◄ Carnets Geol. 25 (6) ►
![]()
Outline:
[1. Introduction]
[2. Geological background and locality]
[3. Material and methods]
[4. Results]
[5. Discussion]
and ...
[Bibliographic references]
Institute of Ecology and Earth Sciences, University of Tartu, Ravila 14A, 50411, Tartu (Estonia)
Institut für Geologie, Universität Hamburg, Bundesstr. 55, 20146 Hamburg (Germany)
The College of Wooster, Department of Earth Sciences, Wooster, OH 44691 (USA)
Institute of Geology, Tallinn University of Technology, Ehitajate tee 5, Tallinn (Estonia)
Published online in final form (pdf) on April 1, 2025
DOI 10.2110/carnets.2025.2506
![]()
[Editor:
Beatriz Bádenas; technical editor: Bruno R.C. Granier]
![]()
Specimens of the rugose coral Streptelasma? sp. are found intergrown with a host bryozoan Oanduellina bella in the lower Katian of Estonia. Rugosa also occur in the bryozoan Proavella proava. This is the earliest and only known record of endobiotic rugose symbionts in cryptostome bryozoans from the Late Ordovician of Baltica. The erect bryozoan colonies provided symbiotic Rugosa with both a higher and more advantageous tier for feeding in the water column and a hard substrate that these encrusting Rugosa required. The rugose corals may have protected the bryozoans with their stinging cells against predators. The lack of malformations in the bryozoan zooid architecture and their normal dimensions around the Rugosa show that the relationship between the Rugosa and bryozoans could have been mutualistic, which is similar to many other Rugosa-bryozoan associations in the Late Ordovician of Baltica, although the exact nature of this association remains uncertain.
• symbiosis;
• bryozoans;
• Rugosa;
• intergrowth;
• lower Paleozoic;
• Baltica
Vinn O., Ernst A., Wilson M.A. & Toom U. (2025).- Symbiosis between bryozoans with erect cribrate colonies and rugose corals from the lower Katian (Upper Ordovician) of Estonia.- Carnets Geol., Madrid, vol. 25, no. 6, p. 147-154. DOI: 10.2110/carnets.2025.2506
Symbiose entre des bryozoaires ŕ colonies cribriformes érigées et des tétracoralliaires (Rugosa) du Katien inférieur (Ordovicien supérieur) d'Estonie.- Des spécimens du tétracoralliaire (Rugosa) Streptelasma? sp. ont été trouvés imbriqués dans un substrat constitué par le bryozoaire Oanduellina bella du Katien inférieur d'Estonie. Les tétracoralliaires sont également observés avec pour hôte le bryozoaire Proavella proava. Il s'agit de la plus ancienne et unique identification de symbiontes tétracoralliaires endobiontes dans des bryozoaires (ŕ opercules) cryptostomes du Dévonien supérieur de Baltica. Les colonies érigées de bryozoaires fournissent aux tétracoralliaires symbiontes une surélévation avantageuse pour s'alimenter dans la colonne d'eau, ainsi qu'un substrat dur nécessaire ŕ ces tétracoralliaires encroűtants. Les tétracoralliaires ont potentiellement protégé les bryozoaires des prédateurs grâce ŕ leurs cellules urticantes. Bien que la nature exacte de cette association entre tétracoralliaires et bryozoaires demeure incertaine, l'absence de malformations dans l'architecture des zoďdes des bryozoaires et leurs dimensions normales autour des tétracoralliaires suggčrent que la relation entre les deux pourrait avoir été mutuellement bénéfique, comme c'est également le cas pour de nombreuses associations similaires dans le Dévonien supérieur de Baltica.
• symbiose ;
• bryozoaires ;
• tétracoralliaires (Rugosa) ;
•
intercroissance ;
• Paléozoďque inférieur ;
• Baltica
Symbiotic interactions between different organisms are rarely preserved in the fossil record. Endobionts embedded (bioimmured) within the living tissues of host organisms hold significant importance (see Taylor, 1990, for review). The earliest microscopic invertebrate symbionts appeared in the Cambrian (Bassett et al., 2004), while macroscopic endobiotic invertebrate symbionts emerged later, becoming common during the Late Ordovician (see Tapanila, 2005, for a summary). In the Silurian and Devonian, rugose corals were frequently bioimmured within bryozoans, stromatoporoid skeletons, crinoid stems, and occasionally in other corals (Berkowski & Klug, 2011; Suárez Andrés et al., 2020; Vinn et al., 2023). These differ from bioclaustrations (Palmer & Wilson, 1988; Słowiński et al., 2020) by having their own skeletons. While several Paleozoic bioclaustrations may have been caused by parasites, it is more accurate to classify them simply as symbionts (Zapalski, 2007, 2011; Zapalski & Hubert, 2011; Taylor, 2015).
The evolution of parasitic relationships has been the focus of recent studies by various researchers (De Baets & Littlewood, 2015; Huntley & De Baets, 2015; De Baets et al., 2011, 2015, 2021a, 2021b; Huntley et al., 2021; Dijk & De Baets, 2021). Increasing evidence suggests that biodiversity influences parasite prevalence and that parasitism has grown throughout the Phanerozoic. Additionally, there is support for the idea that parasitism has intensified alongside increases in biodiversity over the course of life's history (De Baets et al., 2021a, 2021b).
Symbiosis is defined here as any close and long-term biological interaction between interspecific organisms, whether mutualistic, commensalistic, or parasitic. Modern ecologists and biologists follow the "Bary" definition of symbiosis as "the living together of unlike organisms" (Bary, 1879) or an even broader concept where all interspecific biological interactions are considered symbiosis. The more restrictive definition, where symbiosis refers only to mutualism, is no longer commonly used (Martin & Schwab, 2013; Robin, 2021).
In the Paleozoic, bryozoans were primarily colonized by organisms from the Paleozoic fauna (Sepkoski, 1981, 1984), including rugose corals, tabulate corals, conulariids, brachiopods, and cornulitids (Vinn et al., 2023). The earliest bryozoan hosts were Tremadocian trepostomes from South China (Ma et al., 2020), followed by esthonioporates, which were colonized during the Sandbian, and cystoporates, first colonized in the Katian (Vinn et al., 2023). However, there is no previous record of symbiotic endobionts in cryptostome bryozoans from the Ordovician of Baltica. Nevertheless, anomalous structures most probably caused by symbiont embedment from the Ordovician of Sweden were figured by Nils Spjeldnaes (1984). In corals, on the other hand, macrosymbiosis was common in the Ordovician worldwide (Darrell & Taylor, 1993). The Ordovician corals housed cornulitids, worms, and lingulate brachiopods.
The aims of this paper are to: 1) describe the earliest known endobiotic rugose coral symbionts from cryptostome bryozoans of the Late Ordovician of Baltica; and 2) discuss the paleoecology of this Rugosa-bryozoan association.
During the Ordovician period, Baltica transitioned from a temperate climatic zone to a subtropical region (Nestor & Einasto, 1997; Cocks & Torsvik, 2005; Torsvik et al., 2013). By the Sandbian, present-day Estonia was covered by a shallow epicontinental sea with minimal bathymetric variation and very low sedimentation rates (Nestor & Einasto, 1997). Across the ramp, grey argillaceous and calcareous sediments were deposited, with increasing clay content and fewer bioclasts in offshore areas (Nestor & Einasto, 1997).
In the Katian, climatic shifts led to a rise in carbonate production and sedimentation on the carbonate shelf (Nestor & Einasto, 1997). The Dapingian to Hirnantian succession in Estonia is defined by diverse normal marine carbonate rocks (Nestor & Einasto, 1997). In northern Estonia, limestones predominate, formed in the shallow parts of the basin, with marls also present in lesser amounts. The purest limestones are mostly found in the Katian stage of northern Estonia. Carbonate buildups became widespread during the early Katian, beginning with the Keila Regional Stage (Nestor & Einasto, 1997).
The Vasalemma Formation is exclusively found and exposed in an area southwest of Tallinn, near the settlements of Vasalemma and Saku. In this region, the pelmatozoan grainstone facies of the Vasalemma Formation was deposited along a narrow belt, approximately 30 km long (WSW-ENE) and 5 km wide (N-S) (Kröger et al., 2017). Within the massive Vasalemma Formation, which reaches up to 15 meters in thickness, patch reefs developed locally, with diameters of up to 50 meters and similar thicknesses (see Kröger et al., 2014, for review). These reefs are primarily composed of bryozoan and echinoderm framestone to bindstone, with Hemiscosmites holdfasts and encrusting bryozoans as the main metazoan constructors. In some areas, receptaculitids served as important additional bafflers (Kröger et al., 2014). At the top of these reefs, large colonies of Eofletcheria orvikui are commonly found. Recent palaeogeographical reconstructions suggest that these reefs formed in the central areas of the shallow Estonian shelf, which was highly differentiated into local shallow basins and shoals, and likely extended further northwest into areas where Upper Ordovician deposits have since been eroded (Kröger et al., 2014).
The reefs of the Vasalemma Formation in northern Estonia, dating to the late Sandbian of the Late Ordovician, host an exceptionally rich and abundant bryozoan fauna (Kröger et al., 2023). These reefs are part of a global pattern of contemporaneous bryozoan-rich reefs, representing the peak of bryozoan diversification during the Ordovician (Kröger et al., 2023). The organism-mediated nature of these reefs and their substrates highlights the significant role that organisms like bryozoans may have played in ecosystem engineering, contributing to the broader Ordovician diversification (Kröger et al., 2023).
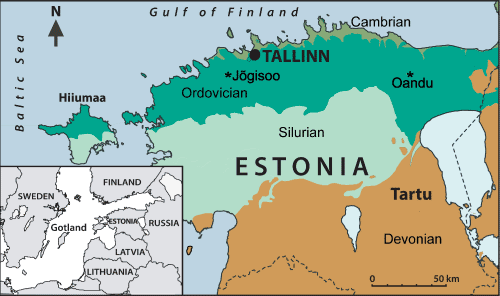
At the Jőgisoo outcrop (GPS coordinates: 59°16'32.8"N,
24°30'27.4"E) limestones of the Keila Regional Stage, Vasalemma
Formation are exposed. At the Oandu outcrop (GPS coordinates: 59°15'44.8"N
27°00'12.4"E),
approximately 0.3 m of highly argillaceous limestone (wackestone-packstone),
abundant in various fossils (such as brachiopods, bryozoans, and trilobites),
is exposed (Männik, 2017). The rocks belong to the Oandu Regional Stage, Hirmuse
Formation (lowermost Katian). The Hirmuse Formation is characterized by marls
and argillaceous limestone. It has a rich, normal, marine fauna including
brachiopods, bryozoans, echinoderms, trilobites, and rugose corals (Hints & Meidla,
1997)
(Fig. 1 ![]() ).
).
|
Figure 1:
Locality map: Jőgisoo and
Oandu indicated with asterisk. |
The collections of Late Ordovician bryozoans deposited at the Tallinn University of Technology, Institute of Geology and Natural History Museum, University of Tartu were searched for intergrown specimens, especially focusing on potential symbiotic Rugosa. Bryozoans with erect cribrate colonies were common in the collection. The bryozoan specimens with symbiotic Rugosa were cleaned and photographed uncoated using a Canon EOS 5Dsr digital camera and apochromatic zoom system Leica Z16 APO. The Rugosa were measured on calibrated photos. Two bryozoan specimens with symbiotic Rugosa are described. They are deposited at the Institute of Geology at Tallinn University of Technology (GIT 537-2322 and GIT 537-5573).
The juvenile subcylindrical solitary coral specimens similar to Streptelasma Hall, 1847, are found in Oanduellina bella Lavrentjeva in Gorjunova & Lavrentjeva, 1993 (GIT 537-2322). They are of small size and have calice diameters from 2.7 mm to 3.0 mm. The septa are long, slender, and not too closely spaced. A deep fossa is developed at the main septum on the concave side of the coral similar to Streptelasma. Dissepiments are absent. Although the juvenile rugose specimens cannot be fully identified, we can tentatively assign them to the genus Streptelasma?.
Three specimens of Streptelasma?
sp. are variably intergrown with their host bryozoan O. bella (GIT 537-2322) so that calices of
the two corals are free of bryozoan encrustation while the calice of the third
coral is partially overgrown, leaving only a slit with erect lip-like edges
open (indicating overgrowth syn vivo) (Fig. 2.A-D
![]() ). The bryozoan has an
erect, cribrate colony composed of bilaminate flattened branches that are
undulate and coalesce to produce lachrymiform lacunae (fenestrules) (Taylor,
2012). The Rugosa are located between lacunae, never crossing them. All
three coral specimens have identical orientation regarding their longitudinal
axis. The edges of two other calices are free of encrustation. Two calices are
oriented almost perpendicular to the plane of the bryozoan colony, while a
single calice is oriented almost parallel to the colony plane. The bryozoan
zooids do not show any malformations or aberrant dimensions near the Rugosa.
In addition to Rugosa, a colony of Corynotrypa
sp. encrusts Proavella proava. There are no signs of overgrowth between the two
bryozoans.
). The bryozoan has an
erect, cribrate colony composed of bilaminate flattened branches that are
undulate and coalesce to produce lachrymiform lacunae (fenestrules) (Taylor,
2012). The Rugosa are located between lacunae, never crossing them. All
three coral specimens have identical orientation regarding their longitudinal
axis. The edges of two other calices are free of encrustation. Two calices are
oriented almost perpendicular to the plane of the bryozoan colony, while a
single calice is oriented almost parallel to the colony plane. The bryozoan
zooids do not show any malformations or aberrant dimensions near the Rugosa.
In addition to Rugosa, a colony of Corynotrypa
sp. encrusts Proavella proava. There are no signs of overgrowth between the two
bryozoans.
A single Proavella
proava (Eichwald, 1842) specimen
(GIT 537-5573)
contains a single partially intergrown unidentified Rugosa (Fig.
2.E
![]() ). The
calice of the Rugosa is free of bryozoan encrustation. The calice is oriented
almost parallel to the surface of the host bryozoan colony and is located
between lacunae.
). The
calice of the Rugosa is free of bryozoan encrustation. The calice is oriented
almost parallel to the surface of the host bryozoan colony and is located
between lacunae.
|
Figure 2:
A, Three specimens of Streptelasma?
sp. are variably intergrown with their host bryozoan Oanduellina
bella from Oandu Regional Stage,
northern Estonia (GIT 537-2322). B-C, Streptelasma? sp. in O. bella (GIT 537-2322).
D. Streptelasma? sp. with partially
overgrown calice (GIT 537-2322). E, A single specimen of rugose coral (Rg) is
partially intergrown with Proavella proava from Keila Regional Stage,
northern Estonia (GIT 537-5573). |
The symbiotic nature of the association is most strongly suggested by the almost complete intergrowth of the Rugosa with their bryozoan host. If there was post-mortem encrustation of the bryozoan by the Rugosa, the latter would not have been overgrown with the skeletal tissues of the bryozoan. Thus, it is clear that the Rugosa encrusted living Oanduellina bella and Proavella proava colonies. Moreover, the uniform orientation of the Rugosa in the Oanduellina bella colony indicates that the settlement took place when the living bryozoan colony was still erect on the sea floor. The encrustation by Corynotrypa sp. could have taken place post-mortem as there are no signs of intergrowth between the cryptostome bryozoan and Corynotrypa sp. However, it is equally possible that the Corynotrypa colony grew on the moribund proximal parts of a living colony.
The colonies of Proavella proava and Oanduellina bella typically arise from a robust base attached to seafloor. The larva of Rugosa settled on the living bryozoan colony, most likely during a single colonization event as indicated by the relatively similar dimensions of all Streptelasma? sp. specimens. Symbiotic cnidarians may exhibit unique characteristics, like modified or simplified structures, potentially as a response to their interactions with hosts (Lopez-Gappa & Liuzzi, 2021). Their small size appears to be a likely adaptation for growing alongside delicate, upright hosts such as cribrate bryozoans. In this scenario, the fact that one of the rugose specimens is bioimmured while the other two are not suggests the possibility of multiple colonization events. The larvae presumably settled on the bryozoan cuticle between the zooids. Later Rugosa overgrew multiple bryozoan zooids, and deactivated them in doing so. This indicates a negative effect and, by some measures, it would suggest that the coral was a parasite. At the same time the bryozoan colony started to overgrow the walls of the Rugosa. The bryozoan encrustation in most cases did not reach over the edges of the rugose calices (N=4), but in a single case the bryozoan competitively overgrew the edges of the rugose calice, leading to the formation of a "lips"-like structure. Overgrowth of the bryozoan host beyond the edges of the calix may indicate that the rugose coral was already dead when the bryozoans finally overgrew the calyx. The overgrowth of the bryozoan host beyond the edges of the calyx may suggest that the rugose coral had already died when the bryozoans eventually overgrew the calyx. Instances of host overgrowth of symbiont bioclaustrations corresponding to dead symbionts have been reported previously. This hypothesis aligns with the possibility that the symbiotic corals settled on the bryozoan host during separate colonization events. The tilt of the calice surfaces from the colony surface varied from subparallel to subperpendicular, indicating that this likely did not influence the efficiency of the feeding in the Rugosa. If it did, one would expect to observe uniform tilt in all calices. The location of Rugosa between the lacunae and not crossing can be interpreted as the best way to have the strongest attachment to the substrate. The orientation of the Rugosa was presumably vertical on the bryozoan colony. Such orientation provided the best opportunity for the Rugosa to feed from the water column. The frond surfaces of cribrate colonies formed regions of inhalant flow and filtered water passed towards the lacunae for expulsion. Exhalent currents did not flow through the lacunae because of the autozooids on both sides of the colony generating converging inhalant currents. Instead, exhalent currents were channeled upwards away from the holes to form diverging exhalent chimneys on opposite sides of the colony (Taylor, 2012). The rugose calices are located on the fronds and are slightly tilted towards the inhalant currents, so that the Rugosa could somewhat benefit from the feeding currents of the bryozoan. The erect bryozoan colonies provided the symbiotic Rugosa with a higher and advantageous tier for feeding in the water column. They also provided a hard substrate that these encrusting Rugosa required (Vinn et al., 2016). The Rugosa, on the other hand, could have protected the bryozoans with their stinging cells against predators (Vinn et al., 2016). The lack of malformations in zooid architecture and their normal dimensions suggest that the infesting Rugosa did not have strong adverse effects on their host. The only harm they caused to their host was deactivating several zooids during the overgrowth of the bryozoan colony. Thus, one scenario is that the relationship between the Rugosa and bryozoans was mutualistic similar to many other Rugosa-bryozoan associations in the Upper Ordovician of Baltica (Vinn et al., 2016, 2017, 2018) and Laurentia (Elias, 1982; McAuley & Elias, 1990) and coral bryozoan symbioses elsewhere, such as the Devonian of North Gondwana (Plusquellec & Bigey, 2011) and the Neogene of Europe (Cadée & McKinney, 1994). Cnidarian-bryozoan intergrowths in living faunas, such as the symbiotic Zancleidae family with certain bryozoan hosts, are typically mutualistic (Lopez-Gappa & Liuzzi, 2021). Only occasionally has predatory behavior by hydroids toward their hosts been documented (Lopez-Gappa & Liuzzi, 2021). The association described here may have been mutualistic, but commensalism is also a plausible explanation. In fact, the most likely outcome of the association was somewhere between mutualism and commensalism: the Rugosa may have helped defend their bryozoan hosts against small predators. However, unlike other Palaeozoic cnidarian-bryozoan associations, the Rugosa were too small relative to the much larger surface area of the cribrate bryozoan colonies to effectively deter predation. Alternatively, living in symbiosis with a host bryozoan could have been suboptimal for the corals, not allowing them to grow to adulthood and possibly to reproduce. If so, then the corals would have derived no benefit from living with a bryozoan and the symbiosis would not have been mutualistic. Moreover, the partial overgrowth of a single rugose calice, may have occurred during the life of the coral, and could indicate a response of the bryozoan to a living symbiont that gave it no benefits, in which case the coral could even be interpreted as a parasite of the bryozoan. The associations between Streptelasma? sp.- Oanduellina bella and Rugosa - Proavella proava were facultative as all species occur solitarily elsewhere in the lower Katian of Estonia (Kaljo, 1961). The Rugosa are rather small for the most common specimens of Streptelasma? sp. and this may result from an adaption to the life in the fragile Oanduellina bella colonies. Nevertheless, such a small size was presumably a phenotypic variation rather than a genetic trait.
All previous records of Rugosa-bryozoan symbiosis known from the Ordovician of Baltica contain only bryozoans with massive, mostly hemispherical colonies. The discovery of symbiotic Rugosa in erect cribrate colonies suggests that the ecology of Rugosa-bryozoan symbioses in the Ordovician of Baltica was much more diverse than we previously knew. The paleoenvironments represented by the Hirmuse and Vasalemma formations have profound differences. The reef environment of the Vasalemma Formation contrasts with the deeper water clay-rich environment of Hirmuse Formation. The occurrence of symbiotic Rugosa in erect cribrate colonies in both environments suggests that this type of symbiotic life mode had a wide environmental distribution and may have offered some universal benefits to the Rugosa and bryozoans.
O.V. was financially supported by a Sepkoski Grant from the Paleontological Society and Estonian Research Council grant PRG PRG2591. We are grateful to two anonymous reviewers for their constructive comments on the manuscript.
Bary A. de (1879).- Die Erscheinung der Symbiose. Vortrag auf der Versammlung der Naturforschung und Ärtze zu Cassel.- Verlag von Karl J. Trubner, Strassburg, p. 1-30.
Bassett M.G., Popov L.E. & Holmer L.E. (2004).- The oldest-known metazoan parasite?- Journal of Paleontology, Tulsa - OK, vol. 78, p. 1214-1216.
Berkowski B. & Klug C. (2011).- Lucky rugose corals on crinoid stems: Unusual examples of subepidermal epizoans from the Devonian of Morocco.- Lethaia, Oslo, vol. 45, no. 1, p. 24-33.
Cadée G.C. & McKinney F.K. (1994).- A coral-bryozoan association from the Neogene of northwestern Europe.- Lethaia, Oslo, vol. 27, p. 59-66.
Cocks L.R.M. & Torsvik T.H. (2005).- Baltica from the late Precambrian to mid-Palaeozoic times: The gain and loss of a terrane's identity.- Earth Science Reviews, vol. 72, p. 39-66.
Darrell J.G. & Taylor P.D. (1993).- Macrosymbiosis in corals: A review of fossil and potentially fossilizable examples.- Courier Forschunginstitut Senckenberg, Frankfurt-am-Main, vol. 164, p. 185-198.
De Baets K. & Littlewood D.T.J. (2015).- The importance of fossils in understanding the evolution of parasites and their vectors.- Advances in Parasitology, vol. 90, p. 1-51.
De Baets K.D., Huntley J.W., Klompmaker A.A., Schiffbauer J.D. & Muscente A.D. (2021a).- The fossil record of parasitism: Its extent and taphonomic constraints. In: De Baets K. & Huntley J.W. (eds.), The evolution and fossil record of parasitism.- Springer, Cham, p. 1-50.
De Baets K., Huntley J.W., Scarponi D., Klompmaker A.A. & Skawina A. (2021b).- Phanerozoic parasitism and marine metazoan diversity: Dilution versus amplification.- Philosophical Transactions of the Royal Society B (Biological Sciences), London, vol. 376, no. 1837, p. 1-13. DOI: 10.1098/rstb.2020.0366
De Baets K., Klug C. & Korn D. (2011).- Devonian pearls and ammonoid-endoparasite co-evolution.- Acta Palaeontologica Polonica, Warsaw, vol. 56, p. 159-180.
De Baets K., Keupp H. & Klug C. (2015).- Parasites of ammonoids. In: Klug C., Korn D., De Baets K., Kruta I. & Mapes R.H. (eds.), Ammonoid paleobiology: From anatomy to ecology.- Springer, Berlin, p. 837-875.
Dijk J. van & De Baets K. (2021).- Biodiversity and host-parasite (co)extinction. In: De Baets K. & Huntley J.W. (eds.), The evolution and fossil record of parasitism: Coevolution and paleoparasitological techniques. Topics in Geobiology, vol. 50, p. 75-97.
Eichwald E. (1842).- Die Urwelt Russlands, durch Abbildungen erlaeutert. Part 2. St. Petersburg vol. 1, p. 1-184.
Elias R.J. (1982).- Latest Ordovician solitary rugose corals of eastern North America.- Bulletins of American Paleontology, Ithaca, vol. 81, p. 1-116.
Hall J. (1847).- Palćontology of New York.- Natural History of New York, Albany - NY, vol. 1, 338 p. (85 Pls.) URL: https://www.biodiversitylibrary.org/page/42447613
Hints L. & Meidla T. (1997).- Oandu Stage. In: Geology and mineral resources of Estonia).- Estonian Academy Publishers, Tallinn, p. 76-79.
Huntley J.W. & De Baets K. (2015).- Trace fossil evidence of trematode-bivalve parasite-host interactions in deep time.- Advances in Parasitology, vol. 90, p. 201-231.
Huntley J., De Baets K., Scarponi D., Linehan L., Epa Y., Jacobs G. & Todd J. (2021).- Bivalve mollusks as hosts in the fossil record. In: De Baets K. & Huntley J.W. (eds.), The evolution and fossil record of parasitism: Coevolution and paleoparasitological techniques.- Topics in Geobiology, vol. 50, p. 251-287.
Kaljo D.L. (1958).- Some new and little-known Baltic tetracorals.- Trudy Instituta Geologii AN ESSR, Tallinn, vol. 3, p. 101-123 [in Russian].
Kaljo D.L. (1961).- Some additional data on the study of Ordovician streptelasmids in Estonia.- Trudy Instituta Geologii AN ESSR, Tallinn, vol. 6, p. 51-67 [in Russian].
Kröger B., Hints L. & Lehnert O. (2014).- Age, facies, and geometry of the Sandbian/Katian (Upper Ordovician) pelmatozoan-bryozoan-receptaculitid reefs of the Vasalemma Formation, northern Estonia.- Facies, Erlangen, vol. 60, p. 963-986.
Kröger B., Hints L. & Lehnert O. (2017).- Ordovician reef and mound evolution: The Baltoscandian picture.- Geological Magazine, Cambridge, vol. 154, no. 4, p. 683-706.
Kröger B., Ernst A., Penny A., Nakrem H.A. & Toom U. (2023).- Scale dependent diversity of bryozoan assemblages in the reefs of the Late Ordovician Vasalemma Formation, Estonia.- Lethaia, Oslo, vol. 56, no. 2, p. 1-16.
Lopez-Gappa J. & Liuzzi M.G. (2021).- An unusual symbiotic relationship between a cyclostome bryozoan and a thecate hydroid.- Symbiosis, Dordrecht, vol. 85, p. 217-223.
Gorjunova R.V. & Lavrentjeva V.D. (1993).- Morphology and system of the cryptostome bryozoans.- Trudy Paleontologischeskogo Instituta, Moscow, vol. 257, p. 1-150 [in Russian].
Ma J., Taylor P.D. & Buttler C.J. (2020).- Sclerobionts associated with Orbiramus from the Early Ordovician of Hubei, China, the oldest known trepostome bryozoan.- Lethaia, Oslo, vol. 54, p. 443-456.
Männik P. (2017).- Conodont biostratigraphy of the Oandu Stage (Katian, Upper Ordovician) in NE Estonia.- Estonian Journal of Earth Sciences, Tallinn, vol. 66, no. 1, p. 1-12.
Martin B.D. & Schwab E. (2013).- Current usage of symbiosis and associated terminology.- International Journal of Biology, Richmond Hill, vol. 5, p. 32-45.
McAuley R.J. & Elias R.J. (1990).- Latest Ordovician to earliest Silurian solitary rugose corals of the east-Central United States.- Bulletins of American Paleontology, Washington, vol. 98, p. 1-82.
Nestor H. & Einasto R. (1997).- Ordovician and Silurian carbonate sedimentation basin. In: Raukas A. & Teedumäe A. (Eds.), Geology and Mineral Resources of Estonia.- Estonian Academy Publishers, Tallinn, p. 192-204.
Palmer T.J. & Wilson M.A. (1988).- Parasitism of Ordovician bryozoans and the origin of pseudoborings.- Palaeontology, Cambridge (UK), vol. 31, p. 939-949.
Plusquellec Y. & Bigey F.P. (2011).- New data on the intergrowth of Rugosa-Bryozoa in the Lower Devonian of North Gondwana.- Carnets Geol., Madrid, vol. 19, no. 18, p. 421-437. DOI: 10.4267/2042/70538
Robin N. (2021).- Importance of data on fossil symbioses for parasite-host evolution. In: De Baets K. & Huntley J.W. (eds.), The evolution and fossil record of parasitism.- Topics in Geobiology, vol. 50, p. 51-73.
Sepkoski J.J. Jr (1981).- A factor analytic description of the Phanerozoic marine fossil record.- Paleobiology, Tulsa - OK, vol. 7, no. 1, p. 36-53. DOI: 10.1371/journal.pone.0242924
Sepkoski J.J. Jr (1984).- A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic families and mass extinctions.- Paleobiology, Tulsa - OK, vol. 10, no. 2, p. 246-267.
Słowiński J., Surmik D., Duda P. & Zatoń M. (2020).- Assessment of serpulid-hydroid association through the Jurassic: A case study from the Polish Basin.- PLoS ONE, San Francisco - CA, vol. 15, no. 12, article e0242924, p. 1-23.
Spjeldnaes N. (1984).- Upper Ordovician bryozoans from Ojl Myr, Gotland, Sweden.- Bulletin of the Geological Institutions of the University of Uppsala (N.S.), vol. 10, p. 1-66.
Suárez Andrés J.L., Sendino C. & Wilson M.A. (2020).- Coral-bryozoan associations through the fossil record: Glimpses of a rare interaction. In: Wyse Jackson P. & Zágoršek K. (eds.), Bryozoan studies 2019.- Czech Geological Survey, Prague, p. 157-168.
Tapanila L. (2005).- Palaeoecology and diversity of endosymbionts in Palaeozoic marine invertebrates: Trace fossil evidence.- Lethaia, Oslo, vol. 38, p. 89-99.
Taylor P.D. (1990).- Preservation of soft-bodied and other organisms by bioimmuration - A review.- Palaeontology, Cambridge (UK), vol. 33, p. 1-17.
Taylor P.D. (2012).- A new bryozoan genus from the Jurassic of Switzerland, with a review of the cribrate colony-form in bryozoans.- Swiss Journal of Palaeontology, Zurich, vol. 131, p. 201-210.
Taylor P.D. (2015).- Differentiating parasitism and other interactions in fossilized colonial organisms.- Advances in Parasitology, vol. 90, p. 329-347.
Torsvik T.H., Cocks L.R.M. & Harper D.A.T. (2013).- New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. In: Servais T. (ed.), Early Palaeozoic biogeography and palaeogeography.- Geological Society Memoirs, no. 38, p. 5-24.
Vinn O. Ernst A. & Toom U. (2016).- Earliest symbiotic rugosans in cystoporate bryozoan Ceramopora intercellata Bassler, 1911 from Late Ordovician of Estonia (Baltica).- Palæogeography, Palæoclimatology, Palæoecology, vol. 461, p. 140-144.
Vinn O., Ernst A. & Toom U. (2017).- Rare rugosan-bryozoan intergrowth from the Late Ordovician of Estonia.- Carnets Geol., Madrid, vol. 17, no. 7, p. 145-151. DOI: 10.4267/2042/62664
Vinn O., Toom U. & Ernst A. (2018).- Intergrowth of Orbignyella germana Bassler, 1911 (Bryozoa) and Lambelasma carinatum Weyer, 1993 (Rugosa) in the pelmatozoan-bryozoan-receptaculitid reefs from the Late Ordovician of Estonia.- Palaeontologia Electronica, Amherst, vol. 21.1.4A, p. 1-7.
Vinn O., Wilson M.A. & Ernst A. (2023).- Macroscopic symbiotic endobionts in Phanerozoic bryozoans.- Palæogeography, Palæoclimatology, Palæoecology, vol. 615, 111453, p. 1-18.
Zapalski M.K. (2007).- Parasitism versus commensalism - the case of tabulate endobionts.- Palaeontology, Cambridge (UK), vol. 50, p. 1375-1380.
Zapalski M.K. (2011).- Is absence of proof a proof of absence? Comments on commensalism.- Palæogeography, Palæoclimatology, Palæoecology, vol. 302, no. 3-4, p. 484-488.
Zapalski M.K. & Hubert B.L. (2011).- First fossil record of parasitism in Devonian calcareous sponges (stromatoporoids).- Parasitology, Cambridge (UK), vol. 138, p. 132-138.