◄ Carnets Geol. 25 (9) ►
![]()
Outline
[1. Introduction]
[2. Material and method]
[3. Systematics]
[4. Discussion and conclusions]
[Bibliographic references] [Plates] and ...
[Appendix]
Corresponding author
Laboratoire du Groupe de recherche en Pal�obiologie et biostratigraphie des
Ammonites (GPA), 65 Grand rue, F-04170 La Mure-Argens (France)
R�serve naturelle nationale g�ologique de Haute-Provence, Conseil
d�partemental des Alpes de Haute-Provence, 13, rue du Docteur-Romieu, CS 70216,
04995 Digne-Les-Bains cedex 9 (France);
Univ. Rennes, CNRS, G�osciences Rennes, UMR 6118, 35000 Rennes (France);
Laboratoire du Groupe de recherche en Pal�obiologie et biostratigraphie des
Ammonites (GPA), 65 Grand rue, F-04170 La Mure-Argens (France)
Published online in final form (pdf) on June 7, 2025
DOI 10.2110/carnets.2025.2509
![]()
[Editor: Christina Ifrim; language editor: Stephen Eagar; technical editor: Bruno
R.C. Granier]
![]()
In the present work, we study 335 specimens of Deshayesitidae (Ammonoidea) from the vicinity of the village of Les Ferres (SE France). The Deshayesitidae is the current key family for the ammonite biostratigraphy of the lower Aptian (Lower Cretaceous). Despite poorly preserved, the studied material could have been identified at species rank in most cases and allowed establishing the biostratigraphic frame of the lower Aptian of the study area. The results are as follows: (1) the studied samples range from the Deshayesites forbesi Zone to the Dufrenoyia furcata Zone; (2) their ontogenetic sequence is described; (3) their evolutionary patterns are consistent with those observed in other samples of Deshayesitidae; (4) no dimorphism is conspicuous; (5) the identified taxa are, from earliest to latest: Deshayesites sp. (Deshayesites forbesi Zone, rounded ventral area probably without smooth siphonal band), Deshayesites multicostatus Swinnerton, 1935 (index of subzone, intermediate smooth siphonal band then rounded ventral area), Deshayesites grandis Spath, 1930 (index of subzone, longer smooth siphonal band then subtabulate ventral area on phragmocone), Dufrenoyia furcata (J. de C. Sowerby, 1836) (index of subzone and zone, even longer smooth siphonal band then subtabulate ventral area, onset of rounded to claviform latero-ventral tubercles in inner whorls) and Dufrenoyia dufrenoyi (Orbigny, 1841) (index of subzone, even more longer smooth siphonal band, latero-ventral tubercles claviform only).
• ammonites;
• Deshayesites;
• Dufrenoyia;
• biostratigraphy;
• intraspecific variability;
• evolution
Bersac S. & Bert D. (2025).- The Deshayesitidae (Ammonoidea, Ancyloceratina) in the lower Aptian (Lower Cretaceous) of the Les Ferres Aptian Basin (southeastern France).- Carnets Geol., Madrid, vol. 25, no. 9, p. 177-200. DOI: 10.2110/carnets.2025.2509
Les Deshayesitidae (Ammonoidea, Ancyloceratina) de l'Aptien inf�rieur (Cr�tac� inf�rieur) du Bassin Aptien de Les Ferres (SE de la France).- Dans le pr�sent travail, nous �tudions 335 sp�cimens de Deshayesitidae (Ammonoidea) provenant du secteur du village de Les Ferres (SE France). Les Deshayesitidae sont une famille cl� pour la bostratigraphie par ammonites de l'Aptien inf�rieur (Cr�tac� inf�rieur). Malgr� un �tat de conservation m�diocre, le mat�riel �tudi� a pu �tre identifi� au rang sp�cifique et a fourni une datation pr�cise dans la plupart des cas. Les r�sultats sont les suivants: (1) les �chantillons �tudi�s s'�tendent de la Zone � Deshayesites forbesi � la Zone � Dufrenoyia furcata ; (2) leur s�quence ontog�nique est d�crite ; (3) leur sch�ma �volutif est coh�rent avec ceux observ�s dans d'autres �chantillons de Deshayesitidae ; (4) aucun dimorphisme n'est mis en �vidence ; (5) les taxons identifi�s sont, du plus ancien au plus r�cent: Deshayesites sp. (Zone � Deshayesites forbesi, r�gion ventrale arrondie probablement sans bande siphonale lisse), Deshayesites multicostatus Swinnerton, 1935 (indice de sous-zone, bande siphonale lisse de longueur interm�diaire puis r�gion ventrale arrondie), Deshayesites grandis Spath, 1930 (indice de sous-zone, bande siphonale lisse plus longue puis r�gion ventrale subtabul�e sur le phragmoc�ne), Dufrenoyia furcata (J. de C. Sowerby, 1836) (indice de sous-zone et de zone, bande siphonale lisse encore plus longue puis r�gion ventrale subtabul�e, apparition de tubercules lat�ro-ventraux arrondis � claviformes dans les tours internes) et Dufrenoyia dufrenoyi (Orbigny, 1841) (indice de sous-zone, bande siphonale lisse encore plus longue, tubercules lat�ro-ventraux claviformes uniquement).
• ammonites ;
• Deshayesites ;
• Dufrenoyia ;
• biostratigraphie ;
• variabilit� intrasp�cifique ;
• �volution
The
Les Ferres Aptian Basin (LFAB) is a small subsident basin presently located in
the vicinity of the village of Les Ferres, in the Est�ron valley
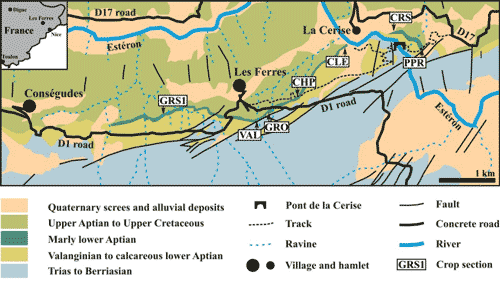
(Alpes-Maritimes department, southeastern France, Fig. 1 ![]() ). A general
presentation of this basin was published in an introductory article by Bersac
and Bert (2019). It is characterized by plurimetric deposits (Fig.
2
). A general
presentation of this basin was published in an introductory article by Bersac
and Bert (2019). It is characterized by plurimetric deposits (Fig.
2 ![]() )
with abundant benthic and nectonic macrofossils, especially ammonites. The age
of the lower Aptian series from this basin extends from the Deshayesites
forbesi Zone (in the sense of Bersac et al.,
2012, see Fig. 3) to the late Dufrenoyia furcata Zone (Dufrenoyia
dufrenoyi Subzone). The LFAB provided many representatives of Deshayesitidae
Stoyanow, 1949 (Bersac & Bert,
2019). This ammonite
family spans the whole lower Aptian and occurs over a wide geographic range (Casey,
1964; Young, 1974; Etayo-Serna,
1979; Renz, 1982; Bogdanova
& Mikhailova, 2004; Dutour,
2005; Ropolo et
al., 2006; Barragan & Szives,
2007; Obata
& Matsukawa, 2007; Garc�a-Mond�jar et
al., 2009; Moreno-Bedmar et
al., 2010, 2012,
2018, 2023; Lehmann et
al., 2015; Rogov & Mironenko,
2016; Lehmann & Bulot,
2020).
It is the key family for the ammonite biostratigraphy of the lower Aptian since
it provided most or all of its index species for this interval of time according
to the local zonal schemes (Baraboshkin, 2004; Casey et
al., 1998; Reboulet et al.,
2018). In our introductory article on the ammonite fauna of the LFAB (Bersac
& Bert, 2019), we only quoted and figured the Deshayesitidae taxa we
recognized in this area: Deshayesites sp., Deshayesites multicostatus
Swinnerton, 1935, Deshayesites grandis Spath,
1930, Dufrenoyia
furcata (J. de C. Sowerby, 1836) and Dufrenoyia dufrenoyi (Orbigny,
1841). The purpose of the present article is to describe our sample of
Deshayesitidae from the LFAB and to argue about their taxonomic assignation.
)
with abundant benthic and nectonic macrofossils, especially ammonites. The age
of the lower Aptian series from this basin extends from the Deshayesites
forbesi Zone (in the sense of Bersac et al.,
2012, see Fig. 3) to the late Dufrenoyia furcata Zone (Dufrenoyia
dufrenoyi Subzone). The LFAB provided many representatives of Deshayesitidae
Stoyanow, 1949 (Bersac & Bert,
2019). This ammonite
family spans the whole lower Aptian and occurs over a wide geographic range (Casey,
1964; Young, 1974; Etayo-Serna,
1979; Renz, 1982; Bogdanova
& Mikhailova, 2004; Dutour,
2005; Ropolo et
al., 2006; Barragan & Szives,
2007; Obata
& Matsukawa, 2007; Garc�a-Mond�jar et
al., 2009; Moreno-Bedmar et
al., 2010, 2012,
2018, 2023; Lehmann et
al., 2015; Rogov & Mironenko,
2016; Lehmann & Bulot,
2020).
It is the key family for the ammonite biostratigraphy of the lower Aptian since
it provided most or all of its index species for this interval of time according
to the local zonal schemes (Baraboshkin, 2004; Casey et
al., 1998; Reboulet et al.,
2018). In our introductory article on the ammonite fauna of the LFAB (Bersac
& Bert, 2019), we only quoted and figured the Deshayesitidae taxa we
recognized in this area: Deshayesites sp., Deshayesites multicostatus
Swinnerton, 1935, Deshayesites grandis Spath,
1930, Dufrenoyia
furcata (J. de C. Sowerby, 1836) and Dufrenoyia dufrenoyi (Orbigny,
1841). The purpose of the present article is to describe our sample of
Deshayesitidae from the LFAB and to argue about their taxonomic assignation.
|
|
Figure 1: Map of the Les Ferres area with location of the sections
quoted in the text. |
|
|
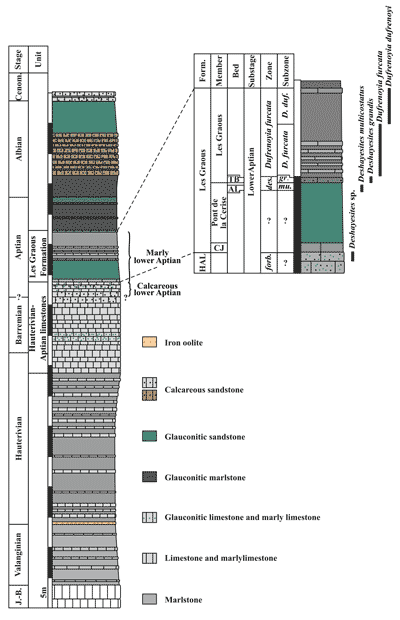
Figure 2: Synthetic
log of the Lower Cretaceous of the Les Ferres area. J.-B.: Jurassic and
Berriasian; Cenom.: Cenomanian; Form.: Formation; HAL: Hauterivian-Aptian
limestones; CJ: Combe de Joinet Member; AL: Ammonitoceras
Level; TB: Toxoceratoides Bed; forb.: Deshayesites forbesi;
des.: Deshayesites deshayesi; mu.: Deshayesites
multicostatus; gr.: Deshayesites grandis; D. duf.:
Dufrenoyia dufrenoyi. |
Material
The
material studied is represented by 335 specimens from 7 sections of the LFAB,
namely Les Graous 1 section (GRS1), La Valliere section (VAL), La Graou section
(GRO), Combe de Joinet section (CHP), Pont de la Cerise section (CLE), Combe de
Marin section (CRS) and Pont Paire section (PPR) (Fig. 1 ![]() ). The
lithostratigraphy, ammonite occurrence, and biostratigraphy of these sections are
described and figured in Bersac and Bert
(2019). The specimens are
preserved as marly limestone internal casts, mostly fragmented and deformed by
compaction.
). The
lithostratigraphy, ammonite occurrence, and biostratigraphy of these sections are
described and figured in Bersac and Bert
(2019). The specimens are
preserved as marly limestone internal casts, mostly fragmented and deformed by
compaction.
The
specimens occur in various lithostratigraphic units from bottom to top (Fig.
2 ![]() ):
the last bed of the Hauterivian-Aptian limestones, the Combe de Joinet Member
(CJ), the Ammonitoceras Level (AL) of
the Pont de la Cerise Member (Deshayesites
multicostatus Subzone, Deshayesites
deshayesi Zone) and the Les Graous Member (Deshayesites grandis
Subzone, D. deshayesi Zone, and D.
furcata Zone) and its basal Toxoceratoides
Bed (TB).
):
the last bed of the Hauterivian-Aptian limestones, the Combe de Joinet Member
(CJ), the Ammonitoceras Level (AL) of
the Pont de la Cerise Member (Deshayesites
multicostatus Subzone, Deshayesites
deshayesi Zone) and the Les Graous Member (Deshayesites grandis
Subzone, D. deshayesi Zone, and D.
furcata Zone) and its basal Toxoceratoides
Bed (TB).
The whole studied material belongs to St�phane Bersac's collection and is housed in the premises of the R�serve naturelle nationale g�ologique de Haute-Provence (RNNGHP, Digne-les-bains, France). The database of the studied specimens is available in the supplementary appendix.
Method
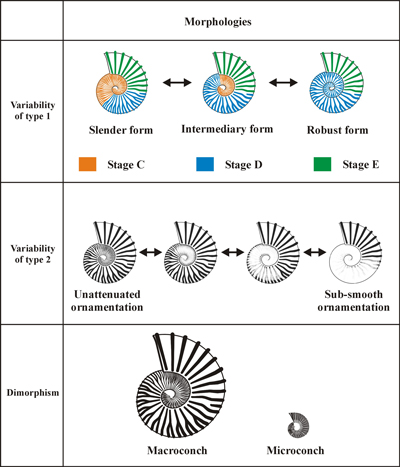
We follow Bersac and Bert (2012, 2015, p. 266, Figs. 1, 3) regarding the ontogenetic sequences of the flanks and the ventral area, the patterns of intraspecific variability and the criteria of taxonomic identification of the Deshayesitidae (based on the population concept of species, e.g., Tintant, 1963; Dzik, 1985). They are summarized above.
The
ontogenetic sequence of the inner whorls and the flanks is represented by 5
successive stages (Fig. 3 ![]() ):
):
the Globular stage A,
the stage B of Kossmatella type in the innermost whorls,
the stage C with thin and dense main ribs, and with numerous intermediate ribs,
the stage D with strong ribs with a more spaced and robust ornamentation, and less intermediate ribs,
the adult stage E with spaced, strong, simple and straight ribs.
Three
main patterns of intraspecific variability are recognised (Fig. 3 ![]() ):
):
variability of type 1. It is determined by the duration of the stages C and D. All the morphologies coexist within an instantaneous population, between extremely robust individuals with absent stage C (and thus long stage D) and extremely slender individuals with a long stage C (and thus short stage D). This variability follows general 'laws' which were subject of several recent studies (Bersac & Bert, 2012; Bert, 2013, 2019; Bert et al., 2023; De Baets et al., 2015, with references).
variability of type 2. It relates exclusively to the intensity of ornamentation/relief of the ribs: the secondary attenuation of ornamentation (see Bert, 2013). All potential intermediates can be observed in a same sample between individuals with ornamentation normally expressed to subsmooth forms. This ornamental attenuation may concern all or part of the shell with exception of the ontogenetic stage E, which is never attenuated.
probable sexual dimorphism with macroconchs and microconchs (these latter never express the stage E).
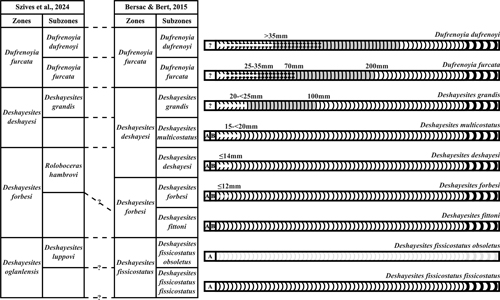
The
evolution of the Deshayesitidae is
characterized by the likely increasing complexity of the septal suture over time
and by the onset of a
new postembryonic stage (stage B) after the stage A in the inner whorls of the populations from
the
Deshayesites fittoni Subzone (Deshayesites forbesi Zone sensu Bersac et al.,
2012) and
of a smooth siphonal band in the populations from the Deshayesites
deshayesi Subzone (Deshayesites deshayesi Zone sensu Bersac
et al., 2012), then of latero-ventral tubercles from the Dufrenoyia
furcata Subzone (Dufrenoyia furcata Zone, Fig. 4 ![]() ). Both these two
last characters evolve through a neotenic process by expanding more and more on
the venter of the shell over time (Dutour, 2005; Bersac & Bert,
2012, 2015). Consequently, the key diagnostic characters of the Deshayesitidae
are all evolutionary characters: the mean value of the end diameter of the
smooth siphonal band (SSB) and of the latero-ventral tubercles measured in an
isochronous population, the shape of these tubercles and the presence of a
subtabulate ventral area (i.e.,
ventral area flattened but without neat limit with the flanks) on the
phragmocone.
). Both these two
last characters evolve through a neotenic process by expanding more and more on
the venter of the shell over time (Dutour, 2005; Bersac & Bert,
2012, 2015). Consequently, the key diagnostic characters of the Deshayesitidae
are all evolutionary characters: the mean value of the end diameter of the
smooth siphonal band (SSB) and of the latero-ventral tubercles measured in an
isochronous population, the shape of these tubercles and the presence of a
subtabulate ventral area (i.e.,
ventral area flattened but without neat limit with the flanks) on the
phragmocone.
An additional pattern of variability of the Deshayesitidae is the intraspecific variability of the evolutionary characters recognized above: in a given isochronous population, these characters are not obviously all identical from a specimen to another, but vary - sometimes considerably - in a way assumed to be Gaussian (character value distribution represented by a bell/Gaussian curve). The species identification is based on the calculation of the mean value of these evolutionary characters in a given isochronous population (population concept of species, see Dzik, 1985).
The measured shell parameters are the diameter of the shell (D), the maximum whorl height (H), the rib density on the last half whorl (R), the end diameter of the smooth siphonal band (SSB) and the end diameter of the latero-ventral tubercles. Because of the deformation of the material, these parameters (given in the supplementary appendix) have to be taken as estimations. Consequently, no statistical analysis was performed apart from the mean values of SSB and R. Due to flattening of most of the specimens, the whorl width was not measured.
The synonymy lists we present only contain the works with the original description of the taxa here recognized, those containing their emended description and those quoting specimens from the LFAB.
|
|
Figure 3: Patterns of intraspecific variability of the
Deshayesitidae, after
Bersac & Bert (2012). |
|
|
Figure 4: Biostratigraphic
chart used in the present work (Bersac & Bert,
2015) with characteristics of the ventral area of its index species,
parallelized with the Standard Ammonite Zonation of the Western Tethys [*] Mediterranean Province (Szives et
al., 2024). Subtabulate ventral area is represented in clear grey. Stages C
and D are not differentiated not to overburden the figure (see text for
explanations). |
Ancyloceratina Wiedmann, 1966
Deshayesitaceae Stoyanow, 1949
Deshayesitidae Stoyanow, 1949
Deshayestites Kazansky, 1914
Deshayesites sp.
Pl. 1 ![]() ,
figs. A-E
,
figs. A-E
? 1982 Deshayesites sp. - Ragazzi, p. 74-75.
? 1982 Deshayesites latelobata (Sinzow) - Ragazzi, p. 75.
v 2019 Deshayesites sp. - Bersac & Bert, p. 154, 157-158, 161, 167, Figs. 5, 10, 12, 14-15, Pl. 1, figs. 4-5.
Material studied: 12 specimens from the uppermost bed of the Hauterivian-Aptian limestones (Deshayesites forbesi Zone) of GRS1 (Bersac & Bert, 2019, Fig. 5), CLE (ibid., Fig. 12) and CRS (ibid., Fig. 14). See the supplementary appendix for the complete list of the specimens with measurements.
Description:
Most of the specimens are whorl fragments. Their shell is discoid to moderately
evolute, with more or less covering whorls. The maximum observed D is 133 mm (Pl. 1 ![]() , fig. A) and the maximum observed H is 63 mm. R was measurable on only two
specimens (SBC.06061-00001/GRS115 and SBC.06061-00007/CLE030) and is
respectively 21 and 27. The umbilical wall is steep and the flanks are flat or
slightly convex. No specimen has its whorls observable below D=25 mm and the
ontogenetic stages A and B cannot consequently be studied. The observable
ontogenetic sequence of the shell flanks starts with the stage C having dense,
more or less flexuous thin ribs, and more or less regular alternation between
primary and intercalary ribs. Intercalary ribs are usually more numerous and
emerge at various heights on the flanks. The stage D, that follows the stage C,
presents thicker, less dense and less flexuous ribs, with a rather regular
alternation between primary and intercalary ribs. No specimen shows a
periumbilical rib thickening. The specimens are too small or fragmentary to
exhibit the adult stage E. The transition between the stages C and D can be more
or less sudden, the ribs are wedge shaped or not and the rib relief is sharp or
more attenuated, especially in the middle part of the flanks. The shape of the
specimens varies between a slender morphology with high rib density, high growth
in whorl height and narrow whorl width and a robust morphology with lower rib
density and lower growth in whorl height.
, fig. A) and the maximum observed H is 63 mm. R was measurable on only two
specimens (SBC.06061-00001/GRS115 and SBC.06061-00007/CLE030) and is
respectively 21 and 27. The umbilical wall is steep and the flanks are flat or
slightly convex. No specimen has its whorls observable below D=25 mm and the
ontogenetic stages A and B cannot consequently be studied. The observable
ontogenetic sequence of the shell flanks starts with the stage C having dense,
more or less flexuous thin ribs, and more or less regular alternation between
primary and intercalary ribs. Intercalary ribs are usually more numerous and
emerge at various heights on the flanks. The stage D, that follows the stage C,
presents thicker, less dense and less flexuous ribs, with a rather regular
alternation between primary and intercalary ribs. No specimen shows a
periumbilical rib thickening. The specimens are too small or fragmentary to
exhibit the adult stage E. The transition between the stages C and D can be more
or less sudden, the ribs are wedge shaped or not and the rib relief is sharp or
more attenuated, especially in the middle part of the flanks. The shape of the
specimens varies between a slender morphology with high rib density, high growth
in whorl height and narrow whorl width and a robust morphology with lower rib
density and lower growth in whorl height.
None of the specimens has its ventral area preserved below D=25 mm. From this diameter, the ventral area is always rounded with no rib attenuation. The septal suture is too poorly preserved to be described and no specimen has its peristome preserved.
Discussion: The range of variability seems to be continuous within the sample and similar to what is generally observed in monospecific samples of Deshayesitidae (Dutour, 2005; Garcia & Moreno-Bedmar, 2010; Bersac & Bert, 2012, 2015; Moreno-Bedmar et al., 2014) and the sample is thus regarded monospecific. This sample exhibits no evidence of dimorphism but its fragmentary state prevents further discussion (expression of sexual dimorphism among Deshayesitidae is still under debate, see discussion in Moreno & Company, 2007; Bersac & Bert, 2012; Moreno-Bedmar et al., 2014; Lehmann & Bulot, 2020; Frau & Delanoy, 2022). The ventral area was not observed in any of the studied specimens before D=25 mm, and similarly, neither the smooth siphonal band, subtabulate ventral area nor latero-ventral tubercles are present. The absence of latero-ventral tubercles allows to assign unambiguously these Deshayesitidae to the genus Deshayesites Kazansky, 1914. The absence of a subtabulate ventral area allows to consider them as representatives of a species earlier than Deshayesites grandis. These specimens could belong to Deshayesites deshayesi (Orbigny, 1841) if they would have SSB<15mm, Deshayesites multicostatus if 15mm ≤ SSB ≤ 20mm or an even earlier species such as Deshayesites forbesi Casey, 1961, Deshayesites fissicostatus (Phillips, 1829), Deshayesites tuarkyricus Bogdanova, 1983, etc. if they do not bear a smooth siphonal band. Because of the impossibility to observe the ventral area in inner whorls and thus to determine if a smooth siphonal band is present or not, the taxonomic identification cannot be carried further and these specimens are left in open nomenclature.
The co-occurring ammonite fauna can nevertheless help to estimate more precisely their biostratigraphic position: these Deshayesites occur with representatives of the genus Procheloniceras Spath, 1923 (Douvilleiceratidae Parona & Bonarelli, 1897) and small Ancyloceras Orbigny, 1842, assigned to Ancyloceras rochi Sarkar, 1955 (Bersac & Bert, 2019, Pl. 1, figs. 2, 6-7). Procheloniceras is known probably up to the top of the Deshayesites forbesi Zone (in the sense of Bersac et al., 2012; see Bersac & Bert, 2018) and small Ancyloceras such A. rochi are present in the D. forbesi Zone (Ropolo et al., 1998). The suggested biostratigraphic position of theses Deshayesites is thus the D. forbesi Zone. The absence of the biostratigraphically significant ammonite subfamily Roloboceratinae Casey, 1961, in the uppermost bed of the Hauterivian-Aptian limestones, if not caused by sample bias or unfavorable paleoenvironment, may suggest a biostratigraphic position within the D. forbesi Zone lower than the Roloboceras hambrovi Subzone (see Ropolo et al., 2008; Moreno-Bedmar et al., 2009, 2010; Bersac & Bert, 2012, 2015; Frau et al., 2017, 2023; Frau, 2020; Delanoy et al., 2022, for discussion on the biostratigraphic position of the Roloboceratinae).
Biostratigraphic distribution: Deshayesites forbesi Zone.
Deshayesites multicostatus Swinnerton, 1935
Pl. 2 ![]() ,
figs. A-C; Pl. 3
,
figs. A-C; Pl. 3 ![]() ,
figs. A-C; Pl. 4
,
figs. A-C; Pl. 4 ![]() ,
figs. A-V; Pl. 5
,
figs. A-V; Pl. 5 ![]() ,
figs. A-H
,
figs. A-H
1935 Deshayesites multicostatus sp. nov. - Swinnerton, Pl. 1, figs. 1a-c.
2015 Deshayesites multicostatus - Bersac & Bert, p. 288.
v 2018 Deshayesites gr. latelobatus/involutus - Delanoy et al., p. 38, Fig. 26B.
v 2018 Deshayesites aff. grandis - Delanoy et al., p. 38, Fig. 9B.
v 2019 Deshayesites multicostatus - Bersac & Bert, p. 154, 161, Figs. 5, 7, 10, 12, 15, Pl. 2, figs. 6-8, 13-15, 19.
Material studied: 71 specimens from the Pont de la Cerise Member: 30 from Bed 405 of VAL (Bersac & Bert, 2019, Fig. 7), 1 from Bed 415 of PPR (ibid., Fig. 15) and 40 from Bed 415 of CHP (ibid., Fig. 10). See the supplementary appendix for the complete list of the specimens with measurements.
Description:
The shell is discoid to moderately evolute, with more or less covering whorls.
The umbilical wall is steep and the flanks are flat or slightly convex. The
largest observed specimen is 273 mm in diameter
(SBC.06061-00006/CHP241) but is deformed (Bersac & Bert,
2019,
Pl. 2, figs. 13-15). R is measurable on 26 specimens: it varies from 16 to 36
with an average value of 24.65. Innermost whorls (D < 10mm) are never
preserved on the studied specimens and the ontogenetic stages A and B cannot
consequently be studied. The observable ontogenetic sequence of the shell flanks
starts with the stage C and is followed by the stage D, which have
characteristics similar to the corresponding stages of Deshayesites sp.
previously described. The stage D is followed by the adult stage E with straight,
simple and thick ribs. The transition between these stages is more or less
sudden, similar to the above described specimens. All intermediate forms are
present between slender specimens with high rib density, high growth in whorl
height and narrow whorl width (Pl. 2 ![]() , fig. A) and robust ones with lower rib
density and lower growth in whorl height (Pl. 3
, fig. A) and robust ones with lower rib
density and lower growth in whorl height (Pl. 3 ![]() , figs. A-B).
, figs. A-B).
The
ventral area below D=30 mm is rarely preserved. When it is the case, a smooth
siphonal band or an absence of smothering of the ornamentation on this area can
be present according to the specimens. The end diameter of the smooth siphonal
band (SSB) is measurable on 4 specimens (see supplementary appendix): it ranges
from 14-19 mm (respectively on the specimens Pl. 4 ![]() , figs. Q-R, and
Pl. 4
, figs. Q-R, and
Pl. 4 ![]() , fig.
P) and the average value is 16.5 mm. The rib relief then progressively reappears
on the ventral area. Throughout the ontogeny, the ventral area is rounded,
except on the specimen SBC.06061-00003/VAL009 (Pl. 4
, fig.
P) and the average value is 16.5 mm. The rib relief then progressively reappears
on the ventral area. Throughout the ontogeny, the ventral area is rounded,
except on the specimen SBC.06061-00003/VAL009 (Pl. 4 ![]() , figs. F-G), where it is
subtabulate in inner whorls.
, figs. F-G), where it is
subtabulate in inner whorls.
The septal suture on the few specimens on which it is observable, is too poorly preserved to be described. No specimen has its peristome preserved.
Discussion:
The range of variability is continuous within the sample and similar to that of
the above studied sample. Consequently, the sample is thus considered
monospecific. Among the 71 specimens of the sample, the key diagnostic SSB is
measurable on only 4 of them. The mean value of this character is 16.5 mm in
this sample, which suggests a
specific assignation to the index species D. multicostatus. However, this
result is in itself of poor significance because it is based on very few
specimens. No specimen has a SSB value larger than 20 mm: this element only
suggests that this sample is earlier than the D. grandis Subzone. After
the smooth siphonal band, the rest of the ventral area of the studied specimens
is rounded with no relief attenuation and no latero-ventral tubercles, which
also corresponds to a character predominant in samples of Deshayesitidae earlier
than the D. grandis Subzone. Only one fragmentary specimen presents a
subtabulate ventral area without latero-ventral tubercles (Pl. 4 ![]() , figs. F-G),
which is a character usually predominant in the D. grandis Subzone. On
the basis of these elements, we consider that the sample has to be assigned to D.
multicostatus. In our opinion, the presence in the sample of only one
specimen with a subtabulate ventral area does not constitute evidence of
polysubzonal time averaging within this sample (i.e.,
a mixture of Deshayesitidae from the D. multicostatus and the D.
grandis subzones). Conspecific samples having specimens with primitive
characters and specimens with derived ones were described in literature in
various groups of Ammonoidea (e.g.,
Tintant, 1963;
Dzik, 1985 � the 'Gauss Bell effect') including
Deshayesitidae (Moreno-Bedmar et
al., 2014). Moreover, the assignation of the sample to the D.
multicostatus Subzone is consistent with the co-occurring ammonite fauna
(see
discussion in Bersac & Bert,
2019).
, figs. F-G),
which is a character usually predominant in the D. grandis Subzone. On
the basis of these elements, we consider that the sample has to be assigned to D.
multicostatus. In our opinion, the presence in the sample of only one
specimen with a subtabulate ventral area does not constitute evidence of
polysubzonal time averaging within this sample (i.e.,
a mixture of Deshayesitidae from the D. multicostatus and the D.
grandis subzones). Conspecific samples having specimens with primitive
characters and specimens with derived ones were described in literature in
various groups of Ammonoidea (e.g.,
Tintant, 1963;
Dzik, 1985 � the 'Gauss Bell effect') including
Deshayesitidae (Moreno-Bedmar et
al., 2014). Moreover, the assignation of the sample to the D.
multicostatus Subzone is consistent with the co-occurring ammonite fauna
(see
discussion in Bersac & Bert,
2019).
Biostratigraphic distribution: Deshayesites multicostatus Subzone, Deshayesites deshayesi Zone.
Deshayesites grandis Spath, 1930
Pl. 5 ![]() ,
figs. I-N;
Pl. 6
,
figs. I-N;
Pl. 6 ![]() ,
figs. A-G
,
figs. A-G
1930 Deshayesites grandis sp. nov. - Spath, Pl. XVII, figs. 2a, b.
2012 Deshayesites grandis - Bersac & Bert, p. 251.
v 2019 Deshayesites grandis - Bersac & Bert, p. 154, 165, Figs. 5, 10, 12, 15, Pl. 3, figs. 10-12.
Material studied: 22 specimens from the Toxoceratoides Bed of the Les Graous Member of GRS1 (Bersac & Bert, 2019, Fig. 5), CHP (ibid., Fig. 10) and CLE (ibid., Fig. 12). See the supplementary appendix for the complete list of the specimens with measurements.
Description:
The shell is discoid to moderately evolute, with more or less covering whorls.
The umbilical wall is steep. The flanks are flat or slightly convex. The maximum
observed diameter is 49 mm (SBC.06061-00001/GRS351,
Pl. 6 ![]() , figs. A-B). R
is measurable on 10 specimens and varies from 18 to 31 with an average value of
23.4. Innermost whorls are never preserved on the studied specimens and the
ontogenetic stages A and B cannot consequently be studied. Ornamentation is
represented by more or less dense and flexuous ribs, with more or less regular
alternation between primary and intercalary ribs. Intercalary ribs are usually
more numerous and emerge at various heights on the flanks. The small size of the
specimens prevents differentiation stage C from the stage D, and to observe the
adult stage E. Periumbilical rib thickening are occasionally present. The ribs
can be wedge shaped or not and the rib relief can be sharp or more attenuated,
especially in the middle part of the flanks. Shape of the specimens vary between
a slender morphology with high rib density, high growth in whorl height (Pl. 5
, figs. A-B). R
is measurable on 10 specimens and varies from 18 to 31 with an average value of
23.4. Innermost whorls are never preserved on the studied specimens and the
ontogenetic stages A and B cannot consequently be studied. Ornamentation is
represented by more or less dense and flexuous ribs, with more or less regular
alternation between primary and intercalary ribs. Intercalary ribs are usually
more numerous and emerge at various heights on the flanks. The small size of the
specimens prevents differentiation stage C from the stage D, and to observe the
adult stage E. Periumbilical rib thickening are occasionally present. The ribs
can be wedge shaped or not and the rib relief can be sharp or more attenuated,
especially in the middle part of the flanks. Shape of the specimens vary between
a slender morphology with high rib density, high growth in whorl height (Pl. 5 ![]() , figs.
M-N) and narrow whorl width and a more robust morphology with lower rib
density and lower growth in whorl height (Pl. 5
, figs.
M-N) and narrow whorl width and a more robust morphology with lower rib
density and lower growth in whorl height (Pl. 5 ![]() , figs. A-C).
, figs. A-C).
When
observable, the ventral area in inner whorls always presents a smooth siphonal
band. SSB (Pl. 6 ![]() , fig. E) was able to be measured on 7 specimens: ranging from
16 to 24 mm with an average value of 20.43 mm. After a brief attenuated siphonal
band, the ventral area is subtabulate except for one poorly preserved specimen,
where it is rounded (SBC.06061-00006/CHP361).
On the few specimens on which it is observable, the septal suture is too poorly
preserved to be described. No specimen has its peristome preserved.
, fig. E) was able to be measured on 7 specimens: ranging from
16 to 24 mm with an average value of 20.43 mm. After a brief attenuated siphonal
band, the ventral area is subtabulate except for one poorly preserved specimen,
where it is rounded (SBC.06061-00006/CHP361).
On the few specimens on which it is observable, the septal suture is too poorly
preserved to be described. No specimen has its peristome preserved.
Discussion: The range of variability is continuous within the sample and similar to that of the above studied samples. Consequently, the sample is considered monospecific. Despite the small size of the specimens, the range of variability of R is similar to that of the sample of D. multicostatus. Taking into account that the studied material is deformed, the average value of SSB tentatively corresponds to the species D. grandis. The ventral area following the smooth siphonal band is subtabulate for most of the specimens, also characteristic of D. grandis. We consequently assign this sample to that species.
Biostratigraphic distribution: Deshayesites grandis Subzone, Deshayesites deshayesi Zone.
Dufrenoyia Kilian & Reboul, 1915
Dufrenoyia furcata (J. de C. Sowerby, 1836)
Pl. 6 ![]() ,
figs. H-X1; Pl. 7
,
figs. H-X1; Pl. 7 ![]() ,
figs. A-Z
,
figs. A-Z
1836 Ammonites furcatus sp. nov. - J. de C. Sowerby in Fitton, Pl. XIV, fig. 17.
? 1971 Dufrenoyia dufrenoyi - Cotillon, 1971, p. 146.
? 1982 Dufrenoyia furcata - Ragazzi, p. 74, 76.
2005 Dufrenoyia furcata - Dutour, Pl. 29, figs. 10-16.
v 2011 Dufrenoyia sp. - Bert & Bersac, Fig. 41.
2012 Dufrenoyia furcata - Bersac & Bert, p. 253.
v 2018 Dufrenoyia dufrenoyi - Delanoy et al., Figs. 8D, 20E-H.
v 2019 Dufrenoyia furcata - Bersac & Bert, p. 154, 165, Figs. 5, 9-10, 12, 15, Pl. 5, figs. 5-9.
Material studied: 210 specimens from the Les Graous Member of GRS1 (Bersac & Bert, 2019, Fig. 5), GRO (ibid., Fig. 9), CHP (ibid., Fig. 10) and PPR (ibid., Fig. 15). See the supplementary appendix for the complete list of the specimens with measurements.
Description:
The shell is discoid to moderately evolute, with whorls covering about 50% of
the flanks. The umbilical wall is steep. The flanks are flat or slightly convex.
The maximum observed diameter is 190 mm (SBC.06061-00001/GRS437, on a poorly
preserved specimen). No fragmentary specimens of potentially larger diameter
seem to be present in the sample (i.e., with a larger H than specimen
SBC.06061-00001/GRS437). R was measured on 96
specimens; it varies from 9 to 30 with an average value of 17.67. Innermost
whorls are never preserved on the studied specimens and the ontogenetic stages A
and B cannot be studied. The observable ontogenetic sequence of the shell flanks
starts with the stage C and is followed by the stage D, which have
characteristics similar to those of the corresponding stages of the Deshayesites
previously described. No specimen of the sample is enough developed to exhibit
the adult stage E. The transition between the stages C and D can be more or less
sudden. We observe a continuous variation between slender specimens with high
rib density, high growth in whorl height and narrow whorl width (Pl. 7 ![]() , figs. H-I) and very robust specimens with very low rib density, low growth in whorl
height and wider whorl width (Pl. 7
, figs. H-I) and very robust specimens with very low rib density, low growth in whorl
height and wider whorl width (Pl. 7 ![]() , figs. R-S).
, figs. R-S).
The
ventral area in the innermost whorls is flat, with a smooth siphonal band and
without latero-ventral
tubercles. These tubercles arise from approximately D=10 mm (Pl. 7 ![]() , figs. V, X-Y). They are very discrete in slender specimens
(Pl. 7
, figs. V, X-Y). They are very discrete in slender specimens
(Pl. 7 ![]() , figs.
T-U) and larger
in the more robust ones (Pl. 6
, figs.
T-U) and larger
in the more robust ones (Pl. 6 ![]() , figs. W1-X1). Their
shape is variable, from conical with oval or rounded base (Pl. 6
, figs. W1-X1). Their
shape is variable, from conical with oval or rounded base (Pl. 6 ![]() , figs. H1-I1),
to claviform (Pl. 6
, figs. H1-I1),
to claviform (Pl. 6 ![]() , figs. S1-T1), this latter shape being less frequent. The
smooth siphonal band is still present between the latero-ventral
tubercles. SSB was measured on 18 specimens, all from beds 100, 102 and 104 of
GRS1 section. It ranges from 16 to 37 mm and its average value is 24.89 mm. At
bed level, SSB
is 24.9 mm on average for Bed 100 (10 specimens), 24.5 mm for Bed 102 (6
specimens) and 26 mm for Bed 104 (2 specimens). There does not appear to be
correlation between the SSB value and the robustness of the specimens. From the
end of the smooth siphonal band, the ribs cross the ventral area between each
pair of tubercles, with more and more relief (Pl. 7
, figs. S1-T1), this latter shape being less frequent. The
smooth siphonal band is still present between the latero-ventral
tubercles. SSB was measured on 18 specimens, all from beds 100, 102 and 104 of
GRS1 section. It ranges from 16 to 37 mm and its average value is 24.89 mm. At
bed level, SSB
is 24.9 mm on average for Bed 100 (10 specimens), 24.5 mm for Bed 102 (6
specimens) and 26 mm for Bed 104 (2 specimens). There does not appear to be
correlation between the SSB value and the robustness of the specimens. From the
end of the smooth siphonal band, the ribs cross the ventral area between each
pair of tubercles, with more and more relief (Pl. 7 ![]() , figs. E-F). Then, the
tubercles vanish and the venter becomes subtabulate without rib attenuation (Pl. 6
, figs. E-F). Then, the
tubercles vanish and the venter becomes subtabulate without rib attenuation (Pl. 6 ![]() , figs. W1-X1). Due to the fragmentary state of the studied material, the end
diameter of the tubercles could have been measured for only two specimens and is
40 and 45 mm (Pl. 7
, figs. W1-X1). Due to the fragmentary state of the studied material, the end
diameter of the tubercles could have been measured for only two specimens and is
40 and 45 mm (Pl. 7 ![]() , figs. C, J). Within the sample, various specimens have
their tubercles present over D=40-50 mm and the few specimens larger than D=80
mm do not exhibit tubercles at this diameter.
, figs. C, J). Within the sample, various specimens have
their tubercles present over D=40-50 mm and the few specimens larger than D=80
mm do not exhibit tubercles at this diameter.
On the few specimens on which it is observable, the septal suture is too poorly preserved to be described. No specimen has its peristome preserved.
Discussion: The range of variability is continuous within this sample and similar to that of the previously studied samples. Such variability has been already observed in different samples of Dufrenoyia (Martin, 2003; Dutour, 2005; Garcia & Moreno-Bedmar, 2010; Lehmann & Bulot, 2020). Consequently, our sample is considered monospecific. It exhibits no evidence of dimorphism, as for the above studied samples. Specimens are abundant in CHP section and from Bed 98 to 105 in GRS1 section. The small value of 24.89 mm of SSB and the mostly rounded to oval latero-ventral tubercles of these specimens are primitive characters in Dufrenoyia (Dutour, 2005; Bersac & Bert, 2012). The end diameter of the latero-ventral tubercles is also a diagnostic character in Dufrenoyia. It has a value of 40 and 45 mm in the only two specimens in which it could have been measured, which is very low (the limit between the primitive species D. furcata and its derived D. dufrenoyi is set at 70 mm for this character according to Bersac and Bert, 2012). On the basis of these elements, we interpret these specimens from CHP and from the interval between the beds 98 and 105 of GRS1 as belonging to the primitive species D. furcata. It implies that some very slender specimens, of which the characters are similar to the taxon Dufrenoyia transitoria Casey, 1961, are here assigned to D. furcata. This is in line with a synonymy between D. transitoria and D. furcata (see discussion about the synonymy between these species in Dutour, 2005; Bersac & Bert, 2012; Lehmann & Bulot, 2020).
The average value of 24.89 mm for SSB in the herein studied sample is very close and consistent with the lowest average value of 25 mm retained for D. furcata according to Bersac and Bert (2012, Fig. 18).
The absence of the adult stage E within the sample is due to the immaturity/fragmentary state of the specimens. Delanoy et al. (2018, Fig. 20E) figured a D=approx. 209mm specimen from their bed 110 of GRS1 section (here Bed 98 in the revised lithology; see Bersac & Bert, 2019, for an explanation). This specimen is the largest Dufrenoyia found in the LFAB so far and exhibits the beginning of the adult stage E (the adult size of a Dufrenoyia seems to range from 250 to 420 mm, see Casey, 1964, 1980; Bersac & Bert, 2012, p. 233).
Biostratigraphic distribution: Dufrenoyia furcata Subzone, Dufrenoyia furcata Zone.
Dufrenoyia dufrenoyi (Orbigny, 1841)
Pl. 7 ![]() ,
figs. A1-B1
,
figs. A1-B1
1841 Ammonites dufrenoyi sp. nov. - Orbigny, Pl. 33, figs. 4-6.
2005 Dufrenoyia dufrenoyi - Dutour, Pl. 29, figs. 10-16
v 2019 Dufrenoyia dufrenoyi - Bersac & Bert, p. 154, 165-166, Fig. 5, Pl. 5, figs. 30-32.
Material studied: 20 specimens from the beds 110 (14 specimens) and 113 (6 specimens) of GRS1 GRS1 (Bersac & Bert, 2019, Fig. 5). See the supplementary appendix for the complete list of the specimens with measurements.
Description: The specimens are
fragmentary and deformed. The largest specimen is estimated to be 37 mm in
diameter (SBC.06061-00001/GRS821, Pl. 7 ![]() , fig.
B1). R varies from 15 to 24. The size of the specimens is too small to
differentiate the stage C from the stage D, and to express the adult stage E.
All the specimens on which the ventral area is preserved exhibit a smooth
siphonal band on the whole shell. Consequently, SSB could not be measured on any
of them and it is obviously higher than 37 mm. The latero-ventral tubercles are
all claviform. The septal suture is never preserved.
, fig.
B1). R varies from 15 to 24. The size of the specimens is too small to
differentiate the stage C from the stage D, and to express the adult stage E.
All the specimens on which the ventral area is preserved exhibit a smooth
siphonal band on the whole shell. Consequently, SSB could not be measured on any
of them and it is obviously higher than 37 mm. The latero-ventral tubercles are
all claviform. The septal suture is never preserved.
Discussion: Despite its poor preservation,
the specimens of this sample have a variability included within the same scheme
of the previously studied samples. We consequently regard these specimens as
being conspecific until proven otherwise. Various elements suggest that this
sample of Dufrenoyia presents derived characters: (1) the smooth siphonal
band is present on the entirety of the preserved part of the shell of all the
specimens, including the two largest ones (SBC.06061-00001/GRS819
with D=34mm and SBC.06061-00001/GRS821, Pl. 7 ![]() , fig. B1
with D= approx. 37 mm), suggesting a high mean value of SSB in the sample
and (2) the latero-ventral tubercles are always claviform. This latter element
is characteristic of the species D. dufrenoyi according to Dutour
(2005), to which we therefore assign the sample. The range of variability of our
sample is narrower than that of D. furcata, which is probably due to its
smaller size.
, fig. B1
with D= approx. 37 mm), suggesting a high mean value of SSB in the sample
and (2) the latero-ventral tubercles are always claviform. This latter element
is characteristic of the species D. dufrenoyi according to Dutour
(2005), to which we therefore assign the sample. The range of variability of our
sample is narrower than that of D. furcata, which is probably due to its
smaller size.
Biostratigraphic distribution: Dufrenoyia dufrenoyi Subzone, Dufrenoyia furcata Zone.
In the present work, we studied a large sample of Deshayesitidae from a relatively wide stratigraphic interval of the LFAB. Despite the deformed and fragmentary state of most of the specimens, we confidently identified several biostratigraphically significant species and their corresponding biostratigraphy at subzonal level in most of the cases.
In the strata older than the D. grandis Subzone, the identification criteria used in the present work are restricted to the inner whorls only (septal suture is absent or too poorly preserved to be studied). If the samples from these levels are poorly preserved, their identification becomes particularly difficult. This is the case for the sample of the top of the Hauterivian-Aptian limestones of the LFAB, that we could not identify at species level (Deshayesites sp.). The age of its original bed was estimated by the co-occurring ammonite fauna. In the Ammonitoceras Level of the LFAB, no septal suture could be studied, but only 4 specimens, of the 71 collected have their inner whorls well enough preserved to be studied, allowing us to consider the sample to belong to the species D. multicostatus. In this case, the co-occurring ammonite fauna only helps to confirm the age and thus the identification of the sample (since D. multicostatus is a subzonal index).
In the more recent subzones, the taxonomic identification of the Deshayesitidae is generally easier because, due to their neotenic evolution, the identification characters occupy a larger part of the shell and thus are more visible and susceptible to be preserved (longer smooth siphonal band, subtabulate ventral area or latero-ventral tubercles on the phragmocone).
All these elements point out the necessity of identifying additional biostratigraphically significant ammonite taxa for the lower Aptian (as it is already the case with the Roloboceratinae), especially for levels below the D. grandis Subzone.
We warmly thank Mikel L�pez�Horgue (University of the Basque Country, Leioa), Antoine Pictet (Mus�um cantonal des sciences naturelles, Lausanne) and an anonymous reviewer for their thorough reviews that improved the manuscript. We are grateful to Steven Eagar for a linguistic correction of the manuscript.
Baraboshkin E.J. (2004).- The Lower Cretaceous ammonite zonal standard of the Boreal Realm.- Bulletin of Moscow Society of Naturalists, t. 79, vol. 5, p. 44-68 [in Russian].
Barrag�n R. & Szives O. (2007).- New records of Mathoceras Casey (Deshayesitidae, Ammonoidea) from the Aptian (Lower Cretaceous) of Mexico and Hungary: Biostratigraphic and paleobiogeographic implications.- Geobios, Villeurbanne, vol. 40, p. 21-30.
Bersac S. & Bert D. (2012).- Ontogenesis, variability and evolution of the Lower Greensand Deshayesitidae (Ammonoidea, Lower Cretaceous, Southern England): Reinterpretation of literature data; taxonomic and biostratigraphic implications.- Annales du Mus�um d'Histoire Naturelle de Nice, Nice, vol. 27, p. 197-270.
Bersac S. & Bert D. (2015).- Two ammonite species under the same name: Revision of Deshayesites deshayesi (d'Orbigny, 1841) based on topotype material (Lower Aptian, Lower Cretaceous, Northeast of France).- Annales de Pal�ontologie, Paris, vol. 101, p. 265-294.
Bersac S. & Bert D. (2018).- Revision of the lower Aptian (Lower Cretaceous) ammonite species Cheloniceras cornuelianum (d'Orbigny, 1841).- Annales de Pal�ontologie, Paris, vol. 104, p. 45-70.
Bersac S. & Bert D. (2019).- The lower Aptian ammonites of the Les Ferres Aptian Basin (Lower Cretaceous, Southeast of France). Part I: Introduction and biostratigraphy.- Carnets Geol., Madrid, vol. 19, p. 149-183. DOI: 10.4267/2042/70292
Bersac S., Bert D. & Matrion, B. (2012).- Revision of the index-species Deshayesites deshayesi (Ammonoidea, lower Aptian, Lower Cretaceous): Taxonomic and biostratigraphic consequences. In: Bert D. & Bersac S. (eds), First meeting of the Research Group for Paleobiology and Biostratigraphy of the Ammonites.- Boletin del Instituto de Fisiografia y Geologia, Rosario, vol. 82, p. 31-33.
Bert D. (2013).- Factors of intraspecific variability in ammonites, the example of Gassendiceras alpinum (d'Orbigny, 1850) (Hemihoplitidae, Upper Barremian).- Annales de Pal�ontologie, Paris, vol. 100, no. 3, p. 217-236
Bert D. (2019).- Les lois de la variabilit� intrasp�cifique chez les ammonites.- Les Cahiers de la R�serve Naturelle de Sainte-Victoire, Aix-en-Provence, num�ro sp�cial congr�s APF 2019, p. 12.
Bert D. & Bersac S. (2011).- Buts et m�thodes du "lev� de coupe".- Fossiles, Saint-Julien-du-Pinet, vol. 6, p. 49-61.
Bert D., Bersac S., Canut L & Beltrand B. (2023).- Can the intraspecific laws of variation be used as a predictive model for understanding species with insufficient data? The example of the early Heteroceratidae in their evolutionary context (heteromorph ammonites, Tethys upper Barremian).- Cretaceous Research, vol. 151, 33 p.
Bogdanova T.N. & Mikhailova I.A. (2004).- Origin, evolution and stratigraphic significance of the superfamily Deshayesitaceae Stoyanow, 1949.- Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre , Brussels, vol. 74, p. 189-243.
Casey R. (1964).- A monograph of the Ammonoidea of the Lower Greensand part. V.- Palaeontographical Society (1963), London, p. 289-398.
Casey R. (1980).- A monograph of the Ammonoidea of the Lower Greensand, part IX.- Palaeontographical Society, London, p. 633-660.
Casey R., Bayliss H.M. & Simpson M.I. (1998).- Observations on the lithostratigraphy and ammonite succession of the Aptian (Lower Cretaceous) Lower Greensand of Chale Bay, Isle of Wight, UK.- Cretaceous Research, vol. 19, p. 511-535.
Cotillon P. (1971).- Le Cr�tac� inf�rieur de l'Arc subalpin de Castellane entre l'Asse et le Var. Stratigraphie et s�dimentologie.- M�moires du Bureau de Recherche G�ologique et Mini�re, Paris, no. 68, 313 p.
De Baets K., Bert D., Hoffmann R., Monnet C., Yacobucci M. & Klug C. (2015).- Ammonoid intraspecific variability. In: Klug C. et al. (eds.), Ammonoid paleobiology: From anatomy to ecology.- Topics in Geobiology, New-York - NY, vol. 43, p. 359-426.
Delanoy G., Baudouin C., Pictet A., Delattre P. & P�nag� E. (2018).- Ancyloceratidae (Ammonoidea, Ancyloceratoidea) nouveaux ou peu connus de l'Aptien inf�rieur des Alpes-Maritimes (Sud-Est de la France).- Annales du Mus�um d'Histoire Naturelle de Nice, vol. 32, p. 35-90.
Delanoy G., Baudouin C., Pictet A., Moreno-Bedmar J., Frau C. & Matrion B. (2022).- The genera Roloboceras Casey, 1954, and Megatyloceras Humphrey, 1949 (Ammonoidea, Ancyloceratina, Douvilleiceratidae), from the Lower Aptian of Ard�che (SE France) - Taxonomic and biostratigraphic implications.- Carnets Geol., Madrid, vol. 22, no. 2, p. 7-109. DOI: 10.2110/carnets.2022.2202
Dutour Y. (2005, unpublished).- Biostratigraphie, �volution et renouvellements des ammonites de l'Aptien sup�rieur (Gargasien) du bassin vocontien (Sud-Est de la France).- PhD thesis, Universit� Claude Bernard, Lyon 1, 280 p.
Dzik J. (1985).- Typologic versus population concept of chronospecies: Implication for ammonite biostratigraphy.- Acta Paleontologica Polonica, Warsaw, vol. 30, no. 1-2, p. 71-92.
Etayo-Serna F. (1979).- Zonation of the Cretaceous of Central Colombia by ammonites.- Publicaciones geologicas especiales del Ingeominas, Bogot�, no. 2, p. 1-186.
Fitton W.H. (1836).- Observations on some of the strata between the Chalk and the Oxford Oolithe in the South-east England. Including: Appendix A, descriptive notes respecting the shells figured in Pls. XI-XXIII by J. de C. Sowerby.- Transactions of the Geological Society, London, vol. IV, no. 2, p. 103-390.
Frau C. (2020).- Stable carbon-isotope chemostratigraphy versus ammonite biostratigraphy: Data from around the Barremian/Aptian boundary (Lower Cretaceous).- STRATA, Gaillac, s�rie 2e, vol. 56, p. 1-31.
Frau C. & Delanoy G. (2022).- Taxonomic notes on some Barremian-Aptian cephalopods from the Station de Cassis section and surrounding sites, Bouches-du-Rh�ne, southern France.- STRATA, Gaillac, s�rie 2e, vol. 58, p. 1-45.
Frau C., Delanoy G. & Baudouin C. (2023).- A new insight into the lower Aptian Roloboceratinae Casey, 1961 (Douvilleiceratidae, Ammonoidea) from southern France.- STRATA, Gaillac, s�rie 2e, vol. 59, p. 1-21.
Frau C., Pictet A., Spangenberg J.E., Masse J.-P., Tendil A.J.-B. & Lanteaume C. (2017).- New insights on the age of the post-Urgonian marly cover of the Apt region (Vaucluse, SE France) and its implications on the demise of the North Provence carbonate platform.- Sedimentary Geology, vol. 359, p. 44-61.
Garcia R. & Moreno-Bedmar J.A. (2010).- Dufrenoyia furcata (Sowerby, 1836), ammonite del Aptiense inferior (Cretacico inferior) del Perello (Tarragona). In: Moreno-Azanza M., Diaz-Martinez I, Gasca J.M., Melero-Rubio M., Rabal-Garc�s R. & Sauqu�, V. (eds.), VIII Encuentro de Jovenes Investigadores en Paleontologia volumen de actas.- Cidaris, Elche, no. 30, p. 129-137.
Garc�a-Mond�jar J., Owen H. G., Raisossadat N., Mill�n M. I. & Fern�ndez-Mendiola P. A. (2009).- The early Aptian of Aralar (northern Spain): Stratigraphy, sedimentology, ammonite biozonation and OAE1.- Cretaceous Research, vol. 30, p. 434-464.
Granier B. (2022).- Tethysian, Tethyan or ... Tethys Ocean and Tethys.- Carnets Geol., Madrid, vol. 22, Editorial Note 1, p. 681-683. DOI: 10.2110/carnets.2022.22EN1
Lehmann J. & Bulot L.G. (2020).- A Dufrenoyia (Ammonoidea, Deshayesitidae) fauna from Germany: Evidence for D. furcata Zone deposits at the lower-upper Aptian boundary.- Cretaceous Research, vol. 116, article 104546, 14 p.
Lehmann J., Bargen D. von, Engelke J. & Cla�en J. (2015).- Morphological variability in response to palaeoenvironmental change � a case study on Cretaceous ammonites.- Lethaia, Copenhagen, vol. 49, no. 1 (2016), p. 73-86
Martin E. (2003).- Intraspecific variation of Dufrenoyia furcata Sowerby (Ammonoidea, Ancylocerataceae, Deshayesitidae) from the Gargasian in the Vaucluse (Southern France).- Mitteilungen aus dem Geologisch-Pal�ontologischen Institut der Universit�t Hamburg, Hamburg, no. 87, p. 115-124.
Moreno J.A. & Company M. (2007).- Caracterizacion de dimorfismo morfodimensional en Deshayesites deshayesi (Leymerie in d'Orbigny) (Ancyloceratina, Ammonoidea).- XXIII Jornadas de la Sociedad Espan�la de Paleontolog�a, Libro de Res�menes, Caravaca de la Cruz (Murcia), p. 153-154.
Moreno-Bedmar J.A., Barragan R., Delanoy G., Company M. & Salas R. (2014).- Review of the early Aptian (Early Cretaceous) ammonoid species Deshayesites deshayesi (d'Orbigny, 1841).- Cretaceous Research, vol. 51, p. 341�360.
Moreno-Bedmar J.A., Casados-Monroy A., Frau C., Pictet A., Chavez Cabello G. & Esquivel-Mac�as C. (2018).- Ammonite biostratigraphy of the Sierra del Patron section (Durango State, Mexico) and its bearing on the lower/upper Aptian boundary of the Central Atlantic Province.- Cretaceous Research, vol. 88, p. 100-110.
Moreno-Bedmar J.A., Company M., Bover-Arnal T., Salas R., Delanoy G., Mart�nez R. & Grauges A. (2009).- Biostratigraphic characterization by means of ammonoids of the lower Aptian Oceanic Anoxic Event (OAE 1a) in the eastern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, vol. 30, p. 864-872.
Moreno-Bedmar J.A., Company M., Bover-Arnal T., Salas R., Delanoy G., Maurrasse F.J., Grauges A. & Mart�nez R. (2010).- Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence.- Geologica Acta, Barcelona, vol. 8, no. 3, p. 281-299.
Moreno-Bedmar J.A., Company M. & Guerrero-Suastegui M. (2023).- Un ammonite del g�nero Juandurhamiceras del Aptiano de Campo Morado, Arcelia, estado de Guerrero, M�xico.- Paleontolog�a Mexicana, Mexico, vol. 12, no. 1, p. 1-8.
Moreno-Bedmar J.A., Company M., Sandoval J., Tavera J.M., Bover-Arnal T., Salas R., Delanoy G., Maurrasse F.J.-M.R. & Martinez R. (2012).- Lower Aptian ammonite and carbon isotope stratigraphy in the eastern Prebetic Domain (Betic Cordillera, southeastern Spain).- Geologica Acta, Barcelona, vol. 10, no. 4, p. 333-350.
Obata I. & Matsukawa M. (2007).- Barremian-Aptian (Early Cretaceous) ammonoids from the Choshi Group, Honshu (Japan).- Cretaceous Research, vol. 28, p. 363�391.
Orbigny A. d' (1841).- Pal�ontologie Fran�aise. Tome premier. Terrains cr�tac�s. C�phalopodes.- Imprimerie Masson, Paris, p. 121-430.
Ragazzi M. (1982, unpublished).- L'Aptien dans les Alpes Maritimes: Stratigraphie, pal�o�cologie, s�dimentologie.- PhD thesis, Universit� de Nice, 224 p.
Reboulet S., Szives O., Aguirre-Urreta M.B., Barragan R., Company M., Frau C., Kakabadze M.V., Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S. N., Va��ček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the "Kilian Group" (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Renz O. (1982).- The Cretaceous ammonites of Venezuela.- Maraven Editor, Caracas, 132 p.
Rogov M.A. & Mironenko A.A. (2016).- Ammonite shell size changes in response of the Early Aptian anoxic event (preliminary data). In: Baraboshkin E.Y. (ed.), Cretaceous system of Russia and CIS countries: Problems of stratigraphy and paleogeography.- Proceeding of the 8th all-Russian meeting (26 September-3 October 2016, Republic of Crimea, Russian Federation).- ChernomorPRESS, Simferopol (Ukraine), p. 236�238.
Ropolo P., Conte G., Gonnet R., Masse J.-P. & Moullade M. (1998).- Les faunes d'ammonites du Barr�mien sup�rieur/Aptien inf�rieur (B�doulien) dans la r�gion stratotypique de Cassis-La B�doule (SE France) : �tat des connaissances et propositions pour une zonation par Ammonites du B�doulien-type.- G�ologie M�diterran�enne, Marseille, t. XXV, no. 3-4, p. 167-175.
Ropolo P., Moullade M., Gonnet R., Conte G. & Tronchetti G. (2006).- The Deshayesitidae Stoyanov, 1949 (Ammonoidea) of the Aptian historical stratotype region at Cassis-La B�doule (SE France).- Carnets Geol., Madrid, Memoir 2006/01 (CG2006_M01), 46 p. DOI: 10.4267/2042/4744
Ropolo P., Moullade M., Conte G. & Tronchetti G. (2008).- About the stratigraphic position of the Lower Aptian Roloboceras hambrovi (Ammonoidea) level.- Carnets Geol., Madrid, Letter 2008/03 (CG2008 L03), 7 p. DOI: 10.4267/2042/18124
Spath L.F. (1930).- On some Ammonoidea from the Lower Greensand.- Annals and Magazine of Natural History (serie 10), London, no. 5, p. 417-464.
Swinnerton H. (1935).- The rocks below the Red Chalk of Lincolnshire and their cephalopod fauna. Quaternary Journal of the Geological Society, London, vol. XCI, 446 p.
Szives O., Moreno�Bedmar J.A., Aguirre�Urreta B., Company M., Frau C., L�pez�Horgue M., Pictet A., Ploch I., Salazar C., Barrag�n R., Latil J.�L., Lehmann J., Robert E & Reboulet S. (2024).- Report on the 7th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Warsaw, Poland, 21st August 2022): State of the art on the current Standard Ammonite Zonation of the Western Tethyan Mediterranean Province.- Cretaceous Research, vol. 153, 14 p.
Tintant H. (1963).- Les Kosmoceratid�s du Callovien inf�rieur et moyen d'Europe occidentale. Essai de pal�ontologie quantitative.- Publications de l'Universit� de Dijon, vol. XXIX, Les Presses Universitaires de France, Paris, 491 p.
Young K. (1974).- Lower Albian and Aptian (Cretaceous) ammonites of Texas.- Geoscience and Man, Baton Rouge - LA, vol. 8, p. 175-228.
Plate
1:
Deshayesites sp. from the last
beds of the Hauterivian-Aptian limestones and of the Combe de Joinet Member (Deshayesites
forbesi Zone) of CRS section. A:
SBC.06061-00008/CRS017, Bed 399. B: SBC.06061-00008/CRS016, Bed 403. C-D:
SBC.06061-00008/CRS019, Bed 400. E: SBC.06061-00008/CRS021, Bed 400. |
|
Plate
2:
Deshayesites multicostatus from the Ammonitoceras Level (top of
the Pont de la Cerise Member, Dehayesites
deshayesi Zone, Deshayesites
multicostatus Subzone) of CHP section's Bed 415. A: SBC.06061-00006/CHP368.
B-C: SBC.06061-00006/CHP373. |
|
Plate
3:
Deshayesites multicostatus from the Ammonitoceras Level (top of
the Pont de la Cerise Member, Dehayesites
deshayesi Zone, Deshayesites
multicostatus Subzone) of CHP section's Bed 415. A-B: SBC.06061-00006/CHP371. C: SBC.06061-00006/CHP292. |
|
Plate
4:
Deshayesites multicostatus from the Ammonitoceras Level (top of
the Pont de la Cerise Member, Deshayesites
deshayesi Zone, Deshayesites
multicostatus Subzone). A-B: SBC.06061-00006/CHP369, CHP section, Bed 415.
C: SBC.06061-00003/VAL037, VAL section, Bed 405. D-E: SBC.06061-00003/VAL040,
VAL section, Bed 405. F-G: SBC.06061-00003/VAL009, VAL section, Bed 405. H-I:
SBC.06061-00003/VAL020, VAL section, Bed 405. J-L: SBC.06061-00003/VAL039,
VAL section, Bed 405. M-N: SBC.06061-00003/VAL018, VAL section, Bed 405. O-P:
SBC.06061-00003/VAL012, VAL section, Bed 405. Q-R: SBC.06061-00003/VAL017, VAL
section, Bed 405. S-T: SBC.06061-00009/PPR004,
PPR section, top of Bed 414. U-V: SBC.06061-00001/GRS115, GRS1 section, top of
Bed 96. |
|
Plate
5:
Black dot: last septal suture. A-H: Deshayesites
multicostatus from the Ammonitoceras Level (top of the Pont de la
Cerise Member, Dehayesites deshayesi Zone,
Deshayesites multicostatus Subzone) of
CHP section's Bed 415. A-B:
SBC.06061-00006/CHP167. C: SBC.06061-00006/CHP294. D: SBC.06061-00006/CHP382.
E-F: SBC.06061-00006/CHP385. G-H: SBC.06061-00006/CHP390. I-N: Deshayesites
grandis from the Toxoceratoides Bed (base of Les Graous Member, Dehayesites
deshayesi Zone, Deshayesites grandis Subzone)
of CHP section's Bed 416. I-J:
SBC.06061-00006/CHP391. K-L: SBC.06061-00006/CHP395. M-N:
SBC.06061-00006/CHP392. |
|
Plate
6:
Black dot: last septal suture. A-G: Deshayesites
grandis from the Toxoceratoides Bed (base of Les Graous Member, Dehayesites
deshayesi Zone, Deshayesites grandis Subzone). A-B: SBC.06061-00001/GRS351, GRS1 section, Bed 97.
C: SBC.06061-00001/GRS352,
GRS1 section, Bed 97. D-E: SBC.06061-00001/GRS353,
GRS1 section, Bed 97. F-G: SBC.06061-00006/CHP118, CHP section, Bed 416. H-X1: Dufrenoyia
furcata from the Les Graous Member (Dufrenoyia furcata Zone, Dufrenoyia
furcata Subzone). H-O: specimens from CHP section. H-I:
SBC.06061-00006/CHP200, Bed 417. J- K: SBC.06061-00006/CHP163, Bed 418. L:
SBC.06061-00006/CHP178, Bed 419. M: SBC.06061-00006/CHP198, Bed 421. N-O:
SBC.06061-00006/CHP218, Bed 426. P: SBC.06061-00005/GRO075, Bed 418, GRO
section. Q-X1: specimens from GRS1 section. Q: SBC.06061-00001/GRS077, Bed 98.
R-S: SBC.06061-00001/GRS375, Bed 99. T: SBC.06061-00001/GRS426, Bed 99.
U-X1:
specimens from Bed 100. U: SBC.06061-00001/GRS084. V-W: SBC.06061-00001/GRS085.
X-Y: SBC.06061-00001/GRS089. A1-B1: SBC.06061-00001/GRS156. C1-D1:
SBC.06061-00001/GRS155. E1-F1: SBC.06061-00001/GRS170. G1:
SBC.06061-00001/GRS168. H1-I1: SBC.06061-00001/GRS205. J1-K1:
SBC.06061-00001/GRS305. L1-M1: SBC.06061-00001/GRS308. N1:
SBC.06061-00001/GRS090. O1-P1: SBC.06061-00001/GRS650. Q1-R1:
SBC.06061-00001/GRS738. S1-T1: SBC.06061-00001/GRS611. U1-V1:
SBC.06061-00001/GRS825. W1-X1: SBC.06061-00001/GRS361. |
|
Plate
7:
A: Dufrenoyia from GRS1 section (Les Graous Member, Dufrenoyia furcata
Zone). Black dot: last septal suture. A-Z: Dufrenoyia
furcata (Dufrenoyia furcata Zone, Dufrenoyia furcata
Subzone).
A: SBC.06061-00001/GRS668, Bed 100. B: SBC.06061-00001/GRS740, Bed 101. C:
SBC.06061-00001/GRS744, Bed 101. D-S: specimens from Bed 102. D:
SBC.06061-00001/GRS048 (in association with Colombiceras crassicostatum).
E-F: SBC.06061-00001/GRS279. G: SBC.06061-00001/GRS272. H-I: SBC.06061-00001/GRS632.
J:
SBC.06061-00001/GRS227. K: SBC.06061-00001/GRS499. L-M: SBC.06061-00001/GRS556.
N: SBC.06061-00001/GRS009. O-P: SBC.06061-00001/GRS275. Q:
SBC.06061-00001/GRS767. R-S: SBC.06061-00001/GRS665. T-Y: specimens from Bed
104. T, U: SBC.06061-00001/GRS525. V: SBC.06061-00001/GRS486. W:
SBC.06061-00001/GRS521. X: SBC.06061-00001/GRS402. Y: SBC.06061-00001/GRS536.
Z:
SBC.06061-00001/GRS494, Bed 106. A1-B1: Dufrenoyia dufrenoyi (Dufrenoyia
furcata Zone, Dufrenoyia dufrenoyi Subzone) from Bed 110. A1:
SBC.06061-00001/GRS751. B1: SBC.06061-00001/GRS821. |
|
Total number of studied specimens: 335.
All specimens: Bersac's collection, deposited in the R�serve naturelle nationale g�ologique de Haute-Provence (RNNGHP, France).
Tables caption: N = number of specimens, D: diameter, H: whorl height, R: last half whorl rib density, SSB: end diameter of the smooth siphonal band, Subtab.: end diameter of the subtabulate ventral area, Tuberc.: end diameter of the latero ventral tubercles.
1. Deshayesites sp.
|
Identification number (N = 12) |
Section |
Bed |
D |
H |
R |
Subtab. |
SSB |
Figuration (N = 5) |
|
SBC.06061-00001/GRS072 |
GRS1 |
96 |
? |
63 |
? |
No |
No |
|
|
SBC.06061-00001/GRS115 |
GRS1 |
96 |
32 |
15 |
21 |
No |
No |
Pl. 4 |
|
SBC.06061-00001/GRS773 |
GRS1 |
95 |
? |
? |
? |
No |
No |
|
|
SBC.06061-00007/CLE029 |
CLE |
400 |
? |
? |
? |
No |
No |
|
|
SBC.06061-00007/CLE030 |
CLE |
400 |
48 |
22 |
27 |
No |
No |
Bersac & Bert, 2019, Pl. 1, figs. 4-5 |
|
SBC.06061-00007/CLE052 |
CLE |
400 |
? |
? |
? |
No |
No |
|
|
SBC.06061-00008/CRS016 |
CRS |
403 |
? |
? |
? |
No |
No |
Pl. 1 |
|
SBC.06061-00008/CRS017 |
CRS |
399 |
133 |
? |
? |
No |
No |
Pl. 1 |
|
SBC.06061-00008/CRS019 |
CRS |
400 |
? |
55 |
? |
No |
No |
Pl. 1 |
|
SBC.06061-00008/CRS020 |
CRS |
400 |
57 |
26 |
? |
No |
No |
|
|
SBC.06061-00008/CRS021 |
CRS |
400 |
? |
52 |
? |
No |
No |
Pl. 1 |
|
SBC.06061-00008/CRS030 |
CRS |
400 |
? |
? |
? |
No |
No |
|
2. Deshayesites multicostatus Swinnerton, 1935
|
Identification number (N = 71) |
Section |
Bed |
D |
H |
R |
Subtab. |
SSB |
Figuration (N = 22) |
|
SBC.06061-00003/VAL008 |
VAL |
405 |
22 |
8.5 |
? |
No |
? |
|
|
SBC.06061-00003/VAL009 |
VAL |
405 |
? |
? |
? |
Yes |
? |
Pl. 4 |
|
SBC.06061-00003/VAL012 |
VAL |
405 |
22 |
8.9 |
26 |
No |
19 |
Pl. 4 |
|
SBC.06061-00003/VAL013 |
VAL |
405 |
21 |
10.5 |
22 |
No |
<20 |
|
|
SBC.06061-00003/VAL014 |
VAL |
405 |
? |
9 |
? |
No |
? |
|
|
SBC.06061-00003/VAL015 |
VAL |
405 |
? |
9 |
? |
No |
? |
|
|
SBC.06061-00003/VAL017 |
VAL |
405 |
24 |
10 |
23 |
No |
14 |
Pl. 4 |
|
SBC.06061-00003/VAL018 |
VAL |
405 |
19 |
9.5 |
26 |
No |
? |
Pl. 4 |
|
SBC.06061-00003/VAL019 |
VAL |
405 |
18 |
8.5 |
? |
No |
16 |
|
|
SBC.06061-00003/VAL020 |
VAL |
405 |
26.5 |
11.8 |
26 |
No |
<15 |
Pl. 4 |
|
SBC.06061-00003/VAL025 |
VAL |
405 |
? |
11.5 |
? |
No |
? |
|
|
SBC.06061-00003/VAL026 |
VAL |
405 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00003/VAL027 |
VAL |
405 |
? |
12 |
? |
No |
? |
|
|
SBC.06061-00003/VAL028 |
VAL |
405 |
28 |
10 |
25 |
No |
<16 |
|
|
SBC.06061-00003/VAL029 |
VAL |
405 |
? |
8 |
? |
No |
? |
|
|
SBC.06061-00003/VAL030 |
VAL |
405 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00003/VAL031 |
VAL |
405 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00003/VAL032 |
VAL |
405 |
? |
14 |
? |
No |
? |
|
|
SBC.06061-00003/VAL033 |
VAL |
405 |
20 |
9.2 |
21 |
No |
17 |
|
|
SBC.06061-00003/VAL034 |
VAL |
405 |
28 |
13 |
22 |
No |
<19 |
|
|
SBC.06061-00003/VAL035 |
VAL |
405 |
? |
8 |
? |
No |
? |
|
|
SBC.06061-00003/VAL036 |
VAL |
405 |
25 |
13 |
? |
No |
? |
|
|
SBC.06061-00003/VAL037 |
VAL |
405 |
27 |
12 |
? |
No |
? |
Pl. 4 |
|
SBC.06061-00003/VAL038 |
VAL |
405 |
? |
14 |
? |
No |
? |
|
|
SBC.06061-00003/VAL039 |
VAL |
405 |
45 |
20 |
? |
No |
<27 |
Pl. 4 |
|
SBC.06061-00003/VAL040 |
VAL |
405 |
52 |
26 |
29 |
No |
? |
Pl. 4 |
|
SBC.06061-00003/VAL041 |
VAL |
405 |
36 |
16 |
21 |
No |
<18 |
|
|
SBC.06061-00003/VAL042 |
VAL |
405 |
13.5 |
|
28 |
No |
>13.5 |
|
|
SBC.06061-00003/VAL050 |
VAL |
405 |
? |
6.5 |
? |
No |
? |
|
|
SBC.06061-00003/VAL064 |
VAL |
405 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP072 |
CHP |
415 |
25 |
9 |
? |
No |
? |
Bersac & Bert, 2019, Pl. 2, fig. 19 |
|
SBC.06061-00006/CHP157 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP167 |
CHP |
415 |
142 |
54 |
36 |
No |
? |
Pl. 5 |
|
SBC.06061-00006/CHP168 |
CHP |
415 |
125 |
65 |
31 |
No |
? |
Bersac & Bert, 2019, Pl. 2, figs. 6-8 |
|
SBC.06061-00006/CHP241 |
CHP |
415 |
273 |
103 |
19 |
No |
? |
Bersac & Bert, 2019, Pl. 2, figs. 13-15 |
|
SBC.06061-00006/CHP255 |
CHP |
415 |
? |
65 |
? |
No |
? |
|
|
SBC.06061-00006/CHP266 |
CHP |
415 |
215 |
? |
22 |
No |
? |
|
|
SBC.06061-00006/CHP290 |
CHP |
415 |
? |
55 |
? |
No |
? |
|
|
SBC.06061-00006/CHP292 |
CHP |
415 |
70 |
34 |
33 |
No |
? |
Pl. 3 |
|
SBC.06061-00006/CHP293 |
CHP |
415 |
? |
70 |
? |
No |
? |
|
|
SBC.06061-00006/CHP294 |
CHP |
415 |
? |
82 |
? |
No |
? |
Pl. 5 |
|
SBC.06061-00006/CHP295 |
CHP |
415 |
62 |
22 |
? |
No |
? |
|
|
SBC.06061-00006/CHP352 |
CHP |
415 |
? |
14 |
? |
No |
? |
|
|
SBC.06061-00006/CHP368 |
CHP |
415 |
248 |
85 |
? |
No |
? |
Pl. 2 |
|
SBC.06061-00006/CHP369 |
CHP |
415 |
175 |
69 |
28 |
No |
? |
Pl. 4 |
|
SBC.06061-00006/CHP370 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP371 |
CHP |
415 |
267 |
95 |
17 |
No |
? |
Pl. 3 |
|
SBC.06061-00006/CHP372 |
CHP |
415 |
190 |
75 |
23 |
No |
? |
|
|
SBC.06061-00006/CHP373 |
CHP |
415 |
? |
? |
? |
? |
? |
Pl. 2 |
|
SBC.06061-00006/CHP374 |
CHP |
415 |
24 |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP375 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP376 |
CHP |
415 |
? |
88 |
? |
No |
? |
|
|
SBC.06061-00006/CHP377 |
CHP |
415 |
155 |
55 |
31 |
No |
|
|
|
SBC.06061-00006/CHP378 |
CHP |
415 |
? |
63 |
? |
No |
? |
|
|
SBC.06061-00006/CHP379 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP380 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP381 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP382 |
CHP |
415 |
31 |
12 |
25 |
No |
? |
Pl. 5 |
|
SBC.06061-00006/CHP383 |
CHP |
415 |
? |
19 |
? |
No |
? |
|
|
SBC.06061-00006/CHP384 |
CHP |
415 |
20 |
12 |
? |
No |
? |
|
|
SBC.06061-00006/CHP385 |
CHP |
415 |
18 |
8.5 |
22 |
No |
? |
Pl. 5 |
|
SBC.06061-00006/CHP386 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP386 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP387 |
CHP |
415 |
27 |
15 |
26 |
No |
? |
|
|
SBC.06061-00006/CHP388 |
CHP |
415 |
32 |
? |
21 |
No |
? |
|
|
SBC.06061-00006/CHP389 |
CHP |
415 |
? |
11 |
? |
No |
? |
|
|
SBC.06061-00006/CHP390 |
CHP |
415 |
25 |
10 |
22 |
No |
< 18 |
Pl. 5 |
|
SBC.06061-00006/CHP394 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP396 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00006/CHP412 |
CHP |
415 |
? |
? |
? |
No |
? |
|
|
SBC.06061-00009/PPR004 |
PPR |
415 |
22 |
8.5 |
? |
No |
<12 |
Pl. 4 |
3. Deshayesites grandis Spath, 1930
|
Identification number (N = 22) |
Section |
Bed |
D |
H |
R |
Subtab. |
SSB |
Figuration (N = 9) |
|
SBC.06061-00001/GRS350 |
GRS1 |
97 |
28 |
13 |
18 |
Yes |
20 |
Bersac & Bert, 2019, Pl. 3, figs. 9-10 |
|
SBC.06061-00001/GRS351 |
GRS1 |
97 |
49 |
23 |
20 |
Yes |
24 |
Pl. 6 |
|
SBC.06061-00001/GRS352 |
GRS1 |
97 |
? |
18 |
? |
Yes |
? |
Pl. 6 |
|
SBC.06061-00001/GRS353 |
GRS1 |
97 |
27.5 |
14 |
24 |
Yes |
20 |
Pl. 6 |
|
SBC.06061-00001/GRS354 |
GRS1 |
97 |
20 |
10 |
? |
? |
>20 |
|
|
SBC.06061-00001/GRS355 |
GRS1 |
97 |
? |
? |
? |
Yes |
? |
|
|
SBC.06061-00001/GRS509 |
GRS1 |
97 |
35 |
16 |
23 |
Yes |
<23 |
|
|
SBC.06061-00006/CHP059 |
CHP |
416 |
? |
13 |
? |
Yes |
? |
|
|
SBC.06061-00006/CHP084 |
CHP |
416 |
? |
13 |
? |
Yes |
? |
|
|
SBC.06061-00006/CHP099 |
CHP |
416 |
? |
18 |
? |
Yes |
? |
|
|
SBC.06061-00006/CHP117 |
CHP |
416 |
13 |
6 |
24 |
? |
>13 |
|
|
SBC.06061-00006/CHP118 |
CHP |
416 |
28.5 |
18 |
25 |
Yes |
<20 |
Pl. 6 |
|
SBC.06061-00006/CHP124 |
CHP |
416 |
36 |
16 |
20 |
Yes |
>21 |
Bersac & Bert, 2019, Pl. 3, figs. 11-12 |
|
SBC.06061-00006/CHP158 |
CHP |
416 |
27 |
13 |
? |
Yes |
20 |
|
|
SBC.06061-00006/CHP361 |
CHP |
416 |
42 |
17 |
? |
No |
16 |
|
|
SBC.06061-00006/CHP391 |
CHP |
416 |
? |
? |
? |
Yes |
? |
Pl. 5 |
|
SBC.06061-00006/CHP392 |
CHP |
416 |
23 |
11.2 |
30 |
Yes |
20 |
Pl. 5 |
|
SBC.06061-00006/CHP393 |
CHP |
416 |
? |
? |
? |
Yes |
? |
|
|
SBC.06061-00006/CHP395 |
CHP |
416 |
15 |
? |
Yes |
? |
Pl. 5 |
|
|
SBC.06061-00007/CLE026 |
CLE |
416 |
12 |
5 |
19 |
? |
>12 |
|
|
SBC.06061-00007/CLE040 |
CLE |
416 |
21 |
9 |
31 |
Yes |
19 |
|
|
SBC.06061-00007/CLE046 |
CLE |
416 |
? |
5 |
? |
? |
? |
|
4. Dufrenoyia furcata (J. de C. Sowerby, 1836)
|
Identification number (N = 210) |
Section |
Bed |
D |
H |
R |
Tuberc. |
SSB |
Figuration (N = 49) |
|
SBC.06061-00001/GRS009 |
GRS1 |
102 |
? |
19 |
? |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS012 |
GRS1 |
102 |
36 |
? |
19 |
>36 |
>36 |
|
|
SBC.06061-00001/GRS017 |
GRS1 |
102 |
? |
13 |
? |
? |
? |
|
|
SBC.06061-00001/GRS024 |
GRS1 |
102 |
30 |
15 |
19 |
>30 |
>30 |
|
|
SBC.06061-00001/GRS042 |
GRS1 |
102 |
65 |
26 |
17 |
? |
? |
|
|
SBC.06061-00001/GRS043 |
GRS1 |
102 |
37 |
18 |
18 |
? |
? |
|
|
SBC.06061-00001/GRS048 |
GRS1 |
102 |
44 |
20 |
18 |
>44 |
29 |
Pl. 7 |
|
SBC.06061-00001/GRS073 |
GRS1 |
100 |
75 |
36 |
19 |
? |
<46 |
Bersac & Bert, 2019, Pl. 5, fig. 5 |
|
SBC.06061-00001/GRS077 |
GRS1 |
98 |
? |
10 |
? |
? |
? |
Pl. 6 |
|
SBC.06061-00001/GRS084 |
GRS1 |
100 |
35 |
14.8 |
11 |
>35 |
? |
Pl. 6 |
|
SBC.06061-00001/GRS085 |
GRS1 |
100 |
27 |
12 |
12 |
>27 |
20 |
Pl. 6 |
|
SBC.06061-00001/GRS086 |
GRS1 |
100 |
? |
15 |
? |
? |
? |
|
|
SBC.06061-00001/GRS087 |
GRS1 |
100 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS089 |
GRS1 |
100 |
23 |
12 |
15 |
>23 |
>23 |
Pl. 6 |
|
SBC.06061-00001/GRS090 |
GRS1 |
100 |
116 |
48 |
21 |
? |
? |
Pl. 6 |
|
SBC.06061-00001/GRS091 |
GRS1 |
99 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS092 |
GRS1 |
100 |
30 |
12 |
16 |
? |
? |
|
|
SBC.06061-00001/GRS093 |
GRS1 |
100 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS094 |
GRS1 |
100 |
26 |
12 |
18 |
? |
? |
|
|
SBC.06061-00001/GRS107 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS113 |
GRS1 |
102 |
27 |
13 |
12 |
>27 |
>27 |
|
|
SBC.06061-00001/GRS120 |
GRS1 |
99 |
? |
11 |
? |
? |
? |
|
|
SBC.06061-00001/GRS124 |
GRS1 |
102 |
25 |
10 |
17 |
? |
? |
|
|
SBC.06061-00001/GRS133 |
GRS1 |
102 |
35 |
13 |
9 |
? |
? |
|
|
SBC.06061-00001/GRS143 |
GRS1 |
100 |
22 |
9 |
? |
>22 |
>22 |
|
|
SBC.06061-00001/GRS154 |
GRS1 |
100 |
? |
14 |
? |
? |
? |
|
|
SBC.06061-00001/GRS155 |
GRS1 |
100 |
25.5 |
11.5 |
13 |
>25.5 |
>25.5 |
Pl. 6 |
|
SBC.06061-00001/GRS156 |
GRS1 |
100 |
39 |
12 |
10 |
>39 |
? |
Pl. 6 |
|
SBC.06061-00001/GRS168 |
GRS1 |
100 |
27 |
12 |
12 |
>27 |
>27 |
Pl. 6 |
|
SBC.06061-00001/GRS169 |
GRS1 |
100 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS170 |
GRS1 |
100 |
26 |
13 |
15 |
>26 |
>26 |
Pl. 6 |
|
SBC.06061-00001/GRS171 |
GRS1 |
100 |
28 |
13 |
12 |
>28 |
24 |
|
|
SBC.06061-00001/GRS172 |
GRS1 |
100 |
26.5 |
14 |
15 |
? |
? |
|
|
SBC.06061-00001/GRS185 |
GRS1 |
100 |
26.4 |
13.2 |
18 |
>26.4 |
>26.4 |
|
|
SBC.06061-00001/GRS189 |
GRS1 |
100 |
25 |
10 |
11 |
>25 |
>25 |
|
|
SBC.06061-00001/GRS190 |
GRS1 |
100 |
28 |
12.6 |
18 |
>28 |
>28 |
|
|
SBC.06061-00001/GRS191 |
GRS1 |
100 |
32 |
12 |
10 |
>32 |
>32 |
|
|
SBC.06061-00001/GRS192 |
GRS1 |
100 |
22 |
11.8 |
17 |
>22 |
20 |
|
|
SBC.06061-00001/GRS194 |
GRS1 |
100 |
29 |
13 |
11 |
>29 |
>29 |
|
|
SBC.06061-00001/GRS195 |
GRS1 |
100 |
36 |
16 |
16 |
>36 |
24 |
|
|
SBC.06061-00001/GRS205 |
GRS1 |
100 |
? |
14 |
? |
? |
? |
Pl. 6 |
|
SBC.06061-00001/GRS212 |
GRS1 |
102 |
? |
20 |
? |
? |
? |
|
|
SBC.06061-00001/GRS214 |
GRS1 |
102 |
? |
55 |
? |
? |
? |
|
|
SBC.06061-00001/GRS227 |
GRS1 |
102 |
118 |
? |
21 |
45 |
? |
Pl. 7 |
|
SBC.06061-00001/GRS228 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS234 |
GRS1 |
102 |
? |
21 |
? |
? |
? |
|
|
SBC.06061-00001/GRS235 |
GRS1 |
102 |
30 |
15 |
24 |
? |
? |
|
|
SBC.06061-00001/GRS241 |
GRS1 |
102 |
17 |
8 |
18 |
>17 |
>17 |
|
|
SBC.06061-00001/GRS272 |
GRS1 |
102 |
40 |
18 |
12 |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS273 |
GRS1 |
102 |
37 |
14 |
15 |
? |
? |
|
|
SBC.06061-00001/GRS274 |
GRS1 |
102 |
18 |
9 |
? |
>18 |
>18 |
|
|
SBC.06061-00001/GRS275 |
GRS1 |
102 |
26 |
10 |
|
>26 |
18 |
Pl. 7 |
|
SBC.06061-00001/GRS276 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS277 |
GRS1 |
102 |
30 |
14 |
15 |
? |
? |
|
|
SBC.06061-00001/GRS278 |
GRS1 |
102 |
53 |
25 |
18 |
>53 |
<30 |
|
|
SBC.06061-00001/GRS279 |
GRS1 |
102 |
34 |
15 |
18 |
? |
16 |
Pl. 7 |
|
SBC.06061-00001/GRS280 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS305 |
GRS1 |
100 |
38 |
18 |
18 |
>38 |
27 |
Pl. 6 |
|
SBC.06061-00001/GRS306 |
GRS1 |
100 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS307 |
GRS1 |
100 |
28 |
16 |
|
>28 |
? |
|
|
SBC.06061-00001/GRS308 |
GRS1 |
100 |
38 |
17 |
15 |
>38 |
37 |
Pl. 6 |
|
SBC.06061-00001/GRS309 |
GRS1 |
100 |
20 |
10 |
? |
>20 |
>20 |
|
|
SBC.06061-00001/GRS314 |
GRS1 |
100 |
19.7 |
8 |
13 |
>19.7 |
>19.7 |
|
|
SBC.06061-00001/GRS316 |
GRS1 |
99 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS317 |
GRS1 |
100 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS318 |
GRS1 |
99 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS332 |
GRS1 |
100 |
? |
17 |
? |
? |
? |
|
|
SBC.06061-00001/GRS333 |
GRS1 |
100 |
? |
14 |
? |
? |
? |
|
|
SBC.06061-00001/GRS356 |
GRS1 |
100 |
24 |
13 |
? |
? |
? |
|
|
SBC.06061-00001/GRS357 |
GRS1 |
100 |
26 |
? |
11 |
>26 |
>26 |
|
|
SBC.06061-00001/GRS361 |
GRS1 |
100 |
41 |
15 |
13 |
>41 |
26 |
Pl. 6 |
|
SBC.06061-00001/GRS363 |
GRS1 |
99 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS375 |
GRS1 |
99 |
30 |
15 |
13 |
>30 |
>30 |
Pl. 6 |
|
SBC.06061-00001/GRS396 |
GRS1 |
99 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS399 |
GRS1 |
104 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS402 |
GRS1 |
104 |
11.5 |
5 |
27 |
? |
>11.5 |
Pl. 7 |
|
SBC.06061-00001/GRS403 |
GRS1 |
104 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS404 |
GRS1 |
104 |
? |
11 |
? |
? |
? |
|
|
SBC.06061-00001/GRS405 |
GRS1 |
104 |
23 |
11 |
25 |
>20 |
>20 |
|
|
SBC.06061-00001/GRS408 |
GRS1 |
104 |
25 |
12 |
14 |
>25 |
>25 |
|
|
SBC.06061-00001/GRS409 |
GRS1 |
104 |
21 |
10 |
22 |
>21 |
>21 |
|
|
SBC.06061-00001/GRS425 |
GRS1 |
99 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS426 |
GRS1 |
|
16 |
7.5 |
? |
? |
? |
Pl. 6 |
|
SBC.06061-00001/GRS431 |
GRS1 |
99 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS433 |
GRS1 |
99 |
124 |
55 |
28 |
? |
? |
|
|
SBC.06061-00001/GRS437 |
GRS1 |
99 |
190 |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS439 |
GRS1 |
104 |
34 |
15 |
? |
? |
? |
|
|
SBC.06061-00001/GRS440 |
GRS1 |
104 |
19 |
10 |
22 |
>19 |
>19 |
|
|
SBC.06061-00001/GRS441 |
GRS1 |
104 |
21 |
9.5 |
30 |
? |
? |
|
|
SBC.06061-00001/GRS442 |
GRS1 |
104 |
27 |
15 |
27 |
>27 |
>27 |
|
|
SBC.06061-00001/GRS443 |
GRS1 |
104 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS445 |
GRS1 |
104 |
30 |
? |
20 |
>30 |
>30 |
|
|
SBC.06061-00001/GRS446 |
GRS1 |
104 |
? |
11 |
? |
? |
? |
|
|
SBC.06061-00001/GRS447 |
GRS1 |
104 |
13 |
6 |
? |
? |
? |
|
|
SBC.06061-00001/GRS448 |
GRS1 |
104 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS450 |
GRS1 |
104 |
26 |
? |
? |
? |
27 |
|
|
SBC.06061-00001/GRS453 |
GRS1 |
104 |
25 |
12 |
19 |
>25 |
>25 |
|
|
SBC.06061-00001/GRS454 |
GRS1 |
104 |
23 |
11.5 |
20 |
>23 |
>23 |
|
|
SBC.06061-00001/GRS455 |
GRS1 |
104 |
31 |
13 |
17 |
? |
? |
|
|
SBC.06061-00001/GRS456 |
GRS1 |
104 |
18 |
7 |
? |
? |
? |
|
|
SBC.06061-00001/GRS459 |
GRS1 |
104 |
26 |
? |
30 |
>26 |
>26 |
|
|
SBC.06061-00001/GRS461 |
GRS1 |
104 |
28 |
? |
18 |
? |
? |
|
|
SBC.06061-00001/GRS462 |
GRS1 |
104 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS470 |
GRS1 |
104 |
23 |
11 |
22 |
>23 |
<13 |
|
|
SBC.06061-00001/GRS476 |
GRS1 |
104 |
22 |
9 |
? |
? |
? |
|
|
SBC.06061-00001/GRS479 |
GRS1 |
104 |
18 |
8.5 |
24 |
>18 |
>18 |
|
|
SBC.06061-00001/GRS485 |
GRS1 |
104 |
? |
9 |
? |
? |
? |
|
|
SBC.06061-00001/GRS486 |
GRS1 |
104 |
25 |
11 |
29 |
>25 |
>25 |
Pl. 7 |
|
SBC.06061-00001/GRS487 |
GRS1 |
104 |
22 |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS494 |
GRS1 |
106 |
? |
? |
? |
? |
|
Pl. 7 |
|
SBC.06061-00001/GRS495 |
GRS1 |
106 |
? |
? |
? |
? |
|
|
|
SBC.06061-00001/GRS499 |
GRS1 |
102 |
80 |
37 |
18 |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS504 |
GRS1 |
104 |
18 |
8 |
20 |
>18 |
>18 |
|
|
SBC.06061-00001/GRS515 |
GRS1 |
102 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS517 |
GRS1 |
102 |
33 |
15 |
? |
? |
? |
|
|
SBC.06061-00001/GRS520 |
GRS1 |
104 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS521 |
GRS1 |
104 |
26 |
? |
? |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS525 |
GRS1 |
104 |
30 |
13.8 |
23 |
>30 |
25 |
Pl. 7 |
|
SBC.06061-00001/GRS536 |
GRS1 |
104 |
16 |
8 |
27 |
>16 |
>16 |
Pl. 7 |
|
SBC.06061-00001/GRS538 |
GRS1 |
104 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS540 |
GRS1 |
104 |
15 |
7 |
19 |
? |
? |
|
|
SBC.06061-00001/GRS546 |
GRS1 |
104 |
? |
10 |
? |
? |
? |
|
|
SBC.06061-00001/GRS551 |
GRS1 |
102 |
25 |
? |
15 |
>25 |
>25 |
|
|
SBC.06061-00001/GRS553 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS556 |
GRS1 |
102 |
? |
? |
? |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS557 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS558 |
GRS1 |
102 |
40 |
16 |
? |
? |
>15 |
|
|
SBC.06061-00001/GRS559 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS560 |
GRS1 |
102 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS563 |
GRS1 |
102 |
29 |
14 |
21 |
>29 |
25 |
|
|
SBC.06061-00001/GRS565 |
GRS1 |
102 |
33 |
15 |
20 |
? |
? |
|
|
SBC.06061-00001/GRS566 |
GRS1 |
102 |
27 |
12 |
? |
>27 |
>27 |
Bersac & Bert, 2019, Pl. 5, fig. 8 |
|
SBC.06061-00001/GRS578 |
GRS1 |
102 |
30 |
14 |
14 |
? |
? |
|
|
SBC.06061-00001/GRS587 |
GRS1 |
109 |
15 |
6 |
26 |
? |
? |
|
|
SBC.06061-00001/GRS588 |
GRS1 |
109 |
22 |
10 |
? |
>22 |
>22 |
|
|
SBC.06061-00001/GRS589 |
GRS1 |
109 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS597 |
GRS1 |
105 |
19 |
8 |
21 |
>19 |
>19 |
Bersac & Bert, 2019, Pl. 5, fig. 9 |
|
SBC.06061-00001/GRS598 |
GRS1 |
105 |
? |
8 |
? |
? |
? |
|
|
SBC.06061-00001/GRS599 |
GRS1 |
105 |
40 |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS604 |
GRS1 |
102 |
35 |
15 |
16 |
>35 |
>35 |
|
|
SBC.06061-00001/GRS608 |
GRS1 |
102 |
? |
25 |
? |
? |
? |
|
|
SBC.06061-00001/GRS609 |
GRS1 |
100 |
30 |
15 |
? |
>30 |
23 |
|
|
SBC.06061-00001/GRS611 |
GRS1 |
100 |
45 |
17 |
14 |
>45 |
<30 |
Pl. 6 |
|
SBC.06061-00001/GRS632 |
GRS1 |
102 |
38 |
16 |
25 |
>38 |
<24 |
Pl. 7 |
|
SBC.06061-00001/GRS633 |
GRS1 |
102 |
36 |
15 |
14 |
>36 |
>36 |
|
|
SBC.06061-00001/GRS635 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS636 |
GRS1 |
102 |
32 |
13 |
10 |
? |
? |
|
|
SBC.06061-00001/GRS650 |
GRS1 |
100 |
27 |
13 |
13 |
>27 |
>27 |
Pl. 6 |
|
SBC.06061-00001/GRS665 |
GRS1 |
102 |
44 |
16 |
9 |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS668 |
GRS1 |
100 |
40 |
20 |
18 |
>40 |
24 |
Pl. 7 |
|
SBC.06061-00001/GRS683 |
GRS1 |
102 |
? |
27 |
? |
? |
? |
Bersac & Bert, 2019, Pl. 5, figs. 6-7 |
|
SBC.06061-00001/GRS738 |
GRS1 |
100 |
35 |
16 |
15 |
? |
24 |
Pl. 6 |
|
SBC.06061-00001/GRS739 |
GRS1 |
99 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS740 |
GRS1 |
101 |
22 |
10 |
? |
? |
? |
Pl. 7 |
|
SBC.06061-00001/GRS741 |
GRS1 |
101 |
18 |
8.5 |
? |
>18 |
>18 |
|
|
SBC.06061-00001/GRS742 |
GRS1 |
101 |
29 |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS743 |
GRS1 |
101 |
? |
16 |
? |
? |
? |
|
|
SBC.06061-00001/GRS744 |
GRS1 |
101 |
45 |
20 |
? |
40 |
? |
Pl. 7 |
|
SBC.06061-00001/GRS745 |
GRS1 |
101 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS756 |
GRS1 |
107 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS757 |
GRS1 |
104 |
23 |
11 |
24 |
>23 |
>23 |
|
|
SBC.06061-00001/GRS758 |
GRS1 |
108 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS759 |
GRS1 |
105 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS760 |
GRS1 |
105 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS763 |
GRS1 |
102 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS764 |
GRS1 |
102 |
25 |
12 |
16 |
>25 |
>25 |
|
|
SBC.06061-00001/GRS765 |
GRS1 |
102 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS766 |
GRS1 |
102 |
? |
5 |
? |
? |
? |
|
|
SBC.06061-00001/GRS767 |
GRS1 |
102 |
44 |
18 |
14 |
>44 |
33 |
Pl. 7 |
|
SBC.06061-00001/GRS768 |
GRS1 |
102 |
? |
8 |
? |
? |
? |
|
|
SBC.06061-00001/GRS769 |
GRS1 |
100 |
32 |
15 |
14 |
? |
? |
|
|
SBC.06061-00001/GRS812 |
GRS1 |
104 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS813 |
GRS1 |
105 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS814 |
GRS1 |
105 |
? |
15 |
? |
? |
? |
|
|
SBC.06061-00001/GRS815 |
GRS1 |
105 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS816 |
GRS1 |
105 |
? |
12 |
? |
? |
? |
|
|
SBC.06061-00001/GRS817 |
GRS1 |
105 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS818 |
GRS1 |
105 |
37 |
17 |
? |
? |
? |
|
|
SBC.06061-00001/GRS823 |
GRS1 |
100 |
30 |
15 |
17 |
>30 |
>30 |
|
|
SBC.06061-00001/GRS824 |
GRS1 |
100 |
40 |
19.5 |
? |
>40 |
? |
|
|
SBC.06061-00001/GRS825 |
GRS1 |
100 |
? |
? |
? |
? |
? |
Pl. 6 |
|
SBC.06061-00001/GRS826 |
GRS1 |
104 |
27 |
? |
26 |
? |
? |
|
|
SBC.06061-00001/GRS827 |
GRS1 |
100 |
32 |
13.5 |
11 |
>32 |
>32 |
|
|
SBC.06061-00001/GRS828 |
GRS1 |
100 |
40 |
15 |
14 |
>40 |
>40 |
|
|
SBC.06061-00001/GRS829 |
GRS1 |
102 |
36 |
17 |
15 |
>36 |
26 |
|
|
SBC.06061-00005/GRO075 |
GRO |
418 |
21 |
8 |
23 |
>21 |
>21 |
Pl. 6 |
|
SBC.06061-00006/CHP037 |
CHP |
418 |
22 |
8 |
|
>22 |
>17 |
|
|
SBC.06061-00006/CHP080 |
CHP |
418 |
25 |
8.2 |
17 |
>25 |
>25 |
|
|
SBC.06061-00006/CHP081 |
CHP |
418 |
19 |
8 |
21 |
>19 |
>19 |
|
|
SBC.06061-00006/CHP163 |
CHP |
418 |
24 |
12 |
14 |
>24 |
>24 |
Pl. 6 |
|
SBC.06061-00006/CHP178 |
CHP |
419 |
12 |
? |
? |
? |
? |
Pl. 6 |
|
SBC.06061-00006/CHP179 |
CHP |
419 |
? |
7 |
? |
? |
? |
|
|
SBC.06061-00006/CHP181 |
CHP |
421 |
35 |
12 |
? |
>35 |
<22 |
|
|
SBC.06061-00006/CHP187 |
CHP |
419 |
37 |
15 |
? |
? |
? |
|
|
SBC.06061-00006/CHP188 |
CHP |
419 |
17 |
7 |
? |
? |
? |
|
|
SBC.06061-00006/CHP195 |
CHP |
421 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP198 |
CHP |
421 |
23 |
10 |
13 |
>23 |
>23 |
Pl. 6 |
|
SBC.06061-00006/CHP200 |
CHP |
417 |
20 |
9 |
22 |
>20 |
>20 |
Pl. 6 |
|
SBC.06061-00006/CHP211 |
CHP |
426 |
18 |
7 |
20 |
>18 |
>18 |
|
|
SBC.06061-00006/CHP213 |
CHP |
426 |
21 |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP214 |
CHP |
426 |
18 |
7 |
14 |
>18 |
>18 |
|
|
SBC.06061-00006/CHP215 |
CHP |
426 |
? |
14 |
? |
? |
? |
|
|
SBC.06061-00006/CHP216 |
CHP |
426 |
27 |
12 |
|
>27 |
>27 |
|
|
SBC.06061-00006/CHP217 |
CHP |
426 |
14 |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP218 |
CHP |
426 |
23 |
12 |
? |
>23 |
>23 |
Pl. 6 |
|
SBC.06061-00006/CHP222 |
CHP |
426 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP223 |
CHP |
426 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP247 |
CHP |
424 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00006/CHP461 |
CHP |
424 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00009/PPR007 |
PPR |
420 |
? |
? |
? |
? |
? |
|
5. Dufrenoyia dufrenoyi (Orbigny, 1841)
|
Identification number (N = 20) |
Section |
Bed |
D |
H |
R |
Tuberc. |
SSB |
Figuration (N = 4) |
|
SBC.06061-00001/GRS592 |
GRS1 |
113 |
? |
15 |
? |
? |
? |
Bersac & Bert, 2019, Pl. 5, fig. 32 |
|
SBC.06061-00001/GRS593 |
GRS1 |
113 |
24 |
11 |
24 |
>24 |
>24 |
|
|
SBC.06061-00001/GRS594 |
GRS1 |
113 |
32 |
14 |
18 |
>32 |
>32 |
|
|
SBC.06061-00001/GRS595 |
GRS1 |
113 |
18 |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS596 |
GRS1 |
113 |
16 |
? |
? |
>16 |
>16 |
|
|
SBC.06061-00001/GRS685 |
GRS1 |
110 |
? |
16 |
? |
? |
? |
Bersac & Bert, 2019, Pl. 5, figs. 30-31 |
|
SBC.06061-00001/GRS746 |
GRS1 |
113 |
? |
15 |
? |
? |
? |
|
|
SBC.06061-00001/GRS747 |
GRS1 |
110 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS748 |
GRS1 |
110 |
19 |
9 |
19 |
>19 |
>19 |
|
|
SBC.06061-00001/GRS749 |
GRS1 |
110 |
? |
14 |
? |
? |
? |
|
|
SBC.06061-00001/GRS750 |
GRS1 |
110 |
12 |
6.5 |
15 |
>12 |
>12 |
|
|
SBC.06061-00001/GRS751 |
GRS1 |
110 |
22 |
10 |
22 |
>22 |
>22 |
Pl. 7 |
|
SBC.06061-00001/GRS752 |
GRS1 |
110 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS753 |
GRS1 |
110 |
23 |
11 |
? |
? |
? |
|
|
SBC.06061-00001/GRS754 |
GRS1 |
110 |
18 |
7 |
24 |
? |
? |
|
|
SBC.06061-00001/GRS755 |
GRS1 |
110 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS819 |
GRS1 |
110 |
34 |
15 |
15 |
>34 |
>34 |
|
|
SBC.06061-00001/GRS820 |
GRS1 |
110 |
? |
? |
? |
? |
? |
|
|
SBC.06061-00001/GRS821 |
GRS1 |
110 |
37 |
19 |
19 |
>37 |
>37 |
Pl. 7 |
|
SBC.06061-00001/GRS822 |
GRS1 |
110 |
? |
? |
? |
? |
? |
|
[*] Editor's note: The wording 'Tethyan,' which was incorrect, was changed to 'Tethys' (see Granier, 2022).