◄ Carnets Geol. 24 (14) ►
![]()
Outline
[1. Introduction]
[2. Geographic and geological setting]
[3. Repository of the material studied]
[4. Systematics]
[5.Conclusions]
[Bibliographic references] and ...
[Plates]
Lerchenauerstr. 167, D-80935 München (Germany)
Museu de Ciències Naturals de Barcelona (Paleontologia), Passeig Picasso s/n, 08003 Barcelona (Spain)
Department of Geology, Yazd University, 89195-741 Yazd (Iran)
Dipartimento di Fisica e Scienze della Terra, Università di Ferrara, via
Saragat 1, 44121 Ferrara (Italy)
Istituto di Scienze Marine (CNR-ISMAR), Area Science Park, Q2 Building, Basovizza, Trieste (Italy)
Published online in final form (pdf) on November 7, 2025
DOI 10.2110/carnets.2025.2514
![]()
[Editor: Bruno
R.C. Granier; language editor: Zev Brook]
![]()
Based on new material from the middle to upper Cenomanian Natih Formation (members A and B) from Oman, the little-known species Gusicella minima (Henson), originally described from the Mishrif Formation of Qatar (Dukhan wells) is redescribed. Late Maastrichtian specimens from the Tarbur Formation of SW Iran previously assigned to the Cenomanian species display a more complex marginal zone and are therefore introduced as Gusicella complexa Schlagintweit & Rashidi sp. nov. Representatives of Gusicella have so far only been reported from the Arabian Plate, showing its endemic character. There is no record of the genus so far from the Turonian to early Maastrichtian interval.
• Late Cretaceous;
• Larger Benthic Foraminifera;
• Orbitolinidae;
• systematics;
• biostratigraphy
Schlagintweit F., Vicedo V., Rashidi K., Frijia G. & Consorti L. (2025).- Gusicella Schlagintweit & Rashidi, 2021 (type species Dictyoconella minima Henson, 1948), Upper Cretaceous Larger Benthic Foraminifer and its species.- Carnets Geol., Madrid, vol. 25, no. 14, p. 283-305. DOI: 10.2110/carnets.2025.2514
Gusicella Schlagintweit & Rashidi, 2021 (espèce-type Dictyoconella minima Henson, 1948), Grand Foraminifère Benthique du Crétacé supérieur et ses représentants.- À partir d'un nouvel échantillonnage de la Formation Natih (membres A et B), Cénomanien moyen à supérieur, en Oman, l'espèce peu connue Gusicella minima (Henson), initialement décrite dans la Formation Mishrif au Qatar (forages pétroliers de Dukhan), est redécrite. Des spécimens du Maastrichtien terminal provenant de la Formation Tarbur, dans le sud-ouest de l'Iran, précédemment attribués à l'espèce cénomanienne, présentent une zone marginale plus complexe et sont donc décrits ici sous le nom de Gusicella complexa Schlagintweit & Rashidi sp. nov. Les représentants du genre Gusicella n'ont jusqu'à présent été signalés que sur la plaque arabique, ce qui témoigne de son caractère endémique. Aucun signalement de ce genre n'existe pour l'intervalle allant du Turonien au Maastrichtien inférieur.
• Crétacé supérieur ;
• Grands Foraminifères Benthiques ;
• Orbitolinidae ;
• systématique ;
• biostratigraphie
In his monograph on Mesozoic and Cenozoic Larger Benthic Foraminifera (LBF) from the area of the Middle East and south-western Asia, Henson (1948) erected the genus Dictyconella for more or less 'compressed-conical' dictyoconids including D. complanata (type species) and D. minima. The latter was described based on a few specimens from Cenomanian strata of Qatar. Both species are, however, structurally completely different, e.g., aligned main partitions in the latter versus alternating structural elements in the former, and together cannot belong either to the same genus nor the same subfamily (Dictyoconinae versus Dictyorbitolininae) (Schlagintweit & Rashidi, 2021; Schlagintweit, 2022). Therefore, the new genus Gusicella was erected by Schlagintweit and Rashidi (2021) with the type species Dictyoconella minima, resulting in the new combination Gusicella minima. Alas, it now turns out that the illustrations of G. minima that were then displayed do not correspond to the species but to a more complex species of the genus. In the mentioned work, specimens showing principally equivalent features (compressed test, aligned partitions) from the upper Maastrichtian Tarbur Formation of SW Iran were considered to belong to the Cenomanian G. minima, thus exhibiting a long stratigraphic range and surviving the Cenomanian/Turonian boundary extinction event (e.g., Parente et al., 2008). This observation echoes a remark by Henson (1948, p. 26) that "a single specimen, possibly of this species [Dictyoconella minima], was observed at Dohuk, Northern Iraq, in Maestrichtian limestones with Orbitoides media d'Arch. and Omphalocyclus macropora (Lmk.)." New material from the Cenomanian of Oman that is in conformity with Henson's taxon 'Dictyoconella minima' clearly shows, however, that it displays a more primitive character, namely a less complex marginal zone, than the Maastrichtian species. The present contribution provides a redescription of the poorly known Cenomanian Gusicella minima from the Cenomanian Natih Formation of Oman and the establishment of a new species for the upper Maastrichtian form from the Iranian Tarbur Formation as G. complexa sp. nov.
2.1 Cenomanian Natih Formation, Oman
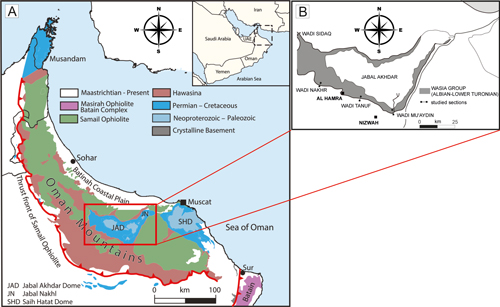
The Natih Formation, of late Albian to middle Turonian age (Philip et al., 1995; Wohlwend et al., 2016; Bromhead et al., 2022), is part of a large Tethys carbonate platform system developed during the Permian-Turonian interval in the Arabian Peninsula. The studied material from Oman was collected from the Natih Formation in three locations.
Wadi
Tanuf (GPS: 23°03'11"N, 57°28'02"E) and Wadi Nakhr
(GPS: 23°09'08"N, 57°12'08"E) are
located in the southern part of Jabal Akhdar mountain in northern Oman,
approximately 150 kilometers from the capital, Muscat (Fig.
1 ![]() ). The studied material
comes from the Natih A informal member which has been dated as late Cenomanian
to middle Turonian (Meister & Piuz,
2015; Wohlwend et al., 2016; Bromhead et
al.,
2022). However, the specimens of Gusicella
minima were found in the lower part of the Natih A of late Cenomanian age
according to biostratigraphic and C-isotope stratigraphic data (Meister
& Piuz, 2015; Wohlwend et
al., 2016; Cardelli,
2024). In the bioclastic packstones, G.
minima is associated with Pararotalia
boixae Piuz & Meister, Pseudorhipidionina casertana
(De Castro), Decastroia sp.,
and Qataria dukhani Henson (?Pastrikella)
sp.
). The studied material
comes from the Natih A informal member which has been dated as late Cenomanian
to middle Turonian (Meister & Piuz,
2015; Wohlwend et al., 2016; Bromhead et
al.,
2022). However, the specimens of Gusicella
minima were found in the lower part of the Natih A of late Cenomanian age
according to biostratigraphic and C-isotope stratigraphic data (Meister
& Piuz, 2015; Wohlwend et
al., 2016; Cardelli,
2024). In the bioclastic packstones, G.
minima is associated with Pararotalia
boixae Piuz & Meister, Pseudorhipidionina casertana
(De Castro), Decastroia sp.,
and Qataria dukhani Henson (?Pastrikella)
sp.
It is worth mentioning that the sections of the Natih Formation exposed in Wadi Mu'Aydin and Wadi Nakhr were previously also studied by Buchem et al. (1996, Fig. 1).
Wadi Mu'Aydin, Oman Mountains (GPS:
22°57'16"N, 57°40'04"E) is located about 115 kilometers southwest of Muscat (Fig.
1 ![]() ). For the lithostratigraphic log of the Wadi
Mu'Aydin section, see Piuz and Vicedo
(2020, Fig. 2). The
microfacies corresponds in most respects principally to the samples obtained
from Wadi Tanuf and Wadi Nakhr. The associated taxa include Rotorbinella
mesogeensis Tronchetti, Decastroia
oblonga Vicedo & Piuz, Qataria
dukhani Henson, Edomia reicheli
Henson, Everticyclammina hensoni
Redmond, Pastrikella sp., Cuneolina sp., Involutinida indet., and Nummoloculinodonta akhdarensis Piuz & Vicedo. The
facies also contains other fossils such as rudists from the Radiolitidae group (e.g.,
Eoradiolites? sp.) and questionable
bryozoans. The samples with Gusicella
minima are from the Natih A and B members of middle to late Cenomanian age.
). For the lithostratigraphic log of the Wadi
Mu'Aydin section, see Piuz and Vicedo
(2020, Fig. 2). The
microfacies corresponds in most respects principally to the samples obtained
from Wadi Tanuf and Wadi Nakhr. The associated taxa include Rotorbinella
mesogeensis Tronchetti, Decastroia
oblonga Vicedo & Piuz, Qataria
dukhani Henson, Edomia reicheli
Henson, Everticyclammina hensoni
Redmond, Pastrikella sp., Cuneolina sp., Involutinida indet., and Nummoloculinodonta akhdarensis Piuz & Vicedo. The
facies also contains other fossils such as rudists from the Radiolitidae group (e.g.,
Eoradiolites? sp.) and questionable
bryozoans. The samples with Gusicella
minima are from the Natih A and B members of middle to late Cenomanian age.
|
|
Figure 1:
(A) Geological overview map of the Oman
Mountains and the Fanja map area after Forbes et al.
(2010) and Callegari et al.
(2022). (B) Close-up map of the Jabal Akhdar dome with the
sampled locations of Gusicella minima (Henson). |
2.2 Late Maastrichtian Tarbur Formation, SW Iran
The material of Gusicella complexa Schlagintweit & Rashidi sp. nov. comes from two sections: the Mandegan and Naghan sections. Various new LBF taxa have already been described from these sections; some references for further reading are provided here (Schlagintweit et al., 2016a, 2016b; Schlagintweit & Rashidi, 2016, 2021).
The Naghan section is located in the folded Zagros belt of southwestern Iran, approximately 50 km southwest of Naghan town near Gandomkar village. The geographical coordinates of the Naghan section base are 31°47'52"N 50°32'53"E. The Mandegan section is situated in the High Zagros Belt, north of Mount Dena, about 65 km south of the town of Semirom (31°25'8.13"N, 51°24'34.58"E). In both sections, G. complexa has a rather wide vertical distribution associated with a variety of LBF such as Loftusia, Omphalocyclus, Gyroconulina columellifera Schroeder & Darmoian, Dicyclina schlumbergeri Munier-Chalmas, Minouxia sp., rotaliids, Palaeoelphidium multiscissuratum (Smout), Pseudorbitolina schroederi Luger, and Dictyoconus bakhtiari Schlagintweit et al. Gusicella complexa occurs preferentially in the lower part of the Tarbur Formation which, evolving from a shallowing-upwards sequence from basinal facies of the Gurpi Formation, reflects an outer platform (ramp) setting.
Gusicella minima: The material figured in this paper from Wadi Mu'Aydin, Oman is housed in the micropaleontological collections of the 'Museu de Ciències Naturals de Barcelona' (MGB numbers) with each specimen having a separate number. The material from Wadi Tanuf (illustrated specimens) and Wadi Nakhr is stored in the University collection of Ferrara. The numbers indicated in the plates refer to the thin-section number.
Gusicella complexa: All the specimens illustrated here refer to Schlagintweit and Rashidi (2021). The holotype of the new species is deposited at the Geosciences Museum of Mashhad (in the Geological Survey of North-Iran East territory) labelled Gmm1404-1 referring to the original sample number 2Ng 168.
Phylum Foraminifera Orbigny, 1826
Class Globothalamea Pawłowski et al., 2013
Order Loftusiida Kaminski & Mikhalevich in Kaminski, 2004
Suborder Loftusiina Kaminski & Mikhalevich in Kaminski, 2004
Superfamily Orbitolinoidea Martin, 1890
Family Orbitolinidae Martin, 1890
Subfamily Dictyorbitolininae Schroeder in Schroeder et al., 1990
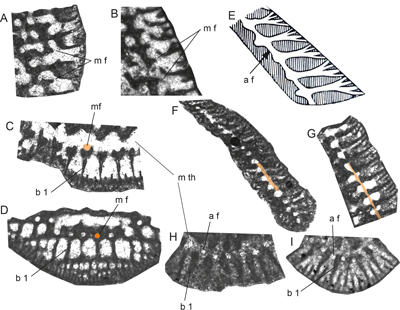
Remarks: Dicytorbitolininids typically include high-conical Orbitolinidae
displaying foramina arranged in a ring at the outer margin of the central zone
towards the marginal zone, with an alignment paralleling the vertical main
partitions (beams) (Schroeder in Schroeder et al.,
1990, p. 196)
(Fig. 2.E-I ![]() ). The aligned beams are
usually well constrained in tangential sections, whereas the peculiar
arrangement of the foramina is often less clear, requiring well-oriented
sections (axial and perpendicular to the cone axis). They are arranged
perpendicular to the septum ("verticaux
par rapport au plancher"), in continuity paralleling the cone mantel line
(e.g., Hottinger, 2006, Fig.
80.E) and were named marginal apertures ("ouvertures
marginales") by Schroeder in Schroeder et
al. (1990, p. 196) (Fig. 2.G
). The aligned beams are
usually well constrained in tangential sections, whereas the peculiar
arrangement of the foramina is often less clear, requiring well-oriented
sections (axial and perpendicular to the cone axis). They are arranged
perpendicular to the septum ("verticaux
par rapport au plancher"), in continuity paralleling the cone mantel line
(e.g., Hottinger, 2006, Fig.
80.E) and were named marginal apertures ("ouvertures
marginales") by Schroeder in Schroeder et
al. (1990, p. 196) (Fig. 2.G ![]() ). They differ from the marginal apertures (foramina)
of the Paleogene taxa which are arranged obliquely to the septum (i.e.,
"about 45o with the mantel line of the
cone") (Hottinger
& Drobne, 1980) (Fig. 2.A-B
). They differ from the marginal apertures (foramina)
of the Paleogene taxa which are arranged obliquely to the septum (i.e.,
"about 45o with the mantel line of the
cone") (Hottinger
& Drobne, 1980) (Fig. 2.A-B ![]() ). While the
'marginal apertures' of
the Dictyorbitolininae are in vertical continuity from chamber to chamber, they
alternate regularly from one chamber to the next in the Paleogene forms (Hottinger
& Drobne, 1980). In addition, they are laterally displaced to each
other, not in linear continuity, but also form a 'single circular row' as
discerned in transverse sections (Hottinger & Drobne,
1980, p.
211). Because of these fundamental structural differences, the term marginal
foramina (with oblique-radial orientation) should be restricted to the
Dictyoconinae (see also Hottinger, 2006, p. 22). For the
Dictyorbitolininae, it is instead proposed to call these 'aligned' apertures
or foramina. In the description of Pseudorbitolina
marthae (= P. schroederi Luger,
see Schlagintweit, 2022), Henson (1948, p. 104, Fig. 16) used the
name tubular apertures. Both types are positioned within a ring-shaped space
named the marginal trough by Davies (1930) or annular passage by Hottinger
(2006, p. 8). In both cases (Dictyorbitolininae and Dictyoconinae), the septa
(or beams?) are bent downwards (towards direction of the apex) at the transition
of the marginal zone to the marginal trough (Fig. 2.A, .G
). While the
'marginal apertures' of
the Dictyorbitolininae are in vertical continuity from chamber to chamber, they
alternate regularly from one chamber to the next in the Paleogene forms (Hottinger
& Drobne, 1980). In addition, they are laterally displaced to each
other, not in linear continuity, but also form a 'single circular row' as
discerned in transverse sections (Hottinger & Drobne,
1980, p.
211). Because of these fundamental structural differences, the term marginal
foramina (with oblique-radial orientation) should be restricted to the
Dictyoconinae (see also Hottinger, 2006, p. 22). For the
Dictyorbitolininae, it is instead proposed to call these 'aligned' apertures
or foramina. In the description of Pseudorbitolina
marthae (= P. schroederi Luger,
see Schlagintweit, 2022), Henson (1948, p. 104, Fig. 16) used the
name tubular apertures. Both types are positioned within a ring-shaped space
named the marginal trough by Davies (1930) or annular passage by Hottinger
(2006, p. 8). In both cases (Dictyorbitolininae and Dictyoconinae), the septa
(or beams?) are bent downwards (towards direction of the apex) at the transition
of the marginal zone to the marginal trough (Fig. 2.A, .G ![]() ). While marginal
foramina appear at the end of the forked radial main partition (Fig.
2.C-D
). While marginal
foramina appear at the end of the forked radial main partition (Fig.
2.C-D ![]() ),
the aligned foramina occupy a position at the tapering end of the chamberlets (between
two main partitions, b 1 in Fig. 2.I
),
the aligned foramina occupy a position at the tapering end of the chamberlets (between
two main partitions, b 1 in Fig. 2.I ![]() )
(Fig. 2.H-I
)
(Fig. 2.H-I ![]() ).
).
|
|
Figure 2:
Marginal foramina of the Dictyoconinae Schubert (A-D) compared to
so-called tubular foramina sensu Henson
(1948) of the Dictyorbitolininae Schroeder (E-I) (intentionally without
scale). (A-B) Marginal foramina in axial sections from Dictyoconus bakhtiari Schlagintweit et al. (upper Maastrichtian Iran) and Dictyoconus sp. cf. D. egyptiensis
(Chapman) (Eocene of Iran). (C) Slightly oblique transverse section with
row of marginal foramina located at the end of the forked main partitions (from Dictyoconus
bakhtiari Schlagintweit et al.,
upper Maastrichtian, Iran). (D) Oblique section (same as C). (E) Schematic
drawing of Pseudorbitolina with aligned foramina (modified from Henson,
1948, Fig. 16A). (F) Axial section of Pseudorbitolina
schroederi Luger (upper Maastrichtian, Iran). (G) Axial section of Gusicella
complexa sp. nov. (upper Maastrichtian, Iran). (H-I) Transverse sections of Gusicella minima (Henson) (upper
Cenomanian, Oman) and Gusicella
complexa sp. nov. (upper Maastrichtian, Iran) showing aligned foramina
arranged in a row at the end of the marginal chamberlets between the distal ends
of the radial main partitions. Fig. 2.I = excerpt from Pl. 6
|
With respect to the vertical structural elements (exoskeletal beams and endoskeleton pillars), Dictyorbitolininae include both taxa where all elements are aligned (genus Paracoskinolina Moullade, 1965) while in others just the beams are aligned with pillars alternating between the chambers (e.g., genera Dictyorbitolinina Cherchi & Schroeder, 1976, Praedictyorbitolinina Schroeder in Schroeder et al., 1990). This means that neither the structure of the embryo nor the disposition of the pillars represent a differentiating criterion as the former can be either simple or complex, and the latter either aligned or alternating from one chamber to the next.
Genus Gusicella Schlagintweit & Rashidi, 2021
Remarks: The genera Dictyoconella and Gusicella were already discussed in Schlagintweit et al. (2016a), Schlagintweit (2020), and Schlagintweit and Rashidi (2021).
The already noted similarities to Carinoconus Cherchi & Schroeder, 1982, are discussed in more details in the subchapter Comparisons for Gusicella minima.
In addition, some additional remarks are provided regarding the often dense, micritic central zone of Gusicella, the embryo, and the arrangement of the pillars. The patchy micritic parts within the central zone of Gusicella are referred to as 'thickened shell material' (Loeblich & Tappan, 1987, for Pfenderina), 'axial fillings', 'secondary deposits' (Hottinger, 2006, for fusulinids and pfenderinids), or 'masse presque opaque' (Schroeder, 1985). In 1948 (p. 26), Henson already noted that in Dictyconella minima "the interseptal structures are sporadic and poorly defined in the central shield" (= central zone). In this context, Henson (ibid.) was obviously referring to the presence of secondarily infilled shell material. In orbitolinids they were reported, for example, from Carinoconus (Schroeder, 1985). These fillings, when extensively present, mask the inner structure namely the individual pillars of the central zone that appear then as a fused almost homogeneous mass. Being present in both Gusicella minima and G. complexa, it can be regarded as a characteristic feature of the genus. This feature was so far only reported from very few Cretaceous orbitolinids and is seemingly lacking in Paleogene forms (e.g., Hottinger & Drobne, 1980).
Concerning the embryo of Gusicella,
no details are known from G. minima.
Some axial sections of G. complexa
might point to a tiny subdivision of short partitions (= complex embryo) (Pl.
5 ![]() ,
figs. E, G). These elements might also belong to the septa of undivided neanic
chambers arranged in a tight whorl around the embryo. Better preserved material
is needed for a final conclusion. In any case, the Dictyorbitolininae includes
both genera with a complex embryo such as Dictyorbitolina Cherchi & Schroeder,
1976, or more
common simple embryo (e.g., Paracoskinolina
Moullade, 1965, or Praedictyorbitolinina
Schroeder in Schroeder et al.,
1990). For Gusicella, it is thought to be simple (see remarks for the
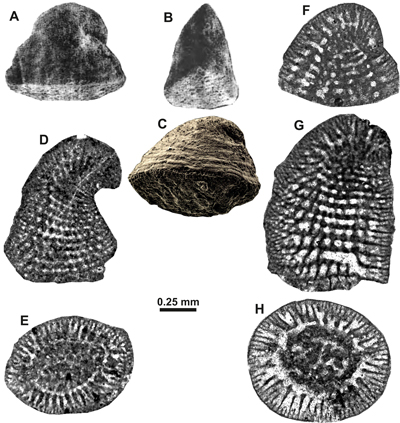
genus below). With respect to the coiling mode of the initial part, Henson
(1948, p. 25) stated that it is "planispiral"
(
) "lying
approximately in the axial plane through the longer diameter of the base".
This observation is, in our opinion, also confirmed by the isolated specimen illustrated by Hewaidy
and Al-Hitmi (1993, Pl. 5, figs. 1-2) (Fig.
3.A-B
,
figs. E, G). These elements might also belong to the septa of undivided neanic
chambers arranged in a tight whorl around the embryo. Better preserved material
is needed for a final conclusion. In any case, the Dictyorbitolininae includes
both genera with a complex embryo such as Dictyorbitolina Cherchi & Schroeder,
1976, or more
common simple embryo (e.g., Paracoskinolina
Moullade, 1965, or Praedictyorbitolinina
Schroeder in Schroeder et al.,
1990). For Gusicella, it is thought to be simple (see remarks for the
genus below). With respect to the coiling mode of the initial part, Henson
(1948, p. 25) stated that it is "planispiral"
(
) "lying
approximately in the axial plane through the longer diameter of the base".
This observation is, in our opinion, also confirmed by the isolated specimen illustrated by Hewaidy
and Al-Hitmi (1993, Pl. 5, figs. 1-2) (Fig.
3.A-B ![]() ), the holotype of Henson
(Fig. 3.C
), the holotype of Henson
(Fig. 3.C ![]() ), as well as some axial sections in the plane of compression (e.g.,
Fig. 3.F
), as well as some axial sections in the plane of compression (e.g.,
Fig. 3.F ![]() ) or perpendicular to it (Pl. 1
) or perpendicular to it (Pl. 1
![]() , figs. C, J).
, figs. C, J).
|
|
Figure 3: Comparison and confrontation of Gusicella minima (Henson) from the Cenomanian of Qatar (A-C), the middle to upper Cenomanian Natih Formation of Oman (C-D), and Gusicella complexa Schlagintweit & Rashidi sp. nov. (E-G) from the upper Maastrichtian Tarbur Formation of Iran. (A-B) Lateral external views parallel to the plane of test compression. (C) Lateral view of A perpendicular to the plane of compression. (D, G) Tangential sections showing aligned arrangement of pillars. (E, H) Slightly oblique transverse sections. (F) Axial section. Fig. 3.A and 3.B from Hewaidy and Al-Hitmi (1993, Pl. 5, figs. 1-2), Fig. 3.C with courtesy of Mike Simmons and permission by NHM, London, Fig. 3.D from Pl. 4, fig. B; Fig. 3.E from Pl. 4, fig. I; Fig. 3.F from Pl. 5, fig. G; Fig. 3.H from Pl. 5, fig. A. |
The last aspect of discussion refers to the arrangement of the pillars of the central zone. In cylindroconical forms such as Paracoskinolina maynci (Chevalier) or Paracoskinolina arcuata (Arnaud-Vanneau), the aligned arrangement of the pillars between subsequent chambers is evident (e.g., Arnaud-Vanneau, 1980). In more flat, conical forms, however, the response to the high rate of increasing chamber diameter can mask some structural features, such as the alignment of pillars. Apart from that, there is, in addition, the presence of a central micritic mass that masks the structure.
Type species. Dictyoconella minima Henson, 1948. Holotype P.35838 in Henson (1948), stored at the Natural History Museum, London as IPC reference M/3441 i NHM P35838 (see https://marinespecies.org/foraminifera/aphia.php?p=image&pic=171946&tid=911119).
Gusicella minima (Henson, 1948)
(Fig. 2.H-I ![]() ; Pls. 1
; Pls. 1
![]() - 2
- 2 ![]() - 3
- 3 ![]() - 4
- 4 ![]() )
)
1948 Dictyoconella minima n. sp. - Henson, p. 25; Pl. 11, figs. 3, 8-10.
1993 Dictyoconella minima Henson - Hewaidy & Al-Hitmi, p. 479; Pl. 5, figs. 1-3.
1998 Dictyoconella minima Henson - Whittaker et al.; Pl. 4, fig. 3; Pl. 5, figs. 1-2; Pl. 48, figs. 1-4.
non 2020 Dictyoconella minima - Mohammed et al.; Figs. 7j, 10f.
non 2021 Gusicella minima (Henson) - Schlagintweit & Rashidi, p.10; Figs. 2c-d, 7-9 (= Gusicella complexa sp. nov.).
non 2022 Gusicella minima (Henson) - Schlagintweit, p. 436; Figs. 2H, N-M, 4A-B, D, F-G, 5A-B (= Gusicella complexa sp. nov.).
Description: Finely agglutinated shell of medium to high-conical morphology; wall thin
(epiderm) and non-canaliculate. Early stage planispiral, offset from the apex. .
The embryonic apparatus could not be characterized due
to the lack of well-centered sections. In one axial section (Pl. 1
![]() ,
fig. A), a structure interpretable as a first chamber or proloculus of
~30-40 µm is observed, suggesting a possible macrospheric specimen. The
preservation of this specimen does not allow ruling out that this structure
represents a dissolution artifact of the test rather than the embryonic first
chamber.
,
fig. A), a structure interpretable as a first chamber or proloculus of
~30-40 µm is observed, suggesting a possible macrospheric specimen. The
preservation of this specimen does not allow ruling out that this structure
represents a dissolution artifact of the test rather than the embryonic first
chamber.
In the adult stage, the chamber arrangement is uniserial and laterally
compressed. As a result, transverse sections of the cone are typically
ellipsoidal in shape, with slightly angular peripheries (Pl. 2 ![]() , fig. N, R).
Axial sections perpendicular to the plane of compression result in a measured
lower test diameter (e.g., Pl. 1
, fig. N, R).
Axial sections perpendicular to the plane of compression result in a measured
lower test diameter (e.g., Pl. 1
![]() , fig. C). Due to the slight lateral
compression of the cone, some slightly oblique transverse sections may give the
impression of a circular morphology (e.g.,
Pl. 2
, fig. C). Due to the slight lateral
compression of the cone, some slightly oblique transverse sections may give the
impression of a circular morphology (e.g.,
Pl. 2 ![]() , fig. K). Adult test with up to 25 chambers subdivided into marginal and
central zones. There are 7 to 9 chambers per 0.5 mm axial length. The marginal
zone consists of one rafter and one to three intercalary beams forming a
subepidermal network. The beams are radially stretching towards the center,
slightly thickening distally with a rounded end. The length of the beams is
roughly three times that of the intercalary beams. Central zone with pillars
that appear aligned between successive chambers rather than with strict
alternating disposition. As the pillars are thickened at the base and top, they
have a knobby appearance in axial sections (e.g., Pl. 1
, fig. K). Adult test with up to 25 chambers subdivided into marginal and
central zones. There are 7 to 9 chambers per 0.5 mm axial length. The marginal
zone consists of one rafter and one to three intercalary beams forming a
subepidermal network. The beams are radially stretching towards the center,
slightly thickening distally with a rounded end. The length of the beams is
roughly three times that of the intercalary beams. Central zone with pillars
that appear aligned between successive chambers rather than with strict
alternating disposition. As the pillars are thickened at the base and top, they
have a knobby appearance in axial sections (e.g., Pl. 1
![]() , fig. E). Often, the central zone contains secondary deposits (or opaque
micritic masses) thereby laterally fusing neighboured pillars and resulting in a
dense micritic zone (Pl. 1
, fig. E). Often, the central zone contains secondary deposits (or opaque
micritic masses) thereby laterally fusing neighboured pillars and resulting in a
dense micritic zone (Pl. 1
![]() , figs. D, O). Marginal zone and central zone are
separated by a narrow marginal interspace (marginal trough sensu Davies,
1930). Here, foramina are
arranged in a concentric row located between the ends of two beams. Beams and
foramina are aligned between successive chambers (e.g., Fig.
3.C, .F
, figs. D, O). Marginal zone and central zone are
separated by a narrow marginal interspace (marginal trough sensu Davies,
1930). Here, foramina are
arranged in a concentric row located between the ends of two beams. Beams and
foramina are aligned between successive chambers (e.g., Fig.
3.C, .F ![]() ; Pl. 1
; Pl. 1
![]() , figs. E-F;
Pl. 3
, figs. E-F;
Pl. 3 ![]() , fig. I). Dimorphism not identified.
, fig. I). Dimorphism not identified.
Dimensions (in mm; data of Henson, 1948 in parentheses):
Test height = up to 1.5 mm (1.0-1.5 mm)
Diameter at base = 0.75 mm to 1.1 mm (0.8-1.8 mm)
Comparisons: Sections of juvenile specimens of G.
minima or sections cutting the initial spire obliquely [Pl. 1
![]() , fig. C (uppermost part), J and M] display striking similarities to Carinoconus
iraqiensis Bernaus & Masse,
2007, described from the
Cenomanian of Iraq. Bernaus and Masse
(2007, p. 4) concluded their generic attribution as
follows: "The presence of an initial
stage with a sharply angular carina was the only criteria used by Cherchi
and Schroeder (1982) for the creation of the new subgenus, now genus,
Carinoconus.
Based on this characteristic, and as no other genus of the family Orbitolinidae has such a distinctive feature, we also assign this new species to the
genus Carinoconus". Bernaus and Masse
(2007) stated that no
rafters were observed in C. iraqiensis,
a view not supported by our observations. Instead, a single rafter per chamber
is visible in Pl. 1
, fig. C (uppermost part), J and M] display striking similarities to Carinoconus
iraqiensis Bernaus & Masse,
2007, described from the
Cenomanian of Iraq. Bernaus and Masse
(2007, p. 4) concluded their generic attribution as
follows: "The presence of an initial
stage with a sharply angular carina was the only criteria used by Cherchi
and Schroeder (1982) for the creation of the new subgenus, now genus,
Carinoconus.
Based on this characteristic, and as no other genus of the family Orbitolinidae has such a distinctive feature, we also assign this new species to the
genus Carinoconus". Bernaus and Masse
(2007) stated that no
rafters were observed in C. iraqiensis,
a view not supported by our observations. Instead, a single rafter per chamber
is visible in Pl. 1
![]() , figs. 3 (right side), 4 (right side), 5 (right below), and
10 (right) of Bernaus and Masse
(2007). This feature provides
another point of comparison with
Gusicella minima.
In addition, Bernaus and MasSe
(2007) noted the presence of two
orders of beams and their alignment in successive chambers like in G.
minima. Possible differences regarding dimensions are not purposeful because
the data provided by Bernaus and Masse
(2007) are assumed to
belong to juvenile forms only. Here we refrain from a direct synonymisation of Carinoconus
iraqiensis and
Gusicella minima, the
former then having nomenclatural priority, as such a conclusion would be
premature without a thorough study of the original material. Due to the possible
identity of the two taxa we prefer to treat Carinoconus
iraqiensis as a species of 'uncertain status', a category used by
Loeblich and Tappan (1987). As further consequence of these
observations, one might think of the late Albian (?to earliest Cenomanian) Carinoconus casterasi (Bilotte et al.,
1973) without rafters representing a possible precursor of
the more complex Gusicella minima
(Schlagintweit & Rashidi, 2021), both having the sharp angled carina
and a pillared central zone with secondary deposits (masking further details in
common). Transverse sections also show foramina arranged in a ring-shaped manner
within a marginal trough (Bilotte et
al., 1973, Pl. 1, fig. 10, above). Regarding the beams, Schroeder
(1985, p. 44) remarked that they are not alternating between subsequent chambers
but could be situated one above the other ("peuvent
être situées l'une au dessous de l'autre"). Gusicella Schlagintweit & Rashidi however appears
to differ from Carinoconus Cherchi
& Schroeder, 1982, because of its inner structure. The (main) beams
of Carinoconus display a thickened
bifurcated distal end (Schroeder, 1985, "l'extrémité de ces cloisonettes et renflée en massue", Pl.
18, figs. 4, 7, above) partly enclosing the outer annular row of foramina
(compare Fig. 2.C
, figs. 3 (right side), 4 (right side), 5 (right below), and
10 (right) of Bernaus and Masse
(2007). This feature provides
another point of comparison with
Gusicella minima.
In addition, Bernaus and MasSe
(2007) noted the presence of two
orders of beams and their alignment in successive chambers like in G.
minima. Possible differences regarding dimensions are not purposeful because
the data provided by Bernaus and Masse
(2007) are assumed to
belong to juvenile forms only. Here we refrain from a direct synonymisation of Carinoconus
iraqiensis and
Gusicella minima, the
former then having nomenclatural priority, as such a conclusion would be
premature without a thorough study of the original material. Due to the possible
identity of the two taxa we prefer to treat Carinoconus
iraqiensis as a species of 'uncertain status', a category used by
Loeblich and Tappan (1987). As further consequence of these
observations, one might think of the late Albian (?to earliest Cenomanian) Carinoconus casterasi (Bilotte et al.,
1973) without rafters representing a possible precursor of
the more complex Gusicella minima
(Schlagintweit & Rashidi, 2021), both having the sharp angled carina
and a pillared central zone with secondary deposits (masking further details in
common). Transverse sections also show foramina arranged in a ring-shaped manner
within a marginal trough (Bilotte et
al., 1973, Pl. 1, fig. 10, above). Regarding the beams, Schroeder
(1985, p. 44) remarked that they are not alternating between subsequent chambers
but could be situated one above the other ("peuvent
être situées l'une au dessous de l'autre"). Gusicella Schlagintweit & Rashidi however appears
to differ from Carinoconus Cherchi
& Schroeder, 1982, because of its inner structure. The (main) beams
of Carinoconus display a thickened
bifurcated distal end (Schroeder, 1985, "l'extrémité de ces cloisonettes et renflée en massue", Pl.
18, figs. 4, 7, above) partly enclosing the outer annular row of foramina
(compare Fig. 2.C ![]() ). In fact, these features are rather delicate and accessible
only in selected sections. With the possibility open pending further data and
studies that Gusicella represents a
junior synonym of Carinoconus,
such a conclusion would result in the lineage C.
casterasi (late Albian to ?earliest Cenomanian; no rafters), C.
minima (middle to late Cenomanian; one rafter), and C. complexa (late
Maastrichtian; two to three rafters). It is worth mentioning that Carinoconus
was not included within the Dictyorbitolininae by Schroeder
in Schroeder
et al. (1990, p. 196).
). In fact, these features are rather delicate and accessible
only in selected sections. With the possibility open pending further data and
studies that Gusicella represents a
junior synonym of Carinoconus,
such a conclusion would result in the lineage C.
casterasi (late Albian to ?earliest Cenomanian; no rafters), C.
minima (middle to late Cenomanian; one rafter), and C. complexa (late
Maastrichtian; two to three rafters). It is worth mentioning that Carinoconus
was not included within the Dictyorbitolininae by Schroeder
in Schroeder
et al. (1990, p. 196).
Finally, juvenile specimens of Gusicella minima may also recall juvenile specimens of Spirocyclina atlasica Saint-Marc & Rahhali, 1982, from the upper Cenomanian of Morocco. In the rich material from Oman, however, the typical peneropliform compressed adult specimens of S. atlasica were not observed.
Stratigraphy: Henson (1948) described 'Dictyoconella' minima "most probably" from the upper Cenomanian or Turonian of Qatar (Dukhan no. 2 and 3 wells; Wasia Group). For the Dukhan oil field of Qatar, Hewaidy and Al-Hitmi (1993) established a late Cenomanian 'Dictyoconella minima Range Zone'. In Oman, Gusicella minima has been identified in Member B and the lower part of Member A of the well-known Natih Formation. The studied material comes from the localities of Tanuf and Wadi Mu'Aydin, the latter of which can also be considered the regional stratotype section for this formation. At Wadi Mu'Aydin, G. minima occurs within a relatively restricted stratigraphic interval, approximately 10 meters thick. In terms of sequence stratigraphy, this G. minima-rich interval lies within the third (III) third-order depositional sequence, which has been extensively described and discussed by Buchem et al. (1996) and Grélaud et al. (2006), among others. This stratigraphic context allows the levels bearing G. minima to be dated to the late middle to late Cenomanian (e.g., Bromhead et al., 2022, Fig. 4).
Gusicella complexa Schlagintweit & Rashidi sp. nov.
(Fig. 3.F-H ![]() ;
Pls.
5
;
Pls.
5 ![]() - 6
- 6
![]() )
)
2016a Dictyoconella? minima Henson - Schlagintweit et al., p. 34; Fig. 3a-b pars, 5-6.
2020 Dictyoconella minima - Mohammed et al.; Figs. 7j, 10f.
2021 Gusicella minima (Henson) - Schlagintweit & Rashidi, p. 10; Figs. 2c-d, 7-9.
2022 Gusicella minima (Henson) - Schlagintweit, p. 436; Figs. 2H, N-M, 4A-B, D, F-G, 5A-B.
Description: G. complexa displays the following specific features:
test medium conical (apical angle from 50 to 90 degrees) and sometimes laterally
slightly compressed (Pl.
5 ![]() , fig. J; Pl. 6
, fig. J; Pl. 6
![]() , fig. J) with both sides and base
slightly convex;
, fig. J) with both sides and base
slightly convex;
acute apex is tilted, marking the early development in a downward turn to the
cone base (e.g., Pl.
5 ![]() , fig. G).
, fig. G).
Early stage:
small spire of few chambers arranged in half a whorl that is closely
attaching (almost parallel) to the side test wall (Pl.
5 ![]() , figs.
D-G);
, figs.
D-G);
embryo represented by a single sub-spherical chamber (protoconch) showing short
septules? at its upper part facing the cone base (Pl.
5 ![]() , figs. E-G).
, figs. E-G).
Adult stage:
marginal zone with one to three rafters (e.g., Pl.
5 ![]() , fig. E, bottom left)
and two to three intercalary beams (e.g., Pl. 6
, fig. E, bottom left)
and two to three intercalary beams (e.g., Pl. 6
![]() , fig. P). In case of two rafters, the upper one is twice as long
as the lower one. In case of three rafters, the middle one is roughly twice as
long as the others. At their inner ends, the main beams are moderately
thickening. In shallow-tangential sections, the sub-epidermal network
displays a pattern of sub-rounded alveolar compartments (Pl.
5
, fig. P). In case of two rafters, the upper one is twice as long
as the lower one. In case of three rafters, the middle one is roughly twice as
long as the others. At their inner ends, the main beams are moderately
thickening. In shallow-tangential sections, the sub-epidermal network
displays a pattern of sub-rounded alveolar compartments (Pl.
5 ![]() , fig. H; Pl. 6
, fig. H; Pl. 6
![]() ,
fig. F). The marginal chamberlets are tapering distally;
,
fig. F). The marginal chamberlets are tapering distally;
outer rim of the marginal trough outlined by the septa slightly bending upwards
and forming a 'buttress on marginal ridge' sensu Davies
(1930)
as seen in sub-axial sections (Pl.
5 ![]() , fig. I bottom; Pl. 6
, fig. I bottom; Pl. 6
![]() , fig. H left
side);
, fig. H left
side);
central zone with pillars [when visible where secondary
deposits (= opaque micritic masse) is not present] comparably narrowly spaced (Pl.
5 ![]() , figs. C-D; Pl. 6
, figs. C-D; Pl. 6
![]() , figs. A,
H);
, figs. A,
H);
up to 18 chambers in the adult stage, with seven to nine chambers per 0.50 mm axial length;
dimorphism possible (high-conical megalospheric and low-conical microspheric forms?).
Remarks:
The species (as a Maastrichtian form of G. minima) has been described in
detail by Schlagintweit and Rashidi
(2021). However, the new form
named herein - G. complexa - differs
from G. minima sensu stricto by
its more complex marginal zone regularly including two rafters and two to four
intercalary beams. There is a maximum of three rafters in the marginal zone, a longer
one in the middle and two shorter rafters of equal length on both sides. As a result of the higher number of rafters, the
greater chamber height of G. complexa
is also worth mentioning. Therefore, there are seven to nine chambers per 0.5 mm
axial length in G. minima,
compared to just six to seven in G.
complexa. The
general high-conical morphology with the angular planispiral initial coil and
the dimensional data, however, are shared by both species (Fig.
3 ![]() ). Both
fulfill partly the requirements for an orbitolinid lineage characterized by increasing
complexity (Schroeder et al., 2002). Ideally, the taxa of such lineages display overlapping ranges, a
requirement that however is not applicable in the present case. Instead, there
is considerable gap spanning the Turonian to early Maastrichtian time interval
without any record of Gusicella.
Further discoveries will hopefully be forthcoming.
). Both
fulfill partly the requirements for an orbitolinid lineage characterized by increasing
complexity (Schroeder et al., 2002). Ideally, the taxa of such lineages display overlapping ranges, a
requirement that however is not applicable in the present case. Instead, there
is considerable gap spanning the Turonian to early Maastrichtian time interval
without any record of Gusicella.
Further discoveries will hopefully be forthcoming.
Dimensions (in mm; data of Henson, 1948 in parentheses):
Test diameter (D): 0.7-1.9 mm. Note: specimens with a test diameter > 1.5 mm appear to be microspheric specimens.
Test height (H): 0.8-1.4 (1.0-1.5 mm)
D/H (megalospheric forms): 0.8-1.35
D/H (microspheric forms): around 1.5
Comparisons: The dimensions (height, diameter) of Gusicella complexa are in the range reported for G.
minima, so differences in size cannot be taken as a distinguishing
criterion. The main difference corresponds to the more complex marginal zone of G.
complexa with two to three rafters per chamber (instead of one in G.
minima) and two to three intercalary beams (instead of one to two in G.
minima) (Fig. 3 ![]() ). As a consequence of the higher number of rafters in G.
complexa, the height of adult chambers is also higher than in G. minima.
). As a consequence of the higher number of rafters in G.
complexa, the height of adult chambers is also higher than in G. minima.
Stratigraphy: Gusicella complexa is the orbitolinid with the greatest vertical distribution in the Naghan section of the Tarbur Formation. It preferentially occurs in wackestone/packstones with rudist shell debris as well as in association with Loftusia spp., Omphalocyclus and several other LBF (Wynd, 1965: Omphalocyclus-Loftusia assemblage zone). The occurrences of Siderolites calcitrapoides Lamarck and/or Canalispina iapygia Robles-Salcedo et al. are in the lower part of several sections of the Tarbur Formation, e.g., in the Mandegan section some meters above the boundary with the underlying Gurpi Formation. This indicates a late Maastrichtian age, since the first occurrence ofC. iapygia is latest early Maastrichtian (according to Robles Salcedo, 2014; Robles Salcedo et al., 2019). Mohammed et al. (2020) reported Dictyoconella minima in their "facies-type 9 Siderolites/Orbitoides packstone" from the upper part of the Maastrichtian Simsima Formation of the United Arab Emirates, associated with Loftusia sp. and Dictyoconus sp. Although Dictyoconus and Dictyoconella were not illustrated, the association clearly recalls the assemblage reported from the upper Maastrichtian Tarbur Formation (Schlagintweit et al., 2016a; Schlagintweit & Rashidi, 2021). However, the illustrated specimen of 'Dictyoconella minima' (Mohammed et al., 2020, Fig. 7j) does not belong to this species but to an indeterminable orbitolinid. Henson (1948, p. 26) noted "a single specimen, possibly of this species" (= Dictyoconella minima) from the Maastrichtian of Iraq.
The present contribution provides an enriched description of Gusicella minima, one of the little-known 'Henson taxa' reported in his important monograph from 1948. The taxonomic inventory includes two species of the genus Gusicella: G. minima (Henson) and G. complexa Schlagintweit & Rashidi, both restricted palaeogeographically to the Upper Cretaceous margins of Neotethys along the northern rim of the Arabian Plate. As the name implies, the latter is characterized by a more complex marginal zone, which is an important criterion for the distinction of species in different orbitolinid lineages (e.g., Montseciella or Simplorbitolina). Gusicella minima and G. complexa are separated by a considerable gap without records that spans the Turonian to early Maastrichtian interval. Both taxa appear at the ends of Global Community Maturation cycles of LBF characterized by high diversities (Hottinger, 1997). Along with Persiconus sarvaki Yazdi-Moghadam & Schlagintweit, Gusicella minima (Henson) represents the second species of Orbitolinidae recorded from upper Cenomanian strata.
Special thanks are extended to the late and much appreciated Prof. Josep Serra-Kiel, as well as to Drs. Carine Grélaud and Philippe Razin, for their logistical support during one of the field seasons in Oman and for generously sharing their extensive knowledge of the geology of Oman. Thanks are also due to Dr. Gerard Lucena, geologist and expert preparator of thin sections at the MCNB. Some remarks provided by an anonymous reviewer and the very detailed review by Hervé Calvez (Lannion) helped to improve the manuscript. The holotype specimen of Dictyconella minima was kindly provided by Mike Simmons (London) with courtesy of the Natural History Museum, London. The language review by Zev Brook (San Diego) is gratefully acknowledged
Arnaud-Vanneau A. (1980).- L'Urgonien du Vercors septentrional et de la Chartreuse.- Géologie Alpine, Mémoire, Grenoble, vol. 11 (3 volumes), 874 p.
Bernaus J.M. & Masse P. (2007).- Carinoconus iraqiensis (Foraminifera), a new orbitolinid from the Cenomanian Mishrif Formation of the oil fields of southeastern Iraq.- Micropaleontology, Flushing - NY, vol. 52, no. 5, p. 471-476.
Bilotte M., Canérot J., Moullade M. & Peybernès B. (1973).- Description et position stratigraphique d'un nouvel Orbitolinidé de l'Albo-Cénomanien Ibérique et Pyrénéen Paracoskinolina casterasi n. sp.- Société de Physique et d'Histoire Naturelle de Genève, vol. 26, no. 2, p. 183-185.
Bromhead A.F., Buchem, F.S.P. van, Simmons M.D. & Davies R.B. (2022).- Sequence stratigraphy, palaeogeography and petroleum plays of the Cenomanian-Turonian succession of the Arabian Plate: An updated synthesis.- Journal of Petroleum Geology, vol. 45, p. 119-162.
Buchem F.S.P. van, Razin P., Homewood P.W., Philip J.M., Eberli G.P., Platel J.-P., Roger J., Eschard R., Desaubliaux G.M.J., Boisseau T., Leduc J.-P., Labourdette R. & Cantaloube S. (1996).- High resolution sequence stratigraphy of the Natih Formation (Cenomanian / Turonian) in Northern Oman: Distribution of source rocks and reservoir facies.- GeoArabia, Manama, vol. 1, p. 65-91.
Callegari I., Scharf A., Mattern F. & Scharf K. (2022).- Tectonic transition from thrusting to polyphase non-confining deformation within the Semail Ophiolite along the sinistral, transtensional Issmaiya Fault Zone (Sultanate of Oman).- Journal of Asian Earth Sciences, vol. 224, article 105007, 16 p.
Cardelli S. (2024, unpublished).- The impact of the Oceanic Anoxic Event 2 on resilient carbonate platforms: A multidisciplinary approach.- PhD thesis, University of Ferrara, 316 p.
Cherchi A. & Schroeder R. (1976).- Dictyorbitolina ichnusae n. gen., n. sp. (Foram.) del Barremiano della Sardegna nord-occidentale.- Bollettino della Società Paleontologica Italiana, Milano, vol. 14, no. 1, p. 47-54.
Cherchi A. & Schroeder R. (1982).- Sobre la edad de la transgresión mesocretácica en Asturias.- Cuadernos de Geología Ibérica, Madrid, vol. 8, p. 219-233. URL: https://dialnet.unirioja.es/servlet/articulo?codigo=264520&orden=1&info=link
Davies L.M. (1930).- XX.-The genus Dictyoconus and its allies: A review of the group, together with a description of three new species from the Lower Eocene beds of northern Baluchistan.- Transactions of the Royal Society of Edinburgh, vol. 56, no. 2, p. 485-505.
Forbes G.A., Jansen H.S.M. & Schreurs J. (2010).- Lexicon of Oman - Subsurface Stratigraphy.- GeoArabia Special Publication, Manama, vol. 5, 371 p.
Grélaud C., Razin P., Homewood P.W., & Schwab A.M. (2006).- Development of incisions on a periodically emergent carbonate platform (Natih Formation, Late Cretaceous, Oman).- Journal of Sedimentary Research, Tulsa - OK, vol. 76, p. 647-669.
Henson F.R.S. (1948).- Larger imperforate Foraminifera of south-western Asia. Families Lituolidae, Orbitolinidae and Meandropsinidae.- Monograph British Museum (Natural History), London, 127 p.
Hewaidy A.G.A. & Al-Hitmi H.A. (1993).- Cretaceous-Early Eocene biostratigraphy of the Dukhan oil field, west Qatar, Arabian Gulf.- Al-Azhar Bulletin of Science, Cairo, vol. 4, no. 2, p. 469-494.
Hottinger L. (1997).- Shallow benthic foraminiferal assemblages as signals for depth of their deposition and their limitations.- Bulletin de la Société Géologique de France, Paris, vol. 168, no. 4, p. 491-505.
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, no. M02 (CG2006_M02), 126 p. DOI: 10.4267/2042/5832
Hottinger L. & Drobne K. (1980).- Early Tertiary conical imperforate foraminifera.- Slovenska Akademija Znanosti in Umetnosti, Classis IV Historia Naturalis, Dissertationes, vol. 22, no. 3, p. 187-276
James G.A. & Wynd J.G. (1965).- Stratigraphic Nomenclature of Iranian Oil Consortium Agreement Area.- AAPG Bulletin, Boulder - CO, vol. 49, no. 12, p. 2218-2232.
Kaminski M.K. (2000).- The new and reinstated genera of agglutinated Foraminifera published between 1986 and 1996. In: Hart M.B., Kaminski M.A. & Smart, C.W. (eds.), Proceedings of the Fifth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Varsaw, vol. 7, p. 185-219.
Kaminski M.A. (2004).- The year 2000 classification of the agglutinated Foraminifera. In: Bubík M. & Kaminski M.A. (eds.), Proceedings of the Sixth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Varsaw, vol. 8, p. 237-255.
Kaminski M.A. (2014).- The year 2010 classification of the agglutinated Foraminifera.- Micropaleontology, Flushing - NY, vol. 60, no. 1, p. 89-108.
Loeblich A.R. Jr & Tappan H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, 2 vol., 970 p. (847 Pls.).
Martin K. (1890).- Untersuchungen über den Bau von Orbitolina (Patellina auct.) von Borneo.- Sammlungen des Geologischen Reichs-Museums in Leiden (ser. 1), vol. 4, p. 209-231.
Meister C. & Piuz A. (2015).- Cretaceous ammonites from the Sultanate of Oman (Adam Foothills).- GeoArabia, Manama, vol. 20, p. 19-74.
Mohammed I.Q., Farouk S., Lawa F.A., Alsuwaidi M. & Morad S. (2020).- Upper Cretaceous wedge-top to foredeep architecture in the United Arab Emirates: Insights from the Faiyah Anticline.- Geological Journal, vol. 56, p. 2602-2624.
Moullade M. (1965).- Contribution au problème de la classification des Orbitolinidae (Foraminiferida, Lituolacea).- Comptes Rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, vol. 260, p. 4031-4034.
Pawłowski J., Holzmann M. & Tyszka J. (2013).- New supraordinal classification of Foraminifera: Molecules meet morphology.- Marine Micropaleontology, vol. 100, p. 1-10.
Parente M., Frijia G., Di Lucia M., Jenkyns H.C., Woodfine R.G. & Baroncini F. (2008).- Stepwise extinction of larger foraminifers at the Cenomanian-Turonian boundary: A shallow-water perspective on nutrient fluctuations during Oceanic Anoxic Event 2 (Bonarelli Event).- Geology, Boulder - CO, vol. 36, no. 9, p. 715-718.
Philip J., Borgomano J. & Al-Maskiry S. (1995).- Cenomanian-Early Turonian carbonate platform of northern Oman: Stratigraphy and palaeoenvironments.- Palæogeography, Palæoclimatology, Palæoecology, vol. 119, nos. 1-2, p. 77-92.
Piuz A. & Vicedo V. (2020).- New Cenomanian "nummoloculinas" of the Natih Formation of Oman.- Cretaceous Research, vol. 107, article 104224, 17 p.
Robles-Salcedo R. (2014).- La Familia Siderolitidae (Macroforaminíferos del Cretácico Superior): Arquitectura de la concha, bioestratigrafía, distribución paleoambiental y palaleobiogeografía.- PhD Thesis, University of Barcelona, 183 p.
Robles-Salcedo R., Vicedo V., Parente M. & Caus E. (2019).- Canalispina iapygia gen. et sp. nov.: The last Siderolitidae (Foraminiferida) from the upper Maastrichtian of southern Italy.- Cretaceous Research, vol. 98, p. 84-94.
Saint-Marc P. & Rahhali I. (1982).- Sur la présence du genre Spirocyclina (foraminifère) dans le Cénomanien supérieur du Maroc.- Revue de Micropaléontologie, Paris, vol. 25, no. 2, 133-140.
Schlagintweit F. (2020).- Maastrichtian larger benthic Foraminifera from the Arabian Plate sensu lato: New data from Somalia, Turkey and Iran.- Acta Palaeontologica Romaniae, Bucharest, vol. 16, no. 2, p. 69-83.
Schlagintweit F. (2022).- Late Cretaceous Orbitolinidae (Larger Benthic Foraminifera): Taxonomy, determination, biostratigraphy and palaeobiogeography.- Newsletters on Stratigraphy, Stuttgart, vol. 55, no. 4, p. 427-450.
Schlagintweit F. & Rashidi K. (2016).- Some new and poorly known benthic Foraminifera from late Maastrichtian shallow-water carbonates of the Zagros Zone, SW Iran.- Acta Palaeontologica Romaniae, Bucharest, vol. 12, no. 1, 53-70.
Schlagintweit F. & Rashidi K. (2021).- Dictyoconella Henson, 1948, Upper Cretaceous Larger Benthic Foraminifera: A taxonomic revision with the establishment of Gusicella gen. nov. (type-species Dictyoconella minima Henson).- Acta Palaeontologica Romaniae, Bucharest, vol. 17, no. 2, p. 3-13.
Schlagintweit F., Rashidi K. & Babadipour M. (2016a).- Orbitolinid Foraminifera from the Late Maastrichtian of the Tarbur Formation (Zagros Zone, SW Iran).- Acta Palaeontologica Romaniae, Bucharest, vol. 12, no. 2, 29-46.
Schlagintweit F., Rashidi K. & Barani F. (2016b).- First record of Gyroconulina columellifera Schroeder & Darmoian, 1977 (larger benthic Foraminifera) from the Maastrichtian Tarbur Formation of SW Iran (Zagros Fold-Thrust-Belt).- Geopersia, Tehran, vol. 6, no. 2, p. 169-185.
Schroeder R. (1985).- Sous-genre Carinoconus Cherchi & Schroeder, 1983. In: Schroeder R. & Neumann M. (coords.), Les grands Foraminifères du Crétacé moyen de la région méditerranéenne.- Geobios, Villeurbanne, mémoire spécial 7, p. 43-44.
Schroeder R., Clavel B. & Charollais J. (1990).- Praedictyorbitolina carthusiana n. gen., n. sp., Orbitolinidé (Foraminiferida) de la limite Hauterivien-Barrémien des Alpes occidentales.- Paläontologische Zeitschrift, vol. 64, nos. 3-4, p. 193-202.
Schroeder R., Clavel B., Cherchi A., Busnardo R., Charollais J. & Decrouez D. (2002).- Lignées phylétiques d'Orbitolinidés dans Hauterivien supérieur - Aptien inférieur du Sud-Ouest de l'Europe et leur importance stratigraphique.- Revue de Paléobiologie, Genève, vol. 21, p. 853-863.
Whittaker J.E., Jones R.W. & Banner F.T. (1998).- Key Mesozoic benthic Foraminifera of the Middle East.- The Natural History Museum, London, 237 p.
Wohlwend S., Hart M. & Weissert H. (2016).- Chemostratigraphy of the Upper Albian to mid-Turonian Natih Formation (Oman) - how authigenic carbonate changes a global pattern.- The Depositional Record, Hoboken - NJ, vol. 2, no. 1, p. 97-117.
Wynd J.G. (1965).- Biofacies of the Iranian Oil Consortium Agreement Area.- IOOC Report, no. 1082, 89 p.
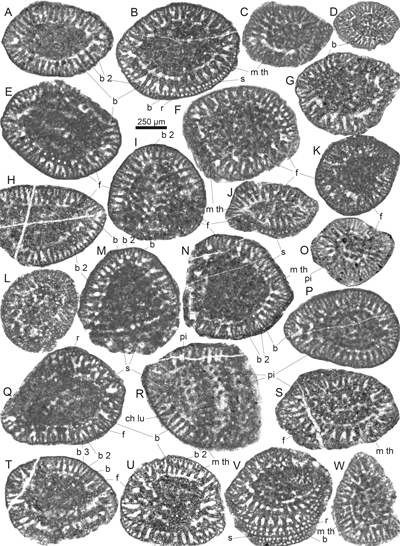
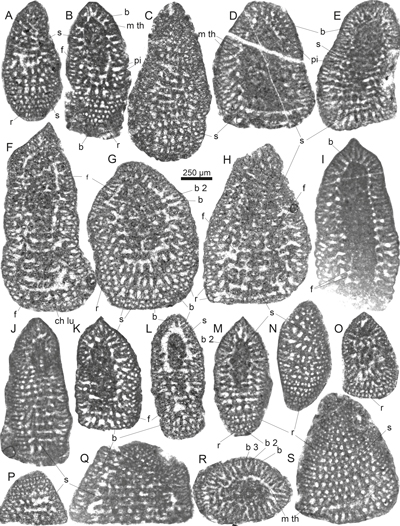
Plate
1:
Gusicella minima (Henson) from the middle to upper Cenomanian
Natih Formation, Wadi Mu'Aydin, Oman
Mountains. All from samples M-47 and M-48 (A-B, D,O) Axial sections in the plane of test compression. Note the dense
central zone in (D) and (O). (C, H, J-K, M, P-Q) (Sub)axial sections
perpendicular to the plane of test compression (especially the early planispire;
e.g., M). (E-G, N, L, R-T) Subaxial sections (unknown orientation towards
the plane of compression). (I) Tangential section. Abbreviations: ch lu = chamber lumen, e = embryo, f = foramen, m th =
marginal trough, op = opaque micritic mass, pi = pillar, r = rafter, s = septum.
Specimen (repository) numbers: A = MGB_59811_LP01.001 B =
MGB_59811_LP03.017, C
= MGB_59811_LP11.001, D = MGB_59812_LP04.004, E = MGB_59811_LP10.001, F=
MGB_59812_LP27.003, G = MGB_59812_LP24.001, H = MGB_59811_LP16.001, I =
MGB_59811_LP09.001, J = MGB_59811_LP06.005', K = MGB_59811_LP07.011, L =
MGB_59811_LP08.011, M = MGB_59811_LP11.006, N = MGB_59811_LP09.002, O =
MGB_59812_LP18.002, P = MGB_59812_LP16.002, Q = MGB_59811_LP10.002, R =
MGB_59811_LP14.002, S = MGB_59811_LP03.007, T = MGB_59811_LP03.005. Specimens
A-C, E, H-N, and Q-T from sample M-47; specimens D, F-G, O-P from sample M-48. |
|
Plate
2: Gusicella minima (Henson) from the middle to upper Cenomanian
Natih Formation, Wadi Mu'Aydin, Oman
Mountains. (A-U, W) Partly oblique transverse sections. Note that there are
round transverse outlines but compression in various degrees exhibiting
subangular margins. In some specimens pillars are well individualized (e.g.,
O, W) while in others are fused and/or with secondary micritic deposits (e.g.,
E, N). (M, V) Oblique section. Abbreviations: b = beam, b 2 = secondary (or
intercalary) beam, b 3 = third order beam, ch lu = chamber lumen, , f = foramen,
m th = marginal trough, pi = pillar, r = rafter, s = septum. Specimen (depository) numbers: A = MGB_59812_LP02.003, B = MGB_59812_LP14.001, C = MGB_59811_LP04.002, D
= MGB_59811_LP16.002, E = MGB_59812_LP17.005, F = MGB_59811_LP07.009, G =
MGB_59811_LP18.001, H = MGB_59812_LP13.003, I = MGB_59812_LP27.002, J =
MGB_59811_LP19.001, K = MGB_59812_LP21.002, L = MGB_59811_LP03.008, M =
MGB_59812_LP21.001, N = MGB_59812_LP04.003, O = MGB_59811_LP01.002, P =
MGB_59812_LP18.003, Q = MGB_59812_LP19.001, R = MGB_59811_LP01.005, S =
MGB_59811_LP19.004, T = MGB_59812_LP30.004, U = MGB_59811_LP18.016, V =
MGB_59812_LP30.005, W = MGB_59811_LP08.001_. Specimens A-B, E, H-I, K, M-N,
P-Q, T, V from sample M-48; specimens C-D, F-G, J, L, O, R-S, U, and W from
sample M-47. |
|
Plate
3:
Gusicella minima (Henson) from the middle to upper Cenomanian
Natih Formation, Wadi Mu'Aydin, Oman
Mountains. (A-S) Tangential, partly oblique sections. Abbreviations: b = beam, b
2 = secondary (or intercalary) beam, b 3 = third order beam, ch lu = chamber
lumen, f = foramen, m th = marginal trough, pi = pillar, r = rafter, s = septum.
Specimen (depository) numbers: A =
MGB_59811_LP02.001, B= MGB_59811_LP06.005, C = MGB_59811_LP14.001, D =
MGB_59812_LP02.001, E = MGB_59811_LP08.006, F = MGB_59811_LP18.011, G =
MGB_59811_LP16.005, H = MGB_59811_LP16.006, I = MGB_59812_LP11.006, J =
MGB_59812_LP23.004, K = MGB_59812_LP29.005, L = MGB_59811_LP18.021, M =
MGB_59812_LP16.001, N = MGB_59812_LP16.004, O = MGB_59811_LP03.006, P =
MGB_59811_LP11.007, Q = MGB_59811_LP08.008, R = MGB_59811_LP04.001, S =
MGB_59812_LP13.001. Specimens A-C, E-H, L, O-R from sample M-47; specimens D,
I-K, M-N, and S from sample M-48. |
|
Plate
4:
Gusicella minima (Henson) from the middle to upper Cenomanian
Natih Formation, Wadi Tanuf, Oman Mountains.
A, F Subaxial sections. B
Tangential section. C, E, I Oblique
transverse section. D, H Oblique
sections. G Transverse section. J
Axial section perpendicular to plane of compression. Note the transverse section
of the initial coil. Abbreviations: a f = aligned foramen, b 1 = primary beams,
b 2 = second order (or intercalary) beam, ch lu = chamber lumen, m th = marginal
trough, pi = pillar, r = rafter, s = septum. Thin-sections: 13 (A,
E-H), S2 (B, J), 14 (C), 24 (D), 9m (I). |
|
Plate
5:
Gusicella complexa Schlagintweit & Rashidi sp. nov.
from the upper Maastrichtian Tarbur Formation of SW Iran (Mandegan section: B;
Naghan section: A, C-J). A, J Oblique
transverse sections. B Tangential-oblique section cutting the marginal (upper
part) and central zones (lower part) displaying irregularly distributed fused
pillars/secondary deposits. C-D, I Subaxial sections. E-G Axial sections showing initial spire and
partial embryo; holotype specimen in E. H Tangential section passing through the
subepidermal network and the aligned beams (centre). Abbreviations: a f =
aligned foramen, b = beam, b 2 = second order beam, b 3 = third order beam, e =
embryo, f = foramen, op = opaque micritic mass, , r = rafter, r 2 second order
rafter, r 3 = third order rafter, s = septum, pi = pillar. Thin sections: 2NG 17
(A, I), Rt 100 (B), Ng 180-1 (C), 2NG 179-1 (D), 2NG
168 (E), 2NG 175 (F), 2NG 174 (G-H), 2NG 177 (J). |
|
Plate
6:
Gusicella complexa Schlagintweit & Rashidi sp. nov.
from the upper Maastrichtian Tarbur Formation of SW Iran (Naghan section: A-B,
J, K; Mandegan section: C-I, L-O). A Subaxial section. B, I, K
Oblique sections. C-E, L-M Tangential sections displaying beams continuous (aligned) from one chamber to the next. F Shallow tangential of the marginal zone section displaying
subepidermal network of beams and rafters. G,
J-K, N-O Different transverse sections, some slightly oblique. Note oval
test outline in J and five marginal
foramina arranged in a tangential row (yellow circles) in O. H Subaxial section. Note alternation of pillars by stairway-like
arrangement of foraminaand opaque
fused pillars/secondary deposits in the central zone. M
Tangential-oblique section. Abbreviations: b = beam, b 2 = second order beam, b
3 = third order beam, f = foramen, a f = aligned foramen,
m th = marginal through, op = opaque micritic mass, pi = pillar, r = rafter, r 2
= second order rafter, s = septum. Thin sections: Ng 196 (A), Ng 192 (B),
Rt 108-3 (C-E, G, M), Rt 104 (F), Rt 96 (H), Rt 108-6 (I),
Ng 187 (J), Ng 186 (K), Rt 107 (L), Rt 102-2 (N), Rt
104 (O). |
|