◄ Carnets Geol. 15 (11) ►
![]()
Contents
[1. Introduction]
[2. Brazilian Cretaceous shells]
[3. Paleobiogeographic discussion]
[4. Conclusions] and ... [Bibliographic references]
Dépt. STU,
Fac. Sci. Tech., UBO, CS 93837, F-29238 Brest (France)
;
Department of Ecology and Evolutionary Biology, The University of Kansas, 1200 Sunnyside Avenue, Lawrence, Kansas 66045 (USA)
UNESP - Universidade Estadual Paulista, Center for
Geosciences Applied to Petroleum (UNESPetro) & Departamento de Geologia
Aplicada, Caixa Postal 178, Av. 24 A, nº 1515, Bela Vista, CEP13506-900 - Rio Claro - SP (Brazil)
Published online in final form (pdf) on July 14, 2015
[Editor: Robert W. ; language editor:
Donald E. ]
![]()
Out of the few records of rudists from the Cretaceous strata of the South Atlantic coastal basins only two refer to Brazilian localities. However, petrographic analyses demonstrate that these shells should be assigned to Ostreids or to Pycnodontids rather than to Rudistids. More specifically, the domain considered herein, north of the Río Grande Rise - Walvis Ridge barrier, was part of the warm-water "tropical" realm, but it was not part of the Mesogean domain because both Rudistids and Orbitolinas are missing. In addition, the scarcity of corals leads us to ascribe the taphonomic assemblage to the Chloralgal facies. Neither generalized hypersalinity or extreme sea-water temperatures seem to account for these biotic peculiarities. Instead, our alternative hypothesis favors the driving role played by oceanic circulation in the dispersal of the benthic organisms.
Rudists; Ostreids; Pycnodontids; corals; Orbitolinids; calcareous algae; Cretaceous; Albian; Cenomanian; South Atlantic; Tethys; Mesogea; Chloralgal; paleobiogeography.
B. & D. (2015).- End of a modern geological myth: there are no rudists in Brazil! Paleobiogeographic implications.- Carnets Geol., Madrid, vol. 15, nº 11, p. 123-136.
Fin d'un mythe géologique moderne : il n'y a pas de rudistes au Brésil ! et ses implications paléobiogéographiques.- Parmi les quelques signalements de rudistes dans les séries crétacées des bassins côtiers de l'Océan Atlantique Sud, deux seulement correspondent à des sites brésiliens. Toutefois, les analyses pétrographiques montrent que ces coquilles ne sauraient être attribuées à des rudistes mais plutôt à des Ostréidés ou à des Pycnodontidés. Plus spécifiquement, les régions situées au N de la barrière Rio Grande Rise-Walvis appartenaient au domaine à eaux "tropicales" chaudes, mais pas au domaine mésogéen parce que rudistes et orbitolines y font défaut. De plus, la rareté des coraux nous a conduit à attribuer l'association taphonomique au faciès Chloralgal. Ni une hypersalinité généralisée, ni des températures particulièrement élevées de l'eau de mer ne semblent rendre compte de ces particularités biotiques. Au lieu de cela, notre hypothèse alternative favorise le rôle moteur joué par la circulation océanique dans la dispersion des organismes benthiques.
Rudistes ; Ostréidés ; Pycnodontidés ; coraux ; Orbitolinidés ; algues calcaires ; Crétacé ; Albien ; Cénomanien ; Atlantique Sud ; Téthys ; Mésogée ; Chloralgal ; paléobiogéographie.
The presence or absence of rudists in mid- and Upper Cretaceous limestones of the Brazilian coastal basins is an episodical issue, recurrent since the 1970's. The myth of Brazilian rudists probably began with the international diffusion of "ready-made" sedimentological models for Cretaceous carbonate environments commonly referred to in the oil industry. In the present publication, we examine some large bioclasts, which were eventually ascribed to rudists, including few that are not even molluscan shells as demonstrated by "quick-look" petrographic analyses. We also take advantage of the opportunity in this review to address paleobiogeographic considerations regarding mid-Cretaceous tropical assemblages.
It is almost impossible to track back the exact date of the first reference to models with the so-called "rudist shoal banks or reefs" in Brazil because this source of further quotes (e.g., the recent et al., 2007) is probably to be found in a confidential and proprietary report of a petroleum company. However, as early as the 80's, authors started publishing papers reporting rudists in Brazil. For instance, et al. (1981) report "rudistids, bryozoans, corals" in their "microfacies 50" of the Macaé Formation (Macaé Group, Quissamã Formation, Albian, as nowadays considered) of the Campos basin, however their illustrations do not provide indisputable rudistid shells. Also a number of paleobiogeographic maps show rudists in the Sergipe Basin (e.g., , 1982: Fig. 6 [*]; , 1987: Fig. 1; , 1999: Fig. 1.1).
Bruno
(the first author of the present paper), the late Pierre-Yves and
the late Alain François (1991b) stated that "L'affleurement
des calcaires à Rudistes et des marnes de la Fazenda Cafuz [?Lower-Middle
Albian, Riachuelo Formation, Sergipe Basin] est (...) l'unique
gisement de Rudistes connu dans les bassins atlantiques de la marge brésilienne"
[the outcrop with rudistid limestones and marls at Fazenda Cafuz (Fig. 1 ![]() , red
star) is the sole locality with rudists known in the Brazilian basins sited on
the South Atlantic margin].
, red
star) is the sole locality with rudists known in the Brazilian basins sited on
the South Atlantic margin].
Click on thumbnail to enlarge the image.
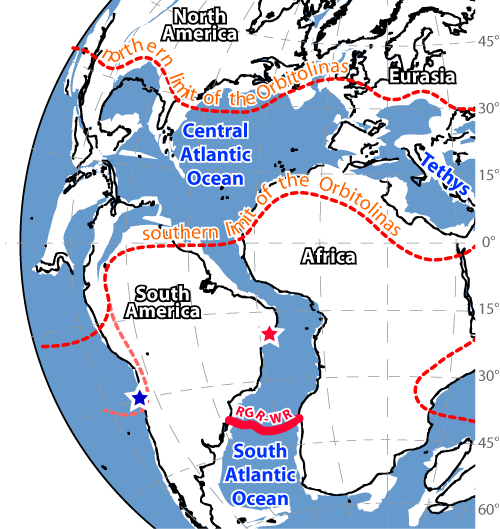
Figure 1: Maximum extension of the Orbitolinas on a Azimuthal map projection with reconstruction of continents for the mid-Cretaceous (circa 100Ma). The red star is for Fazenda Cafuz, Sergipe, Brazil; the blue star is for 's forgotten rudists from the Chañarcillo (Atacama) Basin, Chile. Plate fragments as at 100Ma: grey; present-day shorelines: black; approximate position of the Río Grande Rise - Walvis Ridge barrier (RGR-WR): red. The same base map was used in et al. (2014). It was initially generated using the ODSN (Ocean Drilling Stratigraphic Network) Plate Tectonic Reconstruction Service established by GEOMAR, Research Center for Marine Geosciences at Kiel and the Geological Institute of the University of Bremen (URL: http://www.odsn.de/odsn/services/paleomap/paleomap.html). Parameters used to generate base map were as follows: cartographic projection: Lambert Azimuthal; move plates relative to: Magnetic Ref. Frame; guideline interval 15°; annotation interval 15°; reconstruction age: 100 Ma; map boundaries: 90° North, -120° West, 90° East, 90° South; frame type: thin lines.
There was a concern as regards this
singular occurrence and the uniqueness of this outcrop and a need to re-examine
Pierre-Yves 's material: unfortunately, this material was lost.
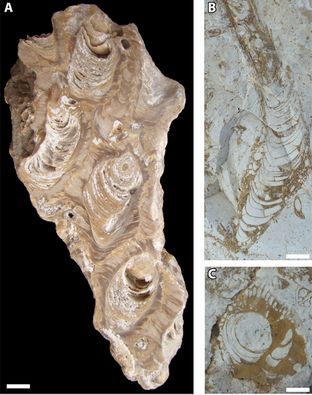
Luckily, Paulo had some specimens from the exact same locality (Fig. 2 ![]() ). At first sight, sections of the shells visible on polished slabs
(Fig. 3.A
). At first sight, sections of the shells visible on polished slabs
(Fig. 3.A ![]() ) evoke sections of rudists. They display foils
(3.A
) evoke sections of rudists. They display foils
(3.A ![]() ) that look similar
to the curved septa observed in some Caprinid rudists (Fig.
3.B-C
) that look similar
to the curved septa observed in some Caprinid rudists (Fig.
3.B-C ![]() ), for example.
Petrographic thin sections should contribute to the analysis and solve the case.
), for example.
Petrographic thin sections should contribute to the analysis and solve the case.

Click on thumbnail to enlarge the image.
Figure 2: Ostreid shells not rudists on the outcrop at Fazenda Cafuz, Sergipe Basin, Brazil; Riachuelo Formation, ?Lower-Middle Albian.
The shells of the rudists, which are
exclusively known from the fossil record, consist of 2 layers: a calcitic outer
layer and an aragonitic inner layer. Some families -- mostly post-Cenomanian --
include calcite-dominated taxa whereas others -- mostly pre-Turonian -- group
aragonite-dominated taxa (, 2002: Fig. 3); the remaining families
consist of taxa with no dominance of calcite over aragonite. In
calcite-dominated shells, aragonite was always present, never scarce as in an
ostreid shell (, 2002: Fig. 2). In contrast, calcite might be
almost non-preserved in aragonite-dominated shells, as in the Offneria
specimens from Lebanon (J.-P. et al., 2015a) illustrated here (Fig.
3.B-C ![]() ).
).
Click on thumbnail to enlarge the image.
Figure
3:
A) Polished slab with a few
large calcite-dominated shells. Fazenda Cafuz, Sergipe Basin, Brazil; Riachuelo
Formation, ?Lower-Middle Albian. B-C) Offneria specimens from Lebanon.
These aragonite-dominated shells were leached and the cavity was later filled by
calcitic or dolomitic cements. Beit Mery, Matn District, Lebanon; Jezzinian,
lowermost Aptian (lower Bedoulian); B) see J.-P. et al.,
2015a:
Fig. 2B ![]() & Pl. 1, fig. I
& Pl. 1, fig. I ![]() ; C) see
J.-P. et al., 2015a: Pl. 1,
fig. G
; C) see
J.-P. et al., 2015a: Pl. 1,
fig. G ![]() . Scale
bars =1 cm.
. Scale
bars =1 cm.
For its part, the shell of the modern Ostrea consists of 4 discrete layers:
Our so-called rudist shells (Figs. 2 ![]() ,
3.A
,
3.A ![]() & 4.A-C
& 4.A-C ![]() ) are valves of an Ostreid similar to those illustrated by
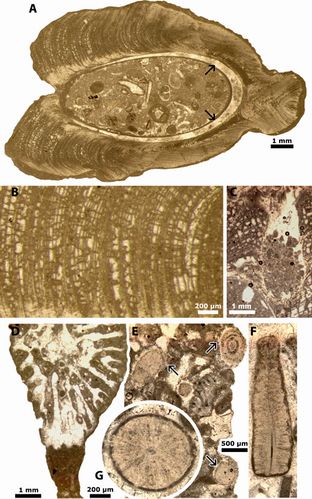
(1969: Pl. 72, fig. 2, to compare with our Fig. 4.A-C
) are valves of an Ostreid similar to those illustrated by
(1969: Pl. 72, fig. 2, to compare with our Fig. 4.A-C ![]() ) showing the "loosely
fabricated latticework of foliated layers forming 'chambers' in the shell".
) showing the "loosely
fabricated latticework of foliated layers forming 'chambers' in the shell".

Click on thumbnail to enlarge the image.
Figure 4: A-C) As noted by (1969), the "loosely fabricated latticework of foliated layers" form "chambers" in this Ostrea shell. The early marine dull cement and the thin micritic "microbial" crusts on the shell fibrous laminae fill original void spaces and were not leached aragonitic parts. The interspaces are now partly or fully filled by drusy calcitic cement (crystals centripetally increasing in dimensions). Fazenda Cafuz, Sergipe Basin, Brazil; Riachuelo Formation, ?Lower-Middle Albian.
More recently, et al.
(2010) reported new "Ocorrência de rudistas em amostras de testemunho
do Albiano inferior da Bacia de Campos" [occurrence of rudists in core
samples from Lower Albian strata of the Campos Basin]. In this case, the
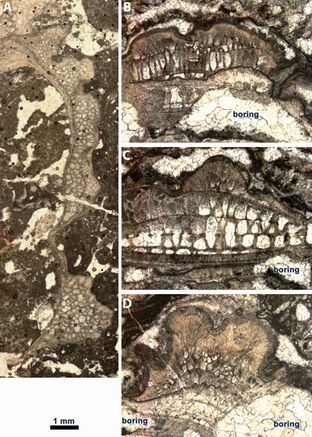
illustrated material seems to be valves of Pycnodonte (Fig.
5.A-C ![]() ),
comparable with those illustrated by (1969: Pl. 72, fig. 1),
with vesicular portions. Additional sections illustrating the skeletal
microstructure of Pycnodontid pelecypods are found in and
(1971: Pl. 35, figs. 1-4). Pycnodonte
vesiculosa (,
1822) is a species well known from Brazil ( & ,
1999, inter alia) but also from Angola (where it is coined either as "Ostrea"
or "Gryphea" by , 1888a: p. 18;
1888b: p. 91-92,
Pl. V, figs. 15-17).
),
comparable with those illustrated by (1969: Pl. 72, fig. 1),
with vesicular portions. Additional sections illustrating the skeletal
microstructure of Pycnodontid pelecypods are found in and
(1971: Pl. 35, figs. 1-4). Pycnodonte
vesiculosa (,
1822) is a species well known from Brazil ( & ,
1999, inter alia) but also from Angola (where it is coined either as "Ostrea"
or "Gryphea" by , 1888a: p. 18;
1888b: p. 91-92,
Pl. V, figs. 15-17).
Click on thumbnail to enlarge the image.
Figure 5: A-D) Pycnodonte shells with vesicular layer, commonly bored.- Sergipe Basin, Brazil; Riachuelo Formation, ?Lower-Middle Albian. A) UPAFSE_0052; B-D) UPAFSE_0129. The thin inner aragonitic layer was replaced by a mosaic of calcite (nearly equant crystals, with ghosts of organic linings); organic pores and borings are filled by drusy calcitic cement (crystals centripetally increasing in dimensions).
Folded shells
are commonly observed in Cretaceous limestones of Brazil (e.g.,
& , 1999) and are usually referred to Lopha-type
shells (Fig. 6.A-F ![]() ). J.H.
(1968: p.
55, Pl. 12, fig. 1) illustrated similar structures from the Comanche Peak
Formation, Albian of Texas. However, he erroneously ascribed them to the algal
genus Clypeina (). The first
author (B.G.) got the opportunity to examine 's
material ( et al., 2013c) and more specifically the thin
section Shell BFP-6459, USNM 42772 (Fig. 6.G-H
). J.H.
(1968: p.
55, Pl. 12, fig. 1) illustrated similar structures from the Comanche Peak
Formation, Albian of Texas. However, he erroneously ascribed them to the algal
genus Clypeina (). The first
author (B.G.) got the opportunity to examine 's
material ( et al., 2013c) and more specifically the thin
section Shell BFP-6459, USNM 42772 (Fig. 6.G-H ![]() ).
The calcitic nature of these "fragments" most definitively disqualifies them
to be ascribed to algae; conversely, the foliated structure of their folded
calcitic layers as observed in thin sections is typically that found in mollusc
shells.
).
The calcitic nature of these "fragments" most definitively disqualifies them
to be ascribed to algae; conversely, the foliated structure of their folded
calcitic layers as observed in thin sections is typically that found in mollusc
shells.

Click on thumbnail to enlarge the image.
Figure 6: A-C) Lopha-like shells.- UPAFSE_015c and UPAFSE_016, Sergipe Basin; Riachuelo Formation, ?Lower-Middle Albian; D-F) Lopha-like shells.- Santos Basin; G-H) Lopha-like shell, identified as "Clypeina species" in (1968: Pl. 12, fig. 1). G: conventional transmitted light, H: cross polarized light.- USNM_42772, Comanche Peak Formation, Texas (U.S.A.). In A-C, E and F-G, the aragonitic layer was leached and the moldic vug was later filled by drusy calcitic cement (crystals centripetally increasing in dimensions).
Some reported
rudist shells may actually be serpulid tubes, which consist
of 2 layers: an aragonitic inner layer and a calcitic outer layer (Fig.
7.A ![]() ).
The outer layer may be alveolar (Fig. 7.A-C
).
The outer layer may be alveolar (Fig. 7.A-C ![]() ) as in Pyrgopolon ,
1808, for instance. For example, the Radiolitidae shells from the Upper
Aptian-Lower Albian of Provence (France) illustrated by P.
(1988: Pl. VI, figs. 1-9) may well be serpulid tubes.
) as in Pyrgopolon ,
1808, for instance. For example, the Radiolitidae shells from the Upper
Aptian-Lower Albian of Provence (France) illustrated by P.
(1988: Pl. VI, figs. 1-9) may well be serpulid tubes.
Click on thumbnail to enlarge the image.
Figure 7: A-C) Serpulid tube.- A) the thin aragonitic inner layer was replaced by mosaic calcite (nearly equant crystals); the thicker calcitic outer layer is locally alveolar; B) in the grid pattern curved rows come to the fore; A-B) 1649.40m, RNS11, Potiguar Basin; C) bored serpulid tube.- UPAFSE_035, Sergipe Basin, Brazil; Riachuelo Formation, ?Lower-Middle Albian; D) Coral bioclast in an oolitic facies.- , 566, Barreirinhas Basin; E) arrows point to alcyonarian sclerites (Pieninia, see , 1986).- , Eo-1+4, Barreirinhas Basin; F-G) alcyonarian sclerites (Pieninia).- , 282, Barreirinhas Basin.
In terms of paleoceanography, though the Tethys Ocean, which opened from East to West, was connected to the northern segment of the young South Atlantic Ocean [we shall use the acronym NYSAO], the latter opened at an angle with respect to the first and, in terms of geodynamics, and NYSAO cannot be considered a branch of the Tethys. However, considering the ecological factors, the question of the scarcity or total absence of some benthic "Tethyan" biota in the proto-South Atlantic Ocean during the "middle" and Late Cretaceous time should be addressed:
Not only the
rudists (as documented above) but also some large benthic foraminifers such as
the Orbitolinas sensu lato are missing in Brazil (,
2000; & ,
2013). In addition, other groups
such as the hermatypic corals are not missing but rather scarcely represented (Fig.
7.D ![]() );
alcyonarians
(Fig. 7.E-G
);
alcyonarians
(Fig. 7.E-G ![]() )
are rare. The situation is almost identical on
the eastern side of the NYSAO. Only two very early records
documented rudists from Angola:
)
are rare. The situation is almost identical on
the eastern side of the NYSAO. Only two very early records
documented rudists from Angola:
a) (1886: p. 155) who described oolitic limestones with rolled pieces of corals and "fragments d'un grand bivalve rappelant le genre Pachyrisma" [fragments of a large bivalve mollusc close to the genus Pachyrisma];
b) (1888a, p. 25) who reports questionable remains of Pachyrisma and two of Requienia (sometimes called Toucasia in old publications). However, we have to be very cautious as such rare finds have never been confirmed since then. Actually there were no records either of rudists or of Orbitolinas from Gabon, Congo-Brazzaville (e.g., P. , 1995), Congo-Kinshasa and Angola where mid-Cretaceous strata have been intensively drilled and cored in the search for post-salt oil and gas reservoirs.
Based on our current knowledge we shall examine what are the possibilities to resolve this paleobiogeographic dilemma.
1) The first approach is based on the analysis of selected elements of the biotic content.
That was the approach of and (1982) who focused on the Early Cretaceous Urgonian (Late Hauterivian-Bedoulian) platforms. These authors assumed that "les éléments les plus caractéristiques en ont été les Rudistes non constructeurs (Requieniés, Monopleuridés) et les Orbitolinidés" [their most characteristic features were rudists that do not form framework structures (Requienids, Monopleurids) and orbitolinids].
Similarly, for the "middle" Cretaceous (Aptian-Cenomanian), (1971: Fig. 3) mapped the extension of the Orbitolinas -- among other foraminifers --; on the other hand, (1973: Fig. 3) combined rudists and Orbitolinas while (1973: Fig. 1), (1987: Fig. 1), & (1988: Fig. 1) and et al. (1993: Fig. 2) used rudists or hermatypic corals. In addition, other groups of organisms were eventually considered to produce similar maps (e.g., actaeonellid gastropods by , 1971, 1987). However, the most frequent combination remains that of rudists and Orbitolinas.
The original idea of using this combination dates back to 1900 and is to be attributed to Henri with the definition of the Mesogean domain:
"Grâce à la considération de ce double élément, Rudistes et Orbitolines ( ), il devient relativement facile de tracer sur une carte l'aire où ces animaux ont vécu aux diverses époques géologiques; ces aires d'habitat ont naturellement varié d'une époque à une autre et leurs variations indiquent les modifications successives éprouvées par le rivage des mers, mais ces modifications sont ordinairement assez faibles." (H. , 1900: p. 222) [The consideration of these two components, rudists and Orbitolinas ( ), makes it relatively easy to map the area where these animals lived at various times in the geological past; these habitat areas have, of course, changed from one period to another and these changes witness to successive shifts of the shoreline, but these changes are usually quite low].
"Ces divers fossiles se trouvent répartis sur une bande régulière que l'on peut suivre d'une manière continue depuis le Mexique et la mer des Antilles à l'ouest jusqu'aux îles de la Sonde à l'est : elle correspond à ce que l'on appelle d'habitude la zone méditerrannéenne. Cette dénomination prête un peu à confusion avec les environs immédiats de la Méditerranée elle-même, et nous proposerons de la remplacer par zone Mésogéenne ou Mésogée." (op. cit.: p. 223) [These various fossils are distributed in a regular realm that can be traced continuously from Mexico and the Caribbean Sea in the West all the way to the Indonesian Sunda Islands in the East; it corresponds to the so-called Mediterranean domain. The name itself leads to confusion with the immediate vicinity of the Mediterranean Sea; thus, we suggest using Mesogean domain or Mesogea instead].
"C'est l'existence de cette mer tropicale continue qui donne à la période crétacée son caractère particulier." (op. cit.: p. 223) [The existence of this contiguous tropical sea determines the unique character of the Cretaceous Period].
"La Mésogée correspond à une phase particulière ( ) de la Téthys de ; c'est uniquement la mer dans laquelle les Rudistes ont vécu et se sont développés." (op. cit.: footnote 2, p. 223) [The Mesogea is a special phase ( ) of the Tethys sensu ; it is only this sea where the rudists have lived and developed].
Actually
plotting on a world map either the rudists or the Orbitolinas or both produces
almost the same outcomes (Fig. 1 ![]() ). Some earlier
authors plotted Orbitolinas -- but not the rudists -- on today's world map: e.g.,
(1971: Fig. 1) combined 's
(1959: Fig. 3) and ' (1960: Fig. 20)
datasets. Note that was the only one of the three
who referred to 's
concept of the Mesogea. Slightly later and
(1982: Fig. 3) who started plotting rudistid facies as well
as Orbitolina localities on palinspastic
world maps (actually on a "mid"-Cretaceous palinspastic world maps for Early
Cretaceous fossils) concluded that it quite consistently constrains the maximal
extension of the Early Cretaceous "Mesogean phenomenon".
). Some earlier
authors plotted Orbitolinas -- but not the rudists -- on today's world map: e.g.,
(1971: Fig. 1) combined 's
(1959: Fig. 3) and ' (1960: Fig. 20)
datasets. Note that was the only one of the three
who referred to 's
concept of the Mesogea. Slightly later and
(1982: Fig. 3) who started plotting rudistid facies as well
as Orbitolina localities on palinspastic
world maps (actually on a "mid"-Cretaceous palinspastic world maps for Early
Cretaceous fossils) concluded that it quite consistently constrains the maximal
extension of the Early Cretaceous "Mesogean phenomenon".
On our map
of the "middle" Cretaceous (Fig. 1 ![]() ), we drew the maximum northern and
southern extent of Orbitolinas, i.e., roughtly the maximum extension of
the Mesogean domain:
), we drew the maximum northern and
southern extent of Orbitolinas, i.e., roughtly the maximum extension of
the Mesogean domain:
This Mesogean domain should not be confused with the Tethys: the Mesogea corresponds to the warm-water biotic province of both the Tethys and the Panthalassa, based upon the sole distribution of distinctive assemblages of organisms (i.e., rudists and Orbitolinas for the Early-"middle" Cretaceous). The NYSAO, which is devoid of both rudists and Orbitolinas, is by definition not part of it.
2) The second approach is still based solely on the analysis of biotic content, but includes calcareous green algae.
Apart from the rudists and Orbitolinas (i.e., the criteria to identify the Mesogean area) we could have considered the hermatypic (= reef-builder) corals and the calcareous green algae (CGA sensu , 2012). That was the approach of J.-P. (1992: Fig. 4) and also that of and (1972) when discussing the factors that control the distribution of modern carbonate platforms. Their starting point is that "hermatypic corals and calcareous green algae live only in warm seas" (op. cit.). and (1972) call "Chlorozoan" (Chlorophyta + Zoantharia with their symbiotic zooxanthellae) the association of skeletal grains derived from photo-autotrophic organisms. Nowadays annual minimum surface water temperature (of at least 14°C) and annual mean temperature (of at least 23°C) are the major physical factors controlling the Chlorozoan distribution ( & , 1972).
As
already mentioned earlier by &
(2013),
hermatypic corals are rather scarcely represented in Brazilian and West-African
mid-Cretaceous series, but CGA are quite common and diverse (Brazil:
et al., 1991a, 1991b, 2008,
2013a, 2013b, 2014;
, 2015; Congo: P. ,
1995).
The maximum extension of the mid-Cretaceous
Chlorozoan association, characterized by the presence of CGA bioclasts,
corresponds roughtly to a marine
warm-water realm with latitudes ranging between circa 35 degrees North and circa 35
degrees South (Fig. 1 ![]() ). During this time the NYSAO,
which was not part of the Mesogean domain, was part of the warm-water, i.e., tropical, realm as herein defined.
). During this time the NYSAO,
which was not part of the Mesogean domain, was part of the warm-water, i.e., tropical, realm as herein defined.
3) The third approach is also based on the analysis of biotic content, but incorporates some non-skeletal grains, i.e., ooids and grain aggregates. and (1972) pointed out that ooids and aggregates are restricted to their Chlorozoan associations but do not necessarily occur in all of them. (1975) advocated that the Chlorozoan association should be split in two: the Chlorozoan sensu stricto and the Chloralgal. The latter "accommodates those sediments containing calcareous green algae but no corals", a definition that fits quite well that of the margins of the NYSAO.
To summarize, the NYSAO, which was not a branch of the Tethys in terms of geodynamics, was located in the southern half of the mid-Cretaceous warm-water realm, i.e., the warm-water realm for the southern hemisphere. It did not belong to the tropical Mesogean domain because it lacked the typical biota, i.e., rudists and Orbitolinas. It was characterized by its Chloralgal, not Chlorozoan sensu (1975), tropical assemblage.
(1975) used a STAR diagram (Salinity Temperature Annual Ranges) to distinguish biofacies. Chlorozoan assemblages are related to a 'normal' salinity of seawater (30 - 40 ) and Chloralgal to hypersaline (>40 ) or brackish (<30 ) environments. In addition, based on our current understanding of coral bleaching phenomenon, extremely warm seawater temperature is one of the causative factors of the dissociation of algal symbionts and their hosts (corals, rudists, large benthic foraminifers). On the basis of a microfacies analysis of lower-middle Albian carbonate rocks from the Campos Basin, i.e., in the southern part of the NYSAO, (1987) suggested that the low foraminiferal diversity could result from high temperatures and hypersalinity. Eight years later, he pointed out that environmental conditions might have been more favorable in Brazilian basins from the northern part of the NYSAO (, 1995). But, another five years later, he applied the scenario of "high temperatures and hypersalinity" to the whole NYSAO (, 2000).
Today,
we can rule out the generalized hypersalinity hypothesis for the entire shallow
water Late Aptian-Albian NYSAO (because, for instance, we have never observed
teratologic specimens of foraminifera like those from the hypersaline Abu Dhabi
coastal lagoons and marshes: Fig. 8 ![]() ). In addition, apart from the corals --
which are rare (but not totally absent in Brazil, nor in West Africa) --, we
have also detected in the mid- and Upper Cretaceous limestones of Brazil some
benthic elements known from the Tethyan realm, such as the foraminifera Coscinoconus
(with an aragonitic test, formerly quoted as Trocholina), Rhapydionina
liburnica (with a porcelaneous test) and Nezzazatinella picardi (with
an agglutinated test), as well as a great diversity of calcareous algae (either
green or red: see et al., 1991a, 1991b,
2008, 2013a, 2013b,
2014; & ,
2013; et al.,
2014;
, 2015).
This assemblage would not be consistent with the high-temperature hypothesis
(for the Late Aptian-Albian interval, see ,
2000), nor with
that of "narrow and temporarily [sic] connections" with the Tethys
(see & ,
2002). In addition, these supposed
narrow and temporary connections never prevented planktonic foraminifers from
entering the NYSAO
(, 2000). The remaining hypothesis is still
valid:
some Mesogean biota was not distributed southward because of unfavorable ocean
currents (
et al.,
2014).
). In addition, apart from the corals --
which are rare (but not totally absent in Brazil, nor in West Africa) --, we
have also detected in the mid- and Upper Cretaceous limestones of Brazil some
benthic elements known from the Tethyan realm, such as the foraminifera Coscinoconus
(with an aragonitic test, formerly quoted as Trocholina), Rhapydionina
liburnica (with a porcelaneous test) and Nezzazatinella picardi (with
an agglutinated test), as well as a great diversity of calcareous algae (either
green or red: see et al., 1991a, 1991b,
2008, 2013a, 2013b,
2014; & ,
2013; et al.,
2014;
, 2015).
This assemblage would not be consistent with the high-temperature hypothesis
(for the Late Aptian-Albian interval, see ,
2000), nor with
that of "narrow and temporarily [sic] connections" with the Tethys
(see & ,
2002). In addition, these supposed
narrow and temporary connections never prevented planktonic foraminifers from
entering the NYSAO
(, 2000). The remaining hypothesis is still
valid:
some Mesogean biota was not distributed southward because of unfavorable ocean
currents (
et al.,
2014).

Click on thumbnail to enlarge the image.
Figure 8: Deformed tests of Peneroplis planatus ( & , 1798) from Mussafah channel section, Abu Dhabi, United Arab Emirates.
Rudists are not present in Brazilian basins. References found in the literature point either to Ostreids ( et al., 1991b) or to Pycnodontids ( et al., 2010).
The NYSAO (the young South Atlantic Ocean
north of the Río Grande Rise - Walvis Ridge barrier), which was not a branch of
the Tethys in terms of geodynamics, belonged to a warm-water / tropical realm,
but not to the Mesogean domain (Fig. 1 ![]() ). Its biotic / taphonomic assemblages are
typically Chloralgal, not Chlorozoan, but the lack or scarcity of corals,
rudists and some large benthic foraminifers (mostly these hosts of algal
symbionts) does not look related to hypersalinity or extreme seawater
temperature. The most probable hypothesis is that, in mid- and even in Late
Cretaceous time, marine currents did not help either (most) coral and (any)
rudist larvae, or the flagellate gametes of some foraminifers (during their
brief phase of free and pelagic life) to penetrate the pathway to the NYSAO.
). Its biotic / taphonomic assemblages are
typically Chloralgal, not Chlorozoan, but the lack or scarcity of corals,
rudists and some large benthic foraminifers (mostly these hosts of algal
symbionts) does not look related to hypersalinity or extreme seawater
temperature. The most probable hypothesis is that, in mid- and even in Late
Cretaceous time, marine currents did not help either (most) coral and (any)
rudist larvae, or the flagellate gametes of some foraminifers (during their
brief phase of free and pelagic life) to penetrate the pathway to the NYSAO.
Elements of this paper were first presented at the 9th International Symposium on the Cretaceous System ( & , 2013). The first author (B.G.) benefited from a Smithsonian Fellowship allowing him to investigate the J. Harlan Collection stored in the premises of the Smithsonian Institution. He would like to thank the staff of the Department of Paleobiology at the Smithsonian National Museum of Natural History and particularly William A. and Jonathan G. for their hospitality and having facilitated his work there. In addition, this research was supported by the "Carbonatos do Brasil Project" linked to the Brazilian Sedimentology/Stratigraphy Net sponsored by Petrobras. The authors thank Jean-Pierre , Pierre , Jose Maria and Robert W. for having discussed some issues. Special thanks go to Paulo who gave us access to the material he collected at Fazenda Cafuz. Last but not least, the first author (B.G.) also thanks Ana Cristina who attempted to locate the Angolese collection of at the Laboratório Nacional de Energia e Geologia, Alfragide (Portugal).
J. (1958).- Quelques observations sur la nature de la coquille chez quelques mollusques.- Annales de la Société royale zoologique de Belgique, Bruxelles, t. LXXXIX, fasc. 9, p. 229-237.
P. (1886).- Note préliminaire sur des
fossiles recueillis par M. Lourenço , dans la province
d'Angola.- Bulletin de la Société Géologique de France, (3ª Série),
t. XV, p. 154-157.
URL: http://jubilotheque.upmc.fr/img-viewer/fonds-bulsgf/GB_000060_001/Contenu/JPEG_HD/viewer.html?ns=GB_000060_
001_J3_0152.jpg
P. (1888a).- Première
partie. Stratigraphie. In: P. & P. de,
Matériaux pour l'étude stratigraphique et paléontologique de la province
d'Angola.- Mémoires de la Société de Physique et Histoire Naturelle de Genève,
t. XXX, nº 2, p. 5-59.
URL: https://archive.org/stream/memoiresdelasoci1888soci#page/n255/mode/2up
P. (1888b).- Deuxième partie.
Description des fossiles crétaciques. Mollusques. In: P.
& P. de, Matériaux pour l'étude stratigraphique et paléontologique
de la province d'Angola.- Mémoires de la Société de Physique et Histoire
Naturelle de Genève, t. XXX, nº 2, p. 61-95 (Pls. I-V).
URL: https://archive.org/stream/memoiresdelasoci1888soci#page/n311/mode/2up
P. & P. de (1888).-
Matériaux pour l'étude stratigraphique et paléontologique de la province
d'Angola.- Mémoires de la Société de Physique et Histoire Naturelle de Genève,
t. XXX, nº 2, 116 p. (VIII Pls.).
URL:
https://archive.org/stream/memoiresdelasoci1888soci#page/n251/mode/2up
A.G. (1973).- Cretaceous Tethyan coral-rudist biogeography related to the evolution of the Atlantic Ocean. In: N.F., ed., Organisms and continents through time.- Palaeontological Association Special Paper 12, p. 169-174.
D. (1987).- A Bacia de Campos no Meso-cretáceo: uma contribuição a paleoceanografia do Atlântico Sul Primitivo.- Revista Brasileira de Geociências, São Paulo, vol. 17, p. 162-167.
D. (1995).- Calcisferas e microfácies em rochas carbonáticas pelágicas mesocretáceas: ocorrência na Margem Atlântico do Brasil impacto na configuração do Tétis.- Tese de Doutorado em Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, 3 vols., 688 p. (+ Atlas).
D. (2000).- Global stratigraphy, paleobiogeography and paleoecology of Albian-Maastrichtian pithonellid calcispheres: impact to Tethys configuration.- Cretaceous Research, vol. 21, nº 2-3, p. 315-349.
W.O. (1925).- Zur Kenntnis der Urgongesteine im südlichen Deutsch-Ostafrika nebst Beschreibung der darin vorkommenden Orbitolinen.- Palaeontographica, Stuttgart, Supplement VII, (Reihe II), Teil I, Lieferung I, p. 25-40 (Pl. 4).
F.C. (1971).- Cretaceous foraminiferal biogeography. In: F.A., P.F. & G., eds., Faunal provinces in space and time. Proceedings of the 17th Inter-University Geological Congress held in Queen Mary College (University of London) - 17, 18, 19 December 1969.- Geological Journal Special Issue, Liverpool, nº 4, p. 169-190.
R.C. (1960).- The foraminiferal genus Orbitolina in North America.- U.S. Geological Survey Professional Paper, Washington, 333, 52 p. (14 Pls.).
H. (1900).- Sur la distribution géographique
des Rudistes, des Orbitolines et des Orbitoïdes.- Bulletin de la Société géologique
de France, Paris, (3ème série), t. XXVIII, p. 222-235.
URL: http://jubilotheque.upmc.fr/img-viewer/fonds-bulsgf/GB_000074_001/Contenu/JPEG_HD/viewer.html?ns=GB_
000074_001_J3_0219.jpg
F.U.H., M.R. & A.V. (1981).- Petroleum geology of the Macaé Formation (Albian-Cenomanian), Campos Basin, Brazil carbonate microfacies depositional and diagenetic models natural and experimental porosity.- Ciência - Técnica - Petróleo, Petrobrás - Cenpes, Rio de Janeiro, vol. 11, 140 p.
W.A. (1973).- Marine life and ocean surface currents in the Cretaceous.- Journal of Geology, Chicago, vol. 81, nº 3, p. 269-284.
B. (1986).- Les Pieninia oblonga & , 1976, sont-elles des sclérites d'Alcyonaires ?.- Revue de Micropaléontologie, Paris, vol. 29, nº 2, p. 103-108.
B. (1992).- Les algues et foraminifères benthiques du Jurassique supérieur et du Crétacé inférieur du Sénégal.- Journal of African Earth Sciences, vol. 14, nº 2, p. 239-253.
B. (2012).- The contribution of calcareous green algae to the production of limestones: a review. In: D. & B., eds., Calcareous algae and the global change: from identification to quantification.- Geodiversitas, Paris, vol. 34, nº 1, p. 35-60.
B. (2015).- Algas calcárias marinhas bentônicas do Cretáceo do Brasil. In: D. & P., eds., Calcários do Cretáceo do Brasil: um Atlas.- IGCE/UNESP/Rio Claro, UNESPetro, Obra 1, p. 518-567.
B., P.-Y. & E. (1991a).- The Dasycladalean algae from the Cretaceous of the New World.- Transactions of the second geological Conference of the geological Society of Trinidad and Tobago, Port-of-Spain (April 2nd-8th, 1990), p. 178-183.
B., P.-Y. & A.F. (1991b).- Constructions bio-sédimentaires laminées, Lithothamnium et Parachaetetes de la Formation Riachuelo (Albien) du bassin de Sergipe (Nord-Est du Brésil).- Geociências, Sâo Paulo, 10, p. 169-181.
B. & D. (2013).- End of a myth: there are no rudists in Brazil!- 9th International Symposium on the Cretaceous System, Ankara (1-5 September 2013), Abstract Book, p. 21.
B., D. & I.I. (2008).- Calcareous algae from Upper AlbianCenomanian strata of the Potiguar basin (NE Brazil).- Geologia Croatica, Zagreb, vol. 61, nº 2-3, p. 311-320.
B., D. & I.I. (2013a).- Brasiliporella, a new mid-Cretaceous dasycladacean genus: the earliest record of the Tribe Batophoreae.- Facies, Erlangen, vol. 59, nº 1, p. 207-220.
B., D. & I.I. (2013b).- A new mid-Cretaceous Neomeris (dasycladacean alga) from the Potiguar Basin, Brazil.- Facies, Erlangen, vol. 59, nº 1, p. 221-230.
B., D. & I.I. (2014).- Clypeina tibanai, sp. nov. (Polyphysacea, Dasycladales, Chlorophyta), mid-Cretaceous green alga from the Potiguar Basin, Brazilian margin of the young South Atlantic Ocean.- Geologica Acta, Barcelona, vol. 12, nº 2, p. 227-237.
B., R. & K. (2013c).- Revision of the Jesse Harlan Collection. Part 1. Some fossil Dasycladales from Guatemala.- Carnets Geol., Madrid, CG2013_A07, p. 58 (Pl. 1), 66 (Pl. 2), 82 (Pl. 3), 88 (Pl. 4), 104 (Pl. 5), 130 (Pl. 6), 208 (Pl. 7), 280 (Pl. 8) & 281-301.
M.B., E.C. & P.P.E. (1979).- A biometric analysis of an Orbitolina fauna from the Cretaceous succession at Wolborough, S. Devon.- Proceedings of the Ussher Society, Exeter, vol. 4, part 3, p. 317-326.
M.B., M.P. & E.A.M. (2007).- The mid-Cretaceous fragmentation of Gondwana.- Geophysical Research Abstracts, vol. 9, 03548.
H.K. (1926).- Foraminifera from Venezuela and Trinidad.- Bulletins of
American Paleontology, Ithaca, vol. 12, nº 47, p. 1-46 (8 Pls.).
URL: https://archive.org/details/bulletinsofameri1247481928pale
A.S. & P.E. (1971).- Introductory petrography of fossils.- Springer-Verlag, Berlin, 302 p. (100 Pls.).
J.H. (1968).- Lower Cretaceous algae from Texas.- Professional Contributions of the Colorado School of Mines, Golden, nº 4, vii + 71 p. (12 Pls.).
E.G. (1973).- Cretaceous bivalvia. In: A., ed., Atlas of paleobiogeography.- Elsevier, Amsterdam, p. 353-383.
E.G. & C.C. (1988).- The morphological and ecological evolution of Middle and Upper Cretaceous reef-building rudists.- Palaios, Lawrence, vol. 3, p. 194-216.
A. (1975).- Possible influence of salinity and temperature on modern shelf carbonate sedimentation.- Marine Geology, vol. 19, nº 3, p. 159-198.
A. & A.T. (1972).- Modern temperate water and warm-water shelf carbonate sediments contrasted.- Marine Geology, vol. 13, nº 5, p. M67-M73.
C.R. (1982).- The Mid-Cretaceous Earth: paleogeography; ocean circulation and temperature; atmospheric circulation.- The Journal of Geology, vol. 90, nº 4, p. 393-413.
O.P. (1969).- Recognition of invertebrate fossil fragments in rocks and thin sections.- International sedimentary petrographical Series, Leiden, vol. XIII, 101 p. (106 Pls.).
J.-P. (1992).- The Lower Cretaceous Mesogean benthic ecosytems: palaeoecologic aspects and palaeobiogeographic implications.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 91, nº 3-4, p. 331-345.
J.-P., S., M., B. & D. (2015a).- Earliest Aptian Caprinidae (Bivalvia, Hippuritida) from Lebanon.- Carnets Geol., Madrid, vol. 15, nº 3, p. 21-30.
J.-P., F.-A. & M. (2015b).- Aptian-Albian rudist bivalves (Hippuritida) from the Chilean Central Andes: Their palaeoceanographic significance.- Cretaceous Research, vol. 54, p. 243-254.
P. (1988).- Évolution de la marge provençale à l'Aptien supérieur et à l'Albien. Sédimentologie, paléotectonique, paléogéographie.- Thèse, Doctorat de l'Université de Provence, Aix-Marseille I, 258 p. (XX Pls.).
P. (1995).- Calcareous algae and microcoprolites of the Albian platform carbonates of the Congo margin.- Bulletin du Centre de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 19, nº 2, p. 301-317.
W. (1959).- Foraminiferal key zones in the Lower Cretaceous of the western hemisphere and the Tethys province. In: A., ed., El Sistema Cretacico. Un symposium sobre el Cretacico en el hemisfero occidental y su correlacion mundial (Primer tomo).- Congreso Geologico International, XXa Sesión - Ciudad de México, 1956, p. 85-111.
F.A., J.-P. & M. (2010).- Precisiones sobre la sistemática y bioestratigrafía de Jerjesia chilensis (d') (Bivalvia; Hippuritoidea), del Cretácico Inferior de Chile. In: A. & S., eds., II Simposio - Paleontología en Chile. Concepcion, 13-15 diciembre 2010, Libro de Resúmenes, Departamento Ciencias de la Terra, Universidad de Concepción, p. 49.
A. d'
(1842).- Voyage
dans l'Amérique méridionale.- P. Bertrand, Paris, tome 3ème, 4ème partie :
Paléontologie, p. 188.
URL: https://archive.org/stream/voyagedanslamr3341844orbi#page/106/mode/2up
B. & R. (1987).- Présence d'une association micropaléontologique de la zone à Praeorbitolina cormyi (Aptien inférieur) dans l'Urgonien de Tanzanie méridionale. Implications paléogéographiques au niveau de la marge indienne de l'Afrique.- Géologie méditerranéenne, Marseille, t. XIV, nº 2, p. 143-151.
P. & A. (1982).- Les plates-formes carbonatées à rudistes (dites Urgoniennes) du Crétacé inférieur et leur environnement.- Cretaceous Research, vol. 3, p. 155-166.
R. & A. (1979).- 18. Upper BarremianLowermost Aptian Orbitolinid foraminifers from the Grand Banks Continental Rise, Northwestern Atlantic (DSDP Leg 43, Site 384). In: B.E., P.R. et al.- Initial Reports of the Deep Sea Drilling Project, Washington, vol. XLIII, p. 575-583.
J. & P. (1999).- Cenomanian oysters from the Sergipe Basin, Brazil.- Cretaceous Research, vol. 15, p. 745-765.
J. & P. (2002).- Palaeobiogeography of the upper Cenomanian-lower Turonian macroinvertebrates of the Sergipe Basin, northeastern Brazil. In: M., ed., Aspects of Cretaceous Stratigraphy and Palaeobiogeography.- Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissionen, Wien, Band 15, p. 151-168.
J.A., R.W. & J.-P. (1993).- Cretaceous carbonate platforms: an overview. In: J.A., R.W. & J.-P., eds., Cretaceous carbonate platforms.- American Association of Petroleum Geologists, Memoir 56, p. 15-24.
N.F. (1971).- North American Cretaceous biotic provinces delineated by gastropods. In: E.L., ed., Proceedings of the North American Paleontological Convention.- Allen Press, Lawrence, p. 1610-1637.
N.F. (1987).- Cretaceous gastropods: Contrasts between Tethys and the temperate provinces.- Journal of Paleontology, vol. 61, nº 6, p. 1085-1111.
T. (2002).- Plate tectonic control on the evolution of Cretaceous platform carbonate production.- Geology, vol. 30, nº 3, p. 259-262.
I.P. (1999).- Rudists and carbonate platform evolution: the Late Cretaceous Maiella carbonate platform margin, Abruzzi, Italy.- PhD, Diss. ETH, Zürich, nº 12494, 131 p. (+ Appendix); Memorie di Scienze geologiche, Padova, vol. 51, nº 2, p. 333-413.
G.J.S., J. & M. (2010).- Ocorrência de rudistas em amostras de testemunho do Albiano inferior da Bacia de Campos.- Boletim de Geociéncias da Petrobras, Rio de Janeiro, vol. 18, nº 2, p. 313-320.
W.J., B. & D. (2014).- Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), a 100 million year old fossil coralline red alga from north-eastern Brazil, and a new Hauterivian record of Sporolithon from Switzerland.- Carnets Geol., Madrid, vol. 14, nº 7, p. 139-158.
H. & S. (1926).- Geological age of Orbitolina-bearing rocks of Japan.- Science Reports of the Tohoku University, (Series 2, Geology), Sendai, vol. IX, nº 1, p. 13-20 (Pls. III-VI).