◄ Carnets Geol. 23 (8) ►
![]()
Outline:
[1. Introduction]
[2. Stratigraphy]
[3. Systematic palaeontology]
[4. Conclusion]
and ...
[Bibliographic references]
Department of
Geography and Geology, The University of the West Indies, Mona, Kingston 7
(Jamaica)
Published online in final form (pdf) on August 27, 2023
DOI 10.2110/carnets.2023.2308
![]()
[Editor: Bruno R.C. Granier]
![]()
The new larger benthic foraminifer, Hanovolepidina browni gen. nov. sp. nov., is described from the Chapelton Formation (mid-Lutetian, middle Eocene, ABZ7) of Jamaica. The genus shows the characteristic transition from a single to double equatorial layer about mid-way across the radius as seen in axial sections that characterises the family Pseudolepidinidae. Equatorial sections show the embryo followed by 4 or 5 spiral rotaliid chambers with the last chamber giving rise to a primary spiral and secondary (counter-) spiral of equatorial chambers. The presence of a long rotaliid stage suggests a more primitive form that Pseudolepidina, and Hanovolepidina gen. nov. differs also from the latter in lacking the extra principal accessory chamber seen in axial section. Hanovolepidina gen. nov. is provisionally suggested as an ancestor of the late Eocene Triplalepidina. The new discovery demonstrates a greater diversity of the family Pseudolpeidinidae in the Caribbean that previously recorded, although occurrences of this family are very restricted stratigraphically suggesting that it may appear in the region due to migration events.
• Pseudolepidinidae;
• Larger
Benthic Foraminifera;
• Lutetian (Middle Eocene);
• Yellow Limestone Group;
• Jamaica
Mitchell S.F. (2023).- A new pseudolepidinid foraminifer, Hanovolepidina browni gen. nov. sp. nov., from the middle Eocene (mid-Lutetian) of Jamaica and its significance.- Carnets Geol., Madrid, vol. 23, no. 8, p. 149-159.
Un foraminifčre pseudolepidinidé nouveau, Hanovolepidina browni gen. nov. sp. nov., de l'Éocčne moyen (Lutétien moyen) de Jamaďque et sa signification.- Un nouveau grand foraminifčre benthique, Hanovolepidina browni gen. nov. sp. nov., est décrit dans la Formation Chapelton (Lutétien moyen, Éocčne moyen, ABZ7) de Jamaďque. Le genre présente la transition caractéristique d'une couche équatoriale simple ŕ double ŕ mi-distance environ du rayon observable en sections axiales, un critčre qui définit la famille Pseudolepidinidae. Les sections équatoriales montrent un embryon suivi de 4 ou 5 chambres spirales rotaliides avec la derničre chambre donnant naissance ŕ une spirale primaire et une (contre-) spirale secondaire de chambres équatoriales. La présence d'une longue phase rotaliide suggčre une forme plus primitive que Pseudolepidina, tandis que Hanovolepidina gen. nov. diffčre également de ce dernier par l'absence de chambre accessoire principale supplémentaire visible en section axiale. Hanovolepidina gen. nov. est considéré provisoirement comme un ancętre du genre fini-éocčne Triplalepidina. Bien que les occurrences de la famille Pseudolepidinidae soient trčs restreintes stratigraphiquement, cette nouvelle découverte dans les Caraďbes illustre une diversité plus grande que celle précédemment enregistrée, suggérant ainsi qu'elle a pu apparaître dans la région ŕ la faveur d'événements migratoires.
•
Pseudolepidinidae ;
•
Grand Foraminifčre Benthique ;
•
Lutétien (Éocčne moyen) ;
• Groupe du Calcaire jaune ;
• Jamaďque
The Eocene foraminiferid family Pseudolepidinidae Mitchell, E. Robinson & Özcan in Mitchell et al., 2022, contains two previously described genera, Pseudolepidina Barker & Grimsdale, 1937, and Triplalepidina Vaughan & Cole, 1938. The family is restricted to the Americas, where there are only sporadic occurrences (Barker & Grimsdale, 1937; Vaughan & Cole, 1938; Adams, 1987; Mitchell et al., 2022). The family contains orbitoidiform foraminifers that are distinguished by the development of a double row of equatorial chamberlets in conjunction with well-developed tiers of lateral chamberlets. In the previously earliest known form Pseudolepidina, from the mid-Lutetian, two Principal Accessory Chambers (PACs) are already developed (together with a possible third PAC that is seen in axial section). PACs only develop within the two subfamilies (Orbitoininae Mitchell, E. Robinson & Özcan in Mitchell et al., 2022, and Lepidocyclininae Scheffen, 1932) of the Lepidocyclinidae Scheffen, 1932, in the late Lutetian (Mitchell et al., 2022), implying that the Pseudolepidinidae are a separate radiation. In this paper the new genus Hanovolepidina is described from the middle Eocene of central and western Jamaica and its significance for understanding the evolution of Pseudolepidinidae is discussed.
Eocene rocks are
widely distributed across Jamaica where they form parts of the Wag Water and
Richmond formations and the Yellow Limestone and White Limestone groups (Zans
et al., 1963; Robinson, 1994; Robinson
& Mitchell, 1999; Mitchell,
2004, 2013a, 2013b,
2016, 2021; Mitchell
et al., 2022). The material described
herein comes from two localities: the Yellow Limestone below Swanswick House,
parish of Trelawny, and the Chapelton Formation, south of the Cretaceous Green
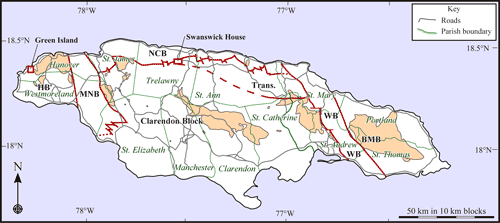
Island Inlier, in the parish of Hanover (Fig. 1 ![]() ).
).
|
Figure 1:
Geographical
map of Jamaica showing parishes, Cretaceous inliers (pale orange), main Cenozoic
blocks (HB, Hanover, BMB, Blue Mountains) and belts (MNB, Montpelier - New
Market, NCB, North Coast, WB, Wag Water) and location of maps where Hanovolepidina
gen. nov. has been collected. |
A single specimen of Hanovolepidina
browni sp. nov., previously figured as Pseudolepidina
trimera Barker & Grimsdale by Mitchell et
al. (2022, Fig. 54.6), was collected from the roadway to Swanswick House
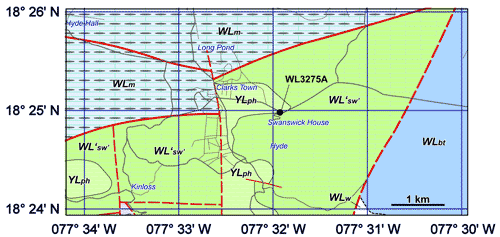
(Fig.
2 ![]() ). The specimen came from sample WL3275A (GPS coordinates: 18°24'58.9"N
77°31'54.9"W) which was collected from a soil sample weathering out of a yellow-brown
limestone in the lower part of the roadway (Fig.
2
). The specimen came from sample WL3275A (GPS coordinates: 18°24'58.9"N
77°31'54.9"W) which was collected from a soil sample weathering out of a yellow-brown
limestone in the lower part of the roadway (Fig.
2 ![]() ). Mitchell et
al. (2022) showed that this was a limestone level developed within the
Preston Hill Formation (Yellow Limestone Group). In this locality Hanovolepidina
gen. nov. is associated with a foraminiferal assemblage including: Operculinoides
jennyi Barker, Nummulites
guayabalensis Barker, Helicostegina
gyralis Barker & Grimsdale, Pseudorlepidina
trimera, Linderina floridana Cole
and Caudriella sp., and can be
assigned to ABZ7 (mid-Lutetian) (Mitchell et
al., 2022; Özcan et al.,
2022).
). Mitchell et
al. (2022) showed that this was a limestone level developed within the
Preston Hill Formation (Yellow Limestone Group). In this locality Hanovolepidina
gen. nov. is associated with a foraminiferal assemblage including: Operculinoides
jennyi Barker, Nummulites
guayabalensis Barker, Helicostegina
gyralis Barker & Grimsdale, Pseudorlepidina
trimera, Linderina floridana Cole
and Caudriella sp., and can be
assigned to ABZ7 (mid-Lutetian) (Mitchell et
al., 2022; Özcan et al.,
2022).
|
Figure 2:
Geology of
the area around Swanswick House (based on unpublished map by SFM) in Trelawny,
Jamaica, showing location of sample WL3275A from which a specimen of Hanovolepidina
browni sp. nov. has been collected. YLph,
Preston Hill Formation (marlstones); WLm,
Montpelier Formation (deep water chalks); WL'sw',
Swanswick Formation sensu stricto; WLw,
Walderston Formation; WLbt,
Browns Town Formation. Alluvium omitted for clarity. |
Infrequent specimens of Hanovolepidina
browni sp. nov. are also found in the Chapelton Formation (sample WL4538;
GPS coordinates: 18°22'17.5"N
78°16'59.3"W) on a trail NW from
the road between Winchester and Phoenix Town, on the edge of the Green Island
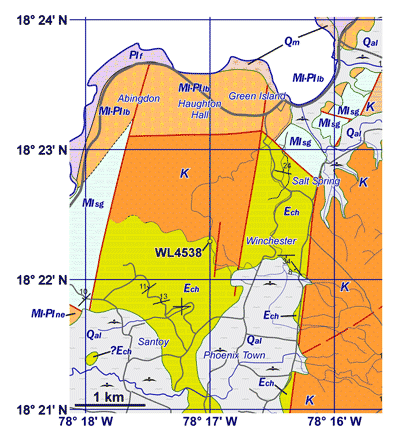
Cretaceous Inlier in the parish of Hanover (Fig. 3 ![]() ). Although abundant
foraminifers occur at this locality, the only other species occurring with Hanovolepidina
browni Mitchell sp. nov. are abundant specimens of O.
jennyi (Fig. 4.1
). Although abundant
foraminifers occur at this locality, the only other species occurring with Hanovolepidina
browni Mitchell sp. nov. are abundant specimens of O.
jennyi (Fig. 4.1 ![]() ) that represent more than 99% of the fauna, and a single
specimen of Planorbitoinella ecuadorensis Hofker (Fig.
4.2
) that represent more than 99% of the fauna, and a single
specimen of Planorbitoinella ecuadorensis Hofker (Fig.
4.2 ![]() ). This sample
is also assigned to ABZ7 largely due to the presence of Operculinoides
jennyi and P. ecuadorensis (Mitchell
et al., 2022). Further south in the
Chapelton Formation north of Phoenix Town, specimens of Yaberinella
cf. hottingeri Robinson are
common suggesting ABZ9-10 (late Lutetian). No calcareous nannofossil data is
available for these samples.
). This sample
is also assigned to ABZ7 largely due to the presence of Operculinoides
jennyi and P. ecuadorensis (Mitchell
et al., 2022). Further south in the
Chapelton Formation north of Phoenix Town, specimens of Yaberinella
cf. hottingeri Robinson are
common suggesting ABZ9-10 (late Lutetian). No calcareous nannofossil data is
available for these samples.
|
Figure 3: Geology
(based on unpublished map by SFM) of the area around Green Island in the parish
of Hanover, Jamaica, showing the location of sample WL4538 in the Chapelton
Formation that has yielded Hanovolepidina
browni sp. nov. K,
Cretaceous rocks in Green Island and Lucea inliers; Ech,
Chapelton Formation (Yellow Limestone Group); MIsg,
Spring Garden Formation of the White Limestone Group; MI-Pl,
various units in the Miocene-Pleistocene Coastal Group; MIf,
Falmouth Formation; Qal,
alluvium; Qm,
mangroves. |
|
Figure 4: Foraminifers
from sample WL4538, Chapelton Formation, south of the Green Island Inlier,
parish of Hanover, Jamaica. 1
(UWIGM.WL4538-13), Operculinoides jennyi
Barker. 2
(UWIGM.WL4538-9), Planorbitoinella
ecuadorensis (Hofker). 3-4
(UWIGM.WL4538-24), Hanovolepidina browni
sp. nov., holotype, external views: 3,
view of pustules on 'convex' side, 4,
view of pustules on 'flat' side. This assemblage (O. jennyi and P. ecuadorensis)
indicates ABZ7 (mid-Lutetian: Mitchell et al., 2022). |
Bulk samples of rock were collected in the field. The samples were disaggregated by washing in water through a 0.5 mm sieve. Larger Benthic Foraminifers (LBF) were then picked from the residues and stored in glass tubes. Specimens were sorted into morphotypes and thin sections were prepared to show orientated (equatorial and axial) sections that show taxonomically significant features. Thin sections were prepared by grinding using 600-grade carborundum grit on a glass plate, with specimens secured to a glass slide using thermoplastic cement to allow specimens to be orientated, and re‑orientated after heating, during sample preparation. The resulting sections were numbered with the sample number and a specimen number (e.g., WL4538-6); specimens are stored in the collections of the University of the West Indies Geological Museum (UWIGM), the national repository for geological samples in Jamaica. Specimens were photographed using a Hayear 4K HDMI Microscope Camera (for 3D specimens) and an AmScope T490B-10MA Digital Compound Trinocular Microscope (for thin sections).
The classification for the larger benthic foraminifers described here follows Loeblich & Tappan (1988) as amended by Mitchell et al. (2022). For details of taxonomic terms see Hottinger (2006) and Mitchell et al. (2022). Material is preserved in the University of the West Indies Geology Museum (the Jamaican repository of geological specimens).
Superfamily Asterigerinoidea Orbigny, 1839
Family Pseudolepidinidae Mitchell, E. Robinson & Özcan in Mitchell et al., 2022
Genus Hanovolepidina gen. nov.
Type species. Hanovolepidina browni sp. nov. from the Chapelton Formation (Lutetian) south of the Cretaceous Green Island Inlier, parish of Hanover, Jamaica.
Diagnosis. Test lenticular, with scattered piles (pustules) on the surface. Equatorial sections of megalospheric specimens show a bilocular embryo with a short spire (4 to 5 chambers, excluding the embryonic chambers), with the last chamber (R) having a retrovert aperture. A spiral and counter spiral of chambers develop from the R-chamber and terminate in a closing chamber. Equatorial chamberlets are developed from the spiral and counter spiral that develop from the R-chamber. Axial sections show that the equatorial layer is initially composed of a single layer of chamberlets, but at a short distance from the embryo it develops into a double layer of equatorial chamberlets that extends to the margin of the test. Axial sections show that most specimens have only a few lateral chambers, but one specimen has much better developed lateral chambers in multiple tiers.
Microspheric specimens are only marginally larger than megalospheric specimens and, in general, it is not possible to separate them unless they are sectioned. The single equatorial section of a microspheric specimen has a very small proloculus (which is not clearly visible) succeeded by about 2˝ whorls of rotaliid chambers), after which orbitoidal equatorial chamberlets are produced. A single axial section shows a largely lamellar wall structure containing scattered, vacuole-like lateral chamberlets, similar to most of the axial sections of megalospheric specimens.
Origin of name. From the parish of Hanover, Jamaica, where the largest population of specimens has been collected. Gender feminine. This represents the type locality.
Discussion. In equatorial section, Hanovolepidina resembles many early members of the Lepidocyclinidae and the Pseudolepidinidae. It is distinguished from all early lepidocyclinids (e.g., Eulinderina Barker & Grimsdale, Eolepidina Tan and Planorbitoinella Mitchell) in possessing a double equatorial layer that begins about half way through the test in axial section. Hanovolepidina gen. nov. is distinguished from Pseudolepidina by its possession of a short spire of chambers following the embryonic chambers; in contrast Pseudolepidina has two PACs, together with the additional 'PAC' which is seen in axial sections. This additional 'PAC' is absent in Hanovolepidina gen. nov. Triplalepidina differs in possessing two PACs and a bilaterally symmetrical embryo, as well as having a compact layer of shell material between the two equatorial layers of chamberlets. Cole (1963) followed Grimsdale (1959) in synonymising Triplalepidina with Lepidocyclina, and T. veracruziana Vaughan & Cole with L. pustulosa Douvillé. Cole (1963) used the stolon structure visible in the equatorial layers of L. ocalana (Cushman) and L. gubernacula Cole to justify this. L. ocalana has multiple stolons (two or three are indicated) between equatorial chamberlets in axial sections, whereas L. gubernacula shows as many as four to six stolons, and in axial sections the equatorial layer flares towards the test margin and develops several to multiple layers of equatorial chamberlets close to the margin of the test. Thus Cole (1963) considered that Triplalepidina was a junior synonym of Lepidocyclina. However, Triplalepidina, Hanovolepidina gen. nov. and Pseudolepidina all develop a double layer of equatorial chamberlets in the middle of the test, whereas in L. gubernacula multiple equatorial chamberlets develop only close to the test margin and no extended interval showing a double layer of equatorial chamberlets is present. The arrangement of equatorial chamberlets seen in L. gubernacula could be generated by the multiple stolons seen in L. ocalana giving rise to additional layers of equatorial chamberlets towards the test margin. However, there is clearly no phylogenetic connection between Pseudolepidina and Hanovolepidina gen. nov. (both are mid-Lutetian) and L. ocalana and L. gubernacula (Bartonian-Priabonian and Priabonian, respectively) and these two groups form separate radiations that are best placed in separate families (Mitchell et al., 2022). The morphology of Triplalepidina mirrors that of Hanovolepidina gen. nov. other than having an isolepidine embryo and it seems best to derive Triplalepidina from Hanovolepidina since both forms lack the additional 'PAC' seen in Pseudolepidina.
Hanovolepidina browni Mitchell sp. nov.
(Figs. 4.3-4 ![]() ,
5.1-6
,
5.1-6 ![]() ,
6.1-6
,
6.1-6 ![]() ,
7.1-4
,
7.1-4 ![]() ,
8.1-2
,
8.1-2 ![]() ,
9.1-3
,
9.1-3 ![]() )
)
2022 Pseudolepidina trimera Barker & Grimsdale; Mitchell et al., p. 478-484, Fig. 54.6 only.
Type material. Twenty specimens: holotype (UWIGM.WL.4538-24) and paratypes (UWIGM.WL4538-5, UWIGM.WL4538-6, UWIGM.WL4538-7, UWIGM.WL4538-8, UWIGM.WL4538-10, UWIGM.WL4538-11); all from the Chapelton Formation (sample WL4538; GPS coordinates: 18°22'17.5"N 78°16'59.3"W), south of the Cretaceous Green Island Inlier, parish of Hanover, Jamaica.
Origin of name. After Charles Barrington Brown who produced the first map of the parish of Hanover in 1867 (Sawkins, 1869).
Diagnosis. As for the genus.
|
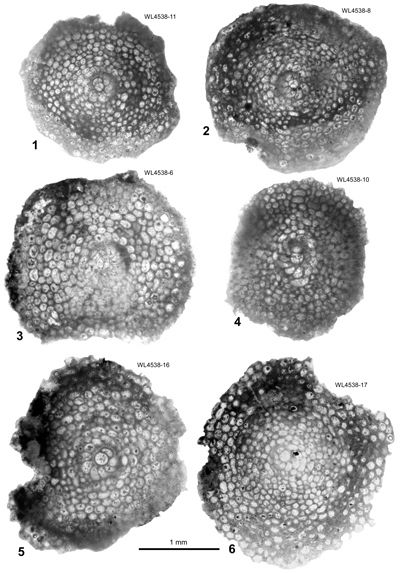
Figure 5: Hanovolepidina
browni sp. nov., megalospheric specimens in equatorial section, from the
Chapelton Formation (ABZ7, mid-Lutetian), south of the Green Island Inlier,
parish of Hanover, Jamaica: 1
(UWIGM.WL4538-11, paratype); 2
(UWIGM.WL4538-8, paratype); 3
(UWIGM.WL4538-6, paratype); 4
(UWIGM.WL4538-10, paratype); 5
(UWIGM.WL4538-16, paratype); 6
(UWIGM.WL4538-17, paratype). |
|
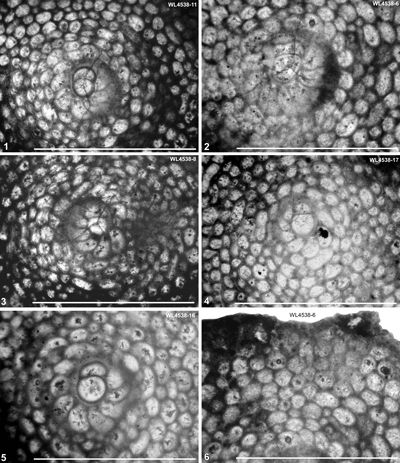
Figure 6:
Hanovolepidina browni
sp. nov. from the Chapelton Formation (ABZ7, mid-Lutetian), south of the Green
Island Inlier, parish of Hanover, Jamaica. 1-5: details of embryo and
rotaliid spire in equatorial sections: 1
(UWIGM.WL4538-11, paratype); 2
(UWIGM.WL4538-6, paratype); 3
(UWIGM.WL4538-8, paratype); 4
(UWIGM.WL4538-17, paratype); 5
(UWIGM.WL4538-16, paratype). 6, detail of equatorial chamberlets in
equatorial section (UWIGM.WL4538-6, paratype). |
Description. Both megalospheric and microspheric specimens occur, but only two microspheric specimens have been identified by sectioning. Megalospheric and microspheric specimens look similar in external view, are of relatively similar diameters, and can only be distinguished when sectioned.
Megalospheric
specimens (Figs. 4.3-4 ![]() ,
5.1-6
,
5.1-6 ![]() ,
6.1-6
,
6.1-6 ![]() ,
7.1-4
,
7.1-4 ![]() ,
8.1-2
,
8.1-2 ![]() ) are the same size or
slightly smaller than microspheric specimens with test diameters of up to 2 mm
and thicknesses of up to 1.1 mm. Orientated equatorial sections show an embryo
with a broadly spherical proloculus and a hemispherical deuteroloculus. The
proloculus has a length of 88-185 µm and a width of 95-177 µm
(excluding the walls). The deuteroloculus has a length of 49-144 µm and a
width of 68-162 µm (excluding the walls). The overall length of the embryo
is 166-305 µm (including the walls). Measurements of individual specimens
are given in Table 1. The embryo is followed by 4 to 5 chambers, with the last
chamber having a retrovert aperture and giving rise to a second spire (Figs.
6.1-3
) are the same size or
slightly smaller than microspheric specimens with test diameters of up to 2 mm
and thicknesses of up to 1.1 mm. Orientated equatorial sections show an embryo
with a broadly spherical proloculus and a hemispherical deuteroloculus. The
proloculus has a length of 88-185 µm and a width of 95-177 µm
(excluding the walls). The deuteroloculus has a length of 49-144 µm and a
width of 68-162 µm (excluding the walls). The overall length of the embryo
is 166-305 µm (including the walls). Measurements of individual specimens
are given in Table 1. The embryo is followed by 4 to 5 chambers, with the last
chamber having a retrovert aperture and giving rise to a second spire (Figs.
6.1-3 ![]() ,
7.1-2
,
7.1-2 ![]() ,
8.1-2
,
8.1-2 ![]() ). The initial spire has a rotaliid-like appearance with
arcuate septa, two spires of more semi-circular equatorial chambers develop from
R. A thickened wall may or may not be present in the primary spire up to R. The
two spires of equatorial chambers that develop from the R chamber coil around
the earlier chambers and end in a closing chamber. Arcuate equatorial
chamberlets develop from the primary and secondary spires giving rise to
orbitoidiform growth. Equatorial chamberlets are generally arranged in short
radial rows. Axial sections of megalospheric forms show that the embryo is
succeeded by a single row of equatorial chambers/chamberlets (chambers and
chamberlets are difficult to distinguish in axial sections). At a distance of
about half way towards the margin of the test the equatorial layer becomes
double, and continues as a double layer until the margin of the test.
Well-developed piles are seen in axial sections. The piles are inwardly conical
and rounded on the test surface with diameters of up to a maximum of 85 µm.
The development of lateral chamberlets is variable. Most axial sections show a
relatively compact laminar test with irregularly developed lateral chamberlets (Fig.
7.2-4
). The initial spire has a rotaliid-like appearance with
arcuate septa, two spires of more semi-circular equatorial chambers develop from
R. A thickened wall may or may not be present in the primary spire up to R. The
two spires of equatorial chambers that develop from the R chamber coil around
the earlier chambers and end in a closing chamber. Arcuate equatorial
chamberlets develop from the primary and secondary spires giving rise to
orbitoidiform growth. Equatorial chamberlets are generally arranged in short
radial rows. Axial sections of megalospheric forms show that the embryo is
succeeded by a single row of equatorial chambers/chamberlets (chambers and
chamberlets are difficult to distinguish in axial sections). At a distance of
about half way towards the margin of the test the equatorial layer becomes
double, and continues as a double layer until the margin of the test.
Well-developed piles are seen in axial sections. The piles are inwardly conical
and rounded on the test surface with diameters of up to a maximum of 85 µm.
The development of lateral chamberlets is variable. Most axial sections show a
relatively compact laminar test with irregularly developed lateral chamberlets (Fig.
7.2-4 ![]() ). One axial section
(Fig. 7.1
). One axial section
(Fig. 7.1 ![]() ) shows up to eight tiers of lateral
chamberlets over the centre of the test; the chamberlets are equal to or thinner
than their walls and floors; semi-regular tiers of lateral chamberlets are
developed in some areas, whereas in others they are more irregular.
) shows up to eight tiers of lateral
chamberlets over the centre of the test; the chamberlets are equal to or thinner
than their walls and floors; semi-regular tiers of lateral chamberlets are
developed in some areas, whereas in others they are more irregular.
|
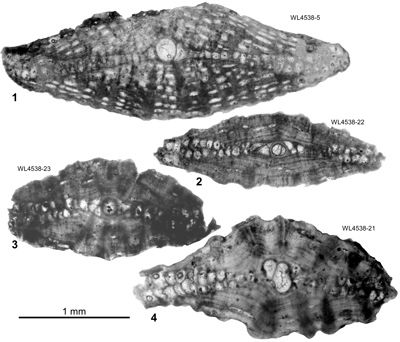
Figure 7: Axial
sections of megalospheric specimens of Hanovolepidina
browni sp. nov. 1
(UWIGM.WL4538-5), showing large numbers of lateral chamberlets. 2
(UWIGM.WL4538-22), 3
(UWIGM.WL4538-23) and 4
(UWIGM.WL4538-21) showing few vacuole-like lateral chamberlets. All from the
Chapelton Formation (ABZ7, mid-Lutetian), south of the Green Island Inlier,
parish of Hanover, Jamaica. |
|
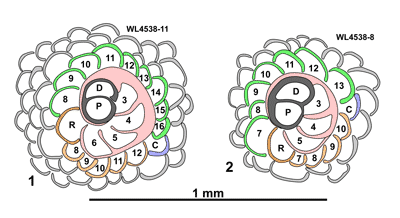
Figure 8: Hanovolepidina
browni sp. nov. from the Chapelton Formation (ABZ7, mid-Lutetian), south of
the Green Island Inlier, parish of Hanover, Jamaica. Drawings showing the
development of the equatorial chamberlets. 1
(UWIGM.WL.4538-11, paratype). 2
(UWIGM.WL.4538-8, paratype). P, proloculus; D, deuteroloculus; R, first chamber
with a retrovert aperture giving rise to a spiral and counter-spiral; C, closing
chamber. |
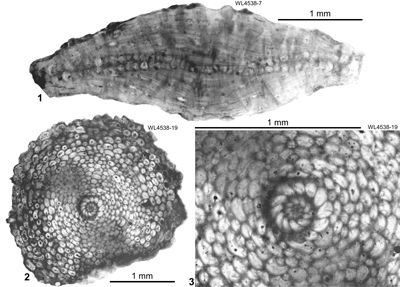
Microspheric specimens (Fig. 9 ![]() )
are lenticular with a diameter of up to 4 mm and a thickness of up to 1.2 mm and
carry an ornament consisting of piles similar to megalospheric specimens. A
single equatorial section (Fig. 9.2-3
)
are lenticular with a diameter of up to 4 mm and a thickness of up to 1.2 mm and
carry an ornament consisting of piles similar to megalospheric specimens. A
single equatorial section (Fig. 9.2-3 ![]() ) shows that the proloculus was small
(although it cannot be definitively seen) and was succeeded by about 2˝ whorls
of rotaliid chambers. Subsequently orbitoidal growth develops. The axial section
(Fig. 9.1
) shows that the proloculus was small
(although it cannot be definitively seen) and was succeeded by about 2˝ whorls
of rotaliid chambers. Subsequently orbitoidal growth develops. The axial section
(Fig. 9.1 ![]() ) shows that the equatorial layer begins initially as a single layer
that becomes double in the outer three-quarters of the radius of the test. The
lateral areas of the test are largely composed of thick-walled, compact, laminar
test material, with irregularly scattered narrow lenticular, widely-spaced
lateral chamberlets.
) shows that the equatorial layer begins initially as a single layer
that becomes double in the outer three-quarters of the radius of the test. The
lateral areas of the test are largely composed of thick-walled, compact, laminar
test material, with irregularly scattered narrow lenticular, widely-spaced
lateral chamberlets.
|
Figure 9: Microspheric
specimens of Hanovolepidina browni sp. nov. from the Chapelton Formation (ABZ7,
mid-Lutetian), south of the Green Island Inlier, parish of Hanover, Jamaica. 1
(UWIGM.WL4538-7), axial section (nearly central). 2-3
(UWIGM.WL4538-19), equatorial section. |
Discussion. The development of lateral chamberlets is variable in axial sections of macrospheric specimens of H. browni sp. nov., with most having relatively few chamberlets and others having larger numbers of chamberlets. The specimen with a large number of lateral chamberlets resembles the lateral chamberlets developed in Pseudolepidina trimera (Mitchell et al., 2022, Fig. 51.1, .3-8), but differs in lacking an additional PCA. The single axial section of a microspheric specimen of Pseudolepidina trimera figured by Mitchell et al. (2022, Fig. 51.2) also shows relatively few lateral chamberlets; it may be that microspheric specimens of Hanovolepidina gen. nov and Pseudolepidina are not possible to separate.
The measurements for the sizes of embryonic chambers and the embryo show a relatively wide degree of variation (Table 1). This is largely due to the presence of UWIGM.WL4538.18 which has unusually small chambers (the thin-section is poor and cannot be figured). If this specimen is ignored, there is less variation in the measurements, and this may represent a rare outlier.
Table 1: Measurements of embryonic chambers (µm) and R values for specimens of Hanovolepidina browni sp. nov. from Jamaica. Proloculus lengths (Pli) and widths (Pwi) and deuteroloculus lengths (Dli) and widths (dwi) exclude walls, whereas embryo length (Ele) includes the walls (see Mitchell, 2022, for details). R chamber includes the two chambers in the embryo.
| Number | Pli | Pwi | Dli | Dwi | Ele | R |
| WL4538-6 | 185 | 176 | 72 | 191 | 305 | 7 |
| WL4538-8 | 119 | 139 | 72 | 140 | 252 | 6 |
| WL4538-10 | 155 | 177 | 89 | 155 | 287 | 7 |
| WL4538-11 | 109 | 127 | 69 | 132 | 211 | 7 |
| WL4538-4 | 126 | 127 | 79 | 128 | 255 | ? |
| WL4538-16 | 167 | 173 | 75 | 162 | 300 | 7 |
| WL4538-20 | 116 | 141 | 144 | 135 | 300 | 6 |
| WL4538-17 | 155 | 177 | 89 | 155 | 287 | 7 |
| WL4538-18 | 88 | 92 | 49 | 68 | 166 | ? |
| Mean | 136 | 148 | 82 | 141 | 263 | |
| Stand. Dev. | 31.4 | 30.1 | 26.1 | 34.4 | 47.4 | |
| WL3275A-18 | 187 | 213 | 117 | 189 | 385 | 4? |
The recognition of a new genus of the Pseudolepidinidae in the mid-Lutetian suggests that there were two different lineages, one including Hanovolepidina gen. nov. that could have evolved into Triplalepidina, and a second lineage including Pseudolepidina. In Jamaica and Mexico, specimens belonging to the family only turn up at distinct stratigraphic intervals (mid-Lutetian for Hanovolepidina Mitchell gen. nov. and Pseudolepidina, and one sample from the ?Priabonian for Triplalepidina) and the appearance of these forms may represent migration events (perhaps from eastern Pacific guyots or accreted east pacific volcanic islands) that allowed faunal exchange, but not the establishment of long-lived evolving lineages. Only further discoveries will allow the unravelling of the history of this little known family of orbitoidiform LBF.
SFM developed the project, collected the material from Jamaica, thin sectioned the LBF, and developed the taxonomy. The main text was prepared by SFM.
I thank Roshaun Brown and Jullian Williams for support in the field collecting samples. Thanks to Johannes Pignatti and an anonymous reviewer for comments that helped improve the original manuscript. Thanks to the editor Bruno Granier for his editorial comment.
Adams C.G. (1987).- On the classification of the Lepidocyclinidae (Foraminiferida) with redescriptions of the unrelated Paleocene genera Actinosiphon and Orbitosiphon.- Micropaleontology, New York - NY, vol. 33, no. 4, p. 289-317.
Barker R.W. & Grimsdale T.F. (1937).- Studies of Mexican fossil Foraminifera.- The Annals and Magazine of Natural History (ser. 10), London, vol. 19, 161-174 (Pls. 5-9).
Cole W.S. (1963).- Illustrations of the conflicting interpretations of the biology and classification of certain larger Foraminifera.- Bulletins of American Paleontology, vol. 46, p. 6-63. URL: https://www.biodiversitylibrary.org/item/40515#page/9/mode/1up
Grimsdale T.F. (1959).- Evolution in the American Lepidocyclinidae (Cainozoic Foraminifcra): An interim review. I-II.- Procedings Koninklijke Nederlandse Akadamie Wetenschappen (ser. B), Amsterdam, vol. 62, no. 1, p. 8-33 (2 text-figs.).
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, no. M02 (CG2006_M02), p. 1-126. DOI: 10.4267/2042/5832
Loeblich A.R. Jr & Tappan H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold Co., New York - NY, 730 p.
Mitchell S.F. (2004).- Lithostratigraphy and palaeogeography of the White Limestone Group. In: Donovan S.K. (Ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Rotterdam, vol. 3, p. 5-29. https://natuurtijdschriften.nl/pub/541700/CR2004003001002.pdf
Mitchell S.F. (2013a).- Stratigraphy of the White Limestone of Jamaica.- Bulletin de la Société Géologique de France, Paris, vol. 184, no. 1-2, p. 111-118.
Mitchell S.F. (2013b).- Lithostratigraphy of the Central Inlier, Jamaica.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 46, p. 31-42. URL: http://caribjes.com/CJESpdf/CJES46-03-MitchellCentral%20InlieR.pdf
Mitchell S.F. (2016).- Geology of the western margin of the Benbow Inlier - implications for the relationship between the Yellow Limestone and White Limestone groups (with the description of the Litchfield Formation, new name).- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 48, p. 19-25. URL: http://caribjes.com/CJESpdf/CJES48-2-Mitchell2016WhiteLimestoneUnconformity.pdf
Mitchell S.F. (2021).- The first Paleogene transgression onto the Clarendon Block (Jamaica).- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 53, p. 1-10. URL: http://caribjes.com/CJESpdf/CJES53-1-Mitchell-Bottom-Leinster-Inlier.pdf
Mitchell S.F., Robinson E., Özcan E., Jiang M.M. & Robinson N. (2022).- A larger benthic foraminiferal zonation for the Eocene of the Caribbean and central American region.- Carnets Geol., Madrid, vol. 22, no. 11, p. 409-565. DOI: 10.2110/carnets.2022.2211
Orbigny A.D. d' (1839).- Foraminifčres. In: Sagra R. de la (Ed.), Histoire physique, politique et naturelle de l'ile de Cuba.- Arthus Bertrand, Paris, 224 p. URL: https://gallica.bnf.fr/ark:/12148/bpt6k975884/f1.item
Özcan E., Yücel A.O., Mitchell S.F., Pignatti J., Simmons M.D., Okay A.I., Erkızan L.S. & Gültekin M.N. (2022).- New records of Caudriella Haman and Huddleston from the middle and late Eocene of Neo-Tethys: Taxonomic and paleobiogeographic implications.- Journal of Foraminiferal Research, Lawrence - KS, vol. 52, p. 21-39.
Robinson E. (1994).- Jamaica. In: Donovan S.K. & Jackson T.A. (Eds.), Caribbean geology an introduction.- University of the West Indies Publisher's Association, Kingston, Jamaica, p. 111-127. URL: http://www.redciencia.cu/geobiblio/paper/1994_K.Donovan_Trevor%20A.%20Jackson_Caribbean%20Geology.pdf
Robinson E. & Mitchell S.F. (1999).- Middle Eocene to Oligocene stratigraphy and palaeogeography in Jamaica: A window on the Nicaragua Rise.- Contributions to Geology, UWI, Mona, Kingston, Jamaica, no. 4, 47 p. URL: http://www.sfmgeology.com/pdfs/MiddleEocenetoOligoceneStratigraphy.pdf
Sawkins J.G. (1869).- Reports on the geology of Jamaica; or, Part II. of the West Indian survey.- Memoirs of the Geological Survey, London, 339 p. URL: https://archive.org/details/reportsongeolog00hoffgoog/page/n43/mode/2up
Scheffen W. (1932).- Zur Morphologie und Morphogenese der "Lepidocyclinen".- Paläontologische Zeitschrift, vol. 14, p. 233-256.
Vaughan T.W. & Cole W.S. (1938).- Triplalepidina veracruziana, a new genus and species of orbitoidal Foraminifera from the Eocene of Mexico.- Journal of Paleontology, Tulsa - OK, vol. 12, p. 167-169.
Zans V.A., Chubb L.J., Versey H.R., Williams J.B., Robinson E. & Cooke D.L. (1963).- Synopsis of the geology of Jamaica an explanation of the 1958 provisional geological map of Jamaica.- Bulletin, Geological Survey Department, Jamaica, Kingston, no. 4, 72 p. [Dated on front cover and title page 1962, printers imprint at bottom of front page is 1963].
| Nomenclatural note: Life Sciences Identifier (LSID) https://zoobank.org/References/da894b08-dc9a-401f-94ef-40ffafde39b8 Genus Group Species Group |