◄ Carnets Geol. 25 (10) ►
![]()
Outline:
[1. Introduction]
[2. Study area and methodology]
[3. Stratigraphic framework]
[4. Biostratigraphic analysis]
[5. Conclusion]
[Bibliographic references] and ...
[List of species]
corresponding author;
AQUABIOTECH laboratory, Cadi Ayyad University, Faculty of Sciences Semlalia
- Marrakesh, P.O. Box 2390, Marrakesh, 40000 (Morocco)
Department of Geology, Faculty of Sciences, Chouaďb Doukkali University, P.O. Box 20, El Jadida (Morocco)
Chemin des Pochattes, n° 4, CH-1055 Froideville (Switzerland)
AQUABIOTECH laboratory, Cadi Ayyad University, Faculty of Sciences Semlalia - Marrakesh, P.O. Box 2390, Marrakesh, 40000 (Morocco)
Institut des mines Touissit / ŕ travers Oujda, 64850 (Morocco)
Geology and Sustainable Mining Institute, Mohammed VI Polytechnic University, Benguerir (Morocco)
Department of Stratigraphy and Paleontology, A.P. Karpinsky Russian Geological Research Institute (VSEGEI), Sredny pr. 74, 199106 Saint-Petersburg (Russia)
Published online in final form (pdf) on October 14, 2025
DOI 10.2110/carnets.2025.2510
![]()
[Editor: Bruno
R.C. Granier; language editors: Simon F. Mitchell
& Stephen Carey]
![]()
The Lower and lower Middle Jurassic carbonates from the southern margin of the Moroccan High Atlas yielded well-preserved foraminifers and calcareous algae. The identification of these microfossils, based on thin-section observations, allowed us to define seven successive micropaleontological assemblages with ages from the early Sinemurian to the early Bajocian. These micropaleontological assemblages, relatively well calibrated by ammonites and brachiopods, were compared with the larger foraminiferal biozonation of Septfontaine (1984) for the Moroccan Tethys domain. A new biozone with Palaeodasycladus mediterraneus is introduced to characterize the upper Pliensbachian (Algovianum Zone) to lower Toarcian interval.
• Lower Jurassic;
• large benthic foraminifers;
• algae;
• biostratigraphy;
• biozones;
• carbonate platform;
• High Atlas;
• Tethys domain;
• Morocco
Ettaki M., Bouaouda M.-S., Septfontaine M., Chellai E.H., Milhi A., El Bamiki R. & Vuks V.J. (2025).- Lower and lower Middle Jurassic foraminiferal assemblages and calcareous algae from the southern margin of the Central High Atlas, Morocco.- Carnets Geol., Madrid, vol. 25, no. 10, p. 201-217. DOI: 10.2110/carnets.2025.2510
Les associations de foraminifčres et d'algues calcaires du Jurassique inférieur et de la base du Jurassique moyen de la marge sud du Haut Atlas central, Maroc.- Les carbonates du Jurassique inférieur et de la base du Jurassique Moyen de la marge méridionale du Haut Atlas marocain ont livré des algues calcaires et des foraminifčres bien conservés. L'identification et l'inventaire de ces microfossiles, basés sur l'observation de lames minces, a permis de définir sept associations micropaléontologiques se succédant du Sinémurien inférieur au Bajocien inférieur. Ces associations micropaléontologiques, relativement bien calibrées par les ammonites et les brachiopodes, ont été comparées ŕ la biozonation ŕ base de foraminifčres de Septfontaine (1984) pour le Domaine téthysien marocain. Nous proposons d'introduire une association ŕ Palaeodasycladus mediterraneus pour caractériser l'intervalle Pliensbachien supérieur (Zone ŕ Algovianum) - Toarcien inférieur.
• Jurassique inférieur ;
• grands foraminifčres benthiques ;
• algues calcaires ;
• biostratigraphie ;
• biozones ;
• plate-forme carbonatée ;
• Haut Atlas ;
• Domaine téthysien ;
• Maroc
For recent depositional environments, foraminifers and calcareous algae are considered good paleontological and environmental bioindicators (Hallock & Glenn, 1986; Kooistra et al., 2002). Fossil assemblages of larger benthic foraminifers (LBF) and calcareous algae are generally considered as indicators of paleoenvironments with a stratigraphical significance (Septfontaine, 1984, 1985, 2020; Kabal & Tasli, 2003; Clark & Boudagher-Fadel, 2004; Velić, 2007; Reolid et al., 2008; Gale, 2014; Sevillano et al., 2020). In some cases, they can play a key role in age-calibrating and refining the biostratigraphic framework for correlation of sedimentary sequences.
The present work deals with biostratigraphic analyses based on the associations of Lower Jurassic calcareous algae and larger benthic foraminifers (LBF) from the southern margin of the Central High Atlas. Most of the previous biostratigraphical and paleontological studies (Dubar, 1952; Dresnay, 1977; Dresnay et al., 1978; Sadki, 1996; El Hariri, 1998; Lachkar, 2000) have been devoted to the northern and the northeastern parts of the Central High Atlas, whereas our study area is located in the southern margin of the central High Atlas. If we exclude the biostratigraphic studies carried by Septfontaine (1984, revised 2020), Ettaki et al. (2011) and those forming part of a PhD Thesis (Ettaki, 2003), this region remains insufficiently studied from a micropaleontological point of view. The micropaleontological work developed by Hottinger (1967), Septfontaine (1984, 1985, 2020), and Bassoullet (1997a, 1997b, in Bassoullet et al., 1999) have been supportive of the study herein.
An inventory of larger benthic foraminifers (LBF) and other microfossils was reported for the first time by Ettaki (2003) in an unpublished PhD thesis. This inventory will be documented and detailed in this paper, providing insights into the proposed assemblages of Lower Jurassic foraminifers and calcareous algae. Furthermore, a biostratigraphic subdivision with high resolution is proposed, helping to trace the evolution of the micropaleontological assemblages from the southern margin of the Central High Atlas during the Lower Jurassic. This subdivision, based on LBF and calcareous algae, is compared to the reference biostratigraphic scheme developed by Septfontaine (1984, revised 2020).The stratigraphic position of some LBF biozones was precisely located from levels with ages constrained by ammonites and brachiopods collected from stratigraphical sections in the Todrha-Dades area.
This study was carried out in the Todrha-Dades area, located about 100 km
NE of Ouarzazate city. This area, which is part of the southern edge of the
Central High Atlas, is bordered by Wadi Dades to the west and Wadi Todrha to the
east, and is limited to the south by the South Atlas Fault (P. & L. Russo,
1934) and to the north by the North Atlas Fault (Roch,
1939)
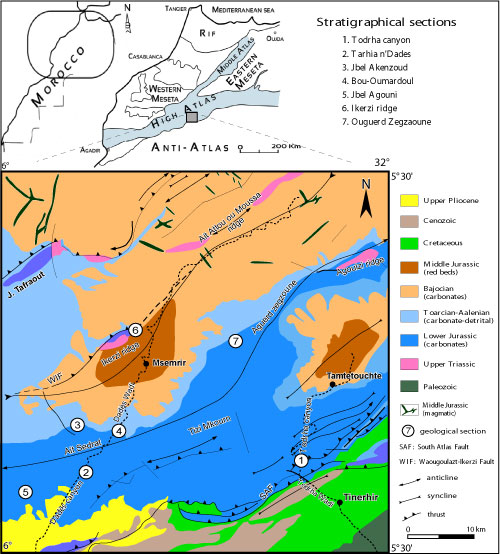
(Fig. 1 ![]() ).
The structural fabric of the Todrha-Dades area is mainly a network of NE-SW and
NNW-SSE faults, inherited from the Hercynian multi-stage orogeny.
).
The structural fabric of the Todrha-Dades area is mainly a network of NE-SW and
NNW-SSE faults, inherited from the Hercynian multi-stage orogeny.
|
Figure 1:
Geological map of the
Todrha-Dades High Atlas. |
For this work, seven stratigraphical sections were logged on the southern margin of the Moroccan High Atlas at localities within the Todrha-Dades area (Table 1):
Table 1: Names and geographical coordinates of the studied sections.
| Locality | Latitude | Longitude |
| 1. Todrha canyon | 31°36'48.60"N | 5°35'55.96"W |
| 2. Tarhia n'Dades | 31°32'47.04"N | 5°55'29.64"W |
| 3. Jbel Akenzoud | 31°37'39.72"N | 5°54'15.48"W |
| 4. Bou-Oumardoul | 31°36'02.52"N | 5°52'21.00"W |
| 5. Jbel Agouni | 31°31'41.16"N | 5°59'16.80"W |
| 6. Ikerzi ridge | 31°44'42.36"N | 5°50'29.76"W |
| 7. Ouguerd Zegzaoune | 31°45'16.92"N | 5°39'44.28"W |
Selective sampling was guided in the field by an initial observation with magnifying hand lenses. Subsequently, the laboratory analysis of microfacies in thin sections allowed for the identification of foraminifers and calcareous algae, as well as the characterization of depositional paleoenvironments. The inventory of the different microfossils found in the Lower Jurassic was compared to previous work by Hottinger (1967), Septfontaine (1984), Milhi (1992, 1997), and Ettaki (2003), carried out in the same area, and with stratigraphic work conducted in other High Atlas regions by Bassoullet et al. (1976) and Ettaki et al. (2000a, 2000b, 2011).
The Lower Jurassic formations of the
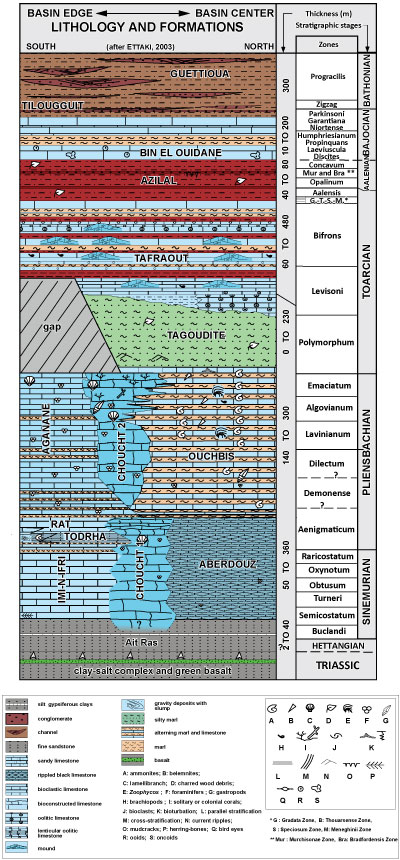
Todrha-Dades High Atlas show a lithological heterogeneity (Fig. 2 ![]() )
related to
active syn-sedimentary tectonics coupled with eustatic variations (Ettaki,
2003; Ettaki et al., 2007a, 2007b, 2008). These
sedimentary successions, often fossiliferous, have evolved in depositional
environments ranging from coastal marginal marine to offshore deeper-water
settings. The
studied sedimentary successions consist of 500 to 800 m-thick carbonate and
siliciclastic sediments,
subdivided into eleven lithostratigraphic formations. These have been
extensively described in unpublished synthetic works elaborated for the whole
central High Atlas by Ettaki et al. (1996) and Ettaki
(2003), and are briefly summarized here:
)
related to
active syn-sedimentary tectonics coupled with eustatic variations (Ettaki,
2003; Ettaki et al., 2007a, 2007b, 2008). These
sedimentary successions, often fossiliferous, have evolved in depositional
environments ranging from coastal marginal marine to offshore deeper-water
settings. The
studied sedimentary successions consist of 500 to 800 m-thick carbonate and
siliciclastic sediments,
subdivided into eleven lithostratigraphic formations. These have been
extensively described in unpublished synthetic works elaborated for the whole
central High Atlas by Ettaki et al. (1996) and Ettaki
(2003), and are briefly summarized here:
3.1. The Aďt Ras Formation (Le Marrec & Jenny, 1980). This formation is represented by alternations of limestone, lithoclastic dolostone, flagstone dolostone, and sandy marlstone (50 to 60 m). The lithology of this formation indicates that it was deposited in a marginal - littoral environment, and yielded only very rare, indeterminate foraminifers and ? Rivulariaceae. This formation is of Hettangian? - early Sinemurian age (Le Marrec & Jenny, 1980).
3.2. The Imi-n-Ifri Formation (Jenny, 1988). This 102 m thick formation consists of laminated limestone, oolitic limestone, lithoclastic limestone, and intraformational breccia levels. This formation, evolved from an intertidal to a supratidal environment, and based on its stratigraphic position is of Sinemurian age (Ettaki, 2003).
|
Figure 2:
Lithostratigraphy of the
Lower and Middle Jurassic in the Todrha-Dades High Atlas (after Ettaki et al.,
1996, 2008;
Milhi et al., 2002;
Ettaki, 2003, compiled). |
3.3. The Choucht Formation (Septfontaine, 1985; Jossen, 1990). This unit corresponds to a set of gravelly, oolitic, and coralligenous sediments. This formation developed within a carbonate platform and has been subdivided into two parts Milhi (1992, 1997):
The lower part (Choucht 1), constituting
the wall of the Todrha canyon, corresponds to approximately 360 m of massive
limestone with cross-bedding and sponge-rich limestone containing: Lithistids (identified by Prof. Bernier,
Claude Bernard Lyon 1 University), calcareous algae including Palaeodasycladus
sp., Tersella alpina, and Thaumatoporella
parvovesiculifera, as well as larger
benthic foraminifers (LBF), including Amijiella
amiji, Everticyclammina sp., Glomospira
sp., Lituosepta recoarensis, Paleomayncina termieri, Pseudopfenderina
butterlini, Redmondoides sp., Siphovalvulina
sp., Spirillina sp., Verneuilinoides
mauritii, and Ophthalmidium martanum
(Fig. 3 ![]() ). Additionally, microproblematic fossils, such as Bacinella
microstructures (Schlagintweit
et al., 2011; Granier, 2021), Cayeuxia
sp., C. piae, C.
aff. kurdistanensis, C. cf.
mediterranea, and Mitcheldeania sp., are present. The association of
algae and LBF indicates an age extending from the early Sinemurian to the early
Pliensbachian (Ettaki, 2003).
). Additionally, microproblematic fossils, such as Bacinella
microstructures (Schlagintweit
et al., 2011; Granier, 2021), Cayeuxia
sp., C. piae, C.
aff. kurdistanensis, C. cf.
mediterranea, and Mitcheldeania sp., are present. The association of
algae and LBF indicates an age extending from the early Sinemurian to the early
Pliensbachian (Ettaki, 2003).
|
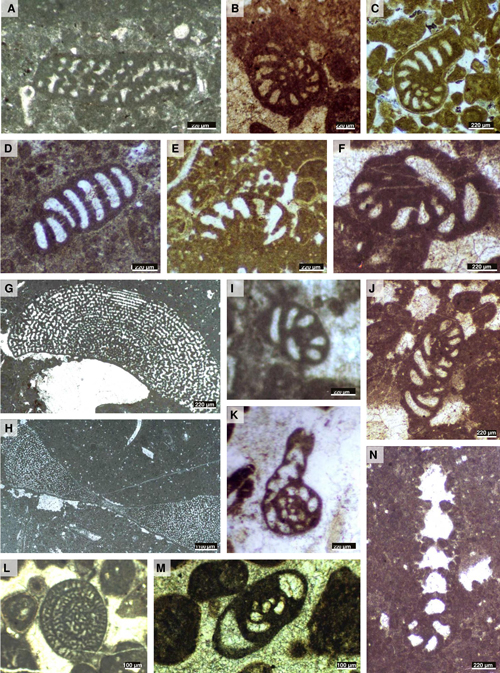
Figure 3:
Examples of Lower Jurassic foraminifers from the
southern margin of the Central High Atlas: A) Lituosepta recoarensis,
sub-axial
section, thin section JR2, Todrha canyon (Akka n'Igoulzane), lower Pliensbachian
(Aenigmaticum
Zone). B-E) Paleomayncina
termieri: B) Equatorial section, thin section N14, Todrha canyon,
Sinemurian (biozone B); C) Equatorial section, thin section Aga19, Todrha canyon
(Akka n'Igoulzane), lower Pliensbachian; D) Sub-axial section,
thin section N17, Todrha canyon, upper Sinemurian - lower Pliensbachian (Aenigmaticum
Zone) (biozone C1); E) Oblique/equatorial section, thin section Aga10, Todrha canyon (Akka n'Igoulzane),
lower Pliensbachian. F, I-J) Pseudopfenderina
butterlini: F) Sub-transverse section, thin section N17, Todrha canyon
(biozone C1); I) Transverse section/oblique, thin section N7, Todrha canyon,
Sinemurian; J) Axial section, thin section N17, Todrha canyon (biozone C1). G-H)
Orbitopsella praecursor: G) Sub-equatorial section of
microspheric form, thin section P20, Todrha canyon (Akka n'Igoulzane), lower Pliensbachian (biozone C2); H) Axial
section of microspheric form preserved in a dark fine micrite, thin section F69,
Tizgui locality, lower Pliensbachian (biozone
C2). K) Bosniella croatica, equatorial section, thin section Rh9,
Ilourhman locality, upper Pliensbachian. L) Haurania
deserta, transverse section, thin section Ta45, Tarhia n'Dades, upper
Pliensbachian Emaciatum Zone
(biozone E). M) Bosniella sp., sub-axial section, thin section
Rh3, Ilourhman locality, upper Pliensbachian. N) Everticyclammina
sp., equatorial section, thin section N18, Todrha canyon (biozone C1). |
The upper part (Choucht 2) is a 190 m
thick peri-reef limestone constituting the dominant
part of the Dades canyon wall (Ettaki, 2003). The fossil content
includes bivalve fragments, echinoid fragments and
radioles, belemnites, brachiopods: Lobothyris
sp., Lobothyris subpunctata, Prionorhynchia
canavarii, and Pr. gignouxi (identified
by Prof. Alméras), ammonites of the genus Emaciaticeras
sp. (identified by Prof. Dommergues), and foraminifers including Amijiella
amiji, Ammobaculites coprolithiformis,
Bosniella sp., Epistomina
sp., Everticyclammina sp., Glomospira
sp., Glomospirella sp., Haplophragmoides
sp., Haurania deserta, Involutina
liassica, Lituosepta cf. compressa, Mesoendothyra sp., Nodosaria sp., Ophthalmidium
martanum, Planisepta compressa, Pseudocyclammina
sp., Redmondoides sp., R. cf. lugeoni,
Siphovalvulina sp., Spirillina
sp., Trochammina sp., Trocholina
sp., Valvulina sp., Verneuilinoides
mauritii, Verneuilina
sp., and encrusting foraminifers such as Coscinophragma
sp. The algae are represented by Dasycladales including Chinianella
ellembergeri, Otternstella sp., Palaeodasycladus
sp., P. elongatus,
P. mediterraneus (Fig. 4.D, .G-H ![]() ), Uragiella liasica, as well as Elianellaceae, Thaumatoporella
parvovesiculifera, and microproblematic organism such as Cayeuxia piae
and Bacinella microstructures.
), Uragiella liasica, as well as Elianellaceae, Thaumatoporella
parvovesiculifera, and microproblematic organism such as Cayeuxia piae
and Bacinella microstructures.
This microfossil content indicates a late Pliensbachian (Algovianum Zone) to the earliest Toarcian age (Ettaki, 2003). The presence of Emaciaticeras allowed the upper part of the Choucht 2 Formation to be attributed to the Emaciatum Zone, Elisa Subzone (Ettaki, 2003).
|
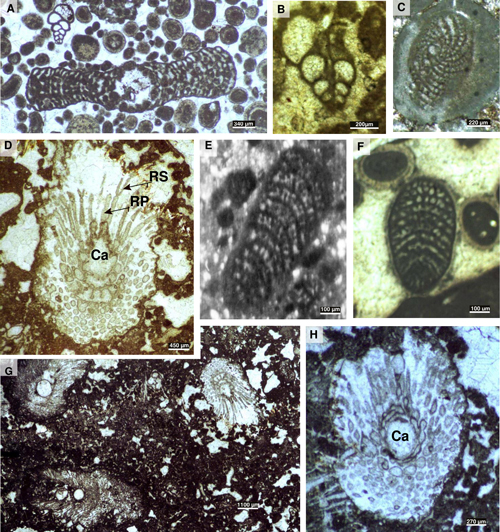
Figure 4:
Examples of Lower Jurassic Algae and foraminifers
from the southern margin of the Central High Atlas: A) Orbitopsella praecursor.
Axial section of a macrospheric form associated to Valvulina sp., thin
section Aga12, Todrha canyon (Akka n'Igoulzane). B) Siphovalvulina
sp. Axial section, thin section GT5, Tarhia n'Dades, upper Pliensbachian (Emaciatum
Zone, Elisa Subzone). C) Timidonella sarda.
Sub-equatorial section, thin section Agz288, Ouguerd Zegzaoune locality, middle
Aalenian - lower Bajocian. D, G-H) Palaeodasycladus
mediterraneus: D) Oblique section (Ca: axial cavity, SL: secondary
laterals, TL: tertiary laterals), thin section Ta44, Tarhia n'Dades, upper Pliensbachian
(Emaciatum Zone, Elisa
Subzone); G) Association of Palaeodasycladus
mediterraneus, thin section Ta44,
Tarhia n'Dades, upper Pliensbachian (Emaciatum Zone,
Elisa Subzone); H) Oblique section, thin section Ta43, Tarhia n'Dades, upper Pliensbachian
(Emaciatum Zone).
E) Valvulina sp.,
Axial section, thin section N15, Todrha canyon, Sinemurian (biozone B). F
Haurania deserta. Axial section, thin section Aga36, Todrha canyon
(Akka n'Igoulzane), upper Pliensbachian (Algovianum
Zone). |
3.4. The Todrha Formation (Milhi, 1992). It consists of 120 m of sediment gravity flow deposits. The limestone microfacies correspond to a biomicrite with lithoclasts, oncoids, bioclasts, and calcareous algae. The foraminiferal content includes Ammobaculites coprolithiformis, Epistomina sp., Everticyclammina sp., Glomospira sp., Pseudopfenderina butterlini, Redmondoides sp., Siphovalvulina sp., Verneuilinoides mauritii, and other small foraminifers with a hyaline wall.
The Todrha Formation, which was attributed to the Sinemurian by Milhi (1992, 1997), yielded Miltoceras taguendoufi (El Hariri et al., 1996), and Metaderoceras gr. apertum (El Hariri et al., 1996) (identified by Prof. Dommergues) in the Akka n'Igoulzane (Todrha Canyon) locality, which made it possible to extend the age of this formation to the Aenigmaticum Zone of the early Pliensbachian (Ettaki, 2003; Ettaki et al., 2011).
3.5. The Jbel Rat Formation (Jenny, 1988). It is composed of a 0 to 60 m-thick sequence of limestones or dolostones with 'bird eyes', oncoids, oolitic limestones with cross-bedding, and dolostones with oncoids and tepee structures. The identified foraminifers are Everticyclammina sp., Lituosepta recoarensis, Redmondoides sp., Siphovalvulina sp., and Verneuilinoides mauritii. The algae include Dasycladales and ? Rivulariaceae (Cayeuxia sp.). This formation was deposited in an intertidal to supratidal environment (Milhi, 1992, 1997; Ettaki, 2003) during the Aenigmaticum Zone of the early Pliensbachian (Ettaki, 2003).
3.6. The Aganane Formation (Jenny, 1985, 1988; Septfontaine, 1985). This 160 m thick formation is a rhythmic sedimentary succession, organized into cycles formed of limestones and dolostones, separated by varicolored marl layers. These facies were deposited in a temporary lagoonal environment within an inner platform (Septfontaine, 1985; Milhi, 1992, 1997; Ettaki, 2003). The faunistic content includes in the lower part of this formation the following benthic foraminifers and other microfossils forms: Ammobaculites sp., Bosniella aff. croatica, Glomospira sp., Paleomayncina termieri, Pseudopfenderina butterlini, Siphovalvulina sp., and archaic forms of Valvulinidae. The upper part contains: Amijiella amiji, Bosniella sp., Orbitopsella praecursor, and Planisepta compressa. The algae include Cayeuxia sp. These microfossils indicate the interval represented by the Demonense Zone to Lavinianum Zone, and corresponding to Septfontaine's (1984) biozones C2 and D.
3.7. The Aberdouz Formation (Stüder, 1980). This formation consists of 30 to 140 m of black micritic limestones, locally associated with flint nodules. This open marine formation includes bivalves, echinoderms, some algae such as Thaumatoporella parvovesiculifera and foraminifers including Ammobaculites gr. coprolithiformis, Dentalina sp., Epistomina sp., Everticyclammina sp., Lenticulina sp., Nodosaria sp., Ophthalmidium martanum, Pseudopfenderina butterlini, Redmondoides sp., Spirillina sp., and Verneuilinoides mauritii. In some sections (e.g., the Bou-Oumardoul locality), rare discoveries of Liospiriferina sp. (brachiopods, identified by Prof. Alméras) allow these layers with brachiopods to be assigned to the Algovianum Zone of the upper Pliensbachian (Ettaki, 2003; Ettaki et al., 2008). The Aberdouz Formation extends from the Sinemurian to the Algovianum Zone of the upper Pliensbachian (Ettaki et al., 2011).
3.8. The Ouchbis Formation (Stüder, 1980). It consists of 140 to 300 m of limestone and marlstone alternations that were deposited in a subtidal to bathyal environment or even on a reef slope (Evans & Kendall, 1977; Dresnay, 1971, 1979; Stüder, 1980; Bernasconi, 1983; Milhi, 1992, 1997; Ettaki, 2003). The macrofauna includes ammonites, belemnites, and bivalves (Bositra). The micropaleontological association is composed of foraminifers: Amijiella amiji, Ammobaculites gr. coprolithiformis, Everticyclammina sp., Paleomayncina sp., Pseudocyclammina aff. liasica, Pseudopfenderina butterlini, aff. Redmondoides lugeoni, Siphovalvulina sp., and other taxa such as Epistomina sp., Glomospirella sp., Glomospira sp., Haurania deserta, Lenticulina sp., Lingulina gr. tenera, Nodosaria sp., Ophthalmidium leischneri, O. martanum, Spirillina sp., Trochamminidae, Verneuilinoides mauritii, and Verneuilinoides sp., as well as algae including Elianellaceae and Thaumatoporella parvovesiculifera. Additionally, microproblematic fossils, such as Cayeuxia piae and Bacinella nodules, are present. These microfossils are often reworked (Ettaki, 2003). The ammonites collected by Ettaki include several species of the genera Metaderoceras, Lytoceras, Protogrammoceras, Fuciniceras, Arieticeras, Reynesocoeloceras, Neolioceras, Emaciaticeras, Canavaria, Tauromeniceras, Fontanelliceras, and Eodactylites, that allow the Ouchbis Formation to be assigned to the interval from the lower Pliensbachian (Demonense Zone) to the lower Toarcian (Polymorphum Zone) (Ettaki, 2003; Ettaki et al., 2011).
3.9. The Tagoudite Formation (Stüder, 1980). It consists of up to 230 m of sandy marlstone and calcareous sandstone. These sediments correspond to turbidite facies that were deposited in a deep depocentre created by block tilting (Ettaki, 2003; Ettaki et al., 2007a, 2008). This formation, dated by stratigraphic framing, is attributed to the lower Toarcian (Polymorphum - Levisoni zones) (Ettaki & Chellai, 2005).
3.10. The Tafraout Formation (Milhi, 1997). This formation is composed of 60 to 400 m of siliciclastic sediments with oolitic and bioclatic limestone layers deposited on a proximal platform (Milhi, 1997). In some sections, the brachiopods collected include: Homoeorhynchia meridionalis, H. batalleri, Telothyris jauberti var. depressa, T. arnaudi, and juvenile forms of Telothyris jauberti (identified by Prof. Ouahhabi). This association indicates an interval within the Toarcian extending from the Serpentinus Zone to the base of the Bifrons Zone (Sublevisoni and probably Lusitanicum subzones) (Ettaki et al., 2011). The upper levels of this formation yielded two poorly preserved ammonite molds corresponding to Hildoceras sp. (identified by Prof. Ouahhabi), indicating the middle Toarcian, Bifrons Zone (Ettaki, 2003).
3.11. The Azilal Formation (Jenny, 1988). This is composed of siltstones, calcareous sandstones and dolostones. The faunistic content includes only rare bivalves, gastropods, brachiopods, and foraminifers. The Azilal Formation is stratigraphically located between the Tafraout Formation (middle Toarcian) and the Bin El Ouidane 1 Formation of a Bajocian age (Milhi et al., 2002). The carbonate horizons at the top of the Azilal Formation yielded Timidonella sarda, indicating that this formation belongs to the upper Toarcian - lower Bajocian interval.
Only some Lower Jurassic formations of the
Todrha-Dades area yielded a significant fauna of ammonites and/or brachiopods
for high precision age-calibration. However, other carbonate-dominated
formations contain well-preserved foraminifers and calcareous algae (Fig. 5 ![]() )
allowing biostratigraphic interpretations. Thus, the Lower Jurassic
micropaleontology can be described and a biostratigraphic subdivision proposed
based on larger benthic foraminifers (LBF) and calcareous algae.
)
allowing biostratigraphic interpretations. Thus, the Lower Jurassic
micropaleontology can be described and a biostratigraphic subdivision proposed
based on larger benthic foraminifers (LBF) and calcareous algae.
In the Todrha-Dades High Atlas, the most recent discoveries of brachiopods and ammonites (Ettaki et al., 2011) assisted with the age-calibration of micropaleontological assemblages and provided constraints regarding the stratigraphy of some biozones previously proposed for the Lower Jurassic of the High-Atlas in the Moroccan Tethys domain (Septfontaine, 1985). In horizons devoid of fossil markers, the age-calibration of micropaleontological assemblages is based on comparisons with the biostratigraphic scale proposed by Septfontaine (1984). The lower Sinemurian - lower Bajocian interval is characterized by seven microfossil assemblages presented here from base to top:
|
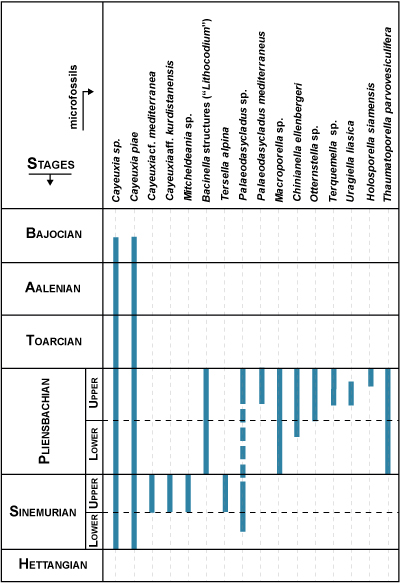
Figure 5:
Stratigraphic
distribution of some algal species in the Lower and Middle Jurassic limestones
of the southern margin of the Central High Atlas. |
4.1. Siphovalvulina sp. and Everticyclammina sp. assemblage: Hettangian? - lower Sinemurian.
This assemblage characterizes the base of the Lower Jurassic series in the study area. It corresponds perfectly to Septfontaine's (1984) biozone A (interval zone). The microfossils identified here are not age-characteristic since they occur in almost all of the overlying assemblages. The absence of typical markers belonging to the orbitopsellids generate a need for further investigations in the field. This assemblage is represented in the Todrha-Dades High Atlas only by Everticyclammina sp., Siphovalvulina sp., and rare ? Rivulariaceae.
4.2. Lituosepta recoarensis and Pseudopfenderina butterlini assemblage: upper Sinemurian.
From the late Sinemurian, new paleoecological conditions led to a flourishing of foraminifers, especially those with complex internal structure.
The base of this assemblage is defined by the appearance of Pseudopfenderina butterlini, whilst its top coincides with the disappearance of Lituosepta recoarensis. These two characteristic forms are associated with other microfossils, among which the most notable are Amijiella amiji, Ammobaculites gr. coprolithiformis, Everticyclammina sp., Ophthalmidium martanum, Paleomayncina termieri, Pseudopfenderina sp., Siphovalvulina sp., Verneuilinoides mauritii, Palaeodasycladus sp., Cayeuxia piae, C. cf. mediterranea, C. aff. kurdistanensis, and Thaumatoporella parvovesiculifera.
This assemblage of late Sinemurian age
corresponds to Septfontaine's (1984) biozone B (lineage zone), defined by the appearance of Lituosepta
recoarensis. In
the Todrha-Dades High Atlas, Pseudopfenderina
butterlini appears before Lituosepta
recoarensis, and
this disagrees with the data presented by Septfontaine
(1984, 2020). This difference
in the vertical distribution of taxa could be related to facies and
paleoenvironmental conditions. It is likely that Lituosepta proliferated in lagoonal facies (Aganane Formation). In
the Todrha canyon, the L. recoarensis
and Pseudopfenderina butterlini
assemblage was recognized within the deposits of the Choucht 1 Formation (Fig.
3.A ![]() ). It should be noted that in the Todrha-Dades High Atlas, the occurrence of
lagoonal facies starts within the Aenigmaticum
Zone of the early Pliensbachian.
). It should be noted that in the Todrha-Dades High Atlas, the occurrence of
lagoonal facies starts within the Aenigmaticum
Zone of the early Pliensbachian.
The chronostratigraphic position of this assemblage is based both on the interpretations of Septfontaine (1984, 2020) and on the occurrence of ammonites (Metaderoceras gr. apertum and Miltoceras taguendoufi) of the lower Pliensbachian (Aenigmaticum Zone) in the overlying horizons (Ettaki, 2003; Ettaki et al., 2011). These occurrences allow the Lituosepta recoarensis and Pseudopfenderina butterlini assemblage to be assigned to the upper Sinemurian.
4.3. Paleomayncina termieri and Pseudopfenderina butterlini assemblage: uppermost Sinemurian - lower Pliensbachian, Aenigmaticum Zone (interval zone).
This interval zone is characterized by the disappearance of Lituosepta recoarensis at its lower limit and by the appearance of Orbitopsella praecursor at its upper limit. This assemblage is based on the association of Paleomayncina termieri and Pseudopfenderina butterlini. The accompanying fauna consists of Bosniella gr. croatica, Everticyclammina sp., aff. Redmondoides lugeoni, and Siphovalvulina sp.. The algae include Dasycladales, Thaumatoporella parvovesiculifera, Cayeuxia piae, and incertae sedis. This association has been recognized within the Aganane and the Choucht 1 formations in Todrha canyon, and within the Aberdouz Formation at Jbel Agouni. This assemblage can be correlated with Septfontaine's (1984) biozone C1. According to this author, the index species of biozone C1 (lineage zone) is Orbitopsella primaeva. This index species has not been identified in the Todrha-Dades High Atlas.
The age of this assemblage is based on the ammonites of the early Pliensbachian (Aenigmaticum Zone): Miltoceras tagendoufi El Hariri et al., 1996, and Metaderoceras gr. apertum El Hariri et al., 1996 (Ettaki, 2003; Ettaki et al., 2011). These species appear in association with Paleomayncina termieri and Pseudopfenderina butterlini. These biostratigraphic calibrations confirm and validate the indirect dating proposed by Septfontaine (1984, 2020) at the scale of the Moroccan Tethys domain.
4.4. Orbitopsella praecursor assemblage: lower Pliensbachian (Demonense and Dilectum zones).
The lower limit of this assemblage is characterized by the appearance of Orbitopsella praecursor. This index species is associated with Paleomayncina termieri and Planisepta compressa. The upper limit is marked by the abrupt disappearance of Orbitopsella praecursor. This is also confirmed in other regions of the Central High Atlas, particularly at its northern border (Souhel, 1996), and in the Middle Atlas (Septfontaine, 1984; Bassoullet et al., 1999). Other taxa, including Ammobaculites gr. coprolithiformis, Bosniella sp., and Everticyclammina sp., are associated with this assemblage, which corresponds perfectly to phylozone C2 of Septfontaine's (1984) biozonation.
4.5. Paleomayncina termieri and Planisepta compressa assemblage: upper Pliensbachian (Lavinianum Zone).
This interval zone is mainly characterized by the association of Paleomayncina termieri and Planisepta compressa. These two species are very abundant and are associated with Amijiella amiji, Bosniella croatica, Everticyclammina sp., and Haurania aff. deserta. The recorded algae are Cayeuxia sp. and Dasycladales. This assemblage is found in the inner platform lagoon-type facies of the Aganane Formation and in the Choucht 2 Formation. The lower limit is denoted by the disappearance of Orbitopsella praecursor, while its upper limit coincides with the appearance of Pseudocyclammina liasica. This assemblage correlates precisely with Septfontaine's (1984) biozone D and is assigned to the upper Pliensbachian (Lavinianum Zone).
4.6. Pseudocyclammina liasica assemblage or Palaeodasycladus mediterraneus assemblage: upper Pliensbachian (Algovianum Zone) - Toarcian
Within the upper Pliensbachian (Algovianum Zone) to Toarcian interval, two assemblages are defined according to facies type:
According to Septfontaine (1984, 2020), this biozone is defined by the presence of Pseudocyclammina liasica. This index species often coexists with Amijiella amiji and Haurania deserta. In the study area, except in the inner platform facies, we noted the scarcity, or even the absence of Pseudocyclammina liasica. In the Todrha-Dades High Atlas, P. liasica is associated, in some sections, with an open sea fauna of the lower to middle Toarcian, allowing its biostratigraphic calibration. Thus, the brachiopods collected at the base are Telothyris arnaudi and many juvenile forms of Telothyris jauberti var. depressa of the lower Toarcian (Levisoni Zone? and base of the Bifrons Zone), while at the top, the taxa identified are Homoeorhynchia batalleri and juvenile forms of Homoeorhynchia meridionalis (identified by Prof. Ouahhabi) indicating the middle Toarcian (Bifrons Zone) (Ettaki, 2003, p. 209-213; Ettaki et al., 2008).
Elsewhere, at several localities of the Atlasic domain and of the south Rifan ridges, Septfontaine's (1984) biozone E (i.e., range zone of Pseudocyclammina liasica) belongs to the upper Pliensbachian (Algovianum Zone). However, in the Todrha-Dades High Atlas, the new biostratigraphic data allow us to reasonably place the upper limit of the biozone within the middle Toarcian (Bifrons Zone).
Palaeodasycladus mediterraneus was first described by Pia in 1920 and subsequently revised by the same author in 1927. In 1967, Praturlon introduced the new variety P. mediterraneus var. elongatulus. Later, Deloffre and Laadila (1990) proposed an emended diagnosis for both the genus and its type species. Additionally, P. elongatulus was later validated in Granier and Deloffre (1993) under the name P. gracilis, which may in fact represent a junior synonym of P. mediterraneus. In their study, Barattolo et al. (1994) offered descriptions that diverged from Pia's original characterization of P. mediterraneus, providing a basis to critically assess the emendation proposed by Deloffre and Laadila (1990). However, in 2003, Granier et al. reinterpreted the original specimens of Deloffre and Laadila as Petrascula iberica, not as a species of Palaeodasycladus.
Other works on calcareous algae (Dasycladales) from the lower Lias, and more precisely from the Pliensbachian, particularly those referencing the genus Palaeodasycladus and its species, were published in the 2000s, notably by Sokač (2001) and Barattolo and Romano (2005). It is noteworthy that P. mediterraneus is widespread in the lower and middle Lower Jurassic of the Tethys domain.
In the Dades High Atlas, P. mediterraneus has been found in peri-reefal and reefal deposits of the Choucht 2 Formation (Ettaki, 2003).
In the peri-reef and reef deposits (Choucht
2 Formation). These facies are characterized by a high abundance of P.
mediterraneus (Fig. 4.G ![]() ). This alga is often associated with Haurania
deserta (in great abundance) and Amijiella
amiji. Other foraminifers, including Everticyclammina
sp., Ophthalmidium leischneri, and O.
martanum, as well as algae such as Cayeuxia piae,
and incertae sedis, are associated
with this characteristic assemblage.
). This alga is often associated with Haurania
deserta (in great abundance) and Amijiella
amiji. Other foraminifers, including Everticyclammina
sp., Ophthalmidium leischneri, and O.
martanum, as well as algae such as Cayeuxia piae,
and incertae sedis, are associated
with this characteristic assemblage.
This assemblage is biostratigraphically calibrated by the occurrence of an ammonite and brachiopod fauna (Ettaki et al., 2000a, 2000b; Ettaki, 2003).
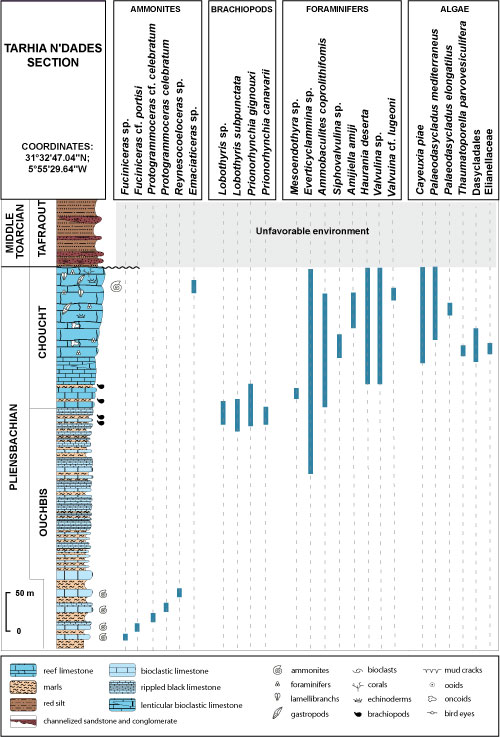
|
Figure 6:
Distribution
of fauna and algae in the Tarhia n'Dades section (after Ettaki,
2003, compiled). |
The
lower horizons can be attributed to the upper Pliensbachian (Algovianum
Zone) by the presence of the brachiopods: Lobothyris sp., L. subpunctata,
and Prionorhynchia gignouxi (Fig. 6 ![]() ). The upper part of the Palaeodasycladus
mediterraneus assemblage is attributed to upper
Pliensbachian (Emaciatum Zone) due
to the occurrence of Emaciaticeras (Ettaki
et al., 2000a, 2000b) (Figs. 7
). The upper part of the Palaeodasycladus
mediterraneus assemblage is attributed to upper
Pliensbachian (Emaciatum Zone) due
to the occurrence of Emaciaticeras (Ettaki
et al., 2000a, 2000b) (Figs. 7 ![]() - 8
- 8 ![]() ).
).
Consequently, at the Atlas domain scale, we propose to include the Palaeodasycladus mediterraneus assemblage, acme zone, in the reef facies to mark the upper Pliensbachian (Algovianum and Emaciatum zones). This P. mediterraneus assemblage can be correlated with the Pseudocyclammina liasica biozone of the reference biostratigraphic scheme developed by Septfontaine (1984, revised 2020), which seems characteristic of lagoonal facies.
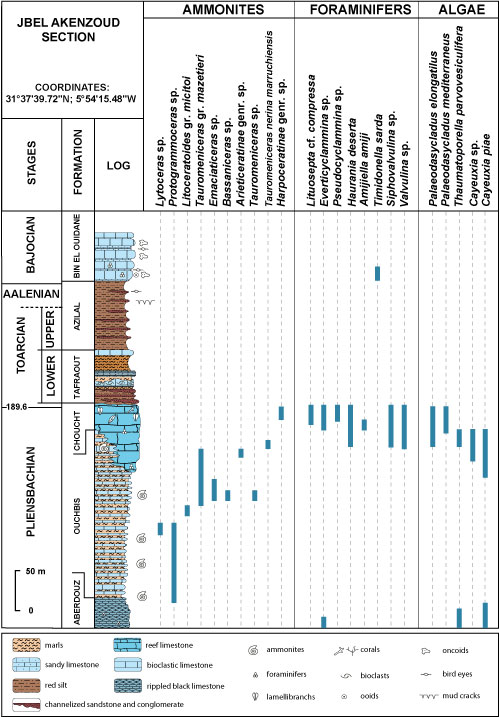
|
Figure 7:
Distribution of fauna and algae in the Jbel
Akenzoud section (after Ettaki, 2003, compiled). |
4.7. Timidonella sarda assemblage
This assemblage is characterized by the occurrence of typical forms of Timidonella sarda. It corresponds to Septfontaine's (1984) biozone F. The earliest occurrences of T. sarda are found within the carbonate beds at the top of the siliclastic Azilal Formation. At Ikerzi, the lower part of this formation yields narrow morphs of Stroudithyris stephanoides (identified by Prof. Ouahhabi), indicating the Aalensis Zone and the transition to the Opalinum Zone (Ettaki, 2003). Timidonella sarda has been found in other areas of the Central High Atlas. It characterizes the middle Aalenian - lower Bajocian interval (Jossen et al., 1987; Septfontaine et al., 1991). This is confirmed in several regions of the Tethys domain, where the stratigraphic distribution of T. sarda is observed within the Middle Jurassic (Bassoullet et al., 1976).
|
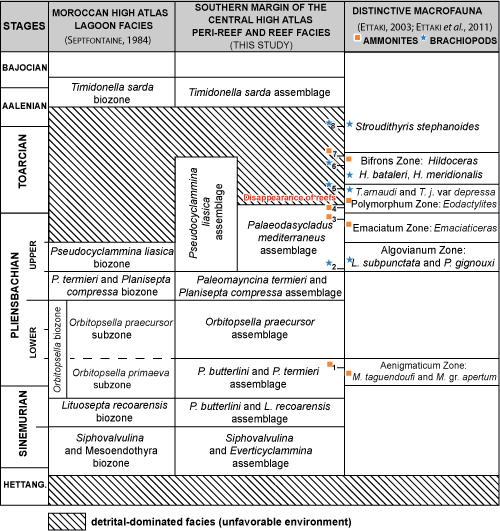
Figure 8:
Micropaleontological assemblages based on foraminifers and the calcareous algae
of the Lower and Middle Jurassic of the Central High Atlas. Numbers refer to
Ammonites and Brachiopods recovered in the southern Central High Atlas (after Ettaki,
2003, compiled): Ammonites, 1:
Miltoceras taguendoufi and
Metaderoceras gr. apertum; 3:
Emaciaticeras; 4: Eodactylites mirabilis; 7: Hildoceras;
Brachiopods, 5:
Telothyris arnaudi and
Telothyris jauberti var. depressa;
2:
Lobothyris subpunctata, Lobothyris
sp., and Priorhynchia gignouxi;
6:
Homeorhynchia batalleri and juvenile
formes of Homeorhynchia meridionalis; 8:
Stroudithyris stephanoides. |
The
biostratigraphic scheme proposed in this work results from the identification of
larger benthic foraminifers, calcareous algae and their associations, as well as
the stratigraphic context of these microfossils. It is the result of a
micropaleontological analysis of several stratigraphic sections studied on the
southern margin of the Central High Atlas. Seven
consecutive assemblages have been recognized based on the stratigraphic
distribution of foraminifers and algae. The
seven identified assemblages (Fig. 8 ![]() ) always occur in the same stratigraphic
order. We partially recognize the different biozones defined in 1984
by Septfontaine
and
revised in 2020; we present observations and provide additional details
regarding the stratigraphic age and succession of the assemblages. We
propose to introduce a new biostratigraphic subdivision: a Palaeodasycladus
mediterraneus assemblage, in the reef facies of the upper Pliensbachian (Algovianum
and Emaciatum zones). This
newly proposed assemblage can be correlated with the Pseudocyclammina
liasica biozone of Septfontaine (1984,
2020), as
defined within low energy lagoon-type facies.
) always occur in the same stratigraphic
order. We partially recognize the different biozones defined in 1984
by Septfontaine
and
revised in 2020; we present observations and provide additional details
regarding the stratigraphic age and succession of the assemblages. We
propose to introduce a new biostratigraphic subdivision: a Palaeodasycladus
mediterraneus assemblage, in the reef facies of the upper Pliensbachian (Algovianum
and Emaciatum zones). This
newly proposed assemblage can be correlated with the Pseudocyclammina
liasica biozone of Septfontaine (1984,
2020), as
defined within low energy lagoon-type facies.
It is noteworthy that Palaeodasycladus mediterraneus is a very common and widely distributed algal species. Its occurrence has been frequently documented in the literature within Jurassic carbonate facies. This species is widespread throughout the lower and middle Lower Jurassic of the Mesogean domain. Numerous studies have reported its presence in various regions, including Italy (Chiocchini & Mancinelli, 1978; Chiocchini et al., 1994, 2008; Barattolo et al., 1994), Spain (Sevillano et al., 2020), Croatia and Slovenia (Sokač, 2001), southern Iran (Bassoullet et al., 1978), as well as Algeria, Tunisia, Turkey, and Oman.
During the Pliensbachian, P. mediterraneus was a significant component of the algal biomass, thriving under favorable environmental conditions on carbonate platforms of the southern margin of the Tethys Ocean. The persistence of favorable environmental conditions sustained the presence of P. mediterraneus until the dislocation of carbonate platforms and the subsequent collapse of reef systems following the syn-Polymorphum crisis (Ettaki, 2003; Ettaki & Chellai, 2005). The disappearance of this microfossil reflects a shift in paleoenvironmental conditions, marking the transition from biostasis to rhexistasis, as observed in the Toarcian Stage (Ettaki & Chellai, 2005; Ettaki et al., 2007a, 2008, 2011). P. mediterraneus appears to be absent in the lower Toarcian (Serpentinus Zone) as well as in the middle and upper Toarcian. The predominantly siliciclastic facies that were deposited following the syn-Polymorphum crisis likely contributed to the absence of P. mediterraneus and larger benthic foraminifera (LBF).
The correlation of certain biozones or assemblages,
as well as some taxa, with the Jurassic stages has been calibrated either
through the association of stratigraphic markers (such as ammonites and
brachiopods) within the assemblages, or by their placement within well-dated
paleontological layers (Fig. 8 ![]() ). This approach enabled us to determine the age
of the Palaeodasycladus mediterraneus assemblage, as well as the Paleomayncina
termieri and Pseudopfenderina butterlini assemblage from Septfontaine's
(1984) biozone C1, and subsequently to define the age of the Pseudocyclammina
liasica biozone.
). This approach enabled us to determine the age
of the Palaeodasycladus mediterraneus assemblage, as well as the Paleomayncina
termieri and Pseudopfenderina butterlini assemblage from Septfontaine's
(1984) biozone C1, and subsequently to define the age of the Pseudocyclammina
liasica biozone.
The biostratigraphic subdivisions proposed contribute to a better stratigraphic knowledge of the southern margin of the Central High Atlas. This publication contributes to the discussion on the spatio-temporal distribution of benthic foraminiferal assemblages and calcareous algal associations throughout the entire Jurassic system, with the aim of establishing a high-resolution regional stratigraphic scale based on microfossils and making correlations with other regions of the Atlas domain and the broader Mediterranean region.
The late Prof. Y. Alméras (Claude Bernard Lyon 1 University) determined the brachiopods, and we gratefully acknowledge his valuable contribution to this study. We would like to express our gratitude to Prof. J.-L. Dommergues (University of Burgundy, Dijon) for determining the ammonite faunas, to Prof. B. Ouahhabi (Mohammed I University, Oujda) for determining the brachiopods, to Prof. L. Boudchich (Mohammed I University, Oujda) for identifying the foraminiferal taxa, and to Prof. M. Mehdi (Ibn Tofail University, Kenitra) for determining the calcareous algae. We are also grateful to Prof. B. Boudad and S. Ait Ami Said (Cadi Ayyad University, Marrakech) for their assistance in producing the figures.
We thank the reviewers, Prof. Ioan I. Bucur and one anonymous reviewer, for their comments and criticisms, which helped to improve the manuscript. Our sincere thanks also go to Dr. Mike Simmons for the time he dedicated to reviewing the first version of this paper. His constructive feedback and the rigor of his analysis greatly contributed to enhancing the quality of this work.
Barattolo F., De Castro P. & Parente M. (1994).- Some remarks on the genera Palaeodasycladus (Pia, 1920) Pia, 1927 and Eodasycladus Cros & Lemoine, 1966 ex Granier & Deloffre, 1994 (Green Algae, Dasycladales).- Beiträge zür Paläontologie, Wien, no. 19, p. 1-11. URL: https://www.zobodat.at/pdf/Beitr-Palaeontologie_19_0001-0011.pdf
Barattolo F. & Romano R. (2005).- Shallow carbonate platform bioevents during the Upper Triassic-Lower Jurassic: An evolutive interpretation.- Bollettino della Societŕ Geologica Italiana, Roma, vol. 124, p. 123-142.
Bassoullet J.-P. (1997a).- Foraminifčres. Les grands foraminifčres. In: Cariou E. & Hantzpergue P. (eds.), Biostratigraphie du Jurassique ouest-européen et méditerranéen. Zonations parallčles et distribution des invertébrés et microfossiles.- Bulletin du Centre de Recherches elf Exploration Production, Mémoire, Pau, no. 17, p. 293-304.
Bassoullet J.-P. (1997b).- Algues dasycladales. Distribution des principales espčces. In: Cariou E. & Hantzpergue P. (eds.), Biostratigraphie du Jurassique ouest-européen et méditerranéen. Zonations parallčles et distribution des invertébrés et microfossiles.- Bulletin du Centre de Recherches elf Exploration Production, Mémoire, Pau, no. 17, p. 339-342.
Bassoullet J.-P., Bernier P., Conrad M.A., Deloffre R. & Jaffrezo M. (1978).- Les algues dasycladales du Jurassique et du Crétacé.- Géobios, Villeurbanne, Mémoire spécial 2, 330 p. (40 Pls.).
Bassoullet J.-P., Boutakiout M. & Echarfaoui H. (1999).- Deux nouveaux genres, Palaeocyclammina et Ijdranella, foraminifčres (Textulariina) d'un niveau liasique ŕ Orbitopsella praecursor (Gümbel) du Moyen Atlas (Maroc).- Revue de Micropaléontologie, Paris, vol. 42, no. 3, p. 231-230.
Bassoullet J.-P., Chabrier G. & Fourcade É. (1976).- Données complémentaires sur la morphologie, la structure interne et la position stratigraphique de Timidonella sarda (Foraminifčre, Lituolidae).- Revue de Micropaléontologie, Paris, vol. 19, no. 1, p. 3-18.
Bernasconi R. (1983, unpublished).- Géologie du Haut-Atlas de Rich (Maroc).- Thčse de Doctorat čs Sciences, Université de Neuchâtel, 107 p.
Chiocchini M. & Mancinelli A. (1978).- Ricerche geologiche sul Mesozoico del Gran Sasso d'Italia (Abruzzo). III. Correlazioni microbiostratigrafiche tra facičs di margine della piattaforma carbonatica e facies pelagiche del Giurassico e Cretaceo inferiore.- Studi geologici Camerti, Comerino, vol. IV, p. 19-36.
Chiocchini M., Farinacci A., Mancinelli A., Molinari V. & Potetti M. (1994).- Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche mesozoiche dell'Appennino centrale (Italia).- Studi Geologici Camerti, Camerino, vol. speciale, parte A: "Biostratigrafia dell'Italia centrale", p. 9-128. URL: http://193.204.8.201:8080/jspui/handle/1336/483
Chiocchini M., Chiocchini R.A., Didaskalou P. & Potetti M. (2008).- Ricerche micropaleontologiche e biostratigrafiche sul Mesozoico della piattaforma carbonatica laziale-abruzzese (Italia centrale).- Memorie per servire alla Descrizione della Carta Geologica d'Italia, Roma, vol. 84 p. 5-170.
Clark G.N. & Boudagher-Fadel M. (2004).- Larger benthic foraminifera and calcareous algae of the Upper Kesrouane Limestone Formation (Middle/Upper Jurassic) in Central Lebanon: Stratigraphy, sedimentology and regional synopsis.- Revue de Paléobiologie, Genčve, vol. 23, no. 2, p. 477-504.
Deloffre R. & Laadila M. (1990).- Un niveau exceptionnel ŕ Palaeodasycladus mediterraneus dans le Lotharingien du Moyen-Atlas (Maroc) - émendation du genre et de l'espčce-type.- Revue de Paléobiologie, Genčve, vol. 9, no. 2, p. 309-320.
Dresnay R. du (1971).- Extension et développement des phénomčnes récifaux jurassiques dans le domaine atlasique marocain, particuličrement au Lias moyen.- Bulletin de la Société Géologique de France (7e série), Paris, t. XIII, fasc. 1-2, p. 46-56.
Dresnay R. du (1977).- Le milieu récifal fossile du Jurassique inférieur (Lias) dans le domaine des chaînes atlasiques du Maroc. In: Second symposium international sur les coraux et récifs coralliens fossiles, Paris 1975.- Mémoire BRGM, Orléans, no. 89, p. 296-312.
Dresnay R. du (1979).- Sédiments jurassiques du domaine des chaînes atlasiques du Maroc. In: Symposium "Sédimentation jurassique Ouest-Européenne".- Association des Sédimentologistes Français, Publication spéciale, Paris, vol. 1, p. 345-365.
Dresnay R. du, Termier G. & Termier H. (1978).- Les Héxactinellides (Lyssakides et Dictionines) du Lias marocain.- Géobios, Villeurbanne, vol. 11, no. 3, p. 269-295.
Dubar G. (1952).- Haut-Atlas central. In: Livret-guide de l'excursion A34.- XIXe Congrčs géologique international, Alger, no. 4, 74 p. (3 Pls.).
El Hariri K. (1998).- Le signal morphologique de l'évolution chez les ammonites du Lias inférieur et moyen dans les contextes stratigraphique, paléogéographiques et paléoécologique du Haut-Atlas Central (Maroc).- Thčse d'État čs-Sciences, Université Cadi Ayyad, Marrakech, 180 p.
Ettaki M. (2003, unpublished).- Étude sédimentologique et stratigraphique du Lias-début du Dogger de la région de Todrha-Dadčs (versant sud du Haut-Atlas Central, Maroc) - Implications géodynamiques.- Thčse de Doctorat, Université Cadi Ayyad, Marrakech, 429 p.
Ettaki M. & Chellai E.H. (2005).- Le Toarcien inférieur du Haut-Atlas de Todrha-Dadčs (Maroc) : Sédimentologie et lithostratigraphie.- Comptes Rendus Géoscience, vol. 337, no. 9, p. 814-823. DOI: 10.1016/j.crte.2005.04.007
Ettaki M., Chellai E.H., Milhi A., Sadki D. & Boudchiche L. (2000a).- Le passage Lias moyen-Lias supérieur dans la région de Todrha-Dadčs : Événements bio-sédimentaires et géodynamiques (Haut-Atlas Central, Maroc).- Comptes Rendus de l'Académie des Sciences (Series IIA - Earth and Planetary Science), Paris, vol. 331, no. 10, p. 667-674.
Ettaki M., Ibouh H. & Chellai E.H. (2007).- Événements tectono-sédimentaires au Lias-Dogger de la frange méridionale du Haut-Atlas Central, Maroc.- Estudios Geológicos, Madrid, no. 63, no. 2, p. 103-125. DOI: 10.3989/egeol.07632196
Ettaki M., Ibouh H., Chellai E.H. & Milhi A. (2007b).- Les structures "diapiriques" liasiques du Haut-Atlas central, Maroc : L'exemple de la ride d'Ikerzi.- Africa Geoscience Review, Paris, vol. 14, no. 1, p. 79-93.
Ettaki M., Milhi A., Chellai E.H., Boudchiche L., & Sadki D. (2000b).- Mise en évidence de la limite Pliensbachien-Toarcien par les ammonites, les foraminifčres et l'interaction tectono-eustatique dans la région de Todrha-Dadčs (Haut-Atlas Central, Maroc).- Revue de Paléobiologie, Genčve, vol. 19, no. 2, p. 299-317.
Ettaki M., Ouahhabi B., Dommergues J.L., Meister C. & Chellai E.H. (2011).- Analyses biostratigraphiques dans le Lias de la bordure sud de la Téthys méditerranéenne : L'exemple de la frange méridionale du Haut-Atlas Central (Maroc).- Bulletin de la Société Géologique de France, Paris, vol. 182, no. 6, p. 523-534.
Ettaki M., Sadki D., Chellai E.H. & Milhi A. (2008).- Interaction eustatisme-tectonique dans le contrôle de la sédimentation au Jurassique inférieur et moyen dans la région de Todrha-Dadčs, versant sud du Haut-Atlas Central, Maroc.- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 527, p. 55-64.
Ettaki M., Sadki D., Milhi A. & Hadri M. (1996).- Les formations lithostratigraphiques jurassiques du Haut-Atlas central marocain. Essai de synthčse. In: 13e Colloque Bassins Sédimentaires Marocains.- Université Cadi Ayyad, Marrakech, Résumé, p. 111-112.
Evans L. & Kendall E.G. (1977).- An interpretation of the depositional setting of some deep-water Jurassic carbonates of the central High Atlas mountains, Morocco. In: Cook H.E. & Enos P. (eds.), Deep water carbonate environments.- SEPM Special Publication, Tulsa - OK, vol. 25, p. 249-261.
Gale L. (2014).- Lower Jurassic foraminiferal biostratigraphy of Podpeč Limestone (External Dinarides, Slovenia).- Geologija, Ljubljana, vol. 57, no. 2, p. 119-146. DOI: 10.5474/geologija.2014.011
Granier B.R.C. (2021).- Bacinella, a discrete type of Mesozoic calcimicrobial structure.- Carnets Geol., Madrid, vol. 21, no. 1, p. 1-25. DOI: 10.2110/carnets.2021.2101
Granier B. & Deloffre R. (1993).- Inventaire critique des algues dasycladales fossiles. IIe partie - les algues dasycladales du Jurassique et du Crétacé.- Revue de Paléobiologie, Genčve, vol. 12, p. 19-65.
Granier B., Poisson A., Ferré B. & Deloffre R. (2003).- New data on Petrascula iberica Dragastan et Trappe 1986, nov. comb., a Liassic club-shaped Dasycladacean. Preliminary note. In: Yuan X. & Mu X. (eds.), Fossil algae and stromatolites.- Acta Micropalaeontologica Sinica, Nanjing, vol. 20, no. 1, p. 67-74.
Hallock P. & Glenn E.C. (1986).- Larger foraminifera: A tool for paleoenvironmental analysis of Cenozoic carbonate depositional facies.- Palaios, Lawrence - KS, vol. 1, no. 1, p. 55-64.
Hottinger L. (1967).- Foraminifčres imperforés du Mésozoďque marocain.- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 209, 168 p.
Jenny J. (1985).- Carte géologique du Maroc au 1/100 000, feuille Azilal.- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 339.
Jenny J. (1988).- Mémoire explicatif de la carte géologique du Maroc au 1/100.000 (feuille d'Azilal, Haut-Atlas central).- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 339 bis, 104 p.
Jossen J.A. (1990).- Carte géologique du Maroc au 1/100.000. Feuille de Zawat ahançal (Haut-Atlas central).- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 355.
Jossen J.A., Milhi A. & Septfontaine M. (1987).- L'horizon repčre ŕ Timidonella sarda de la plate-forme carbonatée du Haut Atlas central au Dogger. Valeur stratigraphique. In: Xe colloque africain de micropaléontologie, Rabat 1987.- Géologie Méditerranéenne, Marseille, t. XIV, no. 2, p. 171. URL: https://www.persee.fr/doc/geolm_0397-2844_1987_num_14_2_1377
Kabal Y. & Tasli K. (2003).- Biostratigraphy of the Lower Jurassic carbonates from the Aydinick area (Central Taurides, S. Turkey) and morphological analysis of Lituolipora termieri (Hottinger, 1967).- Journal of Foraminiferal Research, Lawrence - KS, vol. 33, no. 4, p. 338-351.
Kooistra W.H., Coppejans E. G. & Payri C. (2002).- Molecular systematics, historical ecology, and phylogeography of Halimeda (Bryopsidales).- Molecular Phylogenetics and Evolution, vol. 24, no. 1, p. 121-138.
Lachkar N. (2000).- Dynamique sédimentaire d'un bassin extensif sur la marge sud-téthysienne : Le Lias du Haut-Atlas de Rich (Maroc).- Thčse de Doctorat, Université de Bourgogne, Dijon, 274 p.
Le Marrec A. & Jenny J. (1980).- L'accident de Demnat, comportement synsédimentaire et tectonique d'un décrochement transversal du Haut-Atlas central (Maroc).- Bulletin de la Société Géologique de France (7e série), Paris, t. XXII, fasc. 3, p. 421-427.
Milhi A. (1992).- Les facičs du Lias de la région du Todrha (Tinerhir, versant sud du Haut-Atlas Central, Maroc).- Geologische Rundschau, vol. 81, p. 63-68.
Milhi A. (1997).- Carte géologique du Maroc au 1/100.000. Feuille de Tinerhir (Haut-Atlas Central).- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 377.
Milhi A., Ettaki M., Chellai E.H. & Hadri M. (2002).- Les formations lithostratigraphiques jurassiques du Haut-Atlas Central Marocain : Corrélations et reconstitutions paléogéographiques.- Revue de Paléobiologie, Genčve, vol. 19, no. 2, p. 299-317.
Praturlon A. (1966).- Algal assemblages from Lias to Paleocene in Southern Latium-Abruzzi: A review.- Bollettino della Societŕ geologica italiana, Roma, vol. 85, p. 167-194.
Reolid M., Nagy J., Rodriguez-Tovar F.J. & Oloriz F. (2008).- Foraminiferal assemblages as palaeoenvironmental bioindicators in Late Jurassic epicontinental platforms: Relation with trophic conditions.- Acta Palaeontologica Polonica, Warszawa, vol. 53, no. 4, p. 705-722. DOI: 10.4202/app.2008.0413
Roch E. (1939).- Description géologique des montagnes ŕ l'Est de Marrakech.- Notes et Mémoires du Service Géologique du Maroc, Rabat, no. 51, 438 p.
Russo P. & Russo L. (1934).- Le grand accident sud-atlasien.- Bulletin de la Société Géologique de France (5e série), Paris, t. IV, fasc. 4, p. 375-384 (Pl. XX).
Sadki D. (1996).- Le Haut-Atlas Central (Maroc) : Stratigraphie et paléontologie du Lias supérieur et du Dogger inférieur. Dynamique du Bassin et des peuplements.- Document des Laboratoires de Géologie de Lyon, Villeurbanne, no. 142, 312 p. URL: http://www.persee.fr/doc/geoly_0750-6635_1996_mon_142_1
Schlagintweit F., Bover-Arnal T. & Salas R. (2011).- New insights into Lithocodium aggregatum Elliott 1956 and Bacinella irregularis Radoičić 1959 (Late Jurassic-Lower Cretaceous): Two ulvophycean green algae (?Order Ulotrichales) with a heteromorphic life cycle (epilithic/euendolithic).- Facies, Erlangen, vol. 56, no. 4, p. 509-547.
Septfontaine M. (1984).- Biozonation (ŕ l'aide des foraminifčres imperforés) de la plateforme interne carbonatée liasique du Haut-Atlas (Maroc). Revue de Micropaléontologie, Paris, vol. 27, no. 3, p. 209-229.
Septfontaine M. (1985).- Milieux de dépôt et foraminifčres (Lituolidés) de la plate-forme carbonatée du Lias moyen au Maroc.- Revue de Micropaléontologie, Paris, vol. 28, no. 4, p. 265-289.
Septfontaine M. (2020).- Steps of morphogenesis and iterative evolution of imperforate larger Foraminifera in shallow carbonate shelves during Mesozoic times: Possible relations to symbiotic and abiotic factors.- Chapter 8. In: Guex J., Torday J.S. & Miller W.B. Jr (eds.), Morphogenesis, environmental stress and reverse evolution.- Springer Nature Switzerland, Cham, p. 129-173.
Septfontaine M., Arnaud-Vanneau A., Bassoullet J.-P., Gušić Y., Ramalho M. & Velić I. (1991).- Les foraminifčres imperforés des plates-formes carbonatées jurassiques : État des connaissances et perspectives d'avenir.- Bulletin de la Société vaudoise des Sciences naturelles, Lausanne, Band 80, Heft 3, p. 255-277.
Sevillano A., Septfontaine M., Rosales I., Barnolas A., Bádenas B. & López-García J.M. (2020).- Lower Jurassic benthic foraminiferal assemblages from shallow-marine platform carbonates of Mallorca (Spain): Stratigraphic implications.- Journal of Iberian Geology, vol. 46, p. 77-94.
Sokač B. (2001).- Lower and middle Liassic calcareous algae (Dasycladales) from Mt. Velebit (Croatia) and Mt. Trnovski Gozd (Slovenia) with particular reference to the genus Palaeodasycladus (Pia, 1920) 1927 and its species.- Geologia Croatica, Zagreb, vol. 54, no. 2, p. 133-257 (42 Pls.). URL: https://www.geologia-croatica.hr/index.php/GC/article/view/GC.2001.07
Souhel A. (1996).- Le Mésozoďque dans le Haut-Atlas de Béni-Mellal (Maroc). Stratigraphie, sédimentologie et évolution géodynamique.- Strata (Série 2), Gaillac, no. 27, 249 p.
Stüder M. (1980).- Tectonique et pétrographie des roches sédimentaires, éruptives et métamorphiques de la région de Tounfite-Tirrhist (Haut Atlas central, Maroc).- Thčse de Doctorat, Université Neuchâtel, 102 p. (Tome 1: 95 p.; Tome 2: Illustrations). URL: https://libra.unine.ch/handle/123456789/33724
Velić I. (2007).- Stratigraphy and palaeobiogeography of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe).- Geologia Croatica, Zagreb, no. 60, p. 1-113. URL: https://hrcak.srce.hr/21887
Microfossils
Amijiella amiji (Henson)
Ammobaculites coprolithiformis (Schwager)
Bosniella croatica (Gušić)
Cayeuxia kurdistanensis Elliott
C. mediterranea Herak
C. piae Rech-Frollo
Chinianella ellembergeri (Lebouché & M. Lemoine)
Haurania deserta Henson
Involutina liassica (Jones)
Lingulina gr. tenera Bornemann
Lituosepta cf. compressa (Hottinger)
L. recoarensis Cati
Ophthalmidium leischneri (Kristan-Tollmann)
O. martanum (Farinacci)
Orbitopsella praecursor (Gümbel)
O. primaeva (Henson)
Palaeodasycladus elongatus Praturlon
P. gracilis Cros & M. Lemoine
P. mediterraneus (Pia)
P. mediterraneus var. elongatulus Praturlon
Paleomayncina termieri (Hottinger)
Petrascula iberica (Dragastan & Trappe, 1986)
Planisepta compressa (Hottinger)
Pseudocyclammina liasica (Hottinger)
Pseudopfenderina butterlini (Brun)
Redmondoides lugeoni (Septfontaine)
Tersella alpina Cros & Lemoine
Thaumatoporella parvovesiculifera (Raineri)
Timidonella sarda Bassoullet et al.
Uragiella liasica (Lebouché & M. Lemoine)
Verneuilinoides mauritii (Terquem)
Ammonites
Metaderoceras gr. apertum El Hariri et al.
Miltoceras taguendoufi El Hariri et al.
Brachiopods
Homoeorhynchia batalleri (Dubar)
H. meridionalis (Deslongchamps)
Lobothyris subpunctata (Davidson)
Prionorhynchia canavarii (Jiménez de Cisneros)
Pr. gignouxi (Jiménez de Cisneros)
Stroudithyris stephanoides Alméras & Moulan
Telothyris arnaudi Alméras-Moulan
T. jauberti (Deslongchamps)
T. jauberti var. depressa Dubar