![]()
Contents
[Introduction] [The boundary between ...]
[Considerations regarding the genus Imerites , 1933]
[Conclusions] [Bibliographic references] and ... [Figures]
* Corresponding author
Université de Bourgogne, Laboratoire Biogéosciences, UMR CNRS 5561, 6 bd Gabriel, F-21000, Dijon (France)
Département des Sciences de la Terre, Université de Nice-Sophia-Antipolis, Faculté des Sciences, 28 avenue Valrose, 06108 Nice Cedex 2 (France)
945 route de Gattières, F-06640 Saint Jeannet (France)
Manuscript online since January 17, 2011
![]()
Recent revisions of the genus Imerites make it possible to introduce a new biochronologic horizon to define more precisely the lower boundary of the Giraudi Zone: the Dichotomus Horizon. Using the concept of 'interval zone', this new horizon maintains the current lower boundary of the Giraudi Zone as accepted by authors, and thus contributes to the stabilization of the Barremian zonal system. This stabilization is also strengthened by abandonment of the use of "Crioceras" cristatus d' (nomen dubium) that ought not be used as an index species in detriment of Imerites giraudi (). The classification, origin, and intraspecific variation of the genus Imerites are examined.
Tethyan realm; Upper Barremian; biostratigraphy; Interval zone; southeastern France; ammonites.
D., G. & D. (2011).- The Dichotomus Horizon: proposal for a new biochronologic unit of the Giraudi Zone of the Upper Barremian of southeastern France, and considerations regarding the genus Imerites (Ammonoidea, Gassendiceratinae).- Carnets de Géologie / Notebooks on Geology, Brest, Article 2011/01 (CG2011_A01)
L'horizon à Dichotomus : proposition d'une nouvelle unité biochronologique de la zone à Giraudi du Barrémien supérieur du Sud-Est de la France, et considérations sur le genre Imerites (Ammonitina, Gassendiceratinae).- Les récentes révisions du genre Imerites permettent à présent d'introduire un nouvel horizon biochronologique afin de mieux définir la limite inférieure de la Zone à Giraudi : l'horizon à Dichotomus. Par l'application du concept de la zone d'intervalle, l'utilisation de cet horizon permet de conserver la limite inférieur actuelle de la zone à Giraudi telle qu'elle a été acceptée par les auteurs, et ainsi de contribuer à la stabilisation du schéma zonal du Barrémien. Cette stabilisation est aussi renforcée par l'abandon de l'usage de "Crioceras" cristatus d' (nomen dubium) qui ne devrait pas être utilisé comme espèce indice au détriment de Imerites giraudi (). La classification, l'origine et la variabilité intraspécifique du genre Imerites sont aussi discutées.
Domaine téthysien ; Barrémien supérieur ; biostratigraphie ; zone d'intervalle ; Sud-Est de la France ; ammonites.
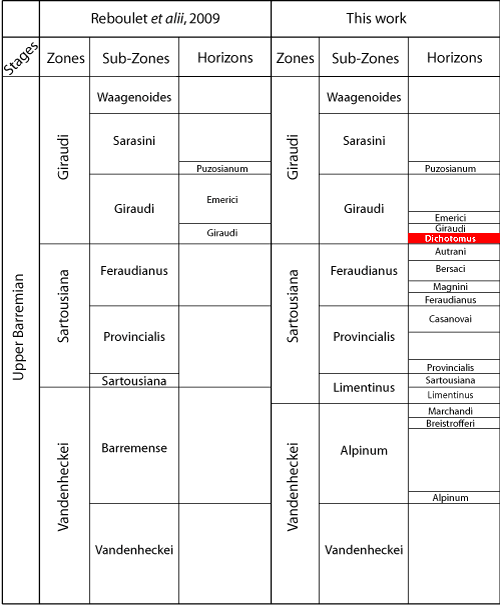
After a period of instability, in recent years several studies have helped to fix the calibration of the Upper Barremian biostratigraphy of southeastern France that increased its value for practical use and increased the degree of its reproducibility by specialists working in discrete localities ( et alii,
2007, 2009 - see et alii, 2008 for a historical account). It now includes the
Vandenheckei, Sartousiana and Giraudi zones (Fig. 1 ![]() ), their limits based on faunal changes commonly associated with sequence boundaries developed in relation to eustasy (, 2005). In accord with the recommendations of the
IUGS Lower Cretaceous Ammonite Working Group, the
Group, stabilization of the boundaries of these zones led to a preference for the use of interval zones rather than zones based on the range of a
taxon. With good reason, for the boundaries of zones based on individual ranges are subject to repeated change in range limits caused by new finds or by tergiversation resulting from a revision of index
taxa. Thus it seems preferable to choose stable and precise horizons for defining the base of zones (, 1997). The introduction of several biochronological horizons (, 1995, 1997, 1998; et alii, 2008; & , 2009; et alii, 2010) has contributed significantly to a refinement of the biostratigraphic pattern of the Tethyan Upper Barremian of southeastern France.
), their limits based on faunal changes commonly associated with sequence boundaries developed in relation to eustasy (, 2005). In accord with the recommendations of the
IUGS Lower Cretaceous Ammonite Working Group, the
Group, stabilization of the boundaries of these zones led to a preference for the use of interval zones rather than zones based on the range of a
taxon. With good reason, for the boundaries of zones based on individual ranges are subject to repeated change in range limits caused by new finds or by tergiversation resulting from a revision of index
taxa. Thus it seems preferable to choose stable and precise horizons for defining the base of zones (, 1997). The introduction of several biochronological horizons (, 1995, 1997, 1998; et alii, 2008; & , 2009; et alii, 2010) has contributed significantly to a refinement of the biostratigraphic pattern of the Tethyan Upper Barremian of southeastern France.
This work revises the definition of the lower boundary of the Giraudi Zone [index species: Imerites giraudi (, 1888)] and is a manifestation of progress in the continuation of the revision of Upper Barremian biostratigraphy in southeastern France through emendation to the ammonite faunas which are a major element of its framework (here Imerites , 1933).
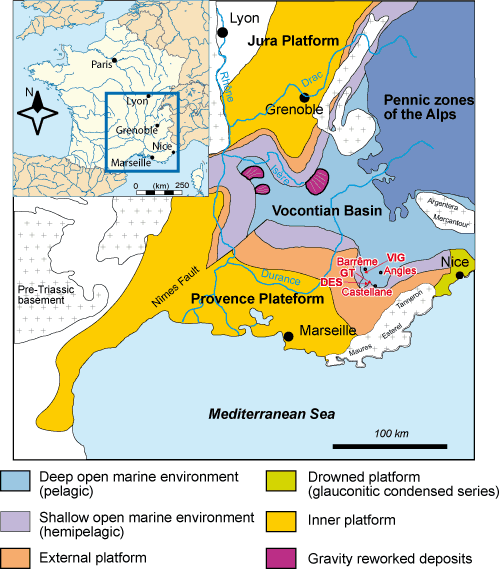
The Lower Cretaceous of southeastern France is marked by the evolution of a large intracratonic subsident area known as the Vocontian Basin (, 1900). The area of the historical Barremian stratotype
(Angles-Barrême-Castellane area Fig. 2 ![]() ), is in the southern part of the Vocontian Basin which is less affected by gravity
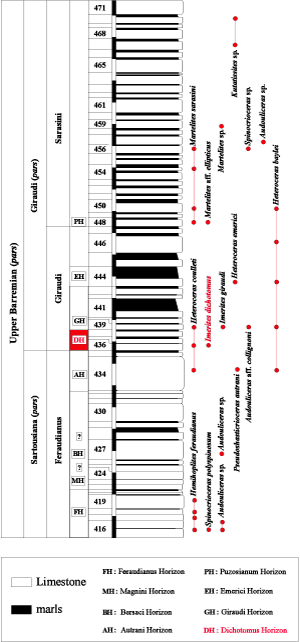
remodelling and Alpine orogeny than its northern portion. The Barremian here is characterized by pelagic sediments, mainly alternation of marls and limestones in decimetric to metric beds. Given the relative continuity of deposits and the paleontological record, it is possible to track in considerable detail the succession and evolution of their ammonite faunas. Thus, following up on the work of (1995, 1998), several sections of this area expose the boundary between the Sartousiana and the Giraudi zones well enough that they can be examined minutely: they are the Vignon section
(VIG, Fig. 3
), is in the southern part of the Vocontian Basin which is less affected by gravity
remodelling and Alpine orogeny than its northern portion. The Barremian here is characterized by pelagic sediments, mainly alternation of marls and limestones in decimetric to metric beds. Given the relative continuity of deposits and the paleontological record, it is possible to track in considerable detail the succession and evolution of their ammonite faunas. Thus, following up on the work of (1995, 1998), several sections of this area expose the boundary between the Sartousiana and the Giraudi zones well enough that they can be examined minutely: they are the Vignon section
(VIG, Fig. 3 ![]() ), the Descouère section (DES,
Fig. 4
), the Descouère section (DES,
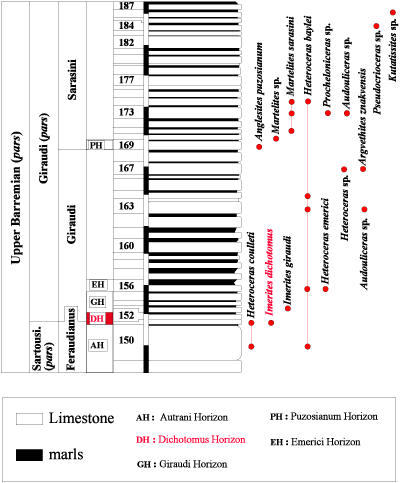
Fig. 4 ![]() ), and the Grande-Terre section
(GT, Fig. 5
), and the Grande-Terre section
(GT, Fig. 5 ![]() ).
).
The base of the Giraudi Zone is characterized by an important phase of marine
transgression (, 2005), marked lithologically by the
"vire marneuse à Heteroceras" of authors. It is associated with a major faunal turnover (, 1990, 1995, 1997, 1998; et alii, 2008). This turnover is relatively progressive in strata at the top of the Feraudianus Subzone and at the base of the Giraudi Subzone. In fact, all change occurs between the lower part of the Feraudianus Subzone and the Emerici Horizon of the Giraudi Subzone
(Fig. 1 ![]() ) where the Hemihoplitinae are progressively replaced quantitatively by the Heteroceratidae. At the base of the Feraudanus Subzone Hemihoplitinae are the major components of the ammonite fauna with a preponderance of the genus Hemihoplites . The Pulcheliidae and the Peirescinae are quite rare, as are the Gassendiceratinae
(genera Gassendiceras , & ,
2006, and Pseudoshasticrioceras ,
1998) which become more numerous at the top of the Subzone (i.e. in the Bersaci and Autrani horizons). The genus Heteroceras d' (with a turriculate morphology) is present at the upper limit of the Feraudianus Subzone (Autrani Horizon) but is extremely rare, for at this level and time it is only a very minor element of the ammonite fauna. In the lower portion of the Giraudi Subzone the situation is reversed: the Hemihoplitinae and the Pulcheliidae are gone and turriculate morphology dominates quantitatively, first briefly with Imerites (Gassendiceratinae), and later with Heteroceras which begin an increase in abundance at the base of the Giraudi Zone and proliferate accompanied by a morphological explosion in the Emerici Horizon (, 1990, 1995, 1997, 1998; & , 2000; & , 2006).
) where the Hemihoplitinae are progressively replaced quantitatively by the Heteroceratidae. At the base of the Feraudanus Subzone Hemihoplitinae are the major components of the ammonite fauna with a preponderance of the genus Hemihoplites . The Pulcheliidae and the Peirescinae are quite rare, as are the Gassendiceratinae
(genera Gassendiceras , & ,
2006, and Pseudoshasticrioceras ,
1998) which become more numerous at the top of the Subzone (i.e. in the Bersaci and Autrani horizons). The genus Heteroceras d' (with a turriculate morphology) is present at the upper limit of the Feraudianus Subzone (Autrani Horizon) but is extremely rare, for at this level and time it is only a very minor element of the ammonite fauna. In the lower portion of the Giraudi Subzone the situation is reversed: the Hemihoplitinae and the Pulcheliidae are gone and turriculate morphology dominates quantitatively, first briefly with Imerites (Gassendiceratinae), and later with Heteroceras which begin an increase in abundance at the base of the Giraudi Zone and proliferate accompanied by a morphological explosion in the Emerici Horizon (, 1990, 1995, 1997, 1998; & , 2000; & , 2006).
Above the Autrani Horizon (Feraudianus Subzone, Sartousiana Zone, Fig. 1 ![]() ), the appearance of the genus Imerites is currently accepted by authors as the valid marker of the lower limit of the Giraudi Zone (see historical account in , 1989; & , 1990; , 1990, 1995, 1998; et alii, 2006, 2007, 2009; et alii, 2008). Revisions of this genus by (1998) and et alii (2009), improved understanding of the stratigraphic distribution of the species of Imerites. Their development over time is coincident with the evolutionary framework of the last Gassendiceratinae , & , 2006. So
in the succession Imerites dichotomus , 1955 appears before Imerites giraudi (,
1888). The use of an interval zone allows a redefinition of the lower boundary of the Giraudi Zone without change of level by use of the new Dichotomus Horizon (defined below). This usage preserves the integrity and stability of Barremian zonation ( et alii, 2008). There is no need to move the lower limit of the Giraudi Zone upward to make it coincident with the appearance of its index species, or to rename the Giraudi Zone as the Dichotomus Zone, because I. dichotomus is the first species of Imerites of which the appearance coincides with the accepted definition of the lower limit of the Giraudi
Zone. In any event, any shift in level would dissociate the base of the Giraudi Zone from the faunal turnover and the sequence boundary with which it is coincident (, 2005); application either of these options would change the level of the base of the zone and return to the instability that has long characterized Barremian zonation and should cease.
), the appearance of the genus Imerites is currently accepted by authors as the valid marker of the lower limit of the Giraudi Zone (see historical account in , 1989; & , 1990; , 1990, 1995, 1998; et alii, 2006, 2007, 2009; et alii, 2008). Revisions of this genus by (1998) and et alii (2009), improved understanding of the stratigraphic distribution of the species of Imerites. Their development over time is coincident with the evolutionary framework of the last Gassendiceratinae , & , 2006. So
in the succession Imerites dichotomus , 1955 appears before Imerites giraudi (,
1888). The use of an interval zone allows a redefinition of the lower boundary of the Giraudi Zone without change of level by use of the new Dichotomus Horizon (defined below). This usage preserves the integrity and stability of Barremian zonation ( et alii, 2008). There is no need to move the lower limit of the Giraudi Zone upward to make it coincident with the appearance of its index species, or to rename the Giraudi Zone as the Dichotomus Zone, because I. dichotomus is the first species of Imerites of which the appearance coincides with the accepted definition of the lower limit of the Giraudi
Zone. In any event, any shift in level would dissociate the base of the Giraudi Zone from the faunal turnover and the sequence boundary with which it is coincident (, 2005); application either of these options would change the level of the base of the zone and return to the instability that has long characterized Barremian zonation and should cease.
In France Imerites dichotomus has a very limited and precise stratigraphic position. By anagenesis ( et alii, 2009) it is the successor of the index species Pseudoshasticrioceras autrani (), and the ancestor of the index species Imerites giraudi (). In the stratotype area (Vocontian Basin, southeastern France Fig. 2 ![]() ) its appearance in the stratigraphic succession is in agreement with its biologic relationships. These facts, and the need for the establishment of a high resolution biostratigraphy for the whole of the Barremian ( et alii, 2006; et alii, 2007; et alii, 2008; et alii, 2009), has impelled us to propose Imerites dichotomus as a new biostratigraphic marker in the Vocontian Basin. This species occurs in strata immediately above the major beds of the Autrani Horizon at the top of the Feraudianus Subzone (Sartousiana Zone), and immediately precedes those of the Giraudi Horizon where Imerites giraudi () occurs
(Figs. 3
) its appearance in the stratigraphic succession is in agreement with its biologic relationships. These facts, and the need for the establishment of a high resolution biostratigraphy for the whole of the Barremian ( et alii, 2006; et alii, 2007; et alii, 2008; et alii, 2009), has impelled us to propose Imerites dichotomus as a new biostratigraphic marker in the Vocontian Basin. This species occurs in strata immediately above the major beds of the Autrani Horizon at the top of the Feraudianus Subzone (Sartousiana Zone), and immediately precedes those of the Giraudi Horizon where Imerites giraudi () occurs
(Figs. 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() ).
).
Index species: Imerites dichotomus , recently revised by et alii (2009).
Status: This horizon is defined by the first appearance of its index species (bed No. 436 in the Vignon [VIG] section,
Fig. 3 ![]() ), and its upper limit is currently set at the base of the Giraudi Horizon (bed No. 439 in the VIG section) with the first appearance of Imerites giraudi (). The Dichotomus Horizon is also present in the sections near La Baume (Castellane area): beds 151 to 152 in the Descouère section (DES,
Fig. 4
), and its upper limit is currently set at the base of the Giraudi Horizon (bed No. 439 in the VIG section) with the first appearance of Imerites giraudi (). The Dichotomus Horizon is also present in the sections near La Baume (Castellane area): beds 151 to 152 in the Descouère section (DES,
Fig. 4 ![]() ), and bed 679 in the Grande-Terre section
(GT, Fig. 5
), and bed 679 in the Grande-Terre section
(GT, Fig. 5 ![]() ) [see also , 1995, 1998].
) [see also , 1995, 1998].
Paleobiogeographic distribution: Imerites dichotomus is present in southeastern France but also in Spain (oral communication of , see , 1998, p. 207), Bulgaria, Romania and Georgia (see et alii, 2009), a distribution that augers an extensive application of the Dichotomus Horizon.
Faunal assemblages: The index species is generally fairly well represented in the French' sections. It is associated with (see , 1995, 1998; et alii, 2008): Macroscaphites yvani () macro- and microconchs, Acantholytoceras pseudoaudouli () macro- and microconchs, Jaubertites collignoni , Protetragonites crebrisulcatus (), Eulytoceras phestus (), Silesites seranonis (d'), Melchiorites melchioris (), Barremites difficilis (d'), Barremites strettostoma (), Phyloceras ponticuli (), and with Heteroceras coulleti , Heteroceras baylei and Spinocrioceras trachyomphalus ().
The genus Imerites has traditionally been classified as a Heteroceratidae, but it is now recognized as a representative of the family Hemihoplitidae (see et alii, 2009 for an historical account) despite the presence of a turricone in the juvenile part of the shell (helicoidal coiling). was the first to classify Imerites (his Escragnolleites) in the Hemihoplitidae (1955, p. 24) because it has a hemihoplitid ancestor (1955, p. 18, 22), although (1907-1913) saw "some similarities" between Heteroceras giraudi (=Imerites) and some Hemihoplitidae [=Ancyloceras (Crioceras) heberti in 's time]. It has been recognized for a long time that helicoidal coiling is not restricted to the Heteroceratidae. Indeed, (1888, 1889) was the first to restrict the genus Heteroceras d' only to those Barremian taxa that have a turricone. But d' classified one Senonian species (1851, p. 222) as a heteroceratid, and (1876) included helicoidal forms from the Upper Cretaceous of the United States of America (Nostoceratidae) in that group, thus indicating broader criteria for their classifications. This type of coiling is known to have developed repeatedly in the evolutionary history of the ammonoids (e.g. Triassic Cochloceras; some Bajocian Spiroceras; Uppermost Barremian Kutatissites; some Lower Cretaceous Leptoceratoidae; Albian Mariella, Turrilitoides, Helicoceras or Pseudhelicoceras; Cenomanian Turrilites, Hypoturrilites, Mesoturrilites and Ostlingoceras; Axonoceras, Jouaniceras, Anaklinoceras and some other Nostoceratidae from the Upper Cretaceous, etc. see et alii, 1957 and et alii, 1996). But these authors did not remark a connection between these homeomorphs and the Heteroceratidae. More recently, (2004, p. 21) made the following observation about certain genera with helicoidal coiling and the absence of any relationship to the Heteroceratidae: "the similarity in the mode of coiling (helicoidal, planispiral or helicoidal, planospiral and uncoiled) is not important for identifying the systematic position as to family". And (1998, p. 184) said about Imerites that "the presence of the turricone would be only the expression of a homeomorphism that affects the early developments of these forms" (translation pars).
et alii (2009) were the first to distinguish and define evolutionary processes in the origin and development of species in the genus Imerites of the Hemihoplitidae. Their work involved study of an evolved "Pseudoshasticrioceras" ornament in Imerites dichotomus (see et alii, 2006). In that stage of development weaker peri-ventral tubercules demonstrate that Imerites is a direct descendant of the genus Pseudoshasticrioceras (Gassendiceratinae , & ). This taxonomic distinction between genera of the same lineage is based on the early appearance of a turricone during the growth of Imerites ( et alii 2009).
We welcome acceptance of the genus Imerites into the Gassendiceratinae but we do not agree with the broader generic limits proposed for Imerites by & (2010, p. 18, 20). Their diagnosis includes some non-turriculate Hemihoplitidae, which would cause problems that preclude its acceptance:
This problem is particularly evident in the taxon "I." stephaniae & , 2010 (found in the Provincialis Subzone: One subzone separates it from the appearance of the first Imerites s. str.) that they attribute to the genus Imerites , but do not compare it with contemporary Hemihoplitids. "I." stephaniae & is a fragmentary specimen ( & , 2010, p. 21, Pl. 1, fig. 7) particularly close in aspect to contemporary Hemihoplites of the group H. casanovai (=H. intermedius - currently under study).
As (in et alii, 2006, p. 138-139), maintained "Crioceras" cristatus d' should be considered synonymous with Imerites giraudi () and according to the principle of priority of the International Code of Zoological Nomenclature (ICZN - Art. 23.1) has seniority. Consequently, for & (2010, p. 20) another amendment of Barremian zonation would be necessary: renaming the Giraudi Zone as the Cristatus Zone [note that this is not obligatory, for the International Stratigraphic Guide (, 1994, p. 67) states that "if it is desirable to continue use of a taxonomic term which is no longer valid, the term should be in quotation marks"].
However new data obtained in the recent revision of Imerites by et alii (2009, p. 32-33) found the syntypes of "Crioceras" cristatus d' of the d''s collection are too fragmented to be identified at a specific level, and the fragments may not be of just one specimen but may even represent more than one species. These facts render this taxon unusable and a nomen dubium. Moreover, d''s (1842, Pl. 115, figs. 4-8) original illustrations are probably a synthetic compilations (i.e. a picture based on several different but fragmentary specimens assembled and restored to appear as if they were a single specimen). They were recognized as such by (1888, 1889). In the original illustration the turricone is replaced by planispiral whorls for the stages of growth of the actual syntypes (particularly fragmented) when compared to the original picture make this obvious. So the rehabilitation of "Crioceras" cristatus d', 1842, to replace Imerites giraudi (, 1888), a long-used, well defined taxon (recognized by many authors), is not desirable. Moreover, the purpose of the ICZN principle of priority is explained by Article 23.2 which states very explicitly that it must foster nomenclatural stability: "it is not intended to be used to upset a long-accepted name in its accustomed meaning by the introduction of a name that is its senior synonym or homonym, or through an action taken following the discovery of a prior and hitherto unrecognized nomenclatural act"; a fortiori when the older name is based on a synthetic compilations and can be considered as a nomen dubium because it differs from characteristics found in its syntypes. On the other hand, the proposal of invalidation by & (2010, p. 17-18) of the d''s syntypes in favour to a more complete neotype (not among the syntypes) does not comply with the provisions of the ICZN (Art. 75, and in particular, § 75.3.4) and therefore cannot be implemented. At most it might be possible to designate one of the original syntypes as a lectotype (n°5405-1 of the d''s collection), but the basic problem remains unchanged...
Whatever the nomenclatural possibilities mentioned earlier ( in et alii, 2006, p. 139), Imerites giraudi () is the type-species of the genus Imerites and we strongly recommend "Crioceras" cristatus d' be considered invalid.
Intraspecific variability in species of Imerites involves ornamentation, some dimensional parameters of the shell and the size and orientation of the turricone
(Fig. 6c ![]() ). The relationship of turricone size and placement to changes in shell morphology has been studied by (1998, p. 53) for the genus Heteroceras and was illustrated by (2004,
Fig. 10) for the Kutatissites. In Imerites, the phenomenon is the same: a larger turricone generally is accompanied by a less regular or triangular coiling before normal involution resumes. However, contrary to the views of & (2010) these purely mechanical constraints are not comparable with the dimorphic differentiation in Imerites dichotomus where tripartite adult microconch [m] are associated with large planospiral adult macroconch [M] (see et alii,
2009, Pl. 2, fig. 2 versus Pl. 1, fig. 1 Fig. 6a
). The relationship of turricone size and placement to changes in shell morphology has been studied by (1998, p. 53) for the genus Heteroceras and was illustrated by (2004,
Fig. 10) for the Kutatissites. In Imerites, the phenomenon is the same: a larger turricone generally is accompanied by a less regular or triangular coiling before normal involution resumes. However, contrary to the views of & (2010) these purely mechanical constraints are not comparable with the dimorphic differentiation in Imerites dichotomus where tripartite adult microconch [m] are associated with large planospiral adult macroconch [M] (see et alii,
2009, Pl. 2, fig. 2 versus Pl. 1, fig. 1 Fig. 6a ![]() ) for several reasons:
) for several reasons:
Recent revisions of the genus Imerites provide an increased understanding of its variability and its evolution, so now permit the introduction of a new biochronologic horizon, the Dichotomus Horizon
(Fig. 1 ![]() ), to define the lower boundary of the Giraudi Zone more precisely. Using the interval zone concept, the addition of this horizon maintains the current lower boundary of the Giraudi Zone as accepted by authors (the appearance of the genus Imerites), and thus contributes to the stabilization of the Barremian zonal scheme. This stabilization is also strengthened by the abandonment of
"Crioceras" cristatus d' (nomen dubium) that ought not be used as index species instead of Imerites giraudi (). Following (1997), in general (as it is for the Barremian) it is preferable to use interval zones rather than the stratigraphic range of any one taxon for example, to avoid constant changes of boundaries that discoveries and systematic reviews of index taxa might impose, thus threatening the stability of the zonation. The introduction of a new biochronologic horizon increases the precision of the biostratigraphic zonation of the Barremian stage in southeastern France which is a requirement for the study in detail of the evolution of ammonites and their populations.
), to define the lower boundary of the Giraudi Zone more precisely. Using the interval zone concept, the addition of this horizon maintains the current lower boundary of the Giraudi Zone as accepted by authors (the appearance of the genus Imerites), and thus contributes to the stabilization of the Barremian zonal scheme. This stabilization is also strengthened by the abandonment of
"Crioceras" cristatus d' (nomen dubium) that ought not be used as index species instead of Imerites giraudi (). Following (1997), in general (as it is for the Barremian) it is preferable to use interval zones rather than the stratigraphic range of any one taxon for example, to avoid constant changes of boundaries that discoveries and systematic reviews of index taxa might impose, thus threatening the stability of the zonation. The introduction of a new biochronologic horizon increases the precision of the biostratigraphic zonation of the Barremian stage in southeastern France which is a requirement for the study in detail of the evolution of ammonites and their populations.
We wish to express our thanks to Mr. Raymond , and to the anonymous reviewer, Mr. Stephane and to Mr. Jaap for their valuable advice. We thank the Réserve Géologique de Haute Provence and Mrs. Myette who have allowed us access to the areas studied. One of us (D.B.) has interesting discussions with Gerd E.G. about stratigraphical nomenclature, and we warmly thank him. We want especially and sincerely to thank Nestor who kindly corrected our English in the final version.
W.J., B. & C.W. (1957).- Mesozoic Ammonoidea. In: R.C. (Ed.), Mollusca 4.- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, p. L80-L437.
H. (2005).- The South-East France Basin (SFB) and its Mesozoic evolution.- Géologie Alpine, Grenoble, (série spéciale "Colloques et Excursions"), n° 7, p. 5-28.
D. & G. (2009).- Pseudoshasticrioceras bersaci nov. sp. (Ammonoidea, Gassendiceratinae), and new ammonite biohorizon for the Upper Barremian of southeastern France.- Carnets de Géologie / Notebooks on Geology, Article 2009/02 (CG2009_A02), 22 p.
D., G. & S. (2006).- Descriptions de représentants nouveaux ou peu connus de la Famille des Hemihoplitidae , 1924 (Barrémien supérieur, Sud-Est de la France) : conséquence taxinomique et phylétiques.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 179-253.
D., G. & S. (2008).- Propositions pour un nouveau découpage biozonal ammonitique, et nouveaux biohorizons pour le Barrémien supérieur.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/03 (CG2008_A03), 18 p.
D., G. & L. (2009).- L'origine des Imerites , 1933 : résultat d'une innovation chez les Gassendiceratinae , 2006 (Ammonoidea, Ancyloceratina).- Annales de Paléontologie, Paris, vol. 95, p. 21-35.
D., R., G. & S. (2010).- Problems in the identity of "Crioceras" barremense , 1895 (Ancyloceratida, Late Barremian), and their proposed resolution.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/01 (CG2010_A01), 17 p.
G. (1990).- Précisions nouvelles sur "Emericiceras" collignoni (Ammonoidea) et proposition d'un nouveau schéma zonal dans le Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 311, p. 259-264.
G. (1995).- Les biozones à Feraudianus, Giraudi et Sarasini du Barrémien supérieur de la région stratotypique d'Angles-Barrême-Castellane (Sud-Est de la France).- Géologie Alpine, Grenoble, Mémoire H.S., n° 20 (1994), p. 279-319.
G. (1997).- Biostratigraphie haute résolution du Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 325, p. 689-694.
G. (1998).- Biostratigraphie des faunes d'Ammonites à la limite Barrémien-Aptien dans la région d'Angles-Barrême-Castellane. Étude particulière de la Famille des Heteroceratidae (Ancyloceratina, Ammonoidea).- Annales du Muséum d'Histoire Naturelle de Nice, t. XII (1997), 270 p.
G. & D. (2006).- Nouvelles unités taxinomiques de la famille des Heteroceratidae , 1924.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 255-295.
G. & L. (2000). Une nouvelle espèce d'Heteroceras: H. mascarelli sp. nov. (Ancyloceratina, Ammonoidea) dans le Barrémien supérieur du S-E de la France.- Annales du Muséum d'Histoire Naturelle de Nice, t. XV, p. 1-17.
M.S. (1955).- Nizhnemelovaja fauna Gruzii (Lower Cretaceous fauna of Georgia).- Institut Geologii i Mineralogii Akademii Nauk G.S.S.R., Monografii, Tbilisi, 6, 224 p. (en russe).
H. avec la collaboration de R., R., G., J.-C., F., B., W.J., J. & H. (eds.) (2006).- Révision critique de la Paléontologie Française d'Alcide d', Vol. IV, Céphalopodes Crétacés, sous la direction de J.C. .- Backhuys Publisher, Leiden, 292 p.
P.J. & L. (reporters), E., R., M., G., M., E., J., I., J.L., L., P.F., J., J.M., J.-P., G., Z. & J. (1990).- Preliminary ammonite zonation for the lower Cretaceous of Mediterranean region (Lower Cretaceous Cephalopod team, Working Group of IGCP, Project 262).- Géologie Alpine, Grenoble, t. 66, p. 123-127.
M.V. (1989).- The Barremian biostratigraphical subdivisions of Georgia and comparison with some western mediterranean regions. In: J. (ed.), Cretaceous of the Western Tethys.- Proceedings 3rd International Cretaceous Symposium, Tubingen (1987), E. Schweizerbart'sche Verlagsbuchandlung, Stuttgart, p. 551-560.
M.V. (2004).- Intraspecific and intrageneric variabilities and their implication for the systematics of the Cretaceous heteromorph ammonites; a review.- Scripta Geologica, Leiden, vol. 128, p. 17-37.
W. (1888).- Sur quelques fossiles du Crétacé inférieur de la Provence.- Bulletin de la Société Géologique de France, Paris, (3ème série), t. XVI, p. 663-691, pls. XVII-XXXI.
W. (1889).- Description géologique de la montagne de Lure.- Thèse, Masson édit., Paris, 458 p.
W. (1907-1913).- Das Mezosoïcum Unterkreide (Palaeocretacium).- Lethaea Geognostica, Stuttgart, Teil III, Bd. 3, p. 234-279.
W. & P. (1915).- I. La faune de l'Aptien inférieur des environs de Montélimar (Drôme) (Carrière de l'Homme d'Armes). In: W. (ed.), Contribution à l'étude des faunes paléocrétacées du Sud-Est de la France.- Mémoires pour servir à l'Explication de la Carte Géologique détaillée de la France, Paris, n° 14, 221 p.
E.V., I.V., M.V. & L.R. (eds., 2005).- Atlas of Early Cretaceous fauna of Georgia.- Proceedings of the Georgian Academy of Sciences, A. Janelidze Geological Institute, Tbilisi, (n. ser.), vol. 120, 788 p.
F.B. (1876).- A report on the invertebrate Cretaceous and Tertiary fossils of the Upper Missouri Country.- Report of the United States Geological Survey of the Territories, Washington, vol. IX, 629 p.
A. d' (1840-1842).- Paléontologie française. Description des mollusques et rayonnés fossiles - Terrains crétacés. Tome Premier. Céphalopodes.- Masson, Paris, 662 p.
A. d' (1851).- Notice sur le genre Heteroceras, de la classe des Céphalopodes.- Journal de Conchyliologie, Paris, tome second, p. 217-222.
M.V. (1900).- Recherches géologiques dans le Diois et les Baronnies orientales.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de l'Université de Grenoble, t. 5, p. 149-556.
S. (reporter), F., D., L., R., G. & J. (2007, unpublished).- Zonation Hauterivien Barrémien.- Compte rendu de la réunion des biostratigraphes français du Group (IUGS Lower Cretaceous Ammonites Working Group), Digne-les-Bains (2 Mai 2007), 14 p.
S. & P.J. (reporters), M.B., P., F., E.Y., M., G., Y., J. J.L., A., V., F.A., I., N., P., J., J.M., Z., J., H., B. & I. (2006).- Report on the second international meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Neuchâtel, Switzerland, 8 September 2005).- Cretaceous Research, London, vol. 27, n° 5, p. 712-715.
S. & J. (reporters), R., M., C., A., S.N., J., O., J.M., Z. & J. (2009).- Report on the 3rd International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Vienna, Austria, 15th April 2008).- Cretaceous Research, London, vol. 30, n° 2, p. 496-502.
J. (1933).- Les ammonites aptiennes de la Géorgie occidentale.- Bulletin de l'Institut géologique de Géorgie, Tbilisi, vol. 1 (1932), n° 3, p. 165-273.
A. (ed., 1994).- International stratigraphic guide. A guide to stratigraphic classification, terminology and procedures.- I.U.G.S. & Geological Society of America, 214 p.
S.S. (1955).- Révision des Ammonites déroulées du Crétacé inférieur du SE de la France.- Mémoires de la Société Géologique de France, Paris, (N.S.), t. XXXIV, n° 72, 176 p.
J. (1997).- La biostratigraphie, du taxon à la biozone et aux échelles biozonales. In: J. (éd.), Stratigraphie, Terminologie Française.- Bulletin des Centres de Recherches Exploration-Production elf-aquitaine, Pau, Mémoire 19, p. 63-89.
J. (2000).- Nouvelles données sur les répartitions stratigraphiques, les évolutions et les classifications de trois familles d'ammonites du Crétacé inférieur.- Géologie Alpine, Grenoble, n° 75, p. 123-132.
J. (2006).- Nouvelle classification à fondement phylogénétique des ammonites hétéromorphes du Crétacé inférieur.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 137-178.
J. & P. (2010). Nouvelles données sur le genre Imerites , 1933 (Ammonidita, Ancyloceratina) ; réponses et précisions apportées au travail de , et , 2009.- Annales de Paléontologie, Paris, vol. 96, p. 13-23.
C.W., J.H. & M.K. (eds., 1996).- Cretaceous Ammonoidea. In: R.L. (ed.), Mollusca 4 (revised).- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, 362 p.
Click on thumbnail to enlarge the image.
Figure 1: Zonal scheme amended after
et alii, 2008, and et alii, 2009. In red the Dichotomus Horizon (new).
Figure 1 : Schéma zonal modifié d'après
et alii, 2008, et et alii, 2009. En rouge, l'horizon à Dichotomus (nouveau).
Click on thumbnail to enlarge the image.
Figure 2:
Barremian paleogeography of southeastern France and location of the area studied (from , 2005, amended).
Figure 2 : Cadre paléogéographique du Sud-Est de la France et localisation du secteur d'étude
(modifiée d'après , 2005).
Click on thumbnail to enlarge the image.
Figure 3: Distribution of the ammonites in the Vignon
(VIG) section pars (Barrême area, Alpes-de-Haute-Provence).
Figure 3 : Répartition des faunes d'ammonites dans la coupe du Vignon
(VIG) pars (secteur de Barrême, Alpes-de-Haute-Provence).
Click on thumbnail to enlarge the image.
Figure 4:
Distribution of the ammonites in the Descouère (DES) section pars (La Baume in the Castellane area,
Alpes-de-Haute-Provence).
Figure 4 : Répartition des faunes d'ammonites dans la coupe du Descouère (DES) pars (La
Baume, secteur de Castellane, Alpes-de-Haute-Provence).
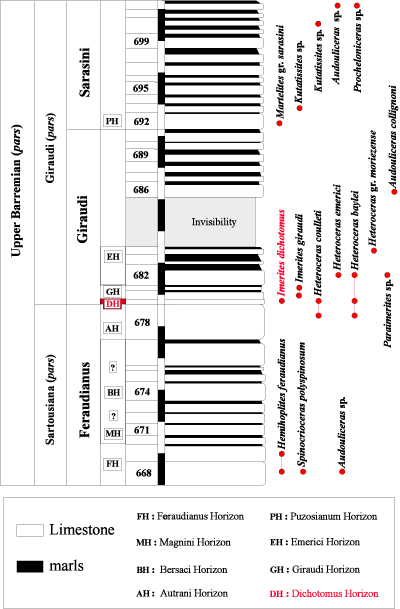
Click on thumbnail to enlarge the image.
Figure 5:
Distribution of the ammonites in the Grande-Terre (GT) section pars (La Baume in the Castellane area,
Alpes-de-Haute-Provence).
Figure 5 : Répartition des faunes d'ammonites dans la coupe de la Grande-Terre
(GT) pars (La Baume, secteur de Castellane, Alpes-de-Haute-Provence).

Click on thumbnail to enlarge the image.
Figure 6:
Dimorphism and variability in shell morphology in Imerites.
Figure 6 : Dimorphisme et variabilité morphologique de la coquille chez Imerites.