![]()
![]()
E.J. (2006).- Patterns of diversification in early eukaryotes. In: P. & E. (eds.), Recent Advances in Palynology.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/01, Abstract 06 (CG2007_M01/06)
![]() Proterozoic;
early eukaryotes; diversification.
Proterozoic;
early eukaryotes; diversification.
![]() Protérozoïque
; premiers eucaryotes ; diversification.
Protérozoïque
; premiers eucaryotes ; diversification.
![]() The Precambrian includes: the Hadean (4.6 to 4 Ga), the period of solar system formation and Earth accretion; the Archean (4 to 2.5 Ga) when life appeared, and the Proterozoic (2.5 to 0.56 Ga) subdivided into the Paleo-, Meso-, and Neoproterozoic. During this
major part of Earth history (about 90%), major environmental changes were
registered in the geological record. These events include the step-wise oxygenation of the atmosphere and oceans, meteoritic impacts, supercontinent formation and breakup, and severe glaciations;
they may have had a profound effect on the early evolution of the eukaryotes. Several lines of evidence from the geological record, the fossil
history and molecular phylogenies can be used to decipher the early record of the domain Eucarya and its evolution.
The Precambrian includes: the Hadean (4.6 to 4 Ga), the period of solar system formation and Earth accretion; the Archean (4 to 2.5 Ga) when life appeared, and the Proterozoic (2.5 to 0.56 Ga) subdivided into the Paleo-, Meso-, and Neoproterozoic. During this
major part of Earth history (about 90%), major environmental changes were
registered in the geological record. These events include the step-wise oxygenation of the atmosphere and oceans, meteoritic impacts, supercontinent formation and breakup, and severe glaciations;
they may have had a profound effect on the early evolution of the eukaryotes. Several lines of evidence from the geological record, the fossil
history and molecular phylogenies can be used to decipher the early record of the domain Eucarya and its evolution.
![]() Genetic material is rarely preserved in the
rock record, so paleontologists have to rely on other features to identify microfossils as members of the domain Eucarya. Fossils provide direct evidence of early cells, and document steps in biological and biochemical innovations. Organisms can be preserved by
a variety of processes in a range of substrates. Early eukaryotic fossils
include: carbonaceous compressions (the organisms are preserved as a thin film of carbon); acritarchs (organic-walled vesicles with unknown biological affinities, they can be extracted from shales using strong acids, or observed in thin sections
of shale, chert or phosphorite); multicellular organic-walled organisms (chert, shale); vase-shaped microfossils; molds and casts in sandstone or shale; skeletons preserved in carbonates or phosphorite; and phylogenetically informative molecules (biomarkers and biopolymers preserved in the rocks
that provide information about past ecosystems and the evolution of biosynthetic pathways).
Genetic material is rarely preserved in the
rock record, so paleontologists have to rely on other features to identify microfossils as members of the domain Eucarya. Fossils provide direct evidence of early cells, and document steps in biological and biochemical innovations. Organisms can be preserved by
a variety of processes in a range of substrates. Early eukaryotic fossils
include: carbonaceous compressions (the organisms are preserved as a thin film of carbon); acritarchs (organic-walled vesicles with unknown biological affinities, they can be extracted from shales using strong acids, or observed in thin sections
of shale, chert or phosphorite); multicellular organic-walled organisms (chert, shale); vase-shaped microfossils; molds and casts in sandstone or shale; skeletons preserved in carbonates or phosphorite; and phylogenetically informative molecules (biomarkers and biopolymers preserved in the rocks
that provide information about past ecosystems and the evolution of biosynthetic pathways).
![]() Molecular phylogenies
yield important information or hypotheses about relationships between clades and
their order of branching. However paleobiological data are essential for testing these trees and for constraining the (minimum age of) timing of diversification. Fossils may also record ancestral forms (and steps in evolution) that might not have any extant relatives. The position of the root of the tree of life is not
yet understood. Within the eukaryotic tree, the eukaryotes are divided into several supergroups whose relationships are not well resolved. Lineages thought to have branched early because they seemed to lack mitochondria, actually host derived mitochondrial organelles (
& , 2006). Nevertheless, calibration of phylogenies
using dates from fossils, biomarkers, and isotopes, shows that a major diversification of extant clades occurred in the Neoproterozoic, preceded by a long evolution of eukaryotic fossils starting in the late Archean
-as suggested by biomarkers- or in the late Paleoproterozoic, when the oldest eukaryotic microfossils are
found (see reviews in
, 2006; et alii,
2006;
, 2004). Superimposing the record of biological innovations and environmental changes
on the fossil record might reveal possible explanations of the pattern of diversification in the middle
Neoproterozoic, long after the origin of the domain and possible early divergence of major clades in the Paleo- and
Mesoproterozoic when eukaryotic fossils of unknown biological affinities are
preserved.
Molecular phylogenies
yield important information or hypotheses about relationships between clades and
their order of branching. However paleobiological data are essential for testing these trees and for constraining the (minimum age of) timing of diversification. Fossils may also record ancestral forms (and steps in evolution) that might not have any extant relatives. The position of the root of the tree of life is not
yet understood. Within the eukaryotic tree, the eukaryotes are divided into several supergroups whose relationships are not well resolved. Lineages thought to have branched early because they seemed to lack mitochondria, actually host derived mitochondrial organelles (
& , 2006). Nevertheless, calibration of phylogenies
using dates from fossils, biomarkers, and isotopes, shows that a major diversification of extant clades occurred in the Neoproterozoic, preceded by a long evolution of eukaryotic fossils starting in the late Archean
-as suggested by biomarkers- or in the late Paleoproterozoic, when the oldest eukaryotic microfossils are
found (see reviews in
, 2006; et alii,
2006;
, 2004). Superimposing the record of biological innovations and environmental changes
on the fossil record might reveal possible explanations of the pattern of diversification in the middle
Neoproterozoic, long after the origin of the domain and possible early divergence of major clades in the Paleo- and
Mesoproterozoic when eukaryotic fossils of unknown biological affinities are
preserved.
![]() As discussed elsewhere (
& , 2006; et alii,
2003, 2004; et alii,
2005), in order to determine the biological affinities of these fossils at the level of domain or beyond, we have defined a set of criteria to differentiate
prokaryotic from eukaryotic microfossils and have formulated a methodology combining microscopy and microchemistry of single acritarchs. Fossils can display morphological and ultrastructural features showing a degree of complexity and/or particular features unknown in prokaryotic organisms, therefore pointing to a eukaryotic affinity. Indeed, the wall structure and ornamentation, the presence of processes that extend from the vesicle wall, the presence of excystment structures (openings through which cyst liberate their content), wall ultrastructure and wall chemistry can clarify the biological affinities of organic-walled microfossils at the level of the domain, and in some
cases even at the level of class. Microchemical analyses such as micro infra-red and Raman spectroscopy, secondary ion mass spectrometry, and other techniques applicable to very small samples such as one acritarch can be used to characterize the chemistry of organic microfossils and might even reveal biomolecules specific
to extant clades.
As discussed elsewhere (
& , 2006; et alii,
2003, 2004; et alii,
2005), in order to determine the biological affinities of these fossils at the level of domain or beyond, we have defined a set of criteria to differentiate
prokaryotic from eukaryotic microfossils and have formulated a methodology combining microscopy and microchemistry of single acritarchs. Fossils can display morphological and ultrastructural features showing a degree of complexity and/or particular features unknown in prokaryotic organisms, therefore pointing to a eukaryotic affinity. Indeed, the wall structure and ornamentation, the presence of processes that extend from the vesicle wall, the presence of excystment structures (openings through which cyst liberate their content), wall ultrastructure and wall chemistry can clarify the biological affinities of organic-walled microfossils at the level of the domain, and in some
cases even at the level of class. Microchemical analyses such as micro infra-red and Raman spectroscopy, secondary ion mass spectrometry, and other techniques applicable to very small samples such as one acritarch can be used to characterize the chemistry of organic microfossils and might even reveal biomolecules specific
to extant clades.
![]() One limitation of this approach is the limited knowledge that we have
about extant organisms producing fossilizable structures and their morphological, ultrastructural and chemical properties. This approach requires investigation of preservable biological properties and comparative actualistic studies of taphonomic processes affecting diverse organisms
in diverse environments ( & ,
2006).
One limitation of this approach is the limited knowledge that we have
about extant organisms producing fossilizable structures and their morphological, ultrastructural and chemical properties. This approach requires investigation of preservable biological properties and comparative actualistic studies of taphonomic processes affecting diverse organisms
in diverse environments ( & ,
2006).
![]() Fossils can inform about
the evolution of biological innovations, regardless of their biological affinities, as briefly summarized below (see reviews in
, 2006; et alii,
2006 and reference therein).
Fossils can inform about
the evolution of biological innovations, regardless of their biological affinities, as briefly summarized below (see reviews in
, 2006; et alii,
2006 and reference therein).
![]() - Biomarkers in 2.7 Ga kerogens of the Fortescue Group, Australia, indicate that contemporaneous cells were able to synthesize sterols, requiring a minimum of oxygen.
- Biomarkers in 2.7 Ga kerogens of the Fortescue Group, Australia, indicate that contemporaneous cells were able to synthesize sterols, requiring a minimum of oxygen.
![]() - Paleo- and Mesoproterozoic macroscopic compressions or mold and cast structures have been compared to algae but this interpretation remains controversial.
- Paleo- and Mesoproterozoic macroscopic compressions or mold and cast structures have been compared to algae but this interpretation remains controversial.
![]() -
The first ornamented acritarchs are populations of Valeria lophostriata recorded
in the Paleoproterozoic of China (~1.8 Ga) and Australia (+1.65 Ga). Early Mesoproterozoic acritarchs Shuiyousphaeridium macroreticulatum, Valeria lophostriata, Tappania plana, and Satka favosa exhibits a complexity of form observed with TEM, SEM, and light microscopy that is unknown in prokaryotes. Prokaryotes can be large, they can have ornamentation, and they can have preservable walls (at least cyanobacterial sheaths), but no prokaryote currently known has all three (large size, ornamentation, preservable acid-resistant walls) at once. Many eukaryotes do. Therefore, these early microfossils display
the characteristics of a eukaryotic grade of organization, and are interpreted as eukaryotes with a sophisticated cytoskeleton. These 1.65-1.3 Ga fossil assemblages record biological innovations such as reproduction by budding, complex ecology, vegetative and resting stages, synthesis of resistant polymers, synthesis of various wall ornamentation including processes, and a moderate diversity.
-
The first ornamented acritarchs are populations of Valeria lophostriata recorded
in the Paleoproterozoic of China (~1.8 Ga) and Australia (+1.65 Ga). Early Mesoproterozoic acritarchs Shuiyousphaeridium macroreticulatum, Valeria lophostriata, Tappania plana, and Satka favosa exhibits a complexity of form observed with TEM, SEM, and light microscopy that is unknown in prokaryotes. Prokaryotes can be large, they can have ornamentation, and they can have preservable walls (at least cyanobacterial sheaths), but no prokaryote currently known has all three (large size, ornamentation, preservable acid-resistant walls) at once. Many eukaryotes do. Therefore, these early microfossils display
the characteristics of a eukaryotic grade of organization, and are interpreted as eukaryotes with a sophisticated cytoskeleton. These 1.65-1.3 Ga fossil assemblages record biological innovations such as reproduction by budding, complex ecology, vegetative and resting stages, synthesis of resistant polymers, synthesis of various wall ornamentation including processes, and a moderate diversity.
![]() - The bangiophyte red alga Bangiomorpha pubescens is
so far the oldest taxonomically resolved eukaryote, and records the evolution of complex multicellularity, cell differentiation, and sexual reproduction, eukaryotic photosynthesis, primary endosymbiosis of a chloroplast ancestor by 1.2-1 Ga. Note that these biological innovations are recorded in this one fossil population of bangiophyte red
algae that chronostratigraphy dates at 1.2 Ga-750 Ma. Chemostratigraphy and
lithostratigraphy indicate an age closer to 1.2 Ga. However other multicellular photosynthetic eukaryotes also appeared around 1 Ga.
- The bangiophyte red alga Bangiomorpha pubescens is
so far the oldest taxonomically resolved eukaryote, and records the evolution of complex multicellularity, cell differentiation, and sexual reproduction, eukaryotic photosynthesis, primary endosymbiosis of a chloroplast ancestor by 1.2-1 Ga. Note that these biological innovations are recorded in this one fossil population of bangiophyte red
algae that chronostratigraphy dates at 1.2 Ga-750 Ma. Chemostratigraphy and
lithostratigraphy indicate an age closer to 1.2 Ga. However other multicellular photosynthetic eukaryotes also appeared around 1 Ga.
![]() - Upper Mesoproterozoic/Lower Neoproterozoic rocks (and possibly Paleoproterozoic rocks) have yielded biomarkers of alveolates (which include dinoflagellates and ciliates, among other groups).
- Upper Mesoproterozoic/Lower Neoproterozoic rocks (and possibly Paleoproterozoic rocks) have yielded biomarkers of alveolates (which include dinoflagellates and ciliates, among other groups).
![]() - Palaeovaucheria, a 1 Ga xanthophyte alga, indicates the appearance of stramenopiles (which include diatoms, xanthophytes, and brown algae) and of secondary symbiosis (involving a red alga-like endosymbiont).
- Palaeovaucheria, a 1 Ga xanthophyte alga, indicates the appearance of stramenopiles (which include diatoms, xanthophytes, and brown algae) and of secondary symbiosis (involving a red alga-like endosymbiont).
![]() - The 750 Ma vase-shaped microfossils provide a firm calibration point for the opisthokonts, the clade that includes animals, fungi and the amoebozoans not to mention direct evidence for heterotrophic eukaryotes and eukaryotic biomineralization, and possibly predation. Cladophorales green algae also appeared, recording again multicellular photosynthetic
eukaryotes, and implying earlier evolution and diversification of green algae,
as clearly underlined by et alii
(1994), (2003)
and other Precambrian paleontologists, but recently misunderstood by
(2006).
- The 750 Ma vase-shaped microfossils provide a firm calibration point for the opisthokonts, the clade that includes animals, fungi and the amoebozoans not to mention direct evidence for heterotrophic eukaryotes and eukaryotic biomineralization, and possibly predation. Cladophorales green algae also appeared, recording again multicellular photosynthetic
eukaryotes, and implying earlier evolution and diversification of green algae,
as clearly underlined by et alii
(1994), (2003)
and other Precambrian paleontologists, but recently misunderstood by
(2006).
![]() - The late Neoproterozoic appearance of animals preserved as calcareous skeletons forming large reefs or as
possible animal embryos in phosphorites added another dimension to ecosystems and predation pressure.
- The late Neoproterozoic appearance of animals preserved as calcareous skeletons forming large reefs or as
possible animal embryos in phosphorites added another dimension to ecosystems and predation pressure.
![]() - Florideophyte red algae at ~600 Ma record the evolution of a tissue-grade organization (cell differentiation in three dimensions to form a tissue with a specific function).
- Florideophyte red algae at ~600 Ma record the evolution of a tissue-grade organization (cell differentiation in three dimensions to form a tissue with a specific function).
![]() So fossils do provide evidence for
the evolution of eukaryotic biology, but their change in diversity might also tell us something about the factors controlling the patterns we see in the fossil record. What are those patterns?
So fossils do provide evidence for
the evolution of eukaryotic biology, but their change in diversity might also tell us something about the factors controlling the patterns we see in the fossil record. What are those patterns?
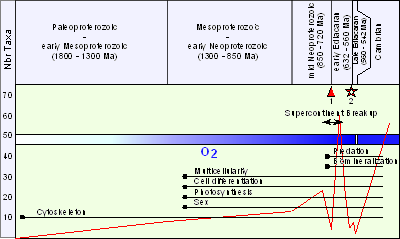
![]() The Figure 1
The Figure 1 ![]() is a schematic curve drawn from a recent compilation of the number of taxa per assemblage throughout the
Proterozoic, including the acritarchs, the VSMs, multicellular microfossils and macroscopic remains (data and references in et alii,
2006). Of course, the fossil record is incomplete and more discoveries will undoubtedly improve our understanding. Nevertheless, there is a clear trend
toward increasing diversity, starting with a low rate of diversity in the late
Paleoproterozoic, a modest rate in the Mesoproterozoic-early Neoproterozoic, followed by a sharp increase in the mid and late Neoproterozoic (or early
Ediacarian). In the late Ediacarian, diversity decreases before rising again at the Phanerozoic boundary. Between
"global" glaciations, the fossil record is sparse and seems to show a big drop in diversity, although this might also result from
insufficient sampling and/or gaps in the fossil record.
is a schematic curve drawn from a recent compilation of the number of taxa per assemblage throughout the
Proterozoic, including the acritarchs, the VSMs, multicellular microfossils and macroscopic remains (data and references in et alii,
2006). Of course, the fossil record is incomplete and more discoveries will undoubtedly improve our understanding. Nevertheless, there is a clear trend
toward increasing diversity, starting with a low rate of diversity in the late
Paleoproterozoic, a modest rate in the Mesoproterozoic-early Neoproterozoic, followed by a sharp increase in the mid and late Neoproterozoic (or early
Ediacarian). In the late Ediacarian, diversity decreases before rising again at the Phanerozoic boundary. Between
"global" glaciations, the fossil record is sparse and seems to show a big drop in diversity, although this might also result from
insufficient sampling and/or gaps in the fossil record.
![]() A closer look at the composition of acritarch assemblages ( et alii,
2006) reveals that earlier assemblages include mostly smooth and ornamented
sphaeromorphs, along with a few forms with asymmetrically distributed processes whereas younger assemblages include more diversely ornamented acritarchs and acritarchs with symmetrically distributed processes. In the Late
Ediacarian, acanthomorphs disappear and large leiospheres dominate assemblages. Cambrian assemblages include a renewed diversity of ornamented forms and small
acanthomorphs, generally assumed to represent planktonic algal cysts.
A closer look at the composition of acritarch assemblages ( et alii,
2006) reveals that earlier assemblages include mostly smooth and ornamented
sphaeromorphs, along with a few forms with asymmetrically distributed processes whereas younger assemblages include more diversely ornamented acritarchs and acritarchs with symmetrically distributed processes. In the Late
Ediacarian, acanthomorphs disappear and large leiospheres dominate assemblages. Cambrian assemblages include a renewed diversity of ornamented forms and small
acanthomorphs, generally assumed to represent planktonic algal cysts.
![]() To understand the factors controlling this
pattern of fluctuations in diversity, we can superimpose on the diversity curve, the timing of environmental changes registered in the rock record and the timing of biological innovations
evinced in the fossil record.
To understand the factors controlling this
pattern of fluctuations in diversity, we can superimpose on the diversity curve, the timing of environmental changes registered in the rock record and the timing of biological innovations
evinced in the fossil record.
![]() Three main factors are generally proposed to explain
changes in diversity: genetic innovations, ecological innovations and environmental changes.
Three main factors are generally proposed to explain
changes in diversity: genetic innovations, ecological innovations and environmental changes.
![]() Since early cells had already all the eukaryotic features typical of their domain by the late Paleoproterozoic and possibly earlier, why did diversity stayed modest until the
mid-Neoproterozoic?
Since early cells had already all the eukaryotic features typical of their domain by the late Paleoproterozoic and possibly earlier, why did diversity stayed modest until the
mid-Neoproterozoic?
![]() Sex, complex multicellularity (and cell differentiation) and eukaryotic photosynthesis appeared around 1.2
Ga, well before the increase in diversity around 850 Ma. Glaciations (poorly constrained at ~720-710 and 650-635 Ma) and possibly a transient shallow-water anoxia event at the Precambrian-Cambrian boundary might have cut down some of the diversity, but most clades (the red, green, xanthophyte algae, the fungi, and the amoebae) survived and diversified again in the
Phanerozoic. Supercontinent breakup might have increased the surface area of epicontinental seas and
modified the cycle of nutrients, thus providing new niches and altering the chemistry
of the early oceans. Animal predation pressure might also have played a role in forcing diversification. Some authors suggested that spines on acritarchs could be a defense mechanism against metazoan grazers, and that the modest early acritarch diversity could have resulted from the absence of animals before the Ediacaran ( & ,
2005). However it is also possible that some of these acritarchs record directly the presence of animals as some of them resemble metazoan eggs ( et alii,
2006), but the wall of at least one species (Tanarium
conoideum) was made of a biopolymer similar to algaenan, thus indicating a green algal affinity ( et alii,
2005). Finally in Australia a meteoritic impact (so-called Acraman impact) occurred at ~570 Ma, and acritarch assemblages before and after the
impact seem to differ (,
2005).
Sex, complex multicellularity (and cell differentiation) and eukaryotic photosynthesis appeared around 1.2
Ga, well before the increase in diversity around 850 Ma. Glaciations (poorly constrained at ~720-710 and 650-635 Ma) and possibly a transient shallow-water anoxia event at the Precambrian-Cambrian boundary might have cut down some of the diversity, but most clades (the red, green, xanthophyte algae, the fungi, and the amoebae) survived and diversified again in the
Phanerozoic. Supercontinent breakup might have increased the surface area of epicontinental seas and
modified the cycle of nutrients, thus providing new niches and altering the chemistry
of the early oceans. Animal predation pressure might also have played a role in forcing diversification. Some authors suggested that spines on acritarchs could be a defense mechanism against metazoan grazers, and that the modest early acritarch diversity could have resulted from the absence of animals before the Ediacaran ( & ,
2005). However it is also possible that some of these acritarchs record directly the presence of animals as some of them resemble metazoan eggs ( et alii,
2006), but the wall of at least one species (Tanarium
conoideum) was made of a biopolymer similar to algaenan, thus indicating a green algal affinity ( et alii,
2005). Finally in Australia a meteoritic impact (so-called Acraman impact) occurred at ~570 Ma, and acritarch assemblages before and after the
impact seem to differ (,
2005).
![]() So far, not
any one event seems to explain the observed pattern of early eukaryote diversification. More precise dating constraints on environmental changes and the fossil record are needed before drawing conclusions, but probably the three factors (genetics, ecology, environmental changes)
were involved.
So far, not
any one event seems to explain the observed pattern of early eukaryote diversification. More precise dating constraints on environmental changes and the fossil record are needed before drawing conclusions, but probably the three factors (genetics, ecology, environmental changes)
were involved.
![]() Early eukaryotes had developed many complex and characteristic cellular and molecular mechanisms by 1.2
Ga, but the diversity of microscopic forms increased only in the middle
Neoproterozoic, and a high diversity of macroscopic forms appeared at the end of the Proterozoic era. Despite the unavoidable incompleteness of the fossil record, the observed pattern seems to reflect natural trends. The causes of this relatively late or delayed diversification are probably multiple, including changes in the chemistry of
the early atmosphere and oceans and a lack of (or reduced) predation pressure until the Ediacarian when increased oxygenation permitted the evolution of animals (possibly already
present) and consequently the complexification of ecosystems (see discussions in ,
2005;
& , 2005; et alii,
2006). et alii
(2006) examined the evolution of variations in the morphology of acritarch vesicles, processes, process tips,
and ornamentation: broadly correlating the observed pattern with environmental (glaciations) and ecological changes (appearance of Ediacaran metazoans and
"Cambrian explosion") but as pointed out above, cause-and-effects relationships
may not be so closely connected. Moreover, the authors do not explain the rise in diversity around 850 Ma also detected in their study.
Early eukaryotes had developed many complex and characteristic cellular and molecular mechanisms by 1.2
Ga, but the diversity of microscopic forms increased only in the middle
Neoproterozoic, and a high diversity of macroscopic forms appeared at the end of the Proterozoic era. Despite the unavoidable incompleteness of the fossil record, the observed pattern seems to reflect natural trends. The causes of this relatively late or delayed diversification are probably multiple, including changes in the chemistry of
the early atmosphere and oceans and a lack of (or reduced) predation pressure until the Ediacarian when increased oxygenation permitted the evolution of animals (possibly already
present) and consequently the complexification of ecosystems (see discussions in ,
2005;
& , 2005; et alii,
2006). et alii
(2006) examined the evolution of variations in the morphology of acritarch vesicles, processes, process tips,
and ornamentation: broadly correlating the observed pattern with environmental (glaciations) and ecological changes (appearance of Ediacaran metazoans and
"Cambrian explosion") but as pointed out above, cause-and-effects relationships
may not be so closely connected. Moreover, the authors do not explain the rise in diversity around 850 Ma also detected in their study.
![]() A multidisciplinary approach combining microscopy and microchemistry on
both fossils and extant
cells may permit a better understanding of the paleobiology of the fossils, and
might clarify this pattern of diversification by identifying members of early and later
clades, such as the late Neoproterozoic acritarchs (were they animal eggs and/or phytoplankton cysts or something else?). One crucial point to elucidate is whether or not the late diversification
reflects diversification between or within clades. Collating the paleobiological data with information from geology and geochemistry
regarding paleoenvironments and their evolution, and with insights from molecular phylogeny, we can better understand the evolution of life on our planet and characterize
the biosignatures needed for paleobiology and astrobiology.
A multidisciplinary approach combining microscopy and microchemistry on
both fossils and extant
cells may permit a better understanding of the paleobiology of the fossils, and
might clarify this pattern of diversification by identifying members of early and later
clades, such as the late Neoproterozoic acritarchs (were they animal eggs and/or phytoplankton cysts or something else?). One crucial point to elucidate is whether or not the late diversification
reflects diversification between or within clades. Collating the paleobiological data with information from geology and geochemistry
regarding paleoenvironments and their evolution, and with insights from molecular phylogeny, we can better understand the evolution of life on our planet and characterize
the biosignatures needed for paleobiology and astrobiology.
![]() This extended abstract has benefited from discussions and/or previous published work in collaboration with A.H. ,
C.P. , and K. .
This extended abstract has benefited from discussions and/or previous published work in collaboration with A.H. ,
C.P. , and K. .
N.J., A.H. & K. (1994).- Paleobiology of the Neoproterozoic Svanbergjellet Formation, Spitsbergen.- Fossils and Strata, Oslo, n° 34, 84 p.
T. & W. (2006).- Eukaryotic evolution, changes and challenges.- Nature, London, vol. 440, n° 7084, p. 623-630.
K. (2005).- Ediacaran Palynology of Australia.- Memoirs of the Association of Australasian Palaeontologists, Canberra, vol. 31, 439 p.
J.W., S. & M. (2005).- 1.3 Billion years of acritarch history: An empirical morphospace approach.- Precambrian Research, Amsterdam, vol. 144, n° 1-2, p. 52-68.
E.J. (2006).- The early eukaryotic fossil record. In: G. (ed.), Origins and Evolution of Eukaryotic Endomembranes and Cytoskeleton.- Landes Biosciences, Austin, 19 p.
E.J., A.H. & M.R. (2003).- Recognizing and interpreting the fossils of Early Eukaryotes.- Origins of Life and Evolution of Biospheres, Amsterdam, vol. 33, n° 1, p. 75-94.
E.J., A.H. & M.R. (2004).- TEM evidence for eukaryotic diversity in mid-Proterozoic oceans.- Geobiology, Malden, vol. 2, n° 3, p. 121-132.
E.J. & C.P. (2006).- A new approach in deciphering early protist paleobiology and evolution: combined microscopy and microchemistry of single Proterozoic acritarchs.- Review of Palaeobotany and Palynology, Amsterdam, vol. 139, n° 1-4, p. 1-15.
A.H. (2003).- Life on a young planet. The first three billion years of evolution on Earth.- Princeton University Press, third edition, 277 p.
A.H., E.J., D. & P. (2006).- Eukaryotic organisms in Proterozoic Oceans.- Philosophical Transactions of the Royal Society of London B, London, vol. 361, n° 1470, p. 1023-1038.
C.P., E.J., A.H. & M.R. (2005).- Combined micro-Fourier transform infrared (FTIR) spectroscopy and micro-Raman spectroscopy of Proterozoic acritarchs: a new approach to Palaeobiology.- Precambrian Research, Amsterdam, vol. 138, n° 3-4, p. 208-224.
K.J. & N.J. (2005).- Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record.- Proceedings of the National Academy of Sciences, Washington, vol. 102, n° 27, p. 9547-9552.
S.M. (2004).- Early eukaryotic diversification. In: J. & B. (eds.), Neoproterozoic-Cambrian biological revolutions.- Paleontological Society Papers, Lawrence, vol. 10, p. 35-50.
B. (2006).- Are the green algae (phylum Viridiplantae) two billion years old?- Carnets de Géologie - Notebooks on Geology, Brest, Article 2006/03 (CG2006_A03), 15 p.
Click on thumbnail to enlarge the image.
Figure 1: Patterns of early eukaryotic diversification, biological innovations and environmental changes (modified from et alii, 2006).
This figure shows the general trend of variation in number of eukaryotic taxa (compilation of acritarchs, VSMs, multicellular fossils and macroscopic compressions, data in et alii, 2006) through the Proterozoic, and the timing of major environmental changes (supercontinent formation and breakup, widespread glaciations, meteorite impact in Australia, and change in oxygenation), and of biological innovations evidenced by the fossil record (, 2006). 1: Glaciations. 2: Acraman impact.