![]()
![]()
![]() The Quaternary, mainly freshwater sediments of the Lower Mesopotamian plain include a thin transgressive marine unit which extends inland some 250 km from the present coastline, the Hammar Formation. The identity and respective areal extent of continental and transgressive units are based mainly on their molluscan faunas. Those faunas reflect the several environments of the Middle Eastern biogeographic province: fluviatile, lacustrine, estuarine and lagoonal. The ecological requirements of the more common subfossil and
living species of the malacofauna were studied during a month of field reconnaissance.
The Quaternary, mainly freshwater sediments of the Lower Mesopotamian plain include a thin transgressive marine unit which extends inland some 250 km from the present coastline, the Hammar Formation. The identity and respective areal extent of continental and transgressive units are based mainly on their molluscan faunas. Those faunas reflect the several environments of the Middle Eastern biogeographic province: fluviatile, lacustrine, estuarine and lagoonal. The ecological requirements of the more common subfossil and
living species of the malacofauna were studied during a month of field reconnaissance.
The freshwater portion comprises only four bivalve species (Corbicula, Unio,
Anodonta and Pseudodontopsis). The principal gastropods include Theodoxus
(Neritaea), Melanopsis, Melanoides, Bellamya, Gyraulus,
Lymnaea (Radix) spp. Corbicula characterizes both fluviatile channels and lacustrine environments;
Melanopsis, Melanoides, Theodoxus and the Unionaceae prefer shallow, sparsely vegetated,
freshwater lakes. The greatest diversity was seen in the slowly flowing water of the channels draining the marshes; small Planorbids are rarely abundant whereas viviparid and lymnaeids may accumulate near the margins of marshes due to floatation during floods.
The brackish water faunas are characterized by Neritina (Dostia), the potamid
Cerithidea (Cerithideopsilla), and Stenothyra. The new combination
Theora mesopotamica (, 1918) is introduced here as the exact equivalent of
Abra cadabra et ,
1957. The abundance of these species is indicative of a restricted, low energy marine environment, confirmed by great
numbers of the foraminiferan Ammonia gr. beccarii and the ostracod
Cyprideis gr. torosa.
Reworked Hammar Fm marine fossils, first recorded by the
(1918) collection from a lacustrine deposit near Nassiriyah, have been observed in river, marsh and lake sediments too, usually in association with scarce foraminifera. The limited occurrence of these macrofossils (abundant in their life environment) indicates reworking.
![]() Iraq; Mesopotamia; Arabian Gulf; Tigris; Euphrates; Shatt al Arab; freshwater; mollusc; brackish water; Holocene; Hammar Formation; 14C dating; ecology; Typha marsh; reed culture.
Iraq; Mesopotamia; Arabian Gulf; Tigris; Euphrates; Shatt al Arab; freshwater; mollusc; brackish water; Holocene; Hammar Formation; 14C dating; ecology; Typha marsh; reed culture.
J.-C., W.R. (2005).- The modern environments of Molluscs in southern Mesopotamia, Iraq: A guide to paleogeographical reconstructions of Quaternary fluvial, palustrine and marine deposits.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2005/01 (CG2005_A01)
![]() Les environments modernes
des Mollusques en Mésopotamie méridionale, Irak : Un guide pour les
reconstitutions paléogéographiques des dépôts quaternaires
fluviatiles, palustres et marins.- Les dépôts quaternaires d'eau douce de la plaine de Mésopotamie comportent
une mince intercalation marine : la Formation Hammar qui a transgressé
d'environ 250 km par rapport au rivage actuel. L'identification et l'extension
des unités sédimentaires continentales et marines sont basées essentiellement
sur les faunes de mollusques. Ces faunes caractérisent les différents
environnements fluviatiles, lacustres, estuariens et lagunaires de la province
biogéographique du Moyen Orient, très peu étudiés du point de vue
écologique. Les conditions de vie des espèces les plus abondantes, vivantes et
fossiles, ont été observées durant une mission de reconnaissance, en 1980.
Les environments modernes
des Mollusques en Mésopotamie méridionale, Irak : Un guide pour les
reconstitutions paléogéographiques des dépôts quaternaires
fluviatiles, palustres et marins.- Les dépôts quaternaires d'eau douce de la plaine de Mésopotamie comportent
une mince intercalation marine : la Formation Hammar qui a transgressé
d'environ 250 km par rapport au rivage actuel. L'identification et l'extension
des unités sédimentaires continentales et marines sont basées essentiellement
sur les faunes de mollusques. Ces faunes caractérisent les différents
environnements fluviatiles, lacustres, estuariens et lagunaires de la province
biogéographique du Moyen Orient, très peu étudiés du point de vue
écologique. Les conditions de vie des espèces les plus abondantes, vivantes et
fossiles, ont été observées durant une mission de reconnaissance, en 1980.
La malacofaune d'eau douce comporte des bivalves peu variés (Corbicula, Unio,
Anodonta et Pseudodontopsis) et les genres de gastéropodes Theodoxus
(Neritaea), Melanopsis, Melanoides, Bellamya, Gyraulus,
Lymnaea (Radix) spp. La
corbicule caractérise aussi bien les chenaux fluviatiles que les environnements
lacustres ; les
Melanopsis, Melanoides, Theodoxus et unionidés préfèrent les
lacs à végétation clairsemée. La plus grande diversité s'observe dans les
chenaux d'eau à faible courant qui draînent les marais ; les petits planorbes
sont rarement abondants tandis que les paludines et les limnées peuvent former
des accumulations en bordure des marais, par flottation et échouage en période
de hautes eaux. La faune saumâtre est caractérisée par une néritine (Dostia),
le potamide
Cerithidea (Cerithideopsilla) cingulata et Stenothyra.
Nous introduisons la nouvelle combinaison Theora mesopotamica (, 1918)
en remplacement de la regrettée
Abra cadabra de et
(1957).
L'abondance de ces espèces caractérise un environnement maritime calme et
confiné, confirmé par le grand nombre de tests du foraminifère Ammonia gr. beccarii et de l'ostracode
Cyprideis gr. torosa.
Le remaniement de fossiles de
la Formation Hammar peut être mis en évidence dans les dépôts lacustres, palustres et fluviatiles. Les mollusques saumâtres ont été signalés pour la
première fois par
(1918) près de Nassiriyah mais mal interprétés.
Leur faible abondance, en mélange avec la malacofaune d'eau douce, caractérise
leur remaniement.
![]() Irak ;
Mésopotamie ; Golfe Arabique ; Tigre ; Euphrate ; Shatt al Arab ; eau douce ;
mollusque ; eau saumâtre ; Holocène ; Formation Hammar ; datation 14C ; écologie ; marais à Typha ; culture des Arabes des marais.
Irak ;
Mésopotamie ; Golfe Arabique ; Tigre ; Euphrate ; Shatt al Arab ; eau douce ;
mollusque ; eau saumâtre ; Holocène ; Formation Hammar ; datation 14C ; écologie ; marais à Typha ; culture des Arabes des marais.
![]()
![]() This contribution is based on a 1980 field and laboratory
study, organized by B.H. as a joint research project of the Laboratory of Sedimentary Petrology and Paleontology
(Paris-Sud University, Orsay) and the Geological Survey of Iraq (Baghdad). The study was made shortly before the onset of the war that destroyed most of the natural environments of Mesopotamian swamps and lakes
(Fig. A
This contribution is based on a 1980 field and laboratory
study, organized by B.H. as a joint research project of the Laboratory of Sedimentary Petrology and Paleontology
(Paris-Sud University, Orsay) and the Geological Survey of Iraq (Baghdad). The study was made shortly before the onset of the war that destroyed most of the natural environments of Mesopotamian swamps and lakes
(Fig. A
![]() ), and consequently the amphibious villages of the Arabian Reed Culture (Fig. B
), and consequently the amphibious villages of the Arabian Reed Culture (Fig. B
![]() ). Its findings were presented at the 1987 Kuwait Conference on Quaternary Sediments in the Arabian Gulf and Mesopotamian
region, the proceedings of which have never been issued. Consequently, the ecological
field data of the Mesopotamian model have not been
available, so we offer this revised version of our
contribution in the hope of inciting greater interest to the environments of the Mesopotamian plain. The collected samples were studied in 1980 and stored in the collections of Orsay
University. However only two subfossil lake samples escaped vandalization in a subsequent
reorganization of storage.
). Its findings were presented at the 1987 Kuwait Conference on Quaternary Sediments in the Arabian Gulf and Mesopotamian
region, the proceedings of which have never been issued. Consequently, the ecological
field data of the Mesopotamian model have not been
available, so we offer this revised version of our
contribution in the hope of inciting greater interest to the environments of the Mesopotamian plain. The collected samples were studied in 1980 and stored in the collections of Orsay
University. However only two subfossil lake samples escaped vandalization in a subsequent
reorganization of storage.
![]()
![]() The Quaternary sediments underlying the alluvial plain of lower Mesopotamia are predominantly fluvial deposits but include a thin Holocene marine unit, the Hammar Formation ( et alii,
1957), that has been reported to extend from Basrah to
Najaf, i.e. more than 450 km inland from the present mouth of the Shatt al Arab estuary. Study of the paleogeographic evolution of the historic "Mesopotamia" (southern Sumeria) began before the Second World War. Figure 23 in
(1980) reproduces for the first time
maps showing these reconstructions as they appeared in the anonymous "Handbook of Iraq and the Persian Gulf" (War office, 1944). Subsequent research on the content and distribution of this marine unit introduced paleontologic studies and elucidated its significance concerning local subsidence and tectonics ( et alii,
1957; & ,
1957; , 1958; & ,
1966; & ,
1978; & ,
1978). The importance of eustacy was emphasized later ( & ,
1988,
Fig. 19). This Late Quaternary marine to lagoonal incursion was the transgressive head of the Arabian-Persian Gulf that attained its highest level during the Holocene Optimum
(Fig. 1
The Quaternary sediments underlying the alluvial plain of lower Mesopotamia are predominantly fluvial deposits but include a thin Holocene marine unit, the Hammar Formation ( et alii,
1957), that has been reported to extend from Basrah to
Najaf, i.e. more than 450 km inland from the present mouth of the Shatt al Arab estuary. Study of the paleogeographic evolution of the historic "Mesopotamia" (southern Sumeria) began before the Second World War. Figure 23 in
(1980) reproduces for the first time
maps showing these reconstructions as they appeared in the anonymous "Handbook of Iraq and the Persian Gulf" (War office, 1944). Subsequent research on the content and distribution of this marine unit introduced paleontologic studies and elucidated its significance concerning local subsidence and tectonics ( et alii,
1957; & ,
1957; , 1958; & ,
1966; & ,
1978; & ,
1978). The importance of eustacy was emphasized later ( & ,
1988,
Fig. 19). This Late Quaternary marine to lagoonal incursion was the transgressive head of the Arabian-Persian Gulf that attained its highest level during the Holocene Optimum
(Fig. 1
![]() ). The marine strata are interfingered between the fluviatile, marsh and lacustrine deposits of the Tigris and Euphrates rivers that progressively built parallel internal deltas NW of their present confluence at Qurna. In a few thousand years the head of the Gulf moved southeastward more than 250 km, replaced south of Basrah by the Shatt al Arab delta, except in the west where a residual marine channel, the Khor Zubair, may be interpreted as
a marginal bay of this delta
(Fig. 2
). The marine strata are interfingered between the fluviatile, marsh and lacustrine deposits of the Tigris and Euphrates rivers that progressively built parallel internal deltas NW of their present confluence at Qurna. In a few thousand years the head of the Gulf moved southeastward more than 250 km, replaced south of Basrah by the Shatt al Arab delta, except in the west where a residual marine channel, the Khor Zubair, may be interpreted as
a marginal bay of this delta
(Fig. 2
![]() ). The Shatt al Arab delta is mainly a contribution of the Karun river,
for the detritus of its flood stages is added to that of the two major Mesopotamian rivers ( et alii,
1982; & ,
1990). The excess of Karun alluvium, derived from the northern Zagros mountains, abuts the coarse-grained Wadi al Batin alluvial fan, the detritus of the Arabian shield. Together, they narrowed the depression south of Basrah and contribute to the
recent southeastward development of the Shatt al Arab delta.
). The Shatt al Arab delta is mainly a contribution of the Karun river,
for the detritus of its flood stages is added to that of the two major Mesopotamian rivers ( et alii,
1982; & ,
1990). The excess of Karun alluvium, derived from the northern Zagros mountains, abuts the coarse-grained Wadi al Batin alluvial fan, the detritus of the Arabian shield. Together, they narrowed the depression south of Basrah and contribute to the
recent southeastward development of the Shatt al Arab delta.
![]() Studies of wells, pits and outcrops of the Hammar Formation show that its upper surface has been affected by neotectonic deformation ranging from -2 to +40 m. The main uplift has occurred on the NW margin of the basin (at Najaf, cf. & ,
1978). As a result of this deformation, marine and estuarine fossils have been found reworked in deposits dominated by modern freshwater organisms. These puzzling faunal assemblages demonstrate the possible influence of early reworking,
a phenomenon which may help in elucidating difficult paleogeographic problems such as those of the Eastern Mediterranean Messinian (cf. et alii,
2000).
Studies of wells, pits and outcrops of the Hammar Formation show that its upper surface has been affected by neotectonic deformation ranging from -2 to +40 m. The main uplift has occurred on the NW margin of the basin (at Najaf, cf. & ,
1978). As a result of this deformation, marine and estuarine fossils have been found reworked in deposits dominated by modern freshwater organisms. These puzzling faunal assemblages demonstrate the possible influence of early reworking,
a phenomenon which may help in elucidating difficult paleogeographic problems such as those of the Eastern Mediterranean Messinian (cf. et alii,
2000).
![]() Accordingly, the interpretation of paleoenvironments merits a discussion based mainly on their molluscan faunas. The ecology of the most common living
molluscs was documented during a one month field reconnaissance (in March 1980), carried out in collaboration with the Geological Survey of Iraq and completed in France by the study of sedimentary and biological samples. Only a part of the findings have been published by et alii
(1982), (1987) and &
(1990).
Accordingly, the interpretation of paleoenvironments merits a discussion based mainly on their molluscan faunas. The ecology of the most common living
molluscs was documented during a one month field reconnaissance (in March 1980), carried out in collaboration with the Geological Survey of Iraq and completed in France by the study of sedimentary and biological samples. Only a part of the findings have been published by et alii
(1982), (1987) and &
(1990).
(data from et alii, 1982 and , 1986)
![]() The lower
Mesopotamian plain, north of 30°N, is in the submediterranean (subtropical) belt, with a local arid character (60-170 mm.yr-1 rainfall) counterbalanced by the fluvial freshwater input from temperate mountains, via the
Tigris and Euphrates rivers, both originating in the Kurdish portion of Turkey. Evaporation fluctuates from 50 to 250/600 mm a month (January and July means), highest in July heat (31-35°C, up to 50°C) but
is still significant in winter (10-12°C, January
mean) because of the strong NW to SE prevailing winds (Shamal). The marsh and lake area is
affected not only by contrasting local climatic conditions (temperature, hygrometry) but also by marked fluctuations in the delivery of exotic freshwater. Prior to the end of the XXth century the discharge rate of the Tigris ranged from 3000 m3.sec-1 to less than 500 m3.sec-1
while that of the Euphrates ranged from 2000 m3.sec-1 to less than 250 m3.sec-1, thus accounting for the variations in T.D.S. (total dissolved salt) of the rivers’
waters (0.4 to 0.75‰ or
g.l-1). The origin of these rivers in mountains of the temperate zone accounts for the fact that their high water stage is restricted to the February-June period during which rain is
abundant and snow melts. This is reflected by variation in the salinity of the Shatt al Arab estuary: at
Abadan salinity remains below 1‰ from February to July while attaining 8‰ in November and December. In accord with these factors marshes and associated lakes had surface water salinities ranging around 1 to 2‰ in March 1980 ( et alii,
1982). A large part of this low area is subject to seasonal flooding and drying; the peripheral subemergent to emergent Typha swamps are grazed by buffalo
(Fig. A
The lower
Mesopotamian plain, north of 30°N, is in the submediterranean (subtropical) belt, with a local arid character (60-170 mm.yr-1 rainfall) counterbalanced by the fluvial freshwater input from temperate mountains, via the
Tigris and Euphrates rivers, both originating in the Kurdish portion of Turkey. Evaporation fluctuates from 50 to 250/600 mm a month (January and July means), highest in July heat (31-35°C, up to 50°C) but
is still significant in winter (10-12°C, January
mean) because of the strong NW to SE prevailing winds (Shamal). The marsh and lake area is
affected not only by contrasting local climatic conditions (temperature, hygrometry) but also by marked fluctuations in the delivery of exotic freshwater. Prior to the end of the XXth century the discharge rate of the Tigris ranged from 3000 m3.sec-1 to less than 500 m3.sec-1
while that of the Euphrates ranged from 2000 m3.sec-1 to less than 250 m3.sec-1, thus accounting for the variations in T.D.S. (total dissolved salt) of the rivers’
waters (0.4 to 0.75‰ or
g.l-1). The origin of these rivers in mountains of the temperate zone accounts for the fact that their high water stage is restricted to the February-June period during which rain is
abundant and snow melts. This is reflected by variation in the salinity of the Shatt al Arab estuary: at
Abadan salinity remains below 1‰ from February to July while attaining 8‰ in November and December. In accord with these factors marshes and associated lakes had surface water salinities ranging around 1 to 2‰ in March 1980 ( et alii,
1982). A large part of this low area is subject to seasonal flooding and drying; the peripheral subemergent to emergent Typha swamps are grazed by buffalo
(Fig. A
![]() ). Permanent lakes are located not only in
the interior of the extensive marshes but also in the barren sabkhas that fringe the southern desert. The high salt content of these southernmost lakes (15/20‰ TDS in March) precludes the existence of a molluscan biota, whereas the extensive palustrine and lacustrine areas harbour the malacofauna reported below.
). Permanent lakes are located not only in
the interior of the extensive marshes but also in the barren sabkhas that fringe the southern desert. The high salt content of these southernmost lakes (15/20‰ TDS in March) precludes the existence of a molluscan biota, whereas the extensive palustrine and lacustrine areas harbour the malacofauna reported below.
![]() The taxonomy of molluscan species and their ecological requirements needs
reexamination because these subjects have been neglected for a considerable period and sometimes misinterpreted (,
1918). The freshwater basin of Lower Mesopotamia includes an extensive association of reeds (Phragmites and Typha)
in the marshlands, controlled by the existence of a ground-water table that is in equilibrium with the flowing rivers. These rivers are almost completely isolated from the adjacent marshes by alluvial levees of several types. The lotic waters of isolated lakes bounded by a reed filter are typical shallow interdistributary (holomictic) lakes. They are characterized by unmixed carbonate sedimentation ( et alii,
1982; & ,
1982).
The taxonomy of molluscan species and their ecological requirements needs
reexamination because these subjects have been neglected for a considerable period and sometimes misinterpreted (,
1918). The freshwater basin of Lower Mesopotamia includes an extensive association of reeds (Phragmites and Typha)
in the marshlands, controlled by the existence of a ground-water table that is in equilibrium with the flowing rivers. These rivers are almost completely isolated from the adjacent marshes by alluvial levees of several types. The lotic waters of isolated lakes bounded by a reed filter are typical shallow interdistributary (holomictic) lakes. They are characterized by unmixed carbonate sedimentation ( et alii,
1982; & ,
1982).
![]() The Mesopotamian freshwater malacofauna was first described by
(1874). He described, but did not illustrate the fauna collected by before 1860. In fact, the majority of the species he considered to be new is synonymous with species described previously.
Subsequent studies of them and their ecology included misinterpretations (,
1918) so their ecological requirements stand
in need of reexamination. Their taxonomic revision was done to some degree, largely from a systematic viewpoint, by
(1918, 1920),
& (1919),
(1924)
and (1969). New collections by
(1961) and the field survey by our group have made possible a general review of the more important species.
The Mesopotamian freshwater malacofauna was first described by
(1874). He described, but did not illustrate the fauna collected by before 1860. In fact, the majority of the species he considered to be new is synonymous with species described previously.
Subsequent studies of them and their ecology included misinterpretations (,
1918) so their ecological requirements stand
in need of reexamination. Their taxonomic revision was done to some degree, largely from a systematic viewpoint, by
(1918, 1920),
& (1919),
(1924)
and (1969). New collections by
(1961) and the field survey by our group have made possible a general review of the more important species.
![]() Bivalve molluscs generally dominate subaquatic environments.
Bivalve molluscs generally dominate subaquatic environments.
![]() Corbicula fluminalis
(Pl. 3, fig. 19-29
Corbicula fluminalis
(Pl. 3, fig. 19-29
![]() ) is a highly variable species ranging from a very thick, triangular morph (C. cor ), to a flatter, elongate morph (C. tigridis ). The ratio of height (umbono-ventral) to length (antero-posterior) dimensions may serve to characterize these morphotypes. It fluctuates between the median values of h/l = 1 (high morphotype) and h/l = 0.875 (elongate morphotype). Their respective distributions do not reflect their
environments during life: the elongate morph occurs and may dominate in all of them: lakes, marshes, and the rapidly flowing Tigris. Neither is it characteristic of any one drainage
system. The elongate morph represents from 30 to 100% of the Corbicula shells in the Tigris channel, versus
13 to 17% in the Euphrates channel, but it dominates in the Euphrates-associated lakes and swamps (up to 100% of Corbicula shells). Dominance of this genus (up to 100% of the assemblage) locally characterizes fluviatile channel environments
for both living and empty shells are buried in the fine sands and muds of this
milieu. It is also abundant in the bottom muds of many lakes and marsh channels but there it is associated with a more varied fauna, enriched by Unionids and gastropods
(Pl. 1, fig. 2-3 & 6
) is a highly variable species ranging from a very thick, triangular morph (C. cor ), to a flatter, elongate morph (C. tigridis ). The ratio of height (umbono-ventral) to length (antero-posterior) dimensions may serve to characterize these morphotypes. It fluctuates between the median values of h/l = 1 (high morphotype) and h/l = 0.875 (elongate morphotype). Their respective distributions do not reflect their
environments during life: the elongate morph occurs and may dominate in all of them: lakes, marshes, and the rapidly flowing Tigris. Neither is it characteristic of any one drainage
system. The elongate morph represents from 30 to 100% of the Corbicula shells in the Tigris channel, versus
13 to 17% in the Euphrates channel, but it dominates in the Euphrates-associated lakes and swamps (up to 100% of Corbicula shells). Dominance of this genus (up to 100% of the assemblage) locally characterizes fluviatile channel environments
for both living and empty shells are buried in the fine sands and muds of this
milieu. It is also abundant in the bottom muds of many lakes and marsh channels but there it is associated with a more varied fauna, enriched by Unionids and gastropods
(Pl. 1, fig. 2-3 & 6
![]() ).
).
![]() The Unionids
(Pl. 2
The Unionids
(Pl. 2
![]() ) were revised and reclassified by
(1969) in a review on a worldwide scale. They are assigned to three genera. Unio tigridis , the most common species, has a solid elongate shell buried obliquely in the mud, its posterior end projecting from the bottom in lake and marsh channels
(Fig. C.4
) were revised and reclassified by
(1969) in a review on a worldwide scale. They are assigned to three genera. Unio tigridis , the most common species, has a solid elongate shell buried obliquely in the mud, its posterior end projecting from the bottom in lake and marsh channels
(Fig. C.4
![]() , C.6
, C.6
![]() , and D.4
, and D.4
![]() ). Pseudodontopsis euphraticus () is a large and solid
rhomboidal flat shell. It burrows in the bottom muds of lakes and marshes
(Pl. 1, fig. 6
). Pseudodontopsis euphraticus () is a large and solid
rhomboidal flat shell. It burrows in the bottom muds of lakes and marshes
(Pl. 1, fig. 6
![]() ). Anodonta
(Anodonta) vescoiana is the rarest and has a large, thin inflated shell. It is indicative of the quietest lacustrine environments (Fig.
C.3
). Anodonta
(Anodonta) vescoiana is the rarest and has a large, thin inflated shell. It is indicative of the quietest lacustrine environments (Fig.
C.3 ![]() ,
C.6
,
C.6 ![]() , and
D.1
, and
D.1 ![]() ). The large size and
post-mortem accumulation (lag deposit) of the shells of Unionacea may cause an overestimation of their number in some marsh channels even though their preferred environments are the quiet (lotic) lake and the mud bottoms of Typha marshes.
). The large size and
post-mortem accumulation (lag deposit) of the shells of Unionacea may cause an overestimation of their number in some marsh channels even though their preferred environments are the quiet (lotic) lake and the mud bottoms of Typha marshes.
![]() The thick-shelled Corbicula, Unio and Pseudontopsis as well as the thinner Anodonta shells are not often transported far from their habitat in life. Only high-energy lake shores and river banks may harbor a few displaced valves of Corbicula
(Pl. 1, fig. 5
The thick-shelled Corbicula, Unio and Pseudontopsis as well as the thinner Anodonta shells are not often transported far from their habitat in life. Only high-energy lake shores and river banks may harbor a few displaced valves of Corbicula
(Pl. 1, fig. 5
![]() ).
).
![]() Gastropods are more diversified. They include 7 well-represented species, and a few rarer forms.
Gastropods are more diversified. They include 7 well-represented species, and a few rarer forms.
![]() Theodoxus (Neritaea) jordani ()
(Pl. 4, fig. 11-20
Theodoxus (Neritaea) jordani ()
(Pl. 4, fig. 11-20
![]() ). The more or less pronounced degree of transverse constriction on the last whorl of this species gave rise to the naming of three varieties (,
1921,
Fig. 1). The species is exceptionally widespread (lakes, marsh channels, fluviatile channels, and estuary). The other Mesopotamian Neritaea species retained by : T. mesopotamicus, T. euphraticus () and T. maccrii () are also varieties or morphs (cf. ,
1975).
). The more or less pronounced degree of transverse constriction on the last whorl of this species gave rise to the naming of three varieties (,
1921,
Fig. 1). The species is exceptionally widespread (lakes, marsh channels, fluviatile channels, and estuary). The other Mesopotamian Neritaea species retained by : T. mesopotamicus, T. euphraticus () and T. maccrii () are also varieties or morphs (cf. ,
1975).
![]() Melanopsis (Melanopsis) praemorsum ()
(Pl. 4, fig. 29-36
Melanopsis (Melanopsis) praemorsum ()
(Pl. 4, fig. 29-36
![]() ) is the name currently given to the diverse nodular, costulate or smooth morphs of North African and Middle Eastern Melanopsis (,
1975; , 1980; ,
1984). The countless species and varieties attributed to the Melanopsis subgenus (or to
Canthidomus for the costulate morphs) are now considered as a single polymorphic species. It too is ubiquitous, living in lakes and inundated marshes
(Fig. C.5
) is the name currently given to the diverse nodular, costulate or smooth morphs of North African and Middle Eastern Melanopsis (,
1975; , 1980; ,
1984). The countless species and varieties attributed to the Melanopsis subgenus (or to
Canthidomus for the costulate morphs) are now considered as a single polymorphic species. It too is ubiquitous, living in lakes and inundated marshes
(Fig. C.5
![]() and D.3
and D.3
![]() ) (mainly on subaquatic plants) or on the mud or gravel intertidal shores of estuaries
(Pl. 1, fig. 1
) (mainly on subaquatic plants) or on the mud or gravel intertidal shores of estuaries
(Pl. 1, fig. 1
![]() ). The nodular morphology is more common in freshwater environments and the smoother form
(Pl. 4, fig. 32
). The nodular morphology is more common in freshwater environments and the smoother form
(Pl. 4, fig. 32
![]() ) appears to be more abundant in estuarine waters.
) appears to be more abundant in estuarine waters.
![]() Melanoides tuberculata ()
(Fig. C
Melanoides tuberculata ()
(Fig. C
![]() ; Pl. 4, fig.
7-10
; Pl. 4, fig.
7-10
![]() ) is an ubiquitous
(African and Euro-Asiatic), euryhaline species generally associated with environments of low salinity (0.2-3‰) (,
1983) where it may predominate, but it tolerates salinities of up to 23‰. It dwells on subaquatic soft bottoms, often concealed in the sediment where its sinuous track is a characteristic token of its presence in freshwater lakes
(Fig. C.5
) is an ubiquitous
(African and Euro-Asiatic), euryhaline species generally associated with environments of low salinity (0.2-3‰) (,
1983) where it may predominate, but it tolerates salinities of up to 23‰. It dwells on subaquatic soft bottoms, often concealed in the sediment where its sinuous track is a characteristic token of its presence in freshwater lakes
(Fig. C.5
![]() ). Empty shells float easily and consequently accumulate on the shores of lakes
(Fig. D.2
). Empty shells float easily and consequently accumulate on the shores of lakes
(Fig. D.2
![]() ; Pl. 1, fig. 7
; Pl. 1, fig. 7
![]() ).
).
![]() Bellamya bengalensis ()
(Pl. 3, fig. 13-18
Bellamya bengalensis ()
(Pl. 3, fig. 13-18
![]() ) is a large viviparid snail, living in quiet fresh water. As it was not recorded by
(1874), (1818) and
& (1921),
(1961) considered it to be a recent immigrant. It lives on the bottom muds of ponds, marshes and marsh channels
(Fig. D.3
) is a large viviparid snail, living in quiet fresh water. As it was not recorded by
(1874), (1818) and
& (1921),
(1961) considered it to be a recent immigrant. It lives on the bottom muds of ponds, marshes and marsh channels
(Fig. D.3
![]() ) but during flood high waters, the empty shells float and are stranded up to the edge of peripheral emergent reliefs in marsh
areas, thus delineating high water marks. It is also present in estuarine shore deposits
(Pl. 1, fig. 5-6
) but during flood high waters, the empty shells float and are stranded up to the edge of peripheral emergent reliefs in marsh
areas, thus delineating high water marks. It is also present in estuarine shore deposits
(Pl. 1, fig. 5-6
![]() ).
).
![]() The small Planorbs - Gyraulus convexiusculus (), G. albus () and G. intermixtus ()
(Pl. 3, fig. 9-12
The small Planorbs - Gyraulus convexiusculus (), G. albus () and G. intermixtus ()
(Pl. 3, fig. 9-12
![]() ) - commonly are not very abundant
and are generally in association with the most diverse of the freshwater assemblages. Their usual habitat is the subaqueous portion of the rooted vegetation of lakes and swamps.
) - commonly are not very abundant
and are generally in association with the most diverse of the freshwater assemblages. Their usual habitat is the subaqueous portion of the rooted vegetation of lakes and swamps.
![]() Lymnaea (Radix) gr. auricularia
(Pl. 3, fig. 1-8
Lymnaea (Radix) gr. auricularia
(Pl. 3, fig. 1-8
![]() ) has been identified under the specific names tenera , euphratica , canalifera (in Mesopotamia) and lagotis (in India, & ,
1925). Probably all are polymorphs of L. (R.) auricularia. This species is mainly lacustrine and palustrine, the empty shells commonly float and are stranded along the shores
(Fig. D.2
) has been identified under the specific names tenera , euphratica , canalifera (in Mesopotamia) and lagotis (in India, & ,
1925). Probably all are polymorphs of L. (R.) auricularia. This species is mainly lacustrine and palustrine, the empty shells commonly float and are stranded along the shores
(Fig. D.2
![]() ; Pl. 1, fig. 7
; Pl. 1, fig. 7
![]() ).
).
![]() The Hydrobids are neither diverse nor abundant.
(1921) recorded the Pomatopsidae Tricula palmyrae (), the Bithyniidae Bithynia badiella and the Hydrobiidae Amnicola (Alocinna) ejecta (). These shells are uncommon (< 1%) and in freshwater assemblages are locally rarer than their opercula. We also collected a Valvata sp. (rare) and a more common Ancylidae in flowing fluvial and marsh channels. Bulinus contortus was not seen in 1980, although it was figured by
(1918) and recorded by
(1874).
The Hydrobids are neither diverse nor abundant.
(1921) recorded the Pomatopsidae Tricula palmyrae (), the Bithyniidae Bithynia badiella and the Hydrobiidae Amnicola (Alocinna) ejecta (). These shells are uncommon (< 1%) and in freshwater assemblages are locally rarer than their opercula. We also collected a Valvata sp. (rare) and a more common Ancylidae in flowing fluvial and marsh channels. Bulinus contortus was not seen in 1980, although it was figured by
(1918) and recorded by
(1874).
![]() Shell assemblages in the marsh-channels
(Pl. 1, fig. 4
Shell assemblages in the marsh-channels
(Pl. 1, fig. 4
![]() ), with active current systems ( et alii,
1982) are significantly more diverse and abundant than those of quiet-water environments: the high concentration of living and empty shells on their floors results probably from the combination of a high rate of production, the winnowing of fine sediment and some transport from adjacent quieter environments
(Bellamya, Pulmonates).
), with active current systems ( et alii,
1982) are significantly more diverse and abundant than those of quiet-water environments: the high concentration of living and empty shells on their floors results probably from the combination of a high rate of production, the winnowing of fine sediment and some transport from adjacent quieter environments
(Bellamya, Pulmonates).
![]() The annual flood, coincident with the beginning of warm weather, tends to limit the increase in salinity caused by evaporation. Thus, the estuarine fauna of the Shatt al Arab consists mainly of freshwater molluscs, except near the mouth: at Fao the salinity
attained 2‰
in March 1980. On the other hand, certain of the previously reviewed freshwater species are euryhaline and can tolerate salinities of more than 2‰ during both winter and summer reductions in the delivery of fresh water. This is true in particular of Melanopsis and Melanoides spp. They can live and even predominate in most of the estuary. Only at its mouth does the malacofauna change with the introduction of
the genera Neritina (Dostia) and Cerithidea (Cerithideopsilla).
The annual flood, coincident with the beginning of warm weather, tends to limit the increase in salinity caused by evaporation. Thus, the estuarine fauna of the Shatt al Arab consists mainly of freshwater molluscs, except near the mouth: at Fao the salinity
attained 2‰
in March 1980. On the other hand, certain of the previously reviewed freshwater species are euryhaline and can tolerate salinities of more than 2‰ during both winter and summer reductions in the delivery of fresh water. This is true in particular of Melanopsis and Melanoides spp. They can live and even predominate in most of the estuary. Only at its mouth does the malacofauna change with the introduction of
the genera Neritina (Dostia) and Cerithidea (Cerithideopsilla).
![]() Neritina (Dostia) schlaeflii
(Pl. 4, fig. 21-29
Neritina (Dostia) schlaeflii
(Pl. 4, fig. 21-29
![]() ). This Nerita-like Neritina is closely related to N. (D.) violacea (= crepidularia) but
(1921) considered it to be a valid species. It forages in the muddy intertidal zone near Fao. It was not recorded in the
buried marine Hammar Formation but we collected it in a near surface fossil assemblage, N of Basrah
(Pl. 4, fig.
27
). This Nerita-like Neritina is closely related to N. (D.) violacea (= crepidularia) but
(1921) considered it to be a valid species. It forages in the muddy intertidal zone near Fao. It was not recorded in the
buried marine Hammar Formation but we collected it in a near surface fossil assemblage, N of Basrah
(Pl. 4, fig.
27
![]() ), thus indicating that it was present relatively early.
), thus indicating that it was present relatively early.
![]() Cerithidea (Cerithideopsilla) cingulata ()
(Pl. 4, fig.
1-5
Cerithidea (Cerithideopsilla) cingulata ()
(Pl. 4, fig.
1-5
![]() ). Until the mid XXth century, this species was erroneously referred to Potamides or Tympanotonus fluviatilis. In the Hammar Formation it is represented by a dwarf morph, less than 2.5 cm in length, as are the living specimens now restricted to the Fao area. During the Holocene its range extended northwestward as far as Nassiriyah.
(1918) supposed that it lived there in slightly saline, lacustrine waters. Later work showed that its range was even more extensive: up to the vicinity of Ur (,
1960) and northward to Amarah ( & ,
1978). We collected great quantities of this species in fossil shoreline accumulations (beach-rocks), between Nassiriyah and south of Lake Logait where the deposits of the Holocene transgression crop out. This intertidal potamid is well known around the Gulf (Iran, Arabian peninsula) as a marine species, tolerating high salinities (40-50‰ metahaline waters). However its extreme euryhalinity enables it to survive in quiet waters below 1.5‰ (,
1989) if these waters are directly connected with the sea (estuarine mud-flats).
). Until the mid XXth century, this species was erroneously referred to Potamides or Tympanotonus fluviatilis. In the Hammar Formation it is represented by a dwarf morph, less than 2.5 cm in length, as are the living specimens now restricted to the Fao area. During the Holocene its range extended northwestward as far as Nassiriyah.
(1918) supposed that it lived there in slightly saline, lacustrine waters. Later work showed that its range was even more extensive: up to the vicinity of Ur (,
1960) and northward to Amarah ( & ,
1978). We collected great quantities of this species in fossil shoreline accumulations (beach-rocks), between Nassiriyah and south of Lake Logait where the deposits of the Holocene transgression crop out. This intertidal potamid is well known around the Gulf (Iran, Arabian peninsula) as a marine species, tolerating high salinities (40-50‰ metahaline waters). However its extreme euryhalinity enables it to survive in quiet waters below 1.5‰ (,
1989) if these waters are directly connected with the sea (estuarine mud-flats).
![]() Potamides conicus () appears to be absent on Iraqi shores although abundant on the Arabian coasts (NE Saudi Arabia, Trucial coast, Oman), usually living in the intertidal zone somewhat closer to shore than the sympatric Cerithidea cingulata. Thus, it tolerates intense stresses but seems to be better adapted to high salinities than to oligo- to mesohaline waters (,
1989, 1993). We did not collect it in the Hammar Formation, but a slender morph of Cerithidea cingulata like the one living in the lagoonal environment of Khor al Hajar, Oman could be confused with it. During late Holocene times P. conicus was abundant on Kuwaiti shores (pers. obs. and unpublished post-Holocene Optimum dates) and therefore seems to have disappeared only recently from the head of the Gulf.
Potamides conicus () appears to be absent on Iraqi shores although abundant on the Arabian coasts (NE Saudi Arabia, Trucial coast, Oman), usually living in the intertidal zone somewhat closer to shore than the sympatric Cerithidea cingulata. Thus, it tolerates intense stresses but seems to be better adapted to high salinities than to oligo- to mesohaline waters (,
1989, 1993). We did not collect it in the Hammar Formation, but a slender morph of Cerithidea cingulata like the one living in the lagoonal environment of Khor al Hajar, Oman could be confused with it. During late Holocene times P. conicus was abundant on Kuwaiti shores (pers. obs. and unpublished post-Holocene Optimum dates) and therefore seems to have disappeared only recently from the head of the Gulf.
![]() Stenothyra iraqensis
et
(1966) is a small hydrobid species of the Stenothyridae family described from the Hammar Formation at Amarah. This genus is easily identified by its aperture smaller than the preceding whorl. We collected it from that shallow marine unit (borehole of Lake Logait) and from partly reworked assemblages supposedly derived from
the same formation (N of Lake Hammar).
Stenothyra iraqensis
et
(1966) is a small hydrobid species of the Stenothyridae family described from the Hammar Formation at Amarah. This genus is easily identified by its aperture smaller than the preceding whorl. We collected it from that shallow marine unit (borehole of Lake Logait) and from partly reworked assemblages supposedly derived from
the same formation (N of Lake Hammar).
![]() Theora mesopotamica () is a tellinimorph bivalve species typifying the Hammar Fm. This banal name unfortunately must replace the amusing Abra cadabra of
& (1957) as
(1918) described and figured it under the name
Corbula (Erodona) mesopotamica, from "subfossil sandy beds at Nasarieh". It was subsequently moved from the genus Abra to the Semelidaean genus Theora by P.G.
(in et alii,
1995,
p. 263) although retaining the erroneous species name, T. cadabra. This material is undoubtedly from the Hammar Formation where we too collected it and where it is associated with a typical brackish oligospecific assemblage. It exists now in the north and northwest portions of the Gulf ( et alii,
1995).
Theora mesopotamica () is a tellinimorph bivalve species typifying the Hammar Fm. This banal name unfortunately must replace the amusing Abra cadabra of
& (1957) as
(1918) described and figured it under the name
Corbula (Erodona) mesopotamica, from "subfossil sandy beds at Nasarieh". It was subsequently moved from the genus Abra to the Semelidaean genus Theora by P.G.
(in et alii,
1995,
p. 263) although retaining the erroneous species name, T. cadabra. This material is undoubtedly from the Hammar Formation where we too collected it and where it is associated with a typical brackish oligospecific assemblage. It exists now in the north and northwest portions of the Gulf ( et alii,
1995).
![]() A small oyster, Saccostrea cuccullata , is also locally abundant in the littoral Hammar Formation deposits. South of
Lake Logait it has been dated
4310 y BP ± 160 ( et alii, 1982).
(1958) recorded it near Najaf and &
(1978) at Amarah.
A small oyster, Saccostrea cuccullata , is also locally abundant in the littoral Hammar Formation deposits. South of
Lake Logait it has been dated
4310 y BP ± 160 ( et alii, 1982).
(1958) recorded it near Najaf and &
(1978) at Amarah.
![]() The freshwater malacofauna of this extensive swamp and lake area is a significant reference for paleoenvironmental reconstructions of Cenozoic successions. It may be directly compared with that of the contemporaneous tropical Lake Chad (,
1972; , 1980) where marshes with salinities of less than 1‰ have a comparable area.
The freshwater malacofauna of this extensive swamp and lake area is a significant reference for paleoenvironmental reconstructions of Cenozoic successions. It may be directly compared with that of the contemporaneous tropical Lake Chad (,
1972; , 1980) where marshes with salinities of less than 1‰ have a comparable area.
![]() Molluscs in Lake Chad do not live in salinities of more than 0.9‰ whereas the Mesopotamian freshwater malacofauna tolerates at least 3‰
(Hammar lake and marsh complex). The benthic African malacofauna too is dominated by Prosobranch gastropods: 83% of the molluscan biomass consists of Bellamya unicolor (45% of all molluscan specimens), the mud dweller Melanoides tuberculata, and Cleopatra bulimoides that replaces another
Thiaridae, our Melanopsis. The occurrence of
Bellamya piled high on the paleodune shores is, as in Iraq, explained by the buoyancy of the empty Viviparid shells. Corbicula africana prefers sandy bottoms, but does not appear to require a specific environment, unlike the unionids Caelatura (current) and Byssanodonta (hard substrate for
attachment). The African Pisidium (< 1%) has no equivalent in the Mesopotamian fauna. Pulmonates living on subaquatic plants are mainly large Planorbids and Bulinus
spp. Thus, despite these minor differences, there is no real inconsistency between the faunas of the Sahelian tropical shallow lakes with aquatic vegetation and the extensive subtropical Mesopotamian marshes and associated lakes.
Molluscs in Lake Chad do not live in salinities of more than 0.9‰ whereas the Mesopotamian freshwater malacofauna tolerates at least 3‰
(Hammar lake and marsh complex). The benthic African malacofauna too is dominated by Prosobranch gastropods: 83% of the molluscan biomass consists of Bellamya unicolor (45% of all molluscan specimens), the mud dweller Melanoides tuberculata, and Cleopatra bulimoides that replaces another
Thiaridae, our Melanopsis. The occurrence of
Bellamya piled high on the paleodune shores is, as in Iraq, explained by the buoyancy of the empty Viviparid shells. Corbicula africana prefers sandy bottoms, but does not appear to require a specific environment, unlike the unionids Caelatura (current) and Byssanodonta (hard substrate for
attachment). The African Pisidium (< 1%) has no equivalent in the Mesopotamian fauna. Pulmonates living on subaquatic plants are mainly large Planorbids and Bulinus
spp. Thus, despite these minor differences, there is no real inconsistency between the faunas of the Sahelian tropical shallow lakes with aquatic vegetation and the extensive subtropical Mesopotamian marshes and associated lakes.
![]() Study of the dried bottom of a recent polder of Lake Hammar
(Fig. C
Study of the dried bottom of a recent polder of Lake Hammar
(Fig. C
![]() ; Pl. 1, fig. 2-3
& 6
; Pl. 1, fig. 2-3
& 6
![]() ) enclosed by causeways a few years before
(Fig. A.3
) enclosed by causeways a few years before
(Fig. A.3
![]() ) showed that, like that of Typha and Phragmites swamps, the most diverse malacofauna exists where the lacustrine bottom has some rooted vegetation.
As we pointed out above, molluscan shells are more abundant on the floor of the marsh channels cleared for boat
traffic than they are anywhere else
(Pl. 1, fig. 4
) showed that, like that of Typha and Phragmites swamps, the most diverse malacofauna exists where the lacustrine bottom has some rooted vegetation.
As we pointed out above, molluscan shells are more abundant on the floor of the marsh channels cleared for boat
traffic than they are anywhere else
(Pl. 1, fig. 4
![]() ), due essentially to winnowing and low rates of sedimentation, good oxygenation and nutrient input, and a moderate importation of light shells from adjacent environments (depending on wind-induced currents), but
the biodiversity of the living population in these channels does not differ from that of the other marsh and lake
subenvironments.
), due essentially to winnowing and low rates of sedimentation, good oxygenation and nutrient input, and a moderate importation of light shells from adjacent environments (depending on wind-induced currents), but
the biodiversity of the living population in these channels does not differ from that of the other marsh and lake
subenvironments.
![]() From a methodological point of view, the local
mixture of fossils from a brackish-marine environment (restricted, lagoon-like shallow waters),
a few thousand years older than the existing modern freshwater assemblages, is a
good example of the errors in interpretation that may be caused by reworking. The resulting symmigies include
discrete fragments of Balanus in marsh deposits (North of Lake Hammar), Balanus, Cerithidea cingulata and Theora mesopotamica shells near
"Nasariyeh" (,
1918) and specimens of Cerithidea cingulata in the vicinity of Ur and at Basrah (,
1961). Some small Foraminifera, especially Miliolids, are reworked too,
as we found a few specimens in all freshwater deposits where terrigenous sedimentation has been active: in fluvial channels (Tigris, as far upstream as
Amarah; Euphrates up to Nassiriyah), and in marshes and channeled lakes, but not in the isolated lakes surrounded by marsh, where bottom sedimentation was unmixed, autochthonous carbonate.
From a methodological point of view, the local
mixture of fossils from a brackish-marine environment (restricted, lagoon-like shallow waters),
a few thousand years older than the existing modern freshwater assemblages, is a
good example of the errors in interpretation that may be caused by reworking. The resulting symmigies include
discrete fragments of Balanus in marsh deposits (North of Lake Hammar), Balanus, Cerithidea cingulata and Theora mesopotamica shells near
"Nasariyeh" (,
1918) and specimens of Cerithidea cingulata in the vicinity of Ur and at Basrah (,
1961). Some small Foraminifera, especially Miliolids, are reworked too,
as we found a few specimens in all freshwater deposits where terrigenous sedimentation has been active: in fluvial channels (Tigris, as far upstream as
Amarah; Euphrates up to Nassiriyah), and in marshes and channeled lakes, but not in the isolated lakes surrounded by marsh, where bottom sedimentation was unmixed, autochthonous carbonate.
![]() On the other hand,
the interpretation of lacustrine foraminifera living occasionally in the salt lakes (e.g.
Lake Logait, 2-8‰ after winter rains) immediately south of the old deltaic complex, cannot be
ignored but remains doubtful.
Indeed, we consider that abundant microfaunas, like that of the lake at Abu Dibbis
(80 km NW of Najaf) recorded by &
(1978), should not be interpreted as "residual" but rather as the natural population of an intracontinental salt lake. Microfaunas of this type are similar to those of the African Holocene paleolakes (Great Western Erg, Algeria, cf. et alii,
1985; et alii, 1987) where a marine-like fauna (Potamides, Cerastoderma, Foraminifera, Ostracods) and microflora (Diatoms) must have been introduced by air (probably by water fowl or pelicans, as a human intervention is unlikely). Birket Karun (Fayum, Egypt) also possesses
many living Potamides, Cerastoderma, Balanus and divers foraminifera and ostracods but, here, human intervention may be suspected as their introduction can be dated, for it occurred during the XXth century (,
1991). One is therefore tempted to interpret the marine fauna of the Mesopotamian plain system as a result of aerial seeding, but there is a fundamental difference: in intracontinental salt lakes the marine-like fauna constitutes the whole or the dominant part of the population and it is especially
abundant. In Mesopotamia, on the contrary, the marine fossils are sparsely scattered among the unaltered remains of freshwater organisms and are commonly worn and broken.
On the other hand,
the interpretation of lacustrine foraminifera living occasionally in the salt lakes (e.g.
Lake Logait, 2-8‰ after winter rains) immediately south of the old deltaic complex, cannot be
ignored but remains doubtful.
Indeed, we consider that abundant microfaunas, like that of the lake at Abu Dibbis
(80 km NW of Najaf) recorded by &
(1978), should not be interpreted as "residual" but rather as the natural population of an intracontinental salt lake. Microfaunas of this type are similar to those of the African Holocene paleolakes (Great Western Erg, Algeria, cf. et alii,
1985; et alii, 1987) where a marine-like fauna (Potamides, Cerastoderma, Foraminifera, Ostracods) and microflora (Diatoms) must have been introduced by air (probably by water fowl or pelicans, as a human intervention is unlikely). Birket Karun (Fayum, Egypt) also possesses
many living Potamides, Cerastoderma, Balanus and divers foraminifera and ostracods but, here, human intervention may be suspected as their introduction can be dated, for it occurred during the XXth century (,
1991). One is therefore tempted to interpret the marine fauna of the Mesopotamian plain system as a result of aerial seeding, but there is a fundamental difference: in intracontinental salt lakes the marine-like fauna constitutes the whole or the dominant part of the population and it is especially
abundant. In Mesopotamia, on the contrary, the marine fossils are sparsely scattered among the unaltered remains of freshwater organisms and are commonly worn and broken.
![]() Accordingly, in the study of the Mesopotamian deposits (surface and
cores) the distribution of reworked marine fossils
will help to delimit the extent of the Hammar Formation. Its shoreline deposits on the Arabian side of Mesopotamia suggests that the existing, relatively deep marine channel of the western Khor Zubair, located near the front of the Dibdibba Formation (Wadi al Batin fan) is a residual expression of Holocene morphology. To the North, the extensive Quaternary gulf (Holocene Optimum transgression) documented by the most inland (Najaf and Amarah) marine deposits, must necessarily have been filled progressively over time, for the last marine deposits in the vicinity of Ur-Nassiriyah are contemporaneous with cultivation of the Garraf delta by Sumerians (Lagash, around 4500 y BP)
(Fig. 2
Accordingly, in the study of the Mesopotamian deposits (surface and
cores) the distribution of reworked marine fossils
will help to delimit the extent of the Hammar Formation. Its shoreline deposits on the Arabian side of Mesopotamia suggests that the existing, relatively deep marine channel of the western Khor Zubair, located near the front of the Dibdibba Formation (Wadi al Batin fan) is a residual expression of Holocene morphology. To the North, the extensive Quaternary gulf (Holocene Optimum transgression) documented by the most inland (Najaf and Amarah) marine deposits, must necessarily have been filled progressively over time, for the last marine deposits in the vicinity of Ur-Nassiriyah are contemporaneous with cultivation of the Garraf delta by Sumerians (Lagash, around 4500 y BP)
(Fig. 2
![]() ) while the more than meter-thick surficial deposits near the Sumerian cities of Uruk, Larsa and
'Oueili contain freshwater molluscs dated 1190 y BP (±100) or less ( & ,
1991). Four of the six 14C dates at our disposal
for the Hammar Formation serve only to confirm the mid-Holocene Optimum age of the transgression
(Fig. 1
) while the more than meter-thick surficial deposits near the Sumerian cities of Uruk, Larsa and
'Oueili contain freshwater molluscs dated 1190 y BP (±100) or less ( & ,
1991). Four of the six 14C dates at our disposal
for the Hammar Formation serve only to confirm the mid-Holocene Optimum age of the transgression
(Fig. 1
![]() ). Dating of samples spaced laterally in succession from the farthest inland trace of the incursion to the existing
shore will be required to determine the precise timing of the presumed stages of the regression.
). Dating of samples spaced laterally in succession from the farthest inland trace of the incursion to the existing
shore will be required to determine the precise timing of the presumed stages of the regression.
![]() Taking into account the tectonic deformation of the lower Mesopotamian basin in Quaternary times, a scenario involving a single regression is not the only hypothesis
possible. The elevations of the youngest marine deposits recorded by &
(1978) and et alii
(1982) demonstrate a major deformation : +40 m at Najaf, +2 m at Amarah,
+1 m S of Lake Logait, -1 m S of Lake Hammar, -4 to -13 m N of Zubair. As the geomorphological analysis of the marsh domain suggests a modern subsident axis from Nassiriyah to Qurna we cannot exclude the possibility that the undated marine deposits around Najaf may be of late Pleistocene age (pre-Hammar Fm: i.e. coincident with the Last Interglacial transgression, MIS 5.5).
Taking into account the tectonic deformation of the lower Mesopotamian basin in Quaternary times, a scenario involving a single regression is not the only hypothesis
possible. The elevations of the youngest marine deposits recorded by &
(1978) and et alii
(1982) demonstrate a major deformation : +40 m at Najaf, +2 m at Amarah,
+1 m S of Lake Logait, -1 m S of Lake Hammar, -4 to -13 m N of Zubair. As the geomorphological analysis of the marsh domain suggests a modern subsident axis from Nassiriyah to Qurna we cannot exclude the possibility that the undated marine deposits around Najaf may be of late Pleistocene age (pre-Hammar Fm: i.e. coincident with the Last Interglacial transgression, MIS 5.5).
![]() As regards the timing of the termination of marine communication between the Mesopotamian depression and the Gulf, we must consider the possibility of a sudden ending to the strait between the Karun delta and the Wadi al Batin fan, caused by the
eustatic (around one meter) drop in sea-level at the end of the Holocene Optimum. This
threshold (?) emersion left a continental depression several meters deep to be filled by alluvium from the two Mesopotamian rivers and by lake deposits. The preceding regression at the end of the Last Interglacial high sea-level episode (MIS 5.5) could not have followed the same scenario because the Karun river and Wadi al Batin fans were not as extensive then as they became later.
As regards the timing of the termination of marine communication between the Mesopotamian depression and the Gulf, we must consider the possibility of a sudden ending to the strait between the Karun delta and the Wadi al Batin fan, caused by the
eustatic (around one meter) drop in sea-level at the end of the Holocene Optimum. This
threshold (?) emersion left a continental depression several meters deep to be filled by alluvium from the two Mesopotamian rivers and by lake deposits. The preceding regression at the end of the Last Interglacial high sea-level episode (MIS 5.5) could not have followed the same scenario because the Karun river and Wadi al Batin fans were not as extensive then as they became later.
![]()
![]() The original, undisturbed environments of Lower Mesopotamia were destroyed twenty years ago but we suggest that the old files of field observations supplemented by new subsurface data would provide the foundation for a reliable reconstruction of the evolution of this fascinating
region. A correct interpretation of the evolution of the deltaic complex of Lower
Mesopotamia, with its important geodynamic and archeological implications, requires more precise dating
and improved mapping
(both in depth and areally) of the limits between marine and freshwater deposits of Holocene
(and possibly Pleistocene) age. Provided that paleoecological interpretations are made
carefully, the malacofauna is the best tool for this purpose, because reworking of shells appears to be
limited.
The original, undisturbed environments of Lower Mesopotamia were destroyed twenty years ago but we suggest that the old files of field observations supplemented by new subsurface data would provide the foundation for a reliable reconstruction of the evolution of this fascinating
region. A correct interpretation of the evolution of the deltaic complex of Lower
Mesopotamia, with its important geodynamic and archeological implications, requires more precise dating
and improved mapping
(both in depth and areally) of the limits between marine and freshwater deposits of Holocene
(and possibly Pleistocene) age. Provided that paleoecological interpretations are made
carefully, the malacofauna is the best tool for this purpose, because reworking of shells appears to be
limited.
![]() The respective assemblages of marshes and associated lakes are difficult
to differentiate. This justifies the notion of grouping under the name "palustrine"
the several environments of the fresh-water areas characterized by a fluctuating
water level (seasonally and interannually). These oscillations cause an intimate
association in one sedimentary unit of subaquatic and emergent conditions, each
marked by a discrete molluscan association and characteristic diagenetic
features.
The respective assemblages of marshes and associated lakes are difficult
to differentiate. This justifies the notion of grouping under the name "palustrine"
the several environments of the fresh-water areas characterized by a fluctuating
water level (seasonally and interannually). These oscillations cause an intimate
association in one sedimentary unit of subaquatic and emergent conditions, each
marked by a discrete molluscan association and characteristic diagenetic
features.
![]() Similarly, the malacofaunas of the major rivers and their common estuarine channel
show very limited differences, except at the mouth of the sea. Therefore
we assume that the presence of scarce marine species in a fresh-water assemblage
is indicative of the
existence of a reworked marine unit rather than their association in life in a
lagoonal or estuarine environment.
Similarly, the malacofaunas of the major rivers and their common estuarine channel
show very limited differences, except at the mouth of the sea. Therefore
we assume that the presence of scarce marine species in a fresh-water assemblage
is indicative of the
existence of a reworked marine unit rather than their association in life in a
lagoonal or estuarine environment.
![]() This brief review of the main subaquatic molluscs of Mesopotamia, compared with those of the Lake Chad malacofauna,
also suggests that the provincial effect does not preclude some useful generalizations regarding current interpretations of the Cenozoic paleoenvironments of the western Old World.
This brief review of the main subaquatic molluscs of Mesopotamia, compared with those of the Lake Chad malacofauna,
also suggests that the provincial effect does not preclude some useful generalizations regarding current interpretations of the Cenozoic paleoenvironments of the western Old World.
![]() From
a biogeographic point of view it is clear that the Mesopotamian freshwater
malacofauna belongs to the Mediterranean palearctic domain. It is accordingly interesting to note some eastern tropical influences in the brackish-water
gastropods (the modern Indo-West Pacific Cerithideopsilla,
Tricula and Stenothyra). Such a biogeographic distribution is not the first record of an
eastern link, the Stenothyridae family being present in European Paleogene
to Middle Miocene deposits (, 1984,
2005).
The Quaternary intruders from the Indo Pacific realm benefited from the warmer
transgressive episode of the Holocene Optimum, in the same way as the reef
and mangrove malacofaunas did when they invaded the northern Red Sea during the Last
Interglacial optimum (MIS 5.5) (, 1995;
et alii,
1998).
From
a biogeographic point of view it is clear that the Mesopotamian freshwater
malacofauna belongs to the Mediterranean palearctic domain. It is accordingly interesting to note some eastern tropical influences in the brackish-water
gastropods (the modern Indo-West Pacific Cerithideopsilla,
Tricula and Stenothyra). Such a biogeographic distribution is not the first record of an
eastern link, the Stenothyridae family being present in European Paleogene
to Middle Miocene deposits (, 1984,
2005).
The Quaternary intruders from the Indo Pacific realm benefited from the warmer
transgressive episode of the Holocene Optimum, in the same way as the reef
and mangrove malacofaunas did when they invaded the northern Red Sea during the Last
Interglacial optimum (MIS 5.5) (, 1995;
et alii,
1998).
![]() This report presents the results of field and laboratory studies (1980-1981) carried out by a team coordinated by B.H.
that included M. , N.H. , F. , K.M. , F. , S.Y. ,
and the
authors. This fruitful cooperation left a feeling of nostalgia for the scientific collaboration
that was possible in times of peace. Both the elaboration of and improvements in this contribution benefited from the help of F.
, B. , N. , P. and an anonymous
reviewer. Their aid is gratefully acknowledged.
This report presents the results of field and laboratory studies (1980-1981) carried out by a team coordinated by B.H.
that included M. , N.H. , F. , K.M. , F. , S.Y. ,
and the
authors. This fruitful cooperation left a feeling of nostalgia for the scientific collaboration
that was possible in times of peace. Both the elaboration of and improvements in this contribution benefited from the help of F.
, B. , N. , P. and an anonymous
reviewer. Their aid is gratefully acknowledged.
Gastropoda, Prosobranchia
Gastropoda, Basommatophora
Bivalvia
![]()
M. (1986).- La sédimentation actuelle sur la plaine de la Basse Mésopotamie (Irak).- Unpublished thesis, Paris-Sud Univ. (Orsay), 832 p., 172 fig., 68 pl.
T.N. (1918).- Freshwater shells from Mesopotamia.- Records of the Indian Museum, Calcutta, t. 15, pp. 159-170, 1 fig., 1 pl.
T.N. (1920).- Report on the freshwater Gastropod Molluscs of Lower Mesopotamia. Part II - The family Planorbidae.- Records of the Indian Museum, Calcutta, t. 18, pp. 147-149, 1 fig.
T.N. & B. (1919).- Report on the freshwater Gastropod molluscs of Lower Mesopotamia. Part 1: The genus Limnaea.- Records of the Indian Museum, Calcutta, t. 18, pp. 103-115.
T.N. & H.S. (1925).- Materials for a revision of the recent Indian Limnaeidae (Mollusca, Pulmonata).- Records of the Indian Museum, Calcutta, t. 27, pp. 137-189, 15 fig.
T.N. & R.B.S. (1921).- The banded pond snail of India: Vivipara bengalensis.- Records of the Indian Museum, Calcutta, t. 22, pp. 217-291, 22 fig., 3 pl.
F. & B.H. (1990).- Modern alluvial fan and deltaic sedimentation in a foreland tectonic setting: the Lower Mesopotamian plain and the Arabian gulf.- Sedimentary Geology, Elsevier, Amsterdam, t. 67, pp. 175-197, 11 fig.
D.T., S.P., R.G. & P.G. (1995).- Seashells of Eastern Arabia.- Motivate Publishing, Dubai-London, 296 p.
D.S. (1980).- Freshwater snails of Africa and their medical importance.- Taylor & Francis, London, 487 p., 153 fig.
S.P. & F.E. (1966).- New molluscs from the recent Hammar Formation of South east Iraq.- Proceedings of the Malacological Society of London, t. 37, pp. 35-43, pl. 2-4.
F.E. & G.D. (1957).- Six new molluscan species from the alluvium of Lake Hammar near Basrah.- Proceedings of the Malacological Society of London, t. 32, pp. 198-203, 2 pl.
J.-C., F., Y., J.-C., P., P.A. & I. (1985).- Freshwater to marine like environments from Holocene lakes in Northern Sahara.- Nature, London, t. 317 (6038), pp. 608-610.
P. & J.-C. (1982).- Continental Carbonate Sedimentation and pedogenesis – Late Cretaceous and Early Tertiary of Southern France.- Contributions to Sedimentology 12 – Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, 213 p., 59 fig., 49 pl.
F., J.-C., J.-C., P., I., P., I., Y. & P. (1987).- Biological remains, geochemistry and stable isotopes for the reconstruction of environmental and hydrological changes in the Holocene lakes from North Sahara.- Palaeogeography, Palaeoclimatology, Palaeoecology, Elsevier, Amsterdam, t. 60, pp. 1-46, 11 fig., 6 tabl., 2 pl.
L. (1924).- Catalogue of the Planorbidae in the Indian Museum (Natural history), Calcutta.- Records of the Indian Museum, Calcutta, t. 21, 210 p., 4 pl.
F. (1969).- Superfamilia Unionacea.- Das Tierreich, 88, Walter de Gruyter & Co, Berlin, pp. 1-663.
R.G.S., F.E. & G.L. (1957).- The fauna of some Recent marine deposits near Basrah, Iraq.- Geological Magazine, London, t. 94, pp. 393-401.
C.E. & G. (1978).- The Holocene geological history of the Tigris-Euphrates-Karun delta.- In: W.C. (ed.), The environmental history of the Near and Middle East since the Last Ice Age.- Academic Press, New York, pp. 227-244, 3 fig., 11 tabl.
P. (1984).- Compléments à la malacofaune oligocène (Stampien) de Gaas (Bassin d’Aquitaine, France). 1. Mollusques saumâtres.- Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, Rotterdam, t. 22 (3), pp. 125-142, 2 fig., 4 pl.
P. (2005).- The European Tertiary Neritilidae (Mollusca, Gastropoda, Neritopsina): indicators of tropical submarine cave environments and freshwater faunas.- Zoological journal of the Linnean Society, London, t. 140, pp. 447-467, 9 fig.
C. (1972).- Mollusques benthiques du Lac Tchad : écologie, étude des peuplements et estimation des biomasses.- Cahiers de ORSTOM, (Hydrobiologie), Paris, 6, pp. 3-45, 17 fig., 37 tabl.
W.A. & C. (1978).- Mesopotamia: the Tigris Euphrate delta and its Holocene Hammar fauna.- Geological Magazine, London, t. 115, pp. 287-300, 1 fig., 1 tabl.
R.C. (1958).- Recent marine deposits near Basrah.- Geological Magazine, London, t. 95 (1957), pp. 84-85.
A. (1874).- Coquilles terrestres et fluviatiles recueillies par M. le Docteur Alex. en Orient.- Journal de Conchyliologie, Paris, t. 22, pp. 5-60.
F., J.-R. & M.-M. (2000).- La transition Messinien-Pliocène en Méditerranée orientale (Chypre) : la période du Lago-Mare et sa signification.- Comptes Rendus de l’Académie des Sciences de Paris, (Sciences de la Terre), Paris, t. 331, pp. 483-490, 3 fig.
J.-C. (1987).- The brackish and fresh water mollusks of southern Mesopotamia, Iraq, and their ecological significance.- Abstracts, Conference on Quaternary Sediments in the Arabian Gulf and Mesopotamian region, Kuwait, pp. 44-45.
J.-C. (1989).- Signification écologique et paléogéographique des peuplements oligotypiques de Potamides (Gastéropodes thalassiques).- Atti 3° Symposio di Ecologia e Paleoecologia delle comunita bentoniche, Università di Catania, Taormina 1985, pp. 25-52, 1 tabl., 11 fig.
J.-C. (1991).- Paleogeographic significance of the Cardium, Potamids and Foraminifera living in intra-continental salt lakes of North Africa (Sahara Quaternary, Egypt Present Lakes).- Journal of African Earth Sciences, Paris, t. 12, pp. 383-389, 3 fig.
J.-C. (1993).- Modern and fossil Potamids (Gastropoda) in saline lakes.- Journal of Paleolimnology, Dordrecht, t. 8, pp. 163-169, 2 fig.
J.-C. (1995).- Modern and fossil mangroves and mangals: their climatic and biogeographic variability.- In: D.W.J. & P.A. (eds.), Marine Paleoenvironmental Analysis from Fossils.- Geological Socociety of London, Special Publication, 83, pp. 73-96.
J.-C., F., A., O., P., F., A. & J.-L. (1998).- Quaternary marine and continental sedimentation in the northern Red Sea and Gulf of Suez (Egyptian coast): influences of rift tectonics, climatic changes and sea level fluctuations.- In: B.H. & D.W.J. (eds.), Sedimentation and Tectonics of Rift Basins : Red Sea – Gulf of Aden.- Chapman & Hall, London, pp. 537-573.
J.-C. & P. (1991).- Données récentes sur la sédimentation tardive dans la plaine de Larsa-'Oueili.- In: J.-L. (ed.), 'Oueili. Travaux de 1985.- Éditions recherche sur les civilisations, Paris, Mémoire 89, pp. 341-343, 1 fig.
H.W. & P.R. (1988).- Eustatic controls on clastic deposition. II – Sequence and systems tract models. In: et alii (eds.), Sea-level changes – an interpreted approach.- Special Publication - Society of Economic Paleontologists and Mineralogists, Tulsa, 42, pp. 125-154.
B. (1921).- Report on the freshwater Gastropod Molluscs of Lower Mesopotamia. III- The families Neritidae, Hydrobiidae and Melaniidae.- Records of the Indian Museum, Calcutta, t. 18, pp. 215-227, 3 fig.
B.H., M., N.H., F., K.M., F., J.-C., S.Y. & W.R. (1982).- Caractères et évolution du complexe deltaïque Tigre-Euphrate.- Mémoire de la Société Géologique de France, (n.s.), 144, pp. 207-216, 5 fig.
J.C. (1983).- Paléofaunes. Mollusques.- In: N. & P. (eds.), Sahara ou Sahel ?.- Lamy, Marseille, pp. 157-171.
J., ed. (1980).- Mesopotamian ecology and destiny.- Monographiae Biologicae, Dr. Junk, The Hague, t. 38, 122 p., 36 fig.
R. (1961).- Contribution à la malacologie de la Mésopotamie (Irak).- Cahiers des Naturalistes, Paris, (n.s.), t. 17, pp. 65-68.
E. (1975).- The molluscs of the Sea of Galilee.- Malacologia, Philadelphia, t. 15, 147-184, 43 fig., 3 tabl.
D. (1984).- The freshwater mollusca of Northern Africa. Distribution, biogeography and palaeoecology.- Developments in Hydrobiology, 25, Dr Junk publ., The Hague, 164 p., 144 fig., 5 tabl.
![]()
Click on thumbnail to enlarge the image.
Figure 1: The alluvial plain of Lower Mesopotamia. Location of the principal localities cited. Unpublished dates (14C – Lab. of Hydrology, Orsay) of fossils from the Hammar Fm (dark green).
Click on thumbnail to enlarge the image.
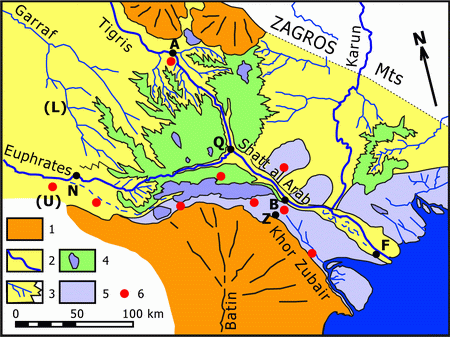
Figure 2: Major morpho-sedimentary units in Lower Mesopotamia (after et alii, 1982, modified). 1- Alluvial fans, 2- Fluviatile channel and levees, 3- Lacustrine delta, 4- Marsh and lake, 5- Sabkha (marine and continental), 6- Hammar Formation fossils (late Quaternary transgression). A: Amarah, B: Basrah, F: Fao, N: Nassiriyah, Q: Qurna, Z: Zubair, (L): Lagash, U: Ur.
Click on thumbnail to enlarge the image.
Figure A: The freshwater marshes, north of Lake Hammar.
1 - The Mesopotamian reed marshes include very large flooded areas covered exclusively by Typha, the reed mace (= bulrush).
2 - The bulrush is the foundation of a cattle breeding culture; seasonal fires
(smoke) are set by the stock–breeding tribes to regenerate fresh green Typha
leaves.
3 - The incipient change in scenery. A network of causeways isolated dried marsh and lake areas (right) from as yet untouched flooded domain. This was a godsend for the working paleoecologist when it was limited in extent
(1980), for it helped the study of molluscan distribution within the polder-like units
(Fig. C
![]() ).
).
4 - Cowsheds of Phragmites reeds in the Typha domain, close to the Phragmites marsh (in the distance). Their platforms are piles of compacted mud taken from the adjacent marsh floor; a peripheral depression (cleared) reveals the water level.
5 - Cattle herd in the Typha. Last year’s cats-tails serve as the origin of the growing green shoots that appeared at the end of
winter.
Click on thumbnail to enlarge the image.
Figure B: The now vanished Lake-and-Reed-Marsh culture of Mesopotamia. Amphibious villages (1-2) were the result of long accomodation to this flooded environment. Each Phragmites house is built on an artificial platform, protected by a wave shield (arrow). The skiff in the foreground is filled with Typha leaves cut for cattle in the sheds. The mud platform is repeatedly added to compensate for natural subsidence, using sediments dredged during the maintenance of the boat-channels. The punt boat (3) holds one-fourth of a cubic meter of silty alluvium (arrow) for that purpose.
Click on thumbnail to enlarge the image.
Figure C: The dried bottom of the Typha marshland and adjacent lake revealed by a polder-like development. This dammed area
(Fig. A.3
![]() ), suddenly cut off from natural flooding, has not been subject to cattle tramping and grazing. The well preserved freshwater malacofauna includes the viviparid
Bellamya (B), Melanopsis (M), Corbicula (C), the unionids Anodonta (A) and Unio (U) and Melanoides (m). There is a minor difference between marsh and lake molluscan assemblages:
Bellamya and Melanopsis are typical of the vegetal substrates of the marsh area and Melanoides is more abundant on lake
floors.
), suddenly cut off from natural flooding, has not been subject to cattle tramping and grazing. The well preserved freshwater malacofauna includes the viviparid
Bellamya (B), Melanopsis (M), Corbicula (C), the unionids Anodonta (A) and Unio (U) and Melanoides (m). There is a minor difference between marsh and lake molluscan assemblages:
Bellamya and Melanopsis are typical of the vegetal substrates of the marsh area and Melanoides is more abundant on lake
floors.
Click on thumbnail to enlarge the image.
Figure D: Contrasting molluscan assemblages of the freshwater subenvironments.
1 - Lacustrine mud bottom with an Anodonta (A) in life position, a bivalved Unio (U), and sparse Melanoides (m) and Corbicula (C).
2 - A high energy shoreline accumulation of floated snails: Lymnaea (L) and Melanoides (m).
3 - The calm Typha marsh bottom includes
Bellamya (B), Melanopsis (M) and Melanoides (m).
4 - Unio tigridis taken from the bottom of a marsh channel with active currents. The buried anterior part of the shell is free of encrusting algae.
![]()
Click on thumbnail to enlarge the image.
Plate 1: Living environment and accumulation of the freshwater and brackish (low salinity) molluscs of Lower Mesopotamia.
Figure 1 - Melanopsis praemorsum crawling on a tidal channel bank of the Shatt al Arab delta. Salinity is below 3‰. The extended snout and tentacles (arrow) show this activity to be subaerial although the species is more animated while submerged, particularly on lake vegetation.
Figure 2 - Corbicula fluminalis in living position (semi-buried) in a lacustrine, mud bottom scattered with Melanopsis and Melanoides. The disassociation of the shells after death may have been caused by rain or by birds. .
Figure 3 – Dried lake floor preserved by a polder enclosure. Corbicula dominated the association that includes only a few (but conspicuous) Unio (U) and smaller Melanoides (m) and Melanopsis (M).
Figure 4 - Underwater photo of the bottom of a marsh channel. The current winnowing reveals accumulation of empty shells and the posterior end of living (buried) Unio.
Figure 5 - Shoreline gravel with displaced shells, Shatt al Arab estuary: floated Bellamya (B) and bottom traction (current, waves) displaced heavier shells (Corbicula - C - and Melanopsis - M). They illustrate a high-energy symmigy (mixture).
Figure 6 - Mud lake floor (polder) with in situ Pseudodontopsis (P - in living position), slightly moved Corbicula valves, a corroded Melanopsis (M) (reworked ?) and a possibly floated Bellamya (B). This assemblage does not depict incontestably the association in life, for it may include contributions from adjacent environments.
Figure 7 - Floated assemblage at a lake shoreline. Apart from marsh derived plants, the dominant Melanoides is associated with Lymnaea (L) and Melanopsis (M). Corrosion and breakage are most evident in Melanoides shells possibly indicating that some of them are redeposited, for the low pH of the sediments hinders fossilization in marsh and lake deposits.
Click on thumbnail to enlarge the image.
Figures 1-5. Unio tigridis . 1 & 3: left valve and umbonal view of a large specimen. 1-2 & 5: marked polymorphism of the shell outline. 4-5: internal and external view of the two valves of a small specimen. Marsh N of the Euphrates river, N of Lake Hammar.
Figures 6-10. Pseudodontopsis euphraticus (). Polymorphism and progressive proportional changes during growth of the shell. 8: umbonal view of the flat bivalve. Lake Hammar.
Figures 11-13. Anodonta (Anodonta) vescoiana . 11-12: Slightly elongated and inflated specimen. 13: internal view of an elongated shell. Lake Hammar.
Scale bars: 1 cm.
Click on thumbnail to enlarge the image.
Figures 1-8. Lymnaea (Radix) gr. auricularia . Highly polymorphic shells from one locality: marsh SW of Qurna.
Figures 9-12. Gyraulus intermixtus (). Marsh SW of Qurna.
Figures 13-18. Bellamya bengalensis (). 14: a young shell with its carinate whorl. 13-17: more or less elongate specimens. 18: aperture of fig. 13, aperture plane placed in the same plane as the photo. Marsh N of Lake Hammar.
Figures 19-29. Corbicula fluminalis (). 19-21: high morphs. 24-29: wide morphs. 22-23: intermediate. See the characteristic striae on lateral teeth of both right and left shells of the same specimen.
Scale bars: 1 cm.
Click on thumbnail to enlarge the image.
Figures 1-6. Cerithidea (Cerithideopsilla) cingulata (). 1-4 & 6: living snails, (1) with an Ostreid shell on its back; (6) axial view of the last whorl. The dorsal half of specimens (3-4 & 6) is eroded by microboring algae; their attack on the lower half is prevented by the ploughing behavior of the snail. Shatt al Arab estuarine mouth, Fao. 5: fossil specimen from the Holocene Hammar Formation. S of Lake Logait.
Figures 7-10. Melanoides tuberculata (). Diverse ornamentations and patterns of whorl development during growth. Lake Hammar.
Figures 11-20. Theodoxus (Neritaea) jordani (). Polymorphism in one locality. Marsh N of Lake Hammar.
Figures 21-29. Neritina (Dostia) schlaeflii . 21-26: polymorphism and diverse color ornamentation. 29: internal view of the operculum. Mouth of the Shatt al Arab estuary, Fao. 27: Holocene fossil, NE of Basrah.
Figures 30-37. Melanopsis (Melanopsis) praemorsum (). 30-33: high degree of polymorphism of an estuarine upper reach population, NE of Basrah. 34-37: lacustrine population of Lake Hammar.
Scale bars: 1 cm.