![]()
Contents
[1. Introduction] [2. Material and methods]
[3. Results]
[4. Variety]
[5. Conclusion]
[Bibliographic references]
[Figures] [Plates] [Video] [Appendix
1] and ... [Appendix 2]
![]()
![]()
Through the use of acetolysis new micropalaeontological studies on the type section of the Campanian-Maastrichtian boundary at Tercis (SW France) we obtained additional information on ostracodes. Acetolysis on hard carbonates levels of the Tercis quarry found 22 species in addition to those of the studies published in 2001. Today, 75 species are recognized: they represent 34 known genera, and 4 currently unidentified. Small species usually rare of the genera Aversovalva, Bythoceratina and Eucytherura were collected. All of the assemblages (of both genera and species) are characteristic of a Campanian-Maastrichtian open carbonate platform environment.
Ostracodes; Campanian; Maastrichtian; stratotype; Tercis; Aquitaine; France; acetolysis.
B. & G.S. (2008).- New micropalaeontological studies on the type section of the Campanian-Maastrichtian at Tercis (SW France): new ostracodes obtained using acetolysis.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/02 (CG2008_A02)
Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : compléments sur les ostracodes extraits par acétolyse.- Une nouvelle étude de la microfaune, obtenue par acétolyse des niveaux indurés dans le Campanien-Maastrichtien de la carrière de Tercis, a permis de récolter 22 espèces additionnelles au regard de l'étude publiée en 2001 et de porter ainsi à 75 espèces, réparties sur 34 genres reconnus et 4 indéterminés, la biodiversité en ostracodes de ce site. Des espèces de petite taille, généralement rares, rapportées aux genres Aversovalva, Bythoceratina et Eucytherura ont par ailleurs été recueillies. Les associations de genres et d'espèces d'ostracodes sont caractéristiques du Campanien-Maastrichtien, dans un environnement de plate-forme carbonatée ouverte.
Ostracodes ; Campanien ; Maastrichtien ; stratotype ; Tercis ; Aquitaine ; France ; acétolyse.
The stratotype section of the Campanian-Maastrichtian boundary at Tercis (Landes, France) was accepted in 2001 ( & ,
2001) following an extensive integrated study (,
2001a) including the documentation of 18 fossil groups illustrated in the second work and later
in et alii (2006). This well-known section
(Fig. 1 ![]() ) was an appropriate choice for the better establishment of the palaeobiodiversity of Upper Cretaceous deposits, at a time when dinosaurs were the top of the biological pyramid just before the Cretaceous-Palaeogene global crisis.
) was an appropriate choice for the better establishment of the palaeobiodiversity of Upper Cretaceous deposits, at a time when dinosaurs were the top of the biological pyramid just before the Cretaceous-Palaeogene global crisis.
Among the possible ways to arrive at a better knowledge, it was suspected that an intensive study directed particularly at the indurated strata comprising the larger portion of the Tercis series
(Figs. 2 ![]() - 3
- 3 ![]() ) would be profitable. In 2001 the study of microfossils, with the exception of the investigation by thin section of planktonic foraminifera, was restricted to the easily disaggregated levels: those richest in clay. At Tercis, the clay content of these levels reached a maximum of 15 % (,
2001c). However, from the environmental point of view, these levels indicate an input of detritus which could influence the development of micro-organisms. The practical selection of the more clayish levels might have discriminated those organisms more tolerant to this detrital input and eliminated those requiring clean waters. This was why we attempted to dissociate rocks using more or less concentrated acetic acid at different temperatures.
) would be profitable. In 2001 the study of microfossils, with the exception of the investigation by thin section of planktonic foraminifera, was restricted to the easily disaggregated levels: those richest in clay. At Tercis, the clay content of these levels reached a maximum of 15 % (,
2001c). However, from the environmental point of view, these levels indicate an input of detritus which could influence the development of micro-organisms. The practical selection of the more clayish levels might have discriminated those organisms more tolerant to this detrital input and eliminated those requiring clean waters. This was why we attempted to dissociate rocks using more or less concentrated acetic acid at different temperatures.
We obtained a satisfying disaggregation of the hard rocks in most of the levels investigated. Picking of the several size fractions of the resulting sands led to the obtention of new forms of previously investigated fossil groups as well as to the discovery of a number of previously unknown forms: microproblematica apparently needing clean waters ( & , 2006; , 2007; et alii, 2007). Bryozoans, holothurian, crinoid and ophiuroid remains, benthic and planktonic foraminifera, ... will be studied later, we hope. The present work reports only the information obtained concerning ostracodes: previously unreported taxa of these microfossils were found and the distribution of known forms was made more nearly exhaustive.
The stratotype section of the Campanian-Maastrichtian boundary is defined in the Grande Carrière at Tercis which was quarried until 1997. Tercis is located in the Landes near Dax, in southern Aquitaine, France, about 10 km north of the first North Pyrenean Thrusts (,
2001). A continuous section about 200 m thick was described ( & ,
1994). It is 8 Ma of deposits bracketing the Campanian-Maastrichtian boundary which is conventionally sited at level 115.2 in accordance with an original approach: this convention involves the use of a combination of 12 biostratigraphical signals pertaining to 6 fossil groups contiguous to a guide event selected by convention (,
2001b; & ,
2001). The homogeneous lithology of the section is shown on
Figure 4 ![]() which identifies 3 units. The d'Avezac Unit is a series of limestones of which the 5 sub-units (A1 to A5) are distinguished according to their respective content of glaucony
(glaucony: facies as opposed to glauconite: mineral according to & ,
1981) that alternates between very low and high. The Les Vignes Unit uncludes a sub-unit with light flint (V1) below and a sub-unit with dark flint (V2) above. The lithological column indicates the presence of glaucony (dotted) and of flint; flints are commonly concentrated in levels of dm thickness with a metric periodicity. The numerical ages are suggested according to 1- magnetostratigraphic and biostratigraphic correlations with radiometrically dated bentonite levels in the United States ( & ,
2001), 2- a uniformity in the rate of sedimentation is supported by the measurable (metric) periodicity of the deposits (orbital in origin), 3- the combination of this consistency with the examination (presence, abundance, and crystallinity) of glauconitic minerals ( & ,
2001). The age of the boundary suggested in 2001 is improved in
accordance with recently established Europe-America correlations using inoceramids ( & ,
2003). The ages indicated alongside the column are estimates based on acceptance of 71.6 Ma as the date of the boundary with an uncertainty of
±0.6 Ma (confidence level 95 %).
which identifies 3 units. The d'Avezac Unit is a series of limestones of which the 5 sub-units (A1 to A5) are distinguished according to their respective content of glaucony
(glaucony: facies as opposed to glauconite: mineral according to & ,
1981) that alternates between very low and high. The Les Vignes Unit uncludes a sub-unit with light flint (V1) below and a sub-unit with dark flint (V2) above. The lithological column indicates the presence of glaucony (dotted) and of flint; flints are commonly concentrated in levels of dm thickness with a metric periodicity. The numerical ages are suggested according to 1- magnetostratigraphic and biostratigraphic correlations with radiometrically dated bentonite levels in the United States ( & ,
2001), 2- a uniformity in the rate of sedimentation is supported by the measurable (metric) periodicity of the deposits (orbital in origin), 3- the combination of this consistency with the examination (presence, abundance, and crystallinity) of glauconitic minerals ( & ,
2001). The age of the boundary suggested in 2001 is improved in
accordance with recently established Europe-America correlations using inoceramids ( & ,
2003). The ages indicated alongside the column are estimates based on acceptance of 71.6 Ma as the date of the boundary with an uncertainty of
±0.6 Ma (confidence level 95 %).
Figure 4 ![]() summarizes the quantitative results obtained in 2001 ( & ,
2001; , 2001). The first represents the collection of 1800 specimens (from about 45 levels) and the second, 2500 specimens (from 28 levels). A synthetic report ( & ,
2001) cited 53 species pertaining to 28 known and 3 unidentified genera. All the genera mentioned were known from the Upper Cretaceous of Europe (mainly from northwestern Europe due to the fact that few studies of the Aquitaine Basin existed) and, secondarily, from North Africa. About ten species known from NW Europe or other areas in Europe have also been found in North America and on the southern margin of the Mediterranean. The conclusion was that no endemism existed in the North Pyrenean appendix of the Atlantic Ocean, the domain where Tercis was though to have been located during the Late Cretaceous.
summarizes the quantitative results obtained in 2001 ( & ,
2001; , 2001). The first represents the collection of 1800 specimens (from about 45 levels) and the second, 2500 specimens (from 28 levels). A synthetic report ( & ,
2001) cited 53 species pertaining to 28 known and 3 unidentified genera. All the genera mentioned were known from the Upper Cretaceous of Europe (mainly from northwestern Europe due to the fact that few studies of the Aquitaine Basin existed) and, secondarily, from North Africa. About ten species known from NW Europe or other areas in Europe have also been found in North America and on the southern margin of the Mediterranean. The conclusion was that no endemism existed in the North Pyrenean appendix of the Atlantic Ocean, the domain where Tercis was though to have been located during the Late Cretaceous.
Acetolysis was employed first on sub-unit A3 of the d'Avezac Unit where
Radotruncana calcarata (, 1927) occurs; this is a recognized and precise biostratigraphical marker used by the experts on Upper Cretaceous planktonic foraminifers. From 100 to 200 g of treated
sample 200 or more specimens of ostracodes were collected from a few levels
(Fig. 4 ![]() ). Acetolyzed samples from the bottom of the quarry, between levels -5 and +25 produced no more fauna than water-washed sediments from those levels so they have not been extensively hand-picked and processed. Between levels 95 and
170 the fauna was meager and poorly preserved so few levels have been investigated. In all, about 1700 valves were collected from 15 selected levels, and these were considered for illustration and identification during the present study.
Figure 5
). Acetolyzed samples from the bottom of the quarry, between levels -5 and +25 produced no more fauna than water-washed sediments from those levels so they have not been extensively hand-picked and processed. Between levels 95 and
170 the fauna was meager and poorly preserved so few levels have been investigated. In all, about 1700 valves were collected from 15 selected levels, and these were considered for illustration and identification during the present study.
Figure 5 ![]() illustrates some interesting forms observed using an optical microscope; they are pictured because we had not seen them in the boundary stratotype before.
illustrates some interesting forms observed using an optical microscope; they are pictured because we had not seen them in the boundary stratotype before.
The collected shells and valves are made fragile by the acetic acid treatment and are sometimes difficult to collect and picture. Their surface is commonly partly dissolved and the ornamentation poorly preserved; specimens obtained from water-washed residues were also poorly preserved.
Diagnoses of the genera, as accepted by the authors, were given in the previous publications on the Campanian-Maastrichtian boundary stratotype at Tercis ( & , 2001; , 2001). Although they might be open to question these diagnoses are necessary in order to inform the reader how and why the forms were segregated and named. The new names are proposed here and our interpretation of the genera is presented below. Generic identifications are based on external morphological characters of the shell; internal characters are not available. The previously identified genera are not described again.
Species added to those in the 2001 publications are illustrated (see Appendix list of species), and the documentation of two previously described species is supplemented.
- Asciocythere , 1952: middle-sized carapace, ovate-triangular in side view, with indistinct angles in lateral view; both ends rounded, turned toward the ventral margin; left valve overlapping all the way around the right; carapace smooth to punctate.
- Cardobairdia , 1960: small- to middle-sized carapace, generally thick, sub-elliptical and smooth; anterior end regularly rounded and turned toward the ventral margin; posterior end rather acuminate; left valve overlaps the right all the way around and more conspicuously on the dorsal and ventral margins.
- Paracyprideis , 1929: middle-sized carapace, sub-elliptical to triangular in lateral view; anterior end regularly rounded, rather turned toward the ventral margin, posterior end triangular near the ventral margin; some denticles or spines on the ends; highest before mid-length; surface generally smooth except when bored with wide pores.
- Eucytherura , 1894: small-sized carapace, rhombic to square in lateral view; dorsal margin straight, ventral margin rounded; anterior end widely rounded, posterior end triangular, with caudal process turned to the dorsal margin; eye spot on the antero-dorsal angle; tuberculate or reticulate surface.
- Pterygocythere , 1954: triangular carapace in lateral view, inflated ventrally, with a strong posteriorly pointed alae in prolongation of the anterior flange, sometimes broken into a row of spines; acuminate posterior margin; surface smooth with some spines or tubercles; costa on the dorsal margin; distinct eye spot; sometimes with spines on the anterior and posterior ends; inflated ventral face with longitudinal costa; simple primary ornament; thick and robust carapace.
- Diogmopteron , 1954: identical in all external morphological features to Pterygocythere, with the exception of the hinge (not distinct here); the right valve overlaps the left on the dorsal margin of Diogmopteron, this character is the opposite on Pterygocythere (not distinct on our individual).
- Rehacythereis , 1973: carapace triangular to rectangular in lateral view; anterior end regularly rounded, posterior end triangular; anterior flange connected with the ventral costa ended with a postero-ventral tubercle; independent dorsal costa, that ends with a postero-dorsal tubercle, and connected with a narrow costa running to the well-marked sub-central tubercle; surface reticulate; right valve overlaps the left particularly on the antero-dorsal angle.
- Genus indet. (Pl. 2 ![]() ,
figs. 22-26): large-sized carapace, sub-rectangular; large frontal lobe; eye-spot on the antero-dorsal angle; anterior margin regularly rounded overtaking the ventral margin; posterior end triangular with concave dorsal margin and convex, rather spinely, ventral margin; discrete independent dorsal and ventral costa; indistinct sub-central tubercle; carapace inflated laterally; the left valve overlaps the right on the antero- and postero-dorsal angles; surface smooth, sometimes with granulation.
,
figs. 22-26): large-sized carapace, sub-rectangular; large frontal lobe; eye-spot on the antero-dorsal angle; anterior margin regularly rounded overtaking the ventral margin; posterior end triangular with concave dorsal margin and convex, rather spinely, ventral margin; discrete independent dorsal and ventral costa; indistinct sub-central tubercle; carapace inflated laterally; the left valve overlaps the right on the antero- and postero-dorsal angles; surface smooth, sometimes with granulation.
During the study preliminary to the designation of the boundary stratotype (, 2001; & , 2001; & , 2001) 53 species of ostracodes were listed. Here, we supplement this first evaluation with 22 additional species. Thus, as a whole the ostracode fauna of the Campanian-Maastrichtian at Tercis comprises a total of 75 species pertaining to 34 known genera, including the 7 additional entities quoted in this work, and 4 non-dentified genera. Disaggregation of the hard carbonate levels using acetolysis aided greatly in the discovery of species of the small genera Aversovalva, Bythoceratina, and Eucytherura. These minute specimens are comparatively rare and may often pass unnoticed in washed residues.
We have identified and named 8 species in this work, among them, 3 are cf. and 4 are aff.. The list is as follows: Eucytherura (Vesticytherura) dorsotuberculata (, 1936), Phacorhabdotus cf. lonsdaleianus lonsdaleianus (, 1849); Xestoleberis cf. northensis , 1966, Bythoceratina cf. umbonata umbonata (, 1848), Paracaudites (Dumontina) aff. puncturata (, 1854), Pterygocythere aff. hilli , 1957, Rehacythereis aff. agedincumensis (, 1964), Spinoleberis aff. macerrima (, 1936).
The stratigraphical locations of the taxa identified in the present study are given in
Figure 6 ![]() . Eucytherura (Vesticytherura) dorsotuberculata is known from the upper Campanian of Dordogne, SW France, the upper Campanian-Palaeocene-Eocene? of Poland, the upper Campanian-Maastrichtian of Germany, the lower Maastrichtian of Belgium and the Netherlands, the Maastrichtian of Denmark (full references
in , 1973, and ,
1983). Phacorhabdotus lonsdaleianus lonsdaleianus is reported from the upper Campanian-Maastrichtian of England, the upper Campanian of the Paris Basin in France, the Campanian-Maastrichtian of Germany, the lower Maastrichtian of Poland, the Maastrichtian of Belgium, the Netherlands, and Denmark (full references
in , 1983). Xestoleberis northensis is known from the upper Campanian-Maastrichtian of Belgium and the Netherlands, the middle Campanian of Dordogne and the upper Campanian of the Paris Basin and the Charentes,
SW France (full references in , 1973). Bythoceratina umbonata umbonata
- sensu 1983) - is a Campanian-Maastrichtian species: upper Campanian of Belgium, Campanian-Maastrichtian of Poland, lower Maastrichtian of the Netherlands and Denmark, Maastrichtian of Germany, Upper Cretaceous of England and Ireland (full references
in , 1973, and ,
1983). Paracaudites (Dumontina) puncturata is collected from the Campanian of Dordogne and Charente, France (,
1973; et alii, 1985), and the Maastrichtian of the Netherlands and Belgium (,
1966). Pterygocythere hilli - sensu
(1983) - is known from the middle Santonian to the lower Campanian of Germany. The species is also known from the Eocene of Belgium and the Netherlands (,
1983). Rehacythereis agedincumensis is dated Santonian-Campanian in the Paris Basin (,
1964; et alii,
1985); the species (or a closely related one, given the discrete stratigraphical location) is also quoted from the upper Cenomanian-lower Turonian of Corbières,
S France ( & ,
2006). Spinoleberis macerrima (, 1936) is known from the Maastrichtian of the Netherlands (,
1966) and the Campanian of Aquitaine (Spinoleberis cf. macerrima
in , 1973).
. Eucytherura (Vesticytherura) dorsotuberculata is known from the upper Campanian of Dordogne, SW France, the upper Campanian-Palaeocene-Eocene? of Poland, the upper Campanian-Maastrichtian of Germany, the lower Maastrichtian of Belgium and the Netherlands, the Maastrichtian of Denmark (full references
in , 1973, and ,
1983). Phacorhabdotus lonsdaleianus lonsdaleianus is reported from the upper Campanian-Maastrichtian of England, the upper Campanian of the Paris Basin in France, the Campanian-Maastrichtian of Germany, the lower Maastrichtian of Poland, the Maastrichtian of Belgium, the Netherlands, and Denmark (full references
in , 1983). Xestoleberis northensis is known from the upper Campanian-Maastrichtian of Belgium and the Netherlands, the middle Campanian of Dordogne and the upper Campanian of the Paris Basin and the Charentes,
SW France (full references in , 1973). Bythoceratina umbonata umbonata
- sensu 1983) - is a Campanian-Maastrichtian species: upper Campanian of Belgium, Campanian-Maastrichtian of Poland, lower Maastrichtian of the Netherlands and Denmark, Maastrichtian of Germany, Upper Cretaceous of England and Ireland (full references
in , 1973, and ,
1983). Paracaudites (Dumontina) puncturata is collected from the Campanian of Dordogne and Charente, France (,
1973; et alii, 1985), and the Maastrichtian of the Netherlands and Belgium (,
1966). Pterygocythere hilli - sensu
(1983) - is known from the middle Santonian to the lower Campanian of Germany. The species is also known from the Eocene of Belgium and the Netherlands (,
1983). Rehacythereis agedincumensis is dated Santonian-Campanian in the Paris Basin (,
1964; et alii,
1985); the species (or a closely related one, given the discrete stratigraphical location) is also quoted from the upper Cenomanian-lower Turonian of Corbières,
S France ( & ,
2006). Spinoleberis macerrima (, 1936) is known from the Maastrichtian of the Netherlands (,
1966) and the Campanian of Aquitaine (Spinoleberis cf. macerrima
in , 1973).
The presence of these additional species confirms the stratigraphical location of the ostracode faunal associations of the Campanian-Maastrichtian. Accordingly, it appears that when compared with the fauna of the Atlantic ocean at this time no endemism exists at Tercis, then sited within a north Pyrenean sound (although the very small number of taxa fully identified at the species level might also be due, in addition to the state of preservation, to the fact that some indication of original fauna is present here). According to the ostracode fauna, during Campanian times the Tercis area was part of the Boreal European Palaeobiogeographical Unit, with temperate warm waters as defined by et alii (2004). This Unit includes the Basco-Cantabrian Basin, the Aquitaine Basin, the Paris Basin, England, Belgium, the Netherlands, Germany, Poland, and Russia. Nevertheless, the genera Dumontina and Limburgina present at Tercis are signs that point to the north-Tethyan margin. A Tethyan or Boreal assignment of the Tercis area thus depends on the criteria considered; & (in et alii, 2001) argue that in Aquitaine the Tethyan domain is present as far North as the Charente because of the presence of large benthic foraminifera and rudists along with the absence of belemnites and of a true chalk facies; all these points are fully valid at Tercis.
The association of the genera Dumontina, Limburgina, Mauritsina, Spinoleberis, Pterygocythere, Diogmopteron, Phacorhabdotus, Imhotepia, Cytherelloidea, Bythoceratina, Cardobairdia, Krithe and Eucytherura, as identified by & (2001), (2001) and & (2001), characterises the euphotic zone of an outer shelf carbonate platform at Tercis. It is intermediate between the two environments (shallow, high energy and a deeper water, low energy) suggested by et alii (2004), and distinguished in the Boreal European Palaeobiogeographical Unit by discrete associations of genera. The shallow high-energy environment is indicated by the genera Dumontina, Kikliocythere, Limburgina, Mauritsina, Mosaeleberis and Spinoleberis, and the deeper water, lower energy environment by the genera Trachyleberidea, Pterygocythere, Alatacythere, Phacorhabdotus, Imhotepia, and species of the Bythocytheridae family.
The present study is part of a comprehensive micropalaeontological investigation undertaken on the hard levels of the Campanian-Maastrichtian boundary stratotype at Tercis (southern Aquitaine). Here we report on ostracodes of which additional taxa were brought to light, and in particular some small forms found but rarely in the samples.
The palaeobiodiversity increased from the 53 species recognised in 2001 to a total of 75 species. Although this number should be regarded with caution due to the real difficulty of compiling a homogeneous terminology of the collected forms due to the nomenclatural differences among authors, these relative numbers (75 versus 53) prove the scientific value of the study of hard levels using acetolysis as a mean for disaggregation of the rocks. With this approach, the actual palaeobiodiversity of the regional basin is better documented and the relationship of this global reference section to that of nearby palaeobiogeographic domains can be better understood.
The genera and species of the ostracode associations at Tercis confirm its Campanian-Maastrichtian age, and a carbonate platform environment; ostracodes also provide unequivocal evidence of relationships with the Boreal Domain while other faunas have shown that Tercis, and the whole Aquitaine Basin, is better interpreted as a portion of the Tethyan domain, at least during the younger portion of the Campanian and thereafter.
One of us (B.A.) thanks T. , LMTG, Université Toulouse III, for pictures using Scanning Electronic Microscope Jeol JSM-6360LV, J.-F. , Marseille, and J.-P. , Cestas, France, for the suggested improvements in
taxonomy. Pictures 40 and 41 and those of
Figure 5 ![]() and Appendix 2
and Appendix 2 ![]() were obtained at the Université Pierre et Marie Curie, Paris. We acknowledge the help of Nestor , Modesto, USA, who is responsible
for improvements in the intelligibility of the English version of this paper.
were obtained at the Université Pierre et Marie Curie, Paris. We acknowledge the help of Nestor , Modesto, USA, who is responsible
for improvements in the intelligibility of the English version of this paper.
An additional specimen of Eucytherura (Vesticytherura) dorsotuberculata was collected from level 120.4, i.e. about 0.2 Ma above the Campanian-Maastrichtian boundary as defined at Tercis.
An additional specimen of Aversvalva sp. was collected from a level located 9.6 m below and consequently less than 0.4 Ma before the Maastrichtian-Danian boundary.
B. (2001).- Ostracodes du site de Tercis les Bains (Landes, France). In: G.S. (ed., 2001a), chap. C6b, p. 402-409.
B. & M. (2006).- Ostracodes du Cénomanien supérieur et du Turonien de la zone sous-pyrénéenne orientale (Corbières méridionales, SE France). Systématique, biostratigraphie et paléobiogéographie.- Revue de Micropaléontologie, Paris, vol. 49, n° 2, p. 55-73.
B. & G.S. (2001).- Synthetic data on the Campanian-Maastrichtian ostracode fauna from Tercis les Bains (Landes, France). In: G.S. (ed., 2001a), chap. C6c, p. 410-413.
J.-F., J.-P. & R. (1985).- Crétacé supérieur. In: H.J., Atlas des Ostracodes de France.- Bulletin des Centres de Recherche Exploration-Production elf-Aquitaine, Pau, Mémoire 9, p. 211-255.
M., G.S. & B. (2001).- Geology and Late Cretaceous palaeogeography of the geological site at Tercis les Bains (Landes, France). In: G.S. (ed., 2001a), chap. A4, p. 47-59.
J.-P. (1973).- Étude stratigraphique et micropaléontologique du Crétacé supérieur de la région de Saint-Cyprien (Dordogne).- Thèse de Doctorat de 3° Cycle, Université Paris VI, 287 p.
B. (1983).- Die Cytheracea (Ostracoda) im Schreibkreide-Richtprofil von Lägerdorf-Kronsmoor-Hemmoor (Coniac bis Maastricht; Norddeutschland).- Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, Heft 54, p. 65-168.
R. (1964).- Contribution à l'étude des ostracodes du Sénonien de Sens (Yonne) : les Trachyleberididae.- Revue de Micropaléontologie, Paris, vol. 7, n° 2, p. 100-110.
R. & G.S. (2001).- Ostracodes du site de Tercis les Bains (Landes, France). In: G.S. (ed., 2001a), chap. C6a, p. 396-401.
G. (1966).- Cytheracea (Ostracodes) du Maastrichtien de Maastricht (Pays-Bas) et des regions voisines; résultats stratigraphiques et paléontologiques de leur étude.- Mededelingen van de geologische Stichting, Maastricht, (Serie C), V2, n° 2, 197 p.
Z. & G.S. (2001).- Magnetostratigraphy across the Campanian-Maastrichtian boundary at Tercis les Bains in comparison with northern Germany, the Apennines (Central Italy) and North America; biostratigraphical and geochronological constraints. In: G.S. (ed., 2001a), chap. B2d, p. 175-183.
G.S., editor (2001a).- The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36; Developments in Palaeontology and Stratigraphy Series, 19, Elsevier Sciences Publ. Amsterdam, xxviii + 881 p.
G.S. (2001b).- The Campanian-Maastrichtian boundary: definition at Tercis (Landes, SW France) principle, procedure, and proposal. In: G.S. (ed., 2001a), chap. F3, p. 820-833.
G.S. (2001c).- Descriptive lithostratigraphy of the Campanian-Maastrichtian succession at Tercis les Bains (SW France). In: G.S. (ed., 2001a), chap. B1a, p. 85-109.
G.S. (2007).- Sur des microfossiles calcaires problématiques du stratotype de la limite Campanien-Maastrichtien de Tercis (Sud-Ouest, France) : les gilianelles.- Comptes rendus Palévol, Paris, vol. 6, n° 3, p. 181-188.
G.S., C., A. & B.
(2006).- Présentation du site géologique de Tercis (Landes,
France) ; 13 affiches pédagogiques informatisées.
a- Le site de la Grande Carrière de Tercis ; haut lieu géologique, témoin des 10 Ma de la fin du Crétacé.
b- Les grands fossiles de la carrière de Tercis; une richesse et une diversité remarquable en macrofossiles.
c- Les bivalves de Tercis ; des fossiles marqueurs de l'environnement et un groupe marqueur de temps (les inocérames).
d- Les ammonites de Tercis ; un groupe disparu depuis 65 Ma, proche des nautiles et des seiches, très utile pour dater les sédiments de plate-forme.
e- Les échinides de Tercis.
f- Les astérides de Tercis; des fossiles communs sur la plate-forme continentale et utiles à la datation relative.
g- Les petits fossiles de la carrière de Tercis.
h- Les nannofossiles calcaires à Tercis.
i- Les kystes de dinoflagellés ; outil stratigraphique de datation relative excellemment illustré à Tercis.
j- Les foraminifères de la Grande Carrière de Tercis; une présence suffisante pour une datation relative acceptable.
l- Datation comparée Continent-Océan à Tercis.
m- Les sites préhistoriques à Tercis ; une grande carrière sur le plus grand site de plein air du sud-ouest de la France.
n- Le site de Tercis, stratotype de référence mondial ; définition du calendrier géologique et application à la limite d'étage Campanien-Maastrichtien.
Available on internet (since 14-VI-06): http://paleopolis.rediris.es/odin/
G.S., C., A. & B.
(2007).- Gilianelles et autres microfossiles problématiques
: contribution à l'étude de la paléobiodiversité et à la biostratigraphie crétacée dans le stratotype de la limite Campanien-Maastrichtien à Tercis (Sud Aquitaine, France).- Journée Hommage à Gérard , Paris 06-XII-2007.
Available on internet (since 21-XI-07): http://paleopolis.rediris.es/odin/images/poster-14.png
G.S. & A. (2001).- Interpretative reading of the Campanian-Maastrichtian deposits at Tercis les Bains: sedimentary breaks, rhythms, accumulation rate, sequences. In: G.S. (ed., 2001a), chap. B1c, p. 120-131.
G.S. & M.A. (2001).- The global Campanian-Maastrichtian stage boundary at Tercis les Bains, Landes, SW France.- Episodes, Beijing, vol. 24, n° 4, p. 229-238.
G.S. & A. (2006).- Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : les gilianelles (microproblematica) extraites par acétolyse.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2006/05 (CG2006_A05), 28 p., 3 videos.
G.S. & A. (1981).- De glauconiarum origine.- Sedimentology, Oxford, vol. 28, n° 5, p. 611-641.
G.S. & P. (1994).- Première description d'une coupe dans la localité aturienne de Tercis (Landes) : contribution à l'étude de la limite Campanien-Maastrichtien.- Géologie de la France, Orléans, n° 2 (1994), p. 31-37.
G.S. & I. (2003).- Sur les inocérames de Tercis (Landes, France) : le meilleur outil corrélatif entre Europe et Amérique du Nord autour de la limite Campanien-Maastrichtien / Inoceramids of the site of Tercis (Landes, France): the best correlative tool between Europe and North America across the Campanian-Maastrichtian boundary.- Comptes rendus Géosciences, Paris, vol. 335, n° 2, p. 239-246.
M. (2001).- The site at Tercis les Bains (Landes), stratotype of the Campanian-Maastrichtian limit: insights from tectonic studies. In: G.S. (ed., 2001a), chap. A6, p. 68-81.
J., J.-P. & G. (2004).- Global Campanian (Upper Cretaceous) ostracod palaeobiogeography.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 213, n° 3-4, p. 379-398.

Click on thumbnail to enlarge the image.
Figure 1: Aerial view of the geological site at Tercis (southern Aquitaine). North is located toward the top; strata are vertical; the Campanian levels are in the South followed to the North by those of the Maastrichtian, then the Danian. The quarry front, to the East/right, is made up of 5 platforms, 170 m wide, perpendicular to the strata, and moved along 400 m during quarrying (picture G.S.O., February 1997).

Click on thumbnail to enlarge the image.
Figure 2: View of platform II, levels 5 to 30, the main outcrop of the Tercis geological site (southern Aquitaine). Carbonate strata are vertical (level 20.0 shown) with a few decimeter-thick marly horizons (picture G.S.O., July 1992).

Click on thumbnail to enlarge the image.
Figure 3: View of platform III, levels 95 to 120, of the main section of the geological site at Tercis (southern Aquitaine). The Campanian-Maastrichtian boundary is shown (green). The flint-bearing strata display marls with low clay content near levels 97 and 118 (picture G.S.O., July 1992).
Click on thumbnail to enlarge the image.
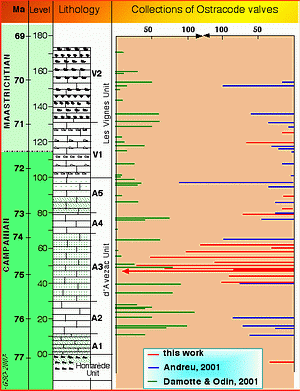
Figure 4: Ostracodes collected from the Tercis section. Previous collections by one of us (B.A.: blue, G.S.O.: green) were made from washed residues; the number of collected specimens was rarely more than 100 either single valves or complete shells (see scale to the top) per level. The new methods of collection used in this study (red bars, G.S.O.) were successful between levels 40 and 60.
Click on thumbnail to enlarge the image.
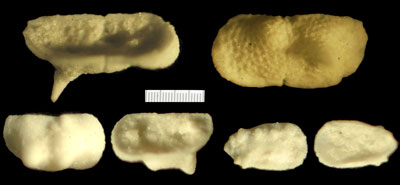
Figure 5:
Optical microscope views of valves obtained using acetolysis. Top: internal ventral view and external profile of a left valve of Bythoceratina sp. 1
(Pl. 3 ![]() ,
figs. 33-34); bottom left: external profile and internal ventral views of
a right valve of a representative of the genus Bythoceratina collected
from level 57.9; bottom right: Eucytherura (Vesticytherura) dorsotuberculata: external and internal profile views of a right valve from level 57.9.
,
figs. 33-34); bottom left: external profile and internal ventral views of
a right valve of a representative of the genus Bythoceratina collected
from level 57.9; bottom right: Eucytherura (Vesticytherura) dorsotuberculata: external and internal profile views of a right valve from level 57.9.
Click on thumbnail to enlarge the image.
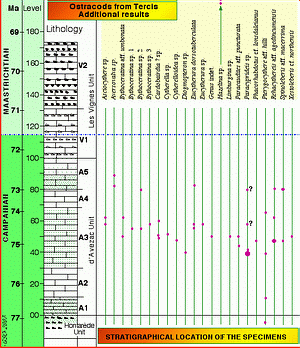
Figure 6: Location of the specimens documented in this paper. Species Genus indet. sp. (synonym of Mauritsina sp.) was documented in (2001); Rehacythereis aff. agedincumensis is a synonym of Spinoleberis sp. 1 documented by (2001); other taxa are new at the Tercis geological site.
Click on thumbnail to enlarge the image.
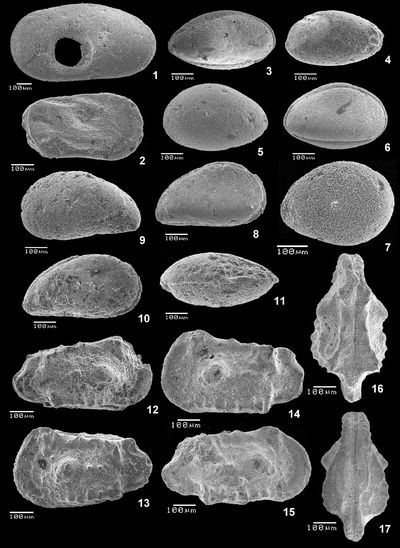
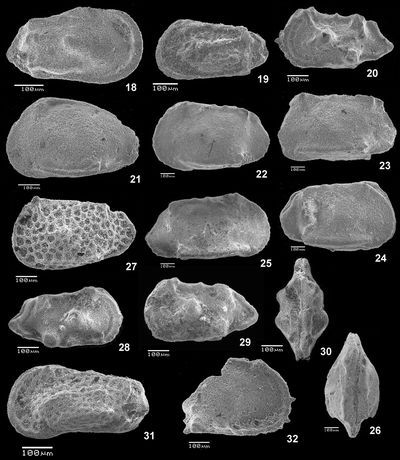
Plate 1: Fig. 1 Trace of gastropod predation on a LV of Cytherella sp., lv (level 52.2). Fig. 2 Cytherelloidea sp., C, Rlv (level 47.2). Figs. 3-4 Cardobairdia ? sp.; 3 C, Rlv (level 49.0); 4 C, Llv (level 50.05 to 50.25). Figs. 5-6 Asciocythere sp.; 5 C, Llv (level 57.9); 6 C, Rlv (level 62.0-62.5). Fig. 7 Xestoleberis cf. northensis , 1966; RV (level 50.05-50.25). Figs. 8-11 Paracyprideis sp.; 8 C, Rlv; 9 C, Llv; 10 C, Rlv; 11 C, dv (level 39.5). Figs. 12-17 Rehacythereis aff. agedincumensis , 1964, 12 C, Rlv; 13 C, Llv (level 79.8); 14 LV, lv; 15 RV, lv; 16 C, dv; 17 C, vv (level 47.2).
Abbreviations: C = carapace, RV = right valve, LV = left valve, R = Right, L = Left, lv = lateral view, dv = dorsal view, vv = ventral view, ind. = individual.
Click on thumbnail to enlarge the image.
Plate 2: Fig. 18 Phacorhabdotus cf. lonsdaleianus (, 1849); C, Rlv (level 47.2). Fig. 19 Hazelina sp., C, Llv (level B-29.5). Figs. 20 & 28-30 Spinoleberis aff. macerrima (, 1936); 20 C, Llv (level 47.2); 28 C, Rlv (level 79.8); 29 C, Llv (level 79.8); 30 C, dv (level 79.8). Fig. 21 Pterygocythere aff. hilli , 1957, sensu (1983), C, Llv (level -5.0). Figs. 22-26 Genus indet. sp.; 22 C, Llv; 23 C, Llv; 24 C, Rlv; 25 RV, lv; 26 C, dv (level 50.05-50.25). Fig. 27 Limburgina sp., C, Llv (level 50.05-50.2). Fig. 31 Paracaudites (Dumontina) aff. puncturata (, 1854) in (1966), C, Llv (level 45.3-43.0). Fig. 32 Diogmopteron sp., portion of RV, lv (level 39.5-41.1).
Abbreviations: C = carapace, RV = right valve, LV = left valve, R = Right, L = Left, lv = lateral view, dv = dorsal view, vv = ventral view, ind. = individual.

Click on thumbnail to enlarge the image.
Plate 3: Figs. 33-34 Bythoceratina sp. 1, 2 portions of LV; 33 anterior part, 34 posterior part, lv (level 54.9). Fig. 35 Eucytherura sp., C, Llv (level 57.9). Fig. 36 Bythoceratina cf. umbonata umbonata (, 1848) sensu (1983), C, LV, lv (level 62-62.5). Fig. 37 Bythoceratina sp. 3, portion of RV, lv (level 57.9). Figs. 38-39 Eucytherura (Vesticytherura) dorsotuberculata (, 1936); 38 C, Rlv (level 54.9); 39 C, Rlv (level 57.9). Figs. 40-41 Bythoceratina sp. 2; 40 LV, lv; 41 LV, dv (level 79.8). Figs. 42-43 Aversovalva sp.; 42 C, Rlv (according to B.A.) or Llv (according to G.S.O.); 43 C, vv (anterior side to the left) of the same individual (level 89.0).
Abbreviations: C = carapace, RV = right valve, LV = left valve, R = Right, L = Left, lv = lateral view, dv = dorsal view, vv = ventral view, ind. = individual.
Asciocythere sp., Pl. 1 ![]() , figs. 5-6; level 57.9 (1 ind.);
62.0-62.5 (1 ind.)
, figs. 5-6; level 57.9 (1 ind.);
62.0-62.5 (1 ind.)
Aversovalva sp., Pl. 3 ![]() , figs. 42-43; level 89.0 (1 ind.); 82.0 (1 ind.)
, figs. 42-43; level 89.0 (1 ind.); 82.0 (1 ind.)
Bythoceratina aff. umbonata umbonata (, 1848) sensu
(1983), Pl. 3 ![]() ,
fig. 36; level 62.0-62.5 (1 ind.); 50.05 (1 ind.)
,
fig. 36; level 62.0-62.5 (1 ind.); 50.05 (1 ind.)
Bythoceratina sp. 1, Pl. 3 ![]() ,
figs. 33-34; level 54.9 (2 ind.)
,
figs. 33-34; level 54.9 (2 ind.)
Bythoceratina sp. 2, Pl. 3 ![]() ,
figs. 40-41; level 79.8 (1 ind.); 54.9 (1 ind.)
,
figs. 40-41; level 79.8 (1 ind.); 54.9 (1 ind.)
Bythoceratina sp. 3, Pl. 3 ![]() ,
fig. 37; level 57.9 (1 ind.)
,
fig. 37; level 57.9 (1 ind.)
Cardobairdia ? sp., Pl. 1 ![]() ,
figs. 3-4; level 60.5 (1 ind.); 49.0 (2 ind.); 50.05-50.25 (2 ind.); note that, according to J.-P.
(personal communication), figs. 3-4 would better illustrate a male of Asciocythere.
,
figs. 3-4; level 60.5 (1 ind.); 49.0 (2 ind.); 50.05-50.25 (2 ind.); note that, according to J.-P.
(personal communication), figs. 3-4 would better illustrate a male of Asciocythere.
Cytherella sp., Pl. 1 ![]() ,
fig. 1; level 52.2 (single perforated individual)
,
fig. 1; level 52.2 (single perforated individual)
Cytherelloidea sp., Pl. 1 ![]() ,
fig. 2; level 47.2 (1 ind.)
,
fig. 2; level 47.2 (1 ind.)
Diogmopteron sp., Pl. 2 ![]() , fig. 32; level 39.5-41.1 (1 ind.)
, fig. 32; level 39.5-41.1 (1 ind.)
Eucytherura (Vesticytherura) dorsotuberculata (, 1936);
Pl. 3 ![]() ,
figs. 38-39; level 54.9 (1 ind.); 57.9 (1 ind.); 82.0 (1 ind.)
,
figs. 38-39; level 54.9 (1 ind.); 57.9 (1 ind.); 82.0 (1 ind.)
Eucytherura sp., Pl. 3 ![]() ,
fig. 35; level 57.9 (2 ind.)
,
fig. 35; level 57.9 (2 ind.)
Genus indet. sp., Pl. 2 ![]() , figs. 22-26; level 50.05-50.25 (8 ind.), synonym of Mauritsina sp. in
(2001)
, figs. 22-26; level 50.05-50.25 (8 ind.), synonym of Mauritsina sp. in
(2001)
Hazelina sp., Pl. 2 ![]() , fig. 19; level B-29.5 (1 ind.)
, fig. 19; level B-29.5 (1 ind.)
Limburgina sp., Pl. 2 ![]() , fig. 27; level 50.05-50.25 (1 ind.)
, fig. 27; level 50.05-50.25 (1 ind.)
Paracaudites (Dumontina) aff. puncturata (, 1854) in
(1966), Pl. 2 ![]() , fig. 31; level 43.0-45.3 (1 ind.); 50.05-50.25 (1 ind.)
, fig. 31; level 43.0-45.3 (1 ind.); 50.05-50.25 (1 ind.)
Paracyprideis sp., Pl. 1 ![]() ,
figs. 8-11; level 39.5 (4 ind.); 41.1 (2
ind.); 79.8 (1 ind.); 57.9 (1 ind. ?)
,
figs. 8-11; level 39.5 (4 ind.); 41.1 (2
ind.); 79.8 (1 ind.); 57.9 (1 ind. ?)
Phacorhabdotus cf. lonsdaleianus (, 1849),
Pl. 2 ![]() , fig. 18; level 47.2 (1 ind.)
, fig. 18; level 47.2 (1 ind.)
Pterygocythere aff. hilli , 1957, sensu
(1983), Pl. 2 ![]() , fig. 21; level -5.0 (1 ind.)
, fig. 21; level -5.0 (1 ind.)
Rehacythereis aff. agedincumensis ,
1964, synonym of Oertliella cf. pulchra in & ,
2001, and of Spinoleberis sp. 1 in
(2001), Pl. 1 ![]() ,
figs. 12-17; level 62.0-62.5 (2 ind.); 47.2 (6 ind.); 79.8 (2 ind.)
,
figs. 12-17; level 62.0-62.5 (2 ind.); 47.2 (6 ind.); 79.8 (2 ind.)
Spinoleberis aff. macerrima (, 1936) in
(1966), Pl. 2 ![]() , figs. 20 & 28-30; level 47.2 (1 ind.); 79.8 (3 ind.)
, figs. 20 & 28-30; level 47.2 (1 ind.); 79.8 (3 ind.)
Xestoleberis cf. northensis ,
1966, Pl. 1 ![]() ,
fig. 7; level 50.05-50.25 (1 ind.)
,
fig. 7; level 50.05-50.25 (1 ind.)
The morphology of the shells referred to the genus Aversovalva in this work is difficult to understand from a single picture. Orientation of the shell itself has been interpreted diversely for the two pictures obtained using the Scanning Electron Microscope (see views 42 and 43 above). In this situation, we have tried to obtain a model showing all aspects of another reasonably well preserved individual. This specimen comes from the acetolysis residue of a sample collected from level 82.0 in the type section of the geological site at Tercis. Figure 1 ![]() in this appendix gives 6 views of this specimen obtained using the optical microscope.
in this appendix gives 6 views of this specimen obtained using the optical microscope.
The model in video 1 was derived from both microscopic observation and pictures in the literature like the one given for Cytheropteron guadalupense , a Recent species illustrated in:
http://www.ucl.ac.uk/GeolSci/micropal/ostracod.html (incorrectly written C. gadaloupense) which shows a number of similar features both morphological and ornamentational. Using this model, we hope that it will be possible to recognise the taxon when it was observed elsewhere and to compare it with other species of the same genus in order to obtain a specific name (not yet suggested).
Compared to the taxon previously called Aversovalva sp. ( & , 2001), and here called Pterygocythere aff. hilli, the most obvious difference is the size: Pterygocythere is 600-700 µm long while Aversovalva sp. of Tercis is only half that size according to the two specimens at hand. At Tercis, Pterygocythere sp. was known from level 39.5 in 2001; it has been found in the acetolytic residues at the same level, at level 11.0 and down to level -5.0 (1 to 2 specimens at each level) and probably at level 65.4 (single valve) too.
N.B.: in a former work ( & , 2001), the specimen of Pterygocythere pictured is incorrectly titled: once as Aversovalva sp. (view 16) and once as Planileberis (view 20 of the same specimen) in the caption of the plate (p. 399).

Click on thumbnail to enlarge the image.
Figure 1: Optical microscope pictures of an ostracode collected from level 82.0 at Tercis. From left to right: lateral right, slightly ventral view; dorsal view, anterior part to the right; anterior ventral view (note the two ridges on the right valve to the left); lateral view of left valve, anterior part to the left; anterior dorsal view, anterior part to the bottom; ventral view, anterior part to the top (note the two ridges on the right valve to the left).
Click on image to play the video file (flv = 1,373 KB)
or click on the icon ![]() to download another version of video file (avi = 2,072 KB)
to download another version of video file (avi = 2,072 KB)
or click on the icon ![]() to open and play another version of video file (mpg = 1,294 KB)
to open and play another version of video file (mpg = 1,294 KB)
or click on the icon ![]() to download another version of video file (wmv = 3,130 KB)
to download another version of video file (wmv = 3,130 KB)
Video: Tridimensional panoramic views of a reconstruction of an individual of the genus Aversovalva. Note that the right valve is smaller than the left one. 3D model realised using software Blender (http://www.blender.org/) a facility already used for another microfauna ( & , 2006).