◄ Carnets Geol. 19 (9) ►
![]()
Outline:
[1. Introduction]
[2. Palaeogeographic, structural and stratigraphic framework]
[3. Historical account of the Aptian ammonite palaeontology in the Les Ferres area]
[4. Methods]
[5. Results]
[6. Conclusion]
[Bibliographic references] and ... [Plates]
Laboratoire du Groupe de recherche en pal�obiologie et
biostratigraphie des Ammonites, Bois-M�sanges, quartier Saint-Joseph, 04170
La Mure-Argens (France)
Laboratoire du Groupe de recherche en pal�obiologie et
biostratigraphie des Ammonites, Bois-M�sanges, quartier Saint-Joseph, 04170
La Mure-Argens (France)
R�serve naturelle nationale g�ologique de Haute-Provence, service
Environnement, Conseil d�partemental des Alpes de Haute-Provence, 13, rue
du Docteur-Romieu, CS 70216, 04995 Digne-Les-Bains cedex 9 (France)
Laboratoire G�osciences,
UMR-CNRS 6118, universit� de Rennes-1, campus Beaulieu, b�timent 15, 35042
Rennes cedex (France)
Published online in final form (pdf) on August 8, 2019
DOI 10.4267/2042/70292
![]()
[Editor: Michel Moullade; language editor:
Simon Mitchell; technical editor: Bruno Granier]
![]()
On the southern margin of the Vocontian Basin (SE France), for taphonomic reasons (fragmentation, reworking, pyritization), lower Aptian (Lower Cretaceous) deposits of the interval between the Deshayesites deshayesi and Dufrenoyia furcata zones are usually not suitable for studying the late ontogenetic developments of ammonites (fossil cephalopods). In the vicinity of the village of Les Ferres (Department of the Alpes-Maritimes, SE France), a relatively thick sedimentary succession with well-preserved ammonites, representative of the zones mentioned above, is found in a small basin called the "Les Ferres Aptian Basin" (LFAB). The 1262 ammonites collected or studied in situ are dated from the Deshayesites forbesi Zone through the top of the Dufrenoyia furcata Zone. This study presents the lithostratigraphy, biostratigraphy derived from these ammonites, and the respective range of these taxa. This is an introductory contribution to any future palaeontological study of the lower Aptian ammonites in the Les Ferres area. The lithologic unit overlying the Hauterivian-Aptian limestones is introduced herein as the Les Graous Formation. It is subdivided in three members, from bottom to top: 1) the Combe de Joinet Member, 2) the Pont de la Cerise Member, and 3) the Les Graous Member. In addition, two remarkable levels are identified: the Ammonitoceras level (outstandingly abundant) and the Toxoceratoides bed.
� Southeast of France;
� lower Aptian;
� biostratigraphy;
� ammonites
Bersac S. & Bert D. (2019).- The lower Aptian ammonites of the Les Ferres Aptian Basin (Lower Cretaceous, Southeast of France). Part I: Introduction and biostratigraphy.- Carnets Geol., Madrid, vol. 19, no. 5, p. 149-183.
Les ammonites de l'Aptien inf�rieur du bassin aptien de Les Ferres (Cr�tac� inf�rieur, SE de la France). I�re partie : Introduction et biostratigraphie.- Sur la marge m�ridionale du Bassin Vocontien (SE de la France), les d�p�ts d'�ge aptien inf�rieur (Cr�tac� inf�rieur) de l'intervalle situ� entre les zones � Deshayesites deshayesi et Dufrenoyia furcata ne sont habituellement pas favorables � l'�tude des d�veloppements ontog�n�tiques tardifs des ammonites (c�phalopodes fossiles) pour des raisons taphonomiques (fragmentation, remaniement, pyritisation). Dans les environs du village de Les Ferres (d�partement des Alpes-Maritimes, SE de la France), des s�diments aptiens relativement �pais et riches en ammonites bien conserv�es relevant de l'intervalle stratigraphique pr�cit� se sont d�pos�s dans un petit bassin structural d�nomm� ici bassin aptien de Les Ferres (LFAB). Les 1262 ammonites collect�es ou �tudi�es in situ sont dat�es de la Zone � Deshayesites forbesi jusqu'au sommet de la Zone � Dufrenoyia furcata. La lithostratigraphie, la biostratigraphie fond�e sur les ammonites et la distribution de leurs taxons respectifs sont d�crits dans ce travail qui constitue une introduction litho- et biostratigraphique � de futurs travaux pal�ontologiques sur les ammonites de l'Aptien inf�rieur du secteur des Ferres. Au-dessus des calcaires hauteriviens � aptiens, une nouvelle formation lithologique est d�crite, la Formation de Les Graous. Elle est divis�e en trois membres, de bas en haut : 1) Membre de la Combe de Joinet, 2) Membre du Pont de la Cerise et 3) Membre de Les Graous. Deux niveaux remarquables y sont identifi�s : le niveau � Ammonitoceras (exceptionnellement abondants) et le banc � Toxoceratoides.
� sud-est de la France ;
� Aptien inf�rieur ;
� biostratigraphie ;
� ammonites
In
the Vocontian Basin (Southeast of France), upper lower Aptian ammonite faunas
(middle Deshayesites deshayesi to Dufrenoyia
furcata zones sensu Bersac et
al., 2012; see Fig. 1 ![]() ) have been
repeatedly described and figured (Sayn,
1920; Thomel, 1963, 1964, 1968; Dauphin, 2002; Dutour,
2005; Joly & Delamette, 2008). Nevertheless, their study is
limited by several factors:
) have been
repeatedly described and figured (Sayn,
1920; Thomel, 1963, 1964, 1968; Dauphin, 2002; Dutour,
2005; Joly & Delamette, 2008). Nevertheless, their study is
limited by several factors:
they are generally preserved as small pyrite nuclei, which makes the study of their ontogeny difficult;
heteromorph ammonites are generally underrepresented in the Vocontian Basin for taphonomical reasons (Dauphin, 2002; Dutour, 2005) and because they were not adapted to a pelagic environment (Westermann, 1996; Lukeneder, 2015);
the conditions are usually unfavorable for preservation of large heteromorph ammonites in the neritic margins of the Vocontian Basin, with the upper lower Aptian either being absent or represented by reworked deposits yielding only fragmented ammonites (Bersac et al., 2010; Pictet et al., 2015).
Most of the large lower Aptian heteromorph ammonites in the Vocontian
Basin that have been figured in the literature are older
than the middle D.
deshayesi Zone (sensu Bersac et
al., 2012; see Fig. 1 ![]() and Roch, 1927; Delanoy, 1998; Delanoy
et al., 2008; Pictet et
al., 2009, 2015). A few of the more recent species that have been reported
include Ammonitoceras
ucetiae Dumas, 1876 (Dumas, 1876) and Toxoceratoides?
sp. inc. "Gigantomorph" godeti Thieuloy,
1990 [Thieuloy, 1990 = Lithancylus
godeti (Thieuloy, 1990), see Conte, 1999 and Mikhailova
& Baraboshkin, 2001] from the Serviers-et-Labaume and
Montaren-et-Saint-M�diers area (Gard department, southeastern France; Dumas,
1876). These ammonites come from the Deshayesites
multicostatus Subzone (D. deshayesi
Zone sensu Bersac et al.,
2012) based on the evolutionary characters of the associated Douvilleiceratinae
Parona & Bonarelli, 1897 (see Bersac & Bert,
2018).
and Roch, 1927; Delanoy, 1998; Delanoy
et al., 2008; Pictet et
al., 2009, 2015). A few of the more recent species that have been reported
include Ammonitoceras
ucetiae Dumas, 1876 (Dumas, 1876) and Toxoceratoides?
sp. inc. "Gigantomorph" godeti Thieuloy,
1990 [Thieuloy, 1990 = Lithancylus
godeti (Thieuloy, 1990), see Conte, 1999 and Mikhailova
& Baraboshkin, 2001] from the Serviers-et-Labaume and
Montaren-et-Saint-M�diers area (Gard department, southeastern France; Dumas,
1876). These ammonites come from the Deshayesites
multicostatus Subzone (D. deshayesi
Zone sensu Bersac et al.,
2012) based on the evolutionary characters of the associated Douvilleiceratinae
Parona & Bonarelli, 1897 (see Bersac & Bert,
2018).
The
Les Ferres area is located in the lower Esteron Valley (Alpes-Maritimes
Department, SE France), in the southern part of the Vocontian Basin. It was
situated at the boundary between the neritic and hemipelatic domains during the
early Aptian (Figs. 2 ![]() - 3
- 3 ![]() ). The
Aptian series were deposited in a small E-W structural basin, within a series of
tilted blocks. This palaeostructure, which is named herein the Les Ferres Aptian
Basin (LFAB), was the site for the deposition of a relatively thick sedimentary
sequence with numerous large ammonites. The interval from the Deshayesites
forbesi to the Dufrenoyia furcata zones does not include any
polyzonal or condensed intervals. The ammonite fauna is represented by the
"classical" taxa described and figured in the pelagic domain (Dufrenoyia
Kilian & Reboul, 1915, Cheloniceras Hyatt, 1903,
Pseudohaploceras Hyatt, 1900, Toxoceratoides Spath,
1924 and Colombiceras Spath, 1923, etc., see for example Dutour,
2005) associated with numerous heteromorph ammonites that belong mainly to the
genera Ammonitoceras Dumas, 1876, Lithancylus Casey,
1960, and Tropaeum J. de C. Sowerby, 1837, which were previously
poorly known in SE France. This material is sufficiently abundant and
stratigraphically constrained to allow the study of evolutionary trends and
intraspecific variability. In a nutshell, the LFAB is a unique locality to study
ammonites of this time interval.
). The
Aptian series were deposited in a small E-W structural basin, within a series of
tilted blocks. This palaeostructure, which is named herein the Les Ferres Aptian
Basin (LFAB), was the site for the deposition of a relatively thick sedimentary
sequence with numerous large ammonites. The interval from the Deshayesites
forbesi to the Dufrenoyia furcata zones does not include any
polyzonal or condensed intervals. The ammonite fauna is represented by the
"classical" taxa described and figured in the pelagic domain (Dufrenoyia
Kilian & Reboul, 1915, Cheloniceras Hyatt, 1903,
Pseudohaploceras Hyatt, 1900, Toxoceratoides Spath,
1924 and Colombiceras Spath, 1923, etc., see for example Dutour,
2005) associated with numerous heteromorph ammonites that belong mainly to the
genera Ammonitoceras Dumas, 1876, Lithancylus Casey,
1960, and Tropaeum J. de C. Sowerby, 1837, which were previously
poorly known in SE France. This material is sufficiently abundant and
stratigraphically constrained to allow the study of evolutionary trends and
intraspecific variability. In a nutshell, the LFAB is a unique locality to study
ammonites of this time interval.
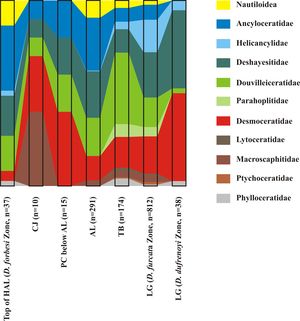
The objective of the present work is to describe the lithostratigraphy and biostratigraphy of the lower Aptian of LFAB, with the aim of supporting our future palaeontological studies of ammonites from this area.
|
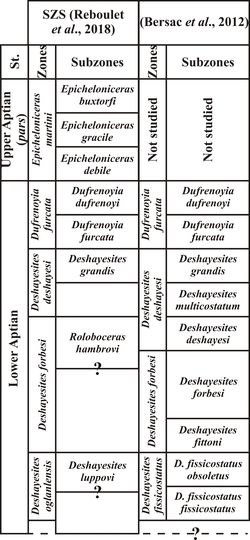
Figure 1:
Biostratigraphic charts used in the present work. SZS: Standard Mediterranean
Zonal Scheme. St.: stages. |
|
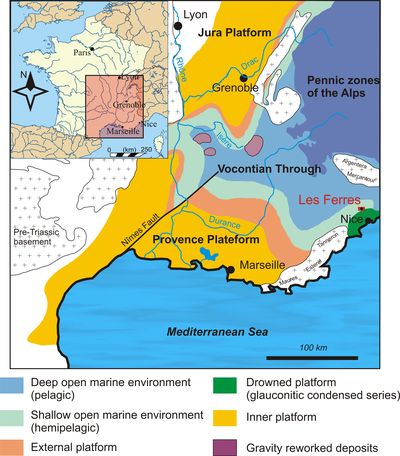
Figure 2:
Location of the Les Ferres area in the Vocontian Basin (figure after Bersac
et al., 2010, amended). |
The
studied outcrops all are located on the northern flank of Cheiron Mountain, in
the vicinity of the village of Les Ferres (Fig. 3 ![]() ).
).
The
study area belongs to the southern subalpine Castellane Arc, at the southern
margin of the Vocontian Basin. It is part of a tectonic structure on the
southern boundary of the Esteron Syncline, at the junction between two tilted
blocks. This tectonic structure is characterized by the presence of WNW-ESE
oriented half-grabens. The (synsedimentary) tectonics were active from the
Valanginian to the Albian, and are responsible for the rapid lateral change in
thickness and the numerous gaps that characterize the Lower Cretaceous
succession in this area (Dardeau, 1987; Dardeau & Graciansky,
1987). This tectonic structure has also resulted in the formation of a small EW-oriented
basin, in which the lower Aptian sediments were deposited. The Lower Cretaceous
strata were deposited in the outer neritic domain. Their facies are intermediary
between those of the hemipelagic units located to the northeast and those of the
condensed neritic series located to the southeast (Cotillon, 1971; Bersac
et al., 2010; Fig. 2 ![]() ).
).
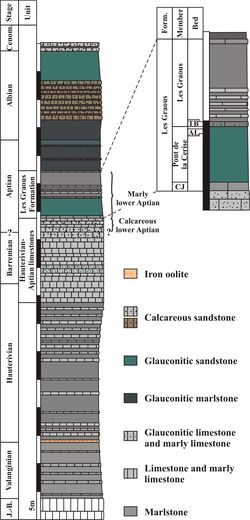
The
Aptian of the LFAB is represented by two main lithological units: a calcareous
unit overlain by a plurimetric marly unit (Cotillon, 1971, 2010; Ragazzi,
1982; Fig. 4 ![]() ); the
calcareous unit and the lower part of the marly unit are referred to the lower
Aptian and the rest of the marly unit to the upper Aptian. These deposits form a
transgressive sequence (Cotillon, 1971) related to the "middle
Bedoulian drowning event" (Br�heret, 1997; Cotillon,
2010; Masse & Fenerci-Masse, 2011, p. 671). The present work
focuses on the marly lower Aptian of the LFAB. Newly identified
lithostratigraphic units are formally described in Chapter 5.2.
); the
calcareous unit and the lower part of the marly unit are referred to the lower
Aptian and the rest of the marly unit to the upper Aptian. These deposits form a
transgressive sequence (Cotillon, 1971) related to the "middle
Bedoulian drowning event" (Br�heret, 1997; Cotillon,
2010; Masse & Fenerci-Masse, 2011, p. 671). The present work
focuses on the marly lower Aptian of the LFAB. Newly identified
lithostratigraphic units are formally described in Chapter 5.2.
|
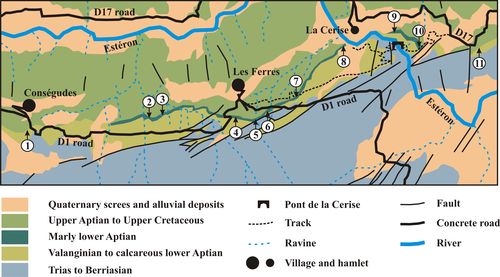
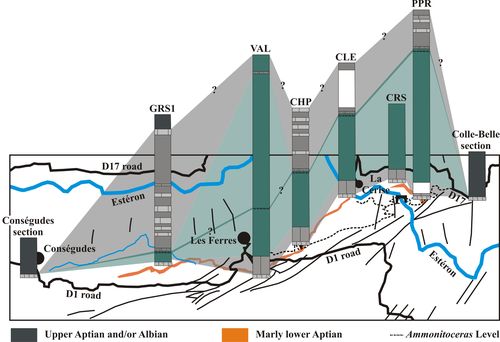
Figure 3: Map of the Les Ferres area with location of the studied or mentioned
sections. 1: Cons�gudes section; 2:
Les Graous 2 section (GRS 2); 3: Les Graous 1 section (GRS 1); 4: unnamed
section of the La Valli�re locality studied by Ragazzi (1982, p. 73),
then Br�heret (1997, p. 267, Figs. 114-115); 5: La Valli�re section
(VAL); 6: La Graou section (GRO); 7: Combe de Joinet section (CHP); 8: Pont de
la Cerise section (CLE); 9: Combe de Marin section (CRS); 10: Pont Pa�r�
section (PPR); 11: Colle-Belle section. |
|
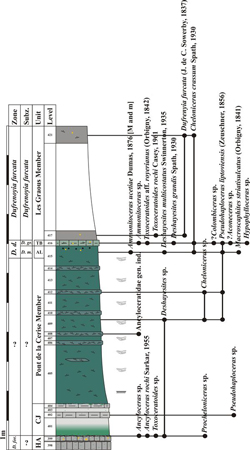
Figure 4:
Synthetic log of the Lower Cretaceous of the Les Ferres area. J.-B.: Jurassic
and Berriasian; Cenom.: Cenomanian; Form.: Formation; CJ: Combe de Joinet
Member; AL: Ammonitoceras Level; TB: Toxoceratoides Bed. |
Fallot
(1885, p. 128) was the first author to indicate the possible presence of Aptian
strata in the vicinity of Les Ferres. Subsequently, Goguel (1944), Cotillon
(1971), Ginsburg et al. (1980), Ragazzi (1982), and Br�heret
(1997) have documented the lithostratigraphy and ammonite biostratigraphy of
this area (see detailed historical accounts in Ragazzi, 1982, and Br�heret,
1997). Cotillon (1971, p. 146), for instance, was the first to mention
the presence of the ammonite genus Dufrenoyia (and consequently the
presence of the D. furcata Zone, see Fig. 1 ![]() ) in the LFAB. However, several occurrences of ammonite taxa reported in these
contributions were not supported by any illustrations. Our own investigations in
the area started in the early 2000s and resulted in preliminary publications (Bert
& Bersac, 2011; Bert, 2014) including illustrations of few
specimens. More recently, Delanoy et al. (2018) revisited the
site. They figured and described these faunas in detail. These latter authors,
focused on large heteromorphic ammonites and described ten new taxa.
) in the LFAB. However, several occurrences of ammonite taxa reported in these
contributions were not supported by any illustrations. Our own investigations in
the area started in the early 2000s and resulted in preliminary publications (Bert
& Bersac, 2011; Bert, 2014) including illustrations of few
specimens. More recently, Delanoy et al. (2018) revisited the
site. They figured and described these faunas in detail. These latter authors,
focused on large heteromorphic ammonites and described ten new taxa.
To
date, the lithostratigraphic units of the LFAB have not been formally described
in the scientific literature. In the present work, we propose a formal
lithostratigraphic scheme for the marly lower Aptian of the LFAB (Fig. 4 ![]() and
see Chapter 5.2). The name of the recognized units is that of the outcrop where
they are visible under the best conditions. Two remarkable levels are recognized
and named according to the characteristics of their respective ammonite faunas.
and
see Chapter 5.2). The name of the recognized units is that of the outcrop where
they are visible under the best conditions. Two remarkable levels are recognized
and named according to the characteristics of their respective ammonite faunas.
The
biostratigraphic scheme used in the present work is that previously proposed by Bersac
et al. (2012) for the lower Aptian, based on a review of the
Deshayesitidae Stoyanow, 1949 (Ammonoidea, Bersac & Bert,
2012, 2015; Fig. 1 ![]() ).
).
Our
objective was to prospect all the Aptian outcrops of the LFAB. Nine sections
were then identified and 8 of them were sampled (Fig. 3 ![]() ). Two
other sections close to the study area were also studied: the Cons�gudes and
the Colle-Belle sections. A total of 1,262 ammonites was bed-by-bed sampled or
studied in situ between 2000 and 2017. This material is deposited
in the collections of the Laboratory of the Research Group for Paleobiology and
Biostratigraphy of the Ammonites (G.P.A., France).
). Two
other sections close to the study area were also studied: the Cons�gudes and
the Colle-Belle sections. A total of 1,262 ammonites was bed-by-bed sampled or
studied in situ between 2000 and 2017. This material is deposited
in the collections of the Laboratory of the Research Group for Paleobiology and
Biostratigraphy of the Ammonites (G.P.A., France).
We were both present in the field when all the material published by Delanoy et al. (2018) was extracted and we reviewed it once prepared. Therefore, we deemed it necessary to include it in our study. Our own field data differ somewhat from those of Delanoy et al. (2018), and we will explain these differences below where necessary. It should be noted that the designation of specimens published by Delanoy et al. (2018) is complicated by the fact that several specimens may bear the same number, or that several numbers may be assigned to the same specimen. The problematic numbers are 28754, 28756, 28767, 28766, 28768, and 28769. Similarly, it should be noted that the holotype of Proaustraliceras bournaudi Delanoy et al. 2018, numbered AP-001 (Delanoy et al., 2018, p. 47 and Fig. 7) does not belong to the Bersac's collection contrary to the statement by these authors.
In
this chapter, we briefly discuss the taxonomic assignments used in our study for
the sole purpose of providing support for the presentation of the
lithostratigraphic and biostratigraphic results. In particular, according to our
data, the new taxa introduced by Delanoy et al. (2018) are all
subjective synonyms of taxa already described. For this reason, these taxa are
not quoted in the stratigraphic logs (Figs. 5 ![]() ,
7
,
7 ![]() ,
9
,
9 ![]() - 10
- 10 ![]() ,
12
,
12 ![]() ,
14
,
14 ![]() - 15
- 15 ![]() ). We will
expose our taxonomic positions in detail in future works.
). We will
expose our taxonomic positions in detail in future works.
5.1.1. The Deshayesitidae and Douvilleiceratidae Parona & Bonarelli, 1897
These
two ammonite families provide lower Aptian zonal and subzonal index species for
different palaeogeographic domains (Casey et al., 1998; Baraboshkin
& Mikhailova, 2002; Reboulet et al., 2018). The
criteria for identifying Deshayesitidae used in this work are those proposed by Bersac
& Bert (2012, 2015). They rely on the morphology of the ventral area
and the suture lines. The taxa we identified in the LFAB are Deshayesites
sp. (Pl. 1 ![]() , figs.
4-5), Deshayesites multicostatus Swinnerton,
1935 (Pl. 2
, figs.
4-5), Deshayesites multicostatus Swinnerton,
1935 (Pl. 2 ![]() , figs. 6-8, 13-15,
19), Deshayesites grandis Spath,
1930 (Pl. 3
, figs. 6-8, 13-15,
19), Deshayesites grandis Spath,
1930 (Pl. 3 ![]() , figs.
9-12), Dufrenoyia furcata (J. de C. Sowerby, 1837)
(Pl. 5
, figs.
9-12), Dufrenoyia furcata (J. de C. Sowerby, 1837)
(Pl. 5 ![]() , figs.
5-9), and Dufrenoyia dufrenoyi (Orbigny, 1841) (Pl. 5
, figs.
5-9), and Dufrenoyia dufrenoyi (Orbigny, 1841) (Pl. 5 ![]() , figs.
30-32). The lower Aptian Douvilleiceratidae of the LFAB are only represented by
Douvilleiceratinae. Their identification is based on the duration of their
ontogenetic stages (see Bersac & Bert, 2018, and references
therein). The identified taxa are Procheloniceras sp. (Pl. 1
, figs.
30-32). The lower Aptian Douvilleiceratidae of the LFAB are only represented by
Douvilleiceratinae. Their identification is based on the duration of their
ontogenetic stages (see Bersac & Bert, 2018, and references
therein). The identified taxa are Procheloniceras sp. (Pl. 1 ![]() , figs.
6-7),
Cheloniceras sp. and Cheloniceras crassum Spath, 1930
(Pl. 2
, figs.
6-7),
Cheloniceras sp. and Cheloniceras crassum Spath, 1930
(Pl. 2 ![]() , figs. 16-17; Pl. 3
, figs. 16-17; Pl. 3 ![]() , figs. 5-6; Pl. 4
, figs. 5-6; Pl. 4 ![]() ,
figs. 8-9).
,
figs. 8-9).
5.1.2. The Parahoplitidae Spath, 1922
The
collected material is assigned to the D. furcata Subzone (D. furcata
Zone) (Pl. 5 ![]() , figs.
10-12). All intermediates can be observed from slender forms
having a Colombiceras crassicostatum-type shape to robust forms
with a Gargasiceras gargasensis-type shape. Consequently we consider Gargasiceras
gargasensis (Orbigny, 1841) as a subjective synonym of Colombiceras
crassicostatum (Orbigny, 1841), as already suggested by Jacob
(1907), Dutour (2005) and Bulot (2010). Possible primitive
representatives of Parahoplitidae occur in the Deshayesites grandis
Subzone (D. deshayesi Zone) and are identified as ?Colombiceras
sp.
, figs.
10-12). All intermediates can be observed from slender forms
having a Colombiceras crassicostatum-type shape to robust forms
with a Gargasiceras gargasensis-type shape. Consequently we consider Gargasiceras
gargasensis (Orbigny, 1841) as a subjective synonym of Colombiceras
crassicostatum (Orbigny, 1841), as already suggested by Jacob
(1907), Dutour (2005) and Bulot (2010). Possible primitive
representatives of Parahoplitidae occur in the Deshayesites grandis
Subzone (D. deshayesi Zone) and are identified as ?Colombiceras
sp.
5.1.3. The Ancyloceratidae Meek, 1876
The genus Ammonitoceras is represented by large ammonites with criocone, aspinoceratic or ancyloceratic coiling, characterized by the presence of a particular ontogenetic stage in the inner whorls: the Ammonitoceras stage. This stage is characterized by the presence of bituberculated main ribs bearing an umbilical tubercle and a lateral tubercle of larger size. In a future article, we will publish a detailed study of Ammonitoceras from the D. deshayesi and D. furcata zones of the LFAB.
Preliminary results from this study suggest that:
the genus Ammonitoceras shows significant intraspecific variability that mainly relates to the duration of the ontogenetic Ammonitoceras stage;
Ammonitoceras is probably sexually
dimorphic, with large crioconic macroconchs (Pl. 2 ![]() , figs.
1-2; Pl. 4
, figs.
1-2; Pl. 4 ![]() , figs.
1-3)
and microconchs with aspinoceratic or ancyloceratic coiling (Pl. 2
, figs.
1-3)
and microconchs with aspinoceratic or ancyloceratic coiling (Pl. 2 ![]() , figs.
3-5);
, figs.
3-5);
Ammonitoceras macroconchs of the D. furcata Zone
(Pl. 4 ![]() , figs.
1-3) have on average an Ammonitoceras stage disappearing later during ontogeny
and a narrower spiral hiatus than those of the D. multicostatus Subzone (Pl. 2
, figs.
1-3) have on average an Ammonitoceras stage disappearing later during ontogeny
and a narrower spiral hiatus than those of the D. multicostatus Subzone (Pl. 2 ![]() , figs.
1-2);
, figs.
1-2);
Ammonitoceras of the D. multicostatus Subzone can be assigned to the species Ammonitoceras ucetiae and those of the D. furcata Zone to Ammonitoceras lahuseni (Sinzow, 1906);
we interpret Ammonitoceras dumasi Delanoy et al., 2018 as the macroconch of A. ucetiae and regard it as a subjective synonym of that species;
we interpret the following species and specimens as microconchs of A. ucetiae: Caseyites esteronensis Delanoy et al., 2018, Caseyites vermeuleni Delanoy et al., 2018, Ammonitoceras madouxi Delanoy et al., 2018, Proaustraliceras bournaudi Delanoy et al., 2018, the specimen identified under ?Proaustraliceras sp. numbered 28766 or 28767 in Delanoy et al. (2018, p. 50-51, Fig. 9.A) and the specimen of Ammonitoceras aff. madouxi numbered 28764 in Delanoy et al. (2018, p. 71-72, Fig. 19);
according to our interpretation, A. leiferrasensis Delanoy et al., 2018, Caspianites ragazziae Delanoy et al., 2018, and the specimen figured under ?Caspianites sp. and numbered 28744 or 28754 in Delanoy et al., (2018, p. 83, Fig. 25.A-B and p. 84) are all macroconchs of A. lahuseni and thus subjective synonyms of that species;
in our view, Caseyites morenobedmari Delanoy et al., 2018 is a microconch of A. lahuseni and consequently a subjective synonym of that species; and
Ammonitoceras specimens from the D. grandis Subzone are too incomplete to be identified at the specific rank.
Lithancylus
of the D. multicostatus Subzone of the LFAB (Pl. 2 ![]() , figs.
11-12) appear
to have significant intraspecific variability, which concerns both their
ontogenetic sequence and adult size. In our opinion, these specimens belong to
the species Lithancylus grandis (J. de C. Sowerby, 1828), and
consequently we interpret Lithancylus bifurcatus Delanoy et al.,
2018 as a subjective synonym of that species.
, figs.
11-12) appear
to have significant intraspecific variability, which concerns both their
ontogenetic sequence and adult size. In our opinion, these specimens belong to
the species Lithancylus grandis (J. de C. Sowerby, 1828), and
consequently we interpret Lithancylus bifurcatus Delanoy et al.,
2018 as a subjective synonym of that species.
5.1.4. The Helicancylidae Hyatt, 1894
On
the basis of the diagnostic characteristics proposed by Bulot et al.
(2017) and Frau et al. (2017), we identified Toxoceratoides
sp., Toxoceratoides aff. royerianus (Orbigny, 1842)
(Pl. 3 ![]() , figs.
13-16) and Toxoceratoides rochi Casey, 1961 (Pl. 5
, figs.
13-16) and Toxoceratoides rochi Casey, 1961 (Pl. 5 ![]() , figs.
1-4).
, figs.
1-4).
5.1.5. The Ptychoceratidae Gill, 1871
The
genus Ptychoceras Orbigny, 1842, is represented by rare specimens
we identified as Ptychoceras emericianum Orbigny, 1842
(Pl. 5 ![]() , fig.
15) on the basis of the revision work of Vermeulen et al.
(2015).
, fig.
15) on the basis of the revision work of Vermeulen et al.
(2015).
5.1.6. The Desmoceratidae Zittel, 1895
We
identified the Desmoceratidae of the LFAB on the basis of the diagnostic
criteria proposed by Dutour (2005), in particular the aspect of the whorl
section. In this respect, identification of the collected specimens at the
specific rank was made difficult by the fact that they are almost always
deformed and crushed. The taxa identified are: Pseudohaploceras sp. (Pl. 1 ![]() , fig.
8), Pseudohaploceras liptoviensis (Zeuschner, 1856)
(Pl. 2
, fig.
8), Pseudohaploceras liptoviensis (Zeuschner, 1856)
(Pl. 2 ![]() , figs. 9-10; Pl. 4
, figs. 9-10; Pl. 4 ![]() , figs.
6-7), Pseudosaynella sp., Aconeceras
nisum (Orbigny, 1841) (Pl. 5
, figs.
6-7), Pseudosaynella sp., Aconeceras
nisum (Orbigny, 1841) (Pl. 5 ![]() , figs.
13-14) and ?Aconeceras sp.
, figs.
13-14) and ?Aconeceras sp.
5.1.7. The Lytoceratidae Neumayr, 1875
Representatives
of this family are rare in the LFAB and are attributed to Lytoceras sp. (Pl. 4 ![]() , figs.
10-11).
, figs.
10-11).
5.1.8. The Macroscaphitidae Hyatt, 1900
Based
on Busnardo�s remarks (in Gauthier, 2006), we identified Macroscaphites
aff. yvani (Puzos, 1832) and Macroscaphites striatisulcatus
(Orbigny, 1841) (Pl. 2 ![]() , fig. 20; Pl. 3
, fig. 20; Pl. 3 ![]() , figs. 3-4;
Pl. 5
, figs. 3-4;
Pl. 5 ![]() , fig.
16).
, fig.
16).
Following
the works of Wright et al. (1996),
Klein et al. (2007) and Va��ček et al. (2014),
we consider Pseudocrioceratites Egoian, 1969, as a subjective
synonym of Acantholytoceras Spath, 1923. The Acantholytoceras
we collected in the LFAB are too fragmented to be identified to specific level (Pl. 4 ![]() , figs.
4-5).
, figs.
4-5).
5.1.9. The Phylloceratidae Zittel, 1884
In
the LFAB, this family is represented only by the subgenus Hypophylloceras
Salfeld, 1924 (Phylloceras Suess, 1865). Joly & Delamette
(2008, Fig. 74) recognized 6 species of Phylloceras (Hypophylloceras)
ranging through the D. deshayesi and D. furcata zones (sensu Reboulet
et al. 2018) in the Vocontian Basin. The diagnostic characters are subtle
variations in ornamentation, section or morphology of the suture line. Our
specimens are not sufficiently well preserved to be identified to the specific
level (Pl. 3 ![]() ,
fig. 1; Pl. 4
,
fig. 1; Pl. 4 ![]() , fig.
12).
, fig.
12).
5.2.1.
Top of the Hauterivian-Aptian limestones (Fig. 4 ![]() )
)
In
the neritic domain of the Castellane Arc, Hauterivian to lower Aptian deposits
are represented by a pluridecametric succession of massive limestone beds. Cotillon
(1971) divided it into a series of formations (formations 17 to 25, Cotillon,
1971, p. 34 and Fig. 12bis). In the LFAB, the Hauterivian-Aptian limestones are
represented by a plurimetric succession of massive decimetric limestone beds
separated by centimetric marly intervals (Fig. 4 ![]() ) that have already been
described in the literature under various names ("Calcaire blanc
compacte" in Fallot, 1885, p. 127, "Barr�mo-B�doulien"
in Ragazzi, 1982, p. 73).
) that have already been
described in the literature under various names ("Calcaire blanc
compacte" in Fallot, 1885, p. 127, "Barr�mo-B�doulien"
in Ragazzi, 1982, p. 73).
The
highest beds in the upper part of this lithological unit contain increasing
amount of sandstone and glauconite (Ragazzi, 1982), and corresponds to Cotillon's
(1971) Formation 25, and these are assigned to the lower Aptian (pars, Ginsburg
et al., 1980; Ragazzi, 1982). The upper limit of the Hauterivian-Aptian
limestones is represented by a firmground-type, with iron crust layers and
extensive bioturbation by Rhizocorallium Zenker, 1836
(Fig. 8.D ![]() ).
This discontinuity surface is correlated with the "early Aptian
unconformity" (Cotillon, 2010, p. 4) that is related to the
"middle Bedoulian drowning event" (Masse & Fenerci-Masse,
2011, p. 671). It corresponds to a large-scale discontinuity surface formed
during the flooding of the Proven�al platform (Br�heret, 1997; Cotillon,
2010; Masse & Fenerci-Masse, 2011). This event is assigned to
the upper D. forbesi Zone (Pictet et al., 2015;
Fig. 1
).
This discontinuity surface is correlated with the "early Aptian
unconformity" (Cotillon, 2010, p. 4) that is related to the
"middle Bedoulian drowning event" (Masse & Fenerci-Masse,
2011, p. 671). It corresponds to a large-scale discontinuity surface formed
during the flooding of the Proven�al platform (Br�heret, 1997; Cotillon,
2010; Masse & Fenerci-Masse, 2011). This event is assigned to
the upper D. forbesi Zone (Pictet et al., 2015;
Fig. 1 ![]() ).
).
We
only sampled the last Hauterivian-Aptian limestone bed: it is represented by a
centimetric level of grey to yellowish sandstone relatively rich in internal
moulds of ammonites, nautiloids and belemnites (Fig. 13.A ![]() ). Ragazzi (1982,
p. 74-75) reported the following ammonite fauna: Ancyloceras audouli (Astier,
1851), Deshayesites latilobatus (Sinzow, 1910) [= �D.
latelobata (Sinzow)' in Ragazzi, 1982, p. 75], Deshayesites
sp., Cheloniceras sp., Procheloniceras sp., Pseudohaploceras
sp., Costidiscus recticostatus (Orbigny, 1841). We identified: Ancyloceras
rochi Sarkar, 1955 (Pl. 1
). Ragazzi (1982,
p. 74-75) reported the following ammonite fauna: Ancyloceras audouli (Astier,
1851), Deshayesites latilobatus (Sinzow, 1910) [= �D.
latelobata (Sinzow)' in Ragazzi, 1982, p. 75], Deshayesites
sp., Cheloniceras sp., Procheloniceras sp., Pseudohaploceras
sp., Costidiscus recticostatus (Orbigny, 1841). We identified: Ancyloceras
rochi Sarkar, 1955 (Pl. 1 ![]() , figs.
1-2), Ancyloceras sp. (Pl. 1
, figs.
1-2), Ancyloceras sp. (Pl. 1 ![]() , fig.
3), Toxoceratoides sp., Deshayesites sp. (Pl. 1
, fig.
3), Toxoceratoides sp., Deshayesites sp. (Pl. 1 ![]() ,
figs. 4-5), Procheloniceras
sp. (Pl. 1
,
figs. 4-5), Procheloniceras
sp. (Pl. 1 ![]() , figs.
6-7), Pseudohaploceras sp. (Pl. 1
, figs.
6-7), Pseudohaploceras sp. (Pl. 1 ![]() ,
fig. 8), Pseudohaploceras
liptoviensis, Hypophylloceras sp. This fauna is associated with
numerous nautiloids [Cymatoceras neckerianus (Pictet, 1847)
(Pl. 1
,
fig. 8), Pseudohaploceras
liptoviensis, Hypophylloceras sp. This fauna is associated with
numerous nautiloids [Cymatoceras neckerianus (Pictet, 1847)
(Pl. 1 ![]() , fig.
9)] and belemnites (Mesohibolites sp.).
, fig.
9)] and belemnites (Mesohibolites sp.).
We
could not identify any Deshayesites from this bed at the specific rank
because the characteristics of their ventral area in the inner whorls and their
suture line could not be observed (Bersac & Bert, 2012, 2015).
Nevertheless, their rounded ventral areas on the phragmocone suggests that they
are older than the D. grandis Subzone (Pl. 1 ![]() , fig.
5).
, fig.
5).
The
Douvilleiceratidae collected in this bed have a relatively long and well
expressed Procheloniceras stage and no Cheloniceras stage was observed (Bersac
& Bert, 2018), but the innermost whorls of the studied specimens are
not preserved: if the Cheloniceras stage occurs, it is restricted to innermost
whorls only (Pl. 1 ![]() , figs.
6-7). The shape of these specimens with more primitive
features than Cheloniceras cornuelianum (Orbigny, 1841) leads us
to attribute them to the genus Procheloniceras Spath, 1923. This
genus seems to occur at the top of the D. forbesi Zone (sensu Bersac
et al., 2012), whereas the genus Cheloniceras succeeds it from the
base of the D. deshayesi Zone (Bersac & Bert, 2018). It
should be noted that we have not collected any representative of the
stratigraphically significant family Roloboceratinae Casey, 1961. Their
representatives occur in the lower part of the D. forbesi Zone (sensu Bersac
et al., 2012) in the Boreal realm and at least from the upper part of the
D. forbesi Zone or from the base of the D. deshayesi Zone (sensu Bersac
et al., 2012) in the Tethysian realm (Casey et al., 1998; Bersac
& Bert, 2012, Fig. 17, 2015, Fig. 15).
, figs.
6-7). The shape of these specimens with more primitive
features than Cheloniceras cornuelianum (Orbigny, 1841) leads us
to attribute them to the genus Procheloniceras Spath, 1923. This
genus seems to occur at the top of the D. forbesi Zone (sensu Bersac
et al., 2012), whereas the genus Cheloniceras succeeds it from the
base of the D. deshayesi Zone (Bersac & Bert, 2018). It
should be noted that we have not collected any representative of the
stratigraphically significant family Roloboceratinae Casey, 1961. Their
representatives occur in the lower part of the D. forbesi Zone (sensu Bersac
et al., 2012) in the Boreal realm and at least from the upper part of the
D. forbesi Zone or from the base of the D. deshayesi Zone (sensu Bersac
et al., 2012) in the Tethysian realm (Casey et al., 1998; Bersac
& Bert, 2012, Fig. 17, 2015, Fig. 15).
Small Ancyloceras Orbigny, 1842, similar to those collected at the top of the Hauterivian-Aptian limestones, were reported in the Deshayesites weissi Zone (= D. forbesi Zone sensu Reboulet et al., 2018) by Ropolo et al. (1998, p. 171) in the Roquefort-la B�doule area (SE France); they were identified as Ancyloceras matheronianum Orbigny, 1842. Pierre Ropolo provided photographs of these specimens to us and, from our point of view, they are similar to the type specimen of Ancyloceras rochi, originally described and figured by Roch (1927, p. 29, Pl. IV, fig. 2) as "Ancyloceras Binelli Astier".
Based
on these considerations, the last Hauterivian-Aptian limestone bed of the LFAB
probably belongs to the lower part of the D. forbesi Zone sensu Bersac
et al. (2012; Fig. 1 ![]() ).
).
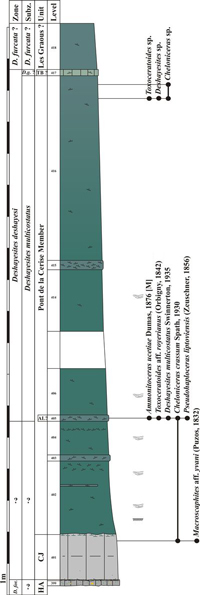
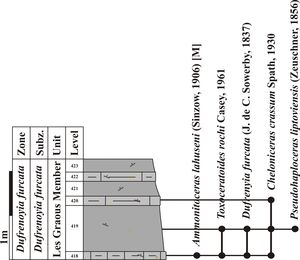
5.2.2. The Les Graous Formation
This
formation is formally defined for the first time in the present work (Fig. 4 ![]() )
and is restricted to the LFAB. It is represented by dark green glauconitic
sandstones in its lower part and grey-blue marls in its upper part. Its lower
limit is the terminal discontinuity of the Hauterivian-Aptian limestones. Its
upper limit is located at the top of a marly-limestone level just below a dark
grey glauconitic level (Fig. 4
)
and is restricted to the LFAB. It is represented by dark green glauconitic
sandstones in its lower part and grey-blue marls in its upper part. Its lower
limit is the terminal discontinuity of the Hauterivian-Aptian limestones. Its
upper limit is located at the top of a marly-limestone level just below a dark
grey glauconitic level (Fig. 4 ![]() ). This glauconitic level yielded the oldest Epicheloniceras
martini (Orbigny, 1842) of the study area and was therefore assigned
to the upper Aptian Epicheloniceras debile Subzone (Epicheloniceras
martini Zone, see Dutour, 2005 and Fig. 1
). This glauconitic level yielded the oldest Epicheloniceras
martini (Orbigny, 1842) of the study area and was therefore assigned
to the upper Aptian Epicheloniceras debile Subzone (Epicheloniceras
martini Zone, see Dutour, 2005 and Fig. 1 ![]() ). The
Les Graous Formation type section is Les Graous 1 (GRS1, Figs. 3
). The
Les Graous Formation type section is Les Graous 1 (GRS1, Figs. 3 ![]() ,
5
,
5 ![]() - 6
- 6 ![]() ,
and see Chapter 5.3.1), where it is observable in its entirety. The Les
Graous Formation is the equivalent of Cotillon's (1971, p. 69) formations
ab1 and ab2 of the neritic domain of the Castellane Arc. At Les Ferres, this
author (ibid., p. 146) named it "[a, b]".
,
and see Chapter 5.3.1), where it is observable in its entirety. The Les
Graous Formation is the equivalent of Cotillon's (1971, p. 69) formations
ab1 and ab2 of the neritic domain of the Castellane Arc. At Les Ferres, this
author (ibid., p. 146) named it "[a, b]".
|
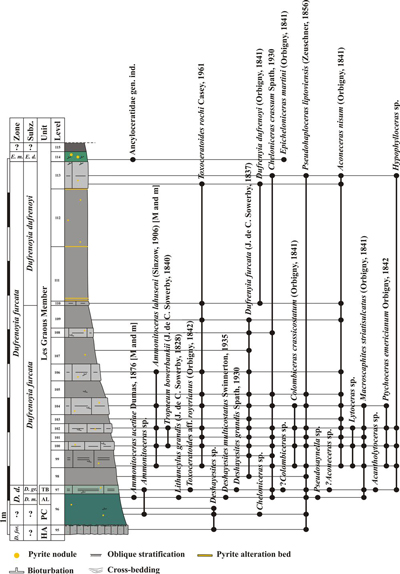
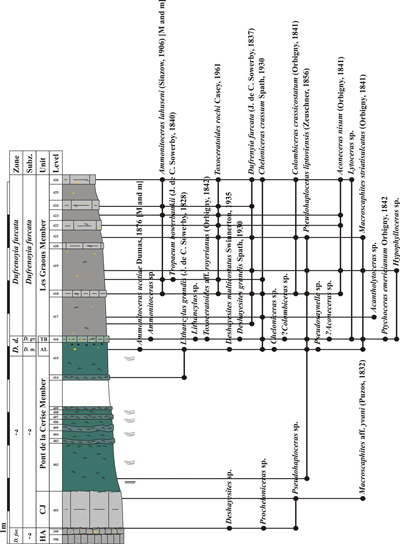
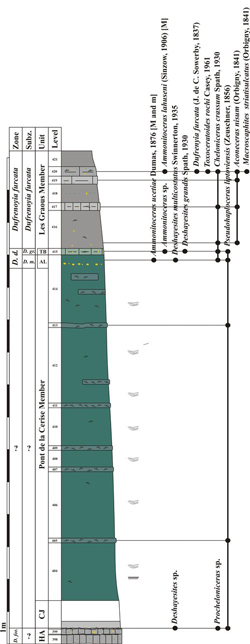
Figure 5: Log of the Les Graous 1 section
(GRS 1) with distribution of the ammonite taxa, see Fig. 4 |
|
Figure 6: Field photographies of the Les Graous 1 section
(GRS 1). Log of this section
is figured in Fig. 5 |
We propose to divide the Les Graous Formation into several lithostratigraphic members, described here for the first time:
This
lithostratigraphic unit is restricted to the LFAB (Fig. 4 ![]() ). It was previously
mentioned by Ragazzi (1982, p. 76), who named it "marnes bleut�es"
("bluish marls").
). It was previously
mentioned by Ragazzi (1982, p. 76), who named it "marnes bleut�es"
("bluish marls").
The
Combe de Joinet Member is represented by 1.10-1.50 m of light grey marly
limestone in which are sometimes intercalated two more indurated decimetric
levels or a glauconitic decimetric level. The lower limit of this member is the
discontinuity surface at the top of the Hauterivian-Aptian limestones. Its upper
limit is an intensely bioturbated discontinuity surface (ichnogenus Thalassinoides
Ehrenberg, 1944; Fig. 13.B ![]() ). The type section of this member is the Combe
de Joinet section (CHP, Figs. 3
). The type section of this member is the Combe
de Joinet section (CHP, Figs. 3 ![]() ,
10
,
10 ![]() - 11
- 11 ![]() ,
and see Chapter 5.3.4). The ammonite
fauna of the Combe de Joinet Member is particularly rare and only represented by
fragmented internal moulds of Ancyloceras sp., Deshayesites sp., Cheloniceras
crassum, Macroscaphites aff. yvani, Pseudohaploceras
liptoviensis and Pseudohaploceras sp. This member crops out in the
following sections: La Valli�re (VAL, see Figs. 3
,
and see Chapter 5.3.4). The ammonite
fauna of the Combe de Joinet Member is particularly rare and only represented by
fragmented internal moulds of Ancyloceras sp., Deshayesites sp., Cheloniceras
crassum, Macroscaphites aff. yvani, Pseudohaploceras
liptoviensis and Pseudohaploceras sp. This member crops out in the
following sections: La Valli�re (VAL, see Figs. 3 ![]() ,
7
,
7 ![]() - 8
- 8 ![]() ,
and Chapter 5.3.2), Combe de Joinet (CHP), Pont de la Cerise (CLE, see Figs. 3
,
and Chapter 5.3.2), Combe de Joinet (CHP), Pont de la Cerise (CLE, see Figs. 3 ![]() ,
12
,
12 ![]() - 13
- 13 ![]() ,
and Chapter 5.3.5) and Combe de Marin (CRS, see Figs. 3
,
and Chapter 5.3.5) and Combe de Marin (CRS, see Figs. 3 ![]() ,
14
,
14 ![]() ,
and Chapter 5.3.6). It also
partly crops out in the Pont Pa�r� section (PPR, see Figs. 3
,
and Chapter 5.3.6). It also
partly crops out in the Pont Pa�r� section (PPR, see Figs. 3 ![]() ,
15
,
15 ![]() - 16
- 16 ![]() ,
and
Chapter 5.3.7).
,
and
Chapter 5.3.7).
|
Figure
7: Log of the La Valli�re section (VAL) with distribution of ammonite
taxa, see Figs. 4 |
|
Figure
8:
Field photographies of the La Valli�re section (VAL). Log of this section is
figured in Fig. 7 |
This
member is restricted to the LFAB (Fig. 4 ![]() ) and is represented by massive
glauconitic sandstones (Ragazzi, 1982; Br�heret, 1997) in which
are often intercalated more or less continuous light grey decimetric beds (Figs. 8.A-B
) and is represented by massive
glauconitic sandstones (Ragazzi, 1982; Br�heret, 1997) in which
are often intercalated more or less continuous light grey decimetric beds (Figs. 8.A-B ![]() ,
16.A
,
16.A ![]() ). The thickness of this member varies from less than one metre in
the Les Graous 1 section (GRS1) to 15 m in the La Valli�re section (VAL, Fig. 17
). The thickness of this member varies from less than one metre in
the Les Graous 1 section (GRS1) to 15 m in the La Valli�re section (VAL, Fig. 17 ![]() ). The lower limit of this member is represented by the discontinuity surface
at the top of the underlying Combe de Joinet Member. Its upper limit is
characterized by the abrupt end of the glauconitic sandstones, which are
overlain by a characteristic, more or less marly, grey limestone bed, here named
the "Toxoceratoides Bed" (which belongs to the overlying Les
Graous Member, see Chapter 5.2.2.3). The type section of the Pont de la Cerise
Member is at the Pont de la Cerise section (CLE, see Figs. 3
). The lower limit of this member is represented by the discontinuity surface
at the top of the underlying Combe de Joinet Member. Its upper limit is
characterized by the abrupt end of the glauconitic sandstones, which are
overlain by a characteristic, more or less marly, grey limestone bed, here named
the "Toxoceratoides Bed" (which belongs to the overlying Les
Graous Member, see Chapter 5.2.2.3). The type section of the Pont de la Cerise
Member is at the Pont de la Cerise section (CLE, see Figs. 3 ![]() ,
12
,
12 ![]() - 13
- 13 ![]() ,
and Chapter 5.3.5). The first decimetres of this member are relatively rich in phosphate
nodules, belemnite fragments and fish teeth. When this member reaches a
plurimetric thickness, the first two meters display cross-stratifications.
Bioturbation is frequent throughout the Pont de la Cerise Member, especially in
the more indurated levels (Figs. 5
,
and Chapter 5.3.5). The first decimetres of this member are relatively rich in phosphate
nodules, belemnite fragments and fish teeth. When this member reaches a
plurimetric thickness, the first two meters display cross-stratifications.
Bioturbation is frequent throughout the Pont de la Cerise Member, especially in
the more indurated levels (Figs. 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() - 9
- 9 ![]() - 10
- 10 ![]() - 11
- 11 ![]() - 12
- 12 ![]() - 13
- 13 ![]() - 14
- 14 ![]() - 15
- 15 ![]() - 16
- 16 ![]() , Thalassinoides in
Fig. 13.C
, Thalassinoides in
Fig. 13.C ![]() ).
).
The glauconitic sandstones of the Pont de la Cerise Member were first mentioned by Goguel (1944, p. 21) who assigned them to the Albian. Ginsburg et al. (1980) and Ragazzi (1982, p. 74) dated them as Aptian. A level remarkably rich in ammonites generally occurs in the upper 0.5 m of this member. It is named here the "Ammonitoceras Level" and will be described below. The Pont de la Cerise Member crops out in the following sections: Les Graous 1 (GRS1), Les Graous 2 (GRS2), La Valli�re (VAL), Combe de Joinet (CHP), Pont de la Cerise (CLE), Combe de Marin (CRS), and Pont Pa�r� (PPR).
|
Figure
9: Log of the La Graou section (GRO) with distribution of ammonite
taxa, see Fig. 5 |
|
Figure
10: Log of the Combe de Joinet section (CHP) with distribution of ammonite
taxa,
see Figs. 4 |
|
Figure
11: Field photographs of the Combe de Joinet section (CHP). Log of this section
is figured Fig. 10 |
Apart
from the Ammonitoceras Level, the ammonite fauna of the Pont de la Cerise
Member is represented by rare and poorly preserved internal moulds (Fig. 13.D ![]() ).
We recognized the following taxa: Ammonitoceras sp., Lithancylus
grandis, Deshayesites sp. Cheloniceras sp., Pseudohaploceras
liptoviensis and a fragmentary Ancyloceratidae unidentifiable at the generic
rank. Belemnites are represented by Neohibolites aptiensis (Stolley,
1913) and Duvalia grasi (Duval-Jouve, 1841). A level one
centimetre above the Ammonitoceras Level yielded small internal phosphate
moulds of Macroscaphites striatisulcatus and Pseudohaploceras
liptoviensis.
).
We recognized the following taxa: Ammonitoceras sp., Lithancylus
grandis, Deshayesites sp. Cheloniceras sp., Pseudohaploceras
liptoviensis and a fragmentary Ancyloceratidae unidentifiable at the generic
rank. Belemnites are represented by Neohibolites aptiensis (Stolley,
1913) and Duvalia grasi (Duval-Jouve, 1841). A level one
centimetre above the Ammonitoceras Level yielded small internal phosphate
moulds of Macroscaphites striatisulcatus and Pseudohaploceras
liptoviensis.
The
Ammonitoceras Level is a 0.20 m-thick level particularly rich in
ammonites. It is located approximately 0.50 m below the upper limit of the Pont
de la Cerise Member (Figs. 4 ![]() ,
11.C-D
,
11.C-D ![]() ). This level provided an abundant
cephalopod fauna mainly consisting of deformed internal moulds of ammonites and
nautiloids, and belemnite rostra (Pl. 2
). This level provided an abundant
cephalopod fauna mainly consisting of deformed internal moulds of ammonites and
nautiloids, and belemnite rostra (Pl. 2 ![]() , figs. 1-17,
19-20). The proportion of
subcomplete representatives of the heteromorphic ammonite family Ancyloceratidae,
especially Ammonitoceras,
is noteworthy (Fig. 11.D
, figs. 1-17,
19-20). The proportion of
subcomplete representatives of the heteromorphic ammonite family Ancyloceratidae,
especially Ammonitoceras,
is noteworthy (Fig. 11.D ![]() ;
Pl. 2
;
Pl. 2 ![]() ,
figs. 1-5). The type section of this reference
level is the Combe de Joinet section (CHP) where it crops out under particularly
favourable conditions (see Chapter 5.3.4). Heteromorph ammonites are deposited
without preferential orientation (Fig. 11.D
,
figs. 1-5). The type section of this reference
level is the Combe de Joinet section (CHP) where it crops out under particularly
favourable conditions (see Chapter 5.3.4). Heteromorph ammonites are deposited
without preferential orientation (Fig. 11.D ![]() ). Bert (2014, p. 393) figured
this level for the first time (Fig. 11.D
). Bert (2014, p. 393) figured
this level for the first time (Fig. 11.D ![]() ), and Delanoy et al.
(2018) described and figured many ammonites. We identified the following taxa in
the Ammonitoceras Level: Ammonitoceras ucetiae macroconchs (Pl. 2
), and Delanoy et al.
(2018) described and figured many ammonites. We identified the following taxa in
the Ammonitoceras Level: Ammonitoceras ucetiae macroconchs (Pl. 2 ![]() , figs.
1-2) and microconchs (Pl. 2
, figs.
1-2) and microconchs (Pl. 2 ![]() , figs.
3-5), Lithancylus grandis (Pl. 2
, figs.
3-5), Lithancylus grandis (Pl. 2 ![]() , figs.
11-12), Deshayesites
multicostatus (Pl. 2
, figs.
11-12), Deshayesites
multicostatus (Pl. 2 ![]() , figs.
6-8, 13-15, 19), Cheloniceras crassum
(Pl. 2
, figs.
6-8, 13-15, 19), Cheloniceras crassum
(Pl. 2 ![]() , figs.
16-17), Cheloniceras sp., Pseudohaploceras liptoviensis
(Pl. 2
, figs.
16-17), Cheloniceras sp., Pseudohaploceras liptoviensis
(Pl. 2 ![]() , figs.
9-10), Pseudosaynella sp., Macroscaphites
striatisulcatus (Pl. 2
, figs.
9-10), Pseudosaynella sp., Macroscaphites
striatisulcatus (Pl. 2 ![]() ,
fig. 20). Belemnites are represented by Neohibolites
aptiensis and Duvalia grasi. Nautiloids are frequent and represented
by Cymatoceras neckerianus, Eucymatoceras plicatum (Fitton,
1836) and Anglonautilus sp. The level 405 of the La Valliere section
(VAL) is a possible local representative of the Ammonitoceras Level (see
Chapter 5.3.2) and additionally provided Toxoceratoides aff. royerianus.
Gastropods, bivalves and ahermatypic corals are relatively rare (Pl. 2
,
fig. 20). Belemnites are represented by Neohibolites
aptiensis and Duvalia grasi. Nautiloids are frequent and represented
by Cymatoceras neckerianus, Eucymatoceras plicatum (Fitton,
1836) and Anglonautilus sp. The level 405 of the La Valliere section
(VAL) is a possible local representative of the Ammonitoceras Level (see
Chapter 5.3.2) and additionally provided Toxoceratoides aff. royerianus.
Gastropods, bivalves and ahermatypic corals are relatively rare (Pl. 2 ![]() ,
fig.
18).
,
fig.
18).
Deshayesitidae
of the Ammonitoceras Level have a smooth siphonal band ending at a
diameter between 15 and 18 mm and their ventral area is not subtabulated on the
phragmocone (Pl. 2 ![]() , figs. 8,
14). These characters correspond to the index
species D. multicostatus (Bersac & Bert, 2015). The
genus Cheloniceras is represented by two distinct morphologies:
, figs. 8,
14). These characters correspond to the index
species D. multicostatus (Bersac & Bert, 2015). The
genus Cheloniceras is represented by two distinct morphologies:
a
relatively frequent one, characterized by a long Cheloniceras stage and a short
Procheloniceras stage. The "intermediate stage" of Cheloniceras
parinodum Casey, 1961, is present in some specimens between the
Cheloniceras and Procheloniceras stages (Bersac & Bert, 2018).
For these reasons, we attributed this morphology to a primitive form of C.
crassum (Pl. 2 ![]() , figs.
16-17);
, figs.
16-17);
a much rarer second morphology, with evolute coiling and with relatively short Cheloniceras stage. This form we left in open nomenclature, resembles the primitive parahoplitid Colombiceras and could be its ancestor.
The
presence of D. multicostatus and C. crassum with primitive
morphology in the Ammonitoceras Level indicates an age corresponding to
the late part of the D. multicostus
Subzone (D. deshayesi Zone) sensu Bersac et al. (2012;
Fig. 1 ![]() ). The morphology of
the Ammonitoceras in the Ammonitoceras Level, with a short
Ammonitoceras ontogenetic stage and an important spiral hiatus in macroconchs (Pl. 2
). The morphology of
the Ammonitoceras in the Ammonitoceras Level, with a short
Ammonitoceras ontogenetic stage and an important spiral hiatus in macroconchs (Pl. 2 ![]() , fig.
1), is consistent with such an age.
, fig.
1), is consistent with such an age.
The absence of Deshayesites bearing a longer smooth siphonal band and a subtabulated ventral area on the phragmocone (corresponding to the index species D. grandis, see Bersac & Bert, 2012), allows us to exclude the presence of the D. grandis Subzone in the Ammonitoceras Level.
The Ammonitoceras Level crops out in the following sections: Les Graous 1 (GRS1), Les Graous 2 (GRS2), La Valli�re (VAL), Combe de Joinet (CHP), Pont de la Cerise (CLE) and Pont Pa�r� (PPR).
|
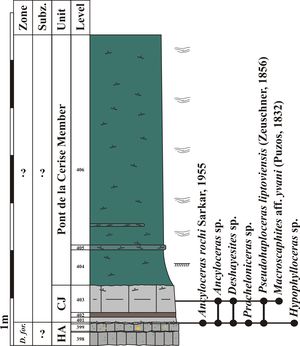
Figure
12: Log of the Pont de la Cerise section (CLE) with distribution of ammonite
taxa, see Figs. 4 |
|
Figure
13: Field photographs of the Pont de la Cerise section (CLE). Log of this
section is available in Fig. 12 |
This
member is restricted to the LFAB (Fig. 4 ![]() ). It starts with a marly limestone
level of about 0.2 m thickness, named here the Toxoceratoides Bed (Figs. 4
). It starts with a marly limestone
level of about 0.2 m thickness, named here the Toxoceratoides Bed (Figs. 4 ![]() ,
11.C
,
11.C ![]() ). The type section of the Les Graous Member is at the Les Graous 1
section (GRS1).
). The type section of the Les Graous Member is at the Les Graous 1
section (GRS1).
The
levels of the Les Graous Member above the Toxoceratoides Bed are
represented in the sections at Les Graous 1 (GRS1) and Les Graous 2 (GRS2) by
blue-grey sandy marls. A bundle of 6 more indurated beds is intercalated in the
lower part of these marls (Figs. 6.B ![]() ,
11.C-D
,
11.C-D ![]() ). Within the Les Graous Member,
pyrite nodules and centimetre-thick pyrite alteration beds are relatively
common. The upper limit of this member can only be observed at the GRS 1 and GRS
2 sections: it corresponds to a discontinuity surface with numerous Thalassinoides
located at the top of a 0.80 m-thick marly limestone bed (GRS 1 and GRS 2 bed
113; Figs. 5
). Within the Les Graous Member,
pyrite nodules and centimetre-thick pyrite alteration beds are relatively
common. The upper limit of this member can only be observed at the GRS 1 and GRS
2 sections: it corresponds to a discontinuity surface with numerous Thalassinoides
located at the top of a 0.80 m-thick marly limestone bed (GRS 1 and GRS 2 bed
113; Figs. 5 ![]() ,
6.D
,
6.D ![]() ).
).
The
Toxoceratoides Bed provided a large number of deformed internal moulds of
ammonites, nautiloids, belemnites (Pl. 3 ![]() , figs.
7-8), bivalves, gastropods,
corals, and fish scales. The ammonite fauna is represented by Ammonitoceras
sp., Lithancylus sp., Toxoceratoides aff. royerianus (Pl. 3
, figs.
7-8), bivalves, gastropods,
corals, and fish scales. The ammonite fauna is represented by Ammonitoceras
sp., Lithancylus sp., Toxoceratoides aff. royerianus (Pl. 3 ![]() , figs.
13-16), Deshayesites grandis (Pl. 3
, figs.
13-16), Deshayesites grandis (Pl. 3 ![]() , figs.
9-12), Cheloniceras
crassum (Pl. 3
, figs.
9-12), Cheloniceras
crassum (Pl. 3 ![]() , figs.
5-6), ?Colombiceras sp., Pseudohaploceras
liptoviensis, ?Aconeceras sp., Macroscaphites striatisulcatus
(Pl. 3
, figs.
5-6), ?Colombiceras sp., Pseudohaploceras
liptoviensis, ?Aconeceras sp., Macroscaphites striatisulcatus
(Pl. 3 ![]() , figs.
3-4), Acantholytoceras sp., Ptychoceras emericianum,
Hypophylloceras sp. (Pl. 3
, figs.
3-4), Acantholytoceras sp., Ptychoceras emericianum,
Hypophylloceras sp. (Pl. 3 ![]() , fig.
1) and Lytoceras sp.
, fig.
1) and Lytoceras sp.
The
Deshayesites in
this level have a well-expressed smooth siphonal band and a subtabulated ventral
area on the phragmocone (Pl. 3 ![]() , figs. 9,
12). The Cheloniceras have a
long Cheloniceras stage and a short or absent Procheloniceras stage (Pl. 3
, figs. 9,
12). The Cheloniceras have a
long Cheloniceras stage and a short or absent Procheloniceras stage (Pl. 3 ![]() , fig.
5). These characters indicate an age corresponding to the D. grandis
Subzone (Fig. 1
, fig.
5). These characters indicate an age corresponding to the D. grandis
Subzone (Fig. 1 ![]() ).
).
Above
the Toxoceratoides Bed, the lower part of the Les Graous Member yielded
an abundant ammonite fauna represented by deformed internal moulds (Fig. 6.C ![]() ).
Within these levels, belemnites (Pl. 5
).
Within these levels, belemnites (Pl. 5 ![]() , figs.
17-21) and fish scales (Pl. 5
, figs.
17-21) and fish scales (Pl. 5 ![]() , fig.
26) are relatively frequent. Nautiloids, echinoids (Pl. 5
, fig.
26) are relatively frequent. Nautiloids, echinoids (Pl. 5 ![]() , figs.
27-28),
brachiopods (Pl. 5
, figs.
27-28),
brachiopods (Pl. 5 ![]() , fig.
29), gastropods (Pl. 5
, fig.
29), gastropods (Pl. 5 ![]() ,
figs. 22-23), bivalves (Pl. 5
,
figs. 22-23), bivalves (Pl. 5 ![]() , fig.
24) and small ahermatypic corals (Pl. 5
, fig.
24) and small ahermatypic corals (Pl. 5 ![]() , fig.
25) are rarer. Fossils are
noticeably more frequent in the more indurated beds. Pyritic internal moulds of
ammonites are extremely rare and poorly preserved. In the lower part of the grey-blue
marls, we identified the following ammonitic taxa: Ammonitoceras lahuseni
macroconchs (Pl. 4
, fig.
25) are rarer. Fossils are
noticeably more frequent in the more indurated beds. Pyritic internal moulds of
ammonites are extremely rare and poorly preserved. In the lower part of the grey-blue
marls, we identified the following ammonitic taxa: Ammonitoceras lahuseni
macroconchs (Pl. 4 ![]() , figs.
1-3) and microconchs, Tropaeum bowerbankii, Toxoceratoides
rochi (Pl. 5
, figs.
1-3) and microconchs, Tropaeum bowerbankii, Toxoceratoides
rochi (Pl. 5 ![]() , figs.
1-4), Dufrenoyia furcata (Pl. 5
, figs.
1-4), Dufrenoyia furcata (Pl. 5 ![]() , figs.
5-9), Cheloniceras
crassum (Pl. 4
, figs.
5-9), Cheloniceras
crassum (Pl. 4 ![]() , figs.
8-9), Colombiceras crassicostatum (Pl. 5
, figs.
8-9), Colombiceras crassicostatum (Pl. 5 ![]() , figs.
10-12), Pseudohaploceras liptoviensis (Pl. 4
, figs.
10-12), Pseudohaploceras liptoviensis (Pl. 4 ![]() , figs.
6-7), Aconeceras
nisum (Pl. 5
, figs.
6-7), Aconeceras
nisum (Pl. 5 ![]() , figs.
13-14), Ptychoceras emericianum (Pl. 5
, figs.
13-14), Ptychoceras emericianum (Pl. 5 ![]() , fig.
15),
Macroscaphites striatisulcatus (Pl. 5
, fig.
15),
Macroscaphites striatisulcatus (Pl. 5 ![]() , fig.
16), Acantholytoceras
sp. (Pl. 4
, fig.
16), Acantholytoceras
sp. (Pl. 4 ![]() , figs.
4-5), Lytoceras
sp. (Pl. 4
, figs.
4-5), Lytoceras
sp. (Pl. 4 ![]() , figs.
10-11), Hypophylloceras sp. (Pl. 4
, figs.
10-11), Hypophylloceras sp. (Pl. 4 ![]() , fig.
12). Nautiles
are represented by Cymatoceras neckerianus
(Pl. 4
, fig.
12). Nautiles
are represented by Cymatoceras neckerianus
(Pl. 4 ![]() , figs.
13-14) and Anglonautilus
sp.
, figs.
13-14) and Anglonautilus
sp.
In
the upper part of the Les Graous Member, we identified Toxoceratoides rochi,
Dufrenoyia dufrenoyi (Pl. 5 ![]() ,
figs. 30-32), Cheloniceras crassum, Pseudohaploceras
liptoviensis, Aconeceras nisum, Hypophylloceras sp.
,
figs. 30-32), Cheloniceras crassum, Pseudohaploceras
liptoviensis, Aconeceras nisum, Hypophylloceras sp.
Belemnite taxa, Neohibolites aptiensis (Pl. 5 ![]() ,
figs. 17-18), Duvalia grasi
(Pl. 5
,
figs. 17-18), Duvalia grasi
(Pl. 5 ![]() ,
figs. 19-20) and Mesohibolites sp. (Pl. 5
,
figs. 19-20) and Mesohibolites sp. (Pl. 5 ![]() , fig.
21), occur
throughout the Les Graous Member.
, fig.
21), occur
throughout the Les Graous Member.
The grey-blue marls of the Les Graous Member are assigned to the D. furcata
Zone (Fig. 1 ![]() ): their lower part is assigned to the D. furcata Subzone and
their upper part to the D. dufrenoyi Subzone.
): their lower part is assigned to the D. furcata Subzone and
their upper part to the D. dufrenoyi Subzone.
|
Figure
14: Log of the Combe de Marin section (CRS) with distribution of ammonite
taxa,
see Figs. 4 |
|
Figure
15: Log of the Pont Pa�r� section (PPR) with distribution of ammonite
taxa,
see Figs. 4 |
|
Figure
16: Field photographies of the Pont de la Cerise section (CLE). Log of this
section is available in Fig. 15 |
Eight
sections were sampled between 2000 and 2017. They are located along an E-W axis
on either side of the village of Les Ferres (Fig. 3 ![]() ). These sections are (from W
to E):
). These sections are (from W
to E):
5.3.1. Les Graous sections (GRS 1 and GRS 2)
These
sections are located about 1.65 km to the WSW of the village of Les Ferres. Les
Graous 1 (GRS 1, Fig. 3 ![]() , lat.:
43�50'31.8"N, long.: 7�4'35.4"E) crops out on the
eastern flank of the Les Graous cuesta and Les Graous 2 (GRS 2, Fig. 3
, lat.:
43�50'31.8"N, long.: 7�4'35.4"E) crops out on the
eastern flank of the Les Graous cuesta and Les Graous 2 (GRS 2, Fig. 3 ![]() , lat.:
43�50'31.0"N, long.: 7�4'27.3"E) on its western flank. Because of their similar
lithostratigraphy, the log of GRS 1 only is figured (Fig. 5
, lat.:
43�50'31.0"N, long.: 7�4'27.3"E) on its western flank. Because of their similar
lithostratigraphy, the log of GRS 1 only is figured (Fig. 5 ![]() ). Field photographs
and log section of GRS 1 were given by Bert & Bersac (2011,
Figs. 30, 32-37, 40-47), but the exact location of this section was not
indicated because fieldwork had not been completed then. In this section, the
marly Aptian can be observed continuously from the top of the Hauterivian-Aptian
limestones to the Albian (Fig. 6.B, .D
). Field photographs
and log section of GRS 1 were given by Bert & Bersac (2011,
Figs. 30, 32-37, 40-47), but the exact location of this section was not
indicated because fieldwork had not been completed then. In this section, the
marly Aptian can be observed continuously from the top of the Hauterivian-Aptian
limestones to the Albian (Fig. 6.B, .D ![]() ). The Combe de Joinet Member is absent
and the Pont de la Cerise Member lies directly on the Hauterivian-Aptian
limestones. The Pont de la Cerise Member is only 0.90 m thick in GRS 1 (Figs. 5
). The Combe de Joinet Member is absent
and the Pont de la Cerise Member lies directly on the Hauterivian-Aptian
limestones. The Pont de la Cerise Member is only 0.90 m thick in GRS 1 (Figs. 5 ![]() ,
6.A
,
6.A ![]() ). A bed of phosphate nodules can be observed at 15 cm above its base. At the
top of this member the Ammonitoceras Level provided some poorly preserved
ammonite fragments. In addition to the ammonite taxa cited in Fig. 5
). A bed of phosphate nodules can be observed at 15 cm above its base. At the
top of this member the Ammonitoceras Level provided some poorly preserved
ammonite fragments. In addition to the ammonite taxa cited in Fig. 5 ![]() , the Pont
de la Cerise Member provided Cymatoceras neckerianus (20 cm above its
base), Anglonautilus sp. (30 cm above its base) and several specimens of Neohibolites
aptiensis.
, the Pont
de la Cerise Member provided Cymatoceras neckerianus (20 cm above its
base), Anglonautilus sp. (30 cm above its base) and several specimens of Neohibolites
aptiensis.
The
Toxoceratoides Bed is present at the base of Les Graous Member (Figs. 5 ![]() ,
6.A
,
6.A ![]() ). It is relatively marly here and yielded a fairly abundant fauna of
ammonites, belemnites and nautiloids. Above the Toxoceratoides Bed, the
first 5 metres of this member are represented by a bundle of 6 sandy and marly
limestone beds separated from each other by marly intervals (Figs. 5
). It is relatively marly here and yielded a fairly abundant fauna of
ammonites, belemnites and nautiloids. Above the Toxoceratoides Bed, the
first 5 metres of this member are represented by a bundle of 6 sandy and marly
limestone beds separated from each other by marly intervals (Figs. 5 ![]() ,
6.B
,
6.B ![]() ). A
rich ammonite fauna assigned to the D. furcata Subzone (Fig. 6.C
). A
rich ammonite fauna assigned to the D. furcata Subzone (Fig. 6.C ![]() ; Pl. 4
; Pl. 4 ![]() , figs. 8-12; Pl. 5
, figs. 8-12; Pl. 5 ![]() , figs.
1-16) was collected mainly from the first 5 sandy and
marly limestone beds and Bed 99 (Fig. 5
, figs.
1-16) was collected mainly from the first 5 sandy and
marly limestone beds and Bed 99 (Fig. 5 ![]() ).
).
The
upper part of the Les Graous Member is attributed to the D. dufrenoyi
Subzone. It is marlier and only the sandy and marly limestone beds yielded
ammonites. Bed 110 yielded the earliest D. dufrenoyi (Fig. 5 ![]() ). The upper
limit of this member is represented by a sandy limestone bed (Bed 113, Fig. 5
). The upper
limit of this member is represented by a sandy limestone bed (Bed 113, Fig. 5 ![]() )
that is relatively rich in ammonites (especially in D. dufrenoyi, Pl. 5
)
that is relatively rich in ammonites (especially in D. dufrenoyi, Pl. 5 ![]() , fig.
32). It is overlain by upper Aptian dark glauconitic marls. All the
material from "les Graoux" published by Delanoy et
al. (2018) was collected from Beds 99, 100 and 102 of the GRS 1 section in
our presence.
, fig.
32). It is overlain by upper Aptian dark glauconitic marls. All the
material from "les Graoux" published by Delanoy et
al. (2018) was collected from Beds 99, 100 and 102 of the GRS 1 section in
our presence.
5.3.2. La Valli�re section (VAL)
Ragazzi
(1982, p. 73) and Br�heret (1997, p. 267, Figs. 114-115) studied a
section located in the La Valli�re area (Fig. 3 ![]() , lat.:
43�50'32.6"N, long.: 7�5'29.9"E). This section is nowadays almost completely covered by vegetation
and located in a private fenced property.
, lat.:
43�50'32.6"N, long.: 7�5'29.9"E). This section is nowadays almost completely covered by vegetation
and located in a private fenced property.
We
studied an outcrop near this section (VAL section, Figs. 3 ![]() ,
7
,
7 ![]() , lat.:
43�50'32.1"N, long.: 7�5'48.4"E). It is located in a half-graben, against a
vertical decametric palaeo-escarpment, about 0.6 km SSW of the village of Les
Ferres (Fig. 8.A
, lat.:
43�50'32.1"N, long.: 7�5'48.4"E). It is located in a half-graben, against a
vertical decametric palaeo-escarpment, about 0.6 km SSW of the village of Les
Ferres (Fig. 8.A ![]() ). A cross-section of this tectonic structure was figured by Dardeau
& Graciansky (1987, Figs. 1-2). In this outcrop, the Pont de la
Cerise Member reaches its maximum thickness (15 m) and is overlain by a 0.2
m-thick sandstone level (Bed 417, Fig. 7
). A cross-section of this tectonic structure was figured by Dardeau
& Graciansky (1987, Figs. 1-2). In this outcrop, the Pont de la
Cerise Member reaches its maximum thickness (15 m) and is overlain by a 0.2
m-thick sandstone level (Bed 417, Fig. 7 ![]() ), which is the probable local
equivalent of the Toxoceratoides Bed (Fig. 8.A
), which is the probable local
equivalent of the Toxoceratoides Bed (Fig. 8.A ![]() ). Bed 405, located at 3.70
m from the base of the Pont de la Cerise Member (Figs. 7
). Bed 405, located at 3.70
m from the base of the Pont de la Cerise Member (Figs. 7 ![]() ,
8.B
,
8.B ![]() ), yielded a
relatively abundant, but fragmentary, ammonite fauna very similar to that of the
Ammonitoceras Level. A level located at the top of the Pont de la Cerise
Member (top of Level 416, Fig. 7
), yielded a
relatively abundant, but fragmentary, ammonite fauna very similar to that of the
Ammonitoceras Level. A level located at the top of the Pont de la Cerise
Member (top of Level 416, Fig. 7 ![]() ) yielded rare representatives of Toxoceratoides,
Deshayesites and Cheloniceras,
but they are too fragmentary to be identified to specific level. Nevertheless,
the lithological and palaeontological characteristics of this last bed are not
equivalent to the Ammonitoceras Level. For this reason, we consider Bed
405 as a local equivalent of the Ammonitoceras Level. This all implies
that the glauconitic sandstones overlying the Ammonitoceras Level of VAL
are therefore considerably more developed here than in other sections (Fig. 17
) yielded rare representatives of Toxoceratoides,
Deshayesites and Cheloniceras,
but they are too fragmentary to be identified to specific level. Nevertheless,
the lithological and palaeontological characteristics of this last bed are not
equivalent to the Ammonitoceras Level. For this reason, we consider Bed
405 as a local equivalent of the Ammonitoceras Level. This all implies
that the glauconitic sandstones overlying the Ammonitoceras Level of VAL
are therefore considerably more developed here than in other sections (Fig. 17 ![]() ).
The Les Graous Member of VAL is almost completely covered by vegetation and thus
could not be studied.
).
The Les Graous Member of VAL is almost completely covered by vegetation and thus
could not be studied.
The
La Graou section (GRO, lat.: 43�50'36.2"N, long.: 7�6'0.0"E) was described by Ragazzi
(1982, p. 73), then by Br�heret (1997, p. 264, Fig. 114). It is located
0.5 km SW to the village of Les Ferres (Figs. 3 ![]() ,
9
,
9 ![]() ). The Lower Aptian is
nowadays almost totally covered by vegetation and only a small part of the Les
Graous Member can be observed.
). The Lower Aptian is
nowadays almost totally covered by vegetation and only a small part of the Les
Graous Member can be observed.
5.3.4. The Combe de Joinet section (CHP)
The
CHP section (lat.: 43�50'42.7"N, long.: 7�6'9.5"E) is located 0.8 km ESE of
the village of Les Ferres, along a track leading to the Pont de la Cerise Figs. 3 ![]() ,
10
,
10 ![]() - 11
- 11 ![]() ). Bert (2014, p. 393) figured the Ammonitoceras Level at
the CHP section for the first time (Fig. 11.D
). Bert (2014, p. 393) figured the Ammonitoceras Level at
the CHP section for the first time (Fig. 11.D ![]() ). Delanoy et al.
(2018, Fig. 2) published a log of this section and described and figured many
ammonites, mainly from the Ammonitoceras Level (Bed 107 in Delanoy
et al., 2018 = Bed 415 in the present work,
Figs. 10
). Delanoy et al.
(2018, Fig. 2) published a log of this section and described and figured many
ammonites, mainly from the Ammonitoceras Level (Bed 107 in Delanoy
et al., 2018 = Bed 415 in the present work,
Figs. 10 ![]() ,
11.C-D
,
11.C-D ![]() ).
Unfortunately, several levels located at the base of the marly Aptian were not
figured in the log of Delanoy et al. (2018, Fig. 2), including the
Combe de Joinet Member. For this reason, we did not use the bed numbering system
of this latter work for our own log (Fig. 10
).
Unfortunately, several levels located at the base of the marly Aptian were not
figured in the log of Delanoy et al. (2018, Fig. 2), including the
Combe de Joinet Member. For this reason, we did not use the bed numbering system
of this latter work for our own log (Fig. 10 ![]() ). At CHP, the Combe de Joinet
Member (Bed 401) crops out in good conditions along the track between the D1
road and the Pont de la Cerise (Figs. 3
). At CHP, the Combe de Joinet
Member (Bed 401) crops out in good conditions along the track between the D1
road and the Pont de la Cerise (Figs. 3 ![]() ,
11.A
,
11.A ![]() ). It is represented by massive
sandy and marly limestones and provided some rare fragments of ammonite. The Ammonitoceras
Level crops out under good conditions, allowing the collecting of abundant
ammonites (Delanoy et al., 2018; Fig. 11.C-D
). It is represented by massive
sandy and marly limestones and provided some rare fragments of ammonite. The Ammonitoceras
Level crops out under good conditions, allowing the collecting of abundant
ammonites (Delanoy et al., 2018; Fig. 11.C-D ![]() ; Pl. 2
; Pl. 2 ![]() , figs.
1-19). Only the lower part of the Les Graous Member is visible, its upper part
being covered with dense vegetation (Fig. 11.B
, figs.
1-19). Only the lower part of the Les Graous Member is visible, its upper part
being covered with dense vegetation (Fig. 11.B ![]() ).
).
5.3.5. The Pont de la Cerise section (CLE)
This
section (lat.: 43�50'42.7"N, long.: 7�6'9.5"E) was first published by Ragazzi
(1982, p. 75) who named it "Pont de la Cerise" despite the fact
that another section closer to the Pont de la Cerise locality was named "Combe
de Marin" by Ragazzi (1982, p. 77 and Fig. 3; Fig. 3 ![]() and see
Chapter 5.3.6). The CLE section crops out on the right bank of the Esteron River
in the locality of the Clot d'Est�ron, 1.95 km east of the village of Les
Ferres (Figs. 3
and see
Chapter 5.3.6). The CLE section crops out on the right bank of the Esteron River
in the locality of the Clot d'Est�ron, 1.95 km east of the village of Les
Ferres (Figs. 3 ![]() ,
12
,
12 ![]() ). The top of the Hauterivian-Aptian limestones, the Combe
de Joinet and Pont de la Cerise members, as well as the Toxoceratoides
Bed crop out under favourable conditions (Fig. 13.A
). The top of the Hauterivian-Aptian limestones, the Combe
de Joinet and Pont de la Cerise members, as well as the Toxoceratoides
Bed crop out under favourable conditions (Fig. 13.A ![]() ). As Ragazzi noticed
(1982, p. 76), the Combe de Joinet Member (Beds 401 to 404 in CHP) is
represented by 1.1 m of marly limestone and two decimetric limestone beds (Figs. 12
). As Ragazzi noticed
(1982, p. 76), the Combe de Joinet Member (Beds 401 to 404 in CHP) is
represented by 1.1 m of marly limestone and two decimetric limestone beds (Figs. 12 ![]() , 13.A
, 13.A ![]() ). The top of this member is intensely bioturbated
(Fig. 13.B
). The top of this member is intensely bioturbated
(Fig. 13.B ![]() ). The
Pont de la Cerise Member crops out in very good conditions and is 5.5 m thick at
that point (Fig. 13.A, .C-D
). The
Pont de la Cerise Member crops out in very good conditions and is 5.5 m thick at
that point (Fig. 13.A, .C-D ![]() ). In its lower and middle parts, rare and
poorly preserved fragments of Ancyloceratidae gen. ind., Deshayesites
sp., Cheloniceras sp., and Pseudohaploceras liptoviensis (Fig. 13.D
). In its lower and middle parts, rare and
poorly preserved fragments of Ancyloceratidae gen. ind., Deshayesites
sp., Cheloniceras sp., and Pseudohaploceras liptoviensis (Fig. 13.D ![]() ) were recorded. The Ammonitoceras Level (Bed 415,
Fig. 12
) were recorded. The Ammonitoceras Level (Bed 415,
Fig. 12 ![]() ) occurs at
the top of this member and its ammonite fauna appears to be slightly less
abundant and less well-preserved than that at CHP (Pl. 2
) occurs at
the top of this member and its ammonite fauna appears to be slightly less
abundant and less well-preserved than that at CHP (Pl. 2 ![]() , fig.
20). Only the Toxoceratoides
Bed (Bed 416, Fig. 12
, fig.
20). Only the Toxoceratoides
Bed (Bed 416, Fig. 12 ![]() ) of the Les Graous Member crops out under good conditions,
the rest of the formation being entirely covered by vegetation.
) of the Les Graous Member crops out under good conditions,
the rest of the formation being entirely covered by vegetation.
5.3.6. The Combe de Marin section (CRS)
This
section was figured by Ragazzi (1982, p. 77 and Fig. 3). It is located on
the left bank of the Esteron River, opposite to the Pont de la Cerise locality,
2.69 km east of the village of Les Ferres (Fig. 3 ![]() ). The CRS section crops out at
the top of a cliff mainly formed by the Hauterivian-Aptian limestones (Fig. 14
). The CRS section crops out at
the top of a cliff mainly formed by the Hauterivian-Aptian limestones (Fig. 14 ![]() ).
Our own observations differ slightly from those of Ragazzi (1982): only
the Combe de Joinet Member and the lower 5.4 m of the Pont de la Cerise Member
are currently observable. The rest of the Aptian marls is covered by vegetation.
The Combe de Joinet Member is represented by 0.80 m of marly limestones, with a
dark brown clay level, 0.10 m thick, in its lower part. The ammonite fauna is
rare and fragmentary, but includes: Ancyloceras sp., Deshayesites
sp., Pseudohaploceras liptoviensis and Macroscaphites aff. yvani.
).
Our own observations differ slightly from those of Ragazzi (1982): only
the Combe de Joinet Member and the lower 5.4 m of the Pont de la Cerise Member
are currently observable. The rest of the Aptian marls is covered by vegetation.
The Combe de Joinet Member is represented by 0.80 m of marly limestones, with a
dark brown clay level, 0.10 m thick, in its lower part. The ammonite fauna is
rare and fragmentary, but includes: Ancyloceras sp., Deshayesites
sp., Pseudohaploceras liptoviensis and Macroscaphites aff. yvani.
5.3.7. The Pont Pa�r� section (PPR)
Ragazzi
(1982) already mentioned this section ("ravin de Pont de Payre"
in Ragazzi, 1982, p. 77) where she observed that the Pont de la Cerise
Member was reduced to a thickness of 3.5 metres. This section (lat.:
43�51'11.6"N,
long.: 7�7'49.0"E, Fig. 15 ![]() ) crops out in a ravine perpendicular to the track
between the D17 road and Pont de la Cerise (Fig. 3
) crops out in a ravine perpendicular to the track
between the D17 road and Pont de la Cerise (Fig. 3 ![]() ). Our own observations differ
from Ragazzi�s (1982): the Pont de la Cerise Member is separated from
the Hauterivian-Aptian limestones by a metre-thick covered section and is 11
m-thick (Figs. 15
). Our own observations differ
from Ragazzi�s (1982): the Pont de la Cerise Member is separated from
the Hauterivian-Aptian limestones by a metre-thick covered section and is 11
m-thick (Figs. 15 ![]() ,
16.A-B
,
16.A-B ![]() ). This member can be observed under good conditions
along the ravine, for a distance of about 100 m. The Ammonitoceras Level
(top of Bed 414 in PPR, see Fig. 15
). This member can be observed under good conditions
along the ravine, for a distance of about 100 m. The Ammonitoceras Level
(top of Bed 414 in PPR, see Fig. 15 ![]() ) is present at its top
(Fig. 16.B
) is present at its top
(Fig. 16.B ![]() ),
immediately above a level particularly rich in large pyrite nodules. The levels
of the Pont de la Cerise Member below the Ammonitoceras Level reach their
maximum thickness in this section. Ammonites are very rare in the Pont de la
Cerise Member in this section, except in the Ammonitoceras Level. Only
the first 3.3 m of the Les Graous Member are observable (Fig. 16.B
),
immediately above a level particularly rich in large pyrite nodules. The levels
of the Pont de la Cerise Member below the Ammonitoceras Level reach their
maximum thickness in this section. Ammonites are very rare in the Pont de la
Cerise Member in this section, except in the Ammonitoceras Level. Only
the first 3.3 m of the Les Graous Member are observable (Fig. 16.B ![]() ); they are
richer in glaucony than in the other sections. The Toxoceratoides Bed is
marlier than in the CHP and CLE sections, and yielded some ammonites (Figs. 15
); they are
richer in glaucony than in the other sections. The Toxoceratoides Bed is
marlier than in the CHP and CLE sections, and yielded some ammonites (Figs. 15 ![]() ,
16.B
,
16.B ![]() ).
).
|
Figure
17:
Lithostratigraphic correlation between the studied sections and the adjacent
sections. |
The
Combe de Joinet Member appears at VAL, where it reaches its maximum thickness
(taking into account that this member could not be measured at PPR). It gets
thinner towards the east (Fig. 17 ![]() ). The Pont de la Cerise Member appears at GRS 1
and GRS 2 with a thickness of only 0.9 m. The interval of this member beneath
the Ammonitoceras Level gradually thicken towards the east (taking into
account that we have no thickness data at CRS, Fig. 17
). The Pont de la Cerise Member appears at GRS 1
and GRS 2 with a thickness of only 0.9 m. The interval of this member beneath
the Ammonitoceras Level gradually thicken towards the east (taking into
account that we have no thickness data at CRS, Fig. 17 ![]() ) reaching their maximum
thickness at PPR. The interval of the Pont de la Cerise Member above the Ammonitoceras
Level are generally a few centimeters-thick, except at VAL where they
exceptionally reach a thickness of 11 m (Figs. 7
) reaching their maximum
thickness at PPR. The interval of the Pont de la Cerise Member above the Ammonitoceras
Level are generally a few centimeters-thick, except at VAL where they
exceptionally reach a thickness of 11 m (Figs. 7 ![]() ,
8.A
,
8.A ![]() ,
17
,
17 ![]() ). This may be due to
the palaeogeographic context of VAL: the presence of a decametric fault palaeo-escarpment
in this area probably allowed an accumulation of a large amount of sediment
above the Ammonitoceras Level (Dardeau & Graciansky,
1987). Our observations differ from those of Ragazzi (1982) who observed
a thickness reduction of the Pont de la Cerise Member ("accumulation
glauconitique" in Ragazzi, 1982, p. 78) at CRS and PPR
(Fig. 17
). This may be due to
the palaeogeographic context of VAL: the presence of a decametric fault palaeo-escarpment
in this area probably allowed an accumulation of a large amount of sediment
above the Ammonitoceras Level (Dardeau & Graciansky,
1987). Our observations differ from those of Ragazzi (1982) who observed
a thickness reduction of the Pont de la Cerise Member ("accumulation
glauconitique" in Ragazzi, 1982, p. 78) at CRS and PPR
(Fig. 17 ![]() )
compared to more westerly sections. The thickness variation of the Les Graous
Member could not be studied because this latter is completely visible only at
GRS 1 and GRS 2.
)
compared to more westerly sections. The thickness variation of the Les Graous
Member could not be studied because this latter is completely visible only at
GRS 1 and GRS 2.
The
nearest outcrop west of GRS 2 (Fig. 3 ![]() ) confirms the westward thickness reduction
of the Pont de la Cerise Member, as already observed by Ragazzi (1982).
This section, named "Cons�gudes" by Ragazzi (1982,
Fig.
3), crops out along the D1 road next to the village of Cons�gudes (lat.:
43�50'26.5"N, long.: 7�2'58.3"E, Fig. 3
) confirms the westward thickness reduction
of the Pont de la Cerise Member, as already observed by Ragazzi (1982).
This section, named "Cons�gudes" by Ragazzi (1982,
Fig.
3), crops out along the D1 road next to the village of Cons�gudes (lat.:
43�50'26.5"N, long.: 7�2'58.3"E, Fig. 3 ![]() ): the "Early Aptian
unconformity" (Cotillon, 2010, p. 4) is located at the top of
the upper Barremian limestones and is directly covered by Albian dark marlstones
(Ragazzi, 1982, p. 73 and Pl. 3; Fig. 17
): the "Early Aptian
unconformity" (Cotillon, 2010, p. 4) is located at the top of
the upper Barremian limestones and is directly covered by Albian dark marlstones
(Ragazzi, 1982, p. 73 and Pl. 3; Fig. 17 ![]() ). The Aptian is therefore
totally absent in this section.
). The Aptian is therefore
totally absent in this section.
East
of the study area, the Aptian is also totally missing in the Colle-Belle section
(Cotillon, 1971, p. 147, lat.: 43�51'10.4"N, long.:
7�8'32.3"E,
Fig. 17 ![]() ), with Albian marlstones lying directly on Barremian limestones (Cotillon,
1971, p. 132 and 147; Fig. 17
), with Albian marlstones lying directly on Barremian limestones (Cotillon,
1971, p. 132 and 147; Fig. 17 ![]() ). The sudden demise of the marly Aptian between
the PPR and the Colle-Belle section is probably related to the presence of a
decametric synsedimentary fault forming the eastern limit of the LFAB.
). The sudden demise of the marly Aptian between
the PPR and the Colle-Belle section is probably related to the presence of a
decametric synsedimentary fault forming the eastern limit of the LFAB.
North of the study area, Aptian deposits can be observed only on the northern flanks of the Roccaforte and Vial mountains and near the village of Tournefort, where they are represented by plurimetric dark marlstones with pyritized ammonites characteristic of the pelagic domain (Ragazzi, 1982, p. 61 and personal observations).
To the south, the Aptian is missing or represented in the Coursegoules and Ruth Mountain areas by thin reworked layers (Cotillon, 1971, p. 145; Bersac et al., 2010; Bert & Bersac, 2011, Figs. 4-6, 20, 67-70; Bert et al., 2013, p. 361, Fig. 9).
The faunal spectrum study of the ectocochleate cephalopods of the LFAB is based on 1,262 ammonites and 58 nautiloids collected or studied in situ, and by 29 ammonites and 1 nautiloid published in Delanoy et al. (2018).
The
most represented ammonites families in the lower Aptian of the LFAB are
Ancyloceratidae, Deshayesitidae, Douvilleiceratidae, and Desmoceratidae (Fig. 18 ![]() ). This faunal spectrum is consistent with the palaeogeography of the LFAB
during the early Aptian times, located in the outer neritic domain.
Ancyloceratidae and Nautiloidea (Ward, 1987; Hewitt, 1989; Westermann,
1990, 1996) have a preferential tropism for the neritic domain, Deshayesitidae
are open oceanic surface organisms, Douvilleiceratidae are opportunistic (Bulot
et al., 2005; Dutour, 2005) and Desmoceratidae, Phylloceratidae
and Lytoceratidae are deeper water environmental representatives (Westermann,
1990, 1996; Bulot et al., 2005).
). This faunal spectrum is consistent with the palaeogeography of the LFAB
during the early Aptian times, located in the outer neritic domain.
Ancyloceratidae and Nautiloidea (Ward, 1987; Hewitt, 1989; Westermann,
1990, 1996) have a preferential tropism for the neritic domain, Deshayesitidae
are open oceanic surface organisms, Douvilleiceratidae are opportunistic (Bulot
et al., 2005; Dutour, 2005) and Desmoceratidae, Phylloceratidae
and Lytoceratidae are deeper water environmental representatives (Westermann,
1990, 1996; Bulot et al., 2005).
The most spectacular character of the lower Aptian ammonite fauna of the LFAB is the abundance of the heteromorphic genus Ammonitoceras in the D. deshayesi and D. furcata Zones, with 80.2% of the Ancyloceratidae of the D. deshayesi Zone and 91.9% for the D. furcata Zone respectively. Such an abundance has never been reported previously in the literature (Kakabadze, 1981; Kakabadze & Hoedemaeker, 2004). It may be related to particularly favourable environmental conditions in the LFAB.
The
scarcity of representatives of the genus Tropaeum in the LFAB may be
related to the fact that this is a boreal taxon (Casey, 1960; Aguirre-Urreta,
1985). Nautiloidea never represent more than 13.5% of the ectocochleate
cephalopod fauna (Fig. 18 ![]() ).
).
The
proportion of each group of ectocochleate cephalopods varies over the
stratigraphic interval considered: excluding data from the Combe de Joinet
Member and the Pont de la Cerise Member below the Ammonitoceras Level due
to the small amount of material collected (respectively 10 and 15 specimens;
Fig. 18 ![]() ), there is a decrease in the proportion of Nautiloidea and
Ancyloceratidae over time and an increase of Deshayesidae and Desmoceratidae
(the latter being related to the "bloom" of the genus Aconeceras
Hyatt, 1903, in the D. furcata Zone). This is strongly consistent
with the transgressive trend of the studied series (see Chapter 2), where open
marine environment taxa become progressively more prevalent. At the top of the
studied series, in the D. dufrenoyi Subzone, this phenomenon leads to a
faunal spectrum similar to that observed in hemipelagic environments, where the
ammonite populations are mainly composed of Deshayesitidae and Desmoceratidae (Dutour,
2005, Fig. 22). The proportion of Helicancylidae is higher when Ancyloceratidae
decrease, as a result of ecological interactions between these two families.
), there is a decrease in the proportion of Nautiloidea and
Ancyloceratidae over time and an increase of Deshayesidae and Desmoceratidae
(the latter being related to the "bloom" of the genus Aconeceras
Hyatt, 1903, in the D. furcata Zone). This is strongly consistent
with the transgressive trend of the studied series (see Chapter 2), where open
marine environment taxa become progressively more prevalent. At the top of the
studied series, in the D. dufrenoyi Subzone, this phenomenon leads to a
faunal spectrum similar to that observed in hemipelagic environments, where the
ammonite populations are mainly composed of Deshayesitidae and Desmoceratidae (Dutour,
2005, Fig. 22). The proportion of Helicancylidae is higher when Ancyloceratidae
decrease, as a result of ecological interactions between these two families.
|
Figure
18:
Nautiloidea and Ammonoidea faunal spectrum of the lower Aptian of les Ferres
area. HAL: Hauterivian-Aptian limestones, n: number of specimens studied, CJ:
Combe de Joinet Member, PC: Pont de la Cerise Member, AL: Ammonitoceras
Level, TB: Toxoceratoides Bed, LG: Les Graous Member. |
The marly deposits of the Les Ferres Aptian Basin (LFAB) are located in one of the few areas of the southern margin of the Vocontian Basin where ammonites from the D. deshayesi � D furcata Zones can be studied under favourable conditions. An abundant bed-by-bed collection of ammonites, constrained at the subzone level and represented by a significant proportion of adult or subadult specimens was collected. The lithostratigraphy and biostratigraphy of the marly Aptian of Les Ferres presented in the present article aims to serve as a basis for our future palaeontological work in this area. The whole marly Aptian of the LFAB is represented by a newly described lithological unit: the Les Graous Formation. This unit is divided into 3 members (from bottom to top): 1) the Combe de Joinet, 2) the Pont de la Cerise and 3) the Les Graous Members. Most of the collected ammonites come from a precise level at the top of the Pont de la Cerise Member, the Ammonitoceras Level (upper D. multicostatus Subzone, D. deshayesi Zone), and from the lower part of the Les Graous Member (D. grandis and D. furcata Subzones). One of the most remarkable features of this fauna is the abundance of the heteromorphic ammonite genus Ammonitoceras (Ancyloceratidae). The abundant collected material allows us to formulate hypotheses on the intraspecific variability and the evolutionary modalities of several taxons of the families Ancyloceratidae, Douvilleiceratidae, Deshayesitidae, Parahoplitidae, and Desmoceratidae, which will be the topic of forthcoming contributions.
We warmly thank Pierre Ropolo for providing data about the lower Aptian Ancyloceras of the Cassis-La B�doule area. The constructive comments of the Editor-in-chief and reviewers Ottilia Szives (Budapest, Hungary) and Yves Dutour (Aix-en-Provence, France) are highly appreciated and greatly improved the manuscript. We are grateful to Simon Mitchell for the improvement of the English manuscript.
Aguirre-Urreta M.B. (1985).- Ancyloceratidos (Ammonoidea) Aptianos de la Cordillera Patagonica Austral, Provincia de Santa Cruz, Argentina.- Boletin de la Academia Nacional de Ciencias, Cordoba, vol. 56, no. 3-4, p. 135-257.
Baraboshkin E.J. & Mikhailova I.A. (2002).- New stratigraphic scheme of the lower Aptian in the Volga River middle courses.- Stratigraphy and Geological Correlation, Moscow, vol. 10, no. 6, p. 603-626 (English translation).
Bersac S. & Bert D. (2012).- Ontogenesis, variability and evolution of the Lower Greensand Deshayesitidae (Ammonoidea, Lower Cretaceous, Southern England): reinterpretation of literature data; taxonomic and biostratigraphic implications.- Annales du Mus�um d'Histoire Naturelle de Nice, vol. XXVII, p. 197-270.
Bersac S. & Bert D. (2015).- Two ammonite species under the same name: revision of Deshayesites deshayesi (d'Orbigny, 1841) based on topotype material (Lower Aptian, Lower Cretaceous, Northeast of France).- Annales de Pal�ontologie, Paris, vol. 101, no. 4, p. 265-294.
Bersac S. & Bert D. (2018).- Revision of the lower Aptian (Lower Cretaceous) ammonite species Cheloniceras cornuelianum (d'Orbigny, 1841).- Annales de Pal�ontologie, Paris, vol. 104, no. 1, p. 45-70.
Bersac S., Bert D. & Delanoy G. (2010).- Description, biostratigraphie et interpr�tation des s�ries condens�es du Cr�tac� inf�rieur de la Montagne de Ruth, � l'extr�mit� sud orientale de l'Arc de Castellane (Alpes-Maritimes, S-E. de la France).- Annales du Mus�um d'Histoire Naturelle de Nice, vol. XXV, p. 1-17.
Bersac S., Bert D. & Matrion B. (2012).- Revision of the index-species Deshayesites deshayesi (Ammonoidea, Lower Aptian, Lower Cretaceous): Taxonomic and biostratigraphic consequences. In: Bert D. & Bersac S. (eds.), First meeting of the Research Group for Paleobiology and Biostratigraphy of the Ammonites.- Boletin del Instituto de Fisiografia y Geologia, Rosario, vol. 82, p. 31-33.
Bert D. (2014, unpublished).- L'influence de la variabilit� intrasp�cifique sur la taxinomie, la biostratigraphie et l'�volution des ammonites: une approche pal�obiologique. Exemples pris dans le Jurassique sup�rieur et le Cr�tac� inf�rieur.- PhD thesis, Universit� de Rennes 1, 523 p.
Bert D. & Bersac S. (2011).- Buts et m�thodes du "lev� de coupe".- Fossiles, Saint-Julien-du-Pinet, no. 6, p. 49-61.
Bert D., Bersac S., Delanoy G. & Canut L. (2013).- Palaeontology, taxonomic revision and variability of some species of the genus Gassendiceras Bert et al., 2006 (Ammonitina, Upper Barremian) from southeastern France.- Acta Geologica Polonica, Warsaw, vol. 63, no. 3, p. 355-397.
Br�heret J.-G. (1997).- L'Aptien et l'Albien de la Fosse vocontienne (des bordures au bassin) : Évolution de la s�dimentation et enseignement sur les �v�nements anoxiques.- Bulletin de la Soci�t� g�ologique du Nord, Lille, vol. 25, 614 p.
Bulot L.G. (2010).- Appendix: Systematic paleontology of Aptian and Albian ammonites from southwest Iran. In: Vincent B., van Buchem F.S.P., Bulot L.G., Immenhauser A., Caron M., Baghbani D. & Huc A.Y. (eds.), Carbon-isotope stratigraphy, biostratigraphy and organic matter distribution in the Aptian-Lower Albian successions of southwest Iran (Dariyan and Kazhdumi formations).- GeoArabia Special Publication, Bahrain, vol. 4, no. 1, p. 139-197.
Bulot L.G., Beaudoin B. & Dauphin L. (2005).- Modification de la structure des populations d'ammonites dans l'Aptien inf�rieur sommital et l'Aptien sup�rieur du domaine vocontien (SE France). In: Grosheny D. (ed.), Impact environnemental sur la dynamique des populations au Cr�tac�. Volume des R�sum�s.- R�unions th�matiques du Groupe Fran�ais du Cr�tac�, Paris, p. 15-17.
Bulot L.G., Frau C. & Pictet A. (2017).- Revision of Toxoceratoides royeri (d'Orbigny, 1842) and its bearing on the systematics of the Aptian Acrioceratidae Vermeulen, 2004 (Ammonoidea, Ancyloceratina, Ancyloceratoidea).- Cretaceous Research, vol. 88, p. 187-196.
Casey R. (1960).- A monograph of the Ammonoidea of the Lower Greensand, part 1.- Palaeontographical Society, London (1959), i-xxxxvi + p. 1-44.
Casey R., Bayliss H.M. & Simpson M.I. (1998).- Observations on the lithostratigraphy and ammonite succession of the Aptian (Lower Cretaceous) Lower Greensand of Chale Bay, Isle of Wight, UK.- Cretaceous Research, vol. 19, p. 511-535.
Conte G. (1999).- Él�ments de la faune de l'Aptien inferieur (B�doulien) de Serviers-la-Baume (Gard).- Bulletin de la Soci�t� d'Étude des Sciences Naturelles de N�mes et du Gard, no. 62, p. 11-15.
Cotillon P. (1971).- Le Cr�tac� inf�rieur de l'arc subalpin de Castellane entre l'Asse et le Var; stratigraphie et s�dimentologie.- M�moires du B.R.G.M., Orl�ans, no. 68, 313 p.
Cotillon P. (2010).- Sea bottom current activity recorded on the southern margin of the Vocontian basin (southeastern France) during the lower Aptian. Evidence for a climatic signal.- Bulletin de la Soci�t� g�ologique de France, Paris, t. 181, no. 1, p. 3-18.
Dardeau G. (1987).- Inversion du style tectonique et permanence des unit�s structurales dans l'histoire m�sozo�que et alpine du basin des Alpes-Maritimes, partie de l'ancienne marge passive de la T�thys.- Comptes Rendus de l'Acad�mie des Sciences (S�rie II), Paris, t. 305, p. 483-486.
Dardeau G. & Graciansky P.-C. de (1987).- Indices d'une tectonique syns�dimentaire distensive d'�ge cr�tac� inf�rieur dans la basse vall�e de l'Est�ron (Alpes-Maritimes) et cons�quences g�odynamiques.- Bulletin de la Soci�t� g�ologique de France (8e S�rie), Paris, t. III, no. 6, p. 1207-1210.
Dauphin L. (2002, unpublished).- Litho-, bio- et chronostratigraphie compar�es dans le bassin vocontien � l'Aptien.- PhD thesis, Universit� de Lille 1, Villeneuve-d'Ascq, 452 p.
Delanoy G. (1998).- Biostratigraphie des faunes d'Ammonites � la limite Barr�mien-Aptien dans la r�gion d'Angles-Barr�me-Castellane. �tude particuli�re de la famille des Heteroceratina Spath, 1922 (Ancyloceratina, Ammonoidea).- Annales du Mus�um d'Histoire Naturelle de Nice, Nice, vol. XII, 270 p.
Delanoy G., Baudouin C., Gonnet R. & Bert D. (2008).- Sur les faunes d'ammonites (Cr�tac� inf�rieur) du niveau glauconieux de la carri�re des Trois-Vernes, pr�s de Crest (Dr�me, Sud-Est de la France).- Annales du Mus�um d'Histoire Naturelle de Nice, Nice, vol. XXIII, p. 11-65.
Delanoy G., Baudouin C., Pictet A., Delattre P. & P�nag� E. (2018).- Ancyloceratidae (Ammonoidea, Ancyloceratoidea) nouveaux ou peu connus de l'Aptien inf�rieur des Alpes-Maritimes (Sud-Est de la France).- Annales du Mus�um d'Histoire Naturelle de Nice, Nice, vol. XXXII, p. 35-90.
Dumas E. (1876).- Statistique g�ologique, min�ralogique, m�tallurgique et pal�ontologique du D�partement du Gard. Deuxi�me partie, constitution g�ologique.- Imprimerie Bertrand, Paris, 695 p.
Dutour Y. (2005, unpublished).- Biostratigraphie, �volution et renouvellements des ammonites de l'Aptien sup�rieur (Gargasien) du bassin vocontien (Sud-Est de la France).- PhD thesis, Universit� Claude-Bernard - Lyon 1, 302 p.
Fallot J.E. (1885).- Étude g�ologique sur les �tages moyens et sup�rieurs du terrain cr�tac� dans le Sud-Est de la France.- Masson Éd., Paris, 268 p.
Frau C., Bulot L.G. & Delanoy G. (2017).- New and poorly known Aptian Acrioceratidae (Acrioceratidae, Ammonoidea) from Cassis-Roquefort-la-B�doule (Bouches-du-Rh�ne, France).- Neues Jahrbuch f�r Geologie und Pal�ontologie Abhandlungen, Stuttgart, vol. 283, no. 3, p. 335-346.
Gauthier H., Busnardo R., Combemorel R., Delanoy G., Fischer J.-C., Guerin-Franiatte F., Joly B., Kennedy W.J., Sornay J. & Tintant H. (2006).- C�phalopodes Cr�tac�s, Volume IV. In: Fischer J.C. (ed.), R�vision critique de la Pal�ontologie Fran�aise d'Alcide d'Orbigny.- Backhuys Publisher, Kerkwerve, 292 p.
Goguel J. (1944).- Contribution � l'�tude pal�og�ographique du Cr�tac� inf�rieur dans le Sud-Est de la France.- Bulletin du Service de la Carte g�ologique de France, Paris, vol. 215, p. 1-62.
Ginsburg L., Montenat C., Baudron J.-C. & Alziar G. (1980).- Carte g�ologique de la France � 1/50 000. Feuille de Roquest�ron.- Bureau des Recherches G�ologiques et Mini�res, Orl�ans, p. XXXVI-42.
Hewitt R.A. (1989).- Outline of research on the ecology and evolution of the Eocene nautilid cephalopods from the London Clay, England.- Tertiary Research, London, vol. 10, p. 65-81.
Jacob C. (1907).- �tudes pal�ontologiques et stratigraphiques sur la partie moyenne des terrains cr�tac�s dans les Alpes fran�aises et les r�gions voisines.- Travaux du laboratoire de g�ologie de la Facult� des Sciences de Grenoble, vol. VIII, p. 280-590.
Joly B. & Delamette M. (2008).- Les Phylloceratoidea (Ammonoidea) aptiens et albiens du bassin vocontien (Sud-Est de la France).- Carnets Geol., Madrid, vol. 8, no. M04 (CG2008_M04), 60 p.
Kakabadze M.V. (1981).- Ancyloceratids of the south of the USSR and their stratigraphic significance.- Trudy geologicheskogo Instituta Akademii Nauk GSSR (new series), vol. 71, 195 p. (in Russian).
Kakabadze M.V. & Hoedemaeker P.J. (2004).- Heteromorphic ammonites from the Barremian and Aptian strata of Colombia.- Scripta Geologica, vol. 128, p. 39-182.
Klein J., Busnardo R., Company M., Delanoy G., Kakabadze M., Reboulet S., Ropolo P., Va��ček Z. & Vermeulen J. (2007).- Lower Cretaceous Ammonites III. Bochianitoidea, Protancyloceratoidea, Ancyloceratoidea, Ptychoceratoidea. In: Riegraf W. (ed.), Fossilium Catalogus I: Animalia.- Backhuys Publishers, Kerkwerve, 381 p.
Lukeneder A. (2015).- Ammonoid Habitats and Life History. In: Klug C., Korn D., De Baets K., Kruta I. & Mapes R.H. (eds.), Ammonoid paleobiology: from anatomy to ecology.- Topics in Geobiology, New York, vol. 43, p. 689-791.
Masse J.P. & Fenerci-Masse M. (2011).- Drowning discontinuities and stratigraphic correlation in plateform carbonates. The late Barremian early Aptian record of southeast France.- Cretaceous Research, vol. 32, no. 6, p. 659-684.
Mikhailova I.A. & Baraboshkin E.Y. (2001).- First Finds of Lithancylus Casey, 1960 (Ammonoidea, Ancyloceratidae) in the Lower Aptian of Ul'yanovsk Povolzhie (Volga Region).- Paleontological Journal, Moscow, vol. 35, no. 4, p. 367-378.
Pictet A., Delanoy G., Baudouin C. & Boselli P. (2009).- Le genre Lithancylus Casey, 1960 (Ammonoidea, Ancyloceratina) dans l'Aptien inf�rieur du Couloir rhodanien (Dr�me, Sud-Est de la France).- Revue de Pal�obiologie, Gen�ve, vol. 28, no. 2, p. 491-509.
Pictet A., Delanoy G., Adatte T., Spangenberg J.E., Baudouin C., Boselli P., Boselli M., Kindler P. & F�llmi K.B. (2015).- Three successive phases of platform demise during the early Aptian and their association with the oceanic anoxic Selli episode (Ard�che, France).- Palæogeography, Palæoclimatology, Palæoecology, vol. 418, p. 101-125.
Ragazzi M. (1982, unpublished).- L'Aptien dans les Alpes Maritimes: stratigraphie, pal�o�cologie, s�dimentologie.- PhD thesis, Universit� de Nice, 224 p.
Reboulet S, Szives O., Aguirre-Urreta B., Barrag�n R., Company M., Frau C., Kakabadze M.V., Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S.N., Va��ček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Roch É. (1927).- Étude stratigraphique et pal�ontologique de l'Aptien inf�rieur de la B�doule (pr�s Cassis) (Bouches-du-Rh�ne).- M�moires de la Soci�t� g�ologique de France (Nouvelle S�rie), Paris t. IV, no. 8, p. 5-37.
Ropolo P., Conte G., Gonnet R., Masse J.-P. & Moullade M. (1998).- Les faunes d'Ammonites du Barr�mien sup�rieur/Aptien inf�rieur (B�doulien) dans la r�gion stratotypique de Cassis-La B�doule (SE France): �tat des connaissances et propositions pour une zonation par Ammonites du B�doulien-type.- G�ologie M�diterran�enne, Marseille, vol. 25, no. 3-4, p. 167-175.
Sarkar S.S. (1955).- R�vision des ammonites d�roul�es du Cr�tac� inferieur du Sud-Est de la France.- M�moires de la Soci�t� g�ologique de France (Nouvelle S�rie), Paris, vol. 72, 176 p.
Sayn
G. (1920).- Les Phylloceras gargasiens du Sud-Est de
Thieuloy J.P. (1990).- Un ancyloceratide g�ant de l'Aptien du Gard (France). Toxoceratoides? sp. inc. "gigantomorphe godeti" nov.- G�ologie Alpine, Grenoble, vol. 66, p. 101-106.
Thomel G. (1963).- Consid�ration sur la faune gargasienne du Sud-Est de la fosse vocontienne.- Travaux du Laboratoire de G�ologie de la Facult� des Sciences de Marseille, Marseille, t. 7, p. 47-58.
Thomel G. (1964).- Les zones d'Ammonites de l'Aptien des Basses-Alpes.- Comptes Rendus de l'Acad�mie des Sciences, Paris, groupe 9, t. 258, p. 4308-4310.
Thomel G. (1968).- À propos de l'Ammonoceratites (Argonauticeras) depereti (Kilian) du Gargasien des Basses-Alpes orientales.- Bulletin de la Soci�t� g�ologique de France (7e S�rie), Paris, t. X, p. 684-687.
Va��ček Z., Rabrenović D., Skupien P., Radulović V. J., Radulović B. V. & Mojsić I. (2014).- Ammonites (Phylloceratina, Lytoceratina and Ancyloceratina) and organic-walled dinoflagellate cysts from the Late Barremian in Boljetin, eastern Serbia.- Cretaceous Research, vol. 47, p. 140-159.
Vermeulen J., Lazarin P., Lepinay P., Leroy L., Mascarelli E. & Pacaud J.-M. (2015).- Nouvelles donn�es sur le genre Ptychoceras Orbigny, 1842 et sur son esp�ce-type (Mollusca, Cephalopoda, Ammonoidea).- Annales du Mus�um d'Histoire Naturelle de Nice, vol. XXX, p. 1-12.
Ward P.D. (1987).- The natural history of Nautilus.- Allen & Unwin eds., Boston, 267 p.
Westermann G.E.G. (1990).- New developments in ecology of Jurassic-Cretaceous ammonoids. In: Pallini G., Cecca F., Cresta S. & Santantonio M. (eds.), Fossili, evoluzione, ambiente.- Atti II Conventione Internationale Pergola, 1987, Tecnostampa, Ostra Vetere, p. 459-478.
Westermann G.E.G. (1996).- Ammonoid life and habitat. In: Landman N.H., Tanabe K. & Davis R.A. (eds.), Ammonoid Paleobiology.- Topics in Geobiology, New York, vol. 13, p. 607-707.
Wright C.W., Callomon J.H. & Howarth M.K. (1996).- Treatise on Invertebrate Paleontology. Part L, Mollusca 4 (revised), Volume 4: Cretaceous Ammonoidea.- The Geological Society of America, Boulder; the University of Kansas, Lawrence, 362 p.
Plate 1:
Cephalopods
of the last bed of the Hauterivian-Aptian limestones (Deshayesites
forbesi Zone). All specimens: Bersac's collection. |
|
||
Plate 2:
Fossils of
the Ammonitoceras Level (top of the Pont de la Cerise Member, Dehayesites
deshayesi Zone, Deshayesites multicostatus Subzone). Scale bar
50 mm for figures 1 to 17. Scale bar 10 mm for figures 18 to 20. All
specimens: Bersac's collection. |
|
||
Plate 3:
Cephalopods
of the Toxoceratoides Bed (base of the Les Graous Member, Dehayesites
deshayesi Zone, Deshayesites grandis Subzone). All specimens: Bersac's
collection. |
|
||
Plate 4:
Cephalopods
of the Les Graous Member (Dufrenoyia furcata Zone, Dufrenoyia
furcata Subzone). All specimens: Bersac's collection. |
|
||
Plate 5:
Fossils of
the Les Graous Member. Figures 1-29: Dufrenoyia furcata Zone, Dufrenoyia furcata Subzone.
Figures 30-32: Dufrenoyia furcata Zone, Dufrenoyia dufrenoyi
Subzone. All
specimens: Bersac's collection. |
|