◄ Carnets Geol. 25 (5) ►
![]()
Outline:
[1. Introduction]
[2. The foundations of the Aptian-Albian biozones based on LBF]
[3. Aptian-Albian sequences along a part of the Mediterranean coast of Alicante]
[4. The ammonite collections of the UCBL1]
[5. Discussion]
[6. The fate of the Aptian-Albian LBF biozones]
[7. Conclusions]
[Bibliographic references] and ... [Plates]
Membre correspondant, Muséum d'histoire naturelle (MHN),
Route de Malagnou 1, 1208 Genève (Switzerland);
Association Carnets de Géologie, 2 impasse Charles Martel, F-29217 Plougonvelin (France)
; [ORCID: 0000-0001-9468-2353]
Laboratoire de Géologie de Lyon, CNRS, Univ Lyon, UCBL, ENSL,
UJM, LGL-TPE, F-69622, Villeurbanne (France)
; [ORCID: 0000-0002-2821-4233]
Department of Geology, Faculty of Science and Technology,
University of the Basque Country (UPV/EHU), Bilbao (Spain)
; [ORCID: 0000-0002-3142-7256]
Published online in final form (pdf) on February 15, 2025
DOI: 10.2110/carnets.2025.2505
![]()
[Editor: Michel Moullade; language editor: Phil Salvador; technical editor: Bruno
R.C. Granier]
![]()
The Aptian-Albian interval of the Costa Blanca (SE Spain) has been studied to provide new insights into the evolution of the neritic domain of carbonate platforms in the Tethys. Three areas have been re-examined: Serra Gelada, Puig Campana, and Cabeçó d'Or. The biozonations of large benthic foraminifers (LBF) have been revised based on ammonite biostratigraphy. Updated ammonite datings and stratigraphic revisions reveal that several LBF and calcareous algae (CA) taxa previously attributed to the late Aptian actually originated in the early Albian. These findings correct and refine the calibration of LBF and CA biozones and ranges with standard ammonite zones. Furthermore, the identification of a significant local hiatus during the middle Albian at Cabeçó d'Or is likely linked to halokinesis.
• Alicante;
• Aptian;
• Albian;
• large benthic foraminifers;
• calcareous algae;
• ammonites;
• biozones
Granier B.R.C., Robert E. & Pérez-Malo J. (2025).- The Aptian-Albian of the Costa Blanca (SE Spain): Implications for identifying the Aptian/Albian boundary in the neritic zone within the Tethys realm.- Carnets Geol., Madrid, vol. 25, no. 5, p. 109-145. DOI: 10.2110/carnets.2025.2505
L'Aptien-Albien de la Costa Blanca (sud-est de l'Espagne) : Implications pour l'identification de la limite Aptien/Albien en zone néritique dans le domaine téthysien.- L'intervalle Aptien-Albien de la Costa Blanca (sud-est de l'Espagne) a été étudié afin de fournir de nouvelles perspectives sur l'évolution du domaine néritique des plates-formes carbonatées dans la Téthys. Trois secteurs ont été réexaminés : Serra Gelada, Puig Campana et Cabeçó d'Or. Les biozonations de grands foraminifères benthiques (LBF) ont été révisées sur la base de la biostratigraphie des ammonites. Les datations par ammonites mises à jour et les révisions stratigraphiques révèlent que plusieurs taxons de LBF et d'algues calcaires (AC), auparavant attribués à l'Aptien supérier, sont en réalité apparus à l'Albien inférieur. Ces résultats corrigent et affinent la calibration des biozones et des répartitions stratigraphiques des LBF et des AC sur la zonation standardisée des ammonites. En outre, une importante lacune locale identifiée au cours de l'Albien moyen au Cabeçó d'Or est probablement liée à l'halocinèse.
• Alicante ;
• Aptien ;
• Albien ;
• grands foraminifères benthiques ;
• algues calcaires ;
• ammonites ;
• biozones
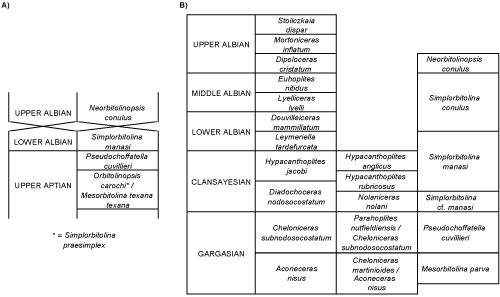
Currently, the Lower
Cretaceous ranges and biozonations of large benthic foraminifers are still based
on early, partially empirical, biozonal schemes proposed by Schroeder
(1963) and Fourcade
(1967, 1970; here: Fig. 1.A ![]() ) more than half a century
ago. Up until the 1980s and even
the 1990s, several authors (e.g., Peybernès, 1976; here: Fig.
1.B
) more than half a century
ago. Up until the 1980s and even
the 1990s, several authors (e.g., Peybernès, 1976; here: Fig.
1.B ![]() ; García
Hernández, 1978; Jaffrezo,
1980; Masse et al., 1992)
refined the original biozonation, eventually introducing some regional schemes
that incorporated a few endemic species. These authors almost reached an
agreement regarding the relative ranges of most key taxa, including overlaps (or
lack thereof) and the relative orders of both first and last occurrences.
However, no significant progress has been made since then in calibrating these
total ranges and related biozones to the ammonite standard scale (Szives
et al., 2024).
; García
Hernández, 1978; Jaffrezo,
1980; Masse et al., 1992)
refined the original biozonation, eventually introducing some regional schemes
that incorporated a few endemic species. These authors almost reached an
agreement regarding the relative ranges of most key taxa, including overlaps (or
lack thereof) and the relative orders of both first and last occurrences.
However, no significant progress has been made since then in calibrating these
total ranges and related biozones to the ammonite standard scale (Szives
et al., 2024).
|
|
Figure 1: Pre-existing
large benthic foraminifer - LBF
biozonal schemes: A) Fourcade, 1970; B) Peybernès,
1976. |
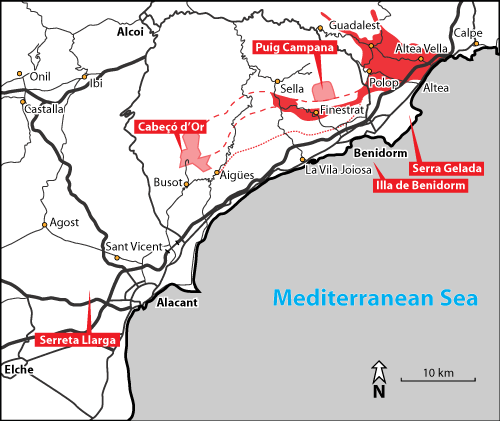
Exposures of Lower Cretaceous
strata in the Costa Blanca (Province of Alicante, SE Spain: Fig.
2 ![]() ) are rather
scattered on the 1:50,000 scale geological maps of Alicante (872), Altea/Benidorm
(848), Elda (871), and Villajoyosa (847). Among these areas, Serra Gelada (Fig.
3
) are rather
scattered on the 1:50,000 scale geological maps of Alicante (872), Altea/Benidorm
(848), Elda (871), and Villajoyosa (847). Among these areas, Serra Gelada (Fig.
3 ![]() ), Puig Campana
(Fig. 4
), Puig Campana
(Fig. 4 ![]() ),
and/or Cabeçó d'Or (Fig. 5
),
and/or Cabeçó d'Or (Fig. 5 ![]() ) have been investigated
to varying extents by numerous researchers since the pioneering work of Nicklès
in 1892 (e.g., Azéma, 1977; Estévez
et al., 1984; and the first author: Granier,
1987). These
three areas form the core of this new study, which focuses exclusively on the
Aptian-Albian interval, comprising both ammonite-rich facies and carbonate
platform facies with large benthic foraminifers (LBF) and calcareous algae (CA).
) have been investigated
to varying extents by numerous researchers since the pioneering work of Nicklès
in 1892 (e.g., Azéma, 1977; Estévez
et al., 1984; and the first author: Granier,
1987). These
three areas form the core of this new study, which focuses exclusively on the
Aptian-Albian interval, comprising both ammonite-rich facies and carbonate
platform facies with large benthic foraminifers (LBF) and calcareous algae (CA).
|
|
Figure 2: Location
map with key localities of the Costa Blanca (Province of Alicante, SE Spain):
Cabeçó d'Or (Sanatorio section), Puig Campana (Cluse, Realet d'Alt, and Rafel
sections), Serra Gelada (Morro San Jordi, Punta de les Caletes, Relais, Calle
Dos Callas, and Camí del Far sections), Illa de Benidorm, and Serreta Llarga. |
According to second author (E.R.), some ammonites collected by the first author (B.R.C.G.) in the eastern Cabeçó d'Or area, which were initially assigned to early Aptian and middle Albian ages, should be reassigned to the late Aptian and the late Albian, respectively. These biostratigraphic observations warrant a revision of earlier conclusions because they imply that the correlation scheme for the Aptian-Albian interval in the Costa Blanca, initially proposed by the first author (Granier, 1987: Fig. 55; 1988: Fig. 1), requires at least slight adjustments. Similarly, as noted by Granier and Perthuisot (2009), the wider lithostratigraphic correlation scheme of Castro et al. (2001, 2008) for the eastern Prebetic Zone erroneously considers the lower Albian strata at Serra Gelada (e.g., 'Cylindroporella oolithes', 'coral biolithites', and 'rudistid limestones') as a member of the non-coeval upper Aptian Seguili Formation.
Finally, given the lack or
scarcity of ammonite data in most earlier works on the large benthic
foraminifers - LBF (e.g.,
Fourcade, 1970; here: Fig. 1.A ![]() ; Peybernès,
1976; here:
Fig. 1.B
; Peybernès,
1976; here:
Fig. 1.B ![]() ; García
Hernández, 1978; Jaffrezo,
1980; Masse et al., 1992),
these revised ammonite datings call into question the reliability of the
calibration of the Aptian-Albian biozones or that of the LBF-specific ranges on
the standard ammonite biozones (Szives et al., 2024). One
goal of this paper is to replace the empirical approach to LBF biozones (which
may also incorporate certain phycological proxies) with a more pragmatic
approach by calibrating their stratigraphic ranges against the ammonite scale.
; García
Hernández, 1978; Jaffrezo,
1980; Masse et al., 1992),
these revised ammonite datings call into question the reliability of the
calibration of the Aptian-Albian biozones or that of the LBF-specific ranges on
the standard ammonite biozones (Szives et al., 2024). One
goal of this paper is to replace the empirical approach to LBF biozones (which
may also incorporate certain phycological proxies) with a more pragmatic
approach by calibrating their stratigraphic ranges against the ammonite scale.
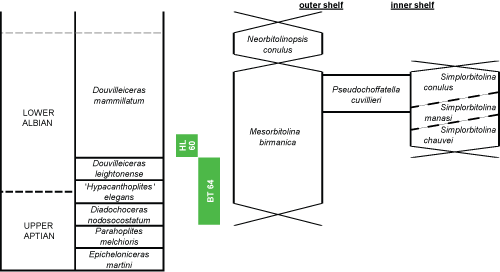
Schroeder (1963, p. 323) and Fourcade (1967, p. 844) are the inventors of a set of biozones based on large benthic foraminifers (LBF) including for the latter from base to top:
a) the Pseudochoffatella cuvillieri biozone, the original definition of which is: 'This biozone, of reduced thickness, constitutes an excellent marker level that I place in the upper Aptian; however, it is not excluded that it may extend partially into the Albian. Simplorbitolina manasi Ciry et Rat, as well as Mesorbitolina texana-texana (Roemer), are also found there' [translated from the French in Fourcade, 1967: 'Cette biozone, d'épaisseur réduite, constitue un excellent niveau repère que nous situons dans l'Aptien supérieur ; il n'est cependant pas exclu qu'elle puisse s'étendre en partie dans l'Albien. On y rencontre également Simplorbitolina manasi Ciry et Rat, ainsi que des Mesorbitolina texana-texana (Roemer)'].
b) the Simplorbitolina manasi biozone, the definition of which is: 'Generally poor in fossils, it has until now been characterized only by this foraminifer and Mesorbitolina texana-texana. We assign this biozone to the lower Albian' [translated from the French in Fourcade, 1967: 'Généralement peu fossilifère, elle n'est caractérisée jusqu'à maintenant que par ce Foraminifère et Mesorbitolina texana-texana. Nous rattachons cette biozone à l'Albien inférieur'].
c) the Neorbitolinopsis conulus biozone, the definition of which is: 'This biozone begins a few meters above levels with ammonites (Knemiceras, which can be compared to K. deserti Douvillé and K. gracile Mahmoud, which in Algeria characterize the lower Albian). The foraminifer that characterizes this biozone is accompanied by an assemblage including Mesorbitolina texana-texana, Cuneolina gr. pavonia-parva Henson, and Trocholina gr. lenticularis Henson' [translated from the French in Fourcade, 1967: 'Cette biozone débute quelques mètres au-dessus de niveaux à Ammonites (Knemiceras que l'on peut rapprocher de K. deserti Douvillé et de K. gracile Mahmoud qui en Algérie caractérisent l'Albien inférieur). Le Foraminifère, qui caractérise cette biozone, est accompagné d'un cortège comprenant : Mesorbitolina texana-texana, Cuneolina gr. pavonia-parva Henson, Trocholina gr. lenticularis Henson'].
Regarding the Pseudochoffatella cuvillieri biozone, Fourcade (1970: p. 131) wrote 'I hypothesize that this zone ( ) is situated in the upper Aptian. ( ) However, the precise age remains uncertain due to the inability to correlate it with an ammonite level' [translated from the French: 'nous situons à titre d'hypothèse cette zone ( ) dans l'Aptien supérieur. ( ) Il subsiste toutefois le problème de son âge précis, faute d'avoir pu le relier à un niveau à ammonites'].
Fourcade (1967, 1970) also stressed that Simplorbitolina manasi was already present in the Pseudochoffatella cuvillieri biozone. The stratigraphic range of this species was considered to extend downward into the upper Aptian. This last hypothesis was not questioned until now. However, a valid alternative could have been to consider that both Simplorbitolina manasi and Pseudochoffatella cuvillieri lived in the early Albian, not in late Aptian times. The range overlap of both species is not necessarily well documented in early publications because the definition of the species Simplorbitolina manasi Ciry & Rat, changed over time, notably with the introduction of new species.
Here, we will examine a pragmatic approach to revising Schroeder's and Fourcade's biozones, which were sometimes based on empirical methods and conclusions.
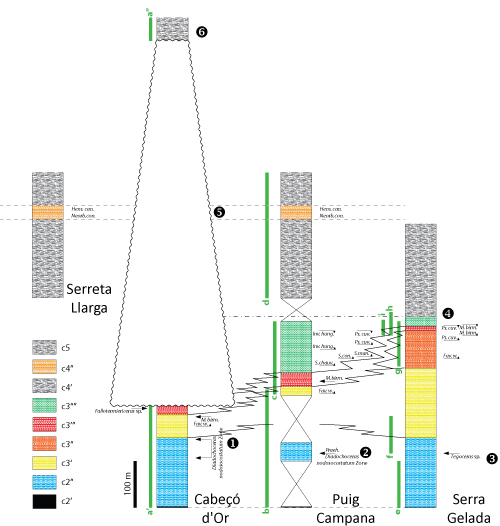
This chapter provides a summary of the lithostratigraphy and biostratigraphy of Serra Gelada, Puig Campana, and Cabeçó d'Or, the three key areas forming the core of the study. All descriptions start at the base of the sedimentary sequences and move upwards.
At Serra Gelada (Fig. 3 ![]() ), the
sedimentary sequence begins above the Jurassic limestones (possibly including
Berriasian strata at their uppermost part: Fig. 3.C
), the
sedimentary sequence begins above the Jurassic limestones (possibly including
Berriasian strata at their uppermost part: Fig. 3.C ![]() ), with a condensed interval
less than 0.5 meter thick, characterized by ferruginous ooids and oncoids
floating in a calcareous muddy matrix (Granier, 1987). It spans part of
the Valanginian to lower Aptian interval and contains numerous late Barremian
and early Aptian ammonites (e.g., Deshayesites deshayesi Zone).
This condensed interval is overlain by a unit approximately 150 meters thick,
containing early and late Aptian ammonites and, as evidenced here for the first time,
early
Albian ammonites (HL 60). This unit consists of a lower succession composed
of alternating argillaceous limestones and marls, and an upper succession that
is predominantly marly. The next unit consists of roughly 150 meters of
'siltstones
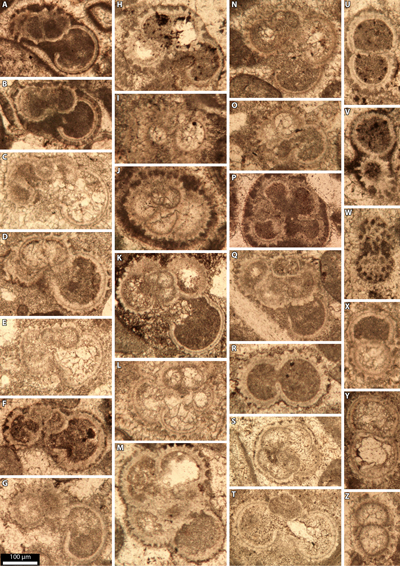
and calcisiltites with Favusella' [Favusella washitensis (Carsey,
1926): Pl. 1
), with a condensed interval
less than 0.5 meter thick, characterized by ferruginous ooids and oncoids
floating in a calcareous muddy matrix (Granier, 1987). It spans part of
the Valanginian to lower Aptian interval and contains numerous late Barremian
and early Aptian ammonites (e.g., Deshayesites deshayesi Zone).
This condensed interval is overlain by a unit approximately 150 meters thick,
containing early and late Aptian ammonites and, as evidenced here for the first time,
early
Albian ammonites (HL 60). This unit consists of a lower succession composed
of alternating argillaceous limestones and marls, and an upper succession that
is predominantly marly. The next unit consists of roughly 150 meters of
'siltstones
and calcisiltites with Favusella' [Favusella washitensis (Carsey,
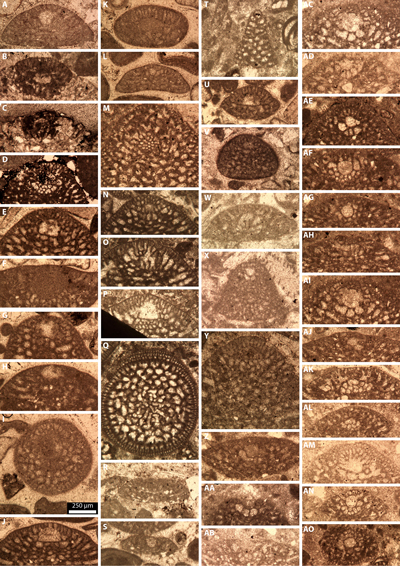
1926): Pl. 1 ![]() , figs. A-E, J-L, N-P, X-Y], forming the main seaside cliff of Serra
Gelada (Fig. 3.D
, figs. A-E, J-L, N-P, X-Y], forming the main seaside cliff of Serra
Gelada (Fig. 3.D ![]() ). This interval is followed by a unit, approximately 90 meters
thick, consisting of 'Cylindroporella
oolithes' (Cylindroporella barnesii Johnson, 1954; here:
Pl. 2
). This interval is followed by a unit, approximately 90 meters
thick, consisting of 'Cylindroporella
oolithes' (Cylindroporella barnesii Johnson, 1954; here:
Pl. 2 ![]() ,
figs. F, K-L) and 'coral biolithites'. The abraded tops of these bioherms pass
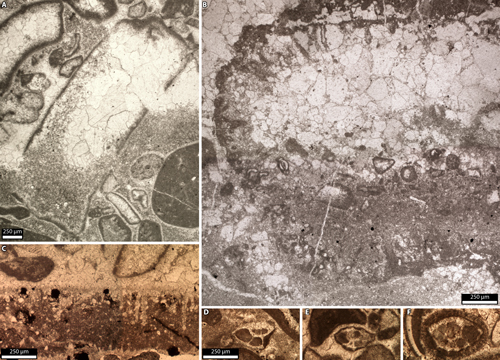
laterally into bored hardgrounds (Fig. 6.A-D
,
figs. F, K-L) and 'coral biolithites'. The abraded tops of these bioherms pass
laterally into bored hardgrounds (Fig. 6.A-D ![]() ). Calcareous microfossils
identified in thin sections include Epistomina spp. [Relais 1 (HL 704) -
Relais 13 (HL 726); Pl. 3
). Calcareous microfossils
identified in thin sections include Epistomina spp. [Relais 1 (HL 704) -
Relais 13 (HL 726); Pl. 3 ![]() , fig. F], Sabaudia minuta (Hofker,
1965),
Pseudochoffatella
cuvillieri Deloffre, 1961 [Relais 12 (HL 722); Pl. 4
, fig. F], Sabaudia minuta (Hofker,
1965),
Pseudochoffatella
cuvillieri Deloffre, 1961 [Relais 12 (HL 722); Pl. 4 ![]() , fig. C], Mesorbitolina
texana (Roemer, 1849) [e.g., Relais 7 (HL 709);
Pl. 5
, fig. C], Mesorbitolina
texana (Roemer, 1849) [e.g., Relais 7 (HL 709);
Pl. 5 ![]() , figs.
A-D, L, R-S], Mesorbitolina
birmanica Sahni,
1937 [e.g., Carabiniers
1 (HL 72); Pl. 5
, figs.
A-D, L, R-S], Mesorbitolina
birmanica Sahni,
1937 [e.g., Carabiniers
1 (HL 72); Pl. 5 ![]() , figs. E-K;
Pl. 6
, figs. E-K;
Pl. 6 ![]() , fig. G],
and Agardhiellopsis cretacea Lemoine, 1966 (Cueva de la ballena,
HL 52). Except for Epistomina spp. and Mesorbitolina texana, these
microfossils are also present in the next unit, which consists of approximately
20 meters of 'rudistid limestones', the top of which marks the maximum of the
progradation of this carbonate platform. According to Granier
(1987) and Granier
and Perthuisot (2009), these microfossil assemblages are characteristic
of the lower Albian, whereas Castro et al. (2008) identify all the
units overlying the Aptian marls as the Helada Member of their Seguilí
Formation and ascribe this interval, more than 250 meters thick, to the late
Aptian. The next unit corresponds to the beginning of a transgression; it
consists of an alternation of 'orbitolinid
calcarenites and marls', approximately 200 meters thick according to Gianetti
et al. (2014). Castro et al.
(2008) identify this last unit
as their Sácaras Formation and its lower boundary as the Aptian/Albian
boundary.
, fig. G],
and Agardhiellopsis cretacea Lemoine, 1966 (Cueva de la ballena,
HL 52). Except for Epistomina spp. and Mesorbitolina texana, these
microfossils are also present in the next unit, which consists of approximately
20 meters of 'rudistid limestones', the top of which marks the maximum of the
progradation of this carbonate platform. According to Granier
(1987) and Granier
and Perthuisot (2009), these microfossil assemblages are characteristic
of the lower Albian, whereas Castro et al. (2008) identify all the
units overlying the Aptian marls as the Helada Member of their Seguilí
Formation and ascribe this interval, more than 250 meters thick, to the late
Aptian. The next unit corresponds to the beginning of a transgression; it
consists of an alternation of 'orbitolinid
calcarenites and marls', approximately 200 meters thick according to Gianetti
et al. (2014). Castro et al.
(2008) identify this last unit
as their Sácaras Formation and its lower boundary as the Aptian/Albian
boundary.
|
|
Figure 3: Serra
Gelada: A-B) Punta de las Caletes section, arrowed (Granier,
1987:
Fig. 22): A) 25/07/1984;
B) Google Earth view © 2024 Airbus. C) lower part of the ammonites marls
and contact with the Jurassic at Morro Sant Jordi, 22/09/2010. D)
Panoramic view from the mirador next to Far de l'Albir, 14/04/1984. E)
The Relais section (Granier,
1987: Fig. 23) with 5 coral bioherms (labeled
A-E) intercalated within 'Cylindroporella oolithes', 09/04/1984.
They pass laterally to hardgrounds, and are abraded and bored at their top. F) Meter-scale sand waves visible
on the cliff face below the Far de l'Albir, 10/1985. F) Decimeter-scale
sand wave on the side of Camí del Far (1986). It exhibits tidal bundles that document the semi-diurnal
lunar control of sedimentation. Figure captions: j: Jurassic limestones
(Berriasian and older strata), c2:
ammonite marls (mostly Aptian), c3': 'siltstones and calcisiltites with Favusella', c3":
'Cylindroporella oolithes',
c3'": 'rudistid limestones'. |
At Puig Campana (Figs. 4 ![]() ,
7
,
7 ![]() ), on
top of lower Valanginian 'Pseudocyclammina calcarenites and sandstones',
the sedimentary sequence begins with a condensed interval, characterized by the
occurrence of ferruginous ooids and oncoids, followed by glauconite grains, all
of them floating in a calcareous muddy matrix (Granier,
1987: Fig. 9).
Almost 2 meters thick, it spans part of the Valanginian to lower Aptian interval
(Fig. 4.B
), on
top of lower Valanginian 'Pseudocyclammina calcarenites and sandstones',
the sedimentary sequence begins with a condensed interval, characterized by the
occurrence of ferruginous ooids and oncoids, followed by glauconite grains, all
of them floating in a calcareous muddy matrix (Granier,
1987: Fig. 9).
Almost 2 meters thick, it spans part of the Valanginian to lower Aptian interval
(Fig. 4.B ![]() ). It is overlain by a poorly exposed marly interval, roughly 40 meters
thick, where pyritous ammonites dated to the latest Aptian (Diadochoceras
nodosocostatum Zone) were collected. These argillaceous strata were likely
tectonically stretched and squeezed, which could explain their relatively
reduced thickness compared to the other areas. The next two units consist of the
'Pieninia calcarenites' and the 'coral biolithites', 20 and 30 meters
thick respectively (Fig. 4.C
). It is overlain by a poorly exposed marly interval, roughly 40 meters
thick, where pyritous ammonites dated to the latest Aptian (Diadochoceras
nodosocostatum Zone) were collected. These argillaceous strata were likely
tectonically stretched and squeezed, which could explain their relatively
reduced thickness compared to the other areas. The next two units consist of the
'Pieninia calcarenites' and the 'coral biolithites', 20 and 30 meters
thick respectively (Fig. 4.C ![]() ). Calcareous microfossils identified in thin
sections include Mesorbitolina
birmanica Sahni,
1937 (Pl. 5
). Calcareous microfossils identified in thin
sections include Mesorbitolina
birmanica Sahni,
1937 (Pl. 5 ![]() , fig. U),
Mesorbitolina sp. (Pl. 5
, fig. U),
Mesorbitolina sp. (Pl. 5 ![]() , figs. V-W),
and Agardhiellopsis cretacea Lemoine, 1966
(Pl. 2
, figs. V-W),
and Agardhiellopsis cretacea Lemoine, 1966
(Pl. 2 ![]() , fig. A).
According to Granier (1987) and Granier and Perthuisot
(2009), based on LBF biozonation, this microfossil assemblage was considered
characteristic of the upper Aptian, a view that is revised here. The overlying 'Simplorbitolina
limestones', which are more than 110 meters thick, consist of lagoonal muddy
facies with Simplorbitolina spp. (Pl. 8
, fig. A).
According to Granier (1987) and Granier and Perthuisot
(2009), based on LBF biozonation, this microfossil assemblage was considered
characteristic of the upper Aptian, a view that is revised here. The overlying 'Simplorbitolina
limestones', which are more than 110 meters thick, consist of lagoonal muddy
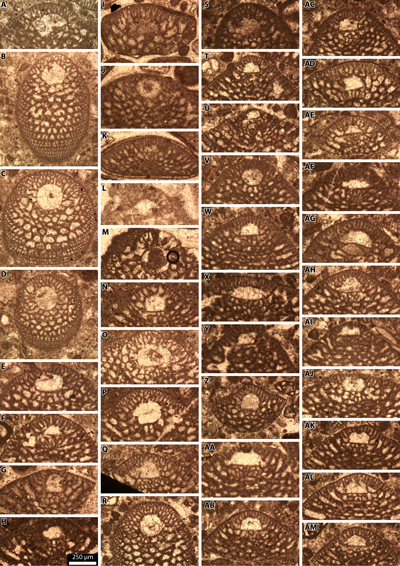
facies with Simplorbitolina spp. (Pl. 8 ![]() , figs. A-Z),
Involutina hungarica (Sidó, 1952), and Pseudochoffatella
cuvillieri (Pl. 4
, figs. A-Z),
Involutina hungarica (Sidó, 1952), and Pseudochoffatella
cuvillieri (Pl. 4 ![]() , figs. D, F-J). Unfortunately, the fault system surrounding the Puig Campana on its western side
interrupts it upward. On the other side of the fault, a new sequence begins with
an alternation of silty marls and argillaceous limestones more than 170 meters
thick followed by about 30 meters of 'Hensonina calcarenites' with Neorbitolinopsis
conulus (Douvillé, 1912) (Pl. 9
, figs. D, F-J). Unfortunately, the fault system surrounding the Puig Campana on its western side
interrupts it upward. On the other side of the fault, a new sequence begins with
an alternation of silty marls and argillaceous limestones more than 170 meters
thick followed by about 30 meters of 'Hensonina calcarenites' with Neorbitolinopsis
conulus (Douvillé, 1912) (Pl. 9 ![]() , figs. A-F, U-X),
and a more than 650-meter-thick alternation of 'Hemiaster marls and
argillaceous limestones', still Albian in age. These 'Hensonina
calcarenites' are also exposed at Serreta Llarga, near the Serra de Fontcalent,
westward of Alicante city (Azéma, 1977).
, figs. A-F, U-X),
and a more than 650-meter-thick alternation of 'Hemiaster marls and
argillaceous limestones', still Albian in age. These 'Hensonina
calcarenites' are also exposed at Serreta Llarga, near the Serra de Fontcalent,
westward of Alicante city (Azéma, 1977).
|
|
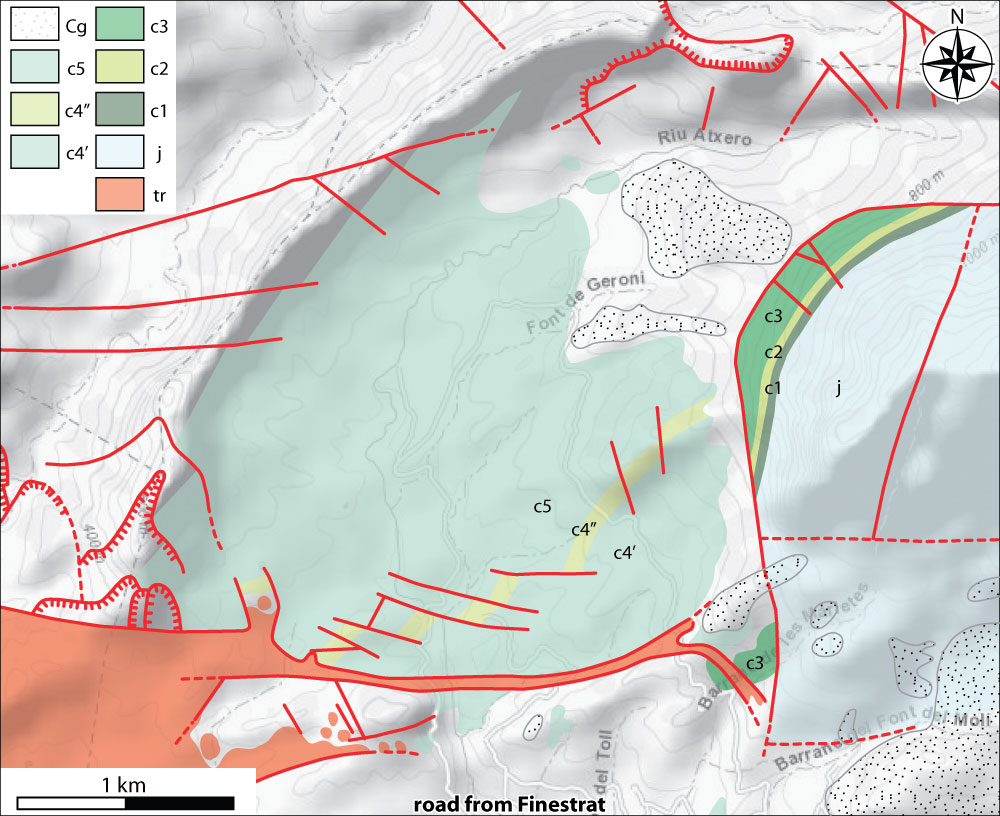
Figure 4: Puig Campana:
A) western side of Puig Campana (Cluse section arrowed), 12/03/1983. B)
condensed section above the 'Pseudocyclammina calcarenites and
sandstones' (lower Valanginian) and below the ammonite
marls (Barremian-Aptian), 15/03/1983. C) Cluse section, 1993. Figure captions: j: Jurassic
limestones (Berriasian and older strata), c1: 'Pseudocyclammina
calcarenites and sandstones' (lower Valanginian), c2: ammonite marls
(Barremian-Aptian), c3: calcisiltites, biolithites,
and rudistid limestones, c5: marls and argillaceous limestones (Albian),
Cg: 'mortadella' conglomerate (Quaternary), dotted lines: faults. |
At Cabeçó d'Or (Fig. 5 ![]() ),
similar to Puig Campana, the lower part of the sedimentary sequence begins on
top of lower Valanginian 'Pseudocyclammina calcarenites and sandstones'
with a condensed interval characterized by the occurrence of ferruginous ooids
and oncoids. Almost 2 meters thick, it spans part of the Valanginian to lower
Aptian interval, and numerous late Barremian and early Aptian ammonites (e.g.,
Deshayesites deshayesi Zone) may be collected there. This condensed
interval is overlain by a marly unit, more than 150 meters thick on the eastern
side of Cabeçó d'Or, containing early and late Aptian ammonites (e.g., Deshayesites
deshayesi, Dufrenoyia furcata, and Diadochoceras nodosocostatum
zones). Approximately 50 meters of 'silty limestones, calcisiltites, and
calcarenites with Favusella' follow. As
demonstrated for the first time in this location, they contain ammonites from
the latest Aptian and possibly the early Albian (BT 64). These are overlain
by approximately 20 meters of 'coral biolithites', which form a second
topographic crest (a first, distinct topographic crest is visible in the
underlying calcarenites, Fig. 5.A-B
),
similar to Puig Campana, the lower part of the sedimentary sequence begins on
top of lower Valanginian 'Pseudocyclammina calcarenites and sandstones'
with a condensed interval characterized by the occurrence of ferruginous ooids
and oncoids. Almost 2 meters thick, it spans part of the Valanginian to lower
Aptian interval, and numerous late Barremian and early Aptian ammonites (e.g.,
Deshayesites deshayesi Zone) may be collected there. This condensed
interval is overlain by a marly unit, more than 150 meters thick on the eastern
side of Cabeçó d'Or, containing early and late Aptian ammonites (e.g., Deshayesites
deshayesi, Dufrenoyia furcata, and Diadochoceras nodosocostatum
zones). Approximately 50 meters of 'silty limestones, calcisiltites, and
calcarenites with Favusella' follow. As
demonstrated for the first time in this location, they contain ammonites from
the latest Aptian and possibly the early Albian (BT 64). These are overlain
by approximately 20 meters of 'coral biolithites', which form a second
topographic crest (a first, distinct topographic crest is visible in the
underlying calcarenites, Fig. 5.A-B ![]() ). A bored erosional subaerially-exposed
surface (Fig. 5.C-D
). A bored erosional subaerially-exposed
surface (Fig. 5.C-D ![]() ) marks the top of these 'coral biolithites' and the base of
several tens of meters of 'Hemiaster marls and argillaceous limestones',
the first meters of which contain early late Albian ammonites.
) marks the top of these 'coral biolithites' and the base of
several tens of meters of 'Hemiaster marls and argillaceous limestones',
the first meters of which contain early late Albian ammonites.
|
|
Figure 5: Cabeçó d'Or:
A-B) Sanatorio section (Granier, 1987: Fig. 43): white arrows: first crest, yellow arrows: second crest,
A) 1993, B) Google Earth view
© 2024 Airbus. C-D) Sanatorio section,
exposure surface at the top of the second Albian crest (GPS coordinates:
38°30'53.5"N, 0°21'51.7"W), C) Photo courtesy of
Eric Monteil,
20/04/2024, D)
20/04/2024. |
Most
ammonites collected by the first author (B.R.C.G.) in the Costa Blanca
(SE Spain) were temporarily
housed in Robert Busnardo's collection in Saint-Didier-au-Mont-d'Or,
awaiting their transfer to the 'Collections de l'Université Claude Bernard Lyon
1'. During this time some were lost. On April 20, 2024, the first author (B.R.C.G.)
and Eric Monteil visited a site near Aigües where the top of the
Sanatorio section is exposed to collect new specimens. All these
ammonites are deposited in the 'Collections
de l'Université Claude Bernard Lyon 1' (with the UCBL-FSL label, as
mentioned in the plate legend).
At
Serra Gelada, specifically at Morro San Jordi (25/07/1984, circa 38°33'16.4"N,
0°03'27.3"W), above the 0.5-meter-thick condensed interval containing late
Barremian and early Aptian ammonites, additional specimens were collected from
the Deshayesites deshayesi Zone (lower Aptian) and the Epicheloniceras
martini Zone (upper Aptian). However, the youngest ammonites from Serra
Gelada were collected in situ at Punta de les Caletes (Fig.
3.A-B ![]() , 13/04/1984, circa
38°31'31.3"N,
0°05'46.3"W). These
specimens (HL 60) include Tegoceras
sp., indicative of the upper part of the Douvilleiceras mammillatum Zone
(lower
Albian) (Robert et al., 2009; Jaillard et al.,
2024).
, 13/04/1984, circa
38°31'31.3"N,
0°05'46.3"W). These
specimens (HL 60) include Tegoceras
sp., indicative of the upper part of the Douvilleiceras mammillatum Zone
(lower
Albian) (Robert et al., 2009; Jaillard et al.,
2024).
The youngest ammonites from Puig Campana ('Acanthohoplites' bigoureti,
'A.'
nolani, A. sp., Aconeceras sp., and Diadochoceras
sp.) were collected in the Cluse section (Fig. 5.C ![]() , Cluse 20 = ZC16503, 20/03/1983; ZC16631, 13/05/1985;
38°35'39.3"N,
0°12'30.2"W). They are
ascribed to the Diadochoceras nodosocostatum Zone of the upper Aptian (Giraud
et al., 2021).
, Cluse 20 = ZC16503, 20/03/1983; ZC16631, 13/05/1985;
38°35'39.3"N,
0°12'30.2"W). They are
ascribed to the Diadochoceras nodosocostatum Zone of the upper Aptian (Giraud
et al., 2021).
At the aforementioned Sanatorio
section of the Cabeçó d'Or,
north of Aigües (Fig. 5.A-B ![]() , 26/04/1985,
starting circa 38°31'07.0"N, 0°22'06.3"W, and ending circa
38°30'53.5"N,
0°21'51.6"W), the same Diadochoceras nodosocostatum Zone is at
least 40 m thick
there (samples BT43 to BT51). Ammonites collected in this interval comprise 'Acanthohoplites'
bigoureti (Seunes, 1887), 'A.' nolani (Seunes,
1887), 'A.' sp., Epicheloniceras sp., Diadochoceras sp. and
Neodufrenoyia sp. Lower in the section (samples BT24 to BT27), the
assemblage consists of 'Acanthohoplites' bigoureti, 'A.'
nolani, 'A.' sp., Aconeceras sp., Colombiceras
sp., Eogaudryceras sp., Epicheloniceras sp., Mathoceras matho
(Pervinquière, 1907), Parahoplites sp., Phylloceras (Hypophylloceras)
sp., ? Valdedorsella sp., and Zuercherella zuercheri (Jacob
et al., 1906), which suggests the Parahoplites melchioris Zone.
Ammonites from samples BT 28 and BT29 do not allow us to discriminate the Parahoplites
melchioris Zone from the Diadochoceras nodosocostatum Zone.
, 26/04/1985,
starting circa 38°31'07.0"N, 0°22'06.3"W, and ending circa
38°30'53.5"N,
0°21'51.6"W), the same Diadochoceras nodosocostatum Zone is at
least 40 m thick
there (samples BT43 to BT51). Ammonites collected in this interval comprise 'Acanthohoplites'
bigoureti (Seunes, 1887), 'A.' nolani (Seunes,
1887), 'A.' sp., Epicheloniceras sp., Diadochoceras sp. and
Neodufrenoyia sp. Lower in the section (samples BT24 to BT27), the
assemblage consists of 'Acanthohoplites' bigoureti, 'A.'
nolani, 'A.' sp., Aconeceras sp., Colombiceras
sp., Eogaudryceras sp., Epicheloniceras sp., Mathoceras matho
(Pervinquière, 1907), Parahoplites sp., Phylloceras (Hypophylloceras)
sp., ? Valdedorsella sp., and Zuercherella zuercheri (Jacob
et al., 1906), which suggests the Parahoplites melchioris Zone.
Ammonites from samples BT 28 and BT29 do not allow us to discriminate the Parahoplites
melchioris Zone from the Diadochoceras nodosocostatum Zone.
The
ultimate ammonite zone of the Aptian, the Hypacanthoplites elegans Zone (Szives
et al., 2023, 2024), was not identified in any of the three areas
examined. However, a questionable Fallotermiericeras sp. (BT 64) was
found in the silty limestones with Favusella within the Sanatorio
section, approximately 50 meters above the Diadochoceras nodosocostatum
Zone. This occurrence corresponds to either the uppermost part of the Diadochoceras
nodosocostatum Zone (uppermost Aptian), the 'Hypacanthoplites'
elegans Zone (marking the Aptian-Albian transition), or the Douvilleiceras
leightonense Zone (lowermost Albian). To explicitly identify the Hypacanthoplites
elegans Zone, further material must be collected from Serra Gelada and Cabeçó
d'Or.
Back
at Cabeçó d'Or, the ammonites collected above the discontinuity at the
top of the Sanatorio section (26/04/1985,
38°30'53.5"N, 0°21'51.6"W), previously illustrated (Granier,
1987: Pl. 47), are indicative of the lower upper Albian. These include Hysteroceras sp., Dipoloceras
sp., and Brancoceras sp. The most
representative specimens are re-illustrated here (Pl. 10 ![]() , figs. A-I). New
ammonites from Aïgues, recently collected by one of us (B.R.C.G.) and Eric Monteil,
include Hysteroceras sp., Dipoloceras
sp., Hamites sp., Anisoceras
sp., and Pusozia (Puzosia) sp. (Pl. 10
, figs. A-I). New
ammonites from Aïgues, recently collected by one of us (B.R.C.G.) and Eric Monteil,
include Hysteroceras sp., Dipoloceras
sp., Hamites sp., Anisoceras
sp., and Pusozia (Puzosia) sp. (Pl. 10 ![]() , figs. J-W). This association confirms an early late Albian age.
, figs. J-W). This association confirms an early late Albian age.
In 1970, in the Province of Valencia (SE Spain),
Fourcade described the
Solana del Alambin section starting
from the ruins of Casa de Anton Julian (38°59'22.1"N, 0°54'48.8"W)
to the summit (1098 m), a few hundreds of meters north of Casa del Alto (38°59'19.5"N,
0°56'03.7"W). He stated that 'This series from the Solana del Alambin
shows that Neorbitolinopsis conulus appears immediately above the Knemiceras'
[translated from the French in Fourcade, 1970:
'Cette série de la Solana del Alambin permet de montrer que les Neorbitolinopsis
conulus apparaissent immédiatement au dessus des Knemiceras'].
He also reported other records of these same ammonites from 1) Casa del Alambín
(presently Casa María Rodevar) at 39°2'9.13"N, 0°51'37.64"W, and 2)
Casa de la Canaleja, 7 km SW (not SE, as stated in the thesis text) of Solana
del Alambín, at 38°56'12.75"N, 0°57'4.76"W. Since then, these
ammonite findings have supported the attribution of Neorbitolinopsis conulus
to the late Albian. However, these ammonites, which are also part of the
'Collections de l'Université Claude Bernard Lyon 1', are not Knemiceras sp.
but Parengonoceras sp. (Pl. 11 ![]() , figs. A-E), based on their
characteristic alternating
elongated ventrolateral clavi (Latil & Jaillard,
2024).
Although its internal calcareous mold is poorly preserved, one specimen retains
the diagnostic morphology of Parengonoceras
ibericum (Arias
& Wiedmann, 1977) (Pl. 11
, figs. A-E), based on their
characteristic alternating
elongated ventrolateral clavi (Latil & Jaillard,
2024).
Although its internal calcareous mold is poorly preserved, one specimen retains
the diagnostic morphology of Parengonoceras
ibericum (Arias
& Wiedmann, 1977) (Pl. 11 ![]() , figs. D-E), which corresponds to the
Douvilleiceras mammillatum Zone (and
possibly the Lyelliceras pseudolyelli
Zone) of the upper lower Albian.
, figs. D-E), which corresponds to the
Douvilleiceras mammillatum Zone (and
possibly the Lyelliceras pseudolyelli
Zone) of the upper lower Albian.
The following paragraphs address some nomenclatural errors or misidentifications from the past (Granier, 1987, 2019; Granier & Perthuisot, 2009). They also integrate recent taxonomic and stratigraphic updates:
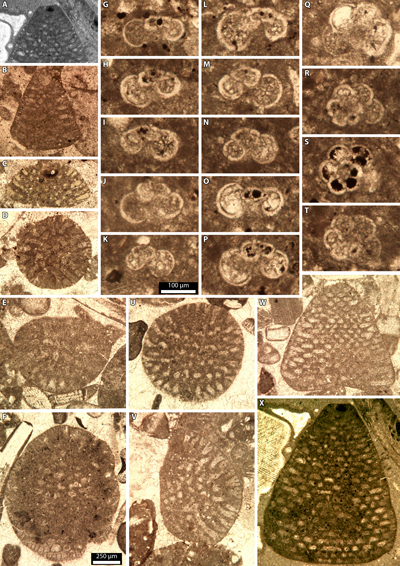
1) Hensonina lenticularis (Henson, 1947) nom. nud., comb. nud., was cited by the first author from both the Puig Campana and Serra Gelada areas, specifically from the Realet d'Alt and Rafel sections in the Puig Campana area, as well as the Relais section in the Serra Gelada area (Granier, 1987, 2019; Granier & Perthuisot, 2009). Recently, both its generic and specific status were examined (Rigaud et al., 2023), leading to the revision of the genus Hensonina Rigaud & Consorti ex Moullade & Peybernès, 2023 (non 1973)[*], and the establishment of the species Hensonina canalicula Rigaud & Consorti in Rigaud et al., 2023.
Earlier reports of this species comprise:
a) in Granier (1987: p. 108, no illustration) or in Granier and Perthuisot (2009: p. 1021-1022, no illustration) in the Albian 'Cylindroporella Oolithes' from the Relais section of the Serra Gelada (Granier, 1987: Fig. 23), samples Relais 10 - HL 712, Relais 12 - HL 722, and Relais 13 (not 14) - HL 726, correspond to Epistomina spp. (Granier, 2019: Pl. 3, figs. A-H). This trochospiral aragonitic foraminiferal genus is a useful marker for paleoenvironments, as it is commonly found in outer platform environments; however, it has no biostratigraphic value. For instance, Epistomina is also known from the Berriasian slope deposits of Busot (Granier et al., 2024).
b) in Granier (1987: p. 58, Pl. 8, figs. f-i) in the Albian 'Simplorbitolina Limestones' from the Realet d'Alt section of the Puig Campana (Granier, 1987: Fig. 12), samples Realet d'Alt 13 - ZC19368 to Realet d'Alt 28 - ZC19351, correspond to Involutina hungarica (Sidó, 1952) (Granier, 2019: Pl. 2, figs. H-AB; Pl. 3, figs. L-AH). According to Rigaud et al. (2023), this species ranges from the upper Aptian to the middle Albian. However, none of these datings are directly correlated with ammonite finds; therefore, they lack rigorous calibration against ammonite biozones.
c) in Granier (1987: as Hensonina lenticularis, p. 63, Pl. 10, figs. a-b, e; 2019: as Involutina hungarica, p. 446, Pl. 3, figs. AI-AJ, Pl. 4, figs. A-V) in the Albian 'Hensonina Calcarenites' from the Rafel section of the Puig Campana (Granier, 1987: Fig. 13), sample Rafel 41 - PC19499, correspond to genuine Hensonina canalicula Rigaud & Consorti, 2023. According to Rigaud et al. (2023), this species is only known from the Albian interval.
2) Mesorbitolina subconcava (Leymerie, 1878) was reported by the first author from the Puig Campana, Serra Gelada, and Cabeçó d'Or areas, specifically in the 'Pieninia Calcarenites' from the Cluse and Realet d'Alt sections in the Puig Campana area, in the 'Cylindroporella Oolites' and 'rudistid limestones' from the Relais and Carabiniers sections in the Serra Gelada area, and in the 'Favusella Calcarenites' from the western Cabeçó d'Or area (Granier, 1987; Granier & Perthuisot, 2009). Although Schroeder (letter to the first author, 26/11/1987) initially ascribed these specimens to M. subconcava (Leymerie, 1878), they should be reascribed to Mesorbitolina birmanica Sahni, 1937, a species that escaped from oblivion (Schlagintweit & Wilmsen, 2014).
3)
According to Schlagintweit (2023), the specimen of Coskinolinoides
texanus Keijzer, 1942, from the 'rudistid limestones' of the Illa de Benidorm, illustrated by Granier
(1987: Pl. 27, fig. c; here Pl. 2 ![]() , fig. M), should be
assigned to the species C. fleuryi (Decrouez & Moullade,
1974). However, since this specimen is an early representative of the genus, we have chosen to retain
Fourcade's
(in Granier, 1987) original attribution.
, fig. M), should be
assigned to the species C. fleuryi (Decrouez & Moullade,
1974). However, since this specimen is an early representative of the genus, we have chosen to retain
Fourcade's
(in Granier, 1987) original attribution.
4) Schlagintweit et al. (2021) document the Simplorbitolina
lineage with S. aquitanica, S. manasi, and S. conulus,
which appears to provide strong support for biozones based on first occurrences
of these species (e.g., Granier, 1987: Fig. 12). They have treated
S. chauvei as a junior synonym of S. aquitanica. However, 'The original description of Orbitolinopsis aquitanica by Schroeder
in Schroeder & Poignant
(1964) was obviously based on an
assemblage of juvenile specimens' (Schlagintweit et al.,
2021: p. 5) or megalospheric specimens (Moullade & Peybernès,
1979). It is assumed that making S. aquitanica and S. chauvei synonymous
would require an in-depth revision of the original material of Schroeder
and Poignant (1964), and that of Fourcade
(1978).
Simplorbitolina spp. are only known from Puig Campana in mud-dominated facies, which are absent or scarce at Cabeçó d'Or and Serra Gelada. The Simplorbitolina lineage with S. chauvei, S. manasi, and S. conulus (Fourcade, 1978), apparently provided strong support for biozones based on the first occurrences of the three species (Schlagintweit et al., 2021). Granier (1987: Fig. 12) used them, combined with the occurrences of Involutina hungarica (as 'Hensonina lenticularis') and Pseudochoffatella cuvillieri, to split the rudistid limestones of the Realet d'Alt section at Puig Campana into 4 intervals:
Nonetheless, subsequent to the recent revision of Granier's (1987) original material during the preparation of this manuscript, species delimitation - based on the presence or absence of intercalary beams and, when the intercalary beams are present, the presence or absence of rafters - has yielded somewhat different results.
The first species of the
evolutionary lineage, S. chauvei, has no intercalary beams and no rafters
(Pl. 2 ![]() , figs. O-R;
Pl. 8
, figs. O-R;
Pl. 8 ![]() , figs. A-B, N). The following species, S. manasi,
is characterized by the presence of intercalary beams, but has no rafters (Pl. 2
, figs. A-B, N). The following species, S. manasi,
is characterized by the presence of intercalary beams, but has no rafters (Pl. 2 ![]() , figs. T-U;
Pl. 8
, figs. T-U;
Pl. 8 ![]() , figs. F-G, O). The last species, S. conulus, is
characterized by the presence of both intercalary beams and rafters (Pl. 2
, figs. F-G, O). The last species, S. conulus, is
characterized by the presence of both intercalary beams and rafters (Pl. 2 ![]() ,
figs. V-W; Pl. 8
,
figs. V-W; Pl. 8 ![]() , figs. C-E, H-L, Q-T). Locally, some intermediate forms may be
found,
e.g., an intermediate form between S. manasi and S. conulus
in sample Realet 14 (Pl. 8
, figs. C-E, H-L, Q-T). Locally, some intermediate forms may be
found,
e.g., an intermediate form between S. manasi and S. conulus
in sample Realet 14 (Pl. 8 ![]() , fig. P). Here, we argue that these transitional
forms should be assigned to the successor species epithet, rather than the
ancestor species epithet, because it cannot be ruled out that, in some cases,
they represent teratological specimens.
, fig. P). Here, we argue that these transitional
forms should be assigned to the successor species epithet, rather than the
ancestor species epithet, because it cannot be ruled out that, in some cases,
they represent teratological specimens.
5) Sabaudia minuta (Hofker, 1965) and related species are quite common in the ?upper Aptian - Albian strata of the studied area. Textulariella minuta Hofker, 1965, is the type species of the genus Sabaudia Charollais & Brönnimann, 1965. Recently, Özdikmen (2009) pointed out that this genus name was initially occupied by Sabaudia Ghigi, 1909, necessitating the adoption of a replacement name. Accordingly, Özdikmen (2009) introduced the new genus name Akcaya to replace Sabaudia Charollais & Brönnimann, 1965. However, Sabaudia liguriae Ghigi, 1909, the type species of the genus Sabaudia Ghigi, 1909, proved to be a junior synonym of Hormiphora australis (Benham, 1907). Consequently, the genus name Sabaudia Ghigi, 1909, which is synonymized with Hormiphora Agassiz, 1860, is no longer in use. The replacement name Akcaya introduced by Özdikmen (2009) is unnecessary because the name Sabaudia Charollais & Brönnimann, 1965, is no longer preoccupied. The genus Sabaudia Charollais & Brönnimann, 1965, is valid and the species Sabaudia minuta (Hofker, 1965) Charollais & Brönnimann, 1965, retains its genus name, as the preoccupation by the earlier Sabaudia Ghigi, 1909, is resolved through synonymization under Hormiphora Agassiz, 1860. The lowest record of Sabaudia minuta is from lower Valanginian strata at 'Les Quatre Confronts', northward of Toulon, Var, France (Masse, 1976: Pl. 14, figs. 20-21). Therefore, its total range spans the Valanginian - Albian (-? lower Cenomanian) interval, not the upper Hauterivian - lower Cenomanian interval, as stated by Arnaud-Vanneau and Chiocchini (in Schroeder & Neumann, 1985).
6) Favusella washitensis (Carsey, 1926) was considered a marker of the Albian stage. However, according to https://www.mikrotax.org/pforams/index.php?id=110109, this species is now thought to span the late Aptian to the middle Cenomanian interval.
7) The calcareous green alga, Cylindroporella barnesii Johnson, 1954, was first described from the Edwards Formation (middle to early late Albian in age, Scott et al., 2019) of the Gillespie County (Texas, U.S.A.). In the Costa Blanca, it is found in the 'Simplorbitolina limestones' in the Realet d'Alt section at Puig Campana (from sample Realet 8 up to sample Realet 28), and in the 'Cylindroporella oolithes' in the Relais section (from sample Relais 2 up to sample Relais 19) and the 'rudistid limestones' in the Carabiniers section (in samples Carabiniers 10 and 13) at Serra Gelada.
8) The calcareous red alga, Agardhiellopsis cretacea Lemoine, 1966, was originally described from P. Feuillée's material collected at Uztegui (Navarra, Spain) in strata ascribed to 'Albien approximatif'. Besides Spain, the author (Lemoine, 1966) mentions several localities in the 'Vimport facies' of southern France, including Ariège, Aude, Basses-Pyrénées, and Les Landes. In the Costa Blanca, this alga is found in the 'Pieninia calcarenites' in the Cluse section (in sample Cluse 30) and in the Realet d'Alt section (in sample Realet 4) at Puig Campana, in the 'orbitolinid calcarenites and marls' in the 'LM' section (in samples LM 11 to 18) at Serra Gelada, and in the 'calcarenites with Favusella', north of Sanatorio, at Cabeçó d'Or (in sample BT 8).
Table 1: List of occurrences of algae and LBF in the studied areas.
| Serra Gelada | Illa de Benidorm | Puig Campana | Cabeçó d'Or | Serreta Llarga | |
| Agardhiellopsis cretacea | X | X | X | ||
| Cylindroporella barnesii | X | ||||
| Coskinolinoides texanus | 1 | 1 | |||
| Involutina hungarica | X | ||||
| Hensonina canalicula | X | X | |||
| Neorbitolinopsis conulus | X | X | |||
| Mesorbitolina birmanica | X | X | X | X | |
| Mesorbitolina texana | X | X | |||
| Pseudochoffatella cuvillieri | X | X | X | ||
| Simplorbitolina aquitanica | X | ||||
| Simplorbitolina chauvei | X | ||||
| Simplorbitolina manasi | X | ||||
| Simplorbitolina conulus | X |
At Puig Campana, Granier's (1987) original material was revised by sticking to the strict rules used
to discriminate the three Simplorbitolina species. S. chauvei, the
oldest species, first occurs in the Realet d'Alt section, approximately 15
meters above the 'coral biolithites' (in samples Realet 8-10). The next species,
S. manasi, first occurs 35 meters above the same unit (in sample Realet
12). Although the youngest species, S. conulus, is present approximately
60 meters above the same unit (in sample Realet 14), it is likely that it first
occurs approximately 30 meters below, i.e., approximately 5 meters below
the first occurrence of S. manasi (in sample Realet 11: Pl. 2 ![]() , fig. S;
Pl. 8
, fig. S;
Pl. 8 ![]() , fig. M). Surprisingly, the Simplorbitolina manasi Zone is not
evidenced here because the first occurrence of S. conulus precedes that
of S. manasi.
, fig. M). Surprisingly, the Simplorbitolina manasi Zone is not
evidenced here because the first occurrence of S. conulus precedes that
of S. manasi.
Involutina hungarica first occurs approximately 50 meters above the 'coral biolithites', in the Simplorbitolina conulus Zone, and its range extends up to the top of the 'Simplorbitolina limestones' in the Realet d'Alt section at Puig Campana where its total range falls within the Simplorbitolina conulus Zone.
Still,
at Puig Campana, Pseudochoffatella cuvillieri first appears in the Realet
d'Alt section, approximately 80 meters above the base of the 'rudistid
limestones', which is approximately 110 meters (sample Realet 14; Pl. 4 ![]() , fig. D)
from the base of the 'Pieninia calcarenites' and 'coral biolithites',
which are, in turn, above strata containing ammonites of the Diadochoceras
nodosocostatum Zone. Consequently, in this area, the first appearance of Pseudochoffatella
cuvillieri is more than 130 meters above these same latest Aptian ammonites
(with sample Realet 14). In addition, the Pseudochoffatella cuvillieri (Sub-)
Zone falls within the Simplorbitolina conulus Zone, not within the Simplorbitolina
manasi Total Range Zone as stated before (e.g.,
Fourcade, 1970; Peybernès,
1976).
, fig. D)
from the base of the 'Pieninia calcarenites' and 'coral biolithites',
which are, in turn, above strata containing ammonites of the Diadochoceras
nodosocostatum Zone. Consequently, in this area, the first appearance of Pseudochoffatella
cuvillieri is more than 130 meters above these same latest Aptian ammonites
(with sample Realet 14). In addition, the Pseudochoffatella cuvillieri (Sub-)
Zone falls within the Simplorbitolina conulus Zone, not within the Simplorbitolina
manasi Total Range Zone as stated before (e.g.,
Fourcade, 1970; Peybernès,
1976).
At Serra Gelada, Pseudochoffatella
cuvillieri first appears in the Relais section (Pl. 4 ![]() , fig. C) approximately
25 meters below the 'rudistid limestones', i.e., approximately 65 meters
from the base of the 'Cylindroporella oolithes', and approximately 215
meters from the base of the 150 meters of 'siltstones and calcisiltites with Favusella',
which is in turn approximately 40 meters above strata containing Tegoceras sp., which is attributed to
the Douvilleiceras mammillatum Zone.
Accordingly, the first appearance of Pseudochoffatella cuvillieri is more
than 250 meters above this early Albian ammonite.
, fig. C) approximately
25 meters below the 'rudistid limestones', i.e., approximately 65 meters
from the base of the 'Cylindroporella oolithes', and approximately 215
meters from the base of the 150 meters of 'siltstones and calcisiltites with Favusella',
which is in turn approximately 40 meters above strata containing Tegoceras sp., which is attributed to
the Douvilleiceras mammillatum Zone.
Accordingly, the first appearance of Pseudochoffatella cuvillieri is more
than 250 meters above this early Albian ammonite.
|
|
Figure 6: Borings from
the hardground, equivalent to the sommital surface of bioherm D.
A-D) Relais 15 (HL 724 bis), Cylindroporella oolithes,
Relais section, Serra Gelada, 01/05/1985. Scale bar (on fig. D) = 250 µm. |
Mesorbitolina texana also
first appears in the Relais section at Serra Gelada (Pl. 5 ![]() , figs. L, R-S), about
60 meters below the 'rudistid limestones', which is approximately 30 meters from
the base of the 'Cylindroporella oolithes'. Thus, the first appearance of
Mesorbitolina texana is approximately 220 meters above the early
Albian ammonite.
, figs. L, R-S), about
60 meters below the 'rudistid limestones', which is approximately 30 meters from
the base of the 'Cylindroporella oolithes'. Thus, the first appearance of
Mesorbitolina texana is approximately 220 meters above the early
Albian ammonite.
Mesorbitolina
birmanica is found
at Serra Gelada in the 'rudistid
limestones' (Pl. 5 ![]() , figs. N-Q;
Pl. 6
, figs. N-Q;
Pl. 6 ![]() , figs. A-D, H-AM;
Pl. 7
, figs. A-D, H-AM;
Pl. 7 ![]() , figs. A-AO), but
also both a few meters below (Pl. 5
, figs. A-AO), but
also both a few meters below (Pl. 5 ![]() , figs. E-K;
Pl. 6
, figs. E-K;
Pl. 6 ![]() , fig. G) or above this
unit (Pl. 5
, fig. G) or above this
unit (Pl. 5 ![]() , fig. M). Therefore, its first appearance is more than 250 meters
above the early Albian ammonites. At Puig Campana, the latter first appears in
the Cluse section (Pl. 5
, fig. M). Therefore, its first appearance is more than 250 meters
above the early Albian ammonites. At Puig Campana, the latter first appears in
the Cluse section (Pl. 5 ![]() , fig. U), only 30 meters above
the latest Aptian ammonites. It is worth noting that the relatively reduced
thickness of the 'ammonites marls' there likely results from the peculiar
structural setting of Puig Campana. At Cabeçó d'Or, this
species first appears in the Sanatorio section (Pl. 5
, fig. U), only 30 meters above
the latest Aptian ammonites. It is worth noting that the relatively reduced
thickness of the 'ammonites marls' there likely results from the peculiar
structural setting of Puig Campana. At Cabeçó d'Or, this
species first appears in the Sanatorio section (Pl. 5 ![]() , fig. Y),
approximately 50 meters above ammonites of the Diadochoceras nodosocostatum Zone
and about 20 meters below a
questionable Fallotermiericeras sp.
(BT 64),
which is attributed to either the uppermost part of
the Diadochoceras nodosocostatum Zone, the Hypacanthoplites elegans Zone, or the Douvilleiceras
leightonense Zone.
, fig. Y),
approximately 50 meters above ammonites of the Diadochoceras nodosocostatum Zone
and about 20 meters below a
questionable Fallotermiericeras sp.
(BT 64),
which is attributed to either the uppermost part of
the Diadochoceras nodosocostatum Zone, the Hypacanthoplites elegans Zone, or the Douvilleiceras
leightonense Zone.
In the Costa Blanca, the green
CA, Cylindroporella barnesii Johnson, 1954, is found at Puig
Campana (Pl. 2 ![]() , figs. E, G-J) and Serra Gelada
(Pl. 2
, figs. E, G-J) and Serra Gelada
(Pl. 2 ![]() , figs. F, K-L); it was not
found at Cabeçó d'Or. The red CA, Agardhiellopsis cretacea Lemoine,
1966, is also found both at Puig Campana (Pl. 2
, figs. F, K-L); it was not
found at Cabeçó d'Or. The red CA, Agardhiellopsis cretacea Lemoine,
1966, is also found both at Puig Campana (Pl. 2 ![]() , fig. A) and Serra Gelada
(Pl. 2
, fig. A) and Serra Gelada
(Pl. 2 ![]() , figs. C-D), as well as at Cabeçó d'Or
(Pl. 2
, figs. C-D), as well as at Cabeçó d'Or
(Pl. 2 ![]() , fig. B).
, fig. B).
Ordering the Simplorbitolina species by their order of appearance should document their lineage (Fourcade, 1978; Schlagintweit et al., 2021), with S. chauvei appearing first, followed by S. manasi and S. conulus, providing strong support for biozones based on the first occurrences of these species (e.g., Granier, 1987: Fig. 12). However, the revision of the succession at Puig Campana slightly disrupted this model, as the S. conulus was identified before the first occurrence of S. manasi. This could be related to sparse sampling intervals.
Considering the youngest
ammonite record at Punta de
les Caletes (Serra Gelada),
the Aptian/Albian boundary likely lies near the top of the ammonite marls, below
the 150-meter-thick siltstones and calcisiltites with Favusella.
Similarly, at Cabeçó d'Or and Puig Campana, it is expected to be near the top
of the ammonite marls, below the calcisiltites and calcarenites with Favusella
and beneath the Pieninia calcarenites, respectively. The shallow-water
facies at Serra Gelada, Puig Campana, and Cabeçó d'Or, which represent the
upper part of a shallowing-upward sequence, are likely of the same early Albian
age. Accordingly, the
presence of most previously mentioned species (i.e., Favusella
washitensis, Mesorbitolina texana, M. birmanica, Agardhiellopsis cretacea, Simplorbitolina chauvei, Cylindroporella
barnesii, Involutina hungarica, Simplorbitolina conulus, S.
manasi, Pseudochoffatella cuvillieri, and Coskinolinoides
texanus, ranged
by time-relative order of appearance) in lower Albian strata is confirmed.
However,
this sorting could be misleading, as it merely reflects the regional
shallowing-upward trend, which causes facies changes from ammonite marls to
rudistid limestones at Serra Gelada, to Simplorbitolina limestones at
Puig Campana, or to coral biolithites at Cabeçó d'Or. Besides evolutionary
processes, the appearance or disappearance of species in a shallow-water marine
sequence are controlled by the presence or absence of the corresponding facies. Favusella
washitensis (Pl. 1 ![]() , figs. A-Z) and Epistomina
spp.
(Pl. 3
, figs. A-Z) and Epistomina
spp.
(Pl. 3 ![]() , figs.
D-F) are predominantly found in outer-shelf mud-dominated facies, while Mesorbitolina
texana and M. birmanica are mostly found in outer-shelf grain-dominated facies. In contrast, Coskinolinoides texanus, Involutina hungarica, Simplorbitolina
chauvei, S. conulus, and S. manasi are primarily found in
inner-shelf mud-dominated facies, which are mainly present at Puig Campana. Agardhiellopsis
cretacea, Cylindroporella barnesii, and Pseudochoffatella
cuvillieri can be found in both environments and in intermediate shoal
facies. These ecological features justify treating Pseudochoffatella
cuvillieri, Mesorbitolina,
and Simplorbitolina zones in separate columns.
, figs.
D-F) are predominantly found in outer-shelf mud-dominated facies, while Mesorbitolina
texana and M. birmanica are mostly found in outer-shelf grain-dominated facies. In contrast, Coskinolinoides texanus, Involutina hungarica, Simplorbitolina
chauvei, S. conulus, and S. manasi are primarily found in
inner-shelf mud-dominated facies, which are mainly present at Puig Campana. Agardhiellopsis
cretacea, Cylindroporella barnesii, and Pseudochoffatella
cuvillieri can be found in both environments and in intermediate shoal
facies. These ecological features justify treating Pseudochoffatella
cuvillieri, Mesorbitolina,
and Simplorbitolina zones in separate columns.
Neorbitolinopsis conulus
(Pl. 9 ![]() , figs. A-F, U-W) and Hensonina canalicula are reported not only
from the 'Hensonina calcarenites' at Puig Campana but also from the
Serreta Llarga (Azéma, 1977). Based on the revision of Fourcade's
'Knemiceras' by the second author (E.R), the first species
should no longer be regarded as an upper Albian marker
but is likely a marker characterizing the upper
lower Albian. Every earlier reference in the literature to a late Albian
dating based on the presence of Neorbitolinopsis conulus should be
reexamined carefully. For instance, in the Albian strata of the 'cove of
Baforeira' section, west of Lisboa (Portugal), Berthou and Schroeder
(1978: Pl. 2, figs. 1, 5-7; Pl. 3, figs. 1-5) documented the occurrence of Neorbitolinopsis
conulus just below a bored surface (ibid.: Pl. 9, fig. 3), which
marks the boundary with Cenomanian strata containing alveolinids. Initially, Berthou
(1971) regarded the entire section as Cenomanian in age, whereas Berthou
and Schroeder (1978) assigned the strata containing Neorbitolinopsis
conulus to the upper substage of the Albian. In light of the new hypothesis
regarding the stratigraphic range of Neorbitolinopsis conulus, the
stratigraphic gap at the Albian/Cenomanian boundary near Lisboa could be more
significant than previously thought.
, figs. A-F, U-W) and Hensonina canalicula are reported not only
from the 'Hensonina calcarenites' at Puig Campana but also from the
Serreta Llarga (Azéma, 1977). Based on the revision of Fourcade's
'Knemiceras' by the second author (E.R), the first species
should no longer be regarded as an upper Albian marker
but is likely a marker characterizing the upper
lower Albian. Every earlier reference in the literature to a late Albian
dating based on the presence of Neorbitolinopsis conulus should be
reexamined carefully. For instance, in the Albian strata of the 'cove of
Baforeira' section, west of Lisboa (Portugal), Berthou and Schroeder
(1978: Pl. 2, figs. 1, 5-7; Pl. 3, figs. 1-5) documented the occurrence of Neorbitolinopsis
conulus just below a bored surface (ibid.: Pl. 9, fig. 3), which
marks the boundary with Cenomanian strata containing alveolinids. Initially, Berthou
(1971) regarded the entire section as Cenomanian in age, whereas Berthou
and Schroeder (1978) assigned the strata containing Neorbitolinopsis
conulus to the upper substage of the Albian. In light of the new hypothesis
regarding the stratigraphic range of Neorbitolinopsis conulus, the
stratigraphic gap at the Albian/Cenomanian boundary near Lisboa could be more
significant than previously thought.
|
|
Figure 7: Geological
map of Puig Campana (excerpt from Granier, 1987). Figure captions: tr: Trias, j: Jurassic limestones (Berriasian and older
strata), c1: 'Pseudocyclammina calcarenites and sandstones' (lower
Valanginian), c2: ammonite
marls (Barremian-Aptian), c3: calcisiltites, biolithites, and rudistid
limestones, c4': silty marls and argillaceous limestones, c4": Hensonina
calcarenites, c5: marls and argillaceous limestones (Albian), Cg: 'mortadella'
conglomerate (Quaternary). |
Although it cannot be excluded that some species (e.g., Favusella washitensis, Mesorbitolina texana, M. birmanica, Agardhiellopsis cretacea, Simplorbitolina chauvei, Cylindroporella barnesii, Involutina hungarica) may have existed in the latest Aptian or earlier in other geographic areas, it is most likely, based on our local finds of ammonites and until further evidence emerges, that they did not appear before the early Albian in the Costa Blanca. In conclusion, although the implementation of biozones from other authors in different geographic areas (e.g., Schroeder, 1963; Fourcade, 1970; Peybernès, 1976; García Hernández, 1978; Jaffrezo, 1980; Masse et al., 1992) proves useful at the start of any new study, the use of their empirical datings is strongly discouraged.
Biozonations and total ranges of large benthic foraminifers (LBF) and calcareous algae (CA) still in use today are partially based on earlier empirical biozonal schemes, some of which date back more than half a century, as proposed by Schroeder (1963), Fourcade (1967), and others. Since then, no significant progress has been made in calibrating LBF and CA ranges (Table 1), or the associated biozones, onto the ammonite standard scale. For instance, the revision of Fourcade's 'Knemiceras' and the subsequent re-dating of strata containing Neorbitolinopsis conulus suggested that the correlation scheme for the Aptian-Albian interval in the Costa Blanca, initially proposed by the first author (Granier, 1987: Fig. 55; 1988: Fig. 1), required revision.
|
|
Figure 8: Correlations
of the sections from Cabeçó d'Or (a: Sanatorio section), Puig Campana
(b: Cluse section, c: Realet d'Alt section, d: Rafel section), Serra
Gelada (e: Morro San Jordi section, f: Punta de les Caletes section, g: Relais
section, h: Calle Dos Callas and
Camí del Far sections), Illa de Benidorm (i, with Serra
Gelada), and Serreta Llarga.
Figure captions: c2': condensed sections, c2": ammonite marls (offshore
facies, below storm wave base), c3': calcisiltites (lower shoreface facies,
above storm wave base and below fair weather wave base), c3": oolites
(upper
shoreface facies, above fair weather wave base), c3'": biolithites,
c3"": rudistid limestones, c4': silty marls and argillaceous
limestones, c4": calcarenites, c5: marls and argillaceous
limestones, Praeh.: Praehedbergella
spp., Fav.w.: Favusella washitensis, M.birm.: Mesorbitolina
birmanica, S.chauv.: Simplorbitolina chauvei, S.man.: S.
manasi, S.con.: S. conulus, S.chauv.: Involutina hungarica,
Ps.cuv.: Pseudochoffatella cuvillieri, Neorb.con.: Neorbitolinopsis
conulus, Hens.can.: Hensonina canalicula. 1-3, uppermost layers with
ammonites below the carbonate platform facies at Cabeçó d'Or, Puig Campana,
and Serra Gelada respectively;
4, the regression-transgression reversal found at Serra Gelada and likely also
occurring at Puig Campana; 5, the Hensonina calcarenites pinch in (i.e.,
thin out) as they approach the Cabeçó d'Or high; 6, lowermost layer with ammonites above the carbonate platform
facies at Cabeçó d'Or. |
Based
on this study, the biozones defined by the total range or the first occurrence
of Mesorbitolina birmanica, Simplorbitolina manasi,
S. conulus, Pseudochoffatella cuvillieri, and Neorbitolinopsis conulus
appear to fall within the early Albian substage (Fig. 9 ![]() ),
rather than in the late Aptian to late Albian interval, as indicated in most
biostratigraphic schemes.
),
rather than in the late Aptian to late Albian interval, as indicated in most
biostratigraphic schemes.
|
|
Figure 9: LBF
biozonal schema calibrated onto the standard ammonite scale for the Costa Blanca
(Alicante). The graphical representation of the evolutionary trend in the
Simplorbitolina lineage shows the coexistence of both an
ancestor and
its direct successor. |
The Albian Global Boundary Stratotype Section and Point were defined a few years ago, and this stage boundary can be identified in most basinal sections of the Tethys realm. However, none of the proxies selected by the Albian Working Group of the International Commission on Stratigraphy is useful for identifying this stage boundary in any nearby shallow-water sections.
The regression-transgression
reversal observed at Serra Gelada also occurs at Puig Campana (Fig.
8.4 ![]() ). At
Serra Gelada, the transgression is gradual, whereas at Puig Campana (north of
Barranc de les Marietes), it was likely less gradual. The thick hemipelagic
succession that follows at Puig Campana contrasts with the incomplete
stratigraphic record at Cabeçó d'Or,
suggesting a collapse of the seabed in the first area, correlated in time with a
ground heave in the second. These significant seabed movements may be related to
vertical shifts within the substratum, driven by the lateral migration of
Triassic salts.
). At
Serra Gelada, the transgression is gradual, whereas at Puig Campana (north of
Barranc de les Marietes), it was likely less gradual. The thick hemipelagic
succession that follows at Puig Campana contrasts with the incomplete
stratigraphic record at Cabeçó d'Or,
suggesting a collapse of the seabed in the first area, correlated in time with a
ground heave in the second. These significant seabed movements may be related to
vertical shifts within the substratum, driven by the lateral migration of
Triassic salts.
Based on the new dating of the
strata with Neorbitolinopsis
conulus, our study
reveals a significant local hiatus at Cabeçó d'Or during Albian times, which is wider than previously suspected,
spanning at least the entire middle Albian, as indicated by evidence of erosion
and non-deposition (Fig. 8 ![]() , in the interval 1 to 6). The Hensonina
calcarenites found in the thick Puig Campana and Serreta Llarga deep-water
sections likely pinch out around a local high at Cabeçó d'Or. The origin of
this high may be related to halokinesis of Triassic salts during Albian times (see
Granier
& Perthuisot, 2009: Fig. 8). In fact, Granier (1987:
Pl. 25, figs. d-h) already reported the presence of reworked Triassic material (Triassic
quartz with salt inclusions) in the lower Albian Cylindroporella oolithes
at Serra Gelada, suggesting evidence of nearby piercement or fault injection.
However, more pieces of evidence are needed to better constrain the halokinetic
hypothesis.
, in the interval 1 to 6). The Hensonina
calcarenites found in the thick Puig Campana and Serreta Llarga deep-water
sections likely pinch out around a local high at Cabeçó d'Or. The origin of
this high may be related to halokinesis of Triassic salts during Albian times (see
Granier
& Perthuisot, 2009: Fig. 8). In fact, Granier (1987:
Pl. 25, figs. d-h) already reported the presence of reworked Triassic material (Triassic
quartz with salt inclusions) in the lower Albian Cylindroporella oolithes
at Serra Gelada, suggesting evidence of nearby piercement or fault injection.
However, more pieces of evidence are needed to better constrain the halokinetic
hypothesis.
This contribution is dedicated to the late Éric Fourcade (1934-2000), thesis supervisor of the first author (B.R.C.G.). Antonio Estévez Rubio assisted in identifying Fourcade's ammonite collection sites in Spain. This work has been supported by the University of the Basque Country (grant number PIF18/105) and by its PhD mobility programme. It is also a contribution to the Research Group IT-1602-22 of the Basque University System, funded by the Basque Government. Fieldwork in SE Spain (April 2024) was made possible thanks to a grant from the Association 'Carnets de Géologie'. Ilene Rex and Eric Monteil kindly hosted the first author during his last stay in Altea. In addition, Eric assisted in collecting new ammonites near Aigües. The UCBL-FSL-numbered ammonites are part of the 'Collections de l'Université Claude Bernard Lyon 1' (Granier Collection), Villeurbanne, France. The studied thin sections will be deposited in the collections of the 'Muséum d'Histoire Naturelle de Genève', Switzerland. The authors gratefully acknowledge the constructive comments and suggestions provided by a group of experts, including Ioan I. Bucur (Cluj-Napoca), Jens Lehmann (Bremen), Josep Anton Moreno-Bedmar (Mexico), and Felix Schlagintweit (Munich). It must be admitted that the manuscript reflects the ideas of its authors, not necessarily those of the referees. Finally, the first author extends his gratitude to Phil Salvador for his assistance in refining the English text of the original manuscript.
Azéma J. (1977).- Étude géologique des zones externes des Cordillères bétiques aux confins des provinces d'Alicante et de Murcie (Espagne).- Thèse de Doctorat d'État, Université Pierre et Marie Curie, Paris, 393 p.
Berthou P.Y. (1971).- Le Cénomanien de l'Estrémadure portugaise.- Serviços geológicos de Portugal, Memória, Lisboa, no. 23, 162 p. (65 Pls.).
Berthou P.Y. & Schroeder R. (1978).- Les Orbitolinidae et Alveolinidae de l'Albien supérieur-Cénomanien inférieur et le problème de la limite Albien/Cénomanien dans le sud-ouest de la région de Lisbonne (Portugal).- Cahiers de Micropaléontologie, Paris, fasc. 1978.3, p. 51-85 (9 Pls.).
Castro J.M., Company M., Gea G.A. de & Aguado R. (2001).- Biostratigraphy of the Aptian-Middle Cenomanian platform to basin domain in the Prebetic Zone of Alicante, SE Spain: Calibration between shallow water benthonic and pelagic scales.- Cretaceous Research, vol. 22, no. 2, p. 145-156.
Castro J.M., Gea G.A. de, Ruiz-Ortiz P.A. & Nieto L.M. (2008).- Development of carbonate platforms on an extensional (rifted) margin: The Valanginian-Albian record of the Prebetic of Alicante (SE Spain).- Cretaceous Research, vol. 29, no. 5-6, p. 848-860.
Charollais J. & Brönnimann P. (1965).- Première note sur les Foraminifères du Crétacé inférieur de la région genevoise. Sabaudia Charollais et Brönnimann, n. gen.- Archives des Sciences, Genève, vol. 18, no. 3, p. 615-624 (Pls. I-III). DOI: 10.5169/SEALS-739229
Decrouez D. & Moullade M. (1974).- Orbitolinidés nouveaux de l'Albo-Cénomanien de Grèce.- Archives des Sciences, Genève, Band 27, Heft 1, p. 75-92 (Pls. I-IV). DOI: 10.5169/seals-739298
Estévez A., García-Hernández M., Pina J.A. & Auernheimer C. (1984).- Nuevas precisiones estratigráficas sobre el Cretácico inferior del Cabeço d'Or ( Provincia de Alicante, Zona Prebética).- I Congreso Español de Geología, Segovia del 9 al 14 de abril de 1984, secc. 23, III, p. 189-197.
Fourcade É. (1967).- Données nouvelles sur le Crétacé inférieur du nord-est de la Cordillère bétique (Espagne).- Atti Accademia Nazionale dei Lincei, Rendiconti (Classe Scienze Fisiche Matematiche Naturali), serie 8, vol. 42, no. 6, p. 842-851. URL: http://www.bdim.eu/item?fmt=pdf&id=RLINA_1967_8_42_6_842_0
Fourcade É. (1970).- Le Jurassique et le Crétacé aux confins des Chaînes bétiques et ibériques (Sud-Est de l'Espagne).- Thèse Doctorat ès Sciences, Université Pierre et Marie Curie, Paris, 415 p. (tomes I-III, 37 Pls.).
Fourcade É. (1978).- Simplorbitolina chauvei n. sp., Orbitolinidae nouveau de l'Albo-Aptien d'Espagne.- Annales Scientifiques de l'Université de Besançon (Géologie, 3ème série), fasc. 29, p. 63-71.
García-Hernández M. (1978).- El Jurasico terminal y el Cretacico inferior en las sierras de Cazorla y del Segura (Zona prebetica).- Tesis doctorales de la Universidad de Granada, 190, 344 p. (XXXV Pls.).
Ghigi A. (1909).- Raccolte planctoniche fatte dalla R. nave 'Liguria' nel viaggio di circonnavigacione del 1903-05 sotti il comando di S.A.R. Luigi di Savoia Duca degli Abruzzi. Vol. II, fasc. I. Ctenofori.- Pubblicazioni del R. Istituto di Studi Superiori Practici e di Perfezionamento in Firenze, Sezione di Scienze Fisiche e Naturali, p. 1-24.
Giannetti A., Monaco P., Corbí H. & Soria J.M. (2014).- Integrated taphonomy in an open-marine platform: The Lower Cretaceous of Sierra Helada (Betic Cordillera, SE Spain).- Cretaceous Research, vol. 51, p. 274-284.
Giraud F., Kassab W.H., Robert E., Jaillard É., Spangenberg J.E., Masrour M., Hammed M.S., Aly M.F. & El Hariri K. (2021).- Integrated stratigraphy of the latest Barremian-early Albian interval in the western part of the Tethyan marin: New data from the Essaouira-Agadir Basin (Western Morocco).- Newsletters on Stratigraphy, vol. 54, no. 1, p. 43-78.
Granier B. (1987).- Le Crétacé inférieur de la Costa Blanca entre Busot et Altea, Alicante (Espagne) : Biostratigraphie, sédimentologie, évolution tectono-sédimentaire.- Thèse, Docteur de l'Université Paris VI (nouveau régime), 23 Novembre 1987; Mémoires des Sciences de la Terre, Université Pierre et Marie Curie, Paris, no. 87-49, vol. I (text): 281 p.; vol. II (plates): 54 pls.
Granier B. (1988).- Algues Chlorophyceae du Jurassique terminal et du Crétacé inférieur en Alicante.- Mediterranea, Alicante, no. 5 (1986), p. 5-96.
Granier B. (2019).- Discussion on some previous records of Involutina hungarica (Sidó, 1952). Revision of the Jesse Harlan Johnson Collection. Part 6.- Carnets Geol., Madrid, vol. 19, no. 20, p. 445-455.
Granier B.R.C. (coordinator), Andreu Rodes J.M., Benzaggagh M., Blau J., Böker É., Estévez Rubio A., Ferré B., Lambert B.C., Marini A., Monteil E., Reboulet S. & Virgone A. (2024).- Rectifying a substantial biostratigraphic error: There are no Hauterivian calpionellids in Busot (Alicante, Spain).- Carnets Geol., Madrid, vol. 25, no. 2, p. 29-72. DOI: 10.2110/carnets.2025.2502
Granier B. & Perthuisot V. (2009).- Discussion of: Development of carbonate platforms on an extensional (rifted) margin: the Valanginian-Albian record of the Prebetic of Alicante (SE Spain), by J.M. Castro et al., Cretaceous Research 29 (2008), 848-860.- Cretaceous Research, vol. 30, no. 4, p. 1019-1026.
Hofker J. Jr (1965).- Some foraminifera from the Aptian-Albian passage of northern Spain.- Leidse Geologische Mededelingen, vol. 33, no. 1, p. 183-189.
Jaffrezo M. (1980).- Les formations carbonatés des Corbières (France) du Dogger à l'Aptien: Micropaléontologie, stratigraphie, biozonation, paléoécologie. Extension des résultats à la Mésogée.- Thèse de Doctorat d'État, Université Pierre et Marie Curie, Paris, 615 p.
Jaillard É., Robert E., Latil J.-L. & Masrour M. (2024).- Albian sedimentation in western Atlas, Morocco.- Cretaceous Research, vol. 159, article 105872, 20 p.
Johnson J.H. (1954).- Cretaceous Dasycladacea from Gillespie county, Texas.- Journal of Paleontology, Cambridge (UK), vol. 25, p. 787-790.
Keijzer F. (1942).- On a new genus of arenaceous foraminifera from the Cretaceous of Texas.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, vol. 45, deel 10, p. 1016-1017. URL: https://dwc.knaw.nl/DL/publications/PU00017848.pdf
Latil J.-L. & Jaillard É. (2024).- Taxonomy, phylogeny and biostratigraphy of the upper Aptian-lower Albian ammonites of the Chott area, southern Central Tunisia.- Cretaceous Research, vol. 154, article 105731, 34 p.
Lemoine M. (1966).- Un nouveau genre d'algue du Crétacé inférieur : Agardhiellopsis nov. gen.- Revue de Micropaléontologie, Paris, vol. 8, no. 4, p. 203-210.
Masse J.-P. (1976).- Les calcaires urgoniens de Provence. Valanginien - Aptien inférieur. Stratigraphie, paléontologie, les paléoenvironnements et leur évolution.- Thèse de Doctorat ès Sciences, Université Aix-Marseille II, 445 + 43 p. (60 Pls.).
Masse J.-P., Arias C. & Vilas L. (1992).- 4.2. Stratigraphy and biozonation of a reference Aptian Albian p.p. Tethyan carbonate platform succession: The Sierra del Carche series (oriental Prebetic zone Murcia, Spain). In: New aspects on Tethyan Cretaceous fossil assemblages.- Schriftenreihe der Erdwissenschaftlichen Kommissionen der Österreichischen Akademie der Wissenschaften, Wien, Band 9, p. 201-221.
Moullade M. & Peybernès B. (1979).- Révision d' "Orbitolinopsis" aquitanica Schroeder 1964 : Orbitolinidé du Gargasien (= Aptien supérieur) pyrénéen.- Archives des Sciences, Genève, vol. 32, no. 1-3, p. 261-266 (2 Pls.)
Nicklès R. (1892).- Recherches géologiques sur les terrains secondaires et tertiaires de la province d'Alicante et du Sud de la province de Valence.- Thèse de Doctorat ès Sciences naturelles de la Faculté des Sciences de Paris (1891); Annales Hébert, Paris, tome I, 219 p. (X Pls.)
Özdikmen H. (2009).- Substitute names for some unicellular animal taxa (Protozoa).- Munis Entomology & Zoology, Ankara, vol. 4, no. 1, p. 233-256.
Peybernès B. (1976).- Le Jurassique et le Crétacé inférieur des Pyrénées franco-espagnoles, entre la Garonne et la Méditerranée.- Thèse de Doctorat ès-Sciences Naturelles, Université Paul-Sabatier, Toulouse III, 459 p. (XLII Pls.).
Rigaud S., Consorti L., Granier B. & Schlagintweit F. (2023).- Foraminifera: Did the first canal systems develop in Involutinida?- Historical Biology, vol. 35, 20 p.
Robert E., Latil J.-L. & Bulot L.G. (2009).- Albian ammonite faunas from South America: The genus Tegoceras Hyatt, 1903.- Revue de Paléobiologie, Genève, vol. 28, no. 1, p. 43-51.
Schlagintweit F. (2023).- Taxonomic revision of Paracoskinolina fleuryi Decrouez & Moullade, 1974, 'primitive' orbitolinid from the early Cenomanian of Greece and its stratigraphic and palaeogeographic records.- Revue de Micropaléontologie, Paris, vol. 78, article 100712, 6 p.
Schlagintweit F., Rosales I. & Najarro M. (2021).- The upper Aptian - middle Albian Simplorbitolina lineage: An example of nepionic acceleration in Lower Cretaceous Orbitolinidae (Foraminifera).- Historical Biology, vol. 34, no. 4, 11 p.
Schlagintweit F. & Wilmsen M. (2014).- Orbitolinid biostratigraphy of the top Taft Formation (Lower Cretaceous of the Yazd Block, Central Iran).- Cretaceous Research, vol. 49, p. 125-133.
Schroeder R. (1963).- Grundlagen einer Orbitoliniden-Biostratigraphie des tieferen Urgons im pyrenäisch-kantabrischen Grenzgebiet (Nordspanien).- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, Jahrgang 1963, Heft 6, p. 320-326.
Schroeder R. & Neumann M. (coordinators, 1985).- Les grands Foraminifères du Crétacé moyen de la région méditerranéenne.- Géobios, Villeurbanne, Mémoire spécial vol. 7, 161 p.
Schroeder R. & Poignant A. (1964).- Orbitolinopsis aquitanica, eine neue Foraminifere der Unterkreide der Aquitaine (SW-Frankreich). (Studien über primitive Orbitolinidae I).- Eclogae Geologicae Helvetiae, Basel, vol. 57, no. 2, p. 553-557 (Pls. I-II). DOI: 10.5169/seals-163151
Scott R.W., Campbell W., Diehl B., Edwards W., Gargili Altintas D., Harlton K., Hojnacki R., Lai X., Porter A., Rush N.,Tan X. & Wang Y. (2019).- Regional upper Albian-lower Cenomanian sequence boundaries, Texas Comanche shelf: Is timing related to oceanic tectonics?- Cretaceous Research, vol. 98, p. 335-362.
Szives O., Latil J.-L., Moreno-Bedmar J.A., Lehmann J., Robert E. & Owen H.G. (2023).- Critical revision and new proposals on the Aptian-Albian zonation of the Standard Mediterranean Ammonite Zonal Scheme.- Newsletters on Stratigraphy, vol. 56, no. 4, p. 423-456.
Szives O., Moreno-Bedmar J.A., Aguirre-Urreta B., Company M., Frau C., López-Horgue M., Pictet A., Ploch I., Salazar C., Barragán R., Latil J.-L., Lehmann J., Robert E. & Reboulet S. (2024).- Report of the 7th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Warsaw, Poland, 21st August 2022): State of the art of the current Standard Ammonite Zonation of the Western Tethyan Mediterranean Province.- Cretaceous Research, vol. 153, article 105716, 14 p.
Plate
1:
Samples
A-E, J-L, N-P, X-Y from Serra Gelada, I, Z
from Puig Campana, and F-H, M, Q-W from Cabeçó d'Or. Favusella washitensis (Carsey, 1926),
A-B: Relais 2 (HL 705), Cylindroporella oolithes, Relais section,
25/04/1985, C-E, K-L, N-O, X-Y: HL 60*, Cylindroporella oolithes,
Punta de les Caletes (13/04/1984,
circa 38°31'31.3"N,
0°05'46.3"W); F, W: BT 61,
calcarenites with Favusella, Sanatorio section, 26/04/1985; G-H, M,
Q-R, U-V: BT 72, calcarenites with Favusella, Sanatorio section,
28/04/1985; I: Rafel 9 (ZC15931), Rafel section, 01/08/1982; J:
Relais 1 (HL 704), Cylindroporella oolithes, Relais section, 25/04/1985; P:
AB 21, Cylindroporella oolithes, Cueva de la ballena, 10/06/1984; S-T:
BT 59, calcarenites with Favusella, Sanatorio section, 26/04/1985; Z:
Realet 2 (ZC19384), Pieninia calcarenites, Realet d'Alt section,
19/04/1983. Scale bar (on fig. G) = 250 µm. |
|
Plate
2:
Samples
A, G-J, O-W from Puig Campana, B, N from Cabeçó d'Or, C-F,
K-L from Serra Gelada, and M from Illa de
Benidorm. Agardhiellopsis cretacea Lemoine, 1966, A:
Realet 4 (ZC19382), Pieninia calcarenites, Realet d'Alt section,
19/04/1983; B: BT 8, calcarenites with Favusella, north of
Sanatorio, 16/04/1985; C-D: HL 48, rudistid limestones, Calle Dos Callas,
'LM' section, 08/04/1984. Cylindroporella barnesii Johnson,
1954, E: Realet 8 (ZC19377), Simplorbitolina limestones,
Realet d'Alt section, 19/04/1983; F, K: Relais 15 (HL 724bis), L:
Relais 13 (HL 725), Cylindroporella oolithes, Relais section, 01/05/1985;
G-I: Realet 22 (ZC19359), J: Realet 19 (ZC19362), Simplorbitolina
limestones, Realet d'Alt section, 09/04/1983. Coskinolinoides
texanus Keijzer,
1942, M:
ISL 2, rudistid limestones, 1984, N: BT 331, north of Sanatorio,
calcarenites with Favusella, 09/05/1985. Simplorbitolina sp. (O-R,
T-U: S. manasi, S, V-W: S. conulus), O-R: Realet 10
(ZC19375), O = Granier, 1987: Pl. 9, fig. d, S: Realet 11
(ZC19374), T-U: Realet 12 (ZC19373), V-W: Realet 14 (ZC19367), Simplorbitolina
limestones, Realet d'Alt section, 09/04/1983. Scale bar = 250 µm. |
|
Plate
3:
A-C: Micritic envelopes with sedimentary infills providing evidence of
subaerial exposure and leaching of the aragonite shells. A: BT DJ1, ca. 38°31'50.4"N, 0°24'20.7"W, coral
biolithites,
Cabeçó d'Or, 14/05/1985 (Granier, 1987: Pl. 46, fig. g); B: BT 331,
calcarenites with Favusella, first crest, Sanatorio section, Cabeçó
d'Or, 09/05/1985 (Granier, 1987: Pl. 46, fig. h); C: BT 456, coral
biolithites, second crest, Sanatorio section, Cabeçó d'Or, 14/05/1985 (Granier,
1987: Pl. 46, fig. i). D-F: Epistomina
spp. D: BT 73, calcarenites
with Favusella, Sanatorio section, Cabeçó d'Or, 28/04/1985; E: Realet 3
(ZC15383), Pieninia calcarenites, Realet d'Alt section, Puig Campana,
19/04/1983; F: Relais 1 (HL 704), Cylindroporella oolithes, Relais
section, Serra Gelada, 25/04/1985. All scale bars = 250 µm (D-F have the same
magnification). |
|
Plate
4:
Samples
A-C from Serra Gelada, D, F-J from Puig Campana, and E from Illa de
Benidorm. Pseudochoffatella cuvillieri Deloffre, 1961,
A: HL 44, B: HL 48, rudistid limestones, Calle Dos Callas, 'LM'
section, 08/04/1984, C: Relais 12 (HL 722), Cylindroporella
oolithes, Relais section, 01/05/1985, E: ISL 4, rudistid limestones,
1984, D: Realet 14 (ZC19367), F: Realet 23 (ZC19357), G: Realet 22 (ZC19359),
Simplorbitolina limestones, Realet d'Alt section,
09/04/1983. Praehedbergella spp., H-J: Cluse 20 (7832), condensed section, Cluse section, 19/04/1983. A,
D-G, same scale bar (on figs. A and G) = 250 µm; B-C, same scale bar (on fig. B) = 500 µm;
H-J, same
scale bar (on fig. J) = 100 µm. |
|
Plate
5:
Samples
A-T from Serra Gelada, U-W from Puig Campana, and X-AE from Cabeçó d'Or. Serra
Gelada:
Mesorbitolina texana (Roemer,
1849), A: AB 5, B: AB 8, C: AB 10A, D: AB 12B, Cylindroporella
oolithes, Cueva de la ballena, 10/06/1984; L: Relais 13 (HL 726), Cylindroporella
oolithes, Relais section, 01/05/1985; R-S:
Relais 7 (HL 709), Cylindroporella oolithes, Relais section, 25/04/1985, Z:
BT 0A-2, AA-AB: BT 0A-1, AC-AE: BT 0A-3, AF: BT 0A-4, AG:
BT 0A-5, AH-AK: BT 0A-7, AL-AO: BT 0A-8, calcarenites with Favusella,
north of Sanatorio, 13/04/1985. Mesorbitolina
birmanica Sahni,
1937, E:
SH-Bb1060, F: SH-Bb1057, G: SH-Bb1055, H: SH-Bb1053, I:
SH-Bb1052, J: SH-Bb1045, Cylindroporella oolithes, Camí del Far,
24/09/2010; K: Relais 13 (HL 726), Cylindroporella
oolithes, Relais section, 01/05/1985; M: HL 141, orbitolinid
calcarenites, Calle Dos Callas, 'LM' section, 12/06/1984; N-Q: HL 78,
rudistid limestones, Punta de les Caletes section, 17/04/1984, U: Cluse 30 (MC19420), coral
biolithites, Cluse section,
19/04/1983, Y:
BT 59, calcarenites with Favusella, Sanatorio section, 26/04/1985. Orbitolinopsis
sp., T: HL 225, rudistid limestones, '3P', 24/07/1984. Mesorbitolina
sp., V: Realet 4 (ZC19382), W: Realet 2
(ZC19384), Pieninia calcarenites, Realet d'Alt section, 19/04/1983, X:
BT 456, coral biolithites, Sanatorio section, 14/05/1985. Scale bar (on fig. I)
= 250 µm. |
|
Plate
6:
All
samples from Serra Gelada. Mesorbitolina birmanica Sahni, 1937,
A-D: HL 225-1, E-F: HL 225-2, rudistid limestones,
'3P', 24/07/1984; G: HL 251, Cylindroporella oolithes, above Camí
del Far, 26/07/1984; H: HL 110, rudistid limestones, Carabiniers 10, Camí
del Far, Carabiniers section, 07/04/1984; I-K: HL 39, rudistid
limestones, Punta de les Caletes section, 07/04/1984; L: HL 42bis, M:
HL 44, rudistid limestones, Calle Dos Callas, 'LM' section, 08/04/1984; N-O:
Relais 24 (HL 740), P: Relais 23 (HL 742), rudistid limestones, Relais
section, 01/05/1985; Q-Z: HL 78A, AA-AM: HL 78B, rudistid
limestones, Punta de les Caletes section, 17/04/1984. Scale bar (on fig. H) =
250 µm. |
|
Plate
7:
Sample
from Serra Gelada. Mesorbitolina birmanica Sahni, 1937, A-S:
HL 225-3, T-AC: HL 225-2, AD-AO: HL 225-1, rudistid limestones,
'3P', 24/07/1984. Scale bar (on fig. K) = 250 µm. |
|
Plate
8:
All
samples from Puig Campana, Simplorbitolina limestones, Realet d'Alt section, 09/04/1983. Simplorbitolina chauvei Fourcade,
1978,
A: Realet 8 (ZC19377), B: Realet 9 (ZC19376), N: Realet 10 (ZC19375). Simplorbitolina conulus
Schroeder, 1965, C-E, H-L, Q-T, AC: Realet 23 ( ZC19357), J = Granier,
1987: Pl. 9, fig. e, F-G: Realet 12 (ZC19373), F = Granier,
1987:
Pl. 9, fig. c, M: Realet 11 (ZC19374). Simplorbitolina manasi Ciry
et Rat, 1953,
O: Realet 12 (ZC19373),? AB: Realet 11 (ZC19374). 'Transitional
form', intermediate form between Simplorbitolina manasi Ciry et Rat,
1953,
and Simplorbitolina conulus Schroeder, 1965, P: Realet 14 (ZC19367). Simplorbitolina
aquitanica (Schroeder
& Poignant, 1964), U-V: Realet 9 (ZC19376), W-Z: Realet 10 (ZC19375), AA:
Realet 23 (ZC19357). Scale bar (on fig. K) = 250 µm. |
|
Plate
9:
All
samples from Puig Campana. Neorbitolinopsis conulus (Douvillé,
1912), A, F, U-V: equivalent Rafel 41 (MC19499), 04/08/1982; B-E,
W: Rafel 41 (BG82813), 04/08/1982 (Granier, 1987: Pl. 10, fig. d); X:
equivalent Rafel 41 (MC19493), 04/08/1982, Hensonina calcarenites, Rafel section. Praehedbergella
spp., G-T: Cluse 20 (7832), condensed section, Cluse section, 19/04/1983.
A-F, U-X, same scale bar
(on fig. F) = 250 µm; G-T, same scale bar (on fig. K) = 100 µm. |
|
Plate
10:
Ammonite
specimens from
Sanatorio Section, Aïgues de Busot, Cabeçó d'Or. A-I first illustrated in Granier
(1987). J-W newly collected in 2024. A-B:
Brancoceras sp. (Granier, 1987: Pl. 47, fig. 4.a-b), C-D: Hysteroceras
sp. (Granier, 1987: Pl. 47,
fig. 6.a-b), E-F: Hysteroceras
sp. (Granier, 1987: Pl. 47,
fig. 1.a-b), G: Dipoloceras sp. (Granier,
1987: Pl. 47, fig.
5.a), H-I: Dipoloceras sp. (Granier, 1987: Pl. 47,
fig. 2.b-c), J-K: Puzosia
(Pusozia) sp. (UCBL-FSL 471979). This specimen displays a constricted
cross-section (more compressed than in Desmoceras, excluding its ascription to
the latter genus) and weakly convex flanks. The involution of the whorls is
difficult to ascertain due to the concealment of the internal whorls. L-M:
Dipoloceras sp. (UCBL-FSL 471989), N-O:
Dipoloceras sp. (UCBL-FSL 471991), P-R: Hysteroceras sp. (UCBL-FSL
471990), S: Hamites
sp. (UCBL-FSL 471983), T-U: Hamites
sp. (UCBL-FSL 471982), V: Hamites
sp. (UCBL-FSL 471984), W:
Anisoceras sp. (UCBL-FSL 471985). All
scale bars = 1 cm. |
|
Plate
11:
Ammonite
specimens from Caroch
Massif, East of Ayora. A-B: Parengonoceras
sp. (UCBL-FSL 471991), C: Parengonoceras
sp. (UCBL-FSL 471997), D-E: Parengonoceras
cf. ibericum (Arias & Wiedmann, 1977), (UCBL-FSL
471994). All scale bars = 1 cm. |
|