◄ Carnets Geol. 15 (20) ►
![]()
Sommaire
[1. Introduction]
[2. Localisation géographique et cadre géologique]
[3. Cadre stratigraphique de l'Aptien des bassins étudiés]
[4. Matériel d'étude]
[5. Systématique]
[6. Discussion]
[7. Conclusion]
[Références bibliographiques] et ... [Planches]
Le Jardin de la Chapelle, Abbaye de Maubec, route d'Allan,
26200 Montélimar (France)
Publié dans sa forme définitive (pdf) le 25 décembre 2015
[Éditeur : Bruno ]
![]()
Des représentants du genre Hemidiadema 1846, provenant de l'Aptien (Crétacé inférieur) d'Espagne – plus précisément des provinces de Castellón, de Tarragone, de Barcelone et de Teruel – sont étudiés. Tous les individus d'Hemidiadema jusqu'ŕ présent cités ou figurés provenant de ces régions avaient été rapportés ŕ l'espčce Hemidiadema rugosum , 1846. Leur révision taxonomique – basée sur le matériel historique et de nouvelles récoltes – démontre que ces individus n'appartiennent pas ŕ l'espčce décrite par . Trois nouvelles espčces sont ainsi proposées et décrites : Hemidiadema batalleri nov. sp., H. malladai nov. sp. et H. forcallensis nov. sp. L'espčce H. rugosum , 1846, n'est connue que par un unique exemplaire provenant de l'Aptien de Castellón. Des remarques sur la paléoécologie et les répartitions géographique et stratigraphique des différentes espéces décrites sont également formulées.
Échinides ; Glyphocyphidae ; Hemidiadema ; nove species ; Aptien ; Espagne ; révision systématique.
A. (2015).- Révision des espčces du genre Hemidiadema , 1846 (Euechinoidea, Camaradonta, Glyphocyphidae), de l'Aptien (Crétacé inférieur) d'Espagne.- Carnets Geol., Madrid, vol. 15, nº 20, p. 279-330.
A review of the species of the genus Hemidiadema , 1846 (Euechinoidea, Camaradonta, Glyphocyphidae), from the Aptian (Lower Cretaceous) of Spain.- Specimens belonging to the genus Hemidiadema , 1846, from Aptian (Lower Cretaceous) strata from the Spanish provinces of Castellón, Tarragona, Barcelona and Teruel –were studied. All specimens of Hemidiadema from these regions cited or described in the literature until now were related to the species Hemidiadema rugosum , 1846. Their taxonomic revision – based on historical material and new collecting – demonstrates that these individuals do not belong to the species described by , but represent new taxa. Three new species are proposed and described: Hemidiadema batalleri nov. sp., H. malladai nov. sp., and H. forcallensis nov. sp). The species H. rugosum , 1846, is known by a single specimen from the Aptian of Castellón. Remarks on the paleoecology and stratigraphic and geographic distributions of these species are given.
Echinoids; Glyphocyphidae; Hemidiadema; nove species; Aptian; Spain.
Les citations du genre Hemidiadema , 1846, dans les terrains du Crétacé d'Espagne sont assez rares et surtout récentes. Une seule espčce y a été jusqu'ŕ présent mentionnée, dans l'Aptien : Hemidiadema rugosum , 1846. Cette espčce est citée dans la région de Morella et de Forcall, province de Castellón (, 1928, p. 149 ; , 1991, p. 213 ; 2003, p. 76 ; & , 2000, p. 18 ; et al., 2007, photo 25 ; , 2008, p. 6, 21, photos 35-36), dans la région de Punta de l'Ŕliga, province de Tarragone (, 1991, p. 213), dans le Massif du Garraf, province de Barcelone ( et al., 2008, p. 159) et Alcoraia en Alicante ( & , 2006, p. 39, 70, Fig. 22).
L'espčce H. rugosum , 1846, a été récemment révisée (, 2013) ŕ partir des examens détaillés de l'holotype et de l'exemplaire décrit et figuré par (1892, p. 70, Pl. IV, figs. 5-6), provenant tous deux des "Grčs vert ferrugineux" de l'Aptien supérieur de Grandpré (Ardennes, France).
Les spécimens de l'Aptien d'Espagne antérieurement étudiés et figurés – provenant de la collection conservée au Musée géologique du Séminaire de Barcelone (, 1991, p. 213, Pl. 11, figs. 4-6 ; 2003, p. 76, Pl. 2, figs. 14-16) – se distinguent nettement de l'espčce décrite par et de toute autre espčce connue ( et al., 2012 ; , 2013). Seul un individu provenant de l'Aptien d'Alicante (dépression de l'Alcoraia) (& , 2006, Fig. 22 ; et al., 2012, p. 3, Fig. 3.1) se rattache bien ŕ H. rugosum et constitue ainsi la seule découverte actuellement connue de cette espčce en Espagne.
Les
bassins d'oů
provient le
matériel étudié – bassins
du Maestrazgo,
de Salou-Garraf et la dépression de
l'Alcoraia (Domaine Prébétique)
(Fig.
1 ![]() ) – situés
ŕ l'Est de l'Espagne se
sont formés au cours du Jurassique
supérieur et
du Crétacé inférieur sur les marges est et nord de la Plaque Ibérique en
relation avec l'ouverture du golfe
de Gascogne et l'Atlantique Centre-Est et du Nord (
et al., 2001).
Le bassin du Maestrazgo,
le plus méridional des trois, montre une paléogéographie interne trčs complexe
et peut ętre subdivisé en sept sous-bassins ( & , 1996). C'est
dans trois
d'entre eux (sous-bassins de Morella, d'El Perelló et de Galve) qu'ont
été récoltés des représentants
du genre Hemidiadema. Le bassin de Salou-Garraf est étroitement lié ŕ
celui du Maestrazgo,
mais expose des facičs
marins généralement plus profonds
liés ŕ sa situation paléogéographique plus distale sur la marge ( et
al., 2001). La dépression
de l'Alcoraia,
prčs d'Alicante, se situe dans la zone Prébétique, large unité tectonique alpine qui
correspond ŕ la partie externe de l'avant-pays plissé de la Cordillčre
Bétique,
au Sud-Est de l'Espagne. Pendant le Crétacé inférieur, ce
domaine tectonique constitue une
vaste plate-forme intracontinentale sur la marge passive méridionale de la
Plaque Ibérique
(
& , 2007).
Plus précisément, la région de l'Alcoraia
se situe dans la partie sud de cette plate-forme (Domaine Prébétique) oů se
sont développés des environnements hémipélagiques.
) – situés
ŕ l'Est de l'Espagne se
sont formés au cours du Jurassique
supérieur et
du Crétacé inférieur sur les marges est et nord de la Plaque Ibérique en
relation avec l'ouverture du golfe
de Gascogne et l'Atlantique Centre-Est et du Nord (
et al., 2001).
Le bassin du Maestrazgo,
le plus méridional des trois, montre une paléogéographie interne trčs complexe
et peut ętre subdivisé en sept sous-bassins ( & , 1996). C'est
dans trois
d'entre eux (sous-bassins de Morella, d'El Perelló et de Galve) qu'ont
été récoltés des représentants
du genre Hemidiadema. Le bassin de Salou-Garraf est étroitement lié ŕ
celui du Maestrazgo,
mais expose des facičs
marins généralement plus profonds
liés ŕ sa situation paléogéographique plus distale sur la marge ( et
al., 2001). La dépression
de l'Alcoraia,
prčs d'Alicante, se situe dans la zone Prébétique, large unité tectonique alpine qui
correspond ŕ la partie externe de l'avant-pays plissé de la Cordillčre
Bétique,
au Sud-Est de l'Espagne. Pendant le Crétacé inférieur, ce
domaine tectonique constitue une
vaste plate-forme intracontinentale sur la marge passive méridionale de la
Plaque Ibérique
(
& , 2007).
Plus précisément, la région de l'Alcoraia
se situe dans la partie sud de cette plate-forme (Domaine Prébétique) oů se
sont développés des environnements hémipélagiques.
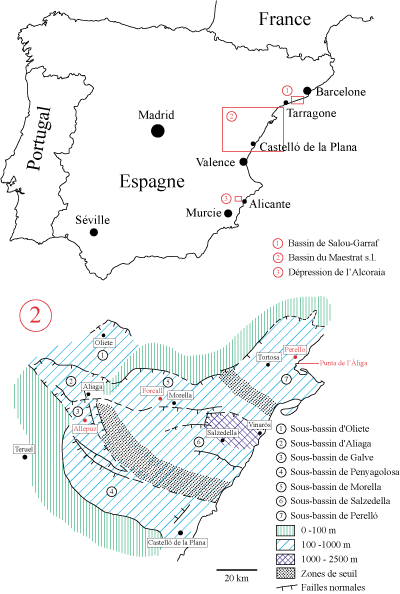
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1: Situation géographique des différentes localités étudiées (carte du Bassin du Maestrat d'aprčs et al., 2001).
La lithostratigraphie suit et al. (2014) et la biozonation ammonitique de & (2014) et et al. (2014).
Dans le bassin du Maestrazgo, les Hemidiadema étudiés proviennent des parties moyenne et supérieure de la Formation des "Marnes de Forcall" ("Forcall Formation") de l'Aptien inférieur. Cette formation se caractérise par une prédominance de marnes et de calcaires marneux. La faune d'ammonites des "Marnes de Forcall", intensivement étudiée ces derničres années ( et al., 2009a, 2010, 2012a, 2014 ; et al., 2013 ; et al., 2013 ; et al., 2014), a permis un découpage biostratigraphique précis.
Dans le sous-bassin de Morella (Fig. 2a ![]() : coupe du Mas del Roc, environs de Forcall ; province
de Castellón) les Hemidiadema
récoltées proviennent des marnes pyriteuses datées de la partie supérieure
de la biozone ŕ Deshayesites forbesi et de la partie basale de la biozone ŕ Deshayesites
deshayesi (Aptien inférieur) (
et al., 2009a, p. 867, Fig. 6 ;
et al., 2014, Figs. 1B, 2 ;
et al., 2014, Fig. 1c).
Deux espčces y ont été reconnues : H.
forcallensis nov.
sp. (Mola d'en Camarŕs, Mas del Roc, environs de Forcall - province de
Castellón) qui y est fréquente et Hemidiadema
sp. A. (Mola d'en Camarŕs,
nord-est de Forcall - province
de Castellón) qui n'est connue que par un seul exemplaire.
: coupe du Mas del Roc, environs de Forcall ; province
de Castellón) les Hemidiadema
récoltées proviennent des marnes pyriteuses datées de la partie supérieure
de la biozone ŕ Deshayesites forbesi et de la partie basale de la biozone ŕ Deshayesites
deshayesi (Aptien inférieur) (
et al., 2009a, p. 867, Fig. 6 ;
et al., 2014, Figs. 1B, 2 ;
et al., 2014, Fig. 1c).
Deux espčces y ont été reconnues : H.
forcallensis nov.
sp. (Mola d'en Camarŕs, Mas del Roc, environs de Forcall - province de
Castellón) qui y est fréquente et Hemidiadema
sp. A. (Mola d'en Camarŕs,
nord-est de Forcall - province
de Castellón) qui n'est connue que par un seul exemplaire.
Dans le sous-bassin
d'El Perelló (Fig. 2b ![]() : coupe
de Cala del Moro,
prčs de Punta de l'Ŕliga, province de Tarragone) de fréquents H. batalleri
nov. sp. ont été récoltés dans les marnes pyriteuses
ŕ Plicatules de la partie inférieure de la biozone Dufrenoyia
furcata (
et al., 2010, Fig. 8).
: coupe
de Cala del Moro,
prčs de Punta de l'Ŕliga, province de Tarragone) de fréquents H. batalleri
nov. sp. ont été récoltés dans les marnes pyriteuses
ŕ Plicatules de la partie inférieure de la biozone Dufrenoyia
furcata (
et al., 2010, Fig. 8).
Dans le sous-bassin de Galve, plus précisément dans la région située entre les villages d'Allepuz et de Jorcas (province de Teruel), des marnes calcaires finement bioclastiques ont fourni de fréquentes H. malladai nov. sp., associées ŕ d'autres échinides [Hyposalenia lardyi (, 1856) en particulier]. Ces marnes calcaires appartiennent aux "Marnes de Forcall" ("Forcall Formation") de l'Aptien inférieur. Cependant, la région de Jorcas / Allepuz n'a pas été incluse dans les travaux de révisions récentes du canevas stratigraphique régional ( et al., 2009, 2010). Il ne nous est pas possible de rattacher les marnes calcaires avec certitude ŕ une biozone ammonitique précise. On ne peut que supposer qu'elles représentent la partie supérieure de la biozone ŕ Deshayesites forbesi ou la partie inférieure de la biozone ŕ Deshayesites deshayesi.
Dans
le bassin de Salou-Garraf (Fig. 2c ![]() :
coupe de
Can Cassanyes,
ŕ l'est de Castellet, province de Barcelone) l'unique
Hemidiadema découverte (coll.
E. ) dans cette région appartient
ŕ l'espčce H. malladai nov. sp. Cet individu provient de
la partie inférieure de l'Aptien supérieur,
plus précisément de la partie inférieure de la biozone ŕ Epicheloniceras martini
de
l'unité des "Marnes de Vallcarca". Cette unité se compose de niveaux marneux et
marno-calcaires, caractérisés par la profusion de
spongiaires et autres
invertébrés marins (,
2007 ;
et
al., 2009b).
:
coupe de
Can Cassanyes,
ŕ l'est de Castellet, province de Barcelone) l'unique
Hemidiadema découverte (coll.
E. ) dans cette région appartient
ŕ l'espčce H. malladai nov. sp. Cet individu provient de
la partie inférieure de l'Aptien supérieur,
plus précisément de la partie inférieure de la biozone ŕ Epicheloniceras martini
de
l'unité des "Marnes de Vallcarca". Cette unité se compose de niveaux marneux et
marno-calcaires, caractérisés par la profusion de
spongiaires et autres
invertébrés marins (,
2007 ;
et
al., 2009b).
Dans le Domaine Prébétique, au niveau de la dépression de l'Alcoraia (province d'Alicante), l'Aptien s'observe au niveau d'une petite colline au sud-ouest de la Serra Mitjana, ŕ 12 km au nord-ouest d'Alicante. Des dépôts alluvionnaires masquant une partie de la coupe, seules les zones ŕ Deshayesites oglanlensis, D. forbesi et D. furcata de l'Aptien inférieur sont visibles ( et al., 2012b, p. 338). Toutefois, c'est un niveau stratigraphiquement plus élevé observable ŕ Loma de Casa lo Bolini et daté du sommet de l'Aptien supérieur qui a fourni une Hemidiadema, la seule Hemidiadema rugosum , 1846, connue actuellement en Espagne (& , 2006, Fig. 22 ; et al., 2012, p. 3, Fig. 3.1). La faune d'ammonites de Casa lo Bolini est essentiellement représentée par des nuclei pyriteux de petites tailles, difficilement identifiables. Dans les niveaux situés en-dessus et en-dessous de celui oů l'H. rugosum a été recueillie cette ammonitofaune est essentiellement constituée par des taxons (Desmoceratidés et Phylloceratidés) ne permettant pas de fournir un âge stratigraphique précis. De petites ammonites hétéromorphes appartenant au genre Ptychoceras sont également assez communes parmi ces ammonites. Les seuls exemplaires spécifiquement déterminables et d'intéręt biostratigraphique appartiennent ŕ l'espčce Neosilesites nepos (, 1917) et ont été collectés au-dessus du niveau ayant fourni l'H. rugosum. La répartition stratigraphique de cette espčce s'étend de la partie supérieure de l'Aptien ŕ l'Albien inférieur selon les auteurs (, 2007 ; et al., 2012). Toutefois, dans une autre étude, il est suggéré que N. nepos est limité ŕ la base de l'Albien inférieur ( et al., 2001). La présence assez fréquente de Ptychoceras rappelle la faune d'ammonites de l'Aptien supérieur étudiée par (1969). Cet auteur y décrit un nouveau genre de l'Aptien supérieur – Neoastieria – qui semble ętre étroitement lié au genre Neosilesites mais sans y avoir reconnu de morphologies typiques des Neosilesites. Cette absence pourrait confirmer que N. nepos soit limité ŕ l'Albien basal comme l'a suggéré et al. (2001). Ŕ la lumičre de toutes ces données, le spécimen d'H. rugosum trouvé ŕ Loma de Casa lo Bolini proviendrait d'un niveau attribuable au sommet de l'Aptien supérieur, les niveaux juste en-dessus appartenant déjŕ ŕ l'Albien basal. Notons que cet âge est le męme que l'âge des niveaux ayant fourni des H. rugosum en France : "Grčs ferrugineux" de l'Aptien supérieur sensu gallico (Clansayésien) de Grandpré (Ardennes) (, 2013) et "Grčs glauconieux ŕ Discoďdes decoratus" de l'Aptien supérieur (Zone ŕ Acanthohoplites nolani, Sous-zone ŕ Diadochoceras nodosocostatum) du Teil prčs de Montélimar (Ardčche) (, in prep.). Les ammonites de la région de l'Alcoraia ont déjŕ été étudiées en détail par et al. (2004) et et al. (2012b) mais dans ces travaux l'étude des niveaux ŕ nuclei pyriteux évoqués ci-dessus n'a pas été incluse.
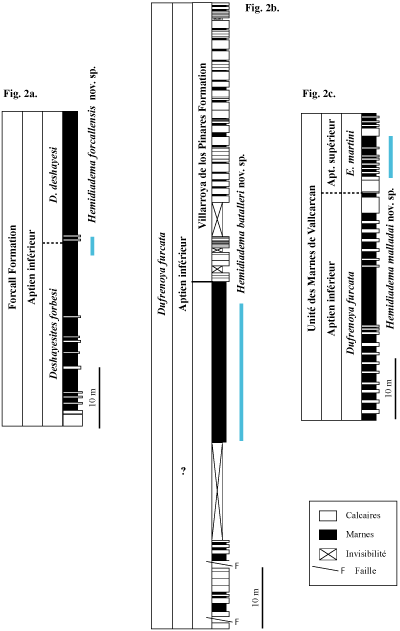
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2:
Coupes
stratigraphiques des localités étudiées et distribution verticale des espčces
d'Hemidiadema décrites.
2a. Coupe du Mas del Roc, Forcall, Bassin du Maestrat (
et al., 2009a, Fig. 6).
2b. Coupe
de Cala del Moro, Punta de l'Ŕliga, Bassin de Perello (
et al., 2010, Fig. 8).
2c. Coupe de
Can Cassanyes, Bassin de Salou-Garraf (
et
al., 2009b, simplifiée).
Le matériel étudié se compose des individus appartenant ŕ la collection de l'auteur du présent travail, d'individus récoltés par Oriol et Josep Anton lors de prospections sur le terrain dans le cadre de leurs travaux respectifs et des échantillons conservés au Musée géologique du Séminaire de Barcelone appartenant ŕ la collection de l'Abbé Josep Ramon . S'y ajoutent un exemplaire gracieusement donné par Brigitte (Cheval-Blanc, France) et deux autres prętés par Carlos (Alicante, Espagne) et Eduard (Barcelone, Espagne), tous les trois amateurs.
Conventions des institutions oů sont déposés les fossiles étudiés (holotypes et paratypes) :
MGSB : Musée géologique du Séminaire de Barcelone, Espagne ;
MNHN.F : Muséum national d'Histoire naturelle, domaine Sciences de la Terre, Paris, France.
Les différentes espčces décrites dans ce travail l'ont été en les comparant aux espčces du Crétacé inférieur de France actuellement connues, en particulier H. rugosum 1846, et H. neocomiense (, 1869) qui ont été récemment révisées (, 2013). La systématique suit & (2010).
Classe Echinoidea , 1778
Sous-classe Euechinoidea , 1860
Infra-classe Acroechinoidea , 1981
Echinacea , 1876
Ordre Camarodonta , 1912
Infra-ordre Temnopleuridea , 2010
Famille Glyphocyphidae , 1889
Genre Hemidiadema in & , 1846
Concernant la diagnose émendée du genre, les remarques d'ordre historique et la nomenclature descriptive utilisée, se reporter au travail de (2013).
Espčce-type. Hemidiadema rugosum , 1846.
Autres espčces associées. Hemidiadema intermedius (, 1864), H. neocomiense (, 1869), H. guebhardi , 1920 [nomen dubium] (= ? H. neocomiense (, 1869)) et les trois nouveaux taxons ici décrits : H. batalleri nov. sp., H. malladai nov. sp. et H. forcallensis nov. sp.
Hemidiadema rugosum in & , 1846
(Pls. 1 ![]() - 2
- 2 ![]() ; Pl. 15
; Pl. 15 ![]() ,
figs. 1-3)
,
figs. 1-3)
1846 – Hemidiadema rugosum : & , p. 351.
1864 – Glyphocyphus rugosus : , p. 533, 543, Pl. 1128, figs. 16-22.
1892 – Hemidiadema rugosum : , p. 69 Fig. 14, 70, 97, Pl. IV, figs. 5-6.
1933 – Glyphocyphus rugosus : , p. 83, Fig. 22c.
1943 – Hemidiadema rugosum : , p. 21 Fig. 18e, p. 22 Fig. 20c, Fig. 21c, p. 28 Fig. 24a, b, c (upper figure), d.
? 1959 – Hemidiadema rugosum : , p. 95, Pl. XI, figs. 9-14.
1966 – Hemidiadema rugosum : & , p. U417, Fig. 312/1h.
? 1977 – Hemidiadema rugosum : , p. 185, 187, 195, 199, Pl. II, figs. 6-7.
non 1991 – Hemidiadema rugosum : , p. 213, Pl. 11, figs. 4-6 = Hemidiadema batalleri nov. sp.
? 1996 – Hemidiadema rugosum : , p. 181, 185, Pl. I, fig. 1, 1 a, 1b.
non 2000 – Hemidiadema rugosum : & , p. 18, Pl. III, figs. 23-24 = Hemidiadema forcallensis nov. sp.
non 2003 – Hemidiadema rugosum : , p. 76, Pl. 2, figs. 14-16 = Hemidiadema forcallensis nov. sp.
non 2004 – Hemidiadema rugosum : et al., p. 600, Fig. 2/7a-c, Fig. 6 = Hemidiadema sp.
2006 – Hemidiadema rugosum : & , p. 39, 70, Fig. 22.
non 2007 – Hemidiadema rugosum : et al., photo nº 25 = Hemidiadema forcallensis nov. sp.
non 2008 – Glyphocyphus rugosum : et al., p. 159 = Hemidiadema forcallensis nov. sp.
non 2008 – Hemidiadema rugosa : , p. 6, 21, photos 35-36 = Hemidiadema forcallensis nov. sp.
2010 – Hemidiadema rugosum : & , p. 221-224, 5 Figs. (non numérotées).
2012 – Hemidiadema rugosum : et al., p. 2-3, Fig. 3.1.
2013 – Hemidiadema
rugosum : , p.
10-14, Fig. 8,
Pls. 1 ![]() - 2
- 2 ![]() - 3
- 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() - 9
- 9 ![]() ;
Pl. 13
;
Pl. 13 ![]() , figs. 1-2, 4, 6, 8, 10 ;
Pl. 14
, figs. 1-2, 4, 6, 8, 10 ;
Pl. 14 ![]() , figs. 1-5 ;
Pl. 15
, figs. 1-5 ;
Pl. 15 ![]() .
.
Matériel étudié. Un seul spécimen (coll. ) provenant du sommet de l'Aptien supérieur de Loma de Casa lo Bolini, dépression de) l'Alcoraia (province d'Alicante).
Conservation. Individu complet, trčs légčrement comprimé latéralement. Un fragment de test manque sur la face supérieure. La surface du test présente une trčs bonne conservation. La face inférieure, en particulier dans la région centrale péristomiale, est encroűtée de gangue.
Mesures et comptages
| Échantillon | Test | Ambulacres | Interambulacres | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. |
Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| Coll. | 7,9 | 5,6 | - | 1,81 | 5 visibles, probable- ment 6 |
5 ŕ 6 | 11 visibles, probable- ment 12 |
2,89 | 2,10 | - |
Nb. pla. : nombre
de plaques par série.
Nb
tub. : nombre de
tubercules primaires par série.
Nb
tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en
dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
Description. Cette espčce ayant fait l'objet d'une étude récente (, 2013), l'étude descriptive de l'exemplaire étudié dans le présent travail sera réduite ŕ une comparaison avec les individus provenant de la localité type de l'espčce (Grandpré, Ardennes, France).
L'individu étudié est tout ŕ fait conforme aux spécimens rencontrés dans les Grčs verts ferrugineux de Grandpré : nombre de tubercules ambulacraires et interambulacraires supérieur ŕ 10 sur chaque série, aires scrobiculaires étroites, zones miliaires interradiales assez bien développées garnies d'assez nombreux et denses granules, granules scrobiculaires difficilement distinguables des granules miliaires, présence de tubercules secondaires microscopiques aux angles des majeures interambulacraires infra-ambitales.
Cet individu se distingue des spécimens de Grandpré par une morphologie plus globuleuse, des zones miliaires interradiales un peu plus larges et par ses tubercules secondaires plus petits et plus discrets. Il ne s'agit que de légčres variations morphologiques intraspécifiques liées sans doute aux conditions paléoenvironnementales.
Hemidiadema batalleri nov. sp.
(Pls. 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7 ; Pl. 15
- 7 ; Pl. 15 ![]() , figs. 6, 8)
, figs. 6, 8)
1991 – Hemidiadema rugosum : , p. 213, Pl. 11, figs. 4-6.
2012 – Hemidiadema nov. sp. 2 : et al., p. 2, Fig. 3.2.
Matériel étudié. Parmi le matériel connu seuls les 6 spécimens les mieux préservés ont été étudiés : MGSB 16823-1, MGSB 17271-2, MGSB 17271-3, MGSB 25112-1, MGSB 25112-2 (coll. ) et MGSB 79924 (coll. ). Tous ces spécimens proviennent du sommet de l'Aptien inférieur (Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata) de Cala del Moro, prčs de Punta de l'Ŕliga, au Sud du village d'El Perelló (province de Tarragone).
Holotype.
MGSB 79924 (Pls. 3 ![]() - 4
- 4 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 1-2, 5-6).
, figs. 1-2, 5-6).
Paratypes. MGSB 25112-1 (Pl. 5 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 3, 7, 9-10 ;
Pl. 15
, figs. 3, 7, 9-10 ;
Pl. 15 ![]() ,
fig. 6), MGSB 25112-2 (Pl. 6
,
fig. 6), MGSB 25112-2 (Pl. 6 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 4, 8).
, figs. 4, 8).
Derivatio nominis. Espčce dédiée ŕ l'Abbé Josep Ramon (1890-1962), ancien directeur du Musée géologique du Séminaire de Barcelone et professeur de Paléontologie ŕ l'Université de Barcelone.
Locus typicus. Cala del Moro, prčs de Punta de l'Ŕliga, au Sud du village d'El Perelló (province de Tarragone).
Stratum typicum. Marnes pyriteuses ŕ Plicatules datées de l'Aptien inférieur, partie inférieure de la Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata.
Mesures et comptages
| Échantillons | Test | Ambulacre | Interambulacre | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. | Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| MGSB 79924 | 10,10 | 6,38 | 30 | 2,10 | 5 | 0 ŕ 4 | 9 | 3,61 | 3,42 | 3,06 |
| MGSB 25112-1 | 10,28 | 6,61 | 27 | 2,52 | 4 ŕ 5 | 0 | 8 ŕ 9 | 3,89 | 3,72 | 3,24 |
| MGSB 25112-2 | 11,33 | 6,20 | 27 | 2,48 | 4 | 4 | 8 ŕ 9 | 4,38 | 3,96 | 3,52 |
| MGSB 16823-1 | 12,50 | 6,55 | 30 / 31 | 2,90 | 5 | 4 ŕ 5 | 9 | 4,41 | 3,74 | 3,39 |
| MGSB 17271-2 | 11,34 | 6,67 | - | 2,60 | 4 | 2 ŕ 3 | 9 | 4,13 | - | 3,25 |
| MGSB 17271-3 | 13,22 | - | - | 2,60 | 4 | 5 ŕ 6 | 9 ŕ 10 | 4,61 | - | - |
| Moyennes | 11,46 | 6,48 | 2,53 | 4,17 | 3,71 | 3,29 | ||||
Nb. pla. : nombre de plaques par série.
Nb tub. : nombre de
tubercules primaires par série.
Nb tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
Diagnose :
des granules scrobiculaires de taille identique, organisés en un cercle autour des aires scrobiculaires des tubercules des zones ambitales et infra-ambitales ;
les plaques des zones porifčres dont la surface est nettement granuleuse par le développement de nombreuses verrues ;
l'absence de zones miliaires adradiales, interradiales et perradiales ;
un disque apical uniformément étendu, nettement relevé sur son pourtour interne ;
une aire périproctale circulaire ŕ sub-circulaire.
Description
de l'holotype MGSB 79924 (Pls. 3 ![]() - 4
- 4 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 1-2, 5-6) :
, figs. 1-2, 5-6) :
Conservation. Individu presque complet (il manque l'ambulacre 5 et l'interambulacre IV), légčrement déformé latéralement. Le test est d'une excellente conservation. La surface du disque apical est en grande partie masquée par un dépôt d'oxyde ;
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Cônes bien développés, crénelés en leur sommet et supportant un mamelon perforé. Aires scrobiculaires étendues, ornées de côtes rayonnantes trčs prononcées. Leur extrémité distale supporte un granule bien marqué surmonté d'un petit mamelon, en particulier dans les zones ambitale et infra-ambitale. Ces granules, de taille homogčne et réguličrement espacés les uns des autres, s'organisent en un cercle entourant l'aire scrobiculaire. En dessous l'ambitus certains de ces granules scrobiculaires sont plus fortement développés et certains montrent une différenciation en tubercules secondaires, avec cône et mamelon perforé bien distincts. Ces tubercules secondaires de trčs petite taille s'observent sur la bordure adradiale des majeures. Dans la zone ambitale et infra-ambitale trois côtes parallčles verticales nettement plus saillantes que les côtes rayonnantes, alternant avec trois fossettes scrobiculaires, se développent en dessous le cône des tubercules primaires. En dessus l'ambitus, ces côtes sont bien moins marquées et peu distinguables des autres côtes rayonnantes scrobiculaires ;
Interambulacres (Pl. 4 ![]() , figs. 1-4). Aires
interambulacraires présentant un arrangement bisérié de 9 tubercules sur
chaque série. Les aires scrobiculaires sont tangentes ou faiblement confluents.
Dans ce dernier cas, il y a atténuation de la taille des granules
scrobiculaires ou disparition de ceux-ci entre deux tubercules consécutifs
d'une męme série. Zones miliaires adradiales inexistantes, les granules
scrobiculaires du côté adradial sont en contact direct avec la zone porifčre.
Zones miliaires interradiales inexistantes, les deux séries de tubercules étant
séparées en dessus l'ambitus par un étroit sillon médian. Quelques trčs
rares granules viennent trčs localement se greffer aux cercles de granules
scrobiculaires au niveau de l'interradius, en dessus de l'ambitus ;
, figs. 1-4). Aires
interambulacraires présentant un arrangement bisérié de 9 tubercules sur
chaque série. Les aires scrobiculaires sont tangentes ou faiblement confluents.
Dans ce dernier cas, il y a atténuation de la taille des granules
scrobiculaires ou disparition de ceux-ci entre deux tubercules consécutifs
d'une męme série. Zones miliaires adradiales inexistantes, les granules
scrobiculaires du côté adradial sont en contact direct avec la zone porifčre.
Zones miliaires interradiales inexistantes, les deux séries de tubercules étant
séparées en dessus l'ambitus par un étroit sillon médian. Quelques trčs
rares granules viennent trčs localement se greffer aux cercles de granules
scrobiculaires au niveau de l'interradius, en dessus de l'ambitus ;
Ambulacres
(Pl. 4 ![]() , figs. 5-6 ;
Pl. 7
, figs. 5-6 ;
Pl. 7 ![]() , figs. 1-2, 5-6). Aires
ambulacraires droites, composées de majeures trisociées ŕ l'exception de la
zone infra-apicale oů elles sont vraisemblablement bisociées. Zones interporifčres. De l'ambitus en se dirigeant vers le péristome s'observe un
arrangement unisérié de 5 tubercules. Ces 5 tubercules sont presque
parfaitement alignés verticalement les uns en dessous les autres, montrant
parfois une légčre disposition alterne (cela dépend de l'ambulacre
considéré).
Chaque tubercule occupe toute la largeur de la zone interporifčre et est généralement
tronqué sur sa bordure adradiale, l'aire srobiculaire étant en contact
direct avec la zone porifčre. Absence de zones miliaires adradiale et
perradiale. La zone
interporifčre supra-ambitale, un peu rétrécie, se caractérise, au niveau de
trois ambulacres (sur quatre préservés) par l'absence de tubercules. Au
niveau des ambulacres II, III et V les tubercules sont remplacés par un granule
émoussé d'aspect et de taille similaire aux granules voisins qui les côtoient
(Pl. 7
, figs. 1-2, 5-6). Aires
ambulacraires droites, composées de majeures trisociées ŕ l'exception de la
zone infra-apicale oů elles sont vraisemblablement bisociées. Zones interporifčres. De l'ambitus en se dirigeant vers le péristome s'observe un
arrangement unisérié de 5 tubercules. Ces 5 tubercules sont presque
parfaitement alignés verticalement les uns en dessous les autres, montrant
parfois une légčre disposition alterne (cela dépend de l'ambulacre
considéré).
Chaque tubercule occupe toute la largeur de la zone interporifčre et est généralement
tronqué sur sa bordure adradiale, l'aire srobiculaire étant en contact
direct avec la zone porifčre. Absence de zones miliaires adradiale et
perradiale. La zone
interporifčre supra-ambitale, un peu rétrécie, se caractérise, au niveau de
trois ambulacres (sur quatre préservés) par l'absence de tubercules. Au
niveau des ambulacres II, III et V les tubercules sont remplacés par un granule
émoussé d'aspect et de taille similaire aux granules voisins qui les côtoient
(Pl. 7 ![]() , figs. 2, 6). Les deux étroits demi-ambulacres recouverts de ces
granules sont séparés par un sillon médian perradial irrégulier.
L'ambulacre I, au contraire, présente des tubercules primaires normalement
développés.
En suivant la disposition des tubercules en partant de l'apex et en se
dirigeant vers le péristome (ce sens est choisi –
et non le sens inverse correspondant ŕ l'ordre d'apparition des tubercules
au cours de la croissance –
pour des raisons de facilité) on s'aperçoit que sur les quatre tubercules
primaires supra-ambitaux théoriques le tubercule 4 est absent, les tubercules
1, 2 et 3 étant normalement développés (Pl. 7
, figs. 2, 6). Les deux étroits demi-ambulacres recouverts de ces
granules sont séparés par un sillon médian perradial irrégulier.
L'ambulacre I, au contraire, présente des tubercules primaires normalement
développés.
En suivant la disposition des tubercules en partant de l'apex et en se
dirigeant vers le péristome (ce sens est choisi –
et non le sens inverse correspondant ŕ l'ordre d'apparition des tubercules
au cours de la croissance –
pour des raisons de facilité) on s'aperçoit que sur les quatre tubercules
primaires supra-ambitaux théoriques le tubercule 4 est absent, les tubercules
1, 2 et 3 étant normalement développés (Pl. 7 ![]() , figs. 1, 5).
Zones
porifčres.
Pores circulaires de grande taille, organisés en paire ŕ disposition oblique,
les deux pores de chaque paire étant trčs rapprochés. Surface des plaques
plane ŕ légčrement bombée (pores ŕ fleur de test), trčs granuleuse par le
développement de nombreuses verrues, ces verrues étant les plus développées
et grandes sur la bordure adapicale (parfaitement visible entre autre sur le
paratype MGSB 25112-1 :
Pl. 7
, figs. 1, 5).
Zones
porifčres.
Pores circulaires de grande taille, organisés en paire ŕ disposition oblique,
les deux pores de chaque paire étant trčs rapprochés. Surface des plaques
plane ŕ légčrement bombée (pores ŕ fleur de test), trčs granuleuse par le
développement de nombreuses verrues, ces verrues étant les plus développées
et grandes sur la bordure adapicale (parfaitement visible entre autre sur le
paratype MGSB 25112-1 :
Pl. 7 ![]() , figs. 9-10). Un petit granule, surmonté d'un minuscule
mamelon,
s'observe sur la bordure adradiale des zones ambitale et infra-ambitale, en
dessus le pore externe ;
, figs. 9-10). Un petit granule, surmonté d'un minuscule
mamelon,
s'observe sur la bordure adradiale des zones ambitale et infra-ambitale, en
dessus le pore externe ;
Fossettes suturales. Des
fossettes scrobiculaires, au nombre de trois, sont présentes sur la bordure
adorale des majeures, en dessous le cône des tubercules. Elles alternent avec
les côtes parallčles verticales saillantes qu'elles mettent ainsi en relief.
En dessus de l'ambitus, les fossettes scrobiculaires internes (interradiales)
s'étendent parfois latéralement par fusion avec des fossettes angulaires peu
développées pour atteindre l'interradius (visible sur la
Pl. 4 ![]() , fig. 1) ;
, fig. 1) ;
Disque apical. Disque apical uniformément étendu, trčs redressé sur son pourtour interne. Un dépôt d'oxyde ne permet pas de décrire en détail les plaques génitales et ocellaires qui le composent. Nous pouvons simplement constater que la taille des différentes plaques est homogčne et qu'une des plaques ocellaires, en partie observable, montre quelques granules (quatre ŕ cinq) ŕ sa surface.
Description
des paratypes MGSB 25112-1 (Pl. 5 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 3, 7, 9-10 ;
Pl. 15
, figs. 3, 7, 9-10 ;
Pl. 15 ![]() , fig. 6) et MGSB 25112-2
(Pl. 6
, fig. 6) et MGSB 25112-2
(Pl. 6 ![]() ;
Pl. 7
;
Pl. 7 ![]() , figs. 4, 8) :
, figs. 4, 8) :
L'étude des deux paratypes permet de compléter la description de l'holotype – notamment concernant le disque apical – et de mettre en évidence certaines variations morphologiques intra-spécifiques au niveau des zones interporifčres :
Zones interporifčres de MGSB 25112-2 (Pl. 7 ![]() , figs. 4, 8). Zones
interporifčres caractérisées par un arrangement unisérié de 8 ŕ 9
tubercules par série. Les tubercules ambitaux et infra-ambitaux, trčs rapprochés
et disposés presque parfaitement verticalement les uns au-dessus
les autres, occupent la totalité de la largeur de la zone interporifčre. La
plupart de ces tubercules sont tronqués sur leur côté adradial, l'aire
scrobiculaire ou la base du cône étant directement en contact avec la zone
porifčre. En dessus l'ambitus les tubercules primaires, au nombre de 4, se
caractérisent par leur réduction progressive de taille en se dirigeant vers
l'apex. Ces tubercules sont excentrés vers les bordures adradiales et leur
disposition est nettement alterne. Quelques granules gravitent autour d'eux.
Zones miliaires adradiales et perradiales inexistantes. Les deux demi-ambulacres
sont séparés par un sillon perradial ;
, figs. 4, 8). Zones
interporifčres caractérisées par un arrangement unisérié de 8 ŕ 9
tubercules par série. Les tubercules ambitaux et infra-ambitaux, trčs rapprochés
et disposés presque parfaitement verticalement les uns au-dessus
les autres, occupent la totalité de la largeur de la zone interporifčre. La
plupart de ces tubercules sont tronqués sur leur côté adradial, l'aire
scrobiculaire ou la base du cône étant directement en contact avec la zone
porifčre. En dessus l'ambitus les tubercules primaires, au nombre de 4, se
caractérisent par leur réduction progressive de taille en se dirigeant vers
l'apex. Ces tubercules sont excentrés vers les bordures adradiales et leur
disposition est nettement alterne. Quelques granules gravitent autour d'eux.
Zones miliaires adradiales et perradiales inexistantes. Les deux demi-ambulacres
sont séparés par un sillon perradial ;
Zones interporifčres de MGSB 25112-1 (Pl. 7 ![]() , figs. 3, 7, 9). La
tuberculation des zones interporifčres en dessus et en dessous l'ambitus étant
sensiblement différente, nous les décrivons successivement. Zone
ambitale et infra-ambitale. L'aspect
et la disposition des tubercules infra-ambitaux et ambitaux sont les męmes que
chez MGSB 25112-2.
Zone
supra-ambitale.
En dessus l'ambitus, immédiatement au-dessus
du gros tubercule ambital, la zone interporifčre se rétrécit et se caractérise
par l'absence totale de tubercules primaires au niveau des cinq ambulacres.
L'étroite zone interporifčre de chaque demi-ambulacre est recouverte de
granules émoussés, serrés entre eux et plus ou moins fusionnés, formant deux
étroites bandes granuleuses grossičres accolées aux zones porifčres
adjacentes et séparés par un sillon médian perradial ;
, figs. 3, 7, 9). La
tuberculation des zones interporifčres en dessus et en dessous l'ambitus étant
sensiblement différente, nous les décrivons successivement. Zone
ambitale et infra-ambitale. L'aspect
et la disposition des tubercules infra-ambitaux et ambitaux sont les męmes que
chez MGSB 25112-2.
Zone
supra-ambitale.
En dessus l'ambitus, immédiatement au-dessus
du gros tubercule ambital, la zone interporifčre se rétrécit et se caractérise
par l'absence totale de tubercules primaires au niveau des cinq ambulacres.
L'étroite zone interporifčre de chaque demi-ambulacre est recouverte de
granules émoussés, serrés entre eux et plus ou moins fusionnés, formant deux
étroites bandes granuleuses grossičres accolées aux zones porifčres
adjacentes et séparés par un sillon médian perradial ;
Disque apical (Pl. 5 ![]() , fig. 8 ;
Pl. 15
, fig. 8 ;
Pl. 15 ![]() , fig. 6). Disque apical bien développé,
uniformément étendu, caractérisé
par son pourtour interne trčs relevé. Son contour interne, et donc celui de
l'aire périproctale, est circulaire. Plaques génitales.
Les cinq plaques génitales sont de forme identique, un peu plus
longue que large. La madréporite est un peu plus développée.
Gonopores circulaires et d'assez grande taille, un peu excentrés vers la
bordure externe, au-dessus
desquels se développe un net granule situé ŕ mi-distance entre le gonopore et
la bordure externe de la plaque. Ce granule est parfois décalé latéralement
et non situé ŕ la verticale du gonopore. Plaques ocellaires.
De formes identiques, les cinq plaques ocellaires présentent un pore circulaire
bien développé, situé ŕ l'extrémité de chaque plaque, sans toutefois
s'y ouvrir latéralement. Présence d'un granule au-dessus du pore et en dessous, ce
dernier étant développé sur la bordure externe de la plaque. La bordure
interne des plaques est granuleuse et présente trois ŕ quatre granules alignés,
nettement plus distincts ;
, fig. 6). Disque apical bien développé,
uniformément étendu, caractérisé
par son pourtour interne trčs relevé. Son contour interne, et donc celui de
l'aire périproctale, est circulaire. Plaques génitales.
Les cinq plaques génitales sont de forme identique, un peu plus
longue que large. La madréporite est un peu plus développée.
Gonopores circulaires et d'assez grande taille, un peu excentrés vers la
bordure externe, au-dessus
desquels se développe un net granule situé ŕ mi-distance entre le gonopore et
la bordure externe de la plaque. Ce granule est parfois décalé latéralement
et non situé ŕ la verticale du gonopore. Plaques ocellaires.
De formes identiques, les cinq plaques ocellaires présentent un pore circulaire
bien développé, situé ŕ l'extrémité de chaque plaque, sans toutefois
s'y ouvrir latéralement. Présence d'un granule au-dessus du pore et en dessous, ce
dernier étant développé sur la bordure externe de la plaque. La bordure
interne des plaques est granuleuse et présente trois ŕ quatre granules alignés,
nettement plus distincts ;
Périprocte. Aire périproctale circulaire ;
Péristome. Aire péristomiale circulaire.
Variations intra-spécifiques. La principale variation intra-spécifique correspond ŕ la présence ou l'absence de tout ou partie des tubercules primaires supra-ambitaux selon les exemplaires considérés.
Les variations intra-spécifiques se traduisent également par des variations de la hauteur du test. Les individus dont seule la partie infra-ambitale des ambulacres présente des tubercules ont une face supérieure plus élevée et plus bombée, ainsi que des zones porifčres un peu plus étroites en dessus l'ambitus (conséquence de l'abscence du développement de tubercules primaires supra-ambitaux) par rapport aux individus possédant des ambulacres tuberculés sur toute leur hauteur.
Hemidiadema malladai nov. sp.
(Pls. 8 ![]() - 9
- 9 ![]() ;
Pl. 15
;
Pl. 15 ![]() , fig. 5)
, fig. 5)
2012 – Hemidiadema nov. sp. 1 : et al., p. 3, Fig. 3.3.
2015 – Hemidiadema sp. 1 : & , Pl. 6, fig. 1a-e.
Matériel étudié. 15 spécimens, appartenant ŕ la collection C: MNHN.F. A50416, A53463, A53464, A53465, A53466, A53467, A53468, A53469, A53470, A53471, A53472, A53473, A53474, A53475, A53476. Tous ces spécimens proviennent des environs d'Allepuz.
Holotype : MNHN.F.
A50416 (Pls. 8 ![]() - 9
- 9 ![]() ; Pl. 15
; Pl. 15 ![]() , fig. 5).
, fig. 5).
Derivatio nominis. Espčce dédiée ŕ Lucas (1841-1921), géologue et paléontologue espagnol.
Locus typicus. Allepuz, province de Teruel.
Stratum typicum. Marnes calcaires trčs finement bioclastiques ŕ échinides et brachiopodes appartenant ŕ la formation des Marnes de Forcall, i.e., "Formación Margas de Forcall" ( et al., 1982, p. 286-288), datée de l'Aptien inférieur, probablement de la partie supérieure de la Zone ŕ Deshayesites forbesi et/ou de la partie basale de la Zone ŕ Deshayesites deshayesi.
Mesures et comptages
| Échantillons | Test | Ambulacre | Interambulacre | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. | Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| MNHN.F. A50416 | 9,86 | 4,73 | 27 | 2,02 | 5 | 4 ŕ 5 | 10 | 3,56 | 2,60 | 3,77 |
| MNHN.F. A53463 | 10,57 | 5,43 | 30 | 2,10 | 5 ŕ 6 | 4 ŕ 5 | 10 ŕ 11 | 3,89 | 3,34 | 3,56 |
| MNHN.F. A53464 | 7,74 | 4,32 | 23 | 1,55 | 5 | 4 | 8 ŕ 9 | 2,77 | 2,61 | 3,01 |
| MNHN.F. A53465 | 8,18 | 3,88 | 24 | 1,78 | 4 ŕ 5 | 3 ŕ 4 | 9 | 2,96 | 2,44 | 3,02 |
| MNHN.F. A53466 | 8,22 | 4,59 | 25 | 1,92 | 4 ŕ 5 | 3 ŕ 4 | 8 ŕ 9 | 2,84 | 2,60 | 3,00 |
| MNHN.F. A53467 | 6,85 | 2,99 | 22 | 1,35 | 4 | 4 | 8 | 2,19 | 1,75 | 2,63 |
| MNHN.F. A53468 | 7,48 | 4,09 | 24 | 1,52 | 4 ŕ 5 | 4 | 9 | 2,77 | 2,31 | 2,98 |
| MNHN.F. A53469 | 7,69 | 3,87 | 22 | 1,62 | 4 | 3 ŕ 4 | 8 ŕ 9 | 2,74 | 2,53 | 2,84 |
| MNHN.F. A53470 | 7,72 | - | 23 | 1,72 | 4 | 3 ŕ 4 | 8 ŕ 9 | 3,06 | 2,29 | - |
| MNHN.F. A53471 | 11,39 | 5,01 | 28 | 2,29 | 5 | 4 | 10 | 4,19 | 3,62 | 4,17 |
| MNHN.F. A53472 | 8,49 | 3,41 | 24 | 1,85 | 4 ŕ 5 | 3 ŕ 4 | 8 ŕ 9 | 2,94 | 2,10 | 2,97 |
| MNHN.F. A53473 | 7,50 | - | 22 | 1,64 | 4 | 4 ŕ 5 | 8 | 2,74 | 1,92 | - |
| MNHN.F. A53474 | 6,88 | 3,25 | 22 | 1,40 | 4 ŕ 5 | 3 ŕ 4 | 8 | 2,45 | 2,12 | 2,70 |
| MNHN.F. A53475 | 6,18 | 2,67 | 20 | 1,45 | 3 | 3 | 7 | 2,38 | 1,81 | 2,29 |
| MNHN.F. A53476 | 5,07 | 2,40 | 18 | 0,97 | 3 | 3 | 6 ŕ 7 | 1,86 | 1,60 | 2,06 |
| Moyennes | 7,96 | 3,91 | 1,68 | 2,88 | 2,36 | 2,98 | ||||
Nb. pla. : nombre
de plaques par série.
Nb
tub. : nombre de
tubercules primaires par série.
Nb
tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en
dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
Diagnose :
la présence de zones interradiales supra-ambitales garnies de granules miliaires épars ;
des fossettes suturales scrobicualaires et angulaires nettement développées, formant de profondes dépressions ŕ la surface du test ;
un disque apical dont la surface est relativement plane, ŕ fleur de test et dont la partie antérieure sub-triangulaire est plus étroite que sa partie postérieure au contour général arrondi ;
une aire périproctale pyriforme.
Description
de l'holotype MNHN.F. A50416 (Pls. 8 ![]() - 9
- 9 ![]() ; Pl. 15
; Pl. 15 ![]() , fig. 5) :
, fig. 5) :
Conservation. Individu complet, trčs légčrement déformé (léger écrasement dorso-ventral avec partie apicale un peu enfoncée), d'excellente conservation ;
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Cônes moyennement développés, crénelés en leur sommet, surmontés d'un net mamelon perforé. Aires scrobiculaires étendues, ornées de côtes rayonnantes nombreuses et bien marquées. En dessous le cône s'observent deux – parfois trois – côtes nettement plus saillantes, parallčles et verticales, dont le relief est accentué par le développement entre celles-ci de profondes fossettes scrobiculaires. Les aires scrobiculaires des tubercules de la région ambitale sont entourées par un cercle assez régulier de granules scrobiculaires de petite taille, généralement mamelonnés et réguličrement espacés. En dessus de l'ambitus ces granules sont plus épars et distribués autour des tubercules sans disposition circulaire évidente. En dessous de l'ambitus, du fait de la convergence des tubercules et de l'augmentation de la taille de ces granules, leur nombre diminue et ils ne s'observent que sur les bordures adradiale et interradiale des aires scrobiculaires. Certains de ces granules scrobiculaires infra-ambitaux, en particulier ceux situés aux angles des majeures, sont un plus développés ;
Interambulacres
(Pl. 9 ![]() , figs. 1, 5, 7). Aires interambulacraires
composées d'un arrangement bisérié de 10 tubercules sur chaque série,
tangents ŕ confluents. Zones miliaires interradiales inexistantes en dessous de
l'ambitus. En dessus de l'ambitus, une faible zone miliaire interradiale se
développe, recouverte sur sa bordure de quelques granules épars, identiques
aux granules scrobiculaires qu'ils côtoient, sa partie centrale en étant en
grande partie dépourvue du fait du développement de profondes dépressions
correspondant aux fossettes suturales. Au niveau et en dessous de l'ambitus,
un sillon interradial trčs étroit sépare les deux séries de tubercules.
Zones miliaires adradiales inexistantes sur toute la hauteur des interambulacres,
les granules scrobiculaires étant en contact direct avec la zone porifčre ;
, figs. 1, 5, 7). Aires interambulacraires
composées d'un arrangement bisérié de 10 tubercules sur chaque série,
tangents ŕ confluents. Zones miliaires interradiales inexistantes en dessous de
l'ambitus. En dessus de l'ambitus, une faible zone miliaire interradiale se
développe, recouverte sur sa bordure de quelques granules épars, identiques
aux granules scrobiculaires qu'ils côtoient, sa partie centrale en étant en
grande partie dépourvue du fait du développement de profondes dépressions
correspondant aux fossettes suturales. Au niveau et en dessous de l'ambitus,
un sillon interradial trčs étroit sépare les deux séries de tubercules.
Zones miliaires adradiales inexistantes sur toute la hauteur des interambulacres,
les granules scrobiculaires étant en contact direct avec la zone porifčre ;
Ambulacres
(Pl. 9 ![]() , figs. 2-4, 7). Aires
ambulacraires droites, composées de majeures trisociées, ŕ l'exception de
la zone infra-apicale oů elles deviennent vraisemblablement bisociées. Zones interporifčres.
Arrangement unisérié de 9 ŕ 10 tubercules. Les tubercules ambitaux sont les
plus gros. En se dirigeant vers le péristome leur taille diminue
progressivement alors qu'en se dirigeant vers l'apex, cette diminution est
plus brusque. En dessous l'ambitus (Pl. 9
, figs. 2-4, 7). Aires
ambulacraires droites, composées de majeures trisociées, ŕ l'exception de
la zone infra-apicale oů elles deviennent vraisemblablement bisociées. Zones interporifčres.
Arrangement unisérié de 9 ŕ 10 tubercules. Les tubercules ambitaux sont les
plus gros. En se dirigeant vers le péristome leur taille diminue
progressivement alors qu'en se dirigeant vers l'apex, cette diminution est
plus brusque. En dessous l'ambitus (Pl. 9 ![]() ,
fig. 7) les tubercules
occupent toute la largeur de la zone interporifčre et sont disposés alignés
verticalement les uns en dessus les autres. Ces tubercules ambitaux et infra-ambitaux
sont tronqués sur leur bordure adradiale, l'aire scrobiculaire venant en
contact direct avec la zone porifčre. Leur côté opposé est intčgre et les
granules scrobiculaires tangents ŕ la zone porifčre. En dessus de l'ambitus (Pl. 9
,
fig. 7) les tubercules
occupent toute la largeur de la zone interporifčre et sont disposés alignés
verticalement les uns en dessus les autres. Ces tubercules ambitaux et infra-ambitaux
sont tronqués sur leur bordure adradiale, l'aire scrobiculaire venant en
contact direct avec la zone porifčre. Leur côté opposé est intčgre et les
granules scrobiculaires tangents ŕ la zone porifčre. En dessus de l'ambitus (Pl. 9 ![]() , fig. 4), du fait de la réduction
de taille des tubercules, ceux-ci sont moins rapprochés et présentent une
disposition alterne plus prononcée. Les cônes de ces tubercules sont
directement accolés ŕ la zone porifčre. La majeure atuberculée associée ŕ
la majeure tuberculée de l'autre série est trčs étroite, recouverte de
quelques granules trčs serrés. Une dépression perradiale discontinue
s'observe plus ou moins bien selon l'ambulacre, résultant de
l'interconnexion de fossettes suturales.
Zones porifčres.
Zones porifčres relativement larges, composées de pores subcirculaires
d'assez grande taille, organisés en paire légčrement oblique. La surface
des plaques ambulacraires est finement granuleuse par le développement de
nombreuses verrues microscopiques. Un granule s'observe fréquemment sur l'angle
adradial des assules, au dessus du pore externe, dans la région ambitale et
infra-ambitale. La bordure adapicale "épaissie" – associée au
granule précédemment cité – tend ŕ former une sorte de petit bourrelet qui
individualise chaque paire de pores ainsi un peu enfoncée ;
, fig. 4), du fait de la réduction
de taille des tubercules, ceux-ci sont moins rapprochés et présentent une
disposition alterne plus prononcée. Les cônes de ces tubercules sont
directement accolés ŕ la zone porifčre. La majeure atuberculée associée ŕ
la majeure tuberculée de l'autre série est trčs étroite, recouverte de
quelques granules trčs serrés. Une dépression perradiale discontinue
s'observe plus ou moins bien selon l'ambulacre, résultant de
l'interconnexion de fossettes suturales.
Zones porifčres.
Zones porifčres relativement larges, composées de pores subcirculaires
d'assez grande taille, organisés en paire légčrement oblique. La surface
des plaques ambulacraires est finement granuleuse par le développement de
nombreuses verrues microscopiques. Un granule s'observe fréquemment sur l'angle
adradial des assules, au dessus du pore externe, dans la région ambitale et
infra-ambitale. La bordure adapicale "épaissie" – associée au
granule précédemment cité – tend ŕ former une sorte de petit bourrelet qui
individualise chaque paire de pores ainsi un peu enfoncée ;
Fossettes suturales (Pl. 8 ![]() , fig. 1). Fossettes suturales
trčs nettement développées au niveau des interambulacres. On distingue :
, fig. 1). Fossettes suturales
trčs nettement développées au niveau des interambulacres. On distingue :
des fossettes suturales scrobiculaires au nombre de trois, parfois quatre, se développant sur la bordure adorale des majeures, en dessous les cônes. Elles sont séparées par des côtes verticales trčs saillantes qu'elles mettent en relief. Les deux fossettes adradial et interradial sont généralement plus développées que la ou les deux fossettes centrales, généralement plus étroites ;
des fossettes suturales angulaires se développant sur l'angle interradial de la bordure adorale de la plupart des majeures supra-ambitales. Celles-ci sont absentes en dessous l'ambitus.
En dessus de l'ambitus, au niveau de la plupart des majeures interambulacraires, la fossette scrobiculaire interradiale et la fossette angulaire adjacente fusionnent, formant ainsi une profonde dépression scrobiculo-angulaire atteignant l'interradius. Le męme phénomčne s'observe entre une fossette scrobiculaire adradiale et sa fossette angulaire adjacente. Au niveau de l'ambitus, les fossettes suturales angulaires "punctiformes" restent indépendantes des fossettes suturales scrobiculaires. En dessous de l'ambitus, les fossettes suturales angulaires sont absentes. Seules les fossettes suturales scrobiculaires persistent, découpant profondément la base du cône et l'aire scrobiculaire en deux ŕ trois côtes étroites trčs saillantes.
Au niveau des ambulacres, en dessus de l'ambitus, ces fossettes sont moins évidentes. Il y a probablement des interconnexions entre fossettes scrobiculaires adradiales et fossettes angulaires adjacentes dessinant formant en partie une dépression perradiale discontinue, plus ou moins visibles selon l'ambulacre considéré ;
Disque apical
(Pl. 9 ![]() , fig. 6 ;
Pl. 15
, fig. 6 ;
Pl. 15 ![]() , fig. 5).
Disque apical ŕ surface relativement plane et ŕ fleur de test, recouvert de trčs
petites verrues éparses et de quelques granules. Sa partie antérieure sub-triangulaire
est plus étroite que sa partie postérieure au contour général arrondi. De ce
fait, son contour interne – et donc celui de l'aire périproctale – est
pyriforme.
Plaques génitales.
Plaques génitales 1, 2, 3 et 4 de forme et de taille identiques, aussi haute
que large. La plaque génitale 5 a un contour externe plus triangulaire et un
contour interne plus arqué. Gonopore circulaire de grande taille, excentré
vers la bordure externe de la plaque. Immédiatement au dessus du pore, sur sa
bordure męme, s'observe un net petit granule excentré, la plaque génitale 5
en étant dépourvu.
Plaques ocellaires.
Plaques ocellaires II, III et V de forme identique. Un petit pore s'ouvre latéralement
sur leur bordure. Leur surface est bosselée et présente généralement dans
leur partie centrale un groupement de 4 ŕ 5 granules. Plaques ocellaires I et
IV sont plus étroites, avec une bordure interne plus redressée, carénée ;
, fig. 5).
Disque apical ŕ surface relativement plane et ŕ fleur de test, recouvert de trčs
petites verrues éparses et de quelques granules. Sa partie antérieure sub-triangulaire
est plus étroite que sa partie postérieure au contour général arrondi. De ce
fait, son contour interne – et donc celui de l'aire périproctale – est
pyriforme.
Plaques génitales.
Plaques génitales 1, 2, 3 et 4 de forme et de taille identiques, aussi haute
que large. La plaque génitale 5 a un contour externe plus triangulaire et un
contour interne plus arqué. Gonopore circulaire de grande taille, excentré
vers la bordure externe de la plaque. Immédiatement au dessus du pore, sur sa
bordure męme, s'observe un net petit granule excentré, la plaque génitale 5
en étant dépourvu.
Plaques ocellaires.
Plaques ocellaires II, III et V de forme identique. Un petit pore s'ouvre latéralement
sur leur bordure. Leur surface est bosselée et présente généralement dans
leur partie centrale un groupement de 4 ŕ 5 granules. Plaques ocellaires I et
IV sont plus étroites, avec une bordure interne plus redressée, carénée ;
Périprocte. Aire périproctale allongée selon l'axe antéro-postérieur, pyriforme ;
Péristome. Aire péristomiale circulaire.
Variations intraspécifiques. Selon les individus les granules miliaires des zones miliaires ambitales et supra-ambitales sont plus ou moins denses et les fossettes suturales angulaires plus ou moins profondes, mais toujours nettement marquées. Il existe également un certain continuum dans la forme de l'aire périproctale, variant de pyriforme ŕ simplement ovale, selon les individus.
Hemidiadema forcallensis nov. sp.
(Pls. 10 ![]() - 11
- 11 ![]() ; Pl. 15
; Pl. 15 ![]() , fig. 7)
, fig. 7)
? 1928 – Hemidiadema rugosum : , p. 149.
2000 – Hemidiadema rugosum : & , p. 18, Pl. III, figs. 23-24.
2003 – Hemidiadema rugosum : , p. 76, Pl. 2, figs. 14-16.
2007 – Hemidiadema rugosum : et al., photo nº 25.
pars 2008 – Glyphocyphus rugosum : et al., p. 159.
2008 – Hemidiadema rugosa : , p. 6, 21 ; photos 35-36.
2012 – Hemidiadema nov. sp. 4 : et al. p. 3, Fig. 3.5.
2015 – Hemidiadema sp. : & , Pl. 5, fig. 1a-e.
Matériel étudié. Les 16 individus les mieux préservés ont été étudiés : MNHN.F. A50417, A53477, A53478, A53479, A53480, A53481, A53482, A53483, A53484, A53485, A53486, A53487, A53488, A53489 (coll. ) ; MGSB 79945, 79946 (coll. ). Tous ces individus proviennent des marnes pyriteuses datées de l'Aptien inférieur, partie supérieure de la Zone ŕ Deshayesites forbesi et de la partie basale de la Zone ŕ Deshayesites deshayesi des environs de Forcall.
Holotype. MNHN.F. A50417 (Pls. 10 ![]() - 11
- 11 ![]() ; Pl. 15
; Pl. 15 ![]() ,
fig. 7).
,
fig. 7).
Derivatio nominis. Fait référence ŕ la région de Forcall (province de Castellón) d'oů proviennent tous les spécimens étudiés.
Locus typicus. Mas del Roc, prčs de Forcall (province de Castellón).
Stratum typicum. Marnes pyriteuses de l'Aptien inférieur, i.e., "Formación Margas de Forcall" ( et al., 1982, p. 286-288), datées de la partie supérieure de la Zone ŕ Deshayesites forbesi et de la partie basale de la Zone ŕ Deshayesites deshayesi.
Mesures et comptages
| Échantillons | Test | Ambulacre | Interambulacre | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. | Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| MNHN.F. 50417 | 5,75 | 2,96 | 20 | 1,25 | 4 ŕ 5 | 2 | 7 | 2,00 | 2,47 | 1,95 |
| MNHN.F. A53477 | 5,33 | 2,60 | 19 | 1,17 | 4 | 1 ŕ 2 | 7 | 1,95 | - | 1,89 |
| MNHN.F. A53478 | 5,35 | 2,50 | 18 | 1,16 | 4 | 1 ŕ 2 | 7 | 1,79 | 1,83 | 1,78 |
| MNHN.F. A53479 | 6,23 | 3,05 | 21 | 1,36 | 4 ŕ 5 | 2 ŕ 3 | 7 ŕ 8 | 2,21 | 2,54 | 2,05 |
| MNHN.F. A53480 | 5,54 | 2,49 | - | 1,20 | 4 | 2 | 7 | 1,93 | 2,03 | - |
| MNHN.F. A53481 | 4,89 | 2,59 | 18 | 1,13 | 4 ŕ 5 | 2 | 7 | 1,76 | 2,18 | 1,75 |
| MNHN.F. A53482 | 4,73 | 2,30 | - | 1,01 | 4 | 1 ŕ 2 | 6 ŕ 7 | 1,63 | 1,73 | - |
| MNHN.F. A53483 | 5,61 | 2,52 | - | 1,03 | 4 | 1 ŕ 2 | 7 | 1,96 | - | 1,97 |
| MNHN.F. A53484 | 4,38 | 2,27 | 17 | 0,85 | 4 | 2 | 7 | 1,48 | 1,62 | 1,55 |
| MNHN.F. A53485 | 4,11 | 1,92 | 15 / 16 | 0,83 | 4 | 1 | 6 | 1,42 | 1,84 | 1,57 |
| MNHN.F. A53486 | 5,39 | 2,40 | 19 | 1,21 | 4 ŕ 5 | 1 ŕ 2 | 7 | 1,86 | 2,06 | 1,80 |
| MNHN.F. A53487 | 5,90 | 2,93 | 20 | 1,27 | 4 | 2 | 7 | 2,00 | 2,09 | 2,13 |
| MNHN.F. A53488 | 6,54 | 3,06 | 21 | 1,32 | 4 ŕ 5 | 2 | 7 | 2,35 | 2,44 | 2,03 |
| MNHN.F. A53489 | 5,24 | 2,21 | - | 1,23 | 4 | 2 | 7 | 1,76 | 1,93 | 1,72 |
| MGSB 79945 | 6,03 | 3,25 | 19 | 1,37 | 4 | 2 ŕ 3 | 7 ŕ 8 | 2,03 | 2,20 | 2,00 |
| MGSB 79946 | 5,94 | 2,95 | 18 | 1,26 | 4 | 2 | 7 | 1,90 | 2,09 | 1,98 |
| Moyennes | 5,37 | 2,62 | 1,18 | 1,88 | 2,07 | 1,87 | ||||
Nb. pla. : nombre
de plaques par série.
Nb
tub. : nombre de
tubercules primaires par série.
Nb
tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en
dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
Diagnose :
sa trčs petite taille, dépassant rarement 6 mm de diamčtre ;
un nombre de tubercules ambulacraires et interambulacraires supra-ambitaux par série trčs faible (1 ŕ 2, rarement 3) ;
le pourtour interne du disque apical est redressé ;
une aire périproctale trčs étendue.
Description
de l'holotype MNHN.F A50417 (Pls. 10 ![]() - 11
- 11 ![]() ; Pl. 15
; Pl. 15 ![]() , fig. 7) :
, fig. 7) :
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Le sommet des cônes est nettement crénelé, ornés de 7 ŕ 8 crénelures fortement développées. Un mamelon arrondi et perforé surmonte le cône. Les aires scrobiculaires sont moyennement développées, elles le sont le plus ŕ l'ambitus. L'aire scrobiculaire est parcourue de côtes rayonnantes épaisses et marquées, principalement dans la zone ambitale. Ces côtes rayonnantes se terminent ŕ leur extrémité distale par un granule. L'ensemble de ces granules, de taille assez homogčne, sont réguličrement espacés les uns des autres et montrent une disposition circulaire autour de l'aire scrobiculaire. Les granules scrobiculaires situés aux angles adradiaux et interradiaux inférieurs et supérieurs de la plupart des majeures ambitales et sous-ambitales sont un peu plus gros et surmontés d'un petit mamelon. En dessus l'ambitus, les granules scrobiculaires sont plus épars. On en compte seulement quelques-uns autour du cône, sans former de cercle. A l'aplomb des cônes, deux ŕ trois côtes, plus étroites, verticales et parallčles entre elles séparent des fossettes suturales scrobiculaires ;
Interambulacres (Pl. 10 ![]() , figs. 1-2, 9-11 ;
Pl. 11
, figs. 1-2, 9-11 ;
Pl. 11 ![]() , figs. 2-3).
Aires interambulacraires composées d'un arrangement bisérié de 6 ŕ 8
tubercules primaires sur chaque série, 7 en moyenne. En dessus l'ambitus, le
nombre de tubercules par série est faible, généralement au nombre de 2. Les
tubercules sont tangents ŕ légčrement confluents, avec disparition ou réduction
de taille des granules scrobiculaires situés entre deux tubercules consécutifs
d'une męme série. Aires miliaires adradiales et interradiales inexistantes,
ŕ l'exception de la région supra-ambitale oů une trčs étroite aire
miliaire interradiale se développe parfois, essentiellement par disparition des
granules scrobiculaires autour des tubercules. Celle-ci est dépourvue de
granules miliaires. Quelques rares granules s'insčrent parfois au niveau de
l'interradius ou s'intercalent entre le côté adradial des tubercules et la
zone porifčre. Les deux demi-ambulacres sont généralement séparés par un trčs
étroit sillon interradial, visible au dessus de l'ambitus uniquement ;
, figs. 2-3).
Aires interambulacraires composées d'un arrangement bisérié de 6 ŕ 8
tubercules primaires sur chaque série, 7 en moyenne. En dessus l'ambitus, le
nombre de tubercules par série est faible, généralement au nombre de 2. Les
tubercules sont tangents ŕ légčrement confluents, avec disparition ou réduction
de taille des granules scrobiculaires situés entre deux tubercules consécutifs
d'une męme série. Aires miliaires adradiales et interradiales inexistantes,
ŕ l'exception de la région supra-ambitale oů une trčs étroite aire
miliaire interradiale se développe parfois, essentiellement par disparition des
granules scrobiculaires autour des tubercules. Celle-ci est dépourvue de
granules miliaires. Quelques rares granules s'insčrent parfois au niveau de
l'interradius ou s'intercalent entre le côté adradial des tubercules et la
zone porifčre. Les deux demi-ambulacres sont généralement séparés par un trčs
étroit sillon interradial, visible au dessus de l'ambitus uniquement ;
Ambulacres
(Pl. 10 ![]() , figs. 3-6 ;
Pl. 11
, figs. 3-6 ;
Pl. 11 ![]() , fig. 4). Aires
ambulacraires droites, composées de majeures trisociées, ŕ l'exception de
la zone infra-apicale oů elles deviennent vraisemblablement bisociées. Zones interporifčres.
Les zones interporifčres sont ornées d'un arrangement unisérié de 6 ŕ 7
tubercules, avec généralement seulement 1 ŕ 2 tubercules en dessus
l'ambitus, rarement plus (3 au maximum). En se dirigeant de l'ambitus vers
le péristome, la taille des tubercules diminue progressivement. Ces tubercules
sont presque parfaitement disposés verticalement les uns au dessus des autres.
Ils occupent toute la largeur de la zone interporifčre et sont tronqués dans
leur partie adradiale, l'aire scrobiculaire venant en contact avec la zone
porifčre. En dessus l'ambitus, la taille des tubercules se réduit plus
brusquement. Ces tubercules supra-ambitaux montrent généralement une
disposition alterne. Zones porifčres.
Les zones porifčres sont composées de paires de pores circulaires ŕ sub-circulaires
de grande taille, assez rapprochés ;
, fig. 4). Aires
ambulacraires droites, composées de majeures trisociées, ŕ l'exception de
la zone infra-apicale oů elles deviennent vraisemblablement bisociées. Zones interporifčres.
Les zones interporifčres sont ornées d'un arrangement unisérié de 6 ŕ 7
tubercules, avec généralement seulement 1 ŕ 2 tubercules en dessus
l'ambitus, rarement plus (3 au maximum). En se dirigeant de l'ambitus vers
le péristome, la taille des tubercules diminue progressivement. Ces tubercules
sont presque parfaitement disposés verticalement les uns au dessus des autres.
Ils occupent toute la largeur de la zone interporifčre et sont tronqués dans
leur partie adradiale, l'aire scrobiculaire venant en contact avec la zone
porifčre. En dessus l'ambitus, la taille des tubercules se réduit plus
brusquement. Ces tubercules supra-ambitaux montrent généralement une
disposition alterne. Zones porifčres.
Les zones porifčres sont composées de paires de pores circulaires ŕ sub-circulaires
de grande taille, assez rapprochés ;
Fossettes suturales. Les seules fossettes suturales observables sont les fossettes suturales scrobiculaires. Celles-ci, trčs étroites, se développent sur la bordure adorale des majeures en dessous les cônes des tubercules ambitaux et infra-ambitaux et sont séparées par 2 ŕ 3 côtes verticales saillantes ;
Disque apical (Pl. 11 ![]() , fig. 1 ;
Pl. 15
, fig. 1 ;
Pl. 15 ![]() , fig. 7).
Disque apical plus étroit dans sa partie antérieure que dans sa partie postérieure
et dont le pourtour interne est redressé. La surface des plaques du disque
apicale est granuleuse par le développement de nombreuses petites verrues. Plaques génitales.
Les plaques génitales 1, 2, 3 et 4 ont une forme identique. Un granule
s'observe au dessus du gonopore. La plaque génitale 5 est plus étroite, sa
bordure interne plus relevée et saillante. Plaques ocellaires.
Les plaques ocellaires II, III et V ont une forme identique, recouvertes par
deux ŕ quatre granules. Les plaques I et IV sont trčs étroites, leur bordure
interne étant plus relevée et saillante. Elle présente également trois ŕ
quatre granules alignés dans leur partie médiane ;
, fig. 7).
Disque apical plus étroit dans sa partie antérieure que dans sa partie postérieure
et dont le pourtour interne est redressé. La surface des plaques du disque
apicale est granuleuse par le développement de nombreuses petites verrues. Plaques génitales.
Les plaques génitales 1, 2, 3 et 4 ont une forme identique. Un granule
s'observe au dessus du gonopore. La plaque génitale 5 est plus étroite, sa
bordure interne plus relevée et saillante. Plaques ocellaires.
Les plaques ocellaires II, III et V ont une forme identique, recouvertes par
deux ŕ quatre granules. Les plaques I et IV sont trčs étroites, leur bordure
interne étant plus relevée et saillante. Elle présente également trois ŕ
quatre granules alignés dans leur partie médiane ;
Périprocte. Aire périproctale trčs étendue, ovale ;
Péristome. Aire péristomiale circulaire.
Hemidiadema sp. A
(Pls. 12 ![]() - 13
- 13 ![]() - 14
- 14 ![]() ; Pl. 15
; Pl. 15 ![]() , fig. 8)
, fig. 8)
2012 – Hemidiadema nov. sp. 3 : et al., p. 3, Fig. 3.4.
Matériel étudié. Un unique individu : MNHN.F. A50418. Cet individu provient des marnes pyriteuses datées de l'Aptien inférieur, partie supérieure de la Zone ŕ Deshayesites forbesi et de la partie basale de la Zone ŕ Deshayesites deshayesi, de Mola d'en Camarŕs, prčs de Forcall (province de Castellón).
Mesures et comptages
| Échantillons | Test | Ambulacre | Interambulacre | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. | Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| MNHN.F. A50418 |
12,96 | - | - | 3,10 | 5 visibles, probable ment 6 ŕ 7 |
4 ŕ 5 | 8 visibles, probable ment 10 |
4,67 | 4,01 | - |
Nb. pla.: nombre
de plaques par série.
Nb
tub. : nombre de
tubercules primaires par série.
Nb
tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en
dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
Description
Conservation. Test complet, écrasé dorso-ventralement, d'excellente conservation ;
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Cônes lisses, bien développés, crénelés en son sommet, supportant un net mamelon muni d'un trčs petit pore. Aires scrobiculaires assez bien développées au niveau des tubercules ambitaux et infra-ambitaux, ornées de côtes rayonnantes épaisses bien marquées. En dessus l'ambitus, les aires scrobiculaires se réduisent trčs sensiblement. L'extrémité de chaque côte rayonnante supporte un granule surmonté d'un net mamelon, toujours bien visible. Ce mamelon est d'autant plus net que l'on se dirige en direction de la face orale. Certains de ces granules émergent par leur plus fort développement, principalement sur la bordure adradiale des scrobicules infra-ambitaux, oů certains tendent ŕ s'individualiser en minuscules tubercules secondaires prčs de la bordure péristomiale. En dessus l'ambitus ces granules, plus épars et moins rapprochés, se distribuent irréguličrement autour du cône sans organisation circulaire nette. A l'aplomb du cône des tubercules primaires ambitaux et infra-ambitaux s'observent généralement 3 ŕ 4 côtes parallčles verticales plus étroites et nettement plus saillantes séparant d'étroites fossettes suturales scrobiculaires. En dessus de l'ambitus, du fait de la réduction des aires scrobiculaires, ces côtes verticales sont bien moins observables, car trčs courtes, trčs rapprochées et moins saillantes ;
Interambulacres
(Pl. 12 ![]() , figs. 6-9 ;
Pl. 13
, figs. 6-9 ;
Pl. 13 ![]() , figs. 1-2 ;
Pl. 14
, figs. 1-2 ;
Pl. 14 ![]() , figs. 3-5).
Aires interambulacraires
composées d'un arrangement bisérié de probablement 10 tubercules primaires
sur chaque série (8 sont parfaitement visibles, deux autres supposés en
bordure péristomiale ne le sont pas ŕ cause de la gangue qui obstrue cette région
du test). En dessus de l'ambitus (Pl. 14
, figs. 3-5).
Aires interambulacraires
composées d'un arrangement bisérié de probablement 10 tubercules primaires
sur chaque série (8 sont parfaitement visibles, deux autres supposés en
bordure péristomiale ne le sont pas ŕ cause de la gangue qui obstrue cette région
du test). En dessus de l'ambitus (Pl. 14 ![]() ,
fig. 3) la taille des tubercules diminue progressivement en se
rapprochant de l'apex (s'accompagnant d'une réduction et disparition de
l'aire scrobiculaire), de męme en dessous de l'ambitus en se dirigeant vers
le péristome mais moins rapidement et les tubercules restant toujours nettement
scrobiculés. Les tubercules sont tangents ŕ légčrement confluents au niveau
de l'ambitus et sous l'ambitus, la taille des granules scrobiculaires entre
deux tubercules consécutifs d'une męme série s'atténue alors trčs
sensiblement. Dans la zone ambitale (Pl. 14
,
fig. 3) la taille des tubercules diminue progressivement en se
rapprochant de l'apex (s'accompagnant d'une réduction et disparition de
l'aire scrobiculaire), de męme en dessous de l'ambitus en se dirigeant vers
le péristome mais moins rapidement et les tubercules restant toujours nettement
scrobiculés. Les tubercules sont tangents ŕ légčrement confluents au niveau
de l'ambitus et sous l'ambitus, la taille des granules scrobiculaires entre
deux tubercules consécutifs d'une męme série s'atténue alors trčs
sensiblement. Dans la zone ambitale (Pl. 14 ![]() ,
fig. 5) et infra-ambitale (Pl. 14
,
fig. 5) et infra-ambitale (Pl. 14 ![]() , fig. 4) il y a absence
de zones miliaires interradiales, les deux séries de tubercules étant séparées par un étroit sillon
interradial lisse. Au dessus de l'ambitus se développe une zone miliaire
interradiale. Quelques granules miliaires épars côtoyant les granules
scrobiculaires s'y observent, mais la majeure partie de la surface des zones
miliaires interradiales est dépourvue de granules et sculptée de dépressions
elliptiques transversales correspondant aux fossettes suturales. Aucune zone
miliaire adradiale n'est observable sur toute la hauteur des interambulacres,
les granules scrobiculaires des tubercules étant en contact avec la zone
porifčre ;
, fig. 4) il y a absence
de zones miliaires interradiales, les deux séries de tubercules étant séparées par un étroit sillon
interradial lisse. Au dessus de l'ambitus se développe une zone miliaire
interradiale. Quelques granules miliaires épars côtoyant les granules
scrobiculaires s'y observent, mais la majeure partie de la surface des zones
miliaires interradiales est dépourvue de granules et sculptée de dépressions
elliptiques transversales correspondant aux fossettes suturales. Aucune zone
miliaire adradiale n'est observable sur toute la hauteur des interambulacres,
les granules scrobiculaires des tubercules étant en contact avec la zone
porifčre ;
Ambulacres (Pl. 12 ![]() , figs. 2-5
; Pl. 13
, figs. 2-5
; Pl. 13 ![]() , figs. 3-6 ;
Pl. 14
, figs. 3-6 ;
Pl. 14 ![]() , figs. 1-2).
Aires ambulacraires droites, relativement larges (largeur A / largeur IA = 0,62)
composées de majeures trisociées qui ŕ partir de la mi-hauteur de la partie
supra-ambitale et jusqu'ŕ l'apex deviennent vraisemblablement bisociées. Zones interporifčres.
Zones interporifčres composées d'un arrangement unisérié de probablement
10, voir 11 tubercules (8 sont parfaitement visibles, 2 ou 3 autres supposés en
bordure péristomiale ne le sont pas ŕ cause de la gangue qui obstrue cette région
du test). Au dessus de l'ambitus (Pl. 12
, figs. 1-2).
Aires ambulacraires droites, relativement larges (largeur A / largeur IA = 0,62)
composées de majeures trisociées qui ŕ partir de la mi-hauteur de la partie
supra-ambitale et jusqu'ŕ l'apex deviennent vraisemblablement bisociées. Zones interporifčres.
Zones interporifčres composées d'un arrangement unisérié de probablement
10, voir 11 tubercules (8 sont parfaitement visibles, 2 ou 3 autres supposés en
bordure péristomiale ne le sont pas ŕ cause de la gangue qui obstrue cette région
du test). Au dessus de l'ambitus (Pl. 12 ![]() , figs. 1-2) la
taille des tubercules se réduit assez brusquement puis progressivement, pour
devenir minuscules pour ceux situés immédiatement en dessous le disque apical.
La disposition de ces tubercules est trčs variable selon l'ambulacre
considéré : tubercules alignés verticalement ou en disposition alterne. Les
tubercules, qu'ils soient tous situés du męme côté ou montrant une
disposition alterne, sont tous accolés ŕ la zone porifčre, ce qui se traduit
par l'absence de zone miliaire adradiale. Les
majeures sans tubercules associées aux majeures tuberculées sont réduites ŕ
leur zone porifčre et ŕ une surface interporifčre
trčs restreinte, recouverte de quelques granules serrés (2 ŕ 3) les uns aux
autres. Dans la zone perradiale supra-ambitale, un sillon plus ou moins régulier
sépare les deux demi-ambulacres, localement élargi par le développement de
fossettes suturales angulaires. Dans la zone ambitale (Pl. 13
, figs. 1-2) la
taille des tubercules se réduit assez brusquement puis progressivement, pour
devenir minuscules pour ceux situés immédiatement en dessous le disque apical.
La disposition de ces tubercules est trčs variable selon l'ambulacre
considéré : tubercules alignés verticalement ou en disposition alterne. Les
tubercules, qu'ils soient tous situés du męme côté ou montrant une
disposition alterne, sont tous accolés ŕ la zone porifčre, ce qui se traduit
par l'absence de zone miliaire adradiale. Les
majeures sans tubercules associées aux majeures tuberculées sont réduites ŕ
leur zone porifčre et ŕ une surface interporifčre
trčs restreinte, recouverte de quelques granules serrés (2 ŕ 3) les uns aux
autres. Dans la zone perradiale supra-ambitale, un sillon plus ou moins régulier
sépare les deux demi-ambulacres, localement élargi par le développement de
fossettes suturales angulaires. Dans la zone ambitale (Pl. 13 ![]() , figs. 3-5)
et infra-ambitale (Pl. 13
, figs. 3-5)
et infra-ambitale (Pl. 13 ![]() , fig. 6)
les tubercules occupent toute la largeur de l'ambulacre, avec une disposition
verticale les uns en dessous les autres ou légčrement alternes, selon
l'ambulacre considéré. Les aires scrobiculaires sont pour la plupart tronquées dans
leur partie adradiale, celles-ci ou la base du cône venant en contact direct
avec la zone porifčre. Zones porifčres
(Pl. 14
, fig. 6)
les tubercules occupent toute la largeur de l'ambulacre, avec une disposition
verticale les uns en dessous les autres ou légčrement alternes, selon
l'ambulacre considéré. Les aires scrobiculaires sont pour la plupart tronquées dans
leur partie adradiale, celles-ci ou la base du cône venant en contact direct
avec la zone porifčre. Zones porifčres
(Pl. 14 ![]() , figs. 1-2,
6).
Zones porifčres larges, avec pores organisés en paires obliques. Les deux
pores de chaque paire sont de taille relativement petite et écartés. Les pores
internes (perradiaux) sont subcirculaires alors que les pores externes (adradiaux)
ont une forme généralement plus allongée et une disposition nettement plus
oblique. La surface des assules est plane ŕ légčrement bombée, densément
recouvertes de verrues microscopiques, en particulier au niveau des bordures
adapicales. Les pores sont ŕ fleur de test. Dans la zone ambitale et infra-ambitale,
en dessus le pore adradial, l'angle supérieur externe de l'assule supporte
généralement un granule ;
, figs. 1-2,
6).
Zones porifčres larges, avec pores organisés en paires obliques. Les deux
pores de chaque paire sont de taille relativement petite et écartés. Les pores
internes (perradiaux) sont subcirculaires alors que les pores externes (adradiaux)
ont une forme généralement plus allongée et une disposition nettement plus
oblique. La surface des assules est plane ŕ légčrement bombée, densément
recouvertes de verrues microscopiques, en particulier au niveau des bordures
adapicales. Les pores sont ŕ fleur de test. Dans la zone ambitale et infra-ambitale,
en dessus le pore adradial, l'angle supérieur externe de l'assule supporte
généralement un granule ;
Fossettes suturales. Dans la zone ambitale et infra-ambitale des aires interambulacraires et ambulacraires, des fossettes suturales scrobiculaires nettement marquées sont observables. Elles se développent sur la bordure adorale des majeures, en dessous chaque cône, au nombre de 3 ŕ 4, séparées par des côtes parallčles verticales saillantes. En dessus l'ambitus ces fossettes et les côtes saillantes associées les séparant sont nettement moins visibles de par la réduction sensible des aires scrobiculaires. On observe par contre sur la bordure adorale interradiale des majeures des dépressions semi-elliptiques peu profondes, sculptant la surface de la zone miliaire interradiale et atteignant l'interradius. Elles correspondent chacune vraisemblablement ŕ la fusion d'une fossette suturale angulaire avec la fossette suturale scrobiculaire interradiale adjacente. Du côté opposé (adradial), ces dépressions, moins étendues, sont également présentes, partant de la base du cône pour atteindre la bordure de la zone porifčre. Au niveau des ambulacres, les phénomčnes de fusion d'une fossette suturale angulaire avec la fossette suturale scrobiculaire perradiale adjacente sont difficilement identifiables ;
Disque apical
(Pl. 13 ![]() , fig. 10 ;
Pl. 15
, fig. 10 ;
Pl. 15 ![]() , fig. 8).
Disque apical ŕ surface plane, ŕ fleur de test. Dans sa partie postérieure le
disque est uniformément étendu et son contour général arrondi. Dans sa
partie antérieure le disque est beaucoup moins étendu du fait de la plus
grande étroitesse des plaques et son contour général est sub-triangulaire.
L'ensemble de sa surface est trčs finement et densément granuleuse par le développement
de trčs nombreuses verrues.
Plaques génitales.
Les plaques génitales 1, 3 et 4 ont une forme identique, percée en leur centre
d'un gonopore circulaire de grande taille, au dessus immédiat duquel
s'observe un net granule. La plaque génitale 2 (madréporite) est d'une
forme sensiblement identique aux plaques précédentes, un peu plus grande et
bombée. Son gonopore est beaucoup plus petit et excentré vers la bordure de la
plaque. La plaque génitale 5 se distingue des autres plaques génitales par sa
forme en "boomerang". Elle est percée en son centre d'un pore génital
mais aucun granule ne s'observe en arričre de celui-ci.
Plaques ocellaires.
Les plaques ocellaires II, III et IV ont une forme identique. Un pore de taille
moyenne s'observe ŕ l'extrémité de la plaque, encadré par deux granules,
l'un situé en arričre immédiat et le second ŕ l'opposé, sur le bord
externe de la plaque. Leur surface supporte quelques granules. Les plaques I et
V sont trčs étroites, leur bordure interne, relevée et saillante, présentant
trois ŕ quatre granules alignés ;
, fig. 8).
Disque apical ŕ surface plane, ŕ fleur de test. Dans sa partie postérieure le
disque est uniformément étendu et son contour général arrondi. Dans sa
partie antérieure le disque est beaucoup moins étendu du fait de la plus
grande étroitesse des plaques et son contour général est sub-triangulaire.
L'ensemble de sa surface est trčs finement et densément granuleuse par le développement
de trčs nombreuses verrues.
Plaques génitales.
Les plaques génitales 1, 3 et 4 ont une forme identique, percée en leur centre
d'un gonopore circulaire de grande taille, au dessus immédiat duquel
s'observe un net granule. La plaque génitale 2 (madréporite) est d'une
forme sensiblement identique aux plaques précédentes, un peu plus grande et
bombée. Son gonopore est beaucoup plus petit et excentré vers la bordure de la
plaque. La plaque génitale 5 se distingue des autres plaques génitales par sa
forme en "boomerang". Elle est percée en son centre d'un pore génital
mais aucun granule ne s'observe en arričre de celui-ci.
Plaques ocellaires.
Les plaques ocellaires II, III et IV ont une forme identique. Un pore de taille
moyenne s'observe ŕ l'extrémité de la plaque, encadré par deux granules,
l'un situé en arričre immédiat et le second ŕ l'opposé, sur le bord
externe de la plaque. Leur surface supporte quelques granules. Les plaques I et
V sont trčs étroites, leur bordure interne, relevée et saillante, présentant
trois ŕ quatre granules alignés ;
Périprocte. Aire périproctale allongée selon l'axe antéro-postérieur, pyriforme ;
Péristome. Non observable car encroűté de gangue.
- d'H. batalleri nov. sp. par ses granules scrobiculaires de taille plus variable et moins organisés autour des aires scrobiculaires ; par la présence d'étroites zones miliaires interradiales garnies de quelques granules miliaires épars et par le plus fort développement de ses fossettes scrobiculaires ; par la présence de trčs nettes fossettes angulaires ; par son disque apical ŕ surface plane ŕ fleur de test, contrairement ŕ celui d'H. batalleri nov. sp. dont le pourtour interne est trčs relevé ; par son aire périproctale pyriforme (circulaire chez H. batalleri nov. sp.) ;
- d'H. neocomiense (, 1869) par la taille de ses tubercules proportionnellement plus petite ; par la présence de zones miliaires ambitales et supra-ambitales (celles-ci étant inexistants chez H. neocomiense) ; par son disque apicale moins "robuste", nettement plus ŕ fleur de test ;
- d'H. rugosum , 1846, par son nombre de tubercules primaires par série plus faible, par ses granules miliaires moins nombreux et moins densément répartis ; par ses zones porifčres proportionnellement plus larges et garnies de paires de pores de bien plus grande taille ; par son aire périproctale pyriforme (circulaire ŕ subcirculaire chez H. rugosum).
D'une maničre générale Hemidiadema forcallensis nov. sp. ne peut ętre confondue avec aucune autre espčce actuellement connue, en particulier en raison du le faible nombre de tubercules primaires ambulacraires supra-ambitaux.
- le nombre de tubercules par série est nettement inférieur ŕ 10 chez H. forcallensis nov. sp. (7 en moyenne) alors qu'il est toujours supérieure ŕ 10 chez H. rugosum (11 en moyenne), cette différence se faisant surtout en dessus de l'ambitus (1 ŕ 2 tubercules primaires supra-ambitaux par série chez H. forcallensis nov. sp. contre 5 ŕ 6 chez H. rugosum) ;
- son disque apical est redressé sur sa bordure interne contrairement au disque apical d'H. rugosum dont la surface bosselée est uniformément plane ;
- l'absence totale de zones miliaires (adradiales, interradiales, perradiales) et ainsi de granules miliaires, contrairement ŕ H. rugosum chez qui des zones miliaires perradiales et surtout interradiales recouvertes de granules miliaires se développent en dessus l'ambitus ;
- l'absence de fossettes suturales angulaires et de larges dépressions interradiales contrairement ŕ H. rugosum oů elles sont fortement développées.
| Espčces [Nb. indiv.] |
Test | Ambulacres | Interambulacres | Périprocte | Péristome | ||||||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. | Largeur ambitale (en mm) |
Nb. tub. infra-A. | Nb. tub. supra-A. |
Nb. tub. | Largeur ambitale (en mm) |
A / IA | Ř (en mm) |
Ř / Ř test | Ř (en mm) |
Ř / Ř test | |
| H.
rugosum (Grandpré (*) + Alcoraia) [7] |
6,90 ŕ 10,23 8,28 |
3,62 ŕ 5,60 4,81 |
25 ŕ 27 | 1,51 ŕ 2,18 1,76 |
5 ŕ 6 | 5 ŕ 6 | 11 ŕ 13 | 2,38 ŕ 3,70 2,86 |
0,62 | 1,80 ŕ 2,84 2,09 |
0,25 | - | - |
| H.
neocomiense (néotype) (*) |
8,48 | 5,14 | 24 ŕ 25 | 1,9 | 5 | 3 ŕ 4 | 9 | 3,1 | 0,61 | 2,12 | 0,25 | 2,95 | 0,35 |
| H.
batalleri [6] |
10,10 ŕ 13,22 11,46 |
6,20 ŕ 6,67 6,48 |
27 ŕ 31 | 2,10 ŕ 2,90 2,53 |
3 ŕ 5 | 0 ŕ 6 | 8 ŕ 10 | 3,60 ŕ 4,61 4,17 |
0,61 | 3,42 ŕ 3,96 3,71 |
0,32 | 3,06 ŕ 3,52 3,29 |
0,29 |
| H.
malladai [15] |
5,07 ŕ 11,39 7,96 |
2,40 ŕ 5,01 3,91 |
18 ŕ 30 | 0,97 ŕ 2,29 1,68 |
3 ŕ 6 | 3 ŕ 5 | 6 ŕ 11 | 1,86 ŕ 4,19 2,88 |
0,58 | 1,60 ŕ 3,62 2,36 |
0,3 | 2,06 ŕ 4,17 2,98 |
0,37 |
| H. forcallensis [16] |
4,11 ŕ 6,54 5,37 |
1,92 ŕ 3,25 2,62 |
15 ŕ 21 | 0,83 ŕ 1,37 1,18 |
4 ŕ 5 | 1 ŕ 3 | 6 ŕ 8 | 1,42 ŕ 2,35 1,88 |
0,63 | 1,62 ŕ 2,47 2,07 |
0,38 | 1,55 ŕ 2,13 1,87 |
0,35 |
| Hemidiadema
sp. A [1] |
12,96 | - | - | 3,1 | 6 ŕ 7 | 4 ŕ 5 | 10 | 4,67 | 0,66 | 4,01 | 0,31 | - | - |
Figure 3: Tableau résumant les différents comptages et mesures réalisés sur les individus étudiés dans le présent travail et dans l'étude de , 2013 (*).
Nb. pla. : nombre
de plaques par série.
Nb
tub. : nombre de
tubercules primaires par série.
Nb
tub. infra-A. : nombre
de tubercules primaires infra-ambitaux (le tubercule ambital - parfois un peu
excentré en
dessus l'ambitus - compris) par série.
Nb
tub. supra-A. : nombre
de tubercules primaires supra-ambitaux par série.
A :
largeur de l'ambulacre au niveau de l'ambitus.
IA :
largeur de l'interambulacre au niveau de l'ambitus.
A
/ IA : rapport entre
les largeurs de l'ambulacre et de l'interambulacre, prises au niveau de
l'ambitus.
Ř : diamčtre.
[Nb. indiv.] : nombre d'individus étudiés et mesurés.
Valeurs
en gras : moyennes.
Hemidiadema rugosum , 1846. L'exemplaire ici décrit – unique exemplaire connu en Espagne – provient du sommet de l'Aptien supérieur de Loma de Casa lo Bolini, dépression de l'Alcoraia (province d'Alicante). En France l'espčce est également connue dans l'Aptien supérieur (Clansayésien) de Grandpré (Ardennes) (, 2013) et du Teil prčs de Montélimar (Ardčche) (, in prep.).
Hemidiadema batalleri nov. sp. n'est actuellement connue que dans les marnes pyriteuses ŕ Plicatules datées du sommet de l'Aptien inférieur (Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata) de Cala del Moro, prčs de Punta de l'Ŕliga, au Sud du village d'El Perelló (province de Tarragone).
Hemidiadema malladai nov. sp. n'est connue que dans les environs d'Allepuz et de Jorcas oů elle n'est pas rare. Tous les individus proviennent de marnes calcaires finement bioclastiques mal datées, mais appartenant aux Marnes de Forcall ("Formación Margas de Forcall"). Un exemplaire (coll. ) a également été trouvé dans la Zone ŕ Epicheloniceras martini du Massif du Garraf (province de Barcelone).
Hemidiadema forcallensis nov. sp. n'est connue actuellement que dans les marnes pyriteuses de l'Aptien inférieur, i.e., "Formación Margas de Forcall" ( et al., 1982, p. 286-288), datées de la partie supérieure de la Zone ŕ Deshayesites forbesi et de la partie basale de la Zone ŕ Deshayesites deshayesi des environs de Forcall et de Morella (province de Castellón).
Hemidiadema sp. A – connue que par un seul individu – provient des niveaux pyriteux ŕ Hemidiadema forcallensis nov. sp. ("Formación Margas de Forcall", partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi) de Mola d'en Camarŕs, prčs de Morella (province de Castellón).
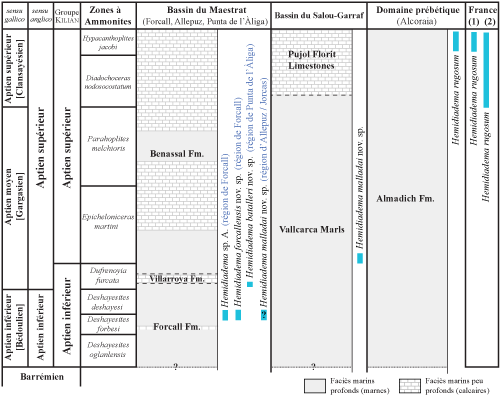
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4:
Distribution verticale et répartition régionale des
différentes espčces étudiées (lithostratigraphie
d'aprčs
et al., 2014, et biozonation ammonitique d'aprčs
& , 2014, et et
al.,
2014).
sensu gallico : division tripartite de l'Aptien (
et
al.,
2004,
2004).
sensu anglico: division bipartite de l'Aptien (
et
al.,
2011).
Groupe : division bipartite de l'Aptien (
& , 2014).
(1) : Le Teil, Ardčche, Sud-Est de la France (,
in prep.).
(2) : Clansayésien de Grandpré, Ardennes, Bassin parisien (, 2013).
La terminologie utilisée pour définir les relations paléoécologiques des espčces en fonction de la nature (texture, granulométrie, …) du substrat suit (1965).
Hemidiadema rugosum , 1846. L'exemplaire connu en Espagne provient d'un niveau clastique (lithoclastes et/ou bioclastes arrondis visibles dans la gangue adhérant au fossile), tout comme les rares exemplaires connus en France ["Minerai de fer du Bois des Loges" prčs de Grandpré (Ardennes) formé de trčs nombreux lithoclastes roulés de silice, de limonite et de glauconie emballés dans un ciment argileux (, 2013) ; "Grčs glauconieux ŕ Discoides decoratus" de l'Aptien supérieur du Teil prčs de Montélimar (Ardčche, France]. Dans ces trois localités oů l'espčce a été trouvée, les formations sédimentaires correspondent toutes ŕ d'anciens substrats oů la fraction de lithoclastes et bioclastes est nettement dominante. Il en résulte de ce constat qu'H. rugosum serait une espčce gravellicole.
Hemidiadema batalleri nov. sp. : Tous les exemplaires connus proviennent de marnes ŕ fossiles pyriteux dépourvues de bioclastes. H. batalleri nov. sp. est ainsi une espčce vasicole stricte.
Hemidiadema malladai nov. sp. : Tous les exemplaires proviennent de marnes calcaires finement bioclastiques. H. malladai nov. sp. est ainsi une espčce vasicole.
Hemidiadema forcallensis nov. sp. : Les individus connus proviennent tous des marnes ŕ fossiles pyriteux des Marnes de Forcall. H. forcallensis nov. sp. est ainsi une espčce vasicole.
Le présent travail a permis de démontrer que, contrairement aux citations antérieures (, 1928 ; , 1991, 2003 ; et al., 2000, 2007 ; , 2008 ; et al., 2008), les Hemidiadema de l'Aptien d'Espagne n'appartiennent pas ŕ l'espčce Hemidiadema rugosum , 1846. L'absence d'études comparatives ainsi qu'une méconnaissance de l'espčce d' liée ŕ une description originale trčs succincte de son auteur ( 1846, p. 351 ; , 2013) ont conduit ŕ des confusions et des attributions spécifiques inappropriées.
L'étude des Hemidiadema récoltés en différentes localités aptiennes d'Espagne et leur comparaison avec les deux principales espčces connues de ce genre [H. rugosum , 1846, et H. neocomiense (, 1869)] du Crétacé inférieur de la bordure orientale du Bassin parisien) permet de reconnaitre trois nouvelles espčces. Parmi ce matériel étudié, un seul spécimen appartient ŕ l'espčce rugosum d'
La diversité des Hemidiadema, jusqu'ŕ présent réduite ŕ quatre espčces nominales [H. rugosum 1846, H. neocomiense (, 1869), H. intermedius (, 1864) et H. guebhardi , 1920], est ainsi mieux appréhendée.
La seule espčce commune ŕ la France et ŕ l'Espagne est Hemidiadema rugosum , 1846, espčce typiquement gravellicole et cantonnée au sommet de l'Aptien supérieur. Les espčces vasicoles (Hemidiadema malladai nov. sp., Hemidiadema forcallensis nov. sp., Hemidiadema batalleri nov. sp., Hemidiadema sp. A) ne sont pas encore connues en dehors des régions espagnoles étudiées.
L'auteur tient ŕ remercier vivement toutes les personnes qui ont rendu possible la recherche (fructueuse ou infructueuse), l'accčs et l'emprunt pour étude des échinides : Sebastián (Musée géologique du Séminaire de Barcelone, Espagne), Maria Paz (Département de Paléontologie, Faculté des Sciences géologiques, Université Complutense, Madrid, Espagne), Brigitte (Cheval-Blanc, France), Carlos (Alicante, Espagne), Joaquín (Caudete, Espagne) et Eduard (Barcelone, Espagne). L'auteur remercie trčs amicalement Josep Anton (Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Mexique) et Oriol Fault Dynamics Research Group, Earth Sciences Department, University of London, Royaume-Uni) pour leurs précieuses aides concernant la rédaction des parties "Cadre géologique" et "Cadre stratigraphique" et le pręt pour étude des échantillons d'Hemidiadema récoltés par eux-męmes. L'auteur remercie également Sylvain et Jean-Michel (Muséum national d'Histoire naturelle de Paris, France) pour l'accčs et l'utilisation des binoculaires du MNHN pour l'étude (observations, comptages et mesures) des échinides. L'auteur remercie enfin les rapporteurs – Loďc (Université Pierre et Marie Curie, Paris, France) et Jaume (Musée des Sciences naturelles de Barcelone, Espagne) – pour leurs remarques constructives ayant permis d'améliorer la qualité de ce travail.
L. & E. (1846-1847).- Catalogue raisonné des espčces, des genres et des familles d'Échinides.- Annales des Sciences naturelles, Paris, série 3, t. VI (1846), p. 306-374 (Pls. 15-16) ; t.VII (1847), p. 129-168 ; t. VIII (1847), p. 5-35 et 355-380.
T., J.A., R., P.W., K. & E. (2010).- Sedimentary evolution of an Aptian syn-rift carbonate system (Maestrat Basin, E Spain): effects of accommodation and environmental change.- Geologica Acta, Barcelona, vol. 8, nº 3, p. 249-280.
T., R., J.A. & K. (2009).- Sequence stratigraphy and architecture of a late Early–Middle Aptian carbonate platform succession from the western Maestrat Basin (Iberian Chain, Spain).- Sedimentary Geology, vol. 219, p. 280-301.
T., R., J. & J.A. (2014).- Deep incision in an Aptian carbonate succession indicates major sea-level fall in the Cretaceous.- Sedimentology, vol. 61, nº 6, p. 1558–1593.
L.G., J-L., A. & A. (2014).- New insight on the genus Nolaniceras , 1961 (Ammonoidea, Cretaceous) and its consequences on the biostratigraphy of the Aptian Stage.- Proceedings of the Geologists' Association, London, vol. 125, p. 227-232.
J., P., G., R. & J. (1982).- Ibérica Central-Maestrazgo. In : El Cretácico de Espańa.- Universidad Complutense, Madrid, p. 273-344.
A. (2013).- Révision des espčces Hemidiadema rugosum , 1846 et H. neocomiense (, 1869) (Euechinoidea, Camarodonta, Glyphocyphidae) du Crétacé inférieur du Bassin parisien (France).- Carnets Geol., Madrid, vol. 13, nº M01 (CG2013_M01), 57 p.
A., O. & J.A. (2012).- El registro de especies de Hemidiadema (Echinoidea) durante el Aptiense (Cretácico Inferior) en las cuencas del Maestrazgo, Salou-Garraf y Organyŕ.- Geo-Temas, Salamanca, vol. 13, p. 1-4.
M., J. & J.M. (2004).- El Barremiense de Fontcalent y L'Alcoraia.- Geo-Temas, Salamanca, vol. 7, p. 217-221.
G. (1862-1867).- Paléontologie française. Terrain Crétacé. Tome 7 : Échinides réguliers.- Masson Ed., Paris, 892 p. (1204 Pls.).
G. (1869).- Échinides nouveaux ou peu connus.- Revue et Magasin de Zoologie pure et appliquée, Paris, 2čme série, t. XXI, p. 238-248 (Pls. 12-13).
G., J.A., J.J. & D. (2013).- Xerticeras gen. nov., a new genus of micromorphic heteromorph ammonite (Ancyloceratina, Ancyloceratidae) from the lower Aptian of Spain.- Carnets Geol., Madrid, vol. 13, nº A02 (CG2013_A02), p. 89-103.
V.L. (1969).- Ammonites from the Clansayesian beds of the Western Caucasus.- Trudy Krasnodarsgogo Filiala Vsesojuznogo Neftegazovogo Nauchno-Issledovatelskogo Instituta, Krasnodar, vol. 19, p. 126-188 [en Russe].
H.B. & D.L. (1966).- Echinacea. In : R.C. (ed.), Treatise on Invertebrate Paleontology, Part U, Echinodermata 3.- The University of Kansas, Lawrence; The Geological Society of America, Boulder, vol. 2, p. U367-U440.
R., J.A., T., M., R., J.-L., J.D., E., L.G., G., R. & A. (2014).- Lower Cretaceous (Hauterivian-Albian) ammonite biostratigraphy in the Maestrat Basin (E Spain).- Journal of Iberian Geology, Madrid, vol. 40, nº 1, p. 99-112.
M. (1933).- Les oursins réguliers fossiles, évolution et classification.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Grenoble, t. XVII, fasc. 2, 125 p. (9 Pls.).
A. & A.B. (2010).- The phylogeny and classification of post-Palaeozoic echinoids.- Journal of Systematic Palaeontology, London, vol. 8, nº 2, p. 147-212.
J. (1892).- Recherches sur les Échinides de l'Aptien de Grandpré.- Bulletin de la Société géologique de France, Paris, (série 3), t. XX, p. 38-100 (Pls. II-IV).
J. (1920).- Note sur quelques échinides du Crétacé inférieur de la Provence.- Notes provençales, Saint-Vallier-de-Thiey, nº 11, p. 1-24 (Pls. II-III).
J. (1928).- Note sur quelques Échinides du Crétacé d'Espagne communiqués par M. le Prof. .- Boletin de la Real Sociedad espańola de Historia natural, Madrid, t. 28, nº 3, p. 147-157 (1 Pl.).
J. & C. (2006).- Equínidos fósiles de la provincia de Alicante.- Memorias del Mupe 2, Museo paleontológico de Elche, 264 p.
J. & B. (2007).- Event stratigraphy of the upper Cretaceous to lower Eocene hemipelagic sequences of the Prebetic Zone (SE Spain): Record of the onset of tectonic convergence in a passive continental margin.- Sedimentary Geology, vol. 197, nº 1-2, p. 141-163.
J.D., G E., T., A., R., J.A., S., M., A., J. & S.L. (2013).- The Upper Aptian to Lower Albian syn-rift carbonate succession of the southern Maestrat Basin (Spain): Facies architecture and fault-controlled stratabound dolostones.- Cretaceous Research, vol. 41, p. 217-236.
M., A. & F. (2004).- Succession des peuplements d'échinides du Crétacé inférieur dans les Haut Atlas Atlantique (Maroc) : révision systématique et intéręt stratigraphique.- Géobios, Lyon, vol. 37, p. 595-617.
M., S.V., F.E., T. & I. (2012).- Early Cretaceous ammonite fauna of Catanduanes Island, Philippines.- Cretaceous Research, vol. 37, p. 261-271.
J. (1977).- Cretaceous echinoids of Stara Planina.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Belgrad, t. XLI, p. 181-202 (4 Pls.).
J. (1996).- Cretaceous Echinoidea from Djebel Djerissa, Central Tunisia.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Belgrad, t. LX, nº 1, p. 169-194.
J.A. (2007).- Bioestratigrafía del Aptiense del macizo del Garraf (NE de la Península Ibérica).- Geogaceta, Salamanca, vol. 41, p. 131-134.
J.A., R., G., M. & R. (2014).- Review of the early Aptian (Early Cretaceous) ammonoid species Deshayesites deshayesi (d', 1841).- Cretaceous Research, vol. 51, p. 341-360.
J.A., T., R. & R. (2012a).- Uppermost Lower Aptian transgressive records in Mexico and Spain: chronostratigraphic implications for the Tethyan sequences.- Terra Nova, Oxford, vol. 24, nº 4, p. 333-338.
J.A., M., T., R., G., R. & A. (2009a).- Biostratigraphic characterization by means of ammonoids of the Lower Aptian Oceanic Anoxic Event (OAE 1a) in the esatern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, vol. 30, p. 864-872.
J.A., M., T., R., G., F.J.-M.R. & A. (2010).- Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence).- Geologica Acta, Barcelona, vol. 8, nº 3, p. 281-299.
J.A., M., J., J.M., T., R., G., F.J.-M.R. & R. (2012b).- Lower Aptian ammonite and carbon isotope stratigraphy in the eastern Prebetic Domain (Betic Cordillera, southeastern Spain).- Geologica Acta, Barcelona, vol. 10, nº 4, p. 333-350.
J.A., O., P., T., M., J., C., R., A., R. & S. (2008).- Bioestratigrafía integrada del Aptiense del Macizo del Garraf (Cadenas Costero-Catalanas, NE de Espańa). Datos preliminares. In : J.I., L. & J.C. (eds.), Libro del resúmenes, XXIV Jornadas de la Sociedad Espańola de Paleontología.- Museo del Jurásico de Asturias (MUJA), Colunga, 15-18 de octubre de 2008, p. 158-159.
J.A., L., M., G., T., L.-G., J.-L. & R. (2009b).– Bioestratigrafía de los ammonites de Can Casanyes, Macizo de Garraf (Barcelona, Espańa).- Batalleria, Barcelona, vol. 14, p. 91-98.
T. (1943).- A monograph of the Echinoidea. Vol. 3, 2 : Camarodonta 1 (Orthopsidae, Glyphocyphidae, Temnopleuridae, and Toxopneustidae).- C.A. Reitzel, Copenhagen, 546 p. (56 Pls.).
M., B. & G. (2011).- The Aptian Stage: Back to fundamentals.- Episodes, Bangalore, vol. 34, nº 3, p. 148-156.
M., G., C. & G. (2006).- Le Gargasien (Aptien moyen) de la Tuiličre - Saint-Saturnin-lčs-Apt (région du stratotype historique de l'Aptien, Vaucluse, SE France) : localisation géographique et corrélations stratigraphiques.- Carnets Geol., Madrid, vol. 6, nº L01 (CG2006_L01), 8 p.
M., G., W., M. & J.-P. (2004).- Le Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de l'Aptien inférieur, SE France) : localisation géographique et corrélations stratigraphiques.- Carnets Geol., Madrid, vol. 4, nº L02 (CG2004_L02), 4 p.
M.-L. (1959).- Contribution ŕ l'étude des Échinides fossiles du Maroc (Jurassique et Crétacé).- Notes et Mémoires du service géologique du Maroc, Rabat, nº 146, 183 p. (Pls. I-XVII).
J. (1965).- Recherches qualitatives sur les biocénoses marines des substrats meubles dragables de la région marseillaise.- Thčse Sc. nat., Marseille, 160 p.
S., O., M.B., R., M., V., M., M., J.A., J., E.J., M.K., I., C., S., A., S.N., P.F. & J.M. (2014).- Report on the 5th International Meeting of the IUGS Lower Cretaceous Working Group, the " Group" (Ankara, Turkey, 31st August 2013).- Cretaceous Research, vol. 50, p. 126-137.
A.E. (2008).- Unterkreide-Lokalitäten im Maestrazgo Nordost-Spanien.- Fundstellen, www.leitfossil.de, 47 p.
E., B. & L.G. (2001).- Caractérisation d'une nouvelle sous-zone d'ammonites au passage Aptien-Albien dans les "Marnes noires ŕ Hypacanthoplites" des Pyrénées espagnoles.- Geobios, Lyon, Vol. 34, nº 1, p. 53-62.
R. & J. (1996).- Rasgos estructurales principales de la cuenca Cretácica Inferior del Maestrazgo (Cordillera Ibérica oriental).- Geogaceta, Salamanca, vol. 20, p. 1704-1706.
R., J., R., C., A. & A. (2001).- Evolution of the Mesozoic Central Iberian Rift System and its Cenozoic inversion (Iberian Chain). In : P.A., W., A.H.F. & S. (eds.), Peri-Tethys Memoir 6: Peri-Tethyan Rift/Wrench Basins and Passive Margins.- Mémoires du Muséum national d'Histoire Naturelle, Paris, t. 186, p. 145-185.
M. & J.A. (2000).- Equínoideos del Aptiense Maestrazgo y zonas limítrofes.- Cidaris, Elche, nº 17-18, p. 10-33.
M. & J.A. (2015).- Carinacea II : Echinacea II - Calycina.- Nomochirus, Onda, nº 6, 330 p. (106 Pls.).
M., J.A. & J. (2007).- Fauna equínida del Maestrat, Cretaci inferior.- Asociación Paleontológica de Onda, 1 poster.
O. (ed.), L., L. & I. (2007).- Aptian-Campanian ammonites of Hungary.- Geologica Hungarica, Budapest, (Series Palaeontologica), vol. 57, p. 1-187.
A. & P. (2010).- Échinides nouveaux, peu ou mal connus.- Annales de la Société d'Histoire naturelle du Boulonnais, Boulogne-sur-Mer, t. IX, fasc. 3, p. 1-24.
M.P. ( 1991, inédit).- Revisión de los equínidos del Cretácico inferior y medio espańol.- Colección Tesis Doctorales, Universidad Complutense Madrid, Facultad de Ciencias geológicas, Madrid, nş 250/93, t. I y II, 714 p. (33 Pls.).
M.P. (2003).- Los equínidos regulares de la provincia de Castellón.- Nemus. Ateneo de Natura, Castelló de la Plana, nº 1 (2002), p. 37-92 (3 Pls.).
Toutes les photographies ont été prises par l'auteur avec un Canon EOS 350D muni d'un objectif 18-55 mm, associé ŕ trois bagues-allonges Kepcor 31 mm, 21 mm et 13 mm. Les échinides ont été éclairés en utilisant des ampoules "day light".
En vues apicale et orale, l'astérisque "*" indique l'interambulacre 2 supportant ŕ son extrémité apicale la madréporite (plaque génitale 2).
All photographs were taken by the author using a Canon EOS 350D equipped with a 18-55 mm lens, together with Kepcor 31 mm, 21 mm and 13 mm extension rings. The echinoids were illuminated using "day light" bulbs.
In apical and oral views, the asterik "*" indicates the interambulacrum 2 bearing the madreporite on his apical end (genital plate 2).
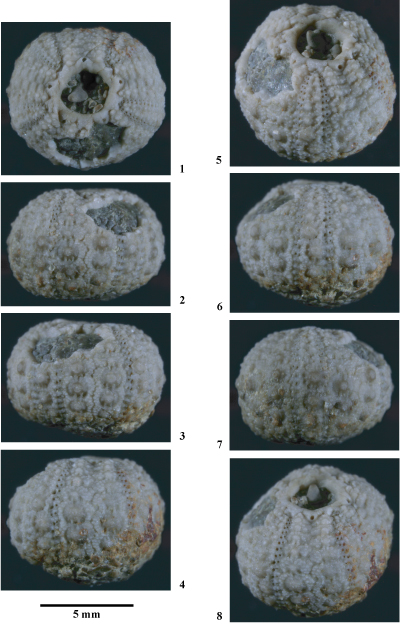
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 1 : Hemidiadema rugosum , 1846
Coll.
Sommet de l'Aptien supérieur ; Loma de Casa lo Bolini, région de l'Acoraia, province d'Alicante, Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 2.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 3.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 5 : Vue supra-ambitale centrée sur l'ambulacre IV.
Fig. 6 : Vue ambitale centrée sur l'ambulacre IV.
Fig. 7 : Vue ambitale centrée sur l'ambulacre II.
Fig. 8 : Vue supra-ambitale centrée sur l'interambulacre 4.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 2 : Hemidiadema rugosum , 1846
Coll.
Sommet de l'Aptien supérieur ; Loma de Casa lo Bolini, région de l'Acoraia, province d'Alicante, Espagne.
Fig. 1 : Interambulacre 3 en vue ambitale.
Fig. 2 : Interambulacre 2 en vue ambitale.
Fig. 3 : Ambulacre IV en vue ambitale.
Fig. 4 : Interambulacre 1 en vue supra-ambitale.
Fig. 5 : Interambulacre 4 en vue supra-ambitale.
Fig. 6 : Ambulacre IV en vue supra-ambitale.
Fig. 7 : Détails d'un tubercule interambulacraire.
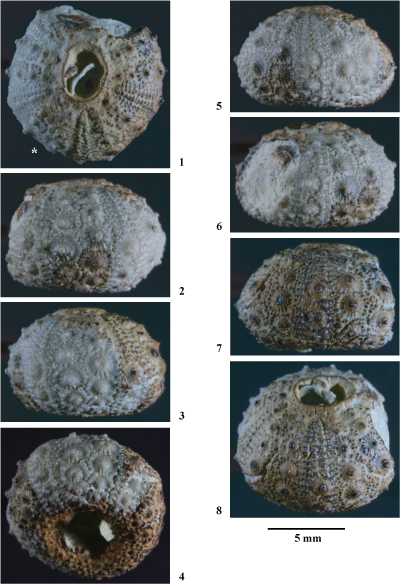
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 3 : Hemidiadema batalleri nov. sp. Holotype
MGSB 79924 ; Coll.
Aptien inférieur, Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata ; Cala del Moro (Punta de l'Ŕliga), Sud d'El Perelló (province de Tarragone), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 1.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 2.
Fig. 4 : Vue infra-ambitale centrée sur l'interambulacre 2.
Fig. 5 : Vue ambitale centrée sur l'ambulacre II.
Fig. 6 : Vue ambitale centrée sur l'ambulacre I.
Fig. 7 : Vue ambitale centrée sur l'ambulacre IV.
Fig. 8 : Vue supra-ambitale centrée sur l'ambulacre IV.
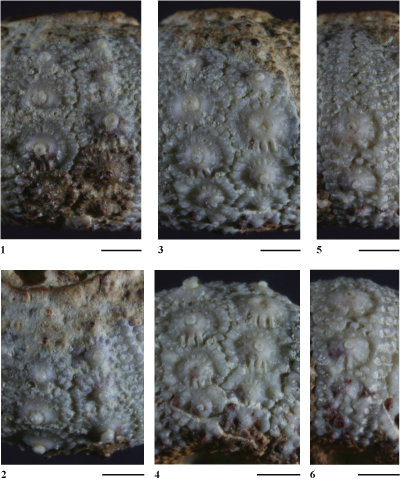
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 4 : Hemidiadema batalleri nov. sp. Holotype
MGSB 79924 ; Coll.
Aptien inférieur, Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata ; Cala del Moro (Punta de l'Ŕliga), Sud d'El Perelló (province de Tarragone), Espagne.
Fig. 1 : Interambulacre 1 en vue ambitale.
Fig. 2 : Interambulacre 1 en vue supra-ambitale.
Fig. 3 : Interambulacre 2 en vue supra-ambitale.
Fig. 4 : Interambulacre 2 en vue infra-ambitale.
Fig. 5 : Ambulacre II en vue ambitale.
Fig. 6 : Ambulacre II en vue infra-ambitale.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 5 : Hemidiadema batalleri nov. sp. Paratype 1
MGSB 25112-1 ; Coll.
Aptien inférieur, Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata ; Cala del Moro (Punta de l'Ŕliga), Sud d'El Perelló (province de Tarragone), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 1.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 4 : Vue supra-ambitale centrée sur l'interambulacre 4.
Fig. 5 : Vue supra-ambitale centrée sur l'ambulacre III.
Fig. 6 : Vue ambitale centrée sur l'ambulacre V.
Fig. 7 : Vue supra-ambitale centrée sur l'ambulacre V.
Fig. 8 : Disque apical.
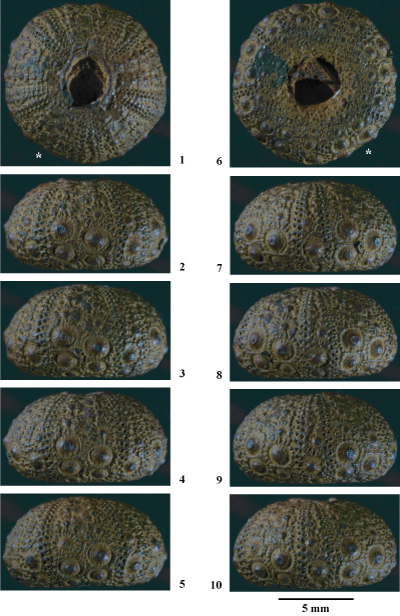
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 6 : Hemidiadema batalleri nov. sp. Paratype 2
MGSB 25112-2 ; Coll.
Aptien inférieur, Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata ; Cala del Moro (Punta de l'Ŕliga), Sud d'El Perelló (province de Tarragone), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 5.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 1.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 2.
Fig. 5 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 6 : Vue générale de la face orale.
Fig. 7 : Vue ambitale centrée sur l'ambulacre I.
Fig. 8 : Vue ambitale centrée sur l'ambulacre II.
Fig. 9 : Vue ambitale centrée sur l'ambulacre III.
Fig. 10 : Vue ambitale centrée sur l'ambulacre V.
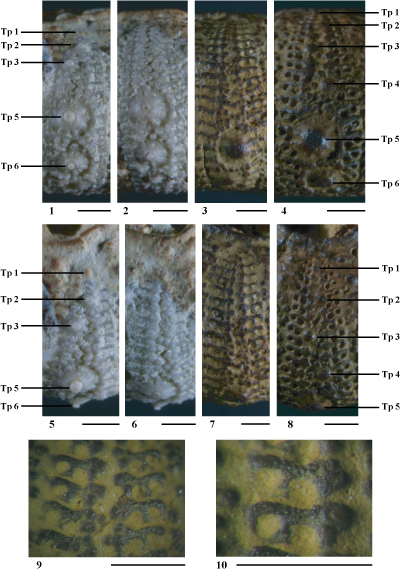
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 7 : Hemidiadema batalleri nov. sp.
Aptien inférieur, Zone ŕ Dufrenoyia furcata, Sous-zone ŕ Dufrenoyia furcata ; Cala del Moro (Punta de l'Ŕliga), Sud d'El Perelló (province de Tarragone), Espagne.
Détails et comparaisons des ambulacres de différents spécimens.
Figs. 1, 2, 5, 6 : Hemidiadema batalleri nov. sp. ; MGSB 79924.
Figs. 1, 5 : Ambulacre I en vues ambitale et supra-ambitale.
Noter l'absence du tubercule primaire supra-ambital 4.
Fig. 2, 6 : Ambulacre II en vue ambitale et supra-ambitale.
Noter l'absence de tous les tubercules primaires supra-ambitaux.
Figs. 3, 7, 9, 10 : Hemidiadema batalleri nov. sp. ; MGSB 25112-1.
Figs. 3, 7 : Ambulacre V en vue ambitale et supra-ambitale.
Noter l'absence de tous les tubercules primaires supra-ambitaux.
Fig. 9 : Détails de la partie supra-ambitale d'un ambulacre.
Fig. 10 : Détails d'assules de la zone porifčre. Noter la présence de granules ŕ la surface de ces assules.
Figs. 4, 8 : Hemidiadema batalleri nov. sp. ; MGSB 25112-2.
Ambulacre IV en vue ambitale et supra-ambitale.
Noter la présence de tous tubercules primaires supra-ambitaux.
Tp : tubercule primaire.
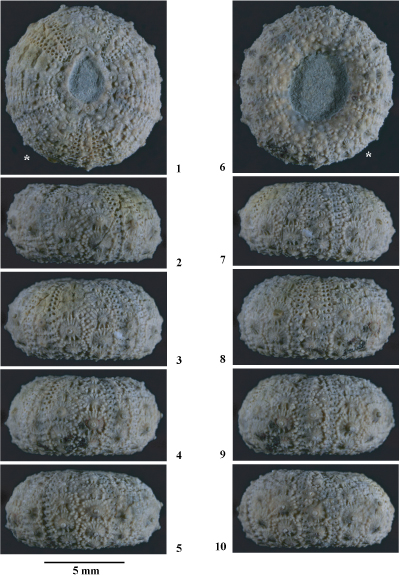
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 8 : Hemidiadema malladai nov. sp. Holotype
MNHN.F. A50416 ; Coll.
Aptien inférieur, Marnes de Forcall (? partie supérieure de la Zone ŕ Deshayesites forbesi et/ou partie basale de la Zone ŕ Deshayesites deshayesi), région située entre Allepuz et Jorcas (province de Teruel), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 5.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 1.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 3.
Fig. 5 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 6 : Vue générale de la face orale.
Fig. 7 : Vue ambitale centrée sur l'ambulacre II.
Fig. 8 : Vue ambitale centrée sur l'ambulacre III.
Fig. 9 : Vue ambitale centrée sur l'ambulacre IV.
Fig. 10 : Vue ambitale centrée sur l'ambulacre V.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 9 : Hemidiadema malladai nov. sp. Holotype
MNHN.F. A50416 ; Coll.
Aptien inférieur, Marnes de Forcall (? partie supérieure de la Zone ŕ Deshayesites forbesi et/ou partie basale de la Zone ŕ Deshayesites deshayesi), région située entre Allepuz et Jorcas (province de Teruel), Espagne.
Fig. 1 : Détails de l'interambulacre 4 en vue ambitale.
Fig. 2 : Détails de l'ambulacre III en vue ambitale.
Fig. 3 : Détails de l'ambulacre IV en vue ambitale.
Fig. 4 : Détails de l'ambulacre II en vue supra-ambitale.
Fig. 5 : Détails de l'interambulacre 5 en vue infra-ambitale.
Fig. 6 : Détails du disque apical.
Fig. 7 : Vue ambitale centrée sur l'ambulacre V.
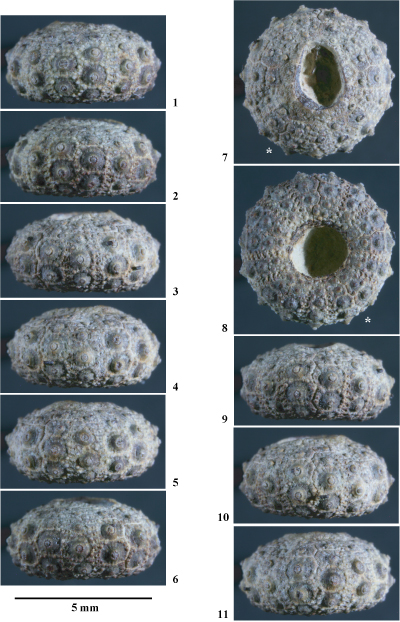
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 10 : Hemidiadema forcallensis nov. sp. Holotype
MNHN.F. A50417 ; Coll.
Aptien inférieur, Marnes de Forcall, partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi ; Mas del Roc prčs de Forcall (province de Castellón), Espagne.
Fig. 1 : Vue ambitale centrée sur l'interambulacre 3.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 3 : Vue ambitale centrée sur l'ambulacre I.
Fig. 4 : Vue ambitale centrée sur l'ambulacre II.
Fig. 5 : Vue ambitale centrée sur l'ambulacre III.
Fig. 6 : Vue ambitale centrée sur l'ambulacre V.
Fig. 7 : Vue générale de la face apicale.
Fig. 8 : Vue générale de la face orale.
Fig. 9 : Vue ambitale centrée sur l'interambulacre 5.
Fig. 10 : Vue ambitale centrée sur l'interambulacre 1.
Fig. 11 : Vue ambitale centrée sur l'interambulacre 2.
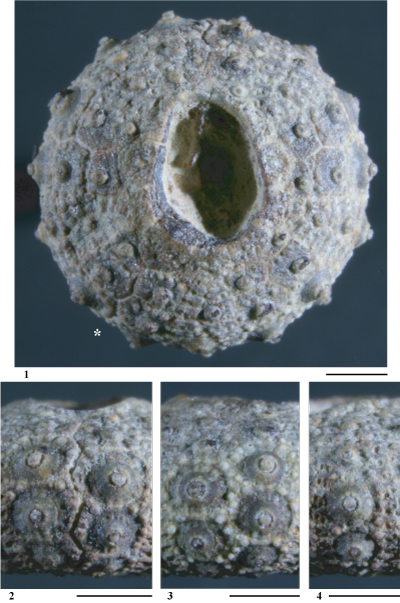
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 11 : Hemidiadema forcallensis nov. sp. Holotype
MNHN.F. A50417 ; Coll.
Aptien inférieur, Marnes de Forcall, partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi ; Mas del Roc prčs de Forcall (province de Castellón), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Détails de l'interambulacre 5 en vue ambitale.
Fig. 3 : Détails de l'interambulacre 3 en vue ambitale.
Fig. 4 : Détails de l'ambulacre I en vue ambitale.
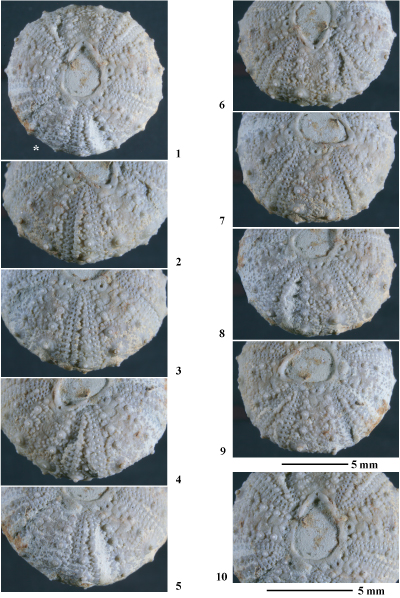
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 12 : Hemidiadema sp. A
MNHN.F. A50418 ; Coll.
Aptien inférieur, Marnes de Forcall, partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi ; Mola d'en Camarŕs prčs de Forcall (province de Castellón), Espagne.
Fig. 1 : Vue générale de la face apicale.
Fig. 2 : Vue supra-ambitale centrée sur l'ambulacre V.
Fig. 3 : Vue supra-ambitale centrée sur l'ambulacre IV.
Fig. 4 : Vue supra-ambitale centrée sur l'ambulacre I.
Fig. 5 : Vue supra-ambitale centrée sur l'ambulacre III.
Fig. 6 : Vue supra-ambitale centrée sur l'interambulacre 5.
Fig. 7 : Vue supra-ambitale centrée sur l'interambulacre 4.
Fig. 8 : Vue supra-ambitale centrée sur l'interambulacre 3.
Fig. 9 : Vue supra-ambitale centrée sur l'interambulacre 1.
Fig. 10 : Vue de la face apicale centrée sur le disque apical.
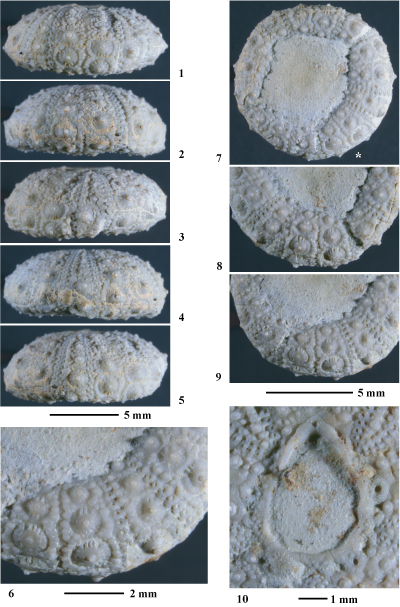
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 13 : Hemidiadema sp. A
MNHN.F. A50418 ; Coll.
Aptien inférieur, Marnes de Forcall, partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi ; Mola d'en Camarŕs prčs de Forcall (province de Castellón), Espagne.
Fig. 1 : Vue ambitale centrée sur l'interambulacre 5.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 4.
Fig. 3 : Vue ambitale centrée sur l'ambulacre I.
Fig. 4 : Vue ambitale centrée sur l'ambulacre V.
Fig. 5 : Vue ambitale centrée sur l'ambulacre V.
Fig. 6 : Vue infra-ambitale centrée sur l'interambulacre 2 et l'ambulacre II.
Fig. 7 : Vue générale de la face orale.
Fig. 8 : Vue infra-ambitale centrée sur l'interambulacre 1.
Fig. 9 : Vue infra-ambitale centrée sur l'interambulacre 2.
Fig. 10 : Disque apical.
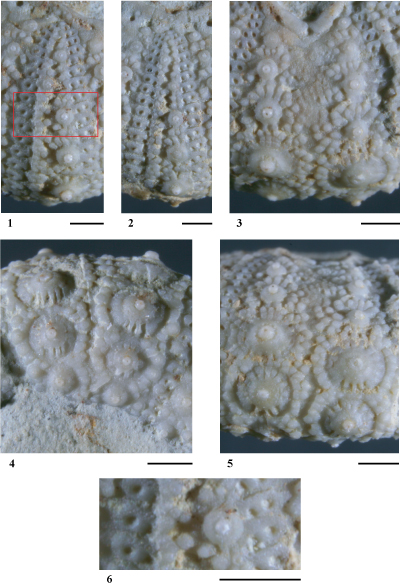
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 14 : Hemidiadema sp. A
MNHN.F. A50418 ; Coll.
Aptien inférieur, Marnes de Forcall, partie supérieure de la Zone ŕ Deshayesites forbesi et partie basale de la Zone ŕ Deshayesites deshayesi ; Mola d'en Camarŕs prčs de Forcall (province de Castellón), Espagne.
Fig. 1 : Détails de l'ambulacre V en vue supra-ambitale.
Fig. 2 : Détails de l'ambulacre IV en vue supra-ambitale.
Fig. 3 : Détails de l'interambulacre 5 en vue supra-ambitale.
Fig. 4 : Détails de l'interambulacre 1 en vue infra-ambitale.
Fig. 5 : Détails de l'interambulacre 5 en vue ambitale.
Fig. 6 : Détails de l'ambulacre V en vue supra-ambitale, centrée sur un tubercule primaire.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 15 : Comparaison des disques apicaux.
Fig. 1 : Hemidiadema rugosum (MNHN.F A33783 ; Coll. ) : Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France.
Fig. 2 : Hemidiadema rugosum (MNHN.F A33782 ; Coll. ) : Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France.
Fig. 3 : Hemidiadema rugosum (coll. ) : Sommet de l'Aptien supérieur, Loma de Casa lo Bolini, région de l'Acoraia, province d'Alicante, Espagne.
Fig. 4 : Hemidiadema neocomiense (ENSGN / MAN, coll. nº384) : Hauterivien inférieur (? Zone ŕ Lyticoceras nodosoplicatum), Ville-sur-Saulx, Meuse, France ; néotype.
Fig. 5 : Hemidiadema malladai nov. sp. (MNHN.F. A50416) ; holotype.
Fig. 6 : Hemidiadema batalleri nov. sp. (MGSB 25112-1, coll. ) ; paratype.
Fig. 7 : Hemidiadema forcallensis nov. sp. (MNHN.F. A50417) ; holotype.
Fig. 8 : Hemidiadema sp. A (MNHN.F. A50418).
Barre d'échelle : 1 mm / Bar scale : 1 mm.