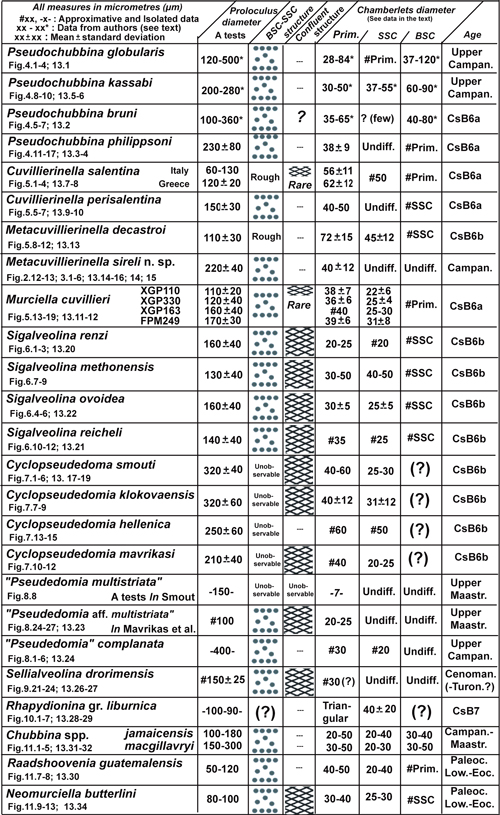
◄ Carnets Geol. 20 (9) ►
![]()
Outline:
[1. Introduction]
[2. Geological srtting]
[3. Test architecture and endoskeleton of the Rhapydioninidae]
[4. Discovering the new taxon]
[5. Looking for the Basal Secondary Chamberlets-Scattered Secondary Chamberlets (BSC-SSC) structure]
[6. The teaching of the search]
[7. Formal establishment of the new taxon]
[8. Conclusion: Distinctive criteria in a prolific family]
[Bibliographic references] and...
[Appending note: A lexicon adapted to the Rhapydioninidae]
118 avenue de Flandre, F-59290 Wasquehal (France)
Turkish Petroleum Corporation (TPAO), Research and Development (Ar-Ge) Center, 06530 Çankaya, Ankara (Turkey)
Published online in final form (pdf) on May 11, 2020
DOI 10.4267/2042/70793
![]()
[Editor: Bruno Granier; language editor: Simon Mitchell]
![]()
The family Rhapydioninidae is a part of the superfamily Alveolinacea. The main
characteristic of this superfamily is its endoskeleton with each chamber
divided into tubular chamberlets,
fundamentally parallel to the coiling direction, and only connected in an
undivided space located in the anterior part of the chamber: The preseptal space. The
family Rhapydioninidae is distinguished by the coexistence of two sets of
chamberlets: Primary chamberlets,
which are isolated by partitions ("cloisonnettes"),
forming a unique layer at the chamber periphery, and secondary chamberlets pierced in a more or less compact mass, the
central
endoskeleton (by no way homologous of the "couche basale", sometimes
called flosculinisation or columella in some Alveolinidae). Two particular
modes of organization of the secondary chamberlets occur, the Basal
Secondary Chamberlets-Scattered
Secondary Chamberlets structure
(BSC-SSC) and the Confluent
structure; they constitute
supplementary features that distinguish this family from other groups.
The BSC-SSC
structure (a new name for a previously well-known organization of
chamberlets in the genus Pseudochubbina
and Cuvillierinella salentina) is the object of a large inventory
undertaken among the known taxa of the Rhapydioninidae. It leads to the
observation that this particular endoskeleton is found in the various
subfamilies on both sides of the Atlantic and cannot be used as a feature of
taxonomic significance within the group. However, it is not observed in
apparently "primitive" taxa equipped with chamberlets of large
isodiametric diameter which display a "fishnet" appearance. The confluent structure is a new name for the helicoidal structure,
which is also widespread within the family.
Metacuvillierinella
sireli n. sp.,
of Campanian age, is described from outcrop and subsurface limestones in
southeast Anatolia, Turkey. The new taxon is a Rhapydioninidae based on its
test architecture and endoskeleton. As a species, it is clearly distinct
because of its initial planispiral coiling of A generation tests, its both
pseudoplanispiral generations with an advolute final stage and its thin
chamberlets showing an obvious BSC-SSC structure. The generic attribution
appears more uncertain: The faint dimorphism between generations and the
persisting pseudoplanispiral-advolute final stage are only known in the genus Metacuvillierinella.
But M. decastroi, the type species,
displays a small proloculus in the A forms, a miliolid juvenile stage, and an
endoskeleton of "fishnet" appearance (cryptic BSC-SSC structure), which
give it a particular character, appearing as being "primitive". This
contrasts with the relatively large proloculus in the A forms,
pseudoplanispiral coiling and the obvious BSC-SSC structure of the new taxon.
Thus, all these features being subjected to evolution, the faint generational
differences and the pseudoplanispiral-advolute coiling seem sufficient to
suggest the affinities between the two taxa. The new taxon is, nevertheless,
clearly more "advanced", which could be interpreted as a clue for a higher
standing, possibly compatible with a new genus. This is not undertaken here,
in consideration of the unknown "radiance" (small variations in several
well-disseminated populations and/or other species of the same kind) of the
new taxon with the present state of knowledge.
Additionally,
with a review of the BSC-SSC structure, the various genera of the family
Rhapydioninidae are revisited, namely Pseudochubbina,
Cuvillierinella, Murciella, Sigalveolina, Cyclopseudedomia, Sellialveolina,
Rhapydionina, Fanrhapydionina, Chubbina, Praechubbina, Raadshoovenia,
Neomurciella, Twaraina; special attention is reserved to the Euro-Asiatic
genus Pseudedomia, of which the
original material and, consequently, the consecutive interpretative
identifications, appear doubtful. New sections of Subalveolina dordonica and Fleuryana
adriatica are figured.
The
conclusion deals mainly with the criteria used for distinguishing various
systematic levels within the family. The classical differentiation between "specific" and "generic"
characters,
if eventually convenient for simple or inadequately known groups, seems
unsuited for a complex and well known family like this one.
A more pragmatic mode of working is proposed,
using any character as a simple element without any meaning by itself, but to
be understood and interpreted among the others, that is to say in the
evolutionary perspective of the whole group.
A lexicon
of the used terms in Rhapydioninidae and closely related taxa in given in an
appendix.
• Foraminifera;
• Alveolinacea;
• Rhapydioninidae;
• Upper Cretaceous;
• Campanian-Maastrichtian;
• Mediterranean area;
• Turkey;
• new species
Fleury J.-J. & Özkan R. (2020).- Metacuvillierinella sireli n. sp., a Campanian Rhapydioninidae (Foraminifera), from southeast Turkey. New considerations on the endoskeleton and particularities of the family, with a specialized lexicon.- Carnets Geol., Madrid, vol. 20, no. 9, p. 165-213.
Metacuvillierinella sireli
n. sp., un nouveau Rhapydioninidae (Foraminifčres) du sud-ouest de la Turquie,
occasion de nouvelles observations sur l'endosquelette et les
particularités
de la famille, avec un lexique spécialisé. La famille des
Rhapydioninidae est une partie de la superfamille des Alveolinacea. La
principale caractéristique de cette superfamille est constituée par son
endosquelette, divisant chaque loge en logettes tubulaires ordinairement
orientées parallčlement ŕ la direction d'enroulement, ne communicant que
dans un espace indivis situé dans la partie antérieure des loges :
L'espace préseptal. Les Rhapydioninidae sont en partie particularisés par
la compression du test dans le plan équatorial et par leur tendance au déroulement
final, contrairement aux Alveolinidae, la famille sśur, qui sont allongés
axialement et ne se déroulent jamais. Les Rhapydioninidae sont en outre
distingués par la coexistence de deux types de logettes : Les logettes
primaires, séparées par les cloisonnettes, formant une unique couche dans la
partie périphérique des loges, et les logettes secondaires constituées par
deux ensembles : Les "Logettes Secondaires Basales" (BSC)
formant une couche accolée au tour précédent et les "Logettes
Secondaires Dispersées" (SSC) percées dans une masse plus ou moins compacte,
l' "endosquelette central" (nullement homologue de la "couche
basale", parfois nommée flosculinisation ou columelle, de certains
Alveolinidae). La présence de piliers préseptaux joignant l'endosquelette
central au septe au travers de l'espace préseptal ainsi que les deux modes
particuliers d'organisation des logettes secondaires : Structure des "BSC-SSC" (et sa variante
"filet de pęche") et "structure confluente"
constituent encore des traits distinctifs de la famille ; ils sont analysés
ci-dessous.
Une nouvelle espčce campanienne, Metacuvillierinella sireli n. sp., est
décrite, en provenance des
calcaires de la formation Sanli, l'unité terminale supposée du groupe
Adiyaman connu dans la région de Mardin, en Turquie (Anatolie sud-orientale).
Le nouveau taxon est un Rhapydioninidae typique par l'architecture de son
test et son endosquelette. C'est une évidente nouvelle espčce par son
enroulement initial planispiralé des tests A, ses deux générations
pseudoplanispiralées ŕ stade final advolute et ses fines logettes trahissant
un endosquelette de type "BSC-SSC". Son attribution générique est plus
douteuse : Le faible dimorphisme de générations et l'enroulement
advolute des tests ne sont connus que chez le genre Metacuvillierinella, alors
que l'organisation de l'endosquelette, non observée chez le type de ce
genre (M. decastroi), rappelle
certains taxons oů cette structure est bien identifiée, tels que Pseudochubbina et Cuvillierinella
perisalentina. Un inventaire général mené au sein des Rhapydioninidae
montre que cette organisation est largement répandue dans toutes les
sous-familles des deux côtés de l'Atlantique et ne peut ętre considérée
comme un critčre distinctif fondamental au sein du groupe; l'un de ses
attributs, l'existence d'une couche de logettes secondaires basales (BSC)
reste cependant indiscernable, pour des raisons géométriques, chez les
taxons comportant des logettes secondaires de fort diamčtre, comme chez M. decastroi en
particulier. Ce critčre, dont l'observation ne dépend
que de la taille des logettes, ne permet donc pas de discriminer
fondamentalement le nouveau taxon de M.
decastroi, dont il serait un descendant, bien qu'il s'en différencie
encore par la grande taille relative de son proloculus A et le stade initial
non miliolin des tests de génération A ; ces critčres, eux-męmes
susceptibles d'interprétation, ne paraissent pas suffisants pour une
distinction d'ordre générique, qui ne pourrait se justifier qu'en
fonction du "rayonnement" que pourrait présenter le nouveau
taxon, par sa dissémination propre ou celle sa parenté-descendance.
Accessoirement, ŕ l'occasion de la recherche effectuée
pour la reconnaissance de la structure "BSC-SSC", tous les genres connus de
la famille dans l'Ancien et du Nouveau Monde sont reconsidérés: Pseudochubbina,
Cuvillierinella, Murciella, Sigalveolina, Cyclopseudedomia, Sellialveolina,
Rhapydionina, Fanrhapydionina, Chubbina, Praechubbina, Raadshoovenia,
Neomurciella, Twaraina. Une attention particuličre est prętée au genre
Euro asiatique Pseudedomia, dont le type et, en conséquence, les interprétations
consécutives, apparaissent peu fiables. De nouvelles sections de Subalveolina
dordonica et Fleuryana adriatica sont figurées.
La conclusion porte principalement sur les critčres de
distinction des divers niveaux systématiques au sein de la famille. On
n'accorde aucun crédit aux classiques distinctions entre caractčres dits "spécifiques" et
"génériques". Un mode de
travail plus pragmatique est proposé, considérant chaque caractčre comme dépourvu
de signification par lui-męme mais nécessitant d'ętre compris et interprété
parmi les autres, c'est-ŕ-dire dans la perspective évolutive de
l'ensemble du groupe.
On propose en appendice un lexique adapté aux
Rhapydioninidae et aux concepts plus ou moins directement associés ŕ cette
famille.
• foraminifčres ;
• Alveolinacea ;
• Rhapydioninidae ;
• Crétacé supérieur ;
• Campanien-Maastrichtien ;
• région méditerranéenne ;
• Turquie ;
• nouvelle espčce
The present work follows three recent papers by Fleury (2014, 2016 and 2018), which bring together most of the previous observations that have been made on the family Rhapydioninidae from the Western Mediterranean area. The reader will find structural investigations on almost all known genera and species, exemplified by studies of several populations of each taxon, which are thoroughly illustrated and described. An attempt to develop a biostratigraphic zonation (CsB zones, first designated by Fleury, 1980) of these taxa was also presented for the Gavrovo-Tripolitza Platform in Greece (Fleury, 2018), where the succession is interrupted by three probable subaerial exposure periods, which limit the observed stratigraphic distribution of the taxa. Thus, this zonation has to be tested in other areas, particularly in the eastern Mediterranean region. The discovery of a new taxon of typical Rhapydioninidae from Turkey gives the first opportunity to begin this task: The taxon occurs in an internal platform limestones situated between two pelagic episodes with globotruncanids. This organism is new, but not without affinities with taxa from further west.
The encountering of a new taxon is always an adventure. The knowledge of a group being never completely understood, any new member gives the opportunity to revise and reappraise the criteria usually admitted for identification at every systematic level. The new species is a typical case of this kind. Although undoubtedly new, it is made up of the association of several combined equivocal features requiring a careful examination of their meaning. Thus, we will be led to revisit the apparently well-known species of the various genera comprising the family Rhapydioninidae. A main surprise arises on the way: The Basal Secondary Chamberlets-Scattered Secondary Chamberlets (BSC-SSC) structure, previously well identified in rare genera (Pseudochubbina, Cuvillierinella), although relatively cryptic in some cases, appears as generalized among members of the family, in the Cretaceous and Cenozoic of the Euro-Asiatic and American provinces.
Turkey, as a part of the
Alpine-Himalayan orogenic belt, is geologically subdivided into three main
tectonic units: The Pontides, the Anatolides-Taurides and the Arabian
Platform. These continental fragments or terranes developed during the
Mesozoic and Cenozoic geologic evolution of the Paleo- and the Neo-Tethysian
systems (Ketin, 1966; Şengör
& Yılmaz, 1981; Okay
& Tüysüz, 1999). Southeast Anatolia, which represents
sedimentary sequences from the Paleozoic to Cenozoic, is located on the
northern Arabian Platform and bounded to the north by the Anatolides-Taurides
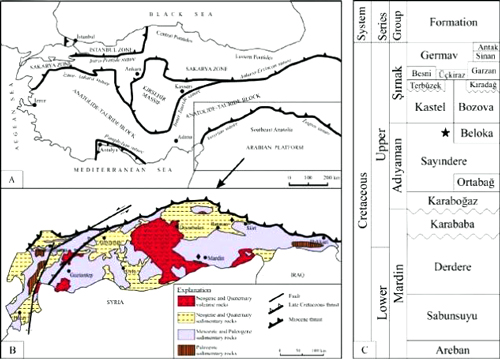
block along the Assyrian and Zagros suture zones (Fig. 1.A ![]() ). The region is mainly covered by Mesozoic and Cenozoic
rocks (Fig. 1.B
). The region is mainly covered by Mesozoic and Cenozoic
rocks (Fig. 1.B ![]() ).
).
From a lithostratigraphic point
of view, the Cretaceous sequence is represented predominately by carbonates
and is divided into three groups, which, stratigraphically from base to top,
are the Mardin, Adıyaman and Şırnak groups (Tuna,
1974; Sungurlu, 1974; Perinçek,
1980; Güven
et al., 1991; Perinçek
et al., 1991; Yılmaz & Duran,
1997; Özkan & Altiner,
2019) (see Fig. 1.C ![]() ). The
Mardin Group comprises the Areban,
Sabunsuyu, Derdere and Karababa
formations. The basal Areban Formation consists mainly of clastic deposits
with thin limestone interbeds. The overlying Sabunsuyu and Derdere formations
are composed mostly of limestones and dolostones with some clastic deposits.
The top unit, the Karababa Formation, is made up mostly of pelagic limestones.
The deposits of the Mardin Group were constrained to have been deposited in a
time interval from the Aptian to Santonian (Özkan
& Altiner, 2019). The Adıyaman
Group, of Campanian age (Güven
et al., 1991; Yılmaz
& Duran, 1997), is
subdivided into four formations: The lowest Karaboğaz Formation is
characterized by chert-bearing pelagic limestones rich in organic matter. The
following Ortabağ and Sayındere formations are composed mainly of
limestones with some clastics of deeper marine environments. The Beloka
Formation, a lateral equivalent unit of Sayındere Formation, and
comprises mostly bioclastic limestones that were deposited in a shallow-marine
environment. The Şırnak Group,
which spans the late Campanian to Maastrichtian (Güven
et al., 1991; Perinçek et al.,
1991; Yılmaz & Duran,
1997), includes Kastel, Bozova, Germav, Üçkiraz, Besni, Garzan and Sinan
formations that are formed of mixed carbonates and clastic deposits of marine
environments. The group also contains the Terbüzek, Kıradağ and
Antak formations that are characterized by terrigenous sediments.
). The
Mardin Group comprises the Areban,
Sabunsuyu, Derdere and Karababa
formations. The basal Areban Formation consists mainly of clastic deposits
with thin limestone interbeds. The overlying Sabunsuyu and Derdere formations
are composed mostly of limestones and dolostones with some clastic deposits.
The top unit, the Karababa Formation, is made up mostly of pelagic limestones.
The deposits of the Mardin Group were constrained to have been deposited in a
time interval from the Aptian to Santonian (Özkan
& Altiner, 2019). The Adıyaman
Group, of Campanian age (Güven
et al., 1991; Yılmaz
& Duran, 1997), is
subdivided into four formations: The lowest Karaboğaz Formation is
characterized by chert-bearing pelagic limestones rich in organic matter. The
following Ortabağ and Sayındere formations are composed mainly of
limestones with some clastics of deeper marine environments. The Beloka
Formation, a lateral equivalent unit of Sayındere Formation, and
comprises mostly bioclastic limestones that were deposited in a shallow-marine
environment. The Şırnak Group,
which spans the late Campanian to Maastrichtian (Güven
et al., 1991; Perinçek et al.,
1991; Yılmaz & Duran,
1997), includes Kastel, Bozova, Germav, Üçkiraz, Besni, Garzan and Sinan
formations that are formed of mixed carbonates and clastic deposits of marine
environments. The group also contains the Terbüzek, Kıradağ and
Antak formations that are characterized by terrigenous sediments.
The rocks hosting Metacuvillierinella
sireli n. sp. are here attributed to Sanlı Formation, even though
this formation is not included in "Stratigraphic Lexicon of southeast
Anatolia" (Yılmaz & Duran,
1997). The Sanlı Formation, known as the "Murciella-bearing
unit", was defined by Çelikdemir
& Dülger (1990) in the
Mardin region for exposures of light-gray, beige, cream-white-colored
limestones. Due to a lack of precise age determination, the stratigraphic
position of this formation within the stratigraphic framework of southeast
Anatolia has been in question. It has been thought to be a lateral equivalent
of the Karababa Formation, the top unit of the Mardin Group sequence, which,
however, was determined to range in age from Aptian through Santonian (Özkan
and Altiner, 2019). In this study, the Sanlı Formation is confirmed to be
Campanian, probably late Campanian, and is proposed to be included in the Adıyaman
Group. It is considered a lagoonal equivalent of the Beloka Formation (see Fig.
1.C ![]() ).
).
|
Figure 1:
A: Tectonic map of Turkey showing the main tectonic
units and sutures (Okay and Tüysüz,
1999). B: Geological map of the
southeast Anatolia (Yılmaz, 1993). C: Generalized columnar
section of the Cretaceous units in the southeast Anatolia (Yılmaz and
Duran, 1997). (*: Sample location;
Stratigraphic location of Metacuvillierinella
sireli n. sp.). |
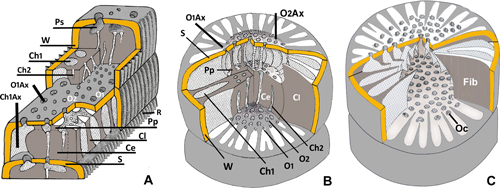
The nomenclature used here was introduced by various authors who have definitively left their mark on the superfamily Alveolinacea, namely P. De Castro, É. Fourcade, L. Hottinger, M. Reichel and A.H. Smout. They are followed here as far as possible, but with increasing knowledge it is sometimes necessary to choose between their divergent options or to ignore part of their contributions. They will mainly be cited in cases where there are unsolved conflicts between different opinions and the actual observations. Anyway, the present work, resulting from a long acquaintance with the group, is an attempt to propose a homogeneous terminology (in large part used by Fleury since 1974), which is missing in classical textbooks or treatises. It is more precisely presented in the final lexicon in the Appendix.
Following Reichel (1936-1937), authors distinguish the exoskeleton ("forming the shell or carapace") from the endoskeleton ("internal deposits"), although they are at the same time independent (various mode of coiling are associated with various types of internal organization) and interdependent (distribution, orientation and even presence of internal elements depend on the shape of chambers and even in the location of the chamber within the test, i.e., the BSC layer is only present in involute part of tests and all secondary chamberlets can be absent in flanges of strongly compressed tests).
3.1 - Exoskeleton (architecture of the test)
The living animal is isolated from the surrounding water by an unperforated porcelaneous wall. The test is divided into successive chambers (more than two in each coil) separated by septa (singular: septum), pierced by openings connecting successive chambers (foramina) and the last one to the outer environment (apertures), without morphological modification. They correspond to the tubular chamberlets (see below: endoskeleton) of the following chamber.
Like many Foraminifera, the family Rhapydioninidae shows a dimorphism of generations; it is restricted to the mode of coiling, the endoskeleton organization remaining unchanged. The megalospheric generation ("A" tests) is ordinary smaller, simpler and less uncoiled than the microspheric one ("B" tests). Various examples can be seen in Fleury, 2018, text-fig. 16.
A
schematic view of successive stages of development of a theoretical A test is
given in Fig. 2.1 ![]() ; the ordinary
succession is as follows:
; the ordinary
succession is as follows:
A first
chamber, ordinary subspherical, is called the proloculus (also
megalosphere in A tests and microsphere in B tests). It is relatively large in
A tests, but very small and rarely observed in B test. In A tests, a tubular
canal (the flexostyle) connects the proloculus to the first ordinary
chamber (see Fig. 2.7 ![]() ):
):
In the
nepionic stage, the chambers are in many cases arranged in a streptospiral
involute coil; miliolid-like or apparently irregular in section around
the proloculus (i.e., genus Cuvillierinella,
see Fig. 2.5 ![]() ). This stage is
sometimes absent in A tests of more advanced genera, wholly planispiral
genera, such as Murciella, Sigalveolina and Cyclopseudedomia
(Figs. 5
). This stage is
sometimes absent in A tests of more advanced genera, wholly planispiral
genera, such as Murciella, Sigalveolina and Cyclopseudedomia
(Figs. 5 ![]() , 6
, 6 ![]() and
7
and
7 ![]() ).
).
The next stage, which is still involute, is either streptospiral or planispiral, depending on genus and even species. This is an evolutionary character: with streptospiral transitioning to planispiral as time goes by, even in a single population. The best example is given by the type population of Cuvillierinella salentina (in Fleury, 2016, text-fig. 5) showing the two modes of coiling independently and variously associated with more or less advanced endoskeletal organization.
In the
adult stage, the chambers tend to adopt a planispiral mode of coiling, involute
at first, then possibly evolute,
forming an "Uncoiled
Uniserial Termination" (abbreviated UUT afterwards), either cylindrical (Fig.
2.3 ![]() ), or bilaterally flattened, flabelliform (Fig.
2.1 and 2.4
), or bilaterally flattened, flabelliform (Fig.
2.1 and 2.4 ![]() ), pseudoevolute when
chambers keep in contact with the involute part, evolute when they become
free. Exceptionally, the evolute chambers may produce an annular stage,
forming a discoidal test (B tests of Cyclopseudedomia
smouti, see Fleury,
2018, text-fig. 11). Another exception is shown by "Pseudedomia" complanata, in which the last chambers of the
pseudoevolute final stage cover the
axial part of the involute young stage (see Fig. 8.5-6 and 8.8
), pseudoevolute when
chambers keep in contact with the involute part, evolute when they become
free. Exceptionally, the evolute chambers may produce an annular stage,
forming a discoidal test (B tests of Cyclopseudedomia
smouti, see Fleury,
2018, text-fig. 11). Another exception is shown by "Pseudedomia" complanata, in which the last chambers of the
pseudoevolute final stage cover the
axial part of the involute young stage (see Fig. 8.5-6 and 8.8 ![]() ); this particular and single example of this mode
of coiling in the Rhapydioninidae is called here "hyperinvolute" or
"archaiasiform" (by analogy with the
soritid genus Archaias).
); this particular and single example of this mode
of coiling in the Rhapydioninidae is called here "hyperinvolute" or
"archaiasiform" (by analogy with the
soritid genus Archaias).
Two particular modes of adult coiling, both observed in the new taxon, have to be considered:
The
streptospiral coiling of the juvenile stage can be more or less preserved in
the adult. Some taxa never reach a perfect planispiral coiling stage, such as Chubbina
(Fig. 11.3 ![]() ), Pseudochubbina
(Fig. 4.2 and 4.15
), Pseudochubbina
(Fig. 4.2 and 4.15 ![]() ), Metacuvillierinella
decastroi (Figs. 3.7
), Metacuvillierinella
decastroi (Figs. 3.7 ![]() and 5.10
and 5.10 ![]() )
and the new taxon (Fig. 3.2 and 3.5
)
and the new taxon (Fig. 3.2 and 3.5 ![]() ):
In every case, even if some sections seem
planispiral, others
maintain the previous streptospiral stage with twisted or sigmoidal axial
sections resulting from a poorly stabilized coiling axis. This particular case
is called "pseudoplanispiral" coiling.
):
In every case, even if some sections seem
planispiral, others
maintain the previous streptospiral stage with twisted or sigmoidal axial
sections resulting from a poorly stabilized coiling axis. This particular case
is called "pseudoplanispiral" coiling.
Some
species adopt another particular mode of coiling: Although the tests are never
evolute, never uncoiled, they present a last coil which does not cover
entirely the preceding one, from which a large umbilicus results. This
particular mode of coiling is called "advolute"
(Fig. 2.2 ![]() ). It is known in Metacuvillierinella
decastroi (Fig. 3.7-9 and 3.11
). It is known in Metacuvillierinella
decastroi (Fig. 3.7-9 and 3.11 ![]() ) and in the new taxon (Fig.
3.1-2 and 3.5-6
) and in the new taxon (Fig.
3.1-2 and 3.5-6 ![]() )
)
|
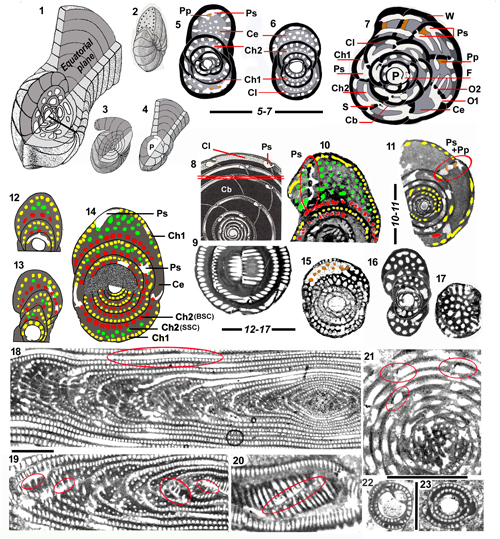
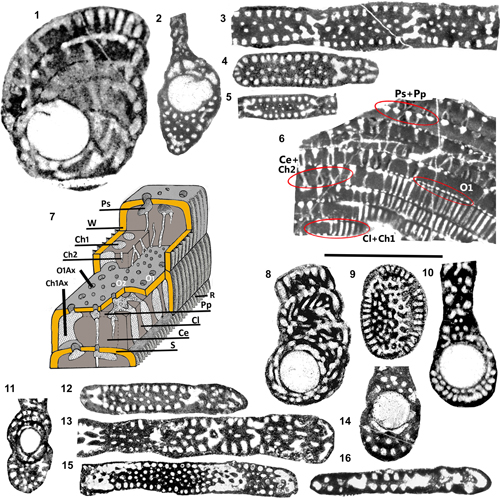
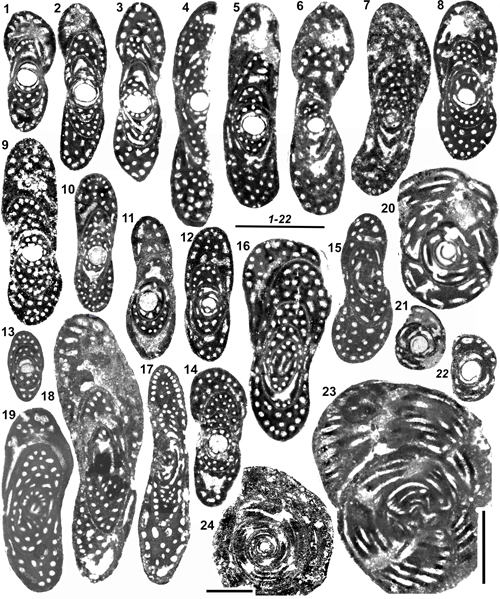
Figure 2:
1-17: Overall
architecture and endoskeletal organization of the Rhapydioninidae. 1-4: Architecture of the tests. 1: Fictitious test combining the main
architectural features of the family: a streptospiral initial part around the A
proloculus followed by planispiral parts, involute, pseudoevolute, then evolute.
2: External aspect of a planispiral or pseudoplanispiral test with an advolute
last coil, resulting in a large umbilicus, but no uncoiling (aspect of M.
sireli n. sp.). 3: Fully planispiral involute test with cylindrical
uniserial uncoiled termination (UUT). 4: Fully planispiral test, with large
evolute terminal flange. 5-7: Terminology
of the endoskeletal elements. 5: Cuvillierinella
salentina; 6-7: C. perisalentina. 8-11:
Comparison between the "couche basale" of sub-spherical Alveolina
sp. (8-9: Equatorial and subaxial sections, with approximate position of
section 9 on 8) and central endoskeleton
of Pseudochubbina bruni (10-11:
Red oval surrounding the preseptal space; central endoskeleton partly micritized
in 11). 12-13: Axial sections of Metacuvillierinella
sireli n. sp. (see Fig. 14.8
and 14.14 |
3.2 - Endoskeleton (internal organization of chambers)
Some
generalized structural schematic reconstructions of the endoskeleton can be
seen in Fleury, 2016, text-fig. 3. The main elements of the ordinary
internal organization of chambers ("structure coaxiale" in Fleury, 2018)
are the cloisonnettes, isolating a unique
layer of primary chamberlets (in the sense of the first materialized and
sometimes remaining the only ones), always oriented in the spiral direction,
forming the peripheral zone beneath the wall. The deeper part of chamber is
occupied by the central endoskeleton, a
more or less compact mass appearing in the space unoccupied by the layer
of primary chamberlets and cloisonnettes when the growing distance between two
successive coils exceeds its thickness. Apparently, this structure is formed
from the fusing of the distal part of the cloisonnettes (see Fleury,
2018, text-fig. 2.16-24); it is pierced by more or less disordered secondary
chamberlets (in the sense of accessory, or subsidiary, lately
produced, sometimes absent), described below. All chamberlets (protoplasmic
columns) merge into an empty space preceding the septum, the preseptal space (preseptal
passage or canal are both
more adapted to Alveolinidae morphology).
This space is nevertheless divided in its periphery by the termination
of the cloisonnettes (triangular in section) which join the septum at the
periphery (i.e., Fleury,
2016, text-fig. 3.E-F; 2018, text-figs. 11.12, 12.16 and 14.26; Fig.
7.16 ![]() ) and in its center by pillars joining the central endoskeleton to the
septum (i.e., Fig. 2.5, 2.7 and 2.15
) and in its center by pillars joining the central endoskeleton to the
septum (i.e., Fig. 2.5, 2.7 and 2.15 ![]() last chamber): These are the preseptal pillars. Each
chamberlet corresponds precisely to an opening
hollowed out in the previous septum; the axis of primary openings being always oblique and slightly shifted from
the corresponding primary chamberlets
(i.e., Fleury, 2014, text-fig. 1.H; 2016, text-figs. 3.C and 7.2;
2018, text-fig. 11.13; Fig. 2.7
last chamber): These are the preseptal pillars. Each
chamberlet corresponds precisely to an opening
hollowed out in the previous septum; the axis of primary openings being always oblique and slightly shifted from
the corresponding primary chamberlets
(i.e., Fleury, 2014, text-fig. 1.H; 2016, text-figs. 3.C and 7.2;
2018, text-fig. 11.13; Fig. 2.7 ![]() :
Several chambers of last coil), while the secondary
openings are directly connected to the following secondary
chamberlets.
:
Several chambers of last coil), while the secondary
openings are directly connected to the following secondary
chamberlets.
This apparatus is peculiar to the Rhapydioninidae, it is roughly resembling the well-known organization of the Alveolinidae, but with some nuance and difference in nomenclature and concepts:
The cloisonnettes were first named by Reichel (1936-1937) in order to characterize the short partitions perpendicular to the wall, which isolate a superficial layer of tubular chamberlets, arranged in the direction of coiling, the primary chamberlets; they are specific to the Alveolinacea and distinguish them from the Soritacea (as seen in DNA investigations of extant species: Holzmann et al., 2001). That is why we do not follow Reichel (1964) who tried to translate the term cloisonnettes into "septula", a general term used in many groups for any secondary dividing wall, without any particular meaning.
The floors
(blades parallel to the surface, delimiting layers of chamberlets) are a
translation of the French "planchers" ("lames parallčles ŕ la
surface, ils délimitent les couches de logettes" in
Reichel, 1936-1937), imported
from the Alveolinidae. This term corresponds effectively to the appearance of
the first division between layers of primary and secondary chamberlets in some
axial (i.e., Figs. 2.15 ![]() , 5.16, 5.18
, 5.16, 5.18 ![]() , 9.13
, 9.13 ![]() and 11.1
and 11.1 ![]() ,
inner coils) and equatorial sections (Figs.
5.13
,
inner coils) and equatorial sections (Figs.
5.13 ![]() and 9.11
and 9.11 ![]() ). But
most of the sections in adult tests with well-developed central endoskeleton
elements do not show this structure. The generalized true organization of the
secondary chamberlets hollowed out in the central endoskeleton corresponds to
the Basal Secondary Chamberlets-Scattered Secondary Chamberlets structure; it
is described below (see §3.3), thoroughly analyzed and illustrated in the
whole group (see §5).
). But
most of the sections in adult tests with well-developed central endoskeleton
elements do not show this structure. The generalized true organization of the
secondary chamberlets hollowed out in the central endoskeleton corresponds to
the Basal Secondary Chamberlets-Scattered Secondary Chamberlets structure; it
is described below (see §3.3), thoroughly analyzed and illustrated in the
whole group (see §5).
The central endoskeleton. This structure comprises the space of the chambers unoccupied by the layer of primary chamberlets, the domain of the secondary chamberlets. Its ordinary organization is described by the Basal Secondary Chamberlets-Scattered Secondary Chamberlets (BSC-SSC) structure (see below). Two variants can be recognized:
A "fishnet"
pattern results from the common diameter of all sets of primary and
secondary chamberlets, separated by thin dividing walls. It is mainly seen in
less specialized taxa, such as Cuvillierinella
and Murciella (Figs. 2.16-17 ![]() and 5.8-9
and 5.8-9 ![]() ).
).
The confluent
structure corresponds to a particular organization of the secondary
chamberlets (BSC excluded). This is a new name (in
Fleury, 2018) for the previous "structure hélicoďdale"
of Fleury (1979), widely
represented among the Rhapydioninidae (see Fleury,
2018, text-fig. 3; Figs. 6.2, 6.6,
6.9, 6.12 ![]() , 7.6
, 7.6 ![]() ,
8.27
,
8.27 ![]() , 9.25
, 9.25 ![]() , 11.13
, 11.13 ![]() and 16
and 16 ![]() ). Rather than an attempt to describe their special setting in a
theoretical case (layers of chamberlets interpenetrated in a cylindrical
chamber), the new name reflects the conjunction and interpenetration of nearly
chamberlets more or less orthogonally oriented, making possible the mixing of
protoplasm without using necessarily the preseptal space. Such a device is
supposed to have favored the free movement of symbionts in their search for
the best light conditions. It will not be the main object of the following
review, but its eventual relation with the BSC-SSC structure will be
considered.
). Rather than an attempt to describe their special setting in a
theoretical case (layers of chamberlets interpenetrated in a cylindrical
chamber), the new name reflects the conjunction and interpenetration of nearly
chamberlets more or less orthogonally oriented, making possible the mixing of
protoplasm without using necessarily the preseptal space. Such a device is
supposed to have favored the free movement of symbionts in their search for
the best light conditions. It will not be the main object of the following
review, but its eventual relation with the BSC-SSC structure will be
considered.
3.3 - The Central Endoskeleton and the Basal Secondary Chamberlets-Scattered Secondary Chamberlets (BSC-SSC) structure: Comparison with the "couche basale" (="basal layer") of the Alveolinidae.
The central endoskeleton is the name given to the domain of the secondary chamberlets resembling superficially, and sometimes confused, with the "couche basale" of the Alveolinidae in its various meanings. Thus, a detailed comparison is necessary.
The "Basal Secondary Chamberlets-Scattered Secondary Chamberlets" (BSC-SSC) structure is intended to describe the general organization of secondary chamberlets. The model is given by the distribution of chamberlets in several species of the genus Pseudochubbina meticulously described by De Castro (1990), and well observable in Cuvillierinella perisalentina (in Fleury, 2016), for which the term Scattered Secondary Chamberlets was created.
The complex detailed terminology used by De Castro cannot be generalized to all genera without damage to nomenclatural homogeneity in the family, but the sequence of the following stages was perfectly distinguished in the text and figures of this author. Around the proloculus, the sections of Pseudochubbina show the following stages:
Stage
with one layer of chamberlets: The primary chamberlets (yellow in Fig.
2.10-14 ![]() );
);
Stage
with two layers of chamberlets: Primary and secondary chamberlets, the last
layer forming the "BSC" (red in Fig. 2.10-14 ![]() );
);
Stage
with the previous two layers of chamberlets and at first a few, and then
numerous, intercalary scattered chamberlets "SSC" (green in Fig.
2.10-14 ![]() ).
).
This
sequence is observable in the sections of the new taxon M. sireli (Fig.
2.12-13 ![]() ), although stage 2 is hidden, because the loose coiling induces a
difference in shape from the first to second coils which creates a wide space
where scattered chamberlets appear as soon as the beginning of the second
coil.
), although stage 2 is hidden, because the loose coiling induces a
difference in shape from the first to second coils which creates a wide space
where scattered chamberlets appear as soon as the beginning of the second
coil.
The "Basal Secondary Chamberlets" (BSC) correspond to the appearance of new chamberlets in stage 2, made up of a layer of well calibrated chamberlets, close to one another, forming a chain parallel to the wall of the preceding coil. This feature persists in the following stages but is missing in chambers of pseudoevolute-evolute stages, indicating that it is probably a purely geometric character, possibly not fundamentally involved in the biologic functioning of the cell. The search for this particular layer will be the main concern of the following review among the whole family.
The "Scattered Secondary Chamberlets" (SSC) term refers specifically to the existence of dispersed chamberlets in stages 3.
The "couche
basale" in its primitive sense was defined by Reichel
(1936-1937) as the deposit laid out on the wall of the preceding coil, forming
the internal coating of the chamber ("dépôt plus ou moins épais qui
s'étend sur la surface du tour précédent et constitue le revętement
interne des loges"). In that sense there is effectively always a "couche
basale" in the Rhapydioninidae tests (see Fig. 2.7 ![]() ), but usually so thin that it is not mentioned (in such case, it could
be called "couche basale sensu stricto"). This French term was
subsequently used in English publications, but was translated by Reichel himself (1964) to "basal layer or basal thickening", mainly to characterize the "enormous basal thickening in
several internal whorls" (with particular reference to "Flosculina",
a synonym of Alveolina, see Smout,
1963, p. 224-225 and Reichel,
1964, p. C506-509). Thus the pair of names couche
basale-basal layer was applied both to the general case (couche basale s.s.)
and the particular example of spherical Alveolina
with thickening equally developed at the base of each whole chamber (Fig.
2.8-9
), but usually so thin that it is not mentioned (in such case, it could
be called "couche basale sensu stricto"). This French term was
subsequently used in English publications, but was translated by Reichel himself (1964) to "basal layer or basal thickening", mainly to characterize the "enormous basal thickening in
several internal whorls" (with particular reference to "Flosculina",
a synonym of Alveolina, see Smout,
1963, p. 224-225 and Reichel,
1964, p. C506-509). Thus the pair of names couche
basale-basal layer was applied both to the general case (couche basale s.s.)
and the particular example of spherical Alveolina
with thickening equally developed at the base of each whole chamber (Fig.
2.8-9 ![]() ). In this example, the thick basal layer is
1) restrained to some
coils, not necessarily the last ones; 2) usually almost deprived of irregular
canals; 3) directly in contact with the previous and following septa, without
interposition of the preseptal space; 4) at the base of chamber, without
interposition of any chamberlets (BSC absent).
). In this example, the thick basal layer is
1) restrained to some
coils, not necessarily the last ones; 2) usually almost deprived of irregular
canals; 3) directly in contact with the previous and following septa, without
interposition of the preseptal space; 4) at the base of chamber, without
interposition of any chamberlets (BSC absent).
More recently, Hottinger (2006, p. 8 and text-fig. 18.H) insisted on an apparent different case, after examining the axially elongate Alveolina tenuis Hott. "showing columella produced by polar thickening of the basal layer" (...) with (...) "tubular passages in the columella, continuous in subsequent chambers, without interruption by preseptal spaces". In this case, the thickening is regularly increasing from the first to last coils and pierced by numerous "tubular passages" which contrasts with the previous example. But the common points 3-4 observed in both cases are decisive when compared with the central endoskeleton as it was defined: The preseptal space occupies the total height of chamber and basal secondary chamberlets are distinctive of this particular structure.
Summarizing,
whatever the case, the lack of both the preseptal space between the couche
basale of two successive chambers and the absence of basal secondary
chamberlets (BSC) in the Alveolinidae indicates there is no supposed homology
between the thickened couche basale of this group and central endoskeleton as
it is known in Pseudochubbina and
the new taxon (Fig. 2.10-14 ![]() ). The
presence of a preseptal space beneath the whole surface of each septum is well
known in the Rhapydioninidae, but demonstrating the general presence of basal
secondary chamberlets in all taxa of the family will be done after a first
consideration of the new taxon.
). The
presence of a preseptal space beneath the whole surface of each septum is well
known in the Rhapydioninidae, but demonstrating the general presence of basal
secondary chamberlets in all taxa of the family will be done after a first
consideration of the new taxon.
The new taxon is known by a rather homogeneous set of sections of bilateral compressed tests, usually larger than 1 mm in diameter, with an average elongation index of about 3. Two groups of sections are easily distinguished.
The
smaller ones (between 1 and 2 mm in diameter) are numerous (Fig.
3.1-4 ![]() ). Many of them, in axial section, show a rounded proloculus,
followed by a test of about 3 to 4 coils, relatively loose and mainly involute,
although the last coil seems to cover only a part of the preceding, showing a
large umbilicus: This is the advolute mode of coiling. The coiling is
apparently planispiral but, as a whole, the tests are slightly twisted in
axial sections: This is the pseudoplanispiral mode of coiling. These are
evidently A tests of a new taxon.
). Many of them, in axial section, show a rounded proloculus,
followed by a test of about 3 to 4 coils, relatively loose and mainly involute,
although the last coil seems to cover only a part of the preceding, showing a
large umbilicus: This is the advolute mode of coiling. The coiling is
apparently planispiral but, as a whole, the tests are slightly twisted in
axial sections: This is the pseudoplanispiral mode of coiling. These are
evidently A tests of a new taxon.
The
larger ones (up to 4 or 5 mm) are rare and mainly known by non-centered
subaxial sections (Fig. 3.5-6 ![]() ).
They never show a proloculus of the preceding type, but no precisely centered
section is visible in our material. They resemble the previous in their
central coiled parts but develop a loose last coil which doubles the diameter
of the preceding part. The last coil is clearly tending to cover only a part
of the preceding, illustrating the advolute mode of coiling. The coiling of
the adult is almost planispiral too, but the overall aspect is fairly twisted,
sometimes strongly contorted: This is the pseudoplanispiral mode of coiling.
These are evidently B tests of the same previous taxon.
).
They never show a proloculus of the preceding type, but no precisely centered
section is visible in our material. They resemble the previous in their
central coiled parts but develop a loose last coil which doubles the diameter
of the preceding part. The last coil is clearly tending to cover only a part
of the preceding, illustrating the advolute mode of coiling. The coiling of
the adult is almost planispiral too, but the overall aspect is fairly twisted,
sometimes strongly contorted: This is the pseudoplanispiral mode of coiling.
These are evidently B tests of the same previous taxon.
The
endoskeletal structure is the same in both types of tests. In axial sections
(see Fig. 3.2 ![]() and 3.5-6
and 3.5-6 ![]() ;
especially Fig. 2.12-13
;
especially Fig. 2.12-13 ![]() ), the successive chambers display a layer of regularly
spaced chamberlets (the primary ones) and, less evident, another layer near
the wall of previous coils (the BSC). Between these two layers, the solid mass
of the central endoskeleton is pierced by scattered holes which are shown as
corresponding to tubular chamberlets in equatorial sections (the SSC). These
sections show that this structure is discontinuous, interrupted by the
preseptal space crossed by rare pillars.
), the successive chambers display a layer of regularly
spaced chamberlets (the primary ones) and, less evident, another layer near
the wall of previous coils (the BSC). Between these two layers, the solid mass
of the central endoskeleton is pierced by scattered holes which are shown as
corresponding to tubular chamberlets in equatorial sections (the SSC). These
sections show that this structure is discontinuous, interrupted by the
preseptal space crossed by rare pillars.
Every reader is certainly aware that the above simplified description is matching an alveolinid and specially a Rhapydioninidae, represented by its two generations.
Some may
have thought that it resembles Metacuvillierinella
decastroi (Figs. 3.7-11 ![]() and
5.8-12
and
5.8-12 ![]() ). In fact, the adult mode of coiling of the advolute type is the
same, and the slight difference between the two generations of both species is
almost identical. But M. decastroi has large chamberlets among which the various kinds are
not easily distinguished.
). In fact, the adult mode of coiling of the advolute type is the
same, and the slight difference between the two generations of both species is
almost identical. But M. decastroi has large chamberlets among which the various kinds are
not easily distinguished.
Others may have recognized the previously described BSC-SSC endoskeleton well known in the various species of Pseudochubbina and Cuvillierinella perisalentina.
The new organism, characterized by an original coiling and an obvious BSC-SSC structure, is thus well distinguished from all others in the family but the question arises of the BSC-SSC meaning. Is it a special structure which indicates a close relationship between the new taxon and Pseudochubbina, for example, or is it a widespread structure shared by several groups, or is it an evolutionary feature, latent in the genetic heritage and potentially present in all members of the family? The generic attribution of the new organism depends in part on the answer to this question. Only a review of the taxa revealing this structure can give the answer.
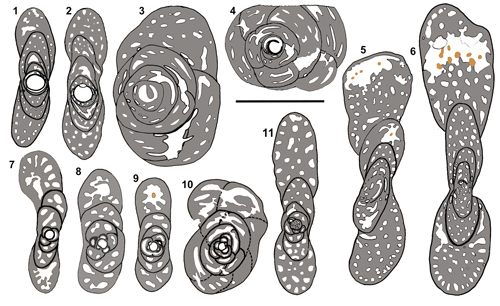
|
Figure 3: Metacuvillierinella decastroi
and
Metacuvillierinella sireli n. sp.,
interpretative drawings for comparison. 1-6:
M. sireli n. sp. 1-4: Axial and
equatorial sections of A tests (see Fig. 14.2-3 and 14.20 |
The
following exploration through the Rhapydioninidae is quite superficial,
concentrating only on the main general characters and endoskeletal
organization in order to reveal the generality of the BSC-SSC structure and to
explain the apparent exceptions. Significant details of the descriptions are
collected together in Fig. 12 ![]() .
.
We follow an order directed by geographic nearness and assumed affinities between taxa.
The Euro-Asiatic taxa examination begins with the Cuvillierinellinae, more or less precisely linked to Cuvillierinella (see Fleury, 2016, text-fig. 13), the first being Pseudochubbina, resembling the new taxon by its endoskeleton; the following are Pseudedomiinae, Sellialveolininae, quite isolated and deserving deeper comments, then finally the Rhapydionininae, particularized by their endoskeleton.
The American taxa are more superficially evocated, although the presence of the BSC-SSC structure confirms it as a fundamental character of the Rhapydioninidae and reinforces the generally supposed kinship between its members on both sides of the Atlantic Ocean.
5.1 - Genus Pseudochubbina De Castro, 1990
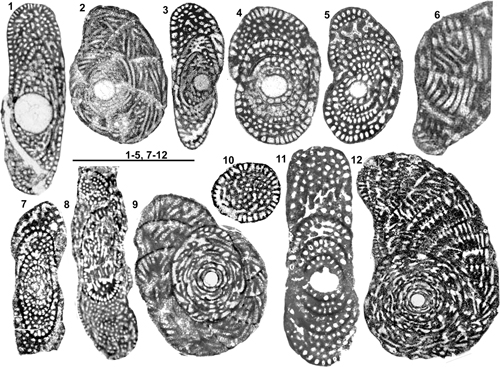
The type species of the genus Pseudochubbina chosen by De Castro was previously known as "Pseudedomia" globularis Smout, 1963, from the Campanian of Iraq. It was then ascribed to Pseudedomia Henson, 1948, the first genus to be recognized as characterized by an association of a true alveolinid endoskeleton elements with an uncoiled test of soritid type (Eames & Smout, 1955). At that time, the other members of the future Rhapydioninidae family were ignored, and any taxon having this combination was open to the same generic attribution; "Pseudedomia" drorimensis Reiss et al., 1964 (a true Sellialveolina as we will see) is another example of this situation. The context was radically changed when De Castro (1972) showed that Rhapydionina Stache, 1913, although showing a differently mode of coiling, was sharing the same type of endoskeletal organization; the way to the understanding of the Rhapydioninidae was open.
In that
perspective, De Castro
(1990), benefactor of new material from the type region of "Pseudedomia" globularis, and discovering in southern Italy a
related species, was able to undertake the revision of this taxon, under the
new genus Pseudochubbina, to which
four species were ascribed. They share globular to nautiloid involute A tests,
pseudoplanispiral as a whole, giving the genus a rather evident homogeneity; a
possible final flabelliform flange exists, apparently prepared in every case
by a sudden looser coiling of late chambers. The B tests, larger and entirely
involute, are only probable in two species (Fig. 4.4 and 4.17 ![]() ,
non-centered sections); they would provide a definitive character for the
genus if confirmed. Thus, the following lines will only concern A tests.
,
non-centered sections); they would provide a definitive character for the
genus if confirmed. Thus, the following lines will only concern A tests.
Pseudochubbina
globularis (Smout, 1963), Fig.
4.1-4 ![]() ,
was well described and illustrated by
Smout, and luxuriously illustrated again by De Castro (1990) based
on new material from Iraq, the type region. The A tests are subspherical to
ovoid, rarely biombilicate (Fig. 4.2
,
was well described and illustrated by
Smout, and luxuriously illustrated again by De Castro (1990) based
on new material from Iraq, the type region. The A tests are subspherical to
ovoid, rarely biombilicate (Fig. 4.2 ![]() );
their diameter ranges from 2 to 2.7 mm, up to 4 mm. We emphasize the sudden
increase in height of the last coil (the holotype of Smout, and Fig.
4.1
);
their diameter ranges from 2 to 2.7 mm, up to 4 mm. We emphasize the sudden
increase in height of the last coil (the holotype of Smout, and Fig.
4.1 ![]() ),
expressing a final tendency to the development of uncoiled A tests; the
sigmoid aspect of axial sections of some tests is to be noticed, indicating
that the coiling axis remains poorly stabilized, which is pseudoplanispiral
coiling (Fig. 4.2
),
expressing a final tendency to the development of uncoiled A tests; the
sigmoid aspect of axial sections of some tests is to be noticed, indicating
that the coiling axis remains poorly stabilized, which is pseudoplanispiral
coiling (Fig. 4.2 ![]() ). The proloculus
diameter is 120 to 500 µm, after De
Castro. The preseptal space is relatively narrow, occupying the whole
height of the chambers, it is equipped with thin and numerous preseptal
pillars (Fig. 4.1 and 4.3
). The proloculus
diameter is 120 to 500 µm, after De
Castro. The preseptal space is relatively narrow, occupying the whole
height of the chambers, it is equipped with thin and numerous preseptal
pillars (Fig. 4.1 and 4.3 ![]() ). The endoskeleton
is typically of the BSC-SSC type, characterized by chamberlets of various
diameters. According to De Castro,
the primary ones (28-84 µm) are much smaller than those forming the BSC
(37-120 µm), but the scattered ones, when present (in the last chambers of
larger tests), are about the same diameter as the primary ones (Figs.
4.1
). The endoskeleton
is typically of the BSC-SSC type, characterized by chamberlets of various
diameters. According to De Castro,
the primary ones (28-84 µm) are much smaller than those forming the BSC
(37-120 µm), but the scattered ones, when present (in the last chambers of
larger tests), are about the same diameter as the primary ones (Figs.
4.1 ![]() and 13.1
and 13.1 ![]() ).
).
Pseudochubbina
kassabi De Castro, 1990, Fig.
4.8-10 ![]() , was found in association with P.
globularis, in De Castro's
material from Iraq. The most important feature is an early tendency to unroll,
giving rise to a large evolute or pseudoevolute final flange; the overall
diameter of the test is up to 15 mm. The proloculus diameter is between 200
and 280 µm (for 3 specimens). The preseptal pillars seem rather thick (Fig.
4.9-10
, was found in association with P.
globularis, in De Castro's
material from Iraq. The most important feature is an early tendency to unroll,
giving rise to a large evolute or pseudoevolute final flange; the overall
diameter of the test is up to 15 mm. The proloculus diameter is between 200
and 280 µm (for 3 specimens). The preseptal pillars seem rather thick (Fig.
4.9-10 ![]() ). The diverse chamberlets appear of uniform diameter in the final
flange (Fig. 4.10
). The diverse chamberlets appear of uniform diameter in the final
flange (Fig. 4.10 ![]() ), but after De
Castro, the primary ones are thinner than the scattered ones (30-50
and 37-55 µm, respectively), those of the BSC are well differentiated and
larger than the others (60-90 µm) (see Fig. 4.9
), but after De
Castro, the primary ones are thinner than the scattered ones (30-50
and 37-55 µm, respectively), those of the BSC are well differentiated and
larger than the others (60-90 µm) (see Fig. 4.9 ![]() ).
This
species, being known by only a few sections, is not well characterized and
stands a little apart from the others. In particular, the mode of coiling, and
the differences between the two generations are still unclear. A resemblance
with Cyclopseudedomia cannot be
denied, especially given the large A test proloculus and the marked tendency
to form a large uncoiled flabelliform final stage, at least in the A
generation. Such characters may correspond to a simple convergence and are
probably not sufficient to build a theory or to imagine a real kinship between
the two taxa, but must not be forgotten when comparing their territories (Iraq
and Periadriatic area).
).
This
species, being known by only a few sections, is not well characterized and
stands a little apart from the others. In particular, the mode of coiling, and
the differences between the two generations are still unclear. A resemblance
with Cyclopseudedomia cannot be
denied, especially given the large A test proloculus and the marked tendency
to form a large uncoiled flabelliform final stage, at least in the A
generation. Such characters may correspond to a simple convergence and are
probably not sufficient to build a theory or to imagine a real kinship between
the two taxa, but must not be forgotten when comparing their territories (Iraq
and Periadriatic area).
Pseudochubbina
bruni De Castro, 1990 (Figs.
2.10-11 ![]() and 4.5-7
and 4.5-7 ![]() ), comes from
the well-known Italian type locality of Cuvillierinella
salentina, of Campanian age (CsB6a zone). The A tests, spherical to ovoid, are smaller than P.
globularis (diameter from 1 to 2 mm up to 2.8 mm); like P.
globularis they are not typically planispiral (with sigmoid aspect, see in
particular De Castro, 1990,
Pl. 20.1) and the last coils increase suddenly in height in the largest tests.
The proloculus diameter ranges between 100 and 360 µm. The preseptal space is
relatively narrow, and contains pillars (Fig. 2.11
), comes from
the well-known Italian type locality of Cuvillierinella
salentina, of Campanian age (CsB6a zone). The A tests, spherical to ovoid, are smaller than P.
globularis (diameter from 1 to 2 mm up to 2.8 mm); like P.
globularis they are not typically planispiral (with sigmoid aspect, see in
particular De Castro, 1990,
Pl. 20.1) and the last coils increase suddenly in height in the largest tests.
The proloculus diameter ranges between 100 and 360 µm. The preseptal space is
relatively narrow, and contains pillars (Fig. 2.11 ![]() ). The central endoskeleton containing the secondary chamberlets is
frequently micritized in the last chambers. The primary chamberlets (35-65 µm)
are typically thinner than the scattered ones, but the two sets can be of
about equal diameter (Fig. 4.5-6
). The central endoskeleton containing the secondary chamberlets is
frequently micritized in the last chambers. The primary chamberlets (35-65 µm)
are typically thinner than the scattered ones, but the two sets can be of
about equal diameter (Fig. 4.5-6 ![]() ),
the basal ones being larger (40-80 µm). A unique section (Fig.
4.7
),
the basal ones being larger (40-80 µm). A unique section (Fig.
4.7 ![]() ) shows an uncommon variation of the chamberlets
orientation, recalling the confluent structure.
) shows an uncommon variation of the chamberlets
orientation, recalling the confluent structure.
Pseudochubbina
philippsoni (Fleury, 1977), Fig.
4.11-17 ![]() ,
previously ascribed with doubt to Chubbina,
comes from the Campanian (CsB6a zone) of Greece. It is moderately but
obviously streptospiral in the early coils, with frequent sigmoidal adult
tests (pseudoplanispiral coiling; see Fig. 4.15
,
previously ascribed with doubt to Chubbina,
comes from the Campanian (CsB6a zone) of Greece. It is moderately but
obviously streptospiral in the early coils, with frequent sigmoidal adult
tests (pseudoplanispiral coiling; see Fig. 4.15 ![]() ). The central coils consist of a biombilicate subspherical test, but
the adult is always nautiloid in form, and is more bilaterally flattened than
in other species. The adult test, without the uncoiled part, shows a final
coil that is much higher than the preceding ones; its diameter is about 2 mm,
twice the axial diameter. The final flange is about 2 to 4 mm in diameter, and
remains probably pseudoevolute. The proloculus is 230 ± 80 µm in diameter.
The preseptal space is narrow, with thick pillars. The chamberlets of the two
sets, measured on part of the type population, are about the same diameter (38
± 9 µm), but those of the BSC are sometimes quite discernable (Fig.
4.11 and 4.14
). The central coils consist of a biombilicate subspherical test, but
the adult is always nautiloid in form, and is more bilaterally flattened than
in other species. The adult test, without the uncoiled part, shows a final
coil that is much higher than the preceding ones; its diameter is about 2 mm,
twice the axial diameter. The final flange is about 2 to 4 mm in diameter, and
remains probably pseudoevolute. The proloculus is 230 ± 80 µm in diameter.
The preseptal space is narrow, with thick pillars. The chamberlets of the two
sets, measured on part of the type population, are about the same diameter (38
± 9 µm), but those of the BSC are sometimes quite discernable (Fig.
4.11 and 4.14 ![]() )
because of their slightly larger diameter and their closeness.
)
because of their slightly larger diameter and their closeness.
In conclusion, three of the species (P. kassabi being relatively apart) constitute a rather homogeneous set by their overall appearance and many important characters, such as the large diameter of the proloculus, the moderate but persisting pseudoplanispiral coiling and the tendency to final uncoiling. In addition to the small variations of these characters from one species to the others, the details of the endoskeleton organization (various diameter of chamberlets) provide another good criterion for distinguishing species. But the discernable BSC-SSC organization is not unique to this genus, as we will show below.
|
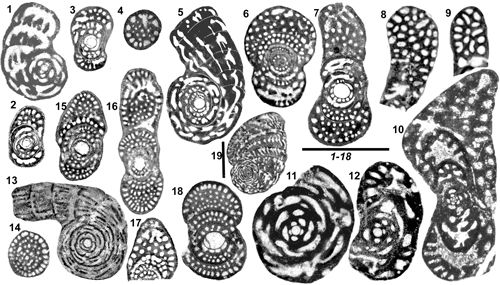
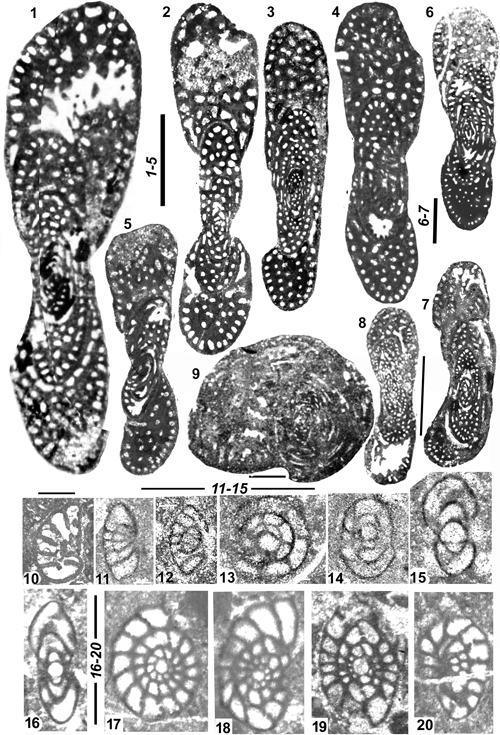
Figure 4: Genus Pseudochubbina.
1-4: P.
globularis. 1-3: Sub-axial, axial and equatorial sections of A tests;
preseptal spaces with preseptal pillars are surrounded in red; 4: Sub-equatorial
section of a supposed B test. 5-7: P. bruni, axial
centered sections of A tests and an off centered equatorial section recalling
the confluent structure. 8-10: P.
kassabi. 8: Oblique centered section of A test (holotype); 9: Off
centered section possibly from the same type of test (see Fig.
13.5 |
5.2 - Genus Cuvillierinella Papetti & Tedeschi, 1965
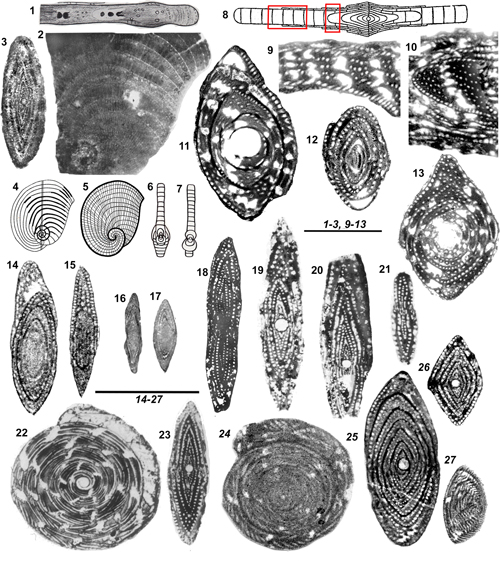
This genus, one of the oldest among the Rhapydioninidae of the Old World, was for a long time confused with Raadshoovenia, a previously described Cenozoic American genus of great resemblance. The main studies of C. salentina Papetti & Tedeschi, 1965, the type species of the genus from its Campanian type locality of southern Italy were presented by De Castro (1988, 1990) and Fleury (2016). This last author provided a historic overview of the genus and concluded that the two genera (Cuvillierinella and Raadshoovenia), with separate evolutionary developments in different provinces and different stratigraphic ranges must not be confused.
Cuvillierinella
salentina (Figs. 2.5 ![]() , 5.1-4
, 5.1-4 ![]() and 13.7-8
and 13.7-8 ![]() ). After Fleury
(2016) who studied and figured several new populations from Spain and Greece,
the species is characterized by relatively small tests (about 1 mm) and small
A proloculus (around 100 or 120 µm in diameter); the coiling is streptospiral
at first, then pseudoplanispiral to planispiral, with occasionally a terminal
cylindrical part in the A generation and cylindrical or flabelliform stages in
the B generation. The diameter of chamberlets, measured on three samples from
the type locality and three samples from Greece, is rather uncertain because
of its variability. The wide endoskeleton mesh obscures the observation of any
BSC-SSC structure in the type species, but the arrangement of the chamberlets
in the last chambers of Fig. 5.2-3
). After Fleury
(2016) who studied and figured several new populations from Spain and Greece,
the species is characterized by relatively small tests (about 1 mm) and small
A proloculus (around 100 or 120 µm in diameter); the coiling is streptospiral
at first, then pseudoplanispiral to planispiral, with occasionally a terminal
cylindrical part in the A generation and cylindrical or flabelliform stages in
the B generation. The diameter of chamberlets, measured on three samples from
the type locality and three samples from Greece, is rather uncertain because
of its variability. The wide endoskeleton mesh obscures the observation of any
BSC-SSC structure in the type species, but the arrangement of the chamberlets
in the last chambers of Fig. 5.2-3 ![]() and the secondary chamberlets of Fig. 5.4
and the secondary chamberlets of Fig. 5.4 ![]() are suggestive of this structure (see interpretation in Fig.
13.7-8
are suggestive of this structure (see interpretation in Fig.
13.7-8 ![]() ). The diameter of primary and basal chamberlets is about
60 µm and the scattered secondary chamberlets around 50 µm. A coarse and
exceptional confluent structure is observed in the last chambers of a B test (Fleury,
2016, text-fig. 6.11).
). The diameter of primary and basal chamberlets is about
60 µm and the scattered secondary chamberlets around 50 µm. A coarse and
exceptional confluent structure is observed in the last chambers of a B test (Fleury,
2016, text-fig. 6.11).
Cuvillierinella
perisalentina Fleury, 2016 (Figs.
2.6-7 ![]() ,
5.5-7
,
5.5-7 ![]() and 13.9-10
and 13.9-10 ![]() ), was found in the vicinity of the type locality of C.
salentina and Pseudochubbina bruni, and is presumably the same age (Campanian:
CsB6a). This species recalls features seen in Pseudochubbina, particularly the involute coiling with poorly
stabilized axis (pseudoplanispiral) and the central endoskeleton. But the
relatively small proloculus (about 150 µm), the overall smaller test, the
uniserial cylindrical final part of the A tests and the flabelliform final
flange of the B tests suggest probably no more than a kinship between this
species and Pseudochubbina. The
layer of primary chamberlets, both sets of secondary chamberlets, BSC and SSC
are clearly distinguished on Figs. 5.6-7
), was found in the vicinity of the type locality of C.
salentina and Pseudochubbina bruni, and is presumably the same age (Campanian:
CsB6a). This species recalls features seen in Pseudochubbina, particularly the involute coiling with poorly
stabilized axis (pseudoplanispiral) and the central endoskeleton. But the
relatively small proloculus (about 150 µm), the overall smaller test, the
uniserial cylindrical final part of the A tests and the flabelliform final
flange of the B tests suggest probably no more than a kinship between this
species and Pseudochubbina. The
layer of primary chamberlets, both sets of secondary chamberlets, BSC and SSC
are clearly distinguished on Figs. 5.6-7 ![]() and 13.9-10
and 13.9-10 ![]() , although the diameter
of chamberlets is almost the same (about 40-50 µm). The confluent structure
is not observed.
, although the diameter
of chamberlets is almost the same (about 40-50 µm). The confluent structure
is not observed.
5.3 - Genus Metacuvillierinella Fleury, 2016
The type
species, M. decastroi Fleury,
2016, is known by several populations of late Campanian (-early Maastrichtian)
age (CsB6b zone) from Greece and Italy. It is close to Cuvillierinella,
and also has a small proloculus in
the A tests, but is characterized by a weak dimorphism between generations and
by its pseudoplanispiral-advolute coiling without a final uncoiled stage. The
endoskeleton mesh is large, with primary chamberlets (about 70 µm in
diameter) slightly larger than the secondary ones (about 45 µm); the BSC is
observable in the last chambers of a few tests (Fig. 13.13 ![]() ). The confluent structure has never been observed.
). The confluent structure has never been observed.
5.4 - Genus Murciella Fourcade, 1966
This genus is not unlike Cuvillierinella, and was even considered as a junior synonym by De Castro (1988) and several followers. But Fleury (2016), studying very rich material kindly provided by P. De Castro from the type locality of C. salentina, showed that the populations contain some tests developing mixed characters described in both genera, in the absence of any individual resembling precisely the type of M. cuvillieri. Furthermore, Fleury (2018) observed that if the two species are occasionally associated in some locations, where they are nevertheless easily distinguishable, each of them is also known alone in various sites.
Murciella
cuvillieri Fourcade, 1966, the type species, was found in several
localities in Spain, Greece and the Dodecanese Astypalian Island (Fleury,
2018); it characterizes, together with C.
salentina, part of the Campanian stage (zone CsB6a). The overall aspects
of both generations of C. salentina
and M. cuvillieri are quite similar,
but the proloculus and test diameters of A tests of M. cuvillieri are a little larger (compare Fleury, 2016, text-fig. 12 and Fleury, 2018, text-fig. 15). The main difference is in the
coiling, quinqueloculine to streptospiral then pseudoplanispiral to
planispiral in Cuvillierinella,
wholly planispiral in M. cuvillieri (at
least in A tests). Another characteristic of this species is the thinner mesh
of the endoskeleton, generalized, but not immediately obvious in some cases.
An extreme example is given by a particular population from Greece: This
population (XGP110 in Fleury,
2018, text-fig. 6.1-14) is represented here by Figs. 5.14, 5.16 ![]() and
13.12
and
13.12 ![]() ; the sections display the ordinary serried primary
chamberlets, with relatively large diameters (about 40 µm) and the smaller
scattered secondary ones (about 25-30 µm), with the well distinguished BSC;
even the first stages are discernible in the central part of Fig.
5.15
; the sections display the ordinary serried primary
chamberlets, with relatively large diameters (about 40 µm) and the smaller
scattered secondary ones (about 25-30 µm), with the well distinguished BSC;
even the first stages are discernible in the central part of Fig.
5.15 ![]() from another population of the same type. These obvious
observations lead to a new understanding of some previously known sections
from the type population of the species; a section (Figs.
5.17
from another population of the same type. These obvious
observations lead to a new understanding of some previously known sections
from the type population of the species; a section (Figs.
5.17 ![]() and 13.11
and 13.11 ![]() ) is given as an example of what can be distinguished on many
figures from other populations showing the same arrangement (see Fleury,
2018, text-fig. 7.1, 7.3, 7.16 and 7.32).
) is given as an example of what can be distinguished on many
figures from other populations showing the same arrangement (see Fleury,
2018, text-fig. 7.1, 7.3, 7.16 and 7.32).
Thus, for the first time a typical BSC-SSC structure is observed in a species in which it was not previously noticed. For the first time too, this structure is observed together with the confluent structure known in several sections of the species, and particularly in the holotype of the type species (a definitive B test, according to Fleury, 2018, text-fig. 5.17), where it is conspicuous. We will have to watch for similar associations in the following examinations.
|
Figure 5: Genera Cuvillierinella,
Metacuvillierinella and Murciella.
1-4: Cuvillierinella salentina (A
tests). 1: Centered equatorial section; 2-3: Axial sections; 4: Transverse
section of the cylindrical terminal part (UUT). 5-7: Cuvillierinella perisalentina
(A tests). 5: Centered equatorial section; 6-7: Axial sections. 8-12:
Metacuvillierinella decastroi. 8-9:
Sections of last chambers of probable B tests with "fishnet" endoskeletal
organization; 10: Centered axial
section of a B test; 11-12: Centered equatorial and oblique sections (A tests).
13-19: Murciella cuvillieri.13:
Centered equatorial section (A test); 14: Transverse section of the cylindrical
UUT (A test); 15-18: Centered axial sections of A tests; 19: Centered equatorial
section of a B test with confluent structure, much resembling the holotype of
the species. Scale bars: 1mm. |
5.5 - Genus Sigalveolina Fleury, 2018
This genus was created in order to distinguish several species from the genus Murciella in which they were previously included. The type species, S. renzi (Fleury, 1979) and the 3 others attributed here to the genus are from Greece, southern Italy, Slovenia and possibly Turkey (Solak et al., 2019, text-fig. 10.J-K), and are probably of late Campanian - early Maastrichtian age (CsB6b zone). Their main common characters are their rather strict planispiral coiling, their usual lack of a final uncoiled stage in both generations, and the narrow diameter of the chamberlets.
Sections
of various species are very revealing (Figs. 6.1, 6.3-5, 6.7-8,
6.11 ![]() and 13.20-22
and 13.20-22 ![]() ):
Every axial one shows, at least in the last coils, the triad constituted by
the regularly disposed beads of the outer primary chamberlets, the median
disordered ones (ordinary a little smaller than the primary) and the BSC stuck
on the wall of the previous coil. But it must be underlined that, at least in
the type species (S. renzi), this
structure is not ordinary obvious because of the rather homogenous diameter of
all chamberlets and their closeness, which is an exception to the ordinary
aspect of this structure. The confluent structure
is known in the four species (Fig. 6.3, 6.6,
6.9 and 6.12
):
Every axial one shows, at least in the last coils, the triad constituted by
the regularly disposed beads of the outer primary chamberlets, the median
disordered ones (ordinary a little smaller than the primary) and the BSC stuck
on the wall of the previous coil. But it must be underlined that, at least in
the type species (S. renzi), this
structure is not ordinary obvious because of the rather homogenous diameter of
all chamberlets and their closeness, which is an exception to the ordinary
aspect of this structure. The confluent structure
is known in the four species (Fig. 6.3, 6.6,
6.9 and 6.12 ![]() ).
).
The conjunction of the BSC-SSC structure and the confluent structure is obvious in this genus.
|
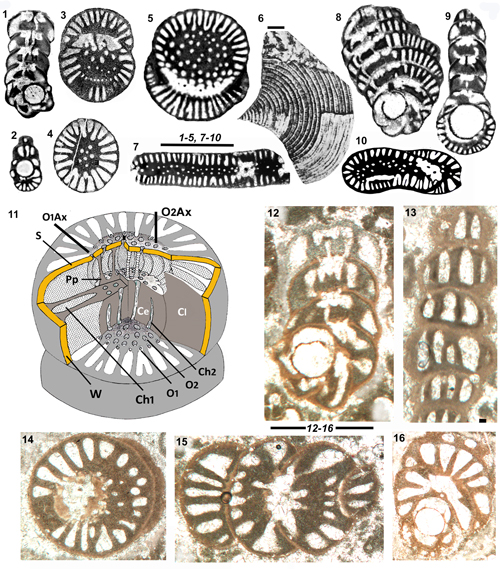
Figure 6: Genus
Sigalveolina. 1-3:
S. renzi and S. aff. renzi
(A tests). 1 and 3: Axial sections showing the regularly disposed BSC in last
chambers; 2: Equatorial section with obvious confluent structure (S. aff. renzi). 4-6: S.
ovoidea. 4-5: Axial sections displaying the various stages of the SSC;
6: Tangential section with confluent structure. 7-9:
S. methonensis. 7-8: Axial sections A and B tests, with BSC
clearly distinguished in chambers of the last coils; 9: Equatorial section
showing the confluent structure. 10-12: S.
reicheli (A tests). 10: Transverse section of a cylindrical UUT; 11:
Axial section with BSC in the chambers of the last coil; 12: Equatorial section
of a large test displaying the confluent structure. Scale bars: 1mm. |
5.6 - Genus Cyclopseudedomia Fleury, 1974
This genus, almost fully planispiral, is one of the ultimate endings of the arborescent genealogy of Rhapydioninidae, the paragon of uncoiling. In the type species, C. smouti Fleury, 1974, the A generation comprises a very large proloculus (up to 360 µm), enveloped by a single involute coil, and a number of pseudoevolute, then quickly evolute semicircular flat chambers; the B generation is mainly made up of annular chambers forming a flat discoid test, up to about 10 mm in diameter. Three other species, occurring in the Campanian-Maastrichtian of the periadriatic region, are similar, hardly less uncoiled, with a flattened or cylindrical final stage in A tests, but always large and flattened (but not discoidal) in B tests (see Fleury, 2018, text-fig. 16). Some resemblance with Pseudochubbina kassabi and Twaraina, both incompletely known, are to be noted.
Although
the flatness of the tests leaves little place for them, the scattered
chamberlets are visible at least in the last chambers of the four species,
especially in C. smouti (Fig. 7.3-4 and 7.6 ![]() ). But the very short coiled stages of A tests do not show the
BSC; a doubtful exception is perhaps represented on Fig. 13.19
). But the very short coiled stages of A tests do not show the
BSC; a doubtful exception is perhaps represented on Fig. 13.19 ![]() . The confluent structure is known in three species:
It is
well developed in C. smouti and klokovaensis,
which have thinner chamberlets, coarse in C.
mavrikasi, and never observed in C.
hellenica, which displays the largest chamberlets (Fig. 12
. The confluent structure is known in three species:
It is
well developed in C. smouti and klokovaensis,
which have thinner chamberlets, coarse in C.
mavrikasi, and never observed in C.
hellenica, which displays the largest chamberlets (Fig. 12 ![]() ).
).
The association of BSC-SSC and confluent structure cannot be confirmed in this genus. The apparent absence of BSC could be due to the difficulty of observations in the extremely short juvenile stage.
|
Figure 7: Genus
Cyclopseudedomia. 1-7:
C. smouti. 1:
Centered equatorial section of a small A test; 2: Axial section of an A test,
with a possible BSC in contact of the proloculus (see Fig.
13.19 |
5.7 - Genus Pseudedomia Henson, 1948
Forewords: About the generic concept of Pseudedomia Henson, 1948
The type
of the genus, P. multistriata Henson,
1948 (a possible B test, although the central part is not seen) is described
after 2 sections of a unique test (Fig. 8.1-2 ![]() ), the holotype. It is known in the Maastrichtian limestones with Lepidorbitoides minor (Schlumb.)
from "a deep borehole at Jebel Dukhan, Qatar Peninsula", according to Smout
(1963, p. 256). One of the sections, after Smout
(op. cit., Pl. I.6, reproduced here Fig. 8.1
), the holotype. It is known in the Maastrichtian limestones with Lepidorbitoides minor (Schlumb.)
from "a deep borehole at Jebel Dukhan, Qatar Peninsula", according to Smout
(1963, p. 256). One of the sections, after Smout
(op. cit., Pl. I.6, reproduced here Fig. 8.1 ![]() ), is "very obscure" and
"little reliance can be placed on this drawing", dixit
Smout himself. The other one (op. cit., Pl. I.1, here Fig.
8.2
), is "very obscure" and
"little reliance can be placed on this drawing", dixit
Smout himself. The other one (op. cit., Pl. I.1, here Fig.
8.2 ![]() ) comprises a small regularly
planispiral part of about 3 coils, followed by about 15 chambers forming a
large flange, according to the original photography and the drawing of Smout
(op. cit., text-fig. 9a, reproduced
here Fig. 8.4
) comprises a small regularly
planispiral part of about 3 coils, followed by about 15 chambers forming a
large flange, according to the original photography and the drawing of Smout
(op. cit., text-fig. 9a, reproduced
here Fig. 8.4 ![]() ). It was interpreted
by Smout (op. cit., text-fig. 9b; here Fig.
8.5
). It was interpreted
by Smout (op. cit., text-fig. 9b; here Fig.
8.5 ![]() )
as belonging to a "hyperinvolute" test of the "P."
complanata type (see below). In fact, no proof is given that
the original description of Henson
(test "evolute, later chambers uniserial") was wrong and two types of
axial sections (Fig. 8.6-7
)
as belonging to a "hyperinvolute" test of the "P."
complanata type (see below). In fact, no proof is given that
the original description of Henson
(test "evolute, later chambers uniserial") was wrong and two types of
axial sections (Fig. 8.6-7 ![]() ) are
still equally credible. Its endoskeleton mesh is very thin. The preseptal
space is narrow, and equipped with numerous pillars. Even the existence of two
sets of chamberlets is questionable, and there is no apparent obliquity in
their arrangement.
) are
still equally credible. Its endoskeleton mesh is very thin. The preseptal
space is narrow, and equipped with numerous pillars. Even the existence of two
sets of chamberlets is questionable, and there is no apparent obliquity in
their arrangement.
The
assumed A generation was described by Smout
(op. cit.). The material consists
partly of specimens labelled as "topotypes" in the legend of his Pl. I (2
and 4), a free test and a partial non-centered section; the figures are quite
precise and show endoskeleton affinities with the holotype by their thin
chamberlets. Nevertheless, the best specimen is a centered axial section (Smout,
1963,
Pl. I.3, reproduced here Fig. 8.3 ![]() ),
which is not a topotype as it comes from southern Iraq; it is totally involute,
with a rather acute periphery, lacks an uncoiled stage, and is almost as large
as the uncoiled supposed corresponding B test (see Fig.
8.2-3
),
which is not a topotype as it comes from southern Iraq; it is totally involute,
with a rather acute periphery, lacks an uncoiled stage, and is almost as large
as the uncoiled supposed corresponding B test (see Fig.
8.2-3 ![]() ). This section shows a very thin illegible central endoskeleton, not
interpretable in terms of BSC-SSC or confluent structure; the diameter of the
proloculus is 150 µm and the diameter of the "primary chamberlets" is 7
µm.
). This section shows a very thin illegible central endoskeleton, not
interpretable in terms of BSC-SSC or confluent structure; the diameter of the
proloculus is 150 µm and the diameter of the "primary chamberlets" is 7
µm.
Summarizing, the interpretation of the type section of the type species of Pseudedomia (B test) is open to discussion, and the assumed corresponding A test does not give rise to full confidence. These considerations are not decisive and it is still possible that the generic concept of Pseudedomia, according to Smout (op. cit.), may be right. It was generally accepted by authors who encountered small tests more or less similar to the "A test" section of Smout, but as we will see, none of them ever described a B test of Henson's original discovery type. Thus, as long as the uncertainty about the classical material is not cleared up, the greatest care is to be observed about the concept of Pseudedomia. That is why all taxa attributed later to the genus Pseudedomia, with the exception of the type specimen, are presented here with speech marks.
About some supposed "Pseudedomia" species
A short review of some populations attributed to Pseudedomia will be informative of the way the authors understood and interpreted the complex situation previously described.
"Pseudedomia"
complanata Eames & Smout,
1955 (Fig.
8.8-13 ![]() ),
from Kuwait, was found in a chalky limestone with occasional gastropods,
lamellibranchia, Archaecyclus
midorientalis Eames
& Smout, 1955,
globotruncanids and various smaller foraminifera. Possessing "many
characters in common with Praealveolina
cretacea" (Eames
& Smout, op. cit.), "P."
complanata is the eldest true member of the Rhapydioninidae. For the first
time, a typical alveolinid endoskeleton was described in a test with soritid
coiling. The new taxon is clearly illustrated and described, with relatively
small A tests (about 2 mm in diameter), "stoutly lenticular" and very
large B tests (up to 35 mm in diameter) "discoidal, complanate, involute,
only part of the last whorl visible externally". Tests of both generations
have very large preseptal spaces (about half of the distance between two
successive septa in B tests) equipped with numerous thin preseptal pillars of
about the same diameter as the chamberlets. The two sets of chamberlets are
quite distinct, although they have almost the same diameter; the primary
chamberlets, separated by thin cloisonnettes, form an obvious continuous row
in section; the secondary ones are scattered in the central endoskeleton and a
BSC can be clearly distinguished in the A tests (Figs. 8.13
),
from Kuwait, was found in a chalky limestone with occasional gastropods,
lamellibranchia, Archaecyclus
midorientalis Eames
& Smout, 1955,
globotruncanids and various smaller foraminifera. Possessing "many
characters in common with Praealveolina
cretacea" (Eames
& Smout, op. cit.), "P."
complanata is the eldest true member of the Rhapydioninidae. For the first
time, a typical alveolinid endoskeleton was described in a test with soritid
coiling. The new taxon is clearly illustrated and described, with relatively
small A tests (about 2 mm in diameter), "stoutly lenticular" and very
large B tests (up to 35 mm in diameter) "discoidal, complanate, involute,
only part of the last whorl visible externally". Tests of both generations
have very large preseptal spaces (about half of the distance between two
successive septa in B tests) equipped with numerous thin preseptal pillars of
about the same diameter as the chamberlets. The two sets of chamberlets are
quite distinct, although they have almost the same diameter; the primary
chamberlets, separated by thin cloisonnettes, form an obvious continuous row
in section; the secondary ones are scattered in the central endoskeleton and a
BSC can be clearly distinguished in the A tests (Figs. 8.13 ![]() and 13.24
and 13.24 ![]() ).
).
Unfortunately and probably because of the lack of an available context, the generic name was chosen by the authors in reference to a previously described taxon from the same area: P. multistriata Henson. A comparison of the endoskeleton of the two taxa "leaves no doubt that there is identity of structure, the difference being only in size and proportions", after Smout (1963, p. 254). Nevertheless, the choice of P. multistriata Henson as a unique reference appears debatable, partly because of the uncertainties about the type, and partly because of the dubious identification of the A generation by Smout.
"Pseudedomia"
was well illustrated with two centered sections by Wagner,
1964 (Fig. 8.22-23 ![]() ), from an unknown
locality. The equatorial section shows a proloculus of comparatively large
diameter (about 200 µm) and a large preseptal space; the axial one shows the
serried primary chamberlets and some scattered secondary
chamberlets, but the
BSC does not appear. Some obliquity of the chamberlets is visible on the
bottom part of the equatorial section.
), from an unknown
locality. The equatorial section shows a proloculus of comparatively large
diameter (about 200 µm) and a large preseptal space; the axial one shows the
serried primary chamberlets and some scattered secondary
chamberlets, but the
BSC does not appear. Some obliquity of the chamberlets is visible on the
bottom part of the equatorial section.
"Pseudoedomia"
is figured by Perconig
& Martin, 1971 (Fig. 8.14-15 ![]() ), from the Oriental Prebetic domain (Spain) in association with
abundant mollusc shells, Orbitoides
media Archiac and other larger foraminifera, in limestones of
Campanian-Maastrichtian age. Two non-centered sections show the usual layer of
serried primary chamberlets and well distinguished scattered secondary ones,
but the BSC is not visible; the preseptal space is apparent.
), from the Oriental Prebetic domain (Spain) in association with
abundant mollusc shells, Orbitoides
media Archiac and other larger foraminifera, in limestones of
Campanian-Maastrichtian age. Two non-centered sections show the usual layer of
serried primary chamberlets and well distinguished scattered secondary ones,
but the BSC is not visible; the preseptal space is apparent.
"Pseudedomia"
hamaouii Rahaghi,
1976 (Fig. 8.18 ![]() ), is associated with globotruncanids, Orbitoides and an
unidentified species of Goupillaudina
Marie from the Campanian of
SW Iran; it is mainly known by free tests, axially compressed (diameter up to
2.7 mm, axial dimension up to 0.5 mm). The diameter of the proloculus in A
tests is less than 75 µm. Equatorial sections show thin chamberlets and a
large preseptal space; the axial one (Fig. 8.18
), is associated with globotruncanids, Orbitoides and an
unidentified species of Goupillaudina
Marie from the Campanian of
SW Iran; it is mainly known by free tests, axially compressed (diameter up to
2.7 mm, axial dimension up to 0.5 mm). The diameter of the proloculus in A
tests is less than 75 µm. Equatorial sections show thin chamberlets and a
large preseptal space; the axial one (Fig. 8.18 ![]() ) displays the serried layer of primary chamberlets and a few
secondary chamberlets isolated in the central endoskeleton, but the BSC is not
visible. Some B tests are figured, and only differ from A tests by their small
proloculus.
) displays the serried layer of primary chamberlets and a few
secondary chamberlets isolated in the central endoskeleton, but the BSC is not
visible. Some B tests are figured, and only differ from A tests by their small
proloculus.
"Pseudedomia"
cf. hamaouii is well illustrated by De Castro
(1988) from the Campanian or Maastrichtian of Abu
Dhabi (Fig. 8.19-21 ![]() ). The A tests,
with a proloculus larger than 100 µm and a relatively loose structure in the
equatorial region, show the usual serried primary chamberlets and a few
scattered secondary ones; a differentiated BSC seems probable in the upper
part of section 20. The assumed B test (Fig. 8.21
). The A tests,
with a proloculus larger than 100 µm and a relatively loose structure in the
equatorial region, show the usual serried primary chamberlets and a few
scattered secondary ones; a differentiated BSC seems probable in the upper
part of section 20. The assumed B test (Fig. 8.21 ![]() ) is very small.
) is very small.
"Pseudedomia" aff. multistriata was discovered by Mavrikas
et al. (1994) in a small
Maastrichtian outcrop of the Gavrovo massif in Greece (Fig.
8.24-27 ![]() ). It is found in poorly stratified limestones with
large rudist shells and rich associations of Maastrichtian foraminifera: Loftusia
sp., Orbitoides apiculata Schlumb.,
O. gensacicus (Leym.), Clypeorbina mamillata Schlumb., Hellenocyclina
beotica Reichel,
Smoutina cruysi Drooger, Siderolites
calcitrapoides Lmk., Nummofallotia
cretacea (Schlumb.) and Sivasella
monolateralis Sirel
& Gündüz. Only A tests were observed, in spite of active
field searches for B tests. The tests are lenticular with an acute periphery;
their proloculus is smaller than 100 µm, equatorial diameters are up to 1.70
mm, and axial diameters up to 0.80 mm. The serried row of primary chamberlets
is clearly distinct; the secondary chamberlets are small in number and
scattered through the poorly developed central endoskeleton; the BSC is
represented by short aligned small chamberlets visible in the nearness of
previous coils (Figs. 8.25
). It is found in poorly stratified limestones with
large rudist shells and rich associations of Maastrichtian foraminifera: Loftusia
sp., Orbitoides apiculata Schlumb.,
O. gensacicus (Leym.), Clypeorbina mamillata Schlumb., Hellenocyclina
beotica Reichel,
Smoutina cruysi Drooger, Siderolites
calcitrapoides Lmk., Nummofallotia
cretacea (Schlumb.) and Sivasella
monolateralis Sirel
& Gündüz. Only A tests were observed, in spite of active
field searches for B tests. The tests are lenticular with an acute periphery;
their proloculus is smaller than 100 µm, equatorial diameters are up to 1.70
mm, and axial diameters up to 0.80 mm. The serried row of primary chamberlets
is clearly distinct; the secondary chamberlets are small in number and
scattered through the poorly developed central endoskeleton; the BSC is
represented by short aligned small chamberlets visible in the nearness of
previous coils (Figs. 8.25 ![]() and 13.23
and 13.23 ![]() ). Tangential sections (Figs.
8.27
). Tangential sections (Figs.
8.27 ![]() ) reveal the presence of the confluent structure.
) reveal the presence of the confluent structure.
"Pseudedomia" sp. and "P." cf. hamaouii were observed by Chiocchini et al. (1994, Pl. 25.10-12) in association with Orbitoides media and Murciella spp. in probable Campanian levels younger than "Raadshoovenia" salentina and older than the Maastrichtian association of Orbitoides apiculata, O. macroporus, S. calcitrapoides and Loftusia sp. The figured sections share the ordinary biconvex profile with sharp periphery and tight coiling of the species classically referred to the A test of "P." multistriata after Smout.
"Pseudedomia"
hekimhanensis Görmüs, 1999, was established from the upper Campanian (?)
to Maastrichtian of central Turkey (Malatya region). The original material
comes from mudstones with gastropods and bivalves, with Goupillaudina daguini Marie,
1958, and various smaller foraminifers. It is considered as upper Maastrichtian
by Solak et al. (2017). It is a small species:
The biconvex A tests are less
than 0.81 mm in equatorial diameter, with a tiny proloculus (less than 50 µm
in diameter). According to Görmüs,
the B generation would be only differentiated by its nepionic stage. The
primary chamberlets have a large diameter (almost half the diameter of the
proloculus in A tests) and the secondary chamberlets seem absent. The
preseptal space is not apparent in the original figures, but the sections
given by Solak et al. (Fig.
8.16-17 ![]() )
show very thin chamberlets and a probable preseptal space.
)
show very thin chamberlets and a probable preseptal space.
"Pseudedomia" sp. aff. complanata, as shown by Luger (2018, Pl. 16.6-9) from Yemen closely resembles the A generation (after Smout) of "P." multistriata, with tight coiling, a sharp periphery without a final uncoiled stage, but with a probably large proloculus.
Conclusion: An apparent fragile taxon
This
review shows that, for all authors, the reference to "Pseudedomia" only refers to the A test section provided by Smout,
1955 (Fig. 8.3 ![]() ), and never to Henson's
(1948) type of the genus; the only B tests observed or interpreted (Rahaghi, De Castro,
Görmüs) do not show a
final pseudoevolute or evolute flange. The ambiguity that was previously
underlined is thus strengthened and the use of brackets seems to be justified
as a safety measure to prevent further taxonomic problems. Nothing significant
can be done until new material from the type region is available.
), and never to Henson's
(1948) type of the genus; the only B tests observed or interpreted (Rahaghi, De Castro,
Görmüs) do not show a
final pseudoevolute or evolute flange. The ambiguity that was previously
underlined is thus strengthened and the use of brackets seems to be justified
as a safety measure to prevent further taxonomic problems. Nothing significant
can be done until new material from the type region is available.
In spite of the taxonomic uncertainties and the difficulty of observations in very compressed tests, the preceding review is not unfavorable to our search. All the so called compressed "Pseudedomia" are provided with scattered chamberlets but the BSC is not proven in every case. Nevertheless, two examples ("P." complanata and "P." aff. multistriata) show a true BSC-SSC organization, even associated with the confluent structure in the second.
|
Figure 8: Pseudedomia multistriata Henson
and other species attributed to "Pseudedomia".
1-7: Pseudedomia multistriata.
1-2: Two sections of the holotype, after Smout
(1963, Pl. I.1 and 1.6); 3: A test, in the sense of Smout
(op. cit., Pl. 1.3); 4: "Reconstruction of the equatorial section of
the holotype, lines actually seen bold, endoskeleton omitted" by Smout,
op. cit., text-fig. 9a, p. 256, with situation of axial sections 6-7; 5:
Interpretation of section 4 by Smout
(text-fig. 9b), mainly inspired by the "hyperinvolute" coiling of "Pseudedomia"
complanata; 6-7: Alternative theoretical axial sections in case of
"hyperinvolute" (6) or involute-pseudoevolute (7) tests, both compatible
with section 4. 8-13:
"Pseudedomia"
complanata Eames & Smout.
8: Idealized axial section of a B test ("hyperinvolute" coiling), after Eames
& Smout (1955, text-fig. 2.B), with situation of Fig.
8.9-10 |
5.8 - Genus Sellialveolina Colalongo, 1963
Sellialveolina was long confused with Pseudedomia, although Fleury & Fourcade (1990) showed that they belong to different subfamilies. The two genera have no more in common than the type of endoskeleton and the time of their descriptions, when the context of the family was still ignored. We will delve into this point after a short description of figured specimens.
A review of the various morphotypes. As almost every population seems to be unique, the distinction of species appears particularly difficult in this widely distributed group; for the present time, waiting for a precise study, the following morphotypes describe the various stages of an apparent complex evolution.
A small
species (Fig. 9.1-5 ![]() ), still
undescribed (but figured in Fleury
& Fourcade, 1990,
text-fig. 5.AB1), is probably not far from the origin of the group. It comes
from a unique but very rich sample from the Gavrovo massif in Greece (around
spot "D", text-fig. 14, in Fleury, 1980). The A tests (less than 0.40 mm in diameter)
are at first either streptospiral or planispiral around a proloculus of about
100 µm or less; the equatorial diameter of the adult is larger than the axial
one; the B test, almost spherical with a small central glomerulus, is up to
0.50 mm in diameter. The wall is thin, the endoskeleton is simple, and there
are no secondary chamberlets. The preseptal space is very large (about one
half of the distance between septa), without visible preseptal pillars.
), still
undescribed (but figured in Fleury
& Fourcade, 1990,
text-fig. 5.AB1), is probably not far from the origin of the group. It comes
from a unique but very rich sample from the Gavrovo massif in Greece (around
spot "D", text-fig. 14, in Fleury, 1980). The A tests (less than 0.40 mm in diameter)
are at first either streptospiral or planispiral around a proloculus of about
100 µm or less; the equatorial diameter of the adult is larger than the axial
one; the B test, almost spherical with a small central glomerulus, is up to
0.50 mm in diameter. The wall is thin, the endoskeleton is simple, and there
are no secondary chamberlets. The preseptal space is very large (about one
half of the distance between septa), without visible preseptal pillars.
"Ovalveolina"
maccagnoae De Castro, 1966 (Fig. 9.6-7 ![]() ), is mainly
known in the Vraconian to middle Cenomanian of Italy (De
Castro, 1988). It is here considered as a Sellialveolina species, because of its nautiloid morphology (with an
equatorial diameter up to 1 mm) and the presence of occasional supplementary
chamberlets in the equatorial plane (Fig. 9.6
), is mainly
known in the Vraconian to middle Cenomanian of Italy (De
Castro, 1988). It is here considered as a Sellialveolina species, because of its nautiloid morphology (with an
equatorial diameter up to 1 mm) and the presence of occasional supplementary
chamberlets in the equatorial plane (Fig. 9.6 ![]() ); the proloculus of the A test is about 100 ± 20 µm. The preseptal
space is large (Fig. 9.7
); the proloculus of the A test is about 100 ± 20 µm. The preseptal
space is large (Fig. 9.7 ![]() ,
last chamber).
,
last chamber).
Forms
resembling both O. maccagnoae and
Sellialveolina viallii (Fig. 9.8-9 ![]() )
do exist in the Albian or Cenomanian of Greece (Varassova massif, in
Fleury, 1971, 1980). They
consist of nautiloid tests, usually smaller than maccagnoae, but with occasional clearly distinct secondary
chamberlets.
)
do exist in the Albian or Cenomanian of Greece (Varassova massif, in
Fleury, 1971, 1980). They
consist of nautiloid tests, usually smaller than maccagnoae, but with occasional clearly distinct secondary
chamberlets.
Sellialveolina
viallii Colalongo, 1963
(Fig. 9.10-17 ![]() ), is considered as of
Vraconian to middle Cenomanian in age (De
Castro, 1980). It appears as a versatile species, with tests either
smaller or larger than maccagnoae in
diameter, always nautiloid with acute or rounded periphery. The proloculus in
A tests is rather small, rarely up to 100 µm. The B tests are rare, and only
distinguished by their nepionic stage. The chamberlets are often numerous,
with the fishnet arrangement, or eventual BSC-SSC structure (Fig.
9.15
), is considered as of
Vraconian to middle Cenomanian in age (De
Castro, 1980). It appears as a versatile species, with tests either
smaller or larger than maccagnoae in
diameter, always nautiloid with acute or rounded periphery. The proloculus in
A tests is rather small, rarely up to 100 µm. The B tests are rare, and only
distinguished by their nepionic stage. The chamberlets are often numerous,
with the fishnet arrangement, or eventual BSC-SSC structure (Fig.
9.15 ![]() and 13.25
and 13.25 ![]() ). The
preseptal space is relatively large (1/3 of the distance between septa, or
more) and the preseptal pillars are frequently present. The type population (Fig.
9.10-11
). The
preseptal space is relatively large (1/3 of the distance between septa, or
more) and the preseptal pillars are frequently present. The type population (Fig.
9.10-11 ![]() )
appears in a median position among the various examples from
Greece and Italy of Fig. 9
)
appears in a median position among the various examples from
Greece and Italy of Fig. 9 ![]() .
.
Transitional
forms between S. viallii to S. drorimensis
(Fig. 9.18-19 ![]() ) are known in likely middle Cenomanian deposits from
Turkey (Bignot & Poisson,
1974) and Spain (S. gutzwilleri
Vicedo et al., 2011). The main character of these morphotypes is their
uncoiled (pseudoevolute) terminal stage and, for the Turkish ones, a very
large preseptal space (half the length of the chambers) equipped with numerous
preseptal pillars, recalling rather precisely S. drorimensis. The SSC-BSC structure is present (Figs.
9.18-19
) are known in likely middle Cenomanian deposits from
Turkey (Bignot & Poisson,
1974) and Spain (S. gutzwilleri
Vicedo et al., 2011). The main character of these morphotypes is their
uncoiled (pseudoevolute) terminal stage and, for the Turkish ones, a very
large preseptal space (half the length of the chambers) equipped with numerous
preseptal pillars, recalling rather precisely S. drorimensis. The SSC-BSC structure is present (Figs.
9.18-19 ![]() and 13.26
and 13.26 ![]() ).
).
Sellialveolina
drorimensis (Reiss et al., 1964), from the upper Cenomanian around the Mediterranean
Sea, was described as a species of
Pseudedomia. We will reject again this generic attribution in the
following paragraph, but it is effectively a very different species from S.
viallii. Fig. 9.21-25 ![]() shows the large tests (larger than 1, up to 2 mm in
equatorial diameter), lenticular, with acute periphery and, above all, the
development of a pseudoevolute to evolute final flange. The proloculus is
about 150 ± 25 µm in diameter according to various authors cited in the
legend to Fig. 9
shows the large tests (larger than 1, up to 2 mm in
equatorial diameter), lenticular, with acute periphery and, above all, the
development of a pseudoevolute to evolute final flange. The proloculus is
about 150 ± 25 µm in diameter according to various authors cited in the
legend to Fig. 9 ![]() ; the endoskeleton
mesh is relatively small and regular; the BSC-SSC structure is clearly
observable in Figs. 9.21
; the endoskeleton
mesh is relatively small and regular; the BSC-SSC structure is clearly
observable in Figs. 9.21 ![]() and 13.27
and 13.27 ![]() ; the preseptal space is large, with numerous preseptal
pillars. A population coming from the type region was extensively figured by De
Castro (1988, Pls. II-III); it fits the main features of the species,
but shows a well-developed confluent structure (Fig. 9.25
; the preseptal space is large, with numerous preseptal
pillars. A population coming from the type region was extensively figured by De
Castro (1988, Pls. II-III); it fits the main features of the species,
but shows a well-developed confluent structure (Fig. 9.25 ![]() ) apparently absent from the type population; it is therefore probably
non-conspecific with the types. Nevertheless, in the lack of a comparative
study, it is not treated separately in Fig. 12
) apparently absent from the type population; it is therefore probably
non-conspecific with the types. Nevertheless, in the lack of a comparative
study, it is not treated separately in Fig. 12 ![]() .
.
The various morphotypes seem to follow a simple trend, possibly a simple and unique track of filiation. This is probably misleading, as Fleury & Fourcade (1990, text-fig. 5.B), tried to show. These authors distinguished two lines of evolution, in order to explain the absence of larger morphotypes in the Greek Gavrovo-Tripolitza carbonates of the restricted internal platform. But it is certainly too simple and an arborescent schema would be probably the only one to give an account of the group complexity, which seems to be poorly understood at present. A last remark can be established about ecology: If S. gr. viallii seems to be confined to internal platform environments, S. drorimensis is probably inhabiting more open marine ones.
About the endoskeleton structure. A progressive change can be observed between the simplest (and oldest) specimens and the more complex (and youngest) specimens considered above.
The
simplest structure is just an alveolinid one (Fig. 9.1-9 ![]() ), with a single layer of (primary) chamberlets and
eventual solitary additional ones in the equatorial plane; these appear either
near the wall, with Y-shaped cloisonnettes (Fig. 9.6
), with a single layer of (primary) chamberlets and
eventual solitary additional ones in the equatorial plane; these appear either
near the wall, with Y-shaped cloisonnettes (Fig. 9.6 ![]() ), or near the preceding coil (Fig.
9.9
), or near the preceding coil (Fig.
9.9 ![]() ), or by division of the former chamberlets by some kind of
"floor" parallel to the wall (Fig. 9.8 and 9.13
), or by division of the former chamberlets by some kind of
"floor" parallel to the wall (Fig. 9.8 and 9.13 ![]() ).
).
The
widespread structure (Fig. 9.10-17 ![]() )
is a mix of the preceding types, giving rise to a fishnet pattern. It can be
noted that in some cases, a BSC can be distinguished (Figs.
9.15
)
is a mix of the preceding types, giving rise to a fishnet pattern. It can be
noted that in some cases, a BSC can be distinguished (Figs.
9.15 ![]() and 13.25
and 13.25 ![]() ,
last chamber).
,
last chamber).
The SSC
structure appears evidently in the largest tests and the differentiation of
the BSC is clear in many cases (Figs. 9.18-19,
9.21, 9.23 ![]() and 13.26-27
and 13.26-27 ![]() ). The
emergence of the confluent structure in a variety of S. drorimensis (Fig.
9.25
). The
emergence of the confluent structure in a variety of S. drorimensis (Fig.
9.25 ![]() )
gives evidence for close affinities with the previously studied younger groups
of the family.
)
gives evidence for close affinities with the previously studied younger groups
of the family.
In
short, if the
Sellialveolininae seems to stand apart from the other groups of the family (at
least by its age and, possibly its origin if the "small species" of Fig.
9.1-5 ![]() can be considered as such), the BSC and the confluent structure in
the most advanced specimens would seem to indicate that only the large
diameter of the chamberlets in primitive specimens hinders the general
identification of the SSC.
can be considered as such), the BSC and the confluent structure in
the most advanced specimens would seem to indicate that only the large
diameter of the chamberlets in primitive specimens hinders the general
identification of the SSC.
Comparison with Pseudedomia. The
simple confrontation of Figs. 8 ![]() and
9
and
9 ![]() is decisive: There is no striking resemblance between the two
genera. But the detailed comparison needs precaution, because of the
complexity of the state of knowledge of both genera. Considered as a unique
entity, despite their diversity, the various figured Sellialveolina
can be described as constituted by subglobular to nautiloid tests with rounded
periphery, the only lenticular ones being finally uncoiled. The coiling is
loose and the endoskeletal mesh is large. The endoskeleton is rather of the
fishnet type in the smaller tests and is reaching the BSC-SSC structure only
in the largest tests with narrower chamberlets. The preseptal space is large,
almost half the distance between 2 successive septa. They differ from:
is decisive: There is no striking resemblance between the two
genera. But the detailed comparison needs precaution, because of the
complexity of the state of knowledge of both genera. Considered as a unique
entity, despite their diversity, the various figured Sellialveolina
can be described as constituted by subglobular to nautiloid tests with rounded
periphery, the only lenticular ones being finally uncoiled. The coiling is
loose and the endoskeletal mesh is large. The endoskeleton is rather of the
fishnet type in the smaller tests and is reaching the BSC-SSC structure only
in the largest tests with narrower chamberlets. The preseptal space is large,
almost half the distance between 2 successive septa. They differ from:
Pseudedomia
multistriata
(considering the holotype, possible B generation, Fig. 8.2 ![]() , comprises mainly a large evolute, pseudoevolute or "hyperinvolute" flange (see Fig.
8.6-7
, comprises mainly a large evolute, pseudoevolute or "hyperinvolute" flange (see Fig.
8.6-7 ![]() )
following a short coiled stage (involute, evolute?); the endoskeleton mesh is
very fine and the preseptal space very narrow.
)
following a short coiled stage (involute, evolute?); the endoskeleton mesh is
very fine and the preseptal space very narrow.
"Pseudedomia"
with various species attributions (more or less resembling the A test
discovered by Smout, 1963, Fig. 8.3 ![]() ), not certainly corresponding to the holotype but accepted as such by
many authors (Fig. 8.14-27
), not certainly corresponding to the holotype but accepted as such by
many authors (Fig. 8.14-27 ![]() ), has a lenticular test with sharp periphery, very
tight coiling and no final uncoiled stage; the endoskeleton comprises a few
chamberlets with small diameters and a narrow preseptal spaces.
), has a lenticular test with sharp periphery, very
tight coiling and no final uncoiled stage; the endoskeleton comprises a few
chamberlets with small diameters and a narrow preseptal spaces.
"Pseudedomia"
complanata (Fig. 8.8-13 ![]() ), with
its larger tests of both generations, the very large proloculus in the A
tests, the special "hyperinvolute" coiling of the B test; the SSC
endoskeleton with very fine BSC chamberlets has no equivalent in Sellialveolina.
), with
its larger tests of both generations, the very large proloculus in the A
tests, the special "hyperinvolute" coiling of the B test; the SSC
endoskeleton with very fine BSC chamberlets has no equivalent in Sellialveolina.
In conclusion, the confusion between the two genera, which cannot be further sustained, is probably due to the association of several causes: 1) the poor knowledge of the family context in the years when the taxa were established; 2) the unfortunate choice of an incomplete single section for the type of the genus Pseudedomia; 3) the rather poor material used for the type of "Pseudedomia" drorimensis; 4) above all, the discontinuity of data acquisition in time and space, which makes one of the main difficulties of our discipline.
|
Figure 9: Genus
Sellialveolina (all presumably A
tests). 1-5: a
small species from Greek Albian (1-4: A tests; 5: B test). 6-7:
"Ovalveolina"
maccagnoae from Albian-Cenomanian
of Italy. 8-9: S. viallii,
"primitive" specimens with few secondary chamberlets from Greece. 10-11: S. viallii, axial and equatorial sections of the type material
(10: holotype). 12-17: S.
viallii, various morphologies and endoskeleton organization from Albian-Cenomanian
of Greece and Italy. 18-20:
S. viallii, transitional
population to S. drorimensis with
better organized endoskeleton (BSC and SSC in 18-19) and pseudoevolute terminal
stage, from Cenomanian of Turkey. 21-25: Sellialveolina drorimensis from Upper Cenomanian of France,
Lebanon, Algeria and Israel, respectively, with well-organized endoskeleton and
frequent final uncoiling; note the BSC-SSC structure in section 21 (see Fig.
13.27 |
5.9 - Genera Rhapydionina Stache, 1913, and Fanrhapydionina Fleury, 2014
Rhapydionina
liburnica (Stache, 1889) is the first Rhapydioninidae named, although it
was understood as such much later by De
Castro (1972) who showed that its endoskeleton is typically of
alveolinid type. Distinction from other genera in the family resulted from
works of several authors, taking into account mainly the early uncoiling of
the A tests and the marked dimorphism between generations (A test, mainly
cylindrical; B test, flabelliform: Fig. 10.1-7 ![]() ; see also Fleury,
2014, text-fig. 1). The older species described by Fleury, 2014 (R.
gr. dercourti), show that these
criteria are perfectly suitable for a genus following a special evolutionary
trend, while faithfully respecting the same proportion in the endoskeleton
between a large peripheral zone (with cloisonnettes) and a central
endoskeleton of comparative small diameter. The same criteria, associated with
bilaterally compressed A tests of Fanrhapydionina
led Fleury (2016) to
restrict the Rhapydionininae to these two genera, and distinguish the Cuvillierinellinae
characterized by a narrow peripheral zone. This criterion is decisive: No
taxon can be attributed to these genera without careful examination of
transverse sections perpendicular to the axis of evolute chambers.
; see also Fleury,
2014, text-fig. 1). The older species described by Fleury, 2014 (R.
gr. dercourti), show that these
criteria are perfectly suitable for a genus following a special evolutionary
trend, while faithfully respecting the same proportion in the endoskeleton
between a large peripheral zone (with cloisonnettes) and a central
endoskeleton of comparative small diameter. The same criteria, associated with
bilaterally compressed A tests of Fanrhapydionina
led Fleury (2016) to
restrict the Rhapydionininae to these two genera, and distinguish the Cuvillierinellinae
characterized by a narrow peripheral zone. This criterion is decisive: No
taxon can be attributed to these genera without careful examination of
transverse sections perpendicular to the axis of evolute chambers.
These
genera are cited here because of the aspect of transversal sections through
the uncoiled part, which appears promising: Primary and secondary chamberlets
are well differentiated and the secondary ones are typically scattered in the
central endoskeleton (see Fleury,
2018, text-fig. 2.19-23 and Fig. 10.3-5 ![]() ).
Nevertheless, the BSC was never undoubtedly observed. It could be possibly
present in a few sections of the initial involute stage (see Figs.
10.2
).
Nevertheless, the BSC was never undoubtedly observed. It could be possibly
present in a few sections of the initial involute stage (see Figs.
10.2 ![]() and 13.28
and 13.28 ![]() ), but the association of such a very short and tight spiral
stage and the large peripheral zone makes this observation highly improbable.
The confluent structure was never seen in the large number of sections of many
rich populations observed from various localities from the Mediterranean area.
), but the association of such a very short and tight spiral
stage and the large peripheral zone makes this observation highly improbable.
The confluent structure was never seen in the large number of sections of many
rich populations observed from various localities from the Mediterranean area.
Accessorily,
Fig. 10.12-16 ![]() shows a rather unusual
apparent differential sensitivity to diagenesis of the test which seems to
support the traditional distinction between exo- and endoskeleton.
shows a rather unusual
apparent differential sensitivity to diagenesis of the test which seems to
support the traditional distinction between exo- and endoskeleton.
|
Figure 10: Genera Rhapydionina and Fanrhapydionina.
1-7: Rhapydionina gr.
liburnica. 1-2: Equatorial and axial centered section of A tests; 3-5:
Transverse sections of the cylindrical evolute part (UUT); 6-7: B tests (6:
External aspect of the flat test; 7: Part of a transverse section through the
flat evolute part). 8-10: Fanrhapydionina
flabelliformis. 8-9: Equatorial and axial centered sections of A tests;
10: Transverse section of evolute part of a presumed A test. 11:
Rhapydionina model of a chamber from evolute cylindrical part of A
test (see legend). 12-16: Rhapydionina
liburnica, A tests. Color pictures showing the apparent differential
sensibility of the exo- and endoskeleton to diagenesis. 12: Centered equatorial
section showing differentiation of a light-brown colored external layer (wall
and septa: exoskeleton) and grey internal partitions (cloisonnettes and central
endoskeleton), the last chamber being wholly grey. 13 and 15: Tangential and
transverse oblique sections of UUT showing again the exoskeleton (light-brown)
and the endoskeleton (grey) with rather sharp limits. 14: Transverse section
through a chamber of a cylindrical UUT, tangential to the septum of previous
chamber; internal part of wall and central endoskeleton pierced by scattered
secondary chamberlets appear grey; external layer of wall and septum (pierced by
openings) of previous chamber appear light-brown. 16: Centered axial section
showing an almost whole light-brown colored test. Scale bars 1-10: 1mm; 12-16:
0.5mm. |
5.10 - Genera Chubbina, Praechubbina, Raadshoovenia, Neomurciella and Twaraina from Central America
Chubbina
Robinson, 1968, from the Campanian - Maastrichtian around the
Gulf of Mexico was described with a correctly interpreted endoskeleton of the
"Pseudedomia" globularis
type (see Pseudochubbina globularis, §5.1), and a coiling mode
"streptospiral in the early
stages, becoming almost planispiral in the last whorls of the microspheric
generation", which is called pseudoplanispiral in this work and seems to be
the "brand mark" of a majority of the American Upper Cretaceous and Lower
Cenozoic Rhapydioninidae branches: The Chubbininae Dilley,
1973, and Neomurciellinae Fleury
& Fourcade, 1990. The
various species of this genus are all characterized by a pseudoplanispiral
coiling in adult A tests and also in the B tests before their large final
uncoiled part (see Fleury
& Fourcade, 1990,
text-fig. 8.B). The proloculus diameters are given on Fig.
12 ![]() after the original description. The two species created by Robinson
(1968) are still difficult to distinguish in detail, but their individual
features are of little interest for our search: All sections show the
characteristic triad of the primary chamberlets and BSC-SSC structure (Figs.
11.1-5
after the original description. The two species created by Robinson
(1968) are still difficult to distinguish in detail, but their individual
features are of little interest for our search: All sections show the
characteristic triad of the primary chamberlets and BSC-SSC structure (Figs.
11.1-5 ![]() and 13.31-32
and 13.31-32 ![]() ). The
diameters of the chamberlets given on Fig. 12
). The
diameters of the chamberlets given on Fig. 12 ![]() were measured on probable populations of the two species (EJF1 for C.
jamaicensis Robinson, 1968;
EJF33 for C. macgillavryi Robinson, 1968); they are not very different from the measures given by Robinson
(1968), who did not distinguish the various types of chamberlets (40-55 µm
for jamaicensis, 25-50 for macgillavryi).
The confluent structure was never observed in several rich populations of the
genus.
were measured on probable populations of the two species (EJF1 for C.
jamaicensis Robinson, 1968;
EJF33 for C. macgillavryi Robinson, 1968); they are not very different from the measures given by Robinson
(1968), who did not distinguish the various types of chamberlets (40-55 µm
for jamaicensis, 25-50 for macgillavryi).
The confluent structure was never observed in several rich populations of the
genus.
Praechubbina
Fourcade & Fleury, 2001. This genus was initially represented by six Campanian species from
Mexico, all of them known by both generations. They are wholly streptospiral
coiled with a relatively small A proloculus (between 60 and 130 µm), and a
short final uncoiled stage rarely developed in B tests. They are considered as
intermediate between Pseudonummoloculina
pecheuxi Fourcade & Fleury,
2001, and the various species of Chubbina
from the same area. The type species, P.
breviclaustra, in particular, shows an aperture system passing from a
"notched" type to a double row of circular foramina (see Fourcade
& Fleury, 2001,
text-fig. 4). In general, the relatively large diameter of the chamberlets is
not favorable to observation of the complete BSC-SSC structure but, by
exception, the holotype of P.
streptospira (Figs. 11.6 ![]() and 13.33
and 13.33 ![]() ), shows a well differentiated layer of basal
chamberlets.
), shows a well differentiated layer of basal
chamberlets.
Raadshoovenia
Van Den Bold, 1946. R.
guatemalensis Van Den Bold, 1946, the type species (Figs. 11.7-8 ![]() and 13.30
and 13.30 ![]() ), was at first only known
by its "early chambers quinqueloculine (...) adult uncoiling (...) labyrinthic
chambers developed in later stages" and some schematic drawings. De Castro
(1971) was the first to describe the true
alveolinid endoskeleton of this American Cenozoic taxon. The endoskeleton
closely resembles A tests of the type species of Cuvillierinella and Murciella,
and De Castro considered
that these two genera are to be considered synonymous of Raadshoovenia. This conception was not followed by Fleury
(2016) who showed that Raadshoovenia
and Cuvillierinella represented different evolutionary lineages on
either sides of the Atlantic, at different periods, and are probably
illustrating a convergence resulting from an ontogenetic recapitulation of
phylogenesis. R. guatemalensis, wholly
streptospiral, is crudely constructed with relatively large chamberlets and shows
an exceptional and slight tendency to a final uncoiled stage. The BSC-SSC
structure is clearly visible; the diameter of primary chamberlets and
chamberlets of the BSC is the same, relatively large (40 to 50 µm on a few
sections in samples EJF26 and 104); the diameter of the scattered chamberlets
is smaller (about 20-40 µm). No confluent structure was observed.
), was at first only known
by its "early chambers quinqueloculine (...) adult uncoiling (...) labyrinthic
chambers developed in later stages" and some schematic drawings. De Castro
(1971) was the first to describe the true
alveolinid endoskeleton of this American Cenozoic taxon. The endoskeleton
closely resembles A tests of the type species of Cuvillierinella and Murciella,
and De Castro considered
that these two genera are to be considered synonymous of Raadshoovenia. This conception was not followed by Fleury
(2016) who showed that Raadshoovenia
and Cuvillierinella represented different evolutionary lineages on
either sides of the Atlantic, at different periods, and are probably
illustrating a convergence resulting from an ontogenetic recapitulation of
phylogenesis. R. guatemalensis, wholly
streptospiral, is crudely constructed with relatively large chamberlets and shows
an exceptional and slight tendency to a final uncoiled stage. The BSC-SSC
structure is clearly visible; the diameter of primary chamberlets and
chamberlets of the BSC is the same, relatively large (40 to 50 µm on a few
sections in samples EJF26 and 104); the diameter of the scattered chamberlets
is smaller (about 20-40 µm). No confluent structure was observed.
Neomurciella Fleury
& Fourcade, 1987. N.
butterlini Fleury & Fourcade,
1987, the type species (Fig. 11.9-13 ![]() ),
is wholly streptospiral to pseudoplanispiral. This American Paleocene to
Eocene species was created when the "helicoidal structure" (called
confluent structure in the present work) was only identified in the European
genus Murciella. This structure, still unknown in other American taxa, is
now observed in several European genera (see Fig. 12
),
is wholly streptospiral to pseudoplanispiral. This American Paleocene to
Eocene species was created when the "helicoidal structure" (called
confluent structure in the present work) was only identified in the European
genus Murciella. This structure, still unknown in other American taxa, is
now observed in several European genera (see Fig. 12 ![]() ) and thus cannot be considered by itself as a definitive
criterion. The species is nevertheless worthy of being the type of a valid
genus, included in a particular lineage comprising Raadshoovenia (see Fleury,
2016, text-fig. 4), probably the Eocene genera Twaraina and (possibly) Yaberinella
as supposed by Fleury & Fourcade
(1987, 1990). The A test has a very small proloculus, and there is a large
flat uncoiled terminal stage in B tests which is especially remarkable. Its
endoskeleton is typically of the rhapydioninid type, with a clear BSC-SSC
structure (Figs. 11.9-10, 11.12
) and thus cannot be considered by itself as a definitive
criterion. The species is nevertheless worthy of being the type of a valid
genus, included in a particular lineage comprising Raadshoovenia (see Fleury,
2016, text-fig. 4), probably the Eocene genera Twaraina and (possibly) Yaberinella
as supposed by Fleury & Fourcade
(1987, 1990). The A test has a very small proloculus, and there is a large
flat uncoiled terminal stage in B tests which is especially remarkable. Its
endoskeleton is typically of the rhapydioninid type, with a clear BSC-SSC
structure (Figs. 11.9-10, 11.12 ![]() and 13.34
and 13.34 ![]() ).
The diameter of chamberlets measured on a few sections of several samples
(EJF101, 104, 106), is quite homogeneous, the primary and basal ones (forming
a well-organized BSC) are a little larger (30 to 40 µm) than the scattered
ones (25-30 µm).
).
The diameter of chamberlets measured on a few sections of several samples
(EJF101, 104, 106), is quite homogeneous, the primary and basal ones (forming
a well-organized BSC) are a little larger (30 to 40 µm) than the scattered
ones (25-30 µm).
Twaraina Robinson,
1993. T. seigliei Robinson,
1993, the type species of the genus from the early Middle Eocene of Nicaragua
develops a large flabelliform test in the A generation, around a gigantic
proloculus (300 to 600 µm in diameter) and a short involute planispiral young
part; B tests are unknown. The endoskeleton is typical of the Rhapydioninidae,
with secondary chamberlets "few in number" (that is to say, according to
the figured section, properly scattered) and "more or less spirally
directed" which could correspond to a rudimentary confluent structure; the
BSC is well differentiated, with chamberlets of small diameter (Fig.
11.14 ![]() ).
).
Concerning these American genera, the BSC-SSC organization in all of them and the typical confluent structure in at least one of them seem to give a new important clue to the probability of a fundamental connection between the Euro-Asiatic and American Rhapydioninidae. But it must not be forgotten that the various resemblances between organisms of the two sides of the Atlantic are most probably only due to their common heritage: They are all included in different local evolutionary sequences (Fleury, 2016, text-fig. 4). This is particularly obvious for the American Cenozoic ones which, at this time, have no equivalent in the Old World.
|
Figure 11:
American genera with BSC-SSC structure.
1-5: Genus Chubbina (not
precisely identified species). 1: Axial section of a possible C.
macgillavryi (A test); 2-5: Axial sections of possible C.
jamaicensis (A tests). 6: Praechubbina streptospira (axial section of A test). 7-8:
Raadshoovenia guatemalensis (axial
sections of A tests). 9-13: Neomurciella butterlini.
9-10, 12: Axial and subaxial sections of A tests; 11: Axial section of a B test;
13: Part of an equatorial section of a probable B test with confluent structure.
14: Twaraina seigliei: Axial
section of an A test. Scale bars: 1-12: 1mm; 13: 0.5mm. |
The main
result of the previous examination of almost all known Rhapydioninidae is
obvious. The BSC-SSC structure, observed in most of them cannot be considered
as a criterion for drawing a distinction between genera. Even if some
particularities of some species or genera (chamberlets too large in C.
salentina and M. decastroi, or involute part too short in Cyclopseudedomia and Rhapydionina-Fanrhapydionina)
prevent the observation of the BSC, the whole structure can be considered as a
fundamental original feature of the group, only dubiously shared in part by
the eccentric genus Subalveolina
(see Reichel, 1936, Pl. IV
and Fig. 2.18-23 ![]() : SSC present and
opened in the preseptal space, but BSC apparently absent) among the
Alveolinidae. The likely generalized presence of the BSC-SSC structure leads
to several considerations about the whole endoskeleton organization of the
family.
: SSC present and
opened in the preseptal space, but BSC apparently absent) among the
Alveolinidae. The likely generalized presence of the BSC-SSC structure leads
to several considerations about the whole endoskeleton organization of the
family.
6.1 - About diameter differentiation of chamberlets.
The diameter of primary chamberlets varies considerable from 7 up to about 80 µm. There is apparently no simple relation between this diameter and observable BSC, but it is interesting to notice that the taxa with primary chamberlets of largest diameter (C. salentina, M. decastroi) coincide with tests in which the secondary chamberlets are also of large diameter and the BSC is apparently absent or difficult to observe. It seems that such a difficulty is not necessarily fundamental and could be a simple geometric question: The small number of distant chamberlets makes it difficult to simultaneously observe in plane section a sufficient number of them to appreciate their eventual presence at the base of chambers. Additionally, it is useful to recall that both species can be considered as "primitive" because of their small sized proloculus and weak dimorphism between generations; in that perspective, a clear differentiation of the BSC would be simply an evolutionary mark linked to the known general decreasing diameter of chamberlets with specialization. Anyway, both cases being doubtful, the differentiation of chamberlet sets would be a fundamental feature of the group's heritage, and only perceptible when the diameter of chamberlets is small enough.
Diameter
differentiation of secondary from primary chamberlets is frequently observed
and does not seem to be related to any other feature. The general rule is a smaller diameter for the secondary ones. The only
exception would be Pseudochubbina
kassabi (see Fig. 4.10 ![]() ), which
is only known from a small number of sections.
), which
is only known from a small number of sections.
Diameter differentiation of basal chamberlets is not the rule. Most species have a BSC with chamberlets of about the same diameter as the SSC ones. Only Pseudochubbina and Chubbina show a BSC made up of chamberlets larger than the primary or even the scattered ones, P. globularis being a noticeable extreme example. Large chamberlets forming the BSC would be a characteristic of some species with a large proloculus, Cyclopseudedomia being an exception because its short involute coiling makes observation impossible.
6.2 - About concomitance of BSC-SSC and confluent structure.
This is the main problem encountered during the preceding review. If the interpenetration of tubular chamberlets separated by thin partitions is easy to conceive, confluence of distant scattered chamberlets embedded in a compact mass is more difficult to understand. Two main examples can be considered:
Sigalveolina
displays clearly both structures (Fig. 6 ![]() ). Distance between chamberlets is relatively short, which marks a
special case. The confluent structure is obvious, resulting from probable
disposition of chamberlets in layers: they are not evident in axial sections,
but equatorial and tangential sections show that equally oriented chamberlets
isolated by thin wall (see Fleury, 2018, text-fig. 3.6;
Figs. 6.3, 6.6
). Distance between chamberlets is relatively short, which marks a
special case. The confluent structure is obvious, resulting from probable
disposition of chamberlets in layers: they are not evident in axial sections,
but equatorial and tangential sections show that equally oriented chamberlets
isolated by thin wall (see Fleury, 2018, text-fig. 3.6;
Figs. 6.3, 6.6 ![]() and 16.5
and 16.5 ![]() )
are forming homogeneous groups which can be referred to distinct layers
oriented orthogonally.
)
are forming homogeneous groups which can be referred to distinct layers
oriented orthogonally.
Cyclopseudedomia
(Fig. 7 ![]() ) does not show the BSC, but
the secondary chamberlets are dispersed without evident order in the compact
mass of the central endoskeleton and particularly do not seem to be arranged
in distinct layers (Fig. 7.3-5, 7.12
and 7.15
) does not show the BSC, but
the secondary chamberlets are dispersed without evident order in the compact
mass of the central endoskeleton and particularly do not seem to be arranged
in distinct layers (Fig. 7.3-5, 7.12
and 7.15 ![]() ).
In this case, the confluence of chamberlets is not generally obvious, but
equatorial sections through the evolute flange show that secondary chamberlets
are organized in two slightly oblique directions permitting (by chance?) some
individual junctions (Fleury,
2018, text-figs. 3.13-14, 11.14, 12.14 and 14.23; Figs. 7.6
).
In this case, the confluence of chamberlets is not generally obvious, but
equatorial sections through the evolute flange show that secondary chamberlets
are organized in two slightly oblique directions permitting (by chance?) some
individual junctions (Fleury,
2018, text-figs. 3.13-14, 11.14, 12.14 and 14.23; Figs. 7.6 ![]() and 16.13-14
and 16.13-14 ![]() ).
).
In conclusion, both structures appear somewhat complementary. The widespread BSC-SSC structure would be fundamental, with some nuance linked to the more or less thick "cloisonnage" (partitioning) of the central endoskeleton. The scarcer confluent structure would be an evolutionary feature, more or less perfectly achieved, according to particularities of the central endoskeleton frame. The presence, more or less well defined, of the confluent structure in all the genera belonging to the Cuvillierinellinae seems to be a characteristic of this subfamily, although it is occasionally observed in the Sellialveolininae, Pseudedomiinae and Neomurciellinae, in very different contexts. Its absence in the Chubbininae and Rhapydionininae, of which "advanced" representative members are well known, remains intriguing.
|
Figure 12: Main features of selected examined taxa.
*Measures after De Castro,
1990: the A proloculus diameter including the wall thickness must be reduced of
about 20-30 µm for comparison with other measures. |
|
Figure 13: Selected
interpretative sections displaying the Basal Secondary
Chamberlets-Scattered
Secondary Chamberlets structure (BSC-SSC)
in the Rhapydioninidae family. 1-6:
Pseudochubbina. 1: P.
globularis (see Fig. 4.1 |
Order Foraminifera Eichwald, 1830
Suborder Miliolina Delage & Hérouard, 1896
Superfamily Alveolinacea Ehrenberg, 1839
Family Rhapydioninidae Keijzer, 1945
Subfamily Cuvillierinellinae Fleury, 2016
Genus Metacuvillierinella Fleury, 2016
7.1 - At generic level.
As discussed before, the choice of a generic assignment for the new taxon is not obvious.
The preceding analysis indicates that the presence of various endoskeletal structures is not useful for generic assignment: The BSC-SSC structure is widespread in the family and cannot be a used for a definite attribution;
The adult coiling of both generations, characterized by pseudoplanispiral-advolute coiling leads strongly to accept the simple choice of an assignment to Metacuvillierinella.
However, the juvenile streptospiral stage of M. decastroi contrasts with the initial planispiral coiling of the new taxon and raises the general question of the passage from one to the other mode of coiling. It is generally accepted that this is a natural and ordinary evolutionary process between "primitive" to "advanced" forms in many lineages. One of the best examples is the likely affiliation between the genera Cuvillierinella and Murciella, which are most probably (in part?) of the same age, sometimes associated (Zone CsB6a, probably Campanian, according to Fleury, 2018, text-fig. 4) and closely resembling one another (see Fleury, 2018, text-fig. 16). Their A tests are involute-ovoid with an incidental cylindrical UUT (see §3.1) whereas their B tests are diverse, resembling the A tests or slightly flabelliform. Their type species differ in the following features (after Fleury, 2016, text-fig. 12 and 2018, text-fig. 15):
C. salentina has slightly smaller proloculus (mean values 70 to 120 µm) in A tests than M. cuvillieri (mean values 110 to 160 µm);
C. salentina shows streptospiral coiling in juvenile tests in the A generation, while M. cuvillieri is wholly planispiral;
C.
salentina has larger chamberlets than M.
cuvillieri (see Figs. 5 ![]() and 12
and 12 ![]() ).
).
In short, these two taxa are clearly distinct at the specific level, but generic discrimination between them is more debatable and somewhat arbitrary. Maintaining a long accepted taxon such as Murciella was partly decided for the sake of nomenclature stability, but mainly because of the wide geographic distribution of both taxa (Spain to Turkey) and the recorded variations around each type: C. salentina is related to several species attributed to the same genus (in Fleury, 2016) and M. cuvillieri is represented by diversified populations (in Fleury, 2018). Thus, the generic distinction between Cuvillierinella and Murciella is a matter of general perspective and particularly linked to their radiance (see §8.2). The facts are not decisive by themselves, but this is the large knowledge of the context which leads to a decision.
The case for the new taxon is different: In the current state of knowledge, it seems isolated and could be a local variation of M. decastroi, the type species of the genus. The discovery of a posterity or a kinship would certainly modify our conclusion but, anyway, nothing in the relatively well-known Rhapydioninidae populations of the western and central Mediterranean area could fit such an eventuality. More eastern regions will provide a decisive choice.
7.2 - As a new species: formal identification of Metacuvillierinella sireli n. sp.
Diagnosis. Species reported with some reserve to the genus Metacuvillierinella Fleury, 2016. A generation: Nautiloid, slightly flattened tests of 1 to 3 mm in equatorial diameter; proloculus subspherical of about 200 µm followed by a flexostyle; first coils mainly planispiral, adult coiling pseudoplanispiral resulting in a sigmoid appearance in axial section; last coil advolute, never uncoiled, surrounding a large bilateral umbilicus. B generation: Tests resembling the A generation, but larger (up to about 5 mm in equatorial diameter); nepionic stage streptospiral, adult pseudoplanispiral with large bulging last coil, leading to a dumbbell-like axial section. Endoskeleton: BSC-SSC organization of chamberlets, with no diameter differentiation, but enlarged in last chambers of B tests; preseptal space relatively narrow, equipped with rare buttresses.
Material and deposition. The material of this study includes both core and
surface samples. The core (core number: 1, depth: 2832-2941 m) samples were
obtained from the well Yemişlik-10 drilled in the Yemişlik oil
field, which is located about 20 km to the southeast of the Town Kozluk of the
City Batman. The GCA14 coded surface samples (Fig. 14.17 ![]() )
have been previously collected in the vicinity of the City Mardin by
colleagues from TPAO (see Fig. 1.C
)
have been previously collected in the vicinity of the City Mardin by
colleagues from TPAO (see Fig. 1.C ![]() for the location of samples). In total, 20 thin sections (TS-1 - TS-20) from
the core samples and 4 thin sections from surface samples (GCA14-100 -
GCA14-104) were examined. Samples and thin sections are housed in the Ar-Ge
Center, TPAO, Ankara.
for the location of samples). In total, 20 thin sections (TS-1 - TS-20) from
the core samples and 4 thin sections from surface samples (GCA14-100 -
GCA14-104) were examined. Samples and thin sections are housed in the Ar-Ge
Center, TPAO, Ankara.
Holotype.
(Figs. 2.13 ![]() and 14.14
and 14.14 ![]() ).
Almost axial section of an A test, of about 1.5 mm in equatorial diameter.
Internal diameter of proloculus in axial section is 200 µm (225 with wall).
Flexostyle clearly distinguished, opened in the proloculus (in its proximal
part probably). Primary chamberlets present in the first sectioned chamber,
not visible in the second (to the bottom), possibly cut through the preseptal
space. The following three coils gradually increase in height, with the
exception of the last one, although the preceding coil is partly covered,
leaving a wide umbilicus on one side of the section. The endoskeleton
comprises chamberlets with rather larger diameters in the first and last coils
than in the coils of middle part.
).
Almost axial section of an A test, of about 1.5 mm in equatorial diameter.
Internal diameter of proloculus in axial section is 200 µm (225 with wall).
Flexostyle clearly distinguished, opened in the proloculus (in its proximal
part probably). Primary chamberlets present in the first sectioned chamber,
not visible in the second (to the bottom), possibly cut through the preseptal
space. The following three coils gradually increase in height, with the
exception of the last one, although the preceding coil is partly covered,
leaving a wide umbilicus on one side of the section. The endoskeleton
comprises chamberlets with rather larger diameters in the first and last coils
than in the coils of middle part.
Etymology. In honor of Ercument Sirel, a well-known Turkish micropaleontologist who specialized in the Meso-Cenozoic larger foraminifera.
Description
Architecture:
A generation (Figs. 2.12-13 ![]() , 3.1-4
, 3.1-4 ![]() , 14.1-15, 14.17
and 14.20-24
, 14.1-15, 14.17
and 14.20-24 ![]() ).
The test, known by about 20 centered axial and rare equatorial sections, is
slightly nautiloid, laterally compressed, of rather constant thickness (0.5 to
0.8 mm), the equatorial diameter being larger than 1 mm and up to about 3 mm.
The proloculus is almost spherical (diameter: inner 186 ± 40, outer 220 ± 40
µm), followed by a distinct flexostyle of about half a coil long. The first
coils apparently almost planispiral are followed by adult coils which are
apparently planispiral, with a tendency of forming twisted tests in axial
sections (pseudoplanispiral coiling). The last coil covers only a part of the
preceding one, leaving a large umbilicus (advolute coiling, see Fig.
2.2
).
The test, known by about 20 centered axial and rare equatorial sections, is
slightly nautiloid, laterally compressed, of rather constant thickness (0.5 to
0.8 mm), the equatorial diameter being larger than 1 mm and up to about 3 mm.
The proloculus is almost spherical (diameter: inner 186 ± 40, outer 220 ± 40
µm), followed by a distinct flexostyle of about half a coil long. The first
coils apparently almost planispiral are followed by adult coils which are
apparently planispiral, with a tendency of forming twisted tests in axial
sections (pseudoplanispiral coiling). The last coil covers only a part of the
preceding one, leaving a large umbilicus (advolute coiling, see Fig.
2.2 ![]() ). B generation (Figs.
3.5-6
). B generation (Figs.
3.5-6 ![]() , 14.16, 14.18-19
, 14.16, 14.18-19 ![]() and 15.1-9
and 15.1-9 ![]() ).
Test larger than the A ones, from about 3, up to more than 5 mm, more or less
twisted as a whole, even distorted with somewhat dumbbell-like axial section
due to the enlarged protruding last coil, leaving a wide umbilicus (advolute
coiling); the proloculus was never seen, but a lot of apparently unorganized (streptospiral
?) small chambers in the center of first coils indicates the presence of the
usual young part of the B tests in the family (a glomerulus); the following
coils resemble the A generation, almost planispiral with the same regular
growth in equatorial and axial directions, but the last coil resulting in a
doubling of the equatorial diameter and thickness.
).
Test larger than the A ones, from about 3, up to more than 5 mm, more or less
twisted as a whole, even distorted with somewhat dumbbell-like axial section
due to the enlarged protruding last coil, leaving a wide umbilicus (advolute
coiling); the proloculus was never seen, but a lot of apparently unorganized (streptospiral
?) small chambers in the center of first coils indicates the presence of the
usual young part of the B tests in the family (a glomerulus); the following
coils resemble the A generation, almost planispiral with the same regular
growth in equatorial and axial directions, but the last coil resulting in a
doubling of the equatorial diameter and thickness.
Endoskeleton.
The primary chamberlets appear as soon as the first coil, around the
proloculus of A tests (Fig. 14.1, 14.3-4, 14.9-11
and 14.14 ![]() ) and remain well differentiated as a distinct layer in all
tests (Figs. 14.14, 14.17
) and remain well differentiated as a distinct layer in all
tests (Figs. 14.14, 14.17 ![]() and 15.2-5
and 15.2-5 ![]() , particularly). The BSC-SSC structure is quite well
represented too, although the BSC is sometimes relatively unclear because of
the distance between chamberlets; but in some cases it is nevertheless
undoubted (i.e., Figs. 2.12-13
, particularly). The BSC-SSC structure is quite well
represented too, although the BSC is sometimes relatively unclear because of
the distance between chamberlets; but in some cases it is nevertheless
undoubted (i.e., Figs. 2.12-13 ![]() , 14.14
, 14.14 ![]() , 15.2
and 15.4
, 15.2
and 15.4 ![]() , last chambers). As a whole, the diameter of all chamberlets in
the same chamber is equal, but this diameter is different in various tests of
the same generation (i.e., Fig. 14.9-10
, last chambers). As a whole, the diameter of all chamberlets in
the same chamber is equal, but this diameter is different in various tests of
the same generation (i.e., Fig. 14.9-10 ![]() ) and the diameter increases from the early coils to the later
ones, reaching its maximum in the last chambers of B tests. The preseptal
space is relatively narrow and obscured by crystallization or micritic
infilling in most of the sections; it can be observed in Fig.
14.20
) and the diameter increases from the early coils to the later
ones, reaching its maximum in the last chambers of B tests. The preseptal
space is relatively narrow and obscured by crystallization or micritic
infilling in most of the sections; it can be observed in Fig.
14.20 ![]() (three chambers west and south-west of proloculus, one chamber
south-east of proloculus, last whorl with probable preseptal pillars) and Fig.
14.23
(three chambers west and south-west of proloculus, one chamber
south-east of proloculus, last whorl with probable preseptal pillars) and Fig.
14.23 ![]() (center of the section, crossed by 5 thin partitions: Preseptal
pillars or possibly end of cloisonnettes in periphery of preseptal space);
section Fig. 15.4
(center of the section, crossed by 5 thin partitions: Preseptal
pillars or possibly end of cloisonnettes in periphery of preseptal space);
section Fig. 15.4 ![]() shows this structure in axial section, with one thin
pillar. Apart from this one, the only undoubted preseptal pillars are from
last chambers of some B tests (Fig. 15.1-2
shows this structure in axial section, with one thin
pillar. Apart from this one, the only undoubted preseptal pillars are from
last chambers of some B tests (Fig. 15.1-2 ![]() ).
).
Comparisons.
The pseudoplanispiral coiling and the advolute last whorl are common to the
new species and M. decastroi (Fig. 3 ![]() );
this last one is at first glance differentiated by a quinqueloculine-like
juvenile stage and chamberlets of large diameter. The new species is well
characterized, even in non-centered sections, by some really distinctive
details: A tests are flat with a relatively large proloculus and B tests have
dumbbell-like axial sections (more or less distorted) and contrasted aspect of
thin chamberlets forming the rosary-like primary and basal layers with the
scattered ones few in number.
);
this last one is at first glance differentiated by a quinqueloculine-like
juvenile stage and chamberlets of large diameter. The new species is well
characterized, even in non-centered sections, by some really distinctive
details: A tests are flat with a relatively large proloculus and B tests have
dumbbell-like axial sections (more or less distorted) and contrasted aspect of
thin chamberlets forming the rosary-like primary and basal layers with the
scattered ones few in number.
|
Figure 14: Metacuvillierinella sireli
n. sp., mainly A
tests. 1-14: Centered axial sections.
The initial coiling of some tests (3-4, 9, 13-14) is almost perfectly
planispiral, but others (1-2, 5-8, 11-14) show various aspects of the first
coils that, as well as faintly twisted overall profile, testify the persisting
streptospiral tendency (pseudoplanispiral coiling). Note the flexostyle in 3, 9,
12, and 14; the first layer of primary chamberlets is already present in the
first coil. 15-19: Off centered
sections displaying the overall twisted profile of sigmoid type (16, and 18-19
could be B tests). 20-22, 24:
Centered equatorial sections. Poor preservation of the tests prevents detailed
observations but the flexostyle is well observable in 20-21 and 22; the
succession of 2 chambers with preseptal space is clear in 20 (next to last coil,
south-west of the proloculus); 24: usual aspect of this type of section, partly
hidden by micritization. 23: Off
centered section of which the central coil shows a preseptal space crossed by
thin probable preseptal pillars; they are much larger in last chamber (see also
20, last coil, south-east of the proloculus). Scale bars: 1mm. |
Range and occurrence. A Campanian ephemeral platform between two pelagic episodes
The Cretaceous succession drilled in the studied well encompasses, from 2600 to 2816 m, Germav, Garzan, Kıradağ and Beloka formations, and finally the unit containing the new taxon M. sireli. The Beloka Formation consists of two informal subdivisions consisting of shallow-marine (Bada Member) and deeper-marine (Dirik Member) carbonates. The Dirik Member comprises 14 m of argillaceous limestones with planktonic foraminifera, especially Globotruncanita elevata (Brotzen, 1934) and Radotruncana calcarata (Cushman, 1927). These species are sometimes found in the same sample (which seems a peculiarity of the Eastern Mediterranean area). Thus, the Beloka Formation is approximately attributed to the Calcarata Zone of the upper Campanian.
The
carbonate unit underlying the Beloka Formation consists of cream, light-gray
colored, about 11 m thick (from 2832 to 2843 m) limestones that contain the
tests of M. sireli n. sp. The whole
unit is made up of predominant wackestone grading rarely in packstone texture.
The microfacies contains abundant miliolid
foraminifers with rare other foraminifers: Cuneolina
sp., possible Nezzazatinella sp.
(Fig. 15.10 ![]() ), Fleuryana?
sp. (Fig. 15.11-15
), Fleuryana?
sp. (Fig. 15.11-15 ![]() , possibly
comparable to F. sp. in
De Castro et
al., 1994, Pl. 4, figs. 9-11, but without observed opening), rotaliids and
discorbids. Common echinoderms and dasycladacean algae as well as rare
ostracods, gastropods and bivalve shells also form the skeletal component of
the rock. The non-skeletal component is formed by abundant very fine- to
fine-grained peloids and rare quartz grains. The matrix is generally
characterized by fine-grained micrite. Dissolution vugs filled with calcite
cement are locally developed. The skeletal association and fine-grained
texture indicate that this facies was deposited in a low-energy, lagoonal
environment. The well is terminated with 2 m thick of dolomite.
, possibly
comparable to F. sp. in
De Castro et
al., 1994, Pl. 4, figs. 9-11, but without observed opening), rotaliids and
discorbids. Common echinoderms and dasycladacean algae as well as rare
ostracods, gastropods and bivalve shells also form the skeletal component of
the rock. The non-skeletal component is formed by abundant very fine- to
fine-grained peloids and rare quartz grains. The matrix is generally
characterized by fine-grained micrite. Dissolution vugs filled with calcite
cement are locally developed. The skeletal association and fine-grained
texture indicate that this facies was deposited in a low-energy, lagoonal
environment. The well is terminated with 2 m thick of dolomite.
On the other hand, the limestones with M. sireli n. sp. crop out in the Mardin region where resting on the pelagic deposits of the Karababa Formation, which, based on planktonic foraminifers, was recently determined to range in age from late middle Turonian to late Santonian (Özkan & Altiner, 2019).
Consequently, in its type region, M. sireli n. sp., can be considered Campanian in age, either older than Calcarata Zone, or possibly Campanian, including part of the Calcarata Zone.
|
Figure 15: Metacuvillierinella sireli
n. sp., B tests,
and some associated foraminifera. 1-8:
Sub-centered axial sections of M. sireli
n. sp. displaying the large umbilicus left by the partial covering of the test
by the last coil. The pseudoplanispiral coiling is only a matter of inference in
the initial parts, but obvious in the overall sigmoid adult tests (especially
evident in 5). Note some rarely observed details: -pillars in the preseptal
space (chambers of the last coil in 1
and 2, possible in 5 and 6),
-triangular termination of the cloisonnettes in 4 (chamber of the next to last
coil, to the bottom). 9: Equatorial
section, a kind of ghost, almost completely micritized. 10:
Nezzazatinella sp. probable. 11-15:
? Fleuryana sp.: 11-12: Oblique non centered sections; 13-15: Equatorial
and axial centered sections. 16-20: Fleuryana
adriatica De Castro et al., 1994, for comparison: Axial, equatorial centered and oblique
off centered sections. Opening can be seen in 16 (last chamber), in 17 and 20
(last coil). Scale bars: 1-7, 9: 1 mm; 8, 10-20: 0.5 mm. |
The new species is an addition to a prolific family. Its study gives the opportunity to review the diversity of the group and leads us to reconsider the criteria which have been used for the distinction of genera. As the understanding of the family has developed gradually, little by little, there is no likelihood for a perfect coherence in the authors' methodology, as can be seen in the following historical review.
8.1 - Historical review of chosen criteria for generic distinctions
Rhapydionina Stache, 1913 (together with "Rhipidionina", the B form of R. liburnica) and Fanrhapydionina Fleury, 2014. In that case, both particularities of architecture (particular dimorphism) and endoskeleton (relative importance of the peripheral zone and central endoskeleton, absence of confluent structure) distinguish these genera from all others in the family and characterize the Rhapydionininae.
Raadshoovenia Van Den Bold, 1946. The almost identical A test morphology of the American R. guatemalensis and the type of the Upper Cretaceous European Cuvillierinella led several authors to follow De Castro (1971) in considering the second as a junior synonym of the first. But the Paleocene-Lower Eocene age of Raadshoovenia was confirmed by Pęcheux (1984) who proposed to give up this synonymy. This fact was taken into account by Fleury & Fourcade (1990) and later by Fleury (2016), who showed that both taxa were parts of separate lineages and are thus fruits of a convergence or, rather, ontogenic recapitulation of recurring intermediate links from a group of Miliolacea to the Alveolinacea. In that case, geographic, chronologic and ontogenic arguments are convergent.
Pseudedomia Henson, 1948. The type species of the genus was indirectly understood as an Alveolinacea by Smout (1963), through its supposed analogies with "Pseudedomia" complanata, as discussed before. The genus was originally justified by Henson in comparison with others genera now classified as Soritacea (Meandropsinidae) which cannot be considered here.
Sellialveolina Colalongo, 1963, was originally recognized as an alveolinid by its endoskeleton. There was at that time no known alveolinid sharing the fine endoskeleton mesh, the small test size and absence of final uncoiling of the type species, C. viallii Colalongo, 1963; consequently the author was led to create the new genus, obviously original. Reiss et al. (1964), ignoring Colalongo's work, created a new species equipped with the alveolinid endoskeleton and a final uncoiling; thus, in those particular conditions, the new species was logically attributed to Pseudedomia, as P. drorimensis Reiss et al., 1964. Unfortunately, this inadvertent error was largely spread, particularly by textbooks, and its consequences took a long time to be appreciated. Anyway, we tried and show previously that the two genera have nothing in common, except at the family level and the time of their description. Moreover, Sellialveolina is Albian-Cenomanian in age and various taxa more or less certainly attributed to Pseudedomia are Campanian-Maastrichtian. In addition to absence of resemblance, the ultimate reason is a reference to the context: There is no probability for a link between them through the Turonian-Senonian times. The uncertainties about the type of Pseudedomia and the following attributions to the genus make an inextricable situation which cannot be solved at present; although there is no true resemblance between them, the chronologic argument takes all its importance.
Cuvillierinella Papetti & Tedeschi, 1965. This genus is here considered as distinct from Raadshoovenia (see above). C. salentina, its type species, resembles in both generations M. cuvillieri, type species of Murciella. They were precisely compared above for the generic assignment of M. sireli n. sp. (§7.2). The conclusion was that, if morphologically very close (although their young stage is of milioline type in Cuvillierinella and planispiral in Murciella), their wide geographic distribution and variations around each type seem to justify their distinction. In this case, the context helps interpretation of the doubtful significance of the morphological features.
Murciella Fourcade, 1966, was created soon after Cuvillierinella which was then poorly known. We just said before what can be concluded at present, giving the full importance to the radiance argument (see below, §8.2).
Chubbina Robinson, 1968, is known by several species difficult to distinguish in detail one from another. Several other older species of more primitive aspect were joined together in Praechubbina, which assigned a local origin to Chubbina (see Fleury, 2016, text-fig. 4). For this genus, this is the conjunction of persistent pseudoplanispiral coiling, absence of confluent structure and close presence of a likely ancestor, together with the geographic distribution which make the distinction from all other taxa.
Cyclopseudedomia Fleury, 1974. Four species from upper Campanian-lower Maastrichtian (CsB6b zone) are known in the periadriatic region. They are all characterized by obvious dimorphism (large proloculus in A tests and strong tendency to develop a large flat uncoiled terminal part in B tests) and full planispiral coiling (except initial part of B tests). The large proloculus in A tests, well-marked dimorphism and extreme uncoiling tendency are here the markers of a particular lineage.
Senalveolina Fleury, 1984. This genus is mainly known by a rich population of late Santonian to early Campanian age from Greece. S. aubouini Fleury, 1984, the type species, resembles the Albian-Cenomanian genus Ovalveolina, with subspherical tests in both generations, but is distinguished by its streptospiral young part and two layers of large chamberlets in last chambers of both generations. The large chamberlets never show a layout of the BSC-SSC type. The distinctive criterion is here the conjunction of a massive architecture of Ovalveolina type with a rudimentary but typical rhapydioninid endoskeleton.
Pseudochubbina
De Castro, 1990. The particularities of this genus was
previously discussed; it comprises four species forming a rather homogeneous
set. Like the type species, P. globularis
(Smout, 1963), the subglobular A tests, with rather large
proloculus (see Fig. 12 ![]() ), are
pseudoplanispiral throughout and show a late tendency to uncoil, well
developed in two species; rare non centered sections of globular tests, most
probably deprived of uncoiled terminal part, could correspond to the B
generation. De Castro (like Fleury,
1977, p. 85; Fleury & Fourcade,
1990, p. 260) was impressed by the homogeneity of a group of species
morphologically not very different from Chubbina
of about the same age, from which they are separated by the Atlantic Ocean. A
possible relationship with Cuvillierinella
perisalentina was presented by Fleury
(2016, p. 221). The (partial?) resemblance between P. kassabi and Cyclopseudedomia
is still tricky. Together with the still problematic B forms of Pseudochubbina
species, the paleogeographic context appears here predominant in contrasting
with Chubbina.
), are
pseudoplanispiral throughout and show a late tendency to uncoil, well
developed in two species; rare non centered sections of globular tests, most
probably deprived of uncoiled terminal part, could correspond to the B
generation. De Castro (like Fleury,
1977, p. 85; Fleury & Fourcade,
1990, p. 260) was impressed by the homogeneity of a group of species
morphologically not very different from Chubbina
of about the same age, from which they are separated by the Atlantic Ocean. A
possible relationship with Cuvillierinella
perisalentina was presented by Fleury
(2016, p. 221). The (partial?) resemblance between P. kassabi and Cyclopseudedomia
is still tricky. Together with the still problematic B forms of Pseudochubbina
species, the paleogeographic context appears here predominant in contrasting
with Chubbina.
Neomurciella
Fleury & Fourcade, 1987. N. butterlini, its type
species, has thinner chamberlets and a looser coiling than Raadshoovenia and develops a very large pseudoevolute to evolute
flat terminal flange in B tests. The confluent structure is well developed (Fig.
11.13 ![]() ). The genus is thus characterized by an association of the confluent
structure and marked tendencies to uncoiling, together with its geographic and
stratigraphic distribution.
). The genus is thus characterized by an association of the confluent
structure and marked tendencies to uncoiling, together with its geographic and
stratigraphic distribution.
Twaraina Robinson, 1993. The type species, T. seigliei Robinson, 1993, from the Central American Eocene closely resembles Cyclopseudedomia in its planispiral A tests with large proloculus and flabelliform flange, although the B form is still unknown. The author envisages a relation with Neomurciella "which may be considered as possibly ancestral to Twaraina". The absence of a well-developed confluent structure is here advocated for discrimination from Neomurciella, but the difference of age and geographic situations for distinction from Cyclopseudedomia are not even mentioned by the author, so much they are evident. We are here confronted with a likely convergence.
Praechubbina Fourcade & Fleury, 2001. This group of 6 species was in part revisited by Vicedo et al. (2013), who created the new genus Chubbinella, founded on difference in openings morphology and degrees of endoskeleton development; the question is much complex and cannot be treated here in the core. Concerning our present search, Vicedo et al. (2013) stated, in particular: "Some characteristics used by Fourcade & Fleury (2001) to differentiate Praechubbina from Chubbina, such as the dimorphism, the size of the megalosphere, the number of medullar" (=secondary) "chamberlets and the first appearance of floors in the chamber lumen must be considered diagnostic criteria of specific rather than generic rank". The sound of such a statement is definitively opposite to the previously recalled tendencies to appreciate the facts in the light of context. In the present point of view, this is precisely the combination of such criteria, together with its particular age and radiance (see §8.2), shared by several species, which ascertain the generic status of this group.
Metacuvillierinella Fleury, 2016. M. decastroi, the type species, looks like Cuvillierinella salentina by its streptospiral juvenile stage, wide endoskeleton mesh and large preseptal space; it differs through its pseudoplanispiral-advolute coiling, absolute resistance to final uncoiling, very low dimorphism of generations and younger age. The relatively wide range of the diameter of the A test proloculus in various populations from Greece and Italy led us to propose an evolutionary ability, of which M. sireli n. sp. would be a possible offshoot.
Sigalveolina Fleury, 2018. Three of the four species attributed to this genus were previously considered as Murciella (in Fleury, 1979). This was the consequence of a hastily interpretation of the "generic" character of the newly understood confluent structure, which was present in these species and discovered at the same time in the holotype of M. cuvillieri. Such an interpretation cannot be maintained any longer, the confluent structure now being known in various taxa. The perfect realization of this structure in the four gathered species of 2018, together with rare uncoiling in both generations give a particular homogeneous aspect to this group. The criteria are here the conjunction of the endoskeleton confluent structure at high degree of completion and an exceptional resistance to final uncoiling in both generations.
8.2 - Discussion of the criteria
Architectural
features. These
features are very varied within the family, linked to changes from
streptospiral to planispiral coiling and
great facilities given by the uncoiling capacity.
It is this character that has long prevented the distinction between
Rhapydioninids from Soritids, which are mainly characterized by the same type of
coiling-uncoiling capacity, but with a very different endoskeleton (i.e.,
comparisons in Fleury & Tronchetti,
1994; Fleury, 1996; Fig. 17 ![]() ).
).
Dimorphism is well distributed in the family, always in the initial part of tests. It is frequent in adult tests, the difference between generations being roughly related to A proloculus diameter, in various modes. B tests are frequently more uncoiled than the A ones, but to various degrees (Cuvillierinella, Murciella, Cyclopseudedomia, Chubbina and Neomurciella); some are not uncoiled (Sigalveolina and possibly Pseudochubbina). The genus Metacuvillierinella stands apart by its absence of final uncoiling in both generations. Thus, this criterion is ambiguous and can be interpreted as a "specific" feature in some cases, but also as a "generic" character, in qualitative and quantitative consideration, according to the circumstances.
The diameter of A tests proloculus has an ambiguous meaning. It is classically reputed to be a "specific" character, but this is only an appearance when the knowledge of a taxon is limited to one or few populations or even specimens. In fact, when a species, identified by other characters, is known by several populations, the result is rather indistinct: For example, 2 species, each of them known by 6 populations, C. salentina and M. decastroi show a proloculus diameter considerably variable (40 to 140 µm and 70 to 170 µm, respectively, for the extreme, see Fleury, 2016, text-fig. 12 and 2018, text-fig. 15). Conversely, some genera known by 4 species, such as Cyclopseudedomia and Sigalveolina show proportionally somewhat more limited variations (180 to 360 µm and 90 to 200 µm, respectively, for the extreme, see Fleury, 2018, text-fig. 15). Certainly, those taxa are interpretative and can be discussed in the details, but the result is nevertheless showing that this criterion is not definitive by itself and must always be confronted to the context.
Initial coiling makes the central point of many debates. The progressive change of the initial coiling from streptospiral to planispiral during evolution is generally accepted, but its significance in taxonomy is controversial and still undecided. For example, on one hand, the genera Cuvillierinella and Murciella are in part distinguished by Fleury (2016) on this criterion but, on the other hand, Cuvillierinella salentina itself and mainly C. fluctuans (in Fleury, 2016) are known to combine jointly both types. The present work is an example of the question: As it was discussed before, is the new species M. sireli to be included in the genus Metacuvillierinella or become the type of a new genus? The example of Cuvillierinella-Murciella invites us to choose the second solution, but the uncertainties about the real radiation and future of the new species carried the decision, while hoping for additional data. For such a criterion, there is no unequivocal solution: The context (to come, in this case) must drive the final decision.
Adult mode of coiling, either pseudoplanispiral or planispiral is to be considered carefully. Pseudoplanispiral coiling could be considered in theory as a passage from streptospiral to planispiral, but it is still well-represented in advanced forms with large proloculi such as the four species of Pseudochubbina, Metacuvillierinella and almost all American Cretaceous and Cenozoic genera, which seem to be well-established taxa. Planispiral coiling is nevertheless the final mode of coiling for many genera and species. Both modes can be alternatively considered as a "specific" or "generic" characters, according to other features, that is to say by considering the context.
Final uncoiling is largely widespread in every subfamily. It can be cylindrical or flabelliform, sometimes present in both modes in the two generations of some species (Cuvillierinella salentina B tests, Cyclopseudedomia mavrikasi A tests) more of less frequent and not necessarily in some taxa, (A tests of Cuvillierinella and Murciella), distinguishing the generations in some cases (Cuvillierinella, Murciella, Cyclopseudedomia and Rhapydionina). Its absence (or rarity) in B tests seems characteristic in some cases (Sigalveolina, and possibly Pseudochubbina). Its absolute absence in both generations, combined with a large umbilicus (advolute coiling), seems particular to Metacuvillierinella. The special coiling of "Pseudedomia" complanata (B tests) is to be recalled, although it represents an extraordinary mode, with no other known representative and thus, in our state of knowledge, a kind of monster.
Endoskeletal features. The endoskeleton features peculiar to the Alveolinacea separate clearly this group from the Soritacea, with which they have important architectural convergence.
Relative importance of the peripheral zone with cloisonnettes (see §3.2) and the central endoskeleton, related to other secondary characters, is considered as most significant; the distinction of the subfamily Rhapydionininae from Cuvillierinellinae being mainly based on this criterion.
Openings make a hard matter to study. They are obviously directly related to the chamberlets of the following chamber and are frequently almost of the same diameter as the chamberlets, which makes them difficult to distinguish in ordinary plane sections. A criterion based upon such a feature would require a much focalized study, which is not in current use. Anyway, their layout is subjected to evolution and the change from a particular arrangement to another one must be expected, even in a single taxon. This feature will be interpretable when it is well observed and illustrated.
Differentiation of chamberlet diameter of the various sets is apparently
characteristic of each species, as it appears especially in a relative
homogeneous genus like Pseudochubbina.
It does not seem linked to any genus (Fig. 12 ![]() ).
).
BSC-SSC
arrangement of the tubular secondary chamberlets, at first of the coaxial
type, would be a fundamental character of the Rhapydioninidae (the BSC being
apparently lacking in the genus Subalveolina
Reichel, classically attributed to Alveolinidae, see
Fig. 2.18-19 ![]() ). As we have seen previously, exceptions to this rule in true
Rhapydioninidae (absence of visible BSC), would be due to the effect of too
large and too widely spaced chamberlets of which the eventual organization
cannot be observed in plane section.
). As we have seen previously, exceptions to this rule in true
Rhapydioninidae (absence of visible BSC), would be due to the effect of too
large and too widely spaced chamberlets of which the eventual organization
cannot be observed in plane section.
Confluent structure seems to be potentially present in several subfamilies.
Its absence in some well-known and well spread genera (Pseudochubbina with possible exception of Fig.
4.7 ![]() , Chubbina, and Rhapydionina)
is nevertheless striking and still unexplained. In the genera where it is
known, it seems related to evolution, although independent from other
features. It appears only exceptionally in apparent relative simple taxa (Cuvillierinella, Murciella), but is absent in C. hellenica, nevertheless well integrated to Cyclopseudedomia. Thus there is no sharp limit for its development
and its more or less important early achievement is submitted to
interpretation in the context.
, Chubbina, and Rhapydionina)
is nevertheless striking and still unexplained. In the genera where it is
known, it seems related to evolution, although independent from other
features. It appears only exceptionally in apparent relative simple taxa (Cuvillierinella, Murciella), but is absent in C. hellenica, nevertheless well integrated to Cyclopseudedomia. Thus there is no sharp limit for its development
and its more or less important early achievement is submitted to
interpretation in the context.
Geographic, chronologic, genealogic features and radiance. Species and genera are not isolated creations; for the evolutionist, they are links in a chain of life which need to be identified. Geography and chronology are important to the understanding of these sequences, but genealogy is essential. Radiance is a new concept linked to spreading of a taxon and its nearby kinship in time and space.
Geographic arguments are uneasy to achieve. Concerning our organisms, in the Euro-Asiatic domain (Tethys and dependencies), the state of knowledge being apparently imperfect, prevents definitive conclusions by Upper Cretaceous and Lower Cenozoic times (Fleury et al., 1985; Goldbeck & Langer, 2009). The role of the Atlantic Ocean as a frontier is more obvious. There are effectively some likeness between the New and Old Worlds (i.e., Chubbina-Pseudochubbina, Raadshoovenia-Cuvillierinella, ...) as it was said before. But, as far as we know now, there is no precise homology between species and even genera from the two sides of the ocean at the same time. The genealogic arguments will be complementary and definitive in several cases of doubt and we must resign to admit that a common ancestor (a group of Miliolacea or a simple "genus" like Pseudonummoloculina Calvez, 1988) succeeded in crossing the Atlantic when it was still time.
Chronologic arguments are not always definitive by themselves too. For example, some authors, in good faith and well informed considered that Raadshoovenia (American Cenozoic) and Cuvillierinella (European Upper Cretaceous), morphologically much resembling, could be confused, which is nevertheless incompatible with their genealogies. Some other examples are less doubtful, such as the likeness of Cyclopseudedomia (European Upper Cretaceous) and Twaraina (American Eocene), resulting from an accountable convergence. Conversely, the same age of various populations reported to Murciella cuvillieri (in Fleury, 2018, text-figs. 6-7) was admitted important to give the genus its validity, although some differences in morphology of populations could be observed.
Genealogic arguments are obviously depending on the degree of knowledge and understanding of a particular group. For example, the question posed by the almost perfect resemblance between Raadshoovenia and Cuvillierinella, partly clarified by the chronologic argument, is definitively solved by knowledge of both genealogies and kinships (Fleury, 2016, text-figs. 4 and 13). The same could be said about Sellialveolina-Pseudedomia, although the resemblance is much smaller. Nevertheless, lack of detailed information still makes difficult the solution of pending questions about origin of some taxa, such as Pseudochubbina, to mention the toughest.
Radiance. This is a rather immaterial and possibly ephemeral argument, depending largely on the momentary knowledge. It is linked to the known importance of spreading of a taxon, its related species and/or genera. It was used above to try and justify the distinction between Murciella from Cuvillierinella, the type species of both being known by several populations from various localities (Fleury, 2018), either associated or dissociated. Other examples of large radiance are given by the genera Sellialveolina, Pseudochubbina, Cyclopseudedomia and Sigalveolina, known by several populations and/or species from large areas. On the contrary, the new taxon M. sireli is considered as a species of Metacuvillierinella, on account of its currently unknown radiance; discovery of several new populations or resembling species in a number of outcrops in a large area would lead us to reconsider this attribution.
8.3 - Conclusive remarks on the family structure
The Rhapydioninidae is remarkable for its diversity, although its stable endoskeleton and particularly the BSC (despite some particular variations) assure the identity. This diversity and the possible relations of the various branches to a common stem are expressed by a diagrammatic model by Fleury & Fourcade (1990, text-fig. 4). The "genus" Pseudonummoloculina (rather an evolutionary stage common to various branches; see Fleury, 2018: Abstract) could give an image of the Miliolacea at the origin of several groups.
The family was widespread during three periods, from Albian to Eocene.
The first period, relatively short (Albian-Cenomanian) and apparently restrained to the western part of the European Tethys is poorly diversified at generic level (Sellialveolina) but comprises a number of largely spread varieties (Sellialveolininae).
The second period (Campanian-Maastrichtian) is the richest and the most diversified, with about 10 genera including 25 to 30 species in Euro-Asiatic Tethysian and Central America realms (Rhapydionininae, Cuvillierinellinae and Chubbininae, the Pseudedomiinae being doubtful for taxonomic reasons, as seen before).
The third period (Lower Cenozoic), American, probably still to be discovered in part, is only known by 4 or 5 presumably monospecific genera (Neomurciellinae).
As we have seen previously, there are no unequivocal features distinguishing species and genera. They are all discriminated by a number of associated characters, independent from one to the other, having their own evolutionary rate and fundamentally not hierarchized. Any taxon is a sort of chimera, or puzzle, in which features are to be selected as making a special set, different from all others (the species), but more or less overall resembling other species sharing a main proper evolutionary tendency (the genera). In each case, one or several features appear predominant, with eventual variations and exceptions due to their fundamental independency, requiring pragmatic interpretation in the light of the context. In short, the evolutionary perspective is the only guide, in its qualitative (various features) and quantitative aspects (degrees in feature fulfillment, radiance of the taxon), which cannot be provided by more or less theoretical procedures founded on approximate knowledge and a priori rigid choices.
Subfamilies are made up of genera supposed as issued from a common ancestor. Age and geography are the best arguments to distinguish two of them (Sellialveolininae and Neomurciellinae). Geographic difference, together with absence of obvious detailed analogies, distinguish the Chubbininae from the Euro-Asiatic subfamilies of same age. Among these, the most successful is the Cuvillierinellinae, from which the Rhapydionininae are distinguished by an unusually stable character of the endoskeleton (relative importance of the peripheral part of chambers). As it was previously explained, the Pseudedomiinae is depending on future discoveries about the type genus.
The Western Mediterranean region seems to be conveniently known at present, but more oriental areas are probably worthy of great interest, as the new taxon from Turkey seems to show. New species, new genera and may be new subfamilies could have proliferated in those regions, if the particular environments of internal restricted platform which our organisms are fond of were sufficiently developed.
Finally, it must be emphasized that knowledge of the general context is necessarily to understand any level in systematics. The various features being independent, no dichotomist key is able to give a convenient image of life complexity. But the proposed demarche is much demanding and cannot be achieved by superficial studies: Any taxon must be completely and properly described, as far as possible through comparisons of several populations in various contexts, with analyzed variability and, at least, some statistics on simple features. Much time and work are necessary, but this is the price to pay for trying to understand a family which deserves much attention, having the privilege to offer so many variations inside such a compact cluster.
This paper is a tribute in honour to Professor Jean Dercourt, who died during the writing of this paper. Although not a micropaleontologist, he was an active supporter of a number of various specialists. In particular, when he was teaching in Lille, he was a generous and open-minded thesis director of one of the present authors.
We thank to TPAO for its support and permission to publish this paper. Thanks to colleague F. Mert Türesin for supplying study materials, and to technician Ömer Çapık for preparing thin sections. We are indebted to Danielle Decrouez and an anonymous reviewer for attentive readings and constructive remarks on the manuscript. Above all, it must be recalled that the periadritic material published here and elsewhere by J.-J. Fleury was collected thanks to support of the French CNRS and the former "Université des Sciences et Techniques de Lille (Lille1)", now "Université de Lille - Faculté des Sciences et Technologies". Special thanks are due to Jessie Cuvelier, from the "UMR 8198 Evo-Eco-Paleo" of the same university, who kindly made part of the pictures of this paper. Last but not least, the authors wish to express their gratitude to Bruno Granier who has greatly helped to get through this work.
Anderson O.R. & Lee J.J. (1991).- Cytology and fine structure. In: Anderson O.R. & Lee J.J. (eds.), Biology of Foraminifera.- Academic Press, Cambridge, MA, p. 7-37.
Bernier P. & Fleury J.-J. (1980).- La plate-forme carbonatée de Gavrovo-Tripolitza (Grčce). Évolution des conditions de sédimentation au cours du Mésozoďque.- Géologie méditerranéenne, Marseille, t. VII, no. 3, p. 247-259.
Bignot G. & Poisson A. (1974).- Le Cénomanien du flanc oriental du Katran Dağ (=Sam Dağ) prčs d'Antalya (Turquie).- Bulletin of the Mineral Research and Exploration Institute of Turkey, Ankara, vol. 82, p. 71-77.
Calvez H. (1988).- Pseudonummoloculina aurigerica n. gen., n. sp. et Dobrogelina ? angulata n. sp. Deux foraminifčres nouveaux de l'Albien calcaire des Pyrénées franco-espagnoles.- Revue de Paléobiologie, Genčve, vol. spécial 2 (Benthos '86), p. 391-399.
Çelikdemir E. & Dülger S. (1990).- Güneydoğu Anadolu'da Mardin Grubu karbonatlarının stratigrafisi, sedimentolojisi ve rezervuar özellikleri.- TPAO Arama Raporu, Ankara, 2665, p. 1-93.
Chiocchini M., Farinacci A., Mancinelli A., Molinari V. & Potetti M. (1994).- Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche mesozoiche dell' Appenninino centrale (Italia).- Studi Geologici Camerti, vol. sp. "Biostratigrafia dell'Italie centrale", p. 9-129.
Colalongo M.L. (1963).- Sellialveolina viallii n. gen., n. sp. di Alveolinid cenomaniano dell'Appenninino meridional.- Giornale di Geologia (Serie 2a), Bologna, vol. XXX (1962), p. 361-371.
De Castro P. (1966).- Contributo alla conoscenza delle alveoline albiano-cenomaniane della Campania.- Bollettino della Societŕ dei Naturalisti in Napoli, vol. 75, p. 219-275.
De Castro P. (1971).- Osservazioni su Raadshoovenia Van Den Bold e i suoi raporti col nuovo genere Scandonea (Foraminiferida, Miliolacea).- Bollettino della Societŕ dei Naturalisti in Napoli, vol. 80, p. 161-236.
De Castro P. (1972).- Osservazioni sui generi Rhapydionina Stache e Rhipidionina Stache (Foraminiferida).- "Atti" Accademia Pontaniana, Napoli, n.s., vol. 21 (1971-1972), p. 1-4.
De Castro P. (1987).- On some foraminifera and algae in Apennine Upper Cretaceous and Paleocene.- Memorie della Societŕ Geologica Italiana, Roma, vol. 40, p. 109-124
De Castro P. (1988).- Les Alvéolines du Crétacé d'Italie.- Revue de Paléobiologie, Genčve, vol. spécial 2 (Benthos '86), p. 401-416.
De Castro P. (1990).- Osservazioni paleontologiche sul cretacico della localitŕ-tipi di Raadshoovenia salentina e su Pseudochubbina n. gen.- Quaderni dell'Accademia pontaniana, Napoli, 10, p. 1-116.
De Castro P., Drobne K. & Gušić I. (1994).- Fleuryana adriatica n. gen., n. sp. (Foraminiferida) from the upermost Maastrichtian of the Brač Island (Croatia) and some other localities on the Adriatic Carbonate Platform.- Razprave IV. razreda SAZU, Ljubljana, vol. 35, no. 8, p. 129-149.
Deloffre R. & Hamaoui M. (1979).- Découverte de Pseudedomia (Foraminifčre) en Aquitaine.- Bulletin du Centre de Recherche Exploration-Production elf-Aquitaine, Pau, vol. 3, no. 1, p. 37-61.
Deloffre R., Fleury J.-J. & Mavrikas G. (1991).- Les algues calcaires du Maastrichtien-Paléocčne de la plate-forme de Gavrovo-Tripolitza (Grčce) : Une floraison liée ŕ un diastrophisme.- Geobios, Villeurbanne, vol. 24, no. 5, p. 515-536.
Dilley F.C. (1973).- Cretaceous Larger Foraminifera. In: Hallam A. (ed.), Atlas of Paleobiogeography.- Elsevier, Amsterdam, p. 403-419.
Drobne K. (1981).- Characteristical foraminifers and their associations in the base of the Danian beds.- Symposium on problems of Danian in Yougoslavia.- Zbornik referatov = proceedings, Ljubljana, p. 85-403.
Eames F.E. & Smout F.G.S. (1955).- LXII. Complanate Alveolinids and Associated Foraminifera from the Upper Cretaceous of the Middle East.- The Annals and Magazine of Natural History (Series 12), London, vol. 8, p. 505-512.
Emberger J., Magné J., Reyre D. & Sigal J. (1955).- Note préliminaire sur quelques Foraminifčres nouveaux ou peu connus dans le Crétacé supérieur de facičs sub-récifal d'Algérie.- Comptes Rendus Sommaires de la Société géologique de France, Paris, no. 5-6, p. 110.
Fleury J.-J. (1970).- Le Sénonien et l'Éocčne ŕ microorganismes benthoniques du Klokova (Zone du Gavrovo, Akarnanie, Grčce continentale).- Revue de Micropaléontologie, Paris, vol. 13, no. 1, p. 30-44.
Fleury J.-J. (1971).- Le Cénomanien ŕ foraminifčres benthoniques du massif du Varassova (Zone du Gavrovo, Akarnanie, Grčce continentale).- Revue de Micropaléontologie, Paris, vol. 14, no. 3, p. 181-194.
Fleury J.-J. (1974).- Contribution ŕ la connaissance des Rhapydionininae (Foraminifčres, Alveolinidae) crétacés.- Geobios, Villeurbanne, vol. 7, no. 4, p. 307-322.
Fleury J.-J. (1977).- Deux Rhapydionininae (Foraminifčres, Alveolinidae) d'affinités américaines dans le Crétacé supérieur de Grčce (Zone de Gavrovo-Tripolitza).- Revue de Micropaléontologie, Paris, vol. 20, no. 2, p. 77-90.
Fleury J.-J. (1979).- Le genre Murciella (Foraminifčre, Rhapydionininae), dans le Crétacé supérieur de Grčce (zone de Gavrovo-Tripolitza).- Geobios, Villeurbanne, vol. 12, no. 2, p. 149-185.
Fleury J.-J. (1980).- Les zones de Gavrovo-Tripolitza et du Pinde-Olonos (Grčce continentale et Péloponnčse du Nord). Evolution d'une plate-forme et d'un bassin dans leur cadre alpin.- Société géologique du Nord, Lille, Publication 4, 651 p.
Fleury J.-J. (1984).- Senalveolina aubouini, n. gen., n. sp., Alveolinidae nouveau du Sénonien de Grčce.- Revue de Micropaléontologie, Paris, vol. 27, no. 3, p. 171-188.
Fleury J.-J. (1996).- Convergence morphologique Alveolinacea - Soritacea : Les genres Rhapydionina Stache et Rhabdorites n. gen.- Revue de Micropaléontologie, Paris, vol. 39, no. 1, p. 41-51.
Fleury J.-J. (2014).- Données nouvelles sur Rhapydionina Stache, 1913 et Fanrhapydionina n. gen., un groupe de Rhapydioninidae (Alveolinacea, Foraminifera) foisonnant en région périadriatique au Campanien-Maastrichtien.- Geodiversitas, Paris, vol. 36, no. 2, p. 173-208.
Fleury J.-J. (2016).- Cuvillierinella salentina (Foraminifera, Rhapydioninidae) and its kinship in the Western Mediterranean during the Campanian-Maastrichtian.- Revue de Micropaléontologie, Paris, vol. 59, no. 2, p. 200-224.
Fleury J.-J. (2018).- Rhapydioninidae du Campanien-Maastrichtien en région méditerranéenne : Les genres Murciella, Sigalveolina n. gen. et Cyclopseudedomia.- Carnets Geol., Madrid, vol. 18, no. 11, p. 233-280.
Fleury J.-J., Bignot G., Blondeau A. & Poignant A. (1985).- Biogéographie de Foraminifčres téthysiens du Sénonien ŕ l'Eocčne supérieur.- Bulletin de la Société géologique de France, Paris, (8e Série), t. I, no. 5, p. 757-770.
Fleury J.-J. & Fourcade É. (1987).- Un nouvel Alveolinidae du Paléocčne-Éocčne inférieur du Mexique. Ses affinités avec les genres Murciella et Yaberinella.- Revue de Micropaléontologie, Paris, vol. 30, no. 2, p. 91-110.
Fleury J.-J. & Fourcade É. (1990).- La super-famille Alveolinacea (foraminifčres) : Systématique et essai d'interprétation phylogénétique.- Revue de Micropaléontologie, Paris, vol. 33, no. 3-4, p. 241-268.
Fleury J.-J. & Tronchetti G. (1994).- Palandrosina taxyae n. gen., n. sp., nouveau foraminifčre (Archaiasidae) du Santonien Provençal.- Revue de Micropaléontologie, Paris, vol. 37, no. 1, p. 31-47.
Fourcade É. (1966).- Murciella cuvillieri n. gen., n. sp., nouveau foraminifčre du sud-est de l'Espagne.- Revue de Micropaléontologie, Paris, vol. 9, no. 3, p. 147-155.
Fourcade É. & Fleury J.-J. (2001).- Origine, évolution et systématique de Praechubbina n. gen., foraminifčre Alveolinacea de Crétacé supérieur du Guatemala et du Mexique.- Revue de Micropaléontologie, Paris, vol. 44, no. 2, p. 125-157.
Goldbeck E.J. & Langer M.R. (2009).- Biogeographic provinces and patterns of diversity in selected Upper Cretaceous (Santonian-Maastrichtian) larger foraminifera. In: Demchuk T.D. & Gary A.C. (eds.), Geologic problem solving with microfossils: A volume in honor of Garry D. Jones.- SEPM Special Publication no. 93, p. 187-232.
Görmüs M. (1999).- Pseudedomia hekimhanensis n. sp. from the late Campanian (?) to Maastrichtian of Hekimhan, NW Malatya, Turkey.- Journal of Foraminiferal Research, Lawrence, vol. 29, no. 3, p. 236-242.
Güven A., Dinçer A., Tuna E.M. & Çoruh T. (1991).- Stratigraphic evolution of the Campanian-Paleocene autochthonous succession of the southeast Anatolia.- Ozan Sungurlu Symposium Proceedings, Ankara, p. 238-261.
Hamaoui M. & Fourcade É. (1973).- Révision des Rhapydionininae (Alveolinidae, Foraminifčres).- Bulletin du Centre de Recherche SNPA, Pau, vol. 7, no. 2, p. 361-435.
Haynes J.R. (1981).- Foraminifera.- Macmillan Publishers, London, 433 p.
Heba G., Prichonnet G. & El Albani A. (2009).- Meteoric diagenesis of Upper Cretaceous and Paleocene-Eocene shallow-water carbonates in the Kruja platform (Albania): Geochemical evidence.- Geologica carpathica, Bratislava, vol. 60, no. 2, p. 165-179.
Henson F.R.S. (1948).- Larger imperforate Foraminifera of south-western Asia.- British Museum of Natural History, London, 127 p.
Holzmann M., Hohenegger J., Hallock P., Piller W.E. & Pawlowski J. (2001).- Molecular phylogeny of large miliolid foraminifera (Soritacea Ehrenberg 1839).- Marine Micropaleontology, vol. 43, no. 1-2, p. 47-74.
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., vol. 6, no. M02 (CG2006_M02), p. 1-126.
Hottinger L., Drobne K. & Caus E. (1989).- Late Cretaceous, large, complex miliolids (Foraminifera) endemic in the Pyrenean faunal province.- Facies, Erlangen, vol. 21, no. 1, p. 99-134.
Kellner S.K, Knappertsbusch M. K., Coster L., Müller B. & Schlulz G. (2019).- Imaging the internal structure of Borelis schlumbergeri Reichel (1937): Advances in high-resolution hard X-ray microtomography.- Palaeontologia Electronica, article 22.1.17, p. 1-19.
Ketin İ. (1966).- Tectonic units of Anatolia.- Bulletin of the Mineral Research and Exploration Institute, Ankara, vol. 66, p. 23-34.
Landrein P., Loreau J.-P. & Fleury J.-J. (2001).- Émersion généralisée intra-maastrichtienne de la plate-forme de Gavrovo-Tripolitza (Grčce), effets sur les populations de foraminifčres Rhapydionininae.- Bulletin de la Société géologique de France, Paris, vol. 172, no. 1, p. 85-98.
Lee J.J., Faber W.W., Anderson O.R. & Pavlowski J. (1991).- Life-cycles of Foraminifera. In: Lee J.J. & Anderson O.R. (eds.), Biology of foraminifera.- Academic press, London, p. 285-328.
Loeblich S. & Tappan H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New-York, 2031 p.
Luger P. (2018).- Mikropaläontologie (Foraminiferen, Ostrakoden), Biostratigraphie und fazielle Entwicklung des Kreide von Nordsomalia mit einem Beitrag zur geodynamischen Entwicklung des östlichen Gondwana im Mesozoikum und frühen Känozoikum.- Documenta Naturae Abhandlungen, Aham, Band 1, Teil 2 (Tafelband), 49 Pls.
Mavrikas G., Fleury J.-J. & Fourcade É. (1994).- Implications paléobiogéographiques de la présence de Pseudedomia (Foraminifčre) dans le Maastrichtien méditerranéen.- Comptes Rendus de l'Académie des Sciences (Série II), Paris, vol. 318, p. 849-855.
Munier-Chalmas E. & Schlumberger C. (1885).- Note sur les Miliolidés trématophorées.- Bulletin de la Société géologique de France, Paris, (3e Série), t. XIII, p. 273-323.
Okay A.I. & Tüysüz O. (1999).- Tethyan sutures of northern Turkey.- Geological Society of London, Special Publications, vol. 156, p. 475-515.
Özkan R. & Altiner D. (2019).- The Cretaceous Mardin Group carbonates in southeast Turkey: Lithostratigraphy, foraminiferal biostratigraphy, microfacies and sequence stratigraphic evolution.- Cretaceous Research, vol. 98, p. 153-178.
Papetti L. & Tedeschi D. (1965).- Nuovo genere di foraminifero del Santoniano superior.- Geologica Romana, vol. 4, p. 119-128.
Pęcheux J.-F. (1984).- Le Sénonien supérieur-Tertiaire du Chiapas (SE Mexique) et ses macroforaminifčres.- Thčse 3e Cycle, Université de Nice, 154 p.
Pęcheux J.-F. (2002).- Des Miliolacea aux Alveolinacea (grands Foraminifčres) : La faune du Crétacé supérieur et du Tertiaire inférieur du Chiapas (S.E. Mexique).- Geobios, Villeurbanne, vol. 35, p. 469-488.
Perconig E. & Martin L. (1971).- Estratigraphia del Cretacico de la zona de Ontur Y Yecla (Zona Prebetica).- Congreso Hispano-Luso-Americano de Geología Económica, Madrid-Lisboa, p. 331-341.
Perinçek D. (1980).- Arabistan kıtası kuzeyindeki tektonik evrimin, kıta üzerine çökelen iftifteki etkileri.- Türkiye 5. Petrol Kongresi Bildirileri, Ankara, p. 77-93.
Perinçek D., Duran O., Bozdoğan N. & Çoruh T. (1991).- Stratigraphy and paleogeographical evolution of the autochthonous sedimentary rocks in the southeast Turkey.- Ozan Sungurlu Symposium Proceedings, Ankara, p. 274-305.
Pomoni-Papaioannou P.A. & Zambetakis-Lekkas A. (2009).- Facies associations of the late Cenomanian carbonate platform of Tripolitza subzone (Vitina, Central Peloponnesus, Greece): Evidence of long-term/terrestrial subaerial exposure.- Italian Journal of Geosciences, Roma, vol. 128, no. 1, p. 123-130.
Rahaghi A. (1976).- Contribution ŕ l'étude de quelques grands foraminifčres de l'Iran.- Société National Iranienne des Pétroles, Tehran, no. 6, p. 1-23.
Reichel M. (1936).- Étude sur les Alvéolines.- Mémoires de la Société paléontologique suisse, Basel, vol. 57, no. 4, p. 1-93.
Reichel M. (1937).- Étude sur les Alvéolines.- Mémoires de la Société paléontologique suisse, Basel, vol. 59, no. 3, p. 95-147.
Reichel M. (1964).- Alveolinidae. In: Loeblich A.R. & Tappan H. (eds.), Sarcodina chiefly "Thecamoebians" and Foraminifera. In: Moore R.C. (ed.), Treatise in Invertebrate Paleontology, Part C, Protista 2.- The Geological Society of America, Tulsa, and The University of Kansas Press, Lawrence, p. C503-510.
Reichel M. (1984).- Le crible apertural de Rhapydionina liburnica Stache du Maastrichtien de Vremski-Britof, Yougoslavie.- Benthos 83, 2nd International Symposium on Benthic Foraminifera (Pau, avril 1983) elf Aquitaine, Esso REP, Total CFP, p. 525-532.
Reiss Z., Hamaoui M. & Ecker A. (1964).- Pseudedomia from Israel.- Micropaleontology, vol. 10, no. 4, p. 431-437.
Robinson E. (1968).- Chubbina, a new Cretaceous Alveolinid genus from Jamaica and Mexico.- Palaeontology, London, vol. 11, no. 4, p. 526-534.
Robinson E. (1993).- Some imperforate Larger Foraminifera from the Paleogene of Jamaica and the Nicaragua Rise.- Journal of Foraminiferal Research, Lawrence, vol. 23, no. 1, p. 47-65.
Saint-Marc P. (1974).- Étude stratigraphique et micropaléontologique de l'Albien, du Cénomanien et du Turonien du Liban.- Notes et Mémoires sur le Moyen-Orient, Paris, vol. 13, 342 p.
Schlagintweit F. & Rashidi K. (2016).- Some new and poorly known benthic foraminifera from Late Maastrichtian shallow-water carbonates of the Zagros zone, SW Iran.- Acta palaeontologica romaniae, vol. 12, no. 1, p. 53-70.
Sengör C. & Yilmaz Y. (1981).- Tethyan evolution of Turkey: A plate tectonic approach.- Tectonophysics, 75, p. 181-241.
Smout A.H. (1963).- The genus Pseudedomia and its phyletic relationships, with remarks on Orbitolites and other complex foraminifera. In: Konigswald E.V. (ed.), Evolutionary trends in Foraminifera.- Elsevier, Amsterdam, p. 224-281.
Solak C., Tasli K. & Koç H. (2017).- Biostratigraphy and facies analysis of the Upper Cretaceous-Danian ? platform carbonate succession in the Kuyucak area, western Central Taurides, S. Turkey.- Cretaceous Research, vol. 79, p. 43-63.
Solak C., Tasli K., Özer S. & Koç H. (2019).- The Madenli (Central Taurides) Upper Cretaceous platform carbonate succession: Benthic foraminiferal biostratigraphy and platform evolution.- Geobios, Villeurbanne, vol. 52, p. 67-83.
Stache G. (1889).- Die Liburnische Stufe und deren Grenzhorizonte.- Abhandlungen des Geologische Reichsanstalt, Wien, Band 13, 170 p.
Stache G. (1913).- Uber Rhipidionina St. und Rhapydionina St.- Jahrbuch des Geologischen Reichanstalt (1912), Wien, Band 62, p. 659-680.
Sungurlu O. (1974).- VI. Bölge kuzeyinin jeolojisi ve petrol imkanları.- Türkiye 2. Petrol Kongresi Bildirileri, Ankara, p. 85-107.
Tuna D. (1974).- VI. Bölge litostratigrafi birimleri adlamasının açıklayıcı raporu.- Türkiye 2. Petrol Kongresi Bildirileri, Ankara, p. 183-192.
Van den Bold W.A. (1946).- Contribution to the study of Ostracoda with special reference to the Tertiary and Cretaceous microfauna of the Carribean region.- Thčse, Rijks Universiteit Utrecht, De Bussy, Amsterdam, 167 p.
Vicedo V., Aguilar-Lekkas M., Caus E. & Hottinger L. (2009).- Fusiform and laterally compressed alveolinaceans (Foraminiferida) from both side of the Late Cretaceous Atlantic.- Neues Jarbuch für geologie und Paleontologie, Frankfurt am Main, vol. 253, no. 2-3, p. 229-247.
Vicedo V., Calonge A. & Caus E. (2011).- Cenomanian Rhapydioninids (Foraminiferida): Architecture of the shell and stratigraphy.- Journal of Foraminiferal Research, Lawrence, vol. 41, no. 1, p. 41-52.
Vicedo V., Caus E. & Frijia G. (2013).- Late Cretaceous alveolinaceans (larger foraminifera) of the Caribbean palaeobioprovince and their stratigraphic distribution.- Journal of Systematic Paleontology, vol. 11, no. 1, p. 1-25.
Wagner C.W. (1964).- Manual of larger Foraminifera.- Bataafse Petroleum Maatschappij, La Haye, 307 p.
Yilmaz Y. (1993).- New evidence and model on the evolution of the southeast Anatolian orogen.- Geological Society American Bulletin, vol. 105, no. 2, p. 251-271.
Yilmaz E. & Duran O. (1997).- Güneydoğu Bölgesi Otokton ve Allokton Birimler Stratigrafisi Adlama Sözlüğü "Lexicon".-TPAO Araştırma Merkezi Eğitim Yayınları, Ankara, vol. 31, 460 p.
The setting up of this lexicon, resulting from a long acquaintance with the group, was suggested by a reviewer. It collects together the terms used in the present paper and several others previously produced by one of the authors. Various other terms, judged as not convenient for the studied group but significant from a more general or historic point of view, are also presented.
The Rhapydioninidae constitute a family well distinguished from all others. They nevertheless share with the Alveolinidae a fundamental organization consisting of tubular divisions of chambers (the chamberlets) and an almost empty space beneath the septum (the preseptal space). These characters were well recognized and named by Reichel (1936-1937) in the Alveolinidae (usually fusiform), later and progressively in the Rhapydioninidae (tending to be axially compressed, eventually uncoiled) by various authors after the enlightening observations of De Castro (1972) on the genus Rhapydionina. These characters have no equivalent in other groups, even in the apparently closest, such as the Soritids. They are thought to be the testimony of a special "way of life", that is to say a particular cellular physiology or behavior (well underlined by DNA studies of recent species of Alveolinidae and Soritids: Holzmann et al., 2001), deserving a particular terminology of the tests morphologic elements.
The main goal of the present attempt is to avoid the confusions resulting from too large a propensity for unification and synthesis. Thus, the present lexicon, dodging through many traps, must be understood as limited to the nomenclature appropriate to the Rhapydioninidae, with no pretention to rule the huge whole domain of foraminifera.
The items effectively used in Rhapydioninidae are presented in bold; others, useful to explain some particularities or evolution of knowledge are in light; suggested deferment to other words in the lexicon are in italics.
Advolute (coiling). An ordinary intermediate stage between the involute and evolute coiling modes, characterized by a large umbilicus showing lateral parts of the previous coils. Almost all Rhapydioninidae tests include this stage, before or preparing the uncoiling one. A rather restrictive meaning is here given to the word, in order to underline the particularity of the genus Metacuvillierinella, of which the two species, in both generations, keep advolute, never reaching the evolute stage. See Fig. 2.2 ![]() and axial sections in Fig. 3
and axial sections in Fig. 3 ![]() .
.
Aperture. Despite some confusions, several authors converge in calling aperture "in chambered shells the ultimate opening of the last chamber cavity into the ambient environment (...) when a new chamber is added, (...) the aperture is transformed into a means of communication between successive chamber lumina and thus its function changes. The transformed aperture is called a (intercameral) foramen" (Hottinger, 2006). Nevertheless, some observations, such as "the distribution of organelles, within intra- and extrashell is continuous through the aperture(s)" (Anderson & Lee, 1991) show that the function change is probably not as fundamental as assumed. Moreover, in our group, although the true apertures and the foramina (not morphologically different), are ordinary observed simultaneously in the same equatorial section, the simplest way to name them together is to use the general term of openings (like in the above cited sentence of Hottinger).
Archaiasiform or hyperinvolute (coiling). Test completely involute, with last whorls made up of strongly recurved chambers reaching the poles after describing several volutions on the flanks. See Eames & Smout, 1955, text-fig. 1; Hottinger, 2006, text-fig. 7.B giving a spatial illustration of one isolated chamber; Fig. 8.5-6 ![]() and 8.8
and 8.8 ![]() .
.
Axial section. Centered or subcentered (almost centered) section perpendicular to the plane (or distorted surface) of coiling, including the coiling axis, or average position of coiling axis in case of pseudoplanispiral coiling.
Axis (of coiling). Theoretical axis of coiling rotation. When the axis remains in the same position for successive coils, the coiling is planispiral, the axial section of the test is bilaterally symmetrical, the equatorial plane being sagittal. When unstable, describing a conical surface, (see pseudoplanispiral coiling) the "axial" section of the test appears sigmoid (S-shaped).
Basal layer. See "couche basale". The main text presents a tentative clarification of this progressively complicated notion. Whatever the interpretation given recently by Hottinger (2006, text-figs. 18.H and 22.E) of the polar thickening in some Alveolina and Borelis, the discovery of the BSC in most of the Rhapydioninidae denies in every case an eventual homology between the basal layer s.s. as traditionally understood and the BSC-SSC structure of the central endoskeleton. Nevertheless, a true thin basal layer s.s. is present in the Rhapydioninidae (see Fig. 2.7 ![]() : Cb).
: Cb).
Basal Secondary Chamberlets (BSC). Part of the central endoskeleton. In fact, a layer of more or less distinctive chamberlets, sometimes differentiated from all others by their diameter, directly in contact with the wall of the previous coil (but separated from it by the ordinary thin basal layer s.s.). Vanished in evolute and pseudoevolute chambers. It would properly be called "basal layer" (of secondary chamberlets) if the name was not beforehand occupied. See interpreted sections of Figs. 2.10-14 ![]() and 13
and 13 ![]() .
.
BSC-SSC structure. Basal Secondary
Chamberlets-Scattered Secondary Chamberlets structure is named after the ontogenetic apparition order of chamberlets in the central endoskeleton. See Fig. 2.14 ![]() .
.
Central endoskeleton. Part of the endoskeleton situated beneath the primary chamberlets layer, comprising the secondary chamberlets: 1) basal ones (BSC) forming a layer parallel to the wall of preceding coil, absent in chambers of evolute or pseudoevolute parts of tests; 2) central ones with fishnet appearance (when distance between chamberlets is smaller than their diameter) or scattered (SSC) in a compact mass (when distance between chamberlets is larger than their diameter).
Central thickening ("Épaississement central"). Proposed by Hamaoui & Fourcade (1973) for the organization of the endoskeleton beneath the layer of primary chamberlets (called "espaces interlamellaires"), admitted to be equivalent to the "basal layer" of the Alveolinidae ("épaississement central equivalent ŕ la "couche basale", p. 372). In fact, the authors rightly deny the presence of floors beneath the layer of primary chamberlets but ignore the real original character of this structure, thus giving an advantage to the central endoskeleton, preferred here.
Chamber. In general case, the space (lumen) limited by the previous coil, the outer wall and two successive septa (wall and septa included). See Fig. 2 ![]() .
.
Chamberlets. Subdivisions of chambers, tubular in most Rhapydioninidae, parallel to direction of coiling (see coaxial structure), communicating only in distal part of chambers, the preseptal space (see primary and secondary chamberlets, Figs. 7.7 ![]() and 10.11
and 10.11 ![]() ). An exception is the confluent structure, a particularity of some species or genera: The secondary chamberlets (the basal ones excluded) taking various orientations, communicate laterally at their crossing in the whole chamber. See Fig. 16
). An exception is the confluent structure, a particularity of some species or genera: The secondary chamberlets (the basal ones excluded) taking various orientations, communicate laterally at their crossing in the whole chamber. See Fig. 16 ![]() ).
).
Cloisonnettes. Name chosen here for the dividing interseptal wall isolating primary chamberlets in the peripheral zone of chambers (see Figs. 7.6-7 ![]() and 10.11
and 10.11 ![]() ). It was created by Reichel (1936-1937) for the Alveolinidae and is well adapted to the Rhapydioninidae sharing the same type of single layer of (primary) chamberlets. Unfortunately, the same author himself (1964), using the term septula, introduced a confusion, because of the rather inaccuracy meaning of this term. Thus the Reichel's French term seems irreplaceable, particularly taking into account that the fusing of their distal parts is likely to give birth to the central endoskeleton. See Fleury, 2018, text-fig. 2.19-24, in particular; Fig. 17
). It was created by Reichel (1936-1937) for the Alveolinidae and is well adapted to the Rhapydioninidae sharing the same type of single layer of (primary) chamberlets. Unfortunately, the same author himself (1964), using the term septula, introduced a confusion, because of the rather inaccuracy meaning of this term. Thus the Reichel's French term seems irreplaceable, particularly taking into account that the fusing of their distal parts is likely to give birth to the central endoskeleton. See Fleury, 2018, text-fig. 2.19-24, in particular; Fig. 17 ![]() .
.
Coaxial structure (of the Rhapydioninidae). The ordinary endoskeletal organization of Rhapydioninidae: All sorts of chamberlets are parallel, oriented in the spiral direction. The term is used in order to distinguish this primitive structure from the confluent structure, where secondary chamberlets are obliquely oriented and interpenetrated with their neighbors.
Coiling. Planispiral coiling is generalized among adult Rhapydioninidae. The nepionic stage is nevertheless usually streptospiral or miliolid, at least in B tests and sometimes in A tests of the simplest ("primitive") species. Pseudoplanispiral coiling is a variant betrayed by sigmoid axial sections
Columella. Term used by Hottinger (2006): "The solid, trochospiral structure formed by the basal walls of spiral chambers coalescing around the coiling axis, as in many gastropod shell, or symmetrically poleward in planispiral-fusiform shells". The examples given by this author: Borelis schlumbergeri (Reichel) and Alveolina tenuis Hott. seem to show that it mainly corresponds to the "basal layer" developed in axial region of these elongate Alveolinidae with no or few
scattered secondary chamberlets. Another use of this word is given by Vicedo et al. (2009), mentioning Subalveolina dordonica Reichel without precise structural details; in the case of this taxon, Fig. 2.19 ![]() shows that the numerous
scattered secondary chamberlets clearly merge in the preseptal space. Thus, the term has no other particular meaning than a morphological comparison with gastropod shell.
shows that the numerous
scattered secondary chamberlets clearly merge in the preseptal space. Thus, the term has no other particular meaning than a morphological comparison with gastropod shell.
Confluent structure (previously called helicoďdal) Fig. 16 ![]() . Structure resulting from interpenetration of neighboring variously oriented secondary chamberlets, at their crossing point. The new connections are thus in competition with the preseptal space for the mixing of protoplasm. It is supposed as favoring the free moving of endosymbionts searching for the optimal illumination. Known in many taxa, from Albian-Cenomanian to Eocene, on both sides of the Atlantic. See Fleury, 2018, text-fig. 3; Figs. 6.2, 6.6, 6.9, 6.12
. Structure resulting from interpenetration of neighboring variously oriented secondary chamberlets, at their crossing point. The new connections are thus in competition with the preseptal space for the mixing of protoplasm. It is supposed as favoring the free moving of endosymbionts searching for the optimal illumination. Known in many taxa, from Albian-Cenomanian to Eocene, on both sides of the Atlantic. See Fleury, 2018, text-fig. 3; Figs. 6.2, 6.6, 6.9, 6.12 ![]() , 7.6-8
, 7.6-8 ![]() , 8.27
, 8.27 ![]() , 9.25
, 9.25 ![]() and 11.13
and 11.13 ![]() .
.
Couche basale (basal layer). According to Reichel (1936-1937) this element is defined as "the deposit laid out on the wall of the preceding coil, forming the internal coating of the chamber". In that sense, there is effectively a couche basale in the Rhapydioninidae, but ordinary so thin that it is not mentioned (Fig. 2.7 ![]() ). The term was afterwards used as such by several English writing authors, but was translated by Reichel himself (1964) in "basal layer or basal thickening, mainly to characterize the enormous basal thickening in several internal whorls" (with reference to "Flosculina", a synonym of Alveolina, see Smout, 1963, p. 224-225; Reichel, 1964, p. 506-509); see Fig. 2.8-9
). The term was afterwards used as such by several English writing authors, but was translated by Reichel himself (1964) in "basal layer or basal thickening, mainly to characterize the enormous basal thickening in several internal whorls" (with reference to "Flosculina", a synonym of Alveolina, see Smout, 1963, p. 224-225; Reichel, 1964, p. 506-509); see Fig. 2.8-9 ![]() . In that case, the main text of this paper quotes 4 reasons to distinguish this basal layer from the central endoskeleton.
. In that case, the main text of this paper quotes 4 reasons to distinguish this basal layer from the central endoskeleton.
Nevertheless, a shade is introduced by Hottinger (2006, p. 8 and text-fig. 18.H), concerning fusiform Alveolinidae in the example of Alveolina tenuis "showing columella produced by polar thickening of the basal layer" (...), with (...) "tubular passage in the columella, continuous in subsequent chambers, without interruption by preseptal spaces". The progressive thickness of the basal layer pierced by tubular passages can be considered as favorable to the supposed homology between the basal layer and the central endoskeleton, but the pointed out lack of preseptal space between the basal layers of two successive chambers and mainly the absence of the BSC in A. tenuis are likely to withdraw such a possibility. Another example is given by Vicedo et al. (2009), with "basal layer pierced by tubular supplementary chamberlets" in Subalveolina dordonica; in the case of this species, the "supplementary chamberlets" open directly in the preseptal space (see Fig. 2.19 ![]() ), but as there is no obvious BSC in various known pictures (Reichel, 1936-1937, Pl. IV, fig. 2; Vicedo et al., 2009, text-fig. 5.1-5; Fig. 2.18-19
), but as there is no obvious BSC in various known pictures (Reichel, 1936-1937, Pl. IV, fig. 2; Vicedo et al., 2009, text-fig. 5.1-5; Fig. 2.18-19 ![]() ), no conclusive comparison can be established in that particular case.
), no conclusive comparison can be established in that particular case.
|
Figure 16: Confluent structure.
1: Idealistic diagrammatical drawing showing part of a cylindrical
chamber with primary chamberlets (Ch1) being parallel to the chamber axis and
secondary chamberlets (Ch2) following two contrasted helixes at an angle to the
chamber axis, justifying the former name of "helicoidal" structure" (after
Fleury, 1979).
Interpenetration of the two families of secondary chamberlets not figured. 2: Diagrammatic figure of the same structure in a flattened chamber
where secondary chamberlets are organized in parallel planes (after Fleury
& Fourcade, 1987).
Interpenetration of secondary chamberlets of the two families not figured. 3-4:
Material model of part of a cylindrical chamber (3) and section (4) by a plane
of which the trace is drawn on side of (3); interpenetration of secondary
chamberlets of adjoining layers create new intralocular communications called
stolons (St). 5-14: Objective
sections as examples of the confluent structure. 5: Drawing of a section of Sigalveolina renzi (in Fleury,
1979). 6: Part of a section of Sigalveolina
aff. renzi displaying layers of
secondary chamberlets parallel to the wall of chamber. 7: Section of a large
test of Cyclopseudedomia? n. sp. (see Fleury,
2018, text-fig. 14.29-37). 8: Sellialveolina
drorimensis (after De Castro, 1988). 9: "Pseudedomia" aff. multistriata (after Mavrikas
et al., 1994). 10: Sigalveolina
ovoidea (see Fig. 6.6 |
Dimorphism (of generations). "Distinct sexually and asexually reproducing generations (...) commonly reflected morphologically in the foraminifers, as differences in size of proloculus, megalospheric and microspheric, in ontogenic development , and in adult test size" (Loeblich & Tappan, 1988, p. 734) is perfectly fitting the Rhapydioninidae. Many examples can be seen in Fleury, 2014, text-fig. 1 and 2018, text-fig. 16. But, this apparent general rule is perhaps too simple, if we consider the variations known in some living Soritids (Lee et al., 1991); Sigalveolina ovoidea would be an example of such complexities (see Fleury, 2018, text-fig. 9).
Ecology (of the Rhapydioninidae). The Gavrovo-Tripolitza carbonate platform of Greece gives a good example of the prevailing surroundings which are preferred by the Rhapydioninidae. The known thick carbonate succession (about 3000 m from Kimmeridgian-Portlandian to Uppermost Cretaceous) is mainly made up of limestones deposited in internal platform environments (restricted lagoon), subtidal to intertidal (Fleury, 1980). Two periods are particularized by emersive tendencies, marked by intertidal to supratidal deposition and frequent vadose diagenesis (Bernier & Fleury, 1980); they are concomitant with presence of Sellialveolininae during Upper Albian-Cenomanian, Cuvillierinellinae and Rhapydionininae during the Campanian-Maastrichtian. Some periods of long term terrestrial emersion are even well characterized in Upper Cenomanian (Pomoni-Papaioannou & Zambetakis-Lekkas, 2009) and Maastrichtian (Mavrikas, 1993; Landrein et al., 2001) in Greece, as well as in Albania (Heba et al., 2009). Among many others, some examples of foraminiferal tests modified by vadose diagenesis are published, i.e., Rhapydionina gr. liburnica (in Fleury, 1970; Bernier & Fleury, 1980), Metacuvillierinella decastroi ("Raadshoovenia guatemalensis" in Fleury, 1977, Pl. 1.1 and 1.13) and Sigalveolina renzi (in Fleury, 1979, Pl. 1.15). The associations of organisms are themselves significant of these conditions, marked by ordinary absence of classic benthic groups linked to more open marine environments, such as mainly Rudists in general (with striking exceptions like the type locality of Cuvillierinella salentina in De Castro, 1990), Orbitolinids and Alveolinids in Albian-Cenomanian, Orbitoids and Siderolitids in Campanian-Maastrichtian (except on platform margins) and even calcareous algae (except in very particular context, see Deloffre et al., 1991), as in the case of the new taxon.
Endoskeleton. The classical and simple definition of this term was summarized by Reichel (1936-1937), who distinguished the exoskeleton ("forming the shell or carapace") from the endoskeleton ("internal deposits"). Hottinger (2006), more concerned about functional meaning and noting that the distribution of openings is reproducing the pattern of chamberlets (thus resulting from the same protoplasmic streaming), included the openings in the list of endoskeleton elements. The observation and its consequences cannot be denied but, in practice, it remains difficult to describe the exoskeletal septum taking no account of the "endoskeletal" holes traversing it. It is thought here that all skeleton elements make a unit, constructed in a moment (an "instar"), which must not be too much dissected, except for practical reasons. Nevertheless, Fig. 10.12-16 ![]() shows some tests of R. liburnica with differentiation between "the shell" and "internal deposits", presumably related to diagenesis, which seems to strengthen the distinction between these two entities.
shows some tests of R. liburnica with differentiation between "the shell" and "internal deposits", presumably related to diagenesis, which seems to strengthen the distinction between these two entities.
Equatorial section. Centered or subcentered section perpendicular to the coiling axis in adult planispiral tests, or the average position of fluctuating axis in pseudoplanispiral tests. All chambers forming the test are visible.
Evolute (coiling). Part of a test made up of chambers only in contact with preceding and following chambers, forming an Uncoiled Uniserial Termination (UUT). This final part can be made of tureen shaped chambers forming a cylindrical or more or less conical UUT, or elongate chambers giving birth to a flat terminal flange. See for example both generations of R. liburnica in Fleury, 2014, text-fig. 1, and Cyclopseudedomia mavrikasi, in Fleury, 2018, text-fig. 14.
Exoskeleton. Wall and septa "forming the shell or carapace", according to Reichel, 1936-1937. See endoskeleton.
Fishnet appearance (of central endoskeleton). A particular aspect of the central endoskeleton due to relatively large chamberlets separated by thin dividing walls (Figs. 2.15-17 ![]() and 5.8-10
and 5.8-10 ![]() ). In this configuration, characterizing rather "primitive" taxa, the various generations of primary and secondary chamberlets (BSC and SSC), sharing approximatively the same (relatively large) diameter, are uneasy to distinguish.
). In this configuration, characterizing rather "primitive" taxa, the various generations of primary and secondary chamberlets (BSC and SSC), sharing approximatively the same (relatively large) diameter, are uneasy to distinguish.
Flabelliform. Fanlike shaped.
Flexostyle. Canal flexostyle (or goulot). "Spiral tube applied like a chestnut sprout on surface of megalospheric proloculus" (translation from Reichel, 1936-1937). Always present in A tests. See Fig. 2.7 ![]() .
.
Floors. Continuous structures, usually observed in some elongate Alveolinidae (namely Praealveolina and Alveolinella), separating several layers of chamberlets which approximatively keep the same diameter when the coils are becoming higher (see Hottinger, 2006, text-fig. 70.F-G). The endoskeleton of Rhapydioninidae (especially in the rounded or faintly flattened tests) shows sometimes some comparable aspects, specifically in equatorial sections of "primitive" taxa (see Murciella, in Fleury, 2018, text-fig. 5.11-13; Sellialveolina, Fig. 9-11 and 9.13 ![]() ). But, as a general rule in the Rhapydioninidae, the only structure of the floor type is the continuous partition separating the primary chamberlets from the first secondary ones (the BSC). See Fig. 2.15
). But, as a general rule in the Rhapydioninidae, the only structure of the floor type is the continuous partition separating the primary chamberlets from the first secondary ones (the BSC). See Fig. 2.15 ![]() , next to last chamber; Figs. 5.18
, next to last chamber; Figs. 5.18 ![]() , 7.10
, 7.10 ![]() and 9.15
and 9.15 ![]() . In fact, the secondary chamberlets are only exceptionally likely separated by apparent continuous structures: Their pattern is either of the fishnet type when they are relatively large to compare with thickness of the dividing wall, either of the BSC-SSC type.
. In fact, the secondary chamberlets are only exceptionally likely separated by apparent continuous structures: Their pattern is either of the fishnet type when they are relatively large to compare with thickness of the dividing wall, either of the BSC-SSC type.
Foramen (plural: Foramina). Opening between successive chambers. See aperture.
Free interseptal blade. A neologism, proposed here in absence of a convenient term underlying the difference between the endoskeletal organization of some Soritids and typical Alveolinacea (see structural models, Fig. 17 ![]() ).
).
Glomerulus. "A nepiont characterized by streptospiral coiling (...) often exclusively in the microspheric generation" after Hottinger, 2006. A potentially useful term to be used in well centered sections.
Helicoidal structure. Previous name of the confluent structure. Corresponding to a rather theoretical image: The secondary chamberlets, being organized in adjacent layers oriented obliquely from one to the next, would constitute a double helix in a cylindrical chamber (see text-fig. 3.1 in Fleury, 2018; Figs. 16.1 ![]() ). Confluent structure is thought to be more expressive by insisting on the function, resulting in easy mixing of the protoplasm in the whole chamber.
). Confluent structure is thought to be more expressive by insisting on the function, resulting in easy mixing of the protoplasm in the whole chamber.
Hyperinvolute. A proposed name for a unique example among the Rhapydioninidae: "Pseudedomia" complanata (B test, Fig. 8.4-6 ![]() ). See archaiasiform.
). See archaiasiform.
Instar. "Single episode of test formation, commonly producing a single chamber" after Loeblich & Tappan, 1988.
Interseptal pillars. More or less cylindrical structure joining two successive septa. Absent in the Alveolinacea.
Involute (coiling). Coils are completely overlapping the preceding ones. See Figs. 2.6, 2.15 ![]() , 4.13, 4.15
, 4.13, 4.15 ![]() , 5.6
, 5.6 ![]() , 6.4-5
, 6.4-5 ![]() , 8.3, 8.13, 8.18-19, 8.23, 8.25
, 8.3, 8.13, 8.18-19, 8.23, 8.25 ![]() , 9.3, 9.6, 9.21
, 9.3, 9.6, 9.21 ![]() , 11.2
, 11.2 ![]() , 11.5, 11.7
, 11.5, 11.7 ![]() , ...)
, ...)
Lenticular test. Small test resembling a lentil, with tight coiling, sharp periphery in axial section and no final uncoiling.
Lumen (plural: Lumina). Inside of chambers, cavity.
Megalosphere. A synonym of A proloculus used in dimorphic species.
Megalospheric generation (A tests). In dimorphic species, generation with large proloculus and small adult test. See dimorphism.
Microsphere. First chamber in B tests (B proloculus). Very rarely observed in Rhapydioninidae because of their small size.
Microspheric generation (B tests). In dimorphic species, generation with small proloculus and large adult test. See dimorphism.
Miliolid or milioline (coiling). An informal term used for more or less well observable central part in some A tests and all B tests. It refers to the mode of coiling common in the Miliolacea: Quinqueloculine, triloculine or eventually pseudotriloculine (see Pęcheux, 2002). They are relatively easy to characterize in simple taxa (see Munier-Chalmas & Schlumberger, 1885, text-figs. 1-8, and modifications by Hottinger, 2006, text-fig. 68), but difficult to distinguish in non-oriented random sections, being progressively changing on their way to reach the planispiral mode of coiling. The term streptospiral is used in a more comprehensive sense, when the milioline mode is not even certain, the coiling being apparently constantly moving in random changing planes.
Models. See structural models.
Nautiloid. Test involute, faintly compressed, with rounded periphery, like the cephalopod Nautilus.
Nepionic stage (nepiont). Young part of the test following the proloculus. Nepiont is the name of this part.
Notched opening. Term presumably created by Calvez (1988), for the genus Pseudonummoloculina: "A complicated slitlike aperture bordered by a series of notches derived from the floor and the roof of the chambers" ("ouverture complexe, en fente arquée, bordée par des crénulations issues du plancher et du toit des loges"). Various structures of this kind were observed by De Castro (1987), Hottinger et al. (1989), Fourcade & Fleury (2001) and Schlagintweit & Rashidi (2016) in diverse taxa. These structures seem to be primitive ones, premonition of more elaborate sets of rounded openings proper to the classical Alveolinacea.
Openings. Useful general term used to designate both apertures and foramina.
Peripheral zone (of chambers). Part of the chamber occupied by the layer of primary chamberlets, underneath the wall.
Planispiral (coiling). Coiling in one plane perpendicular to a permanent axis of coiling.
Porcelaneous test. After Haynes (1981) "wall may resemble porcelain with a shiny white surface (...) composed of three layers: A thick layer of laths in random array with thin inner and outer veneers... arranged parallel to the surface". In section (transmitted light) the ordinary color is grey, but well preserved tests appear light-brown colored; in rare cases, only the wall and septum are colored, the endoskeleton being grey. It is supposed resulting from differential diagenesis, affecting slightly heterogeneous material. See Fig. 10.12-16 ![]() .
.
Postseptal cells. Proper to the Alveolinidae. "Cellules postseptales" in Reichel, 1936, p. 75. Small subspherical chamberlets stuck on the previous septum, just beneath the wall in Subalveolina dordonica. See Fig. 2.20-21 ![]() .
.
Preseptal pillars. In adult tests, more or less cylindrical pillars (or buttresses) joining the central endoskeleton to the septum through the preseptal space (Fig. 17.A-B ![]() and many sections). In most cases, only appearing when the central endoskeleton is well developed, but with apparent exceptions (Figs. 2.15
and many sections). In most cases, only appearing when the central endoskeleton is well developed, but with apparent exceptions (Figs. 2.15 ![]() and 4.1
and 4.1 ![]() ).
).
Preseptal space (or canal, or passage in the Alveolinidae). Fundamental and distinctive element of the Alveolinacea in general and the Rhapydioninidae in particular. Empty volume beneath the septum, where the protoplasmic columns filling the chamberlets are mixing (Figs. 7.7 ![]() and 10.11
and 10.11 ![]() ). In the Rhapydioninidae, the cloisonnettes are extended in the periphery of the preseptal space, forming regular triangular denticles (i.e., Fleury, 2014, text-figs. 1.E, 1.H and 8.P; 2016, text-fig. 9.15 and 9.23; 2018, text-fig. 11.12 and 11.15-16; Figs. 7.16
). In the Rhapydioninidae, the cloisonnettes are extended in the periphery of the preseptal space, forming regular triangular denticles (i.e., Fleury, 2014, text-figs. 1.E, 1.H and 8.P; 2016, text-fig. 9.15 and 9.23; 2018, text-fig. 11.12 and 11.15-16; Figs. 7.16 ![]() and 17.A-B
and 17.A-B ![]() ).
).
Primary chamberlets (or peripheral). In the sense of the first materialized and sometimes remaining the only ones, they constitute a unique layer stuck under the outer wall in the Alveolinacea. Isolated by the cloisonnettes, they constitute the alone chamberlets in "primitive" taxa (see Fig. 9.1-5 ![]() ) and in first coils in almost all members of the Rhapydioninidae. They are tubular cylindrical in the Cuvillierinellinae subfamily and radially elongate in Rhapydionininae (Fig. 17.A-B
) and in first coils in almost all members of the Rhapydioninidae. They are tubular cylindrical in the Cuvillierinellinae subfamily and radially elongate in Rhapydionininae (Fig. 17.A-B ![]() , and many sections).
, and many sections).
Primary openings. Openings corresponding exactly in number (and giving birth) to the primary chamberlets of the next chamber. Their particularity is to be obliquely oriented and slightly shifted from the axis of the corresponding chamberlets (i.e., Fleury, 2014, text-fig. 1H; 2016, text-figs. 3C, 5.20 and 7.2; 2018, text-fig. 11.13; Figs. 2.7 ![]() , 7.8
, 7.8 ![]() , 10.1
, 10.1 ![]() and 17.A-B
and 17.A-B ![]() ).
).
Proloculus (plural: Proloculi). First chamber. In A tests, usually circular in section, followed by the flexostyle. Almost unknown in B tests because of its small size
Pseudoevolute. A particular mode of uncoiling, characterized by partly evolute chambers of which an end keeps in contact with the involute part of the test. It is often an intermediate stage between the involute and evolute stages. Can be confused with true evolute tests in absence of well situated sections.
Pseudoplanispiral (coiling). Some taxa never reach the perfect planispiral coiling. The pseudoplanispiral coiling is revealed by sigmoid (S-shaped) axial sections of adult tests. It is assumed as resulting from regular rotation of the coiling axis following a conical surface with apex coincident with the proloculus. See in particular: Figs. 4.2, 4.15 ![]() , 5.10
, 5.10 ![]() , 6.3
, 6.3 ![]() , 11.9
, 11.9 ![]() , 14.19
, 14.19 ![]() and 15.5
and 15.5 ![]() .
.
Radiance. A reference to the known propagation of taxa and their kinship in time and space. In case of uncertainty between two similar taxa, the knowledge of their radiances, besides their morphologic analogies and dissemblances, may help to choose the most suitable taxonomic treatment. Sometimes a temporary argument, depending on knowledge accuracy on one point.
Residual pillars. A term proposed by Reiss et al., 1964, for a theoretical process of forming the Alveolinidae endoskeleton by lateral fusion of Soritid interseptal structures ("a massive "couche basale" may have arisen through coalescence of interseptal buttresses" (...) the (...) "residual buttresses" resulting from their narrowing in the preseptal canal. Such a theory was perfectly legible at that time, because of the coiling-uncoiling likeness of the two groups, but is now denied by the knowledge of their diverse kinships. Thus, the theory being abandoned, the term is too much linked to it to be preserved.
Ribs. Thin external costa, joining perpendicularly successive chambers sutures, like in some Peneroplis and Dendritina. Exceptionally observed in Cyclopseudedomia smouti and C. mavrikasi (in Fleury, 2018, text-figs. 11.8, 11.12 and 13.8; Figs. 7.7, 7.16 ![]() and 17.A
and 17.A ![]() ).
).
Scattered secondary chamberlets (SSC). Dispersed tubular chamberlets of small diameter imbedded in the massive central endoskeleton. Associated with the BSC in involute coils when the observation is not prevented by too short coiling of this part (Cyclopseudedomia and Rhapydionina). Not to be opposed to the fishnet aspect of the endoskeleton in section (some Cuvillierinella, Metacuvillierinella decastroi and Sellialveolina of the viallii type) resulting from a distance between chamberlets smaller than their diameter. Observed in the presumably Alveolinidae genus Subalveolina (Fig. 2.18-19 ![]() ). Structure not incompatible with the confluent structure, as shown by Murciella (Fig. 5.19
). Structure not incompatible with the confluent structure, as shown by Murciella (Fig. 5.19 ![]() ), Sigalveolina (Fig. 6.12
), Sigalveolina (Fig. 6.12 ![]() ), Cyclopseudedomia (Fig. 7.6-7
), Cyclopseudedomia (Fig. 7.6-7 ![]() ), "Pseudedomia" (Fig. 8.11
), "Pseudedomia" (Fig. 8.11 ![]() and 8.27
and 8.27 ![]() ), Sellialveolina (Fig. 9.25
), Sellialveolina (Fig. 9.25 ![]() ) and Neomurciella (Fig. 11.13
) and Neomurciella (Fig. 11.13 ![]() ).
).
Secondary chamberlets (or central). Appearing secondarily, and sometimes absent in juvenile and even in compressed chambers of adult tests (see Fig. 7.16 ![]() ). Comprising the Basal Secondary Chamberlets (BSC) and the Scattered Secondary Chamberlets (SSC). Rarely represented by an apparent unique set of relatively large chamberlets having a fishnet appearance. (See Figs. 2.15-17
). Comprising the Basal Secondary Chamberlets (BSC) and the Scattered Secondary Chamberlets (SSC). Rarely represented by an apparent unique set of relatively large chamberlets having a fishnet appearance. (See Figs. 2.15-17 ![]() and 5.8-9
and 5.8-9 ![]() ).
).
Septulum (plural: Septula). A general term used by various authors: For example "Secondary partitions extending from chamber roof, and partly subdividing the chambers" for Loeblich & Tappan, 1988; "Endoskeleton wall-like partition extending from the lateral wall into the chamber lumen, dividing it into compartments (chamberlets)" for Hottinger, 2006. Such definitions do not distinguish in particular the peripheral partitions of the Alveolinacea (cloisonnettes) from the radial partitions free at their distal part of the Soritacea (Fig. 17.C ![]() ), in the case of morphologic convergence. See subepidermal partition and free interseptal blade.
), in the case of morphologic convergence. See subepidermal partition and free interseptal blade.
Septum (plural: Septa). The part of the exoskeleton wall covered by the following chamber, through which openings (successively foramina and apertures) are hollowed out.
Sigmoid axial section. S-shaped aspect of tests in "axial" section of some taxa. See pseudoplanispiral coiling.
Slow axial rotation (SAR). The sigmoid aspect of centered "axial" sections is supposed resulting from a regular rotation of the coiling axis, following a conical surface centered on the proloculus. See Metacuvillierinella decastroi (in Fleury, 2016, text-figs. 10-11; "axial" sections in Figs. 3 ![]() , 4.2, 4.15
, 4.2, 4.15 ![]() and 5.10
and 5.10 ![]() ).
).
Streptospiral (coiling). The most comprehensive word for apparently irregular coiling in nepionic stages, when even the fundamental modes of milioline coiling are not distinguished. May be often an aspect resulting from more or less progressive passage from miliolid to planispiral coiling in random sections. See Kellner et al. (2019, text-figs. 4.1-2 and 6.2) for the interpretation of the modern Alveolinidae Borelis schlumbergeri nepiont.
Structural models (Fig. 17 ![]() ). Models A and B summarize the endoskeletal organization of pseudoevolute and evolute chambers of the Cuvillierinellinae (Cyclopseudedomia smouti) and Rhapydionininae (Rhapydionina liburnica) subfamilies, respectively. Differentiations of genera and species from these models in each subfamily are only a matter of detail, such as the size of the preseptal pillars and the diameter of the sets of primary and secondary chamberlets, sometimes distinct or possibly identical, like in the "fishnet" organization (in "primitive" taxa). Absence or presence (in "advanced "taxa) of the confluent structure is another possible variant. The drawings do not show the basal secondary chamberlets, only present in involute parts of tests. The third model (C), given for comparison, is an example of some Soritid (Rhabdorites malatyaensis), underlying the particularity of the structure: Mixing of protoplasmic columns issued from the apertures is possible in the whole chamber; the dividing plates in the marginal parts of the chamber are called "free interseptal blades".
). Models A and B summarize the endoskeletal organization of pseudoevolute and evolute chambers of the Cuvillierinellinae (Cyclopseudedomia smouti) and Rhapydionininae (Rhapydionina liburnica) subfamilies, respectively. Differentiations of genera and species from these models in each subfamily are only a matter of detail, such as the size of the preseptal pillars and the diameter of the sets of primary and secondary chamberlets, sometimes distinct or possibly identical, like in the "fishnet" organization (in "primitive" taxa). Absence or presence (in "advanced "taxa) of the confluent structure is another possible variant. The drawings do not show the basal secondary chamberlets, only present in involute parts of tests. The third model (C), given for comparison, is an example of some Soritid (Rhabdorites malatyaensis), underlying the particularity of the structure: Mixing of protoplasmic columns issued from the apertures is possible in the whole chamber; the dividing plates in the marginal parts of the chamber are called "free interseptal blades".
Subepidermal partitions (lames sous-épidermiques). The term is used by Henson (1948) in a large meaning, including "main partitions and sub-epidermal partitions" This is the name given by Hamaoui & Fourcade (1973) to partitions dividing into compartments the marginal zone of chambers ("compartimentent généralement la zone marginale sous-épidermique, ou périphérique interne des loges"). For Hottinger 2006, the term is "unspecified", corresponding to "all shell elements subdividing external (lateral) parts of the chamber lumen". Thus it seems not far from "septula", considered here too large a term for the particular cloisonnettes of the Alveolinacea. In absence of precise term describing the partitions of the Soritid Rhabdorites (Fig. 17.C ![]() ) the term "free interseptal blade" is proposed here.
) the term "free interseptal blade" is proposed here.
|
Figure 17: Structural
models of evolute chambers in the two subfamilies of the Rhapydioninidae, for
comparison with a Soritidae (genus Rhabdorites
Fleury, 1996)
of superficial comparable general morphology.
A: Cyclopseudedomia smouti, assumed as good representative of the
subfamily Cuvillierinellinae, with narrow peripheral zone of primary chamberlets.
Note that external ribs and secondary chamberlets of confluent structure are not
generalized to all taxa; usual
variations in size and proportions of common elements have no general taxonomic
meaning.
B: Rhapydionina liburnica,
representative of the Rhapydionininae, particularized by the large peripheral
zone of primary chamberlets. C: Rhabdorites malatyaensis (Sirel,
1976) is characterized by interseptal radial partitions free at their
distal ending, restricted to the peripheral part of the chambers and forming an
interlacing underneath the septum, leaving a large (almost) undivided
interseptal axial space (a kind of "pseudo-postseptal space") ; this
structure is here called free interseptal
blades. |
Suture (of chambers). Limit between two successive chambers on external surface of involute and evolute tests, ordinary slightly depressed; easily observable in equatorial sections. See outline of sections Figs. 2.7 ![]() , 4.3
, 4.3 ![]() , 5.1
, 5.1 ![]() and 10.1
and 10.1 ![]() .
.
Taxon (plural: Taxa). Any taxonomic unit, such as family, subfamily, genus, species, ...
Umbilicus. Axial depression in planispiral or pseudoplanispiral tests. Surrounded by the last coil in advolute coiling (Fig. 2.2 ![]() ).
).
Uncoiled uniserial termination (UUT). Evolute part of test, when any chamber is only in contact with the previous and following ones. May be cylindrical to conical (tureen shaped chambers like Rhapydionina A) or flabelliform (banana shaped chambers, like Rhapydionina B) or even discoidal (annular chambers like Cyclopseudedomia smouti B test).
Wall. Outer shell or carapace. Translation of the "muraille" in Reichel, 1936, p. 8 ("nous avons nommé exosquelette celui qui forme carapace. C'est la muraille").
| Nomenclatural note: Life Sciences Identifier (LSID) http://zoobank.org/References/F831BD0E-D7AA-48E6-8C02-04A28A3B7D9F Metacuvillierinella sireli Fleury & Özkan, 2020 http://zoobank.org/NomenclaturalActs/A951BB3A-E899-4E6F-8896-55D66B4745AA |