![]()
Contents
[1. Introduction] [2. Material, Methods and Geologic setting]
[3. Kaugatuma cliff] [4. Results]
[5. Discussion] [Bibliographic references]
Department of Geology, University of Tartu, Ravila 14A, EE-50411 Tartu (Estonia)
Manuscript online since March 12, 2012
[Editor: Christian ]
![]()
Shell repair is common in the late Silurian (Pridoli) encrusting tentaculitoid tubeworm Anticalyptraea calyptrata from Saaremaa, Estonia (Baltica), and is interpreted here as a result of failed predation. A. calyptrata has a shell repair frequency of 29 % (individuals with scars) with 17 specimens. There is probably an antipredatory adaptation, i.e. extremely thick vesicular walls, in the morphology of Silurian Anticalyptraea. The morphological and ecological evolution of Anticalyptraea could thus have been partially driven by predation.
Predation; shell repair; Tentaculita; Anticalyptraea; Pridoli; Silurian; Estonia.
O. (2012).- Shell repair in Anticalyptraea (Tentaculita) in the late Silurian (Pridoli) of Baltica.- Carnets de Géologie [Notebooks on Geology], Brest, Letter 2012/01 (CG2012_L01), p. 31-37.
Réparation de la coquille chez Anticalyptraea (Tentaculita) dans le Silurien supérieur (Pridoli) du bouclier balte (Baltica).- La réparation de coquilles est habituelle chez Anticalyptraea calyptrata, un vers tentaculitoïde encroûtant du Silurien supérieur (Pridoli) à Saaremaa, Estonie (bouclier balte), et est interprétée ici comme la conséquence d'une prédation qui aurait échoué. A. calyptrata a une fréquence de réparation de la coquille de 29% (individus présentant des cicatrices) pour 17 spécimens. Ceci est probablement une adaptation contre la prédation, c'est-à-dire la présence de parois vésiculaires et très épaisses, présentes dans la morphologie de l'Anticalyptraea du Silurien. L'évolution morphologique et écologique d'Anticalyptraea pourrait donc pour partie avoir été provoquée par la prédation.
Prédation ; réparation de la coquille ; Tentaculites ; Anticalyptraea; Pridoli ; Silurien ; Estonie.
There is a single previous record of possible predation on Anticalyptraea from the Pridoli of Saaremaa (Estonia), but the predation frequency or details of the shell repair was unknown (,
2007):
Fig. 1 ![]() . The material for this study was collected from two outcrops
- Kaugatuma cliff and Ohesaare cliff - (Fig. 1
. The material for this study was collected from two outcrops
- Kaugatuma cliff and Ohesaare cliff - (Fig. 1 ![]() ) during five field expeditions in 2006-2010. In addition collections of the Natural History Museum, University of Tartu, were studied. Anticalyptraea is a rare fossil in the Pridoli of Saaremaa. It belongs to encrusting tentaculitoid tubeworms and is closely related to cornulitids, microconchids and tentaculitids ( & ,
2007; , 2010). Anticalyptraea occurs as an encruster on various shelly fossils, such as brachiopods, stromatoporids, corals, but also on hardgrounds ( & ,
2010b).
) during five field expeditions in 2006-2010. In addition collections of the Natural History Museum, University of Tartu, were studied. Anticalyptraea is a rare fossil in the Pridoli of Saaremaa. It belongs to encrusting tentaculitoid tubeworms and is closely related to cornulitids, microconchids and tentaculitids ( & ,
2007; , 2010). Anticalyptraea occurs as an encruster on various shelly fossils, such as brachiopods, stromatoporids, corals, but also on hardgrounds ( & ,
2010b).
Click on thumbnail to enlarge the image.
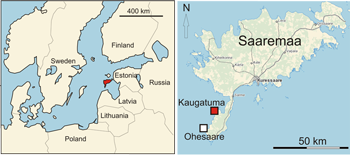
Figure 1: Location of Kaugatuma (lower Pridoli) and Ohesaare (upper Pridoli) cliffs. Locality of Anticalyptraea calyptrata with shell repair marked with red.
The aims of this paper are: 1) to describe in detail the shell repairs in the encrusting tentaculitoid tubeworm Anticalyptraea for the first time; 2) to assess how often Anticalyptraea was attacked by predators in the late Silurian; 3) to describe shell repair mechanisms in Anticalyptraea; 4) to find correlations between shell repair rate and size of Anticalyptraea specimens; and 5) to discuss possible antipredatory adaptations in Anticalyptraea morphology and behavior.
17 specimens from Kaugatuma cliff and five specimens from Ohesaare cliff were examined for shell repair and predatory drillings under an Olympus B061 binocular light microscope. The photography was done with a digital camera mounted on the binocular microscope. In addition, shell repair was studied with a Hitachi S-4300 FE-SEM at the Swedish Museum of Natural History. The diameters of Anticalyptraea shells at the lowest edge of each shell repair was measured and catalogued to record the location of the shell repairs.
There are two common methods of determining shell repair frequencies: 1) the "individuals with scars" method which involves dividing the number of individuals with at least one scar by the total number of individuals in the sample (,
2002; ,
2003: p. 163, and references therein); and 2) the "scar per shell" method which involves dividing the total number of repairs by the total number of individuals in the sample (,
2003: p. 163, and references therein).
(2002: p. 17) and
(2003: p.
163) have advocated standardization of data tables so that data from both methods are available, which is followed in this paper
(Table 1, Fig. 4 ![]() ). All figured specimens are deposited in the Museum of Natural History, University of Tartu (TUG), Estonia.
). All figured specimens are deposited in the Museum of Natural History, University of Tartu (TUG), Estonia.
The Pridoli (uppermost Silurian) rocks (Fig. 2 ![]() ) are distributed in the southwestern part of Saaremaa island (Fig. 1
) are distributed in the southwestern part of Saaremaa island (Fig. 1 ![]() ). The major outcrops are coastal cliffs at the western coast of Sõrve peninsula.
). The major outcrops are coastal cliffs at the western coast of Sõrve peninsula.
| Specimen n° | Shell max. width (mm) | Number of repairs | Shell width at the repair (mm) |
| TUG 1607-1 | 7.0 | 3 | 6.8 - 6.7 - 3.7 |
| TUG 1607-2 | 5.6 | 1 | 5.0 |
| TUG 1607-3 | 5.5 | 1 | 3.5 |
| TUG 1607-4 | 5.1 | 1 | 4.8 |
| TUG 1116-1 | 4.8 | 1 | 4.2 |
| Method | Number |
| individuals with scars | 5 of 17 (29%) |
| Scars per shell | 0.41 |
Table 1: Shell repairs in Anticalyptraea calyptrata , 1860, from the Kaugatuma cliff, lower Pridoli (upper Silurian) of Saaremaa, Estonia.
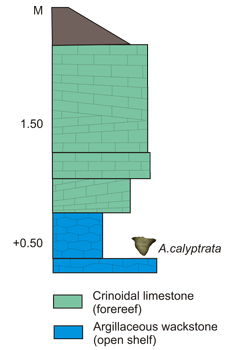
The Kaugatuma cliff (2.5 m high) is situated on the western coast of the Sõrve Peninsula, some kilometers south from its neck and about 100 meters from the sea (58°7'22"N, 22°11'36"E):
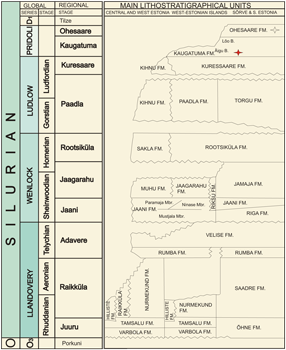
Fig. 3 ![]() . The rocks belong to the middle part of the Äigu Beds of the Kaugatuma Stage (Fig. 2
. The rocks belong to the middle part of the Äigu Beds of the Kaugatuma Stage (Fig. 2 ![]() ):
):
Click on thumbnail to enlarge the image.
Figure 2: Stratigraphy of the Silurian of Estonia (after et al., 2008). Anticalyptraea with shell repair marked with red star.
Click on thumbnail to enlarge the image.
Figure 3: Kaugatuma cliff after (1990). Anticalyptraea calyptrata with shell repair occurs in the lower part of section.
A great number of fossils have been recorded from the Kaugatuma locality, including large crinoid holdfasts and columnals, trilobites, tabulate corals and stromatoporoids. Major predators include acanthodian fish and cephalopods. The following vertebrate remains have been found from the lower beds of the cliff: Nostolepis striata, Gomphonchus sandelensis, Poracanthodes porosus ( et al., 2008).
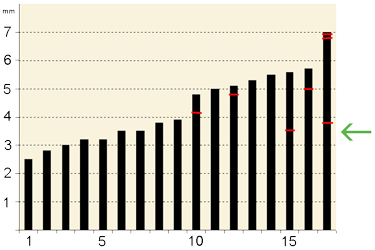
Shell repairs were found in five of 17 specimens from the Kaugatuma cliff
(Table 1, Fig. 4 ![]() ). Most of the shell repairs are very large as compared to the size of Anticalyptraea calyptrata specimens, extending up to 50% of the shell diameter (from 1.5 to 3.5 mm wide):
Figs. 5
). Most of the shell repairs are very large as compared to the size of Anticalyptraea calyptrata specimens, extending up to 50% of the shell diameter (from 1.5 to 3.5 mm wide):
Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() . Studied shell repairs of A. calyptrata display a meandering or zigzag fracture which outlines a jagged piece of the shell surface that was removed and resecreted (Figs. 5
. Studied shell repairs of A. calyptrata display a meandering or zigzag fracture which outlines a jagged piece of the shell surface that was removed and resecreted (Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() ). The regenerated growth lines are oblique to the antecedent growth lines lateral to the fracture in the studied shell repairs. Five specimens (29 %) of A. calyptrata (n=17) from the Kaugatuma cliff (lower Pridoli) of Saaremaa have scars (n=7) of shell repair
(Table 1, Fig. 4
). The regenerated growth lines are oblique to the antecedent growth lines lateral to the fracture in the studied shell repairs. Five specimens (29 %) of A. calyptrata (n=17) from the Kaugatuma cliff (lower Pridoli) of Saaremaa have scars (n=7) of shell repair
(Table 1, Fig. 4 ![]() ). One of the five specimens of A. calyptrata has a multiple shell repairs (three repair marks). No shells smaller than 3.5 mm in diameter have repair marks. There are 0.41 scars per shell
(Table 1).
). One of the five specimens of A. calyptrata has a multiple shell repairs (three repair marks). No shells smaller than 3.5 mm in diameter have repair marks. There are 0.41 scars per shell
(Table 1).
Click on thumbnail to enlarge the image.
Figure 4: Graphic showing size distribution (max. diameter) of studied Anticalyptraea calyptrata specimens (n=17) from Kaugatuma cliff (lower Pridoli) of Saaremaa and location of the shell repairs (red lines). Green arrow points to the minimum diameter of the shells with repair marks.
All shell repairs in A. calyptrata are carried out by resumption of shell growth from underneath the broken edge so that the newly secreted repaired shell lies mostly below the surface of the damaged shell. In one specimen the newly secreted shell is flush with the surface of the damaged shell.
No shell repair was found in five specimens from Ohesaare cliff (upper Pridoli). No predatory drillings were discovered in any Anticalyptraea specimens studied.
Click on thumbnail to enlarge the image.
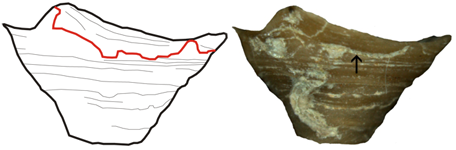
Figure 5: A. calyptrata , 1860, from Kaugatuma cliff (lower Pridoli), lateral view. Arrow points to shell repair. Max. diameter of the shell 7.0 mm.

Click on thumbnail to enlarge the image.
Figure 6: A. calyptrata ,
1860, from Kaugatuma cliff (lower Pridoli), apical view. Arrow points to shell repair. Same as
Figs. 5 ![]() . Max. diameter of the shell 7.0 mm.
. Max. diameter of the shell 7.0 mm.
Click on thumbnail to enlarge the image.
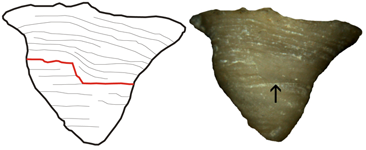
Figure 7: A. calyptrata , 1860, from Kaugatuma cliff (lower Pridoli), lateral view. Arrow points to shell repair. Max. diameter of the shell 5.5 mm.
Click on thumbnail to enlarge the image.
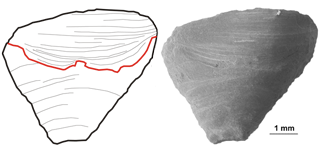
Figure 8: A. calyptrata , 1860, from Kaugatuma cliff (lower Pridoli), lateral view. Arrow points to shell repair.
The morphologies of shell repair in A. calyptrata and sizes of the scars resemble those known in Ordovician cornulitids (, 2009: p. 88, fig. 1A), and Silurian to Devonian tentaculitids (, 1979; et al., 2007). The minute size of A. calyptrata shells makes direct comparison difficult with the larger molluscs and brachiopods. However, even smaller sized shell repairs are known in recent tiny pelagic gastropods ( et al., 2007: p. 411, fig. 4). The shell repair in various organisms has often been interpreted as results of failed predatory attacks (e.g., , 1986; , 1997; , 2003; , 2006). The proposed predatory cause of shell repair in A. calyptrata herefollows from morphological similarities to the predator-caused shell repair in the Paleozoic mollusks and brachiopods, but especially in similar size shells of tentaculitids ( et al., 2007: p. 409, fig. 2, and p. 410, fig. 3) and cornulitids (, 2009: p. 88, fig. 1A). Alternatively, shell repair could have resulted from the natural breakage of shell in intense water movement during the storms. This kind of damage and repair has also been observed in molluscs (, 1968).
The shell repair carried out by resumption of shell growth from underneath the broken edge in Anticalyptraea (Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() ) is characteristic of the encrusting tentaculitoid tubeworms (,
2009) and their probable close relatives the hederelloids (,
2006). A similar method of shell repair has also been reported from free-living tentaculitids (,
1979; et al.,
2007).
) is characteristic of the encrusting tentaculitoid tubeworms (,
2009) and their probable close relatives the hederelloids (,
2006). A similar method of shell repair has also been reported from free-living tentaculitids (,
1979; et al.,
2007).
Recent large gastropod shells usually show higher proportions of shell repair because they have been exposed for longer times to predators as compared to their smaller relatives ( et al.,
1980, 1982). Similar patterns would also be expected for A. calyptrata because their shell growth was similar to that of the molluscs. All shell repairs in A. calyptrata occur only in relatively large specimens (≥3.5 mm in
diameter): Table 1, Fig. 4 ![]() . This could indicate that after a certain size is reached, Anticalyptraea became less vulnerable to lethal attacks and shell repairs accumulated as a result of predation being more likely to fail (,
2002; ,
2003, and references therein). Alternatively, the predators may have been size selective, and only large adult Anticalyptraea specimens contained enough food to attract an attack.
. This could indicate that after a certain size is reached, Anticalyptraea became less vulnerable to lethal attacks and shell repairs accumulated as a result of predation being more likely to fail (,
2002; ,
2003, and references therein). Alternatively, the predators may have been size selective, and only large adult Anticalyptraea specimens contained enough food to attract an attack.
The shell repair frequency of 29% (individuals with scars) in A. calyptrata is comparable to the shell repair frequencies in
Late Ordovician and Silurian gastropods (ranging from 2.4 to 35.7%; ,
1997, 2005; ,
1997; ,
2008):
Table 1, Fig. 4 ![]() . Molluscs were likely among the primary targets of Paleozoic predators. Brachiopods were probably secondary casualties of mistaken or opportunistic attacks ( et al.,
2005). However, the highest recorded shell repair frequencies (26%, individuals with scars) in
Late Ordovician brachiopods (, 1986) are slightly less but still comparable to that observed in A. calyptrata. The relatively high shell repair frequency in A. calyptrata alone does not prove whether the Anticalyptraea was among primary prey for Early Paleozoic predators, but it indicates their interest. The shell repair frequency is an ambiguous indicator that may either indicate a high rate of attacks or a high rate of failed attacks or both.
. Molluscs were likely among the primary targets of Paleozoic predators. Brachiopods were probably secondary casualties of mistaken or opportunistic attacks ( et al.,
2005). However, the highest recorded shell repair frequencies (26%, individuals with scars) in
Late Ordovician brachiopods (, 1986) are slightly less but still comparable to that observed in A. calyptrata. The relatively high shell repair frequency in A. calyptrata alone does not prove whether the Anticalyptraea was among primary prey for Early Paleozoic predators, but it indicates their interest. The shell repair frequency is an ambiguous indicator that may either indicate a high rate of attacks or a high rate of failed attacks or both.
Predation presumably has influenced evolution of cornulitids (, 2009; , 2009) and encrusting tentaculitoid tubeworms in general (, 2010). In Anticalyptraea, the external wall that were in contact with seawater are thick and vesicular, while internal walls are thinner and do not contain vesicles. In cornulitids, the vesicular wall structure may have evolved as an antipredatory adaptation (, 2009, 2010). Thus, the vesicular shell structure of Anticalyptraea could be a protective morphology. In addition to thick vesicular walls, Anticalyptraea has reported to have some preference for the cryptic surfaces when associated with the hardgrounds in Pridoli of Saaremaa that can indicate predation pressure (, 2010b). The latest known Anticalyptraea-like fossils belong to Streptindytes (Middle Devonian to Carboniferous), which lived embedded in the host coral skeleton. If the coral endobiont Streptindytes is a derived Anticalyptraea, it is possible that predation pressure may have led their evolution from encrusting life style to more protected endobiotic one. Similarly, in cornulitids predation pressure may have led to evolution of defensive morphologies and an endobiotic life style (, 2008; , 2009, 2010; , 2010a).
There are just few palaeoecological studies on the skeletal benthos from the Pridoli of Saaremaa. At the present there is no data on the shell repair in other groups of skeletal benthos. The future studies should demonstrate how common the shell repair in the Pridoli of Saaremaa was in general.
I am grateful to the Paleontological Society for a Grant, to Estonian Science Foundation for grant ETF9064 and to the Estonian Ministry of Education and Science for the target project SF0180051s08. I am grateful to Michał and anonymous reviewer for the constructive reviews. Ingrid is thanked for the help during the field work on Saaremaa.
R.R. (1986).- Resistance to and repair of shell breakage induced by durophages in Late Ordovician brachiopods.- Journal of Paleontology, Tulsa, vol. 60, p. 273-285.
R.R. & G.P. (2003).- The fossil record of shell-breaking predation on marine bivalves and gastropods. In: P.H., M. & T.A. (Eds.), Predator-prey interactions in the fossil record.- Kluwer Academic-Plenum Publishers, New York, p. 141-176.
S., J. & P. (2007).- Unsuccessful predation on Middle Paleozoic plankton: shell injury and anomalies in Devonian dacryoconarid tentaculites.- Acta Paleontologica Polonica, Warsaw, vol. 52, p. 407-412.
J.O.R. & J.S. (1997).- Attempted predation and shell repair in Middle and Upper Ordovician gastropods from Sweden.- Journal of Paleontology, Tusla, vol. 71, p. 1007-1019.
J.O.R. & C.A. (2008).- Failed predation in Late Ordovician gastropods (Mollusca) from Manitoulin Island, Ontario, Canada.- Canadian Journal of Earth Sciences, Ottawa, vol. 45, p. 231-241.
, E. (1860).- Lethaea Rossica ou Paléontologie de la Russie.- Schweizerbart, Stuttgart, 1654 p.
R. (1990).- Locality 7:3 Kaugatuma cliff. In: D. & H. (eds). Field meeting, Estonia 1990. An Excursion Guidebook. Institute of Geology, Estonian Academy of Sciences, Tallinn, p. 174-175.
O., L., P. & , T. (eds., 2008).- The Seventh Baltic Stratigraphical Conference.- Abstracts and Field Guide, Geological Society of Estonia, Tallinn, p. 46.
R. (1968).- Zur Morphologie rezenter Mollusken-Bruchschille.- Paläontologische Zeitschrift, Heidelberg, vol. 42, p. 217-235.
M. (2002).- The fossil record of predation: an overview of analytical methods. In: M. & P.H. (Eds.), The Fossil Record of Predation.- Paleontological Society Papers, Pittsburgh, vol. 8, p. 289-317.
M., A.P., T.K. & K. (2005).- Secondary evolutionary escalation between brachiopods and enemies of other prey.- Science, Washington, vol. 308, p. 1774-1777.
K. (1979).- Silurian tentaculitids from Gotland and Scania.- Fossil Strata, Oslo, vol. 11, 180 p.
L.R. (2002).- Inferring predation intensity in the marine fossil record.- Paleobiology, Lawrence, vol. 28, p. 328-342.
A. & J.S. (1997).- Failed predation and shell repair in the gastropod Poleumita from the Silurian of Gotland.- Vĕstník Českého Geologického Ústavu, Prague, vol. 72, p. 115-126.
A. & J.S. (2005).- Repaired injuries and shell form in some Paleozoic pleurotomarioid gastropods.- Acta Palaeontologica Polonica, Warsaw, vol. 50, p. 697-704.
G.J., E. & E.C. (1980).- Predation in time and space: peeling and drilling in terebrid gastropods.- Paleobiology, Lawrence, vol. 6, p. 352-364.
G.J., E. & E.C. (1982).- Breakage-induced shell repair in some gastropods from the Upper Triassic of Italy.- Journal of Paleontology, Tulsa, vol. 56, p. 233-235.
O. (2009).- Attempted predation on Early Paleozoic cornulitids.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 273, p. 87-91.
O. (2010).- Adaptive strategies in the evolution of encrusting tentaculitoid tubeworms.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 292, p. 211-221.
O. & M. (2007).- The tentaculitid affinities of Anticalyptraea from the Silurian of Baltoscandia.- Palaeontology, London, vol. 50, p. 1385-1390.
O. & M.-A. (2008).- The earliest endosymbiotic mineralized tubeworms from the Silurian of Podolia, Ukraine.- Journal of Paleontology, Tulsa, vol. 82, p. 409-414.
O. & H. (2009).- Calcareous tubeworms of the Phanerozoic.- Estonian Journal of Earth Sciences, Tallinn, vol. 58, p. 286-296.
O. & M.A. (2010a).- Endosymbiotic Cornulites in the Sheinwoodian (Early Silurian) stromatoporoids of Saaremaa, Estonia.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 257, p. 13-22.
O. & M.A. (2010b).- Microconchid-dominated hardground association from the late Pridoli (Silurian) of Saaremaa, Estonia.- Palaeontologia Electronica, Amherst, vol. 13, Issue 2, 9A (13.2.9A), 12p.
M.A. & P.D. (2006).- Predatory drillholes and partial mortality in Devonian colonial metazoans.- Geology, Boulder, vol. 34, p. 565-568.