![]()
![]()
![]()
![]() This paper presents a thorough analysis of foraminiferal assemblages ranging in age from the Bedoulian-Gargasian transition to the middle Gargasian in the Cassis-La Bédoule area (SE France), the historical stratotype of the Lower Aptian substage. This region is particularly suitable for detailed studies of Aptian foraminifera owing
to rapid and continuous sediment depositional rates and well-diversified microfaunas. The ranges of benthic forms appear to be fairly stable but some species
(Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) become extinct at the end of the Bedoulian and thus can be used to separate this substage from the Gargasian. The stratotypic area also offers an opportunity to follow the evolution of planktonic forms step by step at a crucial period of their history, when modalities of speciation and phylogenetic relationships appear to be particularly complex. The important morphologic variability of Aptian planktonic foraminifera does not help finding stable stratigraphic markers; nevertheless, we are able to propose a biozonation comprising five zones (Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) for the interval under consideration, usually subdivided into three zones. Our study of foraminiferal assemblages and species took into account the range of variability among populations and not just the characteristics of a single specimen, such as the holotype. As a result this paper provides new taxonomic precisions on certain planktonic species hitherto controversial or possibly of doubtful validity.
This paper presents a thorough analysis of foraminiferal assemblages ranging in age from the Bedoulian-Gargasian transition to the middle Gargasian in the Cassis-La Bédoule area (SE France), the historical stratotype of the Lower Aptian substage. This region is particularly suitable for detailed studies of Aptian foraminifera owing
to rapid and continuous sediment depositional rates and well-diversified microfaunas. The ranges of benthic forms appear to be fairly stable but some species
(Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) become extinct at the end of the Bedoulian and thus can be used to separate this substage from the Gargasian. The stratotypic area also offers an opportunity to follow the evolution of planktonic forms step by step at a crucial period of their history, when modalities of speciation and phylogenetic relationships appear to be particularly complex. The important morphologic variability of Aptian planktonic foraminifera does not help finding stable stratigraphic markers; nevertheless, we are able to propose a biozonation comprising five zones (Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) for the interval under consideration, usually subdivided into three zones. Our study of foraminiferal assemblages and species took into account the range of variability among populations and not just the characteristics of a single specimen, such as the holotype. As a result this paper provides new taxonomic precisions on certain planktonic species hitherto controversial or possibly of doubtful validity.
![]() Early Cretaceous; Aptian; historical stratotype; Foraminifera; biostratigraphy; taxonomy; evolution.
Early Cretaceous; Aptian; historical stratotype; Foraminifera; biostratigraphy; taxonomy; evolution.
M., G., J.-P. (2005).- The Gargasian (Middle Aptian) strata from Cassis-La Bédoule (Lower Aptian historical stratotype, SE France): planktonic and benthic foraminiferal assemblages and biostratigraphy.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2005/02 (CG2005_A02)
![]() Le
Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de
l'Aptien inférieur, SE France) : associations et biostratigraphie des
Foraminifčres benthiques et planctoniques.-
L'analyse approfondie des associations de Foraminifères depuis les termes du passage Bédoulien-Gargasien jusqu'au Gargasien moyen a été réalisée dans le secteur de Cassis-La Bédoule (SE France), stratotype historique du Bédoulien (Aptien inférieur), particulièrement favorable à ce niveau en raison de la continuité et de la dilatation de la série ainsi que de la diversification de la microfaune. L'éventail des formes benthiques se révèle assez stable mais quelques espèces (Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) s'éteignent à la fin du Bédoulien et peuvent être utilisées pour différencier ce sous-étage par rapport au Gargasien. L'aire stratotypique offre aussi la possibilité de suivre pas à pas l'évolution des formes planctoniques à un moment crucial de leur histoire, lorsque les processus de spéciation et les liens phylogénétiques s'avèrent particulièrement complexes. La grande variabilité morphologique des formes planctoniques durant l'Aptien ne facilite pas le choix de marqueurs stratigraphiques
stables; il est cependant possible de proposer une biozonation comportant cinq zones (à Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) pour l'intervalle considéré, habituellement subdivisé en trois zones. L'étude populationnelle, et non pas seulement typologique, des associations nous a enfin permis d'apporter quelques précisions taxinomiques nouvelles, en particulier sur certaines formes planctoniques d'acception controversée ou même de validité douteuse.
Le
Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de
l'Aptien inférieur, SE France) : associations et biostratigraphie des
Foraminifčres benthiques et planctoniques.-
L'analyse approfondie des associations de Foraminifères depuis les termes du passage Bédoulien-Gargasien jusqu'au Gargasien moyen a été réalisée dans le secteur de Cassis-La Bédoule (SE France), stratotype historique du Bédoulien (Aptien inférieur), particulièrement favorable à ce niveau en raison de la continuité et de la dilatation de la série ainsi que de la diversification de la microfaune. L'éventail des formes benthiques se révèle assez stable mais quelques espèces (Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) s'éteignent à la fin du Bédoulien et peuvent être utilisées pour différencier ce sous-étage par rapport au Gargasien. L'aire stratotypique offre aussi la possibilité de suivre pas à pas l'évolution des formes planctoniques à un moment crucial de leur histoire, lorsque les processus de spéciation et les liens phylogénétiques s'avèrent particulièrement complexes. La grande variabilité morphologique des formes planctoniques durant l'Aptien ne facilite pas le choix de marqueurs stratigraphiques
stables; il est cependant possible de proposer une biozonation comportant cinq zones (à Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) pour l'intervalle considéré, habituellement subdivisé en trois zones. L'étude populationnelle, et non pas seulement typologique, des associations nous a enfin permis d'apporter quelques précisions taxinomiques nouvelles, en particulier sur certaines formes planctoniques d'acception controversée ou même de validité douteuse.
![]() Crétacé inférieur ; Aptien ; stratotype historique ; Foraminifères ; biostratigraphie ; taxinomie ; évolution.
Crétacé inférieur ; Aptien ; stratotype historique ; Foraminifères ; biostratigraphie ; taxinomie ; évolution.
![]()
![]() The
geographical location and the stratigraphic relationships of the three sections (La Bédoule-Les
Tocchis; Cassis-carrière Comte; Cassis-carrière de La Marcouline)
included in this micropaleontological study have already been presented in the
two introductory chapters (
& ,
2004; et alii,
2004) of the Memoir in Carnets de Géologie
devoted to the type-Gargasian. In these two papers the reader will also find
information on the lithology and relative
positions of samples in
the La Marcouline section as well as on the circumstances that
justify the re-study of what remains of the Aptian type-sections, employing a
pluridisciplinary approach that uses the methods of integrated stratigraphy.
The
geographical location and the stratigraphic relationships of the three sections (La Bédoule-Les
Tocchis; Cassis-carrière Comte; Cassis-carrière de La Marcouline)
included in this micropaleontological study have already been presented in the
two introductory chapters (
& ,
2004; et alii,
2004) of the Memoir in Carnets de Géologie
devoted to the type-Gargasian. In these two papers the reader will also find
information on the lithology and relative
positions of samples in
the La Marcouline section as well as on the circumstances that
justify the re-study of what remains of the Aptian type-sections, employing a
pluridisciplinary approach that uses the methods of integrated stratigraphy.
![]() The
main purpose of this note is to perfect our knowledge of the distribution of
benthic and planktonic foraminifera during the Bedoulian-Gargasian transition,
based on a detailed analysis of selected sections in the stratotypic area of Cassis-La Bédoule.
In comparison with the Vocontian domain and Italy this region is regarded as
particularly favorable for a study of these levels, because it:
The
main purpose of this note is to perfect our knowledge of the distribution of
benthic and planktonic foraminifera during the Bedoulian-Gargasian transition,
based on a detailed analysis of selected sections in the stratotypic area of Cassis-La Bédoule.
In comparison with the Vocontian domain and Italy this region is regarded as
particularly favorable for a study of these levels, because it:
• has high sedimentation rates and lacks hiatuses, both of which permit increased precision in any investigation regarding the evolution of taxa,
• is rich in macrofossils (particularly in ammonites, cf. , 1994; et alii, 1998), making biostratigraphic correlations easier.
![]() Previous
works concerning Aptian Foraminifera from the western part of Basse-Provence,
and in particular from that of the Bedoulian stratotypic area, are few and most
are already somewhat out-of-date. In 1961,
described several new species of which the type-material, for some at least,
came from this area. et alii
(1965) provided the first extensive analysis of the
distribution of the Bedoulian stratotype microfauna (foraminifers, ostracods),
and correlated it with the distribution of ammonites. In a comparative study of
Aptian planktonic foraminifera from Mexico and SE France,
(1974) included three samples taken from the upper
Bedoulian of the Cassis section. In her thesis devoted to the upper Aptian
stratigraphy and sedimentology of Provence,
(1975) included, for the purpose of biostratigraphic
correlation, data on the micropaleontological content of samples taken from some
sections in the Cassis area studied herein. In his memoir on the Cretaceous
foraminifera of Provence,
(1981) referred frequently to individuals of species
collected from the Cassis-La Bédoule stratotype giving precisions on their
paleontological acceptance and their stratigraphic distribution.
(1987) visited the Bedoulian stratotype using its
well-defined terms to locate more precisely the Bedoulian-Gargasian boundary in
the Subalpine Basin. Then using the ranges of ammonites and foraminifera the
author proposed broader correlations between Basse-Provence and the Vocontian
(Subalpine) basin.
Previous
works concerning Aptian Foraminifera from the western part of Basse-Provence,
and in particular from that of the Bedoulian stratotypic area, are few and most
are already somewhat out-of-date. In 1961,
described several new species of which the type-material, for some at least,
came from this area. et alii
(1965) provided the first extensive analysis of the
distribution of the Bedoulian stratotype microfauna (foraminifers, ostracods),
and correlated it with the distribution of ammonites. In a comparative study of
Aptian planktonic foraminifera from Mexico and SE France,
(1974) included three samples taken from the upper
Bedoulian of the Cassis section. In her thesis devoted to the upper Aptian
stratigraphy and sedimentology of Provence,
(1975) included, for the purpose of biostratigraphic
correlation, data on the micropaleontological content of samples taken from some
sections in the Cassis area studied herein. In his memoir on the Cretaceous
foraminifera of Provence,
(1981) referred frequently to individuals of species
collected from the Cassis-La Bédoule stratotype giving precisions on their
paleontological acceptance and their stratigraphic distribution.
(1987) visited the Bedoulian stratotype using its
well-defined terms to locate more precisely the Bedoulian-Gargasian boundary in
the Subalpine Basin. Then using the ranges of ammonites and foraminifera the
author proposed broader correlations between Basse-Provence and the Vocontian
(Subalpine) basin.
![]() The
most recent study ( et alii,
1998) on Aptian foraminifera from this region deals
mainly with Bedoulian forms. Here, we shall use this work as a starting point
for our paleontological and biostratigraphic study of Gargasian levels. In
addition we shall use the data provided by the latest compilations on
foraminifera (mostly planktonic) of the Tethyan Aptian ( et alii,
2002; & ,
2002, 2003a, 2003b).
The
most recent study ( et alii,
1998) on Aptian foraminifera from this region deals
mainly with Bedoulian forms. Here, we shall use this work as a starting point
for our paleontological and biostratigraphic study of Gargasian levels. In
addition we shall use the data provided by the latest compilations on
foraminifera (mostly planktonic) of the Tethyan Aptian ( et alii,
2002; & ,
2002, 2003a, 2003b).
![]() The
samples upon which the present study is based come from several localities.
Because of the progressive degradation of the outcrops in recent years, in
1997-2002 we could sample only the uppermost Bedoulian beds of the Comte Quarry
section and the lower (but not lowermost) and middle Gargasian beds of the La
Marcouline Quarry section; the latter, as is the case everywhere in the area,
lacks the higher terms of the Gargasian owing to pre-Cenomanian erosion. Both
quarries are very near each other and located within the limits of the town of
Cassis.
The
samples upon which the present study is based come from several localities.
Because of the progressive degradation of the outcrops in recent years, in
1997-2002 we could sample only the uppermost Bedoulian beds of the Comte Quarry
section and the lower (but not lowermost) and middle Gargasian beds of the La
Marcouline Quarry section; the latter, as is the case everywhere in the area,
lacks the higher terms of the Gargasian owing to pre-Cenomanian erosion. Both
quarries are very near each other and located within the limits of the town of
Cassis.
![]() In
addition, we had access to washed residues from and fractional samples of
material collected from 1962 to 1966 by one of us (MM) in the Les Tocchis
section, located in the village of La Bédoule (today Roquefort-La Bédoule),
2 km ENE of the La Marcouline section. This section has now
disappeared because of urbanization but as late as the latter sixties exposed in
a quasi-continuous way the uppermost Bedoulian to lowermost Gargasian
transitional beds, thereby spanning the gap in the continuity of observation of
the sequence between the two almost contiguous quarries at Cassis.
Unfortunately, nowadays this dominantly marly transitional level cannot be seen
anywhere in Basse-Provence.
In
addition, we had access to washed residues from and fractional samples of
material collected from 1962 to 1966 by one of us (MM) in the Les Tocchis
section, located in the village of La Bédoule (today Roquefort-La Bédoule),
2 km ENE of the La Marcouline section. This section has now
disappeared because of urbanization but as late as the latter sixties exposed in
a quasi-continuous way the uppermost Bedoulian to lowermost Gargasian
transitional beds, thereby spanning the gap in the continuity of observation of
the sequence between the two almost contiguous quarries at Cassis.
Unfortunately, nowadays this dominantly marly transitional level cannot be seen
anywhere in Basse-Provence.
![]() The
Bedoulian and Gargasian marly limestones of Provence are relatively indurated
and must be washed with chemicals added. These products not only eliminate the
major part of the biogenic components, thus concentrating the microfauna, but
also clean the microfossils better than water alone, for without the chemicals
the specimens often remain more or less encrusted by an argillocalcareous matrix
that makes their identification difficult.
The
Bedoulian and Gargasian marly limestones of Provence are relatively indurated
and must be washed with chemicals added. These products not only eliminate the
major part of the biogenic components, thus concentrating the microfauna, but
also clean the microfossils better than water alone, for without the chemicals
the specimens often remain more or less encrusted by an argillocalcareous matrix
that makes their identification difficult.
![]() The
samples collected in the sixties were prepared for examination using the
(1962), method. This involves a soaking in sodium
thiosulphate followed by the addition of hydrogen peroxide. The washed residue
is then processed in Bradosol (cationic moistening; see details in et alii,
1965; , 1966).
Unfortunately, these samples were sieved through an 80 µm mesh screen,
thought to be sufficient at that time. Our current knowledge about the size of
the smallest planktonic foraminifera requires that sediments be sieved through a
finer mesh of at least 63 µm, or even of 45 µm.
The
samples collected in the sixties were prepared for examination using the
(1962), method. This involves a soaking in sodium
thiosulphate followed by the addition of hydrogen peroxide. The washed residue
is then processed in Bradosol (cationic moistening; see details in et alii,
1965; , 1966).
Unfortunately, these samples were sieved through an 80 µm mesh screen,
thought to be sufficient at that time. Our current knowledge about the size of
the smallest planktonic foraminifera requires that sediments be sieved through a
finer mesh of at least 63 µm, or even of 45 µm.
![]() After
having been crushed and oven-dried, the
samples collected at the turn of the century were soaked for a minimum of several days in Rewoquat W 3690 (cationic
tenside), then sieved at 45 µm using warm water and rinsed with hydrogen
peroxide. This procedure is time consuming but gives excellent results, for both
concentrating and cleaning microfaunas.
After
having been crushed and oven-dried, the
samples collected at the turn of the century were soaked for a minimum of several days in Rewoquat W 3690 (cationic
tenside), then sieved at 45 µm using warm water and rinsed with hydrogen
peroxide. This procedure is time consuming but gives excellent results, for both
concentrating and cleaning microfaunas.
![]() The
state of preservation of the microfossils varies with its location in the
sequence: from moderately well to rather good (e.g. the more marly middle
Gargasian levels) to mediocre or even poor in the more indurated uppermost
levels of the Bedoulian. These levels are not necessarily more calcareous but
are certainly more siliceous, as shown by the increased occurrence of sponge
spicules, radiolarians and diatoms in the corresponding washed residues. We also
noted the rather constant presence in the residues of a significant fraction of
more or less distorted tests, particularly among the thin walled planktonic
foraminifera (e.g. small few-chambered Globigerinelloides [ex-Blowiella],
small Praehedbergella) but also some agglutinated benthics among them
representatives of the Trochamminidae, Reophax and Recurvoides.
These deformations are probably the result of rather strong compaction.
The
state of preservation of the microfossils varies with its location in the
sequence: from moderately well to rather good (e.g. the more marly middle
Gargasian levels) to mediocre or even poor in the more indurated uppermost
levels of the Bedoulian. These levels are not necessarily more calcareous but
are certainly more siliceous, as shown by the increased occurrence of sponge
spicules, radiolarians and diatoms in the corresponding washed residues. We also
noted the rather constant presence in the residues of a significant fraction of
more or less distorted tests, particularly among the thin walled planktonic
foraminifera (e.g. small few-chambered Globigerinelloides [ex-Blowiella],
small Praehedbergella) but also some agglutinated benthics among them
representatives of the Trochamminidae, Reophax and Recurvoides.
These deformations are probably the result of rather strong compaction.
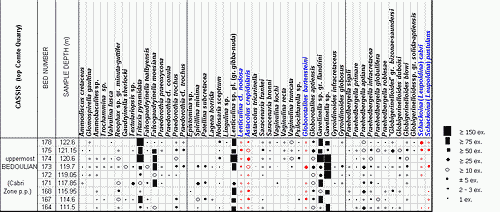
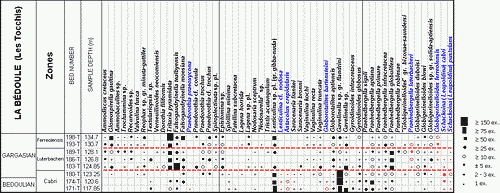
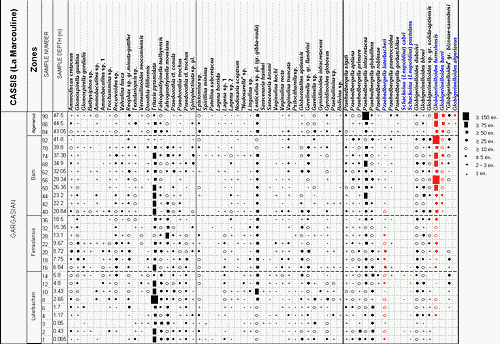
![]() The
foraminiferal content of all the samples (9 from the uppermost levels of the
Comte Quarry section, 8 from the Les Tocchis section and 24 from the La
Marcouline Quarry section) was examined quantitatively; that is, the number of
specimens of each species of foraminifera was counted. This led to the
establishment of frequency classes as follows:
The
foraminiferal content of all the samples (9 from the uppermost levels of the
Comte Quarry section, 8 from the Les Tocchis section and 24 from the La
Marcouline Quarry section) was examined quantitatively; that is, the number of
specimens of each species of foraminifera was counted. This led to the
establishment of frequency classes as follows:
| Category | RR | R | R-F | F | F-C | C | AB | V.AB |
| Specimens numb. | 1 | 2-3 | ±5 | >10 | >20 | >50 | >75 | >150 |
![]() Abundances
are indicated by symbols for each species and each sample in the distribution
tables of Figs. 1
Abundances
are indicated by symbols for each species and each sample in the distribution
tables of Figs. 1 ![]() - 2
- 2 ![]() - 3
- 3
![]() .
.
![]() The
material (remaining fractional sample, washed residues, slides) upon which this
study is based are in the collections of the Paleontology Museum of the
University of Provence in Marseilles.
The
material (remaining fractional sample, washed residues, slides) upon which this
study is based are in the collections of the Paleontology Museum of the
University of Provence in Marseilles.
• Upper Bedoulian: Tritaxia pyramidata, Falsogaudryinellas, "smooth" (i.e. not ornamented: L. gr. gibba-nuda) Lenticulinas, Gavelinellas.
• Gargasian: Tritaxia pyramidata remains (more or less consistently) a dominant species, Falsogaudryinellas and Lenticulinas are slightly less numerous and Gavelinellas much less common than in the Bedoulian; on the other hand planktonic forms are more abundant.
![]() The
number of species increases upwards in the series. Taken as a whole the
type-Bedoulian includes 31 benthic species (14 agglutinated and 17 calcareous)
and 11 planktonic species, i.e. a total of 42 species of foraminifera.
The (lower-middle) Gargasian from the same area provided 45 benthic species (20
agglutinated and 25 calcareous), plus 21 planktonic species, i.e. 66
species; these data are summarized in the table below:
The
number of species increases upwards in the series. Taken as a whole the
type-Bedoulian includes 31 benthic species (14 agglutinated and 17 calcareous)
and 11 planktonic species, i.e. a total of 42 species of foraminifera.
The (lower-middle) Gargasian from the same area provided 45 benthic species (20
agglutinated and 25 calcareous), plus 21 planktonic species, i.e. 66
species; these data are summarized in the table below:
| Species number | Agglut. | Calc. | Plankt. | Total |
| GARGASIAN | 20 | 25 | 21 | 66 |
| BEDOULIAN | 14 | 17 | 11 | 42 |
![]() The
longevity of the great majority of species, both benthics and curiously enough,
planktonics, is rather lengthy. Numerous species have ranges that include a
major part of the Aptian stage. The increasing diversity in the foraminiferal
fauna of the upper Aptian was caused by both a rather low rate of extinction and
by a progressive enrichment in new species.
The
longevity of the great majority of species, both benthics and curiously enough,
planktonics, is rather lengthy. Numerous species have ranges that include a
major part of the Aptian stage. The increasing diversity in the foraminiferal
fauna of the upper Aptian was caused by both a rather low rate of extinction and
by a progressive enrichment in new species.
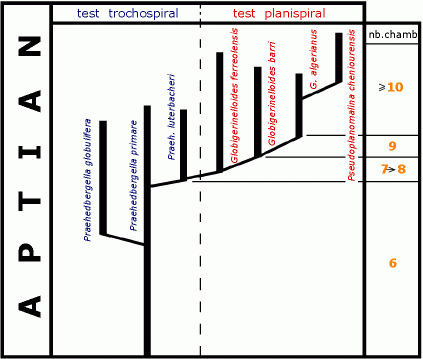
![]() Our
data show that in the Aptian strata of Cassis-La Bédoule the number of
potential markers of both benthic and planktonic forms is rather low (Fig. 4
Our
data show that in the Aptian strata of Cassis-La Bédoule the number of
potential markers of both benthic and planktonic forms is rather low (Fig. 4 ![]() ).
).
![]() In
this category are a few species that are commonly encountered in the Tethys well
before Aptian times, but become extinct at the end of the Bedoulian or range
only slightly past the Bedoulian-Gargasian boundary:
In
this category are a few species that are commonly encountered in the Tethys well
before Aptian times, but become extinct at the end of the Bedoulian or range
only slightly past the Bedoulian-Gargasian boundary:
• Praedorothia praeoxycona
• Lenticulina cf. nodosa
• Astacolus crepidularis
• Globorotalites bartensteini
![]() Benthic
forms that first appear during the Gargasian appear to have little
stratigraphical significance. They are representatives of agglutinated genera
like Bathysiphon, Ammobaculites, Trochammina, Recurvoides,
and the agglutinated species Praedorothia cf. conula. Their
presence at this level may well be related to a change in environment from that
of underlying strata.
Benthic
forms that first appear during the Gargasian appear to have little
stratigraphical significance. They are representatives of agglutinated genera
like Bathysiphon, Ammobaculites, Trochammina, Recurvoides,
and the agglutinated species Praedorothia cf. conula. Their
presence at this level may well be related to a change in environment from that
of underlying strata.
![]() No
planktonic species present in the Bedoulian became extinct during that substage. Schackoina gr. cabri (= S. cabri s.s. + S. pustulans),
which first appeared in the late Bedoulian, is the only species with a very
short range, for it disappeared in the earliest Gargasian. A short distance
above the Bedoulian-Gargasian boundary two new forms appear almost
simultaneously: first Praehedbergella luterbacheri, then Globigerinelloides
ferreolensis. Both taxa are sporadically represented in the lowermost
portion of their range where they coexist with the last Schackoinas, which are
also irregular in occurrence in this level of overlap. From the lower limit of
its acme upward G. ferreolensis becomes an important component of the
middle / upper Gargasian planktonic microfauna.
No
planktonic species present in the Bedoulian became extinct during that substage. Schackoina gr. cabri (= S. cabri s.s. + S. pustulans),
which first appeared in the late Bedoulian, is the only species with a very
short range, for it disappeared in the earliest Gargasian. A short distance
above the Bedoulian-Gargasian boundary two new forms appear almost
simultaneously: first Praehedbergella luterbacheri, then Globigerinelloides
ferreolensis. Both taxa are sporadically represented in the lowermost
portion of their range where they coexist with the last Schackoinas, which are
also irregular in occurrence in this level of overlap. From the lower limit of
its acme upward G. ferreolensis becomes an important component of the
middle / upper Gargasian planktonic microfauna.
![]() Two
other planktonic species, G. barri and G. algerianus, mark the
remainder of the Gargasian section at La Marcouline where they appear
successively, their first occurrences being separated by about 20 meters in a 50
meter section. These species are very useful markers for both have a broad
geographic distribution and their first occurrences have been found to be
consistently reliable in the Tethyan realm. Some other planktonic species also
appear in these levels: Praehedbergella praetrocoidea and then Praehedbergella
gorbatchikae, but they are more sporadic and therefore of lesser value as
markers.
Two
other planktonic species, G. barri and G. algerianus, mark the
remainder of the Gargasian section at La Marcouline where they appear
successively, their first occurrences being separated by about 20 meters in a 50
meter section. These species are very useful markers for both have a broad
geographic distribution and their first occurrences have been found to be
consistently reliable in the Tethyan realm. Some other planktonic species also
appear in these levels: Praehedbergella praetrocoidea and then Praehedbergella
gorbatchikae, but they are more sporadic and therefore of lesser value as
markers.
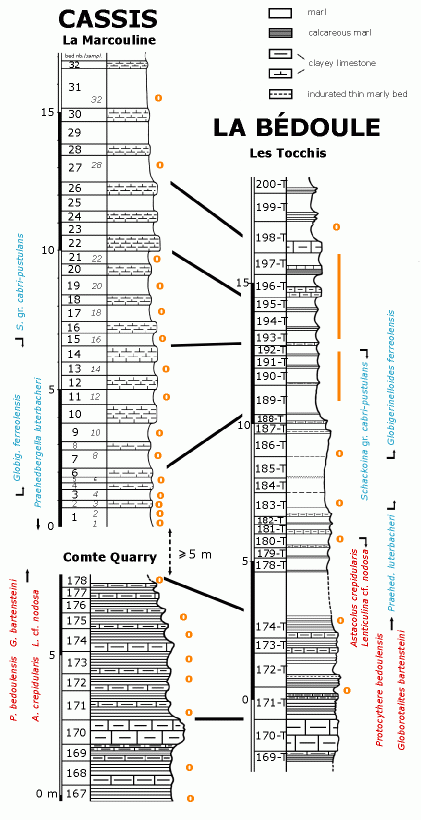
![]() The
detailed inventory of the micropaleontological content of the three sections
mentioned above led first to a refinement in their correlation, which was
initially based on geometrical considerations (Fig. 5
The
detailed inventory of the micropaleontological content of the three sections
mentioned above led first to a refinement in their correlation, which was
initially based on geometrical considerations (Fig. 5 ![]() ).
).
![]() The very short distance (2 km)
between the two areas (Les Tocchis and Comte-La Marcouline)
allows us to eliminate the possibility of any diachronism between the first
occurrences of planktonic forms, such as that recently observed ( et alii,
2004) in the
Cenomanian-Turonian of the Western Interior where
the separation was much greater (600 km). For example the
homology between the "Triplet" of beds 22-24-26 at La Marcouline
(Cassis) and beds 196-T, 197-T, 198-T at Les Tocchis (La Bédoule),
suggested by et alii (2004),
is strongly substantiated. On the basis of the last occurrence of the
Schackoinas and the first occurrence of Globigerinelloides ferreolensis
it is also possible to suggest a correlation between level 14 (Marcouline) and
level 193-T (Tocchis) and between level 6 (Marcouline) and 188-T (Tocchis).
These correlations show that the units of the La Marcouline sequence are
constantly thicker than those of Les Tocchis. As regards the underlying levels,
with the exception of the unquestionable geometric homology between bed 170
(Cassis-Comte Quarry) and bed 170-T (La Bédoule-Les Tocchis), a detailed
correlation is more ticklish; in particular because of the gap mentioned above
in the Cassis section that is not measurable directly and was represented at La Bédoule
by a marly sequence between beds 178-T and 186-T. Thus several datums (the last
occurrences of Astacolus crepidularis and Lenticulina cf. nodosa,
and the first occurrence of Praehedbergella luterbacheri), which are
well-determined at Les Tocchis cannot be precisely located at Cassis for they
are very likely in the zone of no outcrop. However, using the Les Tocchis
section as a guide, the order of magnitude of the Cassis gap can be estimated as
at least 5 meters, even somewhat more if the greater thickness of the series
mentioned above is taken into account. As a result one cannot assume a positive
bed to bed homology between the two sections above level 170; this necessitates
that the numbering of the beds at Les Tocchis be differentiated by the addition
of the suffix "-T".
The very short distance (2 km)
between the two areas (Les Tocchis and Comte-La Marcouline)
allows us to eliminate the possibility of any diachronism between the first
occurrences of planktonic forms, such as that recently observed ( et alii,
2004) in the
Cenomanian-Turonian of the Western Interior where
the separation was much greater (600 km). For example the
homology between the "Triplet" of beds 22-24-26 at La Marcouline
(Cassis) and beds 196-T, 197-T, 198-T at Les Tocchis (La Bédoule),
suggested by et alii (2004),
is strongly substantiated. On the basis of the last occurrence of the
Schackoinas and the first occurrence of Globigerinelloides ferreolensis
it is also possible to suggest a correlation between level 14 (Marcouline) and
level 193-T (Tocchis) and between level 6 (Marcouline) and 188-T (Tocchis).
These correlations show that the units of the La Marcouline sequence are
constantly thicker than those of Les Tocchis. As regards the underlying levels,
with the exception of the unquestionable geometric homology between bed 170
(Cassis-Comte Quarry) and bed 170-T (La Bédoule-Les Tocchis), a detailed
correlation is more ticklish; in particular because of the gap mentioned above
in the Cassis section that is not measurable directly and was represented at La Bédoule
by a marly sequence between beds 178-T and 186-T. Thus several datums (the last
occurrences of Astacolus crepidularis and Lenticulina cf. nodosa,
and the first occurrence of Praehedbergella luterbacheri), which are
well-determined at Les Tocchis cannot be precisely located at Cassis for they
are very likely in the zone of no outcrop. However, using the Les Tocchis
section as a guide, the order of magnitude of the Cassis gap can be estimated as
at least 5 meters, even somewhat more if the greater thickness of the series
mentioned above is taken into account. As a result one cannot assume a positive
bed to bed homology between the two sections above level 170; this necessitates
that the numbering of the beds at Les Tocchis be differentiated by the addition
of the suffix "-T".
![]() The
more detailed data obtained through our analysis of the stratigraphic
distribution of the principal planktonic markers in the Gargasian portion of the
Bedoulian stratotypic area allow us to refine to a somewhat greater degree the
zonation published in et alii
(1998) and, in a more general way that of the
lower-middle Gargasian of the Tethyan region (
& ,
1995; et alii,
2002). We
propose a subdivision of the studied interval into
five rather than three
zones:
The
more detailed data obtained through our analysis of the stratigraphic
distribution of the principal planktonic markers in the Gargasian portion of the
Bedoulian stratotypic area allow us to refine to a somewhat greater degree the
zonation published in et alii
(1998) and, in a more general way that of the
lower-middle Gargasian of the Tethyan region (
& ,
1995; et alii,
2002). We
propose a subdivision of the studied interval into
five rather than three
zones:
|
This work |
|
| Algerianus | Algerianus |
| Ferreolensis | Barri |
| Ferreolensis | |
| Cabri | Luterbacheri |
| Cabri |
![]() In
the previously proposed zonation it was difficult to define with precision the
boundary between the Cabri and Ferreolensis zones. Our findings which confirm
those obtained by
(1974) in Mexico and
&
(1988) in England, show that instead of being mutually
exclusive, as has often been claimed, over a short interval both index species
occur together. Such a concomitance exists over at least 5 meters at La
Marcouline, slightly less at Les Tocchis. In this same interval Schackoina
gr. cabri becomes very rare, and the first Globigerinelloides
ferreolensis are still sparse. In addition, it is not always easy to delimit
precisely the first occurrence of G. ferreolensis. As
&
(1988) foresaw, our observations show that this species
arose from an initially trochospiral form through flattening and progressive
symmetrization of the trochospire. Thus, the ancestor of G. ferreolensis
is represented by Praehedbergella luterbacheri; the process of
symmetrization, gradual but transient, can be detected only on well-preserved
material with well-cleaned ombilici, and involves only a small number of
specimens in a population. Often the degree of symmetrization remains undetected
for its recognition depends on the conditions of fossilization and the methods
of extraction. Globigerinelloides ferreolensis becomes typical and thus
more readily identifiable and more abundant only at the level where Schackoina
gr. cabri becomes extinct. For this reason we choose to use the beginning
of the acme rather than the FAD (First appearance datum) of Globigerinelloides
ferreolensis as the lower limit of the eponymous zone.
In
the previously proposed zonation it was difficult to define with precision the
boundary between the Cabri and Ferreolensis zones. Our findings which confirm
those obtained by
(1974) in Mexico and
&
(1988) in England, show that instead of being mutually
exclusive, as has often been claimed, over a short interval both index species
occur together. Such a concomitance exists over at least 5 meters at La
Marcouline, slightly less at Les Tocchis. In this same interval Schackoina
gr. cabri becomes very rare, and the first Globigerinelloides
ferreolensis are still sparse. In addition, it is not always easy to delimit
precisely the first occurrence of G. ferreolensis. As
&
(1988) foresaw, our observations show that this species
arose from an initially trochospiral form through flattening and progressive
symmetrization of the trochospire. Thus, the ancestor of G. ferreolensis
is represented by Praehedbergella luterbacheri; the process of
symmetrization, gradual but transient, can be detected only on well-preserved
material with well-cleaned ombilici, and involves only a small number of
specimens in a population. Often the degree of symmetrization remains undetected
for its recognition depends on the conditions of fossilization and the methods
of extraction. Globigerinelloides ferreolensis becomes typical and thus
more readily identifiable and more abundant only at the level where Schackoina
gr. cabri becomes extinct. For this reason we choose to use the beginning
of the acme rather than the FAD (First appearance datum) of Globigerinelloides
ferreolensis as the lower limit of the eponymous zone.
![]() We
propose to use the FAD of the ancestor of G. ferreolensis to mark the
base of a new Luterbacheri zone, which thus includes the interval representing
the highest portion of the old Cabri zone i.e the interval of time between the
FAD of the species index and the beginning of the acme in the occurrence of G.
ferreolensis.
We
propose to use the FAD of the ancestor of G. ferreolensis to mark the
base of a new Luterbacheri zone, which thus includes the interval representing
the highest portion of the old Cabri zone i.e the interval of time between the
FAD of the species index and the beginning of the acme in the occurrence of G.
ferreolensis.
![]() In
the same way the upper part of the old Ferreolensis zone can be set up as a
Barri zone, its lower boundary defined par the FAD of G. barri, the
phylogenetic successor of G. ferreolensis. The concept of a Barri zone,
although it was based upon a slightly different definition of the index species,
has already been proposed by (1959).
In
the same way the upper part of the old Ferreolensis zone can be set up as a
Barri zone, its lower boundary defined par the FAD of G. barri, the
phylogenetic successor of G. ferreolensis. The concept of a Barri zone,
although it was based upon a slightly different definition of the index species,
has already been proposed by (1959).
![]() The
first representatives of the phylogenetic successor of G. barri, i.e.
G. algerianus, appear in the last few meters of the La Marcouline
section. Its FAD defines the lower limit of the zone it names, a zone that for
many years has been widely used in the Tethyan domain.
The
first representatives of the phylogenetic successor of G. barri, i.e.
G. algerianus, appear in the last few meters of the La Marcouline
section. Its FAD defines the lower limit of the zone it names, a zone that for
many years has been widely used in the Tethyan domain.
![]() The
zones proposed above in some detail are summarized in
Fig. 4
The
zones proposed above in some detail are summarized in
Fig. 4 ![]() which also indicates the stratigraphic range of the main markers.
which also indicates the stratigraphic range of the main markers.
![]() Taken
as a whole the Aptian appears as the first significant period of radiation among
planktonic foraminifera ( & ,
1983 ; et alii,
1984 ; et alii,
2002). Combined with the published data on the
type-Bedoulian ( et alii,
1998), the results presented here concerning the
Gargasian permit an addition to the particulars involved in the modalities of
this process. Owing to the great detail obtainable because of the rapid rate of
sedimentation in the stratotypic sections, it is possible to see that speciation
during the Aptian, which actually had begun during the late Bedoulian, just before
the anoxic event AOE1a (cf. et alii,
1998) was not a sudden explosion of new forms but a
rather gradual increase in the number of species. The Bedoulian-Gargasian
transitional beds correspond to a brief period of inactivity in planktonic
speciation. This makes it difficult to subdivide them and to delimit the
boundary between the two substages by means of planktonic foraminifera. It
appears easier to approximate this boundary through the use of benthic
foraminifera (Fig. 4
Taken
as a whole the Aptian appears as the first significant period of radiation among
planktonic foraminifera ( & ,
1983 ; et alii,
1984 ; et alii,
2002). Combined with the published data on the
type-Bedoulian ( et alii,
1998), the results presented here concerning the
Gargasian permit an addition to the particulars involved in the modalities of
this process. Owing to the great detail obtainable because of the rapid rate of
sedimentation in the stratotypic sections, it is possible to see that speciation
during the Aptian, which actually had begun during the late Bedoulian, just before
the anoxic event AOE1a (cf. et alii,
1998) was not a sudden explosion of new forms but a
rather gradual increase in the number of species. The Bedoulian-Gargasian
transitional beds correspond to a brief period of inactivity in planktonic
speciation. This makes it difficult to subdivide them and to delimit the
boundary between the two substages by means of planktonic foraminifera. It
appears easier to approximate this boundary through the use of benthic
foraminifera (Fig. 4 ![]() )
and ostracods (cf. & , this
volume).
)
and ostracods (cf. & , this
volume).
![]() The
early / middle Gargasian was a time of moderate revival, in a progressive and
regular rhythm, in the speciation of planktonic foraminifera. Their diversity
continued to increase but only slowly because most species already in existence were
still present at this level.
The
early / middle Gargasian was a time of moderate revival, in a progressive and
regular rhythm, in the speciation of planktonic foraminifera. Their diversity
continued to increase but only slowly because most species already in existence were
still present at this level.
![]() Therefore
the intra-Aptian "radiation" is not a brutal or sudden phenomenon, but
a progressive augmentation marked by two momentary episodes of acceleration in
speciation (anagenetic phases): 1) the lower/middle portion of the late
Bedoulian and 2) Gargasian (lowermost excluded), separated by a brief
stasigenetic period straddling the Bedoulian-Gargasian boundary. During the
period of time considered in this paper this radiation involves only
microperforate (Schackoinas) and finely perforate (Praehedbergella, Globigerinelloides)
planktonic foraminifera; macroperforate species (e.g. Hedbergella
trocoidea) (cf. et alii,
1998) do not appear until the late Gargasian.
Therefore
the intra-Aptian "radiation" is not a brutal or sudden phenomenon, but
a progressive augmentation marked by two momentary episodes of acceleration in
speciation (anagenetic phases): 1) the lower/middle portion of the late
Bedoulian and 2) Gargasian (lowermost excluded), separated by a brief
stasigenetic period straddling the Bedoulian-Gargasian boundary. During the
period of time considered in this paper this radiation involves only
microperforate (Schackoinas) and finely perforate (Praehedbergella, Globigerinelloides)
planktonic foraminifera; macroperforate species (e.g. Hedbergella
trocoidea) (cf. et alii,
1998) do not appear until the late Gargasian.
![]() In
our current state of knowledge it does not appear possible to relate these
biologic processes [a kind of "punctuated gradualism"] to variations
in geochemical and sedimentological parameters. Consequently, a direct causal
relationship between speciation in planktonic foraminifera and external factors
is at best hypothetical.
In
our current state of knowledge it does not appear possible to relate these
biologic processes [a kind of "punctuated gradualism"] to variations
in geochemical and sedimentological parameters. Consequently, a direct causal
relationship between speciation in planktonic foraminifera and external factors
is at best hypothetical.
![]() In
certain portions of the latest Bedoulian/early-middle Gargasian period
planktonic foraminifera show an increasing instability in the distinguishing
characters of theirs tests. This variability is particularly manifest in some
Praehedbergellas, and to a lesser extent in some "small few-chambered Globigerinelloides"
(in
& ,
2003a), and the group of clavate species. Sequences of
evolutive forms showing these fluctuating characters - which are quite apparent
at Cassis-La Bédoule - are imperfectly or incompletely recorded everywhere
else in the world, and the resulting lack of data has led many authors to
phylogenetic and therefore to taxonomic misinterpretations.
In
certain portions of the latest Bedoulian/early-middle Gargasian period
planktonic foraminifera show an increasing instability in the distinguishing
characters of theirs tests. This variability is particularly manifest in some
Praehedbergellas, and to a lesser extent in some "small few-chambered Globigerinelloides"
(in
& ,
2003a), and the group of clavate species. Sequences of
evolutive forms showing these fluctuating characters - which are quite apparent
at Cassis-La Bédoule - are imperfectly or incompletely recorded everywhere
else in the world, and the resulting lack of data has led many authors to
phylogenetic and therefore to taxonomic misinterpretations.
![]() The
detailed study of a rather close sampling of the expanded composite section of
Cassis-La Bédoule allowed us to follow step by step the evolution of
planktonic foraminifera in the beds involving the Bedoulian-Gargasian
transition. From a morphological point of view each lineage shows an evolution
consisting of successive stages of apparent stability alternating with brief
anagenetic sequences. In any case these forms cannot be defined taxonomically on
the bases of stable criteria for they are subject to a constant, more or less
rapid evolutionary slippage, which affects the mode of coiling, the number,
shape and rugosity of chambers,
etc. This variability leads to a relativization of the concept of species
(and even more of genus) in a continuum that drifts continuously.
The
detailed study of a rather close sampling of the expanded composite section of
Cassis-La Bédoule allowed us to follow step by step the evolution of
planktonic foraminifera in the beds involving the Bedoulian-Gargasian
transition. From a morphological point of view each lineage shows an evolution
consisting of successive stages of apparent stability alternating with brief
anagenetic sequences. In any case these forms cannot be defined taxonomically on
the bases of stable criteria for they are subject to a constant, more or less
rapid evolutionary slippage, which affects the mode of coiling, the number,
shape and rugosity of chambers,
etc. This variability leads to a relativization of the concept of species
(and even more of genus) in a continuum that drifts continuously.
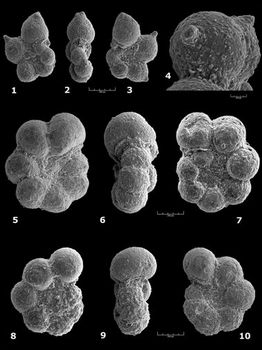
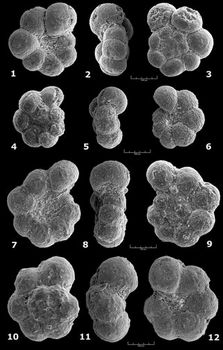
![]() One
particularly well-documented example (Fig. 6
One
particularly well-documented example (Fig. 6 ![]() )
in the Cassis-La Bédoule area involves forms that to begin with are distinctly
trochospiral, with 6 chambers (in the last whorl) that increase in size
moderately; the test shows a very slightly lobate outline. This is the taxon Praehedbergella
primare (cf. taxonomic notes). This relatively long-ranging species appears
in latest Barremian times (,
1966) and remains unchanged morphologically
(stasigenetic phase) until the middle of the upper Bedoulian, where some
specimens evolve by acquiring progressively a more distinctly lobate outline.
These variants were considered as discrete entities taxonomically (in ,
1974, under the name Hedbergella similis, a
junior synonym of Praehedbergella globulifera ( et ),
in , 1971).
Both taxa
(P. primare and P. globulifera), whose delimitation is not always
clear during the interval in which speciation took place, coexist during the
major part of the Gargasian. However, in the lowermost beds of the substage
(more exactly in the brief period of coexistence of Schackoina cabri and Globigerinelloides
ferreolensis) another process of rapid evolution is observed among
populations of 6-chambered Praehedbergellas. The stable forms, regarded as
typical of the two taxa mentioned above, are now a small minority; most of the
specimens, now strongly polymorphic, show a more and more marked tendency toward
evolving from a trochospiral coiling to a planispiral one (the ombilical and
spiral faces are less and less dissimilar, the aperture more and more
peripheral); at about the same time, via numerous intermediate stages, the last
whorl evolves from 6 to 7 chambers. During this short evolutionary phase the
identification and characterization of "species" (and even of genera,
based on the criterion - trochospiral vs. planispiral - mode of coiling) is made
difficult if the usual rules of Linnean nomenclature are applied. The two
terminal stabilized stages of this evolution are described in the literature
respectively under the names Praehedbergella luterbacheri, a very low
trochospiral typically 7-chambered form and Globigerinelloides ferreolensis,
a planispiral 7- and very soon 8-chambered form. Unfortunately some authors,
driven by a strictly typological approach that does not take these processes of
evolutionary slippage into account, have erected species for numerous
intermediate and transitory evolutionary stages selected at random in these
changing and multiform populations. Our position has been to assign names only
to the stabilized morphotypes.
)
in the Cassis-La Bédoule area involves forms that to begin with are distinctly
trochospiral, with 6 chambers (in the last whorl) that increase in size
moderately; the test shows a very slightly lobate outline. This is the taxon Praehedbergella
primare (cf. taxonomic notes). This relatively long-ranging species appears
in latest Barremian times (,
1966) and remains unchanged morphologically
(stasigenetic phase) until the middle of the upper Bedoulian, where some
specimens evolve by acquiring progressively a more distinctly lobate outline.
These variants were considered as discrete entities taxonomically (in ,
1974, under the name Hedbergella similis, a
junior synonym of Praehedbergella globulifera ( et ),
in , 1971).
Both taxa
(P. primare and P. globulifera), whose delimitation is not always
clear during the interval in which speciation took place, coexist during the
major part of the Gargasian. However, in the lowermost beds of the substage
(more exactly in the brief period of coexistence of Schackoina cabri and Globigerinelloides
ferreolensis) another process of rapid evolution is observed among
populations of 6-chambered Praehedbergellas. The stable forms, regarded as
typical of the two taxa mentioned above, are now a small minority; most of the
specimens, now strongly polymorphic, show a more and more marked tendency toward
evolving from a trochospiral coiling to a planispiral one (the ombilical and
spiral faces are less and less dissimilar, the aperture more and more
peripheral); at about the same time, via numerous intermediate stages, the last
whorl evolves from 6 to 7 chambers. During this short evolutionary phase the
identification and characterization of "species" (and even of genera,
based on the criterion - trochospiral vs. planispiral - mode of coiling) is made
difficult if the usual rules of Linnean nomenclature are applied. The two
terminal stabilized stages of this evolution are described in the literature
respectively under the names Praehedbergella luterbacheri, a very low
trochospiral typically 7-chambered form and Globigerinelloides ferreolensis,
a planispiral 7- and very soon 8-chambered form. Unfortunately some authors,
driven by a strictly typological approach that does not take these processes of
evolutionary slippage into account, have erected species for numerous
intermediate and transitory evolutionary stages selected at random in these
changing and multiform populations. Our position has been to assign names only
to the stabilized morphotypes.
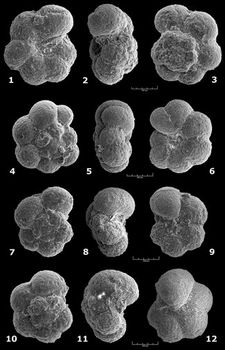
![]() Higher
in the sequence the only noticeable evolution affecting the planispiral forms in
strata of early / middle Gargasian age involves the number and shape of chambers.
Typical 7- or 8-chambered G. ferreolensis persist but fairly soon
variants, always few in number, appear. They include all the stages intermediate
to the 9-chambered (= G. barri) and higher in the succession to ten or
more chambered forms (= G. algerianus). In some individuals of all three
species (ferreolensis, barri, algerianus) there is also a
more or less sporadic tendency toward a lateral widening of the last chambers (=
"Biglobigerinella" trend). A trend toward uncoiling is also
apparent in these three taxa but is most prevalent in G. algerianus. A
great majority of authors are of opinion that neither of these variants is
worthy of generic status. On the other hand, the progressive acquisition of a
kind of peripheral keel by some specimens of G. algerianus has been named
at both the specific and the generic level (Pseudoplanomalina cheniourensis,
cf. et alii,
2002). The keeling process occurs near the upper
Gargasian / Clansayesian boundary but was not seen in the Cassis-La Bédoule area
owing to pre-Cenomanian erosion that removed the terms of the uppermost Aptian.
Higher
in the sequence the only noticeable evolution affecting the planispiral forms in
strata of early / middle Gargasian age involves the number and shape of chambers.
Typical 7- or 8-chambered G. ferreolensis persist but fairly soon
variants, always few in number, appear. They include all the stages intermediate
to the 9-chambered (= G. barri) and higher in the succession to ten or
more chambered forms (= G. algerianus). In some individuals of all three
species (ferreolensis, barri, algerianus) there is also a
more or less sporadic tendency toward a lateral widening of the last chambers (=
"Biglobigerinella" trend). A trend toward uncoiling is also
apparent in these three taxa but is most prevalent in G. algerianus. A
great majority of authors are of opinion that neither of these variants is
worthy of generic status. On the other hand, the progressive acquisition of a
kind of peripheral keel by some specimens of G. algerianus has been named
at both the specific and the generic level (Pseudoplanomalina cheniourensis,
cf. et alii,
2002). The keeling process occurs near the upper
Gargasian / Clansayesian boundary but was not seen in the Cassis-La Bédoule area
owing to pre-Cenomanian erosion that removed the terms of the uppermost Aptian.
![]() A
detailed statistical analysis of the microfauna as well as its relationships
with sedimentary data are beyond the scope of this essentially biostratigraphic
paper. However, some general observations based on foraminiferal distribution
suggest that during the Gargasian the sea was slightly deeper than it had been
in late Bedoulian times (when the paleodepth was estimated to be between one and
two hundred meters, et alii,
1998). The grounds for this presumption of a slight
deepening are species diversity and the planktonic / benthic ratio: both tend to
increase during the time in question. Furthermore, among benthic populations,
the proportion of agglutinated forms, particularly those with an organic wall (Bathysiphon,
Reophax, Recurvoides) tends to become greater during the
Gargasian. Finally, shallow-water species like Spirillina minima, Patellina
subcretacea, which occur more or less commonly in the Bedoulian are
extremely rare or absent during the Gargasian.
A
detailed statistical analysis of the microfauna as well as its relationships
with sedimentary data are beyond the scope of this essentially biostratigraphic
paper. However, some general observations based on foraminiferal distribution
suggest that during the Gargasian the sea was slightly deeper than it had been
in late Bedoulian times (when the paleodepth was estimated to be between one and
two hundred meters, et alii,
1998). The grounds for this presumption of a slight
deepening are species diversity and the planktonic / benthic ratio: both tend to
increase during the time in question. Furthermore, among benthic populations,
the proportion of agglutinated forms, particularly those with an organic wall (Bathysiphon,
Reophax, Recurvoides) tends to become greater during the
Gargasian. Finally, shallow-water species like Spirillina minima, Patellina
subcretacea, which occur more or less commonly in the Bedoulian are
extremely rare or absent during the Gargasian.
![]() This
micropaleontological study of a Gargasian composite section from the Bedoulian
stratotypic area allowed us to follow in great detail the evolution of
foraminiferal assemblages during the period of transition between the two Aptian
substages.
This
micropaleontological study of a Gargasian composite section from the Bedoulian
stratotypic area allowed us to follow in great detail the evolution of
foraminiferal assemblages during the period of transition between the two Aptian
substages.
![]() In
general, the changes observed among the benthic forms appear to be progressive
and reflect in particular a slight deepening upward during that time; several
species extinctions in the benthic microfauna are correlated with the Bedoulian
/ Gargasian boundary.
In
general, the changes observed among the benthic forms appear to be progressive
and reflect in particular a slight deepening upward during that time; several
species extinctions in the benthic microfauna are correlated with the Bedoulian
/ Gargasian boundary.
![]() Many
planktonic species have a relatively long range: some of them that make their
appeareance in the Bedoulian or even in the late Barremian are still present in
the Gargasian. However, some stable markers, their level of first appearance
well-calibrated, permit the division of the interval studied into five zones
instead of the three of the previous zonation. On either side of the
Bedoulian-Gargasian boundary certain groups are affected by a rapid yet
progressive morphologic instability. Such somewhat disordered phases of
evolution have not always been analyzed correctly by authors some of whom merely
multiplied new taxa in this ever-changing background. The greater thickness of
the succession and the absence of hiatuses in the Cassis-La Bédoule
stratotypic area allowed us to observe these processes in great detail and
especially to make certain their stratigraphical continuity. These initial
regional results need substantiation through a similar study of middle Aptian
microfaunas from another privileged stratotypic region, i.e. that of the
Apt-Gargas (work in progress) and, through the Vocontian Basin, to integrate the
data provided by the study of stratotypes into the Tethyan framework.
Many
planktonic species have a relatively long range: some of them that make their
appeareance in the Bedoulian or even in the late Barremian are still present in
the Gargasian. However, some stable markers, their level of first appearance
well-calibrated, permit the division of the interval studied into five zones
instead of the three of the previous zonation. On either side of the
Bedoulian-Gargasian boundary certain groups are affected by a rapid yet
progressive morphologic instability. Such somewhat disordered phases of
evolution have not always been analyzed correctly by authors some of whom merely
multiplied new taxa in this ever-changing background. The greater thickness of
the succession and the absence of hiatuses in the Cassis-La Bédoule
stratotypic area allowed us to observe these processes in great detail and
especially to make certain their stratigraphical continuity. These initial
regional results need substantiation through a similar study of middle Aptian
microfaunas from another privileged stratotypic region, i.e. that of the
Apt-Gargas (work in progress) and, through the Vocontian Basin, to integrate the
data provided by the study of stratotypes into the Tethyan framework.
![]() Presented
below is the minimum of information needed for the identification of the
principal species of planktonic foraminifera whose taxonomy is the most
controversial and thus subject to continual revision. At the generic level we
will use a classification which fundamentally results from the hierarchy of criteria
proposed by et alii
(2002) but as for small Globigerinelloides
includes a few changes adapted from the recent work of
& (2003a).
Complete synonymies, descriptions, more detailed discussions and more numerous
illustrations will be provided in a forthcoming, more paleontologically-oriented
paper, that will combine the observations on benthic and planktonic forms from
Basse Provence, the Aptian stratotypic area (Apt-Gargas basin) and the Vocontian
domain.
Presented
below is the minimum of information needed for the identification of the
principal species of planktonic foraminifera whose taxonomy is the most
controversial and thus subject to continual revision. At the generic level we
will use a classification which fundamentally results from the hierarchy of criteria
proposed by et alii
(2002) but as for small Globigerinelloides
includes a few changes adapted from the recent work of
& (2003a).
Complete synonymies, descriptions, more detailed discussions and more numerous
illustrations will be provided in a forthcoming, more paleontologically-oriented
paper, that will combine the observations on benthic and planktonic forms from
Basse Provence, the Aptian stratotypic area (Apt-Gargas basin) and the Vocontian
domain.
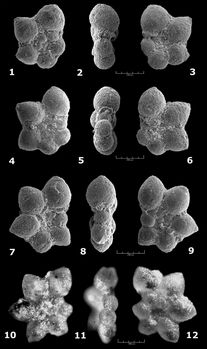
Praehedbergella aptiana (
1965)
Hedbergella aptiana , 1965, p. 347-348, text-figs. 3-6.
Blefuscuiana aptiana (), et , 1988, p. 158, pl. 5, figs. 1-3.
Blefuscuiana aptiana () sensu stricto, , et , 1997, p. 125, pl. 8.2, figs. 1-8.
Blefuscuiana aptiana () orientalis , , , et , 1996, p. 125-126, pl. 8.2, figs. 9-11.
![]() Remarks.
We share fully
& 's opinion (1988)
that the holotype described and illustrated in text-fig. 3 by
(1965) is a distorted specimen and thus paratypes are
more suitable for defining this species (particularly, in our opinion, those of
text-figs. 4 and 6).
Remarks.
We share fully
& 's opinion (1988)
that the holotype described and illustrated in text-fig. 3 by
(1965) is a distorted specimen and thus paratypes are
more suitable for defining this species (particularly, in our opinion, those of
text-figs. 4 and 6).
![]() Occurrence.
Middle Barremian to early Aptian according to
(1965). In the Cassis-La Bédoule area, this
species is rare during the latest Barremian, but moderately abundant in the
Bedoulian and early-middle Gargasian.
Occurrence.
Middle Barremian to early Aptian according to
(1965). In the Cassis-La Bédoule area, this
species is rare during the latest Barremian, but moderately abundant in the
Bedoulian and early-middle Gargasian.
Praehedbergella globulifera (
et , in
1971)
Clavihedbergella globulifera et (in ), 1971, p. 136, pl. 10, figs. 1a-c.
Hedbergella maslakovae , 1974, p. 61-63, pl. 20, figs. 1-3, 14-16; pl. 24, figs. 11-14.
Hedbergella similis , 1974, p. 68-69, pl. 16, figs. 10-21; pl. 18, figs. 12-13; pl. 23, figs. 14-16.
Lilliputianella globulifera et , 1971, , et , 1997, p. 164-165, pl. 9.1, figs. 5-15.
![]() Remarks.
The holotype of Praehedbergella globulifera has 5 chambers in the last
whorl, those of P. maslakovae and P. similis 6 chambers; these
species may also be distinguished typologically by the shape of their last
chambers, slightly more elongate radially in globulifera than in similis
and maslakovae (but not so much as to form cylindriform chambers like
those of P. roblesae). The analysis of populations shows that in fact all
these forms constitute a single but highly variable plexus (as demonstrated in 's
numerous illustrations (1974) of P. similis) and
that the holotypes of P. globulifera, P. maslakovae and P.
similis represent only random morphotypes among many others. Taking
anteriority into account, we are of the opinion that et alii
(1997) were right in considering P. maslakovae
and P. similis as junior synonyms of the species described by
& (1971).
Remarks.
The holotype of Praehedbergella globulifera has 5 chambers in the last
whorl, those of P. maslakovae and P. similis 6 chambers; these
species may also be distinguished typologically by the shape of their last
chambers, slightly more elongate radially in globulifera than in similis
and maslakovae (but not so much as to form cylindriform chambers like
those of P. roblesae). The analysis of populations shows that in fact all
these forms constitute a single but highly variable plexus (as demonstrated in 's
numerous illustrations (1974) of P. similis) and
that the holotypes of P. globulifera, P. maslakovae and P.
similis represent only random morphotypes among many others. Taking
anteriority into account, we are of the opinion that et alii
(1997) were right in considering P. maslakovae
and P. similis as junior synonyms of the species described by
& (1971).
![]() Occurrence.
This plexus first appears in the late Bedoulian, is rather well-represented in
the early Gargasian and progressively decreases in frequency in the middle
Gargasian.
Occurrence.
This plexus first appears in the late Bedoulian, is rather well-represented in
the early Gargasian and progressively decreases in frequency in the middle
Gargasian.
Praehedbergella gorbachikae (
1974)
Pl. 5, figs. 7-12
![]()
Hedbergella gorbachikae , 1974, p. 56-58, pl. 15, figs. 1-16.
![]() Remarks.
This species belongs to the Praehedbergella infracretacea group, in
which it is distinguished by a more protruding adombilical part of the last
chamber.
Remarks.
This species belongs to the Praehedbergella infracretacea group, in
which it is distinguished by a more protruding adombilical part of the last
chamber.
![]() Occurrence.
(1974) proposed this taxon as the index of a new
Gargasian zone, intercalated between the Algerianus and Trocoidea zones. Our
observations led us to reject this proposal because 1) P. gorbatchikae
constitutes a very marginal component of the infracretacea group, 2) the
first appearance of this species coincides with that of Globigerinelloides
algerianus, a more significant marker.
Occurrence.
(1974) proposed this taxon as the index of a new
Gargasian zone, intercalated between the Algerianus and Trocoidea zones. Our
observations led us to reject this proposal because 1) P. gorbatchikae
constitutes a very marginal component of the infracretacea group, 2) the
first appearance of this species coincides with that of Globigerinelloides
algerianus, a more significant marker.
Praehedbergella infracretacea (
1937)
Globigerina infracretacea , 1937, p. 28, text-fig. 1.
Globigerina aptica , 1951, p. 49, pl. 8, figs. 9-11.
Hedbergella delrioensis (), , 1974, p. 54-55 (pro parte : pl. 10, figs. 10-12; pl. 13, figs. 3-5, 15-18).
Praehedbergella luterbacheri (
1974)
Pl. 3, figs. 5-10
![]() ; Pl. 4, figs. 1-6
; Pl. 4, figs. 1-6
![]()
Hedbergella luterbacheri , 1974, p. 61, pl. 19, figs. 21-23; 24-26; pl. 26, figs. 15-17.
Praeglobotruncana modesta , 1959, p. 267 (pro parte : at least mentioned but not illustrated forms from the Biglobigerinella barri zone, Maridale formation).
Hedbergella planispira () 1940, , 1974, p. 64-65 (pro parte : Aptian 7-chambered forms, i.e.: pl. 11, fig. 4-6; pl. 23, figs. 17-18).
Blefuscuiana kuznetsovae et , 1988, p. 156-158, (pro parte : pl. 3, fig. 10; pl. 4, figs. 1-2, 4; non pl. 4, fig. 3).
![]() Remarks.
We follow the author of this species in limiting the definition of Praehedbergella
luterbacheri to low trochospiral 7- (rarely 8-) chambered forms with a
rather slowly opening spire and a moderately lobate outline. The 7-chambered
specimens of P. kuznetsovae, including the holotype, have a spire that
opens slightly less slowly. Taking into account the range of variability within
populations we are led to regard P. kuznetsovae as a junior synonym of P.
luterbacheri.
Remarks.
We follow the author of this species in limiting the definition of Praehedbergella
luterbacheri to low trochospiral 7- (rarely 8-) chambered forms with a
rather slowly opening spire and a moderately lobate outline. The 7-chambered
specimens of P. kuznetsovae, including the holotype, have a spire that
opens slightly less slowly. Taking into account the range of variability within
populations we are led to regard P. kuznetsovae as a junior synonym of P.
luterbacheri.
![]() Our
observations in the Cassis-La Bédoule area show that by a progressive evolution
of the coiling of the initially low trochospiral test towards planispirality, P.
luterbacheri is the immediate ancestor of Globigerinelloides ferreolensis
(cf. Pl. 4, figs.
7-9
Our
observations in the Cassis-La Bédoule area show that by a progressive evolution
of the coiling of the initially low trochospiral test towards planispirality, P.
luterbacheri is the immediate ancestor of Globigerinelloides ferreolensis
(cf. Pl. 4, figs.
7-9
![]() ).
).
![]() Occurrence.
Because it first appears in the upper part of the Cabri zone (sensu lato),
just a few meters above the Bedoulian-Gargasian boundary, this species
constitutes an interesting marker which can be used to approximate the boundary
between the two substages and to be the index of a new zone intercalated between
the Cabri zone (s.s.) and the Ferreolensis zone (see the biostratigraphic
section of this paper). Praehedbergella luterbacheri, most often
represented by few specimens, becomes rare in the upper part of the Ferreolensis
zone and disappears near the Ferreolensis-Algerianus zonal boundary.
Occurrence.
Because it first appears in the upper part of the Cabri zone (sensu lato),
just a few meters above the Bedoulian-Gargasian boundary, this species
constitutes an interesting marker which can be used to approximate the boundary
between the two substages and to be the index of a new zone intercalated between
the Cabri zone (s.s.) and the Ferreolensis zone (see the biostratigraphic
section of this paper). Praehedbergella luterbacheri, most often
represented by few specimens, becomes rare in the upper part of the Ferreolensis
zone and disappears near the Ferreolensis-Algerianus zonal boundary.
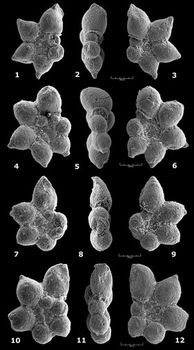
Praehedbergella praetrocoidea (
et 1986)
Pl. 4, figs. 10-12
![]() ; Pl. 5, figs. 1-6
; Pl. 5, figs. 1-6
![]()
Hedbergella praetrocoidea et (in ), 1986, p. 95, pl. 16, figs. 3-5.
![]() Occurrence.
Species sporadically present, always very few in number, in the early-middle
Gargasian.
Occurrence.
Species sporadically present, always very few in number, in the early-middle
Gargasian.
Praehedbergella primare (
et 1986)
Clavihedbergella primare et (in ), 1986, p. 120-121, pl. 25, figs. 2a-3a, 2b-3b.
Hedbergella sp., aff. planispira (), , 1966, p. 94, pl. 8, figs. 1-3.
Hedbergella planispira (, 1940), , 1974, p. 64-65 (pro parte : Aptian 6-chambered forms): pl. 23, figs. 5-7.
Hedbergella aff. planispira auct., , 1979, pl. 2, figs. 1-2.
Blefuscuiana kuznetsovae et , 1988, p. 156-158 (pro parte : pl. 4, fig. 3)
Hedbergella kuznetsovae ( et ), et , 1994, p. 676, fig. 12: 1-12.
Blefuscuiana primare ( et , 1986), , et , 1997, p. 131, pl. 8.10, figs. 1-3.
Praehedbergella kuznetsovae ( et , 1988), , , et , 1998, p. 207-208, pl. 3, figs. 8-9.
![]() Remarks.
& (in , 1986)
defined this
species as having 5.5 to 6 chambers in the last whorl and illustrate a holotype
with 6 chambers. Initially, the rather succinct description and mediocre
illustrations provided by the authors did not give a precise idea of the
determining characteristics of this species.
Remarks.
& (in , 1986)
defined this
species as having 5.5 to 6 chambers in the last whorl and illustrate a holotype
with 6 chambers. Initially, the rather succinct description and mediocre
illustrations provided by the authors did not give a precise idea of the
determining characteristics of this species.
![]() But figures of topotypes in ,
& (1997)
lead us to believe that Praehedbergella primare is in agreement with the
concept of finely perforate, low trochospiral Barremo-Aptian forms, with 6
chambers in the last whorl that increase slowly; forms of this description have
been referred to Hedbergella planispira (as "aff. planispira")
by several authors, but these homeomorphs of H. planispira cannot be
attributed to this Albian species, which has a macroperforate test, nor to Praehedbergella kuznetsovae
for that species is typically 7-chambered with a spire that opens slightly less
slowly.
But figures of topotypes in ,
& (1997)
lead us to believe that Praehedbergella primare is in agreement with the
concept of finely perforate, low trochospiral Barremo-Aptian forms, with 6
chambers in the last whorl that increase slowly; forms of this description have
been referred to Hedbergella planispira (as "aff. planispira")
by several authors, but these homeomorphs of H. planispira cannot be
attributed to this Albian species, which has a macroperforate test, nor to Praehedbergella kuznetsovae
for that species is typically 7-chambered with a spire that opens slightly less
slowly.
![]() Occurrence.
Crimea: late Barremian-early Aptian. In SE France, this species appears in the
latest Barremian and continues (decreasing in frequency) up to the middle
Gargasian (Algerianus zone).
Occurrence.
Crimea: late Barremian-early Aptian. In SE France, this species appears in the
latest Barremian and continues (decreasing in frequency) up to the middle
Gargasian (Algerianus zone).
Praehedbergella roblesae ( 1959)
Globigerina roblesae , 1959, p. 149, pl. 4, fig. 4.
Schackoina gr. cabri (
1952)
Schackoina cabri , 1952, p. 20-21, figs. 1-8.
Schackoina pustulans , 1957, p. 274-275, pl. 1, figs. 1-4, 6-7.
Leupoldina protuberans , 1957, p. 277, pl. 2, figs. 1-13.
Globigerinelloides algerianus (
et 1948)
Globigerinelloides algeriana et , 1948, p. 43, pl. 8, figs. 4-6.
![]() Remarks.
Test planispiral, with at least 10, typically 10-12 or even more chambers in the
last whorl; some specimens show a trend toward uncoiling in the last or the last
few chambers or to widening until final bilobation.
Remarks.
Test planispiral, with at least 10, typically 10-12 or even more chambers in the
last whorl; some specimens show a trend toward uncoiling in the last or the last
few chambers or to widening until final bilobation.
![]() Occurrence.
This Tethyan middle-late Gargasian marker was seen only in the last meters of
the Cassis-La Marcouline section, where it is represented by rather rare
10-11-chambered specimens.
Occurrence.
This Tethyan middle-late Gargasian marker was seen only in the last meters of
the Cassis-La Marcouline section, where it is represented by rather rare
10-11-chambered specimens.
Globigerinelloides gr. aptiensis (
1974)
Globigerinelloides aptiense , 1974, p. 79-80, pl. 4, figs. 9-10; pl. 8, figs. 4-6, 17-18.
Blowiella solida et (in ), 1986, p. 123, pl. 27, fig. 3.
![]() Remarks.
These two planispiral 6-chambered forms differ typologically only in the degree
of opening of the spire, which is very moderate in the solida morphotype,
but slightly more rapid in the aptiensis morphotype; Here, again, between
these extremes many intermediate stages exist; in addition, the stratigraphic
distribution of all these varied and rather sporadic morphotypes is identical.
We are therefore led to follow
& (2003b)
in considering Blowiella solida as a junior synonym of Globigerinelloides
aptiensis. We have observed within this plexus the occurrence of very rare Biglobigerinella-like
specimens; such a trend to the bilobation of the last chamber is more common
among the solida morphotype.
Remarks.
These two planispiral 6-chambered forms differ typologically only in the degree
of opening of the spire, which is very moderate in the solida morphotype,
but slightly more rapid in the aptiensis morphotype; Here, again, between
these extremes many intermediate stages exist; in addition, the stratigraphic
distribution of all these varied and rather sporadic morphotypes is identical.
We are therefore led to follow
& (2003b)
in considering Blowiella solida as a junior synonym of Globigerinelloides
aptiensis. We have observed within this plexus the occurrence of very rare Biglobigerinella-like
specimens; such a trend to the bilobation of the last chamber is more common
among the solida morphotype.
![]() Occurrence.
Rare specimens assignable to this plexus appear in the late Bedoulian, are
sporadically present in the early-middle Gargasian and become even rarer higher
in the series.
Occurrence.
Rare specimens assignable to this plexus appear in the late Bedoulian, are
sporadically present in the early-middle Gargasian and become even rarer higher
in the series.
Globigerinelloides barri (,
et , 1957)
Globigerinelloides barri , et , 1957, p. 25 (pro parte: pl. 1, figs. 17, 18 a-b).
![]() Remarks.
Test planispiral with 9 chambers in the last whorl. Phylogenetic successor of
G. ferreolensis.
Remarks.
Test planispiral with 9 chambers in the last whorl. Phylogenetic successor of
G. ferreolensis.
"Globigerinelloides" gr. bizonae (
1961) – saundersi (,
1959)
Pl. 1, figs. 1-12
![]() ; Pl. 2, figs. 1-12
; Pl. 2, figs. 1-12
![]() ; Pl. 3, figs. 1-4
; Pl. 3, figs. 1-4
![]()
Planomalina saundersi , 1959, p. 262, pl. 20, figs. 9-11.
Hastigerinella bizonae , 1961, p. 34-36, pl. 1, figs. 24-28.
Clavihedbergella bizonae (, 1961), , 1971, p. 231-232, pl. 3, figs. 2 a-c.
Hedbergella kuhryi , 1974, p. 60, pl. 14, figs. 1-6.
non Claviblowiella saundersi (, 1959), , et , 1997, p. 181, pl. 10.2, figs. 7-11.
Blowiella saundersi (, 1959), , , et , 1998, p. 205-206, pl. 4, fig. 6.
Claviblowiella saundersi (, 1959), et , 2004, p. 42, pl. 3, figs. 3a-b.
![]() Remarks.
To make a better comparison of the species named Clavihedbergella bizonae
by
(1961) with that described by
(1959) as Planomalina saundersi, we have examined
the holotype of bizonae (deposited in the collections of the Museum of
Natural History of Paris, N° F62120). It is re-illustrated herein as it appears
under an optical microscope (Pl. 1, figs. 10-12
Remarks.
To make a better comparison of the species named Clavihedbergella bizonae
by
(1961) with that described by
(1959) as Planomalina saundersi, we have examined
the holotype of bizonae (deposited in the collections of the Museum of
Natural History of Paris, N° F62120). It is re-illustrated herein as it appears
under an optical microscope (Pl. 1, figs. 10-12
![]() ). The shape of the last chambers
- ogival and pointed with an apical tapering suggesting that it is the base of a
spine, although almost always found broken at the base- is similar in the two
taxa.
(1961) defined the mode of coiling of his species as
"trochoďde" (= trochospiral), a construction that can be seen on the
figured paratypes; however, the holotype appears to be weakly dissymmetrical and
shows a tendency toward planispirality. The holotype of the species of
is described as planispiral by its author, but the profile shows a slight
asymmetry. Therefore using a strict typologic approach it is hard to find any
objective argument that supports a differentiation of the two species.
). The shape of the last chambers
- ogival and pointed with an apical tapering suggesting that it is the base of a
spine, although almost always found broken at the base- is similar in the two
taxa.
(1961) defined the mode of coiling of his species as
"trochoďde" (= trochospiral), a construction that can be seen on the
figured paratypes; however, the holotype appears to be weakly dissymmetrical and
shows a tendency toward planispirality. The holotype of the species of
is described as planispiral by its author, but the profile shows a slight
asymmetry. Therefore using a strict typologic approach it is hard to find any
objective argument that supports a differentiation of the two species.
![]() The
stratigraphic information in 's
thesis (1960) shows that
the type-population (holotype plus 10 "paratypoïdes" (=
paratypes),
of which only 4 specimens - the most clearly
trochospiral - were illustrated again in his paper of 1961)
comes from only one
sample (labelled MG 781). This sample was taken in the La Marcouline Quarry, precisely
in the more marly uppermost unit
that we have dated middle Gargasian (Algerianus Zone). Furthermore,
(1960) quoted the
occurrence of G. algerianus in levels that include sample 781. The
specimens of "Globigerinelloides" bizonae-saundersi
that we illustrate Pl. 3, figs. 1-4
The
stratigraphic information in 's
thesis (1960) shows that
the type-population (holotype plus 10 "paratypoïdes" (=
paratypes),
of which only 4 specimens - the most clearly
trochospiral - were illustrated again in his paper of 1961)
comes from only one
sample (labelled MG 781). This sample was taken in the La Marcouline Quarry, precisely
in the more marly uppermost unit
that we have dated middle Gargasian (Algerianus Zone). Furthermore,
(1960) quoted the
occurrence of G. algerianus in levels that include sample 781. The
specimens of "Globigerinelloides" bizonae-saundersi
that we illustrate Pl. 3, figs. 1-4
![]() come from this unit.
come from this unit.
![]() A
meticulous analysis of the populations that we have followed step by step in the
sections of the historical stratotype of Cassis-La Bédoule shows
that those from the latest Bedoulian-early Gargasian are composed
predominantly of distinctly trochospiral specimens. However, a part of or even
the complete internal whorl is often visible on the umbilical side, and in
addition its surface is slightly more depressed than that of its dorsal
homologue. As early as the lower Gargasian a trend toward a more symmetrical
coiling appears and is developed in an increasing number of specimens upward in
the series. In the middle Gargasian these subsymmetrical specimens predominate
but they are always accompanied by few more distinctly dissymmetrical
individuals.
A
meticulous analysis of the populations that we have followed step by step in the
sections of the historical stratotype of Cassis-La Bédoule shows
that those from the latest Bedoulian-early Gargasian are composed
predominantly of distinctly trochospiral specimens. However, a part of or even
the complete internal whorl is often visible on the umbilical side, and in
addition its surface is slightly more depressed than that of its dorsal
homologue. As early as the lower Gargasian a trend toward a more symmetrical
coiling appears and is developed in an increasing number of specimens upward in
the series. In the middle Gargasian these subsymmetrical specimens predominate
but they are always accompanied by few more distinctly dissymmetrical
individuals.
![]() Added
to this fluctuation in the mode of coiling is some morphologic variability in
the shape of the chambers, in particular a more or less radial elongation that
can attain the "cepedai" stage, described as a true species by (1959)
but which seems to represent only an extreme variant (very rare in our
populations) of the species of .
Added
to this fluctuation in the mode of coiling is some morphologic variability in
the shape of the chambers, in particular a more or less radial elongation that
can attain the "cepedai" stage, described as a true species by (1959)
but which seems to represent only an extreme variant (very rare in our
populations) of the species of .
![]() Therefore,
taking into account both the features of the holotypes and intra-populational
variability, it appears that
the species described by
(1961) cannot be distinguished from that of
(1959) and must be regarded as a junior synonym.
Therefore,
taking into account both the features of the holotypes and intra-populational
variability, it appears that
the species described by
(1961) cannot be distinguished from that of
(1959) and must be regarded as a junior synonym.
![]() Within
this plexus the fluctuation of characters (e.g. mode of coiling, and the spinose
extension on last chambers), considered of generic significance by a
majority of authors, causes a problem in generic assignment. As a provisional
measure pending additional research we group these forms under the provisional
appellation "Globigerinelloides".
Within
this plexus the fluctuation of characters (e.g. mode of coiling, and the spinose
extension on last chambers), considered of generic significance by a
majority of authors, causes a problem in generic assignment. As a provisional
measure pending additional research we group these forms under the provisional
appellation "Globigerinelloides".
Globigerinelloides gr. blowi (
1959)
Planomalina blowi , 1959, p. 260, pl. 20, figs. 2-3.
Planomalina maridalensis , 1959, p. 261, pl. 20, figs. 4-6.
Globigerinella duboisi , 1961, p. 33, pl. 1, figs. 14-18.
Globigerinella gottisi , 1961, p. 32 (pro parte : pl. 1, figs. 9-11, 13).
Globigerinelloides paragottisi et , 2003, p. 332-333, figs. 6. 7-14, 7. 1-13.
![]() Remarks.
In this few-chambered planispiral plexus, G. duboisi (typically 4
chambers) et G. blowi (5 chambers) are dominant morphotypes, always
present simultaneously among the populations. They are linked by a minority of
individuals comprising many intermediate stages, both in the shape and in the
number of chambers. The G. maridalensis and G. paragottisi
morphotypes are seen only very sporadically so their recognition as species does
not seem worthwhile.
Remarks.
In this few-chambered planispiral plexus, G. duboisi (typically 4
chambers) et G. blowi (5 chambers) are dominant morphotypes, always
present simultaneously among the populations. They are linked by a minority of
individuals comprising many intermediate stages, both in the shape and in the
number of chambers. The G. maridalensis and G. paragottisi
morphotypes are seen only very sporadically so their recognition as species does
not seem worthwhile.
![]() Occurrence.
All the forms cited above are more or less constantly present from late
Bedoulian to late Gargasian times.
Occurrence.
All the forms cited above are more or less constantly present from late
Bedoulian to late Gargasian times.
Globigerinelloides ferreolensis (
1961)
Biticinella ferreolensis , 1961, p. 214, pl. 1, figs. 1-5.
Biglobigerinella sigali , 1961, p. 33, pl. 1, figs. 19-23.
Globigerinelloides macrocameratus , 1974, p. 85-86, pl. 5, figs. 1-6.
![]() Remarks.
Test planispiral, 7-8 chambers in the last whorl. In the lowermost Gargasian
this species evolved from low trochospirally coiled specimens of Praehedbergella
luterbacheri with an increasingly flattened trochospire and a spiral side
differing hardly at all from the umbilical one; the spiral side shows only a
slightly greater part of the chambers of the internal whorl. The aperture too
tends to become more and more symmetrical. The evolution of all these characters
takes place within a progressive but rapid evolution towards planispirality, and
include various intermediate stages. These variants do not justify the erection
of a new species (a kind of "praeferreolensis" stage), which in
any event cannot be attributed to a precise genus since none can be considered
as belonging to typical
Praehedbergella nor to Globigerinelloides.
Remarks.
Test planispiral, 7-8 chambers in the last whorl. In the lowermost Gargasian
this species evolved from low trochospirally coiled specimens of Praehedbergella
luterbacheri with an increasingly flattened trochospire and a spiral side
differing hardly at all from the umbilical one; the spiral side shows only a
slightly greater part of the chambers of the internal whorl. The aperture too
tends to become more and more symmetrical. The evolution of all these characters
takes place within a progressive but rapid evolution towards planispirality, and
include various intermediate stages. These variants do not justify the erection
of a new species (a kind of "praeferreolensis" stage), which in
any event cannot be attributed to a precise genus since none can be considered
as belonging to typical
Praehedbergella nor to Globigerinelloides.
![]() Even
when the coiling mode is stabilized, the intra-populational morphologic
variability of the species ferreolensis appears to be rather extensive.
This may involve the widening rate of the chambers (until they become bilobate,
a feature leading to a "Biglobigerinella" morphotype), the
opening rhythm of the spire, and an increase in the height of the chambers, but
there is no tendency to uncoiling (as in G. algerianus); a very low rate
of increase of chamber height in some specimens does not justify the erection of
the species "macrocameratus" (as described by , 1974).
Even
when the coiling mode is stabilized, the intra-populational morphologic
variability of the species ferreolensis appears to be rather extensive.
This may involve the widening rate of the chambers (until they become bilobate,
a feature leading to a "Biglobigerinella" morphotype), the
opening rhythm of the spire, and an increase in the height of the chambers, but
there is no tendency to uncoiling (as in G. algerianus); a very low rate
of increase of chamber height in some specimens does not justify the erection of
the species "macrocameratus" (as described by , 1974).
![]() Occurrence.
Gargasian species (lowest terms at the base of the substage excluded).
Occurrence.
Gargasian species (lowest terms at the base of the substage excluded).
![]() The authors wish to express their gratitude to Mrs M.-T. (Muséum
National d'Histoire Naturelle, Paris) for having placed at their disposal the
type-material of the 's collection (and
particularly the holotype of "Hastigerinella bizonae", N° F62120), to Mr Omar (Service MEB de l'UFR
de "Sciences de la Terre et évolution des milieux naturels" de
l'Université Pierre & Marie Curie (UPMC, Paris) for the SEM pictures and Mr
Claude (Atelier de Reprographie du
Département de Géologie sédimentaire de l'UPMC, Paris) for the image
processing.
The authors wish to express their gratitude to Mrs M.-T. (Muséum
National d'Histoire Naturelle, Paris) for having placed at their disposal the
type-material of the 's collection (and
particularly the holotype of "Hastigerinella bizonae", N° F62120), to Mr Omar (Service MEB de l'UFR
de "Sciences de la Terre et évolution des milieux naturels" de
l'Université Pierre & Marie Curie (UPMC, Paris) for the SEM pictures and Mr
Claude (Atelier de Reprographie du
Département de Géologie sédimentaire de l'UPMC, Paris) for the image
processing.
![]() They
are also indebted to Nestor J. for his
invaluable assistance in the translation of the French version of this paper,
and to Mesdames M. (University of Fribourg,
Switzerland) and D. (University of
Strasbourg, France) for a critical review of the manuscript.
They
are also indebted to Nestor J. for his
invaluable assistance in the translation of the French version of this paper,
and to Mesdames M. (University of Fribourg,
Switzerland) and D. (University of
Strasbourg, France) for a critical review of the manuscript.
![]()
![]()
D.A. (1951).- Studies of microfauna from Cretaceous deposits of Azerbaydzhan.- In: D.I., D.A. & D.M. (Eds), (Aznefteizdat, Baku) 128 pp. (in Russian)
N.V. (1962).- Désagrégation des roches argilo-marneuses sous l'action simultanée de l'hyposulfite et de l'eau oxygénée.- Voprosy Mikropaleontologii, Moscow, N° 6, pp. 147-148. (in Russian; traduction B.R.G.M. N° 4123).
F.T. & D. (1988).- A review and revision of the Jurassic–Early Cretaceous Globigerinina, with especial reference to the Aptian assemblages of Speeton (North Yorkshire, England).- Journal of Micropaleontology, London, Vol. 7, Part 2, pp. 143–185.
H. (1965).- Taxionomische Revision und Nomenklator zu Franz E. "Standard-Gliederung der Nordwest-deutschen Unterkreide nach Foraminiferen" (1938). Teil 4: Alb. Mit Beschreibungen von Arten aus verschiedenen Unterkreide-Niveaus.- Senckenbergiana Lethaea, Frankfurt am Main, Bd. 46, N° 4/6, pp. 327-366.
H.M. (1957).- The foraminiferal genera Schackoina , emended and Leupoldina, n. gen. in the Cretaceous of Trinidad B.W. I.- Eclogae Geologicae Helvetiae, Basel, Vol. 50, pp. 271–278.
H.M. (1959).- Planktonic foraminifera from the Cretaceous of Trinidad, B.W.I.- Bulletins of American Paleontology, New York, Vol. XXXIX, pp. 257–277.
H.M., A.R. & H. (1957).- Planktonic foraminiferal families Hantkeninidae, Orbulinidae, Globorotaliidae, and Globotruncanidae.- In: A.R. et alii (Eds.), Studies in foraminifera.- United States National Museum, Bulletin, Washington, Vol. 215, pp. 3–50.
M.K., F.T., T.N., M.D. & J.E. (1996).- Evolution in the Early Cretaceous planktonic foraminiferal genus Blefuscuiana.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Bd. 202, pp. 243–258.
M.K., F.T. & J.E. (eds.) (1997).- The early evolutionary history of planktonic foraminifera.- Chapman & Hall, London, 269 p.
M.K., F.T., T.N., M.D. & J.E. (1998).- On the evolution of the Hedbergellidae from the Praehedbergellidae.- Journal of Micropaleontology, London, Vol. 17, pp. 97-103.
G., M. & B. (1965).- Utilisation de l'azote liquide et des mouillants pour l'extraction et la préparation des microfossiles.- Comptes Rendus de l'Académie des Sciences Paris, t. 261, pp. 2383-2386.
M. & P. (1983).- Evolution of early planktonic foraminifers.- Marine Micropaleontology, Amsterdam, Vol. 7, Issue 6, pp. 453-462.
J. (1960).- Contribution à l'étude stratigraphique et micropaléontologique du Crétacé inférieur du Languedoc et de la Provence.- Diplôme d'Etudes Supérieures, Université de Paris, 151 p.
J. (1961).- Quelques nouvelles espèces dans le Crétacé inférieur méditerranéen.- Revue de Micropaléontologie, Paris, 4e Année, N° 1, pp. 30–36.
R. & I. (1994).- Planktonic foraminifera from the Lower Cretaceous of Rio Argos sections (southern Spain) and biostratigraphic implications.- Cretaceous Research, London, Vol. 15, Issue 6, pp. 645–687.
G. (1994).- La limite Bédoulien-Gargasien dans la coupe stratotypique de Cassis-La Bédoule (Bouches-du-Rhône, France).- Géologie Alpine, Grenoble, Mém. H.S., N° 20, p. 321-326.
J.A. & A. (1948).- Globigerinelloides, a new genus of the Globigerinidae.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon, Vol. 24, pp. 42–44.
D., D., B., S. & F. (2004).- Enregistrement à haute résolution des modifications environnementales inscrites dans un cadre téphrochronologique : le bassin du Western Interior au passage Cénomanien-Turonien (USA).- Bulletin de la Société géologique de France, Paris, t. 175, N° 6, pp. 561-572.
S., M. & G. (1965).- Le Bédoulien dans sa région-type, La Bédoule-Cassis (B.-du-Rh.).- In: Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Bureau de Recherches Géologiques et Minières, Mémoires, Orléans, N° 34, pp. 173-179.
G. (1987).- Dynamique du bassin subalpin méridional de l'Aptien au Cénomanien.- École des Mines de Paris, Mémoires des Sciences de la Terre, N° 4, 370 p.
G. (1975).- Contribution à l'étude stratigraphique et sédimentologique de l'Aptien supérieur de Provence.- Thèse, Université de Provence, 92 p.
M.F. (1937).- Planktonforaminiferen aus der Kreide und dem Eozän und ihre stratigraphische Bedeutung.- Etudy po Mikropaleontologii, Paleontologicheskaya Laboratoriya Moskovskogo Gosudarstvennogo Universiteta, 1, pp. 27–46.
T.N. (1971).- On Early Cretaceous foraminifera of the Crimea.- Voprosy Mikropaleontologii, Moscow, N° 14, pp. 125–139 & 211-216, pls. XXI-XXX. (in Russian)
T.N. (1986).- Jurassic and Early Cretaceous planktonic foraminifera of the south of the USSR.- (Akademia Nauk SSSR, Nauka, Moscou), 239 p. (in Russian)
B. (1971).- Lower Cretaceous planktonic Foraminifera from the Miravetes, Argos and Represa Formations (S.E. Spain).- Revista Española de Micropaleontologia, Madrid, Vol. III, N° 3, pp. 219-237.
S. & A. (2004).- Aptian planktonic foraminifera from Israel.- Israel Journal of Earth Sciences, Jerusalem, Vol. 53, N° 1, pp. 27-46.
J.F. (1974).- Stratigraphic, morphologic and taxonomic studies of Aptian planktonic Foraminifera.- Revista Española de Micropaleontologia, Madrid, Numero Extraordinario, pp. 1–107.
F., M. & G. (1984).- Crétacé inférieur : 1.3.6. -Foraminifères planctoniques et benthiques de bassin.- In: S. et alii (Eds.), Synthèse Géologique du Sud-Est de la France. I : Stratigraphie et paléogéographie.- Bureau de Recherches Géologiques et Minières, Mémoires, Orléans, N° 125, pp. 297-299.
M. (1961).- Quelques Foraminifères et Ostracodes nouveaux du Crétacé inférieur des Baronnies (Drôme).- Revue de Micropaléontologie, Paris, 3e Année, N° 4, pp. 213–216.
M. (1966).- Etude stratigraphique et micropaléontologique du Crétacé inférieur de la "fosse vocontienne".- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, N° 15, pp. 1–369.
M. & G. (2004).- Le Gargasien (Aptien moyen) dans les stratotypes historiques de l'Aptien (SE France) : Introduction générale.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/01, pp. 1-3.
M., G., W. & J.-P. (1998).- Les Foraminifères benthiques et planctoniques de la série du Stratotype Historique de l’Aptien inférieur dans la région de Cassis-La Bédoule (SE France).- Géologie Méditerranéenne, Marseille, t. XXV, N° 3-4, pp. 187–225.
M., J.-P. & G. (2002).- Hierarchy of criteria, evolutionary processes and taxonomic simplification in the classification of Lower Cretaceous planktonic foraminifera.- Cretaceous Research, London, Vol. 23, Issue 1, pp. 111–148
M., G., W., M. & J.-P. (2004).- Le Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de l'Aptien inférieur, SE France) : localisation géographique et corrélations stratigraphiques.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/02, pp. 1-4.
J. (1959).- Foraminíferos de la Formación La Peña.- Boletín de la Asociación Mexicana de Geólogos Petroleros, México, Vol. XI, N° 3-4, pp. 135–153.
F. & M. (1995).- Foraminifères planctoniques du Crétacé : commentaire de la zonation Europe-Méditerranée.- Bulletin de la Société Géologique de France, Paris, t. 166, N° 6, pp. 681-692.
P., G., R., J.-P. & M. (1998).- Les faunes d'Ammonites du Barrémien supérieur/Aptien inférieur (Bédoulien) dans la région stratotypique de Cassis-La Bédoule (SE France) : état des connaissances et propositions pour une zonation par Ammonites du Bédoulien-type.- Géologie Méditerranéenne, Marseille, t. XXV, N° 3-4, pp. 167–175.
J. (1952).- Aperçu stratigraphique sur la micropaléontologie du Crétacé.- XIXe Congrès Géologique International, Monographies régionales, (1re Série : Algérie), Alger, N° 26, pp. 3–47.
H. (1940).- Foraminifera from the Grayson Formation of northern Texas.- Journal of Paleontology, Tulsa, Vol. 14, N° 2, pp. 93-126.
G. (1981).- Les Foraminifères crétacés de Provence (Aptien-Santonien) – systématique, biostratigraphie, paléoécologie, paléogéographie.- Thèse, Université de Provence, Travaux du Laboratoire de Géologie Historique et de Paléontologie, Marseille, N° 12, 559 p.
D. & I. (2002).- Early Cretaceous planktonic foraminifera from the Tethys: the genus Leupoldina.- Cretaceous Research, London, Vol. 23, Issue 2, pp. 189-212.
D. & I. (2003a).- Early Cretaceous planktonic foraminifera from the Tethys: the small, few-chambered representatives of the genus Globigerinelloides.- Cretaceous Research, London, Vol. 24, Issue 3, pp. 305-334.
D. & I. (2003b).- Early Cretaceous planktonic foraminifera from the Tethys: the large, many-chambered representatives of the genus Globigerinelloides.- Cretaceous Research, London, Vol. 24, Issue 6, pp. 661-690.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Distribution of Foraminifera in the upper part of the Comte Quarry section (Cassis, SE France).
Click on thumbnail to enlarge the image.
Figure 2: Distribution of Foraminifera in the Les Tocchis section (La Bédoule, SE France).
Click on thumbnail to enlarge the image.
Figure 3: Distribution of Foraminifera in the La Marcouline Quarry section (Cassis, SE France).
Click on thumbnail to enlarge the image.
Figure 4: Stratigraphic range of the principal Aptian markers (benthic and planktonic Foraminifera).
Click on thumbnail to enlarge the image.
Figure 5: Correlation of the three sections (Comte, Tocchis, Marcouline) that permits a reconstitution of the sequence from uppermost Bedoulian to lowermost Gargasian. The boundary between the two substages is fixed at bed 178 (cf. , 1994).
Click on thumbnail to enlarge the image.
Figure 6: Evolution in Aptian times of the mode of coiling and number of chambers in the last whorl in a planktonic foraminiferal lineage with a test initially trochospiral and 6-chambered.
![]()
Click on thumbnail to enlarge the image.
Plate 1:
1-9.
"Globigerinelloides" gr. bizonae () –
saundersi (). 3 specimens showing
various modes of coiling (from trochospiral to subplanispiral). Lowermost
Gargasian, Luterbacheri Zone, Les Tocchis section (La Bédoule), bed 189-T.
10-12. Hastigerinella bizonae, Holotype re-illustrated as it appears under an optical microscope (x 200), 's collection (Muséum National d'Histoire Naturelle, Paris). "lower Gargasian, region of Comte near Cassis", according to the author of the species (1961).
Click on thumbnail to enlarge the image.
Plate 2:
1-12. "Globigerinelloides" bizonae () –
saundersi (). 4 specimens from the
middle Gargasian, Barri Zone, Cassis-La Marcouline section, showing various
stages of dissymmetry in coiling. 1-3. Sample 68. 4-12. Sample 78.
Click on thumbnail to enlarge the image.
Plate 3:
1-4. "Globigerinelloides" bizonae () –
saundersi (). Specimen from the base of
the upper Gargasian, Algerianus Zone, Cassis-La Marcouline section, Sample 90. 4.
Enlargement of the penultimate chamber showing the beginning of the chamber
extension (broken a little above its base).
5-10. Praehedbergella luterbacheri (). Specimens from the lowermost Gargasian, Luterbacheri Zone, Cassis-La Marcouline section, Sample 8.
Click on thumbnail to enlarge the image.
Plate 4:
1-6.
Praehedbergella luterbacheri ().
1-3. lowermost Gargasian, Luterbacheri Zone, Les Tocchis section, bed
183-T. 4-6. Middle Gargasian, Barri Zone, Cassis-La Marcouline section,
Sample 78.
7-9. Transitional specimen between Praehedbergella luterbacheri () and Globigerinelloides ferreolensis (). Lower Gargasian, base of the Ferreolensis Zone, Cassis-La Marcouline section, Sample 16.
10-12. Praehedbergella praetrocoidea ( et ), lower Gargasian, top of the Luterbacheri Zone, Cassis-La Marcouline section, Sample 14.
Click on thumbnail to enlarge the image.
Plate 5:
1-6.
Praehedbergella praetrocoidea (
et ). 1-3. Lower Gargasian,
Ferreolensis Zone, Cassis-La Marcouline section, Sample 22. 4-6. Middle
Gargasian, Barri Zone, Cassis-La Marcouline section, Sample 78.
7-12. Praehedbergella gorbachikae (), base of the upper Gargasian, Algerianus Zone, Cassis-La Marcouline section, Sample 90.
![]()