◄ Carnets Geol. 17 (2) ►
![]()
Contents
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Results]
[5. Discussions and interpretation]
[6. Conclusions]
[Bibliographic references] and ... [Appendix]
Corresponding author
Babeș-Bolyai University, Faculty of Biology and Geology,
Department of Geology, Str. Kogălniceanu 1, 400084, Cluj Napoca (Romania)
Babeș-Bolyai University, Faculty of Biology and Geology,
Department of Geology, Str. Kogălniceanu 1, 400084, Cluj Napoca (Romania)
Babeș-Bolyai University, Faculty of Biology and Geology,
Department of Geology, Str. Kogălniceanu 1, 400084, Cluj Napoca (Romania)
Researcher
ID: J-6072-2014
OMV Petrom S.A. Coralilor 22, 013329, Bucharest 1 (Romania)
Published online in final form (pdf) on March 24,
2017
DOI 10.4267/2042/62041
[Editor: Bruno Granier; language editor:
Stephen Carey]
![]()
Planktonic and benthic foraminiferal assemblages were used for biostratigraphy and paleoenvironmental reconstruction of the marine lower Miocene Chechiș Formation from the Gălp�ia section (Sălaj county, Romania) in the northwestern Transylvanian Basin. Planktonic foraminifera suggest an Eggenburgian (Burdigalian) age for the deposits studied and reveal episodes of high primary productivity and mostly cool surface waters. Benthic foraminiferal assemblages indicate paleoenvironmental deepening from outer shelf to upper bathyal settings. Deltaic influences may be observed at the base of the studied section in outer shelf (possibly upper bathyal) environments with oxygenated bottom water and episodic high primary productivity, as a consequence of nutrient input from the land. Changes in paleobathymetry resulted in reduction of primary productivity. The benthic assemblages from the uppermost part of the section are dominated by tubular agglutinated foraminifera and indicate an upper bathyal setting with low organic flux to the sea floor. The sediments of the studied section were deposited during the late stage of the first early Miocene relative sea-level rise in the Transylvanian Basin.
� Foraminifera;
� paleoecology;
� paleoenvironments;
� early Miocene;
� Transylvanian Basin;
� Central Paratethys.
Sz�kely S-F., Bindiu-Haitonic R., Filipescu S. & Bercea R. (2017).- Biostratigraphy and paleoenvironmental reconstruction of the marine lower Miocene Chechiș Formation in the Transylvanian Basin based on foraminiferal assemblages.- Carnets Geol., Madrid, vol. 17, no. 2, p. 11-37.
Microbiostratigraphie (foraminif�res) et reconstitution pal�oenvironnementale de la Formation marine de Chechiș (Mioc�ne inf�rieur) dans le Bassin Transylvanien.- Les associations de foraminif�res planctoniques et benthiques sont exploit�es � des fins biostratigraphiques et de reconstitution des pal�oenvironnements de la Formation Chechiș du Mioc�ne inf�rieur marin dans la coupe de Gălp�ia (district de Sălaj, Roumanie, NW du Bassin transylvanien). Les foraminif�res planctoniques sugg�rent un �ge eggenburgien (Burdigalien) pour les d�p�ts objets de cette �tude. Ils r�v�lent en outre l'existence d'�pisodes de forte productivit� primaire et la pr�sence d'eaux de surface g�n�ralement froides. Les associations de foraminif�res benthiques indiquent un approfondissement des pal�oenvironnements depuis le bord externe du plateau continental � la partie sup�rieure de l'�tage bathyal. Des influences delta�ques peuvent �tre observ�es � la base de la coupe au sein des environnements du bord externe du plateau continental (peut-�tre m�me dans la partie sup�rieure de l'�tage bathyal) avec des eaux de fond oxyg�n�es et des �pisodes de forte productivit� primaire en raison d'apports de nutriments en provenance des terres �merg�es. Des variations pal�obathym�triques ont entra�n� une diminution de la productivit� primaire. Les associations de formes benthiques de la partie sommitale de la coupe sont domin�es par des foraminif�res tubulaires � test agglutinant, marqueurs de la partie sup�rieure de l'�tage bathyal, avec un faible flux de mati�re organique vers les fonds marins. Les s�diments de la coupe �tudi�e se sont d�pos�s au cours du stade ultime de la premi�re mont�e relative du niveau de la mer enregistr�e au Mioc�ne inf�rieur dans le Bassin transylvanien.
� foraminif�res ;
� pal�o�cologie ;
� pal�oenvironnements ;
� Mioc�ne inf�rieur ;
� Bassin Transylvanien ;
� Parat�thys centrale.
As one of the major basins of the Central Paratethys (Laskarev, 1924; B�ldi, 1969; R�gl, 1998), the Transylvanian Basin evolved as a sag basin during the Oligocene, while its Miocene sediments were deposited in a flexural basin (Kr�zsek & Bally, 2006). Initial marine flooding was followed by major shallowing during the early Miocene, as indicated by well data and regional interpretations (Dicea et al., 1980; Kr�zsek & Bally, 2006; Tischler et al., 2008). The marine lower Miocene is best developed in the northwestern part of the Transylvanian Basin, where various paleoenvironmental settings influenced the distribution of its fossil assemblages.
A detailed micropaleontological study was carried out on the lower Miocene deposits (Gălp�ia section in the Chechiş Formation) in order to provide an accurate biostratigraphic framework and paleoenvironmental reconstructions.
The lithostratigraphy is based on the synthesis of S. Filipescu (2011). The Chechiș Formation (Hofmann, 1879) overlies the nearshore Coruș or Buzaș formations and is represented by a 20 to 80 m thick offshore marine unit consisting of mudstones (Popescu et al., 1995). In some areas, a glauconitic layer is recorded at the base of the Chechiș Formation (Șuraru, 1967, 1968; Rusu, 1969; Popescu, 1970). The upper boundary is transitional to the Hida Formation or is an unconformity overlain by the middle Miocene Dej Tuff.
The fossil assemblages of the Chechiș Formation consist of abundant foraminifera (Hofmann, 1879; Popescu, 1975) and rare molluscs (Rusu, 1977). Popescu (1970) assigned a Burdigalian age to the Chechiș Formation based on planktonic foraminifera. Previous studies of foraminiferal assemblages focused primarily on taxonomy and biostratigraphy (Şuraru, 1952; Rusu & Popescu, 1965; Popescu, 1971, 1975; Popescu & Iva, 1971; Nicorici et al., 1979), while paleoecological and paleogeographic considerations were addressed only recently (Sz�kely et al., 2016).
Continuous sedimentation in the Gălp�ia section allows the description of the biostratigraphy of the marine early Miocene (Eggenburgian or Burdigalian) and of change in the assemblages of benthic and planktonic foraminifera as a result of environmental variation.
|
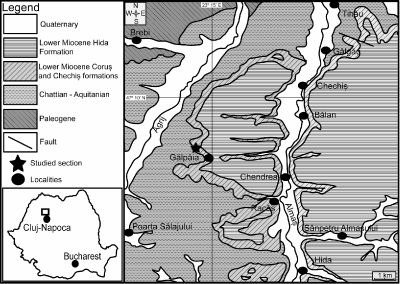
Figure 1:
Geological map and location of the Gălp�ia
section (modified after the Geological Map of Romania, 1:200000, Sheet Cluj;
Petrescu & Drăghici, 1964; Beldean & S. Filipescu,
2011). |
Fourteen
micropaleontological samples were collected from the lower Miocene Chechiș
Formation near Gălp�ia village (47�8'15.30"N
23�14'13.11"E; Fig. 1 ![]() ),
50
km NNW of Cluj-Napoca. The
sampled interval (7 m thick) consists of massive dark grayish mudstone with
very rare laminar fine sandstone (Fig. 2
),
50
km NNW of Cluj-Napoca. The
sampled interval (7 m thick) consists of massive dark grayish mudstone with
very rare laminar fine sandstone (Fig. 2 ![]() ). Clay nodules and coal
have also been identified. Some levels are characterized by moderate
bioturbation (vertical and horizontal burrows). Yellowish-red
centimetric altered mudstone is locally present (Fig. 2
). Clay nodules and coal
have also been identified. Some levels are characterized by moderate
bioturbation (vertical and horizontal burrows). Yellowish-red
centimetric altered mudstone is locally present (Fig. 2 ![]() ).
).
Sediment samples were processed using standard micropaleontological methods. Approximately 300 foraminiferal tests were picked from the >63 �m fraction and identified under the stereomicroscope, while representative specimens were examined in detail using the scanning electron microscope.
Paleoecological proxies are inferred from univariate statistics (such as relative abundance, percentage of microhabitats, diversity indices), multivariate statistics (hierarchical clustering, SIMPER analysis) and agglutinated foraminiferal morphogroups.
The relative abundance of a certain benthic foraminiferal species or groups with similar paleoecological affinities is represented by the ratio between the number of specimens of the selected species (or group) and the total number of benthic specimens per sample.
The diversity indices (Fisher's Alpha and Shannon-Wiener; Hammer & Harper, 2006) were calculated for the benthic assemblages using the software PAST of Hammer et al. (2001).
Identification of the microhabitats of calcareous benthic foraminifera as epifaunal, shallow-infaunal and infaunal was inferred from the morphotypes proposed by Corliss (1985, 1991) and Corliss & Chen (1988) or taken from the literature (e.g., R�gl & Spezzaferri, 2003; Murray, 2006). The abundance of different microhabitats can be used to indicate organic carbon flux to the sea floor and bottom-water oxygenation (Corliss & Chen, 1988; Jorissen et al., 1995; Jorissen, 2002). Agglutinated foraminifer morphogroup analysis is based on the concept that foraminiferal individuals with different test shapes occupy different habitats and have different feeding strategies, so that changes in the relative abundance of morphogroups in fossil assemblages reflect environmental changes (Corliss, 1985; Jones & Charnock, 1985; Murray et al., 2011). Agglutinated foraminifer morphogroup analysis was performed according to the scheme developed by Kaminski et al. (2005).
Multivariate statistics were applied separately to the quantitative data of the benthic and planktonic assemblages using Primer 6 software (Clarke & Warwick, 2001), the analysis including hierarchical clustering. Raw data were double square root-transformed prior to the generation of a resemblance matrix based on the Similarity (Clifford & Stephenson, 1975). With this process, the contribution of most abundant species was limited (Field et al., 1982). Group Average Linking was used for hierarchical agglomerative clustering. Based on the same similarity matrix a SIMPER analysis was included to confirm taxa defining the clusters (Clarke, 1993).
A summary of the paleoecological preferences of the most representative benthic foraminifera (species or groups with similar paleoecological affinities) was compiled for the section studied. The information gathered includes, where available: paleobathymetry, microhabitat, oxygen preferences, temperature, and additional ecological data (Table 1). The paleoecological preferences of planktonic foraminifera with respect to surface water temperature and productivity were also summarized (Table 2).
Table 1. Paleoecological preferences of most common species/genera or groups with similar paleoecological affinities compiled for the Gălp�ia section. References: (a). Corliss & Chen (1988); (b). Spezzaferri & Ćorić (2001); (c). Morkhoven et al. (1986); (d). Murray (2006); (e). De Man (2006); (f). R�gl & Spezzaferri (2003); (g). Spezzaferri et al. (2002); (h). Spezzaferri et al. (2004); (i). Kaminski et al. (2005); (j). Reolid & Nagy (2008); (k). Z�gorsek et al. (2007); (l). Grunert et al. (2012); (m). Kaiho (1994); (n). B�ldi (2006); (o). Almogi-Labin et al. (2000); (p). Miao & Thunell (1993); (q). Schmiedl et al. (2000); (r). Mullins et al. (1985).
| Species\Genera\Group | Paleobathymetry | Microhabitat | Oxygen | Temperature/Comments |
| Alabamina spp. | Epifaunal (e) | Suboxic (e), (m) | ||
| Ammodiscus spp. | High-energy lagoons and estuary (i) | Surficial epifaunal (i) | Active and passive deposit feeding (i) | |
| Amphicoryna spp. | Inner shelf-bathyal (f), 13-3000 m (f) | Low-oxygen (f) | Mud, glacial water (f) | |
| Bathysiphon taurinensis | Bathyal and abyssal (i) | Erect epifaunal (i) | Tranquil environment and low organic matter flux (i) | |
| Bolivina spp. | Inner shelf-bathyal (d) | Infaunal (a), epifaunal-infaunal (d), deep-infaunal (l) | Low-oxygen (g), some species tolerate dysoxia (d), dysoxic (m) | Cold-warm, free, muddy sediment, detritivore? (d) |
| Budashevaella spp. | Shelf to deep marine (i) | Surficial epifaunal (i) | Active deposit feeding (i) | |
| Bulimina elongata | Upper neritic (c), 50-2500 m, abundant down to 80-100 m (h) | Low-oxygen (h) | Mud and muddy sand, river mouths, high organic matter (h), organic carbon preferring (g) | |
| Bulimina spp. | Inner shelf-bathyal (d) | Infaunal (d) | Some species tolerate dysoxia (d) | Cold-temperate, free, mud-fine sand, detritivore? (d), organic carbon preferring (g) |
| Chilostomella oolina | Outer shelf-bathyal (d) | Deep infaunal (d) | Tolerates dysoxia (d), low-oxygen (g) | Free, mud, detritivore (d) |
| Cibicidoides pachyderma | 30-3500 m (h) | Epifaunal, shallow-infaunal (d),(o),(p),(q) | Oxic (h),(q) | Passive suspension feeder, high energy (d), oligotrophic environment, stable physico-chemical conditions (o),(p),(q) |
| Cibicidoides\Heterolepa spp. | Shelf-bathyal (d), 30-3500 m (h) | Epifaunal (d) | Oxic (f),(m) | Cold, clinging (d), hard substrates (d),(f), passive suspension feeder? (d) |
| Glomospira spp. | High-energy lagoons and estuary (i) | Surficial epifaunal (i) | Active and passive deposit feeding (i) | |
| Hansenisca soldanii | Epifaunal (d) | Suboxic, dysoxic (d),(r) | Mud (d) | |
| Haplophragmoides spp. | Marshes-bathyal (d), inner shelf and upper bathyal (i) | Infaunal to surficial (b), epifaunal-shallow infaunal (d),(i) | Cold-temperate, free, mud-sand, detritivore? (d), active deposit feeding (i) | |
| Heterolepa dutemplei | Outer neritic-upper bathyal (c), 50-3000 m (h) | Epifaunal (d) | Oxic (d) | Cold-temperate, clinging?, hard substrates, passive suspension feeder? (d) |
| Hyperammina\Nothia\Rhabdammina\ Rhizammina\Psamosiphonella\Nothia spp. | Bathyal and abyssal (i) | Erect epifaunal (i) | Tranquil environment and low organic matter flux (i) | |
| Karreriella\Karrerulina spp. | Inner shelf - upper bathyal (i) | Deep infaunal (i) | Active deposit feeding and increased organic matter flux (i) | |
| Laevidentalina spp. | Circalittoral-bathyal, 100-4000 m (f) | Suboxic, dysoxic (f) | Mud (f) | |
| Lenticulina spp. | Outer shelf-bathyal (d), from 20 m down (f) | Epifaunal (a),(d),(e),(l) | Suboxic (e),(f),(l),(m), oxic (n) | Cold (d), free, mud, detritivore? (d),(e) |
| Melonis pompilioides | Circalittoral-bathyal, 50-4000 m (f) | Infaunal (d) | Suboxic (f) | Mud, high organic matter (f), high primary productivity (g) |
| Praeglobobulimina pupoides | 30-4000 m (h) | Low-oxygen (g), dysoxic (k) | Stress marker (k) | |
| Praeglobobulimina spp. | Circalittoral to bathyal, preferred depth 80-800 m (b) | Dysoxic (b) | High primary productivity (g) | |
| Recurvoides spp. | Shelf to deep marine (i) | Surficial epifaunal (i), epifaunal to shallow infaunal (j) | Active deposit feeding (i) | |
| Reophax spp. | Inner shelf-upper bathyal (i) | Deep infaunal (i) | Active deposit feeding and increased organic matter flux (i) | |
| Reticulophragmium spp. | Inner shelf-upper bathyal (i) | Surficial epifaunal-shallow infaunal (i) | Active deposit feeding (i) | |
| Saccammina spp. | Bathyal and abyssal (i) | Shallow infaunal (i) | Suspension feeding, passive deposit feeding (i) | |
| Stilostomella\Mylostomella\Neugeborina spp. | 230-2500 m (f) | Suboxic (f) | Mud (f) | |
| Trochammina spp. | Shelf to deep marine (i) | Surficial epifaunal (i) | Active deposit feeding (i) | |
| Uvigerina spp. | 100 to >4500 m, rarely shallower than 100 m (f), shelf-abyssal (d) | Epifaunal-infaunal (d), infaunal (a) | Suboxic (f), (m), low-oxygen (g) | Cold, free, muddy sediments, detritivore? (d), high organic matter (f), high primary productivity (g) |
| Valvulineria spp. | Circalittoral to epibathyal, more abundant between 40-100 m (f) | Suboxic (f), dysoxic (b) | Mud, coastal terrigenous mud, high organic matter (f) |
Table 2. Division of planktonic foraminifera based on water temperature preference (modified after Sz�kely & S. Filipescu, 2016). References: (1). Spezzaferri (1994); (2). Spezzaferri (1995); (3). Spezzaferri & Ćorić (2001); (4). R�gl & Spezzaferri (2003); (5). Bicchi et al. (2003); (6). Bicchi et al. (2006); (7). Pearson et al. (2006); (8). Li et al. (1992); (9). Spezzaferri et al. (2002); (10). Kroon (1988); (11). Roetzel et al. (2006); (12). Amore et al. (2004).
| Latitude \ Temperature indicator |
High \ Cool | High- middle \ Cool- temperate |
Middle \ Cosmopolitan \ No diagnostic |
Middle- low \ Temperate- warm |
Low \ Warm | Productivity | |
| Species\Genera | Cool-temperate | Globigerina anguliofficinalis |
Globigerina anguliofficinalis |
||||
| Globigerina gnaucki (2)(10) | |||||||
| Globigerina lentiana (2)(3)(4)(5)(6)(9) | |||||||
| Globigerina officinalis (1)(2)(4)(5)(6) | Globigerina officinalis (7) | ||||||
| Globigerina ouachitensis (1)(2)(4)(5)(6) | Globigerina ouachitensis (7) | ||||||
| Globigerina ottnangiensis (3)(4)(9) | Globigerina ottnangiensis (2) | High productivity (4) | |||||
| Globigerina praebulloides (1)(2)(3)(4)(5)(6)(9) | |||||||
| Globigerina spp. (2)(4)(5)(6) | Upwelling areas (10) | ||||||
| Globigerina tarchanensis (4) | High productivity (4) | ||||||
| Globoturborotalita connecta (2)(6) | |||||||
| Globoturborotalita woodi (1)(4) | Globoturborotalita woodi (1)(2)(5)(6) | Globoturborotalita woodi (3) | Globoturborotalita woodi (9) | ||||
| Tenuitella clemenciae (1)(4)(8) | |||||||
| Tenuitella gemma (1)(4)(8) | Tenuitella gemma (7) | ||||||
| Tenuitellinata juvenilis (1)(3)(4)(8)(9)(11) | Tenuitellinata juvenilis (1)(2) | Upwelling conditions (11) | |||||
| Tenuitella spp. (1)(2)(4)(8) | |||||||
| Turborotalita quinqueloba (3)(4)(9)(11) | Upwelling conditions (11) | ||||||
| Warm- temperate |
Globigerinoides trilobus (2)(3)(5)(6)(9)(12) | ||||||
| Tenuitellinata angustiumbilicata (3)(9)(11) | Tenuitellinata angustiumbilicata (1)(2)(5)(6)(7) | Upwelling conditions (11) | |||||
| No diagnostic |
Globigerinella obesa (2)(3) | Globigerinella obesa (5) | Globigerinella obesa (9) | ||||
| Paragloborotalia nana (1)(2) | Paragloborotalia nana (1) | Paragloborotalia nana (7) | Paragloborotalia nana (4)(7) | ||||
|
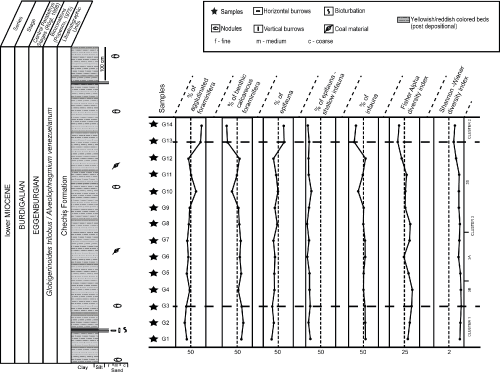
Figure
2:
Graph representing identified biozones, sedimentary log of outcrop
studied and positions of samples, and univariate statistics (P/B ratio,
percent of agglutinated foraminifera, percent of calcareous benthic
foraminifera, microhabitat of benthic foraminifera: percent epifauna,
epifauna - shallow
infauna and infauna, Fisher's alpha and Shannon-Wiener
diversity indices) for Gălp�ia
section. |
4.1. Foraminiferal assemblages
Foraminiferal assemblages are abundant throughout the studied section and the preservation of specimens varies from moderate to good. A total of 139 benthic and 36 planktonic foraminiferal species were identified (Appendix).
The
species diversity values, expressed by the Fisher's alpha diversity index, vary
along the section and range from 13.8 to 37.9, while the Shannon-Wiener
diversity index displays values between 2.6 and 3.6. Both indices are marked
by a decrease in the upper part of the outcrop (Fig. 2 ![]() ).
).
Except for the flattened irregular Ammolagena specimens (M3b morphogroup) all agglutinated foraminiferal morphogroups defined by Kaminski et al. (2005) are present. The graphic distribution of the morphogroups reveals several distinct features within the section: a constant distribution of all the morphogroups from sample G1 to G3, a sharp increase of morphogroup M1 and constant values for the rest of the morphogroups for samples G3-G13 and high abundances of morphogroup M1 (79.83%) and lower values for the rest of the morphogroups in the uppermost part of the section (samples G13-G14).
4.1.1. Statistical data on planktonic foraminiferal assemblages
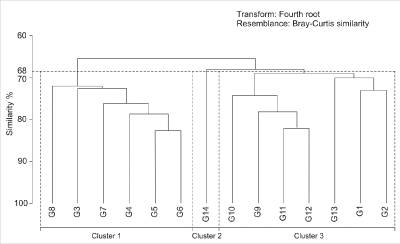
A
quantitative examination of planktonic foraminifera was performed on the samples. Three cluster groups (Cluster 1, Cluster 2,
and Cluster 3) were delimited in the planktonic assemblages at the 68% similarity level (Fig. 4 ![]() ) based on SIMPER analysis
(Table 3) and visual investigation of species composition.
) based on SIMPER analysis
(Table 3) and visual investigation of species composition.
Cluster 1 contains
samples G3 - G8 (Figs. 2 ![]() and 4
and 4 ![]() ).
The most representative species or groups are Globigerina
officinalis, G. spp., G. praebulloides, G. lentiana,
Tenuitellinata angustiumbilicata, Turborotalita
quinqueloba, Globigerina
ottnangiensis, Globoturborotalita
connecta, Globigerina ouachitensis,
G. gnaucki, Tenuitellinata juvenilis,
and Globigerinella obesa (Appendix and Fig. 3
).
The most representative species or groups are Globigerina
officinalis, G. spp., G. praebulloides, G. lentiana,
Tenuitellinata angustiumbilicata, Turborotalita
quinqueloba, Globigerina
ottnangiensis, Globoturborotalita
connecta, Globigerina ouachitensis,
G. gnaucki, Tenuitellinata juvenilis,
and Globigerinella obesa (Appendix and Fig. 3 ![]() ). Twelve species/groups account for 81% of the average
similarity within this cluster (Table 3).
). Twelve species/groups account for 81% of the average
similarity within this cluster (Table 3).
Cluster 2
contains only sample G14 (Figs. 2 ![]() and 4
and 4 ![]() ). The most abundant species or groups
within this cluster are: Globigerina
praebulloides, G. spp., G. lentiana, G. ottnangiensis,
Globoturborotalita connecta and Globigerina
officinalis (Appendix and Fig. 3
). The most abundant species or groups
within this cluster are: Globigerina
praebulloides, G. spp., G. lentiana, G. ottnangiensis,
Globoturborotalita connecta and Globigerina
officinalis (Appendix and Fig. 3 ![]() ).
).
Cluster 3
contains samples G1 - G2 and G9 - G13 (Figs. 2 ![]() and 4
and 4 ![]() ).
The dominant
species or groups within this cluster are: Globigerina
praebulloides, G. ottnangiensis, G.
lentiana, G. spp., G. officinalis, Tenuitellinata
angustiumbilicata, Turborotalita
quinqueloba, Globoturborotalita woodi,
Globigerina anguliofficinalis, and Paragloborotalia
nana (Appendix and Fig. 3
).
The dominant
species or groups within this cluster are: Globigerina
praebulloides, G. ottnangiensis, G.
lentiana, G. spp., G. officinalis, Tenuitellinata
angustiumbilicata, Turborotalita
quinqueloba, Globoturborotalita woodi,
Globigerina anguliofficinalis, and Paragloborotalia
nana (Appendix and Fig. 3 ![]() ). Ten species/groups account for 80% of the
average similarity within this group (Table 3).
). Ten species/groups account for 80% of the
average similarity within this group (Table 3).
|
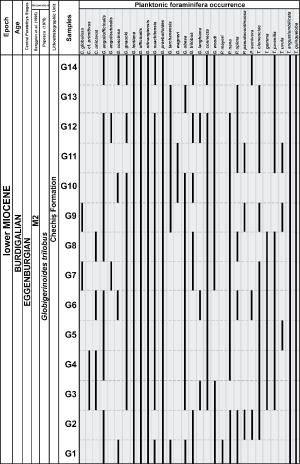
Figure
3: Graph representing occurrences of most
important planktonic foraminiferal species of Gălp�ia section.
Black lines indicate presence of species in respective sample. |
|
|
Figure
4:
Dendrogram showing the hierarchical agglomerative clustering based on the
similarity matrix of planktonic foraminifera. Horizontal dashed
line represents the similarity cut and vertical dashed lines represent
delimitations of resulting clusters. |
4.1.2. Statistical data on benthic foraminiferal assemblages
A
quantitative investigation of benthic foraminifera was performed on the samples. Three cluster groups were separated (Cluster 1, Cluster 2
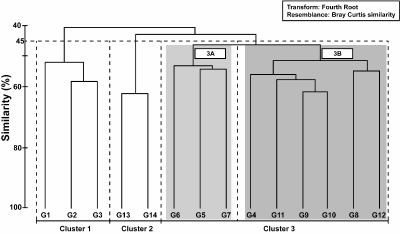
and Cluster 3) at the similarity level of 45% (Fig. 5 ![]() )
based on SIMPER analysis (Table 4) and visual inspection of species composition in benthic assemblages.
)
based on SIMPER analysis (Table 4) and visual inspection of species composition in benthic assemblages.
|
Figure
5:
Dendrogram showing hierarchical
agglomerative clustering based on the similarity matrix of the
benthic foraminifera. Horizontal dashed line represents similarity cut and
vertical dashed lines represent boundaries of resulting clusters. |
Table 3. Similarity-based SIMPER analysis of the resultant clusters derived from planktonic foraminiferal assemblages.
| Group Cluster 1 | Av.Abund | Av.Sim | Contrib% | Cum.% | Group Cluster 3 | Av.Abund | Av.Sim | Contrib% | Cum.% |
| Average similarity: 74.68 |
Average similarity: 71.56 |
||||||||
| Globigerina officinalis | 2.33 | 7.39 | 9.9 | 9.9 | Globigerina praebulloides | 2.44 | 8.43 | 11.8 | 11.8 |
| Globigerina sp. | 2.21 | 6.97 | 9.33 | 19.2 | Globigerina ottnangiensis | 2.4 | 7.9 | 11 | 22.8 |
| Globigerina praebulloides | 2.09 | 6.66 | 8.92 | 28.2 | Globigerina lentiana | 1.98 | 6.86 | 9.58 | 32.4 |
| Globigerina lentiana | 2.05 | 6.36 | 8.52 | 36.7 | Globigerina sp. | 1.93 | 6.75 | 9.43 | 41.8 |
| Tenuitellinata angustiumbilicata | 1.9 | 5.19 | 6.95 | 43.6 | Globigerina officinalis | 1.97 | 6.66 | 9.31 | 51.1 |
| Turborotalita quinqueloba | 1.7 | 4.56 | 6.11 | 49.7 | Tenuitellinata angustiumbilicata | 1.6 | 5.37 | 7.5 | 58.6 |
| Globigerina ottnangiensis | 1.55 | 4.31 | 5.77 | 55.5 | Turborotalita quinqueloba | 1.57 | 4.99 | 6.98 | 65.6 |
| Globoturborotalita connecta | 1.48 | 4.05 | 5.42 | 60.9 | Globoturborotalita woodi | 1.46 | 4.4 | 6.15 | 71.8 |
| Globigerina ouachitensis | 1.36 | 3.92 | 5.24 | 66.2 | Globigerina anguliofficinalis | 1.19 | 3.21 | 4.48 | 76.3 |
| Globigerina gnaucki | 1.32 | 3.84 | 5.14 | 71.3 | Paragloborotalia nana | 1.07 | 2.9 | 4.05 | 80.3 |
| Tenuitellinata juvenilis | 1.22 | 3.63 | 4.86 | 76.2 | Globigerina tarchanensis | 0.98 | 2.58 | 3.6 | 83.9 |
| Globigerinella obesa | 1.23 | 3.6 | 4.82 | 81 | Tenuitella clemenciae | 0.86 | 1.96 | 2.74 | 86.6 |
| Tenuitella gemma | 1.18 | 3.58 | 4.79 | 85.8 | Paragloborotalia opima | 0.77 | 1.88 | 2.63 | 89.3 |
| Tenuitella clemenciae | 1.33 | 3.13 | 4.19 | 90 | Globigerina ouachitensis | 0.85 | 1.67 | 2.33 | 91.6 |
| Paragloborotalia nana | 0.94 | 2.21 | 2.96 | 92.9 | Globoturborotalita connecta | 0.95 | 1.53 | 2.13 | 93.7 |
| Globigerina anguliofficinalis | 0.79 | 1.38 | 1.84 | 94.8 | Globigerinoides trilobus | 0.64 | 1.09 | 1.53 | 95.3 |
| Catapsydrax unicavus | 0.75 | 1.37 | 1.84 | 96.6 | Globigerinella obesa | 0.72 | 1.08 | 1.51 | 96.8 |
| Tenuitella sp. | 0.67 | 0.83 | 1.11 | 97.7 | Tenuitellinata juvenilis | 0.55 | 0.65 | 0.9 | 97.7 |
| Globoturborotalita woodi | 0.63 | 0.78 | 1.04 | 98.7 | Globigerina gnaucki | 0.56 | 0.52 | 0.73 | 98.4 |
| Globigerinoides trilobus | 0.63 | 0.27 | 0.36 | 99.1 | Paragloborotalia pseudocontinuosa | 0.43 | 0.51 | 0.72 | 99.1 |
| Catapsydrax cf. primitivus | 0.4 | 0.24 | 0.32 | 99.4 | Globigerina concinna | 0.29 | 0.17 | 0.24 | 99.4 |
| Globoquadrina langhiana | 0.33 | 0.22 | 0.3 | 99.7 | Globigerina wagneri | 0.29 | 0.16 | 0.22 | 99.6 |
| Paragloborotalia opima | 0.33 | 0.2 | 0.27 | 100 | Tenuitella uvula | 0.31 | 0.16 | 0.22 | 99.8 |
Cluster 1 contains
samples G1 to G3 from the base of the
outcrop (Figs. 2 ![]() and 5
and 5 ![]() ). The benthic
foraminiferal assemblages are composed of
calcareous (dominant) and agglutinated forms (Fig. 2
). The benthic
foraminiferal assemblages are composed of
calcareous (dominant) and agglutinated forms (Fig. 2 ![]() ). The diversity indices
display high values, with the Fisher's alpha index ranging from 30.6 to 36.8 and the Shannon-Wiener
index from 3.4 to 3.6 (Fig. 2
). The diversity indices
display high values, with the Fisher's alpha index ranging from 30.6 to 36.8 and the Shannon-Wiener
index from 3.4 to 3.6 (Fig. 2 ![]() ). Benthic infaunal and
epifaunal microhabitats are relatively well represented, and the
epifaunal-shallow infaunal microhabitat less so (Fig. 2
). Benthic infaunal and
epifaunal microhabitats are relatively well represented, and the
epifaunal-shallow infaunal microhabitat less so (Fig. 2 ![]() ).
).
Table 4. Similarity-based SIMPER analysis of the resultant clusters derived from benthic foraminiferal assemblages.
| Group Cluster 1 | Av.Abund | Av.Sim | Contrib% | Cum.% | Group Cluster 3B | Av.Abund | Av.Sim | Contrib% | Cum.% |
| Average similarity: 54.31 |
Average similarity: 54.68 |
||||||||
| Globigerina officinalis | 2.33 | 7.39 | 9.9 | 9.9 | Globigerina praebulloides | 2.44 | 8.43 | 11.8 | 11.8 |
| Uvigerina popescui | 2.36 | 3.18 | 5.86 | 5.86 | Hyperammina / Rhabdammina sp. | 2.21 | 3.71 | 6.79 | 6.79 |
| Praeglobobulimina pupoides / affinis | 1.79 | 2.5 | 4.61 | 10.47 | Uvigerina popescui | 1.96 | 3.02 | 5.53 | 12.32 |
| Cibicidoides pachyderma | 1.58 | 2.05 | 3.77 | 14.24 | Praeglobobulimina pupoides / affinis | 1.75 | 2.91 | 5.32 | 17.64 |
| Reticulophragmium rotundidorsatum | 1.46 | 1.95 | 3.6 | 17.84 | Reophax sp. | 1.46 | 2.46 | 4.5 | 22.14 |
| Marginulina hirsuta | 1.41 | 1.92 | 3.53 | 21.37 | Cibicidoides pachyderma | 1.45 | 2.38 | 4.36 | 26.5 |
| Lenticulina inornata | 1.52 | 1.9 | 3.5 | 24.87 | Lenticulina arcuatostriata | 1.3 | 2.05 | 3.75 | 30.25 |
| Reticulophragmium venezuelanum | 1.47 | 1.87 | 3.44 | 28.31 | Haplophragmoides sp. | 1.28 | 2.04 | 3.74 | 33.99 |
| Uvigerina farinosa | 1.42 | 1.83 | 3.37 | 31.69 | Hansenisca soldanii | 1.22 | 1.99 | 3.63 | 37.62 |
| Reticulophragmium acutidorsatum | 1.31 | 1.75 | 3.22 | 34.91 | Bathysiphon taurinensis | 1.16 | 1.88 | 3.43 | 41.05 |
| Hansenisca soldanii | 1.29 | 1.69 | 3.11 | 38.02 | Valvulineria palmarealensis | 1.14 | 1.86 | 3.4 | 44.45 |
| Melonis pompilioides | 1.29 | 1.57 | 2.9 | 40.92 | Recurvoides sp. | 1.08 | 1.79 | 3.28 | 47.73 |
| Haplophragmoides sp. | 1.29 | 1.57 | 2.89 | 43.81 | Melonis pompilioides | 1.03 | 1.29 | 2.36 | 50.09 |
| Glomospira charoides | 1.13 | 1.51 | 2.79 | 46.59 | Karreriella chilostoma | 1.04 | 1.27 | 2.33 | 52.42 |
| Nothia sp. | 1.25 | 1.51 | 2.79 | 49.38 | Saccamina grzybowski | 1.06 | 1.24 | 2.27 | 54.69 |
| Bathysiphon taurinensis | 1.17 | 1.51 | 2.78 | 52.16 | Nothia sp. | 0.98 | 1.23 | 2.25 | 56.93 |
| Lenticulina arcuatostriata | 1.23 | 1.51 | 2.78 | 54.94 | Hyperammina rugosa | 0.98 | 1.16 | 2.12 | 59.06 |
| Haplophragmoides vasiceki vasiceki | 1.2 | 1.51 | 2.78 | 57.72 | Bulimina elongata | 0.88 | 1.1 | 2.02 | 61.08 |
| Laevidentalina inornata | 1.17 | 1.51 | 2.78 | 60.5 | Reticulophragmium rotundidorsatum | 0.99 | 1.1 | 2.02 | 63.09 |
| Laevidentalina elegans | 1.06 | 1.42 | 2.62 | 63.11 | Reticulophragmium venezuelanum | 0.96 | 1.08 | 1.98 | 65.08 |
| Karrerulina apicularis | 1.19 | 1.42 | 2.62 | 65.73 | Amphicoryna armata | 0.8 | 1.08 | 1.98 | 67.06 |
| Karrerulina conversa | 1.06 | 1.42 | 2.62 | 68.35 | Hyperammina elongata | 0.8 | 1.07 | 1.96 | 69.01 |
| Reophax sp. | 1.14 | 1.42 | 2.62 | 70.96 | Trochammina sp. | 0.84 | 1.05 | 1.92 | 70.93 |
| Saccamina grzybowski | 1.11 | 1.42 | 2.62 | 73.58 | Ammobaculites agglutinans | 0.8 | 1.04 | 1.91 | 72.84 |
| Stilostomella adolphina | 1.06 | 1.42 | 2.62 | 76.2 | Martinottiella communis | 0.84 | 1.04 | 1.91 | 74.75 |
| Lenticulina sp. | 0.91 | 0.64 | 1.18 | 77.37 | Cibicidoides pygmeus | 0.86 | 1.03 | 1.89 | 76.64 |
| Uvigerina acuminata | 0.91 | 0.64 | 1.18 | 78.55 | Laevidentalina elegans | 0.84 | 1.03 | 1.89 | 78.53 |
| Neugeborina boueana | 0.89 | 0.58 | 1.06 | 79.61 | Lenticulina sp. | 0.78 | 0.65 | 1.19 | 79.72 |
| Uvigerina mantaensis | 0.79 | 0.58 | 1.06 | 80.67 | Praeglobobulimina ovata | 0.79 | 0.61 | 1.12 | 80.84 |
| Rhabdammina sp. | 0.84 | 0.56 | 1.04 | 81.71 | Subreophax sp. | 0.73 | 0.6 | 1.1 | 81.94 |
| Ammodiscus cretaceus | 0.73 | 0.49 | 0.89 | 82.6 | Reticulophragmium acutidorsatum | 0.72 | 0.57 | 1.05 | 82.99 |
| Haplophragmoides horridus | 0.67 | 0.49 | 0.89 | 83.49 | Uvigerina farinosa | 0.7 | 0.56 | 1.03 | 84.02 |
| Group Cluster 2 | Av.Abund | Av.Sim | Contrib% | Cum.% | Group Cluster 3A | Av.Abund | Av.Sim | Contrib% | Cum.% |
| Average similarity: 62.57 |
Average similarity: 53.84 |
||||||||
| Hyperammina / Rhabdammina sp. | 2.54 | 5.71 | 9.12 | 9.12 | Hyperammina / Rhabdammina sp. | 1.93 | 3.79 | 7.04 | 7.04 |
| Hyperammina rugosa | 2.03 | 4.65 | 7.43 | 16.55 | Praeglobobulimina pupoides / affinis | 1.59 | 2.76 | 5.12 | 12.17 |
| Bathysiphon taurinensis | 1.65 | 3.7 | 5.91 | 22.47 | Uvigerina popescui | 1.45 | 2.71 | 5.04 | 17.2 |
| Nothia sp. | 1.65 | 3.7 | 5.91 | 28.38 | Reophax sp. | 1.41 | 2.71 | 5.04 | 22.24 |
| Praeglobobulimina pupoides / affinis | 1.62 | 3.7 | 5.91 | 34.29 | Melonis pompilioides | 1.35 | 2.65 | 4.91 | 27.15 |
| Rhabdammina sp. | 1.49 | 3.34 | 5.34 | 39.63 | Alabamina polita | 1.31 | 2.39 | 4.44 | 31.59 |
| Saccamina grzybowski | 1.55 | 3.34 | 5.34 | 44.97 | Hansenisca soldanii | 1.29 | 2.39 | 4.44 | 36.03 |
| Nothia excelsa | 1.3 | 2.81 | 4.49 | 49.47 | Valvulineria palmarealensis | 1.28 | 2.29 | 4.26 | 40.29 |
| Reophax sp. | 1.3 | 2.81 | 4.49 | 53.96 | Hyperammina rugosa | 1.27 | 2.22 | 4.13 | 44.41 |
| Reticulophragmium acutidorsatum | 1.25 | 2.81 | 4.49 | 58.45 | Saccamina grzybowski | 1.17 | 2.14 | 3.97 | 48.38 |
| Bolivina beyrichi carinata | 1 | 2.36 | 3.78 | 62.23 | Stilostomella adolphina | 1.13 | 2.14 | 3.97 | 52.35 |
| Hansenisca soldanii | 1 | 2.36 | 3.78 | 66 | Bolivina beyrichi carinata | 1.14 | 2.01 | 3.73 | 56.08 |
| Hyperammina elongata | 1.16 | 2.36 | 3.78 | 69.78 | Bulimina alsatica | 1.11 | 2.01 | 3.73 | 59.82 |
| Lenticulina calcar | 1.09 | 2.36 | 3.78 | 73.56 | Cibicidoides pygmeus | 1.11 | 2.01 | 3.73 | 63.55 |
| Melonis pompilioides | 1.09 | 2.36 | 3.78 | 77.34 | Fursenkoina sp. | 1.06 | 2.01 | 3.73 | 67.28 |
| Praeglobobulimina ovata | 1 | 2.36 | 3.78 | 81.11 | Lenticulina inornata | 1.06 | 2.01 | 3.73 | 71.02 |
| Psamosphaera fusca | 1.09 | 2.36 | 3.78 | 84.89 | Nothia sp. | 1.06 | 2.01 | 3.73 | 74.75 |
| Reticulophragmium rotundidorsatum | 1.09 | 2.36 | 3.78 | 88.67 | Uvigerina mantaensis | 1.06 | 2.01 | 3.73 | 78.48 |
| Rhizzamina algaeformis | 1 | 2.36 | 3.78 | 92.45 | Bulimina elongata | 1 | 0.88 | 1.64 | 80.12 |
| Subreophax sp. | 1.25 | 2.36 | 3.78 | 96.22 | Ammosphaeroidina pseudopauciloculata | 0.73 | 0.68 | 1.26 | 81.38 |
| Trochammina sp. | 1 | 2.36 | 3.78 | 100 | Bulimina arndti | 0.73 | 0.68 | 1.26 | 82.64 |
| Cibicidoides pachyderma | 0.77 | 0.68 | 1.26 | 83.91 | |||||
The most
representative benthic calcareous species are Cibicidoides pachyderma, Lenticulina
inornata, Marginulina hirsuta, Praeglobobulimina
pupoides, and Uvigerina popescui,
while the agglutinated foraminiferal assemblages are dominated by Reticulophragmium (Fig. 6 ![]() ) species.
Lower amounts of calcareous benthic forms (Hansenisca soldanii, Laevidentalina
spp., Lenticulina arcuatostriata, Melonis
pompilioides, Neugeborina boueana,
Stilostomella adolphina, Uvigerina
acuminata, Uvigerina farinosa, and Uvigerina mantaensis) and agglutinated species
(Ammodiscus
cretaceus, Bathysiphon taurinensis,
Glomospira charoides, Haplophragmoides
spp., Karrerulina spp., Nothia spp., Reophax spp.,
Rhabdammina spp., and Saccamina
grzybowski) are present in this cluster. Thirty-one species/groups account for 83% of the average
similarity within this group (Table 4).
) species.
Lower amounts of calcareous benthic forms (Hansenisca soldanii, Laevidentalina
spp., Lenticulina arcuatostriata, Melonis
pompilioides, Neugeborina boueana,
Stilostomella adolphina, Uvigerina
acuminata, Uvigerina farinosa, and Uvigerina mantaensis) and agglutinated species
(Ammodiscus
cretaceus, Bathysiphon taurinensis,
Glomospira charoides, Haplophragmoides
spp., Karrerulina spp., Nothia spp., Reophax spp.,
Rhabdammina spp., and Saccamina
grzybowski) are present in this cluster. Thirty-one species/groups account for 83% of the average
similarity within this group (Table 4).
All
agglutinated foraminiferal morphogroups (except M3b) are present in this
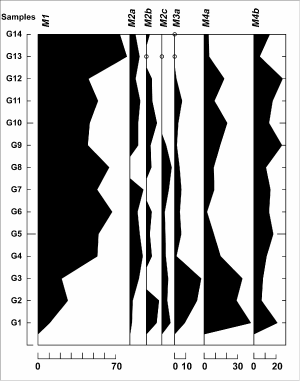
cluster. The tubular agglutinated morphogroup M1 displays an increasing trend (Figs. 6 ![]() and 7
and 7 ![]() ).
Morphogroup M2a
also increases towards the top of the cluster (Fig. 7
).
Morphogroup M2a
also increases towards the top of the cluster (Fig. 7 ![]() ). Agglutinated
foraminifera typical of the shelf (M2c morphogroup) decrease in abundance
towards the top of Cluster 1.
). Agglutinated
foraminifera typical of the shelf (M2c morphogroup) decrease in abundance
towards the top of Cluster 1.
|
Figure
6:
Graph showing relative abundances of most
important genera or groups with similar paleoecological affinities for Gălp�ia
section. |
|
Figure
7:
Graph showingabundance (percent) of agglutinated foraminiferal morphogroups
defined by Kaminski et al. (2005). Abundance of each morphogroup is calculated relative to the total
abundance of agglutinated foraminifera. |
Cluster 2
contains samples from the uppermost part of the outcrop (samples G13 and
G14; Figs. 2 ![]() and 5
and 5 ![]() ). Benthic agglutinated forms clearly dominate these
assemblages. The lowest values of the Fisher's alpha diversity index are found
within this cluster (13.8 and 16.1), while the values of the Shannon-Wiener
diversity index are 2.6 and 2.8 (Fig. 2
). Benthic agglutinated forms clearly dominate these
assemblages. The lowest values of the Fisher's alpha diversity index are found
within this cluster (13.8 and 16.1), while the values of the Shannon-Wiener
diversity index are 2.6 and 2.8 (Fig. 2 ![]() ). Epifaunal forms are relatively well
represented and dominate the benthic assemblages, while the infaunal and
epifaunal-shallow infaunal microhabitats are characterized by low abundance
throughout this cluster (Fig. 2
). Epifaunal forms are relatively well
represented and dominate the benthic assemblages, while the infaunal and
epifaunal-shallow infaunal microhabitats are characterized by low abundance
throughout this cluster (Fig. 2 ![]() ).
).
The dominant calcareous form is Praeglobobulimina pupoides, while among agglutinated foraminifera
the following are common: Bathysiphon
taurinensis, Hyperammina / Rhabdammina
spp., Nothia spp., and Saccamina
grzybowski. Less abundant benthic foraminifera from this cluster are Bolivina
beyrichi carinata, Hansenisca
soldanii, Reophax spp., and Reticulophragmium
acutidorsatum (Fig. 8 ![]() ). Twenty-one species/groups account for 100% of
the average similarity within this group (Table 4).
). Twenty-one species/groups account for 100% of
the average similarity within this group (Table 4).
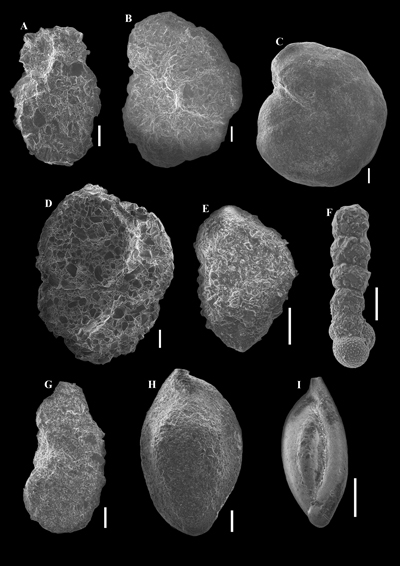
|
Figure
8:
All scales represent 100 μm. |
Cluster 3 was
further divided into two subclusters: 3A
and 3B (Fig. 5 ![]() )
based on the species
composition of the benthic assemblages. Samples of subcluster 3A (G5-G7) are well represented by both calcareous and
agglutinated taxa (Fig. 2
)
based on the species
composition of the benthic assemblages. Samples of subcluster 3A (G5-G7) are well represented by both calcareous and
agglutinated taxa (Fig. 2 ![]() ). The Fisher's alpha diversity indices range between
24.1 and 33.5, and the Shannon-Wiener
diversity indices between 3.2 and 3.5 (Fig. 2
). The Fisher's alpha diversity indices range between
24.1 and 33.5, and the Shannon-Wiener
diversity indices between 3.2 and 3.5 (Fig. 2 ![]() ). Although the epifaunal
microhabitat is well represented, infaunal forms dominate these assemblages (Fig. 2
). Although the epifaunal
microhabitat is well represented, infaunal forms dominate these assemblages (Fig. 2 ![]() ).
).
The dominant benthic calcareous species are Alabamina
polita, Melonis pompilioides, Praeglobobulimina pupoides and Uvigerina
popescui (Fig. 9 ![]() ), while the
agglutinated forms are well represented by species of Hyperammina /
Rhabdammina
and Reophax. Less abundant
benthic foraminifera include Ammosphaeroidina
pseudopauciloculata, Bolivina
beyrichi carinata, Bulimina alsatica, B. arndti, B.
elongata, Cibicidoides pachyderma, C.
pygmeus, Fursenkoina spp., Hansenisca
soldanii, Lenticulina spp.,
Nothia spp., Saccamina grzybowski, Stilostomella
adolphina, Uvigerina mantaensis,
and Valvulineria palmarealensis. Twenty-two
species/groups account for 83% of the average similarity within this group (Table
4).
), while the
agglutinated forms are well represented by species of Hyperammina /
Rhabdammina
and Reophax. Less abundant
benthic foraminifera include Ammosphaeroidina
pseudopauciloculata, Bolivina
beyrichi carinata, Bulimina alsatica, B. arndti, B.
elongata, Cibicidoides pachyderma, C.
pygmeus, Fursenkoina spp., Hansenisca
soldanii, Lenticulina spp.,
Nothia spp., Saccamina grzybowski, Stilostomella
adolphina, Uvigerina mantaensis,
and Valvulineria palmarealensis. Twenty-two
species/groups account for 83% of the average similarity within this group (Table
4).
|
Figure
9:
All scales represent 100 μm. |
Subcluster 3B
includes samples G4 and G8 -
G12 (Figs. 2 ![]() and
5
and
5 ![]() ).
As with subcluster 3A, benthic
foraminiferal assemblages are characterized by both calcareous and agglutinated taxa (Fig. 2
).
As with subcluster 3A, benthic
foraminiferal assemblages are characterized by both calcareous and agglutinated taxa (Fig. 2 ![]() ). The Fisher's alpha diversity indices range between 20.3 and
38, and the
Shannon-Wiener
diversity indices between 2.9 and 3.6 (Fig. 2
). The Fisher's alpha diversity indices range between 20.3 and
38, and the
Shannon-Wiener
diversity indices between 2.9 and 3.6 (Fig. 2 ![]() ). Epifaunal and infaunal forms
are well represented, while the epifaunal-shallow
infaunal microhabitat is uncommon (Fig. 2
). Epifaunal and infaunal forms
are well represented, while the epifaunal-shallow
infaunal microhabitat is uncommon (Fig. 2 ![]() ).
).
The dominant benthic calcareous species are Praeglobobulimina pupoides and Uvigerina popescui, while the agglutinated forms are well represented by species of Hyperammina / Rhabdammina. Less common are Ammobaculites agglutinans, Amphicoryna armata, Bathysiphon taurinensis, Bulimina elongata, Cibicidoides pachyderma, C. pygmeus, Hansenisca soldanii, Haplophragmoides spp., Laevidentalina elegans, Lenticulina spp., Martinottiella communis, Melonis pompilioides, Karreriella chilostoma, Nothia spp., Praeglobobulimina ovata, Recurvoides spp., Reophax spp., Reticulophragmium spp., Saccamina grzybowski, Subreophax spp., Trochammina spp., Uvigerina farinosa, and Valvulineria palmarealensis. Thirty-one species/groups account for 84% of the average similarity within this group (Table 4).
In
Cluster 3, the tubular agglutinated
morphogroup is better represented than Cluster 1 (Figs. 6 ![]() and 7
and 7 ![]() ).
Furthermore, the bathyal- and abyssal-type agglutinated foraminifera
(morphogroup M2a) are more abundant in Cluster 3 than in other clusters (except sample G8 - Fig. 7
).
Furthermore, the bathyal- and abyssal-type agglutinated foraminifera
(morphogroup M2a) are more abundant in Cluster 3 than in other clusters (except sample G8 - Fig. 7 ![]() ). The
shallow-water agglutinated forms (such as Spirorutilus
carinatus, Spiroplectammina pectinata, and Vulvulina haeringensis)
and morphogroup M4a are less abundant than in Cluster 1 (Fig. 7
). The
shallow-water agglutinated forms (such as Spirorutilus
carinatus, Spiroplectammina pectinata, and Vulvulina haeringensis)
and morphogroup M4a are less abundant than in Cluster 1 (Fig. 7 ![]() ).
Morphogroups M3a and M2b are also less commonly represented.
).
Morphogroups M3a and M2b are also less commonly represented.
5.1. Biostratigraphy
Most
of the identified planktonic species from the Gălp�ia section have long
stratigraphic ranges. Possibly reworked individuals of Paragloborotalia
opima were also observed (samples G1 - G2, G6, G8, G11 - G12, and G13).
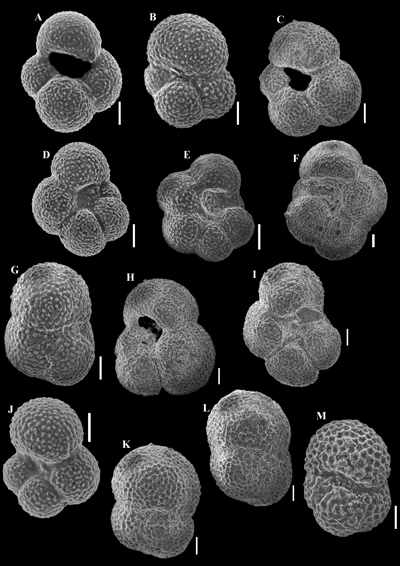
Nevertheless, the presence of the planktonic Globigerinoides
trilobus (in samples G1, G2, G7, G8,
G10, G12, and G14 - Figs. 3 ![]() and 10
and 10 ![]() )
suggests equivalence with the early Miocene (Aquitanian-Burdigalian)
Globigerinoides trilobus Biozone of Popescu
(1975) represented by the stratigraphic interval from the first
appearance of the nominate taxon to the first occurrence of Praeorbulina
glomerosa (Blow, 1956) or the mass occurrence of Globoquadrina
dehiscens (Chapman et al., 1934). The
latter two species of planktonic foraminifera were not identified in the
samples from the Gălp�ia section. More recent
papers (e.g., Wade et al.,
2011) place the Globigerinoides trilobus Zone
(more precisely the Globigerinoides
trilobus Partial-range Zone) in Zone M12 (partial M13a zone of Berggren
et al., 1995). According to Cicha et al.
(1998), the stratigraphic range of the species Cassigerinella
globulosa (Egger, 1857) and Globigerina
ottnangiensis R�gl, 1969, is Eggenburgian to
Karpatian, while Paragloborotalia
semivera (Hornibrook, 1961) is of Egerian to Eggenburgian
age. Based on the identified planktonic foraminiferal species and their
distribution, the age of the deposits from the Gălp�ia
section is probably Eggenburgian (Burdigalian)(R�gl, 1998).
The well-represented agglutinated Reticulophragmium
venezuelanum (Maync, 1952) permits correlation with the Alveolophragmium
venezuelanum Zone of Popescu (1975). Another of Popescu's
(1975) benthic
foraminiferal zones is the Uvigerina
beccarii / Uvigerina galloway
Zone, defined by the occurrence of the two nominate taxa. The species Uvigerina beccarii Fornasini, 1898,
recorded by Popescu
(1975) from the Chechiș Clays, was later described as Uvigerina
popescui R�gl (1998). This calcareous benthic species, initially
considered of Eggenburgian age, is also present in the Oligocene of the Vima
Formation from the Transylvanian Basin (Sz�kely & S. Filipescu,
2015, 2016).
)
suggests equivalence with the early Miocene (Aquitanian-Burdigalian)
Globigerinoides trilobus Biozone of Popescu
(1975) represented by the stratigraphic interval from the first
appearance of the nominate taxon to the first occurrence of Praeorbulina
glomerosa (Blow, 1956) or the mass occurrence of Globoquadrina
dehiscens (Chapman et al., 1934). The
latter two species of planktonic foraminifera were not identified in the
samples from the Gălp�ia section. More recent
papers (e.g., Wade et al.,
2011) place the Globigerinoides trilobus Zone
(more precisely the Globigerinoides
trilobus Partial-range Zone) in Zone M12 (partial M13a zone of Berggren
et al., 1995). According to Cicha et al.
(1998), the stratigraphic range of the species Cassigerinella
globulosa (Egger, 1857) and Globigerina
ottnangiensis R�gl, 1969, is Eggenburgian to
Karpatian, while Paragloborotalia
semivera (Hornibrook, 1961) is of Egerian to Eggenburgian
age. Based on the identified planktonic foraminiferal species and their
distribution, the age of the deposits from the Gălp�ia
section is probably Eggenburgian (Burdigalian)(R�gl, 1998).
The well-represented agglutinated Reticulophragmium
venezuelanum (Maync, 1952) permits correlation with the Alveolophragmium
venezuelanum Zone of Popescu (1975). Another of Popescu's
(1975) benthic
foraminiferal zones is the Uvigerina
beccarii / Uvigerina galloway
Zone, defined by the occurrence of the two nominate taxa. The species Uvigerina beccarii Fornasini, 1898,
recorded by Popescu
(1975) from the Chechiș Clays, was later described as Uvigerina
popescui R�gl (1998). This calcareous benthic species, initially
considered of Eggenburgian age, is also present in the Oligocene of the Vima
Formation from the Transylvanian Basin (Sz�kely & S. Filipescu,
2015, 2016).
|
Figure
10:
All scales represent 100 μm. |
5.2. Paleoecology of planktonic foraminifera
The
planktonic foraminifera from Gălp�ia
are dominated throughout the section by cool-water and high-productivity
indicators (Clusters 2 and 3, Tables 2 and 3). Nevertheless, Cluster 1 contains a higher abundance of the species Globoturborotalita
connecta, which indicates cool-temperate surface waters. At the same
stratigraphic levels, the small five-chambered species Globigerina
ottnangiensis shows a reduced distribution compared to Clusters 2 and 3. Furthermore, episodes of warmer surface waters and
changes in primary productivity could be indicated by the significant increase
in abundance of Globigerinoides trilobus in
samples G7-G8. This trend possibly corresponds with the warming event
documented from the Chechiș Formation (Eggenburgian) by Sz�kely et al.
(2016).
Changes in primary productivity are also reflected by the significant decrease
in abundance of the benthic genus Uvigerina
(Cluster 3A) and a drop in species diversity of the benthic assemblages (Figs. 2 ![]() and 6
and 6 ![]() ).
).
Episodes of high primary productivity are indicated in Clusters 2 and 3 by the small microperforate forms (e.g., Tenuitella), small five-chambered globigerinids (e.g., Globigerina ottnangiensis, G. tarchanensis) and the Globigerina group (see Table 2).
Similar planktonic assemblages with small globigerinids (including Globigerina spp., Tenuitella spp., and Tenuitellinata spp.) and low abundance of Globigerinoides have been reported from the early Miocene (Beldean et al., 2012) and the middle Miocene (S. Filipescu, 2001; S. Filipescu & Silye, 2008; R. & S. Filipescu, 2015) of the Transylvanian Basin. Li & McGowran (1998) and Al-Sabouni et al. (2007) associated these kinds of assemblages with mixing of warm and cold waters and upwelling, respectively.
5.3. Paleoecology of benthic foraminifera
The
presence of the epifaunal
genus Cibicidoides
in Cluster 1 (Fig. 6 ![]() )
suggests
episodes of high-energy well-oxygenated bottom waters (see Table
1). Recent Cibicidoides
pachyderma is characteristic in oligotrophic environments (see Table
1) and
was reported primarily as an upper bathyal
species but is also present in shelf assemblages (Berggren & Haq,
1976).
)
suggests
episodes of high-energy well-oxygenated bottom waters (see Table
1). Recent Cibicidoides
pachyderma is characteristic in oligotrophic environments (see Table
1) and
was reported primarily as an upper bathyal
species but is also present in shelf assemblages (Berggren & Haq,
1976).
Indicators of high
primary productivity are most abundant in this cluster. The
genus Uvigerina
prefers muddy sediments and suggests high organic-matter flux to the sea
floor
and paleodepths greater than 100 m (see Table 1). Similarly, the genus Praeglobobulimina
indicates high primary productivity and poor oxygenation, and it has a preferred depth
range of 80 to 800 m (Spezzaferri & Ćorić,
2001; Spezzaferri et al.,
2002). Both genera are relatively well represented in Cluster 1 (Fig. 6 ![]() ). The less abundant infaunal species Melonis
pompiliodes also supports the interpretation of episodic high primary
productivity (see Table 1). Some authors (e.g., Corliss & Chen,
1988; Gupta & Thomas,
1999; Loubere, 1998; Murray,
2006) associated the genus Pullenia
with high carbon flux rates, variable food flux and low-oxygen
environments. According to R�gl & Spezzaferri
(2003), this
genus is a suboxic indicator present in circum-littoral to bathyal environments.
In particular, the species Pullenia
bulloides has a 200 to 500 m depth range (Spezzaferri et
al., 2004). Episodic decreased bottom-water oxygenation is also suggested by the
presence of suboxic-dysoxic
Lenticulina, the suboxic-dysoxic
Laevidentalina and the mud-preferring
epifaunal Hansenisca soldanii
together with low-oxygen-tolerant taxa such as
species of Bolivina and Bulimina (see Table 1, Fig. 6
). The less abundant infaunal species Melonis
pompiliodes also supports the interpretation of episodic high primary
productivity (see Table 1). Some authors (e.g., Corliss & Chen,
1988; Gupta & Thomas,
1999; Loubere, 1998; Murray,
2006) associated the genus Pullenia
with high carbon flux rates, variable food flux and low-oxygen
environments. According to R�gl & Spezzaferri
(2003), this
genus is a suboxic indicator present in circum-littoral to bathyal environments.
In particular, the species Pullenia
bulloides has a 200 to 500 m depth range (Spezzaferri et
al., 2004). Episodic decreased bottom-water oxygenation is also suggested by the
presence of suboxic-dysoxic
Lenticulina, the suboxic-dysoxic
Laevidentalina and the mud-preferring
epifaunal Hansenisca soldanii
together with low-oxygen-tolerant taxa such as
species of Bolivina and Bulimina (see Table 1, Fig. 6 ![]() ). The
genus Lenticulina is a shallower-water taxon (see Table
1), which has
been reported as a significant component of assemblages from
Eggenburgian-Ottnangian outer-neritic deposits of the Alpine Foreland Basin (Grunert
et al., 2012).
). The
genus Lenticulina is a shallower-water taxon (see Table
1), which has
been reported as a significant component of assemblages from
Eggenburgian-Ottnangian outer-neritic deposits of the Alpine Foreland Basin (Grunert
et al., 2012).
Agglutinated foraminiferal morphogroup M1 is characteristic in bathyal and abyssal
environments with low-organic matter flux. Saccamina
grzybowski (important component of morphogroup M2a in this study) is
known as an outer neritic to abyssal agglutinated form (Kaminski et
al., 2005). The relatively good representation of morphogroup M4b in this
cluster supports the interpretation of episodically increased organic-matter flux to the sea
floor. The deep-water forms of
morphogroup M3 (M3a, Fig. 7 ![]() )
are here represented by flattened surficial epifaunal
species of Glomospira. Glomospira charoides, reported from the
early Eocene North Atlantic,
Mediterranean and Alpine-Carpathian
Flysch (Bindiu & S. Filipescu,
2011), is considered to respond
positively to high organic-matter flux (Kaminski et al.,
2005). Nevertheless, this species was reported also from
oligotrophic intervals in the Mediterranean (De Rijk et
al., 2000). The occurrence of G.
charoides in Cluster 1 may be related to high primary productivity and
the consequent organic-matter flux to the sea floor.
)
are here represented by flattened surficial epifaunal
species of Glomospira. Glomospira charoides, reported from the
early Eocene North Atlantic,
Mediterranean and Alpine-Carpathian
Flysch (Bindiu & S. Filipescu,
2011), is considered to respond
positively to high organic-matter flux (Kaminski et al.,
2005). Nevertheless, this species was reported also from
oligotrophic intervals in the Mediterranean (De Rijk et
al., 2000). The occurrence of G.
charoides in Cluster 1 may be related to high primary productivity and
the consequent organic-matter flux to the sea floor.
Benthic foraminiferal assemblages of Cluster 1 suggest episodic high-energy currents,
intervals with well-oxygenated environments (also
supported by the presence of vertical and horizontal burrows - Fig. 2 ![]() ), and
episodes of high primary productivity with possible reduced oxygen levels as
indicated by the frequent occurrence of Praeglobobulimina
species. These interpretations are supported by the diversity indices,
which show high values (Fig. 2
), and
episodes of high primary productivity with possible reduced oxygen levels as
indicated by the frequent occurrence of Praeglobobulimina
species. These interpretations are supported by the diversity indices,
which show high values (Fig. 2 ![]() ). The occasionally oxygenated, stable
environments and organic-matter flux to the sea floor enabled the
diversification of benthic assemblages in an outer shelf (possibly
upper bathyal) setting. Tubular agglutinated forms, the bathyal-abyssal
morphogroup M2a, and the decrease of shelf-type agglutinated foraminifera (Fig. 7
). The occasionally oxygenated, stable
environments and organic-matter flux to the sea floor enabled the
diversification of benthic assemblages in an outer shelf (possibly
upper bathyal) setting. Tubular agglutinated forms, the bathyal-abyssal
morphogroup M2a, and the decrease of shelf-type agglutinated foraminifera (Fig. 7 ![]() ) indicate a gradual increase in water depth for Cluster 1.
) indicate a gradual increase in water depth for Cluster 1.
Benthic foraminiferal assemblages in Cluster 2
are clearly dominated by agglutinated forms (with the tubular morphogroup
M1 most abundant: Figs. 2 ![]() ,
6
,
6 ![]() and 7
and 7 ![]() ).
Of benthic calcareous
foraminifera, Cibicidoides and Heterolepa
are almost completely absent, while primary productivity indicators (Uvigerina)
decrease in abundance in parallel with the increase of the agglutinated tubular
morphogroup, suggesting low organic flux to the sea floor. This may be related
to the deepening of the environment to an upper bathyal setting, where the
development of calcareous taxa was inhibited. The above-mentioned
paleoecological factors are also reflected by the low species diversity indices (Fig. 2
).
Of benthic calcareous
foraminifera, Cibicidoides and Heterolepa
are almost completely absent, while primary productivity indicators (Uvigerina)
decrease in abundance in parallel with the increase of the agglutinated tubular
morphogroup, suggesting low organic flux to the sea floor. This may be related
to the deepening of the environment to an upper bathyal setting, where the
development of calcareous taxa was inhibited. The above-mentioned
paleoecological factors are also reflected by the low species diversity indices (Fig. 2 ![]() ). Some benthic species have probably been the subject of reworking from
shallower environments, e.g., Hanzawaia
boueana usually found on the inner shelf (preferred depth range of 0 -
50 m -
Spezzaferri & Ćorić,
2001) and shelf-type agglutinated foraminifera such as Spiroplectammina and Vulvulina.
). Some benthic species have probably been the subject of reworking from
shallower environments, e.g., Hanzawaia
boueana usually found on the inner shelf (preferred depth range of 0 -
50 m -
Spezzaferri & Ćorić,
2001) and shelf-type agglutinated foraminifera such as Spiroplectammina and Vulvulina.
Similar
to Cluster 1, subcluster 3A contains
Cibicidoides species (e.g., Cibicidoides
pachyderma, C. pseudoungerianus and
the diminutive C. pygmeus) (Fig. 6 ![]() )
suggesting intervals with high-energy well-oxygenated
bottom waters. The presence of Uvigerina
and Praeglobobulimina together with Melonis
pompiliodes and Valvulineria
palmarealensis are evidence for intervals with high primary productivity
and organic carbon flux to the sea floor and probably lowered oxygenation (see
Table 1), although the significant decrease of Uvigerina
species (Fig. 6
)
suggesting intervals with high-energy well-oxygenated
bottom waters. The presence of Uvigerina
and Praeglobobulimina together with Melonis
pompiliodes and Valvulineria
palmarealensis are evidence for intervals with high primary productivity
and organic carbon flux to the sea floor and probably lowered oxygenation (see
Table 1), although the significant decrease of Uvigerina
species (Fig. 6 ![]() ) and the increase of the tubular agglutinated morphogroup
may suggest shorter episodes. The lower species diversity values (compared
to subcluster 3B) could be the result of the aforementioned conditions.
Furthermore, the presence of the infaunal species Globocassidulina
subglobosa suggests a flux of phytodetritus to the sea floor (Corliss
& Chen, 1988; Gooday,
1994). The highest abundance in the Gălp�ia
section of the low-oxygen
taxa (Bolivina, Bulimina and Praeglobobulimina)
and the sole appearance of the opportunist genus Fursenkoina could be related to abrupt changes in water depth,
decreased bottom water oxygenation as a consequence of high primary
productivity, and a lack of currents to oxygenate the bottom waters.
Alternatively, the above-mentioned genera appeared during more oxygenated
intervals in an environment where the food source was represented by
refractory organic matter. An increase in water depth is also supported by the
significant increase of the tubular agglutinated morphogroup. The species
composition of benthic foraminiferal assemblages from samples included in subcluster
3B is similar to those of subcluster 3A. Nevertheless, the
abundances of low-oxygen
taxa (representatives of Bolivina and
Bulimina) strongly decreases
towards the top of subcluster 3B.
Samples G9 and G12 are characterized by the high abundance of the oxic (such as
Cibicidoides) and primary
productivity (Uvigerina
and Praeglobubulimina) indicators (Fig. 6
) and the increase of the tubular agglutinated morphogroup
may suggest shorter episodes. The lower species diversity values (compared
to subcluster 3B) could be the result of the aforementioned conditions.
Furthermore, the presence of the infaunal species Globocassidulina
subglobosa suggests a flux of phytodetritus to the sea floor (Corliss
& Chen, 1988; Gooday,
1994). The highest abundance in the Gălp�ia
section of the low-oxygen
taxa (Bolivina, Bulimina and Praeglobobulimina)
and the sole appearance of the opportunist genus Fursenkoina could be related to abrupt changes in water depth,
decreased bottom water oxygenation as a consequence of high primary
productivity, and a lack of currents to oxygenate the bottom waters.
Alternatively, the above-mentioned genera appeared during more oxygenated
intervals in an environment where the food source was represented by
refractory organic matter. An increase in water depth is also supported by the
significant increase of the tubular agglutinated morphogroup. The species
composition of benthic foraminiferal assemblages from samples included in subcluster
3B is similar to those of subcluster 3A. Nevertheless, the
abundances of low-oxygen
taxa (representatives of Bolivina and
Bulimina) strongly decreases
towards the top of subcluster 3B.
Samples G9 and G12 are characterized by the high abundance of the oxic (such as
Cibicidoides) and primary
productivity (Uvigerina
and Praeglobubulimina) indicators (Fig. 6 ![]() ).
High-energy currents and episodes of high primary productivity might have
characterized this interval. Simultaneously, agglutinated infaunal
morphogroup M4b is very abundant in samples G9 and G12, supporting high
primary productivity for these levels (Fig. 7
).
High-energy currents and episodes of high primary productivity might have
characterized this interval. Simultaneously, agglutinated infaunal
morphogroup M4b is very abundant in samples G9 and G12, supporting high
primary productivity for these levels (Fig. 7 ![]() ). The existence of such an
environment may also be reflected in the decrease of tubular agglutinated
morphogroup M1 (Fig. 7
). The existence of such an
environment may also be reflected in the decrease of tubular agglutinated
morphogroup M1 (Fig. 7 ![]() ), which suggests low organic-matter flux to the sea
floor (Kaminski et al., 2005).
These intervals may indicate water-depth change (shallowing).
), which suggests low organic-matter flux to the sea
floor (Kaminski et al., 2005).
These intervals may indicate water-depth change (shallowing).
The epifaunal to shallow infaunal (Murray, 2006) Cribrostomoides subglobosus from the middle and upper part of the outcrop is included in morphogroup M2b (Kaminski et al., 2005). Its depth range is considered to be from 400 to 5700 m (Spezzaferri et al., 2004). Another species present in the second part of the outcrop is the outer neritic - abyssal Psammosphaera fusca (Kaminski et al., 2005). The highest abundance of the agglutinated species Karreriella chilostoma (morphogroup M4a) appears in this cluster. According to Van Simaeys et al. (2004), this species is an outer shelf to bathyal indicator.
Subcluster 3B probably represents the transition from outer shelf to upper bathyal settings. Several episodes with high-energy well-oxygenated bottom waters and high primary productivity may mark shallowing events alternating with lower organic flux to the sea floor in deeper settings. The environmental transition is also suggested by the complete absence of the shallow-water agglutinated foraminifera (morphogroup M2c represented by Vulvulina haeringensis, Spiroplectammina carinata and Spirorutilus carinatus) from sample G10 and the general increase of the bathyal - abyssal morphogroup M2a starting with sample G8.
5.4. Succession of paleoenvironments of the Gălp�ia section and the evolution of the northwestern Transylvanian Basin
Based on the data presented above, the rocks at the base of the sequence were probably deposited in an outer shelf (possibly upper bathyal) setting characterized by episodes of higher energy (indicated bythe presence of the genus Cibicidoides), well-oxygenated bottom waters (suggested by Cibicidoides) and intervals with enhanced primary productivity (the strong representation of Uvigerina and Praeglobobulimina species). Planktonic foraminiferal species support the existence of episodes of high primary productivity probably as a consequence of high nutrient flux from the land.
In the middle of the section, the decreasing abundance of Uvigerina and increased tubular agglutinated foraminifera, typical for environments with low organic carbon flux, indicate oscillations in primary productivity. In these environments, the epifaunal benthic forms probably consumed the labile organic matter, while the deep infaunal foraminifera fed on refractory organic matter (more Bolivina and Reophax species) (Jorissen et al., 1995).
The upper part of the sequence is characterized by oscillations in paleoecological factors (such as primary productivity). Towards the top of the outcrop the increase of agglutinated forms indicating bathyal-abyssal environments and the decrease in abundance of calcareous benthic forms suggest the transition to an upper bathyal environment. Besides a low organic-matter flux to the sea floor, transport of some foraminiferal species from shallower environments characterizes the upper bathyal environments of the uppermost part of the section.
The origin of the fine sand laminae may be related to storm activity, longshore drifts or deltaic influences (supported by the presence of coally material). A deltaic influence would support a high nutrient flux to the marine environment (and seasonal high primary productivity), periodic higher current energy and consequent oxygenation of the bottom water. This influence might have diminished during deposition of the sediments in the middle of the sequence. According to Kr�zsek & Bally (2006), the study area (central part of the Transylvanian Basin) occupied a shelf/delta environment during the early Miocene.
In the Transylvanian Basin, the underlying Coruș Formation is considered to be the first
unit deposited during the
early Miocene marine transgression (Popescu et al.,
1995). A glauconite facies
at the base of the Chechiș
Formation was described by Șuraru (1967) and associated in the
neighbouring area (Tihău section) with the maximum flooding surface of the
first early Miocene transgression (Sz�kely et al.,
2016). The deposits above
this glauconite level are considered to have formed during the highstand. The
succession studied in the Gălp�ia section probably represents the middle
and upper parts of the Chechiș Formation and based on the inferred
paleoenvironmental evolution also represents a highstand systems
tract. The sediments belonging to the Chechiș Formation near the Tihău
locality
(Fig. 1 ![]() )
were deposited on a narrow shelf with deltaic influences (Sz�kely et al.,
2016)
while, at Gălp�ia, the shelf was probably characterized by a different
morphology and decreased deltaic influence.
)
were deposited on a narrow shelf with deltaic influences (Sz�kely et al.,
2016)
while, at Gălp�ia, the shelf was probably characterized by a different
morphology and decreased deltaic influence.
(1) Planktonic and benthic foraminiferal assemblages were analyzed from outcrops of the lower Miocene Chechiș Formation in the Gălp�ia section. Biostratigraphic study of the planktonic assemblages allowed the determination of an Eggenburgian/Burdigalian age (Globigerinoides trilobus Biozone of Popescu, 1975) for these deposits.
(2) Planktonic foraminiferal assemblages are mainly dominated by globigerinids and tenuitellids indicating cool surface waters and enhanced primary productivity, while warm-water indicators are rather scarce. Increase in abundance of the species Globigerinoides trilobus in the middle part of the outcrop may suggest a warmer/transgressive interval or changes in primary productivity.
(3) Paleoenvironmental reconstruction based on planktonic and benthic foraminiferal assemblages indicates a general deepening of the environment from outer shelf to upper bathyal. Deltaic influences are suggested by the occurrence of benthic foraminifera originating in shallower environments, with strong bottom currents, high oxygen levels and high primary productivity. Increase in water depth resulted in the retrogradation of the environments, with consequences for primary productivity and a decrease of organic-matter flux to the sea floor. In the second part of the section, a transition was observed from outer shelf to upper bathyal. A mainly low organic flux to the sea floor characterized the upper bathyal setting.
(4) The sedimentary succession studied was probably deposited during the late stage of the relative sea-level rise (highstand systems tract) following the initial early Miocene transgression in the Transylvanian Basin. The species composition and distribution of the foraminiferal assemblages reflect changing paleoecological factors as a consequence of the above-mentioned aspects.
We are thankful to the reviewers, P. Grunert and D. Peryt, for their constructive comments and suggestions on the manuscript. We are grateful to Claudia Beldean for her support during the fieldwork. The research was made possible by the financial support of the Sectorial Operational Programme for Human Resources Development 2007-2013, co-financed by the European Social Fund, under the project POSDRU/159/1.5/S/133391 - "Doctoral and postdoctoral excellence programs for training highly qualified human resources for research in the fields of Life Sciences, Environment and Earth". The Sciex 12.116 Grant, the Grzybowski Foundation, the Brian J. O'Neill Memorial Grant-in-Aid and the S.N.G.N. Romgaz (Contract 18/2011) are also warmly thanked for financial support.
Almogi-Labin A., Schmiedl G., Hemleben C., Siman-Tov R., Segl M. & Meischner D. (2000).- The influence of the NE winter monsoon on productivity changes in the Gulf of Aden, NW Arabian Sea, during the last 530 ka as recorded by foraminifera.- Marine Micropaleontology, vol. 40, p. 295-319.
Al-Sabouni N., Kucera M. & Schmidt D.N. (2007).- Vertical niche separation, control of diversity and size disparity in planktonic foraminifera.- Marine Micropaleontology, vol. 63, no. 1-2, p. 75-90.
Amore F.O., Caffaub M., Massaa B. & Morabito S. (2004).- Late Pleistocene-Holocene paleoclimate and related paleoenvironmental changes as recorded by calcareous nannofossils and planktonic foraminifera assemblages in the southern Tyrrhenian Sea (Cape Palinuro, Italy).- Marine Micropaleontology, vol. 52, p. 255-276.
B�ldi T. (1969).- On the Oligocene and Miocene stages of the Central Paratethys and on the formations of the Egerian in Hungary.- Annales Universitatis Scientarium Budapestensis, (Sectio Geologica), vol. 12, p. 19-28.
B�ldi K. (2006).- Paleoceanography and climate of the Badenian (Middle Miocene, 16.4-13.0 Ma) in the Central Paratethys based on foraminifera and stable isotope (δ18O and δ13C) evidence.- International Journal of Earth Sciences (Geologische Rundschau), vol. 95, p. 119-142.
Beldean C. & Filipescu S. (2011).- "Flysch-type" agglutinated foraminifera from the Lower Miocene of the Transylvanian Basin (Romania). In: Kaminski M.A. & Filipescu S. (eds.), Proceedings of the Eighth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation, Special Publication, no. 16, p. 1-18.
Beldean C., Filipescu S. & Bălc R. (2012).- Paleoenvironmental and biostratigraphic data for the Early Miocene of the north-western Transylvanian Basin based on planktonic foraminifera.- Carpathian Journal of Earth and Environmental Sciences, Baia Mare, vol. 7, no. 1, p. 171-184.
Berggren W.A. & Haq B.U. (1976).- The Andalusian Stage (late Miocene): Biostratigraphy, biochronology and palaeoecology.- Palæogeography, Palæoclimatology, Palæoecology, vol. 20, p. 67-129.
Berggren W.A., Kent D.V., Swisher III C.C. & Aubry M.P.A (1995).- Revised Cenozoic geochronology and chronostratigraphy. In: Berggren W.A., Kent D.V. & Hardenbol J. (eds.), Geochronology, time scale and global stratigraphic correlations: Unified temporal framework for a historical geology.- Society of Economic Paleontologists and Mineralogists, Special Publication, vol. 54, p. 129-212.
Bicchi E., Ferrero E. & Gonera M. (2003).- Palaeoclimatic interpretation based on Middle Miocene planktonic Foraminifera: The Silesia Basin (Paratethys) and Monferrato (Tethys) records.- Palæogeography, Palæoclimatology, Palæoecology, vol. 196, p. 265-303.
Bicchi E., Dela Pierre F., Ferrero E., Maia F., Negri A., Pirini Radrizzani C., Radrizzani S. & Valleri G. (2006).- Evolution of the Miocene Carbonate Shelf of Monferrato (North-western Italy).- Bollettino della Societa Paleontologica Italiana, vol. 45, no. 2-3, p. 171-194.
Bindiu R. & Filipescu S. (2011).- Agglutinated Foraminifera from the Northern Tarcău Nappe (Eastern Carpathians, Romania).- Studia Universitatis Babeș-Bolyai, (Geologia), Cluj-Napoca, vol. 56, no. 2, p. 31-41.
Blow W.H. (1956).- Origin and evolution pf the foraminiferal genus Orbulina d'Orbigny.- Micropaleontology, vol. 2, no. 1, p. 57-70.
Chapman F., Parr W.J. & Collins A.C. (1934).- Tertiary foraminifers of Victoria, Australia: The Balcombian deposits of Port Philip (Pt. III).- Journal of the Linnenan Society of London, vol. 38, no. 262, 569 p.
Cicha I., R�gl F., Rupp C. & Ctyroka J. (1998).- Oligocene - Miocene foraminifera of the Central Paratethys.- Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main, Band 549, 325 p.
Clarke K.R. (1993).- Non-parametric multivariate analyses of changes in community structure.- Austral Ecology, Windsor, vol. 18, p. 117-143.
Clarke K.R. & Warwick R.M. (2001).- Change in marine communities: An approach to statistical analysis and interpretation.- Second Edition, PRIMER-E, Plymouth, 172 p.
Clifford D.H.T. & Stephenson W. (1975).- An introduction to numerical classification.- Academic Press, New York, 241 p.
Corliss B.H. (1985).- Microhabitats of benthic foraminifera within deep-sea sediments.- Nature, vol. 314, p. 435-438.
Corliss B.H. (1991).- Morphology and microhabitat preferences of benthic foraminifera from the northwest Atlantic Ocean.- Marine Micropaleontology, vol. 17, p. 195-236.
Corliss B.H. & Chen C. (1988).- Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications.- Geology, vol. 16, p. 716-719.
De Man E. (2006).- Benthic foraminifera biofacies analysis and stable isotopes of the Middle Eocene to Oligocene successions in the southern North Sea Basin. Tools for stratigraphy and for reconstruction of extreme climate changes.- Unpublished Ph.D. Thesis, University of Leuven, Belgium, 375 p.
De Rijk S., Jorissen F.J., Rohling E.J. & Troelstra S.R. (2000).- Organic flux control on bathymetric zonation of Mediterranean benthic foraminifera.- Marine Micropaleontology, vol. 40, p. 151-166.
Dicea O., Duțescu P., Antonescu F., Mitrea G., Botez R., Donos I., Lungu V. & Moroșanu I. (1980).- Contribution to the knowledge of Maramures's Transcarpathian Zone stratigraphy.- Dări de Seamă Institutul de Geologie și Geofizică, Bucuresti, vol. 65, no. 4, p. 21-85 (in Romanian).
Egger J.G. (1857).- Die Foraminiferen der Mioc�n-Schichten bei Ortenburg in Nieder-Bazern.- Neues Jahrbuch f�r Mineralogie, Geognosie, Geologie, und Petrefakten-Kunde, p. 266-311.
Field J.G., Clarke K.R. & Warwick R.M. (1982).- A practical strategy for analysing multispecies distribution patterns.- Marine Ecology Progress Series, vol. 8, p. 37-52.
Filipescu R. & Filipescu S. (2015).- New data on the Early - Middle Badenian transition in the NW Transylvanian Basin (Romania) revealed by the planktonic foraminifera assemblages.- Studia UBB Geologia, vol. 59, no. 1-2, p. 39-44.
Filipescu S. (2001).- Wielician foraminifera at the western border of the Transylvanian Basin.- Studia Universitatis Babeș-Bolyai, Geologia, vol. 46, no. 2, p. 115-123.
Filipescu S. (2011).- Cenozoic lithostratigraphic units in Transylvania. In: Bucur I.I. & Săsăran E. (eds.), Calcareous algae from Romanian Carpathians.- Field Trip Guide Book, Tenth International Symposium on Fossil Algae, Cluj-Napoca, Romania, 12-18 September, Cluj University Press, p. 37-48.
Filipescu S. & Silye L. (2008).- New biozones of planktonic foraminifera in the Middle Miocene of the Transylvanian Basin as potential opportunities for the Paratethyan stratigraphy.- Geologica Carpathica, vol. 59, no. 6, p. 537-544.
Fornasini C. (1898).- Contributo alla conoscenza della microfauna Terziaria Italiana: Foraminiferi del Pliocene superiore de San Pietro in Lama presso Lecce.- Memorie della Reale Academia delle Scienze dell'Istituto di Bologna vol. (5) 7, p. 205-212
Gooday A.J. (1994).- The biology of deep-sea foraminifera: A review of some advances and their applications in paleoceanography.- Palaios, vol. 9, p. 14- 31.
Grunert P., Soliman A., Ćorić S., Roetzel R., Harzhauser M. & Piller W.E. (2012).- Facies development along the tide-influenced shelf of the Burdigalian Seaway: An example from the Ottnangian stratotype (Early Miocene, middle Burdigalian).- Marine Micropaleontology, vol. 84-85, p. 14-36.
Gupta A.K. & Thomas E. (1999).- Latest Miocene through Pleistocene paleoceanographic evolution of the northwestern Indian Ocean (DSDP Site 219): Global and regional factors.- Paleoceanography, vol. 14, p 62-73.
Hammer �. & Harper D.A.T. (2006).- Paleontological data analysis.- Blackwell Publishing, Oxford, 351 p.
Hammer �., Harper D.A.T. & Ryan P.D. (2001).- PAST: PAleontological STatistics software package for education and data analysis.- Palaeontologia Electronica, vol. 4, no. 1, p. 1-9. URL: http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hofmann K. (1879).- Bericht �ber die im �stlichen Teile des Szil�gyer Comitates wahrend der Sommercampagne 1878 vollf�hrten geologischen Specialaufnahmen.- F�ldtani K�zl�ny, Budapest, vol. 9, no. 5-6, p. 231-283.
Hornibrook N. (1961).- Tertiary foraminifers from Oamaru District (N.Z.), Part 1. Systematics and distribution.- New Zealand Geological Survey Paleontological Bulletin, Wellington, vol. 34, 192 p.
Jones R.W. & Charnock M.A. (1985).- "Morphogroups" of agglutinated foraminifera. Their life positions and feeding habits and potential applicability in (paleo)ecological studies.- Revue de Pal�obiologie, vol. 4, p. 311-320.
Jorissen F.J. (2002).- Benthic foraminiferal microhabitats below the sediment-water interface. In: Sen Gupta B.K. (ed.), Modern Foraminifera, 2nd ed.- Kluwer Academic Publishers, Boston, p. 161-179.
Jorissen F.J., De Stigter H.C. & Widmark J.G.V. (1995).- A conceptual model explaining benthic foraminiferal microhabitats.- Marine Micropaleontology, vol. 22, p. 3-15.
Kaiho K. (1994).- Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean.- Geology, vol. 22, p. 719-722.
Kaminski M.A. & Gradstein F.M. (eds., 2005).- Atlas of Paleogene cosmopolitan deep-water agglutinated Foraminifera.- Grzybowski Foundation, Special Publication, no. 10, 546 + vii p.
Kr�zsek Cs. & Bally A.W. (2006).- The Transylvanian Basin (Romania) and its relation to the Carpathian fold and thrust belt: Insights in gravitational salt tectonics.- Marine and Petroleum Geology, vol. 23, p. 405-442.
Kroon D. (1988).- The planktic ∂13C record, upwelling and climate. In: Brummer G.J.A. & Kroon D. (eds.), Planktonic foraminifers as tracers of ocean-climate history.- Amsterdam (VU Uitgeverij), p. 335-346.
Laskarev V.N. (1924).- Sur les �quivalents du Sarmatien sup�rieur en Serbie. In: Vujević P. (ed.), Receuil de travaux offerts � M. Jovan Cvijić par ses amis et collaborateurs.- Dr�avna �tamparija, Beograd, p. 73-85.
Li Q., Radford S.S. & Banner F.T. (1992).- Distribution of microperforate tenuitellid planktonic Foraminifers in holes 747A and 749B, Kerguelen Plateau. In: Schlich R. & Wise S.W. Jr (eds.), Proceedings of the Ocean Drilling Program.- Scientific Results, College Station, vol. 120, p. 569-594.
Li Q. & McGowran B. (1998).- Oceanographic implications of recent planktonic foraminifera along the southern Australian margin.- Marine and Freshwater Research, vol. 49, p. 439-445.
Loubere P. (1998).- The impact of seasonality on the benthos as reflected in the assemblages of deep-sea foraminifera.- Deep-Sea Research I, vol. 45, p. 409-432.
Maync W. (1952).- Critical taxonomic study and nomenclatural revision of the Lituolidae based on the prototype of the family, Lituola nautiloidea Lamarck 1804.- Contributions from the Cushman Foundation for Foraminiferal Research, vol. 3, no. 2, p. 35-53
Miao Q. & Thunell R.C. (1993).- Recent deep-sea benthic foraminiferal distributions in the South China and Sulu Seas.- Marine Micropaleontology, vol. 22, p. 1-32.
Morkhoven F.P.C.M. van, Berggren W.A. & Edwards A.S. (1986).- Cenozoic cosmopolitan deep-water benthic foraminifera.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, M�moire 11, 423 p.
Mullins H.T., Thompson J.B., McDougall K. & Vercoutere T.L. (1985).- Oxygen minimum zone edge effect: Evidence from the central California coastal upwelling system.- Geology, vol. 13, p. 491-494.
Murray J.W. (2006).- Ecology and Applications of Benthic Foraminifera.- Cambridge University Press, Cambridge, 438 p.
Murray J.W., Alve E. & Jones B.W. (2011).- A new look at modern agglutinated benthic foraminiferal morphogroups: Their value in palaeoecological interpretation.- Palæogeography, Palæoclimatology, Palæoecology, vol. 309, p. 229-241.
Nicorici E., Petrescu I. & M�sz�ros N. (1979).- Contribuţii la cunoaşterea miocenului inferior şi mediu de la Coasta cea Mare (Cluj-Napoca).- Studii şi cercetări de geologie, geofizică, geografie, (Geologie), Bucuresti, vol. 24, p. 103-137 (in Romanian).
Pearson P.N., Olsson R.K., Huber B.T., Hemleben C. & Berggren W.A. (2006).- Atlas of Eocene Planktonic Foraminifera.- Cushman Foundation for Foraminiferal Research, Special Publication 41, 514 p.
Petrescu I. & Drăghici D. (1964).- The Geological Map of Romania, 1:200000, Sheet Cluj.
Popescu G. (1970).- Foraminiferele planctonice din stratele de Hida (nord-vestul Transilvaniei).- Studii şi cercetări de geologie, geofizică, geografie, (Geologie), Bucuresti, vol. 15, p. 240-253 (in Romanian).
Popescu G. (1971).- Biostratigrafia depozitelor Oligo-Miocene de la sud de Preluca, pe bază de foraminifere planctonice.- Dări de Seamă ale Şedinţelor Institutului Geologic, Bucuresti, vol. 58, p. 105-127 (in Romanian).
Popescu G. (1975).- �tudes des foraminif�res du Mioc�ne inf�rieur et moyen du nord-ouest de la Transylvanie.- Institut de G�ologie et de G�ophysique, M�moires, Bucuresti, vol. 23, 121 p.
Popescu G. & Iva M. (1971).- Contribution � la connaissance de la microfaune oligoc�ne des Couches de Valea Lăpuşului.- Institut de G�ologie et de G�ophysique, M�moires, Bucuresti, vol. 14, p. 35-52.
Popescu G., Marunteanu M. & Filipescu S. (1995).- Neogene from Transylvania Depression. In: X. Congress RCMNS, Bucureşti 1995, Guide to excursion A1.- Romanian Journal of Stratigraphy, Bucureşti, vol. 76, p. 1-27.
Reolid M. & Nagy J. (2008).- Jurassic Ttransgressive-regressive cycles in carbonate and siliciclastic shelf facies: Different response of foraminiferal assemblage trends to sea-level changes. In: Kaminski M.A. & Coccioni R. (eds.), Proceedings of the Seventh International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation, Special Publication, no. 13, p. 199-213.
R�gl F. (1969).- Die mioz�ne Foraminiferen-Fauna von Laa an der Thaya in der Molassezone von Nieder�sterreich.- Mitteilungen Geologischen Gesellschaft Wien, vol. 61, p. 63-123.
R�gl F. (1998).- Oligocene-Miocene foraminifera of the Central Paratethys- Systematics and taxonomy. In: Cicha I., R�gl F., Rupp Ch. & Ctyroka J. (eds.), Oligocene-Miocene Foraminifera of the Central Paratethys.- Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main, Band 549, p. 69-78.
R�gl F. & Spezzaferri S. (2003).- Foraminiferal paleoecology and biostratigraphy of the Muhlbach section (Gaindorf Formation, Lower Badenian), Lower Austria.- Annalen des Naturhistorischen Museums in Wien, 104 A, p. 23-75.
Roetzel R., Ćorić S., Galović I. & R�gl F. (2006).- Early Miocene (Ottnangian) coastal upwelling conditions along the southern scarp of the Bohemian Massif (Parisdorf, Lower Austria, Central Paratethys).- Beitr�ge zur Pal�ontologie, Wien, vol. 30, p. 387-413.
Rusu A. (1969).- Sur la limite Oligoc�ne - Mioc�ne dans le Bassin de Transylvanie.- Revue Roumaine de G�ologie, G�ophysique, et de G�ographie, (S�rie de G�ologie), Bucuresti, vol. 13, no. 2, p. 203-216.
Rusu A. (1977).- Stratigrafia depozitelor Oligocene din nord-vestul Transilvaniei (Regiunea Treznea-Hida-Poiana Blenchii).-. Ph.D. Thesis, Facultatea de geologie-geografie, Universitatea din București.- Anuarul Institutului de Geologie și Geofizică, Bucuresti, vol. 51, p. 70-224 (in Romanian).
Rusu A. & Popescu G. (1965).- Contribuţii la stratigrafia Miocenului inferior din nord-vestul Bazinului Transilvaniei.- Studii şi cercetări de geologie, geofizică, geografie, (Geologie), Bucuresti, vol. 10, no. 2, p. 467-473 (in Romanian).
Schmiedl G., Bov�e F.D., Buscail R., Charri�re B., Hemleben C., Medernach L. & Picon P. (2000).- Trophic control of benthic foraminiferal abundance and microhabitat in the bathyal Gulf of Lions, western Mediterranean Sea.- Marine Micropaleontology, vol. 40, p. 167-188.
Spezzaferri S. (1994).- Planktonic foraminiferal biostratigraphy and taxonomy of the Oligocene and lower Miocene in the oceanic record. An overview.- Palaeontographia Italica, Pisa, vol. 81, p. 1-187.
Spezzaferri S. (1995).- Planktonic foraminiferal paleoclimatic implications across the Oligocene-Miocene transition in the oceanic record (Atlantic, Indian and South Pacific).- Palæogeography, Palæoclimatology, Palæoecology, vol. 114, p. 43-74.
Spezzaferri S. & Ćorić S. (2001).- Ecology of Karpatian (Early Miocene) foraminifera and calcareous nannoplankton from Laa an der Thaya, Lower Austria: A statistical approach.- Geologica Carpathica, vol. 200, no. 6, p. 361-374.
Spezzaferri S., Ćorić S., Hohenegger J. & R�gl F. (2002).- Basin-scale paleobiogeography and paleoecology: An example from Karpatian (Latest Burdigalian) benthic and planktonic foraminifera and calcareous nannofossils from the Central Paratethys.- Geobios, M�moire sp�cial, vol. 24, p. 241-256.
Spezzaferri S., R�gl F., Ćorić S. & Hohenegger J. (2004).- Paleoenvironmental changes and agglutinated foraminifera across the Karpatian/Badenian (early/middle Miocene) boundary in the Styrian Basin (Austria, Central Paratethys). In: Bub�k M. & Kaminski M.A. (eds.), Proceedings of the sixth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation, Special Publication, no. 8, p. 423-459.
Sz�kely S.-F. & Filipescu S. (2015).- Taxonomic record of the Oligocene benthic foraminifera from the Vima Formation (Transylvanian Basin, Romania).- Acta Palaeontologica Romaniae, vol. 11, no. 1, p. 25-62.
Sz�kely S.-F. & Filipescu S. (2016).- Biostratigraphy and paleoenvironments of the Late Oligocene in the north-western Transylvanian Basin revealed by the foraminifera assemblages.- Palæogeography, Palæoclimatology, Palæoecology, vol. 449, p. 484-509.
Sz�kely S.-F., Beldean C., Bindiu R., Filipescu S. & Săsăran E. (2016).- Palaeoenvironmental changes in the Transylvanian Basin during the Early Miocene revealed by the foraminifera assemblages.- Geological Quarterly, vol. 60, no. 1, p. 165-178.
Şuraru N. (1952).- Contribuţie nouă la cunoaşterea microfaunei Stratelor de Hida.- Studii şi Cercetări Ştiinţifice, vol. 3, p. 122-130 (in Romanian).
Şuraru N. (1967).- Betr�ge zur Kenntnis des Burdigals im nordwestlichen teil des Siebenb�rger Beckens zwischen Cluj und Surduc (Rum�nien).- Neues Jahrbuch f�r Geologie und Pal�ontologie, Monatshefte, Stuttgart, Heft 8, p. 489-497.
Şuraru N. (1968).- Contribuție la cunoașterea macrofaunei argilelor de Chechiș.- Studia Universitatis "Babeș-Bolyai", (seria Geologie-Geografie), Cluj-Napoca, vol. 13, no. 2, p. 47-58 (in Romanian).
Tischler M., Matenco L., Filipescu S., Gr�ger H.R., Wetzel A. & F�genschuh B. (2008).- Tectonics and sedimentation during convergence of the ALCAPA and Tisza-Dacia continental blocks: The Pienide nappe emplacement and its foredeep (N. Romania). In: Siegesmund S., F�genschuh B. & Froitzheim N. (eds.), Tectonic Aspects of the Alpine-Dinaride-Carpathian System.- Geological Society of London, Special Publications, vol. 298, p. 317-334.
Van Simaeys S., De Man E., Vanderberghe N., Brinkhuis H. & Steurbaut E. (2004).- Stratigraphic and palaeoenvironmental analysis of the Rupelian-Chattian transition in the type region: Evidence from dinoflagellate cysts, foraminifera and calcareous nannofossils.- Palæogeography, Palæoclimatology, Palæoecology, vol. 208, p. 31- 58.
Wade B.S., Pearson P.N., Berggren W. A. & P�like H. (2011).- Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale.- Earth-Science Reviews, vol. 104, p. 111-142
Z�gorsek K., Holcov� K. & Třasoň T. (2007).- Bryozoan event from Middle Miocene (Early Badenian) lower neritic sediments from the locality Kralice and Oslavou (Central Paratethys, Moravian part of the Carpathian Foredeep).- International Journal of Earth Sciences (Geologische Rundschau), vol. 97, p. 835-850.
| Foraminiferal
counts from the Gălp�ia section, Romania |
Samples | |||||||||||||
| Gălp�ia species list | G1 | G2 | G3 | G4 | G5 | G6 | G7 | G8 | G9 | G10 | G11 | G12 | G13 | G14 |
| Benthic foraminifera | ||||||||||||||
| Alabamina polita Becker & Dusenbury | 0 | 0 | 0 | 1 | 6 | 2 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ammobaculites agglutinans (Orbigny) | 2 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 |
| Ammodiscus cretaceus (Reuss) | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ammodiscus miocenicus Karrer | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ammodiscus peruvianus Berry | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ammodiscus sp. | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 2 | 1 | 0 | 2 | 0 | 0 | 1 |
| Ammosphaeroidina pseudopauciloculata (Mjatliuk) | 0 | 0 | 2 | 1 | 0 | 1 | 2 | 0 | 0 | 1 | 2 | 1 | 0 | 0 |
| Amphicoryna armata (Neugeboren) | 0 | 3 | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 |
| Amphicoryna hirsuta (Orbigny) | 4 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Amphicoryna sp. | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Astacolus sp. | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Asterigerinoides guerichi (Franke) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bathysiphon taurinensis Sacco | 2 | 3 | 1 | 3 | 1 | 0 | 0 | 1 | 3 | 2 | 3 | 1 | 6 | 9 |
| Bigenerina agglutinans Orbigny | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Bolivina antiqua Orbigny | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bolivina beyrichi carinata Hantken | 0 | 0 | 0 | 1 | 1 | 4 | 1 | 2 | 2 | 0 | 0 | 0 | 1 | 1 |
| Bolivina crenulata Cushman | 0 | 0 | 1 | 3 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Bolivina dilatata dilatata Reuss | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bolivina fastigia Cushman | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bolivina molassica Hofmann | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bolivina sp. | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Budashevaella laevigata (Voloshinova) | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Budashevaella mulitcamerata (Voloshinova) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulimina alsatica Cushman & Parker | 0 | 0 | 0 | 0 | 3 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Bulimina arndti Hagn | 0 | 2 | 0 | 0 | 0 | 1 | 2 | 0 | 2 | 4 | 1 | 0 | 0 | 0 |
| Bulimina elongata Orbigny | 2 | 0 | 1 | 2 | 3 | 8 | 0 | 2 | 1 | 0 | 1 | 2 | 0 | 0 |
| Bulimina schischkinskayae Samoylova | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulimina sp. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Bulimina striata striata Orbigny | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulimina subulata Cushman & Parker | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bulimina tenera Reuss | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Ceratobulimina contraria (Reuss) | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Chilostomella oolina Schwager | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cibicides amphysiliensis (Andreae) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cibicidoides lopjanicus (Mjatliuk) | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cibicidoides pachyderma (Rzehak) | 11 | 4 | 5 | 2 | 0 | 3 | 1 | 2 | 6 | 5 | 6 | 4 | 0 | 0 |
| Cibicidoides pseudoungerianus (Cushman) | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Cibicidoides pygmeus (Hantken) | 0 | 0 | 0 | 1 | 1 | 1 | 3 | 1 | 3 | 1 | 1 | 0 | 0 | 0 |
| Cibicidoides sp. | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cibicidoides ungerianus ungerianus Orbigny | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cribrostomoides subglobosus (Cushman) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 1 |
| Cylindroclavulina rudis (Costa) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Dentalina leguminiformis (Batsch) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Elphidiella sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Epistominella exigua (Brady) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Frondovaginulina tenuissima (Hantken) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Fursenkoina sp. | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gaudryinopsis megagranosus (Venglinskyi) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Globocassidulina oblonga (Reuss) | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globocassidulina subglogbosa (Brady) | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globulina gibba Orbigny | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Glomospira charoides (Jones & Parker) | 1 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Glomospira gordialis (Jones & Parker) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Guttulina communis (Orbigny) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gyroidinaconstans (Reiser) | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Gyroidinoides umbonatus (Silvestri) | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hansenisca soldanii (Orbigny) | 5 | 2 | 2 | 0 | 2 | 5 | 2 | 2 | 3 | 1 | 2 | 4 | 1 | 1 |
| Hanzawaia boueana (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Haplophragmoides carinatus Cushman & Renz | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Haplophragmoides horridus (Grzybowski) | 0 | 1 | 1 | 0 | 0 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Haplophragmoides sp. | 6 | 3 | 1 | 3 | 3 | 0 | 1 | 1 | 4 | 2 | 3 | 5 | 0 | 1 |
| Haplophragmoides suborbicularis (Grzybowski) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Haplophragmoides vasiceki vasiceki Cicha & Zapletalova | 2 | 1 | 4 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Hemirobulina eximia (Neugeboren) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hemirobulina hantkeni (Bandy) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Heterolepa dutemplei (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 2 | 0 | 0 | 1 |
| Hormosinelloides sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Hyperammina / Rhabdammina sp. | 0 | 0 | 0 | 18 | 12 | 16 | 14 | 19 | 17 | 33 | 28 | 24 | 51 | 34 |
| Hyperammina elongata Brady | 0 | 1 | 1 | 1 | 2 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 3 | 1 |
| Hyperammina rugosa Verdenius & Van Hinte | 0 | 1 | 0 | 6 | 3 | 5 | 1 | 3 | 0 | 4 | 2 | 1 | 19 | 15 |
| Hyperammina sp. | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 1 |
| Karreriella bradyi (Cushman) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Karreriella chilostoma (Reuss) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 3 | 5 | 1 | 0 |
| Karreriella victoriensis (Cushman) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Karrerulina apicularis (Cushman) | 6 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Karrerulina conversa (Grzybowski) | 1 | 1 | 2 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Laevidentalina boueana (Orbigny) | 0 | 2 | 5 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Laevidentalina elegans (Orbigny) | 2 | 1 | 1 | 2 | 2 | 0 | 0 | 1 | 1 | 1 | 2 | 0 | 0 | 0 |
| Laevidentalina inornata (Orbigny) | 2 | 1 | 3 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Lagena sp. | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lagenammina sp. | 0 | 0 | 0 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Lapugyina schmidi Popescu | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 |
| Lenticulina arcuatostriata (Hantken) | 5 | 2 | 1 | 1 | 0 | 0 | 0 | 3 | 6 | 4 | 1 | 2 | 0 | 0 |
| Lenticulina calcar (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 1 |
| Lenticulina cf. crassa (Orbigny) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lenticulina depauperata (Reuss) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lenticulina gibba (Orbigny) | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lenticulina inornata (Orbigny) | 2 | 7 | 9 | 3 | 2 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| Lenticulina limbosa (Reuss) | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Lenticulina melvilli (Cushman & Renz) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lenticulina olianensis (Ruiz de Gaona & Colom) | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lenticulina sp. | 0 | 4 | 3 | 0 | 3 | 0 | 0 | 4 | 0 | 0 | 3 | 2 | 0 | 1 |
| Lenticulina subpapillosa (Nuttall) | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Marginulina behmi (Reuss) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Marginulina hirsuta Orbigny | 3 | 4 | 5 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
| Martinottiella communis (Orbigny) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 2 | 1 | 1 | 0 |
| Melonis pompilioides (Fichtel & Moll) | 1 | 6 | 3 | 2 | 4 | 3 | 3 | 6 | 2 | 2 | 0 | 2 | 2 | 1 |
| Miliammina sp. | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Mylostomella advena (Cushman & Laiming) | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mylostomella recta (Palmer & Bermudez) | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Neolenticulina peregrina (Schwager) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Neugeborina longiscata (Orbigny) | 0 | 0 | 1 | 0 | 3 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 |
| Nodosaria rudis Orbigny | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Nothia excelsa (Grzybowski) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 4 | 0 | 4 | 2 |
| Nothia sp. | 1 | 2 | 6 | 3 | 1 | 1 | 2 | 2 | 3 | 2 | 2 | 0 | 6 | 9 |
| Oridorsalis umbonatus (Reuss) | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 5 | 0 | 0 |
| Pararotalia sp. | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Percultazonaria fragaria (G�mbel) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Placentammina placenta (Grzybowski) | 1 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 | 2 | 0 |
| Planularia kubinyi (Hantken) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Popovia johnrolandi (Preece et al.) | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Popovia sp. | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| Praeglobobulimina ovata (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 6 | 2 | 1 | 1 |
| Praeglobobulimina pupoides (Orbigny) | 11 | 11 | 9 | 12 | 2 | 12 | 9 | 11 | 12 | 5 | 13 | 8 | 6 | 8 |
| Praeglobobulimina pyrula (Orbigny) | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Protobellina vermiculata Luczkowska | 0 | 2 | 0 | 2 | 0 | 1 | 0 | 0 | 1 | 2 | 1 | 0 | 0 | 0 |
| Psamosiphonella cylindrica (Glaessner) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Psamosphaera fusca Schultze | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 2 | 1 |
| Pseudonodosaria inflata (Bornemann) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Pullenia bulloides (Orbigny) | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pullenia salysburii (Stewart & Stewart) | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pyramidulina catesbyi (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Pyramidulina latejugata (G�mbel) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 0 |
| Recurvoides renzi (Asano) | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Recurvoides sp. | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 0 |
| Reophax excentricus Cushman | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Reophax pilulifer Brady | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| Reophax scorpiurus Montfort | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
| Reophax sp. | 4 | 1 | 1 | 3 | 5 | 4 | 3 | 3 | 5 | 5 | 6 | 4 | 2 | 4 |
| Reophax subfusiformis Earland | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Reticulophragmium acutidorsatum (Hantken) | 4 | 2 | 3 | 1 | 0 | 1 | 0 | 1 | 0 | 4 | 0 | 2 | 3 | 2 |
| Reticulophragmium rotundidorsatum (Hantken) | 6 | 3 | 5 | 2 | 0 | 0 | 0 | 0 | 1 | 7 | 3 | 1 | 2 | 1 |
| Reticulophragmium venezuelanum (Maync) | 10 | 3 | 3 | 2 | 0 | 0 | 0 | 0 | 1 | 7 | 1 | 2 | 0 | 0 |
| Rhabdammina sp. | 2 | 3 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 4 | 6 |
| Rhizzamina algaeformis Brady | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
| Sabellovoluta humboldti (Reuss) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Saccamina grzybowski (Schubert) | 1 | 1 | 3 | 3 | 3 | 2 | 1 | 0 | 3 | 5 | 5 | 1 | 8 | 4 |
| Saccamina sphaerica Brady | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Saracenaria sp. | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 |
| Scallopostoma ovicula (Orbigny) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Semivulvulina pectinata (Reuss) | 0 | 1 | 1 | 1 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sigmoilinita tenuis (Czjzek) | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sigmoilopsis schlumbergeri (Silvestri) | 3 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 1 | 0 | 0 |
| Sigmoilopsis sp. | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sigmoilopsis triangularis (Popescu) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siphogenerinoides vasarhelyi (Hantken) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Siphonina reticulata (Czjzek) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 |
| Siphonodosaria consobrina (Orbigny) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spirorutilus carinatus (Orbigny) | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Stilostomella adolphina (Orbigny) | 1 | 1 | 2 | 1 | 2 | 2 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Stilostomella sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Subreophax sp. | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 3 | 1 | 5 |
| Textularia gramen Orbigny | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Textularia nussdorfensis Orbigny | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 1 | 0 |
| Textularia pala Czjzek | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Textularia sp. | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trochammina globigeriniformis (Parker & Jones) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Trochammina kibleri Venglinskyi | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trochammina sp. | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 2 | 1 | 1 | 1 | 1 |
| Uvigerina acuminata Hosius | 0 | 3 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Uvigerina farinosa Hantken | 6 | 2 | 5 | 3 | 0 | 0 | 2 | 3 | 2 | 1 | 0 | 0 | 0 | 0 |
| Uvigerina mantaensis Cushman & Edwards | 0 | 2 | 2 | 5 | 1 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Uvigerina popescui R�gl | 43 | 30 | 23 | 14 | 3 | 4 | 7 | 4 | 16 | 11 | 18 | 40 | 8 | 0 |
| Vaginulinopsis sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| Valvulina flexilis Cushman & Renz | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Valvulineria complanata (Orbigny) | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Valvulineria palmarealensis (Nuttall) | 0 | 0 | 1 | 1 | 1 | 4 | 4 | 1 | 3 | 1 | 2 | 2 | 0 | 1 |
| Virgulinella pertusa (Reuss) | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vulvulina haeringensis (G�mbel) | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Planktonic foraminifera | ||||||||||||||
| Cassigerinella globulosa (Egger) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| Catapsydrax cf. primitivus (Blow & Banner) | 0 | 0 | 1 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Catapsydrax sp. | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Catapsydrax unicavus Bolli et al. | 0 | 0 | 1 | 2 | 0 | 1 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globigerina anguliofficinalis Blow | 2 | 3 | 0 | 1 | 4 | 3 | 0 | 1 | 4 | 2 | 7 | 6 | 0 | 1 |
| Globigerina angulisuturalis Bolli | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 3 | 0 | 0 |
| Globigerina bulloides Orbigny | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 |
| Globigerina concinna Reuss | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 |
| Globigerina diplostoma Reuss | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globigerina gnaucki Blow & Banner | 0 | 0 | 7 | 1 | 3 | 1 | 5 | 5 | 4 | 5 | 0 | 1 | 0 | 3 |
| Globigerina lentiana R�gl | 17 | 23 | 20 | 23 | 25 | 8 | 25 | 12 | 9 | 9 | 16 | 14 | 24 | 29 |
| Globigerina officinalis Subbotina | 8 | 9 | 26 | 31 | 28 | 40 | 19 | 37 | 38 | 15 | 13 | 19 | 15 | 8 |
| Globigerina ottnangiensis R�gl | 9 | 29 | 1 | 4 | 5 | 21 | 4 | 11 | 25 | 49 | 80 | 45 | 26 | 25 |
| Globigerina ouachitensis Howe & Wallace | 3 | 0 | 8 | 4 | 1 | 6 | 5 | 1 | 7 | 1 | 1 | 1 | 0 | 1 |
| Globigerina praebulloides Blow | 19 | 29 | 20 | 15 | 16 | 24 | 26 | 16 | 33 | 61 | 35 | 30 | 56 | 47 |
| Globigerina sp. | 10 | 16 | 25 | 22 | 31 | 32 | 24 | 13 | 14 | 12 | 13 | 10 | 26 | 40 |
| Globigerina tarchanensis Subbotina & Chutzieva | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 2 | 0 |
| Globigerina wagneri R�gl | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 |
| Globigerinella obesa (Bolli) | 1 | 0 | 2 | 4 | 1 | 1 | 6 | 2 | 11 | 2 | 0 | 0 | 1 | 0 |
| Globigerinoides altiaperturus Bolli | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globigerinoides quadrilobatus (Orbigny) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globigerinoides trilobus (Reuss) | 2 | 3 | 0 | 0 | 0 | 0 | 3 | 37 | 0 | 1 | 0 | 1 | 0 | 2 |
| Globoquadrina langhiana Cita & Gelati | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Globorotaloides suteri Bolli | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Globoturborotalita connecta (Jenkins) | 0 | 0 | 4 | 1 | 2 | 4 | 7 | 25 | 7 | 8 | 9 | 7 | 0 | 11 |
| Globoturborotalita woodi (Jenkins) | 1 | 1 | 2 | 0 | 0 | 0 | 2 | 4 | 7 | 21 | 3 | 9 | 4 | 1 |
| Paragloborotalia mayeri (Cushman & Ellisor) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Paragloborotalia nana (Bolli) | 5 | 2 | 0 | 3 | 1 | 3 | 1 | 1 | 2 | 1 | 4 | 2 | 0 | 0 |
| Paragloborotalia opima (Bolli) | 2 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 2 | 1 | 1 | 0 |
| Paragloborotalia pseudocontinuosa (Jenkins) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Paragloborotalia semivera (Hornibrook) | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Tenuitella clemenciae (Bermudez) | 0 | 2 | 2 | 5 | 12 | 5 | 14 | 0 | 3 | 0 | 2 | 3 | 1 | 0 |
| Tenuitella gemma (Jenkins) | 0 | 0 | 1 | 2 | 2 | 1 | 4 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tenuitella liverovskae (Bykova) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Tenuitella sp. | 0 | 0 | 0 | 0 | 4 | 3 | 3 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| Tenuitellinata angustiumbilicata (Bolli) | 2 | 6 | 1 | 14 | 29 | 15 | 40 | 8 | 12 | 5 | 9 | 11 | 6 | 1 |
| Tenuitellinata juvenilis (Bolli) | 0 | 0 | 1 | 2 | 3 | 5 | 3 | 1 | 0 | 0 | 3 | 2 | 3 | 3 |
| Tenuitellinata uvula (Ehrenberg) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 |
| Turborotalia euapertura (Jenkins) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Turborotalia sp. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Turborotalita quinqueloba (Natland) | 7 | 3 | 1 | 1 | 17 | 23 | 15 | 17 | 26 | 8 | 3 | 7 | 2 | 2 |