◄ Carnets Geol. 25 (4) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Systematic palaeontology]
[5. Palaeoecological comments]
[6. Conclusions]
and ...
[Bibliographic references]
Lerchenauerstr. 167, 80935 Munich (Germany)
Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, East Beijing Road 39, Nanjing 210008 (China)
College of Tourism, Henan Normal University, Xinxiang 45300 (China)
Published online in final form (pdf) on February 3, 2025
DOI 10.2110/carnets.2025.2504
![]()
[Editor: George E. PleŞ; technical editor: Bruno R.C. Granier]
![]()
Few stratigraphical and micropalaeontological data are available from Upper Cretaceous shallow-water marine carbonates of the southwestern Tarim Basin, western China. The present study provides new data about the microfauna and calcareous algae of the Cenomanian-Turonian Kukebai Formation and the Campanian Yigeziya Formation. The investigated bioclastic carbonates yield a rather poorly diversified microfauna (benthic foraminifers) contrasting with a relatively well diversified assemblage of dasycladalean green algae comprising 16 taxa. For several taxa, the available material does only allow for an open taxonomic treatment. A new species is described herein as Morelletpora sinica. These calcareous algae are supplemented by debris of halimedacean algae (Arabicodium? sp.). Occurrences of calcareous algae and larger benthic foraminifers in Cenomanian and Campanian carbonates coincide with two major transgressions reported from the Tarim Basin. The observed faunal and phycological elements indicate a direct connection between the Neotethys and the Tarim epicontinental sea. The lack of several typical Cenomanian (e.g., alveolinoids, orbitolinids) and Campanian larger benthic foraminifers in the studied material is striking.
• green algae;
• Late Cretaceous;
•
taxonomy;
• biostratigraphy;
• palaeobiogeography
Schlagintweit F., Xu Y. & Zhang S. (2025).- Calcareous green algae (Dasycladales, Halimedaceae) from the Upper Cretaceous of the western Tarim Basin, NW China: Systematic palaeontology, microfacies, and palaeobiogeographic significance.- Carnets Geol., Madrid, vol. 25, no. 4, p. 89-108. DOI: 10.2110/carnets.2025.2504
Algues vertes calcaires (Dasycladales, Halimedaceae) du Crétacé supérieur de l'ouest du bassin du Tarim, nord-ouest de la Chine : Paléontologie systématique, microfacičs et signification paléobiogéographique.- Peu de données stratigraphiques et micropaléontologiques sont disponibles concernant les carbonates marins peu profonds du Crétacé supérieur du sud-ouest du bassin du Tarim, dans l'ouest de la Chine. Cette étude présente de nouvelles données sur la microfaune et les algues calcaires des formations de Kukebai (Cénomanien-Turonien) et de Yigeziya (Campanien). Les carbonates bioclastiques étudiés montrent une microfaune peu diversifiée (foraminifčres benthiques),, contrastant avec une association relativement diversifiée d'algues vertes dasycladales comprenant 16 taxons, parmi lesquels une nouvelle espčce, Morelletpora sinica, est décrite. Cependant, en raison du matériel disponible, certains taxons sont laissés en nomenclature ouverte. Cette association d'algues calcaires est complétée par des débris d'algues halimédacées (Arabicodium? sp.). Il est ŕ noter que la présence d'algues calcaires et de grands foraminifčres benthiques dans les carbonates cénomaniens et campaniens coďncide avec deux grandes transgressions signalées dans le bassin du Tarim. Les éléments microfauniques et phycologiques observés indiquent une connexion directe entre la Néotéthys et la mer épicontinentale du Tarim. L'absence de plusieurs grands foraminifčres benthiques typiques du Cénomanien (par exemple, les alvéolinidés et les orbitolinidés) et du Campanien dans le matériel étudié est remarquable.
• algues vertes ;
• Crétacé supérieur
;
•
taxinomie ;
• biostratigraphie ;
• paléobiogeographie
The Tarim Basin in western China represents the eastern end (or gulf) of an epicontinental sea
extending from the Mediterranean Neotethys across the eastern Eurasian continent
during Late Cretaceous and Palaeogene times (e.g., Hao & Zeng,
1984) (Fig.
1.1 ![]() ). It is also
referred to as Proto-Paratethys in Paleogene times (see Kaya,
2020, for
details). Being the largest sedimentary basin in China (about 530.000 km2),
it represents an important area for the study of Upper Cretaceous marine strata
in China (e.g., Yang et
al., 1983; Xi et al., 2016,
2019: Fig. 2; Zhang et al.,
2018). Most biostratigraphic data such as dinoflagellate cysts have been
obtained from the marly lithologies (e.g., Zhang et
al., 2022). Data about micropalaeontogical assemblages from thin-sections of
the Upper Cretaceous marine carbonates of the southwestern Tarim Basin are rare.
In his paper about 'Late Cretaceous palaeobiogeography of calcareous algae',
Mu (1993: Table 1) reported several genera of calcareous algae (including
Dasycladales) from the Tarim Basin referring to unpublished data that, according
to our knowledge, still remain unpublished. Recently, larger benthic foraminifers were described by Sun and Schlagintweit
(2024) from
the Campanian Yigeziya Formation. Our paper provides the first description and
illustration of calcareous green algae (mainly Dasycladales) from the Kukebai
and Yigeziya formations. New data about the micropalaeontological inventory
including also some benthic foraminifers, the palaeoenvironment, and the
biostratigraphy as well as the palaeobiogeographical relevance of the
foram-algal assemblages are presented.
). It is also
referred to as Proto-Paratethys in Paleogene times (see Kaya,
2020, for
details). Being the largest sedimentary basin in China (about 530.000 km2),
it represents an important area for the study of Upper Cretaceous marine strata
in China (e.g., Yang et
al., 1983; Xi et al., 2016,
2019: Fig. 2; Zhang et al.,
2018). Most biostratigraphic data such as dinoflagellate cysts have been
obtained from the marly lithologies (e.g., Zhang et
al., 2022). Data about micropalaeontogical assemblages from thin-sections of
the Upper Cretaceous marine carbonates of the southwestern Tarim Basin are rare.
In his paper about 'Late Cretaceous palaeobiogeography of calcareous algae',
Mu (1993: Table 1) reported several genera of calcareous algae (including
Dasycladales) from the Tarim Basin referring to unpublished data that, according
to our knowledge, still remain unpublished. Recently, larger benthic foraminifers were described by Sun and Schlagintweit
(2024) from
the Campanian Yigeziya Formation. Our paper provides the first description and
illustration of calcareous green algae (mainly Dasycladales) from the Kukebai
and Yigeziya formations. New data about the micropalaeontological inventory
including also some benthic foraminifers, the palaeoenvironment, and the
biostratigraphy as well as the palaeobiogeographical relevance of the
foram-algal assemblages are presented.
|
|
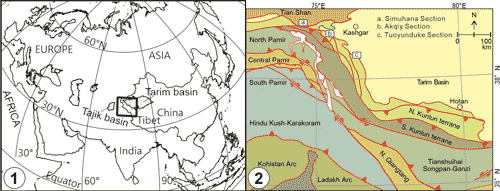
Figure 1: (1) Location of the Tarim Basin and
(2) simplified tectonic map of the Pamir collision zone showing locations of the three studied sections (modified from
Cowgill,
2010, and Zhang
et al., 2018: Fig. 1). |
The study area is located in the western Tarim
Basin (Xinjiang Uygur Autonomous Region, China), bounded by the Tianshan range
to the north, the Pamir to the west-southwest, and the West Kunlun range to the
south (Fig. 1 ![]() ). Due
to the heterogeneity of inherited paleogeomorphology, the Upper Cretaceous
strata in the western Kunlun-Pamir piedmont and the Tianshan piedmont are
different. The Upper Cretaceous strata in the western Kunlun-Pamir piedmont are
better developed, also yielding abundant and diversified marine fossils. The
marine-transitional Upper Cretaceous sequences, defined as the Yingjisha Group,
are composed of the Kukebai, Wuyitake, Yigeziya, and Tuyiluoke formations from
bottom to top (Yang et al., 1983; Tang et al.,
1989, 1992; Hao
et al., 2001; Xi et
al., 2019). Lithostratigraphically, the Kukebai Formation (Cenomanian-Turonian) is dominated by gray-green argillites,
muddy siltstone and bioclastic limestones, interbedded with gypsum and
dolostones (~240
m). The Wuyitake Formation (Coniacian-Santonian) consists of orange-red argillites, gypsiferous argillites, and
siltstones (~50 m). The Yigeziya Formation (Campanian-?early Maastrichtian) is
dominated by white and purple-red bioclastic limestones (~110 m). The Tuyiluoke
Formation ((late) Maastrichtian-Danian) in turn consists of orange-red
argillites and gypsiferous argillites (~20 m). These
formations are attributed to at least two transgressive (Kukebai and Yigeziya
formations) - regressive (Wuyitak and Tuyiluok formations) cycles (Guo et
al., 2015; Zhang et al.,
2018). The algal assemblages described here from three
sections are from carbonate rocks belonging to the two transgressive cycles of
the Kukebai and Yigeziya formations.
). Due
to the heterogeneity of inherited paleogeomorphology, the Upper Cretaceous
strata in the western Kunlun-Pamir piedmont and the Tianshan piedmont are
different. The Upper Cretaceous strata in the western Kunlun-Pamir piedmont are
better developed, also yielding abundant and diversified marine fossils. The
marine-transitional Upper Cretaceous sequences, defined as the Yingjisha Group,
are composed of the Kukebai, Wuyitake, Yigeziya, and Tuyiluoke formations from
bottom to top (Yang et al., 1983; Tang et al.,
1989, 1992; Hao
et al., 2001; Xi et
al., 2019). Lithostratigraphically, the Kukebai Formation (Cenomanian-Turonian) is dominated by gray-green argillites,
muddy siltstone and bioclastic limestones, interbedded with gypsum and
dolostones (~240
m). The Wuyitake Formation (Coniacian-Santonian) consists of orange-red argillites, gypsiferous argillites, and
siltstones (~50 m). The Yigeziya Formation (Campanian-?early Maastrichtian) is
dominated by white and purple-red bioclastic limestones (~110 m). The Tuyiluoke
Formation ((late) Maastrichtian-Danian) in turn consists of orange-red
argillites and gypsiferous argillites (~20 m). These
formations are attributed to at least two transgressive (Kukebai and Yigeziya
formations) - regressive (Wuyitak and Tuyiluok formations) cycles (Guo et
al., 2015; Zhang et al.,
2018). The algal assemblages described here from three
sections are from carbonate rocks belonging to the two transgressive cycles of
the Kukebai and Yigeziya formations.
The Kukebai Fm. was established by the No. 107 Geological Team under the Xinjiang Petroleum Administration in 1976. The type section is located about 20 km west of Ulugqat in the Wuqia County of Xinjiang. It is subdivided either into lower and upper members with the first one attaining greater thickness (Yang et al., 1983) or a tripartite: lower, middle, upper units (e.g., Guo et al., 2015; Xi et al., 2016). Lithologically, the Kukebai Formation represents a series of mixed siliciclastic-carbonatic sediments. Macrofossils from the lower Kukebai Formation include mainly gastropods (Pan, 1990) and rudists (Scott et al., 2010), while the microfossils consist of benthic foraminifers (Hao et al., 2001) and nannoplankton, as well as dinoflagellates from marly lithologies indicating a Cenomanian-Turonian age (Yang et al., 1983; He, 1991; Zhong, 1992; Zhang et al., 2022). Recent isotope analyses also confirmed the presence of the Cenomanian-Turonian boundary in the Kukebai Formation (Zhang et al., 2025).
The Yigeziya Formation was established and defined by the No. 107 team of the department of geological survey under the Xinjiang Petroleum Administration in the year 1976. The type section is located near Yi Geziya about 20 km south of Yingisar county, Xinjiang. It consists mainly of massive neritic carbonates (e.g., Zhang et al., 2022). Macrofossils of the Yigeziya Formation are represented by rudists (X. Rao, 2019; Lan & Wei, 1995; X. Rao et al., 2023), brachiopods, echinoids, and gastropods (Pan, 1990). Among microfossils, dinoflagellates and acritarchs have repeatedly been described (Shaozhi & Norris, 1988; He, 1991; Zhong, 1992; Zhang et al., 2022). Calcareous algae were reported in open nomenclature and without illustration by Mu (1993), but also small benthic foraminifers obtained from washed samples (Hao et al., 2001). The age of the Yigeziya Formation is generally assigned to the Campanian-early Maastrichtian (e.g., Xi et al., 2019: Fig. 7). Recently, the early Maastrichtian age was reinstated referring to the occurrence of the said 'lower Maastrichtian index taxon' Biradiolites boldjuanensis in the upper part of the formation (X. Rao et al., 2023; Gao et al., 2024). In our opinion, the early Maastrichtian age of the upper part of the Yigeziya Formation - though deducible by this rudist species - needs to be reconfirmed by other biogenic groups such as larger benthic foraminifers.
The foraminiferal-algal assemblages described here are from three outcrops:
Simuhana, Akqiy and Tuoyundoke sections (Fig. 1.2 ![]() ). The systematic
part refers essentially to the algae. In the following, some basic information
on each locality is given, comprising the logs (Figs. 2
). The systematic
part refers essentially to the algae. In the following, some basic information
on each locality is given, comprising the logs (Figs. 2 ![]() - 3
- 3 ![]() ), the microfacies of
the algal-bearing samples (Fig. 4
), the microfacies of
the algal-bearing samples (Fig. 4 ![]() ), and some of associated benthic foraminifers
(Fig. 5
), and some of associated benthic foraminifers
(Fig. 5 ![]() ).
).
|
|
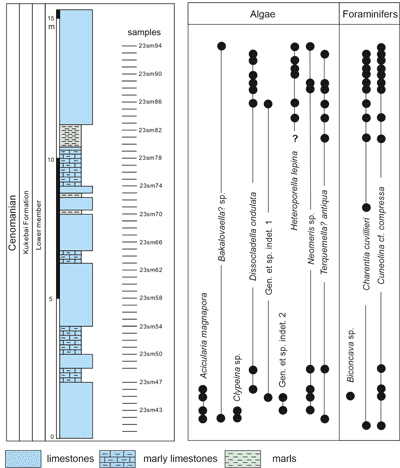
Figure 2: Log
of the lower Kukebai Formation of the Simuhana section showing vertical
distribution of calcareous algae and some benthic foraminifers. |
|
|
Figure 3: Logs
of the Akqiy (1) and Tuoyundoke sections (2) showing vertical distribution of
calcareous algae and some benthic foraminifers (modified from Zhang et
al., 2018: Figs. 8 left side, 9 left side). |
|
|
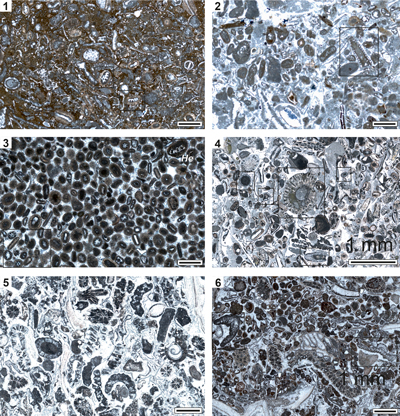
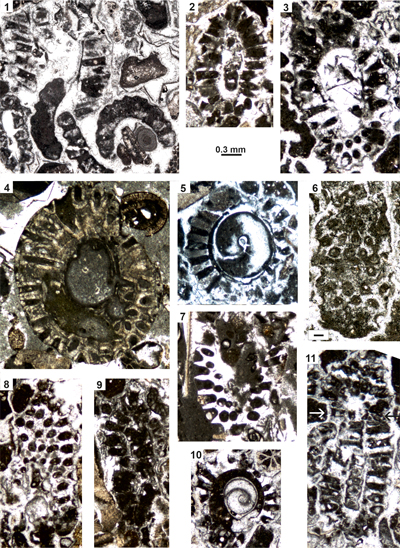
Figure 4:
Microfacies of the algal-bearing samples (examples) from the Upper Cretaceous
Kukebai (1-4) and Yigeziya formations (5-6) of the southwestern Tarim Basin,
China. (1) Packstone with common debris of
dasycladaleans showing oolitic envelopes (rectangles);
Cenomanian of Simuhana section, sample sm43. (2) Packstone with rare
debris of dasycladaleans (rectangle), ooids and a few benthic foraminifers such
as Charentia cuvillieri Neumann
(Ch); Cenomanian of Simuhana section,
sample sm86. (3) Oolitic grain-packstone with dasycladales (Terquemella antiqua (Pia), Heteroporella lepina Praturlon
(He) as core of ooids (black rectangles); Cenomanian of Simuhana
section, sample sm84. (4) Bioclastic packstone with ooids, bryozoans, pelecypod
shells, and rare dasycladaleans (e.g., Morelletpora sinica sp.
nov. in the center) and halimedaceans (rectangles); Turonian? of Akqiy
section, sample AK28. (5) Bioclastic grain-packstone with bryozoans,
pelecypod shells and various sections of Morelletpora
sinica sp. nov.; Campanian of Tuoyunduke
section, sample TY31. (6) Bioclastic packstone with ooids, bryozoans and debris
of halimedaceans (rectangles); Campanian of Tuoyunduke section, sample TY37.
Scale bars 1.0 mm. |
|
|
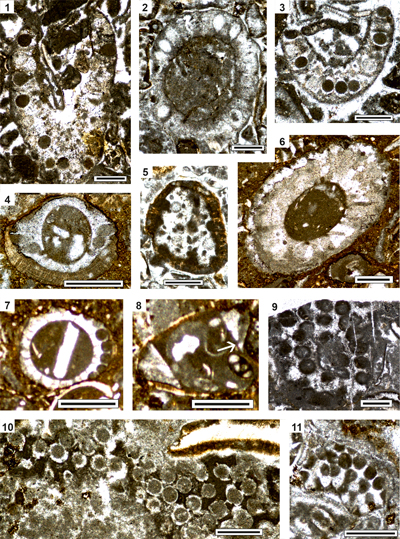
Figure 5:
Benthic foraminifers from the Cenomanian Kukebai
Formation of the Simuhana section, (1-14), Cenomanian-pro parte Turonian? Kukebai
Formation of the Akqiy
section (15, 17-18, 20) and the Campanian Yigeziya Formation of the Tuoyunduke
section (16, 19), SW Tarim Basin, China. (1-5, 8-9) Cuneolina compressa Schlagintweit; transverse sections (1, 5,
8), oblique longitudinal section (2), longitudinal section parallel to plane of
biseriality (3), longitudinal section perpendicular to plane of biseriality (4).
(6, 10-12, 15) Charentia cuvillieri Neumann;
axial section (6), oblique section (10), subaxial sections (11: juvenile
specimen showing oolitic envelope; 15), equatorial section (12). (7) Unknown
planispirally coiled taxon in equatorial section (Biconcava? sp.). (13-14) Comaliamma
sp.; subaxial section (13), oblique section (14). (16, 19) Dictyopselloides? tarimensis
Sun & Schlagintweit; subaxial sections; possible microspheric
specimen in 19. (17, 20) Nezzazatinella
picardi (Henson); oblique sections. (18) Quinqueloculina sp. (right) and Murgeina
apula (Luperto Sinni) (Mu).
Scale bars = 0.30 mm. Samples: sm41 (1-5), sm88 (6), sm45 (7, 9), sm91 (8, 12),
sm84 (10), sm42 (11, 13-14), AK80 (15), TY65 (16), AK58 (17-18, 20), TY31 (19). |
2.1 Kukebai Formation of the Simuhana section
The studied locality is close to the Simuhana 1 section
studied by Xi et al. (2016)
named after the nearby Simuhana village. The GPS coordinates of the section are 39°43'29.335"N, 73°59'34.799"E. The herein investigated carbonates
correspond to "bed six" and "bed eight" in the subdivision of the
section by Xi et al. (2016:
Fig. 6) separated by an intercalation of calcareous shales with abundant
ostracods ("bed seven"). These bioclastic limestones with ooids were
illustrated by Xi et al. (2016:
Fig. 5B) but no microfossils were indicated (Fig. 4.1-4.3 ![]() ). Besides
dasycladalean algae, these carbonates also yielded a poorly diversified
assemblage of benthic foraminifers with Charentia
cuvillieri Neumann, Cuneolina
compressa Schlagintweit, Comaliamma
sp., miliolids, and some not further specified planispirally coiled taxa (Figs. 2
). Besides
dasycladalean algae, these carbonates also yielded a poorly diversified
assemblage of benthic foraminifers with Charentia
cuvillieri Neumann, Cuneolina
compressa Schlagintweit, Comaliamma
sp., miliolids, and some not further specified planispirally coiled taxa (Figs. 2 ![]() ,
5.1-5.14
,
5.1-5.14 ![]() ).
).
2.2 Kukebai Formation of the Akqiy section
The Akqiy section is located about 100 km west of Kashgar City (39°20'20.98"N, 74°56'31.42"E) and has been studied by Zhang et
al. (2018: Fig. 8 left column) (Fig. 3.1 ![]() ). The name refers to the Akqiy
Village, Piaoertuokuoyi Town, Wuqia County, in the Xinjiang Uygur Autonomous
Region. It is located in the hanging wall of the Main Pamir Thrust. The
algal-bearing sample (AK28) represents a bioclastic oolitic packstone with common
to abundant ooids, bryozoans, gastropods and dasycladales (Acicularia
macropora Kuss, one specimen of Morelletpora
sinica sp. nov.) (Figs. 2
). The name refers to the Akqiy
Village, Piaoertuokuoyi Town, Wuqia County, in the Xinjiang Uygur Autonomous
Region. It is located in the hanging wall of the Main Pamir Thrust. The
algal-bearing sample (AK28) represents a bioclastic oolitic packstone with common
to abundant ooids, bryozoans, gastropods and dasycladales (Acicularia
macropora Kuss, one specimen of Morelletpora
sinica sp. nov.) (Figs. 2 ![]() ,
4.4
,
4.4 ![]() ). This
microfacies compares to the type strata of M.
sinica sp. nov. from the Yigeziya
Formation (Fig. 4.5
). This
microfacies compares to the type strata of M.
sinica sp. nov. from the Yigeziya
Formation (Fig. 4.5 ![]() ). The sample is from the upper part of the Kukebai Formation
and might be of Turonian age although direct evidence is missing (see Zhang
et al., 2018: Fig. 8 left column).
). The sample is from the upper part of the Kukebai Formation
and might be of Turonian age although direct evidence is missing (see Zhang
et al., 2018: Fig. 8 left column).
2.3 Yigeziya Formation of the Tuoyunduke section
The Tuoyunduke section is located about 100 kilometers
south of Kashgar City (coordinates: 38°32'00.78"N,
76°16'00.22"E),
and has been examined by Zhang et
al. (2018: Fig. 9, left column) (Fig. 3.2 ![]() ). The section name refers to the
nearby Toyunduk Village and refers to the Tuyiluoke Formation which overlies the
Yigeziya Formation. The Yigeziya Formation in this section is ~87 meters thick
and is characterized by purple-red and grayish bioclastic wackestones/packstones
and bioclastic grainstones. The bioclasts include bivalves, rudists, benthic
foraminifers, crinoids, bryozoans, debris of coralline algae and gastropods.
Laminoid fenestrae and biolaminae are prevalent. The depositional environments
primarily comprising tidal flats, shoal to mid-ramp deposits. The algal-bearing
samples (TY31, TY32, TY37) comprise high-energy open-marine bioclastic
packstones with scattered ooids, common to abundant bryozoans, pelecypod debris
(rudists?), rare gastropods, some benthic foraminifers (e.g., miliolids, Minouxia
sp., Dictyopselloides? tarimensis
Sun & Schlagintweit, Orbitolinidae indet. of Sun & Schlagintweit,
2024: Fig. 7a), and dasycladalean algae (Morelletpora
sinica sp. nov.) (Figs. 4.5
). The section name refers to the
nearby Toyunduk Village and refers to the Tuyiluoke Formation which overlies the
Yigeziya Formation. The Yigeziya Formation in this section is ~87 meters thick
and is characterized by purple-red and grayish bioclastic wackestones/packstones
and bioclastic grainstones. The bioclasts include bivalves, rudists, benthic
foraminifers, crinoids, bryozoans, debris of coralline algae and gastropods.
Laminoid fenestrae and biolaminae are prevalent. The depositional environments
primarily comprising tidal flats, shoal to mid-ramp deposits. The algal-bearing
samples (TY31, TY32, TY37) comprise high-energy open-marine bioclastic
packstones with scattered ooids, common to abundant bryozoans, pelecypod debris
(rudists?), rare gastropods, some benthic foraminifers (e.g., miliolids, Minouxia
sp., Dictyopselloides? tarimensis
Sun & Schlagintweit, Orbitolinidae indet. of Sun & Schlagintweit,
2024: Fig. 7a), and dasycladalean algae (Morelletpora
sinica sp. nov.) (Figs. 4.5 ![]() ,
5.16, 5.19
,
5.16, 5.19 ![]() ). Another variety of this microfacies type (sample TY36) almost completely
lacks the dasycladales element, instead exhibiting common debris of halimedacean
algae (Boueina? sp., Arabicodium?
sp.) (Fig. 4.6
). Another variety of this microfacies type (sample TY36) almost completely
lacks the dasycladales element, instead exhibiting common debris of halimedacean
algae (Boueina? sp., Arabicodium?
sp.) (Fig. 4.6 ![]() ). It is worth mentioning that the Yigeziya Formation of the Akqiy
Formation (see §2.2) also yields inner ramp (tidal flat?) facies with abundant
miliolids and Nezzazatinella picardi (Henson)
but lacks calcareous green algae (Figs. 5.17-5.18
). It is worth mentioning that the Yigeziya Formation of the Akqiy
Formation (see §2.2) also yields inner ramp (tidal flat?) facies with abundant
miliolids and Nezzazatinella picardi (Henson)
but lacks calcareous green algae (Figs. 5.17-5.18 ![]() ). The occurrence of Murgeina
apula (Luperto Sinni) (Fig. 5.18
). The occurrence of Murgeina
apula (Luperto Sinni) (Fig. 5.18 ![]() ) is worth mentioning since it
represents the first record from the Upper Cretaceous carbonates of the Tarim
Basin and at the same time the most eastern occurrence within the Neotethys
realm (Schlagintweit et al., 2023; Simmons & Bidgood,
2023: Fig. 57). M.
apula is indicative of a relatively wide age-range comprising the late early
Cenomanian to early Maastrichtian and thus of no real biostratigraphic
importance. Like several other taxa of (larger) benthic foraminifers, M. apula is also present in Campanian strata of the Tielongtan Group
of the Western Kunlun Mountains (Sha et al., 2020; Sun & Schlagintweit,
2024).
) is worth mentioning since it
represents the first record from the Upper Cretaceous carbonates of the Tarim
Basin and at the same time the most eastern occurrence within the Neotethys
realm (Schlagintweit et al., 2023; Simmons & Bidgood,
2023: Fig. 57). M.
apula is indicative of a relatively wide age-range comprising the late early
Cenomanian to early Maastrichtian and thus of no real biostratigraphic
importance. Like several other taxa of (larger) benthic foraminifers, M. apula is also present in Campanian strata of the Tielongtan Group
of the Western Kunlun Mountains (Sha et al., 2020; Sun & Schlagintweit,
2024).
The described calcareous algae are recorded in 13 thin-sections; 9 from the Kukebai Formation and 4 from the Yigeziya Formation (Table 1). The sample number prefixes are 16AK (Akgiy section), 23sm (Simuhana section) and 15TY (Tuoyunduke section). They are stored at the State Key Laboratory of Mineral Deposits Research, School of Earth Sciences and Engineering, Nanjing University, Nanjing 210023, China.
Table 1: Studied samples and localities studied.
| Localities | Simuhana section | Akqiy section | Tuoyunduke section |
| Reference | Xi et al., 2016 | Zhang et al., 2018 | Zhang et al., 2018 |
| Samples (thin-sections) | sm43 | AK28 | TY31 |
| sm45 | TY32 | ||
| sm46 | TY36 | ||
| sm84 | TY37 | ||
| sm86 | |||
| sm89 | |||
| sm90 | |||
| sm94 | |||
| Total number | 8 | 1 | 4 |
The described
taxa of calcareous algae include green algae of the orders Dasycladales Pascher,
1931, and
Bryopsidales Schaffner, 1922 (Figs. 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() - 9
- 9 ![]() ). Some taxa are treated in open nomenclature (genus
level) and also as gen. and sp. indet. in the cases where the available sections
do not allow a final conclusion about the taxonomic status. The suprageneric classification of the Dasycladales is adopted from Granier
& Bucur in Granier et al.
(2013a). For the biometric data, the standard abbreviations of Bassoullet
et al. (1975) have been used: The abbreviations used are as follows: D = outer thallus diameter, d =
inner thallus diameter, h = vertical spacing of the verticils, l = length
primary laterals, l' = length secondary laterals, l" = length tertiary
laterals, p = diameter primary laterals, p' = diameter secondary laterals,
p" = diameter tertiary laterals, w = number of laterals per verticil. The
occurrences of the individual taxa described from the three localities are
compiled in Table 2.
). Some taxa are treated in open nomenclature (genus
level) and also as gen. and sp. indet. in the cases where the available sections
do not allow a final conclusion about the taxonomic status. The suprageneric classification of the Dasycladales is adopted from Granier
& Bucur in Granier et al.
(2013a). For the biometric data, the standard abbreviations of Bassoullet
et al. (1975) have been used: The abbreviations used are as follows: D = outer thallus diameter, d =
inner thallus diameter, h = vertical spacing of the verticils, l = length
primary laterals, l' = length secondary laterals, l" = length tertiary
laterals, p = diameter primary laterals, p' = diameter secondary laterals,
p" = diameter tertiary laterals, w = number of laterals per verticil. The
occurrences of the individual taxa described from the three localities are
compiled in Table 2.
Table 2: Inventory of dasycladalean taxa in the Kukebai and Yigeziya formations of the southwestern Tarim Basin, China.
| taxa | Kukebai Fm. Cenomanian | Kukebai Fm. ?Turonian | Yigeziya Fm. Campanian |
| Neomeris sp. | X | ||
| Cymopolia sp. | X | ||
| Dissocladella ondulata | X | ||
| Trinocladus? sp. | X | ||
| Morelletpora sinica sp. nov. | X | X | |
| Heteroporella lepina | X | ||
| Bakalovaella? sp. | X | ||
| Acicularia magnapora | X | ||
| Terquemella? antiqua | X | X | |
| Clypeina sp. | X | ||
| Gen. and sp. indet. 1 | X | ||
| Gen. and sp. indet. 2 | X | ||
| Gen. and sp. indet. 3 | X | ||
| Gen. and sp. indet. 4 | X | ||
| Gen. and sp. indet. 5 | X | ||
| Gen. and sp. indet. 6 | X | ||
| Number | 9 | 1 | 8 |
Order Dasycladales Pascher, 1931
Family Dasycladaceae (Kützing, 1843)
Tribe Dasycladeae (Pia, 1920)
Genus Bakalovaella Bucur, 1993
Type species: Cylindroporella elitzae Bakalova, 1971
Bakalovaella? sp.
Remarks: Two oblique sections are here tentatively assigned to Bakalovaella Bucur, 1993, that has recently been revised by Granier and Bucur (2019). The generic reservation results from the verified occurrence of sterile laterals, otherwise the sections could then belong to the genus Holosporella Pia, 1930.
Measurements: D: 0.35-0.64 mm; d: 0.07 mm; h: 0.09 mm; Diameter fertile ampullae: 0.07 mm / 0.11 mm.
Occurrence: Very rare in the Kukebai Formation of the Simuhana section.
Genus Neomeris Lamouroux, 1816
Type species: Neomeris dumetosa Lamouroux, 1816
Neomeris sp.
Remarks: The total diameter (D)
of the thalli varies between 0.80 mm and 1.10 mm while the inner diameter (d)
ranges from 0.49 mm to 0.6 mm. The latter value however only comprises the
preserved calcified part not referable to the main axis that is only seldom
preserved in Neomeris (e.g.,
Granier et al., 2013a). Here,
one possible specimen of Neomeris
shows the preserved main axis and inner portion of the laterals (D: 0.98 mm; d:
0.44 mm leading to a d/D ratio of 0.45) (Fig. 6.6 ![]() ). The subspherical fertile
ampullae range from 0.11 mm to 0.13 mm in diameter. We refrain from any specific
assignments noting the difficulty of addressing all features of thin-section
material especially from lower-mid Cretaceous Neomerideae in general and also
due to the relatively poor material available from the Kukebai Formation.
). The subspherical fertile
ampullae range from 0.11 mm to 0.13 mm in diameter. We refrain from any specific
assignments noting the difficulty of addressing all features of thin-section
material especially from lower-mid Cretaceous Neomerideae in general and also
due to the relatively poor material available from the Kukebai Formation.
Occurrence: Common in the Cenomanian Kukebai Formation of the Simuhana section.
|
|
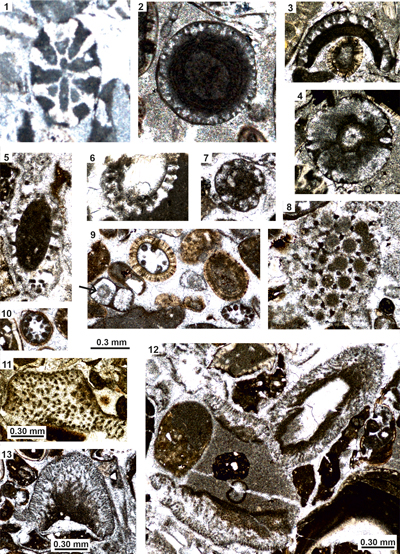
Figure 6:
Dasycladalean algae from the Cenomanian of the lower Kukebai Formation of the
Simuhana section, SW Tarim Basin, China. (1-3, 5, ?6) Neomeris sp. (4, 7-8) Clypeina
sp. (arrow in 8: Connection of lateral with the main axis). (9-11) Gen. and sp.
indet. 1. Scale bars = 0.30 mm. Samples sm94 (1), sm45 (2, 5, 8, 10-11), sm90
(3), sm43 (4, 6-7), sm86 (9). |
Genus Cymopolia Lamouroux, 1816
Remarks: One tangential section of ovoid outline is considered here as
belonging to Cymopolia Lamouroux
(Fig. 9.8 ![]() ). The diameter of the fertile ampullae is
approximately 0.10 mm in average.
Another possible fragment consists of two fertile ampullae (Fig.
9.9
). The diameter of the fertile ampullae is
approximately 0.10 mm in average.
Another possible fragment consists of two fertile ampullae (Fig.
9.9 ![]() ). The
microfacies refers to high-energy deposits as shown in Figure
4.5
). The
microfacies refers to high-energy deposits as shown in Figure
4.5 ![]() .
.
Occurrence: Rare in the Campanian Yigeziya Formation of the Tuoyundoke section.
Family Triploporellaceae (Pia, 1920)
Tribe Dissocladelleae Elliott, 1977
Genus Dissocladella Pia, 1936
Type species: Dissocladella savitriae L.R. Rao & Pia, 1936
Dissocladella ondulata (Raineri, 1922) Pia, 1936
(Fig. 7.9-7.11, 7.12 pars, 7.13-7.14 ![]() )
)
1922 Neomeris cretacea var. ondulata var. nov. - Raineri, p. 75, Pl. 3, figs. 5-11.
1936 Dissocladella undulata (Raineri) - Pia, p. 18, Figs. 12-16.
2005 Dissocladella bonardii n.sp. - Radoičić et al., p. 313, Pl. 1, figs. 4-17 (with synonymy).
2017 Dissocladella ondulata - Granier et al., Fig. 1B-F.
2024 Dissocladella ondulata (Raineri) - Schlagintweit et al., p. 494, Fig. 4A-F.
Remarks: Thalli of D. ondulata represent a common element in oolitic packstones (partly grainstones) associated with rare Bakalovaella? sp. and Neomeris sp., as well as some bryozoans. Benthic foraminifers are represented by a few specimens of Charentia cuvillieri and Cuneolina compressa. For discussion about taxonomic problems associated with the Libyan type material of Dissocladella ondulata see Radoičić et al. (2005) and Granier et al. (2017). D. ondulata is known from Cenomanian to Santonian strata (Barattolo, 2002).
Measurements: D: 0.34-0.45 mm; D: 0.15-0.26 mm; d/D: 0.40-0.65; p: 0.04-0.05 mm; p': 0.03-0.04 mm; w: 8 (1 value); h: 0.07 mm (1 value); Lmax: ~2.0 mm.
Occurrences: Common in the Kukebai Formation of the Simuhana section.
|
|
Figure 7:
Dasycladalean algae from the Cenomanian of the lower Kukebai Formation of the
Simuhana section, SW Tarim Basin, China. (1-3)
Heteroporella lepina (Praturlon);
debris in grain-packstones with ooids, bryozoans (br), Charentia cuvillieri Neumann (Ch), and Neomeris sp. (Ne)
(1-2),
and as core of ooids (3). (4-8)
Acicularia macropora Kuss. (9-14,
17) Dissocladella ondulata (Raineri)
together with Bakalovaella? sp. (in
12). (15-16) Gen. and sp. indet. 2. Scale bars
= 0.30 mm. Samples: sm89 (1-2), sm84 (3), sm45 (4, 7), sm43 (5-6, 8, 15-16),
sm86 (9, 11, 13-14, 17), sm46 (10), sm90 (12). |
Tribe Triploporelleae Pia, 1920
Genus Trinocladus Raineri, 1922
Trinocladus sp.
(Fig. 9.4 ![]() )
)
Remarks: One transverse section (D = 0.70 mm, d = 0.25 mm, d/D = 0.36, w = ?7) with presumably three orders of laterals. The inner part of the skeleton shows micritization thereby masking the primary laterals. Trinocladus sp. occurs in high-energy oolitic shoal facies associated with gen. and sp. indet. 3, 4, and Morelletpora sinica sp. nov. and bryozoans.
Occurrence: Very rare in the Turonian? Kukebai Formation of the Akqiy section.
Tribe Morelletporeae Varma, 1950
Genus Morelletpora Varma, 1950
Type species: Morelletpora nammalensis Varma, 1950
Morelletpora sinica sp. nov.
Origin of the name: Named from the Latin 'Sinica' for 'from China, Chinese'.
Holotype: Fig. 8.11 ![]() , thin-section TY31.
, thin-section TY31.
Type locality: Tuoyunduke section (Fig. 3 ![]() right).
right).
Type stratum: Campanian Yigeziya Formation.
Description: Elongated to club-shaped segments occasionally in connection
and marked by an incision along the longitudinal axis (Fig.
8.11 ![]() ). The
individual segments have a length of 1.8 mm to 3.5 mm and a ratio length/width
of ~1.7 to ~2.4. Calcification includes the main axis and the laterals. The
common type of preservation is represented by a mass of homogeneous sparry (whitish)
calcite. This type appears to be of secondary origin ('secondary collective
sheaths', De Castro, 1997) while the primary type shows a calcification
with an individual calcareous envelope of light-brownish colour covering each
lateral (Fig. 8.4
). The
individual segments have a length of 1.8 mm to 3.5 mm and a ratio length/width
of ~1.7 to ~2.4. Calcification includes the main axis and the laterals. The
common type of preservation is represented by a mass of homogeneous sparry (whitish)
calcite. This type appears to be of secondary origin ('secondary collective
sheaths', De Castro, 1997) while the primary type shows a calcification
with an individual calcareous envelope of light-brownish colour covering each
lateral (Fig. 8.4 ![]() ) ('individual
sheaths', De Castro, 1997). The
laterals are arranged perpendicular to slightly oblique towards the longitudinal
axis and in alternating position (Fig. 8.6
) ('individual
sheaths', De Castro, 1997). The
laterals are arranged perpendicular to slightly oblique towards the longitudinal
axis and in alternating position (Fig. 8.6 ![]() ). Proximally, the laterals are rather
narrow, tubular and connected to the main axis by a pore. Along their length,
they widen continuously and become more enhanced and funnel-like towards the
periphery.
). Proximally, the laterals are rather
narrow, tubular and connected to the main axis by a pore. Along their length,
they widen continuously and become more enhanced and funnel-like towards the
periphery.
Measurements: D: 1.25-2.20 mm (mostly between 1.40 and 1.50 mm); d: 0.64-0.81 mm (approximately 0.70 mm in average); d/D: 0.42-0.56; l: 0.37-0.80 mm; p: 0.15-0.17 mm; h: 0.19-0.23 mm; w: ~15 to ~25.
Comparisons: The genus Morelletpora Varma is known from the Aptian to the Thanetian (Barattolo, 2002; Bucur et al., 2016) including the three species M. nammalensis (Varma, 1950), M. turgida (Radoičić, 1975, non 1965) Barattolo, 2002 (Cenomanian of Croatia), and M. dienii Parente, 1997 (Maastrichtian of Italy). M. nammalensis from the Paleocene of Pakistan shows comparable dimensions, but differs from M. sinica significantly by the shape of the laterals with a rather long and thin tubular part that widens abruptly forming a 'sporangium' with a distal rounded end, well discernible in the specimen shown in Varma (1955: Pl. 1, fig. 2) and his thallus reconstruction of Figs. 18-20 therein. Unfortunately, the specimens from the Yigeziya Formation of the Tarim Basin do not show exactly the morphology of the distal lateral end (widening outwards or forming a rounded end within the calcareous sheeth. In any case, also the size of the segments is different with a length of 4-5 mm (or more?) in M. nammalensis. Based on the biometric data from rich material of Iran (Bucur et al., 2016) M. turgida from can be distinguished from M. sinica by some of its morphometric parameters such as a reduced main axis (d/D about 0.33), or a reduced whorl spacing (h mean 0.136 mm). Also the segment morphology is different; barrel-shaped in M. turgida versus elongated club-shaped in M. sinica. M. turgida was originally described from the Cenomanian of Serbia but has a rather wide distribution in Aptian-Albian strata (see synonymy in Bucur et al., 2016). M. dienii was originally described from the Maastrichtian of southern Italy. This species also shows barrel- to slightly pear-shaped segments with an axial cavity morphologically similar to the external shape. M. dienii has a comparably high number of laterals per whorl (w = 34-41) and a reduced whorl spacing (h = 0.07-0.12 mm).
Occurrence: Turonian? Kukebai Formation of the Akqiy section (rare); Campanian Yigeziya Formation of the Tuoyunduke section (common to abundant).
|
|
Figure 8: Morelletpora sinica sp. nov. from the
Turonian? Kukebai Formation of the Akqiy
section (4) and the Campanian Yigeziya Formation of the Tuoyunduke
section (all others), SW Tarim Basin, China. (1) Debris of diverse sections;
sample TY31. (9, 11) Longitudinal sections; note the incision along the thallus
axis of the holotype (11) marking the conjunction between two segments (arrows);
samples TY32 and TY31. (2-4, 6-7) Oblique sections; samples TY32: 2, 7, TY31: 3,
6; AK28: 4). (5, 10) Slightly oblique transverse sections; sample TY31. (8)
Tangential section; sample TY32. Scale bars = 0.30 mm. Samples: TY31 (1, 3, 5-6,
10-11) TY32 (2, 7-9), AK28 (4). |
Family Thyrsoporellaceae Granier & Bucur in Granier et al., 2013b
Genus Heteroporella (Praturlon, 1967) Granier et al., 1994
Type species: Heteroporella lepina Praturlon, 1967
Heteroporella lepina (Praturlon, 1967) Granier et al., 1994
(Fig. 7.1 pars, 7.2 pars, 7.3 ![]() ).
).
1967 Heteroporella lepina n.gen., n.sp. - Praturlon, p. 202, Pls. 51-52.
1990 Heteroporella lepina Praturlon - Gušić & Jelaska, Pl. 3, fig. 1; Pl. 7, fig. 5.
1992 Heteroporella lepina Praturlon - Schlagintweit, Pl. 2, fig. 10.
1994 Heteroporella lepina Praturlon - Granier et al., p. 129, Pl. 1, figs. 1-8 (with synonymy).
1999 Heteroporella lepina Praturlon - Ensslin & Schlagintweit, p. 478, Pl. 1, figs. 1-3, 5.
2000 Heteroporella lepina Praturlon - J.-P. Masse & Isintak, p. 370, Pl. 1, figs. 7-8.
Remarks: The debris of H. lepina, typically showing the yellowish calcification (Granier,
2012, for details), has been observed as cores of individual ooids in oolitic grainstones (Fig. 7.3 ![]() ) and together with bryozoans and some benthic foraminifers
(e.g., Charentia
cuvillieri) in grainstones-packstones. The fragmentary state of preservation
is not only due to the high-energy facies but also the modular thallus
organization of the alga consisting of individual rhombohedral shields
facilitating breakage (Granier et
al., 1994: Fig. 1). Heteroporella lepina is typically recorded from open lagoonal external
platform facies and occurs also resedimented in slope carbonates (Praturlon,
1967).
) and together with bryozoans and some benthic foraminifers
(e.g., Charentia
cuvillieri) in grainstones-packstones. The fragmentary state of preservation
is not only due to the high-energy facies but also the modular thallus
organization of the alga consisting of individual rhombohedral shields
facilitating breakage (Granier et
al., 1994: Fig. 1). Heteroporella lepina is typically recorded from open lagoonal external
platform facies and occurs also resedimented in slope carbonates (Praturlon,
1967).
Occurrence: Rare to common in the Kukebai Formation of the Simhuana section.
Family Polyphysaceae Kützing, 1843
Forma genus Acicularia Archiac, 1843
Type species: Acicularia pavantina Archiac, 1843
Acicularia cf. macropora Kuss, 1994
(Fig. 7.4-7.8 ![]() )
)
1994 Acicularia macropora n. sp. - Kuss, p. 304, Fig. 6, Pl. 3, figs. 12-16.
Remarks: Large-sized, elongate, often bended spicules
displaying well calcification. The shape of the spicules is often club-shaped
with a thicker rounded and a narrower tapering end (Fig. 7.5 ![]() ). In transverse
section there are 7 to 8 globular sporangial cavities. A. macropora was described by Kuss
(1994) from the upper
Turonian of Egypt. The specimens from the Cenomanian Kukebai Formation display
larger dimensions than the Egyptian type material while other parameters (number
of sporangial cavities) are equal. These differences are here not considered as
being of specific importance.
). In transverse
section there are 7 to 8 globular sporangial cavities. A. macropora was described by Kuss
(1994) from the upper
Turonian of Egypt. The specimens from the Cenomanian Kukebai Formation display
larger dimensions than the Egyptian type material while other parameters (number
of sporangial cavities) are equal. These differences are here not considered as
being of specific importance.
Dimensions (data from Kuss, 1994 in brackets): Length of spicules: up to 1.8 mm (0.8 mm to 1.2 mm); Diameter of spicules 0.40-0.67 mm (0.32-0.48 mm); Diameter of sporangial cavities: 0.14-0.16 mm (0.10-0.14 mm); Number of sporangial cavities: 7-8 (7-8).
Occurrence: Common in the Kukebai Formation of the Simuhana section.
Forma genus Terquemella Munier-Chalmas ex. L. & J. Morellet, 1913
Type species: Terquemella parisiensis Munier-Chalmas ex. L. & J. Morellet, 1913.
Terquemella? antiqua (Pia, 1936)
1936 Acicularia antiqua sp. nov. - Pia, p. 8, Pl. 3, figs. 1-14.
Remarks: Small-sized spiculae (diameter 0.25-0.45 mm) with numerous tiny
sporangial cavities (diameter 0.035-0.045 mm). Occurring in high-energy shoal
facies, the spicules often occur as ooid cores (Fig. 9.9 ![]() ). The forma species is
treated as possibly belonging to Terquemella
(Pia et al., 1937, p. 232)
while others retain it in the original description (Barattolo,
2002). T.?
antiqua is given a general Upper Cretaceous age (Cenomanian-Maastrichtian; Granier
& Deloffre, 1993).
). The forma species is
treated as possibly belonging to Terquemella
(Pia et al., 1937, p. 232)
while others retain it in the original description (Barattolo,
2002). T.?
antiqua is given a general Upper Cretaceous age (Cenomanian-Maastrichtian; Granier
& Deloffre, 1993).
Occurrences: Common in the Kukebai Formation of the Simuhana section and the Yigeziya Formation of the Tuoyunduke section.
|
|
Figure 9:
Dasycladales and Bryopsidales from the Turonian? Kukebai Formation of the
Simuhana section (1-4) and the Campanian Yigeziya Formation of the Tuoyunduke
section (5-12), SW Tarim Basin, China. (1) Gen. and sp. indet. 3. (2-3) Gen. and
sp. indet. 4. (4) Trinocladus
sp. (5-6) Gen et sp. indet. 5. (7) Gen. and sp. indet. 6. (8) Cymopolia?
sp. (9) Terquemella? antiqua (Pia)
(middle and right below) and Cymopolia?
sp. (left below; arrow = fertile ampulla). (11-13) Debris of Arabicodium
sp. Scale bars = 0.30 mm. Samples: AK28 (1-4), TY32 (5-6), TY31 (7), TY36 (8,
12), TY37 (9-11, 13). |
Genus Clypeina Michelin, 1845
Clypeina sp.
Description: The thallus debris consists of isolated
verticils characterized by a large main axis bearing numerous relatively short
tubular (= uncompressed) laterals. They arise from the main axis by means of a
small stalk then widening abruptly and thereby bending
upwards (Fig. 6.8 ![]() ).
).
Remarks: Among the Upper Cretaceous Clypeina species, the form from the Tarim Basin is differentiated from those bearing laterals arranged perpendicular (horizontal) to the axis such as C.? alrawii Radoičić, 1978, C. pastriki Radoičić, 1983, or C. dusanbrstinai Radoičić, 1997. Upper Cretaceous species with inclined laterals include C. croatica Gušić in Deloffre & Radoičić, 1978, and C. sahnii Varma, 1952. The poor material from the Cenomanian Kukebai Formation does not allow more detailed comparisons.
Measurements: D: 0.63-0.88 mm; d: 0.28-0.38 mm; d/D: 0.43-0.64; pmax: 0.064-0.070 mm; w: about 26. These biometric data are based on three specimens only. The outer diameter may be larger than in the preserved fragments and should therefore be considered as tentative.
Occurrence: Rare to common in the Cenomanian Kukebai Formation of the Simuhana section.
Taxa of unknown genera and species
Gen. and sp. indet. 1
Remarks: Thallus fragments showing close set laterals or ampullae (diameter 0.05-0.15 mm) in an alternating position. We note similarities to tangential sections of Brasiliporella nkossaensis (P. Masse, 1995) emend. Granier et al. (2013b) described from Albian-Cenomanian strata of Congo and Brazil (e.g., Granier et al., 2013b: Fig. 5e).
Occurrence: Rare to common in the Kukebai Formation of the Simuhana section.
Gen. and sp. indet. 2
Remarks: One oblique section (D = ~0.82 mm, d = ~0.3 mm) showing laterals of at least two orders. The swollen first order laterals have a diameter of 0.085-0.11 mm; the second order laterals have a diameter of ~0.043 mm. The uncertainty about the number of orders of laterals (two or three) does not allow a detailed discussion. We also note resemblance to the oblique section of Fourcadella sintraensis described by Granier and Berthou (2002: Pl. 1, fig. 4) from the Albian?-Cenomanian of Portugal.
Occurrence: Very rare in the Cenomanian Kukebai Formation of the Simuhana section.
Gen. and sp. indet. 3
(Fig. 9.1 ![]() )
)
Remarks: One oblique section of a well calcified dasycladalean alga exhibiting a narrow main axis (D = 056 mm, d = ~0.08 mm). There is presumably just one order of trumpet-shaped laterals of relatively low number within a whorl (less than 10).
Occurrence: Very rare in the Turonian? Kukebai Formation of the Akqiy section.
Gen. and sp. indet. 4
(Fig. 9.2-9.3 ![]() )
)
Remarks: One complete transverse section and a fragment of a slightly oblique transverse section (D = ~0.78). The alga shows a rather wide central hollow (filled with micrite) that most likely does not correspond to the main axis thus reflecting a secondary preservational stage. There are at least two orders of laterals in high numbers (>20). The material from the Campanian of the Tarim Basin shows some similarities to the Paleocene Dissocladella savitriae Pia in L.R. Rao & Pia, 1936, described from the Paleocene of India.
Occurrence: Rare to common in the Turonian? Kukebai Formation of the Akqiy section.
Gen. and sp. indet. 5
(Fig. 9.5-9.6 ![]() )
)
Remarks: Two oblique sections of a dasycladalean algae (D = 0.45-0.55 mm, d = 0.26-0.29, d/D = 0.53-0.58) of unknown orders of laterals (presumably just one).
Occurrence: Rare in the Yigeziya Formation of the Tuoyunduke section.
Gen. and sp. indet. 6
(Fig. 9.7 ![]() )
)
Remarks: One slightly oblique transverse section (D = 0.41 mm, d = 0.22 mm, d/D = 0.54) with unknown shape and order(s) of laterals. Gen. and sp. indet. 6 occurs in high-energy microfacies with debris of halimedaceans, Terquemella? antiqua (Pia), and bryozoans.
Occurrence: Rare in the Yigeziya Formation of the Tuoyunduke section.
Order Bryopsidales Schaffner, 1922
Family Halimedaceae Link, 1831
Genus Arabicodium Elliott, 1957
Arabicodium sp.
The comparable size of the medullary and cortical filaments discernible in the debris material suggests assignment to the genus Arabicodium Elliott, 1957 (e.g., Bucur, 1994). We refrain from assigning the debris material recovered here to any of the described species (see Table 1 in Bucur, 1994).
Occurrences: Common in the Cenomanian-Turonian Kukebai Formation of the Simuhana and Aksay sections and common to abundant in the Campanian Yigeziya Formation.
Mu (1993) reported 'an important algal flora of Late Cretaceous age' (...) that 'has been found from western Tarim Basin, NW China (ca. 40° N) which is of significance for the delineation of the northern boundary of the Tethyan Realm in Asia. This flora is rather diverse, composed of about 33 species in 22 genera, belonging to the Dasycladales, Halimedaceae, Corallinaceae, Solenoporaceae, Gymnocodiaceae, Peyssonneliaceae, Cyanophyta and other taxa, showing close Tethyan affinities' (op. cit., p. 339). Unfortunately, details of this data have never been published so that the actual assemblages and the lithostratigraphc occurrences (formations) are unknown. Mu (1993: Table 1) only listed genera such as Acicularia, Clypeina, Heteroporella, Neomeris, Salpingoporella, Tarimporella, and Terquemella. Tarimporella reported by Mu (1993) has to be regarded as nomen nudum lacking a taxonomic description in an official publication. It is noteworthy that an identically-named genus has been established decades later by Jiang and Zhang (2022) from the Cambrian of the Tarim Basin as a dasycladalean alga. The poorly preserved material however does not in our opinion allow an accurate description justifying the erection of a new genus. Moreover, the debris of 'Tarimporella' does not refer to dasycladales but halimedaceans. The other genera reported by Mu (1993) are well represented in our material. Alltogether, 16 different taxa of dasycladalean algae are reported herein, including 9 from the Cenomanian and 1 from the Turonian? Kukebai Formation, and 8 from the Campanian Yigeziya Formation with 2 shared taxa (Table 2).
Cenomanian shallow-water carbonates from Neotethys are typically
characterized by highly diverse assemblages of small and larger benthic
foraminifers (Schroeder & Neumann,
1985; Chiocchini et
al. 2012; Simmons & Bidgood,
2023). In fact, with respect to the Phanerozoic diversity of
agglutinated foraminiferal taxa, the Cenomanian stands alongside the Holocene as
the stage with the maximum number of reported taxa (Kaminski et
al., 2008; Schlagintweit & Yazdi-Moghadam,
2023:
'Cenomanian megadiversity'). In contrast hereto, the microfauna recovered
from the lower Kukebai Formation of the Tarim Basin can be considered as
extremely low diversity (Fig. 5 ![]() ). In particular the lack of any typical larger
benthic foraminifers (e.g., alveolinoids, orbitolinids) is striking. The
greatest diversity reported from Cenomanian carbonates as reported above however
is attained in inner platform facies while at least the upper part of the lower
Kukebai Formation containing echinoids, bryozoans, and Heteroporella
lepina (e.g., samples 23sm86 to 94, Fig. 2
). In particular the lack of any typical larger
benthic foraminifers (e.g., alveolinoids, orbitolinids) is striking. The
greatest diversity reported from Cenomanian carbonates as reported above however
is attained in inner platform facies while at least the upper part of the lower
Kukebai Formation containing echinoids, bryozoans, and Heteroporella
lepina (e.g., samples 23sm86 to 94, Fig. 2 ![]() ) can be ascribed to open
marine shoal facies that usually contains only a low diversity microfauna. The
Cenomanian calcareous algae of the Tarim Basin with at least 9 different taxa,
however, can be considered as well diversified against the background of that
from Cenomanian strata elsewhere, where only a reduced number of taxa were
described (Bassoullet et al., 1978; Bucur,
1999; Barattolo, 2002: twenty species; Aguirre
& Riding, 2005). In their analysis of biodiversity through time, Aguirre
and Riding (2005, p. 581) concluded that palaeotemperature and sea-level
were the main influencing factors meaning that
'Dasycladaleans were most diverse when their main habitats—warm shallow
seas—were most extensive'. In fact, the epicontinental Tarim
Gulf must have been a very warm shallow sea evidenced by the occurrences of
gypsum and dolostones in the Upper Cretaceous strata including the Kukebai
Formation (e.g., Xi et al., 2016; Zhang et al.,
2018). Also the calcareous algae of the Yigeziya Formation with at least
8 different taxa can be considered as comparably well diversified bearing in mind
that also from the Campanian a relatively low number of species is known
worldwide (e.g., seven species in Barattolo et al.,
2002). Another
reason might be the poor inventory of Upper Cretaceous limestones and the
related lack of petrographic thin sections.
) can be ascribed to open
marine shoal facies that usually contains only a low diversity microfauna. The
Cenomanian calcareous algae of the Tarim Basin with at least 9 different taxa,
however, can be considered as well diversified against the background of that
from Cenomanian strata elsewhere, where only a reduced number of taxa were
described (Bassoullet et al., 1978; Bucur,
1999; Barattolo, 2002: twenty species; Aguirre
& Riding, 2005). In their analysis of biodiversity through time, Aguirre
and Riding (2005, p. 581) concluded that palaeotemperature and sea-level
were the main influencing factors meaning that
'Dasycladaleans were most diverse when their main habitats—warm shallow
seas—were most extensive'. In fact, the epicontinental Tarim
Gulf must have been a very warm shallow sea evidenced by the occurrences of
gypsum and dolostones in the Upper Cretaceous strata including the Kukebai
Formation (e.g., Xi et al., 2016; Zhang et al.,
2018). Also the calcareous algae of the Yigeziya Formation with at least
8 different taxa can be considered as comparably well diversified bearing in mind
that also from the Campanian a relatively low number of species is known
worldwide (e.g., seven species in Barattolo et al.,
2002). Another
reason might be the poor inventory of Upper Cretaceous limestones and the
related lack of petrographic thin sections.
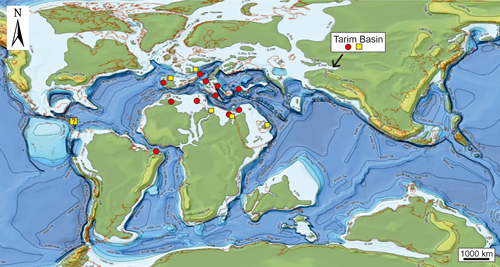
The calcareous algae
from the Upper Cretaceous Tarim Basin compares favourably with other general
Neotethys assemblages, exemplified by the distribution of Heteroporella
lepina and Dissocladella ondulata (Fig. 10 ![]() ).
).
|
|
Figure 10:
Global palaeogeographic map during the Cenomanian (courtesy of Halliburton)
showing the distribution of Heteroporella
lepina (Praturlon) (red dots) and Dissocladella ondulata (Raineri)
(yellow squares). Brazil
(Sergipe-Alagoas Basin, Bengtson & Berthou,
1983), Egypt (Kuss, 1994), Germany (Bavaria, Northern Calcareous Alps, Schlagintweit, 1992),
Italy (type locality, Praturlon, 1967), Croatia (Gušić &
Jelaska, 1990), Jordan (Kuss & Conrad,
1991), Morocco (Ensslin
& Schlagintweit, 1999), Portugal (eastern north Atlantic margin; Lauverjat
& Poignant, 1978), Tarim Basin, China (this work), Tunisia (Bismuth
et al., 1981). Note that for simplicity, the Cenomanian map has been used although some occurrences are from
Turonian-Coniacian strata. |
A well diversified assemblage of calcareous green algae consisting mainly of Dasycladales is described for the first time from Upper Cretaceous strata of the southwestern Tarim Basin, China. The occurrence of the algae, associated with some (larger) benthic foraminifers, corresponds to two carbonate units within two transgressive cycles, the Cenomanian-Turonian Kukebai Formation and the Campanian Yigeziya Formation. The foraminiferal-algal assemblages described here provide some additional data to the present biostratigraphic inventory indicated in the existing literature. It is expected that wider and more detailed investigations of the Upper Cretaceous shallow-water carbonates of the Tarim Basin will identify additional taxa for both groups than are currently known.
Thanks to Mike Simmons (London) for providing the Cenomanian palaeomap and permission for illustration. Helpful remarks were provided by the two reviewers Bruno Granier (Brest) and Ioan Bucur (Cluj-Napoca). Mike Bidgood (Oldmeldrum) assisted with the English.
Aguirre J. & Riding R. (2005).- Dasycladalean algal biodiversity compared with global variations in temperature and sea level over the past 350 myr.- Palaios, Lawrence - KS, vol. 20, no. 6, p. 581-588.
Archiac R. d' (1843).- Description géologique du Département de l'Aisne.- Langlois et Leclercq, Paris, 290 p. URL: https://patrimoine.minesparis.psl.eu/document/G%C3%A9ol_Aisne_1843_texte
Bakalova D. (1971).- Nouvelles espčces de Dasycladaceae (Algae) dans les sédiments urgoniens du Prébalkan central.- Bulletin of the Geological Institute, Bulgarian Academy of Sciences - Committee of Geology (Series Paleontology), Sofia, vol. 20, p. 123-127.
Barattolo F. (2002).- Late Cretaceous-Paleogene Dasycladaleans and the K/T boundary problem, Research advances in calcareous algae and microbial carbonates. In: Bucur I.I. & Filipescu S. (eds.), Proceedings of the 4th IFAA Regional Meeting Cluj Napoca, Romania, 29 August-5 September 2001.- Cluj University Press, p. 17-40.
Bassoullet J.-P., Bernier P., Conrad M. A., Deloffre R. & Jaffrezo M. (1978).- Les algues Dasycladales du Jurassique et du Crétacé.- Géobios, Villeurbanne, Mémoire spécial 2, 330 p.
Bassoullet J.-P, Bernier P., Deloffre R., Génot P., Jaffrezo M., Poignant A.F. & Segonzac G. (1975).- Réflexions sur la systématique des Dasycladales fossiles. Étude critique de la terminologie et importance relative des critčres de classification.- Géobios, Villeurbanne, vol. 8, no. 4, p. 259-290.
Bengtson P. & Berthou P.-Y. (1983).- Microfossiles et échinodermes incertae sedis des dépôts albiens ŕ coniaciens du basin de Sergipe-Alagoas, Brésil.- Cahiers de Micropaléontologie, Paris, no. 3 (1982), p. 13-16.
Bismuth H., Boltenhagen C., Donze P., Le Fčvre J. & Saint-Marc P. (1981).- Le Crétacé moyen et supérieur du Djebel Semmama (Tunisie du Centre-Nord) ; microstratigraphie et évolution sédimentologique.- Bulletin des Centres des Recherches Exploration-Production elf-Aquitaine, Pau, vol. 5, no. 2, p. 193-267.
Bucur I.I. (1993).- Some new or poorly known calcareous algae (Dasycladales, Gymnocodiaceae) in the Lower Cretaceous deposits from the Reşita-Moldava Nouă Zone (Southern Carpathians, Romania).- Revista Espańola de Micropaleontologia, Madrid, vol. 25, no. 1, p. 93-126.
Bucur I.I. (1994).- Lower Cretaceous Halimedaceae and Gymnocodiaceae from Southern Carpathians and Apuseni Mountains (Romania) and the systematic position of the Gymnocodiaceae.- Beiträge zur Paläontologie, Wien, vol. 19, p. 13-37.
Bucur I.I. (1999).- Stratigraphic significance of some skeletal algae (Dasycladales, Caulerpales) of the Phanerozoic. In: Farinacci A. & Lord A.R. (eds.) Depositional episodes and bioevents.- Palaeopelagos, Rome, Spec. Pub. 2, p. 53-104.
Bucur I.I., Schlagintweit F., Rashidi K. & Saberzadeh B. (2016).- Morelletpora turgida (Radoičić, 1975, non 1965) a Tethyan calcareous green alga (Dasycladales): Taxonomy, stratigraphy and paleaogeography.- Cretaceous Research, vol. 58, p. 168-182.
Chiocchini M., Pampaloni M.L. & Pichezzi R.M. (2012).- Microfacies e microfossili delle succession carbonatiche mesozoiche Lazio e Abruzzo.- Memorie per servire descrittive alla Carta Geologica d´Italia, Rome, vol. 17, 223 Pls.
Cowgill E. (2010).- Cenozoic right-slip faulting along the eastern margin of the Pamir salient, northwestern China.- GSA Bulletin, Boulder - CO, vol. 122, vols. 1-2, p. 145-161.
De Castro P. (1997).- Introduzione allo studio in sezione sottile delle Dasicladali fossili (An approach to thin-section study of fossil Dasycladales).- Quaderni dell' Accademia Pontaniana, Napoli, no. 22, 261 p.
Deloffre R. & Radoičić R. (1978).- Algues calcaires (Dasycladales) du Paléocčne de Slovénie (Yougoslavie).- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 2, no. 1, p. 61-95.
Elliott G.F. (1957).- New calcareous algae from the Arabian Peninsula.- Micropaleontology, Flushing - NY, vol. 3, no. 3, p. 227-230.
Ensslin R. & Schlagintweit F. (1999).- Contributions to the knowledge of Upper Cretaceous calcareous algae from the Central Middle Atlas Mountains, Morocco.- Zeitschrift der deutschen geologischen Gesellschaft, Stuttgart, vol. 149, no. 4, p. 473-486.
Gao B., Zhang Q., Rao X. & Ding L. (2024).- Persistence of a shallow-marine environment in the western Kunlun area (northwestern Tibet) until the early Maastrichtian: Evidence from radiolitid rudist bivalves.- Cretaceous Research, vol. 167, article 106035, 18 p.
Granier B. (2012).- The contribution of calcareous green algae to the production of limestones: A review.- Geodiversitas, Paris, vol. 34, no. 1, p. 35-60.
Granier B. & Berthou P.-Y. (2002).- Algues calcaires fossiles, nouvelles ou peu connues, du Portugal. 1čre Partie. In: Bucur I.I. & Filipescu S. (eds.), Proceedings of the 4th IFAA Regional Meeting Cluj Napoca, Romania, 29 August-5 September 2001.- Cluj University Press, p. 117-126.
Granier B. & Bucur I.I. (2019).- Le genre Bakalovaella Bucur, 1993 (Dasycladeae, Dasycladaceae), et description de son plus ancien représentant Crétacé.- Carnets Geol., Madrid, vol. 19, no. 1, p. 1-19. DOI: 10.4267/2042/69540
Granier B., Bucur I. I. & Dias-Brito D. (2017).- About Trinocladus Raineri, 1922: When some Permocalculus (Gymnocodiacean algae) reveal to be Triploporellacean algae (Revision of the Jesse Harlan Johnson Collection. Part 5).- Facies, Erlangen, vol. 63, article 27, 12 p.
Granier B. & Deloffre R. (1993).- Inventaire critique des algues Dasycladales fossiles. IIo Partie - Les Algues Dasycladales du Jurassique et du Crétacé.- Revue de Paléobiologie, Genčve, vol. 12, no. 1, p. 19-65.
Granier B., Dias-Brito D. & Bucur I.I. (2013a).- A new mid-Cretaceous Neomeris (dasycladacean alga) from the Potiguar Basin, Brazil.- Facies, Erlangen, vol. 59, p. 221-230.
Granier B., Dias-Brito D., Bucur I.I. & Tibana P. (2013b).- Brasiliporella, a new mid-Cretaceous dasycladacean genus: The earliest record of the Tribe Batophoreae.- Facies, Erlangen, vol. 59, p. 207-220.
Granier B., Masse J.-P. & Berthou P.-Y. (1994).- Heteroporella lepina Praturlon, 1967, revisited (followed by taxonomic notes on the so-called "Heteroporella" species).- Beiträge zur Paläontologie, Vienna, vol. 19, p. 129-141.
Guo F., Yang D., Eriksson K.A. & Guo L. (2015).- Paleoenvironments, stratigraphic evolution and reservoir characteristics of the Upper Cretaceous Yingjisha Group, southwest Tarim Basin.- Marine and Petroleum Geology, vol. 67, p. 336-355.
Gušić I. & Jelaska V. (1990).- Upper Cretaceous stratigraphy of the Island of Brač.- Djela Jugoslavenska Akademija Znanosti i Umjetnosti Zagreb, vol. 69, 160 p.
Hao Y.C. & Zeng X.L. (1984).- On the evolution of the west Tarim gulf from Mesozoic to Cenozoic in terms of characteristics of foraminiferal fauna.- Acta Micropalaeontologica Sinica, Nanjing, vol. 1, no. 1, p. 1-16.
Hao Y., Zeng X. & Guo X. (1988).- The Marine Cretaceous in the Western part of the Tarim Basin of Xinjiang and its depositional Environments.- Acta Geologica Sinica, Beijing, vol. 1, no. 1, p. 13-27.
Hao Y., Guo X., Ye L., Yao P., Fu D., Li H., Rua P. & Wang D. (2001).- The Boundary between the Marine Cretaceous and Tertiary in the Southwest Tarim Basin.- Geological Publishing House, Beijing, 108 p.
He C. (1991).- Late Cretaceous-Early Tertiary Microphytoplankton from the Western Tarim Basin in Southern Xinjiang, China.- Chinese Science Publishing House, Beijing, 235 p.
Jiang H.-X. & Zhang Y.-Y. (2022).- Earliest calcified green algae from the 520 Ma old Cambrian dolostones in Xinjiang, China.- Biopetrology, Beijing, vol. 1, no. 1, p. 9-18.
Kaminski M.A., Setoyama E. & Cetean C.G. (2008).- Revised stratigraphic ranges and the Phanerozoic diversity of agglutinated foraminiferal genera. In: Kaminski M.A. & Coccioni R. (eds.), Proceedings of the Seventh International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 13, p. 79-106.
Kaya M.Y. (2020).- Cretaceous-Paleogene evolution of the proto-Paratethys Sea in Central Asia: Mechanisms and palaeoenvironmental impacts.- PhD Thesis, University of Potsdam, 237 p. DOI: 10.25932/publishup-48329
Kuss J. (1994).- Cretaceous (Albian-Turonian) calcareous algae from Egypt and Jordan - Systematics, stratigraphy and paleogeography.- Abhandlungen der geologischen Bundesanstalt, Vienna, vol. 50, p. 295-317.
Kuss J. & Conrad M.-A. (1991).- Calcareous algae from Cretaceous carbonates of Egypt, Sinai, and southern Jordan.- Journal of Paleontology, Washington - DC, vol. 65, no. 5, p. 869-882.
Lamouroux J.V.F. (1816).- Histoire des Polypiers coralligčnes flexible vulgairement nommés Zoophytes.- F. Poisson, Caen, 559 p.
Lan X. & Wei J. (1995).- Late Cretaceous-Early Tertiary marine bivalve fauna from the western Tarim basin.- Chinese Science Publishing House, Beijing, 212 p.
Lauverjat J. & Poignant A.F. (1978).- Les algues de la série ŕ Vacocératidés du basin occidental Portugais.- Cahiers de Micropaléontologie, Paris, vol. 3, p. 121-126.
Masse J.-P. & Isintek I. (2000).- Algues Dasycladales de l'Albien de la Péninsule de Karaburun, Turquie.- Revue de Micropaléontologie, Paris, vol. 43, no. 3, p. 365-380.
Masse P. (1995).- Calcareous Algae and microcoprolites of the Albian platform carbonates of the Congo margin.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 19, no. 2, p. 301-317.
Michelin H. (1840-1847).- Iconographie zoophytologique. Description par localités et Terrains des Polypiers fossiles de France et pays environnants.- P. Bertrand, Paris, 348 p.
Morellet L. & Morellet J. (1913).- Les Dasycladacées du Tertiaire parisien.- Mémoires de la Société géologique de France, Paris, t. XXI, fasc. 1, no. 47, 43 p. (Pls. I-III).
Mu X.-N. (1993).- Late Cretaceous palaeobiogeography of calcareous algae. In: Barattolo F., De Castro P. & Parente M. (eds.), Studies on fossil benthic algae.- Bolletino della Societŕ Paleontologica Italiana, spec. vol. 1, p. 333-344.
Pan H.-Z. (1990).- Late Cretaceous gastropod dominated communities of the Western Tarim Basin, Southern Xinjiang, China.- Lethaia, Oslo, vol. 23, no. 3, p. 273-289.
Parente M. (1997).- Dasycladales from the Upper Maastrichtian of Salento Peninsula (Puglia, Southern Italy).- Facies, Erlangen, vol. 36, p. 91-122.
Pia J. (1930).- A new dasycladacea, Holosporella siamensis nov. gen., nov. sp., with a description of the allied genus Aciculella Pia.- Records of the Geological Survey of India, Calcutta, vol. 6, no. 1, p. 177-181.
Pia J. (1936).- Calcareous green algae from the Upper Cretaceous of Tripoli (North Africa).- Journal of Paleontology, Washington - DC, vol. 10, no. 1, p. 3-13.
Pia J., Rao S.R.N. & Rao K.S. (1937).- Dasycladaceen aus Zwischenlagen des Dekkantrapps bei Rajahmundry in Südindien.- Sitzungsberichte der Akademie der Wissenschaften in Wien, vol. 146, p. 227-236.
Praturlon A. (1967).- Heteroporella lepina, new dasyclad species from Upper Cenomanian-Lower Turonian of Central Apennines.- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 5 (1966), no. 2, p. 202-205.
Radoičić R. (1965).- Pianella turgida n. sp. from the Cenomanian of the Outer Dinarids.- Geološki Vjesnik, Zagreb, vol. 18, p. 195-199.
Radoičić R. (1975).- Linoporella buseri sp. nov. from the Liassic of the Julian Alps (a preliminary report).- Bulletin Scientifique, Conseil des Académies des Sciences et des Arts de la R.S.F. de Yougoslavie (Section A: Sciences naturelles, techniques et médicales), Belgrade, vol. 20, no. 9-10, p. 277-278.
Radoičić R. (1978).- Clypeina? alrawii, n. sp., a Dasycladacea from the Upper Cretaceous of Iraq.- Bulletin de l'Académie serbe des Sciences et des Arts (Classe des Sciences naturelles et mathématiques), Belgrade, t. LXI , no. 17, p. 1-4.
Radoičić R. (1983).- Dasyclads Cylindroporella parva n. sp. from the Upper Cretaceous of Libya and Clypeina pastriki n. sp. from the Upper Cretaceous of the Dinarides and Libya.- Bulletin de l'Académie serbe des Sciences et des Arts (Classe des Sciences mathématiques et naturelles), Belgrade, t. CCCXXXV, no. 49, p. 65-80.
Radoičić R. (1997).- New Clypeina (Dasycladales) from Santonian of Grebnik (Mirdita Zone, Yugoslavia).- Annales Géologiques de la Péninsule Balkanique, Belgrade, vol. 61, no. 1, p. 133-141 [in Serbian and English].
Radoičić R., Conrad M.A. & Carras N. (2005).- Observations on Neomeris cretacea var. undulata Raineri followed by Dissocladella bonardii, n. sp. (Dasycladales, green algae).- Revue de Paléobiologie, Genčve, vol. 24, no. 1, p. 311-317.
Raineri R. (1922).- Alghe sifonee fossili della Libia.- Atti della Societa Italiana di Scienze Naturali de Museo Civico, Milano, vol. 61, no. 1, p. 72-86.
Rao L.R. & Pia J. (1936).- Fossil algae from the uppermost Cretaceous beds (the Niniyur group) of the Trichinopoly district, South India.- Palaeontologia Indica, Calcutta, Mémoire, 4, p. 1-49.
Rao X. (2019).- Biradiolites from the Yigeziya Formation of the southwestern Tarim Basin.- Open Journal of Geology, vol. 9, no. 10, p. 562-565.
Rao X., Skelton P.W., Sano S. & Wan B. (2023).- Taxonomy and paleobiogeographic implication of Glabrobournonia Morris and Skelton (Hippuritida, Radiolitidae) from the late Cretaceous Yigeziya Formation, southwestern Tarim Basin.- Palaeoworld, vol. 32, no. 1, p. 136-147.
Schlagintweit F. (1992).- Further record of calcareous algae (Dasycladaceae, Udoteaceae, Solenoporaceae) from the Upper Cretaceous of the Northern Calcareous Alps (Gosau Formation, Branderfleck Formation).- Revue de Paléobiologie, Genčve, vol. 11, no. 1, p. 1-12.
Schlagintweit F. & Yazdi-Moghadam M. (2023).- Pseudocyclammina sarvakensis sp. nov. and Pseudotextulariella brevicamerata sp. nov.: Further evidence for the Cenomanian megadiversity of larger benthic foraminifera from the Sarvak Formation of SW Iran.- Acta Palaeontologica Romaniae, Bucharest, vol. 19, no. 2, p. 3-13.
Schlagintweit F., Yazdi-Moghadam M. & Cvetko-Tesovic B. (2023).- Upper Cretaceous foraminifera Murgeina apula (Luperto Sinni, 1968): A Methusalem and Cenomanian-Turonian boundary survivor taxon.- Acta Palaeontrologica Romaniae, Bucharest, vol. 19, no. 2, p. 25-38.
Schlagintweit F., Omidvar M., Safari A., Yazdi-Moghadam M. & Rashidi K. (2024).- Dasycladales (green algae) and some benthic foraminifera from the Upper Cretaceous Ilam Formation (Late Coniacian-Santonian), SW Iran (Onshore, Offshore).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 130, no. 3, p. 487-506.
Schroeder R. & Neumann M. (coordinators, 1985).- Les grands Foraminifčres du Crétacé moyen de la région méditerranéenne.- Géobios, Villeurbanne, Mémoire spécial vol. 7, 161 p.
Scott R.W., Xiaqiao W., Jingeng S. & Shi-Xuan W. (2010).- Rudists of Tibet and the Tarim Basin, China: Significance to Requieniidae phylogeny.- Journal of Paleontology, Washington - DC, vol. 84, no. 3, p. 444-465.
Sha J., Fabbi S., Cestari R. & Consorti L. (2020).- Stratigraphic and taxonomic considerations on the Late Cretaceous rudist fauna of Aksai Chin (Western Tibet, China) from the De Filippi Collection.- Carnets Geol., Madrid, vol. 20, no. 13, p. 249-272. DOI: 10.2110/carnets.2020.2013
Shaozhi M. & Norris G. (1988).- Late Cretaceous-Early Tertiary dinoflagellates and acritarchs from the Kashi Area, Tarim Basin, Xinjiang Province, China.- Royal Ontario Museum Life Science Division Contributions, Toronto, vol. 150, p. 1-93.
Simmons M.D. & Bidgood M.D. (2023).- "Larger" benthic foraminifera of the Cenomanian. A review of the identity and stratigraphic and palaeogeographic distribution of non-fusiform planispiral (or near-planispiral) forms.- Acta Palaeontologica Romaniae, Bucharest, vol. 19, no. 2, p. 39-169.
Sun Q. & Schlagintweit F. (2024).- First record of Upper Cretaceous (Campanian?) larger benthic foraminifera from the Yigeiya Formation of the southwestern Tarim Basin, China.- Acta Palaeontologica Romaniae, Bucharest, vol. 20, no. 2, p. 131-144.
Tang T., Yang H., Lan X., Yu C., Xue Y., Zhang Y., Hu L., Zhong S. & Wei J. (1989).- Marine Late Cretaceous and Early Tertiary stratigraphy and petroleum geology in western Tarim basin, China.- Chinese Science Publishing House, Beijing, 155 p.
Tang T., Xue Y. & Yu C. (1992).- Marine sedimentary characteristics and environments from Late Cretaceous to Early Tertiary in the west part of Tarim basin of Xinjiang.- Chinese Science Publishing House, Beijing, 138 p.
Varma C.P. (1950).- A new genus of calcareous algae (Dasycladaceae) from the Ranikot beds (Palaeocene) of the Punjab Salt Range.- Current Science, Bangaluru, vol. 19, no. 7, p. 207-208.
Varma C.P. (1952).- Clypeina (Dasycladaceae) from the Cretaceous of South India.- The Palaeobotanist, Lucknow, vol. 1, p. 439-441. DOI: 10.54991/jop.1952.421
Varma C.P. (1955).- Further observations on Morelletpora nammalensis Varma from the Khairabad Limestone (Ranikot) beds of the Nammal Gorge, Punjab Salt Range.- The Palaeobotanist, Lucknow, vol. 4, p. 101-111. DOI: 10.54991/jop.1955.478
Xi D., Cao W., Cheng Y., Jiang T., Jia J., Li Y. & Wan X. (2016).- Late Cretaceous biostratigraphy and sea-level change in the southwest Tarim Basin.- Palæogeography, Palæoclimatology, Palæoecology, vol. 441, p. 516-527.
Xi D., Wan X., Li G. & Li G. (2019).- Cretaceous integrative stratigraphy and timescale in China.- Science China, Earth Sciences, vol. 62, p. 256-286.
Yang H.-R., Tang T.-F., Hu L.-Y., Yu C.-L., Zhang Y.-Y., Zhong Sh.-L. & Wei J.-M. (1983).- A preliminary study of the Upper Cretaceous of the Western Tarim Basin) (South Xinjiang, China) with special reference to its transgressions.- Zitteliana, München, vol. 10, p. 115-121.
Zhang S., Hu X., Han Z., Li J. & Garzanti E. (2018).- Climatic and tectonic controls on Cretaceous-Palaeogene sea-level changes recorded in the Tarim epicontinental sea.- Palæogeography, Palæoclimatology, Palæoecology, vol. 501, p. 92-110.
Zhang M., Du B., Wu Z., Dou L., Zhumahun A., Jiaoba D., Jin P., Du Z., Wang S. & Xia Y. (2022).- Dinoflagellate cyst biostratigraphy on initial Neotethys transgression deposits from the Cenomanian and Turonian in the Tarim Basin, western China.- Marine and Petroleum Geology, vol. 138, article 105531, 16 p.
Zhang M., Dai S., Du B., Wang Y., Liu G., Hong Y., Zhang J., Liu Y. & Cai J. (2025).- High-resolution oceanic anoxic event 2 (OAE2) records from the north of eastern Tethys and evidence for short-term sea regression and wildfire at its early phase.- Marine and Petroleum Geology, vol. 171, article 107180, 14 p.
Zhong S.L. (1992).- Calcareous Nannofossils From the Upper Cretaceous and Lower Tertiary in the Western Tarim Basin, South Xinjiang, China.- Science Press, Beijing, p. 1-34 [in Chinese with English abstract].