bigelowii (Gran & Braarud, 1935) Deflandre, 1947
Pl. 7 ![]() , figs. 20-21
, figs. 20-21
Distribution: The species occurs sporadically in the samples examined.
Genus Micrantholithus Deflandre in Deflandre & Fert, 1954
The highest occurrences of both species of Micrantholithus coincide with the "Nannoconus crisis level" in the materials studied. Bralower et al. (1993) defined the top of Subzone NC7A on the highest occurrence of Micrantholithus hoschulzii, whereas Herrle and Mutterlose (2003) modified the definition of the genus.
Micrantholithus hoschulzii (Reinhardt, 1966) Thierstein, 1971
Each element has a triangular shape.
Micrantholithus obtusus Stradner, 1963
Pl. 8 ![]() , fig. 10;
Pl. 9
, fig. 10;
Pl. 9 ![]() , fig. 14
, fig. 14
The elements have a typical V-shaped outline due to their indented peripheries.
Family CALCIOSOLENIACEAE Kampter, 1937
Genus Calciosolonia Gran, 1912
Calciosolenia fossilis (Deflandre in Deflandre & Fert, 1954) Bown in Kennedy et al., 2000
Pl. 8 ![]() , figs. 21-23
, figs. 21-23
Relative to modern coccolithophores, individual coccolith morphology could vary
from large rhombohedra in the middle part of the cell (Pl. 8 ![]() , fig. 23) to
elongated rhomboedra in the distal part of the cell (Pl. 8
, fig. 23) to
elongated rhomboedra in the distal part of the cell (Pl. 8 ![]() , figs. 21-22).
, figs. 21-22).
Distribution: Sporadic throughout the samples studied.
Family CALYPTROSPHAERACEAE Boudreaux & Hay, 1969
Genus Orastrum Wind & Wise in Wise & Wind, 1977
Orastrum perspicuum Varol in Al-Rifaiy et al., 1990
Pl. 7 ![]() , figs. 23-24
, figs. 23-24
Coccolith composed of a single plate surrounded by a narrow rim. The central plate is less birefringent than the rim.
Distribution: This lowest occurrence of this species is in Subzone NN7B in the material studied. Herrle and Mutterlose (2003) placed this event below the N. truitti acme in Subzone NN7B/C.
Genus Isocrystallithus Verbeek, 1977
Isocrystallithus partitum (Varol in Al-Rifaiy et al., 1990) Bergen, 1998
Pl. 8 ![]() , figs. 17-18
, figs. 17-18
This holococcolith has two central plates separated by a transverse bridge composed of four calcite elements. The original description of this species suggested that these four elements could be the remnants of a short distal process. Bergen (1998) first associated these basal plates with complete specimens having short, tapered distal processes and transferred the species into Isocrystallithus. The genus Orastrum does not have a distal process or central bridge.
Distribution: This species is sporadic in the Gargasian marls.
Genus Calculites Prins & Sissingh in Sissingh, 1977
Calculites dispar Varol in Al-Rifaiy et al., 1990
Pl. 8 ![]() , figs. 15-16;
Pl. 9
, figs. 15-16;
Pl. 9 ![]() , fig. 18
, fig. 18
These small elliptical holococcolith are composed by four unequal plates surrounded by a narrow rim. A longitudinal suture separates the two smaller central plates symmetric across the minor ellipse axis. Obtuse angled sutures extend from the poles of the longitudinal suture to delineate two larger plates symmetric across the major ellipse axis.
Distribution: This species is sporadic in the upper Bedoulian and Gargasian marls.
Genus Zebrashapka Covington & Wise, 1987
The original description and electron photomicrographs of this monopsecific genus by Covington and Wise (1987) indicate that this is a holococcolith.
Zebrashapka vanhintei Covington & Wise, 1987
Pl. 8 ![]() , figs. 3-5
, figs. 3-5
This holococcolith is easily recognizable.
Distribution: Sporadic in rich Gargasian samples.
Family POLYCYCLOLITHACEAE (Forchheimer, 1972) Varol, 1992
Perch-Nielsen (1985) assigned Assipetra Roth, 1973, and Hayesites Manivit, 1971, to this family, but they do not conform to the emended definition of Polycyclolithaceae by Varol (1992) due to their lack of a two-tiered construction. Bown and Young (1997) referred to Assipetra and Hayesites as "uncertain polycycloliths." They are tentatively placed in this family herein because of their radial symmetry and petaloid elements.
Genus Eprolithus Stover, 1966
Eprolithus has a wall constructed of two tiers of vertical to sub-vertical, petaloid elements (5-9) connected by a medial diaphragm.
Eprolithus floralis (Stradner, 1962) Stover, 1966
Pl. 8 ![]() , figs. 6-8;
Pl. 9
, figs. 6-8;
Pl. 9 ![]() , figs. 21-23
, figs. 21-23
Radiolithus planus
Stover, 1966 (Pl. 9 ![]() ,
figs. 21-22) was included within Eprolithus
floralis during sample analyses.
,
figs. 21-22) was included within Eprolithus
floralis during sample analyses.
Distribution: The lowest occurrence of Eprolithus floralis marks the base of Subzone NC7A, which occurs below the lowest sample examined for this study (see Bergen, 1998; Herrle & Mutterlose, 2003; Giraud et al., 2018).
Family LAPIDEACASSACEAE Bown & Young, 1997
Genus Lapideacassis Black, 1971b
Lapideacassis glans Black, 1971b
Pl. 8 ![]() , figs. 11-12;
Pl. 9
, figs. 11-12;
Pl. 9 ![]() , figs. 12-13, 17
, figs. 12-13, 17
This species was distinguished by its low, broad profile and irregular outline.
Lapideacassis mariae Black, 1971b
Pl. 8 ![]() , fig. 13
, fig. 13
This species was distinguished by its high, relatively narrow profile and irregular outline. No intermediate forms have been observed between this species and L. glans.
Distribution: Lapideacassis mariae and Lapideacassis glans are sporadic in the studied samples, including the lower Aptian (Bedoulian). Bergen (1998) did not report the genus from the lower Aptian historical stratotype. Herrle and Mutterlose (2003) placed the lowest occurrences of both species in the Gargasian (Subzone NC7B/C).
Lapideacassis sp.
Pl. 9 ![]() , figs. 11, 16
, figs. 11, 16
Rare specimens with relatively high, narrow profiles and smooth outlines were observed, but more information is needed to interpret their relationship to the other two species of Lapideacassis.
Distribution: This nannofossil was observed only in the lower part of the Clavaillan section (Bedoulian).
Family MICRORHABDULACEAE Deflandre, 1963
Genus Lithraphidites Deflandre, 1963
Lithraphidites carniolensis Deflandre, 1963
Pl. 8 ![]() , fig. 14
, fig. 14
Distribution: This species is sporadic in all the samples examined.
Genus Hayesites Manivit, 1971
Hayesites irregularis (Thierstein in Roth & Thierstein, 1972) Applegate et al. in Covington & Wise, 1987
Pl. 8 ![]() , figs. 19-20
, figs. 19-20
Distribution: This Aptian-Albian species occurs sporadically in the samples examined.
Genus Assipetra Roth, 1973
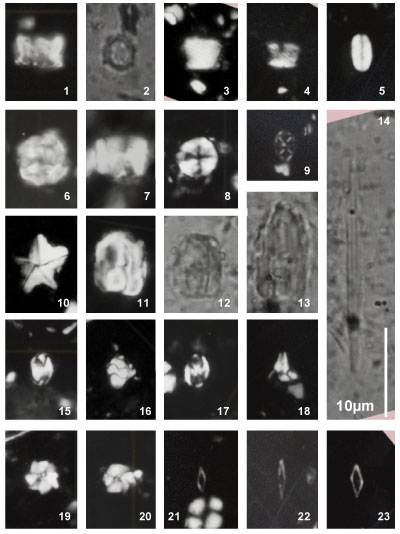
Assipetra infracretacea (Thierstein, 1973) Roth, 1973
Pl. 3 ![]() , fig. 6;
Pl. 10
, fig. 6;
Pl. 10 ![]() , fig.
22
, fig.
22
Thierstein (1973) described this species as "having two sets of flat crystal plates, one set piercing the other at an obtuse angle" and remarked that light microscope specimens having subrectangular to suboval shapes with 4-6 radial sutures. All four specimens illustrated by Thierstein (1973) - transferred between the SEM and LM - appear subrectangular and suboval outlines (op. cit., Pl. 1, figs. 6, 19) could be a consequence of specimen orientation. Aspect ratios of these four original specimens vary between 1.1-1.3 and a maximum diameter of 5-10 μm was given for the species. The diagnosis of A. infracretacea larsonii by Tremolada and Erba (2002) restricted this subspecies to specimens greater than 7.5 μm, but Bown (2005) considered a number of these specimens to be side views of Assipetra terebrodentarius. A size of 10 μm is used herein to separate two morphotypes of A. infracretacea.
Assipetra terebrodentarius (Applegate et al. in Covington & Wise, 1987) Rutledge & Bergen in Bergen, 1994
Pl. 3 ![]() , fig. 12;
Pl. 10
, fig. 12;
Pl. 10 ![]() , figs.
21, 23-24
, figs.
21, 23-24
Applegate et al. (in Covington & Wise, 1987) stated that Assipetra terebrodentarius "is more globular, has more numerous and angular projecting elements, and often displays a regular, spiral construction not observed in A. infracretacea". The diagnosis of A. terebrodentarius youngii by Tremolada and Erba (2002) restricted this subspecies to specimens greater than 7.5 μm. Bown (2005) emended this subspecies for specimens greater than 8.0 μm because the species holotype is 7.7 μm. A size of 10 μm is used herein to separate two morphotypes of A. terebrodentarius (see §3. Materials and methods).
Distribution (including A. infracretacea, Fig. 3 ![]() ): The semi-quantitative analysis of the distribution of
these two species that we carried out (see §3. Materials and methods) shows
that the record of these nannofossils is not continuous with two intervals where
they are relatively frequent:
): The semi-quantitative analysis of the distribution of
these two species that we carried out (see §3. Materials and methods) shows
that the record of these nannofossils is not continuous with two intervals where
they are relatively frequent:
lower Aptian, Deshayesites grandis Zone and
a part of the upper Gargasian, middle part of the Dufrenoyia furcata Zone.
For these two intervals the A.i + A.t large forms / A.i + A.t small forms observed confirm the 2002 Tremolada and Erba' work. The large specimens are clearly more frequent in the lower Aptian (Deshayesites grandis Zone) of the the studied section (i.e., the lower portion the Clavaillan section) compared to the upper Aptian (Dufrenoyia furcata Zone).
Genus Tetralithus Gardet, 1955
The genus Tetralithus has been created in 1955 for Miocene nannofossils probably reworked from Cretaceous sediments. For a discussion of differences between Tetralithus and Quadrum the readers are referred to Prins and Perch-Nielsen (in Manivit et al., 1977).
Tetralithus ? malticus Worsley, 1971
Pl. 9 ![]() , fig. 20
, fig. 20
This small, square nannolith is composed of four elements separated by orthogonal sutures that intersect the midpoints of the peripheries. Individual elements are wedge-shaped, being thicker along the axes offset 45 degrees to the sutures. This form resembles Quadrum, but four species of that genus are restricted to the Upper Cretaceous.
Distribution: This taxon was observed only in the basal portion of the Clavaillan section (Bedoulian).
Genus Conusphaera Trejo, 1969
Conusphaera rothii (Thierstein, 1971) Jakubowski, 1986
Pl. 9 ![]() , fig. 19
, fig. 19
Distribution: This species was observed sporadically in the Clavaillan section (Bedoulian) along with Eprolithus floralis. It is possible that these rare, sporadic specimens are redeposited. Bralower et al. (1993) defined the base of Subzone NC6B on the highest occurrence of Conusphaera rothii and the top of this subzone on the lowest occurrence of Eprolithus floralis. The stratigraphic ranges of these two marker species are separated by approximately 63 m (22 samples) in the historical Bedoulian stratotype (Bergen, 1998).
Family NANNOCONACEAE Deflandre, 1959
Genus Nannoconus Kamptner, 1931
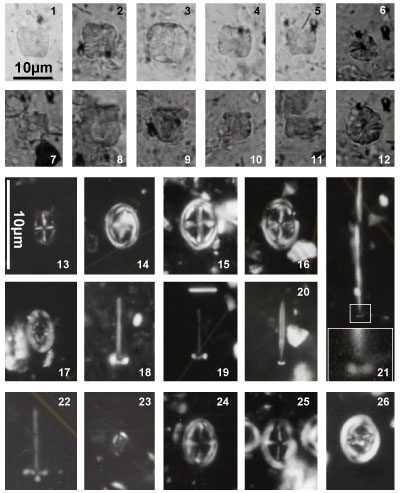
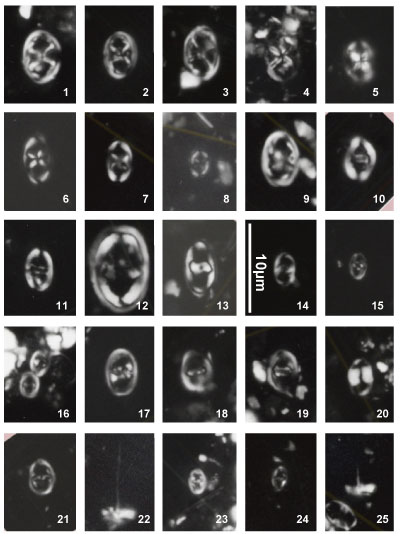
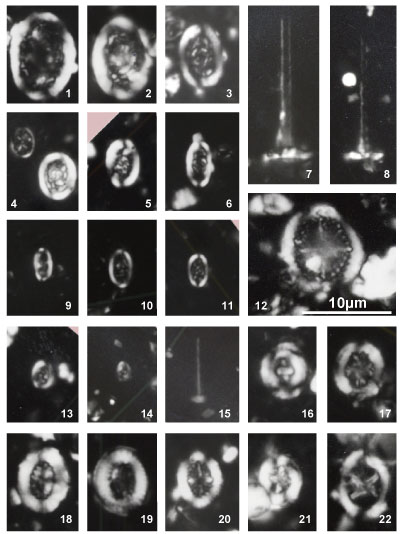
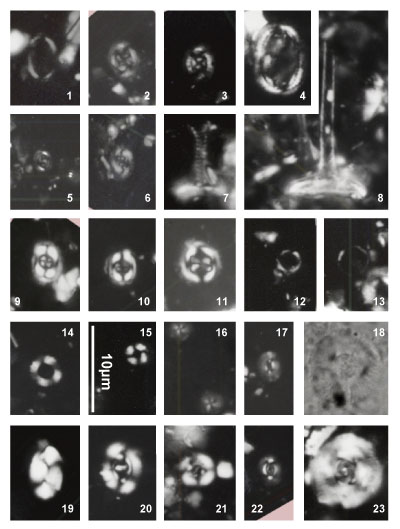
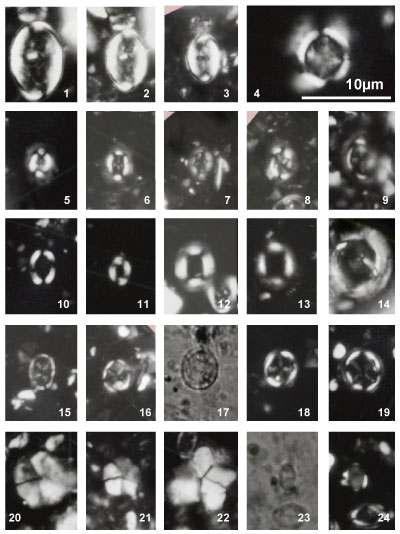
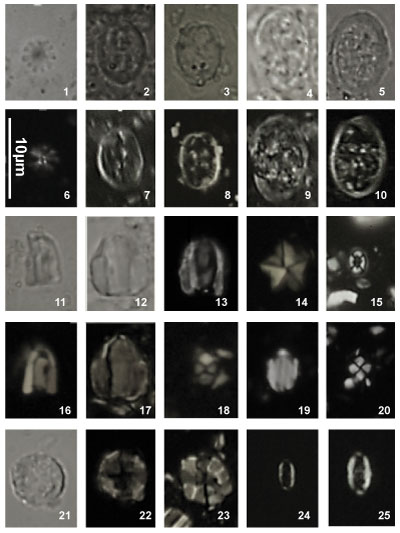
Nannoconus specimens are illustrated both in the photographic plates and as drawn
profiles of actual specimens (Fig. 2 ![]() ). These profiles were drawn from
photographs of the first thirty specimens encountered in each of 29 samples (red
dots in Fig. 2
). These profiles were drawn from
photographs of the first thirty specimens encountered in each of 29 samples (red
dots in Fig. 2 ![]() ). This process has been used to illustrate the significant
morphological variations. These variations make it often difficult the direct
reference to the original diagnosis (holotype). Many Nannoconus specimens
do not fit with the corresponding diagnosis or/and present a lot of intermediate
forms between two type species (see for instance on Fig. 2
). This process has been used to illustrate the significant
morphological variations. These variations make it often difficult the direct
reference to the original diagnosis (holotype). Many Nannoconus specimens
do not fit with the corresponding diagnosis or/and present a lot of intermediate
forms between two type species (see for instance on Fig. 2 ![]() the group B with N.
globulus / N. circularis).
the group B with N.
globulus / N. circularis).
The taxonomic concepts of Deres and Achéritéguy (1980) are followed. Please refer to this publication for more taxonomic information. Fifteen species - including three subspecies - have been organized into five groups based on strong morphological affinities. All have central cavities. They are presented herein relative to these groupings:
Nannoconus steinmannii has a conical outline and is the only species with a central canal.
Group A: constricted outline
Nannoconus boletus Deflandre & Deflandre-Rigaud, 1962
Pl. 1 ![]() , figs. 12, ? 13; Pl. 2
, figs. 12, ? 13; Pl. 2 ![]() , figs. 6, 31, ? 32;
Pl. 3
, figs. 6, 31, ? 32;
Pl. 3 ![]() , figs. ? 1, 2, ? 3, 4-5
, figs. ? 1, 2, ? 3, 4-5
Nannoconus carniolensis Deflandre & Deflandre-Rigaud, 1962
Pl. 3 ![]() , figs. ? 7, 8-11
, figs. ? 7, 8-11
Both species included in this group were first illustrated in 1962 and formally described five years later. The group includes nannoconids with a distinct external restriction at the middle to upper portion of the test, resulting in a more or less mushroom-shaped outline in longitudinal view ("boletus edulis" is a well-known and appreciated mushroom). The distinction between the two species is often subtle. N. carniolensis is shaped more like a pot, whereas N. boletus is more fungiform. Both species are small to medium-sized (< 10 μm), in congruence with specimens first illustrated by Deflandre in 1962. The oldest specimens recovered from the upper part of the Clavaillan section are more rectangular in longitudinal outline with a low relief constrictions and roughly equivalent wall to and central cavity thicknesses.
Distribution: N. boletus occurs sporadically throughout the Gargasian, but is more frequent around the NC7A/NC7B boundary. The smallest forms of N. carniolensis, which are uncommon overall, seem restricted to Nannoconus assemblage zone GIIB.
Group B: rounded outline
Nannoconus circularis Deres & Achéritéguy, 1980
Pl. 1 ![]() , figs. 10-11, 31; Pl. 2
, figs. 10-11, 31; Pl. 2 ![]() , figs. 25-26, 28;
Pl. 10
, figs. 25-26, 28;
Pl. 10 ![]() , fig. 15
, fig. 15
Nannoconus aff. globulus Brönnimann, 1955
Pl. 1 ![]() , figs. 8-9, 30; Pl. 2
, figs. 8-9, 30; Pl. 2 ![]() , figs. 23-24, ? 27
, figs. 23-24, ? 27
This group includes more or less circular to rectangular forms with a large internal cavity and a thin wall (wall thickness << internal cavity). From a semi-quantitative point of view it seems to be very difficult to distinguish the two species which appears in the Aptian sediments and probably represent two poles of a same population. Depending to the preservation (calcite overgrow) is it possible to transform a thin walled with large central cavity N. circularis in a more robust and thick walled with a more narrow central cavity. N. circularis has frequently a quadrangular shape compared to N. aff. globulus with is globular shape.
Distribution: This group is abundant throughout Subzone NC7A, but absent from Subzone NC7B.
Group C: cylindrical outline
Nannoconus elongatus Brönnimann, 1955
Pl. 10 ![]() , figs. 6-7, ? 8
, figs. 6-7, ? 8
This relatively large cylindrical Nannoconus (> 10 μm) has a thick wall and open central cavity approximately equal in width.
Distribution: This rare species was observed only in the lower part of the Clavaillan section.
Nannoconus inconspicuus Deflandre & Deflandre-Rigaud, 1962
Pl. 10 ![]() , fig. 16
, fig. 16
Nannoconus quadriangulus Deflandre & Deflandre-Rigaud, 1962
Pl. 1 ![]() , figs. 14-15, 17-18, 32; Pl. 2
, figs. 14-15, 17-18, 32; Pl. 2 ![]() , figs. 1-2, 7-16, 29, 31
, figs. 1-2, 7-16, 29, 31
Nannoconus truittii Brönnimann, 1955
Pl. 1 ![]() , figs. 16, 33; Pl. 2
, figs. 16, 33; Pl. 2 ![]() , figs. 3-5
, figs. 3-5
Two subspecies of Nannoconus quadriangulus (i.e., N. quadriangulus quadriangulus and N. quadriangulus apertus) were first illustrated in 1962, but formally described in 1967 by Deflandre and Deflandre-Rigaud. Both subspecies are not considered herein. They correspond to small to medium-sized (up to 9 µm) quadrangular forms with a relatively thick wall and a rectangular central cavity (ratio wall thickness / central cavity close to 1). These authors described also a very small species Nannoconus inconspicuus (length around 2 µm). This group includes all the rectangular to subrectangular forms (including the smallest ones) varying from 2/3 to 8/9 µm. N. truitti, another small quadrangular form with a thicker wall, which was first described by Brönnimann in 1955, is also ascribed to this group.
Group D: short, conical outline (horseshoe-shaped)
Nannoconus bucheri Brönnimann, 1955
Pl. 1 ![]() , figs. 2, 5,
28-29; Pl. 10
, figs. 2, 5,
28-29; Pl. 10 ![]() , figs. 12-14
, figs. 12-14
Nannoconus vocontiensis Deres & Achéritéguy, 1980
Pl. 1 ![]() , figs. 1, 3-4, 6-7, 30; Pl. 2
, figs. 1, 3-4, 6-7, 30; Pl. 2 ![]() , figs. 17-22;
Pl. 10
, figs. 17-22;
Pl. 10 ![]() , fig. 25
, fig. 25
Apparently these two species seem to represent a continuous lineage between thicker (wall) forms (N. bucheri) and thinner (wall) forms (N. vocontiensis) both species with a horseshoe type. The largest forms (10-15 µm, N. bucheri) are found mainly in the upper Bedoulian and lower Gargasian and the smaller ones (around 10 µm, N. vocontiensis) in the lower Gargasian.
Distribution: This group is common throughout and restricted to Subzone NC7A.
Group E: elongate, conical outline
Nannoconus bonetii Trejo, 1959
Pl. 10 ![]() , figs. 11, 20
, figs. 11, 20
Nannoconus aff. borealis Perch-Nielsen, 1979
Pl. 10 ![]() , fig. 10
, fig. 10
Nannoconus aff. grandis Deres & Achéritéguy, 1980
Pl. 1 ![]() , fig. 19;
Pl. 10
, fig. 19;
Pl. 10 ![]() , figs. 18-19
, figs. 18-19
Nannoconus kamptneri Brönnimann, 1955
Pl. 1 ![]() , fig. 20;
Pl. 10
, fig. 20;
Pl. 10 ![]() , fig. 17
, fig. 17
Nannoconus wassallii Brönnimann, 1955
Pl. 1 ![]() , figs. 21-27;
Pl. 10
, figs. 21-27;
Pl. 10 ![]() , fig. 9
, fig. 9
In this group we have associated the very large / large forms (up to 25 µm) found in the Clavaillan section Gargasian and in the basal part of the south Pichouraz section. All these species include elongated conical (i.e., Nannoconus aff. grandis and N. kamptneri) to pear-shaped tests with a typical "pear" aspect. We can see a clear relationship between the more elongated species:
Distribution: This group has been mainly observed in the Clavaillan upper part (Gargasian) and in the south Pichouraz two basal samples (i.e., 2270, 2272). However some N. wassalii occur sporadically in the Bedoulian and in the GII Nannoconus Zone.
Other species:
Nannoconus steinmannii Kamptner, 1931
Pl. 10 ![]() , figs. 1-3
, figs. 1-3
Nannoconus cf. steinmannii Kamptner, 1931
Pl. 10 ![]() , figs. 4-5
, figs. 4-5
These conical to sub-conical Nannoconus (i.e., tapers apically) have a very narrow central canal.
Distribution: Very rare specimens were observed in the uppermost Bedoulian of the Clavaillan section (Subzone NC7A) whereas Bralower et al. (1993) and Bergen (1998) placed its highest occurrence in Subzone NC6B.
Many thanks to the people who have supplied helpful assistance and reviewed carefully this work. A special attention to Jean-Michel Villain, former head of TOTAL's micropaleontology service who provide me technical assistance. Special thanks to Bruno Granier and Michel Moullade for their support and patience, this work started in 2005 has been completed in 2019 by the addition of the Clavaillan section (for this last section thanks to Christine Balme and Stéphane Legal from the "Parc Naturel Regional du Luberon"). Many thanks to Fabienne Giraud who first have had the difficult task to review the manuscript. Many thanks to James Bergen who also devoted a lot of his time to reviewing this work. His extensive experience with nannofossils was fundamental for the finalization of this contribution. The substance and the form owe him a lot.
Al-Rifaiy I., Varol O. & Lemone D. (1990).- Middle to Late Albian biostratigraphy of the Cuchillo Formation from Sierra de Sapello, Mexico.- Newsletters on Stratigraphy, Stuttgart, vol. 21, no. 3, p. 187-200.
Atrops F. & Dutour Y. (2005).- Biostratigraphie et faunes d'ammonites de l'Aptien supérieur (Gargasien) de la région stratotypique de Gargas, prčs d'Apt (Sud-Est de la France). In: Grosheny D. (éd.), Impact environnemental sur la dynamique des populations au Crétacé.- Réunions thématiques du Groupe Français du Crétacé (Paris, 5-6 Décembre 2005), Volume des Résumés, p. 12-15 (abstract).
Barrier J. (1977a).- Nannofossiles calcaires des marnes de l'Aptien inférieur type : Bédoulien de Cassis-la-Bédoule (Bouches-du-Rhône).- Bulletin du Muséum national d'Histoire naturelle (Sciences de la Terre), Paris, vol. 59, p. 1-68.
Barrier J. (1977b).- Nannofossiles calcaires du Gargasien stratotypique.- Bulletin du Muséum national d'Histoire naturelle (Sciences de la Terre), Paris, vol. 62, p. 173-228.
Bischoff G. & Mutterlose J. (1998).- Calcareous nannofossils of the Barremian/Aptian boundary interval in NW Europe: Biostratigraphic and paleoecologic implications of a high resolution study.- Cretaceous Research, vol. 19, no. 5, p. 635-661.
Bischoff J.A. (1994).- Berriasian to early Aptian calcareous nannofossils from the Vocontian Trough (SE France) and Deep Sea Drilling Site 534: New nannofossil taxa and a summary low-latitude biostratigraphic events.- Journal of Nannoplankton Research, Seward - NE, vol. 16, no. 2, p. 59-69. URL: http://ina.tmsoc.org/JNR/online/16/Bergen%201994%20JNR%2016-2%20 Berr-Aptian%20[%C2%A7N467].pdf
Bergen J.A. (1998).- Calcareous nannofossils from the lower Aptian historical stratotype at Cassis-La Bédoule. In: Moullade M., Tronchetti G. & Masse J.-P. (éds.), Le stratotype historique de l'Aptien inférieur (Bédoulien) dans la région de Cassis-La Bédoule (S.E. France).- Géologie Méditerranéenne, Marseille, t. XXV, nos. 3-4, p. 227-255. URL: https://www.persee.fr/doc/geol m_0397-2844_1998_num_25_3_1638
Black M. (1968).- Taxonomic problems in the study of Coccoliths.- Palaeontology, Durham, vol. 11, no. 5, p. 793-813.
Black M. (1971a).- Coccoliths of the Speeton Clay and Sutterby Marl.- Proceedings of the Yorkshire Geological Society, Keyworth, vol. 38, no. 3, p. 381-424.
Black M. (1971b).- Problematic microfossils of the Gault Clay.- Geological Magazine, Cambridge (UK), vol. 108, no. 4, p. 325-327.
Black M. (1972).- British Lower Cretaceous coccoliths, I, Gault Clay, Part 1.- Monograph of the Palaeontographical Society, London, vol. 126, no. 534, p. 1-48.
Black M. (1973).- British Lower Cretaceous coccoliths, I, Gault Clay, Part 2.- Monograph of the Palaeontographical Society, London, vol. 127, no. 537, p. 49-112.
Black M. (1975).- British Lower Cretaceous coccoliths, I, Gault Clay, Part 3.- Monograph of the Palaeontographical Society, London, vol. 129, no. 543, p. 113-142.
Black M. & Barnes B. (1959).- The structure of coccoliths from the English Chalk.- Geological Magazine, Cambridge (UK), vol. 96, no. 5, p. 321-328.
Bornemann A. & Mutterlose J. (2006).- Size analyses of the coccolith species Biscutum constans and Watznaueria barnesae from the late Albian "Niveau Breistroffer" (SE France): Taxonomic and palaeoecological implications.- Géobios, Villeurbanne, vol. 39, no. 5, p. 599-615.
Boudreaux J.E. & Hay W.W. (1969).- Calcareous nannoplankton and biostratigraphy of the late Pliocene-Pleistocene-Recent sediments in the Submarex cores.- Revista Espańola de Micropaleontología, Madrid, vol. 1, no. 3, p. 249-292.
Bown P.R. (1987).- Taxonomy, evolution and biostratigraphy of Late Triassic-Early Jurassic calcareous nannofossils.- Special Papers in Palaeontology, Durham, no. 38, 118 p. URL: https://www.palass.org/sites/default/files/media/publications/special_pa pers_in_palaeontology/number_38/SPP38.pdf
Bown P.R. (2005).- Early to mid-Cretaceous calcareous nannoplankton from the northwest Pacific Ocean, ODP Leg 198, Shatsky Rise. In: Bralower T.J., Premoli Silva I. & Malone M.J. (eds.), Leg 198 of the cruises of the Drilling Vessel JOIDES Resolution Yokohama, Japan, to Honolulu, Hawaii. Sites 1207-1214. 27 August-23 October 2001.- Proceedings of the Ocean Drilling Program, Scientific Results, Washington - DC, vol. 198, 82 p. DOI: 10.2973/odp.proc.sr.198.103.2005
Bown P.R. & Cooper M.K.E. (1989).- New calcareous nannofossils from the Jurassic.- Journal of Micropalaeontology, London, vol. 8, no. 1, p. 91-96.
Bown P.R. & Cooper M.K.E. (1998).- 4. Jurassic. In: Bown P.R. (ed.), Calcareous nannofossil biostratigraphy.- Kluwer Academic Publishers, Dordrecht, p. 34-85. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Jurassic/Bown%20&%20 Cooper%201998%20BlackBook%20Ch4%20Jurassic%20[%C2%A7N1317].pdf
Bown P.R. & Young J.R. (1997).- Mesozoic calcareous nannoplankton classification.- Journal of Nannoplankton Research, Seward - NE, vol. 19, no. 1, p. 15-47. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/taxonomy%20synthe ses/Bown%20&%20Young%201997%20JNR%2019-1%20Coccoclassif%20-%20Mesozoic%20[%C2%A7N1053].pdf
Bralower T.J. & Bergen J.A. (1998).- Cenomanian-Santonian calcareous nannofossil biostratigraphy of a transect of cores drilled across the Western Interior Seaway. In: Dean W.E. & Arthur M.A. (eds.), Stratigraphy and paleoenvironments of the Cretaceous Western Interior Seaway, USA.- SEPM Concepts in Sedimentology and Paleontology, Tulsa - OK, vol. 6, p. 59-77.
Bralower T.J., Leckie R.M., Slite W.V. & Thierstein H.R. (1995).- An integrated Cretaceous microfossil biostratigraphy. In: Berggren W.A., Kent D.V.J., Aubry M.-P. & Hardenbol J. (eds.), Geochronology, time scales and global stratigraphic correlation.- SEPM Special Publication, Tulsa - OK, vol. 54, p. 65-79.
Bralower T.J., Monechi S. & Thierstein H.R. (1989).- Calcareous nannofossil zonation of the Jurassic-Cretaceous boundary interval and correlation with the Geomagnetic Polarity Timescale.- Marine Micropaleontology, vol. 14, nos. 1-3, p. 153-235.
Bralower T.J., Sliter W.V., Arthur M.A., Leckie R.M., Allard D. & Schlanger S.O. (1993).- Dysoxic/anoxic episodes in the Aptian-Albian (Early Cretaceous). In: Pringle M.S., Sager W.W., Sliter W.V. & Stein S. (eds.), The Mesozoic Pacific: Geology, tectonics, and volcanism.- Geophysical Monograph, Washington - DC, vol. 77, p. 5-37.
Bramlette M.N. & Sullivan F.R. (1961).- Coccolithophorids and related nannoplankton of the early Tertiary in California.- Micropaleontology, New York - NY, vol. 7, no. 2, p. 291-322.
Bramlette M.N. & Martini E. (1964).- The great change in Calcareous Nannoplankton Fossils between the Maastrichtian and the Danian.- Micropaleontology, New York - NY, vol. 10, no. 3, p. 291-322.
Brönnimann P. (1955).- Microfossils incertae sedis from the Upper Jurassic and Lower Cretaceous of Cuba.- Micropaleontology, New York - NY, vol. 1, no. 1, p. 28-51.
Bukry D. (1969).- Upper Cretaceous coccoliths from Texas and Europe.- The University of Kansas Paleontological Contributions, Lawrence - KS, Article 51 (Protista 2), 79 p. URL: http://hdl.handle.net/1808/3826
Burnett J.A. (1997).- New species and new combinations of Cretaceous nannofossils, and a note on the origin of Petrarhabdus (Deflandre) Wind & Wise.- Journal of Nannoplankton Research, Seward - NE, vol. 19, no. 2, p. 133-146. URL: https:// www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Late%20Cretaceous/Burnett%201997%20JNR%2019-2%20New%20Cr et%20spp%20and%20Petrarhabdus%20[%C2%A7N821].pdf
Burnett J.A. (1998).- Upper Cretaceous. In: Bown P.R. (ed.), Calcareous nannofossil biostratigraphy.- The Micropalaeontological Society Special Publications, Kluwer Academic Publishers, Dordrecht, p. 132-199. URL: https://www.mikrotax.org/PDFs/Nannotax 3/used/nannofossils/Late%20Cretaceous/Burnett%201998%20BlackBook%20Ch6%20Upper%20Cretaceous%20[%C2%A7N8 18].pdf
Busson G. & Noël D. (1991).- Les nannoconidés, indicateurs environnementaux des océans et mers épicontinentales du Jurassique terminal et du Crétacé inférieur.- Oceanologica Acta, vol. 14, no. 4, p. 333-355. URL: https://archimer.ifremer.fr/doc/ 00101/21263/
Caratini C. (1963).- Contribution ŕ l'etude des coccolithes du Cénomanien supérieur et du Turonien de la région de Rouen.- Thčse de 3e Cycle, Publication du Laboratoire de Géologie Appliquée, Faculté des Sciences, Université d'Alger, no. 12, 61 p. (5 Pls.). URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Late%20Cretaceous/Caratini%201963%20Cenom %20France%20[%C2%A7N1819].pdf
Covington J.M. & Wise S.W. Jr (1987).- 16. Calcareous Nannofossil biostratigraphy of a Lower Cretaceous deep-sea fan complex: Deep Sea Drilling Project Leg 93 Site 603, lower continental rise off Cape Hatteras. In: Blakeslee J.H. & Whalen E. (eds.), Leg 93 of the cruises of the Drilling Vessel Glomar Challenger. Norfolk, Virginia, to Norfolk, Virginia. May-June, 1983.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XCIII, p. 617-660. DOI: 10.2973/dsdp.proc.93.116.1987
Crux J.A. (1989).- Biostratigraphy and palaeogeographical applications of Lower Cretaceous nannofossils from north-western Europe. In: Crux J.A. & Heck S.E. van (eds.), Nannofossils and their applications. Proceedings of the 2nd INA Conference, London 1987.- The Micropalaeontological Society Special Publications, Ellis Horwood Ltd, Chichester, p. 143-211. URL: https:// www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Early%20Cretaceous/Crux%201989%20BMSPubl%20ECret%20[% C2%A7N130].pdf
Deflandre G. (1939).- Les Stéphanolithes, représentants d'un genre nouveau de coccolithes du Jurassique supérieur.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 208, no. 17, p. 1331-1333. URL: https:// www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Jurassic/Deflandre%201939%20CRAcadSci%20Stephanoliths%20[%C2% A7N122].pdf
Deflandre G. (1947).- Braarudosphaera, nov. gen., type d'une famille nouvelle de Coccolithophoridés actuels ŕ éléments composites.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 225, p. 439-441. URL: https:// www.mikrotax.org/PDFs/Nannotax3/used/taxonomy-extant/Deflandre%201947%20CRAcadSciParis%20Braarudosphaera%20[% C2%A7N1444].pdf
Deflandre G. (1952).- Classe de Coccolithophoridés. In: Piveteau J. (éd.), Traité de paléontologie, vol. 1.- Masson & Cie, Paris, p. 107-115.
Deflandre G. (1953).- Hétérogénéité intrinsčque et pluralité des éléments dans les coccolithes actuels et fossiles.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 237, no. 3, p. 1785-1787. URL: https://www.mikrotax.org /PDFs/Nannotax3/used/taxonomy-extant/Deflandre%201953%20CRAcadSci%20Extant%20in%20TEM%20[%C2%A7N178].pdf
Deflandre G. (1959).- Sur les nannofossiles calcaires et leur systématique.- Revue de Micropaléontologie, Paris, vol. 2, no. 3, p. 127-152.
Deflandre G. (1963).- Sur les Microrhabdulidés, famille nouvelle de nannofossiles calcaires.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 256, no. 16, p. 3484-3487. URL: https://www.mikrotax.org/PDFs/Nannotax3/ used/nannofossils/Late%20Cretaceous/Deflandre%201963%20CRAcadSci%20%20Microrhabdulus%20[%C2%A7N328].pdf
Deflandre G. & Deflandre-Rigaud M. (1962).- Remarques sur l'évolution des Nannoconidés ŕ propos de quelques nouveaux types du Crétacé inférieur de Haute-Provence.- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, t. 255, no. 20, p. 2638-2640. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Early%20Cretaceous/Deflandre %20&%20Deflandre-Rigaud%201962%20CRAcadSci%20E%20Cret%20nannoconids%20[%C2%A7N1229].pdf
Deflandre G. & Deflandre-Rigaud M. (1967).- Diagnoses de quelques nouveaux Nannoconidés gargasiens de Haute-Provence.- Bulletin du Muséum National d'Histoire Naturelle (2e Série), Paris, t. 39, no. 4, p. 774-778. URL: https://www.mi krotax.org/PDFs/Nannotax3/used/nannofossils/Jurassic/Deflandre%20&%20Deflandre-Rigaud%201967%20BullMusNatHistNat %20Nannoconus%20descriptions%20[%C2%A7N1953].pdf
Deflandre G. & Fert C. (1954).- Observations sur les Coccolithophoridés actuels et fossiles en microscopie ordinaire et électronique.- Annales de Paléontologie, Paris, vol. 40, p. 115-176.
Deres F. & Achéritéguy J. (1980).- Biostratigraphie des Nannoconidés.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 4, no. 1, p. 1-53. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofos sils/Early%20Cretaceous/Deres%20&%20Ach%C3%A9rit%C3%A9guy%201980%20BullElfAq%20Nannoconids%20[%C2 %A7N34].pdf
Doevan P.H. (1983).- Cretaceous nannofossil stratigraphy and paleoecology of the Canadian Atlantic Margin.- Bulletin of the Geological Survey of Canada, Ottawa, no. 356, p. 1-70. DOI: https://doi.org/10.4095/109267
Dutour Y. (2005).- Biostratigraphie, évolution et renouvellements des ammonites de l'Aptien supérieur (Gargasien) du bassin Vocontien (Sud-Est de la France).- Thčse de Doctorat de l'Université Claude Bernard-Lyon 1, Villeurbanne, 302 p. URL: https:// www.pedagogie.ac-aix-marseille.fr/upload/docs/application/pdf/2011-09/these_yves_dutour.pdf
Erba E. (1994).- Nannofossils and superplumes: The early Aptian "nannoconids crisis".- Paleoceanography, vol. 9, no. 3, p. 483-501.
Erba E. (1995).- The Aptian stage. In: Rawson P.F., Dhont A.V., Hanccock J.M. & Kennedy W.J. (eds.), Proceedings Second International Symposium on Cretaceous Stage Boundaries.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique (Sciences de la terre), Bruxelles, vol. 66 - supplement, p. 31-43.
Gale A.S., Kennedy W.J., Burnett J.A., Caron M. & Kidd B.E. (1996).- The late Albian to early Cenomanian succession at Mont Risou near Rosans (Drôme, SE France): An integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes).- Cretaceous Research, vol. 17, no. 5, p. 515-606.
Gardet M. (1955).- Contribution ŕ l'étude des coccolithes des terrains néogčnes de l'Algérie.- Publications du Service de la Carte Géologique de l'Algérie (Nouvelle Série), Bulletin (Travaux des Collaborateurs), Alger, no. 5, p. 477-550.
Gartner S. (1968).- Coccoliths and related calcareous nannofossils from Upper Cretaceous deposits of Texas and Arkansas.- The University of Kansas Paleontological Contributions, Lawrence - KS, Article 48 (Protista 1), 56 p. URL: http://hdl.handle.net/ 1808/3822
Giraud F., Pittet B., Grosheny D., Baudin F., Lécuyer C. & Sakamoto T. (2018).- The paleoceanographic crisis of the early Aptian (OAE 1a) in the Vocontian Basin (SE France). Palćogeography, Palćoclimatology, Palćoecology, vol. 511, p. 483-505.
Górka H. (1957).- Les Coccolithophoridés du Maestrichtien supérieur de Pologne.- Acta Palaeontologica Polonica, Warszawa, vol. II, nos. 3-4, p. 235-284. URL: https://www.app.pan.pl/archive/published/app02/app02-235.pdf
Gran H.H. & Braarud T. (1935).- A quantitative study of the phytoplankton in the Bay of Fundy and the Gulf of Maine (including observations on hydrography, chemistry and turbidity).- Journal of the Biological Board of Canada, Ottawa, vol. 1, no. 5, p. 279-467.
Grün W. & Allemann F. (1975).- The Lower Cretaceous of Caravaca (Spain): Berriasian calcareous nannoplankton of the Miravetes Section (Subbetic Zone, Prov. of Murcia).- Eclogae Geologicae Helvetiae, Basel, vol. 68, no. 1, p. 147-211. URL: https:// www.e-periodica.ch/digbib/view?pid=egh-001%3A1975%3A68%3A%3A6#183
Grün W., Prins B. & Zweili F. (1974).- Coccolithophoriden aus dem Lias epsilon von Holzmaden (Deutschland).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Band 147, Heft 2, p. 294-328.
Grün W. & Zweili F. (1980).- Das kalkige Nannoplankton der Dogger-Malm-Grenze im Berner Jura bei Liesberg (Schweiz). Jahrbuch - Geologische Bundesanstalt, Wien, Band 123, Heft 2, p. 231-341. URL: http://opac.geologie.ac.at/wwwopacx/www opac.ashx?command=getcontent&server=images&value=JB1231_231_A.pdf
Hay W.W. (1977).- Calcareous nannofossils. In: Ramsay A.T.S (ed.), Oceanic micropalaeontology.- Academic Press, London, p. 1055-1120.
Hay W.W. & Molher M.P. (1967).- Calcareous nannoplankton from Early Tertiary rocks at Pont Labau, France, and Paleocene-Early Eocene correlations.- Journal of Paleontology, McLean - VA, vol. 41, no. 2, p. 1505-1541.
Herrle J.O. & Mutterlose J. (2003).- Calcareous nannofossils from the Aptian-Lower Albian of southeast France: Palaeoecological and biostratigraphic implications.- Cretaceous Research, vol. 24, no. 1, p. 1-22.
Herrle J.O., Kossler P. & Bollman J. (2010).- Palaeoceanographic differences of early late Aptian black shale events in the Vocontian Basin (SE France).- Palćogeography, Palćoclimatology, Palćoecology, vol. 297, no. 2, p. 367-376.
Hill M.E. (1976).- Lower Cretaceous calcareous nannofossils from Texas and Oklahoma.- Paleontographica, Abteilung B Palaeophytologie, Stuttgart, Band 156, Lieferung 4-6, p. 103-179.
Jakubowski M. (1986).- New calcareous nannofossil taxa from the Lower Cretaceous of the North Sea.- INA Newsletter, Seward - NE, vol. 8, no. 1, p. 38-42. URL: http://ina.tmsoc.org/JNR/NINA/INANewsl8(1).pdf
Jeremiah J. (1996).- A proposed Albian To Lower Cenomanian nannofossil biozonation for England and the North Sea Basin.- Journal of Micropalaeontology, London, vol. 15, no. 2, p. 97- 129.
Kaenel E. de & Bergen J.A. (1993).- New Early and Middle Jurassic coccolith taxa and biostratigraphy from the eastern proto-Atlantic (Morocco, Portugal and DSDP Site 547B).- Eclogae Geologicae Helvetiae, Basel, vol. 86, no. 3, p. 861-907. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001%3A1993%3A86%3A%3A668#896
Kaenel E. de & Bergen J. A. (1996).- 2. Mesozoic calcareous nannofossil biostratigraphy from sites 897, 899, and 901, Iberia Abyssal Plain: New biostratigraphic evidence. In: Riegel R.N. (ed.), Covering Leg 149 of the cruises of the Drilling Vessel JOIDES Resolution. Balboa Harbor, Panama, to Lisbon, Portugal. Sites 897-901. 10 March-25 May 1993.- Proceedings of the Ocean Drilling Program, Scientific Results, Washington - DC, vol. 149, p. 27-59. DOI: 10.2973/odp.proc.sr.149.206.1996
Kamptner E. (1931).- Nannoconus steinmanni nov. gen., nov. spec., ein merkwürdiges gesteinsbildendes Mikrofossil aus dem jüngeren Mesozoikum der Alpen.- Paläontologische Zeitschrift, Offenbach am Main, vol. 13, no. 4, p. 288-298.
Kamptner E. (1937).- Über Dauersporen bei marinen Coccolithinen.- Sitzungsberichte der Akademie der Wissenschaften in Wien, Mathematisch-Naturwissenschaftliche Klasse, Abt. 1, Band 146, p. 67-76. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/ others/calcareous%20dinos/Kamptner%201937%20SBAWW%20Dauersporen%20[%C2%A7N3487].pdf
Kamptner E. (1950).- Über den submikroskopischen Aufbau der Coccolithen.- Anzeiger der Österreichische Akademie der Wissenschaften, Mathematische-Naturwissenschafliche Klasse, Wien, Band 87, p. 152-158.
Kennedy W.J., Gale A.S., Bown P.R., Caron M., Davey R.J., Groke D. & Wray D.S. (2000).- Integrated stratigraphy across the Aptian-Albian boundary in the Marnes Bleues, at the Col de Pré-Guittard, Arnayon (Drôme), and at Tartonne (Alpes-de-Haute-provence), France: A candidate Global Boundary Stratotype Section and Boundary Point for the base of the Albian Stage.- Cretaceous Research, vol. 21, no. 5, p. 591-720.
Lambert B. (1981).- Étude systématique des nannofossiles calcaires du Crétacé supérieur stratotypique (Coniacien, Santonien, Campanien). Répartition stratigraphique et biozonation.- Thčse de Doctorat de 3e Cycle, Université P. et M. Curie, Paris, 291 p.
Lambert B. (1986).- La notion d'espčce chez le genre Braarudosphaera Deflandre, 1947. Mythe et réalité.- Revue de Micropaléontologie, Paris, vol. 28, no. 4, p. 255-264.
Lambert B. (1987).- Nannofossiles calcaires de l'Albien supérieur et du Vraconnien du Cameroun méridional.- Cahiers de Micropaléontologie (N.S.), Paris, vol. 2, no. 2, p. 33-96.
Lambert B. (1993).- Nannofossiles calcaires de l'Albien supérieur et du Vraconnien du Cameroun méridional (deuxičme partie).- Cahiers de Micropaléontologie (N.S.), Paris, vol. 8, no. 2, p. 183-268.
Lambert B. & Laporte-Galaa C. (1996).- Nannoflore calcaire du puits Mango-1 (offshore, nord Sinaď). In: Jardiné S., Klasz I. de & Debenay J.-P. (éds.), Géologie de l'Afrique et de l'Atlantique Sud. 12e Colloque de micropaléontologie africaine. 2e Colloque de stratigraphie et paléogéographie de l'Atlantique sud. Angers (France), 16-20 juillet 1994.- elf aquitaine édition, Pau, p. 631-645.
Lambert B. & Laporte-Galaa C. (2005).- Discoaster zonation of the Miocene of the Kutei Basin, East Kalimantan, Indonesia (Mahakam Delta offshore).- Carnets Geol., Madrid, vol. 5, no. M01 (CG 2005_M01), p. 1-63. DOI: https://doi.org/10.4267/2042/ 1461
Lees J.A. (2007).- New and rarely reported calcareous nannofossils from the Late Cretaceous of coastal Tanzania: Outcrop samples and Tanzania Drilling Project Sites 5, 9 and 15.- Journal of Nannoplankton Research, Seward - NE, vol. 29, no. 1, p. 39-65. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Late%20Cretaceous/Lees%202007%20JNR%2029- 1%20new%20spp%20TDP%20CRet%20[%C2%A7N1398].pdf
Lees J.A., Bown P.R., Young J.R. & Riding J.B. (2004).- Evidence for annual records of phytoplankton productivity in the Kimmeridge Clay Formation coccolith stone bands (Upper Jurassic, Dorset, UK).- Marine Micropaleontology, vol. 52, nos. 1-4, p. 29-49.
Lees J.A. & Bown P.R. (2016).- New and intriguing calcareous nannofossils from the Turonian (Upper Cretaceous) of Tanzania.- Journal of Nannoplankton Research, Seward - NE, vol. 36, no. 1, p. 83-95. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/ nannofossils/Late%20Cretaceous/Lees%20&%20Bown%202016%20JNR%2036-1%20New%20Tanzania%20Cret%20spp%20[ %C2%A7N1727].pdf
Loeblich A.R. Jr & Tappan H. (1963).- Type fixation and validation of certain calcareous nannoplankton genera.- Proceedings of the Biological Society of Washington, Washington - DC, vol. 76, p. 191-196. URL: https://www.biodiversitylibrary.org/item/107599 #page/223/mode/1up
Lohman H. (1920).- Die Bevölkerung des Ozeans mit Plankton nach den Ergebnissen der Zentrifugenfänge während der Ausreise der Deutschland 1911, zugleich ein Beitrag zur Biologie des Atlantischen Ozeans.- Archiv für Biontologie, Berlin, Band 4, Heft 3, 617 p.
Manivit H. (1965).- Nannofossiles calcaires de l'Albo-Aptien.- Revue de Micropaléontologie, Paris, vol. 8, no. 3, p. 189-201.
Manivit H. (1971).- Les nannofossiles calcaires du Crétacé français : De l'Aptien au Danien, essai de biozonation appuyée sur les stratotypes.- Thčse d'État (Sciences naturelles), Université Paris XI-Orsay, no. 704, 167 p.
Manivit H., Perch-Nielsen K., Prins B. & Verbeek J.W. (1977).- Mid Cretaceous calcareous nannofossil biostratigraphy.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (Series B), Amsterdam, vol. 80, no. 3, p. 169-181.
Manton I. & Oates K. (1985).- Calciosoleniaceae (Coccolithophorids) from the Galapagos Islands: Unmineralized components and coccolith morphology in Anoplosolenia and Calciosolenia with a comparative analysis of equivalents in the unmineralized genus Navisolenia (Haptophyceae = Prymnesiophyceae).- Philosophical Transactions of the Royal Society B (Biological Sciences), London, vol. 309, p. 461-477.
Moshkovitz S. (1972).- Biostratigraphy of the genus Nannoconus in the Lower Cretaceous sediments of the subsurface: Ashqelon-Helez area, central Israel.- Israel Journal of Earth-Sciences, Jerusalem, vol. 21, p. 1-28.
Moullade M., Masse J.-P., Tronchetti G., Kuhnt W., Ropolo P., Bergen J.A., Masure E. & Renard M. (1998).- Le stratotype historique de l'Aptien inférieur (région de Cassis-La Bédoule, SE France) : Synthčse stratigraphique. In: Moullade M., Tronchetti G. & Masse J.-P. (éds.), Le stratotype historique de l'Aptien inférieur (Bédoulien) de Cassis-La Bédoule (S.E. France).- Géologie méditerranéenne, Marseille, vol. 25, no. 3-4, p. 289-298. URL: https://www.persee.fr/doc/geolm_0397-2844 _1998_num_25_3_1641
Moullade M., Tronchetti G., Balme C. & Kouyoumontzakis G. (2006).- Le Gargasien (Aptien moyen) de La Tuiličre - St-Saturnin-lčs-Apt (région du stratotype historique de l'Aptien, Vaucluse, SE France) : Localisation géographique et corrélations des affleurements.- Carnets Geol., Madrid, vol. 6, no. L01 (CG 2006_L01), p. 1-8. DOI: https://doi.org/10.4267/ 2042/4567
Moullade M., Tronchetti G. & Bellier J.-P. (2008).- Associations et biostratigraphie des foraminifčres benthiques et planctoniques du Bédoulien sommital et du Gargasien inférieur de La Tuiličre - St-Saturnin-lčs-Apt (aire stratotypique de l'Aptien, Vaucluse, SE France).- Carnets Geol., Madrid, vol. 8, no. A01 (CG 2008_A01), p. 1-50. DOI: https://doi.org/10.4267/ 2042/14699
Moullade M., Tronchetti G., Balme C., Dutour Y., Ropolo P., Price G., Martinez M. & Granier B. (2017).- Proposition pour l'établissement du GSSP de l'Aptien supérieur dans le Bassin d'Apt (Vaucluse, SE France) : Synthčse des données stratigraphiques.- Carnets Geol., Madrid, vol. 17, no. 9, p. 161-189. DOI: https://doi.org/10.4267/2042/62954
Mutterlose J. (1989).- Temperature-controlled migration of calcareous nannofloras in the north-west European Aptian. In: Crux J.A. & Heck S.E. van (eds.), Nannofossils and their applications. Proceedings of the 2nd INA Conference, London 1987.- The Micropalaeontological Society Special Publications, Ellis Horwood Ltd, Chichester, p. 122-144.
Mutterlose J. (1991).- Verteilungs- und Migrationsmuster des kalkigen Nannoplanktons in der Unterkreide (Valangin-Apt) NW-Deutschlands.- Palaeontographica Abteilung B, Palaeophytologie, Stuttgart, Band 221, p. 27-152.
Mutterlose J. & Wise S.W. Jr (1990).- 23. Lower Cretaceous nannofossil biostratigraphy of ODP Leg 113 Holes 692B and 693A, continental slope off East Antartica, Wedell Sea. In: Kennett D., Masterson A. & Stewart N.J. (Eds.), Leg 113 of the cruises of the Drilling Vessel JOIDES Resolution, Valparaiso, Chile, to East Cove, Falkland Islands, Sites 689-697, 25 December 1986-11 March 1987.- Proceedings of the Ocean Drilling Program, Scientific Results, Washington - DC, vol. 113, p. 325-351. DOI: 10.2973/odp.proc.sr.113.143.1990
Noël D. (1956).- Coccolithes des terrains jurassiques de l'Algérie.- Publications du Service de la Carte Géologique de l'Algérie (Nouvelle Série), Bulletin (Travaux des Collaborateurs), Alger, no. 8, p. 303-385.
Noël D. (1958).- Étude de coccolithes du Jurassique et du Crétacé inférieur.- Publications du Service de la Carte Géologique de l'Algérie (Nouvelle Série), Bulletin (Travaux des Collaborateurs), Alger, no. 8, p. 20, p. 155-196.
Noël D. (1965).- Sur les coccolithes du Jurassique européen et d'Afrique du Nord : Essai de classification des coccolithes fossiles.- Éditions du Centre National de la Recherche Scientifique, Paris, 209 p.
Noël D. (1969).- Arkhangelskiella (coccolithe crétacé) et formes affines du bassin de Paris.- Revue de Micropaléontologie, Paris, vol. 11, no. 4, p. 191-204.
Noël D. (1970).- Coccolithes crétacés. La craie campanienne du Bassin de Paris.- Éditions du Centre National de la Recherche Scientifique, Paris, 129 p.
Noël D. (1972).- Nannofossiles calcaires des sédiments jurassiques finement laminés.- Bulletin du Muséum National d'Histoire Naturelle (3e série), Paris, Sciences de la Terre 14, no. 75, p. 95-156. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/ nannofossils/Jurassic/Noel%201973%20BullMusNatHistNat%20lam%20Jurassic%20[%C2%A7N554].pdf
Perch-Nielsen K. (1968).- Der Feinbau und die Klassification der Coccolithen aus dem Maastrichtien von Dänemark.- Biologiske Skrifter, Kongelige Danske Videnskabernes Selskab, Křbhavn, Bind 16, no. 1, 96 p. (XXXII Pls.). URL: http://www.royalacademy.dk /Publications/Low/319_Perch-Nielsen, Katharina.pdf
Perch-Nielsen K. (1973).- Neue Coccolithen aus dem Maastrichtien von Dänemark, Madagaskar und Ägypten.- Bulletin of the Geological Society of Denmark, Křbenhavn, vol. 22, p. 306-333 (10 Pls.). URL: https://2dgf.dk/xpdf/bull22-04-306-333.pdf
Perch-Nielsen K. (1979).- Calcareous nannofossils from the Cretaceous between the North Sea and the Mediterranean. In: Weidmann J. (ed.), Aspekte der Kreide Europas.- International Union of Geological Sciences Series A, Stuttgart, no. 6, p. 223-272. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Late%20Cretaceous/Perch-Nielsen%201979%20Aspekte derKEuropas%20LateK%20[%C2%A7N1084].pdf
Perch-Nielsen K. (1984).- Validation of new combinations.- INA Newsletter, Seward - NE, vol. 6, no. 1, p. 42-46.
Perch-Nielsen K. (1985).- Mesozoic calcareous nannofossils. In: Bolli H.M., Saunders J.B. & Perch-Nielsen K. (eds.), Plankton stratigraphy.- Cambridge University Press, p. 329-496.
Perch-Nielsen K. (1986).- New Mesozoic and Paleogene calcareous nannofossils.- Eclogae Geologicae Helvetiae, Basel, vol. 79, no. 3, p. 835-847. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001%3A1986%3A79%3A%3A845#855
Reinhardt P. (1964).- Einige Kalkflagellaten-Gattungen (Coccolithophoriden, Coccolithineen) aus dem Mesozoikum Deutschlands.- Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin, Band 6, p. 749-759. URL: https://www.mi krotax.org/PDFs/Nannotax3/used/nannofossils/Late%20Cretaceous/Reinhardt%201964%20MonatDAkad%20%20U%20Cret %20n%20taxa%20[%C2%A7N514].pdf
Reinhardt P. (1965).- Neue Familien für fossile Kalkflagellaten (Coccolithophoriden, Coccolithineen).- Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin, Band 7, p. 30-40. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nan nofossils/Late%20Cretaceous/Reinhardt%201964%20MonatDAkad%20%20U%20Cret%20n%20taxa%20[%C2%A7N5 14].pdf
Reinhardt P. (1966).- Zur Taxionomie und Biostratigraphie der fossilen Nannoplanktons aus dem Malm, der Kreide und des Alttertiär Mitteleuropas.- Freiberger Forschungshefte (Reihe C), Band 196, p. 5-109.
Reinhardt P. (1967).- Fossile Coccolithen mit rhagoidem Zentralfield (Fam. Ahmuellerellaceae, Subord. Coccolithineae).- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, Jahrgang 1967, Heft 3, p. 163-178.
Reinhardt P. (1971).- Synopsis der Gattungen und Arten der Mesozoischen Coccolithen und anderes kalkiger Nannofossilien, Teil III.- Freiberger Forschungshefte (Reihe C), Band 267, p. 19-41.
Reinhardt P. & Górka, H. (1967).- Revision of some Upper Cretaceous coccoliths from Poland and Germany.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Band 129, Heft 3, p. 240-256.
Rood A.P., Hay W.W. & Barnard T. (1971).- Electron microscope studies of Oxford Clay coccoliths.- Eclogae Geologicae Helvetiae, Basel, vol. 64, no. 2, p. 245-272. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001%3A1971%3A64%3A%3A 292#323
Rood A.P., Hay W.W. & Barnard T. (1973).- Electron microscope studies of Lower and Middle Jurassic coccoliths.- Eclogae Geologicae Helvetiae, Basel, vol. 66, no. 2, p. 365-382. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001%3A1973%3A 66%3A%3A277#403
Roth P.H. (1973).- 23. Calcareous nannofossils-Leg 17, Deep Sea Drilling Project. In: Roth P.H. & Herring J.R. (eds.), Leg 17 of the cruises of the Drilling Vessel Glomar Challenger. Honolulu, Hawaii to Honolulu, Hawaii. April-May 1971.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XVII, p. 695-793. DOI: 10.2973/dsdp.proc.17.123.1973
Roth P.H. (1978).- 34. Calcareous nannoplankton biostratigraphy and oceanography of the northwestern Atlantic Ocean. In: Worstell P. (ed.), Leg 44 of the cruises of the Drilling Vessel Glomar Challenger. Norfolk, Virginia to Norfolk, Virginia. August-September 1975.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XLIV, p. 731-759. DOI: 10.2973/ dsdp.proc.44.134.1978
Roth P.H. & Thierstein H. (1972).- 14. Calcareous Nannoplankton: Leg XIV of the Deep Sea Drilling Project. In: Pimm A.C. (ed.), Leg 14 of the cruises of the Drilling Vessel Glomar Challenger. Lisbon, Portugal to San Juan, Puerto Rico. October-December 1970.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XIV, p. 421-485. DOI: 10.2973/dsdp.proc.14.114.1972
Rutledge D. & Bown P.R. (1996).- New names for old: Taxonomic clarification of some Early Cretaceous nannofossil marker species.- Journal of Nannoplankton Research, Seward - NE, vol. 18, no. 2, p. 53-59. URL: https://www.mikrotax.org/PDFs/Nanno tax3/used/nannofossils/Early%20Cretaceous/Rutledge%20&%20Bown%201996%20JNR%2018-2%20E%20cret%20taxa%20[ %C2%A7N443].pdf
Sissingh W. (1977).- Biostratigraphy of Cretaceous calcareous nannoplankton.- Geologie en Mijnbouw, Utrecht, vol. 56, no. 1, p. 37-65. URL: https://drive.google.com/open?id=0B7j8bPm9Cse0cjBMMUdkc0t6MEU
Stover L. (1966).- Cretaceous coccoliths and associated nannofossils from France and Netherlands.- Micropaleontology, New York - NY, vol. 12, no. 2, p. 133-167.
Stradner H. (1961).- Vorkommen von Nannofossilien in Mesozoikum and Alttertiäar.- Erdoel Zeitschrift für Bohr- und Fördertechnik, Gewinnung, Aufbereitung, Transport, Wien, vol. 77, no. 3, p. 77-88.
Stradner H. (1962).- Über neue und wenig bekannte Nannofossilien aus Kreide und Alttertiär.- Verhandlungen der Geologischen Bundesanstalt, Wien, Heft 1962, p. 363-377. URL: http://opac.geologie.ac.at/wwwopacx/wwwopac.ashx?command =getcontent&server=images&value=VH1962_363_A.pdf
Stradner H. (1963).- New contributions to Mesozoic stratigraphy by means of nannofossils.- Proceedings of the Sixth World Petroleum Congress, Frankfurt am Main, Germany, June 1963, Section, Paper 4, p. 167-183.
Street C. & Bown P.R. (2000).- Palaeobiogeography of Early Cretaceous (Berriasian-Barremian) calcareous nannoplankton.- Marine Micropaleontology, vol. 39, nos. 1-4, p. 265-291.
Thierstein H.R. (1971).- Tentative Lower Cretaceous nannoplankton zonation.- Eclogae Geologicae Helvetiae, Basel, vol. 64, no. 3, p. 459-488. URL: https://www.e-periodica.ch/digbib/view?pid=egh-001%3A1971%3A64%3A%3A529#556
Thierstein H.R. (1973).- Lower Cretaceous calcareous nannoplankton biostratigraphy.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 29, p. 1-52. URL: https://www.zobodat.at/pdf/AbhGeolBA_29_0001-0052.pdf
Trejo M. (1959).- Dos nuevas especies del género Nannoconus (Protozoa, inc. saed.).- Cienca (Méx.), Mexico - D.F., vol. 19, nos. 6-7, p. 130-132.
Tremolada F. & Erba E. (2002).- Morphometric analyses of Aptian Assipetra infracretacea and Rucinolithus terebrodentarius nannoliths: Implications for taxonomy, biostratigraphy and paleoceanography._ Marine Micropaleontology, vol. 44, nos. 1-2, p. 77-92.
Tremolada F., Erba E. & Bralower T.J. (2006).- Late Barremian to early Aptian calcareous nannofossil paleoceanography and paleoecology from the Ocean Drilling Program Hole 641C (Galicia Margin).- Cretaceous Research, vol. 27, no. 6, p. 887-897.
Varol O. & Girgis M.H (1994).- New taxa and taxonomy of some Jurassic to Cretaceous calcareous nannofossils.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Band 192, Heft 2, p. 221-253.
Vekshina V.N. (1959).- Coccolithophoridae in Maastrichtian deposits of the West Siberian lowlands.- Trudyi Instituta Geologii i Geogiziki, Sibiriskoe Otlodelenie, Akademiya Nauk SSSR (Nauka), Moscow, Syr'ya no. 2, p. 56-77 [in Russian].
Verbeek J.W. (1977).- Calcareous nannoplankton biostratigraphy of Middle and Upper Cretaceous deposits in Tunisia.- Utrecht Micropaleontological Bulletins, Bull. 16, 157 p. URL: https://www.mikrotax.org/PDFs/Nannotax3/used/nannofossils/Late% 20Cretaceous/Verbeek%201977%20UtrechtMpalBull%20[%C2%A7N14].pdf
Watkins D.K. & Raffi I. (2020).- Subchapter 3F. Calcareous nannofossils. In: Gradstein F.M. (ed.), Chapter 3. Evolution and biostratigraphy. In: Gradstein F.M., Ogg J.G., Schmitz M.D. & Ogg G.M. (eds.), Geologic Time Scale 2020, Volume 1.- Elsevier, p. 69-73.
Wind F.H. & Čepek P. (1979).- 3. Lower Cretaceous calcareous nannoplankton from DSDP Hole 397A (northwest African Margin). In: Laughter F.H. & Fagerberg E.M. (eds.), Leg 47, Part 1 of the cruises of the Drilling Vessel Glomar Challenger. Las Palmas, Canary Islands to Vigo, Spain. March-April 1976.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XLVII, part 1, p. 221-256. DOI: 10.2973/dsdp.proc.47-1.103.1979
Wind F.H. & Wise S.W. Jr (1983).- 22. Correlation of upper Campanian-lower Maastrichtian calcareous nannofossil assemblages in drill and piston cores from the Falkland Plateau, southwest Atlantic Ocean. In: Wise S.W. Jr, Blakeslee J.H. & Lee M. (eds.), Leg 71 of the cruises of the Drilling Vessel Glomar Challenger. Valparaiso, Chile, to Santos, Brazil. January-February, 1980.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXI, p. 551-563. DOI: 10.2973/dsdp.proc.71.122. 1983
Wise S.W. Jr (1983).- 21. Mesozoic and Cenozoic calcareous nannofossils recovered by DSDP Leg 71 in the Falkland Plateau region, Southwest Atlantic Ocean. In: Wise S.W. Jr, Blakeslee J.H. & Lee M. (eds.), Leg 71 of the cruises of the Drilling Vessel Glomar Challenger. Valparaiso, Chile, to Santos, Brazil. January-February, 1980.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. LXXI, p. 481-550. DOI: 10.2973/dsdp.proc.71.121.1983
Wise S.W. Jr & Wind F.H. (1977).- 8. Mesozoic and Cenozoic calcareous nannofossils recovered by DSDP Leg 36 drilling in the Falkland Plateau, southwest Atlantic sector of the Southern Ocean. In: Wise S.W. Jr (ed.), Leg 36 of the cruises of the Drilling Vessel Glomar Challenger. Ushuaia, Argentina to Rio De Janeiro, Brazil. April-May 1974.- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XXXVI, p. 269-491. DOI: 10.2973/dsdp.proc.36.108.1977
Worsley T. (1971).- Calcareous Nannofossil Zonation of Upper Jurassic and Lower Cretaceous sediments from the Western Atlantic. In: Farinacci A. (ed.), Proceedings of the Second Planktonic Conference Roma 1970.- Edizioni Tecnoscienza, Roma, vol. 2, p. 1301-1321.
Young J.R., Bergen J.A., Bown P.R., Burnett J.A., Fiorentino A., Jordan R.W., Kleijne A., Niel B.E. van, Romein A.J.T. & Salis K. von (1997).- Guidelines for coccolith and calcareous nannofossil terminology.- Palaeontology, Durham, vol. 40, no. 4, p. 875-912. URL: https://www.palass.org/sites/default/files/media/publications/palaeontology/volume_40/vol40_part4_pp875-912.pdf
Young J.R., Bown P.R. & Lees J.A. (2022).- Nannotax3 website.- International Nannoplankton Association. URL: https://www. mikrotax.org/Nannotax3/
Plate 1: Association GIIA (sample 2307): fig. 1: Nannoconus vocontiensis Deres & Achéritéguy, 1980; fig. 2: Nannoconus bucheri Brönnimann, 1955; figs. 3-4: Nannoconus vocontiensis Deres & Achéritéguy, 1980; fig. 5: Nannoconus bucheri Brönnimann, 1955; figs. 6-7: Nannoconus aff. N. vocontiensis Deres & Achéritéguy, 1980; figs. 8-9: Nannoconus aff. globulus Brönnimann, 1955; figs. 10-11: Nannoconus circularis Deres & Achéritéguy, 1980; fig. 12: Nannoconus boletus Deflandre & Deflandre-Rigaud, 1962; fig. 13: ? Nannoconus boletus Deflandre & Deflandre-Rigaud, 1962; figs. 14-15: Nannoconus quadriangulus Deflandre & Deflandre-Rigaud, 1962; fig. 16: Nannoconus truittii Brönnimann, 1955; figs. 17-18: Nannoconus quadriangulus Deflandre & Deflandre-Rigaud, 1962. Association GI (sample 2270): fig. 19: Nannoconus sp. aff. grandis Deres & Achéritéguy, 1980; fig. 20: Nannoconus kamptneri Brönnimann, 1955; figs. 21-24: Nannoconus boneti Trejo, 1959; figs. 25-27: Nannoconus wassallii Brönnimann, 1955; figs. 28-29: Nannoconus bucheri Brönnimann, 1955; fig. 30: Nannoconus aff. globulus Brönnimann, 1955 (L), and Nannoconus vocontiensis Deres & Achéritéguy, 1980 (R); fig. 31: Nannoconus circularis Deres & Achéritéguy, 1980; fig. 32: Nannoconus quadriangulus Deflandre & Deflandre-Rigaud, 1962; fig. 33: Nannoconus truittii Brönnimann, 1955 (bottom). For each Nannonus species we illustrated
several specimens in order to show the natural variation in size and shape. See Figure 2 |
|
Plate 2: Association GIII (sample 2303): figs. 1-2, 7-16: Nannoconus quadriangulus Deflandre & Deflandre-Rigaud, 1962; figs. 3-5: Nannoconus truittii Brönnimann, 1955; fig. 6: Nannoconus boletus Deflandre & Deflandre-Rigaud, 1962. Association GIIB
(sample 2323): figs. 17-22: Nannoconus
vocontiensis Deres & Achéritéguy,
1980; figs. 23-24: Nannoconus
aff. globulus Brönnimann, 1955; fig. 25: Nannoconus
circularis Deres & Achéritéguy,
1980; figs. 26, 28: Nannoconus
globulus Brönnimann, 1955; fig. 27: ? Nannoconus
aff. globulus Brönnimann, 1955; figs. 29-30: Nannoconus
quadriangulus Deflandre & Deflandre-Rigaud,
1962; fig. 31: Nannoconus
quadriangulus Deflandre & Deflandre-Rigaud, 1962 (L), and
N. boletus (R) Deflandre & Deflandre-Rigaud,
1962; fig. 32: ? N.
boletus Deflandre & Deflandre-Rigaud, 1962. |
|
Plate 3:
figs. 1, 3: ? Nannoconus
boletus Deflandre & Deflandre-Rigaud, 1962
(sample 2312); figs. 2, 4-5: Nannoconus
boletus Deflandre & Deflandre-Rigaud, 1962
(sample 2312); fig. 6: Assipetra
infracretacea (Thierstein, 1973) Roth,
1973 (sample
2304); fig. 7: ? Nannoconus
carniolensis Deflandre & Deflandre-Rigaud, 1962
(sample 2304); figs. 8-11: Nannoconus
carniolensis Deflandre & Deflandre-Rigaud, 1962
(sample 2304); fig. 12: Assipetra
terebrodentarius (Applegate et al. in Covington
& Wise, 1987) Rutledge
& Bergen in Bergen, 1994 (sample 2324); fig. 13: Staurolithites
handleyi Lees, 2007 (sample 2327); fig. 14: Staurolithites
mutterlosei Crux, 1989 (sample 2327); figs. 15-16: Staurolithites
rectus Black, 1971a (fig. 15, sample 2332; fig. 16, sample
2310); fig. 17: Bukrylithus
ambiguus Black, 1971a (sample 2302); figs. 18-19, 22: Rhabdophidites
parallelus (Wind & Cepek,
1979) Lambert,
1987 (fig. 18, sample 2332; fig. 19, sample 2292; fig. 22, sample 2310); figs. 20-21: Rhabdophidites
moeslensis Manivit, 1971
(fig. 20, sample 2317; fig. 21,
sample 2310); fig. 23: Acaenolithus ? sp. (sample 2316); figs. 24-25:
aff. Misceomarginatus Wind & Wise
in Wise
& Wind, 1977 (fig. 24, sample
2327; fig. 25, sample 2332); fig. 26: Zeugrhabdotus
xenotus (Stover, 1966) Burnett
in Gale et al.,
1996 (sample 2339). |
|
Plate 4:
figs. 1-3, ? 9: Chiastozygus aff. tenuis Black, 1971a (figs. 1, 9, sample 2339;
figs. 2-3, sample 2310); figs. 4-5: Chiastozygus
platyrhetum Hill, 1976 (fig. 4, sample 2284; fig. 5, sample
2299); figs. 6-7: Tegumentum stradneri Thierstein
in Roth & Thierstein, 1972 (fig. 6, sample 2322;
fig. 7, sample 2284); fig. 8: Corollithion
? madagaskarensis Perch-Nielsen, 1973 (sample 2284); figs. 10-11, 18: Zeugrhabdotus cf. diplogrammus (Deflandre
in Deflandre & Fert, 1954) Burnett in Gale et al.,
1996 (fig. 10, sample 2327; fig. 11, sample 2284;
fig. 18, sample 2327); figs. 12-13: Zeugrhabdotus
embergeri (Noël, 1958) Perch-Nielsen, 1985 (sample
2310); fig. 14: Zeugrhabdotus
moulladei Bergen, 1998 (sample 2322); fig. 15: Zeugrhabdotus
choffatii Rood et al., 1973 (sample 2327); figs. 16, 23-25: Zeugrhabdotus
noeliae Rood et al., 1971 (fig. 16, sample
2339; fig. 23, sample 2327; fig. 24, sample 2310; fig. 25, sample 2304); fig. 17: Zeugrhabdotus
howei Bown in Kennedy
et al.,
2000 (sample 2302); fig. 19: Zeugrhabdotus bicrescenticus (Stover,
1966) Burnett in Gale et al.,
1996 (sample 2322);
fig. 20: Tranolithus
gabalus Stover, 1966 (sample 2284); figs. 21-22: Zeugrhabdotus sp. 1 (fig. 21, sample 2310; fig. 22, sample 2304). |
|
Plate 5:
figs. 1-2: Rhagodiscus
asper (Deflandre, 1953) Verbeek,
1977 (fig. 1, sample
2284; fig. 2, sample 2284); fig. 3: Rhagodiscus
splendens (Deflandre, 1953) Verbeek,
1977 (sample 2332); figs. 4-5: Rhagodiscus
asper (Stradner, 1963) Reinhardt,
1967 (fig. 4,
sample 2324; fig. 5, sample 2299); figs. 6, 9: Rhagodiscus
angustus (Stradner, 1963) Rheinardt,
1971 - (fig. 6,
sample 2295; fig. 9, sample 2295); fig. 7: Rhagodiscus
splendens (Deflandre, 1953) Verbeek,
1977 (sample 2304); fig. 8: Rhagodiscus
asper (Stradner, 1963) Reinhardt,
1967 (sample 2304); figs. 10-11: Rhagodiscus
gallagheri Rutledge & Bown,
1996 (fig. 10,
sample 2299; fig. 11, sample 2299); fig. 12: Cretarhabdus
conicus Bramlette & Martini,
1964, large form
(sample 2304); figs. 13-15: Parhabdolithus imperfossus (Black,
1972) Lambert, 1987 (fig. 13, sample 2299; fig. 14, sample 2310;
fig. 15,
sample 2295); fig. 16: Microstaurus
chiastius (Worsley, 1971) Bralower et
al., 1989 (sample 2339); fig. 17: Retecapsa
schizobrachiata (Gartner, 1968) Grün
in Grün & Allemann, 1975
(sample 2327); figs. 18-20: Cretarhabdus
conicus Bramlette & Martini,
1964 (fig. 18,
sample 2295; fig. 19, sample 2310; fig. 20, sample 2302); fig. 21: Microstaurus
quadratus Black, 1971a (sample 2303); fig. 22: Grantarhabdus
meddii Black, 1971a (sample 2342). |
|
Plate 6:
fig. 1: ? Tetrapodorhabdus decorus (Deflandre in Deflandre & Fert,
1954) Wind & Wise
in Wise & Wind, 1977 (sample 2310); figs. 2-3, 6: Tetrapodorhabdus
coptensis Black, 1971a (sample 2304); fig. 4: Axopodorhabdus dietzmannii (Reinhardt,
1965) Wind & Wise, 1983
(sample 2302); fig. 5: ? Tetrapodorhabdus
coptensis Black, 1971a (sample 2304); fig. 7: Tetrapodorhabdus
decorus (Deflandre in Deflandre & Fert,
1954) Wind & Wise
in Wise & Wind, 1977
(sample 2346); fig. 8: ? Axopodorhabdus dietzmannii (Reinhardt,
1965) Wind & Wise, 1983
(sample 2305); figs. 9-11: Flabellites
biforaminis Thierstein, 1973 (fig. 10, sample 2327; figs.
9, 11, sample 2335); figs. 12-13:
Diazomalithus galicianus Kaenel & Bergen,
1996 (fig. 12, sample 2284; fig. 13, sample 2305); figs. 14-15: Diazomatolithus
lehmanii Noël, 1965 (fig. 14, sample 2295; fig. 15, sample 2284);
fig. 16: Discorhabdus
ignotus (Górka, 1957) Perch-Nielsen,
1968
(sample 2339); fig. 17: Biscutum
constans (Górka, 1957) Black,
1968 (sample 2327); figs. 18, 23: Haqius
circumradiatus (Stover, 1966) Roth,
1978 (sample 2324); fig. 19:
Indeterminate, monstruous coccolith without central area? (sample 2284); figs. 20-21: Watznaueria
britannicus (Stradner, 1963) Reinhardt,
1964 (fig.
20, sample 2302; fig. 21, sample 2335); fig. 22: Biscutum
dubium (Noël, 1965) Grün
in Grün et al., 1974
(sample 2327). |
|
Plate 7:
figs. 1-2: Percivalia
hauxtonensis Black, 1973 (sample 2325); fig. 3: Percivalia
fenestratus (Worsley, 1971) Wise,
1983 (sample 2339); fig. 4: Pickelhaube
furtiva (Roth, 1983) Applegate et
al. in Covington
& Wise, 1987 (sample 2304); figs. 5-9: Crucibiscutum
salebrosum (Black, 1971a) Jakubowski,
1986 (fig. 5,
sample 2332; figs. 6, 9, sample 2335; fig. 7, sample 2317; fig. 8, sample 2322);
fig. 10:
Indeterminate, missing central area? (sample 2284); fig. 11: Watznaueria
ovata Bukry, 1969 (sample 2310); figs. 12-13: Tubodiscus
burnettiae Bown in Kennedy et
al.,
2000 (fig. 12, sample 2317; fig. 13, sample 2327); fig. 14: Manivitella
pemmatoidea (Deflandre in Manivit,
1965) Thierstein,
1971 (sample 2317); fig. 15: Stradnerlithus
ellipticus (Bukry, 1969) Perch-Nielsen,
1984 (sample 2335);
figs. 16-19: Stoverius
achylosus (Stover, 1966) Perch-Nielsen,
1986 (fig.
16, sample 2335; figs. 17-18, sample 2302; fig. 19, sample 2304); figs. 20-21: Braarudosphaera
bigelowii (Gran & Braarud,
1935) Deflandre,
1947 (fig. 20, sample 2284; fig. 21, sample 2305); fig. 22: Braarudosphaera
africana Stradner, 1961
(sample 2305); figs. 23-24: Orastrum
perspicuum Varol in Al-Rifaiy et al.,
1990 (sample 2346). |
|
Plate 8:
fig. 1: Rotelapillus sp., large profile shape (sample 2327); fig. 2: Rotelapillus
crenulatus (Noël, 1956) Perch-Nielsen,
1984, normal size natural light sample (sample 2299); figs. 3-5: Zebrashapka
vanhintei Covington & Wise,
1987 (fig. 3, sample
2339, fig. 4, sample 2304; fig. 5, sample 2304); figs. 6-8: Eprolithus
floralis (Stradner, 1962) Stover,
1966 (figs. 6-7,
sample 2335; fig. 8, sample 2284); fig. 9: Stradnerlithus
ellipticus (Bukry, 1969) Perch-Nielsen,
1984 (sample 2304);
fig. 10: Micrantholithus
obtusus Stradner, 1963
(sample 2324); figs. 11-12: Lapideacassis
glans Black, 1971b (sample 2284); fig. 13: Lapideacassis
mariae Black, 1971b (sample 2335); fig. 14: Lithraphidites
carniolensis Deflandre, 1963
(sample 2304); figs. 15-16: Calculites dispar Varol in Al-Rifaiy et
al., 1990 (fig. 15, sample 2302; fig. 16, sample 2295); figs. 17-18: Orastrum
partitum (Varol in Al-Rifaiy et
al., 1990) Bergen, 1998 (fig. 17, sample 2292; fig. 18,
sample 2310); figs. 19-20: Hayesites
irregularis (Thierstein in Roth & Thierstein,
1972) Applegate et
al. in Covington
& Wise, 1987 (fig. 19, sample 2332; fig. 20, sample 2302); figs. 21-23: Calciosolenia (Deflandre in Deflandre & Fert,
1954) Bown
in Kennedy et al.,
2000 (fig. 21, sample 2345; fig. 22, sample 2304;
fig. 23, sample 2322). |
|
Plate 9: Clavaillan section Deshayesites
grandis Zone (Nannofossils association): figs.
1, 6: Discorhabdus serratus Worsley, 1971
(sample 2387); figs.
2-3, 7-8: Parabdolithus infinitus (Worsley, 1971) (figs. 2, 7, sample 2385; figs. 3, 8, sample 2393); fig.
4-5, 9-10: Calcicalathina erbae Bergen, 1998
(sample 2385); figs.
11 (LN), 16 (LP): Lapideacassis sp.
(sample 2395); figs.
12 (LN), 17 (LP): Lapideacassis glans Black,
1971b (sample 2385); fig.
13: Lapideacassis glans Black,
1971b (sample 2387); fig.
14: Micrantholithus obtusus Stradner, 1963
(sample 2385); fig.
15: Eiffellithus hancockii Burnett,
1997 (sample 2389); fig. 18, Calculites dispar Varol
in Al-Rifaiy et al., 1990 (sample 2389); fig.
19: Conusphaera rothii (Thierstein, 1971) Jakubowski,
1986 (sample 2387); fig.
20: Tetralithus ? malticus
Worsley, 1971 (sample 2387); figs.
21-22: Radiolithus planus Stover, 1966
(sample 2386); fig.
23: Eprolithus floralis (Stradner, 1962) Stover,
1966 (sample 2387); fig.
24: Rhagodiscus angustus (Stradner, 1963) Reinhardt, 1971 (tiny specimen, sample 2387); fig.
25: Rhagodiscus angustus (Stradner, 1963) Reinhardt, 1971 (sample 2395). |
|
Plate 10: Clavaillan section Deshayesites grandis Zone (Nannoconus association B): figs. 1-3: N. steinmanii Kamptner, 1931 (fig. 1, sample 2393; fig. 2, sample 2385; fig. 3, sample 2386); figs. 4-5: N. cf. steinmanii Kamptner, 1931 (sample 2387); figs. 6-7: N. elongatus Brönnimann, 1955 (fig. 6, sample 2385; fig. 7, sample 2386); fig. 8: ? N. elongatus Brönnimann, 1955 (sample 2386); fig. 9: N. wassalli Brönnimann, 1955 (sample 2385); fig. 10: Nannoconus aff. borealis Perch-Nielsen, 1979 (sample 2393); fig. 11: N. boneti Trejo, 1959 (sample 2393); fig. 12: N. bucheri Brönnimann, 1955 (large cavity, sample 2388); fig. 13: N. bucheri Brönnimann, 1955 (thin wall specimen, sample 2388); fig. 14: N. bucheri Brönnimann, 1955 (short specimen, sample 2388); fig. 15: N. circularis Deres & Achéritéguy, 1980 (sample 2387). Dufrenoyia
furcata Zone (Nannoconus association GI): fig.16:
N. inconspicuus Deflandre &
Deflandre-Rigaud, 1962 (sample 2407); fig.
17: N. kamptneri Brönnimann, 1955
(sample 2407); figs.
18-19: N. aff. grandis Deres
& Achéritéguy, 1980 (fig. 18, sample 2407; fig. 19, sample 2406); fig.
20: N. boneti Trejo, 1959
(sample 2407); figs.
21-24: Assipetra. 21-22: large forms (above 10µm), 23-24: small forms
(below 10µm); figs.
21, 23-24: Assipetra terebrodentarius (Applegate et
al. in Covington
& Wise, 1987) Rutledge & Bergen
in Bergen, 1994 (sample 2386); fig.
22: Assipetra
infracretacea (Thierstein,
1973) Roth, 1973 (sample 2386); fig.
25: N. vocontiensis Deres &
Achéritéguy, 1980 (sample 2407). |
|