◄ Carnets Geol. 19 (16) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Microfacies]
[5. Facies distribution, micropaleontological assemblages, and biostratigraphy]
[6. Discussions and conclusions] and ...
[Bibliographic references]
Babeş-Bolyai University, Department of Geology, 1 M. Kogălniceanu str., 400084 Cluj-Napoca (Romania)
Babeş-Bolyai University, Department of Geology and Center for Integrated Geological Studies, 1 M. Kogălniceanu str., 400084 Cluj-Napoca (Romania)
Babeş-Bolyai University, Department of Geology, 1 M. Kogălniceanu str., 400084 Cluj-Napoca (Romania)
Corresponding author
Babeş-Bolyai University, Department of Geology, 1 M. Kogălniceanu str., 400084
Cluj-Napoca (Romania)
Published online in final form (pdf) on October 10, 2019
DOI 10.4267/2042/70499
![]()
[Editor: Bruno Granier; language editor:
Simon Mitchell]
![]()
The Hăgimaş Massif provides important data for reconstructing the geological evolution of the Transylvanian Carbonate Platform. This unit is present nowadays in the basement of the Transylvanian Depression and as large-scale olistoliths, which crop out in various locations in the Eastern Carpathians and Apuseni Mountains. They contain a large variety of microfacies as well as microfossil assemblages partly encompassing the Jurassic/Cretaceous boundary. This study presents a detailed account of uppermost Jurassic-lowermost Cretaceous limestones from the Lapoş Valley (Hăghimaş Mountains) containing two distinct successions separated by a fault. The first succession contains platform margin and inner platform deposits (subtidal, intertidal) whereas the second one consists of inner platform deposits (shallow-subtidal to peritidal carbonates). The Upper Jurassic Stramberk-type facies is overlain by lower Berriasian regressive peritidal limestones. The upper Berriasian-? lower Valanginian consists mainly of inner platform deposits. They contain alternating, high and low-energy carbonates with rare calpionellids and calcispheres. The age assignment is based on a rich and diverse micropaleontological assemblage containing similar biota reported from other regions of the Tethysian Realm.
• microfacies;
• algae;
• Foraminifera;
• depositional environments;
• Hăghimaș Massif;
• Romania
Neamţu O., Bucur I.I., Ungureanu R. & Mircescu C.V. (2019).- Upper Jurassic-Lower Cretaceous limestones from the Hăghimaș Massif (Eastern Carpathians, Romania): Microfacies, microfossils and depositional environments.- Carnets Geol., Madrid, vol. 19, no. 16, p. 345-368.
Les calcaires du Jurassique supérieur-Crétacé inférieur du Massif de Hăghimaș (Carpathes orientales, Roumanie) : Microfacičs, microfossiles et environnements de dépôt.- Le Massif de Hăghimaș fournit des données cruciales pour reconstruire l'évolution géologique de la plate-forme carbonatée transylvanienne. Cette unité est présente de nos jours dans le substratum de la dépression transylvanienne sous forme d'olistolites de grandes dimensions qui affleurent en de nombreuses localités des Carpathes orientales et des monts Apuseni. Ils comportent une grande variété de microfacičs, ainsi que des associations micropaléontologiques, qui couvrent en partie la limite Jurassique/Crétacé. Cette étude présente dans le détail un affleurement de calcaires du Jurassique terminal-Crétacé basal de la vallée de Lapos (Monts Hăghimaș) comportant deux séries distinctes séparées par une faille. La premičre série correspond ŕ des dépôts marginaux de plate-forme et de plate-forme interne (subtidaux, intertidaux), tandis que la seconde correspond ŕ des dépôts de plate-forme interne (carbonates peu profonds subtidaux ŕ péritidaux). Les facičs jurassiques supérieurs de type Stramberg sont surmontés par les calcaires péritidaux régressifs du Berriasien inférieur. Le Berriasien supérieur - ? Valanginien inférieur est principalement représenté par des calcaires de plate-forme interne. Ils comportent une alternance de carbonates de haute et de basse énergies avec de rares calpionelles et des calcisphčres. Leur datation se fonde sur un assemblage micropaléontologique riche et diversifié comportant des organismes semblables ŕ ceux décrits dans d'autres régions du domaine téthysien.
• microfacičs ;
• algues ;
• foraminifčres ;
• environments de dépôt ;
• Massif de Hăghimaș ;
• Roumanie
Carbonate microfacies
analysis, as well as micropaleontological associations play a key role in
defining depositional systems by applying sedimentological and paleontological
techniques (Flügel, 2010). This methodology was
applied to the Upper
Jurassic-Lower Cretaceous succession from the Hăghimaş Massif exposed
in a
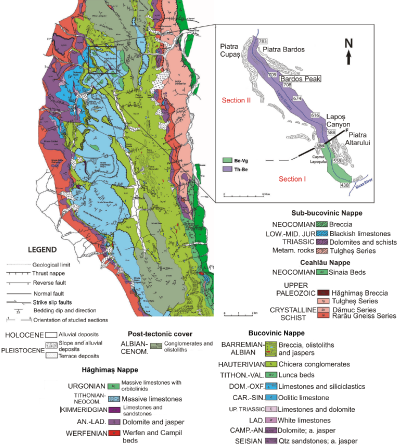
section located on the Lapoş Valley (a left tributary of the Bicaz Valley, Fig. 1 ![]() ).
).
The Hăhgimaş Massif has been studied by many authors since the 19th century (e.g., Herbich, 1866; Neumayr, 1873). It is worth mentioning the contributions of Săndulescu (1967, 1968, 1969, 1975, 1984), Dragastan (1969, 1975a, 1975b, 1981, 2011), Grasu et al. (2010), and D.A. Popescu and L.G. Popescu (2005). Some studies have focused on ammonite associations (Vadasz, 1915; Jekelius, 1921; Preda, 1973; Turculeţ, 1980; Grigore, 2010a, 2010b, 2011), dasycladalean algae (Dragastan, 1971, 1975a; Bucur & Săsăran, 2011; Bucur et al., 2011), foraminifers (Th. Neagu & M. Neagu, 1995), and brachiopods (Lazăr et al., 2011).
Recently, Dragastan (2011) studied the Upper Jurassic-Lower Cretaceous sedimentary succession from the Lapoș Valley, and identified a thin basal package of upper Tithonian limestones that is overlain by thicker, Berriasian-Hauterivian carbonates. Based on microfacies and micropaleontological studies the whole package was divided into three distinct subunits: SU1 - carbonate sandstones with limonite concretions, marly limestones and micrites with algal nodules towards its upper part; SU2 - micrites associated with a basal breccia; SU3 - nodular limestones with algal nodules and interbedded pelsparites and micrites.
Following Dragastan (2011), upper Tithonian limestones are present in the lowermost part of the succession. They contain foraminifers [Anchispirocyclina lusitanica (Egger), Everticyclammina virguliana (Koechlin), Coscinoconus alpinus Leupold, C. elongatus Leupold] and calpionellids [Crassicolaria brevis Remane, C. parvula Remane, Calpionella alpina Lorenz, and Tintinnopsella carpathica (Murgeanu & Filipescu)]. The "Jurassic/Cretaceous boundary" is placed in the basal part of the profile at the confluence with the Bicaz River. Dragastan (2011) mentions a hardground surface between subunits SU1 and SU2 marking the lower-upper Berriasian boundary based on the following microfossil associations: 1) a lower Berriasian assemblage with Calpionella alpina (Alpina B and C zones); 2) an upper Berriasian assemblage with dasycladalean algae [Rajkaella iailaensis (Maslov), Humiella sp. cf. H. sardiniensis (Ott & Flaviani)] and foraminifers [Coscinoconus campanellus (Arnaud-Vanneau et al.), C. cherchiae (Arnaud-Vanneau et al.), Charentia cuvillieri Neumann, Rumanoloculina pseudominima (Bartenstein & Kovatcheva), and Scythiloculina confusa Neagu]. Subunit SU3 is assigned by Dragastan (2011) to the lower Valanginian-Hauterivian interval. The base of this subunit is represented by another hardground surface. The age of the lower part of SU 3 was attributed by Dragastan (2011) to the Lower Valanginian based on the microfossil assemblage with foraminifers [Everticyclammina greigi (Henson); E. kelleri (Henson); Pseudocyclammina lituus (Yokoyama); P. sphaeroidalis Hottinger, Coscinoconus cherchiae, Neotrocholina molesta (Gorbatchik)], algae [Salpingoporella annulata Carozzi, Rajkaella alpina Dragastan, Humiella cataeneformis (Radoičić)], cyanobacteria [(Rivularia piae Frollo, Garwoodia bardosi Dragastan], and calpionellids [Calpionellites allemanni Rehanek and Tintinnopsella carpathica (Murgeanu & Filipescu)]. The middle part of the subunit was assigned to the upper Valanginian starting from the presence of abundant miliolid associations (Decussoloculina, Scythiloculina, and Rumanoloculina) and microbial structures (Lithocodium, Rivularia piae, Garwoodia bardosi). The upper part of SU3 was assigned by Dragastan (2011) to the Hauterivian based on the presence of Neotrocholina molesta, miliolids and Rivularia-like cyanobacteria. Foraminifers and calcareous algae recorded by Dragastan (2011) are not sufficiently diagnostic for such a detailed biostratigraphy, and calpionellids, which could be diagnostic for the age, were not illustrated.
The present study
describes the microfacies and microfossils characteristics of the Mesozoic
deposits from the Lapoș Valley, Bicazului
Gorges (Fig. 1 ![]() ). The main purpose was to
examine the Jurassic-Cretaceous
transition based on detailed sampling and the analysis of relatively-rich
micropaleontological assemblages.
). The main purpose was to
examine the Jurassic-Cretaceous
transition based on detailed sampling and the analysis of relatively-rich
micropaleontological assemblages.
|
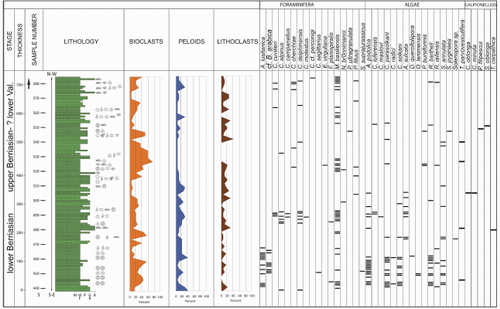
Figure 1: Geological map of the Hăghimaș Syncline (modified from Săndulescu, 1975) with location of the sampled section (right up - detail with the samples location).
|
Lapoș Valley (Fig. 1 ![]() )
is located in
the Hăghimaș
Massif or Hăghimaş syncline (Grasu et
al., 2012). The sedimentary succession from the Hăghimaș Mountains consists of
three distinct tectonic units (Săndulescu, 1967,
1984; Dragastan, 1971; Grasu et al.,
2010): the Bucovinian, the Subbucovinian and the Hăghimaș
Nappes (the last being part of the Transilvanian Nappes).
)
is located in
the Hăghimaș
Massif or Hăghimaş syncline (Grasu et
al., 2012). The sedimentary succession from the Hăghimaș Mountains consists of
three distinct tectonic units (Săndulescu, 1967,
1984; Dragastan, 1971; Grasu et al.,
2010): the Bucovinian, the Subbucovinian and the Hăghimaș
Nappes (the last being part of the Transilvanian Nappes).
The Bucovinian Nappes are important tectonic units of the inner part of the Eastern Carpathians. Their emplacement took place during the Early Cretaceous (Popescu-Voiteşti, 1929; Kräutner, 1980; Săndulescu, 1984). They represent the equivalent of the Getic Nappe from the Southern Carpathians (Săndulescu, 1984). These structures represent fragments of continental crust derived from the European Margin, along the Ceahlău-Severin oceanic rift (Schmid et al., 2008). The Bucovinian Nappe from the Hăghimaş Massif contains a Triassic-Lower Cretaceous sedimentary succession that overlies metamorphic basement. The final term is represented by a Barremian-Aptian "wildflysch" incorporating olistoliths derived from the Transylvanian (Hăghimaş) Nappes (Patrulius, 1969; Săndulescu, 1975, 1984; Ştefănescu, 1976). The Subbucovinian Nappes have a lower position in respect to the Bucovinian ones. They contain similar metamorphic and Triassic-Lower Cretaceous deposits. The Transylvanian Nappes overthrust the Bucovinian Nappes and contain a group of obduction nappes, consisting of sedimentary rocks and ophiolites (Săndulescu, 1984) occurring mostly as olistoliths in the Bucovinian "wildflysh". Their emplacement occurred during the Lower Cretaceous and was triggered by a series of tectonic events preceding the Lower Cretaceous post-tectonic cover (Schmid et al., 2008).
The Hăghimaș Nappe, as part of the Transylvanian nappes, represents a decollement nappe thrust over the Bucovinian Nappe from the west. Thrust was time equivalent to deposition of "wildflysh" in the Bucovinian Nappe. These tectonic processes ended probably in the upper Albian, and were followed by subsequent erosion (Săndulescu, 1975; Bucur et al., 2011). Terrigenous and carbonate Triassic, Jurassic and Lower Cretaceous carbonates (Săndulescu, 1975) form the sedimentary succession in the Bicaz Valley area of the Hăghimaş Mountains. The Triassic contains calcarenites, sandstones, marls, dolomites and cherty limestones (Săndulescu, 1975). Nodular limestones, sandy limestones with Saccoccoma sp., interbedded sandy limestones and marls define the basal Kimmeridgian. They pass upwards into interbedded marls and limestones (Săndulescu, 1975).
The Tithonian-Valanginian-? Hauterivian deposits consist of limestones and marly limestones (Săndulescu, 1975; Dragastan, 1975a). The Kimmeridgian-lower Tithonian marly limestones were dated on various ammonite associations (Platynota, Hypselocyclum, Divizum and Uhlandi zones for the lower Kimmeridgian; Acanthicum, Eudoxus, Beckeri zones for the upper Kimmeridgian; Hybonotum, Vimineus, Semiforme and Falauxi zones for the lower Tithonian) (Grigore, 2011). The ages assigned to the upper Tithonian-?Hauterivian carbonates were based on microfossil assemblages (Dragastan, 1971, 1975a, 2011; Săndulescu, 1975; Bucur et al., 2011).
Fieldwork was performed during the summer
of 2014. A total number of 340 samples was collected and used to prepare 350
thin sections. Sampling was performed at 3-4 m resolution, over a total distance
of two kilometers. The starting point is located fifty meters above the
confluence between the Lapoș and Bicaz rivers (Fig. 1 ![]() ). Microscope analysis was used to describe the microfacies and
microfossil associations. Classification of carbonate rocks follows Dunham
(1962), Embry and Klovan (1971), and Wright
(1992).
). Microscope analysis was used to describe the microfacies and
microfossil associations. Classification of carbonate rocks follows Dunham
(1962), Embry and Klovan (1971), and Wright
(1992).
The studied succession
follows the Lapoș
Valley over a total distance of approximately two kilometers (Fig. 1 ![]() , upper
right). Carbonate beds dip to more than 45 degrees, in areas where bedding
planes are conspicuous (Fig. 2.A-B
, upper
right). Carbonate beds dip to more than 45 degrees, in areas where bedding
planes are conspicuous (Fig. 2.A-B ![]() ). The succession is interrupted by a fault
which is located 700 m from the starting point of the section. This fault
separates two distinct segment that contain different facies characteristics.
Eight microfacies associations were separated from base to top (MFL 1-MFL 8).
). The succession is interrupted by a fault
which is located 700 m from the starting point of the section. This fault
separates two distinct segment that contain different facies characteristics.
Eight microfacies associations were separated from base to top (MFL 1-MFL 8).
|
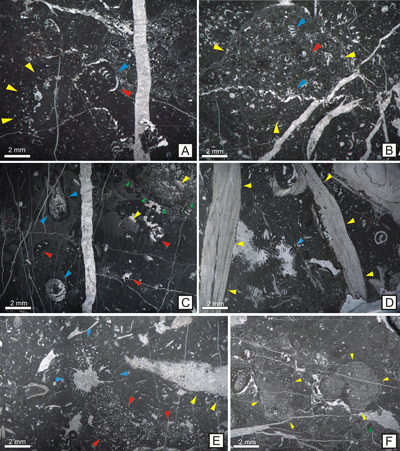
Figure 2:
A - Berriasian peloidal bioclastic
wackestone-packstone carbonates (base of Piatra Altarului); B-Massive Tithonian
reefal limestones with grainstone/rudstone facies types, reefal fragments,
dasycladalean algae and encrusting organisms in Cheile Lapoșului (Lapoş Gorges). |
MFL 1 - Peloidal
bioclastic wackestone (Fig. 3.A-C ![]() ;
Table 1)
;
Table 1)
Angular to well-rounded, abundant peloids
characterize these submicrofacies types. They are uniformly distributed within
the muddy sediment. Sometimes they form aggregated structures. They are
associated with dasycladalean algae (Fig. 3.B-C ![]() ), benthic
foraminifers
(Fig. 3.A
), benthic
foraminifers
(Fig. 3.A ![]() ) and poorly-preserved
calpionellids. Some of the peloids have a biotic
origin. They were produced by complete bioclast micritization. The grains are
coated by a micritic rim (Fig. 3.B-C
) and poorly-preserved
calpionellids. Some of the peloids have a biotic
origin. They were produced by complete bioclast micritization. The grains are
coated by a micritic rim (Fig. 3.B-C ![]() ), and the non-homogeneous matrix is
frequently bioturbated (Fig. 3.B
), and the non-homogeneous matrix is
frequently bioturbated (Fig. 3.B ![]() ). Rare fenestral structures are present.
Sometimes they have a laminar aspect with a well-defined orientation. Fenestrae
shape ranges from oval to circular, irregular or digitated with a planar/sinusoidal
base; they are filled with sparite or geopetal vadose silt (Fig.
3.C
). Rare fenestral structures are present.
Sometimes they have a laminar aspect with a well-defined orientation. Fenestrae
shape ranges from oval to circular, irregular or digitated with a planar/sinusoidal
base; they are filled with sparite or geopetal vadose silt (Fig.
3.C ![]() ).
).
MFL 2 - Bioclastic
oncoidal floatstone (Fig. 3.D-F ![]() ;
Table 1)
;
Table 1)
The bioclastic floatstone
facies is the most abundant subtype. It contains angular to subrounded
centimeter-sized bioclasts, including rudist fragments and other bivalves,
gastropods, coral fragments and echinoderm spines (Fig. 3.D ![]() ). These bioclasts
are encrusted by different organisms (Fig. 3.E
). These bioclasts
are encrusted by different organisms (Fig. 3.E ![]() ).
).
Oncoidal floatstone is
less frequent within this microfacies association. Micritic or porostromatic
oncoids are common. They present growth discontinuities and irregular, ovoidal
or spherical shapes. Bioclastic or lithoclastic cores are frequent. They are
commonly associated with encrusting organisms, benthic foraminifers and
dasycladalean algae. The internal sediment is a peloidal bioclastic wackestone
with benthic foraminifers and/or dasycladalean algae (Fig.
3.D-F ![]() ). Common grain
types include peloids, intraclasts and sometimes extraclasts (terrigenous
quartz). Peloids and intraclasts have variable shapes from angular to well
rounded. They are scattered through the muddy sediment. Angular to subangular
quartz-rich extraclasts are present. This facies association contains grains
with surficial micritic envelopes. They have a clastic or bioclastic origin.
). Common grain
types include peloids, intraclasts and sometimes extraclasts (terrigenous
quartz). Peloids and intraclasts have variable shapes from angular to well
rounded. They are scattered through the muddy sediment. Angular to subangular
quartz-rich extraclasts are present. This facies association contains grains
with surficial micritic envelopes. They have a clastic or bioclastic origin.
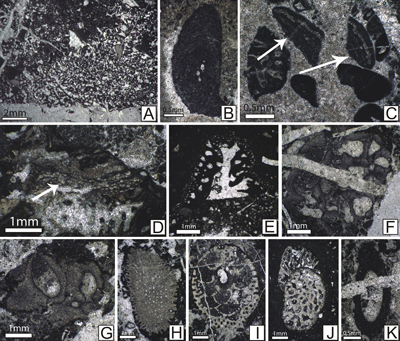
|
Figure 3: MFL 1-MFL 2 (Peloidal
bioclastic wackestone and bioclastic oncoidal floatstone). A: Peloidal wackestone with agglutinated
foraminifers; abundant peloids
with dimensions ranging from 0.1 to 0.8 mm; associated agglutinated foraminifers
(blue arrows), encrusting organisms attached to a lithoclast (yellow arrows) and
bivalve fragments (red arrows); the matrix is non-homogeneous and contains
micrite, microsparite and rare sparite (sample 453). B: Bioclastic wackestone with dasycladalean algae.
Skeletal grains consist of abundant dasycladalean algae (ex: Actinoporella
podolica, Clypeina parasolkani, C. solkani, Rajkaella bartheli) (blue
arrows) and agglutinated foraminifers (red arrows); the matrix is
non-homogeneous displaying bioturbation (yellow arrows); moderately fractured fabric with fractures containing sparite (sample
444). C: Bioclastic wackestone-floatstone with geopetal sediment;
large
fragments of dasycladalean algae (blue arrows) and benthic foraminifers (red
arrows) are present; the matrix is non-homogeneous and contains micrite,
microsparite and rare sparite; voids partially filled with geopetal structures
(yellow arrows) (sample 528). D:
Bioclastic floatstone with rudists; large bioclasts are represented by rudist
fragments (yellow arrows) and coral fragments (blue arrows); all these bioclasts
are encased in a bioclastic wackestone-type matrix with small peloids (sample
562). E: Bioclastic floatstone with microbial structures. It consists of
bioclastic fragments encrusted by different organisms (yellow arrows), microbial
structures (Bacinella sp., Lithocodium
sp.) (red and blue arrows), foraminifers and fragments of dasycladalean algae;
the grains are coated by a micritic rim; these bioclasts are hosted by a
peloidal wackestone matrix (sample 493). F: Microbial floatstone with Rivularia-like
cyanobacteria (yellow arrows); microbial organisms are abundant; they are hosted
by a fine, peloidal wackestone matrix; the green arrow indicate geopetal
structures (sample 712). |
MFL 3 - Peloidal
bioclastic lithoclastic packstone/wackestone (Fig. 4.A-C ![]() ;
Table 1)
;
Table 1)
The peloidal bioclastic
lithoclastic facies contains various amounts of grains. Peloids are the most
abundant whereas bioclasts and lithoclasts are subordinate. Peloid dimension
ranges from 0.1 to 0.8 mm. The peloidal fraction is disseminated within the
micritic/sparitic mass. Their shape ranges from subangular to subrounded (Fig.
4.A-C ![]() ). Bioclasts are represented by benthic
foraminifers
(Fig. 4.A-C
). Bioclasts are represented by benthic
foraminifers
(Fig. 4.A-C ![]() ),
fragments of calcareous algae, sponges (Fig. 4.A
),
fragments of calcareous algae, sponges (Fig. 4.A ![]() ), and mollusks
(Fig. 4.A-C
), and mollusks
(Fig. 4.A-C ![]() ).
Skeletal grains present a surficial micritic rim. Lithoclasts are scarce, with
subangular to subrounded shapes. They are mainly represented by intraclasts
which have the same composition as other carbonate grains. Peloids are uniformly
distributed within the peloidal bioclastic packstone-wackestone
submicrofacies type. They have small dimensions (less than 0.5 mm) are
associated with benthic foraminifers and calcareous algae, and have a moderate
to good sorting. Rare intraclasts have angular to subrounded shapes. Their size
exceeds 1 mm. Silt to sand grade, angular to subangular terrigenous extraclasts
(less than 0.5 mm in dimension) are common. They are disseminated within the
micritic matrix (Fig. 4.B
).
Skeletal grains present a surficial micritic rim. Lithoclasts are scarce, with
subangular to subrounded shapes. They are mainly represented by intraclasts
which have the same composition as other carbonate grains. Peloids are uniformly
distributed within the peloidal bioclastic packstone-wackestone
submicrofacies type. They have small dimensions (less than 0.5 mm) are
associated with benthic foraminifers and calcareous algae, and have a moderate
to good sorting. Rare intraclasts have angular to subrounded shapes. Their size
exceeds 1 mm. Silt to sand grade, angular to subangular terrigenous extraclasts
(less than 0.5 mm in dimension) are common. They are disseminated within the
micritic matrix (Fig. 4.B ![]() ). Ovoidal porostromatic oncoids
are moderately frequent. Their size sometimes exceeds 1 mm. Their core has a bioclastic
origin.
). Ovoidal porostromatic oncoids
are moderately frequent. Their size sometimes exceeds 1 mm. Their core has a bioclastic
origin.
Benthic foraminifers are common in the peloidal bioclastic packstone microfacies type. They are associated with peloids which have an average dimension of 0.3 mm. Intergranular pores contain micrite, microsparite and sparite.
MFL 4 - Bioclastic
peloidal lithoclastic grainstone-packstone (Fig. 4.D-G ![]() ;
Table 2)
;
Table 2)
Bioclastic peloidal
lithoclastic subtype is the most abundant. Peloids have various shapes and
dimensions, ranging from 0.1 to 1 mm. They are scattered through the rock mass,
being associated with various bioclasts. Microorganisms include benthic foraminifers, dasycladalean algae, bivalves and gastropods (Fig.
4.D ![]() ). Most of
the grains have a biotic origin. They have micritic rims and sometimes they
are completely micritised. Associated angular to subrounded lithoclasts have
dimensions which do not exceed 1 mm (Fig. 4.D
). Most of
the grains have a biotic origin. They have micritic rims and sometimes they
are completely micritised. Associated angular to subrounded lithoclasts have
dimensions which do not exceed 1 mm (Fig. 4.D ![]() ).
).
The microbreccia
submicrofacies type contains angular, submillimeter to ruditic fragments of
reworked intraclasts. Subrounded clasts are scarce. Their composition is
relatively homogeneous. This feature points to an intrabasinal source area. The
intraclasts contain coral fragments, Bacinella-like
structures, Rivularia-like
cyanobacteria and small peloids (Fig. 4.E ![]() ). They are encased in a sparitic
cement which may contain iron oxide pigmentations. Such structures may form the
base of Bacinella-like structures.
). They are encased in a sparitic
cement which may contain iron oxide pigmentations. Such structures may form the
base of Bacinella-like structures.
Peloids occupy all the
rock mass, within the peloidal grainstone microfacies subtype (Fig.
4.F ![]() ). They
are defined by good sorting and subrounded to well-rounded shapes and are
commonly associated with benthic foraminifers, dasycladalean algae and bivalve
fragments. These bioclasts are totally or partially micritised. A grain-supported
fabric is common, the intergranular pores being filled with sparite cement.
Another facies subtype is represented by coarse bioclastic grainstone. It
contains coral fragments, sponges, echinoderms, dasycladalean algae, benthic foraminifers, bivalve fragments, gastropods and encrusting organisms (Fig.
4.G
). They
are defined by good sorting and subrounded to well-rounded shapes and are
commonly associated with benthic foraminifers, dasycladalean algae and bivalve
fragments. These bioclasts are totally or partially micritised. A grain-supported
fabric is common, the intergranular pores being filled with sparite cement.
Another facies subtype is represented by coarse bioclastic grainstone. It
contains coral fragments, sponges, echinoderms, dasycladalean algae, benthic foraminifers, bivalve fragments, gastropods and encrusting organisms (Fig.
4.G ![]() ).
Skeletal grains have angular to subrounded shapes. Intergranular pores are
filled with sparitic cement.
).
Skeletal grains have angular to subrounded shapes. Intergranular pores are
filled with sparitic cement.
MFL 5 - Lithoclastic
bioclastic rudstone (Fig. 4.H-I ![]() ;
Table 2)
;
Table 2)
The first subfacies type
contains angular to rounded intraclasts. Some of these fragments have a
microbial origin. They are associated with peloids. Bioclasts are represented by
dasycladalean algae and mollusk fragments (Fig. 4.H ![]() ). Intergranular pores are
filled with sparite, micrite or vadose silt. Bioclastic rudstone with reefal
fragments is the most common submicrofacies type. It contains abundant sponge
fragments (Fig. 4.I
). Intergranular pores are
filled with sparite, micrite or vadose silt. Bioclastic rudstone with reefal
fragments is the most common submicrofacies type. It contains abundant sponge
fragments (Fig. 4.I ![]() ). Other bioclasts include encrusting organisms,
dasycladalean algae, echinoderm plates and various mollusk fragments (Fig.
4.I
). Other bioclasts include encrusting organisms,
dasycladalean algae, echinoderm plates and various mollusk fragments (Fig.
4.I ![]() ).
Intraclasts and extraclasts (subangular quartz fragments) are subordinate. Some
bioclasts are bordered by a surficial micritic rim. Intergranular pores are
filled with sparite or a combination of vadose silt and iron oxides.
).
Intraclasts and extraclasts (subangular quartz fragments) are subordinate. Some
bioclasts are bordered by a surficial micritic rim. Intergranular pores are
filled with sparite or a combination of vadose silt and iron oxides.
|
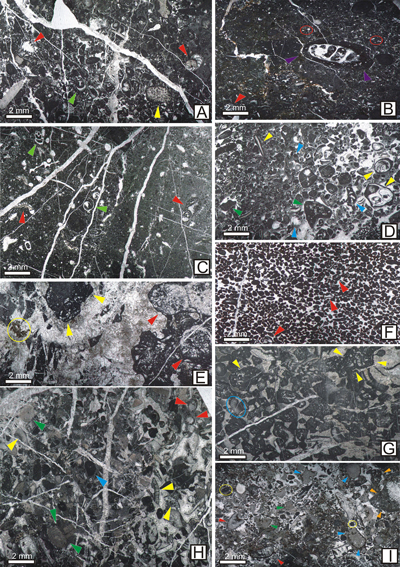
Figure 4:
MFL 3-MFL 5 (Peloidal
bioclastic lithoclastic packstone-grainstone, grainstone, lithoclastic
bioclastic rudstone). A: Peloidal bioclastic lithoclastic packstone-grainstone;
this microfacies contains diverse bioclasts, such as foraminifers (red arrows),
sponge fragments (yellow arrow), gastropod fragments (green arrows) and other
bioclasts; intergranular pores are filled with micrite, microsparite and
sparite;
moderately fractured fabric (sample 487). B: Peloidal bioclastic
packstone-wackestone; abundant peloids incorporate benthic foraminifers (red
arrow), algae fragments, porostromatic oncoids (purple arrows) and quartz
extraclasts (red circles); intergranular pores are filed with micrite and
microsparite; iron oxide pigmentations may occur (sample 578). C: Bioclastic
peloidal packstone with foraminifers; the fabric is dominated by bioclasts and
small peloids; red arrows indicate benthic foraminifers. Dasycladalean algae and
gastropod fragments (green arrows) are present; intergranular pores contain
micrite and microsparite (Sample 783). D: Bioclastic peloidal lithoclastic
grainstone-packstone; bioclasts are represented by fragments of dasycladalean
algae, benthic foraminifers (blue arrows), and mollusks (yellow arrows); peloids
and subangular to subrounded intraclasts are present (green arrows); the
intergranular pores contain equally proportioned sparite and micrite (sample
489). E: Microbrecciated lithoclastic grainstone. Intraclasts are abundant. They
consist of broken, angular fragments yellow arrows encased in a sparitic mass;
some of them exceed 2 mm in dimension; other bioclasts include coral fragments
and microbial organisms (red arrows); the fabric is pigmented with iron oxides
(yellow circle) (sample 641). F: Peloidal grainstone. Well-sorted peloids are
frequent. Coated grains are common (red arrows). The intergranular pores is
filled with sparry cement (sample 670). H: Coarse bioclastic grainstone with
encrusting organisms (Crescentiella morronensis)
(blue arrow), benthic foraminifers (red arrows), calcareous algae (yellow
arrows), echinoderm plates (green arrows); some of these coated grains are
bordered by a surficial micritic rim; small peloids are common and the
intergranular pores contain micrite, microsparite and sparitic cement (sample
603). G: Lithoclastic bioclastic rudstone with calcareous algae (blue circle),
mollusk fragments, microbial-bacinellid structures (yellow arrows) and small
peloids; intergranular pores are filled with sparry cement (sample 700). I:
Bioclastic rudstone; bioclasts are represented by encrusting organisms (Crescentiella
morronensis) (yellow circles), sponges (blue arrows), calcareous algae
(red arrow), echinoderm plates (green arrows). peloids and mm-sized intraclasts
(orange arrows); grains are coated by a micritic rim and the intergranular pores
contain sparite, silt and micrite (sample 594). |
MFL 6 - Microbial
bindstone (Fig. 5.A-B ![]() ;
Table 2)
;
Table 2)
Bacinella-like structures dominate the first subfacies type
(Fig. 5.A ![]() ). These
structures are associated with other microbial organisms and form a solid
framework that binds together all the carbonate clasts. Other bioclasts include
dasycladalean algae, corals, rudist and other mollusk fragments (Fig.
5.A
). These
structures are associated with other microbial organisms and form a solid
framework that binds together all the carbonate clasts. Other bioclasts include
dasycladalean algae, corals, rudist and other mollusk fragments (Fig.
5.A ![]() ).
Intergranular pores are filled with micrite and rare sparitic cement. A rare
subfacies type is represented by microbial bindstone with Rivularia-like
cyanobacteria and other encrusting organisms (Bacinella
type structures). This subtype contains abundant Rivularia-like
cyanobacteria. Small other bioclasts and peloids are associated. Intergranular
pores contain micrite and rare sparitic cement (Fig. 5.B
).
Intergranular pores are filled with micrite and rare sparitic cement. A rare
subfacies type is represented by microbial bindstone with Rivularia-like
cyanobacteria and other encrusting organisms (Bacinella
type structures). This subtype contains abundant Rivularia-like
cyanobacteria. Small other bioclasts and peloids are associated. Intergranular
pores contain micrite and rare sparitic cement (Fig. 5.B ![]() ).
).
MFL 7 - Coral boundstone (Fig. 5.C ![]() ;
Table 2)
;
Table 2)
Corals form a solid
framework which strengthens the rock structure. They are associated with
encrusting microorganisms [(Radiomura
sp., Crescentiella morronensis
(Crescenti)] (Fig. 5.C ![]() ). The clastic components consist of peloids
(smaller than 0.2 mm). Intergranular pores are filled with micrite and rare
sparitic cement.
). The clastic components consist of peloids
(smaller than 0.2 mm). Intergranular pores are filled with micrite and rare
sparitic cement.
MFL 8 - Algal framestone
with Neoteutloporella socialis
(Fig. 5.D ![]() ;
Table 2)
;
Table 2)
This microfacies type is
rare. It contains only one species of dasycladalean algae. This species in
growth position occupies the entire rock mass forming small "patch
reefs" (Fig. 5.D ![]() ).
).
|
Figure 5: MFL 6-8 (Bindstone,
coral boundstone, algal framestone with Neoteutloporella socialis).
A: Bindstone with Bacinella-like
structures and bioclasts; Bacinella
type structures are abundant (outlined by yellow lines) together with large
corals (green arrows), rudists (yellow arrows) and mollusks (blue arrows);
calcareous algae are present (red arrows); intergranular pores contain micrite
and rare sparry cement (sample 782). B: Microbial bindstone with Rivularia- type
cyanobacteria. It also contains Bacinella-like
structures and rare peloids; pores are filled with micrite; fenestral fabric
with sparite infillings (red circles) (sample 518). C: Coral boundstone. Corals
are dominant; rare encrusting organisms are present (Radiomura sp.-yellow circles, Crescentiella
morronensis-blue circles); rare small
peloids are present; the intergranular pores are filled with micrite and sparry
calcite (sample 667). D: Neoteutloporella
boundstone. occupy the entire rock mass. The original growth position can be
clearly observed. Intergranular pores are filled with sparite (sample 681). |
Dragastan (2011) presented new data on the stratigraphy and
micropaleontology of the Lapoş Valley section. The succession is considered
to be of Tithonian-Hauterivian age. The author (Dragastan, 2011) identified
several calpionellid zones, but did not illustrate them. He
also identified two discontinuity surfaces (lower Berriasian-upper Berriasian
and lower Valanginian-Hauterivian). Within the studied section a fault (near
sample 584) (Fig. 1 ![]() ) separates the upper Berriasian-? lower Valanginian deposits
from the upper Tithonian-lower Berriasian carbonates. The microfacies
distribution and micropaleontological data (Figs. 6
) separates the upper Berriasian-? lower Valanginian deposits
from the upper Tithonian-lower Berriasian carbonates. The microfacies
distribution and micropaleontological data (Figs. 6 ![]() ,
7
,
7 ![]() ,
8
,
8 ![]() ) will be presented in
stratigraphic order.
) will be presented in
stratigraphic order.
|
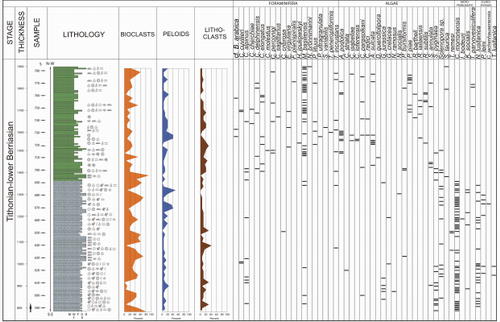
Figure 6: Lithological
column of section II (upper Tithonian - lower Berriasian). The quantitative
abundance of the most important carbonate components and the occurrence of the
most important microfossils is indicated. |
|
Figure
7: Lithological
column of section I (upper Berriasian - ? lower Valanginian). The quantitative
abundance of the most important carbonate components and the occurrence of the
most important microfossils is also indicated. |
|
Figure
8: General
stratigraphic occurence of the most important microfossil identified in the Lapoș
Valley section. |
This interval (samples
585-783) (Fig. 1 ![]() ) contains inner and platform margin deposits with variable
facies types. The first part contains rare boundstone / framestone microfacies
types, rudstone and grainstone with reefal fragments (MFL4, MFL5, , MFL7, MFL8).
The upper part of the interval passes towards restricted facies types with
abundant muddy subcategories (floatstone, packstone, wackestone or bindstone)
(MFL1, MFL2, MFL3, MFL 6). Coarse reefal detritus is present in the lower part
of the interval. It contains sponges, corals, red algae, dasycladalean algae,
benthic foraminifers and abundant, angular lithoclasts. By contrast, the upper
interval contains abundant peloids and microbial structures together with
dasycladalean algae, benthic foraminifers and coated grains (Fig. 6
) contains inner and platform margin deposits with variable
facies types. The first part contains rare boundstone / framestone microfacies
types, rudstone and grainstone with reefal fragments (MFL4, MFL5, , MFL7, MFL8).
The upper part of the interval passes towards restricted facies types with
abundant muddy subcategories (floatstone, packstone, wackestone or bindstone)
(MFL1, MFL2, MFL3, MFL 6). Coarse reefal detritus is present in the lower part
of the interval. It contains sponges, corals, red algae, dasycladalean algae,
benthic foraminifers and abundant, angular lithoclasts. By contrast, the upper
interval contains abundant peloids and microbial structures together with
dasycladalean algae, benthic foraminifers and coated grains (Fig. 6 ![]() ). The
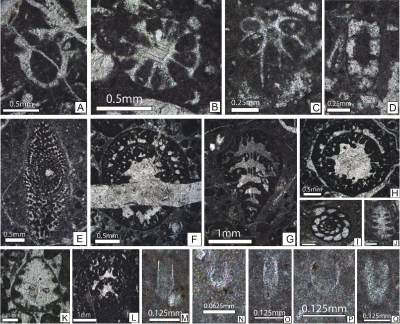
micropaleontological association consists of dasycladalean algae (Fig.
9.A-Q
). The
micropaleontological association consists of dasycladalean algae (Fig.
9.A-Q ![]() ;
Table 1), benthic foraminifers (Fig. 10.A-Q
;
Table 1), benthic foraminifers (Fig. 10.A-Q ![]() ;
Table 1) microproblematic organisms
(Fig. 11.A-G
;
Table 1) microproblematic organisms
(Fig. 11.A-G ![]() , arrows;
Table 1), sclerosponges
(Fig. 11.H-J
, arrows;
Table 1), sclerosponges
(Fig. 11.H-J ![]() , Table 1),
annelid
worm tubes (Fig. 11.K
, Table 1),
annelid
worm tubes (Fig. 11.K ![]() ), coral fragments, echinoderm plates, rudists and other
mollusks.
), coral fragments, echinoderm plates, rudists and other
mollusks.
In terms of biostratigraphy, some of the most representative species are dasycladalean algae [Campbelliella striata (Tithonian cf. Granier & Deloffre, 1993; upper Tithonian-lower Berriasian cf. Bucur et al., 2014), Neoteutloporella socialis (Kimmeridgian-Tithonian cf. Granier & Deloffre, 1993; Bassoullet, 1997; Bucur, 1999), Petrascula bursiformis (Kimmeridgian-Tithonian cf. Granier & Deloffre, 1993), Aloisalthella sulcata (Kimmeridgian-Berriasian cf. Bassoullet, 1997; Bucur, 1999; Kimmeridgian-middle Berriasian cf. Granier, 2019; Granier & Lethiers, 2019), Clypeina parasolkani (Berriasian cf. Farinacci & Radoičić, 1991; Berriasian-Valanginian cf. Bucur & Săsăran, 2005; Bruni et al., 2007; Tithonian-Berriasian cf. Schlagintweit, 2011), Otternstella lemmensis (upper Kimmeridgian-lower Berriasian cf. Granier & Deloffre, 1993), Rajkaella bartheli (Kimmeridgian-Berriasian cf. Granier & Deloffre, 1993; Bucur, 1999; Bucur et. al., 2013)], sponges [Neuropora lusitanica, Thalamopora lusitanica, and Calcistella jachenhausenensis (Tithonian cf. Reitner, 1992; Pleş et al., 2013; Kaya et al., 2015)], foraminifers [Charentia cuvillieri (Berriasian-Cenomanian cf. Neumann, 1965; Bucur et al., 1995; Schlagintweit & Wagreich, 2005), different representatives of the genus Coscinoconus (upper Tithonian-lower Valanginian cf. Arnaud-Vanneau et al., 1988; Neagu, 1994, 1995; Bucur et al., 1995; Bucur & Săsăran, 2005), Haplophragmoides joukowskyi (Berriasian-Hauterivian cf. Charollais et al., 1966; Altiner, 1991; Bucur et al., 1995; Ivanova, 2000), microproblematic organisms [Iberopora bodeuri (Berriasian cf. Granier & Berthou, 2002; Uță & Bucur, 2003; Oxfordian-Berriasian cf. Schlagintweit, 2004)] or the annelid worm Terebella lapilloides (Kimmeridgian-lower Berriasian cf. Kaya & Altiner, 2014). This association indicates an upper Tithonian-Berriasian age for this part of the succession. However, the exact boundary between these stages cannot be identified.
| Depositional environment | Inner platform restricted and high-energy settings |
| Lithofacies association | MFL1-MFL3 |
| Lithofacies | Peloidal bioclastic wackestone, bioclastic wackestone, bioclastic wackestone-floatstone, oncoidal floatstone, peloidal bioclastic packstone-wackestone, bioclastic peloidal packstone |
| Sedimentary structures | Micritic rims, bioturbation, surficial micritic envelopes, fenestral structures |
| Grain types | Angular to well rounded peloids, peloids of biotic origin, oncoids, porostromatic oncoids, intraclasts, extraclasts |
| Biota | Benthic foraminifers [Ammobaculites sp., Anchispirocyclina lusitanica (Egger), Kastamonina abanica Sirel, cf. Bramkampella arabica (Redmond), Charentia cuvillieri (Neumann), C. evoluta, Coscinoconus alpinus, C. campanellus (Arnaud-Vanneau et al.), C. cherchiae, C. delphinensis, C. elongatus, C. molestus, C. cf. perconigi, C. sagittarius (Arnaud-Vanneau et al.), Everticyclammina gr. hedbergi (Maync), E. virguliana, Freixialina planispiralis, Lenticulina sp., Mayncina sp., Mohlerina basiliensis, Nautiloculina broennimanni, Ichnusella infragranulata, Protopeneroplis ultragranulata, Pseudocyclammina lituus, Spiraloconulus suprajurassicus, Troglotella incrustans], algae [Actinoporella podolica, Arabicodium sp., Clypeina cf. loferensis, C. maslovi (Praturlon), C. parasolkani, C. solkani Conrad & Radoičić, Aloisalthella sulcata, Cylindroporella sp., Deloffrella quercifoliipora Granier & Michaud, Felixporidium sp., Holosporella sp., Otternstella lemmensis (Bernier), Permocalculus sp., Petrascula bursiformis, Rajkaella bartheli, R. iailaensis (Maslov), R. subtilis, Russoella sp., Salpingoporella annulata Carozzi, S. pygmaea, Suppiluliumella sp., "Solenopora" jurassica, Thaumatoporella parvovesiculifera], encrusting organisms (Bacinella irregularis, Lithocodium aggregatum, Rivularia sp.), sponges (Cladocoropsis sp.), calpionellids [Calpionellopsis simplex (Colom), Calpionella minuta (Houša), ?Precalpionellites filipescui (Pop), Sturiella oblonga (Borza), ?Tintinnopsella carpathica (Murgeanu & Filipescu)], rare calcispheres [Cadosina minuta (Borza)], and mollusk fragments. |
|
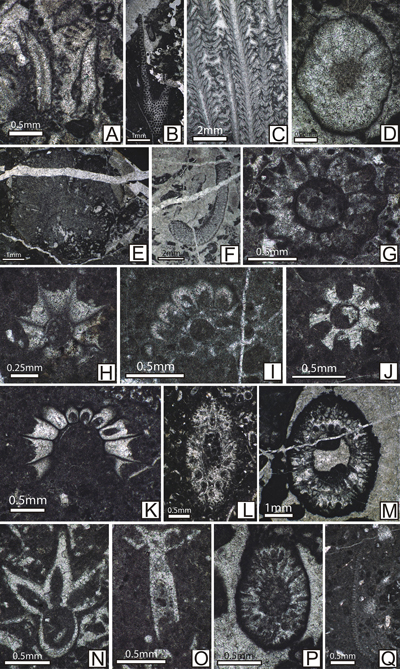
Figure
9: Upper
Tithonian-Berriasian calcareous algae from the Lapoș Valley. A: Campbelliella
striata (Carozzi) (s. [= sample] 618); B: Griphoporella cretacea (Dragastan) (s. 662); C: Neoteutloporella
socialis (Praturlon) (s. 681); D: Nipponophycus
ramosus (Yabe & Toyama) (s. 603); E:
"Solenopora" jurassica (Nicholson & Brown) (s. 598); F:
Triploporella remesi Steinmann (s. 650); G: Actinoporella
podolica (Alth) (s. 737); H: Clypeina
isabellae Masse et al. (s. 728); I: Clypeina
loferensis Schlagintweit et al. (s. 724); J:
Clypeina parasolkani Farinacci & Radoičić (s.
459); K: Aloisalthella sulcata (Alth)
(s. 777); L: Deloffrella quercifoliipora Granier
& Michaud (s. 710); M: Petrascula
bursiformis (Etallon) (s. 701); N: Rajkaella
bartheli (Bernier) (s. 449); O: Rajkaella
subtilis (Dragastan) (s. 726); P: Salpingoporella
pygmaea (Gümbel) (s. 701); Q: Thaumatoporella
parvovesiculifera (Raineri) (s. 767). |
|
Figure
10: Upper
Tithonian-Berriasian foraminifers from the Lapoș Valley. A: Charentia
cuvillieri Neumann (s. 491); B:
Coscinoconus alpinus Leupold
(s. 486); C: C. chiocchinii Mancinelli.
& Coccia (s. 610); D: C.
delphinensis (Arnaud-Vanneau et al.) (s. 711); E: C.
cherchiae (Arnaud-Vanneau et al.) (s. 716); F: C.
elongatus Leupold (s. 699); G: C.
perconigi Neagu (s. 746); H: Mohlerina
basiliensis (Mohler) (s. 487); I: Ichnusella
infragranulata (Noth) (s. 661); J: Troglotella
incrustans Wernli & Fookes (s. 778); K: Everticyclammina
virguliana Koechlin (s. 524); L: Freixialina
planispiralis Ramalho (s. 486); M: Haplophragmoides
joukowsky (Charollais et al.) (s. 722); N: Nautiloculina
broennimanni Arnaud-Vanneau & Peybernčs (s. 489); O: Protopeneroplis
ultragranulata (Gorbachik) (s. 547); P: Pseudocyclammina
lituus (Yokoyama) (s. 582); Q: Siphovalvulina
variabilis Septfontaine (s. 734). |
|
Figure
11: Upper Tithonian-lower Berriasian encrusting microorganisms and sponges from the Lapoș Valley. A: Bacinella
irregularis Radoičić (s. 643); B: Crescentiella
morronensis (Crestenti) (s. 685); C: Iberopora
bodeuri Granier & Berthou (s. 671, arrows); D: Koskinobulina
socialis Cherchi & Schroeder (s. 682, arrow); E: Lithocodium
aggregatum Elliott (p. 522); F: Perturbatacrusta
leini Schlagintweit & Gawlick (p. 617); G: Radiomura cautica Senowbari-Daryan
& Schäfer (s. 589); H: Neuropora
lusitanica G.
Termier et al. (s. 591); I: Thalamopora
lusitanica G.
Termier et al. (s. 616); J: Cladocoropsis
mirabillis Felix (s. 777); K:
Terebella lapilloides Münster
(s. 655). |
The succession
corresponding to this interval shares similar characteristics with the upper
part of the previously described succession. It contains inner platform facies
types with slight tendencies towards peritidal transitions (MFL1-MFL 3) (Fig. 7 ![]() ). Bioclastic fragments
(e.g., mollusks, dasycladalean algae, microbial
organisms, benthic foraminifers) dominate the clast spectrum. They are followed
by peloids and lithoclasts (intraclasts and extraclasts) (Fig. 7
). Bioclastic fragments
(e.g., mollusks, dasycladalean algae, microbial
organisms, benthic foraminifers) dominate the clast spectrum. They are followed
by peloids and lithoclasts (intraclasts and extraclasts) (Fig. 7 ![]() ). The
micropaleontological association consists of calcareous green algae [Fig.
12.A-D
). The
micropaleontological association consists of calcareous green algae [Fig.
12.A-D ![]() ;
Table 2], benthic foraminifers
[Fig. 12.E-L
;
Table 2], benthic foraminifers
[Fig. 12.E-L ![]() ;
Table 2], encrusting organisms and sponges (Table
2), calpionellids (Fig. 12.M-Q
;
Table 2], encrusting organisms and sponges (Table
2), calpionellids (Fig. 12.M-Q ![]() ;
Table 2), rare calcispheres and mollusk
fragments (Table 2).
;
Table 2), rare calcispheres and mollusk
fragments (Table 2).
| Depositional environment | High energy, inner and platform margin settings |
| Lithofacies association | MFL4-MFL8 |
| Lithofacies subtypes | Bioclastic lithoclastic grainstone/packstone, bioclastic grainstone, lithoclastic rudstone, bioclastic rudstone, boundstone, algal framestone |
| Sedimentary structures | Micritic rims, micritised bioclasts, subrounded clasts, grain supported fabric |
| Grain types | Peloids, reworked intraclasts, lithoclasts, extraclasts, skeletal grains |
| Biota | Benthic foraminifers [Ammodiscus sp., cf. Bramkampella arabica (Redmond), Charentia cuvillieri (Gorbachik), Coscinoconus alpinus (Leupold), C. chiocchinii (Mancinelli & Coccia), C. delphinensis (Arnaud-Vanneau et al.), C. cherchiae (Arnaud-Vanneau et al.), C. elongatus (Leupold), C. molestus (Gorbatchik), C. perconigi (Neagu), Coscinophragma cribrosa (Reuss), Lenticulina sp., Mohlerina basiliensis (Mohler), Ichnusella infragranulata (Noth), Reophax sp., Troglotella incrustans (Wernli & Fookes), Ammobaculites sp., Everticyclammina virguliana (Koechlin), Freixialina planispiralis Ramalho, Haplophragmoides joukowsky Charollais et al., Lenticulina sp., Mayncina sp., Nautiloculina broennimanni Arnaud-Vanneau & Peybernčs, Protopeneroplis ultragranulata (Gorbachik), Pseudocyclammina lituus (Yokoyama), Siphovalvulina variabilis Septfontaine], algae [Campbelliella striata (Carozzi), Griphoporella cretacea (Dragastan), G. jurassica (Endo), Neoteutloporella socialis (Praturlon), Nipponophycus ramosus Yabe & Toyama, "Solenopora" jurassica Nicholson & Brown, Triploporella remesi (Steinmann), Actinoporella podolica (Alth), Arabicodium sp., Clypeina isabellae Masse et al., C. loferensis Schlagintweit et al., C. parasolkani Farinacci & Radoičić, Aloisalthella sulcata (Alth), Deloffrella quercifoliipora Granier & Michaud, Linoporella sp., Neomeris sp., Permocalculus sp., Petrascula bursiformis (Etallon), Pseudotrinocladus piae (Dragastan), Rajkaella bartheli (Bernier), R. subtilis Dragastan, Salpingoporella pygmaea (Gümbel), Thaumatoporella parvovesiculifera (Raineri)], encrusting organisms [Bacinella irregularis Radoičić, Crescentiella morronensis (Crestenti), Iberopora bodeuri Granier & Berthou, Koskinobulina socialis Cherchi & Schroeder, Labes atramentosa Eliášova, Lithocodium aggregatum Elliott, Perturbatacrusta leini Schlagintweit & Gawlick, Pseudorothpletzella sp., Radiomura cautica Senowbari-Daryan & Schäfer] Rivularia-like cyanobacteria, sclerosponges (Calcistella jachenhausenensis Reitner, Neuropora lusitanica G. Termier et al., Thalamopora lusitanica G. Termier et al., Cladocoropsis mirabillis Felix), annelid worm tubes (Terebella lapilloides Münster), gastropods, and corals |
Some of these species
indicate the presence of Berriasian [algae: Clypeina
maslovi (Valanginian cf. Granier & Deloffre, 1993; Schindler
& Conrad, 1994; Berriasian-Hauterivian cf. Bucur et
al., 2000), C. solkani (upper Berriasian cf. Masse,
1993;
Malm-Albian,
cf. Granier & Deloffre, 1993; Berriasian-Barremian cf. Bucur
et al., 2000)], Tithonian-Berriasian [benthic foraminifers: Anchispirocyclina lusitanica
(Tithonian-Berriasian cf. Dragastan, 1975a; Sotak,
1989; Schlagintweit
et al., 2005; Tithonian-lower Berriasian cf. Granier,
2019), Kastamonina
abanica (Tithonian-Berriasian, cf. Schlagintweit, 2005]. The
calpionellid association characterizes the upper Berriasian (Calpionellopsis
Zone, Simplex Subzone, Pop, 1997). The entire association
characterizes the upper Berriasian-? lower Valanginian (Fig. 8 ![]() ).
).
|
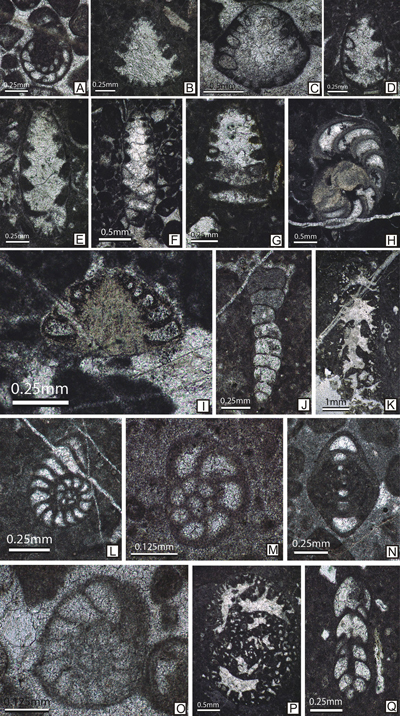
Figure
12: Upper Berriasian-? lower Valanginian microfossils from the Lapoș Valley. A: Clypeina
solkani Conrad & Radoičić (s. 443); B: Otternstella
lemmensis (Bernier) (s. 449); C: Rajkaella
iailaensis (Maslov) (s. 524); D: Salpingoporella
annulata Carozzi (s. 449); E: Anchispirocyclina
lusitanica (Egger) (s. 462); F, H: Kastamonina abanica Sirel (s.
456, 446); G: cf. Bramkampella arabica Redmond
(s. 456); I: Charentia cuvilieri Neumann (s. 563); J: C.
sagittarius (Arnaud-Vanneau et al.) (s. 743); K: Coscinoconus
campanellus (Arnaud-Vanneau et al.) (s. 491); L: Spiraloconulus
suprajurassicus Schlagintweit (s. 452); M: Calpionellopsis oblonga (Cadisch) (s.
505); N: Calpionella minuta Houša (s. 505); O: Sturiella
oblonga Borza (s. 551); P: ?Precalpionellites filipescui Pop
(s. 549); Q: ?Tintinnopsella carpathica (Murgeanu & Filipescu)
(s. 481). |
The studied section
follows an alignment which corresponds to the Lapoș Valley. It consists of two distinct successions
separated by a fault. The micropaleontological and microfacies association
points to an upper Tithonian-lower Berriasian age of the succession situated
above the fault. Upper Berriasian-? lower Valanginian deposits form the lower
part of the succession, below this fault. A preliminary analysis of the most
important facies types indicates that carbonate sediment was accumulating in two
major depositional settings: inner platform areas defined by interbedded
high- and low-energy deposits (upper Berriasian-? lower Valanginian) together
with platform margin areas (dominated by high-energy deposits) (upper
Tithonian-lower Berriasian) (Tables 1 -
2). The upper Tithonian-Berriasian clastic components are
characterized by poor sorting, variable roundness (ranging from angular to well
rounded). This part of the succession contains mainly platform margin high-energy deposits, with rare
interbedded low-energy carbonates. There is a
direct relationship between the existing biota and the environmental conditions.
The vast majority of the identified dasycladalean algae prefer such high energy,
platform margin depositional settings with reefs or bioclastic banks (e.g., Campbelliella
striata, Neoteutloporella socialis, Petrascula bursiformis) (Dragastan,
1975a; Schlagintweit & Ebli,
1999; Bucur et al.,
2005). In addition, the entire association of encrusting organism and calcareous
sponges characterizes such platform margin complexes that were strongly
developed at the Jurassic-Cretaceous transition (Pleş et al.,
2013; Mircescu et al., 2019). Terrigenous quartz fragments are
rare. All the sedimentological features point to a shallow-water, high-energy
depositional environment with abundant, diverse biota. Inner and margin platform
shallow-water carbonates characterize the Tithonian sedimentation and are partly
represented by bioclastic reefal facies types (Fig. 6 ![]() ). The overlying Berriasian
deposits probably accumulated in a regressive depositional context. Peloids,
lithoclasts and micrite make up and increasing proportion of the sediment
components. Grain dimensions are
smaller, their abundance decreases and sorting varies from poor to moderate.
Some of the inner platform deposits accumulated in restricted environments.
They are defined by a transition towards peritidal settings. These tendencies
are indicated by a decrease in the proportion of bioclasts associated with a slight
increase in the proportion of peloids. The upper Berriasian- ? lower Valanginian succession
contains interbedded, high- and low-energy, subtidal and intertidal deposits.
). The overlying Berriasian
deposits probably accumulated in a regressive depositional context. Peloids,
lithoclasts and micrite make up and increasing proportion of the sediment
components. Grain dimensions are
smaller, their abundance decreases and sorting varies from poor to moderate.
Some of the inner platform deposits accumulated in restricted environments.
They are defined by a transition towards peritidal settings. These tendencies
are indicated by a decrease in the proportion of bioclasts associated with a slight
increase in the proportion of peloids. The upper Berriasian- ? lower Valanginian succession
contains interbedded, high- and low-energy, subtidal and intertidal deposits.
The low-energy deposits from the inner platform areas are defined by the presence of abundant carbonate mud and a characteristic biota (Table 2). Such carbonate sediments accumulated under low-energy conditions in restricted, subtidal lagoons. The abundance of cyanobacteria nodules is typical for such restricted environments (Săsăran et al., 2013). In these conditions, micritisation processes were very active, under the action of endolithic bacteria (Bathurst, 1966). Some of the identified dasycladalean algae prefer such isolated, lagoonal environments. Clypeina loferensis was originally described by Schlagintweit et al. (2009) from such shallow-water subtidal lagoons. Salpingoporella annulata is frequently reported from similar depositional environments (Bucur & Săsăran, 2005; Mircescu et al., 2014). Another algae that prefers such lagoonal areas is Clypeina parasolkani (Husinec & Sokač, 2006; Schlagintweit et al., 2009). Pseudocyclammina lituus and Anchispirocyclina lusitanica are two species of foraminifers which are well known from similar deposits (Darga & Schlagintweit, 1991; Dya, 1992). Oncoids are frequent within low-energy subtidal areas. In this case, Bacinella type structures play a key role in forming such structures. The inner platform high-energy deposits were deposited probably in intertidal littoral areas. They contain abundant representatives of the genus Coscinoconus (e.g., C. alpinus, C. sagittarius, C. campanellus). Such foraminifers were described by various authors (Arnaud-Vanneau, 1980; Simmons, 1990) from this type of depositional environments.
The studied succession presents some peculiar characteristics. The low-energy deposits contain occasionally small percentages of calpionellids and calcispheres. Two explanations can be considered, either we have allodapic limestones (resedimented shallow-water carbonates in the basin with calpionellids), or that rare calpionellids were occasionally transported into the platform by storms. Taking into account the general aspect of the carbonate sediments from the succession studied the second hypothesis seems to be more appropriate. Isolated bioconstructions were developed as buildups (Burchette & Wright, 1992). The microfacies and micropaleontological associations are similar with previous data presented by various authors from other outcrop areas in Romania (Median Dacides) (Bucur, 1997; Pleş et al., 2013; Mircescu et al., 2014; Ungureanu et al., 2015; Grădinaru et al., 2016) and other parts of the Tethysian Realm (Gawlick et al., 2004; Rusciadelli & Rici, 2008; Gawlick & Schlagintweit, 2010; Petrova et al., 2011, 2012; Chatalov et al., 2015; Ivanova et al., 2015). They represent valuable correlation tools for future studies at the Jurassic-Cretaceous transition.
The authors would like to thank Daria Ivanova and Felix Schlagintweit for their valuable comments that helped improve the quality of the manuscript.
Altiner D. (1991).- Microfossil biostratigraphy (mainly foraminifers) of the Jurassic-Lower Cretaceous carbonate succession in north-western Anatolia (Turkey).- Geologica Romana, Roma, vol. 27, p. 167‒215.
Arnaud-Vanneau A. (1980).- Micropaléontologie, paléoécologie et sédimentologie d'une plate-forme carbonatée de la marge passive de la Téthys : L'Urgonien du Vercors septentrional et de la Chartreuse.- Thčse de Doctorat čs Sciences; Géologie Alpine, Grenoble, 3 volumes, 876 p.
Arnaud-Vanneau A., Boisseau T. & Darsac C. (1988).- Le genre Trocholina Paalzow 1922 et ses principales espčces au Crétacé.- Revue de Paléobiologie, Genčve, Volume spécial 2 (Benthos '86), p. 353-377.
Bassoullet J-P. (1997).- Les grands foraminifčres. In: Cariou E. & Hantzpergue P. (eds.), Biostratigraphie du Jurassique ouest-européen et Méditeranéen: Zonations parallčles et distribution de microfossiles.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Mémoires, vol. 17, p. 293-304.
Bathurst R.G.C. (1966).- Boring algae, micrite envelopes and lithification of molluscan biosparites.- Geology Journal, vol. 5, no. 1, p. 15-32.
Bruni R., Bucur I.I. & Préat A. (2007).- Uppermost Jurassic-Lower Cretaceous carbonate deposits from Fara San Martino (Maiella, Italy): Biostratigraphic remarks.- Studia UBB Geologia, Cluj Napoca, vol. 52, no. 2, p. 45-54.
Bucur I.I. (1997).- Representatives of the genus Protopeneroplis (foraminifera) in the Jurassic and Lower Cretaceous deposits in Romania. Comparisons with other regions of the Tethyan area.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 1, p. 65-74.
Bucur I.I. (1999).- Stratigraphic significance of some skeletal algae (Dasycladales, Caulerpales) of the Phanerozoic.- Palaeopelagos Special Publication, Roma, vol. 2, p. 53-104.
Bucur I.I., Conrad M.A. & Radoičić R. (1995).- Foraminifers and calcareous algae from the Valanginian limestones in the Jerma River Canyon, Eastern Serbia.- Revue de Paléobiologie, Genčve, vol. 14, no. 2, p. 349- 377.
Bucur I.I., Granier B & Krajewski M. (2014).- Calcareous algae, microbial structures and microproblematica from Upper Jurassic-Lowermost Cretaceous limestones of Southern Crimea.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 10, no. 1-2, p. 61-86.
Bucur I.I., Hoffmann M. & Kolodziej B. (2005).- Uppermost Jurassic-Lowermost Cretaceous benthic algae from Tethys and the European Platform. A case study from Poland.- Revista Espańola de Micropaleontologia, Madrid, vol. 37, no. 1, p. 105-129.
Bucur I.I., Koch R., Kirmaci Z.M. & Tasli K. (2000).- Les algues dasycladales du Crétacé inférieur (Calcaire de Berdiga) de Kirkaova (région de Kale-Gumushane, NE Turkey).- Revue de Paléobiologie, Genčve, vol. 19, p. 435-463.
Bucur I.I. & Săsăran E. (2005).- Micropaleontological assemblages from the Upper Jurassic-Lower Cretaceous deposits of Trascău Mountains and their biostratigraphic significance.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 5, p. 27-38.
Bucur I.I. & Săsăran E. (2011).- Upper Jurassic-Lower Cretaceous algae of Hăghimaș Mountains (Lacul Roșu-Cheile Bicazului area). In: Bucur I.I. & Săsăran E. (eds.), Calcareous algae from Romanian Carpathians.- Field Trip Guidebook, 10th International Symposium on Fossil Algae, Cluj University Press, Cluj Napoca, p. 137.
Bucur I.I., Săsăran E., Lazăr I., Dragastan O.N. & Popa M.E. (2011).- Mesozoic deposits of the Dâmbovicioara Couloir. In: Bucur I.I. & Săsăran E. (eds.), Calcareous algae from Romanian Carpathians.- Field Trip Guidebook, 10th International Symposium on Fossil Algae, Cluj University Press, Cluj Napoca, p. 23-31.
Bucur I.I., Pascariu L. & Săsăran E. (2013).- Calcareous algae from the olistoliths at Poiana Zănoaga northern Piatra Craciului Syncline (Southern Carpathians, Romania).- Berichte der Geologischen Bundesanstalt, Vienna, vol. 99, p. 108-109.
Burchette T.P. & Wright V.P. (1992).- Carbonate Ramp Depositional Systems.- Sedimentary Geology, vol. 79, p. 3-57.
Chatalov A., Bonev N. & Ivanova D.K. (2015).- Depositional characteristics and constraints on the mid-Valanginian demise of a carbonate platform in the intra-Tethyan domain, Circum-Rhodope belt, northern Greece.- Cretaceous Research, vol. 55, p. 84-115.
Charollais J., Broennimann P. & Zaninetti L. (1966).- Troisičme note sur les foraminifčres du Crétacé inférieur de la région genevoise. Remarques stratigraphiques et description de Pseudotextulariella salevensis, n. sp.; Haplophragmoides joukowskyi, n. sp.; Citaella? favrei, n. sp.- Archives des Sciences, Genčve, vol. 19, no. 1, p. 23-48.
Darga R. & Schlagintweit F. (1991).- Mikrofazies, Paläontologie und Stratigraphie der Lerchkogelkalke (Tithon-Berrias) des Dietrichshorns (Salzburger Land, Nördliche Kalkalpen).- Jahresberichte der Geologischen Bundesanstalt, Wien, vol. 134, no. 2, p. 225-226.
Dragastan O. (1969).- Algues calcaires du Jurassique supérieur et du Crétacé inférieur de Roumanie.- Revue de Micropaléontologie, Paris, vol. 12, no. 1, p. 53-62.
Dragastan O. (1971).- New Algae in the Upper Jurassic and Lower Cretaceous in the Bicaz Valley, East Carpathians.- Revista Espańola de Micropaleontologia, Madrid, vol. 3, no. 2, p. 155-192.
Dragastan O. (1975a).- Microfacies of Malm and Lower Cretaceous in the Gorges of Bicaz area. Guide micropaléontologique du Mésozoďque et du Tertiaire des Carpates Roumaines.- 14th European Micropaleontology Colloquium, Institut de Géologie et Géophysique, Bucharest, p. 123-128.
Dragastan O. (1975b).- Upper Jurassic and Lower Cretaceous microfacies from the Bicaz Valley basin (East Carpathians).- Mémoires de l'Institut de Géologie et Géophysique, Bucarest, vol. 21, p. 1-87.
Dragastan O. (1981).- Mesozoic Dasycladaceae from Romania: Distribution and biostratigraphical importance.- Facies, Berlin, vol. 4, p. 165-196.
Dragastan O. (2011).- Early Cretaceous Foraminifera, algal nodules and Calpionellids from the Lapoş Valley, Bicaz Gorges (Eastern Carpathians, Romania).- Analele Științifice ale Universității "Al. I. Cuza" din Iași Seria Geologie, Iași, vol. 57, no. 1, p. 91-113.
Dunham R.J. (1962).- Classification of sedimentary rocks according to depositional structure. In: Ham W.E. (ed.), Classification of carbonate rocks.- A.A.P.G. Memoir, Tulsa, no. 1, p. 235-239.
Dya M. (1992).- Mikropaleontologische und fazielle Unterschungen in Oberjura zwischen Salzburg und Lofer.- PhD Thesis, University of Berlin, 137 p.
Embry A.F. & Klovan J.E. (1971).- A Late Devonian reef tract on Northeastern Banks Island.- Bulletin of Canadian Petroleum Geology, Alberta, vol. 19, p. 730-781.
Farinacci A. & Radoičić R. (1991).- Late Jurassic-Early Cretaceous Dasycladales (Green Algae) from the western Pontides, Turkey.- Geologica Romana, Roma, vol. 27, p. 135-165.
Flügel E. (2010).- Microfacies of carbonate rocks, analysis, interpretation and application.- Springer Verlag, Berlin, 976 p.
Gawlick H.J. & Schlagintweit F. (2010).- The drowning sequence of Mount Bürgl in the Salzkammergut area (Northern Calcareous Alps, Austria): Evidence for a diachronous Late Jurassic to Early Cretaceous drowning of the Plassen Carbonate Platform.- Austrian Journal of Earth Sciences, Vienna, vol. 103, no. 1, p. 58-75.
Gawlick H.J., Schlagintweit F., Ebli O. & Suzuki H. (2004).- Die Plassen Formation (Kimmeridgium) des Krachstein (Steirisches Salzkammergut, Östereich) und ihre Unterlagerung: Neue Daten zur Facies, Biostratigraphie und Sedimentologie.- Zentralblatt für Geologie und Paläontologie, Stuttgart, Teil 1 (2003), vol. 3/4, p. 295-334.
Granier B. (2019).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition. In: Granier B. (ed.), VSI: Jurassic-Cretaceous Transition.- Cretaceous Research, vol. 93, p. 245-274.
Granier B. & Berthou P.Y. (2002).- Algues calcaires fossiles, nouvelles ou peu connues, du Portugal. In: Bucur I.I. & Filipescu, S. (eds.), Research advances in calcareous algae and microbial carbonates.- Proceedings of the 4th IFAA Regional Meeting, Cluj University Press, Cluj Napoca, p. 117-126.
Granier B. & Deloffre R. (1993).- Inventaire critique des algues dasycladales fossiles IIe partie - Les algues dasycladales du Jurassique et du Crétacé.- Revue de Paléobiologie, Genčve, vol. 12, no. 1, p. 19-65.
Granier B. & Lethiers A. (2019).- Aloisalthella, a new genus of fossil Polyphysacean green algae (Chlorophyta, Dasycladales), with notes on the genus Clypeina (Michelin, 1845).- Palaeontologia Electronica, no. 22.2.45, p. 1-20. URL: https://doi.org/10.26879/923
Grasu C., Miclăuş C., Brânzilă M. & Baciu D.S. (2010).- Hăghimaşului Mountains. Geological and phisico-geographical monography.- Editura Universităţii "Alexandru Ioan Cuza", Iaşi, 434 p [in Romanian].
Grasu C., Miclăuş C., Brânzilă M. & Baciu D.S. (2012).- Tulgheș-Hășmaș-Ciuc Mesozoic Syncline. Geological Monography.- Editura Universității "Alexandru Ioan Cuza", Iași, 250 p. [in Romanian].
Grădinaru M., Lazăr I., Bucur I.I., Grădinaru E., Săsăran E., Ducea M.N. & Andrăşanu A. (2016).- The Valanginian history of the eastern part of the Getic Carbonate Platform (Southern Carpathians, Romania): Evidence for emergence and drowning of the platform.- Cretaceous Research, vol. 66, p. 11-42.
Grigore D. (2010a).- Idoceratinae (Presimoceras, Trenerites and Lessiniceras genera) from "Acanthicum beds" of the Haghimaş Mts. (the Eastern Carpathians, Romania).- Muzeul Olteniei Craiova-Oltenia, Studii şi comunicări, Științele Naturii, Craiova, vol. 26, no. 1, p. 287-295.
Grigore D. (2010b).- Idoceratinae (Idoceras and Nebrodites genera) from "Acanthicum beds" of the Haghimaș Mts. (The Eastern Carpathians, Romania).- Muzeul Olteniei Craiova-Oltenia, Studii şi comunicări, Științele Naturii, Craiova, vol. 26, no. 2, p. 303-314.
Grigore D. (2011).- Kimmeridgian-Lower Tithonian ammonite assemblages from Ghilcoş-Hăghimaş massif (Eastern Carpathians, Romania.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 7, p. 177-189.
Herbich F. (1866).- Eine geologische Excursion von Balán an den Vöröstó, nach Békas, Zsedánpatak etc.- Verhandlungen und Mitteilungen des Siebenbürgischen Vereins für Naturwissenschaften zu Hermannstadt, Sibiu, Jahrgang XVII, no. 10, p. 217-230. URL: https://www.biodiversitylibrary.org/item/109980#page/479/mode/1up
Husinec A. & Sokač B. (2006).- Early Cretaceous benthic associations (foraminifera and calcareous algae) of a shallow tropical-water platform environment (Mijet Island, southern Croatia).- Cretaceous Research, vol. 27, no. 3, p. 418-441.
Ivanova D. (2000).- Middle Callovian to Valanginian microfossil biostratigraphy in the west Balkan Mountain, Bulgaria (SE Europe).- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 2, p. 231-238.
Ivanova D., Bonev N. & Chatalov A. (2015).- Biostratigraphy and tectonic significance of lowermost Cretaceous carbonate rocks of the Circum-Rodope Belt (Chalkidiki Peninsula and Thrace region, NE Greece).- Cretaceous Research, vol. 52, p. 25-63.
Jekelius E. (1921).- Der mittlere und obere Jura in Gebiet des Hăghimaşul Mare in Siebeburgen.- Bulletin de la Section Scientifique de l'Académie Roumaine, Bucharest, vol. 7, no. 10, p. 1-127.
Kaya M. Y & Altiner D. (2014).- Terebella lapilloides Münster, 1833 from the Upper Jurassic-Lower Cretaceous İnaltı carbonates, northern Turkey: Its taxonomic position and paleoenvironmental-paleoecological significance.- Turkish Journal of Earth Sciences, Ankara, vol. 23, p. 166-183.
Kaya M. Y. & Altiner D. (2015).- Microencrusters from the Upper Jurassic-Lower Cretaceous İnaltı Formation (Central Pontides, Turkey): Remarks on the development of reefal/peri-reefal facies.- Facies, Berlin, vol. 61, no. 4.
Kräutner H.G. (1980).- Lithostratigraphic correlation of Precambrian of the Romanian Carpathians.- Anuarul Institutului de Geologie şi Geofizică, Bucureşti, vol. 57, p. 229-296.
Lazăr I., Panaiotu C.E., Grigore G., Sandy M.R. & Peckmann J. (2011).- An unusual brachiopod assemblage in a Late Jurassic (Kimmeridgian) stromatactis mud-mound of the Eastern Carpathians (Hăghimaş Mountains), Romania.- Facies, Erlangen, vol. 57, no. 4, p. 627-647.
Masse J.P. (1993).- Early Cretaceous Dasycladales biostratigraphy from Provence and adjacent regions (South of France, Switzerland, Spain). A reference for Mesogean correlations. In: Barattolo F., De Castro P. & Parente M. (eds.), Studies on fossil benthic algae.- Bolletino de la Societa Paleontologica Italiana, Modena, Special Volume 1, p.311-324.
Mircescu C.V., Bucur I.I. & Săsăran E. (2014).- Dasycladalean algae from Upper Jurassic-Lower Cretaceous limestones of Piatra Craiului Massif (South Carpathians, Romania) and their relationship to palaeonvironment.- Studia UBB Geologia, Cluj Napoca, vol. 59, no. 1-2, p. 5-27.
Mircescu C.V., Bucur I.I., Săsăran E., Pleş G., Ungureanu R. & Oprişa A. (2019).- Facies evolution of the Jurassic-Cretaceous transition in the Eastern Getic Carbonate Platform, Romania: Integration of sequence stratigraphy, biostratigraphy and isotope stratigraphy.- Cretaceous Research, vo. 99, p. 71-95.
Neagu Th. (1994).- Early Cretaceous Trocholina group and some related genera from Romania Part I.- Revista Espańola de Micropaleontologia, Madrid, vol. 26, no. 3, p. 117-143.
Neagu Th. (1995).- The Cretaceous Trocholina group and some related genera from Romania.- Revista Espańola de Micropaleontologia, Madrid, vol. 27, no. 2, p. 5-40.
Neagu Th. & Neagu M. (1995).- Smaller agglutinated foraminifera from the acanthicum Limestone (Upper Jurassic), Eastern Carpathians, Romania. In: Kaminski M.A., Geroch S. & Gasinski M.A. (eds.), Proceedings of the Fourth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, no. 3, p. 21 1-225.
Neumann M. (1965).- Contribution ŕ l'étude de quelques Lituolidés du Cénomanien de l'Île Madame (Charente-Maritime).- Revue de Micropaléontologie, Paris, vol. 8, no. 2, p. 90-95.
Neumayr M. (1873).- Die Fauna der Schichten mit Aspidoceras acanthicum.- Abhandlungen der kaiserlich-koeniglichen geologischen Reichsanstalt, Wien, vol. 5, no. 6, p. 141-257.
Patrulius D. (1969).- Geologia Masivului Bucegi și a Culoarului Dâmbovicioara.- Editura Academiei Republicii Socialiste România, București, 321 p.
Petrova S., Lakova L. & Ivanova D.K. (2011).- Berriasian-Valanginian boundary in Bulgaria.- Review of the Bulgarian Geological Society, Sofia, vol. 72, no. 1-3, p. 91-97.
Petrova S., Rabrenovic D., Lakova I., Koleva-Rekalova E., Ivanova D.K., Metodiev L. & Malesevic N. (2012).- Biostratigraphy and microfacies of the pelagic carbonates across the Jurassic-Cretaceous boundary in eastern Serbia (Stara Planina-Poreč Zone).- Geologica Balcanica, Sofia, vol. 41, no. 1-3, p. 53-76.
Pleş G., Mircescu C.V., Bucur I.I. & Săsăran E. (2013).- Encrusting micro-organism and microbial structures in Upper Jurassic limestones from the southern Carpathians (Romania).- Facies, Berlin, vol. 59, no. 1, p. 19-48.
Pop G. (1997).- Tithonian to Hauterivian praecalpionellids and calpionellids bioevents and biozones.- Mineralia Slovaca, Bratislava, vol. 29, no. 4-5, p. 304-305.
Popescu-Voiteşti I. (1929).- Aperçu synthétique sur les structures des régions carpatiques.- Revista Muzeului de Geologie şi Mineralogie, Universitatea Cluj, vol. 1, p. 1-40.
Popescu D.A. & Popescu L.G. (2005).- The Olenekian carbonates of the Bucovinian nappe (the central sector of the Hăghimaş syncline, Eastern Carpathians): Lithology and microfacies.- Studia UBB Geologia, Cluj Napoca, vol. 50, no. 1-2, p. 53-62.
Preda I. (1973).- Variațiile de facies și biostratigrafia Jurasicului superior din Munții Hăghimaș.- Studii și Cercetări de Geologie, Geografie și Biologie, Seria Geologie Geografie, Piatra Neamț, vol. 2, p. 11-21.
Reitner J. (1992).- "Coralline Spongien" Der Versuch einer phylogenetisch-taxonomischen Analyse.- Berliner Geowissenschaftliche Abhandlungen, vol. 1, p. 1-352.
Rusciadelli G. & Ricci C. (2008).- New geological constraints for the extension of the northern Apulia platform margin west of the Maiella Mt. (Central Apeninnes, Italy).-Italian Journal of Geosciences, Roma, vol. 127, p. 375-387.
Săndulescu M. (1967).- La nappe de Hăghimaş - une nouvelle nappe de décollement dans le Carpates Orientales.- Association géologique Carpato-Balkanique, VIIčme Congrčs, Belgrade, Rapports géotectoniques, p. 179-185.
Săndulescu M. (1968).- Probleme tectonice ale sinclinalului Hăghimaş.- Dări de Seamă ale Institutului Geologic şi Geofizic, Bucureşti, vol. 53, no. 3 ( for 1965- 1966), p. 221-240.
Săndulescu M. (1969).- Structura geologică a părţii centrale a sinclinalului Hăghimaş.- Dări de Seamă ale Institutului Geologic şi Geofizic, Bucureşti, vol. 65, no. 3 (for 1966-1967), p. 228-250.
Săndulescu M. (1975).- Studiul geologic al părţii centrale şi nordice a sinclinalului Hăghimaş (Carpaţii Orientali).- Anuarul Institutului Geologic şi Geofizic, Bucureşti, vol. 65, p. 1-160.
Săndulescu M. (1984).- Romania's geotectonics.- Editura Tehnică, Bucureşti, 336 p [in Romanian].
Săsăran E., Pleş G., Mircescu C.V. & Bucur I.I. (2013).- Peritidal cyclical sequences of Kimmeridgian - Berriasian - ? Valanginian limestones from Piatra Craiului Massif (Romania); the role of microbialites and rivulariacean-type cyanobacteria. In: Proceedings of the 11 th Workshop on Alpine Geological Studies & 7th IFAA, Schladming-Dachstein (Austria).- Abstracts Volume, Berichte Geologische B.A., Vienna, p. 116- 117.
Schindler U. & Conrad M.A. (1994).- The Lower Cretaceous Dasycladales from the northwestern Friuli platform and their distribution in chronostratigraphic and cyclostratigraphic units.- Revue de Paléobiologie, Genčve, vol. 13, no. 1, p. 59-96.
Schlagintweit F. (1991).- On the occurrence of the genus Permocalculus Elliott, 1955 (Calcareous algae, Gymnocodiaceae) in the Upper Cretaceous of the Northern Calcareous Alps (Branderfleck Formation, Gosau Formation) with the description of Permocalculus gosaviensis n. sp.- Revue de Paléobiologie, Genčve, vol. 10, no. 1, p. 37-46.
Schlagintweit F. (2004).- Iberopora bodeuri Granier & Berthou 2002 (incertae sedis) from the Plassen Formation (Kimmeridgian-Berriasian) of the Tethyan Realm.- Geologia Croatica, Zagreb, vol. 57, p. 1 1-13.
Schlagintweit F. (2005).- Neogyroporella ? gawlicki n.sp., a new Dasycladale from the Upper Jurassic-Lower Cretaceous "Lärchberg Formation" of the Northern Calcareous Alps, Austria.- Geologia Croatica, Zagreb, vol. 58, no. 2, p. 103-117.
Schlagintweit F. (2011).- The dasycladalean algae of the Plassen Carbonate Platform (Kimmeridgian-Early Berriasian): Taxonomic inventory and palaeogeographical implications within the platform-basin-system of the Northern Calcareous Alps (Austria, p.p. Germany).- Geologia Croatica, Zagreb, vol. 64, no. 3, p. 185-206.
Schlagintweit F., Dieni I. & Radoićić R. (2009).- Two look-alike dasycladalean algae: Clypeina isabellae Masse, Bucur, Virgone & Delmasso, 1999 from the Berriasian of Sardinia (Italy) and Clypeina loferensis sp. n. from the upper Jurassic of the Northern Calcareous Alps (Austria).- Geoloski anali Balkanskog poluostrva, Beograd, vol. 70, p. 43-59.
Schlagintweit F. & Ebli O. (1999).- New results on microfacies, biostratigraphy and sedimentology of Late Jurassic-Early Cretaceous Alps, Part I: Tressenstein Limestone, Plassen Formation.- Geologie ohne Grenzen Festschrift 150 Jahre Geologische Bundesanstalt, Vienna, vol. 56, no. 2, p. 379-418.
Schlagintweit F., Gawlick H.-J. & Lein R. (2005).- Mikropaläontologie und Biostratigraphie der Plassen-Karbonatplattform der Typlokalität (Ober-Jura bis UnterKreide, Salzkammergut, Österreich)/Micropaleontology und biostratigraphy of the Plassen carbonate platform of the type locality (Upper Jurassic to Lower Cretaceous, Salzkammergut, Austria).- Journal of Alpine Geology (Mitteilungen der Gesellschaft der Geologie- und Bergbaustudenten in Österreich), Vienna, vol. 47, p. 11-102
Schlagintweit F. & Wagreich M. (2005).- Micropaleontology of "Orbitolina Beds" of Lower Austria (Branderfleck Formation, Lower Cenomanian).- Jahrbuch der geologischen Bundesanstalt, Vienna, vol. 145, no. 1, p. 115-125
Schmid S.M., Bernoulli D., Fugenschuch B., Matenco L., Schefer R., Schuster R., Tischler M. & Ustaszewski K. (2008).- The Alpine-Carpathian-Dinaridic orogenic system: Correlation and evolution of tectonic units.- Swiss Journal of Geosciences, Basel, vol. 101, p. 139-183.
Simmons M.D. (1990).- Aspects of the micropaleontology and stratigraphy of Cretaceous shelf carbonates from the Oman Mountains.- PhD Thesis, University College of Wales, Swansea, 270 p.
Sotak J. (1989).- Nálezy spodnokriedových zástupcov čelade Pfenderinidae Smout et Sugden (Foraminiferida) vo valúnoch flyšových zlepenkov vonkajšich západných Karpat.- Zbornik z paleontologickei konferencie, Geologicky ustav Dioniza Stura, p. 37-44.
Ștefănescu M. (1976).- O noua imagine a structurii flișului intern din regiunea de curbură a Carpaților.- Dări de seamă ale şedinţelor - Institutul de Geologie şi Geofizică, București, vol. LXII, no. 5, p. 257-259 [in Romanian].
Turculeţ I. (1980).- Acanthicum beds from Mount Ghilcoș (Hăghimaș) - an exceptional paleontological reserve.- Anuarul Muzeului Judeţean Suceava, Ştiinţele Naturii, Suceava, vol. 6, p. 79-87 [in Romanian].
Uță A. & Bucur I.I. (2003).- Microbial structures and microencrusters in the Upper Jurassic-Lower Cretaceous deposits from Buila-Vanturarita Massif (South Carpathians).- Studia Universitatis Babeş-Bolyai, Geologia, Cluj-Napoca, vol. XLVIII, no. 2, p. 3-14.
Ungureanu R., Săsăran E., Bucur I.I., Ungur C.G. & Mircescu C.V. (2015).- The Berriasian- Valanginian and Aptian deposits from the north-western part of the Piatra Craiului Massif: Stratigraphic relationships, facies and depositional environments.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 11, no. 2, p. 59-74.
Vadasz E. (1915).- Geologische Beobachtungen im Persanyer und Nagyhagymas Gebirge.- Mitteilungen aus dem Jahresbericht der koeniglichen ungarischen geologischen Reichsanstalt, Budapest, p. 264-298.
Wright V.P. (1992).- A revised classification of limestone.- Sedimentary Geology, vol. 76, p. 177-186.