◄ Carnets Geol. 20 (8) ►
![]()
Sommaire
[1. Introduction]
[2. Cadre géographique et bref aperçu géologique]
[3. Objectif et méthode]
[4. Description lithostratigraphique et répartition des ostracodes]
[5. Évčnement Anoxique Océanique II (EAO II) ŕ l'échelle globale et dans le bassin du Tinrhert]
[6. Incidence des évčnements sédimentologiques et eustatiques sur l'évolution des ostracodes]
[7. Conclusion : Tendances biogéographiques]
et ...
[Références bibliographiques]
Laboratoire de Recherche nº 10, Université Abou-Bekr Belkaid, Tlemcen (Algérie)
Dame du Lac 213, 3 rue Henri-Barbusse, F-76300 Sotteville-lčs-Rouen (France)
Laboratoire de Recherche nº 25, Université Abou-Bekr Belkaid, Tlemcen (Algérie)
Laboratoire de Recherche nº 25, Université Abou-Bekr Belkaid, Tlemcen (Algérie)
Faculté des Sciences de la Nature et de la Vie, Université de Mascara, 29000 Mascara (Algérie)
Laboratoire de Recherche nº 25, Université Abou-Bekr Belkaid, Tlemcen (Algérie)
Laboratoire de Recherche nº 25, Université Abou-Bekr Belkaid, Tlemcen (Algérie)
Manuscrit en ligne depuis le 11 mai 2020
DOI 10.4267/2042/70792
![]()
[Éditeur : Bruno Granier ; éditeur pour la langue
anglaise : Stephen Eagar]
![]()
Dans le bassin du Tinrhert, au passage Cénomanien/Turonien, les ostracodes sont
rares dans la plupart des niveaux échantillonnés ; ils ne présentent
aucune variabilité, leur fréquence ne dépassant pas 4% de toute la microfaune.
Leur présence optimale, tant qualitative que quantitative, s'observe ŕ la
base du Turonien inférieur oů leurs cortčges sont dominés par les genres Cythereis,
Paracypris et Cytherella.
Afin
de comprendre ces observations, nous avons mené une étude paléoécologique
sur cinq coupes géologiques et reconstitué l'impact des conditions sédimentologiques
(quartz, gypse et pyrite) et celui des variations eustatiques.
• ostracodes ;
• Crétacé ;
• Cénomanien-Turonien ;
• EAO2 ;
• Algérie ;
• bassin
du Tinrhert ;
• paléo-environnement ;
• paléoécologie
Tchenar S., Ferré B., Adaci M., Zaoui D., Benyoucef M., Bensalah M. & Kentri S. (2020).- Incidences de l'Évčnement Anoxique Océanique II sur l'évolution des ostracodes des dépôts cénomano-turoniens du bassin du Tinrhert (SE Algérie).- Carnets Geol., Madrid, vol. 20, nº 8, p. 145-164.
Impact of the Anoxic Oceanic
Event II on the evolution of ostracods in the Cenomanian-Turonian deposits of
the Tinrhert Basin (SE Algeria).- Around the Cenomanian/Turonian
boundary in the Tinrhert Basin, ostracods are seldom found in most of the
sampled levels. The assemblages show no specific diversity, their frequency not
exceeding 4% of the whole microfauna. Their 'climactic' presence, both
qualitatively and quantitatively, is observed at the base of the lower Turonian,
where ostracod assemblages are dominated by the genera Cythereis, Paracypris,
and Cytherella.
In order to interpret this data, we document a paleoecological study supported
by evidence compiled from five geological sections, dealing with the respective
impact of sedimentological conditions (quartz, gypsum and pyrite), and sea-level
fluctuations.
• ostracodes;
• Cretaceous;
• Cenomanian-Turonian;
• OAE2;
• Algeria;
• Tinrhert basin;
• paleo-environment;
• paleoecology
Dans le bassin du Tinrhert, les nombreux travaux réalisés se sont essentiellement intéressés ŕ l'étude lithologique et biostratigraphique. Citons entre autres : Busson (1960, 1971), Busson et al. (1999), Amédro et al. (1996), Tchenar et al. (2015), Zaoui et al. (2016) et Ferré et al. (2016). Le présent travail traite, pour la premičre fois, de la reconstitution paléo-écologique de ce secteur au moyen des ostracodes.
Le
bassin du Tinrhert occupe la partie orientale de la plate-forme saharienne, dans
les confins algéro-tuniso-libyens (Fig. 1 ![]() ).
Sur
le plan géologique, les dépôts sédimentaires datent essentiellement du
Crétacé.
L'Albien y est représenté par des grčs rapportés au "Continental
Intercalaire" et dépourvus de fossiles. Au Cénomanien, les dépôts présentent
un facičs lagunaire de bordure, représentés par des argiles gypseuses et
pyriteuses.
).
Sur
le plan géologique, les dépôts sédimentaires datent essentiellement du
Crétacé.
L'Albien y est représenté par des grčs rapportés au "Continental
Intercalaire" et dépourvus de fossiles. Au Cénomanien, les dépôts présentent
un facičs lagunaire de bordure, représentés par des argiles gypseuses et
pyriteuses.
|
Figure 1 : Situation générale du bassin du Tinrhert (d'aprčs Amédro et al., 1996) et localisation des coupes. BOD : Bordj Omar Driss; TAK : Takouazet; 06 Oh : Ohanet 06 Oh; 07 Oh : Ohanet 07 Oh; In Ad : In Adaoui. General location of the Tinrhert basin
(from Amédro et al., 1996) and position of the studied sections. BOD: Bordj
Omar Driss; TAK: Takouazet; 06 Oh: Ohanet 06 Oh; 07 Oh: Ohanet 07 Oh; In Ad:
In Adaoui. |
Au
passage Cénomanien supérieur/Turonien inférieur, ces dépôts passent ŕ une
sédimentation carbonatée de plate-forme, formant la dalle dite cénomanienne.
Au Turonien inférieur, s'installe une sédimentation d'abord marneuse,
coiffée par des calcaires massifs formant la dalle turonienne. La série se
termine par des argiles gypseuses, rarement dolomitiques, du Coniacien-Santonien.
Les différentes dalles carbonatées arment des corniches qui forment de précieux
repčres visuels dans la topographie (Fig. 1 ![]() ).
).
Le présent travail met en évidence l'incidence des variations paléo-écologiques sur la répartition et l'évolution des cortčges d'ostracodes. Cinq coupes, chacune d'exposition générale NS, ont été levées dans le bassin, selon un transect général d'orientation ENE-WSW. Il s'agit des coupes de Bordj Omar Driss, de Takouazet, de l'Ohanet (coupes 6 Oh et 7 Oh) et d'In Adaoui.
Les niveaux marneux, dont la microfaune est aisément récupérable, ont fait l'objet de 132 prélčvements avec la répartition suivante : 35 pour la coupe de Bordj Omar, 32 ŕ Takouazet, 26 ŕ l'Ohanet 06 Oh, 22 ŕ l'Ohanet 07 Oh et 17 ŕ In Adaoui. Les échantillons marneux (environ 500 g par niveau) ont été lavés ŕ l'eau sur une colonne de deux tamis, respectivement de maille 125μm et 63μm (norme AFNOR, ISO 3310.1). Les résidus de lavage, ainsi récupérés et séchés, ont été triés et étudiés ŕ la loupe binoculaire de type épiscope, afin de recenser et d'identifier la microfaune ŕ des fins biostratigraphiques, puis d'évaluer les différentes fractions minéralogiques et biogéniques, pour reconstituer les conditions paléo-environnementales.
La méthodologie de comptage adoptée est celle préconisée par Arias (1995), Bodergat et al. (1997), Babinot et Colin (2011), Tchenar (2004, 2016) et Tchenar et al. (2015). Ainsi, le nombre se rapporte ŕ l'ensemble de la microfaune présente (100%) et récoltée par niveau. Tous les spécimens d'ostracodes reconnus dans chaque résidu ont été identifiés, triés, comptés et leur pourcentage, calculé.
Quant aux niveaux calcaires pour lesquels l'extraction de la microfaune reste problématique et aléatoire, ils ont fait l'objet de lames minces qui ont permis l'analyse de leur microfacičs respectif et la détermination de leur biophase. Les ostracodes, rarement présents, y sont mal conservés et indéterminables.
Par commodité et pour faciliter la lecture, la terminologie de Busson (1960) a été adoptée pour les différentes unités lithologiques identifiées. Ŕ l'échelle du bassin du Tinrhert, les dépôts du Cénomanien et du Turonien sont représentés par deux membres, comprenant chacun deux termes.
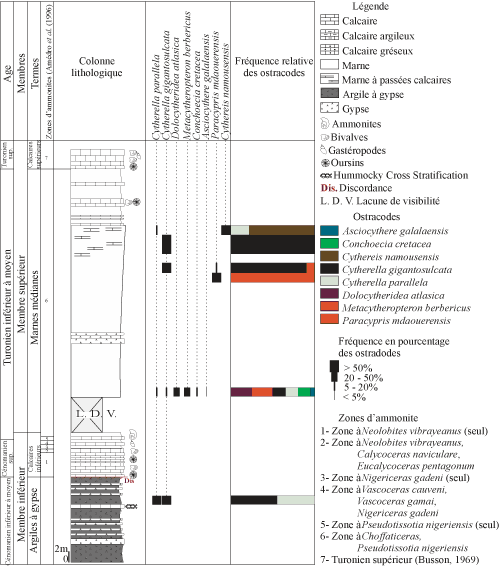
4.
A. Coupe
de Bordj Omar Driss (BOD) (Figs. 2.A ![]() ,
3
,
3 ![]() )
)
Située
dans la région de Bordj Omar Driss, son épaisseur avoisine 73 mčtres (Fig. 3 ![]() ). La série présente les
membres inférieur et supérieur de Busson (1960).
). La série présente les
membres inférieur et supérieur de Busson (1960).
a - membre inférieur
Ce membre repose généralement sur les grčs albiens ; toutefois, ce contact n'est pas observable dans la région étudiée (Busson, 1960). Il comporte les deux termes suivants :
terme A :
les argiles ŕ gypse (Cénomanien inférieur ŕ moyen). Elles
représentent une sédimentation argileuse rougeâtre ŕ niveaux de gypse réticulé
(Fig. 2.F ![]() ). Les résidus de lavage
abondent en gypse et en pyrite.
). Les résidus de lavage
abondent en gypse et en pyrite.
|
Figure 2 : A : Vue panoramique de la coupe de Bordj Omar Driss, depuis le sommet des Calcaires inférieurs ; B : Vue panoramique de la coupe de Takouazet, depuis le sommet des Calcaires inférieurs ; C-D : Calcaires inférieurs de la coupe de Bordj Omar Driss. C - niveau bioturbé et fossilifčre : O. oursin, L. lamellibranche. D - présence de terriers (T) ; E : Calcaires crayeux bioclastiques fossilifčres de la coupe de Takouazet : O. oursin, P. pecten ; F : Argiles ŕ gypse, coupe de Bordj Omar Driss; G : stratifications entrecroisées ("Hummocky Cross Stratifcation"), Argiles ŕ gypse, coupe de Bordj Omar Driss. A: Scenic view of the Bordj Omar
Driss section, from the top of the 'Calcaires inférieurs' (lower limestones);
B: Scenic view of the Takouazet section, from the top of the 'Calcaires
inférieurs'; C-D: 'Calcaires inférieurs' (lower limestones) of the
Bordj Omar Driss section. C - bioturbated and fossiliferous level : O.
sea urchin, L. lamellibranch. D - occurrence of burrows (T); E:
Fossiliferous and bioclastic, chalky limestones of the Takouazet section: O.
sea urchin, P. pecten; F: Clays with gypsum, Bordj Omar Driss section; G:
Hummocky Cross Stratification, Clays with gypsum, Bordj Omar Driss section. |
|
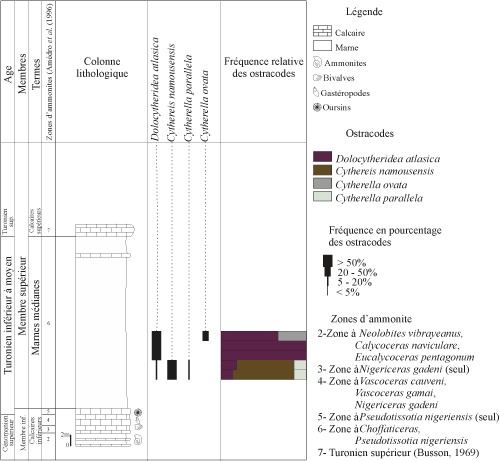
Figure 3 : Distribution et fréquence des ostracodes de la coupe de Bordj Omar Driss. Vertical range and frequency of the
respective ostracode species of the Bordj Omar Driss section. |
La fraction bioclastique, dépourvue de microfaune, est caractérisée par la fréquence des clypéines (algues dasycladacées), de quelques dents de poisson et de rares ossicules d'échinoderme.
Les niveaux calcaires qui s'intercalent parfois dans ces argiles, renferment quelquefois des rognons ferrugineux. Ils se débitent en plaquettes, ou présentent des "Hummocky Cross Stratification" (Fig. 2.G
). Exceptionnellement, les niveaux immédiatement sus-jacents comportent une abondante ostracofaune parmi laquelle figurent Cytherella parallela (Reuss) et C. gigantosulcata Rosenfeld. Ces deux espčces n'y sont représentées que par des individus adultes.
Le microfacičs du premier banc montre une micrite, composée de lamines alternantes sombres et claires, caractéristique d'un milieu lagunaire (Kazi-Tani, 1986) (Fig. 4.A
|
Figure 4 : A : Micrite ŕ lamines alternantes sombres et claires, Argiles ŕ gypse, coupe de Bordj Omar Driss ; B : Calcaires inférieurs de la coupe de Bordj Omar Driss: F. 'filaments', R. Rovéacrinidé ; C : Pelbiomicrite ŕ clypéines et ŕ foraminifčres, présence de cristaux de dolomie secondaire, calcaires inférieurs, coupe de Bordj Omar Driss : C. Clypeina, D. dolomie ; D : Pelbiomicrite ŕ foraminifčres de la coupe de Takouazet : M. Marginotruncana. A:
Micrite with alternating dark-and-light laminations, Clays with gypsum ('Argiles
ŕ gypse'), Bordj Omar Driss section; B: Lower limestones ('Calcaires
inférieurs') of the Bordj Omar Driss section: F. 'filaments', R.
Roveacrinidae; C: Pelbiomicrite with clypeinas and foraminifers,
occurrence of secondary dolomite crystals, Lower limestones ('Calcaires
inférieurs'), Bordj Omar Driss section: C. Clypeina, D. dolomite; D:
Pelbiomicrite with clypeinas and foraminifers of the Takouazet section: M. Marginotruncana.
[B-D photosphopped] |
terme B :
les calcaires inférieurs. Transgressif
sur le terme précédent, il forme la corniche du Cénomanien supérieur. Il
s'agit d'une dalle de calcaires massifs fossilifčres, intercalés de marnes,
en particulier vers son sommet. Ils renferment des oursins, des gastéropodes,
des lamellibranches (exogyres) et comportent également des terriers (Fig.
2.C-D ![]() ).
Les ammonites récoltées [Neolobites
vibrayeanus (Orbigny), Forbesiceras sp.
et Eucalycoceras pentagonum (Jukes-Browne)]
indiquent le Cénomanien supérieur (Amédro et al.,
1996). Le microfacičs montre une bio-micrite ŕ clypéines et
ŕ 'filaments' (Fig. 4.B
).
Les ammonites récoltées [Neolobites
vibrayeanus (Orbigny), Forbesiceras sp.
et Eucalycoceras pentagonum (Jukes-Browne)]
indiquent le Cénomanien supérieur (Amédro et al.,
1996). Le microfacičs montre une bio-micrite ŕ clypéines et
ŕ 'filaments' (Fig. 4.B ![]() ).
).
La partie supérieure de ce terme comporte une alternance marno-calcaire "serrée". Ce sont des calcaires centimétriques de couleur verdâtre ŕ beige. Le microfacičs du dernier banc calcaire montre une pel-bio-micrite ŕ clypéines et foraminifčres (essentiellement benthiques). Les cristaux de dolomie secondaire y sont trčs fréquents (Fig. 4.C
Ces derniers bancs calcaires ont livré une section de foraminifčre planctonique, pouvant se rapporter au genre Marginotruncana. Dans la coupe de Takouazet, la présence de ce genre au sommet des calcaires inférieurs est confirmée.
b - membre supérieur
Il
présente une sédimentation essentiellement marneuse, devenant marno-calcaire
au sommet. Les calcaires renferment des bivalves et des oursins d'âge
Turonien (Busson, 1960). Les marnes sont
compactes. Ce
membre comprend les termes suivants (Fig. 2.F ![]() ) :
) :
terme A : les marnes médianes. Leur base n'est pas visible ŕ Bordj Omar Driss. Elles forment une grande combe marno-gypseuse, riche en veinules de gypse. Dans ces niveaux, l'ostracofaune est représentée par les espčces Dolocytheridea atlasica Bassoullet et Damotte, Metacytheropteron berbericus (Bassoullet et Damotte), Cytherella parallela (Reuss), C. gigantosulcata Rosenfeld, Conchoecia cretacea Pokorny et Asciocythere galalensis Szczechura et al.
Ce terme se poursuit par des niveaux marneux ŕ passées calcaires. Leurs résidus montrent une réduction notable de la fraction minéralogique et des bioclastes. Les ostracodes y sont représentés par Cytherella gigantosulcata Rosenfeld et Paracypris mdaouerensis Bassoullet et Damotte.
Les marnes médianes se terminent par des marnes avec des intercalations de bancs calcaires crayeux, ŕ oursins, gastéropodes et bivalves. Ce sont des marnes compactes, de couleur jaune verdâtre. Leurs résidus montrent une fraction importante d'éléments oxydés et de rares grains de gypse, de pyrite et de quartz. La microfaune est totalement absente.
terme B : les calcaires supérieurs. Il est représenté par des bancs calcaires massifs beiges, riches en lamellibranches, oursins et gastéropodes, d'âge Turonien supérieur (Busson, 1969). Les derniers bancs comportent une croűte ferrugineuse. Ce terme est attribué au Turonien supérieur (Busson, 1969) et constitue la corniche turonienne.
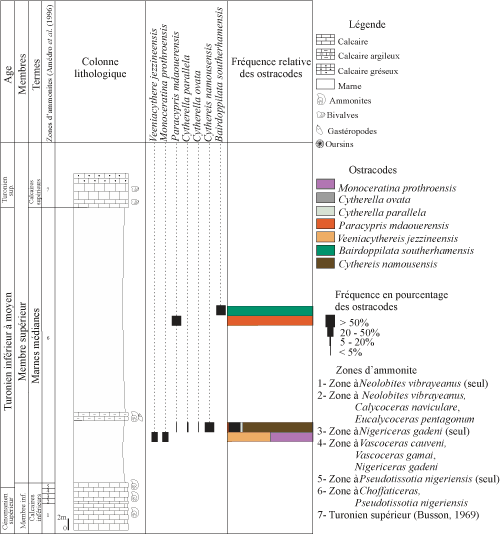
4.
B. Coupe de
Takouazet (Tak) (Fig. 5 ![]() )
)
Elle est distante d'environ 250 km ŕ l'Ouest de la coupe précédente (BOD) et s'étend sur une épaisseur d'environ 63 mčtres.
a - membre inférieur
Il est uniquement formé des calcaires inférieurs.
terme B :
les calcaires inférieurs.
Ils
correspondent ŕ une dalle de calcaires crayeux, compacts et fossilifčres (Fig.
2.E ![]() ). Amédro et al.
(1996) y signalent des ammonites, telles que Neolobites
vibrayeanus (Orbigny), Forbesiceras sp.,
Calycoceras (Calycoceras) naviculare (Mantell)
et Eucalycoceras pentagonum (Jukes-Browne),
du Cénomanien supérieur.
). Amédro et al.
(1996) y signalent des ammonites, telles que Neolobites
vibrayeanus (Orbigny), Forbesiceras sp.,
Calycoceras (Calycoceras) naviculare (Mantell)
et Eucalycoceras pentagonum (Jukes-Browne),
du Cénomanien supérieur.
Le microfacičs des derniers bancs est une pel-bio-micrite ŕ foraminifčres, parmi lesquels figure Marginotruncana (Fig. 4.D
|
Figure 5 : Distribution et fréquence des ostracodes de la coupe lithologique de Takouazet. Vertical range and frequency of the
respective ostracode species of the Takouazet section. |
b -
membre supérieur
(Fig. 5 ![]() )
)
terme A : les marnes médianes
(Figs. 2.B ![]() , 5
, 5 ![]() ).
Il
débute par un banc calcaire ŕ Pseudotissotia
nigeriensis (Woods), daté
du Turonien inférieur (Amédro et al., 1996). Les marnes sont verdâtres et compactes. Leurs résidus
sont riches en pyrite et en calcite, pauvres en quartz, gypse et oxydes de fer.
La fraction biogénique est essentiellement marquée par l'abondance des
bioclastes, des dents de poisson et des clypéines, avec quelques débris de
gastéropodes et des nucléus d'ammonite. La microfaune est absente des
premiers niveaux. Dans les niveaux sus-jacents, les ostracodes sont représentés
par Veeniacythereis jezzineensis (Bischoff),
Monoceratina prothroensis Butler
et Jones, Paracypris mdaouerensis Bassoullet
et Damotte, Cytherella parallela (Reuss),
C ovata (Roemer) et Cythereis
namousensis Bassoullet et Damotte.
La série est interrompue par un banc calcaire marneux, plus ou moins gréseux,
ŕ ammonites et gastéropodes.
).
Il
débute par un banc calcaire ŕ Pseudotissotia
nigeriensis (Woods), daté
du Turonien inférieur (Amédro et al., 1996). Les marnes sont verdâtres et compactes. Leurs résidus
sont riches en pyrite et en calcite, pauvres en quartz, gypse et oxydes de fer.
La fraction biogénique est essentiellement marquée par l'abondance des
bioclastes, des dents de poisson et des clypéines, avec quelques débris de
gastéropodes et des nucléus d'ammonite. La microfaune est absente des
premiers niveaux. Dans les niveaux sus-jacents, les ostracodes sont représentés
par Veeniacythereis jezzineensis (Bischoff),
Monoceratina prothroensis Butler
et Jones, Paracypris mdaouerensis Bassoullet
et Damotte, Cytherella parallela (Reuss),
C ovata (Roemer) et Cythereis
namousensis Bassoullet et Damotte.
La série est interrompue par un banc calcaire marneux, plus ou moins gréseux,
ŕ ammonites et gastéropodes.
La sédimentation marneuse reprend sous l'expression d'une épaisse combe, trčs large de marnes verdâtres, ŕ passées violacées, compactes, parfois ŕ débit conchoďdal ou en plaquettes. La fraction biogénique est marquée par l'abondance des bioclastes : dents de poisson, fragments de bivalves, ossicules d'échinoderme et articles de clypéines. La partie basale de cet intervalle est dépourvue de microfaune (absence des espčces benthiques, foraminifčres et ostracodes). Dans la partie moyenne, les ostracodes, en faible nombre, sont représentés par Paracypris mdaouerensis Bassoullet et Damotte, et Bairdoppilata southerhamensis Weaver. Les marnes médianes datent du Turonien inférieur ŕ moyen.
terme B : les calcaires supérieurs. Il s'agit de calcaires massifs, parfois gréseux, riches en empreintes de bivalves et en gastéropodes. Ils sont gris ŕ jaunâtres, pouvant se débiter en plaquettes massives. Ils représentent la corniche du Turonien supérieur (Busson, 1969).
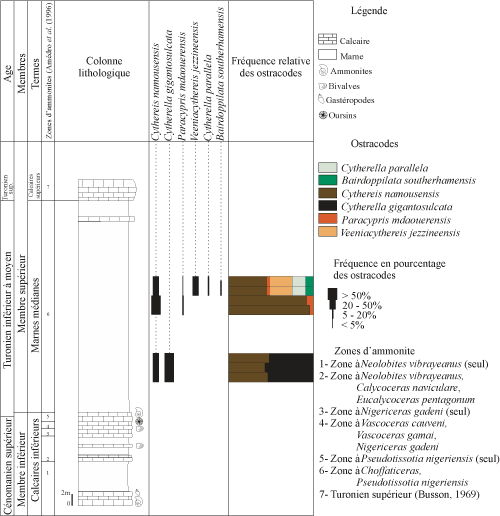
4. C. Coupes de l'Ohanet
Deux coupes ont été levées dans la région de l'Ohanet, au niveau de l'Oued In Adaoui. La coupe 06 Oh se situe ŕ 1 km l'ouest de la coupe 07 Oh.
4.
C.1 - Coupe 06 Oh (Fig. 6 ![]() )
)
Située dans la partie septentrionale du bassin, son épaisseur totale avoisine 58 mčtres. Le membre inférieur n'y est que partiellement représenté par les calcaires inférieurs.
a - membre inférieur
terme B : les calcaires inférieurs. Il correspond ŕ des calcaires crayeux et fossilifčres renfermant des bivalves, des gastéropodes, des oursins et des ammonites. Parmi ces derničres, Amédro et al. (1996) ont reconnu Neolobites vibrayeanus (Orbigny) dans les premiers bancs, puis Vascoceras gamai Choffat, V. cauvini Chudeau, Pseudaspidoceras grecoi Collignon, Fikaites subtuberculatus (Collignon) et F. laffitei (Collignon) dans les niveaux sus-jacents. Ces calcaires peuvent ętre bioturbés, débités ou compacts. Ils alternent avec des marnes dépourvues d'ostracode.
La série se poursuit par des marnes vertes, gypseuses et se débitant en feuillets. Leurs résidus sont riches en éléments oxydés, quartz et gypse. La fraction bioclastique est dominée par les clypéines et les dents de poisson. La microfaune n'a pas été repérée.
Ce terme est coiffé par une dalle de calcaires compacts, crayeux, riches en gastéropodes, oursins, lamellibranches et ammonites du Cénomanien supérieur.
|
Figure 6 : Distribution et fréquence des ostracodes de la coupe lithologique de l'Ohanet (06 Oh). Vertical range and frequency of the
respective ostracode species of the Ohanet section (06 Oh). |
b - membre supérieur
Il est marqué par une sédimentation marno-calcaire répartie en deux termes :
terme A : les marnes médianes. Ce sont des marnes vertes ŕ résidus bioclastiques. La calcite, abondante ŕ la base, diminue de fréquence pour disparaître au sommet. Le gypse y est fréquent. La pyrite et les oxydes de fer sont présents en faible quantité. Nous remarquons une augmentation de fréquence du quartz en allant vers le sommet de ce terme. La fraction bioclastique est représentée par des clypéines et des radioles d'oursin.
Les ostracodes sont présents en faible proportion. Dans les premiers niveaux, ils sont représentés par Cythereis namousensis Bassoullet et Damotte, et Cytherella gigantosulcata Rosenfeld. Dans la partie supérieure, plusieurs espčces apparaissent : Paracypris mdaouerensis Bassoullet et Damotte, Veeniacythereis jezzineensis (Bischoff), Cytherella parallela (Reuss) et Bairdoppilata southerhamensis Weaver.
terme B : les calcaires supérieurs. Ces calcaires sont trčs durs, riches en débris bioclastiques indéterminables, et marquent la corniche du Turonien supérieur.
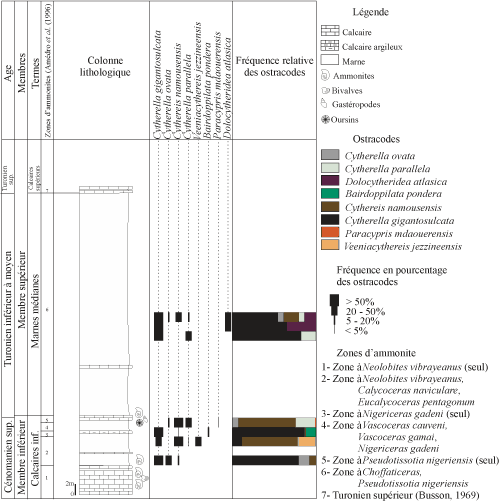
4.
C.2 - Coupe 07 Oh (Fig. 7 ![]() )
)
Localisée ŕ l'est de la coupe 06 Oh, d'une épaisseur de 55 mčtres, elle est généralement marneuse, avec des bancs calcaires fossilifčres livrant des mollusques, des ammonites et des lamellibranches. Elle montre les deux membres suivants :
a - membre inférieur
terme A : les calcaires inférieurs. Les premiers niveaux calcaires sont compacts et comportent des lamellibranches et des gastéropodes. Amédro et al. (1996) y ont signalé Neolobites vibrayeanus (Orbigny). Ils sont surmontés de marnes vertes dont les résidus renferment du quartz, de la pyrite, du gypse et de la calcite. La fraction biogénique est représentée par de nombreuses clypéines, des radioles d'oursin, des ossicules d'échinoderme, des dents de poisson et des fragments de gastéropode. La microfaune n'a pas été repérée.
|
Figure 7 : Distribution et fréquence des ostracodes de la coupe lithologique de l'Ohanet (07 Oh). Vertical range and frequency of the
respective ostracode species of the Ohanet section (07 Oh). |
Les marnes sus-jacentes renferment une microfaune abondante et diversifiée. Les ostracodes y sont représentés par Cytherella gigantosulcata Rosenfeld, C. ovata (Roemer), C. parallela (Reuss), Cythereis namousensis Bassoullet et Damotte, Veeniacythereis jezzineensis (Bischoff) et, dans les derniers niveaux, par Bairdoppilata pondera Jennings et Paracypris mdaouerensis Bassoullet et Damotte.
Ce terme est coiffé par des calcaires ŕ oursins, gastéropodes et ammonites (Choffaticeras ; F. Atrops, in coll.), indiquant le sommet du Cénomanien supérieur. Cet ensemble constitue la corniche du Cénomanien supérieur.
b - membre supérieur
Il comporte une alternance marno-calcaire dont les derniers bancs calcaires renferment des rognons de silex. Deux termes sont reconnus :
terme A : les marnes médianes. Ce sont des marnes vertes trčs épaisses, intercalées par un banc calcaire. Dans les résidus marneux, le gypse est rare ; la pyrite et le quartz, fréquents. Elles sont d'âge Turonien inférieur ŕ moyen.
La fraction bioclastique comprend des radioles d'oursin, des clypéines, des dents de poisson, des ossicules d'échinoderme et des fragments de gastéropode, essentiellement ŕ la base du terme. Ils se raréfient nettement en allant vers le sommet.
La microfaune est présente uniquement dans la partie médiane du terme. Elle se raréfie et disparaît dans les niveaux supérieurs. Parmi les ostracodes, ce terme est marqué par l'apparition de Dolocytheridea atlasica Bassoullet et Damotte, et la disparition de certaines espčces mentionnées dans les niveaux sous-jacents. Seules persistent Cytherella gigantosulcata Rosenfeld, C. ovata (Roemer), C. parallela (Reuss) et Cythereis namousensis Bassoullet et Damotte.
terme B : les calcaires supérieurs. Ils sont indurés et dépourvus de fossile. Les derniers bancs renferment des nodules de silex. Ils forment la corniche du Turonien.
4. D.
Coupe d'In-Adaoui (In Ad) (Fig. 8 ![]() )
)
Elle est située au NE de la coupe 07 Oh, au niveau de l'Oued In Adaoui. Son épaisseur avoisine 39 m. Ŕ In-Adaoui, le Cénomanien supérieur n'est représenté que par les calcaires inférieurs. On rencontre les membres suivants :
|
Figure 8 : Distribution et fréquence des ostracodes de la coupe lithologique d'In Adaoui. Vertical range and frequency of the
respective ostracode species of the In Adaoui section. |
a - membre inférieur
terme A : les calcaires inférieurs. Il s'agit de calcaires crayeux et compacts, riches en oursins, bivalves, gastéropodes et ammonites. Amédro et al. (1996) y signalent la présence d'un niveau repčre ŕ Neolobites vibrayeanus (Orbigny), formant un petit plateau et datant le Cénomanien supérieur. Ces calcaires alternent avec des marnes blanchâtres peu épaisses, dont les résidus n'ont pas livré de microfaune.
Ce terme représente la corniche du Cénomanien supérieur et se termine par des calcaires crayeux compacts ŕ ammonites, oursins et gastéropodes.
b - membre supérieur
Il est représenté par une alternance marno-calcaire, dont les marnes sont épaisses et verdâtres. Il est subdivisé en termes suivants :
terme A : les marnes médianes. Il s'agit d'épaisses marnes vertes contenant de la calcite, du quartz, du gypse, de la pyrite et des oxydes de fer. Pour la fraction biogénique, les dents de poisson et les ossicules d'échinoderme y sont peu fréquentes.
La microfaune est présente uniquement dans la premičre moitié du terme. Elle est faiblement représentée et peu diversifiée. Les ostracodes y sont rares. Ils ne sont représentés que par Dolocytheridea atlasica Bassoullet et Damotte, Cythereis namousensis Bassoullet et Damotte, Cytherella parallela (Reuss) et C. ovata (Roemer).
terme B : les calcaires supérieurs. Il s'agit d'une dalle de calcaires massifs, intercalée par des marnes azoďques. Cet ensemble forme la corniche turonienne.
Schlanger et Jenkyns (1976) introduisent le terme EAO pour désigner les épisodes de distribution globale de dépôts riches en matičre organique, enregistrés dans des séquences pélagiques du Crétacé, en particulier celui du passage Cénomanien/Turonien. Ce dernier (EAO-2) est également appelé "Bonarelli event" (Schlanger et al., 1987 ; Kuhnt et al., 1990). Il est caractérisé par :
l'installation des 'black shales', laminés et riches en matičre organique, représentant des périodes de stockage du carbone organique ŕ l'échelle globale ;
l'installation d'une zone ŕ oxygčne minimum dans des océans relativement chauds, provoquant une crise anoxique (Forster et al., 2007) ;
un réchauffement climatique important (Schlanger et Jenkyns, 1976) ;
une anomalie positive du δC13 (Kuypers et al., 2002) ;
une anomalie géochimique en mangančse (Kuypers et al., 2002) ;
des niveaux riches en pyrite et en phosphate (Kuypers et al., 2002) ;
une faune benthique réduite, voire absente (Kuypers et al., 2002) ;
une faune planctonique abondante, mais peu diversifiée et peu spécialisée (Kuypers et al., 2002) ;
une importante extinction, touchant simultanément la macro- et la microfaune au Cénomanien terminal, elle-męme suivie par de nouvelles apparitions dčs la base du Turonien inférieur (Kuypers et al., 2002) ;
une extension globale (Europe, Afrique, Amérique, Australie et Asie) sur une période allant de -93,5 ŕ -93 Ma (Kuypers et al., 2002) ;
une transgression, rapide et généralisée, qui semble induire ŕ son apogée une limitation du taux sédimentaire (Nzoussi-Mbassani, 2003) et
une expansion du domaine océanique qui aurait entrainé un blocage des apports terrigčnes loin des bassins sédimentaires océaniques, expliquant ainsi la 'non-dilution' de la matičre organique par les apports considérables de constituants minéraux (Nzoussi-Mbassani, 2003).
Chez les foraminifčres et les ostracodes, les études établies dans les bassins téthysiens et atlantiques ont mis en évidence, pour le passage Cénomanien supérieur/Turonien inférieur, les faits suivants :
l'extinction des foraminifčres planctoniques du groupe des Rotalipores (EWGPF, 1979),
la quasi-absence des foraminifčres benthiques et des ostracodes (Tronchetti et Grosheny, 1991 ; Soua, 2005) et
un renouvellement microfaunique chez les foraminifčres et les ostracodes (Sepkoski, 1986).
Dans le bassin du Tinrhert, le Cénomanien supérieur et le Turonien montrent les bio-évčnements suivants (Tchenar et al., 2015 ; Tchenar, 2016) :
une absence totale, ou presque, de microfaune ŕ la base du Cénomanien supérieur (argiles ŕ gypse) et dans la partie sommitale du Turonien inférieur ;
une forte dominance des foraminifčres planctoniques (entre 87% et 95% de la microfaune) ;
une abondance des foraminifčres planctoniques globuleux, notamment les Hétérohélicididés ('Heterohelix event' ; entre 30% et 100% de la population de foraminifčres et entre 25% et 78% de toute la microfaune) ;
une rareté de la microfaune benthique (foraminifčres et ostracodes, ne dépassant pas 4% de toute la microfaune).
une diversité spécifique trčs faible, tant chez les foraminifčres que chez les ostracodes ;
l'absence de foraminifčres dotés de carčne au Cénomanien supérieur. Ils réapparaissent au Turonien inférieur ŕ moyen, avec les espčces du genre Marginotruncana ;
l'absence totale de foraminifčres agglutinés au Cénomanien supérieur et au Turonien ;
un renouvellement de la microfaune dčs la base du Turonien, souligné par l'apparition de nouvelles espčces et
une abondance de 'filaments' dans certains niveaux calcaires du Turonien inférieur ŕ moyen.
Toutefois, dans le bassin du Tinrhert, l'évčnement anoxique océanique montre une hétérochronie de son extinction. Ce décalage serait probablement lié ŕ la paléo-topographie différentielle du bassin. Certains auteurs (Layeb, 1990 ; Layeb et Belayouni, 1999) ont déjŕ invoqué cette hypothčse pour expliquer le dépôt différentiel des facičs riches en matičre organique du passage Cénomanien/Turonien, appelés "black shales".
Dans le bassin du Tinrhert, les ostracodes sont trčs rares, voire absents, dans la plupart des niveaux échantillonnés. En outre, leur diversité spécifique reste faible et leur fréquence ne dépasse pas 4% de toute la microfaune.
Leur présence est trčs irréguličre le long des profils lithologiques. Ils sont absents au Cénomanien, ŕ l'exception d'un seul niveau, dans la partie médiane du Turonien inférieur et ŕ la limite Turonien inférieur/supérieur. Leur présence, tant qualitative que quantitative, s'observe préférentiellement ŕ la base du Turonien inférieur.
Leur présence accrue dans le niveau cénomanien serait due ŕ une immersion temporaire du lagon. Ils seraient restés 'confinés' dans le lagon avec une salinité suffisante ŕ leur survie. L'absence de juvéniles plaide en faveur de conditions défavorables ŕ leur cycle de vie et ŕ une défaillance dans leur stratégie adaptative. La diversité spécifique des ostracodes est liée ŕ la stabilité chimique du milieu tandis que leur densité faunique est généralement rattachée ŕ la richesse trophique (Babinot et Crumičre-Airaud, 1990).
Des études réalisées en domaine océanique (Benson, 1977) ont montré que l'abondance des ostracodes aveugles ŕ architecture externe complexe ou lisse, indique un environnement profond : talus, plateforme externe et bassin proximal (Rosenfeld et Bein, 1978). Les ostracodes ŕ fort tubercule oculaire témoignent, quant ŕ eux, d'une bathymétrie faible ou moyenne, dans la zone photique (Rosenfeld et Bein, 1978).
Les morphes lisses, bien calcifiés, suggčrent un milieu avec un équilibre entre Ca2+ et Mg+. Les Cytherella indiquent plutôt des eaux ŕ salinité normale, peu profondes (Depęche, 1984), littorales et chaudes (Morkhoven, 1963 ; Peypouquet et al., 1981 ; Depęche, 1984 ; Whatley, 1995 ; Bonnet et al., 1999). Les tests réticulés indiquent un enrichissement en Mg+. La présence simultanée de réticulations et de tubercules évoque des apports saisonniers organo-siliceux continentaux (Carbonel et al., 1988 ; Tölderer-Farmer, 1985).
Dans le cadre de notre étude, l'étude paléo-écologique des ostracodes a porté sur :
la diversité spécifique,
le taux de renouvellement spécifique et
le rapport ostracodes ornés/ostracodes lisses.
Les fluctuations de ces indices sont corrélées ŕ celles des fréquences de la pyrite, du gypse, et du quartz, ainsi qu'ŕ celles de la courbe eustatique (Haq et al., 1987).
6. A.
Diversité spécifique et renouvellement microfaunique (Figs. 9 ![]() , 10
, 10 ![]() ,
11
,
11 ![]() ,
12
,
12 ![]() ,
13
,
13 ![]() )
)
Ces deux indices présentent la męme évolution.
Diversité spécifique
Également appelée "sample-based rarefaction" (Gotelli et Colwell, 2001, Colwell et al., 2004, Mao et al., 2004), la diversité spécifique représente le nombre d'espčces d'ostracode identifiées par échantillon. Ses variations matérialisées sur une courbe permettent de visualiser de maničre directe les perturbations environnementales au cours du temps.
Renouvellement microfaunique
Il est calculé grâce au rapport :
| Espčces apparues n1+Espčces disparues n2 |
| Σ Espčces (totales) n1 + n2 |
avec n1 : niveau (ou échantillon) 1, et n2 : niveau sus-jacent ŕ n1. Le rapport ainsi calculé correspond au niveau n2.
L'analyse des figures met en évidence une corrélation positive directe entre la diversité spécifique des ostracodes et l'élévation du niveau marin. De męme, sa décroissance coďncide avec l'augmentation de fréquence de gypse et de pyrite.
Au Turonien inférieur, le renouvellement microfaunique illustre le retour ŕ un milieu favorable ŕ l'épanouissement de la microfaune, en particulier benthique. Les ostracodes se développent, mais en nombre restreint, et ne présentent aucune variabilité qualitative ou quantitative.
Les espčces présentes indiquent toutes un milieu peu profond, probablement de zone néritique. La présence, ŕ Bordj Omar Driss, de Conchoecia cretacea et de Monoceratina prothroensis, vivant dans les zones plus profondes, n'est pas contributive, leur fréquence respective n'étant pas significative.
L'élévation du niveau marin relatif est suivie par une amélioration des conditions écologiques et le retour ŕ la normale de l'oxygénation et de la salinité. L'absence des ostracodes au Cénomanien supérieur est ŕ mettre en relation avec l'installation d'un milieu réducteur (présence de pyrite) et une salinité élevée (présence de cristaux de gypse).
Dans la partie médiane du Turonien inférieur, les cinq coupes documentent un retour ŕ des conditions réductrices, avec une tendance ŕ l'émersion (augmentation de la fréquence du gypse et de la pyrite, des apports terrigčnes, du quartz), expliquant l'absence des ostracodes ŕ cette période. Malgré leur tendance ŕ se développer au sommet des marnes médianes, les conditions restent défavorables ŕ la microfaune benthique et les ostracodes finissent par disparaître.
Au passage Cénomanien supérieur/Turonien inférieur, le renouvellement microfaunique des ostracodes a été étudié en détail par plusieurs auteurs, parmi lesquels : Bassoullet et Damotte (1969), Damotte et Saint-Marc (1972), Grosdidier (1973), Gerry et Rosenfeld (1973), Rosenfeld et Raab (1974), Al-Abdul-Razzaq (1980), Ben Youssef (1980), Al-Abdul-Razzaq et Grosdidier (1981), Bismuth et al. (1981a, 1981b), Al-Furaih (1983), Gargouri-Razgallah (1983), Babinot (1985), Babinot et Basha (1985), Damotte (1985, 1995), Vivičre (1985), Athersuch (1988, 1994), Babinot et Colin (1988), Majoran (1988, 1989, 1996), Andreu-Boussut (1991), Andreu (1993), Abdallah et al. (1995), Gebhardt (1999), Colin et al. (2001), Luger (2003) et Schulze et al. (2004).
|
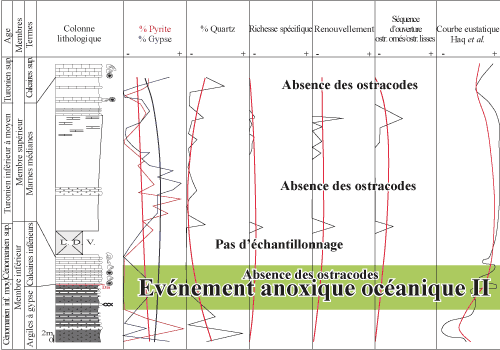
Figure 9 : Corrélations entre diversité spécifique, renouvellement spécifique et séquence d'ouverture des ostracodes, en relation avec la fréquence de pyrite, du gypse et du quartz, et la courbe eustatique au passage Cénomanien/Turonien de la coupe de Bordj Omar Driss. Correlation between species diversity,
species turn-over and marine opening set of ostracodes, in relation to the
occurrence of pyrite, gypsum and quartz, and the sea-level curve around the
Cenomanian/Turonian boundary in the Bordj Omar Driss section. |
|
Figure 10 : Corrélations entre diversité spécifique, renouvellement spécifique et séquence d'ouverture des ostracodes, en relation avec la fréquence de pyrite, du gypse et du quartz, et la courbe eustatique au passage Cénomanien/Turonien de la coupe de Takouazet. Correlation between species diversity,
species turn-over and marine opening set of ostracodes, in relation to the
occurrence of pyrite, gypsum and quartz, and the sea-level curve around the
Cenomanian/Turonian boundary in the Takouazet section. |
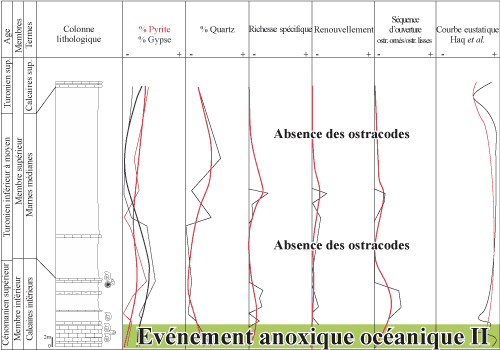
|
Figure 11 : Corrélations entre diversité spécifique, renouvellement spécifique et séquence d'ouverture des ostracodes, en relation avec la fréquence de pyrite, du gypse et du quartz, et la courbe eustatique au passage Cénomanien/Turonien de la coupe de l'Ohanet 06 Oh. Correlation between species diversity,
species turn-over and marine opening set of ostracodes, in relation to the
occurrence of pyrite, gypsum and quartz, and the sea-level curve around the
Cenomanian/Turonian boundary in the Ohanet (06 Oh) section. |
|
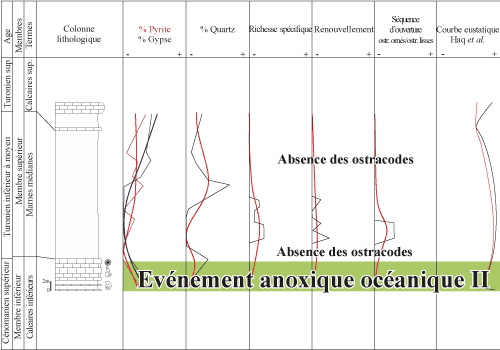
Figure 12 : Corrélations entre diversité spécifique, renouvellement spécifique et séquence d'ouverture des ostracodes, en relation avec la fréquence de pyrite, du gypse et du quartz, et la courbe eustatique au passage Cénomanien/Turonien de la coupe de l'Ohanet 07 Oh. Correlation between species diversity,
species turn-over and marine opening set of ostracodes, in relation to the
occurrence of pyrite, gypsum and quartz, and the sea-level curve around the
Cenomanian/Turonian boundary in the Ohanet (07 Oh) section. |
|
Figure 13 : Corrélations entre diversité spécifique, renouvellement spécifique et séquence d'ouverture des ostracodes, en relation avec la fréquence de pyrite, du gypse et du quartz, et la courbe eustatique au passage Cénomanien/Turonien de la coupe d'In Adaoui. Correlation between species diversity,
species turn-over and marine opening set of ostracodes, in relation to the
occurrence of pyrite, gypsum and quartz, and the sea-level curve around the
Cenomanian/Turonian boundary in the In Adaoui section. |
Ce renouvellement est marqué par la disparition des espčces cénomaniennes et leur remplacement par de nouvelles espčces turoniennes. Dans le bassin du Tinrhert, l'absence d'ostracodes dans les dépôts du Cénomanien supérieur ne permet pas d'établir leur renouvellement au passage Cénomanien/Turonien. Toutefois, cela reste envisageable pour le Turonien inférieur.
Les premičres eaux transgressives dans le bassin du Tinrhert auraient été appauvries en oxygčne, expliquant ainsi l'absence d'ostracodes ŕ la base du Turonien inférieur. La remontée relative continue de la mer aurait amélioré les conditions environnementales qui auraient alors facilité le redéploiement du benthos.
6. B. Rapport ostracodes ornés/ostracodes lisses
Cubaynes (1986) a utilisé ce rapport afin d'établir une séquence d'ouverture ŕ l'océan au moyen des ostracodes du Lias du Quercy (bassin d'Aquitaine, France). Cette séquence s'accroît avec la remontée du niveau marin.
Les résultats obtenus dans le bassin du Tinrhert sont analogues ŕ ceux établis par Cubaynes (1986). Ce rapport montre les taux les plus élevés ŕ la base du Turonien inférieur, ŕ l'occasion du maximum transgressif. Une tendance régressive s'amorce trčs nettement dčs la partie médiane du Turonien inférieur, période pendant laquelle le bassin du Tinrhert montre une affinité océanique plus restreinte. Elle coďncide également avec l'arrivée des apports terrigčnes qui atténuent alors son caractčre marin carbonaté.
Dans le bassin du Tinrhert, le Cénomanien-Turonien est marqué par quatre épisodes sédimentaires, ŕ savoir :
une sédimentation argileuse, riche en gypse et pyrite, datée du Cénomanien inférieur. Elle indique un milieu lagunaire, envahi périodiquement par des dépôts de tempętes. Ce terme est uniquement visible dans la région de Bordj Omar Driss,
une sédimentation de plate-forme carbonatée au Cénomanien supérieur, marquée par une dalle calcaire, trčs fossilifčre, dont le microfacičs montre des 'filaments',
une sédimentation essentiellement marneuse au Turonien inférieur-moyen, dans laquelle se développe la microfaune benthique, notamment les ostracodes et
de nouveau, une sédimentation de plate-forme carbonatée, suite ŕ la régression relative et au comblement du bassin, au Turonien supérieur.
L'étude paléo-écologique des ostracodes met en évidence l'installation d'une zone néritique peu profonde, souvent défavorable au développement de la microfaune benthique.
Les ostracodes sont présents uniquement dans la partie médiane du Turonien inférieur-moyen, suite ŕ la remontée continue du niveau marin et ŕ une meilleure oxygénation des eaux de fond. Au Cénomanien, ils auraient été amenés depuis le large, par immersion occasionnelle, et seraient restés 'confinés' dans le lagon.
La présence et/ou absence des ostracodes, leur répartition et leur diversité démontrent leur étroite dépendance aux conditions environnementales. Ainsi, la fréquence du quartz, gypse, pyrite et les variations eustatiques influent directement sur :
leur fréquence relative,
leur renouvellement,
leur diversité spécifique et
la fréquence relative des ostracodes ornés et des ostracodes lisses.
Les auteurs tiennent ŕ remercier les rapporteurs anonymes pour leurs critiques constructives et leurs précieux conseils sur la version préliminaire de ce texte, ainsi que le rédacteur en chef, Bruno Granier, et le comité éditorial des Carnets Geol. pour leur soutien et leur aide dans la finalisation de cette publication.
Abdallah H., Memmi L., Damotte R., Rat P., Magniez-Jannin F. (1995).- Le Crétacé de la chaîne nord des Chotts (Tunisie du centre-sud) : Biostratigraphie et comparaison avec les régions voisines.- Cretaceous Research, vol. 16, n° 5, p. 487-538.
Al-Abdul-Razzaq S. (1980).- New Eocytheropteron species with reversed valve structure.- Micropaleontology, New York, vol. 26, p. 444-448.
Al-Abdul-Razzaq S. & Grosdidier E. (1981).- Ostracode index species from the Cenomanian of the South Shelf of the Tethys Sea.- Bulletin des Centres de Recherche Exploration-Production elf-Aquitaine, Pau, vol. 5, p. 173-191.
Al-Furaih A.A.F. (1983).- Middle Cretaceous (Cenomanian) Ostracoda from the Wasia Formation of Saudi Arabia.- Paleontological Contributions of the University of Kansas, Lawrence, vol. 108, p. 1-6.
Amédro F., Busson G. & Cornée A. (1996).- Révision des ammonites du Cénomanien supérieur et du Turonien inférieur du Tinrhert (Sahara algérien) : Implications biostratigraphiques.- Bulletin du Muséum national d'Histoire naturelle, Paris, vol. 18, p. 179-232.
Andreu B. (1993).- Associations d'Ostracodes des marges téthysiennes et atlantique marocaines de l'Albien au Turonien.- Géobios, Villeurbanne, vol. 26, p. 69-84.
Andreu-Boussut B. (1991).- Les Ostracodes du Crétacé moyen (Barrémien ŕ Turonien), le long d'une transversale Agadir - Nador (Maroc).- Strata, Toulouse, vol. 14, 765 p.
Arias C. (1995).- Los Ostracodos del Toarciense inferior en la Cordillera Ibérica.- Ph.D. Thesis, Universidad Complutense de Madrid, 521 p.
Athersuch J. (1988).- The biostratigraphy of Cretaceous Ostracods from Oman. In : Hanai T., Ikeya N. & Ishizaki K. (eds.), Evolutionary biology of Ostracoda; its fundamentals and applications. Proceedings of the 9th International Symposium on Ostracoda (Shizuoka, 1985).- Developments in paleontology and stratigraphy, Amsterdam, vol. 11, p. 1187-1206.
Athersuch J. (1994).- The biostratigraphic significance of Cretaceous ostracods from the Arabian Gulf. In : Simmons M.D. (ed.), Micropaleontology and hydrocarbon exploration in the Middle East.- Chapmann et Hall, London, p. 253-265.
Babinot J.-F. (1985).- Paléobiogéographie des ostracodes du Crétacé supérieur des marges ouest-européennes et nord-africaines de la Téthys.- Bulletin de la Société géologique de France (8e série), Paris, t. 1, n° 5, vol. 8, p. 739-754.
Babinot J.-F. & Basha S.H. (1985).- Ostracods from the Early Cenomanian of Jordan, a preliminary report.- Géobios, Villeurbanne, vol 18, p. 257-162.
Babinot J.-F. & Colin J.P. (1988).- Paleobiogeography of Tethyan Cretaceous marine ostracods. In : Hanai T., Ikejya N. & Ishizaki K. (eds.), Evolutionary biology of Ostracoda.- Developments in Palaeontology and Stratigraphy, vol. 11, p. 823-839.
Babinot J.-F. & Colin J.-P. (2011).- Barremian ostracods from the Serre de Bleyton (Drôme, SE France).- Annalen des Naturhistorischen Museums in Wien, vol. 113, p. 735-775.
Babinot J.-F. & Crumičre-Airaud C. (1990).- The effect of global events on the evolution of Cenomanian and Turonian marginal Tethyan ostracod faunas in the Mediterranean region. In : Whatley R. & Maybury C. (eds.), Ostracoda and global events.- Chapman and Hall, London, p. 25-39.
Bassoullet J.-P. & Damotte R. (1969).- Quelques ostracodes nouveaux du Cénomano-Turonien de l'Atlas Saharien occidental (Algérie).- Revue de Micropaléontologie, Paris, vol. 12, p. 130-144.
Benson R.H. (1977).- 36. The Cenozoic ostracode faunas of the Săo Paulo plateau and the Rio Grande rise (DSDP leg 39, sites 356 and 357). In : Perch-Nielsen K., Supko P.R., Neprochnov Y.P., Zimmerman H.B., McCoy F., Kumar N., Thiede J., Bonatti E., Fodor R., Boersma A., Dinkelman M.G. & Carlson R.-L. (eds.).- Initial Reports of Deep Sea Drilling Project, Washington, vol. XXXIX, p. 869-883.
Ben Youssef M. (1980).- Étude stratigraphique et micropaléontologique du Crétacé des Djebels Koumine et Kharroub.- Thčse de Doctorat de 3čme Cycle, Université de Nice, 104 p.
Bischoff G. (1963).- Die Gattung Cythereis in der Unterkreide. Ostracoden-Studien in Libanon 1.- Senckenbergiana lethaea, Frankfurt-am-Main, vol. 44, p. 1-77.
Bismuth H., Boltenhagen C., Donze P., Lefevre J. & Saint-Marc P. (1981a).- Le Crétacé Moyen et Supérieur du Djebel Semmama (Tunisie du Centre-Nord) ; Microstratigraphie et évolution sédimentologique.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 5, p. 213-245.
Bismuth H., Donze, P., Lefčvre J. & Saint-Marc P. (1981b).- Nouvelles espčces d'ostracodes dans le Crétacé Moyen et Supérieur du Djebel Semmama (Tunisie du Centre-Nord).- Cahiers de Micropaléontologie, Paris, vol. 3, p. 51-69.
Bodergat A.-M., Rio M. et Ikeya N. (1997).- Tide versus eutrophication, impact on ostracod populations structure of Mikawa Bay (Japan).- Revue de Micropaléontologie, Paris, vol. 40, p. 3-13.
Bonnet L., Andreu B., Rey J., Cubaynes R., Ruget C., N'Zaba-Makaya O. & Brunei F. (1999).- Fluctuations of environmental factors as seen by means of statistical analyses in micropaleontological assemblages from a Liassic series.- Micropaleontology, New York, vol. 45, p. 399-417.
Busson G. (1960).- Sur la coupe du Crétacé supérieur et de l'Éocčne inférieur du Tinrhert central (Sahara algérien).- Travaux de Recherches sahariennes, Alger, vol. 19, p. 141-149.
Busson G. (1969).- Sédimentation, transgression et paléogéographie sur les grandes plates-formes du Mésozoďque : L'exemple du Cénomanien Turonien du Nord-Est de la plate-forme saharienne et de Berbérie.- Bulletin de la Société géologique de France (7e série), Paris, t. XI, n° 5, p. 687-703.
Busson G. (1971).- Principes, méthodes résultats d'une étude stratigraphique du Mésozoďque saharien.- Thčse Doctorat, Université de Paris VI, 466 p.
Busson G., Dhondt A., Amédro F., Néraudeau D. & Cornée A. (1999).- La grande transgression du Cénomanien supérieur-Turonien inférieur sur la Hamada de Tinrhert (Sahara algérien) : Datations biostratigraphiques, environnement de dépôt et comparaison d'un témoin épicratonique avec les séries contemporaines ŕ matičre organique du Maghreb.- Cretaceous Research, vol. 20, p. 29-46.
Butler E.A. & Jones D.E. (1957).- Cretaceous Ostracoda of Prothro and Rayburns salt domes Bienville Parish, Louisiana.- Bulletin of Geology, Baton Rouge, vol. 32, 165 p.
Carbonel P., Mourguiart P. & Peypouquet J.-P. (1988).- Le polymorphisme induit par l'environnement : Rôle du rythme saisonnier.- Strata, Toulouse, vol. 4, p. 1-12.
Choffat P. (1898).- Recueil d'études paléontologiques sur la faune crétacique du Portugal. Les Ammonées du Bellasien des couches ŕ Neolobites vibrayeanus, du Turonien et du Sénonien.- Commission des Travaux géologiques du Portugal, Lisboa, vol. 2, p. 41-86.
Chudeau R. (1909).- Ammonites du Damergou (Sahara méridional).- Bulletin de la Société géologique de France, Paris, vol. 4, p. 67-71.
Colin J.-P., Platel J.-P., Roger J. & Tambareau Y. (2001).- Nouvelles données sur les faunes d'Ostracodes de l'Albien et du Cénomanien d'Oman : Taxonomie et paléobiogéographie. In : Weiss R.-H. (ed.), Contributions to geology and palaeontology of Gondwana.- Geological Institute, University of Cologne, p. 89-109.
Collignon M. (1965).- Atlas des fossiles caractéristiques de Madagascar (Ammonites). XII (Turonien).- Service géologique, Tananarive, Madagascar, 82 p.
Colwell R.K., Rahbek C. & Gotelli N.J. (2004).- The mid-domain effect and species richness patterns: What have we learned so far.- American Naturalist, Chicago, vol. 163, p. E1-E23.
Cubaynes R. (1986).- Le Lias du Quercy méridional : Étude lithologique, biostratigraphique, paléoécologique et sédimentologique.- Strata, Toulouse, vol. 6, p. 159-481.
Damotte R. & Saint-Marc P. (1972).- Contribution ŕ la connaissance des ostracodes Crétacé du Liban.- Revue Espagnole de Micropaléontologie, Madrid, vol. 4, p. 273-296.
Damotte R. (1985).- Les Ostracodes du Crétacé moyen sud-mésogéen et leur répartition paléogéographique.- Bulletin de la Société géologique de France (8e série), Paris, t. 1, n° 5, p. 733-737.
Damotte R. (1995).- The biostratigraphy and paleobiogeography of the Upper Cretaceous-basal Tertiary ostracods from North Africa, Mali and Congo.- Cretaceous Research, vol. 16, p. 35-366.
Depęche F. (1984).- Les ostracodes d'une plate-forme continentale au Jurassique. Recherches sur le Bathonien du Bassin parisien.- Thčse Doctorat d'État, Université Pierre Marie Curie, Paris, 325 p.
EWGPF (1979).- In : Dabu E.-T. (2005).- Lower Turonian at the Breaza-Comarnic Highway Bridge Pie/rfeet (Prahova Valley).- Acta Palaeontologica Romaniae, Bucarest, vol. 5, p. 87-93.
Ferré B., Benyoucef M., Zaoui D., Adaci M., Tchenar S., Meister C., Mebarki K., Piuz A. & Bensalah M. (2016).- Cenomanian-Turonian roveacrinid microfacies assemblages (Crinoidea, Roveacrinida) from the Tinrhert area (SE Algeria).- Annales de Paléontologie, Paris, vol. 102, p. 225-235.
Forster A., Schouten S., Moriya K., Wilson P.A. & Sinninghe Damsté J.S. (2007).- Tropical warming and intermittent cooling during the Cenomanian/Turonian oceanic anoxic event 2: Sea surface temperature records from the equatorial Atlantic.- Paleoceanography and Paleoclimatology, vol. 22, no. 1, P1219, p. 1-14.
Gargouri-Razgallah S. (1983).- Le Cénomanien de Tunisie centrale, étude paléoécologique, stratigraphique, micropaléontologique et paléogéographique.- Thčse Doctorat čs Sciences, Université Claude Bernard, Lyon, 215 p.
Gebhardt H. (1999).- Cenomanian to Coniacian biogeography and migration of North and West African ostracods.- Cretaceous Research, vol. 20, p. 215-229.
Gerry E. & Rosenfeld R. (1973).- Amphicytherura distincta and Neocyprideis vandenboldi (Ostracoda), new species from the Cenomanian-Turonian of Israel.- Revista espańola de Micropaleontología, Madrid, vol. 5, p. 99-105.
Gotelli N.J. & Colwell R.K. (2001).- Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness.- Ecology Letters, Oxford, vol. 4, p. 379-391.
Grosdidier E. (1973).- Associations d'Ostracodes du Crétacé d'Iran.- Revue de l'Institut Français de Pétrole, Rueil-Malmaison, vol. 28, p. 131-168.
Haq B.-U., Hardenbol J. & Vail P.-R. (1987).- Chronology of fluctuating sea levels, since the Triassic.- Science, Washington, p. 1156-1165.
Jennings P.-H. (1936).- A microfauna from the Monmouth and basal Rancocas groups of New Jersey.- Bulletin of American Paleontology, New York, vol. 23, p. 161-234.
Jukes-Browne A.-J. (1896).- VI. Critical remarks on some of the fossils. In : Jukes-Browne A.J. & Hill H. (eds.), A delimitation of the Cenomanian: Being a comparison of the corresponding beds in southwestern England and northern France.- Quarterly Journal of the Geological Society of London, vol. 52, p. 99-178.
Kazi-Tani N. (1986).- Évolution géodynamique de la bordure nord africaine : Le domaine intraplaque nord algérien, approche séquentielle.- Thčse de Doctorat, Université de Pau, 3 volumes, 871 p.
Kuhnt W., Herbin J.-P., Thurow T. & Wiedman J. (1990).- Distribution of Cenomanian-Turonian organic facies in the western Mediterranean and along the adjacent Atlantic margin. In : Huc A.-Y. (ed.), Depositional of organic facies.- AAPG Studies in Geology, vol. 30, p. 133-160.
Kuypers M.M.M., Pancost R.D., Nijenhuis I.A. & Sinninghe Damsté J.S. (2002).- Enhanced productivity led to increased organic carbon burial in the euxinic North Atlantic basin during the Late Cenomanian oceanic anoxic event.- Paleoceanography, Oxford, vol. 17, p. 1-13.
Layeb M. (1990, inédit).- Étude géologique, géochimique et minéralogique, régionale, des facičs riches en matičre organique de la formation Bahloul d'âge Cénomano-Turonien dans le domaine de la Tunisie Nord-Centrale.- Thčse 3e cycle, Université de Tunis II, 209 p.
Layeb M. & Belayouni H. (1999).- Paléogéographie de la Formation Bahloul (passage Cénomanien - Turonien). In : Les septičmes journées de la géologie tunisienne - Tunis (2-3 Décembre 1999).- Annales des Mines et de la Géologie, Tunis, n° 40, p. 21-44.
Luger P. (2003).- Paleobiogeography of late Early Cretaceous to Early Paleocene marine Ostracoda in Arabia and North to Equatorial Africa.- Palæogeography, Palæoclimatology, Palæoecology, vol. 196, p. 319-342.
Majoran S. (1988).- Comments on a miscellaneous ostracod group from the mid-Cretaceous of the south shelf of the Tethys sea.- Journal of African Earth Sciences, vol. 6, p. 691-702.
Majoran S. (1989).- Mid-Cretaceous Ostracoda of northeastern Algeria.- Fossils and Strata, Oslo, vol. 27, p. 1-67.
Majoran S. (1996).- Mid-Cretaceous "Veeniacythereis" (Ostracoda) from Africa and the Middle East.- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 3, p. 183-192.
Mantell G.A. (1822).- The fossils of the South Downs, or Illustrations of the geology of Sussex.- Lupton Relfe, London, 327 p.
Mao J.-W., Li X.-F., Zhang Z.-H. & Hu H.-B. (2004).- Mesozoic telluride rich gold deposits in eastern China. In : Gold-Silver-Telluride Deposits of the Golden Quadrilateral, South Apuseni Mts., Romania.- IAGOD Guidebook Series, London, vol. 12, p. 232-235.
Morkhoven F.P.C.M. van (1963).- Post-Paleozoic ostracoda. Their morphology, taxonomy and economic use.- Elsevier Publishing Company, vol. 2, 478 p.
Nzoussi-Mbassani P. (2003).- Le Cénomano-Turonien de l'Atlantique nord (bassin du Sénégal) : Environnement de dépôt et évolution diagénétique. Implications pétroličres.- Thčse de Doctorat, Université d'Orléans, 223 p.
Orbigny A. d' (1840-1842).- Paléontologie française : Description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de la France. Terrains crétacés. Tome premier, Céphalopodes.- Masson éditeur, Paris, p. 1-120 (1840) ; p. 121-430 (1841) ; p. 431-662 (1842).
Peypouquet J.-P., Ducasse O. & Rousselle L. (1981).- Morphogenesis and environment. Theoretical and practical aspect from Hammatocythere: Paleogene Ostracods of the Aquitaine Basin. In : Martinell J. (ed.), International Symposium on Concepts and Methods in Paleontology.- Departamento de Paleontologia, Universitat de Barcelona, p. 173-187.
Pokorny V. (1964).- Conchoecia? cretacea n. sp., first fossil species of the family Halocyprididae (Ostracoda, Crustacea).- Acta Universitatis Carolinae - Geologica, Prague, vol. 2, p. 175-180.
Reuss A.E. (1846).- Die Versteinerungen der Böhmischen Kreideformation.- Schweizerbart, Stuttgart, vol. 2, p. 59-148.
Roemer F.A. (1841).- Die Versteinerung des nord deutschen Kreide Gebirges.- Hahnsche Hofbuchhandlung, Hannover, 145 p.
Rosenfeld, A. (1981).- Cytherella gigantosulcata (Ostracoda), a replacement for Cytherella sulcata Rosenfeld, 1974.- Journal of Paleontology, Cambridge, vol. 55, p. 896.
Rosenfeld A. & Bein A. (1978).- A preliminary note on recent ostracodes from shelf to rise sediments off north-east Africa.- Meteor Forschungsergebnisse (Reihe C), Stuttgart, vol. 29, p. 14-20.
Rosenfeld A. & Raab M. (1974).- Cenomanian-Turonian Ostracods from the Judea Group in Israel.- Bulletin of Geologic Survey of Israel, Jerusalem, vol. 62, p. 1-64.
Schlanger S. & Jenkyns H.C. (1976).- Cretaceous oceanic anoxic events: Causes and consequences.- Geologie en Mijnbouw, vol. 55, p. 179-194.
Schlanger S.O., Arthur M.A., Jenkyns H.C. & Scholle A. (1987).- The Cenomanian - Turonian Oceanic Event, I. Stratigraphy and distributions of organic-rich beds and the marine δ13C excursion.- Special Publications of Geologic Society of London, vol. 26, p. 371-399.
Schulze F., Marzouk A.M., Bassiouni A.A. & Kuss J. (2004).- The late Albian-Turonian carbonate platform succession of west-central Jordan: Stratigraphy and crisis.- Cretaceous Research, vol. 25, p. 709-737.
Sepkoski J.J. (1986).- In : Sepkoski J.J. (1989).- Periodicity in extinction and the problem of catastrophism in the history of life.- Journal of the Geological Society, London, vol. 146, p. 7-19.
Soua M. (2005, inédit).- Biostratigraphie de haute résolution des foraminifčres planctoniques du passage Cénomanien Turonien et impact de l'évčnement anoxique EAO-2 sur ce groupe dans la marge sud de la Téthys. Exemple des régions de Jerissa et Bargou.- Mémoire de Mastčre, Université de Tunis El Manar, 169 p.
Szczechura J., Abd-Elshafy E. & Babinot J.-F. (1991).- Late Albian to Early/Mid-Cenomanian Ostracods from Northern Galala Plateau, Egypt.- Acta Paleontologica Polonica, Varsaw, vol. 36, p. 3-38.
Tchenar S. (2004, inédit).- Étude des ostracodes des séries liasiques des Monts des Traras (Mellala, Algérie nord occidentale).- Mémoire de Magister, Université d'Oran, 137 p.
Tchenar S. (2016, inédit).- Évčnements sédimentologiques et évolution de la microfaune des séries cénomano-turoniennes en Algérie - Bassin du Tinrhert, SE algérien.- Thčse Doctorat en Sciences, Université de Tlemcen, 227 p.
Tchenar S., Zaoui D., Adaci M., Bensalah M., Madani B., Kentri T. & Fekih N. (2015).- Le passage Cénomanien-Turonien dans le Bassin du Tinrhert (Sahara algérien) : Lithologie et répartition de la microfaune.- Bulletin du Service géologique de l'Algérie, Alger, vol. 26, p. 23-41.
Tölderer-Farmer M. (1985).- Causalité des variations morphologiques de la carapace chez les ostracodes : Essai d'interprétation sur des populations actuelles et fossiles.- Thčse de Doctorat de 3e Cycle, Université de Bordeaux I, 285 p.
Tronchetti G. & Grosheny D. (1991).- Les assemblages de foraminifčres benthiques au passage Cénomanien-Turonien ŕ Vergons, S-E France.- Géobios, Villeurbanne, vol. 24, p. 13-31.
Whatley R. (1995).- Ostracoda and oceanic palaeoxygen levels.- Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, Hambourg, vol. 92, p. 337-353.
Vivičre J.-L. (1985).- Les Ostracodes du Crétacé supérieur (Vraconien ŕ Campanien basal) de la région de Tébessa (Algérie du Nord-Est) : Stratigraphie, paléoécologie, systématique.- Mémoires des Sciences de Terre, Université Marie Curie, Paris, vol. 85, 261 p.
Weaver P.P.E. (1982).- Ostracoda from the British Lower Chalk and Plenus Marl.- Palaeontographical Society Monographs, London, vol. 562, 127 p.
Woods H. (1911).- The palaeontology of the Upper Cretaceous deposits of northern Nigeria.- Bulletin of Geosciences, vol. 62, p. 63-89.
Zaoui D., Tchenar S., Benyoucef M., Meister C., Adaci M., Piuz A., Mebarki K., Bensalah M., Gabani A. & Mahboubi M. (2016).- Le Cénomano-Turonien dans la Hamada du Tinrhert (Sahara, Algérie) : Résultats préliminaires.- Revue de Paléobiologie, Genčve, vol. 35, p. 541-559.