◄ Carnets Geol. 23 (7) ►
![]()
Outline:
[1. Introduction]
[2. Studied section]
[3. Material and methods]
[4. Lithostratigraphy]
[5. Biostratigraphy]
[6. Discussion]
[7. Conclusions]
and ...
[Bibliographic references]
Dépt. STU, Fac. Sci. Tech., UBO, 6 avenue Le Gorgeu, CS 93837, F-29238 Brest (France)
6D avenue Général de Gaulle, F-05200 Briançon (France)
Department of Geology, Moulay Ismail University, Meknes (Morocco)
Published online in final form (pdf) on June 27, 2023
DOI 10.2110/carnets.2023.2307
![]()
[Editor:
Michel Moullade; language editor: Phil Salvador; technical editor: Bruno Granier]
![]()
Our new study of the Tithonian and lower Berriasian succession of Le Chouet (Les Prčs, La Drôme, France) better characterizes the lithological succession, the macro- and microfacies, and the stratigraphic ranges of some microfossils mostly calibrated on the calpionellid biozonation. On the lithological side, the Tithonian strata are dominantly characterized by thick-bedded breccias representing debris flows and related calciturbidites whereas the Berriasian strata are typically white limestones that also comprises scattered intercalations of thin-bedded breccias and calciturbitides (including cryptic mud calciturbidites). In thin sections, these white limestones display mud- to wackestone textures and their allochems are mostly tiny bioclasts (e.g., radiolarians, calpionellids, saccocomids). Breccias are lithoclastic rudstones and/or floatstones with a matrix similar to the calciturbidites. Their lithoclasts are either extraclasts sensu stricto (i.e., material derived from updip shallow-water areas) or pseudointraclasts, representing reworked subautochthonous material (i.e., mud- and wackestone lithoclasts with radiolarians, saccocomids and/or calpionellids). In addition to the erosional features observed at the bases of the gravity flows, these pseudointraclasts document the intensity of submarine erosion. Locally they help to estimate the depths of erosion updip of the deposit. A number of bioclasts are reworked from updip shallow-water areas; among them, it is worth mentioning the foraminifer Protopeneroplis ultragranulata (Gorbatchik), the first occurrence of which is dated to late early Tithonian. Saccocomids are part of the dominating pelagic biota reported from the lower and lower upper Tithonian interval whereas calpionellids replace them in the uppermost Tithonian to lower Berriasian interval. Intervals with saccocomids characteristic of zones 4-5 and zones 6-7 are respectively ascribed here to the lower Tithonian (4-5) and pro parte to the upper Tithonian (6-7). The biozonation of the calpionellid group sensu lato allows identification of the Boneti Subzone of the chitinoidellids, the Crassicollaria Zone with its four subzones (A0-A3), and the Alpina Zone with its first subzone (B1). On the basis of biostratigraphical and sedimentological data (including the rates of sedimentation), most zonal boundaries are located at the erosional bases of breccia or turbidite layers and thus coincide with hiatuses.
• Tithonian;
• Berriasian;
•
calpionellids;
• saccocomids;
• erosion;
• debris flows;
•
turbidites;
• lithoclasts;
• Vocontian
Trough
Granier B.R.C., Ferry S. & Benzaggagh M. (2023).- Hiatuses and redeposits in the Tithonian-Berriasian transition at Le Chouet (Les Prčs, La Drôme, SE France): Sedimentological and biostratigraphical implications.- Carnets Geol., Madrid, vol. 23, no. 7, p. 123-147.
Lacunes et redéposition ŕ la transition Tithonien-Berriasien au Chouet (Les Prčs, Drôme, SE France) : Implications sédimentologiques et biostratigraphiques.- Notre nouvelle étude de la succession du Tithonien au Berriasien inférieur du Chouet (Les Prčs, Drôme, France) précise la succession lithologique, les macro- et microfacičs, et les répartitions stratigraphiques de quelques microfossiles essentiellement calibrées ici sur la biozonation des calpionellidés. D'un point de vue lithologique, les dépôts du Tithonien sont principalement représentés par des brčches en bancs épais représentant des coulées de débris et par des calciturbidites associées, tandis que les dépôts du Berriasien sont typiquement constitués de calcaires blancs qui recčlent également des intercalations éparses de passées bréchiques et de calciturbitides (y compris des calciturbidites de boue, souvent cryptiques). En lames minces, ces calcaires blancs présentent des textures mud- ŕ wackestone et leurs éléments figurés sont pour la plupart de minuscules bioclastes (comme, par exemple, des radiolaires, des calpionellidés, des saccocomidés). Les brčches sont des rud- et/ou floatstones lithoclastiques avec une matrice comparable aux calciturbidites. Leurs lithoclastes sont soit des extraclastes sensu stricto (c'est-ŕ-dire du matériel remanié de zones peu profondes en amont), soit des pseudo-intraclastes, représentant du matériel sub-autochtone remanié (c'est-ŕ-dire des lithoclastes ŕ textures mud- ŕ wackestone avec des radiolaires, des saccocomidés et/ou des calpionellidés). Outre les structures liées ŕ l'érosion observées ŕ la base des écoulements gravitaires, ces pseudo-intraclastes témoignent de l'intensité de l'érosion sous-marine. Localement, ils permettent d'estimer les profondeurs d'érosion en amont des coulées. Un nombre non négligeable de bioclastes sont remaniés de zones peu profondes en amont, parmi lesquels il convient de mentionner le foraminifčre Protopeneroplis ultragranulata (Gorbatchik), dont la premičre apparition est datée ici du Tithonien inférieur terminal. Les saccocomidés font partie des organismes pélagiques dominants signalés dans l'intervalle Tithonien inférieur ŕ supérieur, tandis que les calpionelles les remplacent dans l'intervalle Tithonien supérieur ŕ Berriasien inférieur. Les intervalles ŕ saccocomidés, caractéristiques des zones 4-5 ainsi que des zones 6-7, sont respectivement attribués ici au Tithonien inférieur (4-5) ainsi que pro parte au Tithonien supérieur (6-7). La biozonation du groupe des calpionelles sensu lato permet l'identification de la Sous-Zone ŕ Boneti des chitinoďdellidés, la Zone ŕ Crassicollaria avec ses quatre sous-zones (A0-A3) et la Zone ŕ Alpina avec sa premičre sous-zone (B1). Sur la base de données sédimentologiques (y compris les taux de sédimentation) et biostratigraphiques, la plupart des limites de zones sont localisées aux bases érosives de couches de brčches ou de turbidites et correspondent donc ŕ des lacunes sédimentaires.
• Tithonien ;
• Berriasien ;
•
calpionellidés ;
• saccocomidés ;
• érosion ;
• coulées de
débris ;
• turbidites ;
• lithoclastes ;
• Fosse vocontienne
In parallel with recent investigations on the former
Berriasian GSSP candidate at Tré Maroua, Le Saix, Hautes-Alpes (Granier et
al., 2020b, 2022, 2023), our group studied another key section of
the Vocontian Trough (SE France) for the Tithonian-Berriasian transition at Le
Chouet, Les Prčs, La Drôme (Ferry & Granier,
2018). This
second section (Figs. 1 ![]() - 2
- 2 ![]() ) was studied by Remane
(1970: Fig. 6) and by
the former Berriasian Working Group (Wimbledon et al.,
2013: Fig.
4, 2020a: Supplement Fig. S1). Its ammonite fauna was first studied by Le Hégarat
(1973) but it did not prove to be useful because the author's log (op.cit.:
Fig. 25) cannot be correlated with any of the other logs. The Le Chouet section
partly spans two lithostratigraphic units: 1) the uppermost part of the "brčches
tithoniennes" (Tithonian breccias) below and 2) the lower part of the
"calcaires blancs vocontiens" (Vocontian white limestones) above.
However, as shown in the logs (Figs. 2
) was studied by Remane
(1970: Fig. 6) and by
the former Berriasian Working Group (Wimbledon et al.,
2013: Fig.
4, 2020a: Supplement Fig. S1). Its ammonite fauna was first studied by Le Hégarat
(1973) but it did not prove to be useful because the author's log (op.cit.:
Fig. 25) cannot be correlated with any of the other logs. The Le Chouet section
partly spans two lithostratigraphic units: 1) the uppermost part of the "brčches
tithoniennes" (Tithonian breccias) below and 2) the lower part of the
"calcaires blancs vocontiens" (Vocontian white limestones) above.
However, as shown in the logs (Figs. 2 ![]() - 3
- 3 ![]() ), facies separation is no so clear cut:
lime mudstone layers sporadically occur between conglomeratic beds of the
Tithonian breccias whereas conglomeratic layers locally occur in the white
limestones. This pattern merely reflects the decreasing intensity of the
resedimentation over the Tithonian-Berriasian transition. Only the dominant type
of macrofacies justifies this subdivision.
), facies separation is no so clear cut:
lime mudstone layers sporadically occur between conglomeratic beds of the
Tithonian breccias whereas conglomeratic layers locally occur in the white
limestones. This pattern merely reflects the decreasing intensity of the
resedimentation over the Tithonian-Berriasian transition. Only the dominant type
of macrofacies justifies this subdivision.
Contrary to Remane (1970), Wimbledon et al. (2013, 2020a) have mostly neglected the sedimentological aspects of the Le Chouet section. More specifically they underestimated the gravity-flow erosion and the redeposition of the derived material that obviously impacted the apparent continuity of the sedimentary record. As will be demonstrated below, although that is not always visible at the scale of temporal resolution of the ammonite zones or that of the calpionellids, the logged section is hiatal. It was already the case for the former Berriasian GSSP candidate at Tré Maroua, Le Saix, Hautes-Alpes (Granier et al., 2020b, 2022, 2023).
|
|
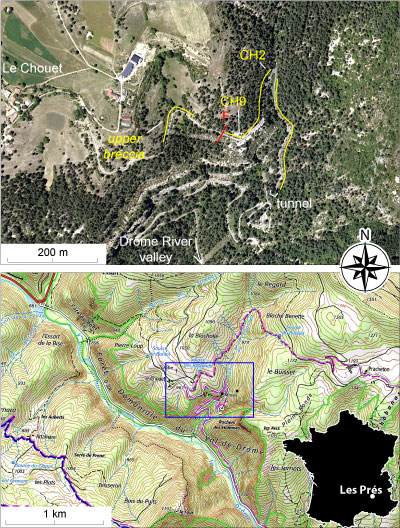
Figure 1: Topographic map of the Le Chouet area (A) and
location of the sections studied (B). |
|
|
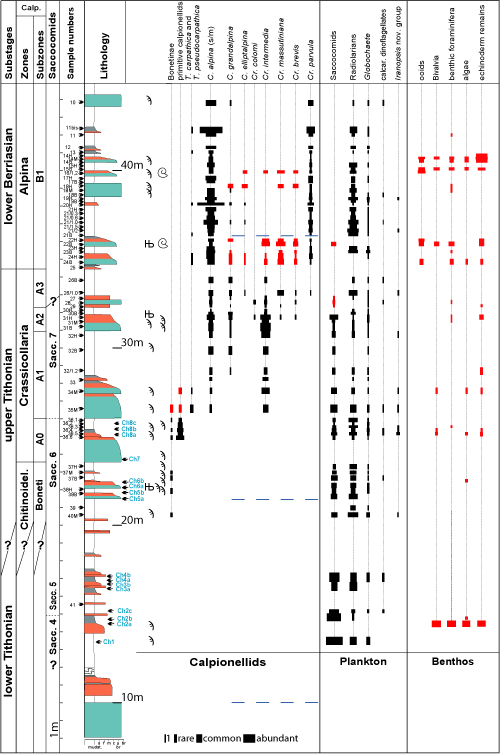
Figure 2: Le Chouet log displaying the lithological
succession, the stratigraphic distribution of the main (micro-) fossil groups of
the Tithonian-lower Berriasian, both the calpionellid and saccocomid biozones,
and the main reworked neritic elements. Autochthonous sedimentation (mud- and
wackestones) is in white; cryptoturbidites
(micropack- and micrograinstones)
in grey; coarse grained turbidites
(pack- and grainstones) in orange; debris flows (float- and rudstones) in
blue-green. The red color in the distribution columns corresponds to reworked
material. Њ: belemnite rostrum; ϡ: aptychus; grain size: s, silt; f, m, c, fine, medium,
coarse sands; µbr, microbreccia; br, breccia. |
|
|
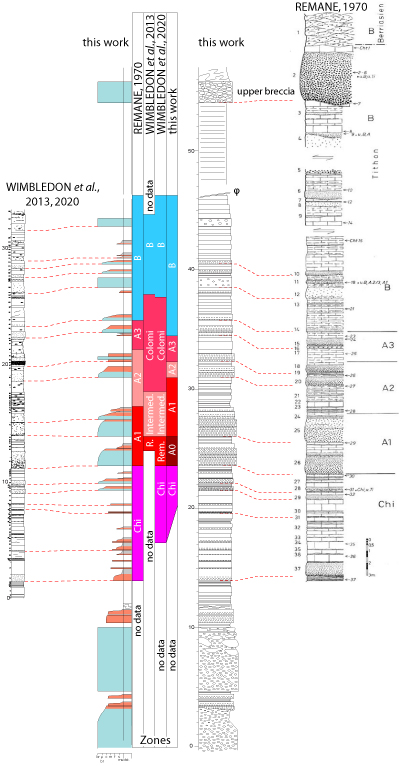
Figure 3: Lithostratigraphic correlations of Remane's
1970 log (right column), Wimbledon et al.'
2013 log (left column), and
this study (center; see caption in Fig. 2) with the various interpretations of
the calpionellid biozones. |
The Le Chouet section is located some tens of meters southeast
of the eponymic farm (Les Prčs municipality, La Drôme department, Fig. 1 ![]() ),
in an area located in the center of the geological map at 1/50,000 scale of
Luc-en-Diois (Flandrin, 1970). A composite section, more than 50 m long (Figs.
2
),
in an area located in the center of the geological map at 1/50,000 scale of
Luc-en-Diois (Flandrin, 1970). A composite section, more than 50 m long (Figs.
2 ![]() - 3
- 3 ![]() ),
was measured along the tarred track from the D306 departmental road to the Le
Chouet farm. It is split in three parts: 1) The lower part, which is sited on
the left side of the stream, starts after a short tunnel section (GPS
coordinates: 44°32'25.8"N 5°33'37.4"E).
It corresponds to the first 10 m of the logged section. 2) The median part
starts at the hairpin bend (GPS coordinates: 44°32'34.3"N 5°33'37.0"E)
where the track crosscuts the Fournet stream, one of the small tributaries of La
Drôme river. This median part, which is sited on the right side of the stream,
ends with a fault at 46 m height on the log (GPS coordinates: 44°32'29.7"N
5°33'32.5"E). The outcrop conditions after the fault become poor;
consequently, 3) the upper part of the section was not logged in detail. The
whole section (Figs. 2
),
was measured along the tarred track from the D306 departmental road to the Le
Chouet farm. It is split in three parts: 1) The lower part, which is sited on
the left side of the stream, starts after a short tunnel section (GPS
coordinates: 44°32'25.8"N 5°33'37.4"E).
It corresponds to the first 10 m of the logged section. 2) The median part
starts at the hairpin bend (GPS coordinates: 44°32'34.3"N 5°33'37.0"E)
where the track crosscuts the Fournet stream, one of the small tributaries of La
Drôme river. This median part, which is sited on the right side of the stream,
ends with a fault at 46 m height on the log (GPS coordinates: 44°32'29.7"N
5°33'32.5"E). The outcrop conditions after the fault become poor;
consequently, 3) the upper part of the section was not logged in detail. The
whole section (Figs. 2 ![]() - 3
- 3 ![]() )
was measured with a Jacob's staff and later correlated with those of Remane
(1970: Fig. 6) and Wimbledon et
al. (2013: Fig. 4, 2020a: Supplement Fig. S1): Figure
3
)
was measured with a Jacob's staff and later correlated with those of Remane
(1970: Fig. 6) and Wimbledon et
al. (2013: Fig. 4, 2020a: Supplement Fig. S1): Figure
3 ![]() .
An eight meter offset exists between the log on Figure 2
.
An eight meter offset exists between the log on Figure 2 ![]() and that on Figure 3
and that on Figure 3 ![]() ;
we shall refer to Figure 2
;
we shall refer to Figure 2 ![]() to give the elevations of the beds and samples on the logged section.
to give the elevations of the beds and samples on the logged section.
The Le Chouet section was sampled twice, the first 16 samples
with labels ranging from Ch1 to Ch8c (collected on the occasion of a first visit
on 2018/05/08 by the second author, S.F., see Fig. 2 ![]() )
and the second 64 samples with labels ranging from 41 to 10 (collected on the
occasion of a second joint visit on 2019/05/12 by the first author, B.R.C.G, and
the second author, S.F., see Fig. 2
)
and the second 64 samples with labels ranging from 41 to 10 (collected on the
occasion of a second joint visit on 2019/05/12 by the first author, B.R.C.G, and
the second author, S.F., see Fig. 2 ![]() ).
The rock samples were cut and the derived slabs scrupulously examined in the
search for mud turbidites, grainy turbidites and debris flows. In addition, sets
of thin sections were successively prepared to validate the identification of
mud turbidites, identify the source of the lithoclasts (parautochthonous or
allochthonous), and study the microfossil contents.
).
The rock samples were cut and the derived slabs scrupulously examined in the
search for mud turbidites, grainy turbidites and debris flows. In addition, sets
of thin sections were successively prepared to validate the identification of
mud turbidites, identify the source of the lithoclasts (parautochthonous or
allochthonous), and study the microfossil contents.
The lower part (up to 2 m on Fig. 2 ![]() )
is comprised of two massive breccia intervals of the uppermost Tithonian
breccia. In the next interval (up to 14 m on the log of Fig. 2
)
is comprised of two massive breccia intervals of the uppermost Tithonian
breccia. In the next interval (up to 14 m on the log of Fig. 2 ![]() )
the yellowish to light grey limestones that form the "background"
basinal sedimentation are irregularly alternating with thin beds of fine-grained
(Fig. 4H
)
the yellowish to light grey limestones that form the "background"
basinal sedimentation are irregularly alternating with thin beds of fine-grained
(Fig. 4H ![]() )
to mud (Fig. 4I
)
to mud (Fig. 4I ![]() )
turbidites. From 14 upward to 28 m on the same log, thick beds made of
coarse-grained graded turbidites (Figs. 4B pars, C, E pars, F-G
)
turbidites. From 14 upward to 28 m on the same log, thick beds made of
coarse-grained graded turbidites (Figs. 4B pars, C, E pars, F-G ![]() ,
5G-I
,
5G-I ![]() )
and breccias, often rudstones of pebbles and cobbles (Fig. 4B pars, D, E
pars
)
and breccias, often rudstones of pebbles and cobbles (Fig. 4B pars, D, E
pars ![]() ),
are common whereas the mud turbidites remain present (Figs. 4A
),
are common whereas the mud turbidites remain present (Figs. 4A ![]() ,
6L
,
6L ![]() ).
The facies of the next and last interval, up to the fault (at 38 m), are
characteristic of the "calcaires blancs" (white limestones) and
include some cryptic mud turbidites (Fig. 5B
).
The facies of the next and last interval, up to the fault (at 38 m), are
characteristic of the "calcaires blancs" (white limestones) and
include some cryptic mud turbidites (Fig. 5B ![]() ).
Additionally, this interval still displays graded turbidites (Fig.
5C-D, F pars
).
Additionally, this interval still displays graded turbidites (Fig.
5C-D, F pars ![]() )
and breccias, often floatstones of pebbles and cobbles (Fig.
5A, E
)
and breccias, often floatstones of pebbles and cobbles (Fig.
5A, E ![]() ).
).
|
|
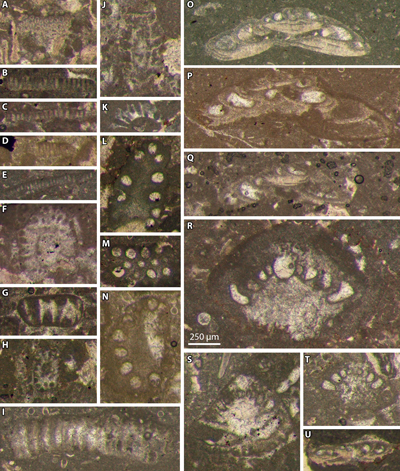
Figure 4:
Lower (I) and upper (A-H) Tithonian facies. A: pinkish cryptoturbidites 25; B:
breccia 28 sandwiched between turbidites 27 (above) and 29 (below); C: breccia
35 and its basal erosion surface; D: breccia Ch7 and its basal erosion surface;
E: breccia Ch7 (top) and graded turbidites Ch6 and Ch5 (bottom); F: enlargement
of E, detail of the basal erosion surface of the graded turbidite Ch6; G:
enlargement of E, detail of the graded turbidite Ch5; H: basal erosion surface
of the turbidite 40; I: thin cryptoturbidite Ch4 with a silty beige base (m)
indicating a faint grading. A, F-G, I: pencil for scale; B, D: Jacob's
staff for scale; C, E: no physical scale; H: finger for scale. |
|
|
Figure 5:
Berriasian facies. A: floatstone breccia 10; B: pinkish cryptoturbidites 12 (m);
C: graded turbidite 14; D: graded turbidite 16; E: enlargement of F, detail of
floatstone breccia 18; F: turbidite 20 (bottom) and floatstone breccia 18 (top);
G: graded turbidite 22; H: graded turbidite 24 (above) eroding fine-grained
turbidite 25 (below); I: enlargement of H, detail of the contact of turbidites
25 (below) and 25 (above). A-B, D, F, I: hammer for scale; C, E: pencil for
scale; H: no physical scale. |
|
|
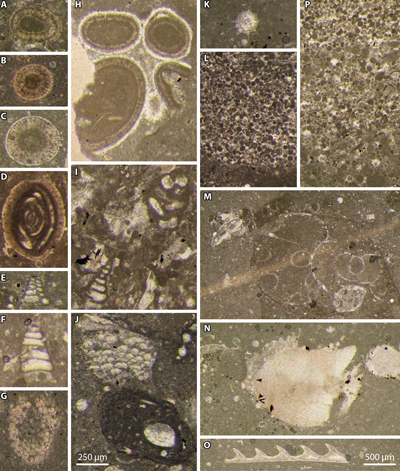
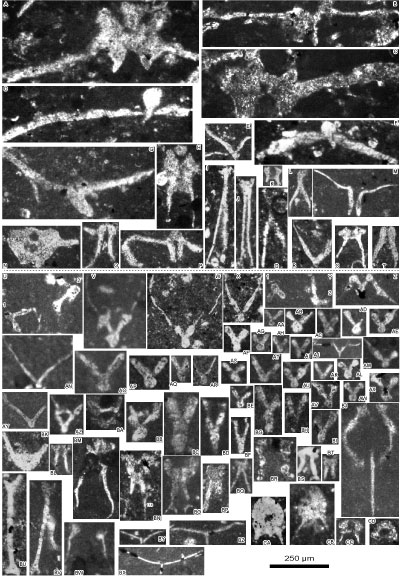
Figure 6: Reworked and in situ material from Le Chouet
section. A-D: concentric radial ooids (a miliolid as a nucleus in D); E-F:
biseriate foraminifers, Textulariidae; G: set of Globochaete
alpina Lombard on a lamella (tangential section); H:
extraclast of oolitic grainstone, reworked from a hardground as evidenced by its
early marine fibrous cementation; I: Redmondoides lugeoni (Septfontaine)
at the top and a biseriate foraminifer (Textulariidae) at the bottom; J: Koskinobullina
socialis Cherchi & Schroeder at the top and Tubiphytes
sp. at the bottom; K: incertae sedis; L: erosional surface at the bottom
of a micrograinstone ("mud turbitidite", cryptoturbidite) with
numerous calpionellid loricae in microlithoclasts; M: Mohlerina basiliensis
(Mohler) at the left top and ammonite phragmocone in
transverse section to the right; N: belemnite rostrum; O: aptychus; P: micrograinstone (fine-grained turbidite) with numerous
calpionellid loricae in microlithoclasts. Photomicrographs
A-K, N with the same scale (scale bar on J = 250 µm), photomicrographs L-M, O-Q
with the same scale (scale bar on O = 500 µm). A: sample 14B; B, H: sample 15B;
C: sample 34M; D: sample Ch8a; E, J: sample Ch6b; F, I: sample Ch8b; G: sample
31M; K: sample 40M; L: sample 30H; M: sample 16; N: sample 14M; Q: sample 22B;
O: sample 31H; P: sample 25. Upper Tithonian: C-G, I-L, O; lower Berriasian: A-B,
H, M-N, P. |
In terms of macro- and microfacies, the rocks naturally fall into two categories: 1) the autochthonous facies and 2) the parautochthonous- and allochthonous-derived facies.
The first category corresponds to background facies related
to the regular sedimentation of pelagic material falling and slowly accumulating
on the seafloor. They consist of mud- and wackestone microfacies with
saccocomids (which are here restricted to Tithonian strata), calpionellids (which
are here restricted to upper Tithonian - lower Berriasian strata), and/or
radiolarians (which may occur here in both Tithonian and lower Berriasian strata).
Besides the pelagic groups of microorganims, which also comprise calcareous
dinoflagellates (Fig. 7AH-AJ ![]() ), Globochaete alpina Lombard
(Figs. 6G
), Globochaete alpina Lombard
(Figs. 6G ![]() ,
7Y-AB
,
7Y-AB ![]() ) and the Iranopsis nov. group
(Fig. 7AC-AG, AK-AN
) and the Iranopsis nov. group
(Fig. 7AC-AG, AK-AN ![]() ), the thin
sections contain various parautochthonous and autochthonous bioclasts, either
smaller (e.g., sponge spicules) or larger ones [e.g., aptychii (Fig.
6O
), the thin
sections contain various parautochthonous and autochthonous bioclasts, either
smaller (e.g., sponge spicules) or larger ones [e.g., aptychii (Fig.
6O ![]() ), ammonite phragmocones
(Fig. 6M pars
), ammonite phragmocones
(Fig. 6M pars ![]() ), belemnite
rostra
(Fig. 6N
), belemnite
rostra
(Fig. 6N ![]() ) and jaws].
) and jaws].
|
|
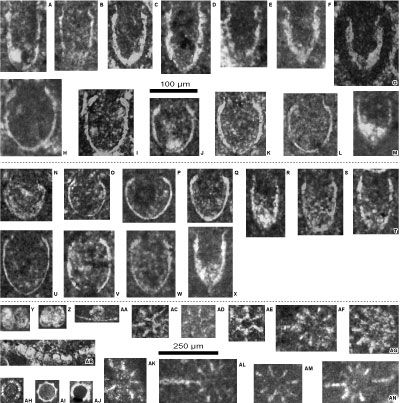
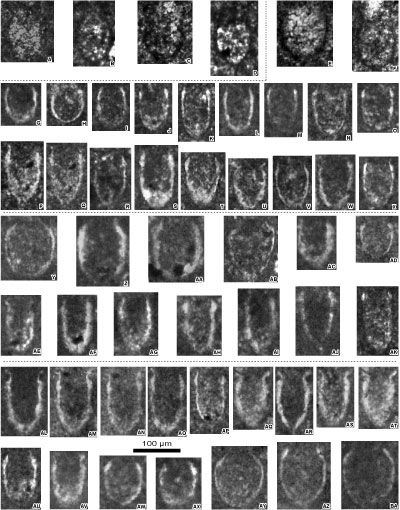
Figure 7: A-X: Calpionellid specimens from the
Crassicollaria Zone (A3 subzone) and the Alpina Zone (B1 subzone) - reworked and/or
in situ material - of the Le Chouet section. A-M: Brevis-Massutiniana
Subzone (A3 subzone), N-T: Alpina-Parvula Subzone (B1 subzone), U-X: specimens from the top of the
Crassicollaria Zone reworked in the Alpina-Parvula Subzone (B1 subzone). A-D: Crassicollaria massutiniana (Colom)
(reworked specimens); E-G: Cr.
brevis Remane (reworked specimens; G: specimen in
a microlithoclast); H: Calpionella
grandalpina Nagy (reworked specimen); I:
C. elliptalpina Nagy (reworked specimen); J:
small-sized C. alpina Lorenz (reworked specimen); K-L:
medium-sized C. alpina Lorenz (reworked specimens); M: Tintinnopsella
pseudocarpathica Benzaggagh et al. (reworked specimen); N-P: Calpionella
alpina Lorenz with ovoid rounded loricae; Q: C. alpina Lorenz
with ovoid elongated lorica; R-T: Crassicollaria
parvula Remane; U-W: Calpionella
elliptalpina Nagy (reworked specimens); X: Crassicollaria
intermedia Durand Delga (reworked specimen). Y-AN: Various
microfossils from Tithonian - lower Berriasian of the Le Chouet section. Y-AB: Globochaete
alpina Lombard:
Y-Z, as pairs; AA, isolated on a lamella; AB, set on a lamella
(transverse section);
AC-AG, AK-AN: Iranopsis nov. group;
AH: Colomisphaera carpathica (Borza);
AI: Parastomiosphaera tuberculata Benzaggagh et al.; AJ: Parastomiosphaera aff. malmica (Borza).
Photomicrographs A-Z with the same scale bar = 100
µm, photomicrographs Y-AN with the same scale bar = 250 µm. A-B, D, AA: sample 22B; C, E-I, K, AB: sample 24B;
J, L: sample 23H;
M: sample 22H;
N-O: sample 10;
P: sample 18M;
Q: sample 19B; R: sample 15H; S-T: sample 21B;
U: sample 18M (reworked specimen); V: sample 21/1.0; W: sample 21B (reworked specimen);
X: sample 16/1.2 (reworked specimen); Y,
AM-AN: sample 36.5; Z: sample 36.4; AC: sample 40M; AD, AI,
AL: sample 26/1.0; AE: sample 38B; AH: sample 18B; AF, AK: sample 19H; AG: sample
21/2.0; AJ: sample Ch2a. Lower
Tithonian: AJ; upper Tithonian: Y-Z, AC-AE, AI, AL-AN; lower Berriasian: A-X,
AA-AB, AF-AI, AK-AL. |
The second category corresponds to gravity-flow deposition
in the form of turbidites and associated debris flows. They consist of pack- and
grainstones that commonly form the matrices of lithoclastic float- and
rudstones. Basal surfaces of
both turbidites and debris flows are commonly
erosional (Fig. 8 ![]() ). Some lithoclasts have been created by the erosive processes
that have affected more or less deeply the underlying strata. Subsequently they
were incorporated into the turbidites and debris flows. These lithoclasts
are not intraclasts, but pseudointraclasts, because they do not result from the in
situ dismantling of the same and single layer but that of several discrete
layers located updip on the slope. Accordingly, a single breccia layer contain
several discrete types of pseudointraclasts (e.g., saccocomid wackestone
lithoclasts, calpionellid wackestone lithoclasts, radiolarian wackestone
lithoclasts, and even microlithoclastic grainstone lithoclasts). Granier et
al. (2020b) referred to them as "extraclasts" whereas Granier
et al. (2023) identified them as "pseudointraclasts"
because their fabrics are similar to that of the background relatively
deep-water sedimentation in contrast to the extraclasts sensu stricto
that correspond to lithoclasts reworked from updip shallow-water areas.
). Some lithoclasts have been created by the erosive processes
that have affected more or less deeply the underlying strata. Subsequently they
were incorporated into the turbidites and debris flows. These lithoclasts
are not intraclasts, but pseudointraclasts, because they do not result from the in
situ dismantling of the same and single layer but that of several discrete
layers located updip on the slope. Accordingly, a single breccia layer contain
several discrete types of pseudointraclasts (e.g., saccocomid wackestone
lithoclasts, calpionellid wackestone lithoclasts, radiolarian wackestone
lithoclasts, and even microlithoclastic grainstone lithoclasts). Granier et
al. (2020b) referred to them as "extraclasts" whereas Granier
et al. (2023) identified them as "pseudointraclasts"
because their fabrics are similar to that of the background relatively
deep-water sedimentation in contrast to the extraclasts sensu stricto
that correspond to lithoclasts reworked from updip shallow-water areas.
|
|
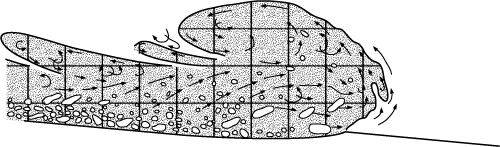
Figure 8: Avalanche model for the formation of debris flows
and related calciturbidites combining erosion and transportation modified from Postma
et al. (1988). |
|
|
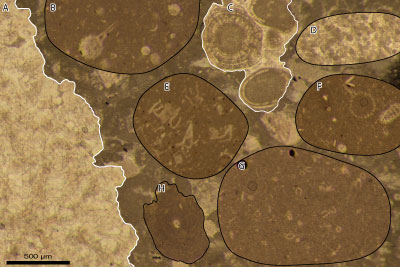
Figure 9: Composite* debris flow microfacies. A: extraclast
of hermatypic coral boundstone; B, F: extraclast of radiolarian wackestone; C:
extraclast of oolitic grainstone; D: extraclast of microclastic grainstone (turbidite)
with calpionellids; E: extraclast of saccocomid wackestone; G: extraclast of
calpionellid wackestone; H: Tubiphytes sp. The matrix is a bioclastic and
extraclastic (pack-) grainstone. The lithoclasts B and D-G, which should be
derived from slope and/or basinal facies, are also referred to as
pseudointraclasts. *: The background as well as clasts B and D-F are patches
copied from another source and pasted here to illustrate a perfect example of
calcareous debris flow (if all types of clasts may well occur in the same thin
section, they hardly occur in the same field of view). The background is derived
from thin section Ch8b-2 whereas the fake clast B is derived from 37H, D from
11, E from Ch1, and F from 34M. Genuine clast A is observed in thin section Ch6
whereas C is found in 14B, G in 18H, and H in Ch8b-2. All photomicrographs with
the same bar = 500 µm. |
The
pseudointraclasts are usually larger and subrounded (Fig. 9B,
D-G ![]() ) because they
were made of mechanically abraded unlithified material whereas the extraclasts sensu
stricto are commonly smaller and subangular (Figs. 6H
) because they
were made of mechanically abraded unlithified material whereas the extraclasts sensu
stricto are commonly smaller and subangular (Figs. 6H ![]() ,
9A, C, H
,
9A, C, H ![]() ).
Extraclasts sensu stricto but also skeletal grains (bioclasts) and
concentric calcitic ooids (Fig. 6A-D, N pars
).
Extraclasts sensu stricto but also skeletal grains (bioclasts) and
concentric calcitic ooids (Fig. 6A-D, N pars ![]() ) are all reworked from
shallow-water environments of a neighbouring platform edge. For instance, these
extraclasts may consists of cemented oolitic grainstones (Fig.
9C
) are all reworked from
shallow-water environments of a neighbouring platform edge. For instance, these
extraclasts may consists of cemented oolitic grainstones (Fig.
9C ![]() ) or of
microbial boundstones. Bioclasts are diverse comprising bryozoans,
calcareous and siliceous sponges, hermatypic corals (Fig. 9A
) or of
microbial boundstones. Bioclasts are diverse comprising bryozoans,
calcareous and siliceous sponges, hermatypic corals (Fig. 9A ![]() ), echinoderm
remains, calcareous algae [among which Salpingoporella pygmaea (Gümbel)
(Fig. 10F
), echinoderm
remains, calcareous algae [among which Salpingoporella pygmaea (Gümbel)
(Fig. 10F ![]() ),
S. annulata Carozzi (Fig.
6N pars
),
S. annulata Carozzi (Fig.
6N pars ![]() )
and Thaumatoporella parvovesiculifera (Raineri) (Fig.
11A-E
)
and Thaumatoporella parvovesiculifera (Raineri) (Fig.
11A-E ![]() ), as
well as the problematic Iberopora bodeuri Granier & Berthou
(Fig. 10S-U
), as
well as the problematic Iberopora bodeuri Granier & Berthou
(Fig. 10S-U ![]() )], and foraminifers. The benthic foraminifers are the most diverse:
ataxophragmiids (Fig. 12AH-AJ
)], and foraminifers. The benthic foraminifers are the most diverse:
ataxophragmiids (Fig. 12AH-AJ ![]() ), Redmondoides lugeoni (Septfontaine)
(Fig. 6I pars
), Redmondoides lugeoni (Septfontaine)
(Fig. 6I pars ![]() ), textulariids
(Fig. 6E-F, I pars
), textulariids
(Fig. 6E-F, I pars ![]() ), lituolids
(Fig. 10N-R, V
), lituolids
(Fig. 10N-R, V ![]() ), miliolids (Fig. 6D
), miliolids (Fig. 6D ![]() ),
lenticulinids, Frentzenella sp. (Fig. 11R-T
),
lenticulinids, Frentzenella sp. (Fig. 11R-T ![]() ), Coscinoconus sp.
(Fig. 10A-F
), Coscinoconus sp.
(Fig. 10A-F ![]() ), Ichnusella spp.
(Fig. 12A-AB
), Ichnusella spp.
(Fig. 12A-AB ![]() ), Mohlerina basiliensis (Mohler) (Figs.
6M pars
), Mohlerina basiliensis (Mohler) (Figs.
6M pars ![]() ,
10J-M
,
10J-M ![]() ,
11O-Q, U
,
11O-Q, U ![]() ), Protopeneroplis ultragranulata (Gorbatchik) (Fig.
12AC-AF, ?AG
), Protopeneroplis ultragranulata (Gorbatchik) (Fig.
12AC-AF, ?AG ![]() ), as well as the incertae sedis Koskinobullina socialis Cherchi
& Schroeder (Fig. 6J pars
), as well as the incertae sedis Koskinobullina socialis Cherchi
& Schroeder (Fig. 6J pars ![]() ) and Tubiphytes sp.
(Figs. 6J pars
) and Tubiphytes sp.
(Figs. 6J pars ![]() ,
9H
,
9H ![]() ).
).
|
|
Figure 10: Reworked benthic foraminifers and calcareous algae
from Le Chouet section. A-F: Coscinoconus sp.; G-I: ? Ataxophragmiidae;
J-M: Mohlerina basiliensis (Mohler); N-O: cf. Anchispirocyclina
lusitanica (Egger); P: sea urchin radiole (bottom right) and subepidermal
meshwork of a large agglutinated foraminiferal test; Q-R: Nautiloculina
sp.; S-U: Iberoporella bodeuri Granier & Berthou,
with Calpionella alpina in U; V:
large agglutinated foraminiferal test. All photomicrographs with the same scale
(scale bar on R = 250 µm). A-B, L, S: sample 14B; C: sample 18H; D, I-K, M, U:
sample Ch8b; E: sample 22M; F: sample 14M; G: sample 15B; H: sample Ch2a; N-O:
sample Ch6b; P-R: sample Ch8a; T: sample 24B; V: sample 16. Lower Tithonian: H;
upper Tithonian: D, I-K, M-R, U; lower Berriasian: A-C, E-G, F, S-T, V. |
|
|
Figure 11: Reworked calcareous algae, sponges, and benthic
foraminifers from Le Chouet (A-M, O-U) and Tré Maroua (N) sections. A-E: Thaumatoporella
parvovesiculifera (Raineri); F-H, ?I: Salpingoporella sp. [F: S.pygmaea
(Gümbel)]; J-K: ? Clypeina sp.; L-N: Perturbatacrusta leini
Schlagintweit & Gawlick, 2011; O-Q, U: Mohlerina basiliensis
(Mohler); R-T: Ichnusella sp.. All photomicrographs with the same
scale (scale bar on R = 250 µm). A: sample Ch2b; B, H, J, L: sample Ch6b; C-D,
G: sample Ch8b; E, T: sample 14B; F: sample 14M; I: sample 15B; K, R: sample
Ch6a; M, S: sample Ch8a; N: Tré Maroua 58; O: sample 31M; P: sample 16; Q:
sample 22M; U: sample 22b. Lower Tithonian: A; upper Tithonian: B-D, G-H, J-M,
O, R-S; lower Berriasian: E-F, I, N, P-Q, T-U. |
|
|
Figure 12: Reworked benthic foraminifers from Le Chouet
section. A-AB: Ichnusella spp. including Ichn. infragranulata (Noth);
AC-AF, ?AG: Protopeneroplis ultragranulata (Gorbatchik); AH-AJ:
triseriate foraminifers, Ataxophragmiidae. All photomicrographs with the same
scale (scale bar on AG = 250 µm). A, I: sample 14B; B, U, AA: sample Ch8a; C-D,
H, S-T: sample 14M; E: sample 15B; F: sample 22B; G, AC: sample Ch2b; K-Q, V,
X-Y, AB, AD-AE, AH-AJ: sample Ch8b; R, W, Z: sample Ch6b; AF-AG: sample 24B.
Lower Tithonian: G, AC; upper Tithonian: B, K-R, U-AB, AD-AE, AH-AJ; lower
Berriasian: A, C-F, H-I, S-T, AF-AG. |
As in the Tré Maroua section (Granier et al.,
2020b, 2023), the Le Chouet section contains a number of
cryptoturbidites, i.e., well-sorted micrograinstones with a very finely
lithoclastic composition. These mud turbidites are hardly detectable with hand
lenses in the field (Granier et al., 2020b, Fig. 5). Their nature
is fully revealed only with thin sections under a standard microscope (Fig.
6L, P ![]() ). Wimbledon et al.
(2013, Fig. 8.2-4, 8.6) have identified
similar microfacies as pelbiomicrosparites or pelbiosparites. As a matter of
fact, most allochems that are less than 100 µm in diameter are not bioclasts or
peloids but mostly small rounded pseudointraclasts (microlithoclasts, Fig.
6L, P
). Wimbledon et al.
(2013, Fig. 8.2-4, 8.6) have identified
similar microfacies as pelbiomicrosparites or pelbiosparites. As a matter of
fact, most allochems that are less than 100 µm in diameter are not bioclasts or
peloids but mostly small rounded pseudointraclasts (microlithoclasts, Fig.
6L, P ![]() ), commonly consisting of a calpionellid lorica filled and coated by micrite.
), commonly consisting of a calpionellid lorica filled and coated by micrite.
5.1. Saccocomid biozones: For practical reasons, saccocomid sections have been named after their geometric shapes (Benzaggagh et al., 2015) as follows: 2Ax.act.br: biaxis with acute branches; 2Ax.brd.br: biaxis with broad branches; 2prl.Br: two parallel branches; 2tn.prl.Br: two thin parallel branches; 3Ax.act.br: triaxis with acute branches; 3Ax.brd.br: triaxis with broad branches; brd.2Ax: broad biaxis; brd.ml.Tt: broad molar tooth thin; cnc.ml.Tt: concave molar tooth; cvx.ml.Tt: convex molar tooth; div.elg.Br: divided elongated branch; elg.Br: elongated branch; elg.ml.Tt: elongated molar tooth; elg.psd-hxg.Hd: elongated pseudo-hexagonal head; elg.Tt: elongated tooth; flt.psd-hxg.Hd: flattened pseudo-hexagonal head; irg.Hd: irregular head; prp.Wg: propeller wings; srd.Wg: serrated wings; tn.Wg: thin wings; /2lat.apd:with two lateral appendices; /ax.tp: with axial tip; /elg.ax.tp: with elongated axial tip; /elg.tp: with elongated tips; /int.cvt: with internal cavity; /ov.ax.tp: with ovoid axial tip; /psd-rtg.ax.tp: with pseudo-rectangular axial tip; /ptd.bs: with pointed basis; /shr.ax.tp: with short axial tip; /smpl.tp: with simple tips; /tk.ts: with thick test; /tn.crw: with thin crown; /tn.flt.crw: with thin and flat crown; /trg.cvt: with triangular cavity.
This microfossil group largely dominates the pelagic
assemblages of the Tethys realm at least from the earliest Tithonian (Hybonotum
Zone) up to its disappearance in the latest Tithonian. However, it has been
largely overlooked except in the outer Rif of Morocco.
Five saccocomid zones (zones 3-7) have been defined for the Tithonian stage (Benzaggagh
et al., 2015: Fig. 17). Four of them, i.e., the saccocomid zones 4
to 7, are identified in the Le Chouet section. The assemblages and successions
at Le Chouet are very similar to those known in the outer Rif with a large
dominance of thick sections in the lowermost Tithonian (Saccocomid zone 4), with
highly diversified smaller sections in both the upper lower Tithonian (Saccocomid
zone 5) and the lower upper Tithonian (Saccocomid zone 6), and finally with less
frequent and undiversified sections dominated by biaxial sections in the middle
upper Tithonian (Saccocomid zone 7). In the Le Chouet section, samples Ch1-Ch2b contain numerous
saccocomid sections (Fig. 13A-CD ![]() ) dominated by large and thick forms
(Fig. 13A-D, F-G
) dominated by large and thick forms
(Fig. 13A-D, F-G ![]() ). The latter become scarcer in samples Ch3a-Ch4b with an
assemblage dominated by sections typical of the upper lower Tithonian (Fig.
13E, H-T
). The latter become scarcer in samples Ch3a-Ch4b with an
assemblage dominated by sections typical of the upper lower Tithonian (Fig.
13E, H-T ![]() ). In the overlying samples (40M to
27), saccocomid sections become less
and less abundant and are dominated by smaller forms (Fig. 13U-CD
). In the overlying samples (40M to
27), saccocomid sections become less
and less abundant and are dominated by smaller forms (Fig. 13U-CD ![]() ). The group
disappears at the base of the Crassicollaria subzone A3 above sample 27. Note
that samples 29-27 were picked in a turbiditic interval and these specimens are
possibly reworked. Sample 22B picked in the second debris flow level above the
base of the calpionellid zone B (i.e., the former Tithonian/Berriasian
boundary) also contains some reworked specimens.
). The group
disappears at the base of the Crassicollaria subzone A3 above sample 27. Note
that samples 29-27 were picked in a turbiditic interval and these specimens are
possibly reworked. Sample 22B picked in the second debris flow level above the
base of the calpionellid zone B (i.e., the former Tithonian/Berriasian
boundary) also contains some reworked specimens.
5.1.1. Saccocomid zone 4 (equivalent to the Darwini
and Semiforme zones, above the Hybonotum Zone) is characterized by an abundance
of thick sections of the srd.Wg/tk.ts, irg.Hd, and flt.psd-hxg.Hd/2lat.apd
types. At the base of the Le Chouet section (Ch1 and Ch2a-b), the typical
saccocomid sections of the zone 3 are missing and these levels are ascribed to
the saccocomid zone 4. The assemblage is dominated by large and thick sections,
e.g., irg.Hd (Fig. 13A-C, H ![]() ), th.Wg/ov.ax.tp
(Fig. 13D, F
), th.Wg/ov.ax.tp
(Fig. 13D, F ![]() ), and th.Wg/shr.ax.tp
(Fig. 13G
), and th.Wg/shr.ax.tp
(Fig. 13G ![]() ). It also comprises sections of the 2Ax.act.br/ptd.bs
(Fig. 13E, R
). It also comprises sections of the 2Ax.act.br/ptd.bs
(Fig. 13E, R ![]() ), flt.psd-hxg.Hd/2lat.apd
(Fig. 13N
), flt.psd-hxg.Hd/2lat.apd
(Fig. 13N ![]() ), 2prl.Br/elg.tp
(Fig. 13M
), 2prl.Br/elg.tp
(Fig. 13M ![]() ), div.elg.Br
(Fig. 13I-J, LF
), div.elg.Br
(Fig. 13I-J, LF ![]() ), elg.Tt
(Fig. 13T
), elg.Tt
(Fig. 13T ![]() ), and cvx.ml.Tt/lrg.crw
(Fig. 13P
), and cvx.ml.Tt/lrg.crw
(Fig. 13P ![]() ) types.
) types.
5.1.2. In
contrast to the previous zone, Saccocomid zone 5 (equivalent to the
Fallauxi Zone) is characterized by the scarcity of thick sections and by the
relative abundance of far less thick sections of the elg.psd-hxg, flt.psd-hxg.Hd,
elg.Br, 2prl.Br/smpl.tp, 3Ax.brd.br, tn.Wg, elg.Tt, ml.Tt, and 2Ax.brd.br/elg.ax.tp
types. In levels Ch2a to Ch4b, the saccocomid assemblage is dominated by
sections of the div.elg.Br (Fig. 13Q ![]() ), elg.Tt
(Fig. 13K, S
), elg.Tt
(Fig. 13K, S ![]() ), and cvx.ml.Tt /lrg.crw
(Fig. 13O
), and cvx.ml.Tt /lrg.crw
(Fig. 13O ![]() ) types.
) types.
5.1.3. Saccocomid zone 6 (equivalent to the Ponti
Zone and, pro parte, the Microcanthum Zone) is rich in sections of tn.Wg,
prp.Wg, elg.psd-hxg, 2prl.Br, 2tn.prl.Br, 3Ax.brd.br, 2Ax.brd.br/trg.cvt/ax.tp,
elg.Br, 2Ax.brd.br, and elg.Tt types, as well as the various sections of the
ml.Tt type. In the Le Chouet section, the assemblage of samples 40M-36.1 is
composed of sections of tn.Wg/shr.ax.tp (Fig. 13BX, BZ ![]() ), flt.psd-hxg.Hd/2lat.apd (Fig. 13CA-CB
), flt.psd-hxg.Hd/2lat.apd (Fig. 13CA-CB ![]() ),
elg.psd-hxg.Hd/2lat.apd (Fig. 13CC
),
elg.psd-hxg.Hd/2lat.apd (Fig. 13CC ![]() ), 2tn.prl.Br
(Fig. 13BW
), 2tn.prl.Br
(Fig. 13BW ![]() ),
brd.2Ax/trg.cvt (Fig. 13BK
),
brd.2Ax/trg.cvt (Fig. 13BK ![]() ),
2Ax.brd.br/trg.cvt/ax.tp (Fig. 13BB
),
2Ax.brd.br/trg.cvt/ax.tp (Fig. 13BB ![]() ),
2prl.Br/elg.tp (Fig. 13BY
),
2prl.Br/elg.tp (Fig. 13BY ![]() ), 3Ax.brd.br
(Fig. 13BD, BF-BG
), 3Ax.brd.br
(Fig. 13BD, BF-BG ![]() ), and
2Ax.act.br/ptd.bs (Fig. 13AN
), and
2Ax.act.br/ptd.bs (Fig. 13AN ![]() )
types, as well as frequent sections of 2Ax.brd.br/rd.ax.tp (Fig.
13U1, X-Y1, W, AB, AD-AE, AL, AP, AU, AW
)
types, as well as frequent sections of 2Ax.brd.br/rd.ax.tp (Fig.
13U1, X-Y1, W, AB, AD-AE, AL, AP, AU, AW ![]() ),
2Ax.brd.br/psd-rtg.ax.tp (Fig. 13U2, Y2, AO, AR-AS, AV, AX-AY
),
2Ax.brd.br/psd-rtg.ax.tp (Fig. 13U2, Y2, AO, AR-AS, AV, AX-AY
![]() ),
2Ax.brd.br/shr.ax.tp (Fig. 13AI, AK, AT
),
2Ax.brd.br/shr.ax.tp (Fig. 13AI, AK, AT ![]() ),
2Ax.brd.br/elg.ax.tp (Fig. 13AF
),
2Ax.brd.br/elg.ax.tp (Fig. 13AF ![]() ), cnc.ml.Tt
(Fig. 13BN-BQ, BT
), cnc.ml.Tt
(Fig. 13BN-BQ, BT ![]() ),
cvx.ml.Tt/tn.crw/int.cvt (Fig. 13BS
),
cvx.ml.Tt/tn.crw/int.cvt (Fig. 13BS ![]() ),
elg.ml.Tt/tn.flt.crw (Fig. 13BL
),
elg.ml.Tt/tn.flt.crw (Fig. 13BL ![]() ), and elg.Br
(Fig. 13BU
), and elg.Br
(Fig. 13BU ![]() ) types.
) types.
5.1.3.
Saccocomid zone 7 (equivalent
to the upper part of the Microcanthum Zone
and the lower part of the Durangites Zone)
contains fewer saccocomid sections. Its assemblage is largely dominated by
sections of 2Ax.brd.br type, with a rounded (Fig. 13AA, AC,
AG-AH, AM, BE ![]() ),
elongated (Fig. 13BI
),
elongated (Fig. 13BI ![]() ) or
pseudo-rectangular (Fig. 13V, Z, AQ
) or
pseudo-rectangular (Fig. 13V, Z, AQ ![]() )
axial apex, and it contains rare sections of 3Ax.brd.br, elg.Br, 2prl.Br/smpl.tp,
tn.Wg, 2Ax.brd.br/trg.cvt/ax.tp (Fig. 13AZ, BA
)
axial apex, and it contains rare sections of 3Ax.brd.br, elg.Br, 2prl.Br/smpl.tp,
tn.Wg, 2Ax.brd.br/trg.cvt/ax.tp (Fig. 13AZ, BA ![]() ),
elg.Tt, and ml.Tt types. At Le Chouet, it also
contains common 2Ax.brd.br/trg.cvt/ax.tp
type and rare sections of cnc.ml.Tt (Fig. 13BM
),
elg.Tt, and ml.Tt types. At Le Chouet, it also
contains common 2Ax.brd.br/trg.cvt/ax.tp
type and rare sections of cnc.ml.Tt (Fig. 13BM ![]() ),
brd.ml.Tt/tn.flt.crw (Fig. 13BR
),
brd.ml.Tt/tn.flt.crw (Fig. 13BR ![]() ),
flt.psd-hxg.Hd/2lat.apd (Fig. 13CD
),
flt.psd-hxg.Hd/2lat.apd (Fig. 13CD ![]() ), 3Ax.brd.br
(Fig. 13BC, BH
), 3Ax.brd.br
(Fig. 13BC, BH ![]() ), and div.elg.Br
(Fig. 13BV
), and div.elg.Br
(Fig. 13BV ![]() ) types.
) types.
|
|
Figure 13: Sections of skeletal segments of Tithonian
saccocomids from the saccocomid zones 4 to 7 in the Le Chouet section. A-C,
H: irg.Hd; D, F: th.Wg/ov.ax.tp;
E, R, AN: 2Ax.act.Br/ptd.bs;
G: th.Wg/shr.ax.tp;
I-J, L, Q: div.elg.Br;
K, S-T: elg.Tt; M:
2prl.Br/elg.tp; N: flt.psd-hxg.Hd/2lat.apd;
O-P: Dn.ml.cvx/crn.lrg; U1,
W, X, Y1, AA, AB, AC, AD, AE, AG, AH, AL, AM, AP, AU, AW, BE: 2Ax.brd.br/rd.ax.tp;
U2, V, Y2, Z, AO, AQ, AS, AV, AX, AY: 2Ax.brd.br/psd-rtg.ax.tp; AF,
AR, BI: 2Ax.brd.br/elg.ax.tp; AI,
AK, AT: 2Ax.brd.br/shr.ax.tp; AJ: 2Ax.act.br/ptd.bs/2lat.apd; AZ,
BA, BB: 2Ax.brd.br/trg.cvt/ax.tp; BC, BD, BF, BG, BH: 3Ax.brd.br; BK:
brd.2Ax/trg.cvt; BL: elg.ml.Tt/tn.flt.crw; BJ: new unnamed morphotype; BM-BQ,
BT: cnc.ml.Tt; BR:
brd.ml.Tt/tn.flt.crw; BS: cvx.ml.Tt/tn.crw/int.cvt; BU:
elg.Br; BV: div.elg.Br; BW:
div.elg.Br; BX, BZ: tn.Wg/shr.ax.tp; BY: 2prl.Br/elg.tp; CA-CB:
elg.psd-hxg.Hd/2lat.apd; CC-CD: flt.psd-hxg.Hd/2lat.apd. All photomicrographs with the same scale bar = 250 µm. A-C: Ch2b; D-J,
L-N, P, R, T: Ch1; K, O, Q, S: Ch4a; U, Y, AL, BD: 36.1; V, BC, BL-BM, BR, BV: 31H; W-X, AP, BE, BK: 38H; Z,
AR, AZ, BG, BT, BW: 35M; AA, AM, AQ, BA, BI, BN, BO, CB: 34M; AB, AK, AV, AX, BX:
36.3; AC: 32B; AD: 36.2; AE-AF, BF: 37B; AG, AH, BH: 31B; AI, AO, AS, BP-BQ, BZ,
BY, CC: 36.4; AJ: Ch8; AN, AU, BB: 37H; AT, AW: 36.5; AY, BS, CA: 38B; BE:
32/1.2; BJ: 36.6; BU: 40M; CD: 32H. Lower Tithonian: A-C, K, O, Q, S;
upper Tithonian: D-J, L-N, P, R, T-CD. Lower Tithonian: A-T; upper Tithonian:
U-CD. |
5.2. Chitinoidella Zone (Boneti Subzone): Contrary
to the Dobeni Subzone that to date has never been identified in any of the
Vocontian Trough sections, the Boneti Subzone is characterized here by the
occurrence of species belonging exclusively to the subfamily Bonetinae (Fig.
14A-F ![]() ), the assemblage of which is similar to those known from both
margins of the Tethys realm (i.e., eastern and western Europe, Turkey,
Iran, Cuba, Mexico, and recently reported from the Blue Nile Basin in Ethiopia, Jain
et al., 2021) and outside it (e.g., in the Neuquén basin of the
eastern Pacific margin of Argentina, Kietzmann, 2017; Kietzmann et
al., 2021). In the Ardescian series, Cecca et al.
(1989) have
already reported the occurrences of "Chitinoidella sp.", "Ch.
boneti Doben", and "Ch. cf. cubensis (Furrazola-Bermúdez)"
whereas, in the Le Chouet section, samples 40M-37M contain Bonetilla boneti
(Doben) (Fig. 14A
), the assemblage of which is similar to those known from both
margins of the Tethys realm (i.e., eastern and western Europe, Turkey,
Iran, Cuba, Mexico, and recently reported from the Blue Nile Basin in Ethiopia, Jain
et al., 2021) and outside it (e.g., in the Neuquén basin of the
eastern Pacific margin of Argentina, Kietzmann, 2017; Kietzmann et
al., 2021). In the Ardescian series, Cecca et al.
(1989) have
already reported the occurrences of "Chitinoidella sp.", "Ch.
boneti Doben", and "Ch. cf. cubensis (Furrazola-Bermúdez)"
whereas, in the Le Chouet section, samples 40M-37M contain Bonetilla boneti
(Doben) (Fig. 14A ![]() ), Furrazolaia insueta (Řehánek)
(Fig. 14B
), Furrazolaia insueta (Řehánek)
(Fig. 14B ![]() ), and F. cristobalensis (Furrazola-Bermúdez)
(Fig. 14C-D
), and F. cristobalensis (Furrazola-Bermúdez)
(Fig. 14C-D ![]() ). Due to a discontinuous sampling from Ch4b to 40M, i.e., a
nearly 5 meter interval, the base of the Boneti Subzone could not be accurately
defined.
). Due to a discontinuous sampling from Ch4b to 40M, i.e., a
nearly 5 meter interval, the base of the Boneti Subzone could not be accurately
defined.
5.3. Calpionellid zones (Benzaggagh, 2020):
Close-spaced sampling of the Le Chouet section allowed characterization of
the four subzones of the Crassicollaria Zone and the first subzone of the Alpina
Zone. The first specimens of calpionellids with microgranular loricae are reported from sample 40M
whereas the first specimens of calpionellids with hyaline tests appear in
sample 36.6. The latters are represented by primitive, often small loricae with
undeveloped or poorly developed collars (Fig. 14G-X ![]() ). They are initially sparse
to moderately abundant but they become abundant and dominate the pelagic
microorganisms from the upper half of the upper Tithonian Crassicollaria Zone
and throughout the lower Berriasian Alpina Zone.
). They are initially sparse
to moderately abundant but they become abundant and dominate the pelagic
microorganisms from the upper half of the upper Tithonian Crassicollaria Zone
and throughout the lower Berriasian Alpina Zone.
5.3.1. Crassicollaria Zone (zone A): In the Vocontian Basin, as in most basins of the Tethys margins, the Crassicollaria Zone is dominated by the genus Crassicollaria. In the Le Chouet section, it spans the interval ranging from sample 36.6 (and probably Ch7) to sample 26B. Its base is marked by an interval containing the last chitinoidellids and the first primitive calpionellids (subzone A0), equivalent to the Remanei and Praetintinnopsella zones of some authors (e.g., Remane et al., 1986; Ölveczká & Reháková, 2022). The rest of the zone is subdivided into three successive assemblage subzones with from bottom to top: 1) one with numerous small to medium-sized Crassicollaria intermedia and some Calpionella grandalpina (subzone A1), 2) another with numerous regular Crassicollaria intermedia, larger in size than those of the underlying subzone, and with few Cr. colomi (subzone A2), and 3) a last one with numerous Cr. brevis and Cr. massutiniana but also with few Calpionella elliptalpina (subzone A3).
5.3.1.1. The Chitinoidellid-primitive calpionellid Subzone (subzone A0) spans
at least the sampling
interval comprised between 36.6 and 36.1. It probably extends downward to the bottom of the breccia Ch7. It is
characterized by the assemblage of the last chitinoidellids and the first
calpionellids with hyaline tests, mostly primitive forms, with or without small
collars. The chitinoidellids comprise representatives of the subfamily Bonetinae,
among which Bonetilla boneti (Doben) and B. sphaerica Benzaggagh.
Calpionellids are represented by primitive forms dominated by Crassicollaria
aff. intermedia Durand Delga, small to medium in size (Fig.
14J-K, M-O, R ![]() ), and also comprise Calpionella
aff. alpina Lorenz,
small in size, with ovoid rounded (Fig. 14G
), and also comprise Calpionella
aff. alpina Lorenz,
small in size, with ovoid rounded (Fig. 14G ![]() ) or ovoid elongated
(Fig. 14I
) or ovoid elongated
(Fig. 14I ![]() )
loricae, and Tintinnopsella aff. carpathica (Murgeanu &
Filipescu) (Fig. 14V-X
)
loricae, and Tintinnopsella aff. carpathica (Murgeanu &
Filipescu) (Fig. 14V-X ![]() ). We did not observe T. remanei Borza
nor Praetintinnopsella andrusovi Borza. Both species are
considered by several authors (e.g., Remane et al., 1986; Ölveczká
& Reháková, 2022) as subzone indexes of the base of the
Crassicollaria Zone, i.e., Remanei Subzone and Praetintinnopsella Subzone
respectively.
). We did not observe T. remanei Borza
nor Praetintinnopsella andrusovi Borza. Both species are
considered by several authors (e.g., Remane et al., 1986; Ölveczká
& Reháková, 2022) as subzone indexes of the base of the
Crassicollaria Zone, i.e., Remanei Subzone and Praetintinnopsella Subzone
respectively.
5.3.1.2. The Tintinnopsella-Intermedia Subzone (subzone A1) spans
the
sampling interval comprised between 35M and 32H. Calpionellids are larger
in size than in the previous subzone. The first typical medium-sized Crassicollaria
intermedia (Fig. 14AE-AF ![]() ) appears there. It also contains medium-sized Cr.
aff. intermedia (Fig. 14AG
) appears there. It also contains medium-sized Cr.
aff. intermedia (Fig. 14AG ![]() ), as well as common Calpionella grandalpina
(Fig. 14Y-AB
), as well as common Calpionella grandalpina
(Fig. 14Y-AB ![]() ), small-sized C. alpina
(Fig. 14AC-AD
), small-sized C. alpina
(Fig. 14AC-AD ![]() ), and Crassicollaria
aff. massutiniana (Fig. 14AH-AK
), and Crassicollaria
aff. massutiniana (Fig. 14AH-AK ![]() ), and few Tintinnopsella
pseudocarpathica Benzaggagh et al. (Fig. 14U
), and few Tintinnopsella
pseudocarpathica Benzaggagh et al. (Fig. 14U ![]() )
with smaller
loricae than those of the genuine T. carpathica (Murgeanu & Filipescu)
from the Berriasian-Valanginian. Bonetilla boneti (Doben) (Fig.
14F
)
with smaller
loricae than those of the genuine T. carpathica (Murgeanu & Filipescu)
from the Berriasian-Valanginian. Bonetilla boneti (Doben) (Fig.
14F ![]() ), B. sphaerica Benzaggagh
(Fig. 14E
), B. sphaerica Benzaggagh
(Fig. 14E ![]() ),
and primitive calpionellids (Fig. 14L, P-Q, S-T
),
and primitive calpionellids (Fig. 14L, P-Q, S-T ![]() )
found in pseudointraclasts of breccia 35 are reworked from the underlying subzone A0.
)
found in pseudointraclasts of breccia 35 are reworked from the underlying subzone A0.
5.3.1.3. The Intermedia-Alpina Subzone (subzone A2) covers the samples 31B to
30H. Typical forms of Crassicollaria intermedia Durand Delga
(Fig. 14AL-AO ![]() ) are dominating. Their sizes are often larger than those of
specimens from the previous subzone. This subzone also commonly contains Cr.
colomi Lorenz (Fig. 14AQ-AT
) are dominating. Their sizes are often larger than those of
specimens from the previous subzone. This subzone also commonly contains Cr.
colomi Lorenz (Fig. 14AQ-AT ![]() ), Calpionella grandalpina Nagy
(Fig. 14AY-BA
), Calpionella grandalpina Nagy
(Fig. 14AY-BA ![]() ), small-sized C. alpina Lorenz (Fig. 14AW-AX), rare Crassicollaria
massutiniana (Colom) (Fig. 14AP
), small-sized C. alpina Lorenz (Fig. 14AW-AX), rare Crassicollaria
massutiniana (Colom) (Fig. 14AP ![]() ), Cr. parvula Remane
(Fig. 14AU
), Cr. parvula Remane
(Fig. 14AU ![]() ), and Cr.
aff. brevis Remane (Fig. 14AV
), and Cr.
aff. brevis Remane (Fig. 14AV ![]() ).
).
5.3.1.4. The Brevis-Massutiniana Subzone (subzone A3) comprises at least the samples 26/1.0 and 26B. It probably extends downward to the bottom of the breccia 29. Typical forms of Crassicollaria massutiniana (Colom) and Cr. brevis Remane dominate. It also commonly contains small-sized to medium-sized Calpionella alpina Lorenz, C. grandalpina Nagy, and few Tintinnopsella pseudocarpathica Benzaggagh et al..
5.3.2. The Alpina
Zone (zone B),
as originally defined by Remane (1963), corresponds to an "acmé"
interval of the fossil index (op.cit., p. 62: "la prédominance du
genre Calpionella"). The base of this biozone is not characterized
by the first occurrence of Calpionella alpina but by the base of its
first acme. Some followers of Remane have focused on identifying
"explosions" of C. alpina. For instance, Wimbledon's
summary Figure (Wimbledon et al., 2011, Fig. 1) identified not
less than "three 'C. alpina' explosions". However, this
"explosion" concept can be misleading as documented by Granier et
al. (2020b, 2023). As a matter of fact, some cryptoturbidites (Fig.
6L, P ![]() ) are
made of well-sorted calpionellid pseudointraclastic micrograinstones, a kind of
dynamic accumulation that is not related to any biological bloom (Granier
et al., 2023).
) are
made of well-sorted calpionellid pseudointraclastic micrograinstones, a kind of
dynamic accumulation that is not related to any biological bloom (Granier
et al., 2023).
At
Le Chouet, the Alpina Zone starts
from breccia 24 (or possibly from turbidite 25 (Fig. 6P ![]() ), which is locally
missing due to the subsequent basal erosion of debris flow 24) upward at least
to breccia 10.
The physical boundary is
aligned with an important change in the calpionellid assemblage that corresponds
to the disappearance of the main species of the Crassicollaria Zone, i.e.,
Crassicollaria brevis (Fig. 7E-G
), which is locally
missing due to the subsequent basal erosion of debris flow 24) upward at least
to breccia 10.
The physical boundary is
aligned with an important change in the calpionellid assemblage that corresponds
to the disappearance of the main species of the Crassicollaria Zone, i.e.,
Crassicollaria brevis (Fig. 7E-G ![]() ),
Cr. colomi, Cr. intermedia, and Cr. massutiniana
(Fig. 7A-D
),
Cr. colomi, Cr. intermedia, and Cr. massutiniana
(Fig. 7A-D ![]() ),
which are still found in pseudointraclasts (Fig. 2
),
which are still found in pseudointraclasts (Fig. 2 ![]() , red rectangles). Cr.
parvula is the only representative of the genus Crassicollaria to
persist in Berriasian times.
, red rectangles). Cr.
parvula is the only representative of the genus Crassicollaria to
persist in Berriasian times.
Only its first subzone, i.e., the Alpina-Parvula
Subzone (B1 subzone), has been identified in the Le Chouet section. The
abundance of calpionellid loricae contrasts with the low specific diversity of
the whole assemblage that consists of -dominating- small-sized Calpionella alpina Lorenz
(Fig. 7J-L, N-P ![]() ),
-common- Crassicollaria parvula (Fig. 7S-T
),
-common- Crassicollaria parvula (Fig. 7S-T ![]() ), and
-rare- Tintinnopsella pseudocarpathica (Fig. 7M
), and
-rare- Tintinnopsella pseudocarpathica (Fig. 7M ![]() ).
).
5.3.2.1. The Alpina-Parvula Subzone
(subzone B1) is largely dominated by small-sized Calpionella alpina,
in particular the forms with ovoid rounded loricae (Fig. 7N-P ![]() ) and ovoid
elongated loricae (Fig. 7Q
) and ovoid
elongated loricae (Fig. 7Q ![]() ). It also contains common Crassicollaria parvula
Remane (Fig. 7R-T
). It also contains common Crassicollaria parvula
Remane (Fig. 7R-T ![]() ) and few Tintinnopsella pseudocarpathica Benzaggagh
et al. Within the subzone B1 of the logged section, the breccias commonly
contains lithoclasts with specimens of Calpionella elliptalpina Nagy
(Fig. 7I, U-W
) and few Tintinnopsella pseudocarpathica Benzaggagh
et al. Within the subzone B1 of the logged section, the breccias commonly
contains lithoclasts with specimens of Calpionella elliptalpina Nagy
(Fig. 7I, U-W ![]() ), C. grandalpina Nagy, Crassicollaria brevis
and Cr. intermedia Durand Delga (Fig. 7X
), C. grandalpina Nagy, Crassicollaria brevis
and Cr. intermedia Durand Delga (Fig. 7X ![]() ), all reworked from the
older strata of the Crassicollaria Zone.
), all reworked from the
older strata of the Crassicollaria Zone.
|
|
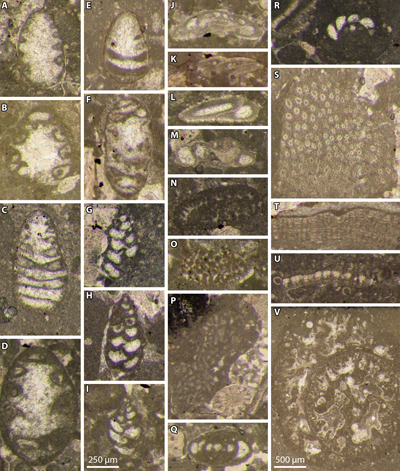
Figure 14: Chitinoidellid and calpionellid specimens
from the upper Tithonian - Boneti Subzone and Crassicollaria Zone (A zone)
strata of the Le Chouet section. A-D: Boneti Subzone; G, I-K, M-O, R, V, X: Chitinoidellid - primitive
calpionellid Subzone (A0 subzone); E-F,
H, L, P-Q, S-T, U, W: material from the Chitinoidellid - primitive
calpionellid Subzone (A0 subzone) reworked in the Tintinnopsella-Intermedia
Subzone (A1 subzone); Y-AK:
Tintinnopsella-Intermedia Subzone (A1 subzone); AL-AP,
AU-BA:
Intermedia-Alpina Subzone (A2 subzone); AQ-AT: Intermedia-Alpina Subzone
(A2 subzone) or ? Brevis-Massutiniana Subzone
(? A3 subzone). A: Bonetilla boneti (Doben); B: Furrazolaia insueta (Řehánek);
C-D: F. cristobalensis
(Furrazola-Bermúdez); E: Bonetilla sphaerica Benzaggagh;
F: B. boneti (Doben); G-H: small-sized Calpionella aff. alpina
Lorenz with rounded loricae; I: small-sized C. aff. alpina Lorenz
with ovoid elongated lorica; J-T: Crassicollaria aff. intermedia Durand
Delga; U: Tintinnopsella pseudocarpathica Benzaggagh et al.;
V-X: T. aff. carpathica (Murgeanu & Filipescu);
Y-AB: Calpionella grandalpina Nagy; AC-AD: small-sized C.
alpina Lorenz; AE-AG: Crassicollaria aff. intermedia Durand
Delga; AH-AK: Cr. aff. massutiniana (Colom);
AL-AO: Cr. intermedia Durand Delga; AP: Cr. massutiniana (Colom); AQ-AT: Cr.
colomi Doben; AU: Cr. parvula; AV: Cr. aff. brevis;
AW-AX: small-sized Calpionella alpina Lorenz; AY-BA: C.
grandalpina Nagy. All
photomicrographs with the same scale bar = 100 µm. A:
sample 40M; B-C: sample 37B; D: sample 38B; E-F, H, L, P-Q, S-U, W:
sample 35M (reworked specimens from the subzone A0); G, I, N, V: sample 36.6; J-K, R: sample
36.4; M: sample 36.2; O: sample 36.5; X: sample 36.3; Y-AA, AF, AI-AJ: sample 32B; AB, AK: sample 32H; AC-AD, AH: sample 33; AE, AG: sample 34M;
AL, AV, BA: sample 31H; AM-AO: sample 31B; AP, AU, AW-AZ: sample 30B; AQ-AT: sample 28
(reworked
specimens from the subzone A2). |
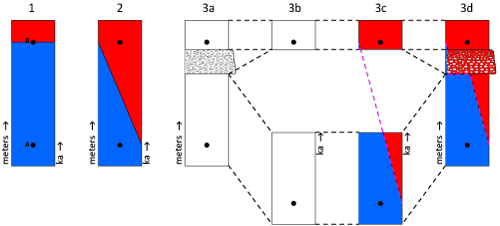
One could argue that a
zone or subzone precisely starts with the first occurrence of the proxy (Fig.
15.1 ![]() ). This assumption can be valid when sampling is dense but it is
debatable when the sampling frequency is low. For instance, in the illustrated
model (Fig. 15.1-2
). This assumption can be valid when sampling is dense but it is
debatable when the sampling frequency is low. For instance, in the illustrated
model (Fig. 15.1-2 ![]() ), two samples A (below) and B (above) were picked 1 meter
apart. Because we deal with a single lithology, it is assumed that the rate of
sedimentation is constant. To simplify the graphical interpretation, the
thickness scales (meters) on the left hand side of the columns and the time
scales (ka) on their right hand side are congruent. The proxy B is observed in
the red zone, not in the blue zone. In this simple model with one lithology (Fig.
15.1-2
), two samples A (below) and B (above) were picked 1 meter
apart. Because we deal with a single lithology, it is assumed that the rate of
sedimentation is constant. To simplify the graphical interpretation, the
thickness scales (meters) on the left hand side of the columns and the time
scales (ka) on their right hand side are congruent. The proxy B is observed in
the red zone, not in the blue zone. In this simple model with one lithology (Fig.
15.1-2 ![]() ), the probability that B belongs to the red zone is 100% whereas
the probability that A belongs to the red zone is nil. Symmetrically, the
probability that A belongs to the blue zone is 100% whereas the probability that
B belongs to the blue zone is nil. Depending of its relative sampling distance
between A and B, the probability that a sample C picked between A and B belongs
to zone B increases when C getting closer to B and decreases when C getting
closer to A. On the basis of this approach, the zonal blue/red boundary is drawn
as an oblique line, not as a horizontal line (Fig. 15.2
), the probability that B belongs to the red zone is 100% whereas
the probability that A belongs to the red zone is nil. Symmetrically, the
probability that A belongs to the blue zone is 100% whereas the probability that
B belongs to the blue zone is nil. Depending of its relative sampling distance
between A and B, the probability that a sample C picked between A and B belongs
to zone B increases when C getting closer to B and decreases when C getting
closer to A. On the basis of this approach, the zonal blue/red boundary is drawn
as an oblique line, not as a horizontal line (Fig. 15.2 ![]() ).
).
For example, the base of the Chitinoidella Zone, i.e., the lower/upper Tithonian boundary, at Le Chouet is drawn as an oblique line from the location of sample CH4b up to that of sample 40M, both samples being picked more than 3 meters apart on the log. In alternative approaches, the boundary is commonly ascribed 1) either to the bottom of the B bed when the latter is found between samples B and A, 2) to a median location between samples B and A (that was the option chosen by Wimbledon et al., 2020a, Supplement Fig. S1), or 3) to a random location close to sample B (that was the option chosen by Wimbledon et al., 2013, p. 444: "The Crassicollaria-Calpionella" (Alpina) "zonal boundary may be placed between 101 and the next sample below", Fig. 4). All these non-probabilistic approaches (either the interpolation or the diagonal boundary) are misleading in the case of Le Chouet (an previously in that of Tré Maroua, Granier et al., 2020b, 2023) when a turbidite or debris flow bed occurs between samples B and A.
As discussed above, identifying a zonal boundary can be straightforward when dealing with a single facies because the main concerns will be the sampling frequency and because, after an initial screening, one can always return to the field and densify the sampling around the boundary to better circumscribe it. However, in the case for most of Le Chouet section, two discrete types of facies differ markedly in their net sedimentation rates, which implies adaptation of the simplistic model. The sedimentation rate of the "calcaires blancs" (saccocomid, calpionellid or radiolarian mud- or wackestones) likely represents a relatively constant background basinal sedimentation of a few cm/ka whereas that of the episodic turbidites and debris flows is often higher than several centimeters (or even tens of centimeters) per hour (see Granier et al., 2013: Fig. 7.B).
Figure 15.3 ![]() illustrates a case with two lithologies
where a slice of debris flow occurs within the basinal facies and between the
locations of samples A and B. The first column (Fig. 15.3a
illustrates a case with two lithologies
where a slice of debris flow occurs within the basinal facies and between the
locations of samples A and B. The first column (Fig. 15.3a ![]() ) corresponds to the
lithology versus the thickness; the second column (Fig. 15.3b
) corresponds to the
lithology versus the thickness; the second column (Fig. 15.3b ![]() ) corresponds to
the lithology versus the time. In a practical case, to convert thickness into
time, the thickness of each interval should be divided by the corresponding rate
of sedimentation. As previously noted for the single lithology case, the
thickness scales (meters) and the time scales (ka) are congruent. The graphical
thicknesses remain the same for the "calcaires blancs" but the
graphical thickness of the debris flow tends toward zero (Fig.
15.3b
) corresponds to
the lithology versus the time. In a practical case, to convert thickness into
time, the thickness of each interval should be divided by the corresponding rate
of sedimentation. As previously noted for the single lithology case, the
thickness scales (meters) and the time scales (ka) are congruent. The graphical
thicknesses remain the same for the "calcaires blancs" but the
graphical thickness of the debris flow tends toward zero (Fig.
15.3b ![]() ), i.e.,
the graphical representation of a few hours on a ka scale would be a horizontal
line. However, the basinal facies above and below the debris flow are not
exactly superposed. There is a time gap between them that represents basinal
sediment (and possibly turbidites and debris flows) that was removed by erosion
at the bottom of the debris flow (Fig. 8
), i.e.,
the graphical representation of a few hours on a ka scale would be a horizontal
line. However, the basinal facies above and below the debris flow are not
exactly superposed. There is a time gap between them that represents basinal
sediment (and possibly turbidites and debris flows) that was removed by erosion
at the bottom of the debris flow (Fig. 8 ![]() ).
).
As stated in Granier et al.
(2023), "There is no relationship" "between the depth
of erosion", i.e., the amount of material removed at the erosion
surface, "at a location and the amount of material accumulated above the
erosion surface at this same location". The duration of such hiatuses is
commonly impossible to determine. Following the previously described simplistic
approach, the zonal blue/red boundary is drawn as an oblique line (Fig.
15.3c ![]() ).
In order to revert to the original figure (Fig. 15.3a
).
In order to revert to the original figure (Fig. 15.3a ![]() ), i.e., to convert
again time into thickness, the duration of each interval should be
multiplied by the rate of sedimentation for the corresponding lithology. On the
new figure, the blue/red boundary is not a straight line anymore (Fig.
15.3d
), i.e., to convert
again time into thickness, the duration of each interval should be
multiplied by the rate of sedimentation for the corresponding lithology. On the
new figure, the blue/red boundary is not a straight line anymore (Fig.
15.3d ![]() ).
This last graphical representation makes it clear how, in this case, the debris
flow very highly probably belongs to the red zone and that the base of this red
zone should be located on the erosional surface at the base of this debris flow.
The probability that the red zone extends below the breccia is not nil but quite
low. For instance, as
documented by Granier et al. (2020b,
2023) for the Tré
Maroua section, the base of the Ferasini Subzone of the Alpina Zone
and the base of the M18r magnetozone should be located at the bottom of the
uppermost debris flow, i.e., number 69, not at its top as shown in Wimbledon
et al. (2020a: Fig. 4).
).
This last graphical representation makes it clear how, in this case, the debris
flow very highly probably belongs to the red zone and that the base of this red
zone should be located on the erosional surface at the base of this debris flow.
The probability that the red zone extends below the breccia is not nil but quite
low. For instance, as
documented by Granier et al. (2020b,
2023) for the Tré
Maroua section, the base of the Ferasini Subzone of the Alpina Zone
and the base of the M18r magnetozone should be located at the bottom of the
uppermost debris flow, i.e., number 69, not at its top as shown in Wimbledon
et al. (2020a: Fig. 4).
|
|
Figure 15:
Models for a confident identification of zonal boundaries based on the
lithologies and the corresponding sedimentary rates. Sampling rate is not
negligible. Erosion rates at the bottom of debris flows and turbitides may vary
significantly. See description in the text. |
Our control on the saccocomid zones or on the Chitinoidella Zone is poor, mainly due to a low sampling frequency in the corresponding interval. For instance, the significant uncertainty on the location of the lower/upper Tithonian boundary is mainly driven by a poor sampling within the interval from sample Ch4b to sample 40M.
In contrast, the bases of the Crassicollaria Zone and subzones and those of the Alpina Zone, hence of its first subzone, are well identified. They systemically correspond to the bottoms of breccia or turbidite layers:
the base of zone A or subzone A0 matches with the bottom of the thick breccia bed Ch7;
that of subzone A1 with the bottom of the thick
breccia bed set 35 to 33. In sample 35M, Bonetilla
boneti (Doben) (Fig. 14F ![]() )
and B. sphaerica Benzaggagh (Fig. 14E
)
and B. sphaerica Benzaggagh (Fig. 14E ![]() ) are found reworked in extraclasts derived from the
preceding subzone (subzone A0);
) are found reworked in extraclasts derived from the
preceding subzone (subzone A0);
that of subzone A2 with the bottom of the graded breccia bed 31;
that of subzone A3 is tentatively located at the bottom of turbidite bed 29 of a set comprising the breccia bed 28 sandwiched between the turbidite beds 29 and 27;
that of zone B or subzone B1 matches with the
bottom of the graded breccia bed 24 or locally to the bottom of turbidite bed 25
when the latter was not eroded by the overlying debris flow 24 (Fig.
5H-I ![]() ).
).
Besides the probabilistic or non-probabilistic options and the sampling frequency, the other key factor that generates inherent discrepancies in the location of the zonal boundaries can be differences in the diverse acceptions of species or zones by the various authors. For instance, Reháková (in Wimbledon et al., 2013, 2020a) does not refer to Tintinnopsella pseudocarpathica Benzaggagh et al., 2012, or Bonetilla sphaerica Benzaggagh, 2021. The various authors do not necessarily use the same subzones (see Benzaggagh, 2020, for discussion).
As
for the calpionellid biozones, all authors generally share the same view regarding
the definitions of the Chitinoidella, Crassicollaria (zone A) and Alpina (zone
B) zones. Due to poor sampling and facies that are not favorable to the
preservation (or identification) of the chitinelloids, i.e, turbidites,
the base of the Chitinoidella Zone cannot be accurately identified: It is
expected to fall between 9 and 12 m on the log of Figure 2 ![]() whereas it would be
near 5 m (below our turbidite Ch2) according to Remane's log
(1970) and
near 9 m
according to Wimbledon et al.' (2020a) log. All the authors agree upon
the base of the Crassicollaria
Zone near 15.5 m (at the bottom of our breccia Ch7) on the log of Figure 2
whereas it would be
near 5 m (below our turbidite Ch2) according to Remane's log
(1970) and
near 9 m
according to Wimbledon et al.' (2020a) log. All the authors agree upon
the base of the Crassicollaria
Zone near 15.5 m (at the bottom of our breccia Ch7) on the log of Figure 2 ![]() . The
main discrepancies occur at the base of the Calpionella alpina acme Zone:
1) Remane
(1970) identifies it at the base of turbidite 22, near 27.7 m; 2) Wimbledon
et al.
near 30 and near 29.5 m in 2013 and in 2020 (2020a) respectively; 3) we
assume that it should be placed at
the base of turbidite(s) 24 (or 25), near 26.5 m on the log of Figure 2
. The
main discrepancies occur at the base of the Calpionella alpina acme Zone:
1) Remane
(1970) identifies it at the base of turbidite 22, near 27.7 m; 2) Wimbledon
et al.
near 30 and near 29.5 m in 2013 and in 2020 (2020a) respectively; 3) we
assume that it should be placed at
the base of turbidite(s) 24 (or 25), near 26.5 m on the log of Figure 2 ![]() .
It is worth mentioning that the marker for this boundary (i.e., the base
of the Calpionella alpina acme Zone) was the candidate proxy for the
Berriasian GSSP proposed by the past Berriasian Working Group (Wimbledon et
al., 2013, 2020a). There are also significant
discrepancies regarding the subzones of the Crassicollaria Zone. The only agreements are found
with 1) the Remanei Subzone of Wimbledon et al.
(2020a) that matches our subzone A0 and 2) the base of the Intermedia
Subzone of Wimbledon et al. (2020a) and that of our subzone A1 because both fall
at the base of turbidite 35, near 18 m on the log
of Figure 2
.
It is worth mentioning that the marker for this boundary (i.e., the base
of the Calpionella alpina acme Zone) was the candidate proxy for the
Berriasian GSSP proposed by the past Berriasian Working Group (Wimbledon et
al., 2013, 2020a). There are also significant
discrepancies regarding the subzones of the Crassicollaria Zone. The only agreements are found
with 1) the Remanei Subzone of Wimbledon et al.
(2020a) that matches our subzone A0 and 2) the base of the Intermedia
Subzone of Wimbledon et al. (2020a) and that of our subzone A1 because both fall
at the base of turbidite 35, near 18 m on the log
of Figure 2 ![]() . The bases of Remane's
(1970) subzones A2 and A3 respectively
fall on top of turbidite 33, at 20.5 m, and above turbidite 27, at 25.5 m on the
log of Figure 2
. The bases of Remane's
(1970) subzones A2 and A3 respectively
fall on top of turbidite 33, at 20.5 m, and above turbidite 27, at 25.5 m on the
log of Figure 2 ![]() . The bases of the Intermedia
and Colomi subzones of Wimbledon et al.
(2013, 2020a) fall between turbidite 33 and breccia 31, at 22 m for the first
one, and below turbidites 24-25, near 26.4 m for the second one. In contrast and
as for our previous zones and subzones, the bases of our subzones A2 and A3
correspond to the erosional bases of turbidites or debris flows, i.e., to
the base of breccia 31, at 23 m for the first one, and to the base of turbidite
29, at 24.5 m for the second one.
. The bases of the Intermedia
and Colomi subzones of Wimbledon et al.
(2013, 2020a) fall between turbidite 33 and breccia 31, at 22 m for the first
one, and below turbidites 24-25, near 26.4 m for the second one. In contrast and
as for our previous zones and subzones, the bases of our subzones A2 and A3
correspond to the erosional bases of turbidites or debris flows, i.e., to
the base of breccia 31, at 23 m for the first one, and to the base of turbidite
29, at 24.5 m for the second one.
From
the outcrops on the side of the tarred track, the erosional bases of the gravity flows
(turbidites and debris flows) at all scales are well exposed (Figs.
4H ![]() ,
5C, F, H
,
5C, F, H ![]() ). In
Figure 5H
). In
Figure 5H ![]() , turbidite 25 is partly eroded at the bottom of the debris flow
24. Many extraclasts and other erosional features such as microscopic erosional
surfaces are also visible under the binocular microscope (see Fig.
6L
, turbidite 25 is partly eroded at the bottom of the debris flow
24. Many extraclasts and other erosional features such as microscopic erosional
surfaces are also visible under the binocular microscope (see Fig.
6L ![]() here or
see Granier
et al., 2023, Fig.
8E at Tré Maroua).
here or
see Granier
et al., 2023, Fig.
8E at Tré Maroua).
According
to Granier et al. (2023), "There is inherently no
relationship between the thickness of a debris flow or a turbidite layer at a
location and the amount of material that was eroded from this same location and
that accumulated downdip". Due to the limited temporal resolution of the
saccocomid and calpionellid biozones, estimating the depth of erosion is
challenging, not to say almost impossible. However, there are a few exceptions.
For instance, at Tré Maroua, Granier
et al. (2020b, 2023) reported saccocomid-bearing
lithoclasts more than 2 meters and even more than 10 meters above the last
saccocomid zone. From the same section, Granier et al.
(2020b, 2023) also documented Crassicollaria-bearing lithoclasts (Granier
et al., 2020b, Pl. 3, figs. B-C) more than 5 meters and even more than 9
meters above the Crassicollaria Zone. At Le Chouet, there are fewer clues. In
breccia 24, at 27 m on
the log of Figure 2 ![]() ,
i.e., at the bottom of the Alpina Zone, Crassicollaria
brevis Remane,
Cr. intermedia Durand Delga (Fig. 7X
,
i.e., at the bottom of the Alpina Zone, Crassicollaria
brevis Remane,
Cr. intermedia Durand Delga (Fig. 7X ![]() ), and Cr. massutiniana
(Colom) are found in extraclasts derived from the preceding
zone (Crassicollaria Zone). This same Crassicollaria assemblage is also observed within the
Alpina Zone in breccia 16, at 31.7 m, more than 5 meters above the Crassicollaria Zone.
), and Cr. massutiniana
(Colom) are found in extraclasts derived from the preceding
zone (Crassicollaria Zone). This same Crassicollaria assemblage is also observed within the
Alpina Zone in breccia 16, at 31.7 m, more than 5 meters above the Crassicollaria Zone.
The Le Chouet section spans a Tithonian-lower Berriasian interval. Saccocomid zones 4 to 7 as well as Chitinoidella, Crassicollaria and Alpina zones of the calpionellids have been documented. However, the Dobeni Subzone of the Chitinoidella Zone has not been identified in its lower part and only the first subzone of the Alpina Zone, i.e., the Alpina Subzone, has been documented at its upper part.
Whereas the location of the lower/upper
Tithonian boundary remains uncertain (possibly close to Ch4, i.e., near 9
m on the log of Figure 2 ![]() ), the Tithonian/Berriasian boundary defined by
calpionellid biostratigraphy coincides with the base(s) of turbidite 25 (or
breccia 24 when the previous one has been eroded, i.e., near
26.5 m on the log of Figure 2
), the Tithonian/Berriasian boundary defined by
calpionellid biostratigraphy coincides with the base(s) of turbidite 25 (or
breccia 24 when the previous one has been eroded, i.e., near
26.5 m on the log of Figure 2 ![]() ):
Figs. 4A
):
Figs. 4A ![]() ,
5H-I
,
5H-I ![]() ,
6B
,
6B ![]() . That pushes the
Tithonian/Berriasian boundary down by almost 1 m relative to Remane
(1970; note that most Crassicollarias found in the interval are here considered
to be reworked) and some 4 m relative to Wimbledon et al.
(2013, 2020a).
. That pushes the
Tithonian/Berriasian boundary down by almost 1 m relative to Remane
(1970; note that most Crassicollarias found in the interval are here considered
to be reworked) and some 4 m relative to Wimbledon et al.
(2013, 2020a).
The foraminifer Protopeneroplis ultragranulata (Gorbatchik)
is the index of the P. ultragranulata Subzone of the Anchispirocyclina
lusitanica Zone (Tithonian-lower Berriasian). This microfossil was supposed
to first occur in the late Tithonian (Bucur, 1997) and the eponymic
subzone to span the upper Tithonian-lower Berriasian interval (Granier,
2019; Granier et al., 2020a). However, its find in upper lower
Tithonian strata (Fig. 12AC ![]() , sample Ch2b) extends its subzone downwards. Granier
(2019) stated that "it is impossible to distinguish the (upper) Tithonian
from the lower Berriasian" in the shallow-water facies of the Tethys realm.
This wider range for the index of the subzone drives the nail deeper into the
coffin. This argument, together with the lack of biological crisis and the
instability of the boundary definition and location, supports the assessment
that the default Tithonian/Berriasian boundary (i.e., base of the Calpionella
alpina "acme" Zone) is a poor system boundary. In contrast, as
stated by Énay
(2020), "the base Valanginian, which corresponds to biotic crises affecting
the ammonites and other groups, is by far the better alternative" for
"the Jurassic/Cretaceous system boundary".
, sample Ch2b) extends its subzone downwards. Granier
(2019) stated that "it is impossible to distinguish the (upper) Tithonian
from the lower Berriasian" in the shallow-water facies of the Tethys realm.
This wider range for the index of the subzone drives the nail deeper into the
coffin. This argument, together with the lack of biological crisis and the
instability of the boundary definition and location, supports the assessment
that the default Tithonian/Berriasian boundary (i.e., base of the Calpionella
alpina "acme" Zone) is a poor system boundary. In contrast, as
stated by Énay
(2020), "the base Valanginian, which corresponds to biotic crises affecting
the ammonites and other groups, is by far the better alternative" for
"the Jurassic/Cretaceous system boundary".
The frequency of gravity-flow (debris flows and turbidites, including cryptoturbidites) led us to reassess the calpionellid stratigraphy with the eyes of sedimentologists. All bases of calpionellid zones and subzones (except those of the Chitinoidella Zone and its Boneti Subzone) are located at basal erosional surfaces of turbidites or debris flows. Accordingly, as in the Tré Maroua section (Granier et al., 2020b, 2023), all these zonal boundaries are probably hiatal here too. As already explained in Granier et al. (2023), not only these two localities but even "a wider region in the Vocontian Trough", which is affected by "paleotectonic instability", should enevitably fail to provide any suitable Berriasian GSSP candidate "in contradiction with the expectations of the 'Colloque sur la limite Jurassique/Crétacé' held in Lyon in 1973 (Flandrin et al., 1975)". It is worth mentioning that corresponding time gaps at erosional surfaces, even if short, artificially emphasise the contrasts between the successive calpionellid assemblages.
The depth of erosion at the bottom of any gravity-flow bed can hardly be estimated in any single site. However, because pebbles and cobbles sourced from the Crassicollaria Zone are reported at Le Chouet some 5 m above the local top of this zone, it is assumed that the depth of erosion could have reached more than 5 m in some updip locations. On the basis of data from the Tré Maroua site (Granier et al., 2020b, 2023), it can even be estimated that updip depth of erosion could have locally reached 10 m or more. Considering published regional data (Courjault, 2011; Courjault et al., 2011; Ferry et al., 2015; Ferry, 2017), such deep erosional features did not result from a single episode but probably from multiple, successive episodes digging each time deeper into the preexisting sedimentary substratum.
Field work was made possible thanks to a grant from the Foundation "Carnets de Géologie". Part of this work was presented on the occasion of JK2018 - International Meeting around the Jurassic/Cretaceous Boundary held in Genčve (Switzerland) from December 5th to 7th, 2018 (Ferry & Granier, 2018), and of the 11th International Cretaceous Symposium held in Warsaw (Poland) from August 22th to 26th, 2022 (Granier et al., 2022). It is a contribution to Project no. 46908YB of the PHC ("Partenariats Hubert Curien") Polonium 2022. The manuscript was originally submitted to Geologica Carpathica and later withdrawn due to requests to remove or modify some key figures. However, the authors are grateful to the two reviewers and the editor for their comments and suggestions that helped improve the original manuscript. Special thanks are also due to Phil Salvador who kindly checked the final version of our English text.
Benzaggagh M. (2020).- Discussion on the calpionellids biozones and proposal of a homogeneous calpionellid zonation for the Tethyan Realm.- Cretaceous Research, vol. 113, article 104184, 24 p.
Benzaggagh M. (2021).- Systematic revision and evolution of the Tithonian family Chitinoidellidae Trejo, 1975.- Carnets Geol., Madrid, vol. 21, no. 2, p. 27-53.
Benzaggagh M., Cecca F., Schnyder J., Seyed-Emami K. & Majidifard M.R. (2012).- Calpionelles et microfaunes pélagiques du Jurassique supérieur-Crétacé inférieur dans les formations Shal et Kolur (Montagnes du Talesh, chaîne de l'Elbourz, Nord-Ouest Iran). Répartition stratigraphique, espčces nouvelles, révision systématique et comparaisons régionales.- Annales de Paléontologie, Paris, vol. 98, no. 4, p. 253-301.
Benzaggagh M., Homberg C., Schnyder J. & Ben Adesselam-Mahdaoui S. (2015).- Description et biozonation des sections de crinoďdes saccocomidés du Jurassique supérieur (Oxfordien-Tithonien) du domaine téthysien occidental.- Annales de Paléontologie, Paris, vol. 101, p. 95-117.
Bucur I.I. (1997).- Representatives of the genus Protopeneroplis (foraminifera) in the Jurassic and Lower Cretaceous deposits from Romania. Comparisons with other regions of the Tethyan realm.- Acta Palaeontologica Romaniae, Cluj-Napoca, vol. 1, p. 65-71 (Pls. 6.I-III).
Cecca F., Énay R. & Le Hégarat G. (1989).- L'Ardescien (Tithonique supérieur) de la région stratotypique : Séries de référence et faunes (ammonites, calpionelles) de la bordure ardéchoise.- Documents des Laboratoires de Géologie de Lyon, Villeurbanne, vol. 107, 115 p.
Courjault T. (2011).- Brčches gravitaires sous-marines du Tithonien subalpin (S-E France).- PhD thesis, Université de Strasbourg, 339 p.
Courjault T., Grosheny D., Ferry S. & Sausse J. (2011).- Detailed anatomy of a deep-water carbonate breccia lobe (Upper Jurassic, French subalpine basin).- Sedimentary Geology, vol. 238, no. 1-2, p. 156-171.
Énay R. (2020).- The Jurassic/Cretaceous System Boundary is at an impasse. Why not go back to Oppel's 1865 original and historic definition of the Tithonian?- Cretaceous Research, vol. 106, article 104241, 20 p. DOI: 10.1016/j.cretres.2019.104241
Ferry S. (2017).- Summary on Mesozoic carbonate deposits of the Vocontian Trough (Subalpine Chains, SE France). In: Granier B. (ed.): Some key Lower Cretaceous sites in Drôme (SE France).- Association Carnets de Géologie, Madrid, CG2017_B01, p. 9-42. DOI: 10.4267/2042/62543
Ferry S. & Granier B. (2018).- 12. Looking for the Jurassic-Cretaceous system boundary in the Vocontian Trough (S-E France): Sedimentological problems. In: Granier B. (ed.), JK2018 - International Meeting around the Jurassic-Cretaceous Boundary.- Association Carnets de Géologie, Madrid, CG2019_B01, p. 24 (abstract). DOI: 10.4267/2042/69811
Ferry S., Grosheny D., Backert N. & Atrops F. (2015).- The base-of-slope carbonate breccia system of Céüse (Tithonian, S-E France): Occurrence of progradational stratification in the head plug of coarse granular flow deposits.- Sedimentary Geology, vol. 317, p. 71-86.
Flandrin J. (1970).- Luc-en-Diois.- Carte géologique détaillée de la France au 1/50 000, Orléans, Feuille 0868, Notice XXXII-38, 19 p., 1 map.
Flandrin J., Schaer J.-P., Énay R., Remane J., Rio M., Kubler B., Le Hégarat G., Mouterde R. & Thieuloy J.-P. (1975).- Colloque sur la limite Jurassique/Crétacé, Lyon-Neuchâtel, 1973. Mémoires du Bureau de Recherches Géologiques et Miničres, Orléans, vol. 86, 393 p.
Granier B. (2019).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition. In: Granier B. (ed.), VSI: The transition of the Jurassic to the Cretaceous: An early XXIth century holistic approach.- Cretaceous Research, vol. 93, p. 245-274.
Granier B., Benzaggagh M. & Ferry S. (2023).- Revised holostratigraphy of the Tithonian-Berriasian transition at Tré Maroua (Le Saix, Hautes-Alpes, SE France): Study of a rejected Berriasian GSSP candidate.- Volumina Jurassica, Varsaw, vol. XXI, p. 1-18.
Granier B., Clavel B., Moullade M., Busnardo R., Charollais J., Tronchetti G. & Desjacques P. (2013).- L'Estellon (Baronnies, France), a "Rosetta Stone" for the Urgonian biostratigraphy.- Carnets Geol., Madrid, vol. 13, no. A04, CG2013_A04, p. 163-207. DOI: 10.4267/2042/51213
Granier B., Énay R. & Charollais J. (2020a).- News and Reviews - Discussion of the paper by Wimbledon et al., 2020b, entitled "The proposal of a GSSP for the Berriasian Stage (Cretaceous System): Part 1" [Volumina Jurassica, XVIII (1)].- Volumina Jurassica, Varsaw, vol. XVIII, no. 2, p. 237-250.
Granier B., Ferry S. & Benzaggagh M. (2020b).- A critical look at Tré Maroua (Le Saix, Hautes-Alpes, France), the Berriasian GSSP candidate section.- Carnets Geol., Madrid, vol. 20, no. 1, p. 1-17. DOI: 10.4267/2042/70714
Granier B., Ferry S. & Benzaggagh M. (2022).- Calpionellid biostratigraphy judged by the yardstick of sedimentology. In: 11th International Cretaceous Symposium.- Abstract Volume, Talk_S10, p. 176-177 (abstract).
Jain S., Mulugeta M., Benzaggagh M., Salamon M.A. & Schmerold R. (2021).- Discovery of chitinoidellids and calpionellids from the Blue Nile Basin and the Jurassic-Cretaceous boundary in Ethiopia.- Cretaceous Research, vol. 132, article 105112, 17 p.
Kietzmann D.A. (2017).- Chitinoidellids from the Early Tithonian-Early Valanginian Vaca Muerta Formation in the Northern Neuquén Basin, Argentina.- Journal of South American Earth Sciences, vol. 76, p. 152-164.
Kietzmann D.A., Iglesia Llanos M.P., González Tomassini F., Lanusse Noguera I., Vallejo D. & Hernán Reijenstein R. (2021).- Upper Jurassic-Lower Cretaceous calpionellid zones in the Neuquén Basin (Southern Andes, Argentina): Correlation with ammonite zones and biostratigraphic synthesis.- Cretaceous Research, vol. 127, article 104950, 28 p.
Le Hégarat G. (1973).- Le Berriasien du Sud-Est de la France.- Thčse Docteur čs Sciences naturelles 149 (1971), Université Claude Bernard - Lyon; Documents du Laboratoire de Géologie de la Faculté de Lyon, Villeurbanne, vol. 43, 576 p.
Ölveczká D. & Reháková D. (2022).- Upper Tithonian Crassicollaria Zone: New data on the calpionellid distribution and subzonal division of the Pieniny Klippen Belt in Western Carpathians.- Acta Geologica Slovaca, Bratislava, vol. 14, no. 1, p. 37-56.
Postma G., Nemec W. & Kleinspehn K.L. (1988).- Large floating clasts in turbidites: A mechanism for their emplacement.- Sedimentary Geology, vol. 58, no. 1, p. 47-61.
Remane J. (1963).- Les calpionelles dans les couches de passage Jurassique-Crétacé de la fosse vocontienne.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Grenoble, vol. 39, p. 25-82.
Remane J. (1970).- Die Entstehung der resedimentären Breccien im Obertithon der subalpinen Ketten Frankreichs.- Dissertation (1969), Göttingen; Eclogae geologicae Helvetiae, Basel, vol. 63, no. 3, p. 685-740.
Remane J., Bakalova-Ivanova D., Borza K., Knauer J., Nagy I., Pop G. & Tardi-Filácz E. (1986).- Agreement on the subdivision of the standard calpionellid zones defined at the Iind planktonic conference, Roma 1970.- Acta Geologica Hungarica, Budapest, vol. 29, no. 1-2, p. 5-14.
Schlagintweit F. & Gawlick H.-J. (2011).- Perturbatacrusta leini n. gen., n. sp. a new microencruster incertae sedis (? sponge) from Late Jurassic to earliest Cretaceous platform margin carbonates of the Northern Calcareous Alps of Austria.- Facies, Erlangen, vol. 57, p. 123-135.
Wimbledon W.A.P., Casellato C.E., Reháková D., Bulot L.G., Erba E., Gardin S., Verreussel R.M.C.H., Munsterman D.K. & Hunt C. (2011).- Fixing a basal Berriasian and Jurassic-Cretaceous (J-K) boundary - Perhaps there is some light at the end of the tunnel?- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 117, no. 2, p. 295-307.
Wimbledon W.A.P., Reháková D., Pszczółkowski A., Casellato C.E., Halásová E., Frau C., Bulot L.G., Grabowski J., Sobień K., Pruner P., Schnabl P. & Čížková K. (2013).- An account of the bio- and magnetostratigraphy of the Upper Tithonian-Lower Berriasian interval at Le Chouet, Drôme (SE France).- Geologica Carpathica, Bratislava, vol. 64, no. 6, p. 437-460.
Wimbledon W.A.P, Reháková D., Svobodová A., Schnabl P., Pruner P., Elbra T., Šifnerová K., Kdýr Š., Frau C., Schnyder J. & Galbrun B. (2020a).- Fixing a J/K boundary: A comparative account of key Tithonian-Berriasian profiles in the departments of Drôme and Hautes-Alpes, France.- Geologica Carpathica, Bratislava, vol. 71, no. 1, p. 24-46.
Wimbledon W.A.P, Reháková D., Svobodová A., Elbra T., Schnabl P., Pruner P., Šifnerová K., Kdýr Š., Dzyuba O., Schnyder J., Galbrun B., Košták M., Vaňková L., Copestake P., Hunt C.O., Riccardi A., Poulton T.P., Bulot L.G., Frau C. & De Lena L. (2020b).- The proposal of a GSSP for the Berriasian Stage (Cretaceous System): Part 1.- Volumina Jurassica, Varsaw, vol. XVIII, no. 1, p. 53-106.