![]()
![]()
![]()
![]() L'analyse approfondie des associations de Foraminifères depuis les termes du passage Bédoulien-Gargasien jusqu'au Gargasien moyen a été réalisée dans le secteur de Cassis-La Bédoule (SE France), stratotype historique du Bédoulien (Aptien inférieur), particulièrement favorable à ce niveau en raison de la continuité et de la dilatation de la série ainsi que de la diversification de la microfaune. L'éventail des formes benthiques se révèle assez stable mais quelques espèces (Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) s'éteignent à la fin du Bédoulien et peuvent être utilisées pour différencier ce sous-étage par rapport au Gargasien. L'aire stratotypique offre aussi la possibilité de suivre pas à pas l'évolution des formes planctoniques à un moment crucial de leur histoire, lorsque les processus de spéciation et les liens phylogénétiques s'avèrent particulièrement complexes. La grande variabilité morphologique des formes planctoniques durant l'Aptien ne facilite pas le choix de marqueurs stratigraphiques
stables; il est cependant possible de proposer une biozonation comportant cinq zones (à Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) pour l'intervalle considéré, habituellement subdivisé en trois zones. L'étude populationnelle, et non pas seulement typologique, des associations nous a enfin permis d'apporter quelques précisions taxinomiques nouvelles, en particulier sur certaines formes planctoniques d'acception controversée ou même de validité douteuse.
L'analyse approfondie des associations de Foraminifères depuis les termes du passage Bédoulien-Gargasien jusqu'au Gargasien moyen a été réalisée dans le secteur de Cassis-La Bédoule (SE France), stratotype historique du Bédoulien (Aptien inférieur), particulièrement favorable à ce niveau en raison de la continuité et de la dilatation de la série ainsi que de la diversification de la microfaune. L'éventail des formes benthiques se révèle assez stable mais quelques espèces (Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis, Globorotalites bartensteini) s'éteignent à la fin du Bédoulien et peuvent être utilisées pour différencier ce sous-étage par rapport au Gargasien. L'aire stratotypique offre aussi la possibilité de suivre pas à pas l'évolution des formes planctoniques à un moment crucial de leur histoire, lorsque les processus de spéciation et les liens phylogénétiques s'avèrent particulièrement complexes. La grande variabilité morphologique des formes planctoniques durant l'Aptien ne facilite pas le choix de marqueurs stratigraphiques
stables; il est cependant possible de proposer une biozonation comportant cinq zones (à Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) pour l'intervalle considéré, habituellement subdivisé en trois zones. L'étude populationnelle, et non pas seulement typologique, des associations nous a enfin permis d'apporter quelques précisions taxinomiques nouvelles, en particulier sur certaines formes planctoniques d'acception controversée ou même de validité douteuse.
![]() Crétacé inférieur ; Aptien ; stratotype historique ; Foraminifères ; biostratigraphie ; taxinomie ; évolution.
Crétacé inférieur ; Aptien ; stratotype historique ; Foraminifères ; biostratigraphie ; taxinomie ; évolution.
M., G., J.-P. (2005).- Le Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de l'Aptien inférieur, SE France) : associations et biostratigraphie des Foraminifères benthiques et planctoniques.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2005/02 (CG2005_A02)
![]() The Gargasian (Middle Aptian) strata from Cassis-La Bédoule (Lower Aptian historical stratotype, SE
France): planktonic and benthic foraminiferal assemblages and biostratigraphy.- This paper presents a thorough analysis of foraminiferal assemblages ranging in age from the Bedoulian-Gargasian transition to the middle Gargasian in the Cassis-La Bédoule area (SE France), the historical stratotype of the Lower Aptian substage. This region is particularly suitable for detailed studies of Aptian foraminifera owing to rapid and continuous sediment depositional rates and well-diversified microfaunas. The ranges of benthic forms appear to be fairly stable but some species
(Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis,
Globorotalites bartensteini) become extinct at the end of the Bedoulian and thus can be used to separate this substage from the Gargasian. The stratotypic area also offers an opportunity to follow the evolution of planktonic forms step by step at a crucial period of their history, when modalities of speciation and phylogenetic relationships appear to be particularly complex. The important morphologic variability of Aptian planktonic foraminifera does not help finding stable stratigraphic markers; nevertheless, we are able to propose a biozonation comprising five zones (Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) for the interval under consideration, usually subdivided into three zones. Our study of foraminiferal assemblages and species took into account the range of variability among populations and not just the characteristics of a single specimen, such as the holotype. As a result this paper provides new taxonomic precisions on certain planktonic species hitherto controversial or possibly of doubtful validity.
The Gargasian (Middle Aptian) strata from Cassis-La Bédoule (Lower Aptian historical stratotype, SE
France): planktonic and benthic foraminiferal assemblages and biostratigraphy.- This paper presents a thorough analysis of foraminiferal assemblages ranging in age from the Bedoulian-Gargasian transition to the middle Gargasian in the Cassis-La Bédoule area (SE France), the historical stratotype of the Lower Aptian substage. This region is particularly suitable for detailed studies of Aptian foraminifera owing to rapid and continuous sediment depositional rates and well-diversified microfaunas. The ranges of benthic forms appear to be fairly stable but some species
(Praedorothia praeoxycona, Lenticulina cf. nodosa, Astacolus crepidularis,
Globorotalites bartensteini) become extinct at the end of the Bedoulian and thus can be used to separate this substage from the Gargasian. The stratotypic area also offers an opportunity to follow the evolution of planktonic forms step by step at a crucial period of their history, when modalities of speciation and phylogenetic relationships appear to be particularly complex. The important morphologic variability of Aptian planktonic foraminifera does not help finding stable stratigraphic markers; nevertheless, we are able to propose a biozonation comprising five zones (Cabri, Luterbacheri, Ferreolensis, Barri, Algerianus) for the interval under consideration, usually subdivided into three zones. Our study of foraminiferal assemblages and species took into account the range of variability among populations and not just the characteristics of a single specimen, such as the holotype. As a result this paper provides new taxonomic precisions on certain planktonic species hitherto controversial or possibly of doubtful validity.
![]() Early Cretaceous; Aptian; historical stratotype; Foraminifera; biostratigraphy; taxonomy; evolution.
Early Cretaceous; Aptian; historical stratotype; Foraminifera; biostratigraphy; taxonomy; evolution.
![]()
![]() La situation géographique et les relations stratigraphiques des trois coupes (La Bédoule-Les
Tocchis; Cassis-carrière Comte; Cassis-carrière de La Marcouline) du secteur faisant l'objet de la présente étude micropaléontologique ont déjà été abordées dans les deux chapitres introductifs (
et ,
2004 ; et alii,
2004) du Mémoire des Carnets de Géologie consacré au Gargasien-type. Le lecteur trouvera également dans ces deux articles des données relatives à la lithologie et au positionnement des échantillons de la coupe de La Marcouline, ainsi que des informations relatives aux circonstances qui justifient une nouvelle étude de ce qui subsiste de nos jours des coupes-types aptiennes, selon une approche pluridisciplinaire faisant appel aux méthodes de la stratigraphie intégrée.
La situation géographique et les relations stratigraphiques des trois coupes (La Bédoule-Les
Tocchis; Cassis-carrière Comte; Cassis-carrière de La Marcouline) du secteur faisant l'objet de la présente étude micropaléontologique ont déjà été abordées dans les deux chapitres introductifs (
et ,
2004 ; et alii,
2004) du Mémoire des Carnets de Géologie consacré au Gargasien-type. Le lecteur trouvera également dans ces deux articles des données relatives à la lithologie et au positionnement des échantillons de la coupe de La Marcouline, ainsi que des informations relatives aux circonstances qui justifient une nouvelle étude de ce qui subsiste de nos jours des coupes-types aptiennes, selon une approche pluridisciplinaire faisant appel aux méthodes de la stratigraphie intégrée.
![]() Dans cette note, nous nous proposons principalement de compléter l'état des connaissances sur la distribution des Foraminifères benthiques et planctoniques au passage Bédoulien-Gargasien, grâce à l'analyse détaillée des coupes choisies dans l'aire stratotypique de Cassis-La Bédoule, considérée comme particulièrement favorable pour ces niveaux (par
rapport au domaine vocontien et à l'Italie) en raison :
Dans cette note, nous nous proposons principalement de compléter l'état des connaissances sur la distribution des Foraminifères benthiques et planctoniques au passage Bédoulien-Gargasien, grâce à l'analyse détaillée des coupes choisies dans l'aire stratotypique de Cassis-La Bédoule, considérée comme particulièrement favorable pour ces niveaux (par
rapport au domaine vocontien et à l'Italie) en raison :
• d'un taux de sédimentation élevé et de l'absence de hiatus, autorisant un suivi en grand détail de l'évolution des taxons,
• des possibilités de corrélation avec la macrofaune (ammonites en particulier, cf. , 1994 ; et alii, 1998).
![]() Les travaux antérieurs sur les Foraminifères de l'Aptien de Basse-Provence occidentale, et du stratotype du Bédoulien en particulier, ne sont pas très nombreux et, pour la plupart, déjà anciens. En
1961, a décrit quelques espèces nouvelles dont, pour
certaines, le matériel-type provenait de cette région. et alii
(1965) ont fourni la première analyse extensive de la distribution du contenu microfossilifère (foraminifères, ostracodes) du stratotype du Bédoulien, en la corrélant avec celle des ammonites. Dans une étude comparative portant sur les Foraminifères planctoniques de l'Aptien du Mexique et du SE de la France,
(1974) a inclus dans le matériel considéré trois échantillons prélevés par ses soins dans le Bédoulien supérieur de la coupe de Cassis. Dans sa thèse consacrée à la stratigraphie et la sédimentologie de l'Aptien supérieur de Provence,
(1975) a incorporé, à des fins de corrélation biostratigraphique, des données sommaires sur le contenu micropaléontologique provenant, pour le secteur de Cassis, d'un échantillonnage de certaines des coupes que nous avons reprises dans le présent travail.
(1981), dans son mémoire sur les Foraminifères crétacés de Provence, fait souvent référence à des spécimens d'espèces recueillies dans le stratotype de Cassis-La Bédoule, en apportant des précisions sur leur définition paléontologique et leur distribution stratigraphique.
(1987) visite le stratotype du Bédoulien dans le but de préciser, à l'aide des ammonites et des foraminifères, la limite Bédoulien-Gargasien dans le bassin subalpin, proposant des corrélations entre les deux domaines.
Les travaux antérieurs sur les Foraminifères de l'Aptien de Basse-Provence occidentale, et du stratotype du Bédoulien en particulier, ne sont pas très nombreux et, pour la plupart, déjà anciens. En
1961, a décrit quelques espèces nouvelles dont, pour
certaines, le matériel-type provenait de cette région. et alii
(1965) ont fourni la première analyse extensive de la distribution du contenu microfossilifère (foraminifères, ostracodes) du stratotype du Bédoulien, en la corrélant avec celle des ammonites. Dans une étude comparative portant sur les Foraminifères planctoniques de l'Aptien du Mexique et du SE de la France,
(1974) a inclus dans le matériel considéré trois échantillons prélevés par ses soins dans le Bédoulien supérieur de la coupe de Cassis. Dans sa thèse consacrée à la stratigraphie et la sédimentologie de l'Aptien supérieur de Provence,
(1975) a incorporé, à des fins de corrélation biostratigraphique, des données sommaires sur le contenu micropaléontologique provenant, pour le secteur de Cassis, d'un échantillonnage de certaines des coupes que nous avons reprises dans le présent travail.
(1981), dans son mémoire sur les Foraminifères crétacés de Provence, fait souvent référence à des spécimens d'espèces recueillies dans le stratotype de Cassis-La Bédoule, en apportant des précisions sur leur définition paléontologique et leur distribution stratigraphique.
(1987) visite le stratotype du Bédoulien dans le but de préciser, à l'aide des ammonites et des foraminifères, la limite Bédoulien-Gargasien dans le bassin subalpin, proposant des corrélations entre les deux domaines.
![]() La plus récente étude sur les Foraminifères aptiens de cette région ( et alii,
1998) concerne essentiellement les formes du Bédoulien. Pour l'analyse paléontologique et biostratigraphique des niveaux du Gargasien, nous partirons de cette mise au point, en y associant les données obtenues à partir des tous derniers travaux synthétiques portant sur les Foraminifères (essentiellement planctoniques) de l'Aptien téthysien ( et alii,
2002 ; et ,
2002, 2003a, 2003b).
La plus récente étude sur les Foraminifères aptiens de cette région ( et alii,
1998) concerne essentiellement les formes du Bédoulien. Pour l'analyse paléontologique et biostratigraphique des niveaux du Gargasien, nous partirons de cette mise au point, en y associant les données obtenues à partir des tous derniers travaux synthétiques portant sur les Foraminifères (essentiellement planctoniques) de l'Aptien téthysien ( et alii,
2002 ; et ,
2002, 2003a, 2003b).
![]() Les échantillons qui ont servi de base à la présente étude ont des origines diverses. Compte tenu de la dégradation actuelle des lieux, il nous a seulement été possible récemment (de 1997 à 2002) d'échantillonner les termes sommitaux du Bédoulien de la coupe de la carrière Comte et le Gargasien inférieur (mais non basal) et moyen de la coupe de la carrière de La Marcouline (coupe incomplète par le
haut : les termes plus élevés du Gargasien sont absents régionalement suite à une érosion ante-cénomanienne). Ces deux carrières, très proches l'une de l'autre, sont situées sur le territoire de la commune de Cassis.
Les échantillons qui ont servi de base à la présente étude ont des origines diverses. Compte tenu de la dégradation actuelle des lieux, il nous a seulement été possible récemment (de 1997 à 2002) d'échantillonner les termes sommitaux du Bédoulien de la coupe de la carrière Comte et le Gargasien inférieur (mais non basal) et moyen de la coupe de la carrière de La Marcouline (coupe incomplète par le
haut : les termes plus élevés du Gargasien sont absents régionalement suite à une érosion ante-cénomanienne). Ces deux carrières, très proches l'une de l'autre, sont situées sur le territoire de la commune de Cassis.
![]() Nous disposions par ailleurs de témoins et/ou de résidus de lavage provenant d'échantillonnages effectués de 1962 à 1966 par l'un de nous (MM) sur la coupe des Tocchis, située sur le territoire de la commune de La Bédoule (aujourd'hui Roquefort-La Bédoule),
à 2 km à l'ENE de la coupe de La Marcouline. Cette coupe, qui a disparu par suite de l'urbanisation du secteur, avait l'avantage de montrer en quasi continuité les termes de passage du Bédoulien au Gargasien, comblant ainsi le hiatus d'observation entre les deux carrières à Cassis. Ces niveaux, à forte dominante marneuse, ne sont malheureusement plus observables en aucun point de la région de Basse-Provence.
Nous disposions par ailleurs de témoins et/ou de résidus de lavage provenant d'échantillonnages effectués de 1962 à 1966 par l'un de nous (MM) sur la coupe des Tocchis, située sur le territoire de la commune de La Bédoule (aujourd'hui Roquefort-La Bédoule),
à 2 km à l'ENE de la coupe de La Marcouline. Cette coupe, qui a disparu par suite de l'urbanisation du secteur, avait l'avantage de montrer en quasi continuité les termes de passage du Bédoulien au Gargasien, comblant ainsi le hiatus d'observation entre les deux carrières à Cassis. Ces niveaux, à forte dominante marneuse, ne sont malheureusement plus observables en aucun point de la région de Basse-Provence.
![]() Les échantillons calcaréo-marneux du Bédoulien et du Gargasien provençal, relativement compacts, nécessitent d'être lavés avec des adjuvants chimiques, à la fois pour éliminer un maximum de composants non biogéniques, concentrant ainsi la microfaune, et aussi pour nettoyer les microfossiles, assez souvent plus ou moins fortement encroűtés par une gangue argilo-calcaire rendant difficile leur identification.
Les échantillons calcaréo-marneux du Bédoulien et du Gargasien provençal, relativement compacts, nécessitent d'être lavés avec des adjuvants chimiques, à la fois pour éliminer un maximum de composants non biogéniques, concentrant ainsi la microfaune, et aussi pour nettoyer les microfossiles, assez souvent plus ou moins fortement encroűtés par une gangue argilo-calcaire rendant difficile leur identification.
![]() Les échantillons historiquement "anciens" avaient ainsi été préparés selon la méthode d'
(1962), comprenant un trempage à l'hyposulfite de sodium suivi d'un ajout d'eau oxygénée, combinée avec un traitement au Bradosol (mouillant
cationique; voir détails sur la méthode in et alii,
1965 ; , 1966). Ces échantillons ne furent cependant tamisés qu'à 80 µm, maille jugée suffisante pour l'époque. L'état actuel des connaissances, en particulier sur les Foraminifères planctoniques, requiert désormais plutôt un tamisage avec maille minimum de 63, voire même 45 µm.
Les échantillons historiquement "anciens" avaient ainsi été préparés selon la méthode d'
(1962), comprenant un trempage à l'hyposulfite de sodium suivi d'un ajout d'eau oxygénée, combinée avec un traitement au Bradosol (mouillant
cationique; voir détails sur la méthode in et alii,
1965 ; , 1966). Ces échantillons ne furent cependant tamisés qu'à 80 µm, maille jugée suffisante pour l'époque. L'état actuel des connaissances, en particulier sur les Foraminifères planctoniques, requiert désormais plutôt un tamisage avec maille minimum de 63, voire même 45 µm.
![]() Les échantillons prélevés ces dernières années ont fait l'objet, après broyage et dessication à l'étuve, d'un trempage préalable de plusieurs jours dans du Rewoquat W 3690 (tenside cationique), et ont ensuite été tamisés (à 45 µm) à l'eau chaude, avec rinçage final à l'eau oxygénée. Le processus est assez lourd mais a conduit à d'excellents résultats, aussi bien pour ce qui est de la concentration que du dégagement des microfaunes.
Les échantillons prélevés ces dernières années ont fait l'objet, après broyage et dessication à l'étuve, d'un trempage préalable de plusieurs jours dans du Rewoquat W 3690 (tenside cationique), et ont ensuite été tamisés (à 45 µm) à l'eau chaude, avec rinçage final à l'eau oxygénée. Le processus est assez lourd mais a conduit à d'excellents résultats, aussi bien pour ce qui est de la concentration que du dégagement des microfaunes.
![]() Selon les niveaux, l'état de conservation des microfossiles aptiens s'est révélé variable, allant de moyen ou même assez bon (niveaux plus marneux du Gargasien moyen) à médiocre, voire mauvais (niveaux plus indurés du Bédoulien terminal, pas forcément plus calcaires mais surtout plus siliceux, comme en témoigne la présence plus marquée de spicules, radiolaires et diatomées dans les résidus de lavage). Il faut aussi noter la présence assez constante, en proportion non négligeable, de tests plus ou moins déformés, sans doute sous l'effet d'une importante compaction, en particulier chez les formes planctoniques à test mince (par
exemple, petits Globigerinelloides
[ex-Blowiella], petites Praehedbergelles) et chez des benthiques agglutinants comme les Trochamminidae, Reophax ou Recurvoides.
Selon les niveaux, l'état de conservation des microfossiles aptiens s'est révélé variable, allant de moyen ou même assez bon (niveaux plus marneux du Gargasien moyen) à médiocre, voire mauvais (niveaux plus indurés du Bédoulien terminal, pas forcément plus calcaires mais surtout plus siliceux, comme en témoigne la présence plus marquée de spicules, radiolaires et diatomées dans les résidus de lavage). Il faut aussi noter la présence assez constante, en proportion non négligeable, de tests plus ou moins déformés, sans doute sous l'effet d'une importante compaction, en particulier chez les formes planctoniques à test mince (par
exemple, petits Globigerinelloides
[ex-Blowiella], petites Praehedbergelles) et chez des benthiques agglutinants comme les Trochamminidae, Reophax ou Recurvoides.
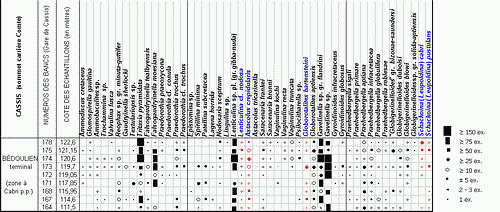
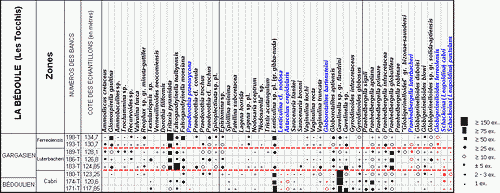
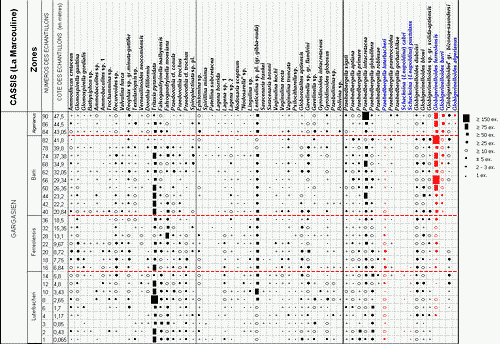
![]() Le contenu de tous les échantillons étudiés (9 pour le sommet de la carrière Comte, 8 aux Tocchis et 24 pour la carrière de La Marcouline) a été considéré sous l'aspect quantitatif, avec comptage systématique des spécimens de chaque espèce de Foraminifères, conduisant à l'établissement de classes de fréquence selon la gradation
suivante :
Le contenu de tous les échantillons étudiés (9 pour le sommet de la carrière Comte, 8 aux Tocchis et 24 pour la carrière de La Marcouline) a été considéré sous l'aspect quantitatif, avec comptage systématique des spécimens de chaque espèce de Foraminifères, conduisant à l'établissement de classes de fréquence selon la gradation
suivante :
| catégorie | RR | R | R-F | F | F-C | C | AB | V.AB |
| nb. spécimens | 1 | 2-3 | ±5 | >10 | >20 | >50 | >75 | >150 |
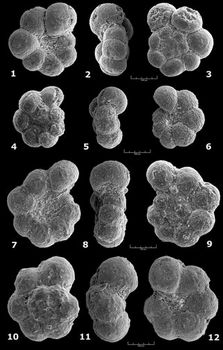
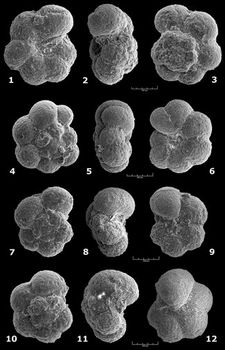
![]() Ces fréquences de représentation sont indiquées pour chaque espèce et chaque échantillon, selon un figuré conventionnel, dans les tableaux de répartition des
Figs. 1
Ces fréquences de représentation sont indiquées pour chaque espèce et chaque échantillon, selon un figuré conventionnel, dans les tableaux de répartition des
Figs. 1 ![]() - 2
- 2 ![]() - 3
- 3
![]() .
.
![]() L'ensemble du matériel ayant fait l'objet de cette étude (témoins d'échantillons, résidus de lavage, cellules) est déposé dans les collections du Musée de Paléontologie de l'Université de Provence à Marseille.
L'ensemble du matériel ayant fait l'objet de cette étude (témoins d'échantillons, résidus de lavage, cellules) est déposé dans les collections du Musée de Paléontologie de l'Université de Provence à Marseille.
• Bédoulien supérieur : Tritaxia pyramidata, Falsogaudryinelles, Lenticulines "lisses" (c'est-à-dire non ornementées : L. gr. gibba-nuda), Gavelinelles.
• Gargasien : Tritaxia pyramidata demeure dominante (avec quelques irrégularités), les Falsogaudryinelles et Lenticulines le sont un peu moins qu'au Bédoulien, les Gavelinelles beaucoup moins, en revanche les formes planctoniques y sont plus abondantes.
![]() On dénote un accroissement du nombre d'espèces en montant dans la série. Pris globalement, le Bédoulien-type comprend 31 espèces de Foraminifères benthiques (dont 14 à test agglutinant, 17 à test calcaire) et 11 espèces de Foraminifères planctoniques, soit en tout 42 espèces de Foraminifères. Le Gargasien (inférieur-moyen) du même secteur comprend 45 espèces benthiques (20 agglutinants et 25 calcaires), plus 21 espèces planctoniques, soit 66 espèces de Foraminifères; ces données sont résumées dans le tableau synthétique
ci-dessous :
On dénote un accroissement du nombre d'espèces en montant dans la série. Pris globalement, le Bédoulien-type comprend 31 espèces de Foraminifères benthiques (dont 14 à test agglutinant, 17 à test calcaire) et 11 espèces de Foraminifères planctoniques, soit en tout 42 espèces de Foraminifères. Le Gargasien (inférieur-moyen) du même secteur comprend 45 espèces benthiques (20 agglutinants et 25 calcaires), plus 21 espèces planctoniques, soit 66 espèces de Foraminifères; ces données sont résumées dans le tableau synthétique
ci-dessous :
| Nombre d'espèces | Agglut. | Calc. | Planct. | Total |
| GARGASIEN | 20 | 25 | 21 | 66 |
| BÉDOULIEN | 14 | 17 | 11 | 42 |
![]() La longévité de la grande majorité des espèces, aussi bien benthiques que, curieusement, planctoniques, est assez importante. Nombreuses sont celles qui s'étendent sur une grande partie de l'Aptien. L'accroissement relatif de la diversité à l'Aptien supérieur résulte à la fois d'un rythme très modéré d'extinctions et d'un enrichissement progressif, régulier, par apparition de nouvelles espèces.
La longévité de la grande majorité des espèces, aussi bien benthiques que, curieusement, planctoniques, est assez importante. Nombreuses sont celles qui s'étendent sur une grande partie de l'Aptien. L'accroissement relatif de la diversité à l'Aptien supérieur résulte à la fois d'un rythme très modéré d'extinctions et d'un enrichissement progressif, régulier, par apparition de nouvelles espèces.
![]() Au sein de l'Aptien de Cassis-La Bédoule, nos données montrent que le nombre des marqueurs potentiels est assez faible, aussi bien chez les formes benthiques que planctoniques
(Fig. 4
Au sein de l'Aptien de Cassis-La Bédoule, nos données montrent que le nombre des marqueurs potentiels est assez faible, aussi bien chez les formes benthiques que planctoniques
(Fig. 4 ![]() ).
).
![]() On remarque chez ce groupe quelques espèces, souvent connues comme apparaissant bien avant l'Aptien dans la Téthys, qui s'éteignent à la fin du Bédoulien ou ne dépassent guère la limite Bédoulien-Gargasien
:
On remarque chez ce groupe quelques espèces, souvent connues comme apparaissant bien avant l'Aptien dans la Téthys, qui s'éteignent à la fin du Bédoulien ou ne dépassent guère la limite Bédoulien-Gargasien
:
• Praedorothia praeoxycona
• Lenticulina cf. nodosa
• Astacolus crepidularis
• Globorotalites bartensteini
![]() Les formes benthiques qui apparaissent au cours du Gargasien ne paraissent pas très significatives stratigraphiquement. Il s'agit de formes au test agglutinant
(Bathysiphon, Ammobaculites, Trochammina, Recurvoides, Praedorothia cf. conula) dont la présence à ce niveau peut tout aussi bien être liée à une modification du milieu par rapport aux termes sous-jacents.
Les formes benthiques qui apparaissent au cours du Gargasien ne paraissent pas très significatives stratigraphiquement. Il s'agit de formes au test agglutinant
(Bathysiphon, Ammobaculites, Trochammina, Recurvoides, Praedorothia cf. conula) dont la présence à ce niveau peut tout aussi bien être liée à une modification du milieu par rapport aux termes sous-jacents.
![]() Aucune espèce planctonique existant déjà au Bédoulien ne disparaît avec ce sous-étage. Schackoina gr. cabri (= S. cabri s.s. + S. pustulans), apparue au cours du Bédoulien supérieur, constitue la seule espèce planctonique qui disparaisse assez rapidement, dans les termes inférieurs du Gargasien. Deux nouvelles formes apparaissent quasi-simultanément un peu au dessus de la limite Bédoulien-Gargasien : Praehedbergella luterbacheri, puis Globigerinelloides ferreolensis. Ces deux taxons, sporadiquement représentés au tout début de leur répartition, coexistent quelque temps avec les dernières Schackoïnes, devenues elles-mêmes très rares en ces niveaux. Ensuite, à partir du début de son acmé, G. ferreolensis devient un constituant important de la microfaune de foraminifères planctoniques du Gargasien
moyen / supérieur.
Aucune espèce planctonique existant déjà au Bédoulien ne disparaît avec ce sous-étage. Schackoina gr. cabri (= S. cabri s.s. + S. pustulans), apparue au cours du Bédoulien supérieur, constitue la seule espèce planctonique qui disparaisse assez rapidement, dans les termes inférieurs du Gargasien. Deux nouvelles formes apparaissent quasi-simultanément un peu au dessus de la limite Bédoulien-Gargasien : Praehedbergella luterbacheri, puis Globigerinelloides ferreolensis. Ces deux taxons, sporadiquement représentés au tout début de leur répartition, coexistent quelque temps avec les dernières Schackoïnes, devenues elles-mêmes très rares en ces niveaux. Ensuite, à partir du début de son acmé, G. ferreolensis devient un constituant important de la microfaune de foraminifères planctoniques du Gargasien
moyen / supérieur.
![]() Le reste de la coupe gargasienne de La Marcouline est marqué par les apparitions successives de G. barri puis, vingt mètres au-dessus, de G. algerianus, pour les formes planctoniques les plus significatives. Ces deux espèces, par leur large distribution géographique et la fiabilité de leur moment d'apparition, sont connues comme d'importants marqueurs téthysiens. Quelques autres espèces, trop sporadiques cependant pour constituer d'utiles marqueurs, apparaissent aussi dans ces
niveaux : Praehedbergella praetrocoidea, puis Praehedbergella
gorbatchikae.
Le reste de la coupe gargasienne de La Marcouline est marqué par les apparitions successives de G. barri puis, vingt mètres au-dessus, de G. algerianus, pour les formes planctoniques les plus significatives. Ces deux espèces, par leur large distribution géographique et la fiabilité de leur moment d'apparition, sont connues comme d'importants marqueurs téthysiens. Quelques autres espèces, trop sporadiques cependant pour constituer d'utiles marqueurs, apparaissent aussi dans ces
niveaux : Praehedbergella praetrocoidea, puis Praehedbergella
gorbatchikae.
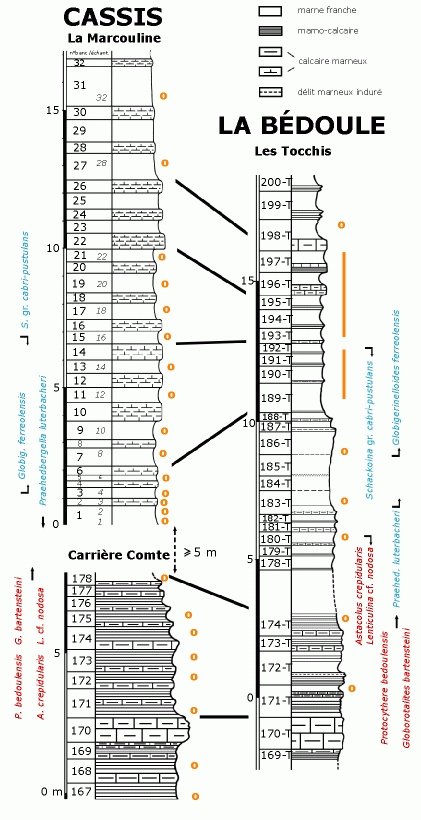
![]() L'inventaire détaillé du contenu micropaléontologique des trois coupes précitées conduit tout d'abord à affiner leur corrélation initialement fondée sur des considérations géométriques
(Fig. 5
L'inventaire détaillé du contenu micropaléontologique des trois coupes précitées conduit tout d'abord à affiner leur corrélation initialement fondée sur des considérations géométriques
(Fig. 5 ![]() ).
).
![]() La très petite distance (2 km) entre les deux secteurs (Les Tocchis et La
Marcouline) permet d'éliminer l'éventualité d'un diachronisme entre les
moments d'apparition des formes planctoniques, tel qu'il a pu être constaté
récemment ( et alii,
2004) dans le Cénomano-Turonien du Western
Interior mais sur des distances beaucoup plus considérables (600 km). Se voit ainsi particulièrement confortée l'homologie suggérée ( et alii,
2004) entre le "triplet" constitué par les bancs 22-24-26 à La Marcouline (Cassis) et les bancs 196-T, 197-T, 198-T aux Tocchis (La Bédoule). Sur la base de la dernière occurrence des Schackoïnes et de la première occurrence de
Globigerinelloides ferreolensis, nous proposons en outre la corrélation du niveau 14 (Marcouline) avec le niveau 193-T (Tocchis), et celle du niveau 6 (Marcouline) avec le niveau 188-T (Tocchis). Il en ressort qu'à La Marcouline la série se revèle constamment plus dilatée qu'aux Tocchis. Pour les niveaux sous-jacents, en dehors de l'homologie géométrique indiscutable du banc 170 (Cassis-carrière Comte) avec le banc 170-T (La Bédoule-Les Tocchis), la corrélation de détail est plus délicate, en particulier à cause de la présence d'une lacune d'observation (non mesurable directement) à Cassis, correspondant à une zone marneuse qui était représentée par la séquence 178-T à 186-T à La Bédoule. Plusieurs datums (LO
d'Astacolus crepidularis et de Lenticulina cf. nodosa, FO de Praehedbergella luterbacheri), bien positionnés aux Tocchis, ne peuvent ainsi être localisés à Cassis, car ils se produisent très vraisemblablement au sein de la zone non affleurante. Sur la base de la coupe des Tocchis, on peut cependant estimer l'épaisseur de cette lacune de visibilité à Cassis à au moins 5 mètres, peut-être un peu plus si l'on tient compte de la dilatation plus forte de la série évoquée ci-dessus. Il en résulte qu'au-delà du banc repère 170, il n'y a pas homologie directe banc à banc entre les deux coupes, ce qui implique de différencier la numérotation des bancs des Tocchis par l'adjonction du suffixe "-T".
La très petite distance (2 km) entre les deux secteurs (Les Tocchis et La
Marcouline) permet d'éliminer l'éventualité d'un diachronisme entre les
moments d'apparition des formes planctoniques, tel qu'il a pu être constaté
récemment ( et alii,
2004) dans le Cénomano-Turonien du Western
Interior mais sur des distances beaucoup plus considérables (600 km). Se voit ainsi particulièrement confortée l'homologie suggérée ( et alii,
2004) entre le "triplet" constitué par les bancs 22-24-26 à La Marcouline (Cassis) et les bancs 196-T, 197-T, 198-T aux Tocchis (La Bédoule). Sur la base de la dernière occurrence des Schackoïnes et de la première occurrence de
Globigerinelloides ferreolensis, nous proposons en outre la corrélation du niveau 14 (Marcouline) avec le niveau 193-T (Tocchis), et celle du niveau 6 (Marcouline) avec le niveau 188-T (Tocchis). Il en ressort qu'à La Marcouline la série se revèle constamment plus dilatée qu'aux Tocchis. Pour les niveaux sous-jacents, en dehors de l'homologie géométrique indiscutable du banc 170 (Cassis-carrière Comte) avec le banc 170-T (La Bédoule-Les Tocchis), la corrélation de détail est plus délicate, en particulier à cause de la présence d'une lacune d'observation (non mesurable directement) à Cassis, correspondant à une zone marneuse qui était représentée par la séquence 178-T à 186-T à La Bédoule. Plusieurs datums (LO
d'Astacolus crepidularis et de Lenticulina cf. nodosa, FO de Praehedbergella luterbacheri), bien positionnés aux Tocchis, ne peuvent ainsi être localisés à Cassis, car ils se produisent très vraisemblablement au sein de la zone non affleurante. Sur la base de la coupe des Tocchis, on peut cependant estimer l'épaisseur de cette lacune de visibilité à Cassis à au moins 5 mètres, peut-être un peu plus si l'on tient compte de la dilatation plus forte de la série évoquée ci-dessus. Il en résulte qu'au-delà du banc repère 170, il n'y a pas homologie directe banc à banc entre les deux coupes, ce qui implique de différencier la numérotation des bancs des Tocchis par l'adjonction du suffixe "-T".
![]() Les précisions qu'apporte notre analyse de la répartition stratigraphique des principaux marqueurs planctoniques dans l'Aptien de l'aire stratotypique du Bédoulien nous permettent d'affiner quelque peu la zonation publiée in et alii
(1998) ainsi que, d'une manière plus générale, la zonation du Gargasien inférieur-moyen téthysien (
et ,
1995 ; et alii,
2002), en subdivisant cet intervalle en cinq zones au
lieu de trois :
Les précisions qu'apporte notre analyse de la répartition stratigraphique des principaux marqueurs planctoniques dans l'Aptien de l'aire stratotypique du Bédoulien nous permettent d'affiner quelque peu la zonation publiée in et alii
(1998) ainsi que, d'une manière plus générale, la zonation du Gargasien inférieur-moyen téthysien (
et ,
1995 ; et alii,
2002), en subdivisant cet intervalle en cinq zones au
lieu de trois :
|
Ce travail |
|
| Algerianus | Algerianus |
| Ferreolensis | Barri |
| Ferreolensis | |
| Cabri | Luterbacheri |
| Cabri |
![]() Dans la zonation précédente, il se posait en effet le problème de la définition de la limite entre les zones à Cabri et à Ferreolensis. Nos résultats, confirmant ceux de
(1974) au Mexique et ceux de
et
(1988) en Angleterre, montrent qu'au lieu de s'exclure, comme il a souvent été admis, les deux marqueurs coexistent fugacement. Cette concomitance se produit sur une épaisseur de terrains d'au moins 5 mètres à La Marcouline, un peu moindre aux Tocchis. Mais dans cet intervalle Schackoina gr. cabri devient très rare, et les premières
Globigerinelloides ferreolensis sont sporadiques. Il n'est en outre pas aisé de délimiter exactement la première apparition de
G. ferreolensis. Nos observations montrent, comme le pressentaient
et
(1988), que cette espèce dérive d'une forme initialement trochospiralée, par aplatissement et symétrisation progressive de la trochospire. L'espèce-souche de G. ferreolensis est ainsi représentée par Praehedbergella
luterbacheri; le processus de symétrisation, graduel mais transitoire, n'est observable que sur du matériel bien conservé, aux ombilics bien dégagés et nettoyés, et ne concerne qu'un nombre réduit de spécimens au sein d'une population. Il peut souvent passer inaperçu, en fonction des conditions de fossilisation et de dégagement de la microfaune. En observation courante,
Globigerinelloides ferreolensis ne devient donc typique, plus aisément identifiable et plus fréquente qu'à partir du niveau d'extinction de
Schackoina gr. cabri. Pour cette raison nous avons choisi d'utiliser le début de l'acmé (plutôt que le FAD) de
Globigerinelloides ferreolensis comme limite inférieure de la zone éponyme.
Dans la zonation précédente, il se posait en effet le problème de la définition de la limite entre les zones à Cabri et à Ferreolensis. Nos résultats, confirmant ceux de
(1974) au Mexique et ceux de
et
(1988) en Angleterre, montrent qu'au lieu de s'exclure, comme il a souvent été admis, les deux marqueurs coexistent fugacement. Cette concomitance se produit sur une épaisseur de terrains d'au moins 5 mètres à La Marcouline, un peu moindre aux Tocchis. Mais dans cet intervalle Schackoina gr. cabri devient très rare, et les premières
Globigerinelloides ferreolensis sont sporadiques. Il n'est en outre pas aisé de délimiter exactement la première apparition de
G. ferreolensis. Nos observations montrent, comme le pressentaient
et
(1988), que cette espèce dérive d'une forme initialement trochospiralée, par aplatissement et symétrisation progressive de la trochospire. L'espèce-souche de G. ferreolensis est ainsi représentée par Praehedbergella
luterbacheri; le processus de symétrisation, graduel mais transitoire, n'est observable que sur du matériel bien conservé, aux ombilics bien dégagés et nettoyés, et ne concerne qu'un nombre réduit de spécimens au sein d'une population. Il peut souvent passer inaperçu, en fonction des conditions de fossilisation et de dégagement de la microfaune. En observation courante,
Globigerinelloides ferreolensis ne devient donc typique, plus aisément identifiable et plus fréquente qu'à partir du niveau d'extinction de
Schackoina gr. cabri. Pour cette raison nous avons choisi d'utiliser le début de l'acmé (plutôt que le FAD) de
Globigerinelloides ferreolensis comme limite inférieure de la zone éponyme.
![]() Le FAD de l'ancêtre de G. ferreolensis nous permet de proposer l'instauration d'une nouvelle zone, à Luterbacheri, correspondant à la partie sommitale de l'ancienne zone à Cabri, pour l'intervalle de temps compris entre le FAD de Praehedbergella luterbacheri et le début de l'acmé de
Globigerinelloides
ferreolensis.
Le FAD de l'ancêtre de G. ferreolensis nous permet de proposer l'instauration d'une nouvelle zone, à Luterbacheri, correspondant à la partie sommitale de l'ancienne zone à Cabri, pour l'intervalle de temps compris entre le FAD de Praehedbergella luterbacheri et le début de l'acmé de
Globigerinelloides
ferreolensis.
![]() La partie supérieure de l'ancienne zone à Ferreolensis peut de même être individualisée sous la forme d'une zone à Barri, dont la limite inférieure est définie par le FAD de
G. barri, successeur phylogénétique de G.
ferreolensis. Le concept d'une zone à Barri, quoique fondé sur une acception un peu différente de l'espèce marqueur, fut déjà proposé par en
1959.
La partie supérieure de l'ancienne zone à Ferreolensis peut de même être individualisée sous la forme d'une zone à Barri, dont la limite inférieure est définie par le FAD de
G. barri, successeur phylogénétique de G.
ferreolensis. Le concept d'une zone à Barri, quoique fondé sur une acception un peu différente de l'espèce marqueur, fut déjà proposé par en
1959.
![]() Dans les tout derniers mètres de la coupe de La Marcouline apparaissent les premiers représentants du successeur phylogénétique de G. barri, soit G. algerianus, dont le FAD définit la limite inférieure de la zone éponyme, largement utilisée depuis nombre d'années dans le domaine téthysien.
Dans les tout derniers mètres de la coupe de La Marcouline apparaissent les premiers représentants du successeur phylogénétique de G. barri, soit G. algerianus, dont le FAD définit la limite inférieure de la zone éponyme, largement utilisée depuis nombre d'années dans le domaine téthysien.
![]() Les propositions qui précèdent sont intégrées dans la
Fig. 4
Les propositions qui précèdent sont intégrées dans la
Fig. 4 ![]() , indiquant la répartition stratigraphique des principaux marqueurs.
, indiquant la répartition stratigraphique des principaux marqueurs.
![]() Considéré dans sa globalité, l'Aptien apparaît comme la première période significative de radiation chez les Foraminifères planctoniques ( et ,
1983 ; et alii,
1984 ; et alii,
2002). Combinés aux données obtenues sur le Bédoulien-type ( et alii,
1998), les résultats ici présentés sur le Gargasien permettent de préciser quelque peu les modalités de ce processus. Au niveau de détail permis par la dilatation des coupes de l'aire stratotypique, on constate que la spéciation aptienne, initiée au cours du Bédoulien supérieur (un peu avant l'événement anoxique AOE1a, cf. et alii,
1998), s'effectue plus par un enrichissement graduel du nombre d'espèces que par un processus brutal et explosif. Les termes de passage du Bédoulien au Gargasien correspondent à une brève période de stabilité, ce qui rend d'ailleurs difficile leur découpage et la caractérisation, au moyen des Foraminifères planctoniques, de la limite entre ces deux sous-étages, au demeurant mieux marquée par les foraminifères benthiques
(Fig. 4
Considéré dans sa globalité, l'Aptien apparaît comme la première période significative de radiation chez les Foraminifères planctoniques ( et ,
1983 ; et alii,
1984 ; et alii,
2002). Combinés aux données obtenues sur le Bédoulien-type ( et alii,
1998), les résultats ici présentés sur le Gargasien permettent de préciser quelque peu les modalités de ce processus. Au niveau de détail permis par la dilatation des coupes de l'aire stratotypique, on constate que la spéciation aptienne, initiée au cours du Bédoulien supérieur (un peu avant l'événement anoxique AOE1a, cf. et alii,
1998), s'effectue plus par un enrichissement graduel du nombre d'espèces que par un processus brutal et explosif. Les termes de passage du Bédoulien au Gargasien correspondent à une brève période de stabilité, ce qui rend d'ailleurs difficile leur découpage et la caractérisation, au moyen des Foraminifères planctoniques, de la limite entre ces deux sous-étages, au demeurant mieux marquée par les foraminifères benthiques
(Fig. 4 ![]() ) et les ostracodes (cf. et , ce volume).
) et les ostracodes (cf. et , ce volume).
![]() Le Gargasien inférieur-moyen correspond à une reprise ménagée, selon un rythme progressif et régulier, de la spéciation chez les formes planctoniques, dont la diversité continue à s'accroître puisque la majeure partie des formes apparues antérieurement est toujours présente à ce niveau.
Le Gargasien inférieur-moyen correspond à une reprise ménagée, selon un rythme progressif et régulier, de la spéciation chez les formes planctoniques, dont la diversité continue à s'accroître puisque la majeure partie des formes apparues antérieurement est toujours présente à ce niveau.
![]() Il apparaît donc que la "radiation" intra-aptienne n'est pas un phénomène soudain et brutal, mais plutôt progressif, et qu'elle peut se décomposer en deux phases anagénétiques : 1) partie inférieure/moyenne du Bédoulien supérieur et 2) Gargasien (extrême base exclue), séparées par un bref stade stasigénétique chevauchant la limite entre Bédoulien et Gargasien. Sur la période considérée dans cet article, elle ne concerne que des formes planctoniques à test microperforé (Schackoïnes) et finement perforé
(Praehedbergella, Globigerinelloides), la première apparition d'une espèce à test macroperforé
(Hedbergella trocoidea) (cf. et alii,
1998) ne s'effectuant qu'au Gargasien supérieur.
Il apparaît donc que la "radiation" intra-aptienne n'est pas un phénomène soudain et brutal, mais plutôt progressif, et qu'elle peut se décomposer en deux phases anagénétiques : 1) partie inférieure/moyenne du Bédoulien supérieur et 2) Gargasien (extrême base exclue), séparées par un bref stade stasigénétique chevauchant la limite entre Bédoulien et Gargasien. Sur la période considérée dans cet article, elle ne concerne que des formes planctoniques à test microperforé (Schackoïnes) et finement perforé
(Praehedbergella, Globigerinelloides), la première apparition d'une espèce à test macroperforé
(Hedbergella trocoidea) (cf. et alii,
1998) ne s'effectuant qu'au Gargasien supérieur.
![]() Dans l'état actuel de nos connaissances, il n'apparaît pas que l'on puisse faire coïncider ces processus biologiques avec les données de la géochimie et de la sédimentologie, rendant ainsi hypothétique une relation directe de causalité entre spéciation et facteurs externes chez les Foraminifères planctoniques.
Dans l'état actuel de nos connaissances, il n'apparaît pas que l'on puisse faire coïncider ces processus biologiques avec les données de la géochimie et de la sédimentologie, rendant ainsi hypothétique une relation directe de causalité entre spéciation et facteurs externes chez les Foraminifères planctoniques.
![]() Durant la période incluant le Bédoulien élevé et le Gargasien inf.-moyen, les Foraminifères planctoniques sont affectés à certains moments par une augmentation de l'instabilité de leurs caractères, ainsi en particulier certaines Praehedbergelles, dans une moindre mesure certains représentants du genre Globigerinelloides ("small few-chambered Globigerinelloides" in
et ,
2003a) et également les formes à loges digitiformes. Ces séquences de formes à caractères fluctuants que l'on peut aisément observer à Cassis-La Bédoule ne sont qu'imparfaitement ou incomplètement enregistrées partout ailleurs dans le monde, ce qui a conduit à des erreurs d'interprétation sur le plan phylogénétique, et donc systématique.
Durant la période incluant le Bédoulien élevé et le Gargasien inf.-moyen, les Foraminifères planctoniques sont affectés à certains moments par une augmentation de l'instabilité de leurs caractères, ainsi en particulier certaines Praehedbergelles, dans une moindre mesure certains représentants du genre Globigerinelloides ("small few-chambered Globigerinelloides" in
et ,
2003a) et également les formes à loges digitiformes. Ces séquences de formes à caractères fluctuants que l'on peut aisément observer à Cassis-La Bédoule ne sont qu'imparfaitement ou incomplètement enregistrées partout ailleurs dans le monde, ce qui a conduit à des erreurs d'interprétation sur le plan phylogénétique, et donc systématique.
![]() L'étude détaillée d'un échantillonnage assez dense portant de plus sur une coupe composite dilatée nous a en particulier permis de suivre, pour ainsi dire pas à pas, l'évolution des Foraminifères planctoniques au niveau de la transition du Bédoulien au Gargasien. Au plan des caractères morphologiques, pour chaque lignée, cette évolution révèle en première analyse une succession de stades de relative stabilité apparente alternant avec de brèves séquences anagénétiques. Chez ces formes il existe de toute manière une difficulté permanente de caractérisation des taxons par des critères stables. On assiste en fait à un glissement évolutif constant, au rythme plus ou moins rapide, affectant aussi bien la nature de l'enroulement, la taille du test, le nombre et la forme des loges, leur rugosité,
etc., qui conduit à relativiser la notion d'espèce (et encore plus de genre) au sein d'un continuum en perpétuelle dérive.
L'étude détaillée d'un échantillonnage assez dense portant de plus sur une coupe composite dilatée nous a en particulier permis de suivre, pour ainsi dire pas à pas, l'évolution des Foraminifères planctoniques au niveau de la transition du Bédoulien au Gargasien. Au plan des caractères morphologiques, pour chaque lignée, cette évolution révèle en première analyse une succession de stades de relative stabilité apparente alternant avec de brèves séquences anagénétiques. Chez ces formes il existe de toute manière une difficulté permanente de caractérisation des taxons par des critères stables. On assiste en fait à un glissement évolutif constant, au rythme plus ou moins rapide, affectant aussi bien la nature de l'enroulement, la taille du test, le nombre et la forme des loges, leur rugosité,
etc., qui conduit à relativiser la notion d'espèce (et encore plus de genre) au sein d'un continuum en perpétuelle dérive.
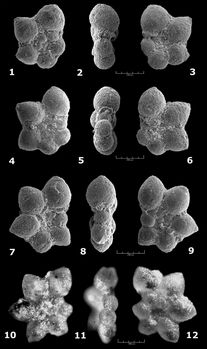
![]() Un exemple (Fig. 6
Un exemple (Fig. 6 ![]() ) que nous avons pu particulièrement bien documenter dans le secteur de Cassis-La Bédoule concerne au départ des formes initialement nettement trochospiralées, à 6 loges au dernier tour, croissant modérément en taille, donnant un contour très peu lobé en vue latérale, qui correspondent au taxon
Praehedbergella primare (cf. appendice taxinomique). Cette espèce à assez grande longévité apparaît au Barrémien terminal (,
1966) et demeure tout d'abord morphologiquement inchangée (phase stasigénétique) jusque dans la partie moyenne du Bédoulien supérieur où, en raison de l'acquisition progressive d'un contour nettement plus découpé, certains spécimens en ont été taxinomiquement isolés (ainsi ,
1974, sous le nom d'Hedbergella similis, synonyme plus récent de Praehedbergella globulifera ( et ),
in , 1971). Ces deux taxons
(P. primare et P. globulifera), qu'il n'est pas toujours aisé de délimiter franchement l'un de l'autre au moment de la phase de spéciation, vont ensuite coexister durant la majeure partie du
Gargasien; mais dès la base du sous-étage (exactement dans la zone où coexistent brièvement Schackoina cabri et Globigerinelloides ferreolensis), on discerne une autre forme d'évolution rapide au sein des populations de Praehedbergelles à 6 loges. Les formes stables, typiques des deux taxons précités y sont devenues très minoritaires, et la majeure partie des spécimens, très polymorphes, montrent une tendance de plus en plus marquée à évoluer de la trochospiralité vers la planispiralité (faces ombilicale et spirale de moins en moins dissemblables, ouverture de plus en plus périphérique); presque simultanément, via des stades intermédiaires, on observe un passage de 6 loges vers 7 loges au dernier tour. Durant cette courte phase, il devient très difficile d'identifier et de caractériser des "espèces" (et même des genres, fondés sur la nature - trochospiralée vs. planispiralée - de l'enroulement), selon les normes habituelles de la nomenclature linnéenne. Les deux stades terminaux, stabilisés, de cette évolution correspondent à deux taxons décrits dans la littérature respectivement sous les noms de Praehedbergella luterbacheri, forme à trochospire très plane, typiquement à 7 loges, et de
Globigerinelloides ferreolensis, forme planispiralée à 7 puis, très vite, 8 loges. Malheureusement certains auteurs, par une approche strictement typologique ne prenant pas en compte ces phénomènes de glissement évolutif, se sont évertués à créer des noms d'espèces distincts pour nombre des stades intermédiaires et transitoires, pris au hasard au sein de ces populations mouvantes et multiformes. Nous avons pour notre part choisi de n'accorder une transcription nomenclaturale qu'aux morphotypes stabilisés.
) que nous avons pu particulièrement bien documenter dans le secteur de Cassis-La Bédoule concerne au départ des formes initialement nettement trochospiralées, à 6 loges au dernier tour, croissant modérément en taille, donnant un contour très peu lobé en vue latérale, qui correspondent au taxon
Praehedbergella primare (cf. appendice taxinomique). Cette espèce à assez grande longévité apparaît au Barrémien terminal (,
1966) et demeure tout d'abord morphologiquement inchangée (phase stasigénétique) jusque dans la partie moyenne du Bédoulien supérieur où, en raison de l'acquisition progressive d'un contour nettement plus découpé, certains spécimens en ont été taxinomiquement isolés (ainsi ,
1974, sous le nom d'Hedbergella similis, synonyme plus récent de Praehedbergella globulifera ( et ),
in , 1971). Ces deux taxons
(P. primare et P. globulifera), qu'il n'est pas toujours aisé de délimiter franchement l'un de l'autre au moment de la phase de spéciation, vont ensuite coexister durant la majeure partie du
Gargasien; mais dès la base du sous-étage (exactement dans la zone où coexistent brièvement Schackoina cabri et Globigerinelloides ferreolensis), on discerne une autre forme d'évolution rapide au sein des populations de Praehedbergelles à 6 loges. Les formes stables, typiques des deux taxons précités y sont devenues très minoritaires, et la majeure partie des spécimens, très polymorphes, montrent une tendance de plus en plus marquée à évoluer de la trochospiralité vers la planispiralité (faces ombilicale et spirale de moins en moins dissemblables, ouverture de plus en plus périphérique); presque simultanément, via des stades intermédiaires, on observe un passage de 6 loges vers 7 loges au dernier tour. Durant cette courte phase, il devient très difficile d'identifier et de caractériser des "espèces" (et même des genres, fondés sur la nature - trochospiralée vs. planispiralée - de l'enroulement), selon les normes habituelles de la nomenclature linnéenne. Les deux stades terminaux, stabilisés, de cette évolution correspondent à deux taxons décrits dans la littérature respectivement sous les noms de Praehedbergella luterbacheri, forme à trochospire très plane, typiquement à 7 loges, et de
Globigerinelloides ferreolensis, forme planispiralée à 7 puis, très vite, 8 loges. Malheureusement certains auteurs, par une approche strictement typologique ne prenant pas en compte ces phénomènes de glissement évolutif, se sont évertués à créer des noms d'espèces distincts pour nombre des stades intermédiaires et transitoires, pris au hasard au sein de ces populations mouvantes et multiformes. Nous avons pour notre part choisi de n'accorder une transcription nomenclaturale qu'aux morphotypes stabilisés.
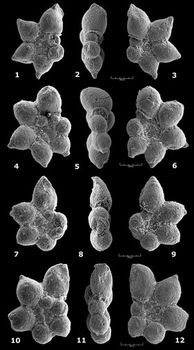
![]() Ultérieurement la seule évolution notable qui affectera les formes planispiralées durant le Gargasien inférieur/moyen concerne le nombre et la forme des loges. G. ferreolensis à 7 ou 8 loges va perdurer en tant que telle, mais apparaissent assez vite, toujours via de rares spécimens constituant autant de stades intermédiaires, des formes à 9 (= G. barri) puis un peu plus tard à au moins 10 (= G. algerianus) loges. Pour ce qui est de la forme des loges, au sein des trois espèces
(ferreolensis, barri, algerianus) on observe une tendance plus ou moins sporadique, chez quelques spécimens, à l'élargissement latéral, pouvant mener à une bilobation, des dernières loges (tendance
"Biglobigerinella"), ou à une amorce de déroulement, présente chez les trois taxons mais surtout chez G. algerianus. Parmi les auteurs, une très grande majorité estime qu'il n'est pas justifié d'accorder un statut générique à ces deux tendances (revue in et alii,
2002). En revanche, l'acquisition progressive (au confins du Gargasien supérieur et du Clansayésien) d'une sorte de carène périphérique parmi les populations de G. algerianus (stade non atteint dans la région de Cassis-La Bédoule, l'érosion ante-cénomaniene y ayant tronqué les termes sommitaux de l'Aptien), a été transcrite nomenclaturalement sur les plans spécifique et générique (= Pseudoplanomalina cheniourensis, cf. et alii,
2002).
Ultérieurement la seule évolution notable qui affectera les formes planispiralées durant le Gargasien inférieur/moyen concerne le nombre et la forme des loges. G. ferreolensis à 7 ou 8 loges va perdurer en tant que telle, mais apparaissent assez vite, toujours via de rares spécimens constituant autant de stades intermédiaires, des formes à 9 (= G. barri) puis un peu plus tard à au moins 10 (= G. algerianus) loges. Pour ce qui est de la forme des loges, au sein des trois espèces
(ferreolensis, barri, algerianus) on observe une tendance plus ou moins sporadique, chez quelques spécimens, à l'élargissement latéral, pouvant mener à une bilobation, des dernières loges (tendance
"Biglobigerinella"), ou à une amorce de déroulement, présente chez les trois taxons mais surtout chez G. algerianus. Parmi les auteurs, une très grande majorité estime qu'il n'est pas justifié d'accorder un statut générique à ces deux tendances (revue in et alii,
2002). En revanche, l'acquisition progressive (au confins du Gargasien supérieur et du Clansayésien) d'une sorte de carène périphérique parmi les populations de G. algerianus (stade non atteint dans la région de Cassis-La Bédoule, l'érosion ante-cénomaniene y ayant tronqué les termes sommitaux de l'Aptien), a été transcrite nomenclaturalement sur les plans spécifique et générique (= Pseudoplanomalina cheniourensis, cf. et alii,
2002).
![]() Une analyse statistique détaillée de la microfaune ainsi que des comparaisons avec les données sédimentaires sortent du cadre de cet article, d'ordre essentiellement biostratigraphique. Mais quelques données globales d'observation fondées sur la distribution des Foraminifères permettent de formuler l'hypothèse d'un léger approfondissement du milieu de dépôt au Gargasien, comparativement au Bédoulien supérieur (durant lequel l'épaisseur de la tranche d'eau a été estimée comme étant comprise entre une à deux centaines de mètres, et alii,
1998). Tout comme la diversité spécifique, le rapport planctoniques/benthiques tend ainsi à s'accroître durant la période considérée. Parmi les formes benthiques, la proportion de formes agglutinantes, en particulier à ciment organique
(Bathysiphon, Reophax, Recurvoides) tend aussi à devenir plus significative au Gargasien. Des formes calcaires de faible profondeur, telles que Spirillina minima, Patellina subcretacea, présentes au Bédoulien, sont rarissimes, voire absentes, au Gargasien.
Une analyse statistique détaillée de la microfaune ainsi que des comparaisons avec les données sédimentaires sortent du cadre de cet article, d'ordre essentiellement biostratigraphique. Mais quelques données globales d'observation fondées sur la distribution des Foraminifères permettent de formuler l'hypothèse d'un léger approfondissement du milieu de dépôt au Gargasien, comparativement au Bédoulien supérieur (durant lequel l'épaisseur de la tranche d'eau a été estimée comme étant comprise entre une à deux centaines de mètres, et alii,
1998). Tout comme la diversité spécifique, le rapport planctoniques/benthiques tend ainsi à s'accroître durant la période considérée. Parmi les formes benthiques, la proportion de formes agglutinantes, en particulier à ciment organique
(Bathysiphon, Reophax, Recurvoides) tend aussi à devenir plus significative au Gargasien. Des formes calcaires de faible profondeur, telles que Spirillina minima, Patellina subcretacea, présentes au Bédoulien, sont rarissimes, voire absentes, au Gargasien.
![]() L'étude micropaléontologique d'une section composite du Gargasien de l'aire stratotypique du Bédoulien a permis de suivre en grand détail l'évolution de la composition des associations de Foraminifères à la période de transition entre les deux sous-étages de l'Aptien.
L'étude micropaléontologique d'une section composite du Gargasien de l'aire stratotypique du Bédoulien a permis de suivre en grand détail l'évolution de la composition des associations de Foraminifères à la période de transition entre les deux sous-étages de l'Aptien.
![]() Les changements observés se révèlent globalement progressifs chez les formes benthiques, qui traduisent surtout un léger approfondissement du milieu en montant dans la série, mais qui, sur la base de plusieurs extinctions, permettent de caractériser stratigraphiquement le Bédoulien par rapport au Gargasien.
Les changements observés se révèlent globalement progressifs chez les formes benthiques, qui traduisent surtout un léger approfondissement du milieu en montant dans la série, mais qui, sur la base de plusieurs extinctions, permettent de caractériser stratigraphiquement le Bédoulien par rapport au Gargasien.
![]() Une part importante des espèces planctoniques montre une assez grande longévité, certaines nées au Bédoulien, voire au Barrémien supérieur, s'observant encore bien au-delà dans le Gargasien. Quelques marqueurs stables, au moment d'apparition bien calibré, permettent cependant de subdiviser la période considérée en cinq zones, au lieu de trois dans les zonations antérieures. Mais de part et d'autre de la limite
Bédoulien-Gargasien certains groupes sont affectés de phénomènes rapides, quoique progressifs, d'instabilité morphologique. Ces phases évolutives quelque peu désordonnées n'ont pas toujours été correctement analysées par les auteurs, certains se contentant de multiplier les créations de nouveaux taxons au sein de cette mouvance. La dilatation de la série et l'absence de hiatus dans la région de Cassis-La Bédoule nous ont permis d'observer ces processus en grand détail et surtout d'en contrôler la continuité stratigraphique. Il reste à conforter ces premiers résultats an procédant à une étude comparative des microfaunes de l'Aptien moyen d'une autre région privilégiée, celle de l'aire stratotypique d'Apt-Gargas (travaux en cours) et, via le "modèle" vocontien, à resituer dans le cadre téthysien les données provenant de l'étude des stratotypes.
Une part importante des espèces planctoniques montre une assez grande longévité, certaines nées au Bédoulien, voire au Barrémien supérieur, s'observant encore bien au-delà dans le Gargasien. Quelques marqueurs stables, au moment d'apparition bien calibré, permettent cependant de subdiviser la période considérée en cinq zones, au lieu de trois dans les zonations antérieures. Mais de part et d'autre de la limite
Bédoulien-Gargasien certains groupes sont affectés de phénomènes rapides, quoique progressifs, d'instabilité morphologique. Ces phases évolutives quelque peu désordonnées n'ont pas toujours été correctement analysées par les auteurs, certains se contentant de multiplier les créations de nouveaux taxons au sein de cette mouvance. La dilatation de la série et l'absence de hiatus dans la région de Cassis-La Bédoule nous ont permis d'observer ces processus en grand détail et surtout d'en contrôler la continuité stratigraphique. Il reste à conforter ces premiers résultats an procédant à une étude comparative des microfaunes de l'Aptien moyen d'une autre région privilégiée, celle de l'aire stratotypique d'Apt-Gargas (travaux en cours) et, via le "modèle" vocontien, à resituer dans le cadre téthysien les données provenant de l'étude des stratotypes.
![]() Nous ne donnerons ici que les strictes précisions nécessaires à une identification des principales espèces planctoniques, dont la systématique est la plus controversée et demeure en quasi-perpétuelle mouvance. Sur le plan générique, nous utiliserons la classification découlant de la hiérarchie des critères préconisée par et alii
(2002), en y incorporant (par exemple pour le statut
des "petits" Globigerinelloides) quelques modifications adaptées de travaux récents de
et (2003a). Des listes synonymiques, descriptions, discussions plus détaillées et figurations plus nombreuses seront fournies dans un article ultérieur, à caractère paléontologique plus affirmé et associant les observations effectuées sur les formes benthiques et planctoniques, à la fois en Basse-Provence, dans l'aire stratotypique aptienne (bassin d'Apt-Gargas) et dans le domaine vocontien.
Nous ne donnerons ici que les strictes précisions nécessaires à une identification des principales espèces planctoniques, dont la systématique est la plus controversée et demeure en quasi-perpétuelle mouvance. Sur le plan générique, nous utiliserons la classification découlant de la hiérarchie des critères préconisée par et alii
(2002), en y incorporant (par exemple pour le statut
des "petits" Globigerinelloides) quelques modifications adaptées de travaux récents de
et (2003a). Des listes synonymiques, descriptions, discussions plus détaillées et figurations plus nombreuses seront fournies dans un article ultérieur, à caractère paléontologique plus affirmé et associant les observations effectuées sur les formes benthiques et planctoniques, à la fois en Basse-Provence, dans l'aire stratotypique aptienne (bassin d'Apt-Gargas) et dans le domaine vocontien.
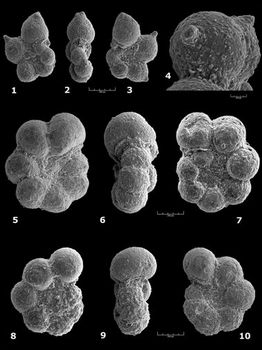
Praehedbergella aptiana (
1965)
Hedbergella aptiana , 1965, p. 347-348, texte-figs. 3-6.
Blefuscuiana aptiana (), et , 1988, p. 158, pl. 5, figs. 1-3.
Blefuscuiana aptiana () sensu stricto, , et , 1997, p. 125, pl. 8.2, figs. 1-8.
Blefuscuiana aptiana () orientalis , , , et , 1996, p. 125-126, pl. 8.2, figs. 9-11.
![]() Remarques. Nous partageons entièrement l'opinion de
et (1988), selon laquelle l'holotype décrit et figuré
texte-fig. 3 par
(1965) concerne un spécimen déformé, et la définition de cette espèce doit plutôt s'appuyer sur les paratypes (en particulier, d'après nous, sur ceux des
texte-figs. 4 et 6).
Remarques. Nous partageons entièrement l'opinion de
et (1988), selon laquelle l'holotype décrit et figuré
texte-fig. 3 par
(1965) concerne un spécimen déformé, et la définition de cette espèce doit plutôt s'appuyer sur les paratypes (en particulier, d'après nous, sur ceux des
texte-figs. 4 et 6).
![]() Occurrence. Barrémien moyen à Aptien inférieur selon
(1965). Dans le secteur de Cassis-La Bédoule, cette espèce est rare au Barrémien terminal, puis se rencontre avec une fréquence modérée durant le Bédoulien et le Gargasien inférieur-moyen.
Occurrence. Barrémien moyen à Aptien inférieur selon
(1965). Dans le secteur de Cassis-La Bédoule, cette espèce est rare au Barrémien terminal, puis se rencontre avec une fréquence modérée durant le Bédoulien et le Gargasien inférieur-moyen.
Praehedbergella globulifera (
et , in
1971)
Clavihedbergella globulifera et (in ), 1971, p. 136, pl. 10, figs. 1a-c.
Hedbergella maslakovae , 1974, p. 61-63, pl. 20, figs. 1-3, 14-16 ; pl. 24, figs. 11-14.
Hedbergella similis , 1974, p. 68-69, pl. 16, figs. 10-21 ; pl. 18, figs. 12-13 ; pl. 23, figs. 14-16.
Lilliputianella globulifera et , 1971, , et , 1997, p. 164-165, pl. 9.1, figs. 5-15.
![]() Remarques. L'holotype de
Praehedbergella globulifera comporte 5 loges, ceux de P. maslakovae et de P. similis 6
loges; on pourrait aussi les distinguer typologiquement par la forme des loges terminales, légèrement plus étirée radialement chez globulifera que chez similis et maslakovae (mais pas au point de former des loges cylindriformes comme chez P. roblesae). L'analyse des populations montre qu'en fait il s'agit d'un même plexus offrant une large variabilité, bien montrée par les nombreuses figurations de P. similis par (1974), et que les holotypes de
globulifera, maslakovae et similis n'y constituent que des morphotypes particuliers parmi bien d'autres. En tenant compte de l'antériorité nous sommes donc d'avis de suivre et alii
(1997), qui considèrent P. maslakovae et
P. similis comme des synonymes plus récents de l'espèce décrite par
et (1971).
Remarques. L'holotype de
Praehedbergella globulifera comporte 5 loges, ceux de P. maslakovae et de P. similis 6
loges; on pourrait aussi les distinguer typologiquement par la forme des loges terminales, légèrement plus étirée radialement chez globulifera que chez similis et maslakovae (mais pas au point de former des loges cylindriformes comme chez P. roblesae). L'analyse des populations montre qu'en fait il s'agit d'un même plexus offrant une large variabilité, bien montrée par les nombreuses figurations de P. similis par (1974), et que les holotypes de
globulifera, maslakovae et similis n'y constituent que des morphotypes particuliers parmi bien d'autres. En tenant compte de l'antériorité nous sommes donc d'avis de suivre et alii
(1997), qui considèrent P. maslakovae et
P. similis comme des synonymes plus récents de l'espèce décrite par
et (1971).
![]() Occurrence. Ce plexus apparaît au Bédoulien supérieur, est assez fréquemment représenté au Gargasien inférieur et voit sa fréquence diminuer progressivement dans le Gargasien moyen.
Occurrence. Ce plexus apparaît au Bédoulien supérieur, est assez fréquemment représenté au Gargasien inférieur et voit sa fréquence diminuer progressivement dans le Gargasien moyen.
Praehedbergella gorbachikae (
1974)
Pl. 5, figs. 7-12
![]()
Hedbergella gorbachikae , 1974, p. 56-58, pl. 15, figs. 1-16.
![]() Remarques.
Cette espèce se situe dans la mouvance du gr. infracretacea, dont elle se distingue par une protrusion plus marquée de la partie adombilicale de la dernière loge.
Remarques.
Cette espèce se situe dans la mouvance du gr. infracretacea, dont elle se distingue par une protrusion plus marquée de la partie adombilicale de la dernière loge.
![]() Occurrence.
(1974) a proposé ce taxon comme index d'une zone du Gargasien, s'intercalant entre la zone à Algerianus et la zone à Trocoidea. Nos observations nous conduisent à ne pas retenir cette proposition, car 1)
Praehedbergella gorbatchikae ne constitue qu'un avatar très marginal au sein de populations du gr.
infracretacea, 2) le moment d'apparition des premiers spécimens relevant de cette espèce coïncide avec celui de
Globigerinelloides algerianus, marqueur beaucoup plus significatif.
Occurrence.
(1974) a proposé ce taxon comme index d'une zone du Gargasien, s'intercalant entre la zone à Algerianus et la zone à Trocoidea. Nos observations nous conduisent à ne pas retenir cette proposition, car 1)
Praehedbergella gorbatchikae ne constitue qu'un avatar très marginal au sein de populations du gr.
infracretacea, 2) le moment d'apparition des premiers spécimens relevant de cette espèce coïncide avec celui de
Globigerinelloides algerianus, marqueur beaucoup plus significatif.
Praehedbergella infracretacea (
1937)
Globigerina infracretacea , 1937, p. 28, texte-fig. 1.
Globigerina aptica , 1951, p. 49, pl. 8, figs. 9-11.
Hedbergella delrioensis (), , 1974, p. 54-55 (pro parte : pl. 10, figs. 10-12 ; pl. 13, figs. 3-5, 15-18).
Praehedbergella luterbacheri (
1974)
Pl. 3, figs. 5-10
![]() ; Pl. 4, figs. 1-6
; Pl. 4, figs. 1-6
![]()
Hedbergella luterbacheri , 1974, p. 61, pl. 19, figs. 21-23, 24-26 ; pl. 26, figs. 15-17.
Praeglobotruncana modesta , 1959, p. 267 (pro parte : au moins formes citées, mais non figurées, de la Zone à Biglobigerinella barri, Maridale formation).
Hedbergella planispira () 1940, , 1974, p. 64-65 (pro parte : formes aptiennes à 7 loges, soit : pl. 11, fig. 4-6 ; pl. 23, figs. 17-18).
Blefuscuiana kuznetsovae et , 1988, p. 156-158, (pro parte : pl. 3, fig. 10 ; pl. 4, figs. 1-2, 4 ; non pl. 4, fig. 3).
![]() Remarques. Nous suivons l'auteur de l'espèce en limitant l'acception de
Praehedbergella luterbacheri aux formes à trochospire basse à 7 (très rarement 8) loges à accroissement lent, avec un test au contour modérément lobé. Les spécimens à 7 loges de
P. kuznetsovae (dont l'holotype) n'en différent que par un rythme d'accroissement très légèrement plus marqué de la hauteur des loges, et la prise en considération de l'éventail de variabilité au sein de populations conduit à considérer cette espèce comme synonyme plus récent de
P. luterbacheri.
Remarques. Nous suivons l'auteur de l'espèce en limitant l'acception de
Praehedbergella luterbacheri aux formes à trochospire basse à 7 (très rarement 8) loges à accroissement lent, avec un test au contour modérément lobé. Les spécimens à 7 loges de
P. kuznetsovae (dont l'holotype) n'en différent que par un rythme d'accroissement très légèrement plus marqué de la hauteur des loges, et la prise en considération de l'éventail de variabilité au sein de populations conduit à considérer cette espèce comme synonyme plus récent de
P. luterbacheri.
![]() Nos observations dans le secteur de La Bédoule-Cassis montrent que par évolution progressive de son test, initialement trochospiralé, vers la planispiralité,
P. luterbacheri constitue l'ancêtre immédiat de Globigerinelloides ferreolensis (cf. Pl. 4, figs.
7-9
Nos observations dans le secteur de La Bédoule-Cassis montrent que par évolution progressive de son test, initialement trochospiralé, vers la planispiralité,
P. luterbacheri constitue l'ancêtre immédiat de Globigerinelloides ferreolensis (cf. Pl. 4, figs.
7-9
![]() ).
).
![]() Occurrence. Par son moment d'apparition, situé au sein de la partie supérieure de la zone à Cabri
(sensu lato), très peu - quelques mètres - au-dessus de la limite Bédoulien-Gargasien, cette espèce constitue un marqueur intéressant pour définir de manière approchée la limite entre les deux sous-étages, de même qu'une nouvelle zone insérée entre la zone à Cabri
(s.s.) et la zone à Ferreolensis (cf. section biostratigraphie de cet article). Sa fréquence de représentation, toujours modérée, décroît dans la partie supérieure de la zone à Ferreolensis et l'espèce disparaît au niveau de la limite entre les zones à Ferreolensis et à Algerianus.
Occurrence. Par son moment d'apparition, situé au sein de la partie supérieure de la zone à Cabri
(sensu lato), très peu - quelques mètres - au-dessus de la limite Bédoulien-Gargasien, cette espèce constitue un marqueur intéressant pour définir de manière approchée la limite entre les deux sous-étages, de même qu'une nouvelle zone insérée entre la zone à Cabri
(s.s.) et la zone à Ferreolensis (cf. section biostratigraphie de cet article). Sa fréquence de représentation, toujours modérée, décroît dans la partie supérieure de la zone à Ferreolensis et l'espèce disparaît au niveau de la limite entre les zones à Ferreolensis et à Algerianus.
Praehedbergella praetrocoidea (
et 1986)
Pl. 4, figs. 10-12
![]() ; Pl. 5, figs. 1-6
; Pl. 5, figs. 1-6
![]()
Hedbergella praetrocoidea et (in ), 1986, p. 95, pl. 16, figs. 3-5.
![]() Occurrence. Espèce très sporadiquement représentée, toujours avec un très petit nombre d'individus, au cours du Gargasien inférieur-moyen.
Occurrence. Espèce très sporadiquement représentée, toujours avec un très petit nombre d'individus, au cours du Gargasien inférieur-moyen.
Praehedbergella primare (
et 1986)
Clavihedbergella primare et (in ), 1986, p. 120-121, pl. 25, figs. 2a-3a, 2b-3b.
Hedbergella sp., aff. planispira (), , 1966, p. 94, pl. 8, figs. 1-3.
Hedbergella planispira (, 1940), , 1974, p. 64-65 (pro parte : formes aptiennes à 6 loges) : pl. 23, figs. 5-7.
Hedbergella aff. planispira auct., , 1979, pl. 2, figs. 1-2.
Blefuscuiana kuznetsovae et , 1988, p. 156-158 (pro parte : pl. 4, fig. 3)
Hedbergella kuznetsovae ( et ), et , 1994, p. 676, fig. 12 : 1-12.
Blefuscuiana primare ( et , 1986), , et , 1997, p. 131, pl. 8.10, figs. 1-3.
Praehedbergella kuznetsovae ( et , 1988), , , et , 1998, p. 207-208, pl. 3, figs. 8-9.
![]() Remarques.
et (in ,
1986) définissent cette espèce comme possédant de 5,5 à 6 loges au dernier tour et figurent un holotype à 6 loges. La description quelque peu sommaire et les illustrations de médiocre qualité de
et (1986) n'ont pas permis initialement de se faire une idée précise de ce à quoi correspondait cette espèce.
Remarques.
et (in ,
1986) définissent cette espèce comme possédant de 5,5 à 6 loges au dernier tour et figurent un holotype à 6 loges. La description quelque peu sommaire et les illustrations de médiocre qualité de
et (1986) n'ont pas permis initialement de se faire une idée précise de ce à quoi correspondait cette espèce.
![]() A la lumière des figurations de topotypes par ,
et (1997), il semble que la dénomination de
Praehedbergella primare corresponde bien au concept de formes barrémo-aptiennes, souvent rapprochées
d'Hedbergella planispira, à test finement perforé, trochospire basse, avec 6 loges au dernier tour,
d'accroissement très modéré. Ces spécimens homéomorphes
d'Hedbergella planispira ne peuvent être attribués à cette espèce albienne, qui possède un test macroperforé, non plus qu'à l'espèce
kuznetsovae, typiquement à 7 loges et à ouverture de la spire un peu plus rapide.
A la lumière des figurations de topotypes par ,
et (1997), il semble que la dénomination de
Praehedbergella primare corresponde bien au concept de formes barrémo-aptiennes, souvent rapprochées
d'Hedbergella planispira, à test finement perforé, trochospire basse, avec 6 loges au dernier tour,
d'accroissement très modéré. Ces spécimens homéomorphes
d'Hedbergella planispira ne peuvent être attribués à cette espèce albienne, qui possède un test macroperforé, non plus qu'à l'espèce
kuznetsovae, typiquement à 7 loges et à ouverture de la spire un peu plus rapide.
![]() Occurrence. Crimée : Barrémien supérieur-Aptien inférieur. Dans le SE de la France, cette espèce apparaît au Barrémien terminal et s'observe (avec une fréquence décroissante) jusque dans le Gargasien moyen (zone à Algerianus).
Occurrence. Crimée : Barrémien supérieur-Aptien inférieur. Dans le SE de la France, cette espèce apparaît au Barrémien terminal et s'observe (avec une fréquence décroissante) jusque dans le Gargasien moyen (zone à Algerianus).
Praehedbergella roblesae ( 1959)
Globigerina roblesae , 1959, p. 149, pl. 4, fig. 4.
Schackoina gr. cabri (
1952)
Schackoina cabri , 1952, p. 20-21, figs. 1-8.
Schackoina pustulans , 1957, p. 274-275, pl. 1, figs. 1-4, 6-7.
Leupoldina protuberans , 1957, p. 277, pl. 2, figs. 1-13.
Globigerinelloides algerianus (
et 1948)
Globigerinelloides algeriana et , 1948, p. 43, pl. 8, figs. 4-6.
![]() Remarques. Test planispiralé, au minimum 10 loges au dernier tour, typiquement 10 à 12, voire plus. Tendance au déroulement ou à la bilobation de la ou des dernières loges chez certains spécimens.
Remarques. Test planispiralé, au minimum 10 loges au dernier tour, typiquement 10 à 12, voire plus. Tendance au déroulement ou à la bilobation de la ou des dernières loges chez certains spécimens.
![]() Occurrence. Ce marqueur du Gargasien moyen-supérieur téthysien ne s'observe que dans les derniers mètres de la coupe de Cassis-La Marcouline, n'y étant d'ailleurs représenté que par d'assez rares spécimens, ne dépassant pas 10-11 loges.
Occurrence. Ce marqueur du Gargasien moyen-supérieur téthysien ne s'observe que dans les derniers mètres de la coupe de Cassis-La Marcouline, n'y étant d'ailleurs représenté que par d'assez rares spécimens, ne dépassant pas 10-11 loges.
Globigerinelloides gr. aptiensis (
1974)
Globigerinelloides aptiense , 1974, p. 79-80, pl. 4, figs. 9-10 ; pl. 8, figs. 4-6, 17-18.
Blowiella solida et (in ), 1986, p. 123, pl. 27, fig. 3.
![]() Remarques. Typologiquement, ces deux formes planispiralées à 6 loges au dernier tour ne diffèrent que par le degré d'ouverture de la spire, très modéré chez le morphotype
solida, un peu plus rapide chez le morphotype aptiensis. Là encore nombre de stades intermédiaires
existent; la distribution stratigraphique (au demeurant très sporadique) des divers morphotypes est la même, et nous sommes donc d'avis de suivre
et (2003b), qui considèrent que
Blowiella solida est un synonyme plus récent de Globigerinelloides aptiensis. Nous avons noté la présence au sein de ce plexus de très rares spécimens biglobigérinelloïdes, la tendance à la bilobation de la dernière loge se produisant plutôt chez le morphotype
solida.
Remarques. Typologiquement, ces deux formes planispiralées à 6 loges au dernier tour ne diffèrent que par le degré d'ouverture de la spire, très modéré chez le morphotype
solida, un peu plus rapide chez le morphotype aptiensis. Là encore nombre de stades intermédiaires
existent; la distribution stratigraphique (au demeurant très sporadique) des divers morphotypes est la même, et nous sommes donc d'avis de suivre
et (2003b), qui considèrent que
Blowiella solida est un synonyme plus récent de Globigerinelloides aptiensis. Nous avons noté la présence au sein de ce plexus de très rares spécimens biglobigérinelloïdes, la tendance à la bilobation de la dernière loge se produisant plutôt chez le morphotype
solida.
![]() Occurrence. De très rares formes relevant de ce plexus apparaissent au Bédoulien supérieur, puis s'observent sporadiquement dans le Gargasien inférieur-moyen, et deviennent encore plus rares ensuite.
Occurrence. De très rares formes relevant de ce plexus apparaissent au Bédoulien supérieur, puis s'observent sporadiquement dans le Gargasien inférieur-moyen, et deviennent encore plus rares ensuite.
Globigerinelloides barri (,
et , 1957)
Globigerinelloides barri , et , 1957, p. 25 (pro parte : pl. 1, figs. 17, 18 a-b).
![]() Remarques. Test planispiralé, 9 loges au dernier tour. Successeur phylogénétique de
G. ferreolensis.
Remarques. Test planispiralé, 9 loges au dernier tour. Successeur phylogénétique de
G. ferreolensis.
"Globigerinelloides" gr. bizonae (
1961) – saundersi (,
1959)
Pl. 1, figs. 1-12
![]() ; Pl. 2, figs. 1-12
; Pl. 2, figs. 1-12
![]() ; Pl. 3, figs. 1-4
; Pl. 3, figs. 1-4
![]()
Planomalina saundersi , 1959, p. 262, pl. 20, figs. 9-11.
Hastigerinella bizonae , 1961, p. 34-36, pl. 1, figs. 24-28.
Clavihedbergella bizonae (, 1961), , 1971, p. 231-232, pl. 3, figs. 2 a-c.
Hedbergella kuhryi , 1974, p. 60, pl. 14, figs. 1-6.
non Claviblowiella saundersi (, 1959), , et , 1997, p. 181, pl. 10.2, figs. 7-11.
Blowiella saundersi (, 1959), , , et , 1998, p. 205-206, pl. 4, fig. 6.
Claviblowiella saundersi (, 1959), et , 2004, p. 42, pl. 3, figs. 3a-b.
![]() Remarques. Dans le but de mieux comparer l'espèce de
(1961) à celle qui fut décrite par
(1959) sous le nom de Planomalina saundersi, nous avons examiné directement l'holotype de
bizonae (déposé dans les collections du Muséum d'Histoire Naturelle de Paris, N°
F62120), refiguré ici en microscopie optique (pl. 1, figs. 10-12
Remarques. Dans le but de mieux comparer l'espèce de
(1961) à celle qui fut décrite par
(1959) sous le nom de Planomalina saundersi, nous avons examiné directement l'holotype de
bizonae (déposé dans les collections du Muséum d'Histoire Naturelle de Paris, N°
F62120), refiguré ici en microscopie optique (pl. 1, figs. 10-12
![]() ). La forme des loges terminales (ogivale avec tendance à un effilement apical, laissant supposer un départ d'épine, pratiquement toujours brisée dès sa base) est semblable chez les deux taxons.
(1961) parle d'un enroulement "trochoïde" (= trochospiral) pour cette espèce, effectivement visible sur les paratypes figurés; mais l'holotype s'avère faiblement dissymétrique et tend vers une certaine subplanispiralité. L'holotype de l'espèce de est décrit comme planispiral par son auteur, mais la vue de profil montre une légère dissymétrie. En stricte approche typologique, il est donc difficile de trouver des arguments objectifs pour différencier les deux espèces.
). La forme des loges terminales (ogivale avec tendance à un effilement apical, laissant supposer un départ d'épine, pratiquement toujours brisée dès sa base) est semblable chez les deux taxons.
(1961) parle d'un enroulement "trochoïde" (= trochospiral) pour cette espèce, effectivement visible sur les paratypes figurés; mais l'holotype s'avère faiblement dissymétrique et tend vers une certaine subplanispiralité. L'holotype de l'espèce de est décrit comme planispiral par son auteur, mais la vue de profil montre une légère dissymétrie. En stricte approche typologique, il est donc difficile de trouver des arguments objectifs pour différencier les deux espèces.
![]() Les documents stratigraphiques du D.E.S. de
(1960) montrent que l'intégralité de la population-type de sa nouvelle espèce (holotype et 10 paratypoïdes, dont seulement 4 exemplaires – les plus nettement trochospiralés – ont été figurés dans l'article de
1961) provient d'un même échantillon (MG 781). Celui-ci se situe dans la coupe de la carrière de La Marcouline, dans la formation sommitale, plus marneuse, que nous avons datée du Gargasien moyen (zone à Algerianus).
(1960) cite d'ailleurs la présence de ce taxon dans les niveaux où se situe son échantillon. Parmi notre matériel, les spécimens que nous figurons Pl. 3, figs.
1-4
Les documents stratigraphiques du D.E.S. de
(1960) montrent que l'intégralité de la population-type de sa nouvelle espèce (holotype et 10 paratypoïdes, dont seulement 4 exemplaires – les plus nettement trochospiralés – ont été figurés dans l'article de
1961) provient d'un même échantillon (MG 781). Celui-ci se situe dans la coupe de la carrière de La Marcouline, dans la formation sommitale, plus marneuse, que nous avons datée du Gargasien moyen (zone à Algerianus).
(1960) cite d'ailleurs la présence de ce taxon dans les niveaux où se situe son échantillon. Parmi notre matériel, les spécimens que nous figurons Pl. 3, figs.
1-4
![]() proviennent de cette formation.
proviennent de cette formation.
![]() L'analyse minutieuse des populations que nous avons suivies pas à pas dans les coupes du stratotype historique de Cassis-La Bédoule montre que celles du celles du Bédoulien terminal-Gargasien inférieur sont composées de manière prédominante de spécimens nettement trochospiralés (mais tout ou partie du tour interne est souvent visible du côté ombilical, par ailleurs un peu plus déprimé que la surface homologue du côté dorsal). Dès le Gargasien inférieur on voit apparaître une tendance vers une certaine symétrisation de l'enroulement, observée chez un nombre croissant de spécimens à mesure que l'on monte dans la série. Au Gargasien moyen ces spécimens subsymétriques deviennent prédominants, mais ils sont toujours accompagnés de quelques spécimens plus nettement dissymétriques.
L'analyse minutieuse des populations que nous avons suivies pas à pas dans les coupes du stratotype historique de Cassis-La Bédoule montre que celles du celles du Bédoulien terminal-Gargasien inférieur sont composées de manière prédominante de spécimens nettement trochospiralés (mais tout ou partie du tour interne est souvent visible du côté ombilical, par ailleurs un peu plus déprimé que la surface homologue du côté dorsal). Dès le Gargasien inférieur on voit apparaître une tendance vers une certaine symétrisation de l'enroulement, observée chez un nombre croissant de spécimens à mesure que l'on monte dans la série. Au Gargasien moyen ces spécimens subsymétriques deviennent prédominants, mais ils sont toujours accompagnés de quelques spécimens plus nettement dissymétriques.
![]() A cette variabilité dans la nature de l'enroulement s'ajoute un certain polymorphisme affectant la forme des loges (concernant en particulier leur degré d'étirement radial, plus ou moins marqué, pouvant aller parfois jusqu'au stade
"cepedai", une espèce décrite par (1959) qui ne semble constituer qu'un variant extrême de l'espèce de , très peu représenté dans nos populations).
A cette variabilité dans la nature de l'enroulement s'ajoute un certain polymorphisme affectant la forme des loges (concernant en particulier leur degré d'étirement radial, plus ou moins marqué, pouvant aller parfois jusqu'au stade
"cepedai", une espèce décrite par (1959) qui ne semble constituer qu'un variant extrême de l'espèce de , très peu représenté dans nos populations).
![]() Par conséquent il semble qu'aussi bien sur le plan strictement typologique que populationnel, l'espèce de
(1961) ne peut être distinguée de l'espèce décrite par
(1959) et doit être considérée comme un synonyme plus récent de cette dernière.
Par conséquent il semble qu'aussi bien sur le plan strictement typologique que populationnel, l'espèce de
(1961) ne peut être distinguée de l'espèce décrite par
(1959) et doit être considérée comme un synonyme plus récent de cette dernière.
![]() Au
sein de ce plexus, la fluctuation de caractères (par exemple, nature de l'enroulement,
voire prolongation épineuse des loges) habituellement considérés comme
d'ordre générique par la plupart des auteurs pose le problème de
l'attribution de ce plexus à un genre déterminé. Dans l'attente du résultat
de nouvelles recherches plus approfondies, nous avons pris le parti de grouper
ces formes sous le vocable provisoire "Globigerinelloides".
Au
sein de ce plexus, la fluctuation de caractères (par exemple, nature de l'enroulement,
voire prolongation épineuse des loges) habituellement considérés comme
d'ordre générique par la plupart des auteurs pose le problème de
l'attribution de ce plexus à un genre déterminé. Dans l'attente du résultat
de nouvelles recherches plus approfondies, nous avons pris le parti de grouper
ces formes sous le vocable provisoire "Globigerinelloides".
Globigerinelloides gr. blowi (
1959)
Planomalina blowi , 1959, p. 260, pl. 20, figs. 2-3.
Planomalina maridalensis , 1959, p. 261, pl. 20, figs. 4-6.
Globigerinella duboisi , 1961, p. 33, pl. 1, figs. 14-18.
Globigerinella gottisi , 1961, p. 32 (pro parte : pl. 1, figs. 9-11, 13).
Globigerinelloides paragottisi et , 2003, p. 332-333, figs. 6. 7-14, 7. 1-13.
![]() Remarques. Au sein de ce plexus planispiralé à 4-5 loges,
G. duboisi (typiquement 4 loges) et G. blowi (5 loges) constituent les morphotypes dominants, toujours simultanément présents et reliés par une minorité de stades intermédiaires, aussi bien pour la forme que pour le nombre des loges. Les morphotypes
G. maridalensis et G. paragottisi ne s'observent que très sporadiquement et leur distinction en tant qu'espèces ne présente guère d'intérêt.
Remarques. Au sein de ce plexus planispiralé à 4-5 loges,
G. duboisi (typiquement 4 loges) et G. blowi (5 loges) constituent les morphotypes dominants, toujours simultanément présents et reliés par une minorité de stades intermédiaires, aussi bien pour la forme que pour le nombre des loges. Les morphotypes
G. maridalensis et G. paragottisi ne s'observent que très sporadiquement et leur distinction en tant qu'espèces ne présente guère d'intérêt.
![]() Occurrence. L'ensemble de ces formes est plus ou moins constamment représenté du Bédoulien supérieur jusqu'au sommet du Gargasien.
Occurrence. L'ensemble de ces formes est plus ou moins constamment représenté du Bédoulien supérieur jusqu'au sommet du Gargasien.
Globigerinelloides ferreolensis (
1961)
Biticinella ferreolensis , 1961, p. 214, pl. 1, figs. 1-5.
Biglobigerinella sigali , 1961, p. 33, pl. 1, figs. 19-23.
Globigerinelloides macrocameratus , 1974, p. 85-86, pl. 5, figs. 1-6.
![]() Remarques. Test planispiralé, à 7-8 loges au dernier tour. Au Gargasien inférieur, on observe la dérivation de cette espèce à partir de spécimens de
Praehedbergella luterbacheri à trochospire de plus en plus plane, avec une face spirale très peu différente de la face
ombilicale : la première montre simplement un peu plus de loges du tour interne. L'ouverture tend corrélativement à être de plus en plus périphérique. L'évolution de tous ces caractères se situe dans un processus progressif mais rapide de cheminement vers la planispiralité, avec nombre de spécimens intermédiaires, dont la variabilité ne nous a pas paru justifier la création d'une nouvelle espèce (sorte de stade
"praeferreolensis", d'attribution générique indéterminée, puisque ne relevant typiquement ni de
Praehedbergella ni de Globigerinelloides).
Remarques. Test planispiralé, à 7-8 loges au dernier tour. Au Gargasien inférieur, on observe la dérivation de cette espèce à partir de spécimens de
Praehedbergella luterbacheri à trochospire de plus en plus plane, avec une face spirale très peu différente de la face
ombilicale : la première montre simplement un peu plus de loges du tour interne. L'ouverture tend corrélativement à être de plus en plus périphérique. L'évolution de tous ces caractères se situe dans un processus progressif mais rapide de cheminement vers la planispiralité, avec nombre de spécimens intermédiaires, dont la variabilité ne nous a pas paru justifier la création d'une nouvelle espèce (sorte de stade
"praeferreolensis", d'attribution générique indéterminée, puisque ne relevant typiquement ni de
Praehedbergella ni de Globigerinelloides).
![]() Même lorsque l'espèce
ferreolensis apparaît stabilisée pour ce qui est de son enroulement, la variabilité morphologique intra-populationnelle demeure relativement importante. Elle peut affecter le taux de croissance en largeur des loges (jusqu'à la bilobation terminale conduisant au morphotype
"Biglobigerinella"), le rythme d'ouverture de la spire et d'accroissement de la hauteur des loges, sans aller jusqu'au déroulement ou, en mode régressif (accroissement très lent), justifier la distinction de l'espèce
"macrocameratus" (, 1974).
Même lorsque l'espèce
ferreolensis apparaît stabilisée pour ce qui est de son enroulement, la variabilité morphologique intra-populationnelle demeure relativement importante. Elle peut affecter le taux de croissance en largeur des loges (jusqu'à la bilobation terminale conduisant au morphotype
"Biglobigerinella"), le rythme d'ouverture de la spire et d'accroissement de la hauteur des loges, sans aller jusqu'au déroulement ou, en mode régressif (accroissement très lent), justifier la distinction de l'espèce
"macrocameratus" (, 1974).
![]() Occurrence. Espèce gargasienne (extrême base du sous-étage exclue).
Occurrence. Espèce gargasienne (extrême base du sous-étage exclue).
![]() Les auteurs expriment leur gratitude à Mme M.-T. (Muséum National d'Histoire Naturelle, Paris) qui a mis à notre disposition le matériel-type de la collection (dont l'holotype de
"Hastigerinella bizonae", N° F62120), Mr Omar (Service MEB de l'UFR de "Sciences de la Terre et évolution des milieux naturels" de l'Université Pierre
et Marie Curie (UPMC, Paris) pour les prises de vues au Microscope Electronique à Balayage, et Mr Claude (Atelier de Reprographie du Département de Géologie sédimentaire de l'UPMC, Paris) pour le traitement des images photographiques numérisées.
Les auteurs expriment leur gratitude à Mme M.-T. (Muséum National d'Histoire Naturelle, Paris) qui a mis à notre disposition le matériel-type de la collection (dont l'holotype de
"Hastigerinella bizonae", N° F62120), Mr Omar (Service MEB de l'UFR de "Sciences de la Terre et évolution des milieux naturels" de l'Université Pierre
et Marie Curie (UPMC, Paris) pour les prises de vues au Microscope Electronique à Balayage, et Mr Claude (Atelier de Reprographie du Département de Géologie sédimentaire de l'UPMC, Paris) pour le traitement des images photographiques numérisées.
![]() Ils tiennent également à
remercier Mmes M. (Université de Fribourg) et D. (Université de
Strasbourg) pour leur relecture critique du manuscrit.
Ils tiennent également à
remercier Mmes M. (Université de Fribourg) et D. (Université de
Strasbourg) pour leur relecture critique du manuscrit.
![]()
![]()
D.A. (1951).- Studies of microfauna from Cretaceous deposits of Azerbaydzhan.- In : D.I., D.A. & D.M. (Eds), (Aznefteizdat, Baku) 128 pp. (en russe)
N.V. (1962).- Désagrégation des roches argilo-marneuses sous l'action simultanée de l'hyposulfite et de l'eau oxygénée.- Voprosy Mikropaleontologii, Moscou, N° 6, pp. 147-148 (en russe ; traduction B.R.G.M. N° 4123).
F.T. & D. (1988).- A review and revision of the Jurassic–Early Cretaceous Globigerinina, with especial reference to the Aptian assemblages of Speeton (North Yorkshire, England).- Journal of Micropaleontology, London, Vol. 7, Part 2, pp. 143–185.
H. (1965).- Taxionomische Revision und Nomenklator zu Franz E. "Standard-Gliederung der Nordwest-deutschen Unterkreide nach Foraminiferen" (1938). Teil 4: Alb. Mit Beschreibungen von Arten aus verschiedenen Unterkreide-Niveaus.- Senckenbergiana Lethaea, Frankfurt am Main, Bd. 46, N° 4/6, pp. 327-366.
H.M. (1957).- The foraminiferal genera Schackoina , emended and Leupoldina, n. gen. in the Cretaceous of Trinidad B.W. I.- Eclogae Geologicae Helvetiae, Basel, Vol. 50, pp. 271–278.
H.M. (1959).- Planktonic foraminifera from the Cretaceous of Trinidad, B.W.I.- Bulletins of American Paleontology, New York, Vol. XXXIX, pp. 257–277.
H.M., A.R. & H. (1957).- Planktonic foraminiferal families Hantkeninidae, Orbulinidae, Globorotaliidae, and Globotruncanidae.- In : A.R. et alii (Eds.), Studies in foraminifera.- United States National Museum, Bulletin, Washington, Vol. 215, pp. 3–50.
M.K., F.T., T.N., M.D. & J.E. (1996).- Evolution in the Early Cretaceous planktonic foraminiferal genus Blefuscuiana.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Bd. 202, pp. 243–258.
M.K., F.T. & J.E. (eds.) (1997).- The early evolutionary history of planktonic foraminifera.- Chapman & Hall, London, 269 p.
M.K., F.T., T.N., M.D. & J.E. (1998).- On the evolution of the Hedbergellidae from the Praehedbergellidae.- Journal of Micropaleontology, London, Vol. 17, pp. 97-103.
G., M. & B. (1965).- Utilisation de l'azote liquide et des mouillants pour l'extraction et la préparation des microfossiles.- Comptes Rendus de l'Académie des Sciences Paris, t. 261, pp. 2383-2386.
M. & P. (1983).- Evolution of early planktonic foraminifers.- Marine Micropaleontology, Amsterdam, Vol. 7, Issue 6, pp. 453-462.
J. (1960).- Contribution à l'étude stratigraphique et micropaléontologique du Crétacé inférieur du Languedoc et de la Provence.- Diplôme d'Etudes Supérieures, Université de Paris, 151 p.
J. (1961).- Quelques nouvelles espèces dans le Crétacé inférieur méditerranéen.- Revue de Micropaléontologie, Paris, 4e Année, N° 1, pp. 30–36.
R. & I. (1994).- Planktonic foraminifera from the Lower Cretaceous of Rio Argos sections (southern Spain) and biostratigraphic implications.- Cretaceous Research, London, Vol. 15, Issue 6, pp. 645–687.
G. (1994).- La limite Bédoulien-Gargasien dans la coupe stratotypique de Cassis-La Bédoule (Bouches-du-Rhône, France).- Géologie Alpine, Grenoble, Mém. H.S., N° 20, p. 321-326.
J.A. & A. (1948).- Globigerinelloides, a new genus of the Globigerinidae.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon, Vol. 24, pp. 42–44.
D., D., B., S. & F. (2004).- Enregistrement à haute résolution des modifications environnementales inscrites dans un cadre téphrochronologique : le bassin du Western Interior au passage Cénomanien-Turonien (USA).- Bulletin de la Société géologique de France, Paris, t. 175, N° 6, pp. 561-572.
S., M. & G. (1965).- Le Bédoulien dans sa région-type, La Bédoule-Cassis (B.-du-Rh.).- In : Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Bureau de Recherches Géologiques et Minières, Mémoires, Orléans, N° 34, pp. 173-179.
G. (1987).- Dynamique du bassin subalpin méridional de l'Aptien au Cénomanien.- École des Mines de Paris, Mémoires des Sciences de la Terre, N° 4, 370 p.
G. (1975).- Contribution à l'étude stratigraphique et sédimentologique de l'Aptien supérieur de Provence.- Thèse, Université de Provence, 92 p.
M.F. (1937).- Planktonforaminiferen aus der Kreide und dem Eozän und ihre stratigraphische Bedeutung.- Etudy po Mikropaleontologii, Paleontologicheskaya Laboratoriya Moskovskogo Gosudarstvennogo Universiteta, 1, pp. 27–46.
T.N. (1971).- On Early Cretaceous foraminifera of the Crimea.- Voprosy Mikropaleontologii, Moscow, N° 14, pp. 125–139 & 211-216, pls. XXI-XXX. (en russe)
T.N. (1986).- Jurassic and Early Cretaceous planktonic foraminifera of the south of the USSR.- (Akademia Nauk SSSR, Nauka, Moscou), 239 p. (en russe)
B. (1971).- Lower Cretaceous planktonic Foraminifera from the Miravetes, Argos and Represa Formations (S.E. Spain).- Revista Española de Micropaleontologia, Madrid, Vol. III, N° 3, pp. 219-237.
S. & A. (2004).- Aptian planktonic foraminifera from Israel.- Israel Journal of Earth Sciences, Jerusalem, Vol. 53, N° 1, pp. 27-46.
J.F. (1974).- Stratigraphic, morphologic and taxonomic studies of Aptian planktonic Foraminifera.- Revista Española de Micropaleontologia, Madrid, Numero Extraordinario, pp. 1–107.
F., M. & G. (1984).- Crétacé inférieur : 1.3.6. - Foraminifères planctoniques et benthiques de bassin.- In : S. et alii (Eds.), Synthèse Géologique du Sud-Est de la France. I : Stratigraphie et paléogéographie.- Bureau de Recherches Géologiques et Minières, Mémoires, Orléans, N° 125, pp. 297-299.
M. (1961).- Quelques Foraminifères et Ostracodes nouveaux du Crétacé inférieur des Baronnies (Drôme).- Revue de Micropaléontologie, Paris, 3e Année, N° 4, pp. 213–216.
M. (1966).- Etude stratigraphique et micropaléontologique du Crétacé inférieur de la "fosse vocontienne".- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, N° 15, pp. 1–369.
M. & G. (2004).- Le Gargasien (Aptien moyen) dans les stratotypes historiques de l'Aptien (SE France) : Introduction générale.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/01, pp. 1-3.
M., G., W. & J.-P. (1998).- Les Foraminifères benthiques et planctoniques de la série du Stratotype Historique de l’Aptien inférieur dans la région de Cassis-La Bédoule (SE France).- Géologie Méditerranéenne, Marseille, t. XXV, N° 3-4, pp. 187–225.
M., J.-P. & G. (2002).- Hierarchy of criteria, evolutionary processes and taxonomic simplification in the classification of Lower Cretaceous planktonic foraminifera.- Cretaceous Research, London, Vol. 23, Issue 1, pp. 111–148
M., G., W., M. & J.-P. (2004).- Le Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de l'Aptien inférieur, SE France) : localisation géographique et corrélations stratigraphiques.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/02, pp. 1-4.
J. (1959).- Foraminíferos de la Formación La Peña.- Boletín de la Asociación Mexicana de Geólogos Petroleros, México, Vol. XI, N° 3-4, pp. 135–153.
F. & M. (1995).- Foraminifères planctoniques du Crétacé : commentaire de la zonation Europe-Méditerranée.- Bulletin de la Société Géologique de France, Paris, t. 166, N° 6, pp. 681-692.
P., G., R., J.-P. & M. (1998).- Les faunes d'Ammonites du Barrémien supérieur/Aptien inférieur (Bédoulien) dans la région stratotypique de Cassis-La Bédoule (SE France) : état des connaissances et propositions pour une zonation par Ammonites du Bédoulien-type.- Géologie Méditerranéenne, Marseille, t. XXV, N° 3-4, pp. 167–175.
J. (1952).- Aperçu stratigraphique sur la micropaléontologie du Crétacé.- XIXe Congrès Géologique International, Monographies régionales, (1re Série : Algérie), Alger, N° 26, pp. 3–47.
H. (1940).- Foraminifera from the Grayson Formation of northern Texas.- Journal of Paleontology, Tulsa, Vol. 14, N° 2, pp. 93-126.
G. (1981).- Les Foraminifères crétacés de Provence (Aptien-Santonien) – systématique, biostratigraphie, paléoécologie, paléogéographie.- Thèse, Université de Provence, Travaux du Laboratoire de Géologie Historique et de Paléontologie, Marseille, N° 12, 559 p.
D. & I. (2002).- Early Cretaceous planktonic foraminifera from the Tethys: the genus Leupoldina.- Cretaceous Research, London, Vol. 23, Issue 2, pp. 189-212.
D. & I. (2003a).- Early Cretaceous planktonic foraminifera from the Tethys: the small, few-chambered representatives of the genus Globigerinelloides.- Cretaceous Research, London, Vol. 24, Issue 3, pp. 305-334.
D. & I. (2003b).- Early Cretaceous planktonic foraminifera from the Tethys: the large, many-chambered representatives of the genus Globigerinelloides.- Cretaceous Research, London, Vol. 24, Issue 6, pp. 661-690.
![]()
Cliquer sur la miniature pour agrandir l'image.
Figure 1 : Répartition des Foraminifères dans la coupe du sommet de la carrière Comte (Cassis, SE France).
Cliquer sur la miniature pour agrandir l'image.
Figure 2 : Répartition des Foraminifères dans la coupe des Tocchis (La Bédoule, SE France).
Cliquer sur la miniature pour agrandir l'image.
Figure 3 : Répartition des Foraminifères dans la coupe de la carrière de La Marcouline (Cassis, SE France).
Cliquer sur la miniature pour agrandir l'image.
Figure 4 : Répartition stratigraphique des principaux marqueurs aptiens (Foraminifères benthiques et planctoniques).
Cliquer sur la miniature pour agrandir l'image.
Figure 5 : Corrélation des trois coupes (Comte, Tocchis, Marcouline) permettant de reconstituer la séquence du passage du Bédoulien sommital au Gargasien basal. Limite entre les deux sous-étages fixée au banc 178 (cf. , 1994).
Cliquer sur la miniature pour agrandir l'image.
Figure 6 : Évolution au cours de l'Aptien du mode d'enroulement et du nombre des loges chez une lignée de Foraminifères planctoniques initialement à test trochospiralé et 6 loges au dernier tour.
![]()
Cliquer sur la miniature pour agrandir l'image.
Planche 1 :
1-9. "Globigerinelloides" gr. bizonae () –
saundersi (). 3 spécimens montrant divers modes d'enroulement (de trochospiralé à subplanispiralé). Gargasien basal, Zone à Luterbacheri, Coupe des Tocchis (La Bédoule), Niveau 189-T.
10-12. Hastigerinella bizonae, Holotype refiguré en microscopie optique (x 200), collection (Muséum National d'Histoire Naturelle, Paris). "Gargasien inférieur, région de Comte près Cassis", selon l'auteur de l'espèce (1961).
Cliquer sur la miniature pour agrandir l'image.
Planche 2 :
1-12. "Globigerinelloides" bizonae () –
saundersi (). 4 spécimens du Gargasien moyen, Zone à Barri, coupe de Cassis-La Marcouline, montrant également divers stades dans la dissymétrie de l'enroulement.
1-3. Échantillon 68. 4-12. Échantillon 78.
Cliquer sur la miniature pour agrandir l'image.
Planche 3 :
1-4. "Globigerinelloides" bizonae () –
saundersi (). Spécimen de la base du Gargasien supérieur, Zone à Algerianus, coupe de Cassis-La Marcouline, Échantillon 90.
4. détail de l'avant-dernière loge, montrant le départ du prolongement loculaire (brisé un peu au dessus de sa base).
5-10. Praehedbergella luterbacheri (). Spécimens du Gargasien basal, Zone à Luterbacheri, coupe de Cassis-La Marcouline, Échantillon 8.
Cliquer sur la miniature pour agrandir l'image.
Planche 4 :
1-6. Praehedbergella luterbacheri ().
1-3. Gargasien basal, Zone à Luterbacheri, coupe des Tocchis, niveau 183-T.
4-6. Gargasien moyen, Zone à Barri, coupe de Cassis-La Marcouline, Échantillon 78.
7-9. Exemplaire de transition entre Praehedbergella luterbacheri () et Globigerinelloides ferreolensis (). Gargasien inférieur, Base de la Zone à Ferreolensis, coupe de Cassis-La Marcouline, Échantillon 16.
10-12. Praehedbergella praetrocoidea ( et ), Gargasien inférieur, sommet de la Zone à Luterbacheri, coupe de Cassis-La Marcouline, Échantillon 14.
Cliquer sur la miniature pour agrandir l'image.
Planche 5 :
1-6. Praehedbergella praetrocoidea (
et ). 1-3. Gargasien inférieur, Zone à Ferreolensis, coupe de Cassis-La Marcouline, Échantillon 22.
4-6. Gargasien moyen, Zone à Barri, coupe de Cassis-La Marcouline, Échantillon 8.
7-12. Praehedbergella gorbachikae (), base du Gargasien supérieur, Zone à Algerianus, coupe de Cassis-La Marcouline, Échantillon 90.
![]()