◄ Carnets Geol. 16 (24) ►
![]()
Sommaire
[1. Introduction]
[2. Cadres géographique et géologique]
[3. Systématique des ostracodes]
[4. stratigraphie]
[5. Paléoécologie]
[6. Paléobiogéographie]
[7. Conclusion]
[Références bibliographiques] et ...
[Planches]
GET (Géosciences Environnement Toulouse),
Université Paul Sabatier, Toulouse III, CNRS, IRD, OMP, Thématique Couplage
Lithosphčre-Océan-Atmosphčre (LOA), 14 avenue É. Belin, 31400 Toulouse
(France)
Département
de Géologie, Faculté des Sciences, Université Mohammed 1er, B.P.
524, 60 000 Oujda (Maroc)
13 Lot. Terrasses de la Figuičre, 30140 Anduze (France)
Manuscrit en ligne depuis le 12 décembre 2016
[Éditeur : Bruno Granier]
![]()
Les marnes de la partie supérieure de la Formation du Tigri du Crétacé supérieur des Hauts Plateaux méridionaux, Maroc oriental, ont livré des faunes d'ostracodes non-marins, dulçaquicoles ŕ oligohalins. Malgré la mauvaise conservation des individus, 23 espčces ont été reconnues, qui appartiennent ŕ 14 genres. Les associations génériques montrent une répartition paléobiogéographique globale ŕ la fois sur la Laurasia et sur le Gondwana.
• ostracodes non-marins ;
• Crétacé supérieur ;
• Hauts Plateaux méridionaux ;
• Maroc oriental.
Andreu B., Haddoumi H. & Charričre A. (2016).- Ostracodes non marins du Crétacé supérieur des Hauts Plateaux méridionaux, Maroc oriental.- Carnets Geol., Madrid, vol. 16, no. 24, p. 569-590.
Upper Cretaceous non-marine ostracods from the southern High Plateaus, eastern Morocco.- The marls of the upper part of the Tigri Formation in the Upper Cretaceous of the southern High Plateaus, eastern Morocco, reveal non-marine ostracod faunas from freshwater to oligohaline environments. Despite the poor specimen preservation, 23 species belonging to 14 genera have been recognized. The generic assemblages show a global paleobiogeographic distribution both on Laurasia and Gondwana.
• non-marine ostracods;
• Late Cretaceous;
• southern High Plateaus;
• Eastern Morocco.
Depuis quelques années, les "couches rouges" des facies purbeckiens-wealdiens et du Crétacé supérieur continental du Maroc livrent des associations d'ostracodes et de charophytes (Andreu et al., 2003 ; Colin et Tabuce, 2004 ; Haddoumi et al., 2008 ; Mojon et al., 2009) qui permettent d'une part de dater ces dépôts continentaux, d'autre part de préciser la paléoécologie des faunes et des flores ainsi que leur distribution paléobiogéographique locale et globale.
Les ostracodes présentés dans ce travail proviennent des dépôts du Crétacé supérieur, ou Sénonien rouge continental, des Hauts Plateaux méridionaux, du Maroc oriental. Ils ont été récoltés dans la partie supérieure de la Formation du Tigri (Haddoumi et al., 2015).
Le
Maroc oriental est occupé par une vaste zone de Hauts Plateaux s'étendant du sud
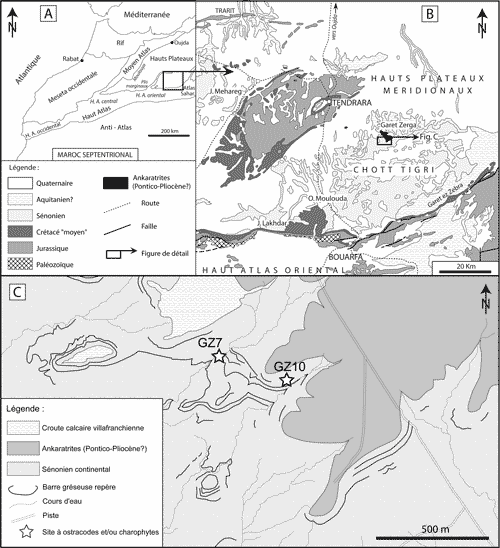
d'Oujda aux chaînons du Haut Atlas oriental (Fig. 1.A ![]() )
et se
prolongeant vers l'est par la Meseta Oranaise. Les Hauts Plateaux méridionaux
marocains constituent une large gouttičre synclinale bordée, ŕ l'est et au
nord,
par l'axe Mechkakour-Trarit et, au sud, par la chaîne bordičre du Haut Atlas
oriental. Cette gouttičre est compliquée par le large bombement du dôme de
Tendrara (Fig. 1.B
)
et se
prolongeant vers l'est par la Meseta Oranaise. Les Hauts Plateaux méridionaux
marocains constituent une large gouttičre synclinale bordée, ŕ l'est et au
nord,
par l'axe Mechkakour-Trarit et, au sud, par la chaîne bordičre du Haut Atlas
oriental. Cette gouttičre est compliquée par le large bombement du dôme de
Tendrara (Fig. 1.B ![]() ). Au
sud de ce dernier, le Chott Tigri constitue une dépression
d'environ 60 km d'W en E et de 30 km du N au S, sculptée dans la partie sud
des Hauts Plateaux marocains jusqu'aux premiers reliefs atlasiques.
). Au
sud de ce dernier, le Chott Tigri constitue une dépression
d'environ 60 km d'W en E et de 30 km du N au S, sculptée dans la partie sud
des Hauts Plateaux marocains jusqu'aux premiers reliefs atlasiques.
Les
terrains crétacés des Hauts Plateaux sont discordants sur le Bajocien ou le
Bathonien marins. Ils débutent avec la trilogie classique du "Mésocrétacé"
(Choubert et Faure-Muret, 1960-62) : Infracénomanien, Cénomanien
et Cénomano-Turonien. Les calcaires de plate-forme du Cénomano-Turonien sont
suivis en concordance par une série détritique rouge, souvent azoďque et généralement
attribuée au "Sénonien régressif".
Sur le rebord nord de la dépression de Tigri, cette unité est ravinée et
recouverte en discordance soit par les ankaratrites de Garet Zerga, attribuées
au Pliocčne (Gautier, 1914) ou au Pontico-Pliocčne (Médioni,
1970), soit par un ensemble de bancs de grčs rose, ŕ calcédoine et accidents
siliceux, attribué ŕ l'Aquitanien (Médioni, 1970), recouverts d'une
croűte villafranchienne (Fig. 1.C ![]() ).
).
Le substratum du Chott Tigri est ainsi majoritairement constitué par la série détritique rouge post-turonienne définie (Médioni, 1970) comme "Formation des grčs et conglomérats du Chott Tigri" ou plus simplement nommée "Formation du Tigri" (Choubert, 1956). Elle est subdivisée en deux membres (Haddoumi et al., 2015) :
membre inférieur (100 ŕ 150 m) constitué de silts ou de marnes rouges, souvent gypsifčres, comportant plusieurs intercalations de barres carbonatées ŕ faune marine ou laguno-marine ;
membre supérieur (250 ŕ 350 m) comprenant une série relativement homogčne formée
de barres gréseuses alternant avec des niveaux silteux. La partie supérieure
de cet ensemble s'est révélée fossilifčre en deux points, dans le secteur
de Garet Zerga (Fig. 1.C ![]() ). Le gisement supérieur (GZ10) a livré des
charophytes et le gisement inférieur (GZ7), de nombreux ostracodes associés ŕ
divers autres microfossiles.
). Le gisement supérieur (GZ10) a livré des
charophytes et le gisement inférieur (GZ7), de nombreux ostracodes associés ŕ
divers autres microfossiles.
|
Figure 1 : A) Carte simplifiée du Maroc septentrional et localisation de l'aire étudiée dans les Hauts Plateaux méridionaux ; B) Carte géologique simplifiée de la région de Tigri-Tendrara dans les Hauts Plateaux méridionaux ; C) Carte géologique de secteur de Tigri et position des sites fossilifčres. Figure 1: A) Simplified map of Northern Morocco and location of the studied area in the Southern High Plateaus; B) Simplified geological map of the Tigri-Tendrara area in the Southern High Plateaus; C) Geological map of the Tigri area and location of the fossiliferous sites. |
À Garet Zerga, ŕ une cinquantaine de mčtres au-dessous des ankaratrites, s'observe la barre de grčs la plus épaisse (10 ŕ 15 m) de la série ; elle est suivie par 5 m de marnes dont la partie supérieure (gisements GZ7) est riche en charophytes (Mesochara, Stephanochara) et ostracodes.
Sur
deux coupes détaillées
(Fig. 2 ![]() )
dans le niveau GZ7 (échantillons GZ7a
- j), on note de bas en haut la succession suivante : 1) ostracodes mal conservés,
avec oogones de charophytes et un fragment de coquille de Dinosaurien ; 2)
ostracodes et oogones de charophytes mal conservés, présence de gypse ; 3)
ostracodes et oogones de charophytes plus ou moins bien conservés et radioles
d'oursins localement trčs abondants (éch. GZ7d). La séquence sus-jacente débute
avec des grčs ŕ rides de courant, rides linguoďdes et fentes de dessiccation.
)
dans le niveau GZ7 (échantillons GZ7a
- j), on note de bas en haut la succession suivante : 1) ostracodes mal conservés,
avec oogones de charophytes et un fragment de coquille de Dinosaurien ; 2)
ostracodes et oogones de charophytes mal conservés, présence de gypse ; 3)
ostracodes et oogones de charophytes plus ou moins bien conservés et radioles
d'oursins localement trčs abondants (éch. GZ7d). La séquence sus-jacente débute
avec des grčs ŕ rides de courant, rides linguoďdes et fentes de dessiccation.
D'aprčs Haddoumi et al. (2015), "L'ootaxon Pseudomegaloolithus atlasi est défini dans le Maastrichtien supérieur marocain d'Achlou2 (Moyen Atlas) et se trouve associé ŕ des charophytes du Campanien-Maastrichtien ŕ Tendrara (Hauts Plateaux). En conséquence, au moins le niveau GZ7 peut ętre considéré comme Maastrichtien. L'extension du genre Stephanochara ne permet pas une datation précise du niveau GZ10. Cependant, ce genre y est associé avec une espčce du genre Mesochara, genre connu seulement du Crétacé. Il en ressort qu'un âge Crétacé supérieur peut ętre proposé pour la partie supérieure de la Formation de Tigri".
|
Figure 2 : Deux coupes détaillées du site GZ7, avec composition des échantillons. Figure 2: Two detailed sections in the GZ7 outcrops, with composition of the samples. |
La classification adoptée est celle de Maddocks (1982) recommandée par Meisch (2000).
La répartition stratigraphique et paléobiogéographique
des genres est synthétisée sur les Figures 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() .
.
Classe Ostracoda Latreille, 1806
Sous-Classe Podocopa Sars, 1866
Super-Ordre Podocopomorpha Kozur, 1972
Ordre Podocopida Sars, 1866
Sous-Ordre Cytherocopina Baird, 1850
Super-Famille Cytheroidea Baird, 1850
Famille Limnocytheridae Klie, 1938
Sous-Famille Timiriaseviinae (Mandelstam, 1960) Colin et Danielopol, 1980
Genre Vecticypris Keen, 1972
Vecticypris sp. 1
(Pl. 1 ![]() , figs. 1-2)
, figs. 1-2)
Matériel : 2 carapaces.
Dimensions en mm : L = 0,75-0,85; l = 0,53 ; h = 0,40.
Description sommaire : carapace de forme ovale en vue latérale, peu élevée mais étalée ventralement ; en vue dorsale, forme losangique, extrémité antérieure moins arrondie que la postérieure.
Répartition locale : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7c), Hauts Plateaux méridionaux, Maroc.
Vecticypris sp. 2
(Pl. 1 ![]() , figs. 3-4)
, figs. 3-4)
Matériel : 2 carapaces.
Dimensions en mm : L = 0,63-0,66; l = 0,40 ; h = 0,34.
Description sommaire : carapace de forme ovale en vue latérale, peu élevée mais étalée ventralement ; en vue dorsale, forme losangique, les deux extrémités montrant ŕ peu prčs le męme arrondi.
Paléoécologie : Vecticypris, comme le genre Metacypris, colonise généralement les milieux dulçaquicoles (Keen, 1972) ŕ oligohalins. Trabelsi et Colin (2013) l'ont récolté dans un milieu limnique peu profond et permanent.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7i), Hauts Plateaux méridionaux, Maroc.
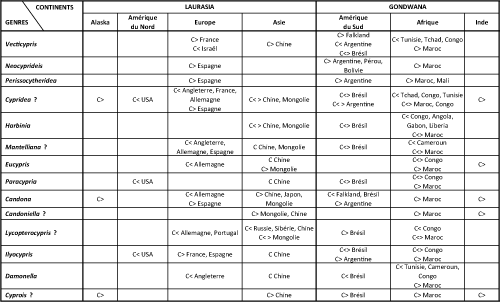
Répartition paléogéographique du genre : Vecticypris, considéré comme un Metacypris primitif (Keen, 1972), est connu dans le Cénomanien de France (Colin, 1974), Albien de Tunisie (Trabelsi et Colin, 2013), Crétacé basal du bassin nord des Îles Falkland (Ayress et Whatley, 2014), Crétacé inférieur d'Israël (Rosenfeld et Raab, 1984 ; Trabelsi et Colin, 2013), Aptien-Albien du Tchad (Colin et Dépęche, 1997), Wealdien-Albien du Congo (Metacypris polita, in Grékoff, 1957, 1960), Campanien supérieur-Maastrichtien inférieur d'Argentine (Carignano et Varela, 2011 ; Whatley, 2012), Turonien-Santonien (?) du Brésil (Dias-Brito et al., 1998), et Crétacé supérieur du Maroc (ce travail).
Famille Cytherideidae Sars, 1925
Sous-Famille Cytherideinae Sars, 1925
Genre Neocyprideis Apostolescu, 1956
Neocyprideis sp.
(Pl. 1 ![]() , fig. 5)
, fig. 5)
Matériel : 1 carapace.
Dimensions en mm : L = 1.00 ; h = 0,63.
Description sommaire : espčce de grande taille, allongée et ovale en vue latérale externe ; extrémités réguličrement arrondies, bordées par un bourrelet ; carapace trčs enflée latéralement, bombée dans la région postéro-ventrale ; flancs faiblement réticulés ; valve gauche légčrement plus grande que la droite.
Paléoécologie : genre oligohalin ŕ polyhalin voire euhalin (euryhalin de 0 ŕ 35 ‰, in Bilotte et Andreu, 2006), plutôt mésohalin, mais jamais d'eau douce (Keen, 1972 ; Mebrouk et al., 2011).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7f), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : Neocyprideis est présent depuis le Cénomanien en Europe du Sud et au Moyen Orient (Israël ; Colin et Carbonel, 1990a ; Bassiouni, 2002). Au Crétacé supérieur, il est reconnu en Europe du Sud (Maastrichtien ; Colin et al., 1990b ; Whatley et Bajpai, 2006 ; Mebrouk et al., 2011), au Maroc (ce travail) ; en Argentine (Uliana et Musacchio, 1978 ; Mebrouk et al., 2011 ; Whatley, 2012), au Pérou et en Bolivie (Colin et al., 1990b).
Famille Cytheridae Baird, 1850
Sous-Famille Cytherinae Baird, 1850
Tribu Perissocythereidini van den Bold, 1963
Genre Perissocytheridea Stephenson, 1938
Perissocytheridea sp.
(Pl. 1 ![]() , fig. 6)
, fig. 6)
Matériel : 1 carapace.
Dimensions en mm : L = 0,50 ; h = 0,30.
Description sommaire : carapace ovale en vue latérale externe ; bord dorsal subrectiligne, bord ventral légčrement concave ; extrémité antérieure réguličrement arrondie, tournée vers le bas ; extrémité postérieure triangulaire, tournée vers l'arričre ; les deux extrémités sont situées au-dessous de la mi-hauteur ; flancs bombés latéralement ; présence d'un léger sulcus oblique dans la partie antéro-centrale des flancs, souligné par l'ornementation ; réticulation grossičre ; carapace parcourue de muri proéminents, concentriques sur les bordures et se prolongeant du haut vers le bas le long du sulcus, alignés horizontalement dans la région centrale ; mailles de la réticulation peu apparentes ; les deux valves semblent subégales.
Affinités : Perissocytheridea ? laminensis Andreu et Ettachfini, 1994, du Cénomanien moyen ŕ supérieur du Bassin d'Essaouira, se différencie par la présence d'un bombement ventral associé ŕ une côte.
Paléoécologie : genre plutôt mésohalin, mais euryhalin (oligohalin ŕ polyhalin, Mebrouk et al., 2011).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7g), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : Perissocytheridea est connu dans l'Aptien de Tunisie (Trabelsi et al., 2015), et plus généralement depuis le Cénomanien, en Afrique, dont au Maroc (du Cénomanien au Santonien, in Andreu, 2002 ; dans le Crétacé supérieur (ce travail) et au Mali (Colin et al., 1996 ; Whatley et Bajpai, 2006), en Amérique du Sud (Maastrichtien, Musacchio, 1981, 1990 ; Whatley et Bajpai, 2006 ; Whatley, 2012), dont au Brésil (Viviers et al., 2000 ; Piovesan et al., 2014a, 2014b), en Grčce, et au Moyen-Orient (Mebrouk et al., 2011).
Famille Cyprideidae Martin, 1940
Sous-Famille Cyprideinae Martin, 1940 (cf. Horne et Colin, 2005)
Genre Cypridea Bosquet, 1852, emend.
Cypridea ? sp.
(Pl. 1 ![]() , fig. 7)
, fig. 7)
Matériel : 1 carapace.
Dimensions en mm : L = 0,50 ; h = 0,32.
Description sommaire : espčce appartenant avec doute au genre Cypridea ; individu de taille moyenne qui ne semble pas déformé, ovale en vue latérale externe ; extrémité antérieure haute, pourvue d'un trčs long rostre, qui dépasse le bord ventral, suivi postérieurement d'une légčre incision rostrale ; extrémité postérieure basse, réguličrement arrondie et tournée vers le bas ; flancs lisses ; valve droite dépassant la gauche sur les bords dorsal et ventral.
Affinités : nous n'avons pas trouvé, dans la littérature, une espčce proche de celle-ci.
Paléoécologie : genre d'eau douce ŕ saumâtre (Hou et al., 2002), dulçaquicole ŕ oligohalin (Andreu et al., 2003).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : cosmopolite (Trabelsi et Colin, 2013), qui montre une distribution globale (Do Carmo et al., 2008). Il est présent aussi bien sur la Laurasia que sur le Gondwana : dans les facies purbeckiens et wealdiens (Tithonien inférieur-Aptien) d'Europe (Angleterre, France, Allemagne ; Martin, 1940 ; Anderson et Barker, 1966 ; Anderson, 1967 ; Colin et Oertli, 1985) ; il est connu aussi en Amérique du Nord (USA, in Swain, 1999) et du Sud (Aptien-Turonien du Brésil, in Nicolaidis et al., 2011 ; Trabelsi et Colin, 2013 ; Maastrichtien d'Argentine (Musacchio, 1981, 1990 ; Whatley et Bajpai, 2006), en Alaska (Maastrichtien, Brouwers et De Deckker, 1993 ; Whatley et Bajpai, 2006), en Chine et Mongolie (Tithonien inférieur-Crétacé supérieur, in Szczechura, 1971, 1978 ; Horne et Colin, 2005 ; Whatley et Bajpai, 2006 ; Trabelsi et Colin, 2013), en Afrique (Crétacé inférieur-Wealdien, in Grékoff, 1957, et Crétacé supérieur du Congo, in Grékoff, 1960 ; Tithonien-Albien d'Afrique du Nord : Maroc et Tunisie, in Andreu et al., 2003 ; Haddoumi et al., 2008 ; Mojon et al., 2009 ; Crétacé inférieur du Tchad (Trabelsi et Colin, 2013) ; Crétacé supérieur du Maroc (ce travail), en Inde (Maastrichtien, Khosla et Sahni, 2000 ; Khosla et al., 2011 ; Whatley, 2012). Hou et al. (2002) donnent la répartition globale suivante : Europe, Amériques, Asie, Afrique, du Jurassique moyen au Cénozoďque.
Sous-Ordre Cypridocopina Jones, 1901
Super-Famille Cypridoidea Baird, 1845
Famille Cyprididae Baird, 1845
Sous-Famille Cypridinae Baird, 1845
Genre Harbinia (Tsao, 1959) Hou, 1984
Harbinia sp. 1
(Pl. 1 ![]() , fig. 8)
, fig. 8)
Matériel : 1 carapace, usée et légčrement écrasée.
Dimensions en mm : L = 0,73 ; h = 0,46.
Description sommaire : carapace ovale en vue latérale externe ; extrémité antérieure réguličrement arrondie et située au-dessous de la mi-hauteur ; extrémité postérieure dissymétrique, en position basse et aplatie (?) latéralement ; bord dorsal en arc de cercle, angle dorsal trčs ouvert ; plus grande hauteur ŕ mi-longueur ; bord ventral rectiligne ; flancs lisses ; valve gauche recouvre la droite sur la périphérie (?).
Paléoécologie : genre lacustre ŕ lagunaire (Do Carmo et al., 2008), mixohalin ŕ hyperhalin (Andreu et al., 2003 ; Mojon et al., 2009 ; Antonietto et al., 2012). Les formes brésiliennes, citées par Do Carmo et al., ont été considérées comme appartenant au genre Pattersoncypris par Poropat et Colin (2002 ; Cabral, A.C., communication personnelle).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7c), Hauts Plateaux méridionaux, Maroc.
Harbinia sp. 2
(Pl. 1 ![]() , fig. 12)
, fig. 12)
Matériel : 1 carapace.
Dimensions en mm : L = 0,66 ; h = 0,40.
Description sommaire : carapace ovale en vue latérale externe ; extrémité antérieure réguličrement arrondie et située au-dessous de la mi-hauteur, tournée vers le bas ; extrémité postérieure réguličrement arrondie, tournée vers l'arričre ; les deux extrémités sont situées au-dessous de la mi-hauteur ; bord dorsal en arc de cercle, angle dorsal trčs ouvert, plus grande hauteur ŕ l'avant ; bord ventral rectiligne ŕ légčrement concave ; flancs lisses ; valves subégales.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7c), Hauts Plateaux méridionaux, Maroc.
Harbinia sp. 3
(Pl. 1 ![]() , figs. 9-11)
, figs. 9-11)
Matériel : une dizaine de carapaces.
Dimensions en mm : L = 0,60-0,65 ; h = 0,42-0,46.
Description sommaire : carapace ovale en vue latérale externe, mais plus arrondie que chez Harbinia sp. 1 et sp. 2 ; extrémité antérieure réguličrement arrondie et située au niveau de la mi-hauteur, tournée vers l'avant ; extrémité postérieure réguličrement arrondie, tournée vers le bas, située au-dessous de la mi-hauteur ; bord dorsal arrondi, angle dorsal trčs ouvert, plus grande hauteur ŕ l'avant ; bord ventral légčrement concave ; flancs lisses ; valves subégales.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7c et GZ7i), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : décrit dans le Crétacé de Chine (in Hou et al., 2002 ; Do Carmo, 2008), reconnu dans le Crétacé supérieur du Brésil (Hourcqia in Dias-Brito et al., 2001 ; Nicolaidis et al., 2011 ; Antonietto et al., 2012), dans le Crétacé inférieur (plutôt Aptien-Albien) de l'Afrique de l'Ouest : Congo, Angola, Gabon, Liberia (synonymes genres Hourcqia et Pattersoncypris in Antonietto et al., 2012 ; Poropat et Colin (2002) considčrent que ces deux genres ne sont pas synonymes), dans le Crétacé inférieur (Mojon et al., 2009 ; genre Hourcqia in Andreu et al., 2003) et dans le Crétacé supérieur (ce travail) du Maroc.
Genre Mantelliana Anderson, 1966
Mantelliana ? sp.
(Pl. 1 ![]() , figs. 13-15)
, figs. 13-15)
Matériel : 8 (?) carapaces, plus ou moins écrasées.
Dimensions en mm : L = 0,65-0,68 ; h = 0,33-036.
Description sommaire : espčce appartenant avec doutes au genre Mantelliana ; forme ovale et arquée en vue latérale externe ; extrémité antérieure arrondie avec présence d'un angle antéro-dorsal peu marqué ; extrémité postérieure réguličrement arrondie ; les deux extrémités sont tournées vers le bas et situées au-dessous de la mi-hauteur ; bord dorsal en arc de cercle, bord ventral rectiligne ŕ convexe ; flancs lisses ; valves subégales.
Paléoécologie : genre dulçaquicole (Colin et al., 1992) ŕ oligohalin (Andreu et al., 2003).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7f1), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : décrit dans les facies purbeckiens et wealdiens d'Angleterre (Anderson in Anderson et Barker, 1966) ; retrouvé dans le Tithonien-Berriasien d'Allemagne (Schudack, 1994), Barrémien-Aptien d'Espagne (Brenner, 1976), Crétacé de Chine et de Mongolie (Hou et al., 2002), Crétacé inférieur du Cameroun (Colin et al., 1992), Aptien-Albien (Tomé et Filho, 2010) et Crétacé supérieur du Brésil (Dias-Brito et al., 2001), Crétacé inférieur (Hauterivien supérieur-Barrémien inférieur ; Andreu et al., 2003) et Crétacé supérieur (ce travail) du Maroc.
Sous-Famille Eucypridinae Bronstein, 1947
Genre Eucypris Vávra, 1891
Eucypris sp. 1
(Pl. 1 ![]() , figs. 16-19)
, figs. 16-19)
Matériel : une dizaine de carapaces, souvent écrasées.
Dimensions en mm : L = 0,83-0,88 ; h = 0,46-0,53.
Description sommaire : espčce d'assez grande taille, triangulaire ŕ ovale en vue latérale externe ; extrémités antérieure et postérieure réguličrement arrondies, tournées vers le bas, la postérieure semblant plus aplatie latéralement ; bord dorsal arrondi, présentant un angle dorsal trčs ouvert, en avant de la mi-longueur ; bord ventral rectiligne ; flancs lisses ; valves subégales (?).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7b, GZ7c, GZ7f2), Hauts Plateaux méridionaux, Maroc.
Eucypris sp. 2
(Pl. 2
![]() , figs. 1-2)
, figs. 1-2)
Matériel : une quinzaine de carapaces, parfois écrasées.
Dimensions en mm : L = 0,66-0,69 ; h = 0,40-0,42.
Description sommaire : espčce ovale en vue latérale externe ; extrémités antérieure et postérieure réguličrement arrondies, tournées vers le bas, au-dessous de la mi-hauteur ; extrémité antérieure moins élevée que la postérieure ; hauteur maximale juste en avant de la mi-longueur ; bord dorsal en arc de cercle, bord ventral légčrement concave ; flancs lisses ; valve droite plus grande que la gauche (?) avec recouvrement sur le bord dorsal, de part et d'autre de la hauteur maximale.
Paléoécologie : genre dulçaquicole (Szczechura, 1971 ; Khosla et Sahni, 2000 ; Haddoumi et al., 2008) ŕ oligohalin (Neale, 1988).
Répartition locale : Albien-Cénomanien (?) du Haut Atlas oriental (Eucypris ?, in Haddoumi et al., 2008) ; Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7c, GZ7d), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : Purbeckien-Wealdien d'Allemagne (Martin, 1940), Wealdien (Crétacé inférieur anté-Aptien supérieur, Grékoff, 1957) et Crétacé supérieur du Congo (Grékoff, 1960), Crétacé de Chine (Hou et al., 2002) et de Mongolie (Maastrichtien ; Whatley, 2012), Albien-Crétacé supérieur du Maroc (ci-dessus et ce travail), Crétacé supérieur (Maastrichtien) d'Inde (Khosla et Sahni, 2000 ; Whatley et Bajpai, 2006 ; Khosla et al., 2011 ; Whatley, 2012).
Famille Candonidae Kaufmann, 1900
Sous-Famille Paracypridinae Sars, 1923
Tribu Thalassocypridini Hartmann et Puri, 1974
Genre Paracypria Sars, 1910
Paracypria sp.
(Pl. 2
![]() , fig. 3)
, fig. 3)
Matériel : une carapace.
Dimensions en mm : L = 1.75 ; h = 0,90.
Description sommaire : espčce de grande taille ; carapace ovale en vue latérale externe ; extrémité antérieure arrondie, légčrement tournée vers le bas, aplatie latéralement ; extrémité postérieure réguličrement arrondie, tournée vers l'arričre ; extrémités au-dessous de la mi-hauteur ; bord dorsal arrondi ŕ l'avant de la hauteur maximale, rectiligne ŕ l'arričre ; hauteur maximale en avant de la mi longueur ; bord ventral presque rectiligne ; flancs lisses ; la valve gauche recouvre la droite sur le bord postéro-dorsal et sur le bord ventral, peut-ętre sur la périphérie ?
Paléoécologie : genre dulçaquicole (Do Carmo et al., 2013). Actuellement, le genre Paracypria peut vivre dans un environnement anchihalin, dans des mares n'ayant aucun lien de surface avec la mer, contenant du sel ou de l'eau saumâtre, et dont la salinité fluctue au gré des marées (Maddocks et al., 1993).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7d), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : présent dans le Crétacé inférieur anté Aptien supérieur du Congo (Grékoff, 1960), Crétacé inférieur (Aptien) du Brésil (Do Carmo et al., 2013) et des USA (Swain, 1999), Crétacé supérieur du Maroc (ce travail), Crétacé supérieur du Brésil (Dias-Brito et al., 2001), Crétacé de Chine (Hou et al., 2002). Ces derniers auteurs donnent la répartition suivante : Afrique, Asie, Europe, Océanie, Nouvelle Zélande, du Jurassique supérieur ŕ l'Actuel.
Sous-Famille Candoninae Kaufmann, 1900
Genre Candona Baird, 1845
Candona sp.
(Pl. 2
![]() , fig. 4)
, fig. 4)
Matériel : une carapace entičre non déformée.
Dimensions en mm : L = 0,86 ; h = 0,42.
Description sommaire : espčce d'assez grande taille, ovoďde en vue latérale externe ; extrémité antérieure arrondie, haute, tournée vers le bas ; hauteur maximale située prčs de l'avant ; extrémité postérieure triangulaire, tournée vers l'arričre, beaucoup moins développée que l'antérieure ; bord dorsal peu arrondi de la hauteur maximale ŕ l'angle postéro-dorsal ; bord ventral concave ; flancs lisses ; valves subégales.
Paléoécologie : genre généralement dulçaquicole (Szczechura, 1971 ; Do Carmo et al., 2013), qui peut vivre dans les milieux oligohalin et mésohalin (Meisch, 2000).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7g), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : cosmopolite, global (Hou et al., 2002), présent dans les facies purbeckiens-wealdiens d'Allemagne (Martin, 1940), dans le Crétacé basal du bassin nord des Îles Falkland (Ayress et Whatley, 2014), dans le Crétacé inférieur (Aptien) du Brésil (Do Carmo et al., 2013), dans le Crétacé supérieur de Chine et du Japon (Cao, 1996 ; Hou et al., 2002), de Mongolie (Szczechura et Blaszyk, 1969 ; Szczechura, 1971, 1978), d'Inde (Maastrichtien ; Khosla et Sahni, 2000 ; Khosla et al., 2011 ; Whatley, 2012), d'Alaska (Maastrichtien, Brouwers et De Deckker, 1993 ; Whatley et Bajpai, 2006), d'Amérique du Sud (Argentine ; Whatley, 2012), et dans le Crétacé supérieur du Maroc (ce travail).
Genre Candoniella Schneider, 1956
Candoniella ? sp.
(Pl. 2
![]() , figs. 5-9)
, figs. 5-9)
Matériel : 8 carapaces.
Dimensions en mm : L = 0,66-0,80 ; h = 0,30-0,40.
Description sommaire : espčce appartenant avec doutes au genre Candoniella ; ovoďde et arquée en vue latérale externe ; extrémité antérieure arrondie, tournée vers le bas, située au niveau de la mi-hauteur ; extrémité postérieure plutôt arrondie, tournée vers le bas, moins développée que l'antérieure, et située au-dessous de la mi-hauteur ; hauteur maximale au niveau de la mi-longueur ; bord dorsal en arc de cercle ; bord ventral légčrement concave ; flancs lisses ; valves subégales.
Paléoécologie : genre dulçaquicole (Szczechura, 1971 ; Swain, 1999).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7, GZ7f1, GZ7i), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : cité dans le Paléozoďque (Swain, 1999) et connu dans le Crétacé supérieur de Mongolie (Szczechura et Blaszyk, 1969 ; Szczechura, 1971, 1978 ; Whatley et Bajpai, 2006) et de Chine (Whatley, 2012), dans le Maastrichtien d'Inde (Whatley, 2012), et dans le Crétacé supérieur du Maroc (ce travail). Hou et al. (2002) donnent la répartition suivante : Europe, Asie, Amériques, du Crétacé ŕ l'Actuel.
Genre Lycopterocypris Mandelstam, 1956
Lycopterocypris ? sp.
(Pl. 2
![]() , fig. 10)
, fig. 10)
Matériel : 1 carapace.
Dimensions en mm : L = 1.66 ; h = 0,84.
Description sommaire : espčce appartenant avec doutes au genre Lycopterocypris, de grande taille, ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers l'avant, située au niveau de la mi-hauteur ; extrémité postérieure arrondie, mais présence d'un angle postérieur, tournée vers l'arričre et située au niveau de la mi-hauteur ; hauteur maximale au niveau de la mi-longueur ; bord dorsal en arc de cercle, légčrement dissymétrique et moins élevé ŕ l'arričre ; bord ventral légčrement concave ; flancs lisses ; valves subégales mais dépassement de la valve droite au niveau de l'angle antéro-ventral. Malgré des différences dans le contour latéral externe, nos individus se rapprochent le plus du genre Lycopterocypris.
Paléoécologie : genre généralement dulçaquicole (Szczechura, 1971 ; Andreu et al., 2003), mais associé ŕ des genres d'ostracodes euryhalins (Cabral, 1995 ; Cabral et al., 2008).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : décrit dans le Crétacé inférieur de la Transbaďkalie (ou Daourie, ŕ l'est du Lac Baďkal), Russie, en Sibérie et Mongolie (Mandelstam, 1956 ; Lyubimova, 1955, 1956 ; Whatley et Bajpai, 2006 ; Cabral et al., 2008). Il est connu aussi dans le Crétacé inférieur Européen : France (Donze, 1964 ; Donzeau et al., 1997), Portugal (Cabral, 1995 ; Cabral et al., 2008), et Africain : Congo (dans le Wealdien, Crétacé inférieur anté Aptien supérieur, in Grékoff, 1960), Maroc (Andreu et al., 2003), dans le Crétacé supérieur du Brésil (Dias-Brito et al., 2001), dans le Crétacé supérieur du Maroc (ce travail), dans le Crétacé inférieur de Chine (Dequan, 1990), et dans le Crétacé supérieur-Paléocčne de Mongolie (Szczechura, 1971, 1978).
Famille Ilyocyprididae Kaufmann, 1900
Sous-Famille Ilyocypridinae Kaufmann, 1900
Genre Ilyocypris Brady et Norman, 1889
Ilyocypris sp.
(Pl. 2
![]() , fig. 11)
, fig. 11)
Matériel : 1 carapace.
Dimensions en mm : L = 0,66 ; h = 0,32.
Description sommaire : espčce de petite taille pour le genre, ovale ŕ sub-quadrangulaire en vue latérale externe, comprimée latéralement ; extrémités antérieure et postérieure réguličrement arrondies, tournées respectivement vers l'avant et l'arričre ; plus grande longueur au niveau de la mi-hauteur ; bord dorsal rectiligne avec des angles antérieur et postérieur marqués ; bord ventral concave avec un angle postérieur marqué ; double sulcus en position antérieure, dépressions séparées par un trčs fort tubercule ; le sulcus antérieur descend verticalement sur les flancs jusqu'ŕ mi-hauteur, le postérieur, en position médiane, sur plus des 2/3 de la hauteur ; surface des flancs recouverte de fossettes d'oů émergent quelques rares tubercules ŕ l'arričre ; valves subégales.
Paléoécologie : genre généralement dulçaquicole (Do Carmo et al., 2013) pouvant vivre en milieu oligohalin (Meisch, 2000).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7d), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : cosmopolite et global (Hou et al., 2002), connu dans le Crétacé inférieur-Cénozoďque des USA (Swain, 1999), dans le Crétacé inférieur (Aptien) du Brésil (Do Carmo et al., 2013 ; Tomé et al., 2014), dans le Crétacé inférieur (Wealdien) et supérieur du Congo (Grékoff, 1957, 1960), dans le Crétacé de Chine (Hou et al., 2002), dans le Crétacé supérieur d'Europe (France et Espagne, in Rodríguez-Lázaro et Martín-Rubio, 2005) et d'Amérique du Sud (Maastrichtien d'Argentine, in Musacchio, 1973, 1981, 1990 ; Musacchio et Simeoni, 1989 ; Whatley et Bajpai, 2006), et dans le Crétacé supérieur du Maroc (ce travail).
Genre Damonella Anderson, 1966
Damonella aff. buchaniana Anderson, 1966
(Pl. 2
![]() , fig. 12)
, fig. 12)
1966 Damonella buchaniana n. sp. - Anderson, p. 441, Figs. 17 et 21 p. 440, Fig. 24 p. 444.
Matériel : 1 carapace.
Dimensions en mm : L = 0,60 ; h = 0,45.
Description sommaire : espčce ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, peu tournée vers le bas, située au niveau de la mi-hauteur ; extrémité postérieure arrondie, tournée vers l'arričre, et située au niveau de la mi-hauteur ; hauteur maximale au niveau de la mi-longueur ; bord dorsal en arc de cercle, légčrement dissymétrique et moins élevé ŕ l'arričre ; bord ventral concave sur sa partie médiane, la concavité maximale étant située au niveau de la hauteur maximale ; angles latéraux absents ; flancs lisses ; valve gauche plus grande que la droite, la recouvrant au niveau du bord ventral.
Affinités et différences : Damonella buchaniana montre un contour latéral externe symétrique de part et d'autre de la hauteur maximale, et des dimensions bien inférieures.
Paléoécologie : genre dulçaquicole, limnique (Trabelsi et Colin, 2013).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7), Hauts Plateaux méridionaux, Maroc. Damonella buchaniana est décrite dans le Jurassique (Purbeckien supérieur) d'Angleterre (Anderson, 1966, in Anderson et Barker, 1966).
Répartition paléogéographique du genre : connu dans le Jurassique (Purbeckien)-Crétacé (Wealdien) d'Angleterre (Anderson et Barker, 1966), dans le Jurassique supérieur-Crétacé de Chine (Hou et al., 2002), dans le Crétacé inférieur (Aptien-Albien inférieur) du Brésil (Tomé et Filho, 2010 ; Reconcavona ? ultima Krommelbein et Weber, 1971, in Tomé et al., 2014), dans l'Albien de Tunisie (Trabelsi et Colin, 2013), dans le Crétacé inférieur du Cameroun (Tomé et al., 2014) et du Congo (facičs wealdiens, in Grosdidier, 1967), et dans le Crétacé supérieur du Maroc (ce travail).
Damonella sp. 1
(Pl. 3 ![]() , figs. 1-12)
, figs. 1-12)
Matériel : une vingtaine de carapaces.
Dimensions en mm : L = 0,62-0,73 ; h = 0,33-0,41.
Description sommaire : espčce ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers le bas, située au-dessous de la mi-hauteur ; extrémité postérieure trčs réguličrement arrondie, tournée vers l'arričre, plus haute et développée que l'antérieure, située au niveau de la mi-hauteur ; hauteur maximale au niveau de la mi-longueur ; bord dorsal réguličrement arrondi, légčrement dissymétrique et plus élevé ŕ l'arričre ; bord ventral rectiligne, légčrement concave en avant de la hauteur maximale ; angles latéraux absents ; flancs lisses ; valve gauche plus grande que la droite, la recouvrant sur la périphérie. La mauvaise conservation des individus ne permet pas de mettre en évidence un éventuel dimorphisme sexuel ; les mâles correspondraient aux individus les plus longs et proportionnellement les moins hauts.
Affinités et différences : Damonella sp. 1 montre quelques ressemblances morphologiques avec Damonella grandiensis Tomé et al., 2014, de l'Aptien du Brésil. Chez cette derničre, le contour latéral externe est plus carré, l'extrémité antérieure plus élevée, les dimensions inférieures.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7, GZ7b, GZ7d, GZ7f1, GZ7g), Hauts Plateaux méridionaux, Maroc.
Damonella sp. 2
(Pl. 2
![]() , figs. 17-20)
, figs. 17-20)
Matériel : une vingtaine de carapaces.
Dimensions en mm : L = 0,60-0,80 ; h = 0,36-0,45.
Description sommaire : espčce ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers le bas, située au-dessous de la mi-hauteur ; extrémité postérieure plutôt angulaire, tournée vers l'arričre, et située au-dessous de la mi-hauteur, plus développée en hauteur que l'antérieure en vue de droite ; hauteur maximale en avant de la mi-longueur ; bord dorsal quasi rectiligne de part et d'autre de l'angle dorsal ; bord ventral rectiligne et concave sur sa partie médiane, la concavité maximale étant située au niveau de la hauteur maximale ; flancs lisses ; dépassement de la valve gauche sur la périphérie de la droite.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7, GZ7c, GZ7f1, GZ7g, GZ7 lat.), Hauts Plateaux méridionaux, Maroc.
Damonella sp. 3
(Pl. 2
![]() , figs. 13-16)
, figs. 13-16)
Matériel : une vingtaine de carapaces.
Dimensions en mm : L = 0,53-0,68 ; h = 0,33-0,40.
Description sommaire : espčce ovoďde et arquée en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers le bas, située au niveau de la mi-hauteur ; extrémité postérieure arrondie, tournée vers l'arričre et située au niveau de la mi-hauteur ; hauteur maximale au niveau de la mi-longueur ; bord dorsal en arc de cercle, symétrique de part et d'autre de la hauteur maximale ; bord ventral concave sur sa partie médiane, la concavité maximale étant située au niveau de la hauteur maximale ; angles latéraux absents ; flancs lisses ; valve gauche plus grande que la droite.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7, GZ7f1, GZ7g), Hauts Plateaux méridionaux, Maroc.
Famille Notodromadidae Kaufmann, 1900
Sous-Famille Cyproidinae Hartmann, 1963
Genre Cyprois Zenker, 1854
Cyprois ? sp.
(Pl. 3 ![]() , figs. 13-16)
, figs. 13-16)
Matériel : 4 carapaces.
Dimensions en mm : L = 0,44-0,53 ; h = 0,33-0,41.
Description sommaire : espčce appartenant avec doutes au genre Cyprois, sub-carrée en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers l'avant, située au niveau de la mi-hauteur, trčs haute ; extrémité postérieure arrondie, dissymétrique, tournée vers le bas, et située juste au-dessus du bord ventral ; hauteur maximale ŕ peu prčs au niveau de la mi-longueur ; bord dorsal courbe, dissymétrique de part et d'autre de la hauteur maximale, la partie antérieure faisant un angle de quelques degrés avec l'horizontale, et la partie postérieure, un angle proche de 45 degrés ; bord ventral rectiligne ; angles antérieurs, dorsal et ventral, nets ; angles postérieurs absents ; flancs lisses ; la valve droite recouvre la gauche sur sa périphérie.
Paléoécologie : genre Pléistocčne-Récent dulçaquicole, pouvant accepter des eaux trčs légčrement oligohalines (Meisch, 2000).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7g, GZ7i), Hauts Plateaux méridionaux, Maroc.
Répartition paléogéographique du genre : généralement connu dans le Cénozoďque (Swain, 1999 ; Meisch, 2000). Il a été reconnu dans le Crétacé supérieur du Maroc (ce travail), dans le Maastrichtien d'Inde (Whatley et Bajpai, 2006 ; Khosla et al., 2011 ; Whatley, 2012), d'Alaska (Maastrichtien, Brouwers et De Deckker, 1993 ; Whatley et Bajpai, 2006), de Chine (Whatley, 2012) et d'Amérique du Sud (Carignano et Varela, 2011). Hou et al. (2002) donnent la répartition suivante : Asie, Europe, Amérique du Nord, du Crétacé ŕ l'Actuel.
Genres indéterminés
Genre indét. sp. 1
(Pl. 3 ![]() , fig. 17)
, fig. 17)
Matériel : une carapace.
Dimensions en mm : L = 0,63 ; h = 0,30.
Description sommaire : espčce ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, située au niveau de la mi-hauteur ; extrémité postérieure arrondie, située au niveau de la mi-hauteur ; hauteur maximale ŕ l'avant, au 1/3 de la mi-longueur ; bord dorsal courbe, dissymétrique de part et d'autre de la hauteur maximale ; bord ventral convexe, la convexité maximale étant située ŕ l'avant, au niveau de la hauteur maximale ; angles latéraux absents ; flancs lisses ; valve gauche recouvrant la droite sur les parties antérieure et ventrale.
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (éch. GZ7), Hauts Plateaux méridionaux, Maroc.
Genre indét. sp. 2
(Pl. 3 ![]() , fig. 18)
, fig. 18)
Matériel : une carapace.
Dimensions en mm : L = 0,95 ; h = 0,51.
Description sommaire : espčce de grande taille, ovoďde en vue latérale externe ; extrémité antérieure réguličrement arrondie, tournée vers le bas, située au-dessous de la mi-hauteur ; extrémité postérieure triangulaire, tournée vers l'arričre, et située au niveau de l'antérieure ; hauteur maximale au niveau de la mi-longueur ; bord dorsal courbe, symétrique de part et d'autre de la hauteur maximale ; bord ventral légčrement concave ; angles latéraux absents ; flancs lisses ; valves subégales (?).
Répartition stratigraphique : Crétacé supérieur du membre supérieur de la Formation du Tigri (échantillons GZ7, GZ7f1, GZ7g), Hauts Plateaux méridionaux, Maroc.
Vingt-trois espčces ont été reconnues, appartenant ŕ 12 genres nominés et deux genres indéterminés. Les déterminations sont réalisées sur la base des ressemblances morphologiques au niveau générique surtout ; pour cette raison, la plupart des espčces sont laissées en nomenclature ouverte. Les 12 genres reconnus sont les suivants : GZ7 (90 Carapaces) : Candoniella ?, Cypridea ?, Damonella, Lycopterocypris ? ; GZ7b (20 C) : Damonella, Eucypris ; GZ7c (90 C) : Damonella, Eucypris, Harbinia, Vecticypris ; GZ7d (15 C) : Damonella, Eucypris, Ilyocypris, Paracypria ; GZ7f1 (110 C) : Candoniella ?, Damonella, Mantelliana ?, Neocyprideis ; GZ7f2 (50 C) : Eucypris ; GZ7g (70 C) : Candona, Cyprois, Damonella, Perissocytheridea ; GZ7i (30 C) : Candoniella ?, Cyprois, Harbinia, Vecticypris ; GZ7lat (10 C) : Damonella.
Les associations d'ostracodes présentent peu d'intéręt stratigraphique dans l'état actuel des connaissances au Maroc ; elles caractériseraient le Crétacé supérieur (?).
Les barres gréseuses abondantes sur l'ensemble de la Formation du Tigri constituent des remplissages de chenaux anastomosés divaguant sur une plaine d'inondation ŕ facičs silteux ou argilo-pélitique. La présence d'épisodes marneux dans la partie supérieure de la Formation traduit l'installation d'une sédimentation limnique temporaire ŕ charophytes et ostracodes (Haddoumi et al., 2015).
La
plupart des genres d'ostracodes récoltés sont caractéristiques de milieu dulçaquicole,
avec une salinité de 0 ŕ 0.5 ‰, voire oligohalin, salinité de 0.5 ŕ 5 ‰ (Fig. 3 ![]() , d'aprčs Szczechura,
1971 ; Maddocks et al.,
1993 ; Swain, 1999 ; Meisch,
2000 ; Do
Carmo et al., 2008, 2013 ; Andreu et al.,
2003 ; Haddoumi et al.,
2008 ; Mojon
et al., 2009 ; Mebrouk et
al., 2011 ; Nicolaidis et al.,
2011 ; Antonietto et al.,
2012).
, d'aprčs Szczechura,
1971 ; Maddocks et al.,
1993 ; Swain, 1999 ; Meisch,
2000 ; Do
Carmo et al., 2008, 2013 ; Andreu et al.,
2003 ; Haddoumi et al.,
2008 ; Mojon
et al., 2009 ; Mebrouk et
al., 2011 ; Nicolaidis et al.,
2011 ; Antonietto et al.,
2012).
Les associations d'ostracodes non-marins, charophytes et radioles d'oursins parfois abondants, soulignent la proximité de la côte et un mélange d'eaux douces et d'eaux salées (Haddoumi et al., 2015).
|
Figure 3 : Intervalles de salinité, suivant la classification des salinités de Venise (1958), des genres d'ostracodes reconnus dans le Crétacé supérieur du membre supérieur de la Formation du Tigri, Hauts Plateaux méridionaux, Maroc oriental. Figure 3: Salinity ranges of the ostracode genera, according to the Venice Salinity Classification (1958), recognized in the Upper Cretaceous of the upper member of the Tigri Formation, southern High Plateaus, eastern Morocco. |
Les répartitions paléobiogéographiques des genres ne sont fournies, généralement, que pour le Crétacé, avec un minimum de références bibliographiques, les plus générales et les plus complčtes.
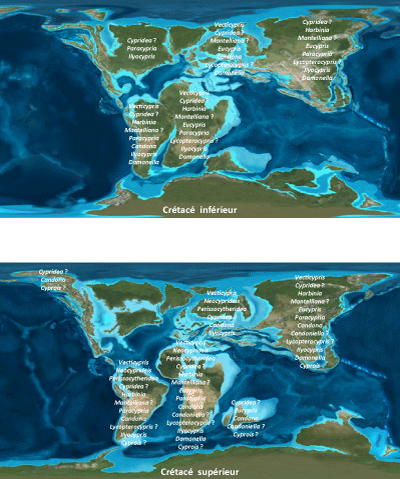
Nous
avons reconnu, dans le Crétacé supérieur du Maroc (ce travail), 14 genres
(incluant 23 espčces dont les individus sont mal conservés, usés et déformés),
de milieu dulçaquicole ŕ oligohalin, dont la répartition sur le globe est
fournie sur les Figures 4 ![]() - 5
- 5 ![]() ; parmi ces
genres, 9 sont déjŕ présents au Crétacé inférieur.
; parmi ces
genres, 9 sont déjŕ présents au Crétacé inférieur.
|
Figure
4 : Répartition des genres
d'ostracodes continentaux sur les deux supercontinents (Laurasia avec Alaska, Amérique
du Nord, Europe et Asie ; Gondwana avec Amérique du Sud, Afrique et Inde) au Crétacé
inférieur et supérieur (le Crétacé indifférencié est compté ŕ la fois
pour le Crétacé inférieur et supérieur). Figure 4: Distribution of continental ostracode genera on both supercontinents (Laurasia with Alaska, North America, Europe and Asia; Gondwana with South America, Africa and India) in the Early and Late Cretaceous (undifferentiated Cretaceous includes both Early and Late Cretaceous). |
|
Figure 5 : Nombre de genres d'ostracodes continentaux présents sur les deux supercontinents (Laurasia avec Alaska, Amérique du Nord, Europe et Asie ; Gondwana avec Amérique du Sud, Afrique et Inde) au Crétacé (le Crétacé indifférencié est compté ŕ la fois pour le Crétacé inférieur et pour le Crétacé supérieur). Figure 5: Number of continental ostracode genera present on both supercontinents (Laurasia with Alaska, North America, Europe and Asia; Gondwana with South America, Africa and India) in the Cretaceous (undifferentiated Cretaceous includes both the Early Cretaceous and the Late Cretaceous). |
Les diversités générique et spécifique sont moyennes. Pour donner un point de comparaison, Whatley (2012) a relevé, dans la littérature, 39 genres non-marins, incluant 133 espčces, dans son travail sur le Maastrichtien de l'Inde ; il a recensé, au total, 72 genres non-marins, dans le Maastrichtien du globe.
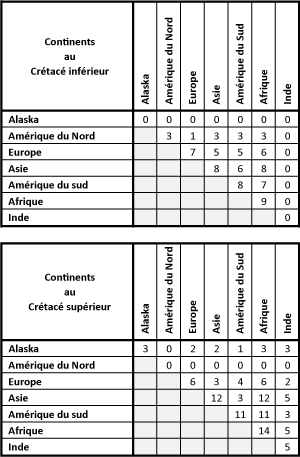
Au
Crétacé inférieur (Figs. 4 ![]() - 5
- 5 ![]() et
7
et
7 ![]() ), sur la Laurasia, 3 genres sont présents en Amérique du Nord (Cypridea
?, Paracypria, Ilyocypris), 7 en Europe (Vecticypris,
Cypridea ?, Mantelliana ?, Eucypris, Candona,
Lycopterocypris ?, Damonella)
et 8 en Asie (Cypridea ?, Harbinia,
Mantelliana ?, Eucypris, Paracypria,
Lycopterocypris ?, Ilyocypris
et Damonella). Sur le Gondwana, on
compte 8 genres en Amérique du Sud (Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Paracypria,
Candona, Ilyocypris et Damonella),
et 9 genres en Afrique (au Maroc ; Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Lycopterocypris
?, Ilyocypris et Damonella).
Le nombre de genres communs entre les différents continents est fourni sur la Figure 6
), sur la Laurasia, 3 genres sont présents en Amérique du Nord (Cypridea
?, Paracypria, Ilyocypris), 7 en Europe (Vecticypris,
Cypridea ?, Mantelliana ?, Eucypris, Candona,
Lycopterocypris ?, Damonella)
et 8 en Asie (Cypridea ?, Harbinia,
Mantelliana ?, Eucypris, Paracypria,
Lycopterocypris ?, Ilyocypris
et Damonella). Sur le Gondwana, on
compte 8 genres en Amérique du Sud (Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Paracypria,
Candona, Ilyocypris et Damonella),
et 9 genres en Afrique (au Maroc ; Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Lycopterocypris
?, Ilyocypris et Damonella).
Le nombre de genres communs entre les différents continents est fourni sur la Figure 6 ![]() ; il varie de 1 ŕ 5 sur la Laurasia et de 3 ŕ 8 sur le
Gondwana.
; il varie de 1 ŕ 5 sur la Laurasia et de 3 ŕ 8 sur le
Gondwana.
Au
Crétacé supérieur (Figs. 4 ![]() - 5
- 5 ![]() et 7
et 7 ![]() ), sur la Laurasia, 3 genres sont reconnus en Alaska (Cypridea
?, Candona, Cyprois ?), 6 en Europe (Vecticypris,
Neocyprideis, Perissocytheridea, Cypridea
?, Candona, Ilyocypris), et 12 en Asie (Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Candona, Candoniella
?, Lycopterocypris ?, Ilyocypris,
Damonella et Cyprois ?). Sur le Gondwana, apparaissent 11 genres en Amérique du
Sud (Vecticypris, Neocyprideis,
Perissocytheridea, Cypridea
?, Harbinia, Mantelliana ?, Paracypria,
Candona, Lycopterocypris ?, Ilyocypris
et Cyprois ?), 14 genres en Afrique (Vecticypris,
Neocyprideis, Perissocytheridea,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Candona, Candoniella
?, Lycopterocypris ?, Ilyocypris,
Damonella et Cyprois ?) et 5 genres en Inde (Cypridea
?, Eucypris, Candona, Candoniella ? et Cyprois
?). Le nombre de genres communs entre les différents continents est fourni sur
la Figure 6
), sur la Laurasia, 3 genres sont reconnus en Alaska (Cypridea
?, Candona, Cyprois ?), 6 en Europe (Vecticypris,
Neocyprideis, Perissocytheridea, Cypridea
?, Candona, Ilyocypris), et 12 en Asie (Vecticypris,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Candona, Candoniella
?, Lycopterocypris ?, Ilyocypris,
Damonella et Cyprois ?). Sur le Gondwana, apparaissent 11 genres en Amérique du
Sud (Vecticypris, Neocyprideis,
Perissocytheridea, Cypridea
?, Harbinia, Mantelliana ?, Paracypria,
Candona, Lycopterocypris ?, Ilyocypris
et Cyprois ?), 14 genres en Afrique (Vecticypris,
Neocyprideis, Perissocytheridea,
Cypridea ?, Harbinia, Mantelliana ?, Eucypris,
Paracypria, Candona, Candoniella
?, Lycopterocypris ?, Ilyocypris,
Damonella et Cyprois ?) et 5 genres en Inde (Cypridea
?, Eucypris, Candona, Candoniella ? et Cyprois
?). Le nombre de genres communs entre les différents continents est fourni sur
la Figure 6 ![]() ; il varie de 2 ŕ 3 sur la Laurasia et de 1 ŕ 12 sur le
Gondwana.
; il varie de 2 ŕ 3 sur la Laurasia et de 1 ŕ 12 sur le
Gondwana.
Pendant le Crétacé, tout genre, déterminé au Maroc dans ce travail, est connu soit sur la Laurasia, soit sur le Gondwana, soit sur ces deux supercontinents. Les associations de genres sont globales et n'apparaissent donc pas comme caractéristiques d'un seul super continent. Le nombre de genres communs entre les cinq continents, Amérique du Nord - Europe - Asie, sur la Laurasia, Amérique du Sud - Afrique, sur le Gondwana, laisse ŕ penser ŕ des échanges aisés entre eux, soit par zoochorie (vertébrés, oiseaux, insectes), soit par anémochorie, soit le long de ponts continentaux, mais les voies de communications restent encore trčs floues. Ces échanges se réalisent d'autant plus aisément que le climat est tempéré chaud aux hautes latitudes et tropical ŕ aride aux basses latitudes, sur les deux supercontinents, au Crétacé inférieur et supérieur (Scotese, 2000), les températures élevées facilitant les migrations.
|
Figure 6 : Nombre de genres d'ostracodes continentaux communs entre les masses continentales (Alaska, Amérique du Nord, Europe, Asie, Amérique du Sud, Afrique, Inde), au Crétacé inférieur et supérieur. Figure 6: Number of common continental ostracode genera between landmasses (Alaska, North America, Europe, Asia, South America, Africa, India), in Early and Late Cretaceous. |
|
Figure 7 : Répartition globale des genres d'ostracodes continentaux reconnus au Maroc, sur les cartes paléogéographiques (Earth History) du Crétacé inférieur et supérieur, proposées par Scotese (2000). Figure 7: Global distribution of continental ostracode genera recognized in Morocco, noted on paleogeographic maps (Earth History) from the Early and Late Cretaceous, proposed by Scotese (2000). |
Les marnes de la Formation du Tigri, Hauts Plateaux méridionaux, Maroc oriental, ont livré 23 espčces d'ostracodes appartenant ŕ 14 genres.
Les barres gréseuses de la Formation du Tigri se sont déposées dans des chenaux anastomosés d'une plaine alluviale. Elles sont associées ŕ des argiles ou ŕ des marnes de plaine d'inondation et ŕ des dépôts limniques, comme en témoignent les associations d'ostracodes non-marins et de charophytes. La découverte ponctuelle de radioles d'oursins démontre des mélanges entre eaux douces et eaux salées lors d'évčnements exceptionnels et donc la proximité de la côte.
Dans l'état actuel des connaissances sur les ostracodes du Maroc, les associations caractériseraient le Crétacé supérieur (?).
Les genres d'ostracodes reconnus sont caractéristiques d'un milieu dulçaquicole ŕ oligohalin.
Leur répartition paléobiogéographique est globale, aussi bien au Crétacé inférieur qu'au Crétacé supérieur, et leurs associations ne caractérisent pas, particuličrement, la Laurasia ou le Gondwana. Les échanges d'ostracodes entre les deux supercontinents semblent faciles et favorisés par un climat chaud.
Les auteurs remercient M. Thierry Aigouy, CNRS, et Mme Sophie Gouy, GET, Toulouse, pour la réalisation des photographies au MEB, ainsi que les deux reviewers, Mme Cristina Cabral et M. Jacques Sauvagnat, pour leurs conseils avisés et remarques pertinentes.
Anderson F.W. (1967).- Ostracods from the Weald Clay of England.- Bulletin of the Geological Survey of Great Britain, vol. 27, p. 237-269.
Anderson F.W. & Barker D. (1966).- Some British Jurassic and Cretaceous Ostracoda.- Bulletin of the British Museum (Natural History), Geology, vol. 11, nº 9, p. 433-487.
Andreu B. (2002).- Cretaceous ostracode biochronology of Morocco.- Eclogae geologicae Helvetiae, vol. 95, p. 133-152.
Andreu B., Colin J.-P., Haddoumi M. & Charričre A. (2003).- Les ostracodes des "Couches Rouges" du Synclinal d'Aďt Attab, Haut Atlas Central, Maroc. Systématique, biostratigraphie, paléoécologie, paléobiogéographie.- Revue de Micropaléontologie, Paris, vol. 46, p. 193-216.
Andreu B. & Ettachfini El M. (1994).- Nouvelles espčces d'ostracodes du Cénomanien du Bassin d'Essaouira (Maroc). Implications paléoécologiques.- Revue de Micropaléontologie, Paris, vol. 37, nº 1, p. 3-29.
Antonietto L.S., Gobbo S.R., Do Carmo D.A., Assine M.L., Castanheira Cordeiro Fernandes M.A.M. & Lima E Silva J.E. (2012).- Taxonomy, ontogeny and paleoecology of two species of Harbinia Tsao, 1959 (Crustacea, Ostracoda) from the Santana Formation, Lower Cretaceous, Northeastern Brazil.- Journal of Paleontology, vol. 86, nº 4, p. 659-668.
Ayress M.A. & Whatley R.C. (2014).- Early Cretaceous non-marine ostracoda from the north Falkland Basin, South Atlantic.- Palaeontology, p. 1-33.
Bassiouni M.A.A. (2002).- Mid-Cretaceous (Aptian-early Turonian) Ostracoda from Sinai, Egypt.- Neue Paläontology Abhandlungen, Dresden, vol. 5, p. 1-123.
Bilotte M. & Andreu B. (2006).- Les Marnes d'Auzas (Maastrichtien supérieur sous-pyrénéen). Stratigraphie et paléoenvironnements, associations d'ostracodes.- Revista Espańola de Micropaleontología, Madrid, Volumen homenaje a Enrico Perconig, vol. 49, p. 55-73.
Brenner P. (1976).- Ostracoden und Charophyten des Spanischen Wealden.- Palaeontographica A, Stuttgart, vol. 152, p. 113-201.
Brouwers E.M. & De Deckker, P. (1993).- Late Maastrichtian and Danian ostracode faunas from Northern Alaska: Reconstructions of environnement and paleogeography.- Palaios, Lawrence, vol. 8, p. 140-154.
Cao M.-Z. (1996).- Nonmarine Cretaceous Ostracods from Inner Zone of SW Japan.- Bulletin Kitakyushu Museum Natural History, vol. 15, p. 13-30.
Cabral M.C. (1995).- Ostracodos do Cretácico inferior do Algarve e da Regiăo de Lisboa : Sistemática, biostratigrafia, aspectos paleoecológicos e paleobiogeográficos.- Unpublished Ph.D. Thesis, University of Lisbon, Portugal, 137 p.
Cabral M.C., Colin J.-P. & Azerędo A.C. (2008).- Taxonomy and palaeoecology of new brackish ostracod species from the middle Cenomanian of Lousa, Lisbon region, Portugal.- Palæogeography, Palæoclimatology, Palæoecology, vol. 264, p. 250-262.
Carignano A.P. & Varela J.A. (2011).- Ostrácodos (Crustacea) de la Formación Allen (Cretácico tardío), Cuenca Neuquina, Argentina.- Revista Brasileira de Paleontologia, Porto Alegre, vol. 14, nº 2, p. 169-178.
Choubert G. (1956).- Lexique stratigraphique du Maroc (en collaboration avec A. Faure-Muret).- Notes et Mémoires du Service Géologique du Maroc, t. 134, p. 1-164.
Choubert G. & Faure-Muret A. (1960-62).- Évolution du domaine atlasique marocain depuis les temps paléozoďques. In : Livre ŕ la mémoire du Professeur Paul Fallot.- Mémoire hors-série de la Société Géologique de France, Paris, t. 1, p. 447-527.
Colin J.-P. (1974).- Nouvelles espčces des genres Metacypris et Theriosynoecum (Ostracodes lacustres) dans le Cénomanien de Dordogne (S.O. France).- Revista Espańola de Micropaleontología, Madrid, vol. VI, nº 2, p. 183-189.
Colin J.-P. & Dépęche F. (1997).- Faunes d'ostracodes lacustres des bassins intra-cratoniques d'âge albo-aptien en Afrique de l'Ouest (Cameroun, Tchad) et au Brésil : Considérations d'ordre paléoécologique et paléographique.- Africa Geoscience Review, Malakoff, vol. 4, p. 431-450.
Colin J.-P. & Carbonel P. (1990a).- Phylogenetical affinities of Cytherissa with other Cytherideinae (Vernoniella, Fabanella, Neocyprideis, Cyprideis). A paleontological approach. In : Danielopol D.L., Carbonel P. & Colin J.-P. (eds.), Cytherissa (Ostracoda) - the Drosophila of Paleolimnology.- Bulletin de l'Institut de Géologie du Bassin d'Aquitaine, Talence, vol. 47-48, p. 83-96.
Colin J.-P., Carbonel P. & Olteanu R. (1990b).- A review of the paleobiogeography and paleoecology of the closest group of Cytherissa : from the Mesozoďc Fabanella and Vernoniella to the Cenozoďc Cyprideis. In : Danielopol D.L., Carbonel P. & Colin J.-P. (eds.), Cytherissa (Ostracoda) - the Drosophila of Paleolimnology.- Bulletin de l'Institut de Géologie du Bassin d'Aquitaine, Talence, vol. 47-48, p. 119-134.
Colin J.-P., Brunet M., Congleton J.D., Dejax J., Flynn L.J., Hell J. & Jacobs L. (1992).- Ostracodes lacustres des bassins d'âge Crétacé inférieur du nord Cameroun : Hama-Koussou, Koum et Babouri-Figuil.- Revue de Paléobiologie, Genčve, vol. 11, nº 2, p. 357-372.
Colin J.-P. & Oertli H.J. (1985).- Atlas des ostracodes de France : Purbeckien.- Bulletin des Centres de Recherche Exploration-Production elf-Aquitaine, Pau, Mémoire 9, p. 147-161.
Colin J.-P., Tambareau Y. & Krasheninikov V.A. (1996).- Ostracodes limniques et lagunaires dans le Crétacé supérieur du Mali (Afrique de l'Ouest) : Systématique, paléoécologie et affinités paléobiogéographiques.- Revue de Micropaléontologie, Paris, vol. 39, nº 3, p. 211-222.
Colin J.-P. & Tabuce R. (2004).- Ostracodes limniques de la Formation d'Irbzer, Crétacé terminal du Moyen Atlas, Maroc : Taxinomie, biostratigraphie, paléoécologie, paléobiogéographie.- Revue de Micropaléontologie, Paris, vol. 47, p. 103-109.
Dequan Y. (1990).- The geological and exploration significance of Cretaceous non-marine Ostracoda from the Hailaer Basin, northwestern China. In : Whatley R. & Maybury C. (eds.), Ostracoda and global events.- Chapman and Hall, London, p. 251-259.
Dias-Brito D., Musacchio E.A., Maranhăo M.S.A.S., Suárez J.M. & Rodrigues R. (1998).- Cretaceous non-marine calcareous microfossils from the Adamantina Formation (Bauru Group), Western Săo Paulo, Brasil. In : 3th Annual Conference, Project 381 (IUCP-IUGS), Annual Conference of the South Atlantic Mesozoic Correlation Project 381-3, Argentina, 1998.- Abstracts, IGCP-IUGS, Boletim 2 (Edición Especial), Comodoro Rivadavia, p. 8-10.
Dias-Brito D., Musacchio E.A., Castro J.C. de, Maranhăo M.S.A.S., Suárez J.M. & Rodrigues R. (2001).- The Bauru group: A continental Cretaceous unit in Brazil - Concepts based on micropaleontological, oxygen isotope and stratigraphical data.- Revue de Paléobiologie, Genčve, vol. 20, nº 1, p. 245-304.
Do Carmo D.A., Coimbra J.C., Whatley R.C., Antonietto L.S. & De Paiva Citon R.T. (2013).- Taxonomy of limnic Ostracoda (Crustacea) from the Alagamar Formation, Middle-Upper Aptian, Potiguar Basin, Northeastern Brazil.- Journal of Paleontology, vol. 87, nº 1, p. 91-104.
Do Carmo D.A., Whatley R.C., De Queiroz Neto J. V. & Coimbra J. C. (2008).- On the validity of two Lower Cretaceous non-marine ostracode genera: Biostratigraphic and paleogeographic implications.- Journal of Paleontology, vol. 82, p. 790-799.
Donze P. (1964).- Ostracodes berriasiens des Massifs Subalpins septentrionaux (Bauges et Chartreuse).- Travaux du Laboratoire de la Faculté des Sciences de Lyon, Villeurbanne, (N.S.), vol. 11, p. 103-158.
Donzeau M., Wernli R., Charollais J. & Monjuvent G. (1997).- Feuille Saint-Julien-en-Genevois (653). Notice explicative.- Carte géologique de la France (1/50 000), BRGM, Orléans, 144 p.
Gautier E.F. (1914).- Un volcan pliocčne au chott Tigri.- Comptes Rendus de l'Académie des Sciences de Paris, vol. 159, nº 15, p. 580-582.
Grékoff N. (1957).- Ostracodes du Bassin du Congo. I Jurassique supérieur et Crétacé inférieur du nord du bassin.- Annales du Musée Royal du Congo Belge, Tervuren, vol. 19, p. 1-97.
Grékoff N. (1960).- Ostracodes du Bassin du Congo. II Crétacé.- Annales du Musée Royal du Congo Belge, Tervuren, vol. 35, p. 1-70.
Grosdidier E. (1967).- Quelques Ostracodes nouveaux de la série anté-salifčre ("wealdienne") des bassins côtiers du Gabon et du Congo.- Revue de Micropaléontologie, Paris, vol. 10, nº 2, p. 107-118.
Haddoumi H., Charričre A., Andreu B. & Mojon P.-O. (2008).- Les dépôts continentaux du Jurassique moyen au Crétacé inférieur dans le Haut Atlas oriental, Maroc. Paléoenvironnements successifs et signification paléogéographique.- Carnets Geol., Madrid, vol. 8, nº A06 (CG2008_A06), p. 1-29.
Haddoumi H., Mebrouk F., Andreu B., Tabuce R., Vianey-Liaud M., Capetta H., Bensaad K. & Charričre A. (2015).- Nouvelles données sédimentologiques et micropaléontologiques (charophytes, ostracodes), sur la Formation du Tigri (Sénonien des Hauts Plateaux méridionaux, Maroc oriental) ; paléoenvironnements et évolution paléogéographique.- Revue de Paléobiologie, Genčve, vol. 34, nº 1, p. 85-111.
Horne D. J. & Colin J.-P. (2005).- The affinities of the ostracod genus Cypridea Bosquet, 1852, and its allies, with consideration of implications for the phylogeny on non-marine cypridoidean ostracods.- Revue de Micropaléontologie, Paris, vol. 48, p. 25-34.
Hou Y.-T., Gou Y. & Chen D.-Q. (2002).- Fossil Ostracoda of China Vol. 1- Superfamilies Cypridacea and Darwinulidacea.- Science Press, Beijing, 1090 p. (en chinois).
Keen M.C. (1972).- The Sannoisian and some other Upper Palaeogene ostracoda from north-west Europe.- Palaeontology, vol. 15, nº 2, p. 267-325.
Khosla S.C., Rathore A.S., Nagori M.L. & Jakhar S.R. (2011).- Non-marine Ostracoda from the Lameta Formation (Maastrichtian) of Jabalpur, Madhya Pradesh and Nand-Dongaraon Basin, Mararashtra.- Revista Espańola de Micropaleontología, Madrid, vol. 43, nº 3, p. 209-260.
Khosla A. & Sahni A. (2000).- Late Cretaceous (Maastrichtian) Ostracodes from the Lameta Formation, Jabalpur Cantonment Area, Madhya Pradesh, India.- Journal of the Palaeontological Society of India, Lucknow, vol. 45, p. 57-78.
Krömmelbein K. & Weber R. (1971).- Ostracoden des "Nordost-Brasilianischen wealden".- Beihefte zum Geologischen Jahrbuch, Hannover, vol. 115, 93 p.
Lyubimova P.S. (1955).- Ostracods from the Mesozoic sediments of the Middle Volga Region and Obshchii Syrt Plateau. In : Ostrakody mezozoiskikh otlozhenii Volgo Ural'skoi oblasti (Ostracods from the Mesozoic sediments of the Volga-Ural Region).- Trudy Vsesoyuznogo Neftyanogo Nauchno-Issledovatelskogo Geologo - Razvedochnogo Instituta (VNIGRI), Leningrad, (Novaya Seriya), vol. 84, p. 3-190 (en russe).
Lyubimova P.S. (1956).- Ostracody Melovykh Otlozheniy Vostochnoy Chasti Mongolskoy Narodnoy Respubliki i ikh Znachenie dlya Stratigrafii (Ostracoda from Cretaceous deposits of the eastern part of the People's Republic of Mongolia and their significance for stratigraphy).- Trudy Vsesoyuznogo Neftyanogo Nauchno-Issledovatelskogo Geologo - Razvedochnogo Instituta (VNIGRI), Leningrad, (Novaya Seriya), vol. 93, p. 1-174 (en russe).
Mandelstam M.I (1956).- Order Ostracoda. In : Mandelstam M.I, Shneyder G.F. & Zanina J.E. (eds.), New families and genera.- All-Union Scientific Research, Geological Institute (VSEGEI), Moscow, vol. 12, p. 87-144 (en russe).
Maddocks R.M. (1982).- Ostracoda. In : Bliss D.E. (ed.), The Biology of Crustacea. Vol. 1, Systematics, the fossil record and biogeography.- Academic Press, New-York, p. 221-239.
Maddocks R.F., Iliffe T.M. & Sarbu S. (1993).- Anchialine podocopid Ostracoda of New Caledonia. In : McKenzie K.G. & Jones P.J. (eds.), Ostracoda in the earth and life sciences.- A.A. Balkema, Rotterdam, p. 439-450.
Martin G.P.R. (1940).- Ostracoden des norddeutschen Purbeck und Wealden.- Senckenbergiana, Frankfurt am Main, vol. 22, nº 5-6, p. 275-361.
Mebrouk F., Colin J.-P. & Hennache F. (2011).- Un gisement d'ostracodes non-marins dans l'Éocčne inférieur du Djebel Amour, Atlas saharien central, Algérie : Taxonomie, paléoécologie et paléogéographie.- Carnets Geol., Madrid, vol. 11, nº A04 (CG2011_A04), p. 83-96.
Médioni R. (1970).- Le Chott Tigri (Hauts-Plateaux du Maroc oriental) : Stratigraphie, tectonique et évolution morphogénique ŕ la lumičre d'observations nouvelles.- Notes et Mémoires du Service géologique du Maroc, vol. 30, nº 225, p. 11-20.
Meisch C. (2000).- Freshwater Ostracoda of Western and Central Europe.- Spektrum Akademischer Verlag, Berlin, 522 p.
Mojon P.-O., Haddoumi H. & Charričre A. (2009).- Nouvelles données sur les Charophytes et Ostracodes du Jurassique moyen-supérieur - Crétacé inférieur de l'Atlas marocain.- Carnets Geol., Madrid, vol. 9, nº M03 (CG2009_M03), p. 1-39.
Musacchio E.A. (1973).- Charophytas y ostrácodos no marinos del Grupo Neuquén (Cretácico Superior) en algunos afloramientos de las provincias de Río Negro y Neuquén, República Argentina.- Revista del Museo de La Plata, (nueva serie), Paleontología, vol. 8, nº 48, p. 1-37.
Musacchio E.A. (1981).- South American Jurassic and Cretaceous Foraminifera, Ostracoda and Charophyta, of Andean and sub-Andean regions. In : Volkheimer W. & Musacchio E.A. (eds.), Cuencas sedimentarias del Jurásico y Cretácico de America del Sur.- VIII Congreso Geológico Argentino, Comité Sudamericano del Jurásico y Cretácico, Buenos Aires, vol. 2, p. 461-498.
Musacchio E.A. (1990).- Non-marine Cretaceous ostracods from Argentina and their palaeobiogeographical relationships. In : Whatley R. & Maybury C. (eds.), Ostracoda and global events.- Chapman and Hall, London, p. 557-569.
Musacchio E.A. & Simeoni M. (1989).- Cretaceous non-marine cypridacean Ostracoda from central and northern Argentine Patagonia.- Courier Forschungsinstitut Senckenberg, Frankfurt am Main, vol. 113, p. 77-88.
Neale J.W. (1988).- Ostracods and paleosalinity reconstruction.- In : De Deckker P., Colin J.-P. & Peypouquet J.-P. (eds.), Ostracoda in Earth Sciences.- Elsevier, Amsterdam, p. 125-156.
Nicolaidis D.D., Piovesan E.K., Fauth G. & Viviers M.C. (2011).- Non-marine, transitional and marine ostracode associations from the NeoAptian-Turonian of Santos Basin, Brazil.- Paleontologia : Cenŕrios de vida, Rio de Janeiro, p. 301-312.
Piovesan E.K., Cabral M.C., Colin J.-P., Fauth G. & Trecastro Bergue C. (2014a).- Ostracodes from the Upper Cretaceous deposits of the Potiguar Basin, northeastern Brazil : Taxonomy, paleoecology and paleobiogeography. Part 1: Turonian.- Carnets Geol., Madrid, vol. 14, nº 12, p. 211-252.
Piovesan E.K., Cabral M.C., Colin J.-P., Fauth G. & Trecastro Bergue C. (2014b).- Ostracodes from the Upper Cretaceous deposits of the Potiguar Basin, northeastern Brazil : Taxonomy, paleoecology and paleobiogeography. Part 2: Santonian-Campanian.- Carnets Geol., Madrid, vol. 14, nº 15, p. 315-351.
Poropat S.F. & Colin J.-P. (2012).- Early Cretaceous ostracod biostratigraphy of eastern Brazil and western Africa: An overview.- Gondwana Research, vol. 22, p. 772-798.
Rodríguez-Lázaro J. & Martín-Rubio M. (2005).- Pliocene Ilyocyprididae (Ostracoda) from the Ebro Basin (N. Spain).- Revue de Micropaléontologie, Paris, vol. 48, p. 37-49.
Rosenfeld A. & Raab M. (1984).- Lower Cretaceous Ostracodes from Israel and Sinaď.- Israel Journal of Earth Sciences, vol. 33, nº 3, p. 85-134.
Schudack U. (1994).- Revision, Dokumentation und Stratigraphie der Ostracoden des nordwestdeutschen Oberjura und Unter-Berriasium.- Berliner Geowissenschaftliche Abhandlungen, (Reihe E), vol. 11, p. 1-193.
Scotese C. (2000).- Paleomap Project : http://www.scotese.com/ ; Earth History : http://www.scotese.com/earth.htm ; Climate History : http://www.scotese.com/climate.htm.
Swain F.M. (1999).- Fossil non-marine Ostracoda of the United States.- Developments in Palaeontology and Stratigraphy, Amsterdam, vol. 16, 401 p.
Szczechura J. (1971).- Fresh-water Ostracoda from the Paleocene of the Nemegt Basin, Gobi Desert, Mongolia. In : Kielan-Jaworowska Z. (ed.), Results of the Polish-Mongolian palaeontological expeditions, Part III.- Palaeontologia polonica, Warszawa, vol. 25, p. 85-97.
Szczechura J. (1978).- Results of the Polish-Mongolian palaeontological expeditions - Part VIII. Fresh-water ostracodes from the Nemegt Formation (Upper Cretaceous) of Mongolia.- Palaeontologia Polonica, Warszawa, vol. 38, p. 65-121.
Szczechura J. & Blaszyk J. (1969).- Fresh-water ostracoda from the Upper Cretaceous of the Nemegt Basin, Gobi Desert.- Palaeontologia Polonica, Warszawa, vol. 21, p. 107-124.
Tomé M.E.T.R. & Filho M.F. de L. (2010).- Ostracodes da Bacia de Cedro, Andar Alagoas, Estado de Pernambuco, NE Brasil : Implicacoőes paleoambientais e bioestratigráficas.- Estudos Geológicos, Recife, vol. 20, nº 2, p. 49-63.
Tomé M.E.T.R., Filho M.F. de L. & Neumann V.H.M.L. (2014).- Taxonomic studies of non-marine ostracods in the Lower Cretaceous (Aptian-Lower Albian) of post-rift sequence from Jatobá and Araripe basins (Northeast Brazil): Stratigraphic implications.- Cretaceous Research, vol. 48, p. 153-176.
Trabelsi K. & Colin J.-P. (2013).- Découverte d'ostracodes limniques dans l'Albien de la Formation Kebar de Tunisie centrale.- Revue de Paléobiologie, Genčve, vol. 32, nº 2, p. 531-540.
Trabelsi K., Sames B., Salmouna A., Piovesan E.K., Ben Rouina S., Houla Y., Touir J. & Soussi M. (2015).- Ostracods from the marginal coastal Lower Cretaceous (Aptian) of the Central Tunisian Atlas (North Africa): Paleoenvironment, biostratigraphy and paleobiogeography.- Revue de Micropaléontologie, Paris, vol. 58, nº 4, p. 309-331.
Uliana M.A. & Musacchio E.A. (1978).- Microfosiles calcareos no marinos del Cretacico Superior en El Zampal, Provincia de Mendoza, Argentina.- Ameghiniana, Buenos Aires, vol. 15, p. 11-135.
Viviers M.C., Koutsoukos E.A.M., Silva-Telles A. Jr & Bengtson P. (2000).- Stratigraphy and biostratigraphic affinities of the Late Aptian-Campanian ostracods of the Potiguar and Sergipe basins in northeastern Brazil.- Cretaceous Research, vol. 21, p. 407-455.
Whatley R.C. (2012).- The "out of India" hypothesis: Further supporting evidence from the extensive endemism of Maastrichtian non-marine ostracoda from the Deccan volcanic region of peninsular India.- Revue de Paléobiologie, Genčve, Vol. spéc. 11, p. 229-248.
Whatley R. & Bajpai S. (2006).- Some aspects of the palaeoecology and distribution of non-marine Ostracoda from Upper Cretaceous Intertrappean deposits of the Lameta Formation of Peninsular India.- Journal of the Palaeontological Society of India, Lucknow, Golden Jubilee Volume 50, p. 61-76.
Les photographies ont été prises par T. Aigouy, CNRS, et S. Gouy, GET, Toulouse, ŕ l'aide du JEOL 6360 LV de l'Université Paul Sabatier, Toulouse III. Les grossissements sont fournis ŕ 5 % prčs.
The photographs were taken by T. Aigouy, CNRS, and S. Gouy, GET, Toulouse, with the JEOL 6360 LV of the Paul-Sabatier University, Toulouse III, France. The enlargements are carried out with a factor of about 5%.
Abréviations : C = Carapace, VD = Valve Droite, VG = Valve Gauche, éch. = échantillon.
Abbreviations: C = Carapace, VD = Right Valve, VG = Left Valve, éch. = sample.
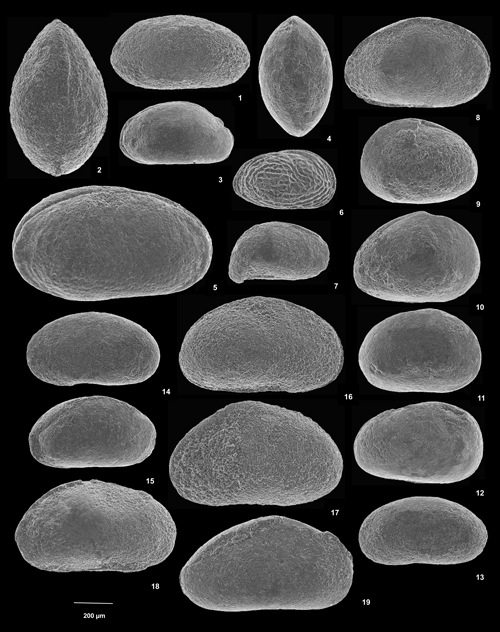
Cliquer sur la vignette pour agrandir l'image.
Planche 1 : 1-2) Vecticypris sp. 1. 1) C vue de gauche, éch. GZ7c ; 2) C vue dorsale, éch. GZ7c. 3-4) Vecticypris sp. 2. 3) C vue de gauche, éch. GZ7i ; 4) C vue dorsale, éch. GZ7i. 5) Neocyprideis sp. C vue de droite, éch. GZ7f1. 6) Perissocytheridea sp. C vue de droite, éch. GZ7g. 7) Cypridea ? sp. C vue de gauche, éch. GZ7. 8) Harbinia sp. 1. C vue de droite, éch. GZ7c. 9-11) Harbinia sp. 3. 9) C vue de gauche, éch. GZ7i ; 10) C vue de droite, éch. GZ7i ; 11) C vue de droite, éch. GZ7i. 12) Harbinia sp. 2. C vue de droite, éch. GZ7c. 13-15) Mantelliana ? sp. 13) C vue de gauche, éch. GZ7f1 ; 14) C vue de gauche, éch. GZ7f1 ; 15) C vue de gauche, légčrement écrasée latéralement, éch. GZ7f1. 16-19) Eucypris sp. 1. 16) C vue de gauche, éch. GZ7f2 ; 17) C vue de gauche, éch. GZ7b ; 18) C vue de gauche, légčrement écrasée latéralement, éch. GZ7c ; 19) C vue de droite, légčrement écrasée latéralement, éch. GE7c [Échelle graphique 200 µm].
Plate 1: 1-2) Vecticypris sp. 1. 1) C left view, sample GZ7c; 2) C dorsal view, sample GZ7c. 3-4) Vecticypris sp. 2. 3) C left view, sample GZ7i; 4) C dorsal view, sample GZ7i. 5) Neocyprideis sp. C right view, sample GZ7f1. 6) Perissocytheridea sp. C right view, sample GZ7g. 7) Cypridea ? sp. 7 C left view, sample GZ7. 8 Harbinia sp. 1. 8) C right view, sample GZ7c. 9-11) Harbinia sp. 3. 9) C left view, sample GZ7i; 10) C right view, sample GZ7i; 11) C right view, sample GZ7i. 12) Harbinia sp. 2. C right view, sample GZ7c. 13-15) Mantelliana ? sp. 13) C left view, sample GZ7f1; 14) C left view, sample GZ7f1; 15) C left view, slightly compressed laterally, sample GZ7f1. 16-19) Eucypris sp. 1. 16) C left view, sample GZ7f2; 17) C left view, sample GZ7b; 18) C left view, slightly compressed laterally, sample GZ7c; 19) C right view, slightly compressed laterally, sample GE7c [Graphical scale 200 µm].
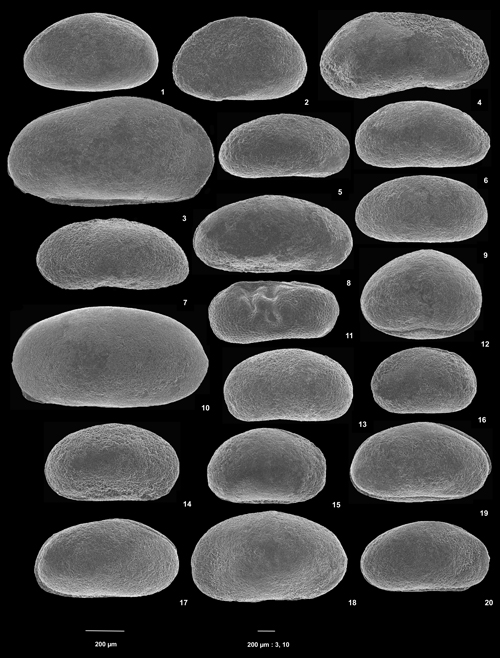
Cliquer sur la vignette pour agrandir l'image.
Planche 2 : 1-2) Eucypris sp. 2. 1) C vue de gauche, éch. GZ7d ; 2) C vue de gauche, éch. GZ7c. 3 Paracypria sp. 3) C vue de droite, éch. GZ7d. 4) Candona sp. C vue de gauche, éch. GZ7g. 5-9) Candoniella ? sp. 5) C vue de droite, éch. GZ7 ; 6) C vue de gauche, éch. GZ7 ; 7) C vue de gauche, éch. GZ7f1 ; 8) C vue de droite, éch. GZ7i ; 9) C vue de droite, éch. GZ7. 10) Lycopterocypris ? sp. C vue de gauche, éch. GZ7. 11) Ilyocypris sp. C vue de gauche, éch. GZ7d. 12) Damonella aff. buchaniana Anderson, 1966. C vue de droite, éch. GZ7. 13-16) Damonella sp. 3. 13) C vue de gauche ?, éch. GZ7 ; 14) C vue de droite, éch. GZ7g ; 15) C vue de droite, éch. GZ7f1 ; 16) C vue de gauche, éch. GZ7f1. 17-20) Damonella sp. 2. 17) C vue de droite, éch. GZ7f1 ; 18) C vue de gauche, éch. GZ7c ; 19) C vue de droite, éch. GZ7 ; 20) C vue de droite, éch. GZ7f1 [Échelle graphique 200 µm].
Plate 2: 1-2) Eucypris sp. 2. 1) C left view, sample GZ7d; 2) C left view, sample GZ7c. 3) Paracypria sp. 3) C right view, sample GZ7d. 4) Candona sp. C left view, sample GZ7g. 5-9) Candoniella ? sp. 5) C right view, sample GZ7; 6) C left view, sample GZ7; 7) C left view, sample GZ7f1; 8) C right view, sample GZ7i; 9) C right view, sample GZ7. 10) Lycopterocypris ? sp. C left view, sample GZ7. 11) Ilyocypris sp. C left view, sample GZ7d. 12) Damonella aff. buchaniana Anderson, 1966. C right view, sample GZ7. 13-16) Damonella sp. 3. 13) C left view ?, sample GZ7; 14) C right view, sample GZ7g; 15) C right view, sample GZ7f1; 16) C left view, sample GZ7f1. 17-20) Damonella sp. 2. 17) C right view, sample GZ7f1; 18) C left view, sample GZ7c; 19) C right view, sample GZ7; 20) C right view, sample GZ7f1 [Graphical scale 200 µm].
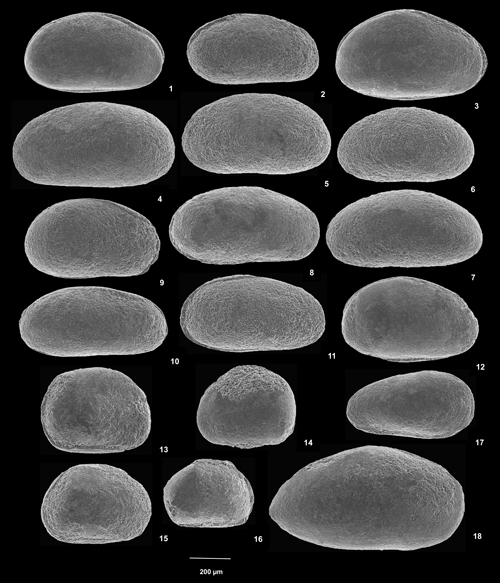
Cliquer sur la vignette pour agrandir l'image.
Planche 3 : 1-12) Damonella sp. 1. 1) C vue de droite, éch. GZ7d ; 2) C vue de gauche, éch. GZ7f1 ; 3) C vue de droite, éch. GZ7d ; 4) C vue de gauche, éch. GZ7c ; 5) C vue de gauche, éch. GZ7 ; 6) C vue de gauche, éch. GZ7f1 ; 7) C vue de gauche, éch. GZ7f1 ; 8) C vue de droite, éch. GZ7f1 ; 9) C vue de droite, éch. GZ7c ; 10) C vue de droite, éch. GZ7f1 ; 11) C vue de droite, éch. GZ7 ; 12) C vue de gauche, éch. GZ7d. 13-16) Cyprois ? sp. 13) C vue de gauche, éch. GZ7g ; 14) C vue de droite, éch. GZ7g ; 15) C vue de gauche, éch. GZ7g ; 16) C vue de gauche, éch. GZ7g. 17) Genre indét. sp. 1. C vue de droite, éch. GZ7. 18) Genre indét. sp. 2. C vue de droite, éch. GZ7i [Échelle graphique 200 µm].
Plate 3: 1-12) Damonella sp. 1. 1) C right view, sample GZ7d; 2) C left view, sample GZ7f1; 3) C right view, sample GZ7d; 4) C left view, sample GZ7c; 5) C left view, sample GZ7; 6) C left view, sample GZ7f1; 7) C left view, sample GZ7f1; 8) C right view, sample GZ7f1; 9) C right view, sample GZ7c; 10) C right view, sample GZ7f1; 11) C right view, sample GZ7; 12) C left view, sample GZ7d. 13-16) Cyprois ? sp. 13) C left view, sample GZ7g; 14) C right view, sample GZ7g; 15) C left view, sample GZ7g; 16) C left view, sample GZ7g. 17) Genus indet. sp. 1. C right view, sample GZ7. 18) Genus indet. sp. 2. C right view, sample GZ7i [Graphical scale 200 µm].