![]()
Sommaire
[1. Introduction] [2. Cadre de
l'étude]
[3. Résultats]
[4. Variété, répartition ...]
[5. Conclusion]
[Références bibliographiques]
[Figures] [Planches] [Annexe
1] [Annexe 2] et ... [Animation]
![]()
![]()
Une nouvelle étude de la microfaune, obtenue par acétolyse des niveaux indurés dans le Campanien-Maastrichtien de la carrière de Tercis, a permis de récolter 22 espèces additionnelles au regard de l'étude publiée en 2001 et de porter ainsi à 75 espèces, réparties sur 34 genres reconnus et 4 indéterminés, la biodiversité en ostracodes de ce site. Des espèces de petite taille, généralement rares, rapportées aux genres Aversovalva, Bythoceratina et Eucytherura ont par ailleurs été recueillies. Les associations de genres et d'espèces d'ostracodes sont caractéristiques du Campanien-Maastrichtien, dans un environnement de plate-forme carbonatée ouverte.
Ostracodes ; Campanien ; Maastrichtien ; stratotype ; Tercis ; Aquitaine ; France ; acétolyse.
B. & G.S. (2008).- Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : compléments sur les ostracodes extraits par acétolyse.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/02-fr (CG2008_A02 (fr))
New micropalaeontological studies on the type section of the Campanian-Maastrichtian at Tercis (SW France): new ostracodes obtained using acetolysis.- Through the use of acetolysis new micropalaeontological studies on the type section of the Campanian-Maastrichtian boundary at Tercis (SW France) we obtained additional information on ostracodes. Acetolysis on hard carbonates levels of the Tercis quarry found 22 species in addition to those of the studies published in 2001. Today, 75 species are recognized: they represent 34 known genera, and 4 currently unidentified. Small species usually rare of the genera Aversovalva, Bythoceratina and Eucytherura were collected. All of the assemblages (of both genera and species) are characteristic of a Campanian-Maastrichtian open carbonate platform environment.
Ostracodes; Campanian; Maastrichtian; stratotype; Tercis; Aquitaine; France; acetolysis.
La coupe stratotypique de la limite Campanien-Maastrichtien à Tercis (Landes, France) a été désignée en 2001 ( & ,
2001) à la suite d'une étude coopérative détaillée (,
2001a) comportant des observations sur 18 groupes fossiles illustrés dans l'ouvrage cité comme postérieurement ( et alii,
2006). Cette
coupe (Fig. 1 ![]() ), déjà bien étudiée, était toute désignée pour servir de support à des compléments d'étude réalisés pour mieux établir et mettre en valeur la paléobiodiversité crétacée à un moment situé juste avant la crise de la limite Crétacé-Paléogène, cet événement global qui a marqué la fin de l'Ère secondaire et d'un équilibre biologique dont la pyramide était sommée par les dinosaures.
), déjà bien étudiée, était toute désignée pour servir de support à des compléments d'étude réalisés pour mieux établir et mettre en valeur la paléobiodiversité crétacée à un moment situé juste avant la crise de la limite Crétacé-Paléogène, cet événement global qui a marqué la fin de l'Ère secondaire et d'un équilibre biologique dont la pyramide était sommée par les dinosaures.
Parmi les possibilités susceptibles d'améliorer nos connaissances, figurait l'étude des niveaux indurés qui constituent une portion majeure de la
coupe de Tercis
(Figs. 2 ![]() - 3
- 3 ![]() ). En effet, en 2001, l'étude de la microfaune, à l'exception notable d'une intense étude des foraminifères planctoniques en lames minces, s'était restreinte aux niveaux dissociables aisément par lavage c'est à dire les plus riches en argile. À
Tercis, les teneurs en argiles de ces niveaux dépassaient rarement 15 % (,
2001c). Cependant, sur le plan environnemental, ces niveaux traduisent un apport détritique qui influe notoirement sur le développement de la vie des micro-organismes. La sélection pratique des niveaux les plus argileux risquait donc de nous avoir fait manquer les organismes ne supportant que des eaux claires. C'est, entre autres, dans cette optique que des essais de dissociation par acétolyse (immersion du sédiment dans l'acide acétique pur ou concentré à chaud ou à froid) ont été réalisés ; ils ont abouti de façon satisfaisante quant à la dissociation elle-même de la plupart des niveaux soumis à l'expérimentation. L'examen et la séparation de divers groupes fossiles ont été entrepris ensuite dans ces sédiments dissociés, en particulier pour les formes qui n'avaient pas encore été identifiées dans les travaux précédents ou pour des niveaux qui avaient été moins bien caractérisés.
). En effet, en 2001, l'étude de la microfaune, à l'exception notable d'une intense étude des foraminifères planctoniques en lames minces, s'était restreinte aux niveaux dissociables aisément par lavage c'est à dire les plus riches en argile. À
Tercis, les teneurs en argiles de ces niveaux dépassaient rarement 15 % (,
2001c). Cependant, sur le plan environnemental, ces niveaux traduisent un apport détritique qui influe notoirement sur le développement de la vie des micro-organismes. La sélection pratique des niveaux les plus argileux risquait donc de nous avoir fait manquer les organismes ne supportant que des eaux claires. C'est, entre autres, dans cette optique que des essais de dissociation par acétolyse (immersion du sédiment dans l'acide acétique pur ou concentré à chaud ou à froid) ont été réalisés ; ils ont abouti de façon satisfaisante quant à la dissociation elle-même de la plupart des niveaux soumis à l'expérimentation. L'examen et la séparation de divers groupes fossiles ont été entrepris ensuite dans ces sédiments dissociés, en particulier pour les formes qui n'avaient pas encore été identifiées dans les travaux précédents ou pour des niveaux qui avaient été moins bien caractérisés.
C'est lors de ces tris qu'ont été découvertes une cinquantaine de formes de microproblematica ( & , 2006 ; , 2007 ; et alii, 2007) ainsi que des restes encore non répertoriés sur le site de Tercis appartenant à divers groupes connus comme les bryozoaires, les holothuries, les crinoïdes, les astérides, les ophiures, les foraminifères, ... Le présent travail rapporte les informations recueillies à propos des ostracodes, microfossiles pour lesquels des espèces non déterminées dans les travaux précédents ont été récoltées tandis que la répartition d'espèces déjà connues a été précisée et qu'une illustration nouvelle a pu être constituée.
Le stratotype de la limite Campanien-Maastrichtien a été défini dans une carrière, dont l'exploitation a cessé en 1997, située sur le territoire de la commune de Tercis, près de Dax, département des Landes, en Aquitaine méridionale, à une dizaine de km au nord des premiers charriages nord-pyrénéens (, 2001). Une coupe continue de près de 200 m a été décrite en détail ( & , 1994). Elle représente à peu près 8 Ma de dépôts entourant la limite Campanien-Maastrichtien définie par convention à la cote 115,2 à la suite d'une approche originale : cette définition implique la combinaison de 12 signaux biostratigraphiques pris dans 6 groupes fossiles et entourant un événement guide accepté par convention (, 2001b ; & , 2001).
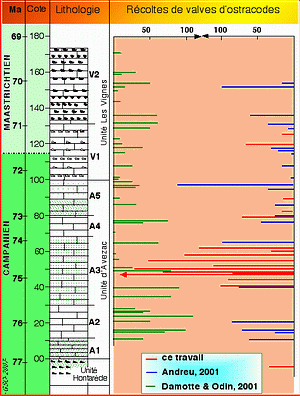
La lithologie de la coupe est schématisée dans la Figure 4 ![]() qui identifie 3 unités. L'Unité Hontarède est constituée de calcaires à silex glauconieux au sommet. L'Unité Avezac est une succession homogène de calcaires dont les 5 sous unités (A1 à A5) ne se distinguent que par des teneurs en glauconie alternativement plus élevées et quasi nulles. L'Unité Les Vignes comprend une sous unité à silex clairs (V1) à la base et une sous unité à silex foncés (V2) au sommet. La colonne lithologique note la présence de glauconie (pointillés) et celle de silex presque toujours en niveaux décimétriques bien séparés avec une périodicité métrique.
qui identifie 3 unités. L'Unité Hontarède est constituée de calcaires à silex glauconieux au sommet. L'Unité Avezac est une succession homogène de calcaires dont les 5 sous unités (A1 à A5) ne se distinguent que par des teneurs en glauconie alternativement plus élevées et quasi nulles. L'Unité Les Vignes comprend une sous unité à silex clairs (V1) à la base et une sous unité à silex foncés (V2) au sommet. La colonne lithologique note la présence de glauconie (pointillés) et celle de silex presque toujours en niveaux décimétriques bien séparés avec une périodicité métrique.
La stratigraphie en âges numériques est proposée d'après des corrélations magnéto.- et biostratigraphiques avec des niveaux de bentonites datés aux États-Unis ( & , 2001), une vitesse de sédimentation constante illustrée par une périodicité orbitale des dépôts, constance tempérée par l'examen (présence et cristallinité) des minéraux glauconitiques ( & , 2001). La correction de l'âge de la limite proposée en 2001 a été améliorée grâce aux corrélations Europe-Amérique établies à l'aide des inocérames ( & , 2003). Ces âges s'entendent, pour une limite située à 71,6 Ma, avec une incertitude de ±0,6 Ma (intervalle de confiance 95 %).
La Figure 4 ![]() regroupe les résultats quantitatifs obtenus en 2001 ( & ,
2001 ; , 2001). Au cours de ces deux études, 1800 spécimens (pour environ 45 niveaux) et 2500 spécimens (pour 28 niveaux) avaient été récoltés
respectivement ; 36 et 61 valves avaient été illustrées respectivement. Dans le rapport de synthèse ( & ,
2001), 53 espèces étaient ainsi identifiées, appartenant à 28 genres connus et 3 non déterminés. Les genres cités étaient tous connus dans le Crétacé
Supérieur d'Europe (Europe du Nord-Ouest d'abord puisqu'il n'y avait guère d'études sur l'Aquitaine à l'époque) et, secondairement, d'Afrique du Nord. Une dizaine d'espèces étaient signalées en Europe du Nord-Ouest ou autres régions d'Europe et secondairement en Amérique du Nord et sur la marge méditerranéenne méridionale. Ceci était considéré comme dénotant une absence d'endémisme dans le diverticule océanique nord pyrénéen ouvert sur l'Atlantique où se situait Tercis au Crétacé
Supérieur.
regroupe les résultats quantitatifs obtenus en 2001 ( & ,
2001 ; , 2001). Au cours de ces deux études, 1800 spécimens (pour environ 45 niveaux) et 2500 spécimens (pour 28 niveaux) avaient été récoltés
respectivement ; 36 et 61 valves avaient été illustrées respectivement. Dans le rapport de synthèse ( & ,
2001), 53 espèces étaient ainsi identifiées, appartenant à 28 genres connus et 3 non déterminés. Les genres cités étaient tous connus dans le Crétacé
Supérieur d'Europe (Europe du Nord-Ouest d'abord puisqu'il n'y avait guère d'études sur l'Aquitaine à l'époque) et, secondairement, d'Afrique du Nord. Une dizaine d'espèces étaient signalées en Europe du Nord-Ouest ou autres régions d'Europe et secondairement en Amérique du Nord et sur la marge méditerranéenne méridionale. Ceci était considéré comme dénotant une absence d'endémisme dans le diverticule océanique nord pyrénéen ouvert sur l'Atlantique où se situait Tercis au Crétacé
Supérieur.
Les essais de dissociation se sont d'abord concentrés sur la sous-unité A3 de l'Unité Avezac qui avait livré des indices de la zone d'existence à Radotruncana calcarata (, 1927), repère biostratigraphique classique et précis du Campanien supérieur pour les foraminifères planctoniques. Pour des quantités de sédiment traitées de l'ordre de 100 à 200 g, environ 200 spécimens ou plus ont été récoltés à divers niveaux
(Fig. 4 ![]() ). Les essais entrepris pour les niveaux inférieurs, notamment entre les cotes -5 et +25, ne livrant pas plus de faune que les lavages antérieurs, ont été peu triés. De même, entre les cotes 95 et 170, la faune étant pauvre et les restes en assez mauvais état, les tris ont été limités à quelques niveaux.
). Les essais entrepris pour les niveaux inférieurs, notamment entre les cotes -5 et +25, ne livrant pas plus de faune que les lavages antérieurs, ont été peu triés. De même, entre les cotes 95 et 170, la faune étant pauvre et les restes en assez mauvais état, les tris ont été limités à quelques niveaux.
Les expérimentations de dissociation ont permis de recueillir environ 1700 valves pour la quinzaine de niveaux qui se sont révélés raisonnablement productifs, niveaux dont les faunes ont été prises en compte pour l'étude de reconnaissance et la nouvelle illustration. La
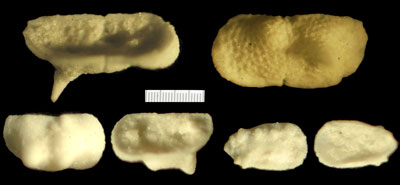
Figure 5 ![]() illustre, en microscopie optique, quelques exemplaires intéressants d'ostracodes ayant attiré notre attention sur des taxons non encore cités à Tercis.
illustre, en microscopie optique, quelques exemplaires intéressants d'ostracodes ayant attiré notre attention sur des taxons non encore cités à Tercis.
Les carapaces et les valves d'ostracodes récoltées sont très fragilisées par l'acétolyse et supportent difficilement les manipulations. Leur surface a souvent été partiellement dissoute et à l'origine malheureusement de la disparition d'une partie de l'ornementation. On notera toutefois que les spécimens obtenus par lavage sur niveaux peu indurés lors des études précédentes n'étaient généralement pas très bien préservés non plus.
Dans les études publiées en 2001 ( & , 2001 ; , 2001), les diagnoses des genres, tels qu'ils étaient conçus dans le travail, étaient données. Cette présentation avait paru indispensable puisque les différents auteurs ne considèrent pas toujours les mêmes critères. Ainsi les noms utilisés trouvaient leur justification même si une autre conception pouvait être adoptée. Des noms sont proposés ici, il est raisonnable de dire sur quels critères ils reposent. Dans cette optique, la diagnose adoptée ici pour les genres est donnée ci-dessous.
Ces déterminations génériques reposent, sur les caractéristiques morphologiques externes de la carapace, les caractères internes n'étant pas accessibles. Les genres décrits en 2001 ne sont pas repris ; en revanche, les espèces additionnelles, nouvelles par rapport au travail de 2001, sont figurées (voir Annexe 1) et l'illustration de deux espèces est complétée.
- Asciocythere , 1952 : carapace de taille moyenne, ovale à triangulaire avec angles généralement non à peu apparents en vue latérale externe ; extrémités arrondies, tournées plutôt vers le bas ; recouvrement de la valve gauche sur la périphérie de la droite ; carapace lisse à ponctuée.
- Cardobairdia , 1960 : carapace de taille petite à moyenne, généralement épaisse, sub-elliptique et lisse ; extrémité antérieure régulièrement arrondie et tournée vers l'avant ; extrémité postérieure plutôt acuminée ; la valve gauche recouvre la valve droite sur la périphérie et plus particulièrement sur les bords dorsal et ventral.
- Paracyprideis , 1929 : carapace de taille moyenne, sub-elliptique à triangulaire en vue latérale externe ; extrémité antérieure régulièrement arrondie, plutôt tournée vers le bas, extrémité postérieure triangulaire et proche du bord ventral ; les extrémités portent parfois des denticules ou des épines ; hauteur maximale en avant de la mi-longueur ; surface généralement lisse excepté lorsqu'elle est percée de larges pores.
- Eucytherura , 1894 : carapace de petite taille, sub-losangique à carrée en vue latérale externe ; bord dorsal rectiligne, bord ventral arrondi ; extrémité antérieure largement arrondie, extrémité postérieure triangulaire, à processus caudal tournée vers le bord dorsal ; bouton oculaire présent dans l'angle antéro-dorsal ; carapace tuberculée ou réticulée.
- Pterygocythere , 1954 : carapace triangulaire en vue latérale externe, enflée ventralement et possédant une expansion alaire pointue à l'arrière, dans le prolongement du bourrelet antérieur, parfois réduite à une rangée d'épines ; bord postérieur acuminé ; surface des flancs lisse pouvant supporter quelques épines ou tubercules ; présence d'une côte sur le bord dorsal ; bouton oculaire apparent ; extrémités antérieure et postérieure parfois épineuses ; face ventrale renflée, portant des côtes longitudinales ; ornementation généralement simple ; le matériel constituant la carapace est épais et cette dernière est robuste.
- Diogmopteron , 1954 : les genres Pterygocythere et Diogmopteron présentent les mêmes caractères morphologiques externes ; ils se distinguent par des différences dans la charnière (non apparente dans ce travail) ; cependant, extérieurement, ces différences se manifestent ; chez Diogmopteron, la valve droite dépasse la gauche sur le bord dorsal, ce caractère étant inversé chez Pterygocythere (ce caractère n'apparaît pas sur notre individu).
- Rehacythereis , 1973 : carapace triangulaire à rectangulaire en vue latérale externe ; extrémité antérieure régulièrement arrondie, postérieure triangulaire ; bourrelet antérieur se poursuivant par une côte ventrale terminée par un tubercule postéro-ventral ; côte dorsale indépendante et terminée par un tubercule postéro-dorsal ; bouton oculaire net, suivi par une fine côte dirigée vers le tubercule musculaire subcentral ; ce dernier est net ; réticulation sur la surface des valves ; recouvrement de la valve gauche sur la valve droite particulièrement dans l'angle antéro-dorsal.
- Genre indét. (Pl. 2 ![]() ,
figs. 22-26) : carapace de grande taille,
sub-rectangulaire ; lobe frontal prononcé ; bouton oculaire dans l'angle antéro-dorsal ; extrémité antérieure régulièrement arrondie dépassant le bord
ventral ; extrémité postérieure triangulaire à bord dorsal concave, bord ventral convexe et plus ou moins épineux ; côtes dorsale et ventrale nettes et indépendantes ; tubercule musculaire subcentral à peine esquissé ; flancs bombés ; recouvrement de la valve gauche sur la droite au niveau des angles antéro- et postéro-dorsal ; surface lisse portant parfois quelques granules de petite taille.
,
figs. 22-26) : carapace de grande taille,
sub-rectangulaire ; lobe frontal prononcé ; bouton oculaire dans l'angle antéro-dorsal ; extrémité antérieure régulièrement arrondie dépassant le bord
ventral ; extrémité postérieure triangulaire à bord dorsal concave, bord ventral convexe et plus ou moins épineux ; côtes dorsale et ventrale nettes et indépendantes ; tubercule musculaire subcentral à peine esquissé ; flancs bombés ; recouvrement de la valve gauche sur la droite au niveau des angles antéro- et postéro-dorsal ; surface lisse portant parfois quelques granules de petite taille.
La liste des espèces additionnelles, associées à 2 espèces déjà figurées en 2001, est donnée en
Annexe avec les références des illustrations, les échantillons (éch.) dans lesquels elles ont été reconnues et le nombre d'individus (ind.) récoltés dans ces échantillons. La
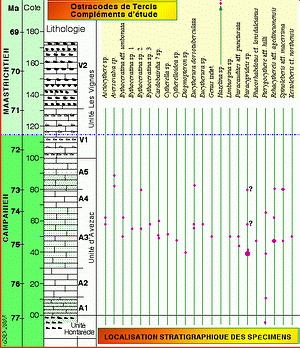
Figure 6 ![]() illustre la provenance stratigraphique des spécimens des taxons discutés dans ce travail ; ces nouveaux taxons ont tous été observés dans la portion campanienne de la
coupe.
illustre la provenance stratigraphique des spécimens des taxons discutés dans ce travail ; ces nouveaux taxons ont tous été observés dans la portion campanienne de la
coupe.
Une première étude (, 2001 ; & , 2001 ; & , 2001) avait permis de reconnaître 53 espèces d'ostracodes. Nous complétons ce travail avec 22 espèces additionnelles. Apparaissent donc au total, dans le Campanien-Maastrichtien de Tercis, 75 espèces qui se répartissent dans 34 genres reconnus, dont 7 additionnels décrits dans ce travail et 4 indéterminés.
La dissociation des bancs calcaires par acétolyse a permis de mettre à jour notamment des espèces appartenant aux genres Aversovalva, Bythoceratina et Eucytherura, espèces de petite taille et relativement rares qui peuvent passer souvent inaperçues dans les lavages.
Dans ce travail, nous avons identifié 8 espèces nommées parmi lesquelles 3 conférées et 4 affines : Eucytherura (Vesticytherura) dorsotuberculata (, 1936), Phacorhabdotus cf. lonsdaleianus lonsdaleianus (, 1849) et Xestoleberis cf. northensis , 1966, Bythoceratina cf. umbonata umbonata (, 1848), Paracaudites (Dumontina) aff. puncturata (, 1854), Pterygocythere aff. hilli , 1957, Rehacythereis aff. agedincumensis (, 1964) et Spinoleberis aff. macerrima (, 1936).
La localisation stratigraphique des espèces examinées dans le présent travail est donnée dans la
Figure 6 ![]() . La
localisation connue auparavant est résumée ci-après. Eucytherura (Vesticytherura) dorsotuberculata est connue dans le Campanien supérieur de Dordogne, France, le Campanien supérieur-Paléocène-Éocène ? de Pologne, le Campanien supérieur-Maastrichtien d'Allemagne, le Maastrichtien inférieur de Belgique et des Pays-Bas, le Maastrichtien du Danemark (bibliographie complète
dans , 1973, et ,
1983). Phacorhabdotus lonsdaleianus lonsdaleianus est décrite dans le Campanien supérieur-Maastrichtien d'Angleterre ; elle est reconnue dans le Campanien supérieur du Bassin de Paris, France, le Campanien-Maastrichtien d'Allemagne, le Maastrichtien inférieur de Pologne, le Maastrichtien de Belgique, des Pays-Bas et du Danemark (bibliographie complète
dans , 1983). Xestoleberis northensis apparaît dans le Campanien supérieur-Maastrichtien de Belgique et des Pays-Bas, le Campanien moyen de Dordogne et supérieur du Bassin de Paris et des Charentes, France (bibliographie complète
dans , 1973). Bythoceratina umbonata umbonata
- sensu (1983) - est une espèce du Campanien-Maastrichtien : Campanien supérieur de Belgique, Campanien-Maastrichtien de Pologne, Maastrichtien inférieur des Pays-Bas et du Danemark, Maastrichtien d'Allemagne, Crétacé
Supérieur d'Angleterre et d'Irlande (bibliographie complète
dans , 1973, et ,
1983). Paracaudites (Dumontina) puncturata est récoltée dans le Campanien de Dordogne et des Charentes, France (,
1973 ; et alii,
1985), et le Maastrichtien des Pays-Bas et de Belgique (,
1966). Pterygocythere hilli
- sensu (1983) -
se distribue du Santonien moyen au Campanien inférieur d'Allemagne. Elle est connue en outre dans
l'Éocène de Belgique et des Pays-Bas (,
1983). Rehacythereis agedincumensis est datée du Santonien-Campanien du Bassin de Paris (,
1964 ; et alii,
1985) ; elle a été retrouvée (elle-même ou une espèce proche) dans le Cénomanien supérieur-Turonien inférieur des Corbières, France ( & ,
2006). Spinoleberis macerrima (, 1936) est connue dans le Maastrichtien des Pays-Bas (,
1966) et dans le Campanien d'Aquitaine (Spinoleberis cf. macerrima
dans , 1973).
. La
localisation connue auparavant est résumée ci-après. Eucytherura (Vesticytherura) dorsotuberculata est connue dans le Campanien supérieur de Dordogne, France, le Campanien supérieur-Paléocène-Éocène ? de Pologne, le Campanien supérieur-Maastrichtien d'Allemagne, le Maastrichtien inférieur de Belgique et des Pays-Bas, le Maastrichtien du Danemark (bibliographie complète
dans , 1973, et ,
1983). Phacorhabdotus lonsdaleianus lonsdaleianus est décrite dans le Campanien supérieur-Maastrichtien d'Angleterre ; elle est reconnue dans le Campanien supérieur du Bassin de Paris, France, le Campanien-Maastrichtien d'Allemagne, le Maastrichtien inférieur de Pologne, le Maastrichtien de Belgique, des Pays-Bas et du Danemark (bibliographie complète
dans , 1983). Xestoleberis northensis apparaît dans le Campanien supérieur-Maastrichtien de Belgique et des Pays-Bas, le Campanien moyen de Dordogne et supérieur du Bassin de Paris et des Charentes, France (bibliographie complète
dans , 1973). Bythoceratina umbonata umbonata
- sensu (1983) - est une espèce du Campanien-Maastrichtien : Campanien supérieur de Belgique, Campanien-Maastrichtien de Pologne, Maastrichtien inférieur des Pays-Bas et du Danemark, Maastrichtien d'Allemagne, Crétacé
Supérieur d'Angleterre et d'Irlande (bibliographie complète
dans , 1973, et ,
1983). Paracaudites (Dumontina) puncturata est récoltée dans le Campanien de Dordogne et des Charentes, France (,
1973 ; et alii,
1985), et le Maastrichtien des Pays-Bas et de Belgique (,
1966). Pterygocythere hilli
- sensu (1983) -
se distribue du Santonien moyen au Campanien inférieur d'Allemagne. Elle est connue en outre dans
l'Éocène de Belgique et des Pays-Bas (,
1983). Rehacythereis agedincumensis est datée du Santonien-Campanien du Bassin de Paris (,
1964 ; et alii,
1985) ; elle a été retrouvée (elle-même ou une espèce proche) dans le Cénomanien supérieur-Turonien inférieur des Corbières, France ( & ,
2006). Spinoleberis macerrima (, 1936) est connue dans le Maastrichtien des Pays-Bas (,
1966) et dans le Campanien d'Aquitaine (Spinoleberis cf. macerrima
dans , 1973).
La présence de ces espèces additionnelles confirme la position stratigraphique des associations, caractéristiques du Campanien-Maastrichtien avec une absence d'endémisme pour ce diverticule de l'Océan Atlantique à Tercis. La région de Tercis appartient, au Campanien, à l'Unité Paléobiogéographique Européenne Boréale (Boreal European Palaeobiogeographical Unit), à eaux chaudes tempérées, définie par et alii (2004), par la présence des associations d'ostracodes et qui inclut notamment les pays suivants : Allemagne, Belgique-Pays Bas, Angleterre, Bassin de Paris, Bassin d'Aquitaine, Bassin Vasco-Cantabrique, Pologne et Russie. Il faut noter, néanmoins, que les genres Dumontina et Limburgina présents à Tercis sont typiques de la marge nord-téthysienne. Cette attribution, téthysienne ou boréale, dépend donc des critères retenus ; ainsi, & (in et alii, 2001) font-ils remonter jusqu'en Charente le domaine téthysien caractérisé par les grands foraminifères benthiques, les rudistes, l'absence de bélemnites et du véritable faciès craie, ces deux derniers points étant clairs à Tercis.
L'environnement, caractérisé à Tercis par les associations des genres Dumontina, Limburgina, Mauritsina, Spinoleberis, Pterygocythere, Diogmopteron, Phacorhabdotus, Imhotepia, Cytherelloidea, Bythoceratina, Cardobairdia, Krithe et Eucytherura, genres reconnus dans & (2001), (2001) et & (2001), est typique de l'étage circalittoral d'une plate-forme carbonatée externe, à tranche d'eau relativement importante, en zone photique. Il est intermédiaire entre les deux environnements proposés par et alii (2004) et distingués dans l'Unité Paléobiogéographique Européenne Boréale par des associations génériques caractéristiques, un environnement peu profond de haute énergie (avec les genres Dumontina, Kikliocythere, Limburgina, Mauritsina, Mosaeleberis et Spinoleberis) et un environnement profond de plus faible énergie (avec les genres Trachyleberidea, Pterygocythere, Alatacythere, Phacorhabdotus, Imhotepia, et des espèces de la famille des Bythocytheridae).
Partie d'une étude micropaléontologique complémentaire des niveaux indurés du stratotype de la limite Campanien-Maastrichtien à Tercis (Aquitaine méridionale), ce compte rendu sur les ostracodes rapporte la récolte d'espèces additionnelles et particulièrement d'espèces de petite taille, généralement rares dans les échantillons.
La paléobiodiversité augmente ; le nombre d'espèces d'ostracodes passe de 53, reconnues en 2001, à 75 au total, ce qui prouve l'intérêt de l'acétolyse sur les niveaux carbonatés indurés pour établir un bilan plus complet de la biodiversité de ce bassin et permet d'enrichir la connaissance de cette coupe de référence mondiale.
Les associations de genres et d'espèces d'ostracodes sont caractéristiques de l'intervalle Campanien-Maastrichtien, dans un environnement de plate-forme carbonatée ouverte.
L'un des auteurs (B.A.) tient à remercier T. , LMTG, Université Toulouse III, pour la réalisation des photographies au microscope électronique à balayage Jeol JSM-6360LV, J.-F. , Marseille, et J.-P. , Cestas, France, pour les améliorations proposées. Les vues 40 et 41, celles de la
Figure 5 ![]() , et l'illustration de
l'Annexe 2
, et l'illustration de
l'Annexe 2 ![]() ont été obtenues à l'Université Pierre-et-Marie Curie.
ont été obtenues à l'Université Pierre-et-Marie Curie.
En dernière minute, un nouvel individu de Eucytherura (Vesticytherura) dorsotuberculata a été récolté à la cote 120,4, c'est-à-dire à la base du Maastrichtien de Tercis.
Un nouvel individu de Aversovalva sp. a été récolté 9,6 m sous (moins de 0,4 Ma avant) la limite Crétacé-Paléogène à Tercis.
B. (2001).- Ostracodes du site de Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. C6b, p. 402-409.
B. & M. (2006).- Ostracodes du Cénomanien supérieur et du Turonien de la zone sous-pyrénéenne orientale (Corbières méridionales, SE France). Systématique, biostratigraphie et paléobiogéographie.- Revue de Micropaléontologie, Paris, vol. 49, n° 2, p. 55-73.
B. & G.S. (2001).- Synthetic data on the Campanian-Maastrichtian ostracode fauna from Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. C6c, p. 410-413.
J.-F., J.-P. & R. (1985).- Crétacé supérieur. In : H.J., Atlas des Ostracodes de France.- Bulletin des Centres de Recherche Exploration-Production elf-Aquitaine, Pau, Mémoire 9, p. 211-255.
M., G.S. & B. (2001). Geology and Late Cretaceous palaeogeography of the geological site at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. A4, p. 47-59.
J.-P. (1973).- Étude stratigraphique et micropaléontologique du Crétacé supérieur de la région de Saint-Cyprien (Dordogne).- Thèse de Doctorat de 3° Cycle, Université Paris VI, 287 p.
B. (1983).- Die Cytheracea (Ostracoda) im chreibkreide-Richtprofil von Lägerdorf-Kronsmoor-Hemmoor (Coniac bis Maastricht; Norddeutschland).- Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, Heft 54, p. 65-168.
R. (1964).- Contribution à l'étude des ostracodes du Sénonien de Sens (Yonne) : les Trachyleberididae.- Revue de Micropaléontologie, Paris, vol. 7, n° 2, p. 100-110.
R. & G.S. (2001).- Ostracodes du site de Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. C6a, p. 396-401.
G. (1966).- Cytheracea (Ostracodes) du Maastrichtien de Maastricht (Pays-Bas) et des régions voisines ; résultats stratigraphiques et paléontologiques de leur étude.- Mededelingen van de geologische Stichting, Maastricht, (Serie C), V2, n° 2, 197 p.
Z. & G.S. (2001).- Magnetostratigraphy across the Campanian-Maastrichtian boundary at Tercis les Bains in comparison with northern Germany, the Apennines (Central Italy) and North America; biostratigraphical and geochronological constraints. In : G.S. (réd., 2001a), chap. B2d, p. 175-183.
G.S., rédacteur (2001a).- The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, 19, Elsevier Sciences Publ. Amsterdam, xxviii + 881 p.
G.S. (2001b).- The Campanian-Maastrichtian boundary: definition at Tercis (Landes, SW France) principle, procedure, and proposal. In : G.S. (réd., 2001a), chap. F3, p. 820-833.
G.S. (2001c).- Descriptive lithostratigraphy of the Campanian-Maastrichtian succession at Tercis les Bains (SW France). In : G.S. (réd., 2001a), chap. B1a, p. 85-109.
G.S. (2007).- Sur des microfossiles calcaires problématiques du stratotype de la limite Campanien-Maastrichtien de Tercis (Sud-Ouest, France) : les gilianelles.- Comptes rendus Palévol, Paris, vol. 6, n° 3, p. 181-188.
G.S., C., A. & B.
(2006).- Présentation du site géologique de Tercis (Landes, France) ; 13 affiches pédagogiques informatisées.
a- Le site de la Grande Carrière de Tercis ; haut lieu géologique, témoin des 10 Ma de la fin du Crétacé.
b- Les grands fossiles de la carrière de Tercis ; une richesse et une diversité remarquable en macrofossiles.
c- Les bivalves de Tercis ; des fossiles marqueurs de l'environnement et un groupe marqueur de temps (les inocérames).
d- Les ammonites de Tercis ; un groupe disparu depuis 65 Ma, proche des nautiles et des seiches, très utile pour dater les sédiments de plate-forme.
e- Les échinides de Tercis.
f- Les astérides de Tercis ; des fossiles communs sur la plate-forme continentale et utiles à la datation relative.
g- Les petits fossiles de la carrière de Tercis.
h- Les nannofossiles calcaires à Tercis.
i- Les kystes de dinoflagellés ; outil stratigraphique de datation relative excellemment illustré à Tercis.
j- Les foraminifères de la Grande Carrière de Tercis ; une présence suffisante pour une datation relative acceptable.
l- Datation comparée Continent-Océan à Tercis.
m- Les sites préhistoriques à Tercis ; une grande carrière sur le plus grand site de plein air du sud-ouest de la France.
n- Le site de Tercis, stratotype de référence mondial ; définition du calendrier géologique et application à la limite d'étage Campanien-Maastrichtien.
Disponible sur internet (depuis le 14-VI-06) : http://paleopolis.rediris.es/odin/
G.S., C., A. & B.
(2007).- Gilianelles et autres microfossiles problématiques : contribution à l'étude de la paléobiodiversité et à la biostratigraphie crétacée dans le stratotype de la limite Campanien-Maastrichtien à Tercis (Sud Aquitaine, France).- Journée Hommage à Gérard , Paris 06-XII-2007.
Disponible sur internet (depuis le 21-XI-07) : http://paleopolis.rediris.es/odin/images/poster-14.png
G.S. & A. (2001).- Interpretative reading of the Campanian-Maastrichtian deposits at Tercis les Bains: sedimentary breaks, rhythms, accumulation rate, sequences. In : G.S. (réd., 2001a), chap. B1c, p. 120-131.
G.S. & M. A. (2001).- The global Campanian-Maastrichtian stage boundary at Tercis les Bains, Landes, SW France.- Episodes, Beijing, vol. 24, n° 4, p. 229-238.
G.S. & A. (2006).- Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : les gilianelles (microproblematica) extraites par acétolyse.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2006/05 (CG2006_A05), 28 p., 3 vidéos.
G.S. & P. (1994).- Première description d'une coupe dans la localité aturienne de Tercis (Landes) : contribution à l'étude de la limite Campanien-Maastrichtien.- Géologie de la France, Orléans, n° 2 (1994), p. 31-37.
G.S. & I. (2003).- Sur les inocérames de Tercis (Landes, France) : le meilleur outil corrélatif entre Europe et Amérique du Nord autour de la limite Campanien-Maastrichtien / Inoceramids of the site of Tercis (Landes, France): the best correlative tool between Europe and North America across the Campanian-Maastrichtian boundary.- Comptes rendus Géosciences, Paris, vol. 335, n° 2, p. 239-246.
M. (2001).- The site at Tercis les Bains (Landes), stratotype of the Campanian-Maastrichtian limit: insights from tectonic studies. In : G.S. (réd., 2001a), chap. A6, p. 68-81.
J., J.-P. & G. (2004).- Global Campanian (Upper Cretaceous) ostracod palaeobiogeography.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 213, n° 3-4, p. 379-398.

Cliquer sur la vignette pour agrandir l'image.
Figure 1 : Vue aérienne du site géologique de Tercis (Aquitaine méridionale). Le Nord est situé vers le haut ; les couches sont verticales, la série campanienne est au Sud et se poursuit au Maastrichtien puis au Danien vers le Nord. Le front de carrière, à l'Est, est large de 170 m et a été exploité sur une longueur de 400 m (photo G.S.O., Février 1997).

Cliquer sur la vignette pour agrandir l'image.
Figure 2 : Vue du palier II, cotes 5 à 30, affleurement principal du site de Tercis (Aquitaine méridionale). Les couches sont verticales et carbonatées avec quelques passages décimétriques plus argileux (photo G.S.O., Juillet 1992).

Cliquer sur la vignette pour agrandir l'image.
Figure 3 : Vue du palier III autour du niveau de la limite Campanien-Maastrichtien : cotes 95 à 120 ; affleurement principal du site de Tercis (Aquitaine méridionale). Les couches de calcaires à silex comportent des passées peu argileuses vers les cotes 97 et 118 (photo G.S.O., Juillet 1992).
Cliquer sur la vignette pour agrandir l'image.
Figure 4 : Récoltes d'ostracodes le long de la coupe de Tercis. Les récoltes anciennes rapportées ici (vert à gauche et bleu à droite) sont celles réalisées par l'un de nous (B.A. : bleu, G.S.O. : vert) dans des lavages ; le nombre de spécimens dépassait rarement 100 valves ou carapaces (échelle en haut) par niveau. Les nouvelles récoltes soumises à la présente étude (barres rouges, G.S.O.) ont été fructueuses entre les cotes 40 et 60.
Cliquer sur la vignette pour agrandir l'image.
Figure 5 :
Clichés en microscopie optique de valves obtenues par acétolyse. En haut : vues cavalière ventrale interne et de profil externe de valve gauche de l'espèce Bythoceratina sp. 1
(Pl. 3 ![]() ,
figs. 33-34) ; en bas à gauche : vues de profil externe et cavalière ventrale interne d'une valve droite récoltée à la cote 57,9 d'une espèce du genre Bythoceratina ; en bas à droite : vues de profil externe et interne d'une valve droite récoltée à la cote 57,9 de l'espèce Eucytherura (Vesticytherura) dorsotuberculata.
,
figs. 33-34) ; en bas à gauche : vues de profil externe et cavalière ventrale interne d'une valve droite récoltée à la cote 57,9 d'une espèce du genre Bythoceratina ; en bas à droite : vues de profil externe et interne d'une valve droite récoltée à la cote 57,9 de l'espèce Eucytherura (Vesticytherura) dorsotuberculata.
Cliquer sur la vignette pour agrandir l'image.
Figure 6 : Localisation des spécimens illustrés dans le présent travail. L'espèce Genus indet. sp., synonyme de Mauritsina sp., était déjà illustrée par (2001) ; Rehacythereis aff. agedincumensis est synonyme de Spinoleberis sp. 1 illustrée par (2001) ; les autres taxons sont nouveaux pour le site de Tercis.
Cliquer sur la vignette pour agrandir l'image.
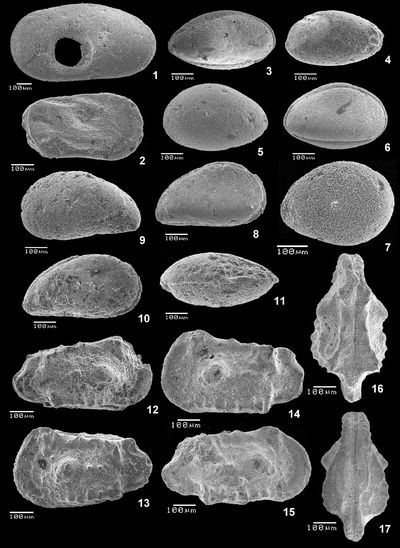
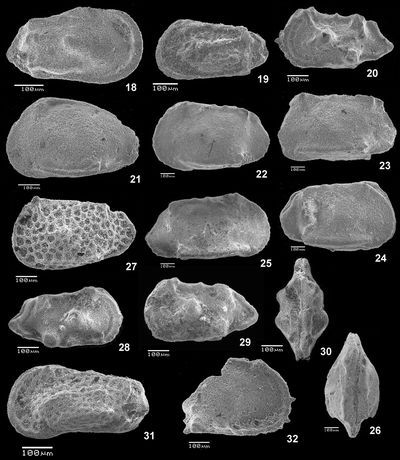
Planche 1 : Fig. 1 Trace de prédation de gastéropode sur une VG d'une Cytherella sp. ; vl (cote 52,2). Fig. 2 Cytherelloidea sp. ; C, vlD (cote 47,2). Figs. 3-4 Cardobairdia ? sp. ; 3 C, vlD (cote 49,0) ; 4 C, vlG (cote 50,05 à 50,25). Figs. 5-6 Asciocythere sp. ; 5 C, vlG (cote 57,9) ; 6 C, vlD (cote 62-62,5). Fig. 7 Xestoleberis cf. northensis , 1966 ; VD (cote 50,05-50,25). Figs. 8-11 Paracyprideis sp. ; 8 C, vlD ; 9 C, vlG ; 10 C, vlD ; 11 C, vd (cote 39,5). Figs. 12-17 Rehacythereis aff. agedincumensis (, 1964) ; 12 C, vlD ; 13 C, vlG (cote 79,8) ; 14 VG, vl ; 15 VD, vl ; 16 C, vd ; 17 C, vv (cote 47,2).
Légende : C = carapace, VD = valve droite, VG = valve gauche, D = Droite, G = gauche, vl = vue latérale, vd = vue dorsale, vv = vue ventrale, ind. = individu.
Cliquer sur la vignette pour agrandir l'image.
Planche 2 : Fig. 18 Phacorhabdotus cf. lonsdaleianus (, 1849) ; C, vlD (cote 47,2). Fig. 19 Hazelina sp. ; C, vlG (cote B-29,5). Figs. 20 & 28-30 Spinoleberis aff. macerrima (, 1936) ; 20 C, vlG (cote 47,2) ; 28 C, vlD (cote 79,8) ; 29 C, vlG (cote 79,8) ; 30 C, vd (cote 79,8). Fig. 21 Pterygocythere aff. hilli , 1957, sensu (1983) ; C, vlG (cote -5,0). Figs. 22-26 Genus indet. sp. ; 22 C, vlG ; 23 C, vlG ; 24 C, vlD ; 25 VD, vl ; 26 C, vd (cote 50,05-50,25). Fig. 27 Limburgina sp. ; C, vlG (cote 50,05 à 50,2). Fig. 31 Paracaudites (Dumontina) aff. puncturata (, 1854) in (1966) ; C, vlG (cote 45,3 à 43,0). Fig. 32 Diogmopteron sp. ; fragment de VD, vl (cote 39,5 à 41,1).
Légende : C = carapace, VD = valve droite, VG = valve gauche, D = Droite, G = gauche, vl = vue latérale, vd = vue dorsale, vv = vue ventrale, ind. = individu.

Cliquer sur la vignette pour agrandir l'image.
Planche 3 : Figs. 33-34 Bythoceratina sp. 1 ; 2 fragments de VG ; 33 partie avant, 34 partie arrière, vl (cote 54,9). Fig. 35 Eucytherura sp. ; C, vlG (cote 57,9). Fig. 36 Bythoceratina cf. umbonata umbonata (, 1848) sensu (1983) ; C, VG, vl (cote 62-62,5). Fig. 37 Bythoceratina sp. 3 ; fragment de VD, vl (cote 57,9). Figs. 38-39 Eucytherura (Vesticytherura) dorsotuberculata (, 1936) ; 38 C, vlD (cote 54,9) ; 39 C, vlD (cote 57,9). Figs. 40-41 Bythoceratina sp. 2 ; 40 VG, vl ; 41 VG, vd (cote 79,8). Figs. 42-43 Aversovalva sp. ; 42 C, vlD (d'après B.A.) ou vlG (d'après G.S.O.) ; 43 C, vv (avant à gauche) du même ind. (cote 89,0).
Légende : C = carapace, VD = valve droite, VG = valve gauche, D = Droite, G = gauche, vl = vue latérale, vd = vue dorsale, vv = vue ventrale, ind. = individu.
Asciocythere sp., Pl. 1 ![]() ,
figs. 5-6 ; cote 57,9 (1 ind.) ; 62,0-62,5 (1 ind.)
,
figs. 5-6 ; cote 57,9 (1 ind.) ; 62,0-62,5 (1 ind.)
Aversovalva sp., Pl. 3 ![]() ,
figs. 42-43 ; cote 89,0 (1 ind.) ; 82,0 (1 ind.)
,
figs. 42-43 ; cote 89,0 (1 ind.) ; 82,0 (1 ind.)
Bythoceratina aff. umbonata umbonata (, 1848) sensu
(1983), Pl. 3 ![]() , fig. 36 ; cote 62 à 62,5 (1 ind.) ; 50,05 (1 ind.)
, fig. 36 ; cote 62 à 62,5 (1 ind.) ; 50,05 (1 ind.)
Bythoceratina sp. 1, Pl. 3 ![]() ,
figs. 33-34 ; cote 54,9 (2 ind.)
,
figs. 33-34 ; cote 54,9 (2 ind.)
Bythoceratina sp. 2, Pl. 3 ![]() ,
figs. 40-41 ; cote 79,8 (1 ind.) ; 54,9 (1 ind.)
,
figs. 40-41 ; cote 79,8 (1 ind.) ; 54,9 (1 ind.)
Bythoceratina sp. 3, Pl. 3 ![]() , fig. 37 ; cote 57,9 (1 ind.)
, fig. 37 ; cote 57,9 (1 ind.)
Cardobairdia ? sp., Pl. 1 ![]() ,
figs. 3-4 ; cote 60,5 (1 ind.) ; 49,0 (2 ind.) ; 50,05-50,25 (2 ind.) ; pour J.-P. (communication personnelle), les
figs. 3-4 illustreraient un mâle de Asciocythere.
,
figs. 3-4 ; cote 60,5 (1 ind.) ; 49,0 (2 ind.) ; 50,05-50,25 (2 ind.) ; pour J.-P. (communication personnelle), les
figs. 3-4 illustreraient un mâle de Asciocythere.
Cytherella sp., Pl. 1 ![]() ,
fig. 1 ; cote 52,2 (1 seul individu percé)
,
fig. 1 ; cote 52,2 (1 seul individu percé)
Cytherelloidea sp., Pl. 1 ![]() , fig. 2 ; cote 47,2 (1 ind.)
, fig. 2 ; cote 47,2 (1 ind.)
Diogmopteron sp., Pl. 2 ![]() , fig. 32 ; cote 39,5 à 41,1 (1 ind.)
, fig. 32 ; cote 39,5 à 41,1 (1 ind.)
Eucytherura (Vesticytherura) dorsotuberculata (, 1936);
Pl. 3 ![]() ,
figs. 38-39 ; cote 54,9 (1 ind.) ; 57,9 (1 ind.) ; 82,0 (1 ind.)
,
figs. 38-39 ; cote 54,9 (1 ind.) ; 57,9 (1 ind.) ; 82,0 (1 ind.)
Eucytherura sp., Pl. 3 ![]() , fig. 35 ; cote 57,9 (2 ind.)
, fig. 35 ; cote 57,9 (2 ind.)
Genus indet. sp., Pl. 2 ![]() ,
figs. 22-26 ; cote 50,05 à 50,25 (8 ind.) ; synonyme de Mauritsina sp. in
(2001)
,
figs. 22-26 ; cote 50,05 à 50,25 (8 ind.) ; synonyme de Mauritsina sp. in
(2001)
Hazelina sp., Pl. 2 ![]() ,
fig. 19 ; cote B-29,5 (1 ind.)
,
fig. 19 ; cote B-29,5 (1 ind.)
Limburgina sp., Pl. 2 ![]() ,
fig. 27 ; cote 50,05 à 50,25 (1 ind.)
,
fig. 27 ; cote 50,05 à 50,25 (1 ind.)
Paracaudites (Dumontina) aff. puncturata (, 1854) in
(1966), Pl. 2 ![]() ,
fig. 31 ; cote 43,0 à 45,3 (1 ind.) ; 50,05-50,25 (1 ind.)
,
fig. 31 ; cote 43,0 à 45,3 (1 ind.) ; 50,05-50,25 (1 ind.)
Paracyprideis sp., Pl. 1 ![]() ,
figs. 8-11 ; cote 39,5 (4 ind.) ; 41,1 (2 ind.) ; 79,8 (1 ind.) ; 57,9 (1 ind. ?)
,
figs. 8-11 ; cote 39,5 (4 ind.) ; 41,1 (2 ind.) ; 79,8 (1 ind.) ; 57,9 (1 ind. ?)
Phacorhabdotus cf. lonsdaleianus (, 1849),
Pl. 2 ![]() ,
fig. 18 ; cote 47,2 (1 ind.)
,
fig. 18 ; cote 47,2 (1 ind.)
Pterygocythere aff. hilli , 1957, sensu
(1983), Pl. 2 ![]() ,
fig. 21 ; cote -5,0 (1 ind.)
,
fig. 21 ; cote -5,0 (1 ind.)
Rehacythereis aff. agedincumensis (,
1964), Pl. 1 ![]() ,
figs. 12-17 ; cote 62,0-62,5 (2 ind.) ; 47,2 (6 ind.) ; 79,8 (2 ind.) ; synonyme de Oertliella cf. pulchra
in &
(2001), et de Spinoleberis sp. 1 in
(2001)
,
figs. 12-17 ; cote 62,0-62,5 (2 ind.) ; 47,2 (6 ind.) ; 79,8 (2 ind.) ; synonyme de Oertliella cf. pulchra
in &
(2001), et de Spinoleberis sp. 1 in
(2001)
Spinoleberis aff. macerrima (, 1936) in
(1966), Pl. 2 ![]() ,
figs. 20 & 28-30 ; cote 47,2 (1 ind.) ; 79,8 (3 ind.)
,
figs. 20 & 28-30 ; cote 47,2 (1 ind.) ; 79,8 (3 ind.)
Xestoleberis cf. northensis ,
1966, Pl. 1 ![]() ,
fig. 7 ; cote 50,05-50,25 (1 ind.)
,
fig. 7 ; cote 50,05-50,25 (1 ind.)
La forme en cur ailé de l'ostracode attribué au genre Aversovalva dans ce travail est difficile à imaginer d'après une photographie. L'orientation même des prises de vues du spécimen photographié au microscope électronique (vues 42-43 ci-dessus) a pu être diversement interprétée. Afin de donner une représentation ne laissant pas de point d'ombre sur la forme de la carapace, nous avons reconstitué un spécimen prélevé à la cote 82,0, dans le Campanien supérieur du site géologique de Tercis ; sa préservation est raisonnablement bonne. La Figure 1 ![]() de cette annexe propose 6 vues au microscope optique de cet individu.
de cette annexe propose 6 vues au microscope optique de cet individu.
Le modèle de l'animation 1 combine les observations au microscope optique et des données de la littérature dont une photographie au microscope électronique de Cytheropteron guadalupense , une espèce récente illustrée dans
http://www.ucl.ac.uk/GeolSci/micropal/ostracod.html (avec l'orthographe incorrecte C. gadaloupense). Cette photographie révèle de nombreux points communs avec notre spécimen tant pour la morphologie que pour l'ornementation. Nous espérons qu'à partir de ce modèle, il sera possible de reconnaître ce taxon s'il est observé ailleurs et de le comparer avec d'autres espèces appartenant à ce même genre pour en proposer une détermination spécifique qui n'a pas encore pu être obtenue.
Comparé au taxon antérieurement nommé Aversovalva sp. ( & , 2001), qui est interprété ici comme Pterygocythere aff. hilli, la différence la plus évidente est la taille ; Pterygocythere est longue de 600-700 µm et Aversovalva sp., récoltée à Tercis, deux fois plus petite d'après les deux spécimens observés. À Tercis, Pterygocythere sp. était connue dans le niveau 39,5 en 2001 ; elle a été trouvée à la même cote dans la présente étude ainsi qu'aux cotes 11,0 et 5,0 (1 à 2 spécimens dans chaque niveau) de même, probablement, qu'à la cote 65,4 (valve seule).
N.B. : dans un travail antérieur ( & , 2001), le spécimen illustré de Pterygocythere est incorrectement noté, une fois Aversovalva sp. (vue 16) et une fois Planileberis (vue 20 obtenue sur le même individu), dans la légende des planches, p. 399.

Cliquer sur la vignette pour agrandir l'image.
Figure 1 : Photographies au microscope optique d'un ostracode récolté au niveau 82,0 à Tercis ; de gauche à droite : vue latérale droite légèrement ventrale ; vue dorsale, avant à droite ; vue ventrale antérieure (noter les deux rides sur la valve droite de l'individu située à gauche de la vue) ; vue latérale gauche, avant à gauche ; vue dorsale antérieure, avant en bas ; vue sub-ventrale, avant en haut (noter encore les deux rides sur la face ventrale de la valve droite située à gauche).
Cliquer sur la vignette pour lire la vidéo (flv = 1.373 KB)
ou cliquer sur l'icone ![]() pour télécharger une autre version de la vidéo (avi = 2.072 KB)
pour télécharger une autre version de la vidéo (avi = 2.072 KB)
ou cliquer sur l'icone ![]() pour ouvrir et lire une autre version de la vidéo (mpg = 1.294 KB)
pour ouvrir et lire une autre version de la vidéo (mpg = 1.294 KB)
ou cliquer sur l'icone ![]() pour télécharger une autre version de la vidéo (wmv = 3.130 KB)
pour télécharger une autre version de la vidéo (wmv = 3.130 KB)
Animation : Panorama tridimensionnel d'une reconstitution d'un spécimen d'Aversovalva en coeur ailé récolté au niveau 82,0 à Tercis. Noter que la valve droite est plus petite que la valve gauche. Modèle 3D réalisé avec le logiciel libre Blender (http://www.blender.org/). Une application de l'utilisation de ce logiciel est présentée par & (2006).