![]()
Sommaire - Contents
[Abridged English version ![]() ]
]
[Introduction] [Material and methods]
[Biostratigraphic results]
[Discussion]
[Paleocecological ... remarks]
[Taxonomy]
[Conclusion]
[Bibliographic references] [Figures] and ... [Plates]
[Version française ![]() ]
]
[Introduction] [Matériel et méthodes]
[Résultats biostratigraphiques]
[Discussion]
[Remarques paléoécologiques ...]
[Notes taxinomiques]
[Conclusion]
[Références bibliographiques] [Figures] et ... [Planches]
![]()
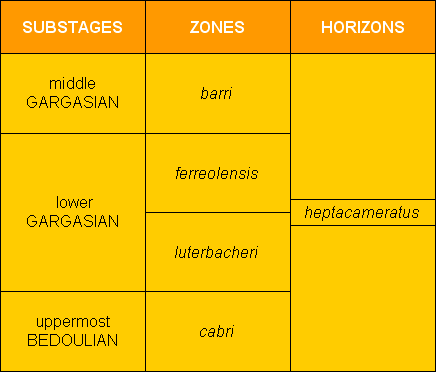
Faisant suite aux recherches sur le Bédoulien et le Gargasien du secteur de Cassis-La Bédoule (B. du Rh., SE France), l'étude de quatre coupes en relais dans l'aire stratotypique de l'Aptien, situées dans le secteur du Hameau de la Tuilière (commune de Saint-Saturnin-lès-Apt, Vaucluse, SE France) a permis l'analyse détaillée des associations de foraminifères benthiques (30 taxons) et planctoniques (15 taxons) du Bédoulien supérieur et du Gargasien inférieur. Descriptions, figurations et répartition stratigraphique sont fournies pour les formes les plus significatives, avec individualisation de plusieurs marqueurs, benthiques et surtout planctoniques.
L'étude de ces derniers dans le cadre privilégié de coupes dilatées et dépourvues de hiatus a permis de conforter la zonation définie antérieurement dans le stratotype de l'Aptien inférieur de Cassis-La Bédoule (succession des zones à cabri, luterbacheri, ferreolensis, barri, algerianus) et même de l'affiner, par la mise en évidence d'un horizon à Globigerinelloides ferreolensis heptacameratus nov. subsp., chevauchant la limite entre les zones à luterbacheri et à ferreolensis. Nos résultats montrent également la quasi-coïncidence temporelle 1) du toit de la zones d'ammonites à Furcata, 2) du changement lithologique dit du "changement de couleur" et 3) d'un triple datum micropaléontologique (FAD de Praehedbergella luterbacheri, LAD de Protocythere bedoulensis et Lenticulina cf. nodosa) dont l'importance avait déjà été signalée dans le stratotype de Cassis-La Bédoule. Ces nouvelles données peuvent constituer des éléments pour la définition de la limite Bédoulien-Gargasien.
Durant le Gargasien inférieur (au sein de la Zone à luterbacheri), un épisode probablement d'origine volcanique, que nous sommes tentés d'assimiler au Niveau Van Gogh (initialement détecté dans le domaine vocontien), est marqué par une forte crise de diversité affectant les foraminifères épibenthiques, planctoniques, ainsi que les ostracodes.
Sur le plan paléoécologique et paléobiogéographique, la comparaison des associations de foraminifères benthiques et planctoniques du secteur de La Tuilière avec celles des régions voisines fait apparaître des spécificités liées à la paléo-profondeur et des affinités paraissant plus marquées avec les microfaunes de certaines régions boréales, comme l'Allemagne du Nord, qu'avec le domaine téthysien.
Aptien ; stratotype historique ; SE France ; Foraminifères ; benthiques ; planctoniques ; biostratigraphie ; systématique.
M., G. & J.-P. (2008).- Associations et biostratigraphie des Foraminifères benthiques et planctoniques du Bédoulien sommital et du Gargasien inférieur de La Tuilière - St-Saturnin-lès-Apt (aire stratotypique de l'Aptien, Vaucluse, SE France).- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/01 (CG2008_A01)
Planktonic and benthic foraminiferal assemblages and biostratigraphy of the uppermost Bedoulian and lower Gargasian of La Tuilière - St-Saturnin-lès-Apt (area of the Aptian stratotype, Vaucluse, SE France).- Research on the Bedoulian and Gargasian of the Cassis-La Bédoule (Bouches du Rhône, SE France) area is extended, by a study of four overlapping sections in the stratotypic area of the Aptian stage in the hamlet of La Tuilière (commune of Saint-Saturnin-lès-Apt, Vaucluse, SE France). This new information permitted a detailed analysis of upper Bedoulian and lower Gargasian benthic (30 taxa) and planktonic (15 taxa) foraminiferal assemblages. Here we describe, illustrate and give the ranges and distribution of the most significant forms, and call attention to several benthic and planktonic species that may serve as extra-regional markers.
The study of the planktonics in these expanded and continuous sections made it possible to strengthen the zonation which had been established in the Lower Aptian stratotype of Cassis-La Bédoule, i.e. a succession of five zones (cabri, luterbacheri, ferreolensis, barri, algerianus); this zonal scheme is improved by the addition of a horizon with Globigerinelloides ferreolensis heptacameratus nov. subsp., which straddles the boundary between the luterbacheri and ferreolensis zones. We also show the temporal quasi-coincidence of: 1) the top of the Furcata ammonite Zone, 2) the lithological change named "change of color" and 3) a triple micropaleontological datum (FAD of Praehedbergella luterbacheri, LAD of Protocythere bedoulensis and Lenticulina cf. nodosa) of which the importance in the stratotype of Cassis-La Bédoule had already been pointed out. All these data may be valuable in the definition of the Bedoulian-Gargasian boundary.
Within the luterbacheri Zone of the early Gargasian, an episode probably of volcanic origin that we interpret as the equivalent of the Van Gogh Level (initially detected in the Vocontian domain), is marked by a strong crisis in the diversity of epibenthic and planktonic foraminifers, as well as in that of the ostracodes.
From a paleoecological and paleobiogeographical point of view, a comparison of the benthic and planktonic foraminiferal assemblages of La Tuilière with those of neighbouring areas reveals specific features related to paleodepth and shows affinities which appear to be closer with the microfaunas of certain boreal areas (e.g. northern Germany) than with the Tethysian realm.
Aptian; historical stratotype; SE France; Foraminifera; benthic; planktonic; biostratigraphy; taxonomy.
In a series of recently published papers, we detailed the microfaunal content (benthic and planktonic Foraminifera) of the lower (
et alii, 1998) and middle ( et alii,
2005) Aptian beds of the historical stratotype of the Bedoulian substage at Cassis-La Bédoule (Bouches-du-Rhône, SE France). This article
is a continuation of the series which together will constitute a collective Memoir for the
Notebooks on Geology concerning lower and middle Aptian stratotypes.
It includes a report on the distribution of Foraminifera collected in a succession of sections located in and very near the hamlet of La Tuilière (parish of Saint-Saturnin-lès-Apt, Vaucluse, SE France)
(Fig. 1 ![]() ). There, four well-exposed sections (Pichouraz, La Tuilière W, Les Gays 1 & 2)
(Fig. 2
). There, four well-exposed sections (Pichouraz, La Tuilière W, Les Gays 1 & 2)
(Fig. 2 ![]() ) allowed us to investigate with continuity and in great detail the uppermost Bedoulian as well as the lower Gargasian, very poorly exposed in the historical type-locality of Gargas, which is only a few kilometers from La Tuilière
(Fig. 1
) allowed us to investigate with continuity and in great detail the uppermost Bedoulian as well as the lower Gargasian, very poorly exposed in the historical type-locality of Gargas, which is only a few kilometers from La Tuilière
(Fig. 1 ![]() ).
).
The location, lithological description and geometrical correlation of the sections, as well as the geological and historical background of the sector of La Tuilière have already been discussed in two preliminary papers ( & ,
2004;
et alii, 2006); therefore, these are not reviewed in this article. But we include some additional lithostratigraphic
(Fig. 3 ![]() ) and biostratigraphic precisions based on the results of progress in the study of the microfaunas.
) and biostratigraphic precisions based on the results of progress in the study of the microfaunas.
The 87 samples (mainly marls) upon which this study is based were taken in 2003 and 2004, at spacings of one (sometimes even less) to two meters. They were treated by Rewoquat W 3690 (see details in et alii, 2005) before sieving through a 45 microns screen.
The state of preservation of the Aptian microfossils from La Tuilière is variable, ranging from poor to moderate. All the specimens of each species were counted. In making the tables of distribution
(Figs. 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() ) we used the frequency-classes defined for the Cassis-La Bédoule area ( et
alii, 2005).
) we used the frequency-classes defined for the Cassis-La Bédoule area ( et
alii, 2005).
The material studied (remainders of samples, washed residues, slides) is preserved in the collections of the Museum of Paleontology of the University of Provence in Marseilles. The types and illustrated specimens are deposited in the micropaleontological collections of the National Museum of Natural History in Paris.
Globorotalites bartensteini
At Cassis-La Bédoule we had seen that this Bedoulian species did not cross the Bedoulian-Gargasian boundary, so its extinction might constitute an additional benthic marker in defining the boundary between the two substages. This datum cannot be confirmed in the sector of La Tuilière, as the genus Globorotalites is very sparsely represented there.
Astacolus crepidularis tricarinella
At Cassis-La Bédoule, in Les Tocchis and in the Comte Quarry sections, this form depasses by only few meters the Bedoulian-Gargasian boundary. In the Pichouraz section its extinction occurs 0.5 m above this limit. This subspecies is also absent in the lower Gargasian succession of the Les Gays 1 section, with the surprising and inexplicable exception of a single sample, more than forty meters above the boundary, containing not less than 13 specimens.
Lenticulina cf. nodosa
This species occurs constantly in the Bedoulian and appears to constitute the only reliable benthic foraminifer that is useful in locating the Bedoulian-Gargasian boundary because of its sharp extinction at this level at Cassis-La Bédoule and Pichouraz, and only 0.5 m above it at La Tuilère W.
Saracenaria sp. aff. frankei
This form, absent in the Bedoulian and basal Gargasian, appears in the upper part of the luterbacheri Zone, where it is rare, and becomes more common in the ferreolensis Zone (Les Gays 1 and 2 sections). It is found more sporadically at similar levels in the La Marcouline section (Cassis).
Oolina apiculata
This species, always rare, is present only in the uppermost beds of the luterbacheri Zone and in the ferreolensis Zone (Les Gays 1 and 2 sections).
Schackoina gr. cabri-pustulans
Data obtained at both Cassis-La Bédoule and La Tuilière (Figs. 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() )
confirm that the various "species" created by some authors within this morphological plexus all have precisely the same stratigraphic distribution. We thus choose to continue to consider them as one group, which appears in the upper Bedoulian with an acme in the uppermost Bedoulian and the extreme base of the Gargasian. The group is then more sporadic in the lower Gargasian and becomes extinct at the beginning of the acme of Globigerinelloides ferreolensis.
)
confirm that the various "species" created by some authors within this morphological plexus all have precisely the same stratigraphic distribution. We thus choose to continue to consider them as one group, which appears in the upper Bedoulian with an acme in the uppermost Bedoulian and the extreme base of the Gargasian. The group is then more sporadic in the lower Gargasian and becomes extinct at the beginning of the acme of Globigerinelloides ferreolensis.
Praehedbergella luterbacheri
In the section of Pichouraz (Fig. 4 ![]() ), P. luterbacheri first occurs in the sample 2367, 2 m above the extinction of the ostracode Protocythere bedoulensis (
et alii, 2006; et alii,
2007) and 1 m above the extinction of the benthic foraminifer Lenticulina cf. nodosa. In the section of La Tuilière W, sampled in greater detail, the appearance of P. luterbacheri coincides with the extinction of P. bedoulensis and precedes by
0.4 m the extinction of
L. cf. nodosa. The datums based either on the extinction of the two benthic forms or on the appearance of the planktonic one are thus almost synchronous and these occurrences are but a very small distance above the lithostratigraphic key level named "change of color", as seen at Pichouraz
(Fig. 4
), P. luterbacheri first occurs in the sample 2367, 2 m above the extinction of the ostracode Protocythere bedoulensis (
et alii, 2006; et alii,
2007) and 1 m above the extinction of the benthic foraminifer Lenticulina cf. nodosa. In the section of La Tuilière W, sampled in greater detail, the appearance of P. luterbacheri coincides with the extinction of P. bedoulensis and precedes by
0.4 m the extinction of
L. cf. nodosa. The datums based either on the extinction of the two benthic forms or on the appearance of the planktonic one are thus almost synchronous and these occurrences are but a very small distance above the lithostratigraphic key level named "change of color", as seen at Pichouraz
(Fig. 4 ![]() ).
).
Globigerinelloides ferreolensis heptacameratus nov. subsp.
In the Les Gays 1 section, the 7-chambered forms of the ferreolensis group first appear in sample 2328,
i.e. approximately 12 m before the 8-chambered ones occur (Fig. 6 ![]() ). In the first 6 meters of this distribution, which correspond to the uppermost levels of the luterbacheri
Zone,
G. ferreolensis heptacameratus nov. subsp. is rather rare and not always easy to distinguish from its direct phylogenetic ancestor, P. luterbacheri (
et alii, 2002, 2005). It is more numerous in the next 6 meters, before the first occurrence of the 8-chambered forms
(= G. ferreolensis ferreolensis).
). In the first 6 meters of this distribution, which correspond to the uppermost levels of the luterbacheri
Zone,
G. ferreolensis heptacameratus nov. subsp. is rather rare and not always easy to distinguish from its direct phylogenetic ancestor, P. luterbacheri (
et alii, 2002, 2005). It is more numerous in the next 6 meters, before the first occurrence of the 8-chambered forms
(= G. ferreolensis ferreolensis).
Globigerinelloides ferreolensis ferreolensis
In the Les Gays 1 & 2 sections, the 8-chambered subspecies of the ferreolensis group appears 6 m above the level at which the acme of the group begins,
i.e. at the level of the extinction of Schackoina gr. cabri (Figs. 6 ![]() - 7
- 7 ![]() ).
).
Globigerinelloides barri
In the La Tuilière area, we found only one individual of this species, in the topmost sample (n° 2301) of the Les Gays 2 section. This may indicate that at the upper limit of the outcrop there the boundary between the ferreolensis and barri zones is just touched.
In the sector of La Tuilière the study of the upper Bedoulian-lower Gargasian beds allowed us to confirm the existence of the zonal sequence: cabri-luterbacheri-ferreolensis as determined previously at Cassis-La Bédoule ( et alii, 2005). In addition, a greater sedimentation rhythm and overlapping continuous outcrops at La Tuilière made it possible to establish that the first appearance datum set by the first appearance of Praehedbergella luterbacheri is not just an approximation of the Bedoulian-Gargasian boundary, as previously suggested ( et alii, 2005), but coincides exactly with this limit. Moreover at La Tuilière (Les Gays 1 and 2 sections) the more abundant and undisturbed sedimentation permitted an excellent comprehension of the span of time involved between the appearances of the 7-chambered and 8-chambered forms of Globigerinelloides ferreolensis plexus. This made it possible to define and to insert within this zonation a horizon with G. ferreolensis heptacameratus (Table 1).
The main purpose of this article is to present an updated inventory of the foraminiferal microfaunas in sections largely straddling the levels of transition from the Bedoulian to the Gargasian in the type area of the Aptian stage. So we do not intend to delve very far into the recurring problem of the determination of the boundary between these substages. However, we can at least attempt a comparison of our micropaleontological data with the results of the most recent published works ( & , 2002, 2005; , 2005; , 2002; et alii, 2006) that deal with the ammonites from this interval in the same area and its surroundings.
In the Pichouraz section, the 0.50 m more indurated level (="NB", that includes our samples 2281-2282)
(Fig. 3 ![]() ) which we (
et alii, 2006) had suggested to compare with the Vocontian "Niveau Blanc", seems to correspond to the uppermost of four calcareous beds intercalated in marly sequences labelled "3a" and "5a-5b", as drawn by
(2005, fig. 18).
) which we (
et alii, 2006) had suggested to compare with the Vocontian "Niveau Blanc", seems to correspond to the uppermost of four calcareous beds intercalated in marly sequences labelled "3a" and "5a-5b", as drawn by
(2005, fig. 18).
In addition, taking into consideration its position in the series, the interruption that the author defines with a line between his levels 5b and 6a, just 6 m above the last of the four calcareous beds, could correspond to our "Change of Color"
(labelled "CO", Fig. 3 ![]() ). It is at this level that
(2005) places the upper limit of his Furcata
s.s. Zone.
). It is at this level that
(2005) places the upper limit of his Furcata
s.s. Zone.
The data that we obtained in Cassis-La Bédoule ( et alii, 2005; et alii, 2006) show that the micropaleontological boundary defined by the triple datum with Protocythere bedoulensis, Lenticulina cf. nodosa, and Praehedbergella luterbacheri is coincident with the top of the Furcata ammonite Zone, as defined in the Bedoulian stratotype.
For the moment, despite the recommendations of & (2002, 2005) to use the Furcata Zone as the basal subdivision of the middle Aptian (Gargasian), the quasi-consensus established at the last meetings of the Lower Cretaceous ammonite Working Group ("Kilian" Group) led rather to the proposal that the Furcata Zone be considered as the final zone of the lower Aptian (Bedoulian). We have adopted this position since our earlier work on the type-Bedoulian ( et alii, 1998) and it is followed here. So at La Tuilière the micropaleontological limit between Bedoulian and Gargasian is slightly (approximately 2 m) above the "Change of Color". It is interesting to note that in the Vocontian realm (2002) observed that this lithostratigraphic key-level is coincident with "the extinction of the genus Dufrenoyia".
In summary, our results show that 1) the micropaleontological limit as mentioned above, 2) the top of the Furcata Zone and 3) the "Change of Color" are practically coincident (± 1 m) in the stratotypic area of La Tuilière. The two (macro- and microfaunal) bio-events are also contemporaneous in the Bedoulian stratotype where however, probably because of the poor outcrops the colorimetric change observed at La Tuilière and in the Vocontien realm was not detected.
This lithostratigraphic key-level, mentioned as level "d" in our general presentation of the sector of La Tuilière ( et alii, 2006), is in the luterbacheri Zone near the bottom of the Les Gays 1 section and in those of La Tuilière E and W. We had suggested that it might correspond to the "Van Gogh" level described from the Vocontian Gargasian (, 2002). In La Tuilière it appears as an insert in the marls for almost ten meters: a sequence of brownish centimetric layers made up of pyritic nodules, each layer separated from the other by a marly interval of several tens of centimeters to one meter.
Our micropaleontological study shows that this episode is accompanied by a strong decrease in the abundance and diversity of the exobenthic (foraminifers, ornate ostracodes) and planktonic forms, whereas the endobenthic forms (Gavelinellas, "Valvulineria",
Coryphostoma, Cytherella gr. ovata-parallela) are much less affected
(Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() ).
).
If our proposal for a correlation between the Vocontian Van Gogh level and our level "d" (here labelled "VG" too) is valid, it seems that the episode (supposedly of a volcanic nature [, 2002]) produced more important repercussions and is better recorded at La Tuilière than in the Vocontian Basin. (2002) raised the question of the location of the site from which the material dispersed in the SE of France came. In the light of our data, it seems that this source may have been closer to La Tuilière than to the Vocontian Basin.
Most of the benthic forms observed in the Gargasian of Cassis-La Marcouline are also found in the sector of La Tuilière. However, some of them are curiously absent, or occur with very different frequencies of representation. Thus Spiroplectinata spp., Globorotalites spp., Falsogaudryinella moesiana, all abundant at La Marcouline, are extremely rare or absent at La Tuilière. In addition, the specimens of La Tuilière are in general of a size definitely smaller than that of their representatives in the Cassis sector.
Apart from these few differences, the overall paleobiogeographic affinities of the benthic stock represented at La Tuilière are strongly reminiscent of the microfaunas of Basse Provence. This stock does not have many similarities with the Vocontian assemblages which, however, are closer geographically. This might be explained by differences in bathymetry and/or a geographical barrier on or near the Ventoux-Lure axis.
As for the "Boreal" relationships, affinities are weak with the Paris Basin but curiously more obvious with Northern Germany (cf. similarities with the material described by , 1841, 1842; , 1863; & , 1962; and above all , 1995).
The crisis of diversity during the major part of the luterbacheri Zone of the La Tuilière region had neither been suspected nor mentioned until now in this area or anywhere else for that interval of time. The continuity of an exceptionally expanded section and dense sampling was necessary to detect and to measure its effects. Most presumably, the rhythmic spreading of a great amount of volcanic ash in the area of sedimentation seriously disturbed the life of epibenthic and planktonic microfaunas.
See the chapter entitled "Notes taxinomiques" included below in the French detailed version of this paper.
Because of an abundant, continuous and undisturbed sedimentation during the transition from Bedoulian to Gargasian, deposits in La Tuilière area recorded in great detail a succession of bio-events which had not previously been so precisely detected in any other area of the world. Our study enabled us to constitute a detailed inventory of its microfaunal content, which leads to the following results:
confirmation of the stratigraphic value of some benthic and planktonic markers;
strengthening of the cogency of the zonation by means of planktonic foraminifers which had been defined previously in Basse-Provence; this zonal scheme is now completed by the addition of a biohorizon (with Globigerinelloides ferreolensis heptacameratus nov. subsp.) which straddles the transition of the luterbacheri to ferreolensis zones;
quasi-coincidence of the triple micropaleontological datum (LAD of Protocythere bedoulensis - Lenticulina cf. nodosa / FAD of Praehedbergella luterbacheri) with the lithostratigraphic change called "change of color" together with the upper limit of the Furcata ammonite Zone; this limit is generally recognized as marking the boundary between the Bedoulian and Gargasian substages;
discovery of a strong decline in the diversity and abundance of epibenthic and planktonic Foraminifera during an important part of the luterbacheri Zone. This oligospecific crisis appears to be related to an episode of intense sedimentation of volcanic ash, which could correspond to the Van Gogh Level of the Vocontian Basin.
The richness of the collected material has also allowed us to provide additional taxonomic precisions for a certain number of benthic foraminiferal species; most of which are very rare or unknown elsewhere than the Aptian of Basse-Provence but, curiously, often present in certain sectors of the Boreal realm (e.g. N Germany).
The authors express their gratitude to Isabella (University of Milan), Silvia (University of Parma) and Danièle (University of Strasbourg) for their critical reading and suggestions which helped us to improve our manuscript.
They are also indebted to Nestor J. for his invaluable assistance in the writing of the abridged English version of this paper.
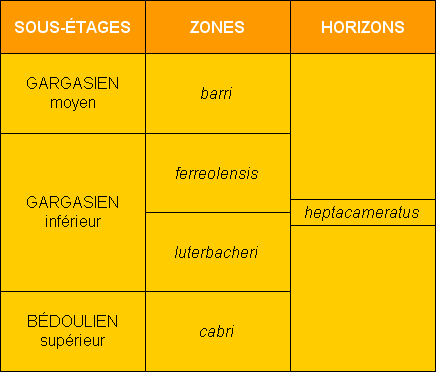
Dans une série d'articles récemment publiés, nous avons détaillé le contenu microfaunique (Foraminifères benthiques et planctoniques) des couches de l'Aptien inférieur (
et alii, 1998) et de l'Aptien moyen (
et alii, 2005) du stratotype historique du Bédoulien, à Cassis-La Bédoule (Bouches-du-Rhône, SE France). Dans la continuité de cette série, réalisée au sein d'un mémoire collectif des Carnets
de Géologie consacré plus particulièrement aux stratotypes de l'Aptien inférieur
et moyen, le présent article inclut l'étude de la distribution des Foraminifères recueillis dans une succession de coupes situées dans le secteur du hameau de La Tuilière (commune de Saint-Saturnin-lès-Apt, département du Vaucluse, SE France)
(Fig. 1 ![]() ). Quatre coupes principales (Pichouraz, La Tuilière W, Les Gays 1 & 2)
(Fig. 2
). Quatre coupes principales (Pichouraz, La Tuilière W, Les Gays 1 & 2)
(Fig. 2 ![]() ) nous ont permis d'observer en continuité et en grand détail les termes sommitaux du Bédoulien ainsi que le Gargasien inférieur, autrefois peu visibles dans la localité-type historique de Gargas, située à quelques kilomètres de La Tuilière.
) nous ont permis d'observer en continuité et en grand détail les termes sommitaux du Bédoulien ainsi que le Gargasien inférieur, autrefois peu visibles dans la localité-type historique de Gargas, située à quelques kilomètres de La Tuilière.
La localisation, la description lithologique et la corrélation géométrique des coupes, ainsi que le contexte géologique et historique du secteur de La Tuilière, ont déjà été exposés en deux notes préliminaires ( & ,
2004 ;
et alii, 2006) et ne seront donc pas repris en détail dans le présent article. Nous ajouterons seulement ici des précisions complémentaires, notamment lithostratigraphiques
(Fig. 3 ![]() ) et biostrati-graphiques, découlant des derniers résultats d'une étude plus avancée des microfaunes. Nous procéderons également aux mises à jour taxinomiques nécessaires, à la fois pour les formes benthiques et pour les formes
planctoniques.
) et biostrati-graphiques, découlant des derniers résultats d'une étude plus avancée des microfaunes. Nous procéderons également aux mises à jour taxinomiques nécessaires, à la fois pour les formes benthiques et pour les formes
planctoniques.
Les 87 échantillons qui ont servi de base à la présente étude ont été prélevés en 2003 et 2004, avec une maille d'un (voire moins) à deux mètres, juste avant que les secteurs de La Tuilière s.s. et de Pichouraz fassent l'objet d'une accélération de leur processus d'urbanisation. Pour le moment seul le secteur des Gays (non aedificandi) demeure relativement préservé, bien que son usage comme piste de moto-cross ainsi qu'une intense fréquentation par des scolaires et des collectionneurs de fossiles conduisent à une dégradation notoire de la qualité des affleurements. Nos tentatives auprès des édiles responsables pour faire protéger l'ensemble du site, d'un grand intérêt géologique, n'ont malheureusement pas été entendues, bien au contraire.
Les échantillons, à prédominance marneuse pour la plupart, ont d'abord été préparés selon la méthode d' (1962), comprenant un trempage à l'hyposulfite de sodium suivi d'un ajout d'eau oxygénée. Les résultats n'étant pas excellents avec cette méthode (persistance de l'encro¹tement des tests), nous avons utilisé avec plus de succès le traitement par le Rewoquat W 3690 (voir détails in et alii, 2005), suivis d'un tamisage à 45 Ąm.
L'état de conservation des microfossiles aptiens de La Tuilière s'est révélé variable, allant de médiocre à moyen. Le plus souvent les tests ont subi une forte dissolution diagénétique, succédant à un remplissage syn-sédimentaire (en fait surtout constitué de nannofossiles calcaires), parfois plus ou moins pyritisé. Dans de telles conditions de fossilisation, il n'a pas été possible d'observer les détails microstructuraux, parfois même (foraminifères agglutinants complètement épigénisés) la nature de l'ouverture ou la disposition des loges.
Le contenu microfaunique des échantillons étudiés a été considéré sous l'aspect quantitatif, avec comptage systématique des spécimens de chaque espèce de Foraminifères. Nous avons utilisé dans les tableaux de répartition
(Figs. 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() ) les mêmes classes de fréquence que celles qui ont été définies pour le secteur de Cassis-La Bédoule (
et alii, 2005).
) les mêmes classes de fréquence que celles qui ont été définies pour le secteur de Cassis-La Bédoule (
et alii, 2005).
L'ensemble du matériel (témoins d'échantillons, résidus de lavage, cellules) est conservé dans les collections du Musée de Paléontologie de l'Université de Provence à Marseille. Les types et figurés sont déposés dans les collections de micropaléontologie du Muséum National d'Histoire Naturelle à Paris.
Les espèces de Foraminifères benthiques sont en général caractérisées par une répartition stratigraphique s'étageant sur des périodes assez longues. Leur présence ou leur absence, leurs moments d'apparition et d'extinction sont aussi réputés être plus dépendants des conditions de milieu que de la chronologie. Le secteur de La Tuilière, qui n'expose que les termes sommitaux du Bédoulien et le Gargasien inférieur, soit un intervalle stratigraphique relativement bref, ne constitue donc pas a priori l'endroit idéal pour apporter des indications significatives sur la répartition temporelle de ces microfossiles. Il pourra s'avérer nécessaire de procéder à des comparaisons avec d'autres domaines (Basse-Provence, Bassin Vocontien, voire Bassin de Paris ou NW Allemagne et même Caraïbes) pour juger du bien-fondé et de la valeur de certains "datums" de formes benthiques mis en évidence dans le stratotype, au demeurant jusqu'à présent très peu étudié (, 1965 ; , 1975) sur ce plan.
En ce qui concerne les Foraminifères planctoniques, nos travaux récents nous avaient conduit à densifier une zonation déjà très fine pour cette période, marquée par une forte anagenèse. Le secteur de Cassis-La Bédoule ( et alii, 2005) nous avait ainsi permis de mieux définir un enchaînement de trois zones de foraminifères (cabri, luterbacheri et ferreolensis) pour découper l'intervalle stratigraphique qui fait l'objet de la présente étude. Il subsistait cependant quelques incertitudes à Cassis, découlant d'une relative condensation sédimentaire et d'une exposition discontinue au niveau des termes chevauchant la limite Bédoulien-Gargasien. Nous verrons ci-après que les résultats obtenus à la Tuilière, où la série correspondante est plus dilatée et intégralement exposée à ce niveau ( et alii, 2006), permettent de compléter et de confirmer les données du secteur de Cassis-La Bédoule, voire même d'affiner encore le découpage zonal.
Globorotalites bartensteini
À Cassis-La Bédoule nous avions observé que cette espèce bédoulienne ne franchissait pas la limite Bédoulien-Gargasien et que par son extinction elle pouvait donc constituer un marqueur benthique d'appoint jalonnant la limite entre les deux sous-étages. Cette donnée n'a pas pu être confirmée dans le secteur de La Tuilière, car les Globorotalites n'y sont pratiquement pas représentés. G. aptiensis est totalement absente, et trois exemplaires seulement de G. bartensteini ont été recueillis, en trois échantillons distincts (deux à Pichouraz : à 0,5 m sous la limite précitée et 11, 5 m au-dessus ; un à La Tuilière W : 1,5 m au-dessus de la limite).
Astacolus crepidularis tricarinella
À Cassis-La Bédoule, aussi bien dans la coupe des Tocchis que dans celle de la Carrière Comte, cette forme ne franchit que de quelques mètres la limite Bédoulien-Gargasien. Dans la coupe de Pichouraz son extinction se produit 0,5 m au dessus de ladite limite. Mais elle "réapparaît" brusquement de manière assez inexplicable, en 13 exemplaires (!), dans un seul échantillon de la coupe des Gays 1, situé à plus d'une quarantaine de mètres au dessus de la limite. Un aussi grand nombre d'exemplaires, concernant une seule espèce, conduit à exclure l'hypothèse d'une contamination accidentelle ou d'un remaniement sédimentaire. Cette surprenante donnée, qui demande cependant à être confirmée en d'autres coupes, amoindrit quelque peu l'intérêt de cette forme comme marqueur stratigraphique.
Lenticulina cf. nodosa
Cette espèce, constante dans le Bédoulien, paraît constituer le seul foraminifère benthique vraiment fiable pour déterminer la limite Bédoulien-Gargasien, avec une extinction nette, juste à ce niveau à Cassis-La Bédoule et à Pichouraz, et à 0,5 m seulement au dessus de la limite à La Tuilière W.
Saracenaria sp. aff. frankei
Cette forme, absente au Bédoulien et au Gargasien basal, apparaît dans la partie supérieure de la Zone à luterbacheri, où elle est rare, et devient plus fréquente dans la Zone à ferreolensis (coupes des Gays 1 et 2). On la retrouve, plus sporadiquement, dans des niveaux similaires de la coupe de La Marcouline (Cassis).
Oolina apiculata
Cette espèce, toujours rare, n'est présente que dans l'extrême sommet de la Zone à luterbacheri et dans la Zone à ferreolensis (coupes des Gays 1 et 2).
Schackoina gr. cabri-pustulans
Les données obtenues aussi bien à Cassis-La Bédoule qu'à La Tuilière
(Figs. 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() ) confirment que les "espèces" individualisées par certains auteurs au sein de ce plexus morphologique ont toutes exactement la même répartition stratigraphique. Nous avons donc choisi de continuer à les réunir taxinomiquement en un seul groupe, qui apparaît au Bédoulien supérieur, avec un acmé correspondant au Bédoulien terminal et à l'extrême base du
GargasienĀ; il devient ensuite plus sporadique, et son extinction se produit au niveau du début de l'acmé de Globigerinelloides ferreolensis.
) confirment que les "espèces" individualisées par certains auteurs au sein de ce plexus morphologique ont toutes exactement la même répartition stratigraphique. Nous avons donc choisi de continuer à les réunir taxinomiquement en un seul groupe, qui apparaît au Bédoulien supérieur, avec un acmé correspondant au Bédoulien terminal et à l'extrême base du
GargasienĀ; il devient ensuite plus sporadique, et son extinction se produit au niveau du début de l'acmé de Globigerinelloides ferreolensis.
Praehedbergella luterbacheri
Dans la coupe de Pichouraz (Fig. 4 ![]() ), P. luterbacheri est présente à partir de l'échantillon n°2367, 2 m au dessus de l'extinction de l'ostracode Protocythere bedoulensis (
et alii, 2006 ; et alii,
2007) et 1 m au dessus de l'extinction du foraminifère benthique Lenticulina cf. nodosa. Dans la coupe de La Tuilière W, échantillonnée en plus grand détail, l'apparition de P. luterbacheri coïncide avec l'extinction de P. bedoulensis et précède de 0,4 m l'extinction de
L. nodosa. Les datums fondés soit sur l'extinction des deux formes benthiques soit sur l'apparition du taxon planctonique sont donc quasiment synchrones et se placent très peu au dessus du repère lithostratigraphique dit "changement de couleur", tel qu'il a pu être observé à Pichouraz
(Fig. 4
), P. luterbacheri est présente à partir de l'échantillon n°2367, 2 m au dessus de l'extinction de l'ostracode Protocythere bedoulensis (
et alii, 2006 ; et alii,
2007) et 1 m au dessus de l'extinction du foraminifère benthique Lenticulina cf. nodosa. Dans la coupe de La Tuilière W, échantillonnée en plus grand détail, l'apparition de P. luterbacheri coïncide avec l'extinction de P. bedoulensis et précède de 0,4 m l'extinction de
L. nodosa. Les datums fondés soit sur l'extinction des deux formes benthiques soit sur l'apparition du taxon planctonique sont donc quasiment synchrones et se placent très peu au dessus du repère lithostratigraphique dit "changement de couleur", tel qu'il a pu être observé à Pichouraz
(Fig. 4 ![]() ).
).
Globigerinelloides ferreolensis heptacameratus nov. subsp.
Dans la coupe des Gays 1, les formes à 7 loges au dernier tour du gr. ferreolensis apparaissent les premières, au niveau de l'échantillon 2328, soit environ 12 m avant l'apparition des formes à 8 loges
(Fig. 6 ![]() ). Sur les 6 premiers mètres de cette répartition, qui correspondent à l'extrême sommet de la Zone à luterbacheri,
G. ferreolensis heptacameratus nov. subsp. est assez rare et pas toujours facile à différencier de P. luterbacheri, son ancêtre phylogénétique direct (
et alii, 2002, 2005). Elle est plus fréquente sur les 6 m suivants, avant l'apparition des formes à 8 loges (=
G. ferreolensis ferreolensis).
). Sur les 6 premiers mètres de cette répartition, qui correspondent à l'extrême sommet de la Zone à luterbacheri,
G. ferreolensis heptacameratus nov. subsp. est assez rare et pas toujours facile à différencier de P. luterbacheri, son ancêtre phylogénétique direct (
et alii, 2002, 2005). Elle est plus fréquente sur les 6 m suivants, avant l'apparition des formes à 8 loges (=
G. ferreolensis ferreolensis).
Globigerinelloides ferreolensis ferreolensis
Dans les coupes des Gays 1 et 2, la sous-espèce à 8 loges au dernier tour du gr. ferreolensis apparaît 6 m au dessus du début de l'acmé du groupe, soit au niveau de l'extinction des Schackoïnes du gr. cabri
(Figs. 6 ![]() - 7
- 7 ![]() ).
).
Globigerinelloides barri
Dans le secteur de La Tuilière, nous n'avons observé qu'un seul exemplaire de cette espèce, dans le dernier échantillon (n° 2301) de la coupe des Gays 2. Cet échantillon constitue le dernier point de prélèvement possible dans une zone de marnes terreuses de plus en plus altérées, passant ensuite à la terre végétale, située à proximité des colluvions quaternaires qui viennent régionalement tronquer les marnes gargasiennes. Ces mauvaises conditions d'affleurement nous conduiraient à douter que cet unique exemplaire de G. barri puisse signifier que la zone éponyme soit atteinte dans la coupe des Gays 2. Il faut toutefois noter qu'à La Marcouline (Cassis-La Bédoule) les premiers G. barri apparaissent environ 12 m au dessus de la base de la Zone d'acmé de Globigerinelloides ferreolensis. Dans la coupe des Gays 2, appartenant à une série dans l'ensemble beaucoup plus dilatée qu'à Cassis, environ 25 m séparent ce même datum à ferreolensis de l'échantillon 2301. Il s'agit là d'un ordre de grandeur qui n'est donc pas incompatible avec une limite entre les deux zones juste touchée au sommet de la séquence échantillonnable.
L'intervalle stratigraphique étudié dans le secteur de La Tuilière nous permet de confirmer la succession zonale cabri-luterbacheri-ferreolensis déterminée à Cassis-la Bédoule ( et alii, 2005). La plus grande dilatation de la série et une meilleure continuité d'affleurement à La Tuilière permettent en outre de constater que le datum de l'apparition de Praehedbergella luterbacheri ne constitue pas une approximation de la limite Bédoulien-Gargasien, comme il avait été suggéré précédemment ( et alii, 2005), mais qu'il coïncide avec elle. De plus à la Tuilière (coupes des Gays 1 et 2), la sédimentation à la fois plus abondante et non perturbée à conduit à une excellente appréhension du décalage d'apparition entre les formes à 7 loges et les formes à 8 loges au dernier tour du plexus Globigerinelloides ferreolensis. Nous avons ainsi pu définir et insérer au sein de cette zonation un horizon repère à G. ferreolensis heptacameratus (Tableau 1).
Le terme d'horizon est utilisé ici dans son acception biostratigraphique, soit "un intervalle fossilifère pourvu d'une épaisseur temporelle (...) nettement distinct" (, 1997, p. 76). Cet horizon mesure 12 mètres aux Gays et seulement 6 mètres dans la coupe de La Marcouline à Cassis-La Bédoule.
L'objectif principal du présent article est surtout de présenter un inventaire actualisé des microfaunes de Foraminifères provenant d'un ensemble de coupes chevauchant largement les niveaux de transition du Bédoulien au Gargasien dans la région-type de l'étage Aptien. Nous n'avons donc pas l'intention d'entrer très avant dans le problème récurrent de la détermination de la limite entre les deux sous-étages, d'autant plus que jusqu'à présent il ne nous a pas été possible de travailler directement sur le terrain en compagnie d'un des nombreux spécialistes d'ammonites qui ont effectué, de leur côté, des récoltes de fossiles dans ces mêmes gisements.
Pour ce qui est de la biostratigraphie des ammonites du stratotype, les travaux (publiés) de révision les plus récents et les plus complets sont l'£uvre d' & (2002, 2005) et de
(2005). Nous nous sommes rendu compte à l'usage que la corrélation de détail (épaisseurs, lithostratigraphie) entre leurs coupes et les nôtres est assez difficile, sauf peut-être pour la coupe de Pichouraz : il est ainsi pratiquement certain que le faisceau plus induré (= "NB", incluant nos échantillons 2281-2282,
Fig. 3 ![]() ), épais de 0,50 m, que nous avons suggéré d'assimiler au "Niveau Blanc" vocontien (
et alii, 2006), doit correspondre au dernier des quatre niveaux calcaires figurés par
(2005, fig. 18). Au sein de sa coupe dite de Pichouras, il s'agit de bancs qui s'intercalent entre des passées marneuses portant la mention "3a" et "5a-5b". D'autre part, par sa position dans la série, la coupure que l'auteur matérialise par un trait entre ses niveaux 5b et 6a, 6 m au dessus du faisceau calcaire précité, mais sans préciser si elle correspond à un repère lithologique ou colorimétrique particulier, pourrait peut-être correspondre à notre "changement de couleur" (niveau "CO",
Fig. 3
), épais de 0,50 m, que nous avons suggéré d'assimiler au "Niveau Blanc" vocontien (
et alii, 2006), doit correspondre au dernier des quatre niveaux calcaires figurés par
(2005, fig. 18). Au sein de sa coupe dite de Pichouras, il s'agit de bancs qui s'intercalent entre des passées marneuses portant la mention "3a" et "5a-5b". D'autre part, par sa position dans la série, la coupure que l'auteur matérialise par un trait entre ses niveaux 5b et 6a, 6 m au dessus du faisceau calcaire précité, mais sans préciser si elle correspond à un repère lithologique ou colorimétrique particulier, pourrait peut-être correspondre à notre "changement de couleur" (niveau "CO",
Fig. 3 ![]() ). Cependant nous avons mesuré une épaisseur de 9,5 m, et non de 6 m, entre ces deux niveaux-repères. À chaque fois qu'une corrélation précise est possible, il nous est apparu que nos mesures d'épaisseur étaient assez systématiquement supérieures (d'un coefficient de ± 1,3) à celles qui sont fournies par
(2005). En tenant compte de ce biais, la corrélation précitée devient plausible. Retenons que c'est à ce même niveau que
(2005) place le toit de sa Zone à Furcata
s.s..
). Cependant nous avons mesuré une épaisseur de 9,5 m, et non de 6 m, entre ces deux niveaux-repères. À chaque fois qu'une corrélation précise est possible, il nous est apparu que nos mesures d'épaisseur étaient assez systématiquement supérieures (d'un coefficient de ± 1,3) à celles qui sont fournies par
(2005). En tenant compte de ce biais, la corrélation précitée devient plausible. Retenons que c'est à ce même niveau que
(2005) place le toit de sa Zone à Furcata
s.s..
Les données que nous avons obtenues à Cassis-La Bédoule ( et alii, 2005 ; et alii, 2006) montrent que la limite micropaléontologique fondée sur le triple datum à Protocythere bedoulensis, Lenticulina cf. nodosa, et Praehedbergella luterbacheri, coïncide pratiquement avec le toit de la Zone d'ammonites à Furcata, telle que définie dans le stratotype du Bédoulien. Pour le moment, en dépit des préconisations d'& (2002, 2005), qui souhaitent faire de la Zone à Furcata la subdivision basale de l'Aptien moyen (Gargasien), le quasi-consensus établi lors des dernières réunions du Working Group sur les ammonites du Crétacé inférieur (Groupe "Kilian"), sous l'égide de la Sous-Commission de Stratigraphie du Crétacé de l'U.I.S.G., aboutit plutôt à la proposer comme la zone terminale de l'Aptien inférieur (Bédoulien). C'est la position que nous suivons depuis nos travaux sur le Bédoulien-type ( et alii, 1998) et que nous avons également adoptée dans le présent article. Dans ces conditions, il apparaît qu'à la Tuilière la limite micropaléontologique entre Bédoulien et Gargasien se situe peu au dessus (2 m environ) du "changement de couleur". Il est enfin intéressant de noter que dans le domaine vocontien (2002) fait coïncider ce repère lithostratigraphique avec "l'extinction du genre Dufrenoyia".
En résumé, nos résultats montrent que notre limite micropaléontologique précitée, le toit de la Zone à Furcata et le changement de couleur sont pratiquement coïncidents (à ± 1 m près) dans l'aire stratotypique de la Tuilière. Les deux bio-événements le sont également dans le stratotype du Bédoulien où, probablement à cause des conditions d'affleurement, le changement colorimétrique observé à La Tuilière et dans le domaine Vocontien n'a pu être détecté.
Dans notre article de présentation générale du secteur de La Tuilière ( et alii, 2006), nous avons évoqué la possibilité que ce repère lithostratigraphique (mentionné comme niveau "d" dans notre coupe), observé au sein de la Zone à luterbacheri vers la base de la coupe des Gays 1 et dans les coupes de La Tuilière E et W, puisse correspondre au niveau "Van Gogh" du Gargasien Vocontien (, 2002). À la Tuilière il apparaît, sur près d'une dizaine de mètres, comme une séquence de lits centimétriques brunâtres de nodules pyriteux, distants les uns des autres de plusieurs dizaines de centimètres à un mètre, insérés au sein de la série marneuse.
Sur le plan micropaléontologique cet épisode s'accompagne d'une forte chute d'abondance et de diversité des formes exobenthiques (Foraminifères, Ostracodes ornés) et planctoniques, les formes endobenthiques (Gavelinelles, "Valvulineria",
Coryphostoma, Cytherella gr. ovata-parallela) étant beaucoup moins affectées. Dans le détail
(Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() ), il apparaît que la crise de diversité des microfaunes précède de quelques mètres l'épisode détecté macroscopiquement et continue bien au delà, sur une vingtaine de mètres, avec deux rémissions plus ponctuelles. En fait la crise précitée s'étage sur une bonne part, médiane et épaisse d'une quarantaine de mètres, de la Zone à luterbacheri.
), il apparaît que la crise de diversité des microfaunes précède de quelques mètres l'épisode détecté macroscopiquement et continue bien au delà, sur une vingtaine de mètres, avec deux rémissions plus ponctuelles. En fait la crise précitée s'étage sur une bonne part, médiane et épaisse d'une quarantaine de mètres, de la Zone à luterbacheri.
Si notre proposition de corrélation entre le repère vocontien (niveau Van Gogh) et notre niveau VG est valable, il semble que l'épisode (présumé de nature volcanique [, 2002]) qui lui a donné naissance induit des répercussions plus importantes et est enregistrée de manière plus démultipliée à La Tuilière que dans le Bassin vocontien. Sur la base des données vocontiennes, (2002) s'est posé la question du lieu d'origine du matériel ainsi épandu dans le SE de la France. À la lumière de nos données, il semblerait que ce dernier soit encore plus proche de La Tuilière que du Bassin vocontien. Des études plus spécifiques portant sur nos échantillons sont en cours afin de tenter de mieux élucider la nature (physique, chimique, etc.) du phénomène générateur des perturbations affectant plus ou moins périodiquement la microfaune.
Parmi les formes benthiques observées dans le Gargasien de Cassis-La Marcouline, un grand nombre se retrouve dans le secteur de La Tuilière. Toutefois quelques unes en sont curieusement absentes, ou alors avec des fréquences de représentation très différentes. Ainsi Spiroplectinata spp., Globorotalites spp., Falsogaudryinella moesiana, abondants à La Marcouline, sont rarissimes ou absents à La Tuilière. D'autre part, les spécimens de La Tuilière sont en général d'une taille nettement plus petite que celle de leurs homologues du secteur de Cassis. Quoiqu'il en soit de ces différences, les affinités paléobiogéographiques du stock benthique représenté à la Tuilière sont globalement fortes avec les microfaunes de Basse-Provence. Ce stock ne présente pas beaucoup de similitudes avec les associations du domaine vocontien, pourtant proche géographiquement. Dans ce cas, à la fois la profondeur (bien plus importante dans le Bassin vocontien que dans le Bassin d'Apt ou le "Sillon" de Cassis-La Bédoule), ainsi que l'existence possible d'une barrière géographique au niveau de (ou proche de) l'axe Ventoux-Lure, peuvent expliquer ces différences. Pour ce qui est des régions "boréales", les affinités sont peu marquées avec le Bassin de Paris mais curieusement plus évidentes avec l'Allemagne du Nord (cf. le matériel décrit par , 1841, 1842 ; , 1863 ; & , 1962 ; et surtout , 1995).
La crise de diversité qui s'instaure pendant la majeure partie de la Zone à luterbacheri dans le secteur de la Tuilière n'avait jamais été soupçonnée ni signalée jusqu'à présent dans la région ni même ailleurs à cette époque. Il fallait la continuité d'une coupe exceptionnellement dilatée et un échantillonnage dense pour en mesurer les effets. On peut présumer que l'épandage rythmique d'une grande quantité de cendres volcaniques dans le milieu de sédimentation ait sérieusement perturbé la microfaune épibenthique et planctonique.
Ammobaculites sp. gr. reophacoides/parvispira-subcretaceus
Pl. 1 ![]() , figs. 4-11
, figs. 4-11
Ammobaculites subcretacea & , 1930, p. 6, Pl. 2, figs. 9-10.
Ammobaculites parvispira , 1950, p. 10, Pl. I, fig. 8 a-b.
Ammobaculites reophacoides , 1952, p. 307, Fig. 1 a-b.
Les spécimens attribuables au genre Ammobaculites sont assez régulièrement représentés, avec une fréquence non négligeable, tout au long des coupes du Bédoulien terminal-Gargasien inférieur du secteur de La Tuilière. Leur importante variabilité porte sur les caractères suivants :
l'enroulement initial, qui peut être peu marqué, car n'impliquant que les trois premières loges, ou plus net, cinq à six loges étant concernées, avec des stades intermédiaires,
le nombre de loges du stade rectilinéaire (cinq en moyenne),
le degré d'accroissement en largeur des loges du stade rectilinéaireĀ: de quasi-nul (d'où un test à faces subparallèles) à un peu plus marqué, d'où une forme de test plus évasée,
la forme de la dernière loge (au demeurant souvent partiellement brisée) : chez les rares spécimens où elle est bien conservée, le sommet est en général arrondi, avec un rétrécissement sommital pouvant esquisser une sorte d'amorce de col.
Le test, de section normalement sub-circulaire, peut aussi avoir été plus ou moins aplati lors de la fossilisation, ce qui altère ses caractéristiques.
Au sein des populations étudiées on peut isoler deux morphotypes, minoritairesĀ: l'un à la spire initiale discrète
(Pl. 1 ![]() , figs.
9-11), évoquant A. reophacoides ,
1952 (qui est peut-être un synonyme plus récent
d'A. parvispira , 1950), l'autre avec un départ spiralé plus important
(Pl. 1
, figs.
9-11), évoquant A. reophacoides ,
1952 (qui est peut-être un synonyme plus récent
d'A. parvispira , 1950), l'autre avec un départ spiralé plus important
(Pl. 1 ![]() , figs.
4-6,
etc.), comme chez
A. subcretaceus & ,
1930 ; ces deux morphotypes sont reliés par de nombreuses formes intermédiaires (cf.
Pl. 1
, figs.
4-6,
etc.), comme chez
A. subcretaceus & ,
1930 ; ces deux morphotypes sont reliés par de nombreuses formes intermédiaires (cf.
Pl. 1 ![]() , figs.
7-8).
, figs.
7-8).
Haplophragmoides? sp.
Pl. 3 ![]() , figs. 1-8
, figs. 1-8
Des formes au test planispiralé involute et d'apparence générale symétrique, avec 8 à 10 loges au dernier tour, constituent un stock assez constant parmi les populations de foraminifères au test agglutinant du Bédoulien terminal ¢ Gargasien inférieur du secteur de La Tuilière. En raison des conditions de fossilisation et de diagenèse, leur test est souvent plus ou moins déformé. La paroi semble également pratiquement dissouteĀ; l'on n'observe en fait que des moules internes, formés par une gangue de remplissage argilo-calcaire. À cause de ce remplissage, en dépit de tentatives d'observation après immersion dans la glycérine, nous n'avons pu discriminer avec certitude la nature de l'enroulement interne initial. La mauvaise conservation de ces tests (dont la face aréale de la dernière loge est souvent abrasée) ne permet pas non plus de se faire une idée précise de la nature et de la disposition de l'ouverture. Pour ces raisons nous n'avons pu formellement exclure une attribution de ces spécimens au genre Recurvoides, en particulier pour ceux qui montrent une très légère dissymétrie au stade du dernier tour.
Leur assignation à une espèce précise n'a pas non plus été possible. Des Haplophragmoides plus ou moins déformés (H. subagrestis, H. granifer) décrits de l'Aptien inférieur de la région de Krasnodar par (in et alii, 1964) évoquent nos formes mais possèdent nettement moins de loges (5).
Falsogaudryinella moesiana (, 1965)
Pl. 3 ![]() , figs. 9-11
, figs. 9-11
Uvigerinammina moesiana , 1965, p. 5, Pl. 2, figs. 11-18.
Falsogaudryinella moesiana ()Ā; et alii, 1998, p. 201, Pl. 1, figs. 1-4.
Cette espèce, pourtant bien représentée dans le Bédoulien et le Gargasien de Cassis-La Bédoule ( et alii, 1998, 2005), est curieusement absente à la fois dans le Bédoulien terminal et le Gargasien basal (Zone à luterbacheri) dans le secteur de La Tuilière. On ne l'y observe, rare et très sporadique, qu'au sommet de la Zone à luterbacheri et dans la Zone à ferreolensis.
Falsogaudryinella tealbyensis (, 1956)
Pl. 3 ![]() , figs. 12-15
, figs. 12-15
Gaudryinella tealbyensis , 1956, p. 513-514, Fig. 3 ; Pl. 3, fig. 63.
Falsogaudryinella tealbyensis ()Ā; et alii, 1998, p. 202, Pl. 1, figs. 5-10.
Contrairement à F. moesiana, F. tealbyensis est assez constamment présente à La Tuilière, avec une fréquence généralement modérée à moyenne, voire épisodiquement forte dans les quelques mètres précédant le datum de l'apparition de Globigerinelloides ferreolensis.
Gaudryinella sherlocki , 1952
Pl. 1 ![]() , figs. 20-22
, figs. 20-22
Gaudryinella sherlocki , 1952, p. 268, Pl. 1, figs. 1-5.
Gaudryina pschechaensis & in et alii, 1964, p. 43, Pl. 6, figs. 7a-b.
Gaudryinella sherlocki Ā; & , 1977, p. 547, Pl. 1, figs. 21-22.
Gaudryinella sherlocki ahlumensis , 1995, p. 81, Pl. 3, figs. 7-9.
Gaudryinella sherlocki sherlocki Ā; , 1995, Pl. 3, fig. 6.
Gaudryinella sherlocki ; et alii, 1998, p. 202, Pl. 2, figs. 8-9.
Cette espèce barrémo-aptienne (plutôt barrémo-bédoulienne) est rare et assez sporadique dans le Bédoulien terminal et le Gargasien basal du secteur de La Tuilière, où elle ne dépasse pas la partie inférieure de la Zone à luterbacheri. Quand il est bien conservé, le test montre une section subcirculaire, mais les spécimens écrasés par suite de la compaction diagénétique dominent. Comme souvent chez cette espèce, l'ouverture terminale est peu ou pas discernable. Nous ne suivons pas (1995), qui a créé une nouvelle sous-espèce (Gaudryinella sherlocki ahlumensis) sur la base d'un nombre de loges plus faible que chez la forme-type, mais reconnaît lui-même que les deux sous-espèces sont des types extrêmes, coexistant avec des cas intermédiaires. L'excellent état de conservation de son matériel permet de se faire une idée précise de la terminaison aperturale, la loge se rétrécissant progressivement, dans sa partie apicale, en une sorte de col.
Textulariopsis sp.
Textulariopsis sp., et alii, 1998, p. 203.
L'on retrouve sporadiquement, dans le Bédoulien sommital et le Gargasien basal du secteur de La Tuilière, quelques rares exemplaires de cette forme, déjà identifiée à Cassis-La Bédoule ( et alii, 1998). Sa rareté et son mauvais été de conservation (tests souvent brisés, dépourvus du stade initial, mince et comprimé, donc fragile) ne nous ont pas permis d'obtenir des figurations présentables à partir du matériel de La Tuilière. Cette forme est mieux représentée, mais pas mieux conservée, durant tout le Bédoulien et le Gargasien inférieur-moyen dans le secteur de Cassis-La Bédoule.
Genre Praedorothia & , 1987
Il est très difficile de répartir et d'attribuer à des espèces précises les morphotypes relevant de la famille des Dorothiidae dans le matériel aptien de La Tuilière. Tous les stades intermédiaires peuvent être observés entre un morphotype très "acuminé", au diamètre croissant lentement à mesure de l'ontogénie, et un morphotype au diamètre croissant rapidement, donnant un aspect beaucoup plus évasé au test. Durant l'ontogénie, le rythme d'accroissement du diamètre du test peut être régulier ou bien montrer des stades alternant à vitesse d'accroissement différente (par exempleĀ: stade initial assez rapide ¢ stade médian plus lent ¢ stade final plus rapide), d'où un aspect en ligne brisée de la génératrice du cône. Les deux dernières loges du stade bisérié peuvent aussi être plus ou moins renflées ou au contraire présenter un rebord plus ou moins abrupt, la face aperturale pouvant ainsi être plus ou moins convexe ou concave. Cette dernière variation morphologique (loges à septa convexes vs loges à septa concaves) a même été proposée par & (1987) comme critère distinctif de rang générique (Praedorothia versus Protomarssonella).
Ce critère fondé sur l'aspect des septa est peu applicable dans la réalité : 1) il nécessite de procéder à une section longitudinale du test pour sa discrimination, car il arrive fréquemment que la forme de la face supérieure des deux dernières loges, seule visible en épiscopie, ne reflète pas celle des loges plus anciennes, qui est seulement visible en sectionĀ; 2) là encore des stades intermédiaires existent entre les extrêmes utilisés par & (1987) pour justifier la séparation en deux genres. Nous préférons donc n'en utiliser qu'un seul, le plus ancien par priorité de page dans la publication des auteurs, soit Praedorothia, décrit en p. 18, Protomarssonella ne l'étant qu'en p. 24.
La fluctuation des caractéristiques, supposées d'ordre spécifique et générique, de ces formes aptiennes a également été remarquée par (1995) dans le Gargasien du NW de l'Allemagne. Elles ont malheureusement incité beaucoup d'auteurs à multiplier, sur des bases le plus souvent purement typologiques, les descriptions de nouveaux taxons. Leur utilisation s'avère illusoire au vu de la variabilité montrée par les populations de notre matériel. Au plan spécifique nous ne retiendrons en fait que trois morphogroupes à grande longévité, que nous labelliserons par les noms de la ou des "espèces" paraissant correspondre au type le plus représentatif, et une espèce qui paraît mieux s'individualiser et dont la distribution semble plus limitée dans le tempsĀ:
Praedorothia sp. aff. kummi, pour les tests de forme étroite, effilée,
P. sp. gr. hechti-subtrochus, pour les tests moyennement évasés,
P. sp. aff. trochus, pour les tests fortement évasés.
P. sp. aff. pupa.
Praedorothia sp. aff. kummi (, 1961)
Pl. 2 ![]() , figs. 1-4
, figs. 1-4
Marssonella kummi , 1961, p. 31, Pl. 7, figs. 1a-c.
P. kummi a été décrite initialement à partir d'un matériel d'âge hauterivien d'Allemagne du N, pour des formes éocrétacées jusque là rapportées par les auteurs à l'espèce oxycona (du Crétacé supérieur) et mentionnées sous ce nom dans le Néocomien-Barrémien de maints secteurs de la Tethys. Dans le Bédoulien terminal-Gargasien inférieur de la Tuilière, faute de mieux, nous regroupons sous le vocable "aff. kummi" de rares Praedorothia au test résolument conique, effilé, à accroissement lent en diamètre, de section subcirculaire. Des formes analogues, mais pas tout à fait identiques, ont été observées dans l'Aptien inférieur de Cassis-La Bédoule ("Protomarssonella cf. kummi" in et alii, 1998). Nous avions par ailleurs dénommé comme "Praedorothia cf. conula" des spécimens du Gargasien inférieur-moyen de Cassis ( et alii, 2005), qui après réexamen paraissent bien s'inscrire dans la mouvance de ceux de La Tuilière. Nous préférons rapprocher cet ensemble aptien de l'espèce de , qui montre un aspect général "en pointe" un peu plus marqué que chez P. conula et dont la morphologie évoque ainsi mieux celle de nos populations.
Praedorothia sp. gr. hechti (?) - subtrochus
Pl. 2 ![]() , figs. 7-11
, figs. 7-11
Valvulina D 1 , 1938, Pl. 3a, figs. 46-50 ; Pl. 6a, figs. 39-40.
Marssonella cf. trochus (d') ; et alii, 1957, p. 20-21, Pl. 3, figs. 44-45a-b.
Marssonella subtrochus , 1962, p. 137-139, Pl. 15, figs. 3-5.
? Dorothia hechti & , 1966, p. 106-107, Pl. II, figs. 17-22 ; Pl. X, figs. 4-8.
Praedorothia cf. trochus (sensu et alii) ; et alii, 1998, p. 203.
Praedorothia cf. trochus, et alii, 2005, in tableaux-fig. 3.
Ce groupe intermédiaire entre les deux morphotypes extrêmes que constituent P. sp. aff. kummi et P. sp. aff. trochus se révèle comme le plus polymorphe au sein des Praedorothia du Crétacé inférieur. Il concentre par conséquent un maximum de problèmes d'ordre nomenclatural.
et alii (1957) décrivent et figurent sous la dénomination "Marssonella cf. trochus" des formes du Barrémien de La Trinité (domaine caraïbe), en mentionnant qu'elles sont identiques à celles de l'Aptien et de l'Albien (pro parte) d'Europe. Au sein de ces dernières ils citent la "Valvulina D 1" du NW de l'Allemagne, figurée par en 1938. La description fournie par et alii (1957) est sommaire et consiste surtout en une énumération des différences que cette forme présente avec M. oxycona (rebords de la surface aperturale plus anguleux, test en forme de cône plus tranchant, surface aperturale plus plate, uniformément ronde, en forme d'assiette). Les figurations montrent un test en forme de cône régulier plutôt évasé, avec une surface aperturale plutôt plane.
En 1962, décrit longuement une nouvelle espèce, Marssonella subtrochus. Les informations découlant des figurations, de la liste synonymique, de la description et de la discussion sont contradictoires. Le but avéré de l'auteur, par la création de cette espèce, est de donner un nom à un concept de formes évasées ("Breite der Mundfläche größer als die Gehäuselange" : largeur de la surface aperturale supérieure à celle de la longueur du test) du Crétacé inférieur, que jusque là la plupart des auteurs ont appelé du vocable spécifique "trochus", espèce décrite du Campanien à l'origine (d', 1840). Les figures qui accompagnent la description de (1962) montrent des exemplaires déformés, dont l'accroissement ontogénétique du diamètre du test paraît rapide, plus que celui des spécimens figurés à l'appui de la description de M. cf. trochus, sensu et alii (1957). Toutefois (1962) inclut dans sa liste synonymique la "Valvulina D 1" de (1938), déjà évoquée dans et alii (1957). Il va même jusqu'à désigner comme holotype de sa nouvelle espèce l'un des spécimens figurés par . Enfin il place dans la synonymie de M. subtrochus une "Marssonella cf. trochus" non figurée mais seulement mentionnée dans un tableau en 1962 par & , en limitant toutefois l'analogie aux spécimens possédant un test à agglutinat quartzeux assez grossier ! De tout ce cheminement compliqué il ressort qu'il n'est pas clairement établi que M. cf. trochus, sensu et alii (1957) et M. subtrochus , 1962, soient conspécifiques, et quoi qu'il en soit le concept de subtrochus demeure quelque peu flou.
Les choses vont se compliquer encore un peu plus en raison de la création de Dorothia hechti (Valanginien supérieur-Hauterivien de Sardaigne) par & (1966). À l'exception de l'holotype, les populations de la description originelle offrent un aspect très similaire à celui des spécimens figurés comme M. cf. trochus par et alii (1957), sauf en qui concerne la surface aperturale, concave au lieu d'être plane. L'holotype de D. hechti évoque plutôt D. praeoxycona (1966), qui montre toutefois des loges un peu plus renflées. & (1966) signalent l'analogie de leur nouvelle espèce avec D. trochus, et incluent aussi dans leur liste synonymique des références (partielles) à des formes de (1938), mais pas les mêmes que celles qui figurent (, 1962) dans la synonymie de M. subtrochus.
Nos spécimens de l'Aptien de La Tuilière, tout comme ceux de l'Aptien du stratotype de Cassis, que nous avions déterminés comme "Praedorothia cf. trochus", sensu et alii (1957), montrent une forme générale qui correspond en gros à celle des figurations de Marssonella cf. trochus ou de Dorothia hechti (holotype excepté), mais avec des loges un peu plus renflées. La plupart d'entre eux peuvent aussi s'inscrire sans trop de difficultés dans la variabilité des populations du Barrémo-Aptien de Roumanie abondamment figurées par (1972, 1975), respectivement sous les noms de D. hechti et de D. subtrochus.
Toutes ces péripéties montrent que nous avons affaire à un plexus à forte variabilité, et qu'il est sans doute illusoire de vouloir y discriminer l'un ou l'autre de ces taxons sur des bases purement typologiques.
Praedorothia sp. aff. trochus (d', 1840)
Pl. 2 ![]() , figs. 5-6
, figs. 5-6
Textularia trochus d', 1840, p. 45-46, Pl. IV, figs. 25-26.
Trochamminella saxonica , 1995 (pro parteĀ: seulement certaines formes macrosphériques ¢ au sens de l'auteur ¢), p. 79-80, Pl. 1, figs. 3-4 & 6, non figs. 1-2 & 5.
Protomarssonella minuta , 1995, p. 80-81, Pl. 1, figs. 8-10.
Praedorothia trochus (d')Ā; et alii, 1998, p. 203.
Praedorothia trochus et alii, 2005, in tableaux-fig. 3.
La brève définition fournie par et alii (1998, p. 203) résume la principale caractéristique des spécimens infracrétacés rapportés à l'espèce trochus par maints auteursĀ: "forme très conique (...) résultant de l'accroissement en largeur très rapide des loges". La figuration de d' (1840) (qui concerne une espèce d'âge campanien) paraît quelque peu théorique, mais dans sa description l'auteur mentionne une "coquille (...) très courte, trochoïde, plus large que haute (...) arrondie sur les côtés, qui sont évidés" et "des loges (...) non saillantes (...) les deux supérieures concaves". Par rapport à la description et la figuration de d', les formes du Crétacé inférieur (y compris notre matériel aptien des stratotypes) montrent des loges plus distinctement saillantes, avec une génératrice du cône un peu moins concave mais plutôt rectiligne, au moins dans la partie juvénile du test.
Comme pour les deux groupes de Praedorothia précédents, on observe au sein des populations rapportées à P. sp. aff. trochus une multitude de stades intermédiaires. Ils correspondent à des formes plus ou moins évasées, en fonction du degré d'accroissement du diamètre, plus ou moins rapide, du test, ou/et d'un nombre variable de tours (pouvant aller de 3 à 6) pour le stade bisérié.
(1995) a décrit (très sommairement) sous le nom de Protomarssonella
minuta trois spécimens à faible nombre de tours, qui sont très semblables à celui que nous figurons ici
(Pl. 2 ![]() ,
figs. 5-6). Des stades plus avancés (nombre de tours plus élevé) figurent par ailleurs dans le matériel (incluant l'holotype) qui accompagne sa description de
Trochamminella saxonica ; L'auteur les interprète comme étant la forme macrosphérique de sa nouvelle espèce. D'une part, à l'opposé des formes supposées microsphériques
(Pl. 1, figs. 1-2 & 5 in , 1995), il n'est pas certain que ses formes macrosphériques aient un départ spiralé et relèvent donc du genre Trochaminella. Elles évoquent plutôt les stades avancés de nos Praedorothia sp. aff. trochus. De plus, au sein de nos populations aptiennes, de tels stades apparaissent reliés aux formes à stade bisérié court (cf. P.
minuta) par nombre d'intermédiaires.
,
figs. 5-6). Des stades plus avancés (nombre de tours plus élevé) figurent par ailleurs dans le matériel (incluant l'holotype) qui accompagne sa description de
Trochamminella saxonica ; L'auteur les interprète comme étant la forme macrosphérique de sa nouvelle espèce. D'une part, à l'opposé des formes supposées microsphériques
(Pl. 1, figs. 1-2 & 5 in , 1995), il n'est pas certain que ses formes macrosphériques aient un départ spiralé et relèvent donc du genre Trochaminella. Elles évoquent plutôt les stades avancés de nos Praedorothia sp. aff. trochus. De plus, au sein de nos populations aptiennes, de tels stades apparaissent reliés aux formes à stade bisérié court (cf. P.
minuta) par nombre d'intermédiaires.
Praedorothia sp. aff. pupa (, 1860)
Pl. 2 ![]() , figs. 12-14
, figs. 12-14
Textilaria pupa , 1860, p. 232, Pl. 13, figs. 4a-b & 5a-b.
Les spécimens que nous rapportons, avec doute compte tenu du hiatus stratigraphique, à l'espèce décrite du Sénonien d'Allemagne du Nord par (1860), possèdent une morphologie tout à fait comparable aux exemplaires figurés par l'auteur. Le test de celui que nous figurons ne présente pas de déformation, à l'encontre de la majorité de ceux qui composent notre matériel, qui sont le plus souvent plus ou moins comprimés, voire aplatis. Cependant le spécimen figuré ici est affecté d'un artefact résultant probablement d'une abrasion différentielle du test, qui pourrait laisser croire à la présence d'une ouverture circulaire, avec bref col, située à cheval sur la suture séparant les deux dernières loges. Aucun autre spécimen n'a montré une telle "structure". À La Tuilière, cette forme n'apparaît qu'au Gargasien, dans la partie supérieure de la coupe des Gays, au sein de la Zone à ferreolensis.
Tritaxis (?) cf. paleofusca , 1975
Pl. 3 ![]() , figs. 16-20
, figs. 16-20
Valvulina fusca ()Ā; et alii, 1966, p. 145, Pl. 1, figs. 72-75.
"Valvulina fusca" & 1951 (not )Ā; , 1972, p. 201, Pl. 4, figs. 7-9.
Tritaxis paleofusca , 1975, p. 31, Pl. XII, figs. 16-22 ; Pl. XIII, figs. 27-29.
? Trochamminella saxonica , 1995 (pro parteĀ: seulement certaines formes microsphériques ¢ au sens de l'auteur ¢), p. 79-80, Pl. 1, figs. 1-2, non figs. 3-6.
Notre matériel aptien des stratotypes de Cassis-La Bédoule et de la Tuilière renferme assez constamment de rares spécimens à test spiralé fortement comprimé, constitué de loges en forme de croissant. Leur médiocre état de conservation ne permet pas de se faire une idée très précise des caractéristiques de leur ouverture. Ce critère est pourtant déterminant pour l'attribution générique (Tritaxis vs Trochamminella) de telles formes, très communément dénommées "Valvulina fusca" jusqu'aux mises au point de (1972, 1975). Sur le plan spécifique nous rapprochons nos spécimens de l'espèce du Néocomien-Barrémien de cet auteur, sans pouvoir envisager une assimilation totale, dans la mesure où P. paleofusca possède 2-3 loges au dernier tour, alors que nos exemplaires aptiens en montrent 4.
Oolina apiculata (, 1851)
Pl. 4 ![]() , fig. 1
, fig. 1
Oolina apiculata , 1851, p. 22, Pl. 1, fig. 1.
Cette espèce, comme beaucoup d'autres Nodosariidae uniloculaires, a longtemps et communément été attribuée au genre Lagena.
Au sein de ce groupe, à côté de Lagena s.s., & (1988) ont retenuĀ:
Reussoolina, créé par (1956) comme sous-genre de Lagena, avec apiculata (décrite originellement comme Oolina par , 1851) comme espèce-type,
Oolina d', 1839 (espèce-type O. laevigata).
Sur la base des diagnoses compilées par & (1988), il nous apparaît que les différences pouvant exister entre Reussoolina et Oolina sont fort subtiles, voire inexistantes, et en tout cas pas d'ordre générique. Elles sont un peu plus marquées, bien que pas vraiment fondamentales, entre Lagena (surface striée ou costulée, ouverture avec col net) et Oolina (surface lisse ou au plus très finement striée, ouverture avec col court et pouvant être radiée). Si l'on s'en tient strictement à ces diagnoses, l'espèce apiculata doit être considérée comme relevant du genre Oolina.
Dans notre matériel, O. apiculata n'est présente (toujours rare) que dans l'extrême sommet de la Zone à luterbacheri et dans la Zone à ferreolensis (coupes des Gays 1 et 2).
Ramulina sp. gr. aculeata , 1886
Pl. 4 ![]() , fig. 2
, fig. 2
Ramulina aculeata , 1886, p. 331, Pl. 26, fig. 11.
Ramulina aculeata Ā; et alii, 1966, p. 159-161, Pl. 4, figs. 315-339.
Nous suivrons la position de et alii (1966), selon laquelle il est inutile de pulvériser en de nombreuses "espèces" (= R. aptiensis, R. fusiformis, R. novaculeata, etc.), sans signification biologique ni stratigraphique, ces Polymorphinidae aux loges plus ou moins fusiformes, à la surface hispide. Les stolons qui les relient se brisent facilement lors de la fossilisation, au point que dans notre matériel nous n'observons que des loges dissociées.
R. sp. gr. aculeata est assez constamment présente dans le Bédoulien terminal et le Gargasien inférieur de La Tuilière, avec une fréquence très modérée.
Genre Vaginulina d', 1826
Sous-genre Psilocitharella & , 1986
Pendant 160 ans, jusqu'à la création du genre Psilocitharella par & (1986), l'attribution au genre Vaginulina du groupe de Vaginulinidae jurassico-crétacés au test comprimé, en forme de harpe plus ou moins évasée et à section subquadrangulaire (bords tronqués), a semblé aller de soi et n'a guère été un sujet de discussion. La création de Psilocitharella résulte du fait que l'espèce-type de Vaginulina (Nautilus legumen , 1758) possède une morphologie (départ rectilinéaire, section ovalaire à lenticulaire, allure plus proche d'une Dentalina) qui la singularise par rapport aux autres espèces du plexus mésozoïque évoqué ci-dessus.
En fait jusque là les difficultés soulevées au sujet de ce groupe concernaient plutôt la définition des espèces. Presque dès le départ, la pauvreté des descriptions de (1841) et la multiplication excessive de nouveaux taxons par (1860, 1863) ont compliqué le problème. Des efforts méritoires dans le sens d'une clarification ont été ultérieurement tentés (, 1880 ; , 1952 ; , 1975). Cependant, en raison de la grande variabilité intra-populationnelle de notre assez riche matériel aptien stratotypique, ces mises au point successives ne nous ont pas permis de dégager une hiérarchie cohérente des critères spécifiques, et de répartir ces populations entre des taxons stables et bien définis. Beaucoup de noms ("trop", disait déjà en 1880) sont à disposition dans la littérature, et il est aisé de se rendre compte qu'ils sont souvent utilisés avec des acceptions très différentes selon les auteurs. Enfin aussi bien notre matériel que celui de certains de nos prédécesseurs (ex. , 1952) laissent percevoir des problèmes liés à l'existence probable d'un dimorphisme. Nous avons donc pris le parti :
pour des raisons d'usage, de conserver l'acception "élargie", longtemps reconnue, pour le genre Vaginulina,
de ne considérer Psilocitharella que comme un sous-genre de Vaginulina s.l.,
au plan spécifique, de ne décrire et figurer que quelques morphotypes, choisis parmi les plus significatifs et/ou les plus fréquemment représentés au sein de notre matériel,
de laisser pour le moment certains de ces morphotypes en nomenclature ouverte, dans l'attente d'une révision plus exhaustive.
Vaginulina (Psilocitharella) cf. recta , 1863
Pl. 4 ![]() , figs. 8-9
, figs. 8-9
Vaginulina recta , 1863, p. 48, Pl. 3, figs. 14-15a-b.
Le bord dorsal nettement rectiligne de ce morphotype incite à le rapprocher de Vaginulina recta . Il est modérément abondant et sporadiquement représenté dans les quatre coupes du secteur de La Tuilière.
Vaginulina (Psilocitharella) cf. bicostulata , 1860
Pl. 4 ![]() , figs. 12-13
, figs. 12-13
Vaginulina bicostulata , 1860, p. 202-203, Pl. 8, figs. 5a-b.
Morphotype à la distribution assez irrégulière dans notre matériel, un peu plus fréquent dans le Bédoulien terminal que dans le Gargasien inférieur. La forme générale, et surtout la forte épaisseur du test, nettement moins comprimé que chez tous les autres morphotypes ici décrits, évoquent quelque peu V. bicostulata . Cependant le médiocre état de conservation de notre matériel ne nous a permis de discerner à la surface du proloculus les deux petites rides ("zwei kurzen, schmalen, etwas gebogenen Rippchen") mentionnées par (1860). De plus chez nos formes le nombre de loges est supérieur (7-8 au lieu de 4) et les crêtes suturales se fragmentent en allant vers le bord dorsal, tandis qu'elles apparaissent continues sur les figurations de (1860).
Vaginulina (Psilocitharella) cf. procera , 1952
Pl. 4 ![]() , figs. 10-11
, figs. 10-11
Vaginulina procera , 1952, p. 80-82, Figs. 4-9 ; Pl. 4, figs. 1-4.
Ce morphotype élancé, souvent de grande taille, aux loges nombreuses (au moins une quinzaine) évoque l'espèce Vaginulina procera décrite par (1952). Cependant, chez cette dernière le départ est le plus souvent résolument rectilinéaire, alors que nos spécimens montrent une légère, mais nette, courbure initiale. Ils se rapprochent également beaucoup de certains exemplaires (ex. Pl. 14, fig. 30) du large plexus dénommé V. kochii truncata par (1975), ou de l'exemplaire (Pl. 80, figs. 17a-b) attribué à Vaginulina kochii par (1943). Dans les deux cas il ne s'agit pas de l'espèce V. kochii de (1841), et l'attribution spécifique de nos formes, au demeurant assez rares et très irrégulièrement présentes dans notre matériel, demeure quelque peu incertaine.
Vaginulina (Psilocitharella) sp. 1
Pl. 4 ![]() , fig. 16
, fig. 16
Cette forme constitue le "taxon" le plus fréquent et le plus constamment représenté dans notre matériel des quatre coupes du secteur de La Tuilière. Sa forme générale en triangle plus ou moins évasé, "harpiforme", évoque celle d'espèces fréquemment citées dans la littérature, comme V. kochii, V. truncata, V. arguta. Toutefois ces "espèces" (d'ailleurs plus moins mises en synonymie entre elles, de manières différentes, par divers auteurs) sont dépourvues des fines et discrètes costulations qui chez nos spécimens apparaissent transversalement sur les loges. Nos formes pourraient donc plutôt relever de l'espèce V. striolata , chez qui ces costulations sont cependant plus marquées.
Vaginulina (Psilocitharella) sp. 2
Pl. 4 ![]() , figs. 18-19
, figs. 18-19
Formes de petite taille caractérisées par un proloculus bien développé suivi d'une à deux (voire plus rarement trois) loges. Leur répartition est curieuseĀ: elles sont assez constantes dans le Bédoulien terminal, rarissimes dans le Gargasien basal (Zone à luterbacheri), à nouveau assez présentes dans la Zone à ferreolensis. Il ne nous a pas été possible de discerner si elles constituent des formes macrosphériques ou des stades juvéniles [de quelle (s) espèce (s)?], ou une espèce à part entière. Quelques auteurs ont figuré des spécimens analogues, en les interprétant par exemple comme la forme macrosphérique de Vaginulina kochii (, 1943, Pl. 80, fig. 18), ou comme "un échantillon jeune" de Vaginulina truncata (, 1880, Pl. 1, fig. 28 a-b).
Frondicularia hastata , 1842
Pl. 4 ![]() , fig. 17
, fig. 17
Frondicularia hastata , 1842, p. 272, Pl. 7B, figs. 5a-c.
Souvent représentée dans notre matériel par des spécimens de grande taille (plusieurs mm de longueur). Espèce couramment citée au Crétacé inférieur, particulièrement du Néocomien à l'Aptien inclus. Assez constamment présente dans l'Aptien stratotypique, que ce soit dans le secteur de Cassis-La Bédoule ou dans celui de La Tuilière.
Citharina cf. acuminata (, 1863)
Pl. 4 ![]() , figs. 14-15
, figs. 14-15
Vaginulina acuminata , 1863, p. 49, Pl. IV, fig. 1.
Citharina acuminata () ; & , 1973, p. 230 (pro parte : Pl. 4, figs. 62-63, non fig. 64).
? Vaginulina raristriata, sensu , 1943 (non Nodosaria (Dentalina) raristriata , 1893), p. 501, Pl. 80, figs. 23-24.
Nous rapportons, avec réserve, à l'espèce décrite du Barrémien du NW de l'Allemagne par (1863) cette élégante Citharina à la fine et assez dense costulation sub-longitudinale. Nos exemplaires ont une forme générale plus arquée que celle de l'unique spécimen figuré par . En ce sens ils ressemblent beaucoup à ceux des figs. 62-63 (in Pl. 4) des illustrations fournies par & (1973), tandis que celui de leur fig. 64 correspond beaucoup mieux au type de . Nos Citharina ressemblent aussi un peu à celles de l'Aptien-Albien de l'Oklahoma et du Texas que (1943) a assimilées, très probablement à tort, à l'espèce raristriata du Gault d'Angleterre. Décrite par (1893) comme Nodosaria (Dentalina) "élégante et filiforme", celle-ci paraît présenter une section plutôt cylindrique, alors qu'elle est triangulaire pour les formes texanes (selon ) et, après un bref départ cylindriforme, plutôt en ovale aplati chez nos exemplaires.
Citharina cf. acuminata se montre assez rare et sporadique dans le Bédoulien terminal de la coupe de Pichouraz et rarissime dans l'extrême base du Gargasien des coupes de La Tuilière W et des Gays I. Elle est n'est plus représentée ensuite, soit dans la majeure partie de la Zone à luterbacheri et dans la Zone à ferreolensis.
Lingulina sp. gr. loryi (, 1880)
Pl. 4 ![]() , fig. 4
, fig. 4
Frondicularia loryi , 1880, p. 60-61, Pl. IV, figs. 5a-b.
Lingulina furcillata , 1880, p. 65, Pl. IV, figs. 6a-c.
Lingulina praelonga , 1946, p. 576, Pl. 88, figs. 12a-b.
La plus grande part des morphotypes observés dans notre matériel comportent 6 loges, comme chez L. loryi. Quelques rares variants, au demeurant morphologiquement identiques, en présentent un plus grand nombre, pouvant aller jusqu'à 9, ce qui fait entrer ces spécimens dans la mouvance de L. praelonga (dont son auteur reconnaît qu'elle est liée à L. furcillata ). L'holotype de L. furcillata, refiguré à partir des collections de (1880) par (1954), est considéré par ce dernier comme relevant de l'espèce loryi, tandis que (1975) conserve furcillata comme une espèce distincte. L'ensemble de ces morphotypes nous paraît devoir être regroupé au sein d'une acception élargie, non typologique, de L. loryi.
Dans les stratotypes aptiens de Cassis-La Bédoule et du secteur de la Tuilière, Lingulina sp. gr. loryi, toujours assez rare, n'est présente qu'à partir du Gargasien.
Lenticulina cf. nodosa (, 1863)
Pl. 4 ![]() , figs. 20-22
, figs. 20-22
Robulina nodosa , 1863, p. 78, Pl. 9, fig. 6.
Lenticulina nodosa ()Ā; , 1966, p. 51-52, Pl. 4, figs. 9-12.
Lenticulina cf. nodosa ()Ā; et alii, 1998, tabl. fig. 1, p. 189, Pl. 7, fig. 2.
Cette espèce à grande longévité (elle est présente dès le Valanginien, avec un "premier" acmé au Valanginien inférieur-moyen) est citée jusque dans l'Aptien inférieur dans maints gisements téthysiens. Dans les stratotypes aptiens de Cassis-La Bédoule et de La Tuilière, elle montre curieusement un deuxième acmé correspondant au Bédoulien supérieur, et disparaît brusquement à la limite Bédoulien-Gargasien. Il s'agit peut-être d'un phénomène d'homéomorphie entre deux taxons successifs, mais aucun critère objectif de différenciation ne nous permet de distinguer les formes néocomiennes des formes aptiennes.
Saracenaria sp. aff. frankei sensu (1973)
Pl. 5 ![]() , figs. 1-2
, figs. 1-2
non Saracenaria frankei , 1946, p. 573, Pl. 88, fig. 1.
Cristellaria bononiensis (non ), 1894, p. 652-653, Pl. X, figs. 9a-b.
Lenticulina |Saracenaria| aff. frankei (, 1946) ; in & , 1973 (pro parte), p. 32, Fig. 20, Pl. 4, figs. 17-18.
Test de petite taille, élancé, s'évasant modérément dans la partie subrectilinéaire (légèrement arquée) ; section tricarénée, chaque angle étant prolongé par une sorte de mince frange diaphane courant sur toute la longueur du testĀ; surface lisse, d'aspect souvent translucide. Nos exemplaires s'inscrivent dans la variabilité des populations figurées par (in & , 1973), mais sont dans l'ensemble un peu plus élancés, de section plus étroite. Des morphologies analogues, mais non identiques dans les détails, ont été figurées (au moyen de dessins un peu théoriques) par (1943) sous les noms de S. bononiensis striatolipa, S. callitoecha et S. duckcreekensis. S. callitoecha constitue l'espèce dont la morphologie générale ressemble le plus à celle de nos spécimens, mais elle est pourvue d'une fine costulation longitudinale qui manque à nos exemplaires. Le test de S. bononiensis striatolipa est lisse, mais sa forme générale s'éloigne un peu plus de celle de nos Saracenarias. Nous pensons que le matériel du secteur de La Tuilière relève d'une espèce nouvelle, distincte de S. frankei.
Cette forme semble également présenter un certain intérêt sur le plan stratigraphiqueĀ: absente au Bédoulien terminal et au Gargasien basal, rare dans la partie supérieure de la Zone à luterbacheri, elle devient plus fréquente dans la Zone à ferreolensis. Ceci est confirmé par un réexamen de notre matériel du Gargasien de Cassis, coupe de La Marcouline ( et alii, 2005) ; nous y avons en effet découvert qu'elle figure, rare, au sein de populations que nous avions alors regroupées sous le nom de S. frankei, et ce dans le même intervalle stratigraphique qu'à La Tuilière.
Astacolus crepidularis subsp. tricarinella (, 1863)
Pl. 4 ![]() , figs. 23-24
, figs. 23-24
Cristellaria tricarinella , 1863, p. 68-69, Pl. 7, fig. 9.
Lenticulina (Astacolus) tricarinella ()Ā; et alii, 1957, p. 30, Pl. 3, fig. 56a-bĀ; Pl. 4, figs. 84-85.
et alii (1957) soulignent les difficultés d'interprétation résultant du fait que (1863) a redéfini à sa manière l'espèce crepidularis, très imparfaitement décrite à l'origine par (1842), tout en compliquant encore un peu plus les choses par la création simultanée d'une nouvelle espèce (tricarinella), aux caractéristiques très voisines. La distinction entre les deux taxons, tous deux fortement comprimés et à l'enroulement plus lâche que chez Lenticulina, semble résider principalement dans l'aspect du rebord ventral et la nature des crêtes suturales. A. crepidularis (d'après , 1863) possède un rebord caréné et des crêtes suturales fines, modérément proéminentes. Le rebord serait plus tronqué chez tricarinella, avec des crêtes plus développées, plus saillantes. Selon et alii (1957), s'ajoute chez tricarinella la présence d'une crête centrale similaire aux deux latérales et axée sur la carène. Cette caractéristique n'est pas vraiment mentionnée par (1863) dans sa description ni n'apparaît sur son illustration, mais on peut dire qu'elle est implicitement signifiée en raison du nom qu'il attribue à sa nouvelle espèce.
et alii (1957) insistent également sur la répartition stratigraphique différente des deux taxons : Dogger à Aptien pour A. tricarinella, essentiellement Valanginien supérieur-Hauterivien (devenant rare au Barrémien) pour A. crepidularis.
Les caractéristiques de nos spécimens des stratotypes aptiens, que jusqu'à présent nous rapportions à l'espèce A. crepidularis, relèvent en fait d'A. tricarinella, que nous sommes plutôt enclins à considérer comme une sous-espèce, très proche d'A. crepidularis s.s.. À Cassis, de même que dans le secteur de la Tuilière, A. crepidularis tricarinella est cantonnée au Bédoulien ; son extinction (moins franche que celle de L. nodosa) coïncide plus ou moins avec la limite Bédoulien-Gargasien (voir ci-dessus chapitre "résultats biostratigraphiques").
Praebulimina sp. aff. elongata (, 1995)
Pl. 5 ![]() , figs. 3-5
, figs. 3-5
Gubkinella aff. elongata n. sp. , 1995, p. 83-84, Pl. 11, fig. 4.
Des populations relativement abondantes de petits Buliminidae trisériés, d'une taille égale ou inférieure à 150-200 Ąm, forment l'un des constituants majeurs parmi les Foraminifères benthiques de la fraction fine de l'ensemble des échantillons prélevés dans les quatre coupes de l'Aptien de La Tuilière. Parmi les rares auteurs à avoir observé et décrit des Buliminidae plus ou moins similaires en des niveaux anté-albiens, on peut citer et alii (1964) (Barrémien-Aptien de la région de Krasnodar) et (1995) (Aptien du NW de l'Allemagne). Ce sont les illustrations fournies par ce dernier, pour une forme qu'il rapporte à une "Gubkinella" nouvelle, qui nous ont paru les plus proches de nos spécimens. La description de Gubkinella elongata par se limite à une très courte diagnose ("espèce du genre, caractérisée par une forme étroite et allongée"). Elle est complétée par de brèves remarques, où l'auteur dit en substance que d'autres commentaires seraient superflus car cette nouvelle espèce se distingue de manière évidente de toutes les autres Gubkinellas rencontrées dans son matériel et figurées sur la même planche. En fait, étant trisériée jusqu'au stade final, G. elongata ne relève pas du genre Gubkinella, chez qui le test, plurisérié au départ, comporte 4 loges au dernier tour, mais plutôt du genre Praebulimina. À noter que dans le même article (1995) décrit une Praebulimina n. sp. a, avec laquelle notre matériel présente quelques affinités, mais nos spécimens en diffèrent par une forme moins arrondie, moins renflée, de la partie sommitale des dernières loges.
Coryphostoma? sp.
Pl. 5 ![]() , figs. 9-10
, figs. 9-10
Les populations de Buliminidae trisériés [Praebulimina sp. aff.
elongata ()] observées dans la fraction fine de notre matériel aptien de La Tuilière sont assez constamment accompagnées par des formes bolivinoïdes bisériées, élancées et plutôt comprimées, à tendance ontogénétique de plus en plus alternante, avec loges terminales quasiment superposées (cf.
Pl. 5 ![]() , fig. 10). À noter cependant que ce groupe est plus rarement représenté que P. sp. aff.
elongata pendant l'épisode de forte chute de la diversité microfaunique qui survient dans la partie supérieure de la
Zone à luterbacheri, tandis que la situation s'inverse dans la Zone à ferreolensis.
, fig. 10). À noter cependant que ce groupe est plus rarement représenté que P. sp. aff.
elongata pendant l'épisode de forte chute de la diversité microfaunique qui survient dans la partie supérieure de la
Zone à luterbacheri, tandis que la situation s'inverse dans la Zone à ferreolensis.
Des formes morphologiquement voisines ou analogues ont été figurées et décrites sous des dénominations génériques et spécifiques différentes selon les auteurs :
Neobulimina boliviniformis & , 1964, in et alii (1964), de l'Hauterivien-Barrémien de la région de Krasnodar ;
Bolivina kuznetsovae , 1964, in et alii (1964), de l'Aptien de la région de Krasnodar ;
Coryphostoma sp., ("tentatively", faute d'en élucider la structure interne) par (1974) (Albien du Leg DSDP 27, Océan Indien) ;
Cassidella viscida (, 1950), sensu , 1975, de l'Albien supérieur de l'Aube. Cette forme a été décrite initialement comme une Virgulina par (1950). Son attribution générique est incertaine, la structure interne n'étant pas décelableĀ; de plus (1975) mentionne une torsion plus ou moins importante de la partie initiale du test, caractéristique que nous n'avons pas observée au sein de notre matériel ;
Coryphostoma sp., par (1992) (Aptien-Albien du Leg ODP 123, Océan Indien, sans voir la structure interne) ;
Ahlumia n. gen. brunsvicensis n. sp. (Aptien inférieur du NW de l'Allemagne) par & (1995), qui la différencient spécifiquement du taxon viscida de (1950) au motif qu'elle possède plus de loges. D'autre part les auteurs appuient la création de leur nouveau genre par une description ainsi que des figurations précises et d'excellente qualité, montrant bien la présence d'une structuration interne complexe (sorte d'endosiphon tubulaire connectant entre elles les loges successives et paraissant dédoublé au niveau de son raccordement avec l'ouverture apicale, elle-même simple, sans dent). & (1995) se posent la question de l'appartenance possible du taxon viscida de (1950) ainsi que de Coryphostoma sp. sensu (1974) au nouveau genre Ahlumia, mais ne peuvent conclure faute de connaître la nature de leur endostructure ;
Coryphostoma sp. 1, & , 1997 (Aptien du Site 260 et -Albien des Sites 256, 257, 763 et 766, forages DSDP-ODP dans l'Océan Indien). Pas de données sur la structure interne ;
Fursenkoina viscida () sensu et alii (2001) (Albien inférieur du Leg ODP 171B, Site 1049, au large de la Floride). Le matériel ainsi dénommé, bien illustré mais non décrit, possède une morphologie très semblable à celle du taxon brunsvicensis créé par & (1995). D'autre part les données visibles sur la structure interne (amorce de départ de deux tubes sous-jacents à l'ouverture), bien qu'incomplètes, pourraient laisser envisager une probabilité d'endostructure analogue à celle du genre Ahlumia, sans toutefois permettre d'exclure totalement la simple possibilité de "toothplates" et non d'un endosiphon.
Compte tenu de ce qui précède, la question se pose de savoir 1) si sur le plan morphologique l'espèce de , au statut générique incertain, n'a pas tout simplement été redécrite sous un nouveau vocable spécifique (brunsvicensis) par & (1995) : la différenciation fondée simplement sur le nombre de loges nous paraît en effet insuffisante ; elle oppose tout au plus des stades ontogénétiques différents au sein d'une même population, ce qu'illustre bien notre propre matériel 2) si les diverses formes mentionnées ci-dessus, semblables par leur morphologie externe, relèvent soit du genre Coryphostoma (endostructure limitée à une "toothplate"), soit du genre Ahlumia, ou bien encore d'un autre genre.
Aussi bien d'une manière générale que pour ce qui est de notre matériel, nous ne pouvons donc conclure de manière décisive, faute de données comparatives sur la structure interne.
En effet nos spécimens paraissent identiques morphologiquement aux formes figurées par & (1995) et par et alii (2001). Malheureusement leur état de conservation, plutôt médiocre, ne nous a pas permis de nous faire une idée de leur endostructure, voire même de l'aspect externe de leur ouverture. Nous en sommes donc réduits à ne pouvoir fournir à leur sujet d'attribution générique ni même spécifique précise.
Epistomina spp.
Pl. 5 ![]() , figs. 6-8
, figs. 6-8
Les Épistomines représentent un constituant assez constant, d'abondance modérée à moyenne, de la fraction grossière (> 250 Ąm) des échantillons du Bédoulien et du Gargasien basal du secteur de La Tuilière ; leur fréquence s'atténue ensuite quelque peu au cours du Gargasien inférieur
(Zone à ferreolensis). La presque totalité des spécimens recueillis est à l'état de moules internes pyritisés ou limonitisés, ce qui rend impossible toute détermination spécifique. De rares formes montrent des traces résiduelles, mal conservées, du test initialement aragonitique. La mauvaise conservation permet quand même de discerner un certain polymorphisme au sein de ces
populationsĀ: test plus ou moins bi-évolute mais toujours dissymétrique, profil plus ou moins biconvexe mais asymétrique, rarement subsymétrique à faces planes parallèles (cas de l'exemplaire de grande taille illustré ici
Pl. 5 ![]() , figs. 6-8), sutures intercamérales et spirales plus ou moins en relief, paraissant mieux présentes dans les tours internes ; il peut cependant s'agir là d'un artefact de conservation différentielle, la partie ombilicale étant mieux protégée de l'abrasion par le débord des tours plus externes. Compte tenu du contexte diagénétique, nous n'avons pas été en mesure de transcrire ces différences en terme de nomenclature.
, figs. 6-8), sutures intercamérales et spirales plus ou moins en relief, paraissant mieux présentes dans les tours internes ; il peut cependant s'agir là d'un artefact de conservation différentielle, la partie ombilicale étant mieux protégée de l'abrasion par le débord des tours plus externes. Compte tenu du contexte diagénétique, nous n'avons pas été en mesure de transcrire ces différences en terme de nomenclature.
"Valvulineria" sp.
Pl. 5 ![]() , figs. 11-13
, figs. 11-13
Gyroidinoides loetterlei () sensu , 1995, Pl. 5, figs. 6-7Ā; Pl. 9, figs. 10-11Ā; Pl. 10, figs. 9-11.
[non Gyroidina loetterlei , 1940]
Gyroidinoides sp., et alii, 1998, p. 204-205, Pl. 6, figs. 1-3.
Des populations de Valvulineria/Gyroidinoides de petite taille sont constamment et assez abondamment présentes dans les fractions les plus fines de notre matériel de l'Aptien du secteur de la Tuilière. Nous les avions déjà observées dans le Bédoulien et le Gargasien du secteur de Cassis-la Bédoule (
et alii, 1998, 2005). Le test, peu comprimé, subsymétrique à légèrement dissymétrique, à rebord arrondi, possède en général 5-6 loges assez globuleuses au dernier
tourĀ; la face dorsale, évolute, est plutôt plane, la face ventrale, involute, est légèrement bombée. Dans notre matériel, d'une conservation plutôt médiocre, les structures aperturales ("umbilical flaps") sont plus ou moins discernables. La fig. 13 de la
Pl. 5 ![]() laisse entrevoir la présence possible d'une sorte de prolongement adombilical surmontant l'ouverture.
laisse entrevoir la présence possible d'une sorte de prolongement adombilical surmontant l'ouverture.
Par la forme générale et le nombre des loges, nos spécimens sont tout à fait semblables à ceux (tout particulièrement les formes d'âge aptien inférieur) que (1995) a figurés sous la dénomination Gyroidinoides loetterlei. Cette espèce a été décrite par (1940) de l'Albien supérieur du Texas ; selon le texte de la description originelle, elle possède 6 à 7 loges au dernier tour, mais l'holotype (seul figuré) en montre 8 sur la face ventrale et 8,5 sur la face dorsale. Nous pensons que ni le matériel illustré par (1995) ni nos formes éo-aptiennes ne sont conspécifiques avec la forme de l'Albien du Texas, pas plus d'ailleurs qu'avec les nombreux taxons fréquemment cités dans la littérature (crassa, nitida, globosa, lenticula, parva, infracretacea, etcģ), et attribués à divers genres (Gyroidina, Gyroidinoides, Valvulineria).
La relativement médiocre conservation de notre matériel ne nous permet pas d'entrer très avant dans la question de son attribution générique, d'autant plus que par suite d'une utilisation différente des critères distinctifs possibles, d'ailleurs peu tranchés, la différenciation entre les genres Valvulineria et Gyroidinoides varie selon les auteurs (, 1967 ; , 1975 ; & , 1988 ; , 1995). Une révision critique dépasserait très largement le cadre restreint de la présente étude. Pour le moment nous nous bornerons à attribuer, avec quelque réserve, ces petites formes aptiennes au genre le plus anciennement décrit, à savoir Valvulineria.
Gavelinella sp. gr. flandrini , 1960
Pl. 5 ![]() , figs. 18-20
, figs. 18-20
Gavelinella flandrini , 1960, p. 137, Pl. 2, figs. 10-14.
Gavelinella flandrini ; et alii, 1998, p. 204, Pl. 5, figs. 7-8Ā; Pl. 7, fig. 3.
Les Gavelinelles représentent le constituant le plus abondant de la microfaune de Foraminifères benthiques du secteur de la Tuilière, en particulier au Bédoulien supérieur et au Gargasien basal. Affectées ici, tout comme à Cassis-La Bédoule ( et alii, 1998, 2005) d'un polymorphisme important, elles gravitent autour du morphotype flandrini, mieux caractérisé dans le domaine vocontien, où l'espèce fut initialement décrite.
Gavelinella sp. aff. flandrini , 1960
Pl. 5 ![]() , figs. 14-17
, figs. 14-17
Gavelinella sp. cf. gr. barremiana-flandrini et alii, 1998, p. 204.
Ce morphotype possède en vue latérale l'aspect général de G. flandrini mais en diffère surtout par le profil, plus épais et plus nettement plan-convexe. De telles formes sont beaucoup plus rares à La Tuilière (où elles disparaissent dès le Gargasien basal) qu'à Cassis-La Bédoule, où elles se prolongent d'ailleurs plus haut dans le Gargasien. Un tel morphotype n'est pas observé dans le domaine vocontien.
Praehedbergella sigali (, 1966)
Pl. 6 ![]() , figs. 1-5
, figs. 1-5
Hedbergella (Hedbergella) sigali , 1966, p. 87, Pl. 7, figs. 20-25.
Praehedbergella sigali ()Ā; et alii, 2002, p. 128.
Cette espèce typiquement à 4 loges, d'accroissement rapide à mesure qu'elles s'ajoutent, est plus ou moins constamment représentée par quelques spécimens (un peu plus fréquents dans le Bédoulien supérieur que dans le Gargasien inférieur) dans la plupart des échantillons du secteur de La Tuilière. On observe aussi çà et là de rares exemplaires à 4+/4,5 loges, morphologiquement mais, sans doute pas phylogénétiquement, transitionnels vers P. infracretacea ().
Lignée Praehedbergella aptiana-globulifera-roblesae
Comme nous l'avons déjà mentionné antérieurement ( et alii, 2005), l'extrême polymorphisme manifesté durant la transition Bédoulien/Gargasien par les Praehedbergelles possédant 5-6 loges au dernier tour rend malaisée et illusoire la désignation en leur sein d'espèces uniquement fondées sur des bases typologiques. Ce groupe doit en fait être considéré comme un plexus unique, à forte variabilité morphologique. Les abondantes figurations fournies par (1974) en rendaient déjà largement compte. Nous en donnons d'autres exemples dans les Pls. 6 et 7 du présent travail.
Praehedbergella aptiana (, 1965)
Pl. 6 ![]() , figs. 9-12Ā;
Pl. 7
, figs. 9-12Ā;
Pl. 7 ![]() , fig. 1
, fig. 1
Praehedbergella aptiana , 1965, p. 347-348, Figs. 3-6.
Praehedbergella aptiana ()Ā; et alii, 2005, p. 9.
Trochospire basse, typiquement 5 loges au dernier tour, profil modérément comprimé, contour moyennement lobé. Des formes transitionnelles à 5+/5,5 loges, au contour de plus en plus lobé, mènent progressivement à P. globulifera ( & ).
Dans le secteur de La Tuilière, cette espèce constitue un composant assez constant et assez fréquent de la microfaune planctonique du Bédoulien supérieur et du Gargasien inférieur. Elle est rare au Barrémien supérieur puis modérément présente dans le Bédoulien et le Gargasien inférieur et moyen de Cassis-La Bédoule ( et alii, 2005).
Praehedbergella globulifera ( & , 1971)
Pl. 7 ![]() , figs. 2-7, 9-11 & 15-17
, figs. 2-7, 9-11 & 15-17
Clavihedbergella globulifera & (in ), 1971, p. 136, Pl. 10, figs. 1a-c.
Praehedbergella globulifera ( & )Ā; et alii, 2005, p. 9-10.
Relativement rare et sporadique dans le Bédoulien supérieur de la coupe de Pichouraz, P. globulifera est plus fréquente et plus constante au Gargasien inférieur. Cette espèce, ainsi que P.
similis () et P. maslakovae () que, tout comme
et alii (1998), nous considérons comme ses synonymes plus récents, possèdent typiquement 6 loges au dernier tour, à l'accroissement plus ou moins progressif et régulier ; le contour du test est assez découpé en vue latérale par suite d'une lobation marquée des loges. En fait, à partir de morphotypes aptiana, par accentuation progressive de la lobation au sein du plexus globulifera tous les stades intermédiaires existent, menant insensiblement à des exemplaires (ex.
Pl. 7 ![]() , figs. 15-17) qui font la transition avec le stade extrême représenté par P. roblesae ().
, figs. 15-17) qui font la transition avec le stade extrême représenté par P. roblesae ().
Praehedbergella roblesae (, 1959)
Globigerina roblesae , 1959, p. 149, Pl. 4, fig. 4.
Praehedbergella roblesae ()Ā; et alii, 2002, p. 128.
Ce stade ultime de la lignée pour ce qui est de l'étirement des loges, qui montrent un aspect cylindriforme, est beaucoup plus rare et sporadique dans notre matériel aptien du secteur de La Tuilière qu'il ne l'était à Cassis-La Bédoule.
Lignée Praehedbergella infracretacea-praetrocoidea
Ce groupe de formes à 5 et 6 loges au dernier tour se distingue nettement de la lignée précédemment mentionnée, par un profil nettement moins comprimé, des loges plus triangulaires et non trapézoïdales en vue ventrale, disposition qui conduit en outre à une plus grande réduction de l'ombilic. Selon les spécimens, la trochospire peut se décoller plus ou moins dorsalement au niveau des premier tours.
Praehedbergella infracretacea (, 1937)
Pl. 6 ![]() , figs. 13-21
, figs. 13-21
Globigerina infracretacea , 1937, p. 28, Fig. 1.
5 ou 6 loges au dernier tour, les formes à 6 loges devenant progressivement plus nombreuses à mesure que l'on s'élève dans la série. Surface du test plus grossière, avec "perforation cones" mieux individualisés, que dans la lignée de P. aptiana. Un peu moins fréquente que P. aptiana dans le Bédoulien supérieur et le début de la Zone à luterbacheri, P. infracretacea fait ensuite jeu égal avec elle, puis la dépasse en fréquence dans la Zone à ferreolensis.
Praehedbergella praetrocoidea ( & , 1986)
Pl. 7 ![]() , figs. 12-14
, figs. 12-14
Hedbergella praetrocoidea & (in ), 1986, p. 95, Pl. 16, figs. 35.
Cette espèce, rare, évolue un peu avant la limite Bédoulien-Gargasien à partir de formes à 6 loges de P. infracretacea, dont elle se distingue au départ par une taille plus importante et une rugosité du test un peu plus accentuée. Elle est sporadique (trop pour que l'on puisse l'utiliser comme marqueur) et ne dépasse guère les dix premiers mètres de la Zone à luterbacheri dans le secteur de La Tuilière. De plus, dans la coupe de la carrière de La Marcouline à Cassis ( et alii, 2005), nous l'avons observée, tout aussi sporadique, dans les zones à luterbacheri, ferreolensis et barri, et elle atteint même la Zone à algerianus. et alii (1998) ont suggéré que cette espèce était l'ancêtre d'Hedbergella trocoidea, qui apparaît au Gargasien supérieur.
Praehedbergella primare ( & , 1986)
Pl. 6 ![]() , figs. 6-8 ;
Pl. 7
, figs. 6-8 ;
Pl. 7 ![]() , fig. 8
, fig. 8
Clavihedbergella primare & (in ), 1986, p. 120-121, Pl. 25, figs. 2a-3a & 2b-3b.
Praehedbergella kuznetsovae ( & ) sensu et alii, 1998, p. 207-208, Pl. 3, figs. 8-9.
Praehedbergella primare ( & )Ā; et alii, 2005, p. 10-11.
La principale caractéristique de cette Praehedbergelle à trochospire basse, à 6 loges au dernier tour, est la quasi absence d'accroissement en hauteur de ces dernières à mesure qu'elles s'ajoutent, ainsi qu'un contour du test (en vue latérale) faiblement lobé. Cette espèce est présente, avec une fréquence modérée, dans pratiquement tous les échantillons du Bédoulien supérieur-Gargasien inférieur du secteur de La Tuilière.
Praehedbergella luterbacheri (, 1974)
Pl. 8 ![]() , figs. 1-6
, figs. 1-6
Hedbergella luterbacheri , 1974, p. 61, Pl. 19, figs. 21-26 ; Pl. 26, figs. 15-17.
Praehedbergella luterbacheri ()Ā; et alii, 2005, p. 10, Pl. 3, figs. 5-10 ; Pl. 4, figs. 1-6.
Pour la définition de cette espèce, nous renvoyons le lecteur aux descriptions, figurations et synonymies fournies par (1974) et et alii (2005). L'un des résultats significatifs de notre étude sur les Foraminifères de l'Aptien du secteur de La Tuilière est d'avoir pu localiser avec une précision encore plus grande qu'à Cassis-La Bédoule le moment d'apparition de ce marqueur (cf. la section biostratigraphie du présent article), à partir de formes transitionnelles avec P. primare ( & ). Toujours rare dans la zone éponyme, aussi bien à Cassis que dans les coupes des Gays 1 et 2, l'espèce devient un peu plus fréquente dans la partie sommitale de la zone ainsi que dans la partie inférieure de la Zone à ferreolensis, puis amorce une décroissance. À Cassis elle devient plus rare dans la Zone à barri et disparaît juste à la limite avec la Zone à algerianus ( et alii, 2005).
Lignée Globigerinelloides duboisi-blowi-aptiensis/solidus
Ce plexus réunit les "petites" formes planispiralées à 4, 5 ou 6 loges au dernier tour. Là encore, on observe au sein des populations de nombreux stades transitionnels entre des morphotypes particuliers qui ont été artificiellement figés, à partir de données strictement typologiques, en "espèces" dans le cadre de la nomenclature linnéenne.
Globigerinelloides sp. gr. duboisi-blowi
Pl. 9 ![]() , figs. 1-12
, figs. 1-12
Planomalina blowi , 1959, p. 260, Pl. 20, figs. 2-3.
Globigerinella duboisi , 1961, p. 33, Pl. 1, figs. 14-18.
Nous renvoyons le lecteur à nos précédentes mises au point ( et alii, 2002, 2005) concernant le polymorphisme intra-populationnel au sein de ce groupe et ses répercussions sur sa nomenclature. Dans le secteur de La Tuilière, le morphotype duboisi domine numériquement au Bédoulien supérieur et dans la Zone à luterbacheri, ensuite (Zone à ferreolensis) la répartition entre les deux morphotypes s'équilibre quantitativement. Cette égalité va se poursuivre au-delà, au moins jusqu'à la base de la Zone à algerianus, comme l'ont montré les données obtenues à Cassis dans la coupe du Gargasien de La Marcouline ( et alii, 2005). La distinction entre ces deux espèces ne présente donc pas grand intérêt au plan biostratigraphique. Nous avons également renoncé à individualiser au sein de ce plexus les formes "clavates" (cf. Hastigerina alpina 1959, ou les espèces/sous-espèces décrites par & , 2005), polymorphes, très peu stables et à distribution sporadique. De tels morphotypes nous paraissent s'intégrer dans la variabilité intra-populationnelle du groupe duboisi-blowi.
Globigerinelloides sp. gr. aptiensis - solidus
Pl. 9 ![]() , figs. 13-22 & 26-29
, figs. 13-22 & 26-29
Globigerinelloides aptiense , 1974, p. 79-80, Pl. 4, figs. 9-10 ; Pl. 8, figs. 4-6 & 17-18.
Globigerinelloides solida & (in ), 1986, p. 123, Pl. 27, fig. 3.
Les deux morphotypes "aptiense" et "solida" (nom. corr. : aptiensis et solidus, respectivement) ne constituent probablement que des variants typologiques au sein d'une même espèce, essentiellement caractérisée par un test planispiralé avec six loges au dernier tour. Ces morphotypes ne se distinguent que par une légère différence du rythme d'accroissement des loges, un peu plus rapide chez le premier (aspect plus "évolute" du test en vue latérale) que chez le second, où il est quasi nul. Inutile de préciser qu'il existe des stades intermédiaires au sein des populations afférentes. Il semble toutefois que la tendance à la "biglobigerinellisation" (cf. nos spécimens de la
Pl. 9 ![]() ,
figs. 26-29), qui se manifeste dans ce groupe au niveau de la Zone à ferreolensis, soient plus fréquente chez les morphotypes solidus. Dans le secteur de La Tuilière, les deux morphotypes coexistent : toujours rares au Bédoulien supérieur et au Gargasien basal, ils deviennent un peu plus fréquents à partir de la partie supérieure de la
Zone à luterbacheri et surtout dans la Zone à ferreolensis. Ces données confirment nos observations précédentes effectuées dans le secteur de Cassis-La Bédoule, où ce groupe persiste, assez bien représenté, dans la
Zone à barri et atteint, en devenant plus rare, la Zone à algerianus.
,
figs. 26-29), qui se manifeste dans ce groupe au niveau de la Zone à ferreolensis, soient plus fréquente chez les morphotypes solidus. Dans le secteur de La Tuilière, les deux morphotypes coexistent : toujours rares au Bédoulien supérieur et au Gargasien basal, ils deviennent un peu plus fréquents à partir de la partie supérieure de la
Zone à luterbacheri et surtout dans la Zone à ferreolensis. Ces données confirment nos observations précédentes effectuées dans le secteur de Cassis-La Bédoule, où ce groupe persiste, assez bien représenté, dans la
Zone à barri et atteint, en devenant plus rare, la Zone à algerianus.
Lignée Globigerinelloides ferreolensis-barri-algerianus
Cette lignée, dérivée de formes trochospiralées (Praehedbergella luterbacheri) évoluant progressivement vers la planispiralité (cf. et alii, 2005), inclut les "grands" Globigerinelloides, à 7 loges et plus au dernier tour. Nous retenons trois espèces (la première avec deux-sous espèces, dont l'une nouvelle) au sein de ce groupe. La distinction entre ces taxons repose, selon nous, uniquement sur le nombre de loges au dernier tour, à l'exclusion de très faibles disparités morphologiques, qui relèvent plutôt de la variabilité intra-populationnelle et ont conduit certains auteurs à des créations "d'espèces" superflues. Ainsi définis sur la base de leur nombre de loges au dernier tour, les trois taxons précités présentent en outre un réel intérêt biostratigraphique, en permettant d'établir une zonation homophylétique fine et fiable, qui s'échelonne au sein du Gargasien.
Globigerinelloides ferreolensis heptacameratus n. subsp.
Pl. 8 ![]() , figs. 7-8
, figs. 7-8
HolotypeĀ: exemplaire de la Pl. 8, fig. 8, déposé dans les collections de micropaléontologie du Muséum National d'Histoire Naturelle à Paris (n° F62184).
ParatypeĀ: exemplaire de la Pl. 8, fig. 7 (coupe des Gays 1, Gargasien inférieur, échantillon 2340), déposé dans les collections de micropaléontologie du Muséum National d'Histoire Naturelle à Paris (n° F62185).
Localité-type : Hameau de La Tuilière (commune de Saint-Saturnin-lès-Apt), près de la ferme des Gays.
Stratum typicumĀ: Coupe des Gays 2 (Gargasien moyen basal ?, échantillon 2301).
Origine du nomĀ: sept loges au dernier tour.
DiagnoseĀ: cette nouvelle sous-espèce regroupe les formes à 7 loges au dernier tour au sein de l'espèce Globigerinelloides ferreolensis s.l.
RemarquesĀ: G. ferreolensis a été originellement décrite du domaine vocontien (,
1961) comme incluant des formes à 7 et 8 loges au dernier tour. L'holotype et le paratype ont chacun 8 loges, ce que nous avons pu vérifier directement (collect. ). Dans sa diagnose originelle, l'auteur mentionne déjà l'existence d'un décalage d'apparition en fonction du nombre de
logesĀ: "Les loges, au nombre de 7 (formes du Gargasien inférieur de base) à 8 (chez les exemplaires de la partie supérieure du Gargasien inférieur)"
(op. cit., p. 214). Nous avons retrouvé ce décalage à La Tuilière (coupes des Gays 1 et 2), où nous avons pu le déterminer avec une plus grande précision que dans le domaine vocontien grâce à la forte dilatation de la série et à la densité de l'échantillonnage
(Figs. 6 ![]() - 7
- 7 ![]() ).
).
RépartitionĀ: dans le secteur de La Tuilière, Globigerinelloides ferreolensis heptacameratus n. subsp., à 7 loges seulement au dernier tour, apparaît en effet plus précocement dans le Gargasien inférieur (mais non basal, comme le croyait en 1961) que G. ferreolensis ferreolensis (à 8 loges). La création de cette nouvelle sous-espèce nous paraît d'autant plus se justifier que nos recherches montrent qu'elle constitue un repère biostratigraphique précis ("horizon à heptacameratus") situé à la transition entre les zones à luterbacheri et ferreolensis (cf. chapitre "résultats biostratigraphiques"). À la partie terminale de la Zone à luterbacheri, Globigerinelloides ferreolensis heptacameratus n. subsp. coexiste, sur environ 6 m, avec les derniers (rares) représentants de Schackoina gr. cabri. À l'extrême base de la Zone d'acmé de Globigerinelloides ferreolensis, seules ces formes à 7 loges sont présentes, les formes à 8 loges (G. ferreolensis ferreolensis) apparaissant seulement 6 m au dessus de la limite zonale. Les deux sous-espèces coexistent ensuite jusqu'au sommet de la série visible dans le secteur de la Tuilière, qui pourrait atteindre l'extrême base de la Zone à barri (Gargasien moyen). À Cassis (coupe de La Marcouline), dans la partie basale de la Zone à algerianus, soit la base du Gargasien supérieur avec laquelle se termine la coupe, G. ferreolensis heptacameratus constitue encore 20% des populations relevant de l'espèce ferreolensis s.l.. Des vérifications dans le domaine vocontien seront nécessaires pour voir si les deux sous-espèces disparaissent simultanément, ou avec un décalage, dans le Gargasien supérieur/Clansayésien.
Globigerinelloides ferreolensis ferreolensis (, 1961)
Pl. 8 ![]() , figs. 9-11
, figs. 9-11
Biticinella ferreolensis , 1961, p. 214, Pl. 1, figs. 1-5.
Globigerinelloides macrocameratus , 1974, p. 85-86, pro parte : paratype Pl. 5, figs. 1 & 5-6.
Globigerinelloides ferreolensis ()Ā; et alii, 2002, p. 131-133.
Cette sous-espèce de G. ferreolensis inclut les formes à 8 loges au dernier tour, quelle que soit leur forme ou leur rythme d'accroissement. Pour plus de détails, nous renvoyons le lecteur à nos travaux antérieurs (, 1961, 1966 ; et alii, 1998, 2002, 2005).
Globigerinelloides barri (, & , 1957)
Pl. 8 ![]() , figs. 12-14
, figs. 12-14
Biglobigerinella barri , & , 1957, p. 25, pro parte : Pl. 1, figs. 18a-b (holotype) & 17, non figs. 13-16.
Globigerinelloides macrocameratus , 1974, p. 85-86, pro parte : holotype, Pl. 5, figs. 2-4.
Globigerinelloides barri (, & )Ā; et alii, 2002, p. 133.
Un seul exemplaire de cette espèce a été recueilli dans notre matériel, dans l'échantillon n° 2301, situé au sommet de la coupe des Gays 2. L'état de conservation (test jaunâtre) de ce spécimen diffère de celui de tous les autres foraminifères planctoniques du même niveau. La question de sa présence à ce niveau a été discutée ci-dessus dans le chapitre consacré aux "résultats biostratigraphiques".
Genre Schackoina , 1932
Sous-genre Leupoldina , 1957
Leupoldina a été séparé de Schackoina par (1957) sur la base de légères différences dans la morphologie de la partie distale des loges. Nous considérons cette différenciation comme tout au plus d'ordre sub-générique. Ces deux taxons sont cependant considérés comme deux genres distincts par & (2005), non plus en raison de critères morphologiques mais sur la base d'une légère différence de microstructure du test. Selon ces auteurs, la paroi du test de Schackoina est microperforée, tandis que celle de Leupoldina est finement perforée.
Schackoina (Leupoldina) sp. gr. cabri
Pl. 10 ![]() , figs. 1-9
, figs. 1-9
Schackoina cabri , 1952, p. 20-21, figs. 1-8.
Schackoina pustulans , 1957, p. 274-275, Pl. 1, figs. 1-4 & 6-7.
Leupoldina protuberans , 1957, p. 277, Pl. 2, figs. 1-13.
Bien représenté dans le Bédoulien supérieur et encore assez fréquent à l'extrême base du Gargasien, ce groupe se raréfie ensuite progressivement, devenant sporadique dans le reste de la Zone à luterbacheri. Son extinction est un élément d'appoint dans la caractérisation de la limite entre les zones à luterbacheri et à ferreolensis.
Pseudoschackoina saundersi (, 1959)
Pl. 9 ![]() , figs. 23-25
, figs. 23-25
Planomalina saundersi , 1959, p. 262, Pl. 20, figs. 9-11.
"Globigerinelloides" gr. bizonae (, 1961) ¢ saundersi (, 1959) ; et alii, 2005, p. 11-12, Pl. 1, figs. 1-12 ; Pl. 2, figs. 1-12 ; Pl. 3, figs. 1-4.
Pseudoschackoina saundersi () ; & , 2005, p. 257-258, Figs. 5.13-15 & 6.1-14.
Nos observations sur le matériel de l'Aptien de Cassis ( et alii, 2005) nous ont incités à mettre en synonymie l'espèce de (1961) avec celle de (1959). L'évolution progressive dans le temps de certains caractères, en particulier d'un mode d'enroulement trochospiralé au Bédoulien supérieur vers un enroulement subplanispiralé au Gargasien, nous avait conduits à nous interroger sur l'attribution générique de telles formes. Entre temps & (2005) ont résolu la question en créant pour ce taxon le genre Pseudoschackoina, position que nous suivons dans ce travail.
À La Tuilière, P. saundersi est toujours rare et sporadique au Bédoulien sommital et dans la Zone à luterbacheri, devenant rarissime dans la Zone à ferreolensis. Elle est un peu mieux représentée à Cassis, apparaissant assez tôt dans le Bédoulien supérieur, plus irrégulièrement présente au Gargasien inférieur et à nouveau assez constante dans la partie supérieure de la Zone à barri et la base de la Zone à algerianus. Il est possible que ces variations de fréquence de représentation soient liées à des fluctuations des conditions environnementales, comme cela a été suggéré d'une manière générale pour les formes planctoniques à loges étirées radialement ( et alii, 2006 : "radially elongated chambered planktonic foraminifera").
En raison d'une sédimentation abondante, continue et non perturbée, le secteur de la Tuilière a enregistré en grand détail, pour ce qui est de la période de transition du Bédoulien au Gargasien, une succession de bio-événements qu'il n'avait pas été possible jusqu'à présent de détecter aussi finement en n'importe quelle autre région du globe. Son étude nous a permis de constituer un inventaire détaillé du contenu microfaunique, avec comme principaux résultats :
une confirmation de la valeur stratigraphique de certains marqueurs, benthiques et planctoniques,
un confortement du bien-fondé de la zonation par foraminifères planctoniques précédemment définie en Basse-Provence, avec en outre l'adjonction d'un biohorizon (à Globigerinelloides ferreolensis heptacameratus nov. subsp.) marquant le passage de la Zone à luterbacheri à la Zone à ferreolensis ;
la mise en évidence de la quasi-coïncidence du triple datum micropaléontologique (LAD de Protocythere bedoulensis ¢ Lenticulina cf. nodosa / FAD de Praehedbergella luterbacheri) avec le changement lithostratigraphique dit du "changement de couleur" ainsi qu'avec le toit de la Zone d'ammonites à Furcata, marquant la limite entre les sous-étages Bédoulien et Gargasien ;
la mise en évidence d'une forte chute de la diversité et de l'abondance des foraminifères épibenthiques et planctoniques durant une part importante de la Zone à luterbacheri. Cette crise oligospécifique paraît être liée à un épisode d'intense sédimentation de cendres volcaniques, qui pourrait correspondre au Niveau Van Gogh du domaine vocontien.
La richesse du matériel recueilli nous a également permis d'apporter des précisions taxinomiques sur un certain nombre d'espèces de foraminifères benthiques, pour une grande part peu ou pas connues en dehors de l'Aptien de Basse-Provence mais, curieusement, souvent représentées dans certains secteurs du domaine boréal (ex.Ā: Allemagne du Nord).
Les auteurs tiennent à exprimer leur gratitude à Isabella (Université de Milan), Silvia (Université de Parme) et Danièle (Université de Strasbourg) pour leur relecture critique et leurs suggestions qui nous ont permis d'améliorer notre manuscrit.
J. (1952).- Taxonomie und Entwicklung einiger Arten von Vaginulina d' aus dem Barrême bei Hannover (Foram.).¢ Mitteilungen aus dem Geologisches Staatsinstitut in Hamburg, Hft. 21, p. 75-112.
Z.A., T.A. A.G. & O.M. (1964).- Foraminifères néocomiens et aptiens dans l'inter-fleuve Pchecha-Ubin (Caucase NW). In : V.L (éd.), Problèmes de stratigraphie et de lithologie concernant les dépôts mésozoïques et cénozoïques de la région de Krasnodar.- Publications V.N.N.I., n° 12, Éditions "Nedra", Moscou, p. 3-72. (en russe)
F. & Y. (2002).- Nouvelles données biostratigraphiques sur l'Aptien moyen et supérieur du Sud-Est de la France, à la lumière de la succession des ammonites du domaine vocontien.- In : STRATI 2002 - 3ème Congrès Français de Stratigraphie, Lyon, 8-10 juillet 2002. Volumes de résumés.- Documents des Laboratoires de Géologie de Lyon, n° 156, p. 23-24.
F. & Y. (2005).- Biostratigraphie et faunes d'ammonites de l'Aptien supérieur (Gargasien) de la région stratotypique de Gargas, près d'Apt (Sud-Est de la France). In : Impact environnemental sur la dynamique des populations au Crétacé.- Réunions thématiques du Groupe Français du Crétacé (Paris, 5-6 Décembre 2005), Volume des Résumés, p. 12-15 (abstract).
N.V. (1962).- Désagrégation des roches argilo-marneuses sous l'action simultanée de l'hyposulfite et de l'eau oxygénée.- Voprosy Mikropaleontologii, Moscou, n° 6, p. 147-148 (en russeĀ; traduction B.R.G.M. n° 4123).
J.-F., M. & G. (2007).- Les Ostracodes du Bédoulien supérieur et du Gargasien inférieur du stratotype de l'Aptien : Systématique et corrélations biostratigraphiques.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2007/05-fr (CG2007_A05 (fr)), 36 p.
H. (1952).- Taxionomische Revision und Nomenklator zu Franz E. "Standard-Gliederung der Nordwest-deutschen Unterkreide nach Foraminiferen", Teil 2: Barrême.- Senckenbergiana, Frankfurt am Main, Bd. 33, n° 4/6, p. 297-312.
H. (1954).- Revision von 's Mémoire 1880 über die Alb-Foraminiferen von Montcley.- Senckenbergiana Lethaea, Frankfurt am Main, Bd. 35, n° 1/2, p. 37-50.
H. (1956).- Zur Mikrofauna des englischen Hauterive.- Senckenbergiana Lethaea, Frankfurt am Main, Bd. 37, n° 5/6, p. 509-533.
H. (1962).- Neue Foraminiferen aus Unterkreide und Oberkeuper NW-Deutschlands und der Schweiz.¢ Senckenbergiana Lethaea, Frankfurt am Main, Bd. 43, n° 2, p. 135-149.
H. (1965).- Taxionomische Revision und Nomenklator zu Franz E. "Standard-Gliederung der Nordwest¢deutschen Unterkreide nach Foraminiferen" (1938). Teil 4: Alb.- Senckenbergiana Lethaea, Frankfurt am Main, Bd. 46, n° 4/6, p. 327-366.
H. & F. (1962).- Marine Unterkreide (Boreal und Tethys). In : Leitfossilien der Mikropaläontologie.- Verlag Gebrüder Borntraeger, Berlin, p. 225-297.
H. & H. M. (1977).- The Foraminifera in the Lower Cretaceous of Trinidad, W. I. ¢ Part 4: Cuche Formation, upper PartĀ; Leupoldina protuberans Zone.¢ Eclogae Geologicae Helvetiae, Bâle, vol. 70, n° 2, p. 543-573.
H. & E. (1951).- Mikropaläontologische Untersuchungen zur Stratigraphie des nordwestdeutschen Valendis.¢ Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Stuttgart, n° 485, p. 239-336.
H. & M. (1973).- Die Unterkreide von Helgoland und ihre mikropaläontologische Gliederung.¢ Senckenbergiana Lethaea, Frankfurt am Main, Bd. 54, n° 2/4, p. 207-264.
H., F. & H. M. (1957).- Die Foraminiferen der Unterkreide von Trinidad, B.W.I. ¢ Erster Teil: Cuche- und Toco-Formation.¢ Eclogae Geologicae Helvetiae, Bâle, vol. 50, n° 1, p. 5-67.
H., F. & H. M. (1966).- Die Foraminiferen der Unterkreide von Trinidad, W.I. ¢ Zweiter Teil: Maridale-Formation (Typlokalität).¢ Eclogae Geologicae Helvetiae, Bâle, vol. 59, n° 1, p. 129-177.
G. (1880).- Mémoire sur les Foraminifères fossiles de l'étage Albien de Montcley (Doubs).- Mémoires de la Société Géologique de France, Paris, sér. 3, vol. 1, n° 5, p. 1-84.
F. (1952).- Stratigraphisch wichtige Foraminiferen-Arten aus dem Barrême vorwiegend Nordwest-Deutschlands.- Senckenbergiana, Frankfurt am Main, Bd. 33, n° 4/6, p. 263-295.
H.M. (1957).- The foraminiferal genera Schackoina , emended and Leupoldina, n. gen. in the Cretaceous of Trinidad B.W.I.- Eclogae Geologicae Helvetiae, Bâle, vol. 50, p. 271¢278.
H.M. (1959).- Planktonic Foraminifera from the Cretaceous of Trinidad, B.W.I.- Bulletin of American Paleontology, Ithaca, N.Y., vol. 39, n° 179, p. 257-277.
H.M., A.R. & H. (1957).- Planktonic foraminiferal families Hantkeninidae, Orbulinidae, Globorotaliidae, and Globotruncanidae. In : A.R. et alii (éds.), Studies in foraminifera.- United States National Museum, Bulletin, Washington, Vol. 215, p. 3¢50.
M.K., F.T., T.N., M.D. & J.E. (1998).- On the evolution of the Hedbergellidae from the Praehedbergellidae.- Journal of Micropaleontology, London, vol. 17, p. 97-103.
F. (1893).- The Foraminifera of the Gault of Folkestone.- Journal of the Royal Microscopical Society, Londres, pt. IV, p. 579-595.
F. (1894).- The Foraminifera of the Gault of Folkestone.- Journal of the Royal Microscopical Society, Londres, pt. VII, p. 645-654.
J. (1961).- Quelques nouvelles espèces dans le Crétacé inférieur méditerranéen.- Revue de Micropaléontologie, Paris, vol. 4, n° 1, p. 30¢36.
G. (1956).- Los foraminiferos del Burdigaliense de Mallorca.- Memorias de la Real Academia de Ciencias y Artes de Barcelona, vol. 32, n° 5, p. 7-140.
R., V. & A. (2006).- Cretaceous oceanic anoxic events and radially elongated chambered planktonic foraminifera: Paleoecological and paleoceanographic implications.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 235, p. 66-92.
J.A. & C.I. (1930).- Some Vaginulinas and other foraminifera from the Lower Cretaceous of Texas.¢ Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon, vol. 6, pt. 1, p. 1-10.
R. & F. (1973).- Ostracodes et Foraminifères de l'Aptien inférieur du sondage du Bois du Perchois (Aube).- Bulletin d'Information des Géologues du Bassin de Paris, n° 36, p. 3-47.
C.H. von & E. (1995).- Ahlumia brunsvicensis n. gen. n. sp., eine neue Foraminifere des nordwestdeutschen Unter-Apt.- Geologisches Jahrbuch, Hannover, Abt. 141, p. 225-233.
L. (2002).- Litho-, bio-, et chronostratigraphie comparées dans le Bassin vocontien à l'Aptien.- Thèse Université de Lille 1, 451 p.
D. & F.T. (1987).- The evolution of Early Cretaceous Dorothiinae (Foraminiferida).¢ Journal of Micropaleontology, Londres, vol. 6, n° 2, p. 13-27.
L. & F. (1966).- I foraminiferi del Valanginiano superiore di Orosei (Sardegna).¢ Palaeontographia Italica, Pisa, Vol. LXI (n. ser. Vol. XXXI)- anno 1966, p. 7-186.
Y. (2005).- Biostratigraphie, évolution et renouvellements des ammonites de l'Aptien supérieur (Gargasien) du bassin vocontien (Sud-Est de la France).- Thèse Université Claude Bernard-Lyon 1, 302 p.
G. (1975).- Contribution à l'étude stratigraphique et sédimentologique de l'Aptien supérieur de Provence.- Thèse Doctorat 3ème cycle, Marseille, 86 p.
M.F. (1937).- Planktonforaminiferen aus der Kreide und dem Eozän und ihre stratigraphische Bedeutung.- Etudy po Mikropaleontologii, Paleontologicheskaya Laboratoriya Moskovskogo Gosudarstvennogo Universiteta, Moscou, 1, p. 27¢46.
T.N. (1971).- On Early Cretaceous foraminifera of the Crimea.- Voprosy Mikropaleontologii, Moscou, n° 14, p. 125¢139 & 211-216. (en russe)
T.N. (1986).- Jurassic and Early Cretaceous planktonic foraminifera of the south of the USSR.- Akademia Nauk SSSR, Nauka, Moscou, 239 p. (en russe)
D.W. (1992).- Aptian-Albian foraminifers from Site 766, Cuvier Abyssal Plain, and comparison with coeval faunas from the Australian region.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station (Texas), vol. 123, p. 271-297.
F.E. (1938).- Standard-Gliederung des nordwest-deuschen Unterkreide nach Foraminiferen.- Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main, n° 443, p. 1-42.
A.E.L. & M.A. (1997).- Lower Cretaceous deep-water benthic foraminifera of the Indian Ocean.- Grzybowski Foundation Special Publication, Cracovie, n° 4, 172 p.
A., W. & J. (2001).- Benthic foraminifers from Lower Albian Black Shales (Site 1049, ODP Leg 171): Evidence for a non "uniformitarian" record.- Journal of Foraminiferal Research, Lawrence (Kansas), vol. 31, n° 1, p. 60-74.
F. (1967).- Les Valvulineria de l'Albien de l'Aube.- Revue de Micropaléontologie, Paris, vol.10, n° 3, p. 153-178.
E. (1995).- Abfolge und Verbreitung der Foraminiferen im Barrêrme und Apt Nordwest-Deutschlands. In : E. (éd.), Die Wende Barrême/Apt ¢ Untersuchungen an Profilen des Borealgebietes.- Geologisches Jahrbuch, Reihe A, Hannover, Heft 141, p. 63-111.
M.H. (1950).- On some new Foraminifera from the Lower Gault of Southern England.- Journal of the Royal Microscopical Society, Londres, sér. 3, vol. 70, p. 268-279.
C. von (1758).- Systema naturae. Tomus I, Regnum animale.- Holmiae: Laurentii Salvii, Stockholm, 823 p.
A.R. & H. (1986).- Some new and redefined genera and families of hyaline calcareous Foraminiferida (Protozoa).- Transactions of the American Microscopical Society, Lawrence, vol. 105, p. 239-265.
A.R. & H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold Company, New York, vol. 1, 970 p.; vol. 2, 847 pls.
J.F. (1974).- Stratigraphic, morphologic and taxonomic studies of Aptian planktonic Foraminifera.- Revista Española de Micropaleontologia, Madrid, Numero Extraordinario, p. 1¢107.
F. (1975).- Les Foraminifères de l'Albien de l'AubeĀ: Paléontologie, stratigraphie, écologie.¢ Cahiers de Paléontologie, Paris, 360 p.
M. (1960).- Sur quelques Foraminifères du Crétacé inférieur des Baronnies (Drôme).- Revue de Micropaléontologie, Paris, vol. 3, n° 2, p. 131-142.
M. (1961).- Quelques Foraminifères et Ostracodes nouveaux du Crétacé inférieur vocontien.- Revue de Micropaléontologie, Paris, vol. 3, n° 4, p. 213-216.
M. (1965).- Révision des stratotypes de l'Aptien. II- Gargas (Vaucluse). In : Colloque sur le Crétacé inférieur, Lyon, 1963.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, n║ 34, p. 201-214.
M. (1966).- Étude stratigraphique et micropaléontologique du Crétacé inférieur de la "Fosse vocontienne".- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, n° 15, 369 p.
M., J.-P. & G. (2002).- Hierarchy of criteria, evolutionary processes and taxonomic simplification in the classification of Lower Cretaceous planktonic foraminifera.- Cretaceous Research, London, vol. 23, n° 1, p. 111¢148.
M. & G. (2004).- Le Gargasien (Aptien moyen) dans les stratotypes historiques de l'Aptien (SE France) : Introduction générale.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/01 (CG2004_L01), 3 p.
M., G., C. & G. (2006).- Le Gargasien (Aptien moyen) de La Tuilière - St-Saturnin-lès-Apt (région du stratotype historique de l'Aptien, Vaucluse, SE France) : localisation géographique et corrélations des affleurements.- Carnets de Géologie / Notebooks on Geology, Brest, Note brève 2006/01 (CG2006_L01), 8 p.
M., G. & J.-P. (2005).- The Gargasian (Middle Aptian) strata from Cassis-La Bédoule (Lower Aptian historical stratotype, SE France): planktonic and benthic foraminiferal assemblages and biostratigraphy.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2005/02 (CG2005_A02), 20 p.
M., G., W. & J.-P. (1998).- Les foraminifères benthiques et planctoniques du stratotype historique de l'Aptien inférieur dans la région de Cassis - La Bédoule (SE France).¢ Géologie Méditerranéenne, Marseille, vol. XXV, n° 3-4, p. 187-225.
T. (1965).- Albian Foraminifera of the Romanian Plain.- Micropaleontology, New York, vol. 11, n° 1, p. 1-38.
T. (1972).- The Eo-Cretaceous foraminiferal fauna from the area between the Ialomitza and Prahova valleys (Eastern Carpathians).- Revista Española de Micropaleontologia, Madrid, vol. 4, n° 2, p. 181-224.
T. (1975).- Monographie de la faune des Foraminifères éocrétacés du couloir de la Dîmbovicioara, de Codlea et des Monts Persani (Couches de Carhaga).- Mémoires de l'Institut de Géologie et Géophysique, Bucarest, n° 25, 141 p.
J. (1959).- Foraminíferos de la Formación La Peña.- Boletín de la Asociación Mexicana de Geólogos Petroleros, México, vol. XI, n° 3-4, p. 135¢153.
A. d' (1826).- Tableau méthodique de la classe des Céphalopodes.- Annales des Sciences Naturelles, Paris, vol. 7, p. 245-314.
A. d' (1839).- Voyage dans l'Amérique méridionale ¢ Foraminifères, vol. 5, pt. 5.- Paris & Strasbourg, P. Bertrand, 86 p.
A. d' (1840).- Mémoire sur les Foraminifères de la Craie Blanche du Bassin de Paris.- Mémoire de la Société Géologique de France, t. IV, n°1, 51 p.
A.M. (1851).- Die Foraminiferen und Entomostraceen des Kreidemergels vom Lemberg. ¢ Haidinger's Naturwissenschaftliche Abhandlungen, Wien, vol. 4 (1850), p. 17-52.
A.M. (1860).- Die Foraminiferen der westphälischen Kreideformation..¢ Sitzungsberichte der mathematisch-naturwissenschaftliche Klasse der Kayserliche Akademie der Wissenschaften in Wien, vol. 8 (1859), p. 147-238.
A.M. (1863).- Die Foraminiferen des norddeutschen Hils und Gault.¢ Sitzungsberichte der mathematisch-naturwissenschaftliche Klasse der Kayserliche Akademie der Wissenschaften in Wien, vol. 46, sect. I (1862), p. 5-100.
F.A. (1841).- Die Versteinerungen der norddeutschen Kreidegebirges.- Hannover, 145 p.
F.A. (1842).- Neue Kreide-Foraminiferen.- Neues Jahrbuch für Mineralogie Geogn. Geologie Petref.-Kunde, Stuttgart, p. 272-273.
P., M., R., G. & G. (2006).- The Deshayesitidae , 1949 (Ammonoidea) of the Aptian historical stratotype region at Cassis-La Bédoule (SE France).- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2006/01 (CG2006_M01), 46 p.
V. (1974).- Aptian-Albian benthonic foraminifers from DSDP leg 27, Sites 259, 260 and 263, Eastern Indian Ocean.- Initial Reports of the Deep Sea Drilling Project, Washington, vol. 27, p. 697-741.
J. (1952).- Aperçu stratigraphique sur la micropaléontologie du Crétacé.- XIXe Congrès Géologique International, Monographies régionales, (1ère Série : Algérie), Alger, n° 26, p. 3¢47.
J. (1959).- Notes micropaléontologiques alpines. Les genres Schackoina et Leupoldina dans le Gargasien vocontien. Étude de morphogenèse.- Revue de Micropaléontologie, Paris, vol. 2, p. 68-79.
H. (1940).- Foraminifera from the Grayson Formation of northern Texas.- Journal of Paleontology, Tulsa, vol. 14, n° 2, p. 93-126.
H. (1943).- Foraminifera from the Duck Creek Formation of Oklahoma and Texas.- Journal of Paleontology, Tulsa, vol. 17, n° 5, p. 476-517.
A. (1946).- Arenaceous Foraminifera and Lagenidae from the Neocomian (Lower Cretaceous) of the Netherlands.¢ Journal of Paleontology, Tulsa, vol. 20, n° 6, p. 570-577.
A. (1950).- Les Foraminifères de l'Albien des Pays-Bas.¢ Mémoires de la Société géologique de France, Paris, (N.S.), t. XXIX, n° 63, p.1-67.
J. (1997).- V- La Biostratigraphie.- In : J. (éd.), Stratigraphie. Terminologie française.- Bulletin du Centre de Recherches Exploration Production elf-Aquitaine, Pau, Mémoire n° 19, p. 1-164.
D. & I. (2005).- Early Cretaceous planktonic foraminifera from the Tethys. The Upper Aptian, planispiral morphotypes with elongate chambers.- Cretaceous Research, London, vol. 26, p. 239-259.
J. (1886).- Cretaceous Foraminifera of Keady Hill, Co. Derry.- Belfast Naturalists Field Club, Report Proceedings, 1885-1886, Appendix, p. 327-332.
B. (1961).- Stratigraphische Verbreitung und Phylogenie von Foraminiferen des nordwestdeutschen Oberhauterive.- Paläontologische Zeitschrift, Stuttgart, vol. 35, n° 1/2 p. 28-61.
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Carte générale du secteur étudié.
General map of the area studied.
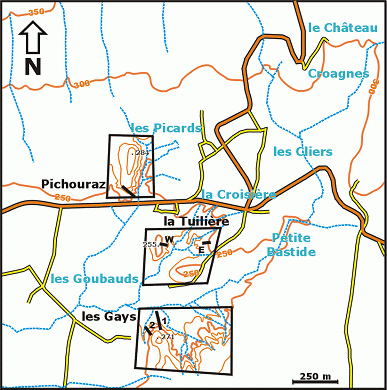
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : Carte de détail du secteur étudié.
Detailed map of the area
studied.
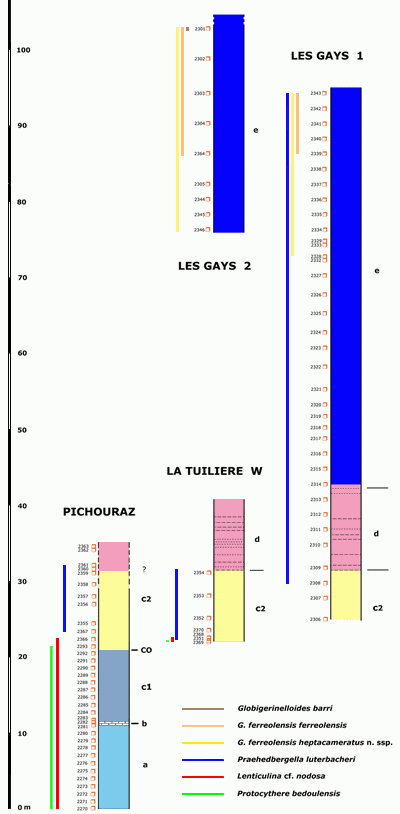
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 3 :
Positionnement des échantillons et présence des principaux marqueurs dans les coupes de Pichouraz, de La Tuilière W et des Gays. Modifié d'après
et alii (2006), où est détaillée la lithologie des niveaux "a",
"b", "c1", "c2", "d" et "e".
Location of the samples collected and distribution of the main markers in the sections of Pichouraz, la Tuilière W and Les Gays. Modified after
et alii (2006), where the lithology of the levels "a",
"b", "c1", "c2", "d" and "e" is detailed.
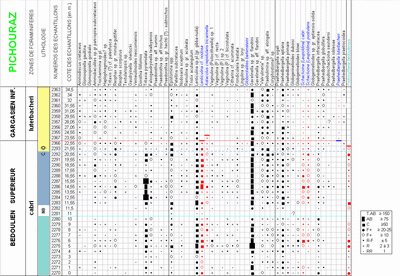
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 :
Répartition des Foraminifères dans la coupe de Pichouraz.
Distribution of Foraminifera in the Pichouraz section.
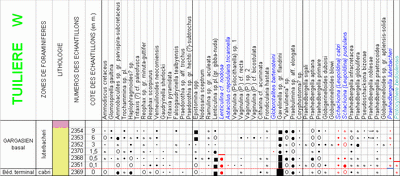
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 5 :
Répartition des Foraminifères dans la coupe de La Tuilière W.
Distribution of Foraminifera in the La Tuilière W section.
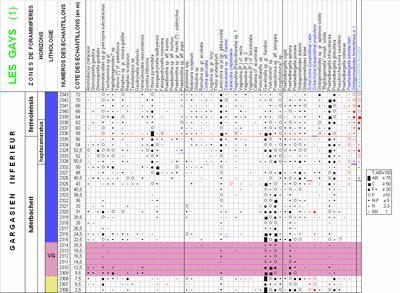
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 6 :
Répartition des Foraminifères dans la coupe des Gays 1.
Distribution of Foraminifera in the Les Gays 1 section.
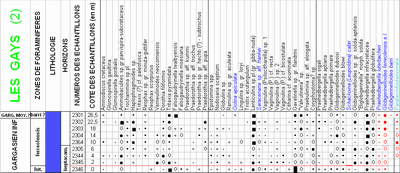
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 7 :
Répartition des Foraminifères dans la coupe des Gays 2.
Distribution of Foraminifera in the Les Gays 2 section.
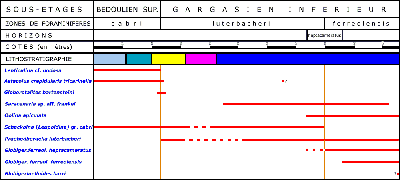
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 8 :
Répartition des principaux marqueurs (Foraminifères) dans le Bédoulien supérieur-Gargasien inférieur du secteur de La Tuilière
(s.l.).
Distribution of the principal markers (Foraminifera) in the upper Bédoulian-lower Gargasian in La Tuilière
(s.l.) area.
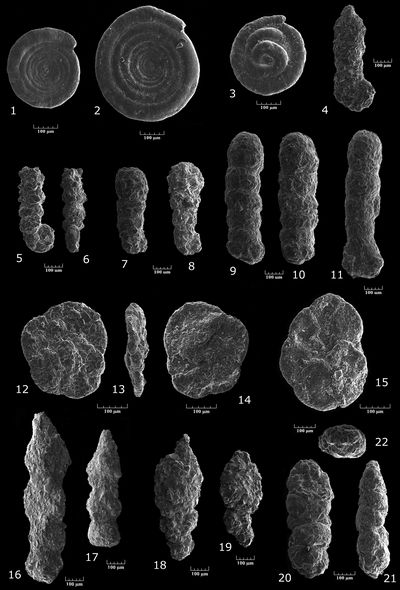
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 1 : 1-2. Ammodiscus cretaceus (). Bédoulien supérieur, coupe de Pichouraz. 1 : échantillon 2288. 2 : éch. 2292. 3. Glomospirella gaultina (). Bédoulien terminal, Pichouraz, éch. 2366. 4-11. Ammobaculites sp. gr. reophacoides/parvispira-subcretaceus. 4-7. Bédoulien supérieur, Pichouraz (4 : éch. 2290 ; 5-6 : éch. 2291) (5 : vue latérale, 6 : profil). 7 : éch. 2366. 8. Gargasien inférieur, Pichouraz, éch. 2358. 9-11. Gargasien inférieur, Les Gays 1, éch. 2332 ; 9 : vue latérale, 10 : profil. 12-15. Trochammina sp. gr. vocontiana ¢ concava. Bédoulien terminal, Pichouraz, éch. 2366. 16-17. Reophax sp. gr. minuta-guttifer. Bédoulien supérieur, Pichouraz, éch. 2284. 18-19. Reophax scorpiurus (). Bédoulien supérieur, Pichouraz ; 18 : éch. 2284 ; 19 : éch. 2290. 20-22. Gaudryinella sherlocki . Bédoulien terminal, Pichouraz, éch. 2366. 20 : vue latérale ; 21 : vue de profil ; 22 : vue apicale.
Plate 1 : 1-2. Ammodiscus cretaceus (). Upper Bedoulian, Pichouraz section. 1: sample 2288. 2: sample 2292. 3. Glomospirella gaultina (). Uppermost Bedoulian, Pichouraz, sample 2366. 4-11. Ammobaculites sp. gr. reophacoides/parvispira-subcretaceus. 4-7. Upper Bedoulian, Pichouraz (4: sample 2290; 5-6: sample 2291) (5: lateral view, 6: profile). 7: sample 2366. 8. Lower Gargasian, Pichouraz, sample 2358. 9-11. Lower Gargasian, Les Gays 1, sample 2332; 9: lateral view, 10: profile. 12-15. Trochammina sp. gr. vocontiana ¢ concava. Uppermost Bedoulian, Pichouraz, sample 2366. 16-17. Reophax sp. gr. minuta-guttifer. Upper Bedoulian, Pichouraz, sample 2284. 18-19. Reophax scorpiurus (). Upper Bedoulian, Pichouraz; 18: sample 2284; 19: sample 2290. 20-22. Gaudryinella sherlocki . Uppermost Bedoulian, Pichouraz, sample 2366. 20: lateral view; 21: profile; 22: apical view.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 2 : 1-4. Praedorothia sp. aff. kummi (). Bédoulien supérieur, coupe de Pichouraz, échantillon 2284. 1, 3 : faces latérales, 2 : profil, 4 : face aperturale. 5-6. Praedorothia sp. aff. trochus (d'). Bédoulien supérieur, coupe de Pichouraz, éch. 2286. 5 : vue latérale, 6 :face aperturale. 7-11. Praedorothia sp. gr. hechti (?) ¢ subtrochus. Bédoulien supérieur, coupe de Pichouraz (cinq spécimens distincts). 7-8 : éch. 2286. 9-11 : éch. 2292. 7-10 : vues latérales, 11 : vue de profil. 12-14. Praedorothia sp. aff. pupa (). Gargasien inférieur, coupe des Gays 2, éch. 2305. 12 : vue latérale, 13 : profil, 14 : vue aperturale. 15-18. Dorothia filiformis (). Gargasien inférieur, coupe des Gays 1. 15-17 : éch. 2332 (15-16 : vues latérales, 17 : profil du même spécimen) ; 18 : éch. 2329, vue latérale. 19, 23-26. Verneuilinoides neocomiensis (). 19. Bédoulien terminal, coupe de La Tuilière W, éch. 2369. 23-26. Bédoulien supérieur, coupe de Pichouraz, éch. 2286. 23 : vue apicale, 24 : vue latérale du même spécimen ; 25 : vue apicale, 26 : vue latérale (idem). 20-22. Tritaxia pyramidata . Gargasien inférieur, coupe des Gays 1. 20 : éch. 2336 ; 21-22 : éch. 2328 (deux spécimens distincts).
Plate 2: 1-4. Praedorothia sp. aff. kummi (). Upper Bedoulian, Pichouraz section, sample 2284. 1, 3: lateral faces, 2: profile, 4: apertural face. 5-6. Praedorothia sp. aff. trochus (d'). Upper Bedoulian, Pichouraz section, sample 2286. 5: lateral view, 6: apertural face. 7-11. Praedorothia sp. gr. hechti (?) ¢ subtrochus. Upper Bedoulian, Pichouraz section (five distinct specimens). 7-8: sample 2286. 9-11: sample 2292. 7-10: lateral views, 11: profile. 12-14. Praedorothia sp. aff. pupa (). Lower Gargasian, Les Gays 2 section, sample 2305. 12: lateral view, 13: profile, 14: apertural view. 15-18. Dorothia filiformis (). Lower Gargasian, Les Gays 1 section. 15-17: sample 2332 (15-16: lateral views, 17: profile of the same specimen); 18: sample 2329, lateral view. 19, 23-26. Verneuilinoides neocomiensis (). 19. Uppermost Bedoulian, La Tuilière W section, sample 2369. 23-26. Upper Bedoulian, Pichouraz section, sample 2286. 23: apical view, 24: lateral view of the same specimen; 25: apical view, 26: lateral view (idem). 20-22. Tritaxia pyramidata . Lower Gargasian, Les Gays 1 section. 20: sample 2336; 21-22: sample 2328 (two distinct specimens).
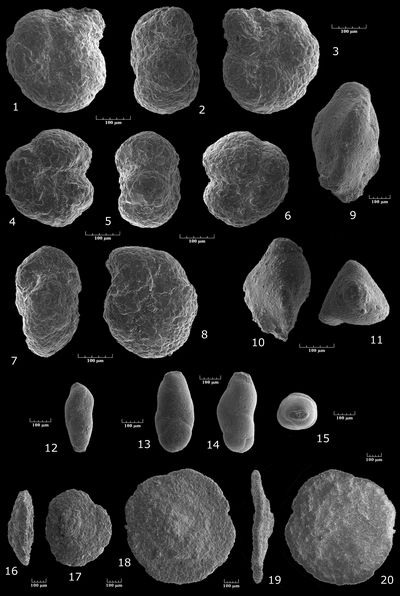
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 3 : 1-8. Haplophragmoides? sp. 1-3 : Bédoulien supérieur, coupe de Pichouraz, échantillon 2285 (1, 3 : faces latérales, 2 : profil). 4-8 : Gargasien inférieur, coupe des Gays 1. 4-6 : éch. 2333 ; 7-8 : éch. 2332. 9-11. Falsogaudryinella moesiana (). Gargasien inférieur, coupe des Gays 1, éch. 2336 (deux spécimens distincts ; 9 : vue latérale ; 10 : vue latérale, 11 : vue aperturale). 12-15. Falsogaudryinella tealbyensis (). 12 : Gargasien inférieur, coupe de La Tuilière W, éch. 2352. 13-15 : Gargasien inférieur, coupe des Gays 1, éch. 2327 (un seul spécimen ; 13-14 : vues latérales, 15 : vue apicale). 16-20. Tritaxis (?) cf. paleofusca . 16-17 : Gargasien inférieur, coupe de La Tuilière W, éch. 2353 ; 16 : profil, 17 : face spirale ; 18-20 : Gargasien inférieur, coupe des Gays 1, éch. 2340 ; 18 : face spirale, 19 : profil, 20 : face ombilicale.
Plate 3: 1-8. Haplophragmoides? sp. 1-3: upper Bedoulian, Pichouraz section, sample 2285 (1, 3: lateral faces, 2: profile). 4-8: lower Gargasian, Les Gays 1 section. 4-6: sample 2333; 7-8: sample 2332. 9-11. Falsogaudryinella moesiana (). Lower Gargasian, Les Gays 1 section, sample 2336 (two distinct specimens; 9: lateral view; 10: lateral view, 11: apertural view). 12-15. Falsogaudryinella tealbyensis (). 12: lower Gargasian, La Tuilière W section, sample 2352. 13-15: lower Gargasian, Les Gays 1 section, sample 2327 (single specimen; 13-14: lateral views, 15: apical view). 16-20. Tritaxis (?) cf. paleofusca . 16-17: lower Gargasian, La Tuilière W section, sample 2353; 16: profile, 17: spiral face; 18-20: lower Gargasian, Les Gays 1 section, sample 2340; 18: spiral face, 19: profile, 20: ombilical face.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 4 : 1. Oolina apiculata (). Gargasien inférieur, coupe des Gays 1, échantillon 2339. 2. Ramulina sp. gr. aculeata . Gargasien inférieur, coupe de La Tuilière W, éch. 2368. 3. Globulina sp.Ā: spécimen avec ouvertures "aulostomelles" (sensu , 1880). Gargasien inférieur, coupe des Gays 1, éch. 2339. 4. Lingulina sp. gr. loryi (). Gargasien inférieur, coupe des Gays 2, éch. 2345. 5. Nodosaria sceptrum . Gargasien inférieur, coupe de Pichouraz, éch. 2359. 6-7. Tristix acutangulum (). Gargasien inférieur, coupe des Gays 1, éch. 2334 ; 6 : vue latérale, 7 : vue apicale. 8-9. Vaginulina (Psilocitharella) cf. recta . Bédoulien terminal, coupe de La Tuilière W, éch. 2369. 8 : vue latérale, 9 : profil. 10-11. Vaginulina (Psilocitharella) cf. procera . Gargasien inférieur, coupe des Gays 1, échantillon 2328. 10Ā: vue latérale, 11Ā: profil. 12-13. Vaginulina (Psilocitharella) cf. bicostulata . Bédoulien supérieur, coupe de Pichouraz, éch. 2292. 14-15. Citharina cf. acuminata (). Bédoulien supérieur, coupe de Pichouraz, éch. 2291. 16. Vaginulina (Psilocitharella) sp. 1. Bédoulien supérieur, coupe de Pichouraz, éch. 2276. 17. Frondicularia hastata . Bédoulien supérieur, coupe de Pichouraz, éch. 2293. 18-19. Vaginulina (Psilocitharella) sp. 2. Bédoulien supérieur, coupe de Pichouraz, éch. 2286. 20-22. Lenticulina cf. nodosa (). Bédoulien supérieur, coupe de Pichouraz. 20-21 : éch. 2288 ; 20 : vue latérale, 21 : profil. 22 : éch. 2293. 23-24. Astacolus crepidularis subsp. tricarinella (). Bédoulien supérieur, coupe de Pichouraz, éch. 2284.
Plate 4: 1. Oolina apiculata (). Lower Gargasian, Les Gays 1 section, sample 2339. 2. Ramulina sp. gr. aculeata . Lower Gargasian, La Tuilière W section, sample 2368. 3. Globulina sp.: specimen with "aulostomelle" (sensu , 1880) apertures. Lower Gargasian, Les Gays 1 section, sample 2339. 4. Lingulina sp. gr. loryi (). Lower Gargasian, Les Gays 2 section, sample 2345. 5. Nodosaria sceptrum . Lower Gargasian, Pichouraz section, sample 2359. 6-7. Tristix acutangulum (). Lower Gargasian, Les Gays 1 section, sample 2334; 6: lateral view, 7: apical view. 8-9. Vaginulina (Psilocitharella) cf. recta . Uppermost Bedoulian, La Tuilière W section, sample 2369. 8: lateral view, 9: profile. 10-11. Vaginulina (Psilocitharella) cf. procera . Lower Gargasian, Les Gays 1 section, sample 2328. 10: lateral view, 11: profile. 12-13. Vaginulina (Psilocitharella) cf. bicostulata . Upper Bedoulian, Pichouraz section, sample 2292. 14-15. Citharina cf. acuminata (). Upper Bedoulian, Pichouraz section, sample 2291. 16. Vaginulina (Psilocitharella) sp. 1. Upper Bedoulian, Pichouraz section, sample 2276. 17. Frondicularia hastata . Upper Bedoulian, Pichouraz section, sample 2293. 18-19. Vaginulina (Psilocitharella) sp. 2. Upper Bedoulian, Pichouraz section, sample 2286. 20-22. Lenticulina cf. nodosa (). Upper Bedoulian, Pichouraz section. 20-21: sample 2288; 20: lateral view, 21: profile. 22: sample 2293. 23-24. Astacolus crepidularis subsp. tricarinella (). Upper Bedoulian, Pichouraz section, sample 2284.
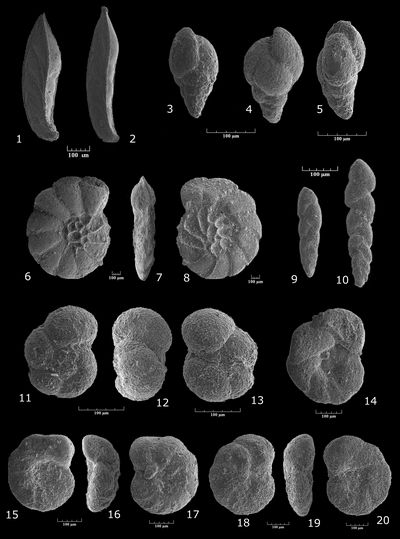
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 5 : 1-2. Saracenaria sp. aff. frankei sensu . Gargasien inférieur, coupe des Gays 1. 1Ā: échantillon 2337 ; 2Ā: éch. 2339. 3-5. Praebulimina sp. aff. elongata (). Bédoulien supérieur, coupe de Pichouraz. 3, 5Ā: éch. 2283 ; 4Ā: éch. 2286. 6-8. Epistomina sp. Bédoulien supérieur, coupe de Pichouraz, éch. 2289. 6Ā: vue ventrale, 7 : profil, 8 : vue dorsale. 9-10. Coryphostoma? sp. 9Ā: Bédoulien supérieur, coupe de Pichouraz, éch. 2291. 10Ā: Gargasien inférieur, coupe de La Tuilière W, éch. 2354. 11-13. "Valvulineria" sp. Gargasien inférieur, coupe de Pichouraz, éch. 2358. 11 : vue dorsale, 12 : profil, 13 : vue ventrale. 14-17. Gavelinella sp. aff. flandrini . Gargasien inférieur, coupe de La Tuilière W, éch. 2368. 14 : vue dorsale. 15 : vue dorsale, 16 : profil, 17 : vue ventrale. 18-20. Gavelinella sp. gr. flandrini . Gargasien inférieur, coupe de Pichouraz, éch. 2359.
Plate 5: 1-2. Saracenaria sp. aff. frankei sensu . Lower Gargasian, Les Gays 1 section. 1: sample 2337; 2: sample 2339. 3-5. Praebulimina sp. aff. elongata (). Upper Bedoulian, Pichouraz section. 3, 5: sample 2283; 4: sample 2286. 6-8. Epistomina sp. Upper Bedoulian, Pichouraz section, sample 2289. 6: ventral view, 7: profile, 8: dorsal view. 9-10. Coryphostoma? sp. 9: upper Bedoulian, Pichouraz section, sample 2291. 10: lower Gargasian, La Tuilière W section, sample 2354. 11-13. "Valvulineria" sp. Lower Gargasian, Pichouraz section, sample 2358. 11: dorsal view, 12: profile, 13: ventral view. 14-17. Gavelinella sp. aff. flandrini . Lower Gargasian, La Tuilière W section, sample 2368. 14: dorsal view. 15: dorsal view, 16: profile, 17: ventral view. 18-20. Gavelinella sp. gr. flandrini . Lower Gargasian, Pichouraz section, sample 2359.
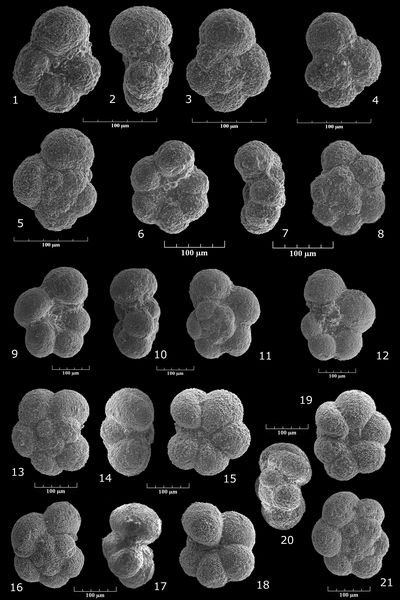
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 6 : 1-5. Praehedbergella sigali (). Bédoulien supérieur, coupe de Pichouraz, échantillon 2291. 1 : vue ventrale, 2 : profil, 3 : vue dorsale ; 4-5 : vues dorsales (deux spécimens distincts). 6-8. Praehedbergella primare ( & ). Bédoulien supérieur, coupe de Pichouraz, éch. 2293. 6 : vue ventrale, 7 : profil, 8 : vue dorsale. 9-12. Praehedbergella aptiana (). 9-11 : Bédoulien supérieur, coupe de Pichouraz, éch. 2290 ; 9 : vue ventrale, 10 : profil, 11 : vue dorsale. 12 : Gargasien inférieur, coupe des Gays 1, éch. 2340, vue ventrale. 13-21. Praehedbergella infracretacea (). 13-15 : Gargasien inférieur, coupe des Gays 1, éch. 2334. 16-21 : Gargasien inférieur, coupe des Gays 2 ; 16-18 : éch. 2364. 19-21 : éch. 2345. 13, 16, 21 : vues dorsales ; 14, 17, 20 : profils ; 15, 18-19 : vues ventrales.
Plate 6: 1-5. Praehedbergella sigali (). Upper Bedoulian, Pichouraz section, sample 2291. 1: ventral view, 2: profile, 3: dorsal view; 4-5: dorsal views (two distinct specimens). 6-8. Praehedbergella primare ( & ). Upper Bedoulian, Pichouraz section, sample 2293. 6: ventral view, 7: profile, 8: dorsal view. 9-12. Praehedbergella aptiana (). 9-11: upper Bedoulian, Pichouraz section, sample 2290; 9: ventral view, 10: profile, 11: dorsal view. 12: lower Gargasian, Les Gays 1 section, sample 2340, ventral view. 13-21. Praehedbergella infracretacea (). 13-15: lower Gargasian, Les Gays 1 section, sample 2334. 16-21: lower Gargasian, Les Gays 2 section; 16-18: sample 2364. 19-21: sample 2345. 13, 16, 21: dorsal views; 14, 17, 20: profiles; 15, 18-19: ventral views.
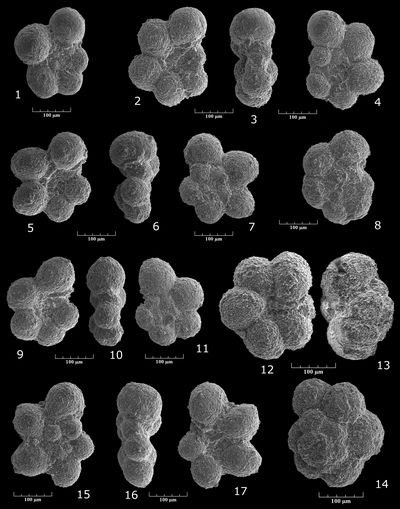
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 7 : 1. Praehedbergella aptiana (). Bédoulien supérieur, coupe de Pichouraz, échantillon 2290. 2-7, 9-11, 15-17. Praehedbergella globulifera ( & ) (série de stades transitionnels). 8. Praehedbergella primare ( & ). Bédoulien supérieur, coupe de Pichouraz, éch. 2293. 12-14. Praehedbergella praetrocoidea ( & ). Gargasien basal, coupe de Pichouraz, éch. 2367. 12 : vue ventrale, 13 : profil, 14 : vue dorsale.
Plate 7: 1. Praehedbergella aptiana (). Upper Bedoulian, Pichouraz section, sample 2290. 2-7, 9-11, 15-17. Praehedbergella globulifera ( & ) (successive transitional stages). 8. Praehedbergella primare ( & ). Upper Bedoulian, Pichouraz section, sample 2293. 12-14. Praehedbergella praetrocoidea ( & ). Lowermost Gargasian, Pichouraz section, sample 2367. 12: ventral view, 13: profile, 14: dorsal view.
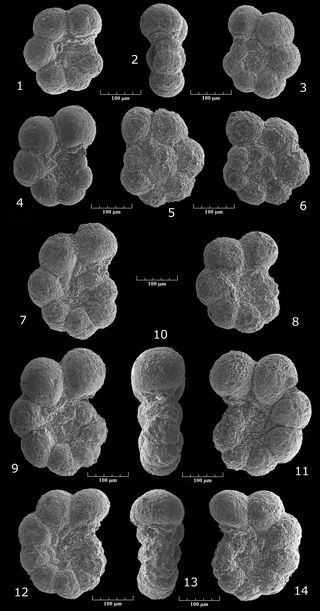
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 8 : 1-6. Praehedbergella luterbacheri (). 1-3. Gargasien inférieur, coupe de La Tuilière W, éch. 2368 ; 1 : vue ventrale, 2 : profil, 3 : vue dorsale. 4. Gargasien inférieur, coupe de Pichouraz, éch. 2358 ; vue ventrale. 5. Gargasien inférieur, coupe de La Tuilière W, éch. 2368 ; vue dorsale. 6. Gargasien basal, coupe de Pichouraz, éch. 2367 ; vue dorsale. 7-8. Globigerinelloides ferreolensis heptacameratus n. subsp. 7 : Paratype, Gargasien inférieur, coupe des Gays 1, éch. 2340. 8 : Holotype, Gargasien moyen?, coupe des Gays 2, éch. 2301. 9-11. Globigerinelloides ferreolensis ferreolensis (). Gargasien inférieur, coupe des Gays 1, éch. 2340. 9, 11 : vues latérales, 10 : profil. 12-14. Globigerinelloides barri (, & ). Gargasien moyen?, coupe des Gays 2, éch. 2301. 12, 14 : vues latérales, 13 : profil.
Plate 8 : 1-6. Praehedbergella luterbacheri (). 1-3. Lower Gargasian, La Tuilière W section, sample 2368 ; 1: ventral view, 2: profile, 3: dorsal view. 4. Lower Gargasian, Pichouraz section, sample 2358 ; ventral view. 5. Lower Gargasian, La Tuilière W section, sample 2368 ; dorsal view. 6. Lowermost Gargasian, Pichouraz section, sample 2367 ; dorsal view. 7-8. Globigerinelloides ferreolensis heptacameratus n. subsp. 7: Paratype, lower Gargasian, Les Gays 1 section, sample 2340. 8: Holotype, middle Gargasian ?, Les Gays 2 section, sample 2301. 9-11. Globigerinelloides ferreolensis ferreolensis (). Lower Gargasian, Les Gays 1 section, sample 2340. 9, 11: lateral views, 10: profile. 12-14. Globigerinelloides barri (, & ). Middle Gargasian?, Les Gays 2 section, sample 2301. 12, 14: lateral views, 13: profile.
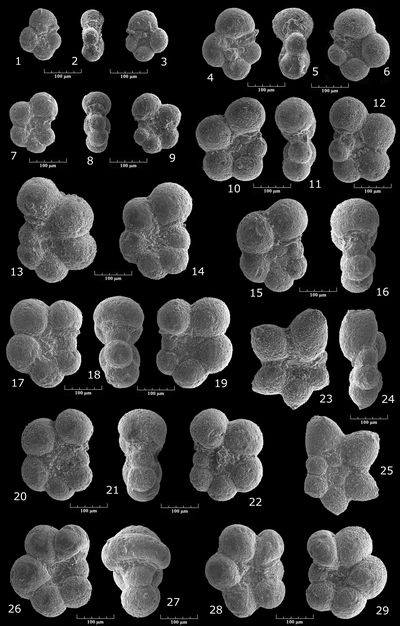
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 9 : 1-12. Globigerinelloides sp. gr. duboisi-blowi (série de formes transitionnelles entre les deux "espèces"). 1-3 : morphotype duboisi, à 4 loges au dernier tour. Bédoulien terminal, coupe de La Tuilière W, échantillon 2369. 4-6 : morphotype duboisi, à 4+ loges. Gargasien inférieur, coupe des Gays 2, éch. 2305. 7-9 : morphotype blowi, à 5,5 loges. Gargasien inférieur, coupe de La Tuilière W, éch. 2352. 10-11 : morphotype blowi, à 5+ loges. Gargasien inférieur, coupe des Gays 2, éch. 2364. 13-22, 26-29. Globigerinelloides sp. gr. aptiensis-solidus. 13-16 : morphotypes aptiensis ; 17-22 : morphotypes solidusĀ; 26-29 : morphotypes solidus à tendance "biglobigerinelloïde". 13 : Gargasien inférieur, coupe des Gays 1, éch. 2328 ; 14 : Gargasien inférieur, coupe des Gays 1, éch. 2336 ; 15-16 : Gargasien inférieur, coupe des Gays 1, éch. 2338 ; 17-19 : Gargasien inférieur, coupe des Gays 1, éch. 2336 ; 20-22 : Gargasien inférieur, coupe des Gays 2, éch. 2305Ā; 26-29 : Gargasien inférieur, coupe des Gays 2, éch. 2345 (deux spécimens distincts : 26-27 [26 : vue latérale, 27 : profil] [28-29 : vues latérales]). 23-25. Pseudoschackoina saundersi (). Bédoulien supérieur, coupe de Pichouraz, éch. 2286. 23 : vue ventrale, 24 : profil, 25 : vue dorsale.
Plate 9: 1-12. Globigerinelloides sp. gr. duboisi-blowi (succession of transitional form between the two "species"). 1-3: 4-chambered morphotype duboisi. Uppermost Bedoulian, La Tuilière W section, sample 2369. 4-6: 4+-chambered morphotype duboisi. Lower Gargasian, Les Gays 2 section, sample 2305. 7-9: 5.5-chambered morphotype blowi. Lower Gargasian, La Tuilière W section, sample 2352. 10-11: morphotype blowi, à 5+ loges. Lower Gargasian, Les Gays 2 section, sample 2364. 13-22, 26-29. Globigerinelloides sp. gr. aptiensis-solidus. 13-16: morphotypes aptiensis; 17-22: morphotypes solidus; 26-29: morphotypes solidus showing a "biglobigerinelloid" trend. 13: lower Gargasian, Les Gays 1 section, sample 2328; 14: lower Gargasian, Les Gays 1 section, sample 2336; 15-16: lower Gargasian, Les Gays 1 section, sample 2338; 17-19: lower Gargasian, Les Gays 1 section, sample 2336; 20-22: lower Gargasian, Les Gays 2 section, sample 2305; 26-29: lower Gargasian, Les Gays 2 section, sample 2345 (two distinct specimens: 26-27 [26: lateral view, 27: profile] [28-29: lateral views]). 23-25. Pseudoschackoina saundersi (). Upper Bedoulian, Pichouraz section, sample 2286. 23: ventral view, 24: profile, 25: dorsal view.
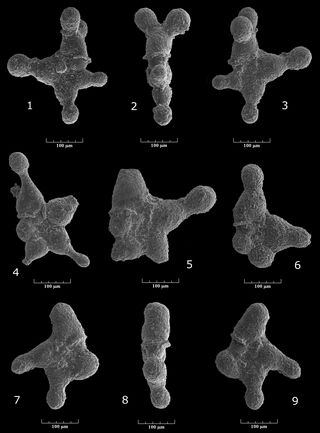
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 10 : 1-9. Schackoina (Leupoldina) sp. gr. cabri (). 1-4 : morphotypes cabriĀ; 5-9 : morphotypes pustulans. 1-3 : Bédoulien terminal, coupe de Pichouraz, échantillon 2366 ; 1, 3 : vues latérales, 2 : profil. 4 : Bédoulien terminal, coupe de La Tuilière W, éch. 2369 ; 5. Bédoulien supérieur, coupe de Pichouraz, éch. 2292 ; 6. Gargasien inférieur, coupe de Pichouraz, éch. 2356 ; 7-9. Bédoulien supérieur, coupe de Pichouraz, éch. 2292 ; 7, 9 : vues latérales, 8 : profil.
Plate 10: 1-9. Schackoina (Leupoldina) sp. gr. cabri (). 1-4: morphotypes cabri; 5-9: morphotypes pustulans. 1-3: uppermost Bedoulian, Pichouraz section, sample 2366; 1, 3: lateral views, 2: profile. 4: uppermost Bedoulian, La Tuilière W section, sample 2369; 5. Upper Bedoulian, Pichouraz section, sample 2292; 6. Lower Gargasian, Pichouraz section, sample 2356; 7-9. Upper Bedoulian, Pichouraz section, sample 2292; 7, 9: lateral views, 8: profile.