![]()
![]()
J., É., F. & J. (2007).- Palynology of Baudour Clays Formation (Mons Basin, Belgium): correlation within the "stratotypic" Wealden. In: P. & E. (eds.), Recent Advances in Palynology.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/01, Abstract 03 (CG2007_M01/03)
![]() This article
is a palynological study of four clay samples from the dinosaur-bearing Wealden facies at Baudour (Belgium).
Their microflora is compared to that found at Bernissart. The palynomorphs of
the four samples are of continental origin. The botanical affinities of the palynological taxa are mainly ferns, gymnosperms and freshwater "algae". Also encountered are stratigraphically
important angiospermous-like pollen grains (biorecord Superret-croton, probable paleotaxon Superret-subcrot and biorecord Retisulc-dentat); the aperture of the former is questioned ("variant forms").
This article
is a palynological study of four clay samples from the dinosaur-bearing Wealden facies at Baudour (Belgium).
Their microflora is compared to that found at Bernissart. The palynomorphs of
the four samples are of continental origin. The botanical affinities of the palynological taxa are mainly ferns, gymnosperms and freshwater "algae". Also encountered are stratigraphically
important angiospermous-like pollen grains (biorecord Superret-croton, probable paleotaxon Superret-subcrot and biorecord Retisulc-dentat); the aperture of the former is questioned ("variant forms").
![]() Belgium; Wealden facies; Baudour; palynology; angiosperm pollen; dinosaur.
Belgium; Wealden facies; Baudour; palynology; angiosperm pollen; dinosaur.
![]() L'analyse palynologique du faciès wealdien à dinosaures de Baudour (Belgique) est exposée et la microflore est comparée à celle de Bernissart. Les palynomorphes des quatre échantillons d'argile sont d'origine continentale. Les affinités botaniques des taxons palynologiques se partagent essentiellement entre les fougères, les gymnospermes et les "algues"
d'eau douce. Des grains de pollen d'affinité angiospermienne possédant une importance stratigraphique ont été également observés
(biorecord Superret-croton, probables
paleotaxon Superret-subcrot et biorecord Retisulc-dentat) ; la nature de l'ouverture germinative du premier est discutée ("formes variantes").
L'analyse palynologique du faciès wealdien à dinosaures de Baudour (Belgique) est exposée et la microflore est comparée à celle de Bernissart. Les palynomorphes des quatre échantillons d'argile sont d'origine continentale. Les affinités botaniques des taxons palynologiques se partagent essentiellement entre les fougères, les gymnospermes et les "algues"
d'eau douce. Des grains de pollen d'affinité angiospermienne possédant une importance stratigraphique ont été également observés
(biorecord Superret-croton, probables
paleotaxon Superret-subcrot et biorecord Retisulc-dentat) ; la nature de l'ouverture germinative du premier est discutée ("formes variantes").
![]() Belgique ; faciès wealdien ; Baudour ; palynologie ; pollen d'angiosperme ; dinosaure.
Belgique ; faciès wealdien ; Baudour ; palynologie ; pollen d'angiosperme ; dinosaure.
![]() The Wealden facies of the Mons Basin (Belgium)
is well-known because of the exceptional fossil found in a coal mine at Bernissart
during the years 1878-1881. At depths of 322 and 356 meters at least 29 complete specimens of Iguanodon
were found, along with many fish, turtles, crocodiles, amphibians, insects and fragments of plants (see & ,
1990, for a complete list of references). The palynology of these sediments
has been reported on previously by and
(1955, 1956, 1959a,
1959b) and et alii
(1963). Recently et alii
(2005, 2006) and et alii
(in press) analyzed the palynological assemblage of the 322
meter level at Bernissart and focused their study on angiosperm pollen, found for the first time in
the Belgian Wealden facies. Bernissart has long been considered as the only site yielding dinosaur bones
from a Belgian Wealden facies. Recently, however, a fragment of dinosaur bone has been identified in the Wealden facies of the Baudour quarry, located in the Mons
Basin near Bernissart. We offer here the first palynological study of the Wealden facies
of the Baudour site, and we compare its palynological assemblage with that at Bernissart.
The Wealden facies of the Mons Basin (Belgium)
is well-known because of the exceptional fossil found in a coal mine at Bernissart
during the years 1878-1881. At depths of 322 and 356 meters at least 29 complete specimens of Iguanodon
were found, along with many fish, turtles, crocodiles, amphibians, insects and fragments of plants (see & ,
1990, for a complete list of references). The palynology of these sediments
has been reported on previously by and
(1955, 1956, 1959a,
1959b) and et alii
(1963). Recently et alii
(2005, 2006) and et alii
(in press) analyzed the palynological assemblage of the 322
meter level at Bernissart and focused their study on angiosperm pollen, found for the first time in
the Belgian Wealden facies. Bernissart has long been considered as the only site yielding dinosaur bones
from a Belgian Wealden facies. Recently, however, a fragment of dinosaur bone has been identified in the Wealden facies of the Baudour quarry, located in the Mons
Basin near Bernissart. We offer here the first palynological study of the Wealden facies
of the Baudour site, and we compare its palynological assemblage with that at Bernissart.
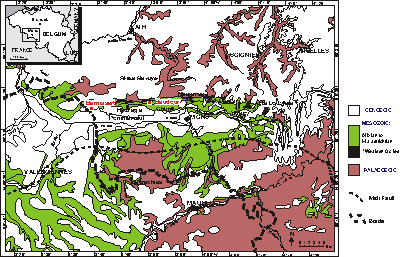
![]() The Baudour quarry is in the Mons Basin,
Belgium (Fig. 1
The Baudour quarry is in the Mons Basin,
Belgium (Fig. 1 ![]() ), which can be regarded as the
northeastern part of the Paris Basin
(see , 1970). The sedimentary succession of the Mons Basin begins with the Wealden facies and
continues with Middle-Upper Albian, Upper Cretaceous and Cenozoic deposits
(, 1970). The Wealden facies of Baudour is attributed to the Baudour Clays Formation
( et alii, 2001).
), which can be regarded as the
northeastern part of the Paris Basin
(see , 1970). The sedimentary succession of the Mons Basin begins with the Wealden facies and
continues with Middle-Upper Albian, Upper Cretaceous and Cenozoic deposits
(, 1970). The Wealden facies of Baudour is attributed to the Baudour Clays Formation
( et alii, 2001).
![]() For the purpose of palynological analysis, we processed four samples of grey clays from the collections of the Royal Belgian Institute of Natural Sciences (Brussels). The treatment
involves the destruction of minerals with hydrofluoric acid (70 %), followed by a filtration
using a 9 µm-mesh sieve.
For the purpose of palynological analysis, we processed four samples of grey clays from the collections of the Royal Belgian Institute of Natural Sciences (Brussels). The treatment
involves the destruction of minerals with hydrofluoric acid (70 %), followed by a filtration
using a 9 µm-mesh sieve.
![]() The morphological classification and nomenclature of and
(1954 and subsequent papers), as improved by
(1963), are followed herein. For the purpose in hand, another nomenclature sensu and collaborators (see complete reference list in ,
1994) was also used for four taxa, two of which are discussed below.
The morphological classification and nomenclature of and
(1954 and subsequent papers), as improved by
(1963), are followed herein. For the purpose in hand, another nomenclature sensu and collaborators (see complete reference list in ,
1994) was also used for four taxa, two of which are discussed below.
![]() The palynological assemblage of Baudour quarry is very well preserved, suggesting that 1) most of the mother-plants grew close to the place where the palynomorphs were buried, and 2) diagenesis was minor. This assemblage is
entirely continental in origin, quite similar to that at Bernissart, with the exception
of differences in the relative abundances of some taxa. Fern spores (e.g. Cicatricosisporites brevilaesuratus, C. hallei, Deltoidospora minor, Dictyophyllidites harrisii and Trilobosporites hannonicus) and bisaccate pollen grains are a main feature of both
assemblages; Sciadopityaceae (biorecord Hauterivian-cactisulc, alias Cerebropollenites sp.) and ephedroid pollen grains (Ephedripites montanaensis) are less
numerous, but the angiospermous biorecord Superret-croton and its "variant forms" are more abundant than in Bernissart assemblage; freshwater "algae" zygospores (Schizosporis reticulatus) are also present.
The palynological assemblage of Baudour quarry is very well preserved, suggesting that 1) most of the mother-plants grew close to the place where the palynomorphs were buried, and 2) diagenesis was minor. This assemblage is
entirely continental in origin, quite similar to that at Bernissart, with the exception
of differences in the relative abundances of some taxa. Fern spores (e.g. Cicatricosisporites brevilaesuratus, C. hallei, Deltoidospora minor, Dictyophyllidites harrisii and Trilobosporites hannonicus) and bisaccate pollen grains are a main feature of both
assemblages; Sciadopityaceae (biorecord Hauterivian-cactisulc, alias Cerebropollenites sp.) and ephedroid pollen grains (Ephedripites montanaensis) are less
numerous, but the angiospermous biorecord Superret-croton and its "variant forms" are more abundant than in Bernissart assemblage; freshwater "algae" zygospores (Schizosporis reticulatus) are also present.
![]() Many
previous studies dealt with the palynology of the Wealden facies of northern Europe ( & ,
1955, 1956, 1959a,
1959b; , 1955,
1976, 1994; ,
1958; , 1958; et alii,
1963; , 1964; ,
1964, 1965, 1966; ,
1966; & ,
1967; , 1969; ,
1970; , 1971; ,
1973; et alii, 1976; ,
1977; et alii,
1979; & ,
1987, 1990, 1994). The palynological assemblage of
the Baudour quarry is typically "Wealden" in aspect: many of the palynomorphs identified have been reported elsewhere by and
(1955), et alii
(1963) and others.
Many
previous studies dealt with the palynology of the Wealden facies of northern Europe ( & ,
1955, 1956, 1959a,
1959b; , 1955,
1976, 1994; ,
1958; , 1958; et alii,
1963; , 1964; ,
1964, 1965, 1966; ,
1966; & ,
1967; , 1969; ,
1970; , 1971; ,
1973; et alii, 1976; ,
1977; et alii,
1979; & ,
1987, 1990, 1994). The palynological assemblage of
the Baudour quarry is typically "Wealden" in aspect: many of the palynomorphs identified have been reported elsewhere by and
(1955), et alii
(1963) and others.
![]() Because the biorecord Superret-croton, its "variant forms", the probable paleotaxon Superret-subcrot and
the probable biorecord Retisulc-dentat are discussed below, a reminder of
meanings of biorecord and paleotaxon is pertinent. Some palynomorphs have names
that do not conform with classic palynological terminology: they are biorecords, sensu
(1976) and et alii
(1979), reiterated by
(1991). Morphological taxa and biorecords are not true
Linnean taxa, but para-taxa or morphotypes which are sporae dispersae
with no specific relationship to a mother-plant; in both biorecord and
paleotaxon the forms are classified in an organizing chart developed in
accordance with a descriptive pattern (a biorecord is generally more rigorously defined). A biorecord is considered here as a fundamental reference taxon, equivalent to
the taxa defined through the use of the conventional morphological scheme (,
1976, p. 26; et alii,
1979, p. 515); the word paleotaxon is here used in accordance with '
(1994) meaning, as qualifying a less rigorously defined taxon.
Because the biorecord Superret-croton, its "variant forms", the probable paleotaxon Superret-subcrot and
the probable biorecord Retisulc-dentat are discussed below, a reminder of
meanings of biorecord and paleotaxon is pertinent. Some palynomorphs have names
that do not conform with classic palynological terminology: they are biorecords, sensu
(1976) and et alii
(1979), reiterated by
(1991). Morphological taxa and biorecords are not true
Linnean taxa, but para-taxa or morphotypes which are sporae dispersae
with no specific relationship to a mother-plant; in both biorecord and
paleotaxon the forms are classified in an organizing chart developed in
accordance with a descriptive pattern (a biorecord is generally more rigorously defined). A biorecord is considered here as a fundamental reference taxon, equivalent to
the taxa defined through the use of the conventional morphological scheme (,
1976, p. 26; et alii,
1979, p. 515); the word paleotaxon is here used in accordance with '
(1994) meaning, as qualifying a less rigorously defined taxon.
![]() The goal
of these concepts is to designate precisely what is recognized under the microscope,
and so to make it useful for stratigraphic purposes; though the philosophies of "palynological language" may appear different on
either side of the English Channel - as they concern the Lower Cretaceous - these "languages" have been so useful and
so firmly established after several decades that there is no reason to reject one for the other, all the more that both may comply with the procedures
of the Linnean Code [see
(1976, p. 29), who concludes as follows: "the two can be separate and complementary"].
The goal
of these concepts is to designate precisely what is recognized under the microscope,
and so to make it useful for stratigraphic purposes; though the philosophies of "palynological language" may appear different on
either side of the English Channel - as they concern the Lower Cretaceous - these "languages" have been so useful and
so firmly established after several decades that there is no reason to reject one for the other, all the more that both may comply with the procedures
of the Linnean Code [see
(1976, p. 29), who concludes as follows: "the two can be separate and complementary"].
![]() These three taxa (biorecord Superret-croton, probable paleotaxon Superret-subcrot and probable biorecord Retisulc-dentat) are of stratigraphic importance in the Baudour palynological assemblage.
These three taxa (biorecord Superret-croton, probable paleotaxon Superret-subcrot and probable biorecord Retisulc-dentat) are of stratigraphic importance in the Baudour palynological assemblage.
![]() Biorecord Superret-croton
(Figs. 2
Biorecord Superret-croton
(Figs. 2 ![]() ,
3
,
3 ![]() ,
4
,
4 ![]() ,
5
,
5 ![]() ,
6
,
6 ![]() ,
7
,
7 ![]() &
8
&
8 ![]() ) was defined by et alii
(1979). In the "stratotypic" Wealden facies of the Weald and Wessex
sub-basins, which is dated by interbeds with ammonites and dinoflagellate cysts (,
1986, 1990), the stratigraphic distribution of biorecord Superret-croton is attributed to MCT (Monosulcate Columellate Tectate) phases 3 to 5, ranging from the middle Barremian to the earliest Aptian (,
1994). et alii
(2004) therefore suggested a middle Barremian to earliest Aptian age for the sediments of Bernissart (at a depth of 322 m) and the same dating is proposed here for the Baudour Clays Formation. In the description of the species Stellatopollis hughesii, from the Upper Barremian (?) of Egypt,
(1986) regarded it equivalent to biorecord Superret-croton.
) was defined by et alii
(1979). In the "stratotypic" Wealden facies of the Weald and Wessex
sub-basins, which is dated by interbeds with ammonites and dinoflagellate cysts (,
1986, 1990), the stratigraphic distribution of biorecord Superret-croton is attributed to MCT (Monosulcate Columellate Tectate) phases 3 to 5, ranging from the middle Barremian to the earliest Aptian (,
1994). et alii
(2004) therefore suggested a middle Barremian to earliest Aptian age for the sediments of Bernissart (at a depth of 322 m) and the same dating is proposed here for the Baudour Clays Formation. In the description of the species Stellatopollis hughesii, from the Upper Barremian (?) of Egypt,
(1986) regarded it equivalent to biorecord Superret-croton.
![]() Another peculiar palynomorph, very rare, is the probable paleotaxon Superret-subcrot (Fig. 16
Another peculiar palynomorph, very rare, is the probable paleotaxon Superret-subcrot (Fig. 16 ![]() ), which was defined by et alii
[1979, sic: SUPERRET-(CAND)SUBCROT] and
reexamined by
(1994), using a scanning electron microscope. It is difficult to recognize with certainty under light microscopy; however its morphology appears to
make it identifiable (shape, size, wall stratification and very small, numerous and closely packed crotonoid sculptural elements which
are at the limit of the microscope's resolving power, so it is very difficult to produce convincing photographs). In the English Wealden, paleotaxon Superret-subcrot is reported from MCT phase 4, apparently restricted to one locality (Kingsclere borehole at a depth of 474 feet). Nevertheless, its scarcity renders impossible a more refined stratigraphic correlation than that mentioned above.
), which was defined by et alii
[1979, sic: SUPERRET-(CAND)SUBCROT] and
reexamined by
(1994), using a scanning electron microscope. It is difficult to recognize with certainty under light microscopy; however its morphology appears to
make it identifiable (shape, size, wall stratification and very small, numerous and closely packed crotonoid sculptural elements which
are at the limit of the microscope's resolving power, so it is very difficult to produce convincing photographs). In the English Wealden, paleotaxon Superret-subcrot is reported from MCT phase 4, apparently restricted to one locality (Kingsclere borehole at a depth of 474 feet). Nevertheless, its scarcity renders impossible a more refined stratigraphic correlation than that mentioned above.
![]() Lastly, another peculiar palynomorph, also very rare, is the probable biorecord Retisulc-dentat (Fig. 17
Lastly, another peculiar palynomorph, also very rare, is the probable biorecord Retisulc-dentat (Fig. 17 ![]() ), which was defined by et alii
(1979) then again by (1994). In the English Wealden, the stratigraphic distribution of biorecord Retisulc-dentat (and similar forms) is attributed to MCT phases 2 to 4; its occurrence in the Baudour Clays Formation could be an argument to rule out the MCT phase
5. However, because of its rarety such a conclusion would be rash.
), which was defined by et alii
(1979) then again by (1994). In the English Wealden, the stratigraphic distribution of biorecord Retisulc-dentat (and similar forms) is attributed to MCT phases 2 to 4; its occurrence in the Baudour Clays Formation could be an argument to rule out the MCT phase
5. However, because of its rarety such a conclusion would be rash.
![]() et alii
(1979) and (1994) highlighted the angiospermid characters of these three reference
taxa: essentially they are the tectum structure (continuous reticulum) and the supratectal elements (particularly the crotonoid pattern exhibited by biorecord
Superret-croton and paleotaxon Superret-subcrot). The crotonoid supratectal pattern is known
in the following extant families: Euphorbiaceae (,
1952; , 1962; ,
1966; , 1975), Buxaceae and Thymeleaceae (,
1952; , 1966), Atherospermataceae ( et alii,
1975) and Liliaceae (, 1952; ,
1966; , 1970).
Last, the biorecord Retisulc-dentat is very similar to pollen grains of the extant
Chloranthaceae family (see ,
1987).
et alii
(1979) and (1994) highlighted the angiospermid characters of these three reference
taxa: essentially they are the tectum structure (continuous reticulum) and the supratectal elements (particularly the crotonoid pattern exhibited by biorecord
Superret-croton and paleotaxon Superret-subcrot). The crotonoid supratectal pattern is known
in the following extant families: Euphorbiaceae (,
1952; , 1962; ,
1966; , 1975), Buxaceae and Thymeleaceae (,
1952; , 1966), Atherospermataceae ( et alii,
1975) and Liliaceae (, 1952; ,
1966; , 1970).
Last, the biorecord Retisulc-dentat is very similar to pollen grains of the extant
Chloranthaceae family (see ,
1987).
![]() After examination of numerous grains (several
hundred) from the Baudour quarry samples, the presence of a definite sulcus is now
certain, at least in a significant number: biorecord Superret-croton is undoubtedly monosulcate (Figs. 2
After examination of numerous grains (several
hundred) from the Baudour quarry samples, the presence of a definite sulcus is now
certain, at least in a significant number: biorecord Superret-croton is undoubtedly monosulcate (Figs. 2 ![]() ,
3
,
3 ![]() ,
4
,
4 ![]() ,
5
,
5 ![]() ,
6
,
6 ![]() &
7
&
7 ![]() ). However, other grains
with the same overall appearance (similar size and crotonoid pattern) differ
with respect to the germinal aperture; these grains are here designed as "variant forms":
). However, other grains
with the same overall appearance (similar size and crotonoid pattern) differ
with respect to the germinal aperture; these grains are here designed as "variant forms":
![]() - a large proportion of these grains are
undoubtedly inaperturate, their shape being almost spherical instead of ellipsoidal
(Figs. 9
- a large proportion of these grains are
undoubtedly inaperturate, their shape being almost spherical instead of ellipsoidal
(Figs. 9 ![]() ,
10
,
10 ![]() ,
11
,
11 ![]() &
12
&
12 ![]() );
);
![]() - some very rare grains seem to be tricolpate
(Figs. 13
- some very rare grains seem to be tricolpate
(Figs. 13 ![]() & 14
& 14 ![]() )
or are clearly trichotomosulcate (Fig. 15
)
or are clearly trichotomosulcate (Fig. 15 ![]() ); these two early "variant forms" herald the diversification
of apertures that began in strata of Early Aptian age.
); these two early "variant forms" herald the diversification
of apertures that began in strata of Early Aptian age.
![]() These three "variant forms" may
be due to some morphological intra-specific variability (,
1987), or on the contrary may indicate the existence of different
species of mother-plant.
These three "variant forms" may
be due to some morphological intra-specific variability (,
1987), or on the contrary may indicate the existence of different
species of mother-plant.
![]() In
any event, as the grains are not monosulcate, these three "variant forms" cannot
be referred to Stellatopollis hughesii, or to the
Superret- genusbox (whichever "palynological language" is used). In order to remain logical and to
avoid confusion, ' data-handling system (1976 and subsequent papers) must obviously be
applied; these "variant forms" require discrete genusboxes which respectively
are: Inapert- [proposed here, in accordance with the recommendations of et alii
(1979)], and Retitri- and Trichoto- (both established by ,
1991). Logically too, trivial names can remain as -croton; thus, these "variant forms" belong respectively to these new taxa: biorecord
Inapert-croton, biorecord (cand) Retitri-croton and biorecord (cand)
Trichoto-croton ["cand" (for candidate) between brackets means that too few specimens are available to be acceptable as a full
biorecord, after et alii
(1979) and (1991)].
In
any event, as the grains are not monosulcate, these three "variant forms" cannot
be referred to Stellatopollis hughesii, or to the
Superret- genusbox (whichever "palynological language" is used). In order to remain logical and to
avoid confusion, ' data-handling system (1976 and subsequent papers) must obviously be
applied; these "variant forms" require discrete genusboxes which respectively
are: Inapert- [proposed here, in accordance with the recommendations of et alii
(1979)], and Retitri- and Trichoto- (both established by ,
1991). Logically too, trivial names can remain as -croton; thus, these "variant forms" belong respectively to these new taxa: biorecord
Inapert-croton, biorecord (cand) Retitri-croton and biorecord (cand)
Trichoto-croton ["cand" (for candidate) between brackets means that too few specimens are available to be acceptable as a full
biorecord, after et alii
(1979) and (1991)].
![]() In the Mons Basin, the Wealden facies occur ( et alii,
2001):
In the Mons Basin, the Wealden facies occur ( et alii,
2001):
![]() - in kilometric outcrops (also called "pockets")
and in the northern part of this basin as shallow subcrops,
- in kilometric outcrops (also called "pockets")
and in the northern part of this basin as shallow subcrops,
![]() -
as the infilling of several sinkholes developed in carbonates and siliciclastic sediments (for example at
Bernissart),
-
as the infilling of several sinkholes developed in carbonates and siliciclastic sediments (for example at
Bernissart),
![]() - in the
east as white sands and sandstones containing lignite and glauconitic material.
- in the
east as white sands and sandstones containing lignite and glauconitic material.
![]() Recently et alii
(2005, 2006) demonstrated that the Wealden facies of the natural pit of Bernissart are
of middle Barremian to earliest Aptian in age. A similar age is proposed for the Wealden facies of the
"Baudour pocket". If so, subsidence responsible for both the Baudour pocket and the natural pit of Bernissart
was contemporaneous and perhaps caused by the same genetic conditions. Deep boreholes drilled in the Mons Basin demonstrated partial and local dissolution of buried anhydrites immediately below the natural pits ( et alii,
1982; , 1989; & ,
1989; et alii, 1993). As in other areas, the result of this partial dissolution of deeply buried anhydrites produces local
repeated collapse in overlying rocks; this may cause geomorphological depressions
or sinkholes at the surface (,
1966; , 1996; ,
1999). So natural pits and "pockets" may be the surface
expression of deep dissolution.
Recently et alii
(2005, 2006) demonstrated that the Wealden facies of the natural pit of Bernissart are
of middle Barremian to earliest Aptian in age. A similar age is proposed for the Wealden facies of the
"Baudour pocket". If so, subsidence responsible for both the Baudour pocket and the natural pit of Bernissart
was contemporaneous and perhaps caused by the same genetic conditions. Deep boreholes drilled in the Mons Basin demonstrated partial and local dissolution of buried anhydrites immediately below the natural pits ( et alii,
1982; , 1989; & ,
1989; et alii, 1993). As in other areas, the result of this partial dissolution of deeply buried anhydrites produces local
repeated collapse in overlying rocks; this may cause geomorphological depressions
or sinkholes at the surface (,
1966; , 1996; ,
1999). So natural pits and "pockets" may be the surface
expression of deep dissolution.
![]() The palynological content of the dinosaur-bearing Wealden facies of the Baudour Clays Formation is reported for the first time. The palynomorphs are of continental origin. Ferns and gymnosperms are the dominant floristic groups. The occurrence of pollen grains of the angiospermous biorecord
Superret-croton, probable paleotaxon Superret-subcrot and probable biorecord Retisulc-dentat [sensu et alii
(1979) and (1994)]
denotes a middle Barremian to earliest Aptian age for these sediments, and so
they are confirmed to be of the same age as the Wealden facies sediments in the natural pit of
Bernissart. This identity in age suggests that the natural pit of Bernissart and the "pocket" of Baudour
are the result of the same mechanisms of subsidence, probably thus demonstrating
a genetic relationship between natural pits and relatively large "pockets" in the Mons Basin.
The palynological content of the dinosaur-bearing Wealden facies of the Baudour Clays Formation is reported for the first time. The palynomorphs are of continental origin. Ferns and gymnosperms are the dominant floristic groups. The occurrence of pollen grains of the angiospermous biorecord
Superret-croton, probable paleotaxon Superret-subcrot and probable biorecord Retisulc-dentat [sensu et alii
(1979) and (1994)]
denotes a middle Barremian to earliest Aptian age for these sediments, and so
they are confirmed to be of the same age as the Wealden facies sediments in the natural pit of
Bernissart. This identity in age suggests that the natural pit of Bernissart and the "pocket" of Baudour
are the result of the same mechanisms of subsidence, probably thus demonstrating
a genetic relationship between natural pits and relatively large "pockets" in the Mons Basin.
![]() This study has been partly founded by
F.N.R.S. (Fonds National de la Recherche Scientifique de Belgique). The authors wish to thank Marcella for
processing the rock samples, Christiane for her assistance with the scanning electron
microscope and Nestor for the improvement of
this English text.
This study has been partly founded by
F.N.R.S. (Fonds National de la Recherche Scientifique de Belgique). The authors wish to thank Marcella for
processing the rock samples, Christiane for her assistance with the scanning electron
microscope and Nestor for the improvement of
this English text.
![]() The slides and glass stubs will be housed in the Institut Royal des Sciences Naturelles (Brussels).
The slides and glass stubs will be housed in the Institut Royal des Sciences Naturelles (Brussels).
![]() Note about
Figs. 2
Note about
Figs. 2 ![]() ,
3
,
3 ![]() ,
4
,
4 ![]() ,
5
,
5 ![]() ,
6
,
6 ![]() ,
9
,
9 ![]() ,
13
,
13 ![]() ,
14
,
14 ![]() ,
15
,
15 ![]() and
16
and
16 ![]() :
interferencial- differential contrast after , unmodified photographs; the position of the nine illustrated palynomorphs on each slide is given after the "England Finder".
:
interferencial- differential contrast after , unmodified photographs; the position of the nine illustrated palynomorphs on each slide is given after the "England Finder".
H.W.J. Van, G.F.W. & B.J. (1976).- Palaeobotanical and palynological investigation with notes on the microfauna of some core samples from the Lower Cretaceous in the West Netherlands Basin.- Mededelingen Rijks geologischen Dienst, Maastricht, vol. 27, n° 2, p. 41-79.
D.B. (1966).- The exine stellate sculpture of pollen grains (in Russian).- In: The importance of palynological analysis for the stratigraphical and paleofloristic investigations. 2nd International palynological Conference (Utrecht).- SSSR Academy of Sciences "Nauka", Moscow, p. 22-26.
D.J. (1973).- Palynology of Early Cretaceous soil beds and associated strata.- Palaeontology, London, vol. 16, n° 2, p. 399-424.
D. (1966).- Palynology of uppermost Jurassic and lowermost Cretaceous strata in the eastern Netherlands.- Leidse geologische Mededelingen, Leiden, vol. 35, p. 209-276.
J.L. (1987).- Comparison of Chloranthaceae pollen with the Cretaceous "Clavatipollenites complex". Taxonomic implications for palaeopalynology.- Pollen et Spores, Paris, vol. 29, n° 2-3, p. 249-272.
R.A. (1958).- British Mesozoic microspores and pollen grains, a systematic and stratigraphic study.- Palaeontographica, Stuttgart, vol. B 103, n° 4-6, p. 75-179.
J., D. & J. (in press).- Palynology of the dinosaur-bearing Wealden facies in the natural pit of Bernissart (Belgium).- Review of Palaeobotany and Palynology, Amsterdam.
A. & G. (1955).- Les spores et grains de pollen du Wealdien du Hainaut.- Mémoires de la Société belge de Géologie, de Paléontologie et d'Hydrologie, Brussels, vol. 5, p. 1-73.
A.F. & G. (1956).- Présence d'Eucommiidites dans une coupe du Wealdien de Baudour.- Bulletin de la Société belge de Géologie, Paléontologie et Hydrologie, Brussels, vol. 65, p. 375-380.
A. & G. (1959a).- Spores, grains de pollen, Hystrichosphères et Péridiniens dans le Wealdien de Féron-Glageon.- Annales de la Société Géologique du Nord, Lille, vol. 79, p. 29-64.
A. & G. (1959b).- Curieuse association de microfossiles dans le Wealdien de Féron-Glageon (France).- Bulletin de la Société belge de Géologie, Paléontologie et Hydrologie, Brussels, vol. 68, p. 122-125.
A.F., M.E. & N.F. (1963).- Revision of some Lower Cretaceous microspores from Belgium.- Palaeontology, London, vol. 6, p. 282-292.
A. (1989).- Description, genèse et âge des "puits naturels" du Hainaut.- Tunnels et ouvrages souterrains, Brussels, vol. 93, p. 114-116.
A., V., R. & F. (1982).- La géothermie en Hainaut et le sondage de Ghlin (Mons, Belgique).- Annales de la Société géologique du Nord, Lille, vol. 101, p. 189-206.
M.E. (1963).- Upper Mesozoic microfloras from south-eastern Australia.- Proceedings of the Royal Society of Victoria, Melbourne, vol. 77, n° 1, p. 1-148.
G. (1977).- Palynologie und Stratigraphie der Bückeberg - Formation (Berriasium - Valanginium) in der Hilsmulde (NW - Deutschland).- Geologisches Jahrbuch, Stuttgart, vol. A 42, p. 3-122.
H. (1964).- Trilete Sporen aus dem Oberen Jura und dem Wealden Norddeutschlands.- Geologie, Berlin, vol. 13, n° 9, p. 1099-1129.
H. (1965).- Die sporenpaläontologische Gliederung des Wealden in Westmecklenburg (Struktur Werle).- Geologie, Berlin, vol. 14, n° 47, p. 1-118.
H. (1966).- Sporenstratigraphischer Vergleich zwischen dem Wealden Norddeutschlands und Südenglands.- Geologie, Berlin, vol. 15, n° 55, p. 102-129.
J., M. & B. (1975).- Observations on exine structure of Eucommiidites and Lower Cretaceous Angiosperm pollen.- Pollen et Spores, Paris, vol. 17, n° 3, p. 429-486.
C. & S. (1989).- Tectonique et karstification profonde: un modèle de subsidence original pour le Bassin de Mons.- Annales de la Société géologique de Belgique, Brussels, vol. 112, p. 479-487.
G. (1952).- Pollen morphology and plant taxonomy - Angiosperms. An introduction to palynology.- Almqvist & Wiskell, Stockholm, 539 p.
F. (1996).- Gypsum karstification induced subsidence: effects on alluvial systems and derived geohazards (Calatayud Graben, Iberian Range, Spain).- Geomorphology, Amsterdam, vol. 16, p. 277-293.
I.C. (1986).- An Early Cretaceous dinocyst assemblage from the Wealden of Southern England.- Special Papers in Palaeontology, London, vol. 35, p. 95-109.
I.C. (1990).- A dinocyst calibration of the European Boreal Barremian.- Palaeontographica, Stuttgart, vol. B 218, p. 1-76.
G.F.W. (1971).- Palynology of a Wealden section (Lower Cretaceous) in the "Carrière de Longueville", the Boulonnais (France).- Review of Palaeobotany and Palynology, Amsterdam, vol. 12, p. 271-302.
N.F. (1955).- Wealden plant microfossils.- Geological Magazine, London, vol. 92, n° 3, p. 201-217.
N.F. (1976).- Palaeobiology of angiosperm origins.- Cambridge University Press, Cambridge, 242 p.
N.F. (1994).- The enigma of angiosperm origins.- Cambridge University Press, Cambridge, 303 p.
N.F. & , A.B. (1987).- Records of angiospermid pollen entry into the English Early Cretaceous succession.- Review of Palaeobotany and Palynology, Amsterdam, vol. 50, p. 255-272.
N.F. & A.B. (1990).- New Wealden correlation for the Wessex Basin.- Proceedings of the Geologists' Association, London, vol. 101, p. 85-90.
N.F. & A.B. (1994).- Search for antecedents of Early Cretaceous monosulcate columellate pollen.- Review of Palaeobotany and Palynology, Amsterdam, vol. 83, p. 175-183.
N.F. & J.C. (1967).- Palynological facies and correlation in the English Wealden.- Review of Palaeobotany and Palynology, Amsterdam, vol. 1, p. 259-268.
N.F., G.E. & J.F. (1979).- Barremian earliest angiosperm pollen.- Palaeontology, London, vol. 22, n° 3, p. 513-535.
E.M. (1970).- Aptian and Albian miospores from southern England.- Palaeontographica, Stuttgart, vol. B 131, n° 1-4, p 73-143.
J. (1958).- Étude palynologique de quelques échantillons mésozoïques du Dorset (Grande-Bretagne).- Revue de l'Institut français du pétrole, Paris, vol. 13, n° 6, p. 917-943.
R. (1970).- Géologie du Bassin de Mons.- Annales de la Société géologique du Nord, Lille, vol. 40, n° 4, p. 171-189.
F. & P. (1990).- The Iguanodons of Bernissart.- Publication de l'Institut royal des Sciences naturelles de Belgique, Brussels, 51 p.
G. (1970).- Palynological evidence on early differenciation of Angiosperms.- Biological Reviews of the Cambridge Philosophical Society, Cambridge, vol. 45, p. 417-450.
G. (1969).- Miospores from the Purbeck Beds and marine Upper Jurassic of Southern England.- Palaeontology, London, vol. 12, n° 4, p. 574-620.
J.H.J. (1986).- An Early Cretaceous angiosperm pollen assemblage from Egypt.- Special Papers in Palaeontology, London, vol. 35, p. 121-134.
J.H.J. (1991).- Early Cretaceous angiosperm pollen from the borehole Mersa Matruh 1, North West Desert, Egypt.- Palaeontographica, Stuttgart, vol. B 222, p. 31-88.
R. & G. (1954).- Die Gattungen der paläozoischen Sporae dispersae und ihre Stratiraphie.- Geologisches Jahrbuch, Hannover, vol. 69, p. 111-194.
W. (1962).- Pollen morphology of the Euphorbiaceae with special reference to taxonomy.- Wentia (Acta Botanica Neerlandica), Amsterdam, vol. 7, 116 p.
F., A. & J.W.M. (2001).- Cretaceous lithostratigraphic units (Belgium). In: P. & L. (eds.), Guide to a revised lithostratigraphic scale of Belgium.- Geologica Belgica, Brussels, vol. 4, n° 1-2, p. 121-134.
J.-M., E. & A. (1993).- Dislocation des formations évaporitiques par la tectonique et la dissolution: le modèle des évaporites dinantiennes du domaine varisque franco-belge.- Bulletin de la Société géologique de France, Paris, vol. 164, p. 39-50.
R.J. (1966).- The solution brecciation process.- Bulletin of the Geological Society of America, Boulder, vol. 77, p. 843-848.
G. (1964).- Eine unterkretazische Mikroflora aus dem nördlichen Sauerland.- Fortschritte in der Geologie von Rheinland und Westfalen, Krefeld, vol. 12, p. 117-168.
J. (1999).- Evaporites.- Blackwell Press, London, 438 p.
J., J., D., C. & P. (2005).- Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique).- Comptes Rendus Palevol, Paris, vol. 4, p. 135-150.
J., J., D., L. & P. (2006).- The iguanodons of Bernissart (Belgium) are middle Barremian to earliest Aptian in age.- Bulletin de l'Institut des Sciences Naturelles de Belgique, Brussels, vol. 76, p. 91-95.
J., D. & J. (2004).- Palynological study of the dinosaurs-bearing Wealden facies sediments of Bernissart (Belgium).- XI Palynological International Congress, Granada.- Pollen, Granada, vol. 14, p. 177-178.
J., P., C., M., S., J.-M., J.-P., P., M. & C. (2005).- Description et implications géologiques préliminaires d'un forage carotté dans le "Cran aux Iguanodons" de Bernissart.- Geologica Belgica, Brussels, vol. 8, p. 43-49.
J.P. (1975).- Observation du pollen de Manihot sp. (Euphorbiaceae) au microscope électronique à balayage.- Bulletin de la Société botanique de France, Paris, vol. 122, p. 131-133.
The taxa named here follow the morphological scheme instituted by and (1954) and subsequently amplified by several palynologists. Biorecords and subordinate records are included, for in the authors' opinion there is no incompatibility .
Anteturma Proximegerminantes , 1970
![]() Turma Triletes-Azonales (, 1881) et , 1954
Turma Triletes-Azonales (, 1881) et , 1954
![]()
![]()
![]() Subturma Azonotriletes (,1935) , 1963
Subturma Azonotriletes (,1935) , 1963
![]()
![]()
![]()
![]() Infraturma Laevigati, Quasilaevigati ( et , 1886) , 1956
Infraturma Laevigati, Quasilaevigati ( et , 1886) , 1956
Biretisporites potoniaei ( et , 1955) , et , 1963
Deltoidospora australis (, 1953) , 1970
Deltoidospora minor (, 1953) , 1970
Dictyophyllidites harrisii , 1958
Matonisporites equiexinus , 1958
![]()
![]()
![]()
![]() Infraturma Apiculati ( et , 1886) , 1956
Infraturma Apiculati ( et , 1886) , 1956
![]()
![]()
![]()
![]()
![]() Subinfraturma Baculati et , 1957
Subinfraturma Baculati et , 1957
Pilosisporites trichopapillosus (, 1949) et , 1955
![]()
![]()
![]()
![]()
![]() Subinfraturma Granulati, Scabrati et , 1957
Subinfraturma Granulati, Scabrati et , 1957
Concavissimisporites verrucosus ( et , 1955) , et , 1963
![]()
![]()
![]()
![]()
![]() Subinfraturma Verrucati et , 1957
Subinfraturma Verrucati et , 1957
Leptolepidites tumulosus (, 1964) , 1975
![]()
![]()
![]()
![]() Infraturma Murornati et , 1954
Infraturma Murornati et , 1954
Cicatricosisporites baconicus , 1963
Cicatricosisporites brevilaesuratus (, 1958) , 1970
Cicatricosisporites hallei et , 1955
Cicatricosisporites hughesi , 1963
Foveotriletes subtriangularis (, 1963) et , 1970
Klukisporites foveolatus , 1964
Retitriletes austroclavatidites (, 1953) , , et in , 1963
![]()
![]()
![]() Subturma Lagenotriletes et , 1954
Subturma Lagenotriletes et , 1954
![]()
![]()
![]()
![]() Infraturma Trifoliati, Barbati , 1970
Infraturma Trifoliati, Barbati , 1970
Dijkstraisporites helios (Dijkstra, 1951) , 1956
![]() Turma Triletes-Zonales ( et , 1886) , 1956
Turma Triletes-Zonales ( et , 1886) , 1956
![]()
![]()
![]() Subturma Auritotriletes et , 1954
Subturma Auritotriletes et , 1954
![]()
![]()
![]()
![]() Infraturma Auriculati (, 1938) ,1963
Infraturma Auriculati (, 1938) ,1963
Rouseisporites reticulatus , 1962
Trilobosporites apiverrucatus , 1958
Trilobosporites hannonicus ( et , 1955) , 1956
![]()
![]()
![]()
![]() Infraturma Appendiciferi , 1956
Infraturma Appendiciferi , 1956
Appendicisporites erdtmanii , 1965
Appendicisporites stylosus (, 1954) , 1963
![]()
![]()
![]()
![]() Infraturma Tricrassati , 1963
Infraturma Tricrassati , 1963
Camarozonosporites insignis , 1967
Sestrosporites pseudoalveolatus (, 1958) , 1963
![]()
![]()
![]() Subturma Zonotriletes , 1935
Subturma Zonotriletes , 1935
![]()
![]()
![]()
![]() Infraturma Cingulati ( et , 1954) , 1963
Infraturma Cingulati ( et , 1954) , 1963
Foraminisporis asymmetricus ( et , 1958) , 1963
![]()
![]()
![]()
![]()
![]() Subinfraturma Euzonati, Coronati , 1970
Subinfraturma Euzonati, Coronati , 1970
Aequitriradites spinulosus ( et , 1958) et , 1961
![]() Turma Vestitriletes , 1970
Turma Vestitriletes , 1970
![]()
![]() Suprasubturma Perinotrilites (, 1947) , 1963
Suprasubturma Perinotrilites (, 1947) , 1963
Crybelosporites pannuceus (, 1963) , 1975
Anteturma Variegerminantes , 1970
![]() Turma Saccites , 1947
Turma Saccites , 1947
![]()
![]()
![]() Subturma Monosaccites (, 1951) et , 1954
Subturma Monosaccites (, 1951) et , 1954
![]()
![]()
![]()
![]() Infraturma Saccizonati , 1957
Infraturma Saccizonati , 1957
Applanopsis dampieri (, 1957) , 1961
![]()
![]()
![]() Subturma Disaccites , 1947
Subturma Disaccites , 1947
Parvisaccites radiatus , 1958
![]() Turma Aletes et Kryptoperturates , 1966
Turma Aletes et Kryptoperturates , 1966
![]()
![]()
![]() Subturma Azonoletes (, 1935) et , 1954
Subturma Azonoletes (, 1935) et , 1954
![]()
![]()
![]()
![]() Infraturma Granulonapiti, Retinapiti (, 1947) , 1975
Infraturma Granulonapiti, Retinapiti (, 1947) , 1975
Araucariacites australis (, 1947) , 1953
Biorecord Inapert-croton (this study)
![]()
![]()
![]()
![]() Infraturma Circumpollini (, 1953) , 1960
Infraturma Circumpollini (, 1953) , 1960
Classopollis sp.
![]() Turma Plicates (, 1939) , 1960
Turma Plicates (, 1939) , 1960
![]()
![]()
![]() Subturma Costates , 1970
Subturma Costates , 1970
![]()
![]()
![]()
![]() Infraturma Costati , 1970
Infraturma Costati , 1970
Ephedripites montanaensis , 1968
Ephedripites zaklinskaiae et , 1974
![]()
![]()
![]() Subturma Trichotomocolpates, Trichotomosulcates (, 1954) , 1970
Subturma Trichotomocolpates, Trichotomosulcates (, 1954) , 1970
![]()
![]()
![]()
![]() Infraturma Eutrichotomosulcati , 1970
Infraturma Eutrichotomosulcati , 1970
Biorecord (cand) Trichoto-croton (this study)
![]()
![]()
![]() Subturma Monocolpates (Monosulcites) & Zonocolpates , 1970
Subturma Monocolpates (Monosulcites) & Zonocolpates , 1970
![]()
![]()
![]()
![]() Infraturma Quasilaevigati & Microsculptati , 1970
Infraturma Quasilaevigati & Microsculptati , 1970
Cycadopites minimus (, 1947) , 1970
![]()
![]()
![]()
![]() Infraturma Reticulati , 1988
Infraturma Reticulati , 1988
Biorecord Retisulc-dentat (in , et , 1979)
![]()
![]()
![]()
![]() Infraturma Sculptati , 1970
Infraturma Sculptati , 1970
Biorecord Hauterivian-cactisulc (in et , 1987)
Biorecord Superret-croton (in , et , 1979)
Paleotaxon Superret-subcrot (in , et , 1979)
![]()
![]()
![]() Subturma Tricolpates, Triptyches , 1970
Subturma Tricolpates, Triptyches , 1970
![]()
![]()
![]()
![]() Infraturma Heterotricolpati, Praecolpati , 1970
Infraturma Heterotricolpati, Praecolpati , 1970
Eucommiidites minor et , 1960
Eucommiidites sp. in , 1970
![]()
![]()
![]()
![]() Infraturma Isotricolpati , 1970
Infraturma Isotricolpati , 1970
Biorecord (cand) Retitri-croton (this study)
Ovoidites parvus ( et , 1959) , 1966
Schizosporis reticulatus ( et , 1959) , 1976
Dictyothylakos sp. in , 1964
Click on thumbnail to enlarge the image.
Figure 1: Simplified geological map of the Mons Basin. Location of Baudour, Bernissart and of the "pockets" filled with Wealden facies sediments (in black) on the northern border of the basin.
Click on thumbnail to enlarge the image.
Figure 2: Biorecord Superret-croton, presumed distal face (slide Bau 2A coordinates G37 - 1a, 2b and 2c: respectively high, less high and middle focus). Scale-bar is 10 µm.

Click on thumbnail to enlarge the image.
Figure 3: Biorecord Superret-croton, presumed distal face (slide Bau 3B coordinates N44-4). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 4: Biorecord Superret-croton (slide Bau 4B coordinates T27-U27 4a, 4b and 4c: respectively high, average and low focus). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 5: Biorecord Superret-croton (slide Bau 4B coordinates D28-2 5a and 5b: respectively low and high focus). Scale-bar is 10 µm.

Click on thumbnail to enlarge the image.
Figure 6: Biorecord Superret-croton, presumed distal face (slide Bau 2B coordinates Q24-4/Q25-3). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
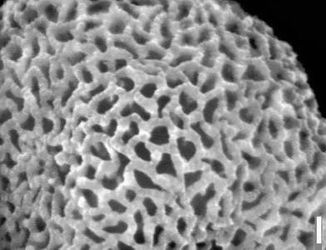
Figure 7: Biorecord Superret-croton, presumed distal face (glass stub 2/1 7a and 7b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).
Click on thumbnail to enlarge the image.
Figure 8: Biorecord Superret-croton, presumed proximal face (glass stub 2/1 8a and 8b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).

Click on thumbnail to enlarge the image.
Figure 9: Inaperturate "variant form": biorecord Inapert-croton, usual wrinkled specimen (slide Bau 4B coordinates D43-E44). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 10: Inaperturate "variant form": biorecord Inapert-croton (glass stub 2/1 10a and 10b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).
Click on thumbnail to enlarge the image.
Figure 11: Inaperturate "variant form": biorecord Inapert-croton (glass stub 2/2 11a and 11b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).
Click on thumbnail to enlarge the image.
Figure 12: Inaperturate "variant form": biorecord Inapert-croton (glass stub 2/2 12a and 12b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).

Click on thumbnail to enlarge the image.
Figure 13: Tricolpate "variant form": biorecord (cand) Retitri-croton (slide Bau 2A coordinates N33-2). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 14: Tricolpate "variant form": biorecord (cand) Retitri-croton (slide Bau 4B coordinates F27-1 14a and 14b: respectively low and high focus). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 15: Trichotomosulcate "variant form": biorecord (cand) Trichoto-croton (slide Bau 2B coordinates J31/K31 15a and 15b: respectively average and low focus). Scale-bar is 10 µm.

Click on thumbnail to enlarge the image.
Figure 16: Probable paleotaxon Superret-subcrot (slide Bau 4B coordinates L32-L33 assemblage of two photographs with different foci). Scale-bar is 10 µm.
Click on thumbnail to enlarge the image.
Figure 17: Biorecord Retisulc-dentat, presumed proximal face (glass stub 2/2 17a and 17b: whole specimen and detail). Scale-bar is 10 µm, except higher SEM magnification (1 µm).
 a
a
 b
b
 c
c a
a
 b
b
 c
c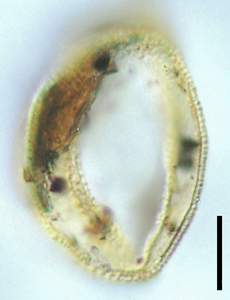 a
a 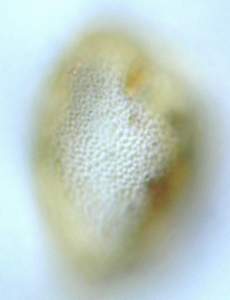 b
b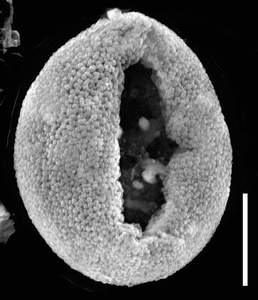 a
a
 b
b a
a
 b
b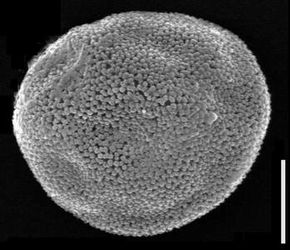 a
a
 b
b a
a
 b
b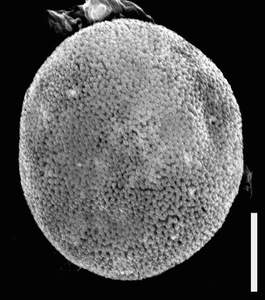 a
a
 b
b a
a
 b
b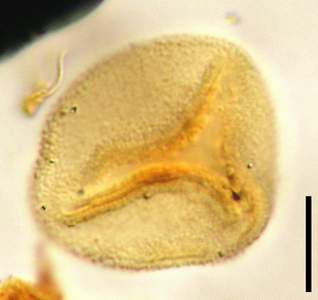 a
a
 b
b a
a
 b
b