![]()
Contents
[1. Introduction] [2. Location of outcrops]
[3. Stratigraphy]
[4. Regional changes in lithology and thickness] [5. Environment of sedimentation]
[6. Regional correlation of the Hasa series] [7. Paleontology]
[Bibliographic references] and
... [Plates]
Modesto CA 95355-9171 (U.S.A.)
Manuscript online since May 11, 2012
[Editors: Christian , Bruno and Michel ]
![]()
The beds of early Tertiary age in eastern Saudi Arabia are in large part chemical precipitates laid down in a shallow epicontinental sea. The sequence is divided into three formations, each clearly differentiated in both lithology and fauna. From bottom to top they have been named the Umm er Radhuma, Rus and Dammam formations. Ecologic conditions remained more or less uniform for considerable lengths of time. Consequently, the fauna, made up mainly of foraminifera, became well adapted to the environment, as demonstrated by the abundance of some species. The great number of individuals of these species have a range of variability much broader that that seen in other areas of their occurrence. The detailed study of these variations has demonstrated that species have often been defined on too small a number of individuals. The abundance of specimens has allowed evolutionary trends in species to be traced even to the extent that in the end some variants must be classed as a discrete genus.
Arabian peninsula; Paleogene; Umm er Radhuma; Rus; Dammam; larger foraminifera.
N.J. (2012).- Paleontologic and stratigraphic overview of the Paleogene in eastern Saudi Arabia.- Carnets de Géologie [Notebooks on Geology], Brest, Article 2012/04 (CG2012_A04), p. 53-92.
Aperçu paléontologique et stratigraphique du Paléogène en Arabie Séoudite orientale.- Les couches du début du Tertiaire en Arabie Séoudite orientale sont, pour la plupart, des dépôts chimiques d'une mer épicontinentale peu profonde. La séquence se divise en trois formations, chacune nettement différenciée par sa lithologie et sa faune. On les a nommées, de bas en haut, les Formations de Umm er Radhuma, Rus et Dammam. Les conditions écologiques sont restées plus ou moins inchangées pendant des périodes considérables. Par conséquent, la faune, surtout constituée par des foraminifères, a pu s'adapter au milieu, comme le montre la grande prolifération de quelques espèces. De ce fait, les individus de chacune de ces espèces sont très nombreux et montrent une gamme de variations bien plus large que celle rencontrée dans d'autres régions. L'étendue de la variation a été étudiée minutieusement afin de montrer que les définitions des espèces sont souvent bien trop limitées, étant établies à partir d'un nombre trop restreint d'individus. Le grand nombre d'échantillons nous a permis de montrer les lignes d'évolution entre espèces et même de suivre celle des formes aboutissant à un autre genre.
Péninsule arabique ; Paléogène ; Umm er Radhuma ; Rus ; Dammam ; grands foraminifères.
ForewordThe text and illustrations that follow are in their entirety extracts from a 'Thèse d'État' titled: "La stratigraphie de l'Éocène le long du rivage occidental du Golfe Persique", presented 5 March 1952 at the University of Paris. A large portion of the text and more than half of the plates and figures are not reproduced here. Professor J. has suggested that nevertheless it would be interesting to summarize the principal conclusions and to print some of the descriptions of microfossils, even though this work was presented ten years ago [sixty years ago today!]. Indeed, as the thesis was not printed it is unknown to most geologists. Some of the new species and the new genus described in the thesis (1952) and in this note appear in 's thesis of 1954 under other names. |
Since Cambrian times the Arabian peninsula (except Oman) has not been subject to large-scale orogenic movements, and, because of this relative stability many lithologic units consist of fine-grained sediments, among which limestones and evaporites are major constituents. Early Tertiary formations follow this rule. They are almost entirely limestones, marls, and evaporites laid down in the littoral of the Tethys sea.
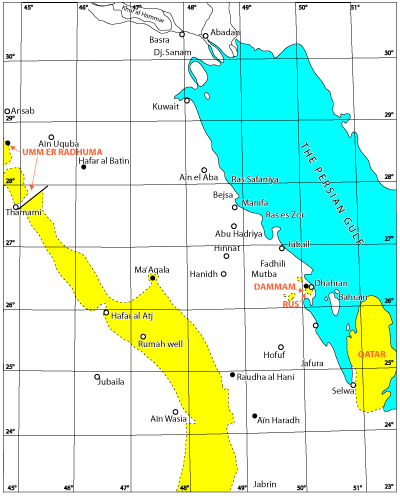
The area included in this study lies between 22° and 30° North. It occupies the broad area between the shore of the Persian Gulf and a NW trending line joining Ain Wasia and Themami
(Fig. 1  ). Outcrops of Eocene age occupy only a small part of this 34,000 square kilometers, because most of this area is covered either by Neogene sediments or sand. However, marine strata assigned the Eocene underlie the entire region east of the outcrops.
). Outcrops of Eocene age occupy only a small part of this 34,000 square kilometers, because most of this area is covered either by Neogene sediments or sand. However, marine strata assigned the Eocene underlie the entire region east of the outcrops.
Some 250 km west of the shoreline in the western portion of the Summan, rocks of Paleocene-Eocene age crop out in a band ranging in width from 25 to 120 km. This band, although partially covered by the Dahana, (moving sand) forms an arc convex to the East. The arc occupies much of Saudi Arabia north of the 'Rub al Khali and crosses the northern border into Iraq where Paleogene outcrops also occupy extensive tracts. To the South the outcrop belt disappears under a younger Tertiary cover. In outcrop the Paleogene is some 330 m thick of which the basal 160 m represent the Paleocene.
In the coastal region of Al Hasa outcrops of Eocene age are few. They exist only on Dammam dome, in Bahrain, on the Qatar peninsula and in the Haradh area. Where topographic relief exists (Dammam, Bahrain and Qatar) erosion has exposed sections complete enough to make their study worthwhile. However these outcrops have been found to be markedly thinner than the succession present in the coastal subsurface, undoubtedly because of the local absence of evaporites, a slower rate of sedimentation over sea-floor highs and more intense compaction later, all due to continual uplift during and after deposition.
The lower part of the outcrop sequence is more or less strongly altered in many localities, sufficiently so that the fauna is for the most part unidentifiable or unrecognizable.
Click on thumbnail to enlarge the image.
Figure 1: Outcrops of the lower Tertiary. West of the Persian Gulf (• RUS: location of type sections).
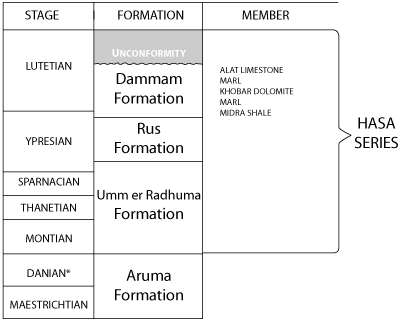
Strata of Paleocene and Eocene age present offshore and west of the Persian Gulf are designated by the term Hasa Series, taken from the name of the province of al Hasa where they crop out. In Saudi Arabia itself this series includes strata that have been correlated with European stages from the base of the Montian through much of the Lutetian. North and east of Saudi Arabia upper stages of the Eocene epoch may exist ( et al., 1951) but in Al Hasa the series ends with an incomplete representation of the Lutetian stage.
ARAMCO geologists divide the Hasa Series into three formations - each has been mapped as such. The basal unit has been named the Umm er Radhuma Formation. The name was chosen because of the site of the type section in an erosional escarpment near a well of that name; limestone and dolomite predominate, but the formation includes small quantities of marl and calcareous shale. The formation appears to rest conformably on Cretaceous limestones of similar aspect. This lower boundary is placed at the top of a thin bed of dark colored dolomitic clay shale, under which Maestrichtian fossils, predominantly neritic, occur. Not far above the shale a shallow water, mainly benthonic Paleocene microfauna exists, its representatives persisting for tens of meters of section. In the upper part of the Umm er Radhuma Formation, in poorly bedded strata lithologically similar to those below, is a fauna dated early Eocene by geologists working in India ( & , 1937; , 1925a). Portions of this fauna continue to the upper limit of the formation where there is a rather abrupt transition to anhydrite, limestone, and varicolored clay shale, characteristics of a shallow desiccating basin.
This environment produced strata easily identifiable in both well and outcrop. They are named the Rus Formation from the outcrop on the southeast flank of Jebel Umm er Rus. Anhydrite, a characteristic of the sequence encountered in many wells of coastal Al Hasa, is absent in this locality.
The uppermost lithologic entity of the Hasa series on land is the Dammam Formation named from its occurrence on the western flank of Dammam dome. It marks the return of a normal marine environment in marls and shales with a benthonic marine fauna thought to be of Ypresian age that grade upward to limestones with a Lutetian microfauna. At the upper limit of some outcrops is a thin bed of silicified limestone containing the leached impressions of small molluscs.
The great lateral persistence of even the smallest recognizable units making up the Hasa Series confirm that it was formed at a time when epeirogeny was the principal mechanism of diastrophism active in the region with continual slow upward movement of salt bodies the only significant modifier of the stable regime.
Figure 2
shows the approximate correlation of the Paleogene sequence in Saudi Arabia with the subdivisions of geologic time (stages) established in Europe. Currently, the Danian stage is placed in the Cenozoic because its fauna of planktonic microfossils, not present in the epeiric sea of eastern Saudi Arabia, has Tertiary affinities.
Click on thumbnail to enlarge the image.
Figure 2: Relations
of formations to standard stages.
* In 1952, the Danian was considered to represent the uppermost Cretaceous. No fossils characteristic of the stage have been found to date.
These formations were named and described in a symposium of the American Association of Petroleum Geologists ( et al., 1958). However a more extensive discussion of the succession is in the thesis.
The Umm er Radhuma Formation is not as well exposed at the type locality as it is at Wadi al Batin, 108 km to the southeast. There the sequence referred to this formation is 217 meters thick. The basal unit is 15 meters of very strongly dolomitized limestone on which lie 74 meters of softer chalky limestone and 128 meters of gray to cream crystalline limestone, of which the uppermost 40 meters are partially silicified. In the chalky levels of some outcrops a poorly preserved Paleocene fauna can be recognized while at Wadi al Batin the uppermost 50 meters includes an Eocene microfauna. In no locality has the fauna been well preserved.
However, some of the wells that drilled through the Umm er Radhuma Formation found a large number of microfossils in an excellent state of preservation. Consequently, it has been possible to make lithologic and faunal correlations between well and outcrop. The lithologies of the Umm er Radhuma seen in wells are similar to those of the outcrop, but less altered: they are an essentially unbroken sequence of light colored limestones with scattered intercalations of marl and impure limestone.
It is possible to make subdivisions in this fauna characteristic of warm, shallow water. It includes a number of forms identical to those found in India in the upper Ranikot and lower Laki formations.
At the type locality the Rus Formation is made up only of light colored limestones, chalk, and marls about 55 meters thick. It usually comprises three units:
At its type locality the Dammam Formation is only 28 meters thick, but includes all of the levels that characterize it. Near the coast, the basal element is a gray crystalline, pyritic limestone with a small number of foraminifera as fauna. Above it are marls and clays that range widely in thickness, but always include a distinctive brown argillaceous limestone. This limestone is present everywhere near the coast but cannot be recognized far to the southwest. Above the Midra shale (named from its occurrence in Jebel Midra Shemali on Dammam Dome) interbedded in lighter-colored shales are one, occasionally two limestone units that reach a maximum thickness of eight meters each. This limestone, light yellow to brownish yellow in color, is soft, porous, and slightly impure. It has a fauna of foraminifera poor in species but rich in individuals, mainly Alveolina subpyrenaica . The shales above this Alveolina limestone are more calcareous locally than those below it and grade upward to a gray marl with abundant nummulites.
This transition to a more limy regime continues upward with marly limestones, commonly dolomitized near their upper limit which is marked by an abrupt change to dolomitic clay or marl. Dolomitization of the limestone is commonly almost complete and the crystalline texture of the altered beds is recognized in the name "Khobar dolomite". The name is taken from the occurrence of a fossiliferous outcrop of the unit near the village of Al Khobar where it includes an abundant microfauna of Lutetian age.
The marl or shale above the Khobar dolomite is capped by gray, finely crystalline limestone, frequently dolomitic, called the "Alat limestone" from its outcrop near the well Alat n° 1. Over extensive tracts this unit has been made porous through the dissolution of the shells of small pelecypods and gastropods, generally indeterminate. Fortunately some molds were silicified, thus making it feasible to demonstrate that beds of late Eocene age are absent in Al Hasa province.
Near the coast overlying marine deposits are marls and shaly clays that rest with slight angular discordance on Eocene strata. Lower Miocene marine beds may extend some twenty kilometers inland. They are thinner and more sandy with distance from the present coast. Where marine Miocene is absent Eocene strata are covered by fresh water limestones, clays or gravels that, at least in part, are continental equivalents of the marine beds to the east.
Less information is available concerning regional variation in the thickness and lithology of the Umm er Radhuma Formation than for overlying units. Wells scattered over a large part of coastal Arabia for the most part do not reach its base. In outcrop, exposures of the entire sequence are few because of the low dip and massive character of the beds. Nevertheless, in the Wadi al Batin channel it is possible to measure its thickness which is about 217 meters. Thickness increases southward so that near Ma'agala it is some 275 m, but it thins again farther south to less than 150 m at Wadi Sahaba. A few tens of km farther south the whole of the Paleogene sequence is covered by Neogene continental deposits.
The thickest section drilled through is along the coast in northern Al Hasa — 490 m. Southward a gradual thinning occurs, comparable to that seen in outcrop. Near Dhahran the thickness is but 386 m; still farther south and toward the interior it is only 217 m. Although a part of the thinning may be local and caused by salt tectonics, a substantial portion must result from regional movements of an epeirogenic character.
The Rus and Dammam formations have been drilled through many times by widely dispersed wells. At first glance it is clear that Eocene strata thicken markedly to the northeast and that the progression is in a southwest-northeast direction. Detailed study shows that almost all lithologic entities thicken in the same proportion. Erosion at the unconformable upper limit seems to have been more or less consistent over the entire area, but because to the south and west the succession is thinner, older beds form the erosional surface. The maximum thickness of lower and middle Eocene strata known is in a well at Abu Hadriya where it is 361 m. Farther north and east the thickness is about the same, at least 330 m. The thinnest section known is south of Wadi Sahaba — 28 m.
The regional continuity of lithologies has a number of exceptions. Anhydrite is present in the Rus Formation only a considerable distance down dip from the outcrop, its distribution poorly defined. The limestone with Alveolina subpyrenaica above the Midra shale and under the Khobar dolomite disappears in the far south. Apparently the arm of the sea in which it was deposited did not extend as far as that which laid down the gray marls above and below it.
A similar phenomenon may have influenced the distribution of the Midra shale, but the source of this earthy dark-colored entity, in part finely clastic, may have been in part a water course or courses of which the discharge had a distribution more restricted than that of precipitates of calcium carbonate.
Variations in the make up of the Khobar dolomite are due mainly to the degree of dolomitization. When undolomitized the unit is a fine-grained soft limestone. Where the invasion of magnesium was complete it produced a porous, coarsely crystalline dolomite. Its upper limit, an abrupt change to a clay or dolomitic marl, is easy to recognize wherever the sequence is complete. In the south erosion cuts below the contact.
The aspect of the Alat limestone changes little wherever it is found. Any changes are due principally to the presence or absence of the porosity caused by the almost universal and complete solution of the shells of the small molluscs when they are present in large numbers. Other variations are due to change in the number of intercalations of marl in the predominant limestone.
Color changes are common in all three formations so color cannot be used to identify them. However, texture, the sequence of the succession of rock types and the faunas permit easy recognition of equivalency from one locality to another. For example, the dolomitic marl or clay just above the Khobar dolomite may be dark brown, blue, gray, orange or yellow but it is always in the same position between a dolomitic limestone above and a dolomite or limestone below. Where this lower unit is not completely dolomitized it exhibits a characteristic middle Eocene fauna.
Certain color changes have been attributed to the position of a given bed above or below the regional piezometric level. In general it appears that the more brilliantly colored and more variegated sediments are above the water table.
In eastern Saudi Arabia the restricted number of lithologies and their lateral continuity over the whole area suggests that the depositional environment must have more or less uniform from early Paleocene into medial Eocene times. The most acceptable explanation for this uniformity is that these strata were laid down in a broad shallow epicontinental sea, with its greatest depth not over 200 m. Nearly all of the sediments deposited by it were chemical or organic precipitates in the form of calcareous muds with occasional small quantities of fine clastics introduced by water courses either with an extremely low gradient or with the point of discharge at a great distance from this part of the sea.
A slight decrease in depth and an interruption in direct communication with the open sea during the time of deposition of the Rus Formation would have been sufficient to produce evaporation, a concentration of salts and eventually an important body of anhydrite.
In general, the sedimentary rocks discussed here thin over tracts that are now structural highs. This thinning is greater than that which would be anticipated from compaction alone; more so because anticlinal slopes average less than three degrees. Removal of sediment by currents does not seem to have been in effect because there are almost no detrital or oolitic elements.
Granted that most of the succession was made up of chemical precipitates — then, if the rate of precipitation were uniform throughout the water column, water depth would be the main factor controlling the quantity of sediment formed and thinner sequences would be deposited over sea floor highs. Renewed uplift or continual slow uplift would maintain the differential in water depth and thus accentuate the differences in thickness between highs and lows caused not only by the reduced rate of sedimentation over highs but also by the greater degree of compaction on them. The accentuated structures thus developed are called supratenuous folds.
The fauna confirms that the sea was shallow. It has many elements in common with faunas of the same age in India and a lesser number are shared with those of British Somaliland. The microfaunas of these two regions are considered by & (1937), (1926a), and & (1931) as indicative of a littoral and warm water milieu. [The sparse macrofaunas do not controvert this opinion]
In some portion of southern Iraq lithologies similar to those of eastern Saudi Arabia exist along with vestiges of the fauna, but by far the major portion of the forms described here have not, according to current knowledge [1952] reached this northern extension of the Tethyan sea. [Drilling along Tapline -the Trans-Arabian Pipe Line- has shown that the sequence of lithologies and the faunas described here disappear rapidly near the northern boundary of Saudi Arabia]
The relationships of the fauna of the Hasa series to those of other regions is mentioned above. The following paragraphs enlarge on and complete the discussion of the regional distribution of the invertebrate fauna of the series.
Most of the identified molluscs from Bahrain (one gastropod and eight pelecypods) occur also in the middle Eocene of northwest India [now for the most part Pakistan], Somalia, and Egypt, but the presence of Ostrea turkestanensis indicates a linkage with central Asia too. All of the species from Bahrain (described by ) have been found on the mainland, almost all in the form of internal molds difficult to identify. Several other small gastropods and pelecypods have been found in silicified limestones at the upper limit of the Dammam Formation. None of them invalidate (and a few support) 's conclusions regarding age.
In short, the molluscan fauna of the Dammam Formation has much in common with those of the Kirthar and Laki formations of India, with the middle Eocene of Somalia and a more tenuous relationship with the upper Libyan and lower Mokattam of Egypt.
The foraminiferal fauna adequately confirms the middle Eocene age of the Dammam Formation, or more precisely the greater part of it, and has a close affinity with those present in strata of the same age in Somalia and western India [now Pakistan] as well as some links with Egypt. Larger foraminifera found in both the Dammam Formation and the middle Eocene of Somalia include: Nummulites somaliensis, N. discorbina var. major, Dictyoconus aegyptiensis, Dictyoconoides kohaticus, Linderina buranensis, Alveolina subpyrenaica, and Dictyoconus indicus. Species in common with those of Egypt include Nummulites beaumonti, Orbitolites complanata and Alveolina subpyrenaica. Nummulites gizehensis, so abundant in Egypt, is absent in Saudi Arabia and Bahrain. This may indicate that rocks of late Lutetian age were not laid down in the Persian Gulf region, or perhaps that the distribution of this species was limited regionally.
In northern Al Hasa other species of nummulites are present. They appear identical to certain species described in western Europe and found in Syria. They have not been reported south of 27°30'N.
The Rus Formation is almost barren of identifiable fossils, but the Umm er Radhuma Formation is rich in microfauna, at least locally. Like the lower Eocene of Somalia and India the upper Umm er Radhuma contains Lockhartia tipperi, Miscellanea miscella, Rotalia trochidiformis, and Flosculina globosa.
However, the faunas of the Ranikot and Laki beds of India have even more in common with that of the Umm er Radhuma because of the presence of Sakesaria cotteri, Lockhartia conditi, Lockhartia haimei, Operculina cf. canalifera, Nummulites lahirii and Nummulites cf. irregularis.
In addition, Alveolina frumentiformis and Alveolina lepidula found in the upper Umm er Radhuma are common in the lower and middle Eocene of Egypt (, 1930).
The abundance of individuals of the genera Lockhartia and Miscellanea in the lower Umm er Radhuma occurs also in rocks of the same age in northwest India, although a number of Operculines and Assilines in the Paleocene of India as described by have not been seen in the Persian Gulf region. With the help of the faunal assemblage discussed above an attempt was made to correlate the formations of Paleogene age along the western shore of the Persian Gulf with those of Egypt and India
(Fig. 3 ).
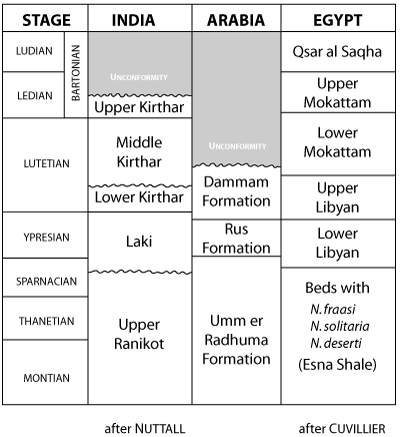
Click on thumbnail to enlarge the image.
The study of the fauna of the Hasa series was made primarily to establish as precisely as possible its position in the standard time scale. The lower limit was determined with ease because of the presence of an indubitable Paleocene microfauna immediately above an unquestionable Maestrichtian assemblage. The upper limit too is relatively well established by the presence of large nummulites that commonly occur only in strata of Lutetian age. They are found just below the erosional (Oligocene absent) upper boundary of the series.
The precise relationship of the boundaries of the lithologic subdivisions of the series to the time scale is more difficult to determine. In fact, the limits indicated in the stratigraphic portion of this study should be regarded only as an exercise of judgment because it is difficult to make exact correlations between widely separated areas of which the faunas have few elements in common. The correlations made here attempted to use other places (Egypt, Somalia, India) in which more or less precise time relationships have been established, to set up grosso modo, an equivalency to the European standard.
The study was not hampered by a lack of microfaunas. Sampling from wells and outcrop in most of eastern Saudi Arabia provided adequate material. Difficulties were presented by the fact that a number of species differ sufficiently from those described elsewhere to make identity uncertain. But, as was done with nummulites, stratigraphic position may be estimated based on the degree of evolution required to reach an equivalent stage. In spite of these problems it is likely that the correlations indicated are close to being exact.
Methods used to study faunas comprised:
All those who participated in the identification of molluscs agree that the beds from which they were obtained are of middle Eocene age, although M. P. points out that Ostrea turkestanensisis perhaps a little older. This species comes from the Midra shale and therefore could well be of early Eocene age, but not older, while all the other molluscs come from higher levels in the Dammam Formation.
| Eastern Arabia |
India | Egypt | Somalia | Europe | elsewhere | |||||||||||||
| L | M | U | L | M | U | L | M | U | L | M | U | L | M | U | L | M | U | |
| Terebellum carcassense | x | x | x | x | ||||||||||||||
| Rimella fissurella | x | |||||||||||||||||
| Lyria turgidula | x | |||||||||||||||||
| Lucina metableta | x | x | x | x | x | |||||||||||||
| Lucina pharaonis | x | x | x | x | x | x | x | x | x1 | x1 | ||||||||
| Spondylus radula | x | x | x | x | x | x | x | x | x4 | |||||||||
| Euphenax jamaicensis | x | x | x2 | |||||||||||||||
| Ostrea turkestanensis | x | x3 | ||||||||||||||||
| Mytilus cleopatrae | x | x | x | |||||||||||||||
| Corbula subexarata | x | x | x | x | x | x | x5 | |||||||||||
| Chama calcarata | x | x | x | x | x | x | x | |||||||||||
Figure 4
shows the distribution of some of these macrofossils and the age of the strata in which they
occur [1. Algeria-Tunisia; 2. Jamaica; 3. Asia; 4. Southern Arabia (); 5.
Burma].
As indicated above, a large part of the microfauna of the Hasa series exists in widely separated areas of the province of Al Hasa. Identical species and associations of species and genera are found in wells and in outcrops separated by some 500 km. The wide distribution of these associations all of which appear to have been restricted to shallow warm water indicates a certain uniformity of ecologic conditions and suggests that the milieu in which sediments were deposited was also relatively stable throughout a great area — a suggestion supported by similar conclusions regarding lithologic entities in widely separated tracts — see "Environment of Deposition" above.
Certain relatively thin lithologic entities, mainly limestones with a very considerable areal extent, have a distinctive fauna, very rich in individuals but limited in the number of species. Elsewhere these beds are replaced by strata of the same age but in discrete lithologic and faunal facies. Thus some species are confined to a narrow range of environments. Consequently the very large area of occurrence of these stenobionomic species shows even more positively the relative uniformity of ecologic and lithologic environments in Al Hasa during the time that the Umm er Radhuma and Dammam formations were being laid down.
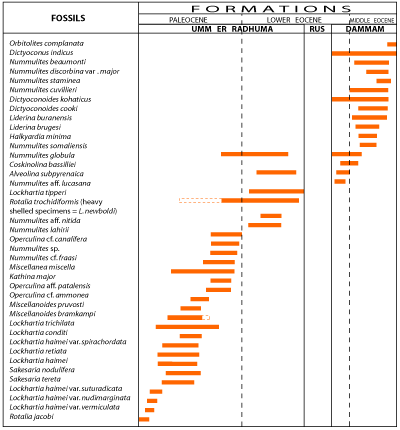
With the exception of the nearly azoic Rus Formation, most of the marls and limestones of the Hasa series are highly fossiliferous with great numbers of large benthonic foraminifera as well as smaller forms, they too mostly benthonic, and a number of ostracods. Based on the restricted stratigraphic ranges of a good number of these forms it has been possible to establish zones based on the latest occurrence of a given species.
Click on thumbnail to enlarge the image.
Figure 5
lists a number of species whose upper limit is sharply defined and, from what has been
observed, indicates the presumed lower limit of their occurrence.
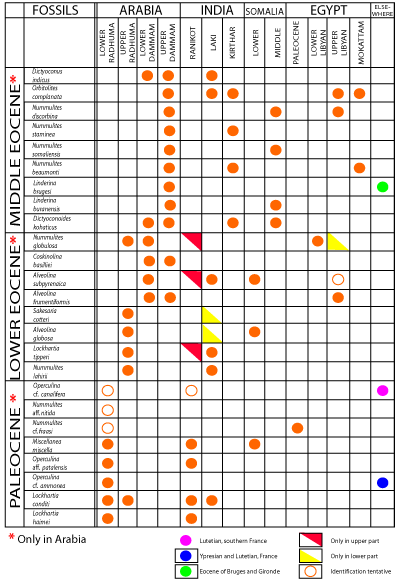
Click on thumbnail to enlarge the image.
Figure 6
indicates what is known concerning the occurrence of some of the species of Figure 5
in other regions. The two charts are the basis for the tentative assignment of the units of the Hasa series to the European stages.
Because of the great number of species found in the Dammam and Umm er Radhuma formations it was not possible to discuss those that seemed identical to those already known. However, the species that seem to be valuable as stratigraphic markers in Arabia are figured and their principal characteristics sketched.
New species are described in detail. Each is illustrated by several photographs. The specimens photographed were chosen to illustrate the range of variation for each species as defined by the author.
The synonymy is not complete. The reader desiring more detailed references may consult the works cited in the bibliography and in the synonymies. Holotypes, paratypes and cotypes have been deposited in the collection of the Laboratory of Paleontology of the University of Paris. Some paratypes and topotypes have been placed in the collections of the American Museum of Natural History, New York, and in the collections of the Arabian American Oil Company. [The specimens at the AMNH have been transferred to the National Museum (Smithsonian) in Washington, D.C., under Brian ]
Order Foraminifera d'
Family Nummulitidae , 1861
Genus Nummulites , 1801 (1)
(1) Established by Decision N° 192, International Committee on Rules of Nomenclature.
Genotype: Nummulites laevigata (2) (, 1792)
(2) Nummulites was considered a feminine word by ; modern latinists confirm this view ( & , 1934).
The following characters have been used to define the several species of Nummulites found in Arabia: (1) Size and general form; (2) type of septal filaments and their abundance; (3) presence, dimensions and placement of granules; (4) characteristics of the spire, dimensions of the step, the degree of regularity and the thickness of the spiral lamina and the number of whorls; (5) form, type and placement of septa; (6) shape of the chambers; (7) size and shape of the proloculum in megalospheric specimens; (8) presence and degree of prominence of the marginal cord, the existence and placement of pillars; (9) character of the test in axial section.
In order to present the several dimensional relationships of the species discussed the following rapid and convenient method of notation has been adopted. The first expresses in the form of a fraction the relationship of diameter to thickness.
For example: d/t : 3.8/1.3 à 4.7/2.6. [measurements in the fractions are given in mm]
The second relationship (w/d) is that between the number of whorls and the diameter: 8.0/3.4; the third one (s/w) is the number of septa in a quadrant of the given whorl: 9-10/3, 16-17/7.
Because much of the fauna of older beds appears to belong to the Indo-Pacific province it was presumed that the nummulites too would be species found in this vast region. Most of them are not. Only a small part of the nummulite species of Al Hasa are unquestionably Indo-Pacific — N. lahirii and N. staminea. The others are more or less typically European, except N. somaliensis, which, although resembling N. laevigata, is clearly different. The same comment applies to the new species of a granular appearance like N. cuvillieri and N. cizancourti.
Nummulites cuvillieri n.sp., Form B
Pl. 1 , figs. 1-3, 9, 11-13
1952. — Nummulites cuvillieri n.sp., Forme B, , p. 115-117, Pl. V, figs. 1-3, 9, 11-13.
Test small, the diameter/thickness (d/t) ratio ranging from 2.1/0.7 to 2.6/1.1, averaging 2.4/0.9. It is thus relatively thick, the two faces nearly parallel but depressed near the poles and sloping abruptly near the edge so the periphery is obtusely rounded. Large granules (0.1 to 0.16 mm in diameter) cover nearly the entire surface. In some rare specimens the granules are almost completely absent. The granules are close together thus forming a spiral at the edge of the whorls, but this arrangement is not obvious in many individuals because of the great size of the granules. Their diameter is greatest in the central depressed region, where they cover most of the surface. This region occupies about one third of the diameter of the test. After weathering or special preparation septal filaments are visible near the periphery. There are 9-10 per quadrant; they are not visible in the central depression. The granules lie on the filaments or touch them. In certain examples are represented on the last whorl only by thickenings of the filaments themselves. Tangential sections show that the filaments are sinuous between the granules and are not necessarily unbroken in their continuations to the periphery.
The spire is regular, the relation w/d is 6.0/2.5; the step increases regularly in the first three whorls, and decreases only slowly thereafter. The mean step in the outer whorls is 0.25 mm. It decreases slightly in the last whorl of some specimens. The thickness of the spiral lamina is a quarter to a third of the height of the step, varying slightly in the same specimen. Septa are numerous (s/w : 4/3, 6-7/4, 9-10/5, 10-11/6) at the base inclined ten to fifteen degrees, then gently curved backward in the upper two-thirds thus forming an angle of 30 to 40 degrees with the outer spiral lamina. In the inner whorls the angle may be less. A tiny prolongation like the tip of a shoe pointing forward exists at the base of every septum. The chambers are nearly twice as high as long, except in the last whorl where length and height may be nearly the same (1 to 1.5).
The axial section shows clearly the approximate dumbbell shape of the test: the central area depressed and the extremities much thicker. The three early whorls are regularly involute with interlamellar spaces clearly evident except in the polar region where the third whorl melds with a thick lamina, depressed in the center, against which the spiral lamina forming the last three whorls abut. Interlamellar spaces exist between these outer whorls. Where these outer whorls join the thick polar lamina there are three large pillars, that exist at other points along the lamina of the outer whorls.
Nummulites cuvillieri n.sp., Form A
Pl. 1 ,
figs. 4-8, 10
1952. — Nummulites cuvillieri n.sp., Forme A, , p. 117-118, Pl. V, figs. 4-8, 10.
The test is small (the d/t ratio is from 1.4/0.6 to 1.8/1, averaging 1.550/0.725), weakly lenticular with a small polar depression present on some specimens. The periphery is sharp, but the test faces are rather sharply angled where they approach the edge. Granules range widely in size and placement with the individual. Their diameter varies from about 0.08 to 0.10 mm. Septal filaments are visible only after weathering, 6-8 by quadrant, and with a sinuous course like that of the microspheric form. The proloculum is double: the two chambers form a flattened oval, the second slightly smaller than the first. The larger diameter of the initial chamber ranges from 0.05 to 0.075 mm. The spire is regular, the step increasing slowly to the periphery (w/d : 4.0/1.6). The spiral lamina is thick, on the average half as high as the chambers. The septa are thick, at an angle of 15 to 20 degrees, with the upper half curved smoothly backward. Chambers weakly sickle-shaped, one and third times higher than long on the average, upper rear angle acute.
The axial section is dumbbell-shaped like that of the microspheric form, but the central depression is less marked. The first and second whorls are normally involute but the thick spiral lamina of the third and fourth meld. It appears that the fourth whorl is even more thickened; its inner edge butts against the large granules developed at the periphery of the depression, where its lamina thins and melds with that of the third whorl. Large pillars are present at the edge of the depressed region, and in lesser number continue to the edge of the test. They may be present in both of the outer whorls, or only in the fourth.
Range: Dammam Formation, Khobar dolomite member.
Nummulites cizancourti n.sp., Form A
Pl. 1 ,
figs. 14-17
1952. — Nummulites cizancourti n.sp., Forme A, , p. 119-121, Pl. III, figs. 16-19.
Test small thick, flatly lenticular with a rather heavy marginal cord that produces an obtusely rounded periphery. The central portion of the test is slightly depressed in some specimens. The surface is covered with large granules (0.1 to 0.2 mm) arranged in a poorly defined spiral, but their number, distribution and arrangement vary. They are not larger in the central area. Septal filaments are visible only on worn tests; there are about 10 per quadrant. The granules are either on the filaments or touch them so the filaments are slightly wavy. Sometimes they thicken near the periphery. Proloculum very small 0.07 to 0.11 mm in diameter, double. The two chambers are a flattened oval, the first slightly larger than the second.
The spire is regular. The step increases rapidly in the first two and a half whorls, more slowly thereafter. In some specimens the step decreases slightly in the outermost whorls. The spiral lamina ranges in thickness from a quarter to a half the height of the chambers; it is constant, its thickness varying only slightly after the first one and a half whorls. The inclination of the septa is variable, sometimes they are vertical, sometimes they are at an angle of 10 to 15 degrees to the spiral lamina of the preceding whorl; commonly they are curved backward at 30 to 40 degrees in the upper third; the angle may be less in the inner whorls. At the base of each septum is a small, forward-pointing projection. Septa are numerous, spaced very regularly (s/w : 4/2, 5-6/3, 7-8/4, 9-10/5). The chambers are higher than long, but may be almost isometric in the outer whorls. The average ratio of height to length is 1.25 to 1. The chamber roofs are weakly arched, the posterior superior angle obtusely rounded.
The axial section shows that the spiral lamina and the interlamellar spaces are both rather wide, but decrease markedly in the central flattened area. Prismatic structure exists in the marginal cord, which is flattened and obtuse only in the last whorl. Pillars are sharply defined, but it is only rarely that they extend between two whorls. They may be either a long oval or pear shaped. In the outer whorl the spiral lamina is thinner and depressed in the central region where it is in contact with the preceding one. Some of the rows of granules are linked by lamellae. The interior whorls are swollen centrally because of the megalosphere.
This species resembles Form B of N. cuvillieri n.sp., but can be easily recognized externally because of the absence of a marked central depression and in section by the megalosphere. It is much larger than Form A of N. cuvillieri.
Range: Lower Dammam Formation, Al Hasa province.
Family Miscellaneidae , 1952
Genus Miscellanoides n.gen. (3)
(3) This genus was described by (1954) under the name Daviesina.
The distinguishing characteristics of the genus Miscellanoides are defined in the following diagnosis: test nummulitoid in general aspect, but asymmetric, completely involute on one side, one or more outer whorls evolute on the other. The microstructure of the test is similar to that of Miscellanea, with a finely prismatic spiral lamina. The test being both involute and evolute causes it to be asymmetric (Convex on the involute side, concave or flat on the evolute side). The Miscellanea-like microstructure is, at least in the genotype, more clearly expressed on the convex side. As in Miscellanea there is no marginal cord, but it differs in that it there are no septal filaments.
Miscellanoides bramkampi n.sp., Form B
Pl.
2 ,
figs. 1-2, 5-7, 14,
16
1952. — Miscellanoides bramkampi n. gen. et sp., Forme B, , p. 140-142, Pl. VIII, figs. 1-2, 5, 7, 9, 17-18.
Test lenticular, asymmetric (younger specimens are more strongly asymmetric), one side slightly concave, rarely convex, the other always convex. The dimensions of fifteen presumably adult specimens ranges between 2.9/0.7 to 4.0/1.3, averaging 3.2/1.0. In the central region of the convex face there are 8-10 large granules that in some individuals form a poorly defined spiral. They may attain 0.25 mm in diameter and rise about 0.1 mm above the surface, although in the largest specimens they are considerably lower. In general ornamentation is less conspicuous on large individuals than on the smaller ones. Smaller granules (0.8 to 0.10 mm) continue the spiral from the polar area, completing up to a half or three quarters of a full circle or even more in small specimens, thus marking the junction between whorls. The granules may be elongated radially, and in any event there diameter decreases as the spire continues toward the periphery. Finally they cannot be distinguished as such from the small granules that cover the remainder of the convex surface. These abundant small granules are about 0.03 mm in diameter, and seem to be distributed radially, at least in certain specimens. If the granules are slightly larger in some of these radial files, the surface of the test may appear to be grooved.
The polar region of the flatter face has 12 to 15 projecting, slightly asymmetric granules elongated in the direction of coiling and succeeded by progressively smaller granules also arranged spirally at whorl junctions for a total of 20 to 25. In many individuals here is no other ornamentation on this face, but in others radial lines of small granules mark the position of the septa. The periphery is undulate and quite sharp, particularly in large specimens. In general large granules are less well developed but surface granulation is more marked in the earlier representatives of this species.
The spire is operculiniform (w/d : 3.0/2.8, 3.3/4.2), somewhat irregular, elongated in one axial direction. The spiral lamina is thin. A microstructure similar to that of Miscellanea is visible in sections made slightly off center on the convex side, but never on the concave side. Rather thin septa are numerous, 19 in the second whorl (s/w : 4-5/2), 30-32 in the third whorl (s/w : 7-8/3, 12-14/4). The septa are irregularly spaced, curved backward to 30 or 40 degrees in the upper third of their length; they are almost straight in some specimens. The step is regular, but in the outermost whorl the chambers may be up to five times as high as long. The roof the chambers appears rounded, the upper posterior angle more or less acute.
The axial section shows the spiral lamina to be finely prismatic as in Miscellanea. The big granules and papillae of the two sides seem to be coarse prismatic pillars. The central ones continue inward to the first whorl, but the smaller ones occupy only one whorl. The axial section shows again that that the test is thicker on the convex side, although the heavy pillars tend to obscure this feature, and that the successive whorls are involute. On the flatter more concave side, the last whorl covers only a portion of the preceding one as in Operculina. The first and second whorls are involute on both sides, but the spiral lamina of the third whorl is thinner on the flat side and overlaps only partially the preceding whorl, with pillars formed commonly where they join, so the surface of the second pillar is visible. Sometimes a lamella of the third whorl may overlap the second but is not a common occurrence.
Miscellanoides bramkampi n.sp., Form A
Pl.
2 ,
figs. 3-4, 8-13,
15
1952. — Miscellanoides bramkampi n. gen. et sp., Forme A, , p. 143-144, Pl. VIII, figs. 3-4, 6, 8, 10-16.
Test lenticular, markedly asymmetric, one face strongly convex, the other either slightly concave, or more commonly weakly convex. The d/t relationships of 20 specimens range from 1.6/0.8 to 3.4/1.5, as an average 2.6/1.1. Note that comparatively the megalospheric generation is thicker than the microspheric. In large part this is due to the strong development of granules on both faces. In the central region of the convex face is a series of granules arranged in a poorly defined spiral. These granules attain 0.25 mm in diameter and rise above the surface of the test up to 0.1 mm. In small specimens the central pillar is larger. Smaller granules (0.05 to 0.08 mm in diameter start from the central region as a spiral following the boundary between whorls. They make up from a half to three-fourths of a complete circle on each whorl, even more on small specimens. They may be stretched out radially but become progressively smaller as they approach the periphery. In a number of specimens the remainder of the convex surface is marked by low radial ridges that give the surface a grooved appearance. In others the surface is less strongly marked by rows of granules less than 0.03 mm in diameter, but they too are arranged radially.
In the polar area of the flatter face is a group of 5 to 12 strongly projecting papillae, the largest generally over the proloculum, the others in a spiral along the junction of the whorls. They are succeeded by granules following the same course and decreasing in size, up to a total of 20 to 25. The papillae and granules may fuse to form a more or less continuous coating. The papillae rise up to 0.02 mm above the test surface and there mean diameter is 0.17 mm. Many are elongated in the direction of coiling. The rest of this face is free of ornamentation in most specimens, but a few have radial lines mark the trace of the septa. The periphery may be more or less rounded or sharp.
The proloculum is bilocular. Commonly the two chambers form a slightly flattened oval. The second is slightly larger with thick walls. The largest diameter of the first chamber ranges from 0.83 to 1.2 mm and the smallest from two thirds to three quarters of the other. The spire is somewhat irregular, operculiniform (w/d : 2.5/2, 3/3.2). The spiral lamina is thin. Septa are almost perpendicular at the base, in the upper third curved backward more or less abruptly to form an angle at a maximum of 30 degrees with the spiral lamina of the preceding whorl. They are spaced rather regularly: 10 to 12 in the first complete whorl, in the second 17 to 32.
The posterior superior angle is acute and the chambers may be three times as high as long in the third whorl (s/w : 3-4/1, 4-6/2). The characteristics of the axial section are like those of the microspheric generation with very large central pillars. They penetrate the interior whorls while the smaller ones occupy only one. The Miscellanea-like prismatic structure of the walls is somewhat less evident than in M. s.s.
Range: Middle Umm er Radhuma Formation, Al Hasa province.
Miscellanoides pruvosti n.sp., Form A
Pl.
3 ,
figs. 1-12
1952. — Miscellanoides pruvosti n.sp., Forme A, , p. 145-147, Pl. IX, figs. 1-12.
Test asymmetrically lenticular, varying from biconvex to plano-convex. Generally the convex side is involute and the flatter side more or less evolute. The dimensions vary between 1.3/0.5 and 3.6/1.3, averaging 2.1/0.8. The periphery is rounded but rather acutely so in most specimens. On the convex side ornamentation consists of one to five rather large granules (up to 0.2 mm diameter) situated near the center. Smaller granules arranged in a weak spiral meld with the larger ones to form radial ridges giving a channeled appearance to the surface. The flatter side also has large granules near the center, set at the junction of the whorls and decreasing in size toward the periphery. Smaller granules. Sometimes joined to form ridges mark the trace of the septa in some individuals. Ornamentation varies widely on both faces.
The proloculum is bilocular, rather thick-walled; the two chambers are both flattened ovals. The larger diameter of the first chamber ranges from 0.13 and 0.18 mm. The second chamber is slightly smaller. The spire is irregular; the step increases rapidly but not consistently (w/d : 2.1/2.6, 3.0/2.4, 2.0/1.5). The spiral lamina is rather thick, clearly prismatic in the outer whorls. The septa are straight at the base and nearly perpendicular to the spiral lamina; they may either be straight throughout or curved backward at 20 to 30 degrees in the upper half. Their spacing is somewhat irregular (s/w : 3-4/1, 4-5/2). The chambers are higher than long, their proportions varying from twice to four times as high as long.
The axial section shows the test to be thick and heavy with one or two massive polar pillars commonly extending to the proloculum, while the others are confined to one whorl. The spiral lamina of the last whorl does not ordinarily reach the central pillars on the flatter side, resembling M. bramkampi in this respect, but some specimens are involute on both sides.
Range: Upper part of the lower Umm er Radhuma Formation.
Family Rotaliidae , 1860
Genus Rotalia , 1804
Rotalia jacobi n.sp.
Pl.
3 ,
figs. 13-20
1952. — Rotalia jacobi n.sp., , p. 153-155, Pl. IX, figs. 13-15, 17-19, 21-22.
Test small, about 1.1 mm in diameter and 0.7 mm in height. The dorsal side is strongly convex. The central portion of the flat to slightly convex ventral side is occupied by a large number of tightly appressed pillars variable in shape and about 0.08 mm in diameter. The pillars are smaller near the periphery where a narrow imperforate band about 0.08 mm wide marks the edge. The pillars cover the junctions of the chambers in most specimens, but occasionally linear radial depressions mark their location. When this occurs the resemblance of the ventral side to that of Rotalia trochidiformis is striking. The aperture is a low arched slit, situated between the periphery and the center of the test. Generally it is covered by a thick prolongation of pillar-like material over the slight depression at the end of the final whorl. In a very well preserved specimen, a flattened oval opening formed by a prolongation of the pillars near the peripheral band leads to the aperture.
The spire is made up of three whorls; the proloculum is double, made up of an oval chamber about 0.08 by 0.05 mm and a second slightly larger chamber somewhat flattened; the two are bounded by a thick wall. In the peripheral whorl of a large specimen here are 10 to 12 chambers, each about 0.25 mm high, 0.24 mm long, and 0.20 mm wide. These dimensions decrease gradually toward the top of the spire. The septa curve gently back from their point of attachment in the direction of the preceding whorl. The test is coarsely perforate, the pore having a mean diameter of about 0.10 mm. The lamina is perforated everywhere, the septa not at all. The lateral walls are about 0.10 mm thick, thicker at the top where they reach 0.20 mm in the most robust specimens. In section the thick walls are clearly visible, but because of recrystallization this section gives little information concerning the inner walls of the chambers, and it was not possible to determine the manner in which the pillars and the chamber walls join. But it appears that there is no sharp break between them.
Ornamentation is variable. Its diverse forms reflect those of the variants of Lockhartia haimei. Some individuals have a vermicular network on the top, with no other ornament. In others the network is reduced in height and the sutures are limbate. Another variant with a low spire has papillae on top; these are similar in aspect to L. haimei. One variant is almost entirely without ornamentation of any kind. Another has a broad and heavy spiral bourrelet at the whorl junctions, the sutures being slightly depressed. All of these variants appear together and the first occurrence of the species is not above that of L. haimei. All of the characteristics of Rotalia jacobi suggest that it may be ancestral to the Lockhartia haimei group, and perhaps also, in view of the high rounded spire and large pores, to L. retiata.
Range: A few meters at the base of the Umm er Radhuma Formation concurrent with the appearance of L. haimei var. vermiculata.
Genus Lockhartia , 1932
Characteristics of the genus Lockhartia: The genus Lockhartia was established by (1932, p. 406-407) to satisfy the need for a change in the classification of rotalid foraminifera to accommodate those forms that resemble the genus Dictyoconoides but do not develop intercalary whorls in the spire.
defines the genus as follows: (4) "The test is calcareous, tubulated, trochoid, varying in outline from a low arch to convex on one side and flat on the other. Like Dictyoconoides, in the umbilical region where the tightly spaced pillars are in contact with the walls of the chambers, the spire is sharply differentiated. Umbilical pillars are continuous as in Dictyoconoides and cross the successive layers."
(4) The quoted paragraphs are from the Transactions of the Royal Society of Edinburgh, vol. 57, pt. 2, n° 13 (1932). (1954) modifies the definitions of Rotalia and Lockhartia.
Other characteristics shared with Dictyoconoides:
"All of the whorls are visible on the upper surface and only the terminal ends of the whorls are visible round the periphery on the lower surface. The whorls are multilocular, the chambers being separated from each other by almost straight double septa, each pierced by a single oblong to half-moon slot where it abuts against the wall of the preceding whorl. (...) The chamber walls are partially embracing (as in the Assilines) on the dorsal surface, but are open toward the umbilicus where their terminal ends are claw-shaped and recurved upward. Lateral structures which seem to take the form of irregular filaments of shell material are developed on both surfaces of the test. On the upper surface they are best developed at the pole, where they serve to cover the protoconch fairly thickly, but they apparently cease to be developed on the outer whorls. On the lower surface they continue regularly to be developed to the end of growth."
differentiates Rotalia from Dictyoconoides and Lockhartia by four distinctive structural characteristics:
The whorls are typically enveloping on the dorsal face whereas in Lockhartia they overlap only slightly.
The lower wall of the chamber develops transverse folds that result in depressions on the ventral surface that separate the umbilical part of each chamber from the peripheral portion, thus forming "astral lobes".
Umbilical pillars are discontinuous, while in Lockhartia and Dictyoconoides they are usually continuous.
The lower walls of the chambers extend inward toward the center of the test and in the early whorls may even cross the umbilicus, so that there is an intermediate region between the umbilicus and the spire, while in Lockhartia and Dictyoconoides the two are sharply separated.
Using these criteria (1932) distinguished four species of Lockhartia: the genotype L. haimei () and L. tipperi () both typical of the genus, then L. conditi () and L. newboldi (d' & ). In one of his discussions seems to doubt in some measure that the last mentioned has all the characteristics of Lockhartia; indicates that the species is only a variant of Rotalia trochidiformis . In any case, L. conditi differs from the genotype in that the pillars seem to join the chamber walls instead of ending sharply; the umbilical region is filled with pillars larger and more closely appressed than in other Lockhartia. In addition the dorsal surface is completely unornamented.
Most of the following discussion is based on the examination of cuttings, but the stratigraphic conclusions are supported by the findings derived from core and outcrop. The suggested lineages of species and varieties are obviously only interpretations, for they are based solely on variations in the form and ornamentation of tests. However, the interpretations are the result of the study of a large number of specimens. The stratigraphic levels of maximum abundance and the first appearance of each species and variety are facts and the evolutionary lines suggested are in accordance with these facts.
The oldest species of Lockhartia in the Umm er Radhuma is found in strata just above the lower limit of the formation. It appears to be related to L. haimei. The general form, the relatively flat ventral surface, the moderately convex dorsal surface, the coarsely porous texture, the tendency toward the development of an imperforate bourrelet at the junction of the dorsal and ventral walls of the chambers of the last whorl, the large number of small pillars that appear on the ventral surface, the shape of the chambers, all of these are in common with L. haimei. Because this variant has value as an indicator of stratigraphic position it has been named L. haimei var. vermiculata in accordance with the nature of the dorsal ornamentation.
This ornamentation is characteristic: thin walls encircle the large pores of the dorsal surface; they may be thickened and thus provide considerable relief on the test surface, and in some specimens form complex networks. This ornamentation varies greatly in detail. In many individuals the thickening of the walls is confined to the central part of the dorsal surface; in others it is covered completely. In some specimens these thickened walls coalesce; they form grains, elongated granules and U-shaped lines, thus presaging the more precisely defined patterns seen in slightly younger variants, while in others there is no coalescence and the thickenings are intricately interlaced.
Slightly above the succession in which L. haimei var. vermiculata is predominant another variety appears very abundantly. It appears to have developed from one of several less well-defined variants for intermediaries exist in an intermediate stratigraphic position. In this variety the mean diameter is greater than that of the preceding dominant variant although the proportions of the test remain about the same. The name L. haimei var. nudimarginata is proposed for this variety, easily recognizable from its dorsal ornamentation confined to the summit. The pores of the outer whorls thus open to view are smaller than those of the vermiculata variant and are not encircled. In most specimens ornamentation consists of granules or comma-shaped granules, or in U-shaped or elongate lines that tend toward a radial disposition. In some individuals the sutures are limbate, have some relief, and are imperforate.
This variety appears to have been the source of many others. One appears just above its level of its maximum abundance. It is called L. haimei var. suturadicata because the whorl junctions are marked by a thick imperforate bourrelet. In most individuals ornament is much more regular and uniform than in the varieties preceding it in dominance. Usually it is a series of elongate granules in the summit area, arranged more or less radially and attached to the spiral bourrelet. All ornamentation has a low relief. Almost all of the representatives of this variant are large in diameter and have a low spire. In its turn this variant gives way rapidly to others that are considered to lie within the range of variation of L. haimei s.s.
Although extremely polymorphic, L. haimei can be can be distinguished because of certain constant characters. Both elongated and round granules are confined to the central part of the dorsal surface, where, commonly, they are linked to the spiral bourrelet and form a radiating design. All ornamentation is heavy, coalescent and in high relief. In an extreme case it is 12 to 15 rounded and discrete papillae ranged on the spiral suture that is faintly marked by a narrow bourrelet.
Individuals with ornamentation identical to that of the holotype figured by (1932, Pl. II, fig. 5) are infrequent and not much more common are specimens like those figured in his paper of 1937 (Pl. III, figs. 9, 11-12). Those that came from an argillaceous marl are most similar to those figured by .
Thus the type of Lockhartia haimei represents but a small part of the variations grouped under that name here; nevertheless, all are easily recognized by the coarse and heavy ornamentation, not easily confounded with those described above. Many of these variations are in the direction of higher spires and the average size of these specimens is slightly greater than that of other variants of L. haimei, and they are also distinguishable through the presence of a very heavy bourrelet.
These higher-spired forms occur only in the upper portion of the levels occupied by L. haimei and, because of this restricted range have been named: L. haimei var. spirachordata.
All of the many variations here included under L. haimei occur only in the lower half of the Umm er Radhuma Formation. The lower limit of occurrence of all named varieties is not known precisely, but the upper limit is reasonably well established. Their usefulness is demonstrated by the fact that the sequence of their occurrence was found to be the same in areas 200 km apart. The sketch
(Fig. 7 ) suggests possible relationships among the variants and their relative abundance at a given level, not only for L. haimei but also for the other species of Lockhartia found in the Umm er Radhuma Formation.
A little above the level of the appearance of L. haimei s.s. another species of Lockhartia occurs, its range nearly the same. This is L. retiata, described below. The variation of this species with the longest range appears first and is easily recognized by its high rounded dorsal contour and its thick reticulate network around the pores, hence the name. The summit area is covered by thick raised and coalescent striae that may form papillae or interwoven bars. As in L. haimei, in the outer whorls of some variants the network around the pores is low and the pores themselves are smaller. In other variants with a tendency to finer, less gross ornament, the sutures between whorls are marked by narrow, imperforate raised striae, but a true bourrelet occurs only in those high-spired forms referred to the genus Sakesaria.
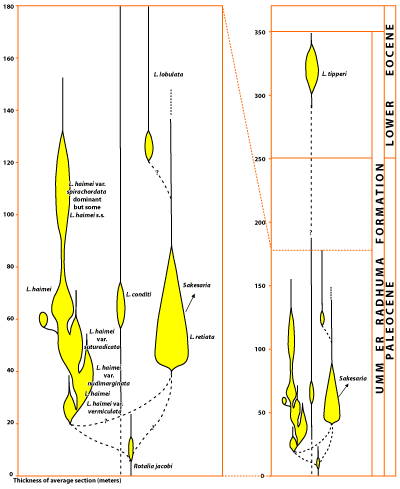
Click on thumbnail to enlarge the image.
Figure 7: Stratigraphic position and relative abundance of species of Lockhartia in the Umm er Radhuma Fm.
Another species with a slightly lobulate rather than a circular or weakly ovate periphery and with ornamentation reduced in importance is named Lockhartia lobulata nov. sp. Its range extends above those of L. haimei and L. retiata and it differs from them in several other significant features.
Lockhartia conditi () has a long range — approximately equivalent to that of L. haimei s.s. and that of the named variety L. haimei var. spirachordata. It has not been found higher in Arabia although (1937) reports that it exists throughout the Laki Formation of northwest India. Certain variations suggest that this species is ancestral to L. tipperi, however.
Lockhartia tipperi s.s. occurs only in the upper third of the Umm er Radhuma Formation, with its maximum of abundance in the portion considered to be of early Eocene age. Certain rare variants found lower stratigraphically are perhaps primitive representatives of the species, but their relationships to the typical form are obscure.
In summary: The species of the genus Lockhartia found in the Umm er Radhuma Formation include Lockhartia haimei (), with four newly named varieties; L. conditi (); L. tipperi (); L. retiata n.sp., and L. lobulata n.sp. Representatives of "Lockhartia newboldi" (d' & ) are present also, but here they are considered to be large and heavy-shelled variant of Rotalia trochidiformis .
Lockhartia haimei (, 1927)
Pl.
5 ,
figs. 1-37
1927. — Dictyoconoides haimei , p. 280; Pl. 21, figs. 13-15; Pl. 22, fig. 6.
1932. — Lockhartia haimei (), , p. 407, Pl. II, figs. 4-6.
1937. — Lockhartia haimei (), & , p. 45-46, Pl. VII, figs. 9, 13, 15.
1952. — Lockhartia haimei , , p. 167-170, Pl. XI, figs. 1-38.
The type of Lockhartia haimei comes from the upper Ranikot Formation of India. It is obvious, according to his descriptions and figures, that saw only a small part of the very extensive variation in form and ornamentation among specimens referred to this species in Saudi Arabia. Four abundant varieties with a restricted vertical range have been added to the type as described by . Furthermore there are other variants that might have been cited if it had been possible to determine the limits of their ranges. The named varieties are L. haimei var. vermiculata n.var., L. haimei var. nudimarginata n.var., L. haimei var. suturadicata n.var., and L. haimei var. spirachordata n.var.
L. haimei s.s. and its varieties are distinguishable by differences in ornamentation, and some have discrete mean height/diameter parameters. The adult test of L. haimei is fairly large in the gamut of forms taken to comprise the species. In fifty specimens, the average d/h is 1.39/0.82 or 1.70. The relationship between the dimensions of fifty other specimens, taken a little higher stratigraphically is 1.61/0.90 or 1.79. Higher levels show more variation in size. They range from 1.1 to 2.1 mm in diameter and from 0.55 to 1.45 mm in height while those of the lower group are less variable ranging from d/h of 1.1/0.5 to 1.8/1.1. The dorsal face is convex, the ventral one flat, or slightly convex. ' description of internal structure agrees with that of the Arabian specimens, except that in Arabia some individuals are more inflated (e.g. L. haimei var. spirachordata). The large bilocular proloculum is followed by two and a half to three whorls made up of chambers which increase in size gradually. The other characters of the spire are similar to those figured by (1937). The ventral face is covered by pillars, the largest about 0.1 mm wide, but the length and degree of appression vary; they decrease in size toward the periphery which is marked by an imperforate bourrelet that extends onto the ventral surface for about 0.1 mm. The aperture is not visible, probably partly covered by one or more of the pillars, but in a few specimens a low slit exists, almost covered by pillars, at the base of the outer face of the last whorl near the periphery. This may be the aperture.
An invariable part of the dorsal ornamentation of Lockhartia haimei s.s. is an imperforate bourrelet at the junction of the whorls. It may or may not be raised above the surface. At the apex of the spire are papillae or radial bars commonly with a tendency toward coalescence. They are thickened, arranged like beads, and commonly form a spiral. This occurs because most of the elements of the ornamentation are sited where septa are in contact with the preceding whorl. However, this spiral arrangement may be unrecognizable. In almost all variants the trace of the septa is indicated by a thin band of imperforate material.
The variations considered to fall within the bounds of L. haimei s.s. are as follows:
All of these varieties as well as the form chosen by as the type of the genus are linked by intermediates, and all exist at the same larger stratigraphic level.
The variations of Lockhartia haimei to which names have been assigned have been found to have a restricted range and thus can be used as markers. They are discussed below in the order of their appearance starting with the oldest.
Lockhartia haimei var. vermiculata n.var.
Pl.
5 ,
figs. 10-15
1952. — Lockhartia haimei var. vermiculata nov., , p. 170-171, Pl. XI, figs. 10-15.
This is the earliest form of Lockhartia haimei to appear. It is found in the lowest fifth of the Umm er Radhuma Formation. It is never very abundant. Its dorsal ornamentation is the key to its recognition. The test is of moderate size. The relationship between dimensions of ten specimens, some immature, varies from the average being 1.32/0.78 or a ratio of 1.69. If a larger number of adults had been available the mean size would have increased. In general, the spire is fairly low, the convex base compensating in part for a low diameter/height ratio. Dorsal ornamentation is a complex series of narrow interlaced ridges around the pores, rising only slightly above the general surface (0.05 mm). The ridges may be coarse, sometimes thickened and coalescent giving the impression of worn-holed wood. This ornamentation may extend over the whole of the dorsal surface, or be confined to the first two whorls. If so, the pores of the last two whorls are encircled by even lower ridges. Occasionally the coalescence of the ornament near the summit is carried so far that the ridges appear as rather long broken lines irregular in their course. The pores average 15 microns in diameter, but vary considerably. Note that there is no spiral lamina, or any thickenings where it might have been present.
Lockhartia haimei var. nudimarginata n.var.
Pl.
5 ,
figs. 16-21,
35-36
1952. — Lockhartia haimei var. nudimarginata nov., , p. 171-172, Pl. XI, figs. 16-21.
Lockhartia haimei var. nudimarginata appears near the upper limit of the range of L. haimei var. vermiculata and persists for about twenty meters; the precise figure depends on the total thickness of the formation in any one locality, but in any event its duration in time was limited.
The dimensions of 25 adult specimens ranged from 1.2/0.6 to 2.3/1.3, the average being 1.68/0.82 or 2.04. Thus this variety has a lower spire than the type but the difference may not be appreciable if the whole population were included. The distinctive characters of this variety are:
Lockhartia haimei var. suturadicata n.var.
Pl.
5 ,
figs. 22-24, 37
1952. — Lockhartia haimei var. suturadicata nov., , p. 172-174, Pl. XI, figs. 22-25.
Lockhartia haimei var. suturadicata appears at a level near the upper limit of the range of L. haimei var. nudimarginata. It seems clear that it is ancestral to L. haimei s.s. for the two have many characteristics in common, especially ornamentation. The type species first occurs near the upper limit of the range of this variety.
The average of the diameter/height relationships is 1.64/0.77 or 2.13. The range of sizes is from 1.3/0.6 to 2.1/1.1. In most individuals the base is flat; in a few it is slightly convex. In addition to some peculiarities in ornamentation this variety has a low dorsal contour. The dorsal surface rises abruptly from the periphery, but is flattened at the summit as may be seen from the mean diameter/height ratio — greater than either of those described previously, or that of the species s.s. as it is defined here.
Ornamentation is distinct from that of the two varieties already described. It consists of a spiral bourrelet at the whorl junctions and limbate sutures on the septa. The remainder of the ornamentation is a few slightly elongate and flattened granules near the summit or a small number of papillae in the same area located where septa meet whorl junctions. In this arrangement of granules and papillae the variety approaches a weakly ornamented L. haimei s.s. but is clearly differentiated by the flatter dorsal contour and the less prominent ornamentation.
L. haimei s.s. occurs with this variety in the upper part of its range. Higher in the sequence the genotype becomes very abundant in most of the rest of the lower half of the Umm er Radhuma Formation. Most of the extreme variants of L. haimei s.s. given letter designations occur only in lower levels, while forms more like the designated holotype predominate higher in the succession. Only one variant, probably derived from the variant "C" mentioned above is as common as the type form and is even more abundant in the higher part of the occurrence. It is L. haimei var. spirachordata.
Lockhartia haimei var. spirachordata n.var.
Pl.
5 ,
figs. 23-30
1952. — Lockhartia haimei var. spirachordata nov., , p. 174-175, Pl. XI, figs. 26-31.
L. haimei var. spirachordata is recognizable at once because if its relatively high and swollen spire and one aspect of its ornamentation: a thick, wide and continuous spiral lamina. Measurements on fifty specimens showed the range of size to be 1.1/0.6 to 2.6/1.6 with an average of 1.67/1.04 or 1.60. Commonly the base is almost flat, only rarely strongly convex; Normally the dorsal face is arched and full and the ornamentation is a wide spiral lamina, slightly raised in the early whorls, less so in the outer whorls. A few elongated granules and low papillae mark the summit area. Most sutures are limbate This variety includes the largest representatives of the species.
All variations included, L. haimei ranges in size from 1.0/0.55 to 2.6/1.6. The mean d/h ratio for 216 specimens is thus 1.84, indicating that the average arching of the spire of the entire species is slightly less than its average in L. haimei s.s.
This species of Lockhartia has been reported only in India and in Arabia, although other species present in the upper portion of its range have a broader distribution. L. conditi and L. tipperi are among these species, for both are reported from British Somaliland. In any case the range of L. haimei s.s. appears to be restricted to equivalents of the Montian and Thanetian stages.
Lockhartia retiata n.sp.
Pl.
4 ,
figs. 1-21
1952. — Lockhartia retiata n.sp., , p. 179-183, Pl. X, figs. 1-22.
Lockhartia retiata appears at about the same level as L. haimei s.s. It is entirely possible that it has the same lineage as L. haimei, either through L. haimei var. vermiculata or directly from Rotalia jacobi. Some of the variants of L. haimei var. vermiculata have a relatively high spire, and if they had been slightly more rounded and fuller would be almost impossible to distinguish externally from L. retiata.
The character of the test that defines this species is a network of raised imperforate ridges around the pores. Where not complicated by thickening or coalescence, most commonly in the outer whorl, the low ridges form a pentagonal or rectangular rim around each pore. But the net is subject to great modification in several of the varieties described below. The d/h dimensions of fifty individuals ranged between 1.1/0.8 and 1.9/1.7, averaging 1.49/1.15 or 1.29. The ventral face is flat to weakly convex. It is occupied by numerous pillars of uniform size. Most are irregularly rounded and appressed; most ranging between 0.05 and 0.08 mm in diameter, rarely larger; and a number are malformed, the exposed contour drawn out into a comma or U shape. The periphery is marked by an imperforate bourrelet, somewhat irregular in width, ranging in most specimens between 0.1 and 0.2 mm. The edge of the bourrelet on the dorsal side does not extend toward the apex uniformly, thus differing from L. haimei where the bourrelet is more or less uniform in width everywhere. In all of the specimens examined no aperture could be found.
The dorsal face of the specimen selected as the type is a high arch, with the portion near the periphery inclined inward from the vertical 15 to 20 degrees; in some specimens the dorsal face begins with a curvature toward the exterior before swinging inward. In the inner whorls the sharply defined ridges around the pores become thickened, some coalescent into papillae or interlaced in complicated patterns. The greater the amount of coalescence the simpler the pattern of ornamentation becomes. In some specimens this coalescence affects almost all of the dorsal surface, but commonly the outer whorl is not involved.
The proloculum of L. retiata is double, large in all of the examples sectioned. The first chamber is almost globular, from 0.12 to 0.20 mm in diameter; the second is 0.12 to 0.15 mm, and may be slightly teardrop-shaped upward. The dorsal wall is thickened and pierced by tubes up to 18 microns in diameter; they are separated by intervals about as wide as the tubes, occasionally even greater. The average thickness of the wall is about 0.20 mm, but in the final whorl may be reduced to about 0.05, which suggests that the thickening occurs after the wall is first constructed.
The ventral wall of each whorl is imperforate and is slightly thinner than the outer wall. The wall is flattened on the inner side and a small hook-shaped process extends dorsally, undoubtedly touching the base of the preceding whorl, which itself may have a downward extension not always discernible in the sections. The inner portion of the spire is filled by a mass of test material which appears to be mainly solid pillars elongated more or less vertically and more numerous near the base. Mixed with these pillars are masses of test material generally somewhat plate-like, arranged at various angles with many more or less horizontal. The whole assemblage is clearly discrete from the spire itself. In one section the granules on the ventral surface appear to be hollow, but this may be only a stage of growth.
The septa are rather thick, curved backward dorsally, thus forming an angle of about 35 degrees with the spire; communication between chambers is through an elongate opening near the inner edge of the septum of which the contour follows that of the outer wall of the preceding whorl. In the last whorl this opening is about 0.16 mm long and 0.05 mm wide, sometimes larger at the base and thus triangular. Opposite these openings that permit communication between chambers is a smaller oval to round hole that opens on the central pillar-filled mass. The greater diameter of this opening is from 0.030 to 0.050 mm for it may be elongated vertically.
The chambers of the last whorl are 0.3 to 0.4 mm wide and 0.4 to 0.7 mm high varying according to the size and complexity of the specimen. The chambers of the inner whorls are progressively smaller toward the proloculum. The reduction in size may be due in part to the addition of material to the inner surface of the outer wall as growth proceeds.
The spire is regular: 6 to 7 chambers in the first whorl, 11 to 12 in the second and 12 to 15 in the third. The whorls overlap more or less in concordance with the height of the spire. High-spired specimens may have four whorls, but the average is three and a half.
Variation in several directions took place in this species as in all of the other forms of Lockhartia. In one variety ornamentation is reduced to a comparatively simple arrangement: thick papillae and bars at the center; an intermediate zone on the second whorl of fine ridges around the pores and the third whorl with smaller pores and thin, imperforate lace-like sutures marking the position of the septa. In this group the spire remains as it is in the type. In a second variant the last whorl is unornamented except in a few specimens that are weakly limbate and the pores are large (up to 18 microns). In both of these the central ornamentation is highly coalescent and strong, often with a spiral design. At the same time the test elongates, the first and second whorls being "pulled out" so that the arch of the dorsal surface becomes more acutely angled. There is a tendency toward the development of a spiral lamina which is thin and discontinuous but thickened into nodules where the septa meet the edge of the preceding whorl. At the same time the thin walls around the pores become prominent in the first two whorls and project even more in the third. With an elongation of the spire a little more marked and with several whorls added in the adult, this variety would become Sakesaria nodulifera n.sp.
In still another variation, all of the summit ornament is coalescent. The spire elongates somewhat, but in this form the coalescence is expressed as a spiral lamina marked with ridges and papillae, forming a more or less complex ornamentation. All is in high relief, and in the most diverse form a trellis made up of small ridges appears between the heavy spiral lamina. With secondary ornamentation reduced, together with an increase in the prominence of the spiral lamina at the whorl junctions and a slight augmentation of height in the spire (still rounded) would lead to Sakesaria tereta.
Probably Sakesaria trichilata developed from a variant of this type, with the secondary ornamentation a heavier trellis and stronger horizontal bars, but the intermediate forms that would confirm this hypothesis were not found.
Range: Lower half of the Umm er Radhuma Formation.
Lockhartia lobulata n.sp.
Pl.
3 ,
figs. 21-27
1952. — Lockhartia lobulata n.sp., , p. 184-185, Pl. X, figs. 23-29.
This species may have been derived from a source common to it and L. haimei, but the two share no characteristics except for similar ornamentation in the central portion of the dorsal region. The same remark can be made regarding L. retiata, but here the relationship is closer because of the existence of large pores in both. The geographic distribution of this species is restricted and only a few specimens were available for study, although it is fairly common in some localities. In eight individuals the d/h relationship ranged from 1.2/0.7 to 1.7/1.0, averaging 1.35/0.82 or 1.64. The specimens to hand have three and a half to four whorls with 14 to 15 chambers in the outer whorl in the larger specimens. The larger examples are all microspheric and in general their ornamentation is more complex than that of the smaller ones most of which are megalospheric (from a study of Libyan representatives of the species). The ventral surface is weakly concave in most individuals, the central portion covered by round granules rather widely separated and ranging up to 0.1 mm in diameter. Some of the granules are smaller near the periphery and the area adjacent to it is free of them. This peripheral region differs greatly from that of L. haimei and L. retiata in that there is no imperforate bourrelet; in this absence it resembles L. tipperi.
The dorsal surface is arched, but the flanks are almost flat so that the summit is angled rather than rounded. However, the junction of the dorsal and ventral surfaces is a well-rounded curve. Ornamentation near the summit is a series of low radial ridges and papillae, on the whole somewhat irregular and coalescent. The remainder of the dorsal surface with the exception of the last whorl is covered by a coarse rectangular or pentagonal, slightly raised net of imperforate material grosso modo arranged like a honeycomb, the spaces between the meshes ranging up to 0.07 mm in the greater dimension. The outer whorl has a finer net around each pore, about 0.02 mm maximum diameter. Sutures in the outer whorls are marked by a slight depression and an increase in the size of the meshes, thus resembling somewhat the lacelike ridges on the sutures of some variants of L. retiata. In most specimens the depression at the sutures gives a weakly lobate aspect to the periphery, hence the name.
A single axial section shows that the dorsal and the ventral walls of the chambers are pierced by tubes. With only a few specimens to hand it was not prudent to attempt more sections. The dorsal wall is thinner in the outer whorl and the ventral wall is thickened. A proloculum is lacking but the form of the test suggests that it must be large (5). The central mass of pillars is not clearly defined, the most that can be said is that it seems more ordered than that of L. retiata. The chambers are clearly separate from the central cavity.
(5) Libyan specimens show the proloculum to be "double" in the megalospheric form and very large, each chamber 0.2 to 0. 27 mm in diameter.
Range: The upper part of the lower Umm er Radhuma Formation.
Lockhartia tipperi (), B and A Forms
Pl.
4 , figs. 25-26,
33-36
1926. — Conulites tipperi n.sp., , p. 247-248, Pl. 18, fig. 8.
1931. — Dictyoconoides tipperi (), & , p. 56-57, Pl. III, figs. 14-17.
1932. — Lockhartia tipperi (), , p. 407.
1937. — Lockhartia tipperi (), & , p. 48-49, Pl. VI, figs. 14-16; Pl. VII, fig. 12.
1952. — Lockhartia tipperi () Forme A, , p. 186-189, Pl. XI, figs. 47-51.
1952. — Lockhartia tipperi () Forme B, , p. 186-189, Pl. XI, figs. 52-53.
This rather large species is highly polymorphic, as much in form as in structure and ornamentation. Form A specimens are always strongly biconvex, but Form B may be flattened on both dorsal and ventral faces. Including both forms, the diameter ranges between 0.7 and 3.8 mm, thickness from 0.4 to 2.2 mm. Form B appears to have a mean d/h of about 2.4/1.38 an average slightly less than that seen in India and British Somaliland. Near the upper limit of the range of this species which is that of the Umm er Radhuma Formation, the A Form predominates
A specimen 2.6 mm in diameter has six whorls; larger ones have 8 or 9 whorls. The step increases rapidly; chamber height is 0.06 mm in the first whorl, 0.22 in the third whorl, but slows thereafter: 0.24 in the fifth and from 0.26 to 0.28 in the final whorls. In the sixth whorl of the specimen discussed there are 20 chambers, each about 0.4 mm long, 0.26 mm high and 0.52 mm wide; the spiral lamina is about 0.045 mm wide in the outer whorls, less in the inner ones.
The dorsal surface is ornamented. There are a variable number of rounded papillae from 0.06 to 0.18 mm in diameter arranged along whorl junctions, so they form a spiral, apparent only in some specimens because the papillae are so large.
The papillae of Form B are higher than those of the megalospheric form and may attain a relief of 0.15 mm. They are present only rarely on the outer whorls of the adult. In general they occupy only half the diameter of the test; in the A Form they occupy three-quarters or more of the test surface. In some specimens near the bottom of the range there are striae extending from the papillae over the septa. Most of these early representatives have small papillae (0.06 mm) so their arrangement in a spiral is evident.
The ventral face is always convex, more strongly so in the A Form. The ventral surface of the chambers of the last whorl forms a marginal belt around a circular central area filled with pillars. This belt occupies more than half of the radius in small specimens, but only a tenth of it on very large individuals. The pillars vary widely in outline; rounded, rectangular, oblong, square, U-shaped. Their major axis is between 0.10 and 0.20 mm in length, slightly less near the marginal belt.
A meridian section reveals the simple nature of the spire: whorls are added almost without overlap on the dorsal side and on the ventral side are clearly separate from the umbilical region, filled with pillars and transverse layers, together making up a compact mass.
In accordance with a remark made by M. J. the chambers of small and large specimens were investigated in more detail; it was evident, as indicated above, that Lockhartia tipperi has both megalospheric and microspheric forms, a fact unknown previously. The proloculum of Form B is small, round unilocular, the mean diameter less than 0.04 mm, whereas the proloculum of Form A is "double", relatively large, the two chambers together having a diameter of about 0.12 mm. Taken separately the first chamber attains 0.06 mm, the second may be slightly the larger in some specimens.
The two forms can be distinguished without recourse to sectioning. The mean diameter of twenty specimens of Form B taken from one 2 meter sequence is 2.39 mm, the height 1.73. In actual size the d/h ranged from 1.8/0.9 to 3.0/2.0. Form A which is much smaller and slightly higher in proportions and with larger granules on the dorsal face, the d/h of twenty specimens from the same locality varied from 0.9/0.7 to 1.7/1.2, averaging 1.41/0.95 or 1.48, thus almost a millimeter smaller in diameter and a half millimeter in height than Form B and averaging 0.25 less in ratio, thus demonstrating that the megalospheric generation is proportionately thicker than the microspheric one.
Range: Upper Umm er Radhuma Formation, upper lower Eocene, equivalent in age to the lower Laki Formation of India.
Genus Dictyoconoides , 1925b
Dictyconoides kohaticus (), Form B
1861. — Conulites cooki (pars) , Pl. XV, Figs. 7, 7-7a-b-7d, 7f-g.
1924. — Conulites kohaticus , p. 223.
1925b. — Dictyoconoides cooki (), , p. 387.
1926. — Conulites kohaticus , p. 240-246, Pl. 16, figs. 1-6.
1930. — Rotalia kohaticus (), C. van , Pl. 135.
1930. — Dictyoconoides kohaticus (), , p. 73-74, Pl. X, fig. 5.
1931. — Dictyoconoides kohaticus (), & , p. 54-56, Pl. III, figs. 9-13.
1932. — Dictyoconoides kohaticus (), , p. 405-406, Pl. I, figs. 2, 5, 8, 10, 11, 12 (less c and e) 13, 14; Pl. II, fig. 1.
1952. — Dictyoconoides kohaticus () Forme B, , p. 190-191, Pl. XII, fig. 31.
Dictyoconoides kohaticus is well represented in Arabian deposits; it is very abundant in certain outcrops of the lower Dammam Formation, somewhat less so in the upper portion. Arabian specimens are varied in dimension and proportions, the largest found being very flat and thin with a d/h of 14.5/2.1. In general the diameter is about a centimeter and the height around 2.0 mm. In some variants that appear relatively high in the succession the dorsal surface is covered with large granules, more numerous near the apex and somewhat elongated radially, while in others at the same levels an unweathered dorsum is smooth. Perhaps the granulose specimens should have been named because the test seems to be slightly thicker than that of the typical forms, but in the individuals seen there are no other distinctive characteristics.
Most specimens are not distinguishable from the type described by . Their description need not be repeated here.
Dictyoconoides kohaticus (), Form A nov.
Pl.
4 ,
figs. 22-24
1952. — Dictyoconoides kohaticus Forme A nov., , p. 192-193, Pl. XII, figs. 32-35.
This form may occur with the microspheric generation, but is often found alone. The test is conical, with a slightly rounded base. The d/h ratio ranges between 1.4/0.9 and 2.0/1.3, with the average 1.7/1.1. Some specimens occurring with the granulose microspheric form are ornamented with irregularly rounded granules; commonly they are near the apex, but continue down the slope a variable distance; they attain 0.2 mm, but most are smaller; the largest granules are over the proloculum; a few individuals have only one or two in that area. Some granules are elongated radially.
The central portion of the ventral face is occupied by a large number of small pillars, slightly elongated in one direction, but well separated, averaging about 0.1 mm in diameter. The ventral surface of the last whorl extends 0.2 to 0.3 mm into the ventral area forming a belt around the pillars. The peripheral region is not thickened by an imperforate band. The septa are sometimes marked by sutural depressions visible on both the dorsal and ventral faces.
The proloculum is large, 'double,' eight-shaped because the two chambers are ovals flattened on one side. The greater diameter of the first chamber ranges from 0.13 to 0.18 mm, and the second chamber is slightly larger. The proloculum walls are thick. In axial section the sectioned specimens have six whorls. The chambers of the sixth whorl are 0.33 mm high, 0.20 mm wide and 0.37 mm long. Near the apex chambers are shorter and consequently relatively higher. The inner side of the chamber wall is thickened and curves upward in a hook that joins the preceding whorl at a considerable distance from its base. At the top the dorsal wall is about 0.2 mm thick, but decreases to about 0.05 mm in the sixth whorl. It is very finely porous.
The umbilical region is occupied by ten to fifteen very large continuous pillars (visible in the section) that fill the space below the spire and are not completely discrete, for horizontal lamina unite them and the chamber walls. Like the dorsal wall, the ventral wall too is finely porous.
Range: Dammam Formation, more common in the lower portion.
Genus Sakesaria , 1937
The author's definition of the species is as follows [retranslation phraseology questionable]:
Structure similar to that of Lockhartia, from which the genus differs only in the extreme elongation of the axis, representing an excessive degree of specialization about one pole. Umbilical space much reduced, the number of umbilical pillars greatly decreased.
This definition is amplified in the reference cited ( 1937, p. 49-50) in his discussion of the genotype, Sakesaria cotteri. This species is abundant in the lower Eocene portion of the Umm er Radhuma Formation, along with a number of variants, all of which occur together. These are discussed in some detail in the description of the species as it exists in Arabia. As says, Sakesaria cotteri is clearly related to Lockhartia, but among the variants found there is no real evidence of the lineage. Perhaps this is due to an environment in Saudi Arabia unfavorable to both Lockhartia and Sakesaria (Rus Fm.) before Sakesaria could flourish there again.
Study of an abundant and well-preserved fauna demonstrated the close relationships of Lockhartia and Sakesaria. Lockhartia retiata nov. sp. includes variants that progressively approach Sakesaria. They seem to have been forerunners of two discrete species of this genus. They are described below, but it may be helpful to summarize these findings here. Illustrations of some variants are on
Pls.
4 ![]() and 5 .
and 5 .
Shortly after L. retiata appears, variants are found in which the coarse mesh around the pores does not cover all of the dorsum. Instead finer pores without a mesh exist on the final whorl. On some of these specimens the sutures become limbate with a narrow but heavy stria on each septum and in others a lacelike 'remnant' of such striae occurs.
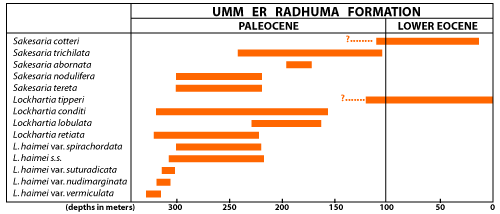
Click on thumbnail to enlarge the image.
Figure 8: Ranges of the species of Lockhartia and Sakesaria in the Umm er Radhuma Formation.
In still others the network is heavier on the last whorl and at the summit the striae tend to coalesce and to form a radial but spiral ornament as in L. haimei. This variability of ornamentation is accompanied by changes in the height of the spire, but the relationship of one to the other is slight, although most of the higher-spired forms have relatively simple ornamentation. Some specimens with a heavy, simple ornament and a relatively high spire have a rudimentary bourrelet at the junction of the last two whorls. Transitions link this variant to incontestable Sakesaria, high-spired and with a complete bourrelet on the whole length of the spiral suture. In this form, named Sakesaria nodulifera nov. sp. every whorl overlaps the preceding one slightly. An irregularly-shaped nodule at the junction of whorl and septum is always present, but its shape and size vary widely.
Another, slightly different but discrete species of Sakesaria occurs at the same levels. In this variant ornamentation is reduced. It consists only of a thickened bourrelet along with the coarse reticulation reduced in importance. In the intermediaries to this species increase in the height of the spire appears to be accompanied by a thicker bourrelet, and at the same time a reduction in other ornamentation. The resulting Sakesaria has a lower spire than S. nodulifera and is easily distinguishable at every stage of its growth. This relatively low-spired entity is named Sakesaria tereta n.sp.
Sakesaria cotteri is most abundant in the upper fourth of the Umm er Radhuma Formation and is not found in association with the two species named above. In Arabia the course of its evolution cannot be traced but is nearly certain that it, like the others, was derived from the genus Lockhartia, perhaps through an intermediate form of Sakesaria.
The form, size and ornamentation of S. cotteri vary within broad limits in the diameter / height relationship as well as in ornamentation. Creation of named varieties is not required for all are found at every level. and named variants would have no stratigraphic significance. In general large specimens (height/diameter almost 3 to 1) lose ornamentation on the last two whorls.
The oldest specimens found show the complete range of variation in the height/diameter gamut, but ornamentation is restricted to striae on the septa and there is no bourrelet. A little higher in the succession some variants have heavier striae that form ridges but in others they disappear completely. Farther up all individuals are heavily ornamented and the pattern becomes irregular (discontinuous striae and papillae), or, on the contrary form a continuous bourrelet. Near the upper limit of the species ornamentation is restricted, striae may be replaced by isolated papillae and no bourrelet is present.
Thus it is possible to get a vague impression of stratigraphic position through the dominant type of ornamentation, but the other variants are always present and the transition from one to another is gradual.
A third group of Sakesaria exists in the Paleocene-lower Eocene of coastal Saudi Arabia with a more extensive geographic distribution than those already mentioned. It appears at about the same level as Sakesaria nodulifera. It too seems to have originated with Lockhartia retiata but no intermediate forms that trace the transition.
The distinctive characters of this species are: its form grosso modo more elongate than those mentioned (Increase in height of successive whorls more pronounced); its ornamentation, which although coalescent in some specimens, is in the form of a trellis that is commonly confined to the early whorls. In many individuals the last whorl is finely porous, and in a considerable proportion of these, lacy limbate sutures are concurrent; this tendency exists also in Lockhartia retiata. The range in the proportions of the test is large.
The upper limit of this species, Sakesaria trichilata n.sp., extends into the lower part of that occupied by Sakesaria cotteri. One of its many varieties occurs in almost half of the Umm er Radhuma Formation, excluding the upper and lower fourths.
Another species of Sakesaria occurs in the Umm er Radhuma Formation. It is conical, diameter and height almost equal, and thus easily recognized by this shape and its weak ornamentation. The surface is finely perforate and all but smooth, with some low irregular nodules, perhaps traces of heavy suture delineation that may have been present in its ancestors. The spiral suture is a little depressed in most specimens. This species, Sakesaria abornata nov. sp. is restricted both geographically and in range. It appears over a few meters at the upper limit of the range of Lockhartia haimei. It has been found only rarely in the coastal portion of Al Hasa province.
Sakesaria tereta n.sp.
Pl.
5 ,
figs. 42-45
1952. — Sakesaria tereta n.sp., , p. 199-200, Pl. XII, figs. 6-9.
Sakesaria tereta was derived from Lockhartia retiata through elongation of the axis of enrollment together with a reduction in ornamentation. Numerous intermediaries link the two genera. The species is recognizable at once because of the obtuse rounded apex, never acute as is common in other species of this genus, by the relatively low spire and the thick, heavy bourrelet that maintains a uniform width throughout its course on the spiral suture. The height/diameter relationship measured on forty specimens ranged from 1.0/0.9 to 1.8/1.6, average 1.42/1.33 or 1.06. As these figure show, the shape is squat and broad as the spire reaches nearly its maximum width in the third whorl. Most adults have only four whorls. The base is moderately convex and has an imperforate peripheral thickening; the pillars reach it without an appreciable decrease in diameter (average about 0.08 mm), with the proviso that the arrangement is not complicated by an increase in the length of some pillars that intertwine and become U-shaped as in certain Lockhartia.
The dorsal ornament is a heavy bourrelet, continuous from top to bottom, occasionally supplemented by elongate thickened granules or papillae arranged radially on the apex. Other less conspicuous motifs are of two types: grooves marking the position of the septa and a raised net around the pores. This secondary ornamentation is present only near the summit and rarely extends past the third whorl. There are 12 to 14 chambers in the third and fourth whorls of a large specimen; they are 0.5 mm high, 0.3 mm long and 0.3 mm wide. The septa are at an angle of 30 to 40 degrees to the vertical, inclined backward and slightly curved inward. The double wall of the septum separates where it joins the base of the preceding whorl.
The axial section shows that the test is coarsely perforate and that the dorsal wall is thick (up to 0.2 mm), thinner in the third and fourth whorls. The lower wall of the chambers is 20 to 30 degrees from horizontal and slightly thickened interiorly. A tangential section shows a 'double' proloculum of moderate size. The two chambers, both a flattened oval, are almost equal in size with the greater diameter about 0.1 mm.
Range: Lower third of the Umm er Radhuma Formation.
Sakesaria nodulifera n.sp.
Pl.
5 ,
figs. 46-50
1952. — Sakesaria nodulifera n.sp., , p. 201-202, Pl. XII, figs. 1-5.
As mentioned, Sakesaria nodulifera is undoubtedly related to Lockhartia retiata. Ornamentation is identical, but the spire is much higher. As in all species of Sakesaria dimensions vary considerably, but in general the spire is rather high, particularly in those specimens from near the upper limit of the range. The dimensional relationships of fifty specimens varies from 1.1/0.8 to 3.0/1.6 or an average h/d of 1.84/1.17 or 1.57. Calculated inversely as was the rule for Lockhartia the ratio is always less than one: in this species 0.64.
In most specimens the spire grows rapidly for three whorls, the rate of increase so great that in some individuals the spiral lamina appears step-like. In the fifth and six whorls growth almost ceases. On the periphery is a thick imperforate band up to 0.25 mm wide. The base may be strongly convex or almost flat, according to the stage of growth; in most adults bases are strongly convex. Pillars may or may not be arranged in a complex manner; in general they are more complex in individuals from the upper end of the range; their diameter averages 0.08 mm when they are not elongated.
Ornamentation is mainly heavy nodules placed at the junction of septa with the spiral suture. If the nodules are heavy enough they may join to make a scalloped ridge at the whorl junctions. In addition the septa may be marked by ridges that link with the bourrelet. However, ornamentation is usually simple and is never very complex. As the septa are inclined at about 25 degrees the ridges on them are at the same angle. There are 12 to 14 chambers in the fourth whorl of a large specimen. There are openings between the chambers: they are rather large half-circles (0.1 mm diameter) on the inner wall of the septa; the ventral walls of the chambers appear to be in contact with the central mass of pillars; only a thin wall separates pillars from the inner side of the chambers, so that the openings in the septa form a continuous spiral tunnel around the pillars. This thin wall has an opening into each chamber as in L. retiata. The septa are double-walled and the walls separate dorsally in a regular curve to join the preceding whorl.
The axial section shows a small round proloculum (about 0.09 mm in diameter) with thin lateral walls (0.05 mm thick); in contrast, the ventral walls of the chambers are very thick (0.25 mm) and on their distal ends are the projections giving the impression of very rapid growth in each successive whorl. In fact each whorl is set slightly outside the preceding one. The thick ventral walls of the chambers seem to be imperforate. The central mass is well defined in the figure, with some fairly long pillars in the lower part.
Range: Lower third of the Umm er Radhuma Formation.
Sakesaria trichilata n.sp.
Pl.
5 ,
figs. 38-41
1952. — Sakesaria trichilata n.sp., , p. 203-204, Pl. XII, figs. 17-20.
As mentioned, Sakesaria trichilata too may have been derived from Lockhartia retiata. Some specimens of L. retiata have ornamentation similar to that of this new species, named for its curious pattern which resembles a trellis. As in other species of Sakesaria the proportions of the test are extremely variable, but in general the spire is comparatively low. Measurements of twenty specimens range from 1.0/0.8 to 3.6/2.0, with the average h/d of 1.76/1.25 or 1.43. The first four whorls of the spire grow rapidly, but it is constant thereafter or decreases slightly in the final whorl of very large specimens. The form varies from that of a rather pointed cone (diameter/height ratio 1.87) to that of a low, squat cone (diameter/height ratio 1.11). The ornamentation is equally variable, but in the pattern considered typical thick vertical ribs cover the entire test, except in the last whorl where nodules extend laterally from the spiral lamina. These ribs are linked by less prominent horizontal or diagonal ribs. In another pattern two sets of bars are equally developed: one common variant shows a quadrilateral or pentagonal on which the vertical component is thickened over the septa in the early whorls. In others the diagonal components of the trellis are the most prominent. There are many other variations, but the robust test and the generally reticular aspect of the ornamentation form an easily recognized whole. In this species of Sakesaria the ventral pillars (like those of S. cotteri) are very elongated, interlaced, and at the ventral surface form convoluted shapes similar to those in Lockhartia. On the last whorl of some few individuals there are traces of thickenings resembling those of L. retiata. The axial section reveals that more than half of the diameter of the test is filled by a central confused mass of pillars. The dorsal wall is thick (0.12 to 0.17 mm) the thickness decreasing slightly in the lower whorls. The chambers are much higher than long, the approximate proportion in the second whorl being two to one. The ventral wall of the chambers is thick, imperforate (0.10 mm), and short, inclined at a downward angle of about fifty degrees and enlarged toward the periphery. A thin wall apparently separates the inner edge of the chambers from the area and is joined to the dorsal wall of the chambers, but this connection is not well defined. The proloculum is large, "double". The two chambers are about equal in size, oval, with the greater diameter about 0.11 mm.
Range: Middle third of the Umm er Radhuma Formation.
Sakesaria abornata n.sp.
Pl.
5 , fig.
51
1952. — Sakesaria abornata n.sp., , p. 205, Pl. XII, fig. 16.
This species is rare so only two specimens are to hand. It differs from all others described here. Height and diameter are about the same, each slightly greater than 1.2 mm. The spire is cone-shaped, the spire growing rapidly throughout the four whorls; the base is almost flat, the proloculum slightly off center. Ornamentation is meager. However several unfigured specimens have low, poorly developed nodules at the junction of the spiral lamina and the sutures. There are 12 chambers in the last whorl of large specimens; each is about one-and a-half times as high as long; a slight depression marks the position of the sutures in the final whorl. The septa are inclined toward the origin of the spire at about thirty-five degrees to the vertical. In some specimens the conical shape is twisted in the last whorl.
The ventral surface is covered by many small pillars, not over 0.5 mm in diameter, but widely spaced and somewhat irregular in shape. A marginal region exists in which the base of the last chambers is visible. It varies in width, but the maximum is 0.2 mm. The lack of ornament and the small size of this species make it easy to recognize.
Range: Upper portion of the lower half of the Umm er Radhuma Formation.
Sakesaria cotteri , 1937
Pl.
4 ,
figs. 27-32
1937. — Sakesaria cotteri , & , p. 49-50, Pl. VII, figs. 18, 21-24, text-fig. 2.
1952. — Sakesaria trichilata , , p. 206-209, Pl. XII, figs. 10-15.
defined this extremely variable species from a small number of specimens, or at most from a selection of varieties that do not demonstrate the great diversity of the Arabian fauna. He hardly mentions the wide range of height/width dimensions or the enormous variety of ornamentation apparent at first glance in Arabian examples. As mentioned, a number of trends in variation may be traced, but all are linked by intermediate forms. Stratigraphic position can be estimated grosso modo on the relationships and relative importance of some varieties; but it is unlikely that this could be done in widely separated areas for it is doubtful that evolutionary processes would occur with the same rhythm and in precisely the same trends in discrete geographic provinces.
The first Sakesaria cotteri to appear have a low spire, the form of the test depending on the diameter of successive whorls and the degree of overlap between them. All are a relatively obtuse cone, which sometimes appears to be slightly truncated. The base varies in degree of curvature, but is commonly rounded, flatly obtuse. Ornamentation consists mainly of low, imperforate striae, and sutures more or less in relief, on occasion linked and continuous on the early whorls. Thirty-five specimens of this type with an origin near the first appearance of the species have a height/diameter relationship of 1.1/0.7 to 2.2/1.4, an average of 1.45/1.08. The maximum h/d ratio is 2.00, the minimum 1.08, the mean 1.34.
At a little higher stratigraphic level, ornamentation on the chambers is more pronounced on almost all specimens; it has a high relief and may join between whorls to make continuous striae over the length of the test. In others, beads and nodules occur on the early whorls and the relief of the striae lessens on the final whorl. The height/diameter relationship increases markedly and some specimens show an extreme elongation. In most of these elongated forms the relief is lower on the last two whorls. In addition to the general tendency to elongation of the axis of coiling, the diameter of succeeding whorls may decrease; in other words the summit angle becomes even more acute. The h/d dimensions of fifty specimens of this type range from 1.3/0.9 to 3.7/1.4, an average of 2.08/1.11. The height/diameter ratio is thus between 1.36 and 2.84 with the average 1.86.
In the upper part of its range ornamentation becomes discontinuous; it resembles that of the earliest variants, but with more prominent nodes and beads. Forms with striae over the length of the test are rare. In general, the summit is covered by raised papillae with a relief of 0.12 mm and a mean diameter of 0.08 mm. On the larger whorls short and thick striae of variable relief occur grading in one direction toward thick nodules and in another to low thickenings on the sutures. In addition the two with tendencies toward discrete forms of ornament also vary in dimensions, the one toward a low spire, the other to a high spire, but intermediates to both tendencies exist.
Fifty specimens of the low-spired variant have an h/d relationship between 1.1/0.9 and 2.3/1.4, the average 1.47/1.15 or 1.28, a figure almost the same as that obtained from the oldest specimens. The highest figure for this variant was 1.82, the lowest, 0.84.
Measurement of thirty individuals with high spires gave a height/diameter ratio of 2.0/1.0 to 4.2/1.6 with the average 3.60/1.36 or a ratio of 2.65. For this variant the highest ratio was 2.70, the lowest 1.83.
If the population is taken as a whole with no interior divisions the two groups together have an average h/d of 2.07/1.23 or 1.69.
All of these figures serve to show the range of the dimensions of Sakesaria cotteri. Of the 165 individuals measured the actual size varied from 1.1/0.7 to 4.2/1.6. And the height/diameter ratio is 0.84 to 2.70.
The mean height/diameter ratio for the whole population is 1.68. These figures should express with fair precision those of the species as it is represented in coastal Saudi Arabia.
In summary, the variations in ornamentation are as follows:
There are several other tendencies in ornamentation; individuals possessing them are fewer in number and certain of the tendencies seem related to unusual dimensional characteristics. In one of these the shape is heavy and thick (h/d : 3.7/1.5) with an obtuse angle at the summit and a rapid increase in the diameter of the whorls. These characters appear to be linked to a reduction in ornamentation. All that remain are more or less well-developed traces of low striae on the sutures, along with longitudinal striae also of low relief; this variant has a punctate aspect owing to the development of low pentagonal or quadrilateral ridges around the pores. A host of combinations of form and ornamentation occur, but some are more common. As the number of variations and combinations is so great it was not possible to describe every variant, but only the tendencies of groups of variants.
Range: Upper third of the Umm er Radhuma Formation.
L.R. (1936).- Fossil Mollusca from southern Persia and Bahrein.- Memoirs of the Geological Survey of India: Palaeontologia Indica, Calcutta, (N.S.), vol. XXII, n° 2, 69 p. (VIII pls.)
J. (1930).- Révision du Nummulitique égyptien.- Thèses présentées à la Faculté des Sciences de Paris, Cairo, E. & R. Schindler.- Mémoires présentés à l'Institut d'Égypte, Le Caire, vol. XVI, 371 p. (25 pls.)
L.M. (1861).- XXVII.- On the structure of the larger Foraminifera.- Annals and Magazine of Natural History, London, (serie III), vol. VIII, n° 45, p. 246-251.
L.M. (1924).- Notes on the geology of Kohat, with reference to the homotaxial position of the Salt Marl at Bahadhur Khel.- Journal of the Royal Asiatic Society of Bengal, Calcutta, (N.S.), vol. XX, p. 207-224. (pls. 8-9)
L.M. (1926).- Remarks on 's genus Conulites - Dictyoconoides with descriptions of some new species from the Eocene of North-West India.- Records of the Geological Survey of India, Calcutta, vol. LIX, pt. 2, p. 237-253. (5 pls.).
L.M. (1927).- The Ranikot beds at Thal (North-West Frontier Provinces of India).- Quarterly Journal of the Geological Society, London, vol. 83, p. 260-290.
L.M. (1930).- The fossil fauna of the Samana Range and some neighbouring area. Part VI: The Paleocene Foraminifera.- Memoirs of the Geological Survey of India: Palaeontologia Indica, Calcutta, (N.S.), vol. XV, p. 67-79. (pl. X)
L.M. (1932).- The genera Dictyoconoides , Lockhartia nov., and Rotalia . Their type species, generic differences, and fundamental distinction from the Dictyoconus group of forms.- Transactions of the Royal Society of Edinburgh, vol. LVII, pt. II, n° 13, p. 397-428. (IV pls.)
L.M. & E.S. (1937).- The Eocene Beds of the Punjab Salt Range.- Memoirs of the Geological Survey of India: Palaeontologia Indica, Calcutta, (N.S.), vol. XXIV, n° 1, 79 p. (VII pls.)
W.L.F. (1925a).- The Stratigraphy of the Laki Series (Lower Eocene) of parts of Sind and Baluchistan (India) with a description of the larger Foraminifera contained in these beds.- Quarterly Journal of the Geological Society of London, vol. 81, pt. 3, p. 417-453. (pls. XXIII-XXVII)
W.L.F. (1925b).- XLVIII.- Two species of Eocene Foraminifera from India. Alveolina elliptica and Dictyoconoides cooki.- Annals and Magazine of Natural History, London, (serie IX), vol. XVI, n° 94, p. 378-388.
W.L.F. (1926a).- The larger Foraminifera of the Upper Ranikot Series (Lower Eocene) of Sind, India.- Geological Magazine, Cambridge, vol. LXIII, n° 3, p. 112-121. (pls. X-XI)
W.L.F. (1926b).- The zonal distribution and description of the larger Foraminifera of the middle and lower Kirthar zones (Middle Eocene) of western India.- Geological Magazine, Cambridge, vol. LXIII, n° 11, p. 495-504.
W.L.F. (1931).- The stratigraphy of the Upper Ranikot Series (Lower Eocene) of Sind, India.- Records of the Geological Survey of India, Calcutta, vol. LXV, p. 306-313.
W.L.F. & A.G. (1931).- Larger Foraminifera from the Tertiary of Somaliland.- Geological Magazine, Cambridge, vol. LXVIII, n° 2, p. 49-65. (IV pls.)
C. van (1930).- IX.- Some remarks on Dictyoconoides, (=Conulites, =Rotalia, ).- Annals and Magazine of Natural History, London, (serie X), vol. V, n° 25, p. 116-135.
N.J. (1952).- 1ère thèse : La stratigraphie de l'Éocène le long du rivage occidental du Golfe Persique ; 2ème thèse : Essai biométrique sur les variations des genres Lokhartia et Sakesaria.- Thèses, Université de Paris, La Sorbonne (5 mars 1952), 256 p. (27 pls.)
N.J. (1962).- Aperçu paléontologique et stratigraphique du Paléogène en Arabie Séoudite orientale.- Revue de Micropaléontologie, Paris, vol. 5, n° 9, p. 3-40.
F.C., P.E. & A.N. (1951).- Stratigraphical exploration surveys in southwest Persia.- Proceedings of the Third World Petroleum Congress, The Hague, section 1, p. 141-157.
A.H. (1954).- Lower Tertiary Foraminifera of the Qatar Peninsula.- British Museum of Natural History, Monograph, London, 96 p. (I-XV pls.)
M., R.A. & N.J. (1958).- Stratigraphic relations of Arabian Jurassic oil. In: L.G. (ed.), Habitat of oil: A symposium.- The American Association of Petroleum Geologists, Tulsa, p. 1294-1329.

![]()
Click on thumbnail to enlarge the image.
1-3. Nummulites cuvillieri n.sp., Form B. (All specimens figured are from Oqair N° 1, 85-86 meters).
1. Surface of the cotype illustrating variant with few granules (Pl. V, fig. 1 in , 1952).
2. Tangential section showing the relationship between septal filaments and granules (Pl. V, fig. 2 in , 1952).
3. Equatorial section (Pl. V, fig. 3 in , 1952).
4-8. Nummulites cuvillieri n.sp., Form A.
4-6. Exterior (Pl. V, figs. 4-6 in , 1952).
7-8. Equatorial sections (Pl. V, figs. 7-8 in , 1952).
9. Nummulites cuvillieri n.sp., Form B. Exterior of cotype (Pl. V, fig. 9 in , 1952).
10. Nummulites cuvilllieri n.sp., Form A. Axial section (Pl. V, fig. 10 in , 1952).
11-13. Nummulites cuvillieri n.sp., Form B.
11. Axial section (Pl. V, fig. 11 in , 1952).
12. Peripheral view (Pl. V, fig. 12 in , 1952).
13. Equatorial section (Pl. V, fig. 13 in , 1952).
14-17. Nummulites cizancourti n.sp., Form A. (All specimens figured from well S-50, 164-166 meters).
14. Exterior (Pl. III, fig. 16 in , 1952).
15. Axial section (Pl. III, fig. 17 in , 1952).
16-17. Equatorial sections (Pl. III, figs. 18-19 in , 1952).

![]()
Click on thumbnail to enlarge the image.
Plate 2: Miscellanoides bramkampi n.gen., n.sp.
1. Genotype. Form B. Convex face (Pl. VIII, fig. 1 in , 1952). Dammam N° 6, 156-161 m.
2. Genotype. Form B. Concave face (Pl. VIII, fig. 7 in , 1952).
3. Form A. Convex face (Pl. VIII, fig. 11 in , 1952). Abqaiq N° 6, 390-402 m.
4. Form A. Concave face of fig. 3 (Pl. VIII, fig. 15 in , 1952).
5. Form B. Equatorial section (Pl. VIII, fig. 18 in , 1952). Abqaiq N° 1, 387-389 m.
6. Form B. Axial section (Pl. VIII, fig. 2 in , 1952). Abqaiq N° 1, 387-389 m.
7. Form B. Axial section (Pl. VIII, fig. 5 in , 1952). Abqaiq N° 1, 387-389 m.
8. Form A. Concave face (Pl. VIII, fig. 8 in , 1952). Abqaiq N° 6, 399-402 m.
9. Form A. Convex face of fig. 8 (Pl. VIII, fig. 12 in , 1952). X 21.
10. Form A. Concave face (Pl. VIII, fig. 16 in , 1952). Abqaiq N° 6, 399-402 m.
11. Form A. Axial section (Pl. VIII, fig. 3 in , 1952). Abqaiq N° 1, 387-389 m.
12-13. Form A. Equatorial sections (Pl. VIII, figs. 10 & 14 in , 1952). Abqaiq N° 6, 441-445 m.
14. Form B. Axial section (Pl. VIII, fig. 6 in , 1952). Abqaiq N° 1, 387-389 m.
15. Form A. Axial section (Pl. VIII, fig. 4 in , 1952). Abqaiq N° 1, 387-389 m.
16. Form B. Axial section (Pl. VIII, fig. 9 in , 1952). Abqaiq N° 1, 387-389 m.
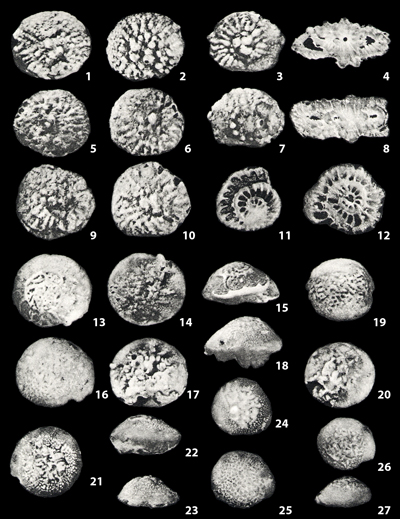
![]()
Click on thumbnail to enlarge the image.
1-12. Miscellanoides pruvosti n.sp., Form A.
1-3. Cotypes. Convex face (Pl. IX, figs. 1-3 in , 1952). Abqaiq N° 1, 344-346 m.
4 and 8. Axial sections (Pl. IX, figs. 4 & 8 in , 1952). Abqaiq N° 1, 344-346 m.
5-7. Cotypes. Concave face (Pl. IX, figs. 5-7 in , 1952).
9-10. Cotype. Concave and convex faces (Pl. IX, figs. 9-10 in , 1952). Abqaiq N° 1, 344-346 m.
11-12. Equatorial sections (Pl. IX, figs. 11-12 in , 1952). Abqaiq N° 1, 344-346 m.
13-20. Rotalia jacobi n.sp.
13-15. Dorsal, ventral and lateral views of cotype (Pl. IX, figs. 13-15 in , 1952). Al Alat N° 1, 582-585 m.
16-18. Dorsal, ventral and lateral views of cotype (Pl. IX, figs. 17-19 in , 1952). Al Alat N° 1, 582-585 m.
19-20. Dorsal and ventral views of cotype (Pl. IX, figs. 21-22 in , 1952). Al Alat N° 1, 582-585 m.
21-27. Lockhartia lobulata n.sp.
21-22. Dorsal and lateral views of cotype (Pl. IX, figs. 23 & 27 in , 1952). Al Alat N° 1, 423-424 m.
23-25. Lateral, dorsal and ventral views of cotype (Pl. IX, figs. 25 & 23-24 in , 1952). Al Alat N° 1, 423-424 m.
26-27. Dorsal and lateral views of cotype (Pl. IX, figs. 28-29 in , 1952). Al Alat N° 1, 423-424 m.
![]()
Click on thumbnail to enlarge the image.
1-15. Lockhartia retiata n.sp. (Pl. X, figs. 1-15 in , 1952). Abqaiq N° 1, 454-457 m.
1-3. Dorsal, lateral and ventral views of a specimen.
4-5 Dorsal and lateral views of another specimen.
6-8. Dorsal, lateral and ventral views of a specimen.
9-10. Dorsal and lateral views of another specimen.
11-13. Dorsal, lateral and ventral views of a specimen.
14-15. Dorsal and lateral views of a specimen.
16-20. Lockhartia retiata n.sp. (Pl. X, figs. 16-20 in , 1952). Dammam N° 3, 214-216 m. Lateral views showing the changes in the height of the spire and in ornamentation in the direction of the genus Sakesaria.
21. Lockhartia retiata n.sp. (Pl. X, fig. 21 in , 1952). Abqaiq N° 1, 454-457 m. Axial section showing the proloculum.
22-24. Dictyoconoides kohaticus (). Form A. nov. (Pl. XII, figs. 35 & 32-33 in , 1952). Al Alat N° 1, 116-121 m.
22. Lateral view.
23. Axial section.
24. Dorsal view.
25-26. Lockhartia tipperi (). Form B (Pl. XI, figs. 53-52 in , 1952). Abqaiq N° 6, 234-236 m. Dorsal and lateral views of a specimen.
27-32. Sakesaria cotteri (). Abqaiq N° 2, 234-236 m.
27. Specimen with conical spire (Pl. XII, fig. 15 in , 1952).
28. Specimen with thin spire (Pl. XII, fig. 14 in , 1952).
29 and 31. Ornamentation reduced to striae (Pl. XII, figs. 11-10 in , 1952).
30 and 32. Ornamentation of gross papillae (Pl. XII, figs. 13-12 in , 1952).
33-36. Lockhartia tipperi (). Form A. nov. (Pl. XI, figs. 47-48 & 50-51 in , 1952).
33. Dorsal view.
34. Ventral view.
35. Lateral view.
36. Ventral view.
37-39. Lockhartia conditi () (Pl. XI, figs. 39- & 41-40 in , 1952). Dorsal, lateral and ventral views.
![]()
Click on thumbnail to enlarge the image.
1-9. Lockhartia haimei () (Pl. XI, figs. 1-9 in , 1952). Abqaiq N° 1 451-454 m.
1-5. Dorsal views.
6. Ventral view.
7-9. Lateral views.
10-15. Lockhartia haimei var. vermiculata n.var. (Pl. XI, figs. 10-15 in , 1952). Abqaiq N° 1, 478-479 m. Dorsal and lateral views of three specimens.
16-21. Lockhartia haimei var. nudimarginata n.var. (Pl. XI, figs. 16-21 in , 1952). Abqaiq N° 1, 472-475 m. Dorsal and lateral views of three specimens.
22-24. Lockhartia haimei var. suturadicata n.var.; (Pl. XI, figs. 22-24 in , 1952). Abqaiq N° 1, 472-475 m. Dorsal views of two specimens and a lateral view of one of them.
25-30. Lockhartia haimei var. spirachordata n.var. (Pl. XI, figs. 26-31 in , 1952). Abqaiq N° 1, 457-460 m. Dorsal and lateral views of three specimens.
31-32. Lockhartia haimei. Unnamed variation. (Pl. XI, figs. 25 & 32 in , 1952). Abqaiq N° 1, 454-457 m. Dorsal and lateral views of a specimen ornamented only with papillae.
33. Lockhartia haimei. Unnamed variation. (Pl. XI, fig. 34 in , 1952). Abqaiq N° 1, 454-457 m. Dorsal view of specimen with ornament leading to L. haimei var. spirachordata.
34. Lockhartia haimei. Unnamed variation. (Pl. XI, fig. 35 in , 1952). Abqaiq N° 1, 472-475 m. Dorsal view showing localized ornament leading to L. haimei var. nudimarginata.
35. Lockhartia haimei var. nudimarginata n.var. (Pl. XI, fig. 36 in , 1952). Abqaiq N° 6, 472-475 m. Tangential section near summit.
36. Lockhartia haimei var. nudimarginata n.var. (Pl. XI, fig. 37 in , 1952). Dammam N° 2, 227-228 m. Axial section showing the proloculum.
37. Lockhartia haimei var. suturadicata n.var. (Pl. XI, fig. 38 in , 1952). Abqaiq N° 1, 457-458 m. Axial section.
38-41. Sakesaria trichilata n.sp.
38. Axial section (Pl. XII, fig. 17 in , 1952).
39-41. Lateral views (Pl. XII, figs. 20 & 18-19 in , 1952).
42-45. Sakesaria tereta n.sp. (Pl. XII, figs. 6-7 & 9-8 in , 1952). Lateral views.
46-50. Sakesaria nodulifera n.sp.
46-47. Lateral views (Pl. XII, figs. 2-3 in , 1952). Dammam N° 2, 213-217 m.
48. Axial section (Pl. XII, fig. 1 in , 1952). Dammam N° 2, 213-217 m.
49-50. Lateral views (Pl. XII, figs. 5-4 in , 1952). Abqaiq N° 6, 472-475.
51. Sakesaria abornata n.sp. (Pl. XII, fig. 16 in , 1952) Al Alat N° 1, 445-446 m. Lateral view of holotype.